Embed Size (px)
Citation preview

Marine seaweed invasions Impacts and biotic resistance in native ecosystems
Josefin Sagerman

II
© Josefin Sagerman, Stockholm University 2015
Illustrations by the author
Cover photo: Codium fragile ssp. tomentosoides in a red algal
meadow of cf. Bonnemaisonia hamifera, © Joakim Hansen
ISBN 978-91-7649-176-8
Printed in Sweden by Holmbergs, Malmö 2015
Distributor: Department of Ecology, Environment and Plant Sciences

III
Till Farmor

IV
Populärvetenskaplig sammanfattning
Marina makroalger (som t.ex. tång, kelp eller rödslick) utgör grunden i ett av
de mest produktiva ekosystemen på jorden, grunda vegetationsklädda klipp-
bottnar. Makroalgerna spelar en nyckelroll i näringsflödet vid kusten och
utgör viktiga barnkammare för fisk och kräftdjur. Mänskliga aktiviteter är
ofta intensiva i kustområden, vilket utsätter de marina ekosystemen för på-
frestningar. Detta har orsakat storskaliga förändringar i var havets växter och
djur kan leva. En av de mest iögonfallande av dessa förändringar är intro-
duktion av arter till nya regioner utanför deras naturliga utbredningsområde,
så kallade främmande arter.
Målsättningen med den här avhandlingen har varit 1) att studera hur främ-
mande makroalger påverkar ekosystemfunktioner så som produktivitet och
nedbrytning av vissnade växtdelar och 2) att studera hur samspelet mellan en
främmande makroalg och inhemska växter och djur kan påverka om främ-
mande arter blir framgångsrika. Jag har använt mig av en kombination av
experiment utförda i laboratoriemiljö, i akvarier i utomhus, samt ute i havet.
Artikel I och II visar att de två främmande rödalgerna japanplym (Hetero-
siphonia japonica) och japantofs (Bonnemaisonia hamifera) har mycket
olika effekt på produktivitet och nedbrytning i invaderade algsamhällen, på
Svenska västkusten. Japanplymen hade en mycket stark effekt på produktivi-
teten på grund av artens höga tillväxttakt. Under experimentets gång produ-
cerade japanplymen nio gånger mer biomassa än de inhemska algerna, när
arterna växte var och en för sig. När japanplymen växte tillsammans med
inhemska alger ökade samhällets biomassa med upp till fem gånger jämfört
med algsamhällen med enbart inhemska alger. Japantofs hade i motsats till
japanplym, en relativt långsam tillväxt och påverkade därmed inte produkti-
viteten i rödalgssamhället. Däremot bröts japantofsen ned betydligt lång-
sammare än de inhemska algerna. Det kan bero på att japantofsen innehåller
ämnen (bl.a. bromoheptanon) som gör den oaptitlig för inhemska växtätare
och förhindrar bakteriell tillväxt. Snabb tillväxt och kemiskt försvar mot
växtätare är antagligen viktiga faktorer som kan förklara varför japanplymen
och japantofsen har kunnat bli så framgångsrika i sitt nya utbredningsområde
i Europa. Mina resultat visar att egenskaper som är förknippade med inva-
sionsframgång även kan avgöra vilka effekter främmande arter får på eko-
systemet där de etablerar sig. Förändringar i ekosystemfunktioner kan vara

V
av stor betydelse eftersom levande och död algbiomassa utgör basen i en
omfattande födoväv, och hastigheten av produktion och nedbrytning påver-
kar flödet av näring genom algsamhället. Japanplymen och japantofsen
kommer båda från nordvästra Stilla havet och finns nu spridda längs hela
Europas atlantkust.
Artikel III visar att även den snabbväxande japanplymen undviks som föda
av inhemska växtätare, vilket sannolikt gör det möjligt för arten att överleva
under årets kallare månader när den inte kan växa så snabbt. Att japanply-
men är oaptitlig beror på artens kemiska egenskaper, men det var inte möj-
ligt att avgöra om orsaken var näringssammansättning eller kemiskt försvar.
Artikel IV visar att den begränsade invasionsframgången hos den främman-
de brunalgen ishavstång (Fucus evanescens) åtminstone delvis kan förklaras
av att den är en sämre konkurrent än inhemska alger. Ishavstången växte
dåligt när den var omgiven av den inhemska släktingen blåstång (Fucus ve-
siculosus). Växtätande strandsnäckor (Littorina obtusata) orsakade betydligt
större skador på blåstången än på den oaptitliga ishavstången, men detta gav
inte ishavstången någon fördel i konkurrensen med blåstång. Resultaten från
min studie pekar på att konkurrens från inhemska makroalger har förmågan
att reducera framgången för främmande makroalger. För att minska risken
för att främmande makroalger får stor spridning kan det därmed vara viktigt
att skydda inhemska makroalger från påfrestningar så som miljögifter och
övergödning.
Sammantaget visar den här avhandlingen att främmande makroalger skiljer
sig markant i hur de påverkar ekosystemfunktioner. Kunskap om vilka egen-
skaper som är vanliga bland framgångsrika främmande arter och hur dessa
egenskaper är länkade till olika typer av effekter kan hjälpa oss att få en bätt-
re förståelse för invasionseffekter i marin miljö. Avhandlingen visar även att
konkurrens från inhemska arter kan ha en viktig begränsande inverkan på
invasionsframgång för främmande makroalger.

VI

VII
Contents
Contents ..................................................................................................... VII
List of papers ............................................................................................ VIII
Introduction .................................................................................................. 1 Impacts by non-native seaweeds ........................................................................... 1 Biotic resistance in seaweed communities ............................................................ 3 Aim ............................................................................................................................... 4
Scientific methods ....................................................................................... 5 Study area .................................................................................................................. 5 Study species ............................................................................................................. 5 Mesocosm and field experiments ........................................................................... 8
Invasion effects on community production (Paper I) .................................. 8 Invasion effects on decomposition (Paper II) .............................................. 8 Effects of herbivory and competition on invader success (Paper IV) ... 9
Laboratory assays ..................................................................................................... 9 Survival of juvenile isopods (Paper I) ............................................................ 9 Feeding assays (Paper III) ............................................................................ 10
Results and discussion ............................................................................. 11 Impacts on ecosystem processes ......................................................................... 11 Biotic resistance ....................................................................................................... 14
Conclusions and future perspectives ..................................................... 17
Acknowledgements ................................................................................... 18
References .................................................................................................. 19
Thank you ................................................................................................... 26

