Embed Size (px)
Citation preview

Developmental Cell
Article
Making the Connection:Ciliary Adhesion ComplexesAnchor Basal Bodies to the Actin CytoskeletonIoanna Antoniades,1,2 Panayiota Stylianou,1,2 and Paris A. Skourides1,*1Department of Biological Sciences, University of Cyprus, P.O. Box 20537, Nicosia 2109, Cyprus2These authors contributed equally to this work
*Correspondence: [email protected]://dx.doi.org/10.1016/j.devcel.2013.12.003
SUMMARY
Cilia have been associated with diverse develop-mental and physiological processes, and defects incilia underlie a number of genetic conditions. Severallines of evidence support a critical role of the actincytoskeleton in ciliogenesis and ciliary function.Here,we show thatwell-characterized focal adhesion(FA) proteins, including FAK, Paxillin, and Vinculin,associate with the basal bodies of multiciliated cellsand form complexes (CAs) that interact with theactin cytoskeleton. FAK downregulation leads tociliogenesis defects similar to those observed whenthe actin cytoskeleton is disrupted, including defectsin basal body migration, docking, and spacing,suggesting that CAs link basal bodies to the actincytoskeleton. The important role of FA proteins inciliogenesis leads us to propose that evolutionarilyFA proteins, many of which are found in primitiveflagellated unicellular eukaryotes,mayhave originallyevolved to perform functions at flagella and werelater co-opted for use in cell adhesion.
INTRODUCTION
Cilia are microtubule-based organelles extending from basal
bodies on the surface of vertebrate cells. There are two main
types of cilia; primary and motile cilia. While primary cilia can
be found on almost every cell type, motile cilia are restricted to
specialized epithelia and are essential for generating fluid flow.
Such epithelia include the airways, the ventricles of the brain,
and the oviducts (Knowles and Boucher, 2002; Sawamoto
et al., 2006; Shah et al., 2009; Worthington and Cathcart,
1963). Motile cilia can be found in single copy, as motile mono-
cilia, or multicilia (Goetz and Anderson, 2010; Pazour and
Witman, 2003; Pedersen et al., 2008; Roy, 2009), and the disrup-
tion of ciliary function has been shown to be responsible for
a variety of human diseases termed ciliopathies (Afzelius,
1976, 2004; Baker and Beales, 2009; Bisgrove and Yost, 2006;
Hildebrandt et al., 2011;Wallingford andMitchell, 2011; Zariwala
et al., 2007). Perhaps the most striking results regarding the
importance of cilia come from the fly, where DSas-4 mutant
larvae, which lack centrioles, develop into morphologically
70 Developmental Cell 28, 70–80, January 13, 2014 ª2014 Elsevier In
normal adults, which die shortly after birth because their sensory
neurons lack cilia (Basto et al., 2006).
The dependence of ciliogenesis on the actin cytoskeleton is
well documented, and both components of the actin cyto-
skeleton, as well as regulators of actin dynamics, have been
shown to play important roles in various aspects of ciliogenesis
(Bershteyn et al., 2010; Boisvieux-Ulrich et al., 1990; Dawe et al.,
2007; Ioannou et al., 2013; Klotz et al., 1986; Lemullois et al.,
1988; Pan et al., 2007; Ravanelli and Klingensmith, 2011;
Tamm and Tamm, 1988). In multiciliated cells, basal bodies
form de novo deep within the cytoplasm (Sorokin, 1968) and
are subsequently transported, via an actin-myosin-based mech-
anism, to the apical surface where they dock (Boisvieux-Ulrich
et al., 1990; Dawe et al., 2007; Klotz et al., 1986; Lemullois
et al., 1987). In addition to the proposed role of the actin cyto-
skeleton in the transport of basal bodies, the apical surface of
multiciliated cells is enriched with a dense meshwork of actin
composed of two distinct pools, the apical and the subapical
actin networks. Loss of the apical actin network leads to prob-
lems with basal body localization and polarity and appears to
be necessary for basal body docking (Boisvieux-Ulrich et al.,
1990; Park et al., 2006, 2008; Werner et al., 2011). Once basal
bodies dock at the apical surface, they must be polarized in
order for the cilia to beat in a synchronized manner and create
directional fluid flow. The subapical actin network has been
shown to be important for basal body spacing and metachronal
synchrony. This network appears to connect basal bodies
through the striated rootlets. Interestingly, disruption of the sub-
apical actin network during basal body docking results in severe
spacing issues, suggesting that spacing is determined by this
actin network, while its disruption after ciliogenesis is completed
leads to loss of metachornal synchrony (Werner et al., 2011). The
actin cytoskeleton and its regulators have also been shown to
influence the formation of primary cilia. Although the precise
role of actin on primary cilia formation is not clear, studies
suggest that modulation of the actin cytoskeleton regulates
basal body positioning and primary cilium development overall,
suggesting that primary cilia formation is promoted through the
disassembly of certain pools of highly dynamic filamentous actin
(Bershteyn et al., 2010; Kim et al., 2010; Sharma et al., 2011; Yan
and Zhu, 2013; Yin et al., 2009). However, a mutation in Talpid3
has been shown to elicit defects in primary cilia formation and
actin organization in the chicken. In these mutants, docking of
basal bodies to the apical cell membrane is defective, despite
the fact that mature basal bodies form (Yin et al., 2009). These
c.

Figure 1. FA Proteins FAK, Paxillin, and Vin-
culin Associatewith Basal Bodies in Ciliated
Cells
(A and A0 ) Multiciliated cell expressing GFP FAK
and centrin2 RFP. Each basal body is associated
with a GFP FAK punctum at its posterior (in relation
to the tadpole’s anterior-posterior axis). GFP FAK
signal displays a gradient within the cell (in respect
to the anterior-posterior axis), with elevated signal
at the basal bodies localized at the cell’s posterior.
(B and B0) Multiciliated cell expressingGFP Paxillin
and centrin2 RFP. GFP Paxillin exhibits a similar
localization and distribution throughout the cell,
like GFP FAK.
(C and C0) Multiciliated cell expressing GFP Vin-
culin and centrin2 RFP. Localization and distribu-
tion of GFP Vinculin resembles that of GFP FAK
and GFP Paxillin.
(D) Immunofluorescence staining with a p-S732
FAK antibody in centrin2 CFP-injected embryos.
Endogenous FAK, associated with the basal
bodies, is phosphorylated on Serine 732.
(E) Immunofluorescence staining of endogenous
Vinculin in centrin2 RFP-expressing multiciliated
cells, gives a similar localization as GFP Vinculin.
(F and F0) GRP ciliated cells of a stage 17 embryo
expressing GFP FAK and centrin2 RFP, immuno-
stained for acetylated tubulin.
(G and G0) Neural tube cross section of a stage 24
embryo coexpressing GFP FAK and centrin2 RFP
and immunostained for acetylated tubulin. GFP
FAK associates with the basal bodies in both GRP
and neural tube ciliated cells.
(H and I) Immunofluorescence staining of endog-
enous Serine 732 phosphorylated FAK (H) and
Paxillin (I) in NIH 3T3 cells, costained for g tubulin
and acetylated tubulin. Endogenous FAK and
Paxillin localize at the base of primary cilia next to
the basal body in mammalian ciliated cells.
See also Figure S1.
Developmental Cell
CAs Link Basal Bodies to the Actin Network
results suggest that formation of the apical actin network affects
the attachment of basal bodies to the cell membrane and that
certain aspects of the role of the actin network may be shared
between motile and primary cilia.
In addition to the studies in animals, actin has also been
shown to be important for the control of flagellar length in
Chlamydomonas, while knockdown of actin in Giardia results
in defects in the organism’s flagella and their positioning, sug-
gesting that the requirement of actin for normal ciliogenesis
extends to flagellated unicellular eukaryotes (Dentler and
Adams, 1992; Paredez et al., 2011). Despite the critical role
of the actin cytoskeleton in ciliogenesis and ciliary function,
how the basal bodies interact with the actin cytoskeleton is
not known. Here, we show that four well-characterized focal
adhesion (FA) proteins are found associated with basal bodies
and form what we termed ‘‘ciliary adhesions’’ (CAs), connecting
basal bodies to the actin cytoskeleton of multiciliated cells.