VIII
List of papers
The thesis is based on the following papers, referred to in the text by their
roman numerals:
I Sagerman J, Enge S, Pavia H, Wikström
SA (2014). Divergent eco-
logical strategies determine different impacts on community produc-
tion by two successful non-native seaweeds. Oecologia 175:937-
946.©2014 Springer
II Sagerman J, Staberg N, Pavia H, Wikström SA. Decomposition
rates differ between two seaweed invaders with different
ecophysiological traits. (manuscript)
III Sagerman J, Enge S, Pavia H, Wikström
SA. Low feeding prefer-
ence of native herbivores for the successful non-native seaweed
Heterosiphonia japonica. (submitted)
IV Sagerman J, Pavia H, Wikström SA. Competition between the non-
native Fucus evanescens and the native congener Fucus vesiculosus.
(manuscript)
Paper I is reprinted in the thesis with permission by Springer Sci-
ence+Business Media.
My contributions to the papers were: Participating in the planning of the
experimental design (I), in charge of planning the experimental design (II &
III), carrying out the experiments (I–IV), the main part of data analysis and
writing the papers (I–IV).
Additional publication by the author of this thesis:
Hansen JP, Sagerman J, Wikström SA (2010) Effects of plant morphology
on small-scale distribution of invertebrates. Marine Biology 157:2143–2155

1
Introduction
Marine seaweed systems are among the most productive ecosystems on the
planet (Mann 1973). They have a key function in nutrient cycling within
temperate costal zones and as a habitat, seaweeds harbor a rich diversity of
fauna and serve as a nursery for commercially important fish (Rangeley and
Kramer 1995; Costanza et al. 1998). Human activities are intense in coastal
areas and marine communities are under pressure from a multitude of stress-
ors, which has resulted in large scale changes in the abundance and distribu-
tion of species (Lotze et al. 2006; Worm et al. 2006).
Species introductions to new areas outside of their natural ranges belong to
one of the most conspicuous of these changes (Grosholz 2002; Lotze et al.
2006; Worm et al. 2006). A large number of seaweeds have been introduced
to new areas, mostly as a result of accidental introductions through transpor-
tation of ballast water in the ocean trade, or import of organisms for aquacul-
ture (Williams and Smith 2007). The majority of seaweeds that have suc-
ceeded to establish in new areas only have a limited occurrence in the receiv-
ing ecosystem (Nyberg and Wallentinus 2005), but a few have become
abundant and dominate invaded communities. These highly successful in-
vaders have the potential to cause strong ecological effects, such as suppres-
sion of native species and alterations of ecosystem functions (Schaffelke and
Hewitt 2007; Vye et al. 2014).
Impacts by non-native seaweeds
Non-native seaweeds may cause ecological impact at different organization-
al level, i.e. impacts ranges from the level of individual organisms to the
scale of ecosystem processes. The most commonly reported impact is dis-
placement of native seaweeds and reduced native seaweed abundance (re-
viewed by Schaffelke and Hewitt 2007). A wide variety of mechanisms ena-
ble non-native primary producers to gain competitive advantage over native
species, such as preemption, allelopathy, apparent competition, alterations of

2
disturbance regimes and resource depletion (Caldwell et al. 1987; Gordon et
al. 1989; Britton-Simmons 2004; Callaway and Ridenour 2004; Coffman et
al. 2010; Enge et al. 2013; Svensson et al. 2013). For instance, the intro-
duced brown alga Sargassum muticum reduces the growth of native under-
story vegetation due to its high shading canopy (Britton-Simmons 2004;
White and Shurin 2011). Another example is the non-native green alga
Codium fragile ssp. tomentosoides, which displaces native kelp through
preemption of space, after grazing sea urchins have created barrens in the
kelp forest (Scheibling and Gagnon 2006). Non-native seaweeds may also
have negative effects on species richness (Schaffelke and Hewitt 2007),
which is in accordance with the general view that species invasions are a
threat to diversity. This view has, however, been challenged (Davis 2003).
Most studies of invasion impacts on species diversity are conducted at a
local scale and are focused on the effects of highly successful invaders. But
the negative impacts on species richness decreases with increasing spatial
scale (Powell et al. 2011). Further, Sax and Gaines (2003) points out that on
a regional scale non-native species have instead increased species diversity.
The authors are giving a more comprehensive picture by including all non-
native species (of which most will not have a negative impact on species
diversity even at local scale). Thus, introductions of non-native seaweeds
affect diversity, but the effect probably differs depending on the spatial
scale.
When native vegetation is displaced by a non native species, ecosystem
functions such as primary productivity and decomposition may be altered
(reviewed by Liao et al. 2008). These impacts may be profound, since living
and dead seaweed biomass is the base of extensive food webs (Mann 1973;
Mann 1988), and the rate at which primary biomass is produced and subse-
quently broken down affects the flux of nutrients through the community
(Cebrian 1999; Moore et al. 2004; Gessner et al. 2010). Invasion impacts on
ecosystem functions have primarily been studied on vascular plants, while
there are few studies of the subject from the seaweed system. The difference
in research focus between the plant and the seaweed system is obvious when
comparing review-studies from the two systems. Williams and Smith (2007)
report six cases (single response variable in single study) where the effect of
a non-native seaweed on a ecosystem function has been tested. As a contrast
Vilà et al. (2011) reports 368 corresponding cases from invaded vascular
plant communities (despite making a considerably more narrow data selec-
tion). The number of studies of ecosystem impacts by seaweed invaders has

3
increased since the review by Williams and Smith from four to about ten.
But the knowledge is still very limited and to a large part based on observa-
tional studies, which impede the possibility to draw general conclusions.
Meta-analyses from terrestrial environment show that leaf traits are the most
important determinants of both productivity and decomposition rates in plant
communities (Wright et al. 2004; Cornwell et al. 2008). Thus, species traits
can be expected to affect the rate of productivity and decomposition in sea-
weed communities too, and abundant invaders with traits that differ from
those of native species will have potential to impact on ecosystem processes.
Biotic resistance in seaweed communities
Researchers have spent large efforts trying to explain why some non-native
species tend to dominate invaded communities, while others only persist in
small numbers in the new range. It has become well established that compe-
tition and consumption exerted by native species has the potential to limit
the success of non-native species, which is termed ‘biotic resistance’ (Elton
1958; Levine et al. 2004; Mitchell et al. 2006). In seaweed communities
there is an intense competition for limited resources such as space, light and
nutrients (reviewed by Carpenter 1990). Strict competitive hierarchies fre-
quently determine the distribution of native seaweeds (Chapman and John-
son 1990) and several studies show that the establishment of non-native
seaweeds are facilitated when native seaweeds are removed (Ambrose and
Nelson 1982; Levin et al. 2002; Valentine and Johnson 2003). Thus, compet-
itive interactions in seaweed communities have the potential to affect in-
vaders. However, few studies have examined the role of competition in re-
stricting invasion success of species that do not become very abundant and
dominant.
Herbivory could also potentially decrease the success of non-native sea-
weeds. The grazing pressure exerted by generalist herbivores is high in ma-
rine seaweed communities (Poore et al. 2012) and can have large effects on
native seaweed species composition (Duffy and Hay 2000). However, herbi-
vores may facilitate the invasion by unpalatable non-native seaweeds if the
herbivores cause more damage to palatable native competitors (e.g. Bulleri
2009; Enge et al. 2013). Low palatability is common among non-native sea-
weeds (e.g. Tomas et al. 2011; Enge et al. 2012; Hammann et al. 2013), but