Similar complexes are also found in monociliated cells of the
gastrocel roof plate (GRP) and primary cilia. CA protein FAK
interacts with both basal bodies and actin, while its downregu-
lation leads to defects in ciliogenesis related to actin-based
processes, suggesting that CAs functionally link basal bodies
to the actin cytoskeleton.
Deve
RESULTS
FAK, Paxillin, Vinculin, and Talin Are Associatedwith the Basal Bodies in Ciliated CellsFAs are large, dynamic protein complexes through which the
actin cytoskeleton connects to the extracellular matrix. FAK
and Paxillin are well-characterized FA proteins with critical roles
in the assembly and disassembly of FA complexes. The fact that
both FAK and Paxillin have been previously detected in associa-
tion with centrosomes raised the possibility that FA proteins also
associate with basal bodies (Herreros et al., 2000; Park et al.,
2009). Using green fluorescent protein (GFP) fusions of FAK,
Paxillin, and Vinculin in combination with live imaging, we noted
that all three proteins localize in close association with the basal
bodies, visualized using centrin2 red fluorescent protein (RFP),
in Xenopus multiciliated cells (Figures 1A–1C0). Control cells
expressing GFP alone show that GFP by itself does not localize
at the basal bodies (Figures S1A and S1A0 available online), and
western blotting showed that expression levels of GFP Paxillin
and GFP Vinculin were similar to those of the endogenous
proteins (Figures S1B and S1C). In addition, a fourth FA protein,
Talin, fused to FusionRed also localizes at the basal bodies of
multiciliated cells (Figure S1D). To further verify the GFP fusion
lopmental Cell 28, 70–80, January 13, 2014 ª2014 Elsevier Inc. 71

Figure 2. FA Proteins Present an Anterior-
Posterior Gradient in Multiciliated Cells
(A–B) Intensity color-coded maximum intensity
projections of centrin2 RFP (A) and GFP Vinculin
(A0) reveals the presence of a Vinculin gradient
in multiciliated cells. The signal of centrin2 RFP
is uniform (A), while the GFP Vinculin signal is
stronger at the cell’s posterior (A0 ). In (B), multi-
ciliated cells coexpressing GFP Vinculin and
centrin2 RFP are shown. The anterior-posterior
gradient is maintained at the tissue level. The
arrows indicate cells’ posterior where the signal
for GFP Vinculin appears elevated.
(C) Multiciliated cell coexpressing mKate2 FAK,
clamp GFP, and centrin2 CFP. FAK is localized
next to the basal body at the end of the region
marked by clamp GFP.
(D and E) Confocal optical sections of a multi-
ciliated cell showing that GFP Paxillin and mKate2
FAK colocalize. GFP Paxillin is more concentrated
at the apical-most region of the basal bodies (D),
while mKate2 FAK is more concentrated slightly
below (E).
(F–G0 ) Intensity coding of ROIs of (D) and (E).
Paxillin exhibits highest intensity at 0.00 mm, as
shown in (F) and (F0 ), whereas FAK exhibits highest
intensity at 0.38 mm, as shown in (G) and (G0).
Developmental Cell
CAs Link Basal Bodies to the Actin Network
localizations, we went on to stain endogenous FAK and Vinculin
using previously characterized antibodies against these two
proteins. Since FAK associated with centrosomes was shown
to be phosphorylated on Serine 732, we used a phospho-Serine
732-specific FAK antibody in tadpoles expressing centrin2 cyan
fluorescent protein (CFP), and, as shown, the staining confirms
that endogenous FAK is associated with basal bodies and is
phosphorylated on Serine 732 (Figure 1D). In addition, immuno-
fluorescence experiments using a monoclonal Vinculin antibody
in centrin2 RFP-expressing tadpoles gives very similar results
to the GFP fusion (Figure 1E).
We went on to express GFP FAK in neural tissues to examine
the possibility that FA proteins also associate with basal bodies
in the monociliated cells of the GRP and the primary cilia of the
neural tube. As shown in Figures 1F and 1G, GFP FAK localizes
adjacent to the basal bodies of both motile monocilia of the
GRP (Figures 1F and 1F0) and primary cilia of the neural tube
(Figures 1G and 1G0). The localization of FAK and Paxillin fusion
constructs at the basal bodies of primary cilia was confirmed
using antibodies to image the endogenous proteins in NIH 3T3
cells after serum-starvation-induced ciliogenesis (Figures 1H
and 1I). These results suggest that the aforementioned FA
proteins associate with the basal bodies in all types of cilia.
FAK, Paxillin, and Vinculin Display Polarity withinMulticiliated Cells of the Xenopus EpidermisIn addition to the strong localization at the basal bodies, FAK,
Paxillin, and Vinculin display polarity within the cell. Two types
of polarity have been identified in multiciliated cells, both
controlled by the planar cell polarity pathway: rotational polarity
and tissue-level polarity (Mitchell et al., 2007, 2009; Park et al.,
2008; Wallingford, 2010). Rotational polarity refers to the align-
ment of basal bodies within each cell, and the aforementioned
FA proteins are clearly rotationally polarized (Figures 1A–1C).
72 Developmental Cell 28, 70–80, January 13, 2014 ª2014 Elsevier In
Tissue-level polarity, on the other hand, refers to the coordina-
tion of the multiciliated cells across the tissue. However, FAK,
Paxillin, and Vinculin also display a third type of polarity, with
stronger signal emanating from basal bodies at the cell posterior
and weaker signal from basal bodies at the front of the cell
(in relation to the tadpole’s anterior-to-posterior axis) (Figures
1A–1C). This gradient is sharper in the case of Vinculin, in which
the first row of basal bodies, in close proximity to the cortical
actin cytoskeleton, shows a much higher level of Vinculin
compared to the second row, which is not interacting with the
cell cortex (Figures 1C and 1C0). This gradient becomes clearer
in images where both centrin as well as Vinculin are presented
using intensity coding, revealing a strong gradient in the cell (Fig-
ures 2A and 2A0). Imaging ciliated cells at lower magnifications
shows that this polarity is retained at the tissue level (Figure 2B).
The signal for all four proteins localizes immediately adjacent to
each basal body and is rotationally polarized, but its relation to
accessory structures, like the basal foot and the rootlet, cannot
be determined without additional markers.
To better define this region, we expressed mKate2 FAK with
clamp GFP, to mark the striated rootlets, and centrin2 CFP, to
mark the basal bodies. As shown in Figure 2C, the FAK signal
is concentrated at the end of the region marked by clamp
GFP, roughly, the area where the basal foot forms and an
area where electron microscopy analysis identified interactions
between basal bodies and the actin cytoskeleton (Chailley
et al., 1989). This shows that the gradient within each cell
described earlier, is an anterior-to-posterior gradient with higher
concentrations at the basal bodies at the back of each cell
(in relation to the tadpole’s anterior-to-posterior axis).
Despite the fact that the localization of the three proteins was
nearly identical, we noticed that Paxillin and Vinculin were always
appearing simultaneously with the basal bodies in z stacks, while
FAK appeared immediately after. This suggests that Paxillin and
c.

Figure 3. FA Proteins Form CA Complexes
Connecting Basal Bodies to the Actin
Network
(A) Basal bodies (labeled with centrin2 RFP) are
docked at the apical surface of multiciliated cells,
in the same plane as apical actin (visualized with
GFP utrophin). Each basal body is found at the
center of an actin ring.
(B) mKate2 FAK is in contact with the apical actin
network.
(C) Coexpression of centrin2 RFP and GFP FAK
followed by phalloidin staining suggests that FAK
connects basal bodies to the apical actin network.
(D and D0) Multiciliated cell expressing GFP FAK,
marking CAs, and mKate2 actin, showing the
apical actin network, before (D) and after (D0)acceptor photobleaching.
(D00) mKate2 (acceptor) intensity drops after
photobleaching, while GFP (donor) intensity rises,
showing that FRET is taking place between GFP
andmKate2 and suggesting that FAK is interacting
with actin.
(E and E0) Multiciliated cell expressing centrin GFP
andmKate2 actin before (E) and after (E0) acceptorphotobleaching.