4
there are still few studies that have tested whether this defense releases the
non-native species from competition in the new range.
Aim
The general aims of this thesis were to test (1) if the effect of non-native
seaweeds on ecosystem functions differs between species with different eco-
logical traits and (2) how biotic interactions with native species affects a
non-native seaweed that has not become abundant or dominant in the new
range. The specific aims have been to:
test how net-primary production and litter decomposition rates are
affected by two highly successful non-native seaweeds;
Bonnemaisonia hamifera and Heterosiphonia japonica (Paper I-II)
test the palatability of H. japonica to native herbivores and investi-
gate possible mechanisms for herbivore preferences (Paper III)
test how intra- and interspecific competition interacts with
herbivory to affect growth of the non-native Fucus evanescens and
the native congener Fucus vesiculosus (Paper IV)

5
Scientific methods
This chapter describes the study area and the model organisms (Fig. 1). It
also provides an overview of the experimental methods used in the thesis.
More detailed descriptions of the methods are given in each paper.
Study area
The studies were conducted at the Tjärnö Laboratory (Sven Lovén Center for
Marine Sciences) on the Swedish Skagerrak coast (58°52’N, 11°08’E). The
Tjärnö archipelago is characterized by a high proportion of rocky shores, a
surface salinity that fluctuates between 15-30 PSU and a small tidal variation
(0.1-0.3 m). The low salinity in the area is caused by the flow of brackish
water from the Baltic Sea to the Atlantic Ocean. The zone of fucoid sea-
weeds is typically submerged due to the low tidal variation. Below the fu-
coid zone there is generally a kelp zone, which is in turn followed by exten-
sive red algal communities often dominated by filamentous seaweeds.
Study species
Heterosiphonia japonica is a filamentous red alga that originates from the
coastal areas of northern China, Korea and Japan (Bjærke 2004). The species
was first observed outside of its native range in Brittany, France in 1984
(Sjøtun et al. 2008). Heterosiphonia japonica has since then rapidly spread
along the European Atlantic coastline and is now present from Portugal to
Norway, as well as in the Mediterranean Sea and on the American east coast
(Sjøtun et al. 2008; Schneider 2010). The introduction of the species to Eu-
rope is likely a result of import of aquaculture material from Korea or Japan
(Sjøtun et al. 2008). A large amount of seaweeds have been transferred from
the North Western Pacific to Europe in this manner by growing on the shells
of oysters (Williams and Smith 2007). In 2002, H. japonica reached the
Swedish west coast (Axelius and Karlsson 2004) and is now an abundant

6
component in the red algal communities in the area. The species often domi-
nates in terms of surface cover on sheltered rock bottoms and gravel (Husa et
al. 2004; Moy and Christie 2012). Heterosiphonia japonica has an efficient
asexual reproduction through fragmentation and the spread within the intro-
duced range is likely mediated by boat traffic (Husa and Sjøtun 2006; Sjøtun
et al. 2008). Besides these details little has previously been known of factors
that explain the success of the species. Further, there is only one previous
study that has addressed ecological effects of H. japonica on native commu-
nities. It is a study by Husa et al. (2008) that compared native seaweed di-
versity before and after the species had established on the Norwegian coast.
The study detected no impact by the H. japonica, but effects by the species
on for instance, ecosystem processes have not been addressed so far.
Bonnemaisonia hamifera is another non-native filamentous red alga that
shares the area of origin with H. japonica. However, B. hamifera has a con-
siderably longer history within Europe. The species was first observed on the
south coast of England in the 1890s (Maggs and Stegenga 1998).
Bonnemaisonia hamifera was probably transferred to Europe in ballast water
or as a fouling organism on a ship hull (Maggs and Stegenga 1998). The
species is now widespread in Europe and has become one of the most abun-
dant seaweeds in the Skagerrak region (Thomsen et al. 2007; Moy and
Christie 2012). It grows side by side with H. japonica in sheltered to semi
sheltered areas (Moy and Christie 2012). The reason why B. hamifera has
become so successful in the invaded range is most likely due to that the in-
vader produces potent brominated defense compounds (Enge et al. 2012).
The chemical defense deters herbivores and reduces the growth and survival
of seaweed propagules and microbes that settles on its surface (Nylund et al.
2005; Enge et al. 2012; Svensson et al. 2013). Bonnemaisonia hamifera is
also favored by having an intricate morphology, which serves as a refuge to
herbivorous crustaceans that feed on more palatable native neighboring sea-
weeds (Enge et al. 2013). However, ecological impacts of B. hamifera have
not previously been studied.
Fucus evanescens differs in several ways from the other two non-native
seaweeds in this thesis. The species is a brown alga that belong to the
fucoids, canopy-forming seaweeds that form ecologically important commu-
nities on shallow rock bottoms (Chapman 1995). Fucus evanescens has a
circumpolar distribution and is native to the north Atlantic, north Pacific and
the Arctic Ocean (Powell 1957a). In the north-western Atlantic, the native

7
range stretch over the Faeroes, Iceland and northern Norway (north of
Trondheim fjord). The species was introduced to the Oslo fjord in 1890s,
probably by shipping, and has since spread southward (Simmons 1898;
Wikström et al. 2002). Fucus evanescens is now present in the British Isles,
southern Norway, the Swedish west coast, and in the western Baltic Sea
(Powell 1957b; Schueller and Peters 1994; Wikström et al. 2002). In the
introduced range Fucus evanescens has a modest distribution and primarily
occurs in areas that are under human influence, such as boat harbors, where
the native fucoids are growing sparingly. The species is therefore not ex-
pected to have any major ecological effects on native communities
(Wikström 2004). Fucus evanescens is known to be of low palatability to
native herbivores; a common trait among successful, high impact invaders in
seaweed communities (e.g. Gollan and Wright 2006; Monteiro et al. 2009;
Nejrup et al. 2012). However, it is so far unknown how interactions with
native biota affect the success of F. evanescens.
Fig. 1 Top left; Bonnemaisonia hamifera, bottom left; Heterosiphonia japonica and to the right; young frond of Fucus evanescens.