(E00) mKate2 intensity drops after photobleaching,
but the GFP intensity remains unchanged, sug-
gesting that no FRET is taking place between GFP
and mKate2, indicating the absence of a direct
interaction between centrin and actin.
See also Figure S2.
Developmental Cell
CAs Link Basal Bodies to the Actin Network
Vinculin display different distributions along the apicobasal axis
compared to FAK. In an effort to confirm this, we coexpressed
GFP Paxillin, mKate2 FAK, and centrin2 yellow fluorescent
protein (YFP). As shown in Figures 2D and 2E, FAK and Paxillin
colocalize but have a slightly different distribution along the
z axis. Paxillin appears to have the highest density at the api-
cal-most region of the basal bodies (Figures 2F and 2F0), while
FAK has a maximal density slightly below the apical-most region
of each basal body (Figures 2G and 2G0), as shown in the two
adjacent optical sections of Figures 2F–2G0.
CAs Link Basal Bodies to the Actin CytoskeletonThe presence of four FA proteins in close association with basal
bodies in a region shown to have interactions with the actin
cytoskeleton raised the possibility that these proteins are form-
ing a complex connecting basal bodies to the actin network.
As shown in Figure 3A, each basal body is in the center of a
ring of actin. On the other hand, FAK partially colocalizes with
the apical actin network, as shown in Figure 3B, suggesting
that FAK, Paxillin, Talin, and Vinculin are forming a complex
linking each basal body to the actin network. Coexpression of
GFP FAK with centrin2 RFP and staining with phalloidin shows
that FAK is in contact with both the basal bodies and the
actin network (Figure 3C). In order to confirm an interaction
between FAK and the actin network, we coexpressed GFP
FAK (donor) with mKate2 actin (acceptor) and carried out
acceptor photobleaching experiments to examine the possibility
of intermolecular fluorescence resonance energy transfer
(FRET). These experiments show that FRET is taking place
between GFP and mKate2 in ciliated cells (Figures 3D–3D00).Control experiments using centrin GFP as the donor andmKate2
Deve
actin as an acceptor fail to detect FRET, as expected, and show
that centrin does not interact with actin, in agreement with the
localization data (Figures 3E–3E00). Spot bleaching experiments
confirm that FRET is restricted to the areas containing basal
bodies (Figures S2A–S2A0 and Figure S2B). These results pro-
vide evidence that FA proteins form complexes that we termed
CAs and that CAs are connecting each basal body to the actin
network.
Given the proposed role of actin in basal body transport and
the close association of basal bodies and actin during this pro-
cess, we wanted to explore the possibility that CAs are present
during basal body migration (Boisvieux-Ulrich et al., 1990;
Ioannou et al., 2013). Imaging basal bodies (centrin2 RFP) during
ciliated cell intercalation revealed that CAs (marked by GFP Pax-
illin) are present during basal body migration, suggesting that
CAs are likely connecting basal bodies to the internal actin
network thought to be responsible for basal body transport
(Figures 4A–4A00 0 and 4B) (Ioannou et al., 2013).
Since basal bodies also connect to the subapical actin
network through the ciliary rootlets, we went on to examine
whether FA proteins are also involved in making this connection.
As shown in Figure 4C0, we found a second complex forming
below the basal bodies, in close association with the subapical
actin, suggesting that CAs are responsible for the connection
between the ciliary rootlets and the subapical actin network.
Coexpression of GFP FAK with clamp RFP and centrin2 CFP
confirms that FAK is found both in association with the basal
bodies, at the apical surface, as well as at the end of the ciliary
rootlets (Figures 4D and 4D0).In order to get a better resolution of the association of the
CAs with the actin cytoskeleton, both apical and subapical,
lopmental Cell 28, 70–80, January 13, 2014 ª2014 Elsevier Inc. 73

Figure 4. CA Complexes Link Basal Bodies
and Ciliary Rootlets to the Subapical Actin
Network
(A–A00 0) Optical sections of an intercalating multi-
ciliated cell expressing GFP Paxillin and centrin2
RFP showing that Paxillin is associated with basal
bodies during their migration to the apical surface.