8
Mesocosm and field experiments
Three mesocosms and one field experiment were used to 1) test how H. ja-
ponica and B. hamifera affect net primary production and decomposition
rates in invaded red algal communities, 2) explore how herbivory and com-
petition interacts to affect the invasion success of F. evanescens.
Invasion effects on community production (Paper I)
An outdoor mesocosm experiment was set up to separately test for the effect
of H. japonica and B. hamifera on community production. A replacement
design was used (all replicates had the same starting biomass) and the same
experimental treatments where applied for both invaders, i.e. i) a mix of
three native red algae, ii) a mix of two natives and one invader and iii) a mix
of three natives and one invader. The first treatment represents a pristine
seaweed community. The second and third treatment represents two possible
outcomes of invasion; that an invader displaces a native species or that an
invader adds to the species richness at local scale. The seaweeds were at-
tached to negatively buoyant plastic mesh and each synthetic community
was added to a 1.5 L bucket supplied with a constant flow of seawater
(n=12). The herbivorous isopod Idotea granulosa was added to half of the
replicates at natural density. The duration of the experiment was three
weeks.
Invasion effects on decomposition (Paper II)
The decomposition of three native red algae and the two non-native sea-
weeds (B. hamifera and H. japonica) was studied in an outdoor mesocosm
system. A repeated measures design was used with seaweed species and time
(decomposition after ten, sixteen and twenty days) as factors (n = 10 per
species and sampling time). Decomposition of the different seaweed species
were studied separately in different aquaria. One replicate mesh bag with
decomposing seaweeds was sampled from each aquarium at each sampling
time. Besides the replicate mesh bags, each aquarium contained a natural
decomposer community (i.e. 1.5 L of sediment and 0.6 L decomposing sea-
weed litter with associated detritivore fauna) gathered in the field.

9
Effects of herbivory and competition on invader success
(Paper IV)
The separate and interactive effects of competition and herbivory on the
growth of the two Fucus species were tested in a mesocosm experiment.
Three competition regimes were established for each species: i) no competi-
tion (one single frond of F. evanescens/F. vesiculosus), ii) intraspecific com-
petition (one frond of F. evanescens/F. vesiculosus with conspecific fronds)
and iii) interspecific competition (one frond of F. evanescens/F. vesiculosus
with fronds of the other Fucus species). In addition, two levels of herbivory
were applied in a factorial design; no herbivores and herbivory of Littorina
obtusata. In addition, a field experiment was set up to test the effect of com-
petition in the presence of a natural herbivore community. It included the
same competition regimes as the mesocosm experiment. Each treatment in
every experiment was replicated five times.
Laboratory assays
Four laboratory assays were performed to 1) explore the quality of H. japon-
ica and B. hamifera as food to native isopods, and explain a reduced survival
in monocultures of B. hamifera in the primary production experiment, and 2)
to test the palatability of and the presence of a potential grazer deterrent in
H. japonica.
Survival of juvenile isopods (Paper I)
To explore the growth and survival of juvenile I. granulosa on a diet of H.
japonica, B. hamifera and four native seaweeds, fronds of each species were
placed in individual plastic beakers filled with 80 ml filtered sea water
(n=20). A control treatment without food (but with a plastic mesh for the
isopods to cling to) was also included. One juvenile I. granulosa was intro-
duced to each beaker. The isopods were photographed at the start and end of
the experiment, to measure their body length, using the picture analysis
software ImageJ (Abramoff et al. 2004). The animals were monitored daily
and dead individuals were recorded. The experiment was terminated after 35
days when only one individual in the control treatment was still alive.

10
Feeding assays (Paper III)
The three herbivores I. granulosa, Gammarus locusta and Gammarellus
angulosus were allowed to feed on H. japonica and four native seaweeds, in
pairwise choice experiments. For each comparison, one shoot of H. japonica
and a native alga were placed in 200-ml containers filled with seawater. One
of the herbivore species (3-8 individuals depending on size and species), was
added to half of the containers. The other half of the containers was used as
controls for autogenic changes in the algae. The experiment was replicated
10 times for each algal pair and grazer species. The experiment was termi-
nated and the remaining algal pieces re-weighed when 25–50 % of the algal
biomass was eaten or at the latest when 48 h had passed.
To test whether the feeding preference of the generalist grazers was affected
by differences in physical structure between the algae, and to test for the
presence of a chemical grazer deterrent in H. japonica, similar experiments
were set up with homogenized algae in agar disks. For the test of a chemical
deterrent, extract from H. japonica was applied to a powder of the palatable
seaweed Ceramium virgatum that was incorporated in agar disks.

11
Results and discussion
Impacts on ecosystem processes
To explore invasion effects on ecosystem functions I used the two successful
non-native red algae; Heterosiphonia japonica and Bonnemaisonia
hamifera. In paper I, I found that H. japonica increased community net
primary production when included in mixed cultures with native seaweeds,
while B. hamifera had no effect (Fig. 2). Neither herbivory nor whether the
invader replaced a native species or added to the species diversity, changed
the effect of the two invaders.
Fig. 2 Biomass change of red algal communities [means ± 95 % confidence intervals
(CI95)] grown for three weeks in a polyculture experiment with introduction of
Heterosiphonia japonica or Bonnemaisonia hamifera, with and without grazing by
the crustacean Idotea granulosa. The experiment had three levels of introduction: no
introduction with three native seaweeds (NATIVE 3), introduction where the non-
native seaweed randomly replaced a native seaweed (INTRO 2+1), and introduction
where the non-native seaweed was present in addition to the native seaweeds (IN-
TRO 3+1), n = 10-12. The data set with H. japonica as the invader were square root
transformed and converted to normal scale. Means with a mutual letter are not sig-
nificantly separated in pairwise comparisons (Tukey´s HSD test). Native seaweeds
in the experiment were Ceramium virgatum, Brongniartella byssoides, Lomentaria
clavellosa and Spermothamnion repens. (Figure from paper I)
12
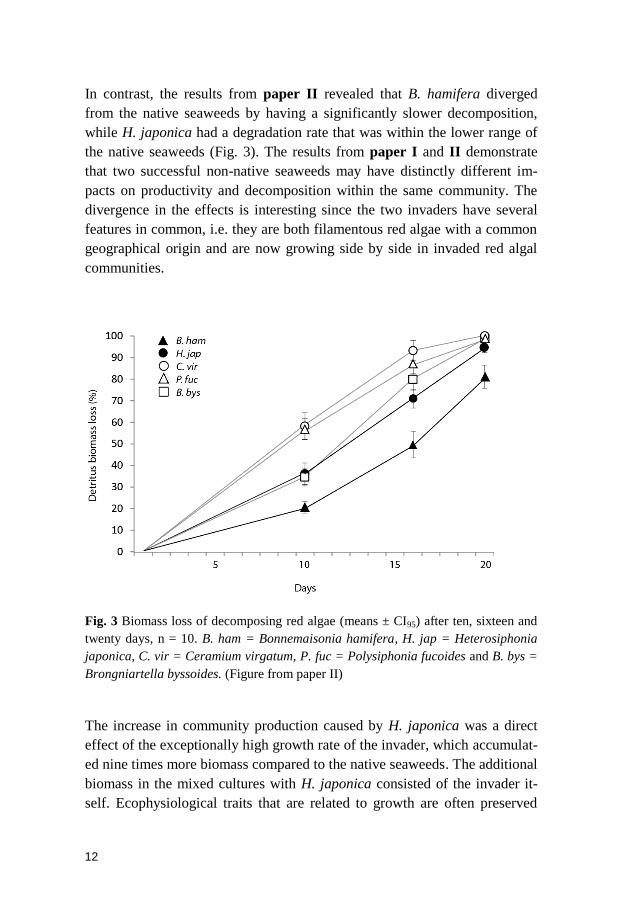
In contrast, the results from paper II revealed that B. hamifera diverged
from the native seaweeds by having a significantly slower decomposition,
while H. japonica had a degradation rate that was within the lower range of
the native seaweeds (Fig. 3). The results from paper I and II demonstrate
that two successful non-native seaweeds may have distinctly different im-
pacts on productivity and decomposition within the same community. The
divergence in the effects is interesting since the two invaders have several
features in common, i.e. they are both filamentous red algae with a common
geographical origin and are now growing side by side in invaded red algal
communities.
Fig. 3 Biomass loss of decomposing red algae (means ± CI95) after ten, sixteen and
twenty days, n = 10. B. ham = Bonnemaisonia hamifera, H. jap = Heterosiphonia
japonica, C. vir = Ceramium virgatum, P. fuc = Polysiphonia fucoides and B. bys =
Brongniartella byssoides. (Figure from paper II)
The increase in community production caused by H. japonica was a direct
effect of the exceptionally high growth rate of the invader, which accumulat-
ed nine times more biomass compared to the native seaweeds. The additional
biomass in the mixed cultures with H. japonica consisted of the invader it-
self. Ecophysiological traits that are related to growth are often preserved

13
throughout senescence and may have a large effect on decomposition
(Cornwell et al. 2008; Liao et al. 2008). Paper II – III contains four chemi-
cal analyses that reveal important physiological features of H. japonica. Pa-
per III shows that the species has several traits that are inherent to rapidly
growing primary producers, i.e. high levels of nitrogen (N) and low structur-
al investment. These traits render plant tissue attractive to primary consum-
ers, promotes rapid decomposition and is associated with high N release in
seaweed litter (Smith and Foreman 1984; Mann 1988; Wright et al. 2004;
Hättenschwiler et al. 2005). A high amount of N leached from the litter of H.
japonica when the species started to decompose. This probably contributed
to that the remaining litter had a degradation rate that was lower or equal to
the native seaweeds (paper II). However, N that is released into the sur-
rounding water will be available for uptake by organisms such as microbes,
filter feeders and primary producers. The high N accumulation, rapid growth
and subsequent N release at senescence indicates that H. japonica may in-
crease N cycling in invaded communities; an impact by non-native primary
producers with rapid growth that is frequently observed in terrestrial and
wetland systems (reviewed by Liao et al. 2008).
In contrast to H. japonica, B. hamifera had a growth rate that was within the
lower range of the native seaweeds and therefore had no significant effect on
community production. But B. hamifera has another important feature that
makes the species diverge from native co-occurring seaweeds; it produces a
potent chemical defense that efficiently deters native herbivores, and has
anti-microbial effects (Enge et al. 2012; Svensson et al. 2013). Defense
compounds residing in litter may (like nutritional properties) influence de-
composition, since the activity of microbes and detritivore fauna can be af-
fected (Hättenschwiler et al. 2005; Gessner et al. 2010). The litter of B.
hamifera had low N content (which probably reflects the slow growth rate of
the species). It also contained secondary metabolites, although the concentra-
tion was much lower than in living seaweeds (paper II). The fact that B.
hamifera degraded slower than the native seaweeds may therefore be a con-
sequence of residues of the chemical defense and/or low levels of tissue N.
The slow decomposition rate of B. hamifera compared to native filamentous
red algae indicates that the invader has the potential to slow the cycling of
nutrients in invaded communities.
High growth rate and chemical defense are likely to be important factors in
explaining how H. japonica and B. hamifera have become abundant constit-

14
uents of invaded communities. Several studies have found general correla-
tions between growth rate and the success of non-native primary producers
(Grotkopp et al. 2002; Pyšek and Richardson 2007; Van Kleunen et al.
2010), and in the case of B. hamifera experimental studies show that the
chemical defense gives the species competitive advantage through apparent
competition and chemical inhibition of native seaweeds´ recruits (Enge et al.
2013; Svensson et al. 2013). Ecophysiological traits may be important de-
terminants of invasion success and much effort has been spent on trying to
identify traits that characterize invasive species (Grotkopp et al. 2002; Pyšek
and Richardson 2007; Van Kleunen et al. 2010). But considerably less atten-
tion has been directed to how invader traits may affect impacts on native
communities. My studies demonstrate that the same traits that promote inva-
sion success may determine invasion effects on ecosystem functions.
Biotic resistance
Results from paper IV show that the non-native Fucus evanescens and the
native Fucus vesiculosus differ in competitive ability. Fucus evanescens
accumulated less biomass when growing together with its native congener,
while there was no corresponding effect of interspecific competition on F.
vesiculosus. Further, the grazer Littorina obtusata caused considerably more
damage to F. vesiculosus than to F. evanescens, but the herbivory did not
alter the competitive relationship between the two seaweeds despite high
grazer densities in the experiment. The results indicate that at least in adult
competition F. evanescens is a weak competitor compared to the native F.
vesiculosus, and that low palatability to native herbivores does not seem to
relieve the invader from the competitive disadvantage.
In the area of its origin, F. evanescens is an abundant constituent of fucoid
seaweed communities, which contrasts to the non-native range where the
species primarily is recorded from harbors and other sites that have low den-
sities of native fucoids (Powell 1957b; Schueller and Peters 1994; Wikström
et al. 2002). There are a number of native fucoids (such as, F. vesiculosus,
Fucus serratus and Ascophyllum nodosum) growing within the ecological
niche of F. evanescens in the new range. The results from paper IV support
previous suggestions that the non-native population of F. evanescens might
be restricted by competition with native fucoids (e.g. Powell 1957b; Bokn
and Lein 1978; Wikström et al. 2002).