(B) 3D reconstruction (y-z) of optical sections of
the intercalating cell in (A)–(A00 0).(C and C0) Multiciliated cell expressing GFP FAK
and stained with phalloidin. GFP FAK exhibits the
highest density at two focal planes. The first one
corresponds to the plane of the apical actin
network (C), while the second one appears slightly
below, at the plane of the subapical actin network
(C0). GFP FAK is in close association with both
pools of actin.
(D) Maximum intensity projection of a region of
a multiciliated cell coexpressing centrin2 RFP
(to label basal bodies), clamp RFP (to label the
striated rootlet), and GFP FAK. GFP FAK exhibits
two intensity maxima: one at the level of the cell’s
apical surface, adjacent to the basal bodies, and
a second one deeper, at the end of the ciliary
rootlets.
(D0) Confocal image from a mechanically
sectioned (along the apicobasal axis) multiciliated
cell coexpressing centrin2 CFP, clamp RFP, and
GFP FAK. The arrows mark the two maxima of
GFP FAK, one associated with the basal body and
one associated with the end of the striated rootlet.
(E) Optical section of a multiciliated cell coexpressing centrin2 CFP and mKate2 FAK and labeled with phalloidin (apical surface is at the top). The cell was initially
mechanically sectioned, approximately along its anterior-posterior axis. The subapical actin, known to connect each basal body with the end of the striated
rootlet of the cilium behind (Werner et al., 2011), appears to project from the CAs (red-labeled with mKate2 FAK) and form the characteristic discontinuous
network.
(F) Optical section of a multiciliated cell expressing mKate2 FAK and labeled with phalloidin (apical surface is at the top). The cell was initially mechanically
sectioned, approximately along its left-to-right axis. The subapical actin appears to connect CAs of neighboring basal bodies (along the left-right axis), creating
continuous loops. Again, the subapical actin appears to project from the CAs.
(G) Multiciliated cell coexpressing GFP FRNK and centrin2 RFP. GFP FRNK associates with the basal bodies in a similar way as the full-length protein. It is
localized at the posterior site of each basal body and exhibits an anterior-posterior gradient within the cell.
(H) Multiciliated cell coexpressing Paxillin C GFP and centrin2 RFP. Paxillin C GFP localizes at the posterior site of basal bodies and displays a gradient with
respect to the anterior-posterior axis of the cell.
Developmental Cell
CAs Link Basal Bodies to the Actin Network
we sectioned embryos expressing mKate2 FAK and stained
them with phalloidin. This led to an improvement of the resolu-
tion in the apicobasal axis, since lateral resolution is higher than
axial in confocal systems. We observed that the subapical
network in these cells is projecting from the CAs basally and
toward the back of the cell. We also observed that the subapi-
cal network, as described elsewhere (Werner et al., 2011),
is connecting each basal body to the one immediately behind
and in front through the rootlet, creating the characteristic
discontinuous actin network shown in Figure 4E. However,
we also noted an additional subapical network, rotated 90�
with respect to the first, creating loops between the CAs of
adjacent basal bodies (Figure 4F). This network appears to
connect basal bodies in the left-to-right direction and is
probably responsible for the spacing of basal bodies in this
direction, while the discontinuous front-to-back subapical
network is responsible for the spacing of basal bodies front
to back. Both networks appear to be originating at apical
CAs and are linked subapically at a nexus, presumably the
subapical region showing elevated FAK concentrations at the
end of the clamp-positive domain.
74 Developmental Cell 28, 70–80, January 13, 2014 ª2014 Elsevier In
FAK is known to localize to FAs through its C-terminal region
and, specifically, the FAT domain. This localization is believed
to be strongly dependent on FAK’s interaction with Paxillin,
whichmaps at the C terminus of FAK (Hayashi et al., 2002; Hilde-
brand et al., 1993, 1995). We went on to explore the domains
required for FAK localization at the basal bodies. As shown in
Figure 4G, the FAK C terminus, including the two proline-rich
sequences and the FAT domain, (FRNK: FAK-Related Non-
Kinase) is sufficient for the basal body localization of the protein.