15
Competition is important in determining the distribution and colonization of
native seaweed species (Chapman and Johnson 1990; Olson and Lubchenco
1990; Worm and Chapman 1996; Arenas et al. 2006), which implies that the
biotic resistance against species invasions exerted by native competitors
could be high in the seaweed system. Several studies have also shown that
the establishment of non-native seaweeds are facilitated when native sea-
weeds are removed, supporting the assumption that native competitors are an
important part of biotic resistance (Ambrose and Nelson 1982; Levin et al.
2002; Valentine and Johnson 2003). However, a recent meta-analysis found
that competition has no general effect on non-native marine primary produc-
ers (Kimbro et al. 2013). One possible reason for the lack of effect of com-
petition is that research on seaweed invasions has had a large focus on suc-
cessful high impact invaders. Little is, however, known about non-native
seaweeds with a modest occurrence. A strong bias towards studying success-
ful invaders may lead to an underestimation of the biotic resistance in sea-
weed communities. Paper IV gives support to competition being important
in reducing the success of non-native seaweeds. I suggest that more studies
of non-native seaweeds that have been incorporated in native communities
without dominating them may contribute to a better understanding of how
native communities affect invasion success. This would also contribute to a
more balanced picture of the effects of seaweed invasions.
Paper IV shows in accordance with several previous studies, that F.
evanescens largely escapes herbivory in the invaded range. Paper III simi-
larly shows that the fast-growing H. japonica is low preferred food to all
three investigated herbivores, which may be important for survival of the
invader when the growth rate is reduced during cold seasons. The chemically
defended B. hamifera is also avoided as food by native herbivores (Enge et
al. 2012; Enge et al. 2013). There are two factors that may explain why
many seaweed invaders are of low palatability. First, the grazing pressure
exerted by generalists is high in seaweed communities (Duffy and Hay 2000;
Poore et al. 2012), which reduces the probability of escaping predation. Se-
cond, most seaweed introductions are accidental (Williams and Smith 2007),
meaning that many seaweed invaders are likely to originate from relatively
few individuals. These conditions may cause a selection for low palatability.
Grazer avoidance has therefore been suggested to be an important contribu-
tor to the success of a number of non-native seaweeds that has become dom-
inant in their new range (e.g. Gollan and Wright 2006; Monteiro et al. 2009;
Nejrup et al. 2012). The case of F. evanescens shows that low palatability

16
can also occur in less successful non-native seaweeds, which implies that the
link between low palatability and invasion success cannot be taken for grant-
ed.

17
Conclusions and future perspectives
One of the main findings of this thesis is that successful non-native seaweeds
have potential to affect ecosystem functions in different ways depending on
their ecophysiological traits. Rapidly growing invaders, like Heterosiphonia
japonica, tend to accumulate a high amount of nitrogen in their tissues. They
may increase primary productivity in invaded communities due to their fast
growth, and when they decompose their litter may release a high amount of
nitrogen. Hence, they have potential to increase the flux of nutrients in the
receiving environment. Slow-growing invaders, that are producing costly
defense compounds, like Bonnemaisonia hamifera are on the other hand
unlikely to increase primary production. But they may slow litter decomposi-
tion rates and have potential to slow down nutrient cycling due to low tissue
nitrogen content and secondary metabolites in their litter, deterring
detritivores.
The link between ecophysiological traits and effects on ecosystem functions
is well known in general ecology, but in invasion biology there has been
little focus on how traits relates to invasion impacts. My studies show that
traits promoting invasion success may determine impacts on ecosystem func-
tions. There is plenty of information available on traits of specific non-native
primary producers, since there has been a relatively large focus on identify-
ing traits that characterize successful invaders. Hence, an interesting next
step is to explore if ecophysiological traits of plant and seaweed invaders can
be used to predict invasion impacts on ecosystem processes.
My thesis also shows that native seaweeds may reduce the success of non-
native seaweeds. The results comply with previous findings that emphasize
the importance to reduce anthropogenic stressors on native seaweed commu-
nities to decrease the risk of non-native seaweeds becoming abundant in
native ecosystems.

18
Acknowledgements
I would like to thank Sofia Wikström, Johan Eklöf, Joakim Hansen, Stina
Tano and Nils Kautsky for their valuable comments and help during the
writing of this synthesis.

19
References
Abramoff MD, Magalhaes P, Ram S (2004) Image processing with ImageJ. Biophotonics International 11:36-42
Ambrose RF, Nelson BV (1982) Inhibition of giant kelp recruitment by an introduced brown alga. Bot Mar 25:265–268.
Arenas F, Sánchez I, Hawkins SJ, Jenkins SR (2006) The invasibility of marine algal assemblages: role of functional diversity and identity. Ecology 87:2851–2861.
Axelius B, Karlsson J (2004) Japanplym, ny rödalg för Sverige. Sven Bot Tidskr 98:269.
Bjærke MR (2004) Molecular and ecological studies on introduced marine macroalgae in Norwegian waters. Dr. Scient. thesis, Faculty of Mathematics and Natural Sciences, University of Oslo
Bokn T, Lein TE (1978) Long term changes in fucoid association of the in-ner Oslofjord, Norway. Norwegian Journal of Botany 25:9–14
Britton-Simmons KH (2004) Direct and indirect effects of the introduced alga Sargassum muticum on benthic, subtidal communities of Wash-ington State, USA. Mar Ecol Prog Ser 277:61–78.
Bulleri F (2009) Facilitation research in marine systems: state of the art, emerging patterns and insights for future developments. J Ecol 97:1121–1130. doi: 10.1111/j.1365-2745.2009.01567.x
Carpenter RC (1990) Competition among marine macroalgae: a physiologi-cal perspective. J Phycol 26:6–12.
Cebrian J (1999) Patterns in the fate of production in plant communities. Am Nat 154:449–468.
Chapman ARO (1995) Functional ecology of fucoid algae: twenty-three years of progress. Phycologia 34:1–32.

20
Chapman ARO, Johnson CR (1990) Disturbance and organization of macroalgal assemblages in the Northwest Atlantic. Hydrobiologia 192:77–121.
Cornwell WK, Cornelissen JH, Amatangelo K, et al (2008) Plant species traits are the predominant control on litter decomposition rates with-in biomes worldwide. Ecol Lett 11:1065–1071.
Costanza R, d’ Arge R, Groot R de, et al (1998) The value of the world’s ecosystem services and natural capital. Nature 387:253–260
Davis MA (2003) Biotic Globalization: Does Competition from Introduced Species threaten Biodiversity? BioScience 53:481.
Duffy JE, Hay ME (2000) Strong impacts of grazing amphipods on the or-ganization of a benthic community. Ecol Monogr 70:237–263. doi: 10.1890/0012-9615(2000)070[0237:SIOGAO]2.0.CO;2
Elton C (1958) The ecology of invasions by animals and plants. Methuen, London, UK.
Enge S, Nylund GM, Harder T, Pavia H (2012) An exotic chemical weapon explains low herbivore damage in an invasive alga. Ecology 93:2736–2745. doi: 10.1890/12-0143.1
Enge S, Nylund GM, Pavia H (2013) Native generalist herbivores promote invasion of a chemically defended seaweed via refuge-mediated ap-parent competition. Ecol Lett 16:487–492. doi: 10.1111/ele.12072
Gessner MO, Swan CM, Dang CK, et al (2010) Diversity meets decomposi-tion. Trends Ecol Evol 25:372–380.
Gollan JR, Wright JT (2006) Limited grazing pressure by native herbivores on the invasive seaweed Caulerpa taxifolia in a temperate Australian estuary. Mar Freshw Res 57:685–694. doi: 10.1071/MF05253
Grosholz E (2002) Ecological and evolutionary consequences of coastal invasions. Trends Ecol Evol 17:22–27.
Grotkopp E, Rejmánek M, Rost TL (2002) Toward a causal explanation of plant invasiveness: seedling growth and life-history strategies of 29 pine (Pinus) species. Am Nat 159:396–419. doi: 10.1086/338995
Hammann M, Wang G, Rickert E, et al (2013) Invasion success of the sea-weed Gracilaria vermiculophylla correlates with low palatability.