Similarly, the C terminus of Paxillin is also sufficient to drive FA
localization of the protein (Brown et al., 1996) and, as shown in
Figure 4H, is also sufficient for basal body localization, showing
that, for both proteins, the determinants for CA complex locali-
zation are similar to those for FA localization, further emphasizing
the similarity between the two complexes.
Morpholino-Based FAK Downregulation BlocksCiliogenesisAlthough the localization and FRET data are strongly sugges-
tive of the function of the CA complex, we wanted to determine
what effects the disruption of the complex would have on
c.
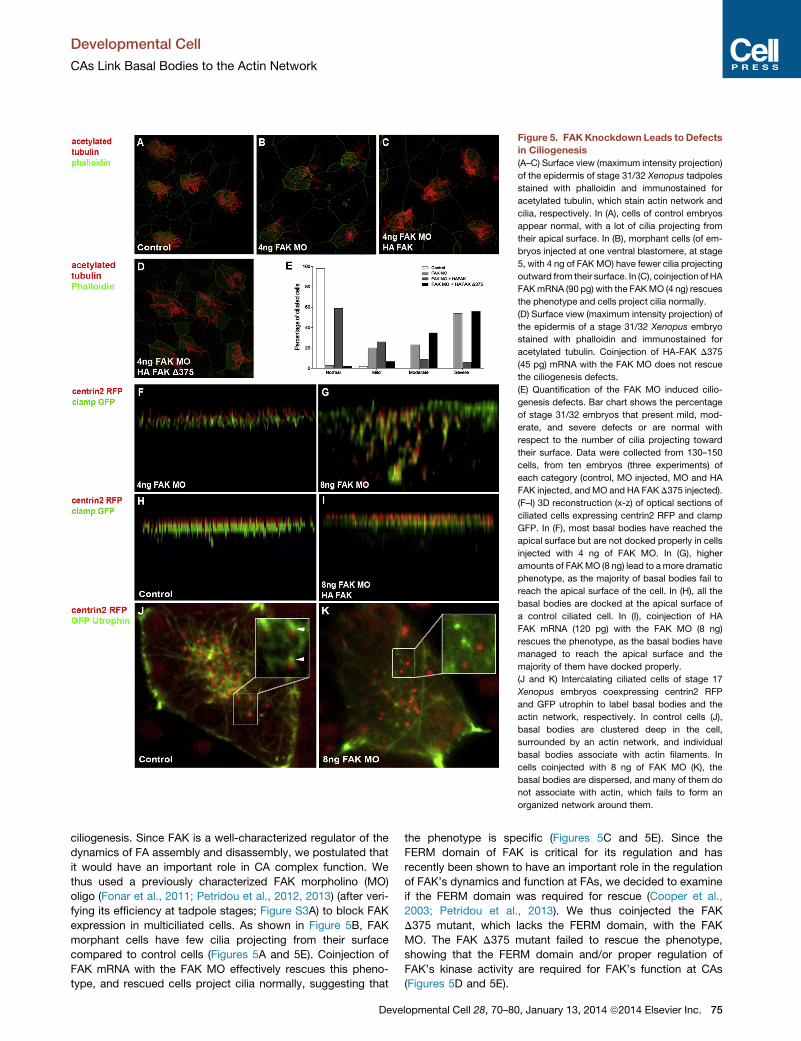
Figure 5. FAKKnockdown Leads to Defects
in Ciliogenesis
(A–C) Surface view (maximum intensity projection)
of the epidermis of stage 31/32 Xenopus tadpoles
stained with phalloidin and immunostained for
acetylated tubulin, which stain actin network and
cilia, respectively. In (A), cells of control embryos
appear normal, with a lot of cilia projecting from
their apical surface. In (B), morphant cells (of em-
bryos injected at one ventral blastomere, at stage
5, with 4 ng of FAK MO) have fewer cilia projecting
outward from their surface. In (C), coinjection ofHA
FAKmRNA (90 pg) with the FAKMO (4 ng) rescues
the phenotype and cells project cilia normally.
(D) Surface view (maximum intensity projection) of
the epidermis of a stage 31/32 Xenopus embryo
stained with phalloidin and immunostained for
acetylated tubulin. Coinjection of HA-FAK D375
(45 pg) mRNA with the FAK MO does not rescue
the ciliogenesis defects.