21
Hättenschwiler S, Tiunov AV, Scheu S (2005) Biodiversity and litter de-composition in terrestrial ecosystems. Annu Rev Ecol Evol Syst 36:191–218.
Husa V, Sjøtun K (2006) Vegetative reproduction in “ Heterosiphonia ja-ponica” (Dasyaceae, Ceramiales, Rhodophyta), an introduced red alga on European coasts. Bot Mar 49:191–199. doi: 10.1515/BOT.2006.024
Husa V, Sjøtun K, Brattenborg N, Eiliv Lein T (2008) Changes of macroalgal biodiversity in sublittoral sites in southwest Norway: impact of an introduced species or higher temperature? Mar Biol Res 4:414–428. doi: 10.1080/17451000802232874
Husa V, Sjøtun K, Lein TE (2004) The newly introduced species Heterosiphonia japonica Yendo (Dasyaceae, Rhodophyta): geo-graphical distribution and abundance at the Norwegian southwest coast. Sarsia 89:211–217. doi: 10.1080/00364820410006600
Kimbro DL, Cheng BS, Grosholz ED (2013) Biotic resistance in marine environments. Ecol Lett 16:821–833. doi: 10.1111/ele.12106
Levine JM, Adler PB, Yelenik SG (2004) A meta-analysis of biotic re-sistance to exotic plant invasions. Ecol Lett 7:975–989.
Levin PS, Coyer JA, Petrik R, Good TP (2002) Community-wide effects of nonindigenous species on temperate rocky reefs. Ecology 83:3182–3193.
Liao C, Peng R, Luo Y, et al (2008) Altered ecosystem carbon and nitrogen cycles by plant invasion: a meta-analysis. New Phytol 177:706–714. doi: 10.1111/j.1469-8137.2007.02290.x
Lotze HK, Lenihan HS, Bourque BJ, et al (2006) Depletion, degradation, and recovery potential of estuaries and coastal seas. Science 312:1806–1809. doi: 10.1126/science.1128035
Maggs CA, Stegenga H (1998) Red algal exotics on North Sea coasts. Helgoländer Meeresunters 52:243–258. doi: 10.1007/BF02908900
Mann KH (1973) Seaweeds: Their Productivity and Strategy for Growth. Science 182:975–981.
Mann KH (1988) Production and use of detritus in various freshwater, estua-rine, and coastal rnarine ecosystems. Limnol Oceanogr 33:910–930.

22
Mitchell CE, Agrawal AA, Bever JD, et al (2006) Biotic interactions and plant invasions. Ecol Lett 9:726–740. doi: 10.1111/j.1461-0248.2006.00908.x
Monteiro CA, Engelen AH, Santos ROP (2009) Macro- and mesoherbivores prefer native seaweeds over the invasive brown seaweed Sargassum muticum: a potential regulating role on invasions. Mar Biol 156:2505–2515. doi: 10.1007/s00227-009-1275-1
Moore JC, Berlow EL, Coleman DC, et al (2004) Detritus, trophic dynamics and biodiversity. Ecol Lett 7:584–600.
Moy FE, Christie H (2012) Large-scale shift from sugar kelp (Saccharina latissima) to ephemeral algae along the south and west coast of Norway. Mar Biol Res 8:309–321. doi: 10.1080/17451000. 2011.637561
Nejrup LB, Pedersen MF, Vinzent J (2012) Grazer avoidance may explain the invasiveness of the red alga Gracilaria vermiculophylla in Scan-dinavian waters. Mar Biol 159:1–10. doi: 10.1007/s00227-012-1959-9
Nyberg CD, Wallentinus I (2005) Can species traits be used to predict ma-rine macroalgal introductions? Biol Invasions 7:265–279. doi: 10.1007/s10530-004-0738-z
Nylund G, Cervin G, Hermansson M, Pavia H (2005) Chemical inhibition of bacterial colonization by the red alga Bonnemaisonia hamifera. Mar Ecol Prog Ser 302:27–36. doi: 10.3354/meps302027
Olson AM, Lubchenco J (1990) Competition in seaweeds: linking plant traits to competitive outcomes. J Phycol 26:1–6.
Poore AGB, Campbell AH, Coleman RA, et al (2012) Global patterns in the impact of marine herbivores on benthic primary producers. Ecol Lett 15:912–922. doi: 10.1111/j.1461-0248.2012.01804.x
Powell HT (1957a) Studies in the genus Fucus LI Fucus distichus L. emend. Powell. J Mar Biol Assoc U K 36:407–432.
Powell HT (1957b) Studies in the genus Fucus L. II. Distribution and ecolo-gy of forms of Fucus distichus L. emend. Powell in Britain and Ire-land. J Mar Biol Assoc U K 36:663–693.
Powell K, Chase J, Knight T (2011) A synthesis of plant invasion effects on biodiversity across spatial scales. Am J Bot 98:539–548. doi: 10.3732/ajb.1000402

23
Pyšek P, Richardson DM (2007) Traits associated with invasiveness in alien plants: where do we stand? In: Nentwig W (ed) Biological invasions. Springer–Verlag, Berlin Heidelberg, pp 97–125
Rangeley RW, Kramer DL (1995) Use of rocky intertidal habitats by juve-nile pollock Pollachius virens. Mar Ecol Prog Ser 126:9–17.
Sax D, Gaines S (2003) Species diversity: from global decreases to local increases. Trends Ecol Evol 18:561–566. doi: 10.1016/S0169-5347(03)00224-6
Schaffelke B, Hewitt C (2007) Impacts of introduced seaweeds. Bot Mar 50:397–417. doi: 10.1515/BOT.2007.044
Scheibling RE, Gagnon P (2006) Competitive interactions between the inva-sive green alga Codium fragile ssp tomentosoides and native cano-py-forming seaweeds in Nova Scotia (Canada). Mar Ecol Prog Ser 325:1–14. doi: 10.3354/meps325001
Schneider CW (2010) Report of a new invasive alga in the Atlantic United States: Heterosiphonia japonica in Rhode Island. J Phycol 46:653–657. doi: 10.1111/j.1529-8817.2010.00866.x
Schueller GH, Peters AF (1994) Arrival of Fucus evanescens (Phaeophyceae) in Kiel Bight (Western Baltic). Bot Mar 37:471–478.
Simmons H (1898) Algologiska notiser. II. Einige Algenfunde bei Drøbak. Bot Not 117–123.
Sjøtun K, Husa V, Peña V (2008) Present distribution and possible vectors of introductions of the alga Heterosiphonia japonica (Ceramiales, Rhodophyta) in Europe. Aquat Invasions 3:377–394. doi: 10.3391/ai.2008.3.4.3
Smith BD, Foreman RE (1984) An assessment of seaweed decomposition within a southern Strait of Georgia seaweed community. Mar Biol 84:197–205.
Svensson JR, Nylund GM, Cervin G, et al (2013) Novel chemical weapon of an exotic macroalga inhibits recruitment of native competitors in the invaded range. J Ecol 101:140–148. doi: 10.1111/1365-2745.12028
Thomsen MS, Wernberg T, Staeaehr P, et al (2007) Alien macroalgae in Denmark - a broad-scale national perspective. Mar Biol Res 3:61–72. doi: 10.1080/17451000701213413

24
Tomas F, Box A, Terrados J (2011) Effects of invasive seaweeds on feeding preference and performance of a keystone Mediterranean herbivore. Biol Invasions 13:1559–1570. doi: 10.1007/s10530-010-9913-6
Valentine JP, Johnson CR (2003) Establishment of the introduced kelp Undaria pinnatifida in Tasmania depends on disturbance to native algal assemblages. J Exp Mar Biol Ecol 295:63–90.
Valentine JP, Magierowski RH, Johnson CR (2007) Mechanisms of inva-sion: establishment, spread and persistence of introduced seaweed populations. Bot Mar 50:351–360.
Van Kleunen M, Weber E, Fischer M (2010) A meta-analysis of trait differ-ences between invasive and non-invasive plant species. Ecol Lett 13:235–245. doi: 10.1111/j.1461-0248.2009.01418.x
Vilà M, Espinar JL, Hejda M, et al (2011) Ecological impacts of invasive alien plants: a meta-analysis of their effects on species, communities and ecosystems. Ecol Lett 14:702–708. doi: 10.1111/j.1461-0248.2011.01628.x
Vye SR, Emmerson MC, Arenas F, et al (2014) Stressor intensity determines antagonistic interactions between species invasion and multiple stressor effects on ecosystem functioning.
White LF, Shurin JB (2011) Density dependent effects of an exotic marine macroalga on native community diversity. J Exp Mar Biol Ecol 405:111–119. doi: 10.1016/j.jembe.2011.05.024
Wikström SA (2004) Marine seaweed invasions; the ecology of introduced Fucus evanescens. Dr. Scient. thesis, Department of botany, Stock-holm university
Wikström SA, Wachenfeldt T von, Kautsky L (2002) Establishment of the Exotic Species Fucus evanescens C. Ag. (Phaeophyceae) in öresund, Southern Sweden. Bot Mar 45:510–517. doi: 10.1515/BOT. 2002.054
Williams S, Smith J (2007) A global review of the distribution, taxonomy, and impacts of introduced seaweeds. Annu Rev Ecol Evol Syst 38:327–359. doi: 10.1146/annurev.ecolsys.38.091206.095543
Worm B, Barbier EB, Beaumont N, et al (2006) Impacts of biodiversity loss on ocean ecosystem services. Science 314:787–790. doi: 10.1126/science.1132294

25
Worm B, Chapman ARO (1996) Interference competition among two inter-tidal seaweeds: Chondrus crispus strongly affects survival of Fucus evanescens recruits. Mar Ecol Prog Ser 1996:297–301.
Wright IJ, Reich PB, Westoby M, et al (2004) The worldwide leaf econom-ics spectrum. Nature 428:821–827. doi: 10.1038/nature02403

26
Thank you
First of all I want to thank my supervisors; Sofia Wikström, Henrik Pavia
and Nils Kautsky.
Sofia, you are a skilled ecologist and a good writer, which has been a great
source of inspiration during my studies. However, I am most of all grateful
for your patience as supervisor, and that you always emphasized that the
PhD-education is a period of learning.
Henrik, thank you for your encouragement and excellent feedback on vari-
ous projects, experimental designs and manuscripts. This thesis would not
have been the same without you.
Nils, you showed your support by sitting at the front row during my presen-
tations at various conferences (independent of whether you had heard the
talks before or not). You also opened up your home for me at Tjärnö, when
project finances where running low. I appreciate this tremendously.
In addition, I am grateful to all brilliant people that I have had the opportuni-
ty to meet while working on this thesis.
A special thanks to Nichlas Staberg, Gundula Kolb, Linda Eggertsen,
Rebecka Wikström and Anna Pavia, for working with me in the field. I wish
the sun will always shine on you.
I also want to thank Kjersti Sjøtun, Jan Rueness, Vivian Husa and Stein
Fredriksen, and show my gratitude to Poul Møller Pedersen. The days of
studying seaweeds at Espegren field station are still an important source of
inspiration to me, and your faith in me and Jenny at the start of our PhD-
studies meant more to me than you probably know.
Thank you Johan Eklöf, for good advice given during long road trips to
Tjärnö and for excellent times during teaching.
I also want to thank my colleagues both at SU and at Tjärnö for moral and
practical support, as well as my sisters in arms Stina Tano and Jenny Zie, I
would have been lost without you.
Further, I want to thank Peter Hambäck for statistical advice and Ove Eriks-
son for advice on future career plans.

27
Jag vill också tacka min familj. Ni har bidragit genom att stötta mig på en
mängd olika sätt.
– Mamma, du var den som lyckades överlista mig när jag som barn ansåg att
det där med att läsa och skriva var ett onödigt påfund som de vuxna hade
kommit på.
– Johan, du hjälpte mig att läsa upp mina mattebetyg, så att jag kunde bli
antagen till biologiprogrammet på Södertörn högskola.
– Pappa, det är dig jag anförtror tekniska problem i mina experimentanlägg-
ningar.
– Lena, du gav mig min första växtpress, den är fortfarande flitigt använd.
– Erik, det är till dig jag kommer när livet är komplicerat. Du lyssnar och ger
råd utan att döma.
Till sist, vill jag tacka min partner Joakim. Du har stått vid min sida i sol och
regn, trots att jag är långt ifrån den perfekta flickvännen. Jag älskar dig mer
än jag kan uttrycka i ord.





























![Sitt,Hans - Scale Studies (CD Sheet Music)[1]](https://img.pdfslide.us/doc/110x75/563db9d1550346aa9aa03934/sitthans-scale-studies-cd-sheet-music1.jpg)