(E) Quantification of the FAK MO induced cilio-
genesis defects. Bar chart shows the percentage
of stage 31/32 embryos that present mild, mod-
erate, and severe defects or are normal with
respect to the number of cilia projecting toward
their surface. Data were collected from 130–150
cells, from ten embryos (three experiments) of
each category (control, MO injected, MO and HA
FAK injected, andMO and HA FAK D375 injected).
(F–I) 3D reconstruction (x-z) of optical sections of
ciliated cells expressing centrin2 RFP and clamp
GFP. In (F), most basal bodies have reached the
apical surface but are not docked properly in cells
injected with 4 ng of FAK MO. In (G), higher
amounts of FAKMO (8 ng) lead to amore dramatic
phenotype, as the majority of basal bodies fail to
reach the apical surface of the cell. In (H), all the
basal bodies are docked at the apical surface of
a control ciliated cell. In (I), coinjection of HA
FAK mRNA (120 pg) with the FAK MO (8 ng)
rescues the phenotype, as the basal bodies have
managed to reach the apical surface and the
majority of them have docked properly.
(J and K) Intercalating ciliated cells of stage 17
Xenopus embryos coexpressing centrin2 RFP
and GFP utrophin to label basal bodies and the
actin network, respectively. In control cells (J),
basal bodies are clustered deep in the cell,
surrounded by an actin network, and individual
basal bodies associate with actin filaments. In
cells coinjected with 8 ng of FAK MO (K), the
basal bodies are dispersed, and many of them do
not associate with actin, which fails to form an
organized network around them.
Developmental Cell
CAs Link Basal Bodies to the Actin Network
ciliogenesis. Since FAK is a well-characterized regulator of the
dynamics of FA assembly and disassembly, we postulated that
it would have an important role in CA complex function. We
thus used a previously characterized FAK morpholino (MO)
oligo (Fonar et al., 2011; Petridou et al., 2012, 2013) (after veri-
fying its efficiency at tadpole stages; Figure S3A) to block FAK
expression in multiciliated cells. As shown in Figure 5B, FAK
morphant cells have few cilia projecting from their surface
compared to control cells (Figures 5A and 5E). Coinjection of
FAK mRNA with the FAK MO effectively rescues this pheno-
type, and rescued cells project cilia normally, suggesting that
Deve
the phenotype is specific (Figures 5C and 5E). Since the
FERM domain of FAK is critical for its regulation and has
recently been shown to have an important role in the regulation
of FAK’s dynamics and function at FAs, we decided to examine
if the FERM domain was required for rescue (Cooper et al.,
2003; Petridou et al., 2013). We thus coinjected the FAK
D375 mutant, which lacks the FERM domain, with the FAK
MO. The FAK D375 mutant failed to rescue the phenotype,
showing that the FERM domain and/or proper regulation of
FAK’s kinase activity are required for FAK’s function at CAs
(Figures 5D and 5E).
lopmental Cell 28, 70–80, January 13, 2014 ª2014 Elsevier Inc. 75

Figure 6. Apical Actin Enrichment and
Rotational Polarity Are Unaffected in FAK
Morphants
(A–C) Confocal images of phalloidin-stained con-
trol (A), morphant (B), and rescued (C) ciliated cells
expressing centrin2 RFP plus enlarged close-ups
of selected regions, shown in (A0), (B0), and (C0 ),and a respective deeper optical section of each, in
(A00), (B00), and (C00), showing details of apical and
subapical actin networks. Morphants (B), unlike
controls (A) and rescued (C) cells, display reduced
numbers of basal bodies at the cell surface with
large areas completely devoid of basal bodies.
The apical actin network is present both in mor-
phants and controls; however, it appears less
organized and less dense in regions devoid of
basal bodies. The subapical actin of morphants
(B00) is completely missing in areas devoid of basal
bodies and appears disorganized in areas with
basal bodies.
(D and E) Centrin2 RFP- and clamp GFP-ex-
pressing control (D) and FAK morphant (E) multi-
ciliated cells. Spacing is disrupted, with regions
of the apical surface devoid of basal bodies in the
morphant cell. However, overall rotational polarity
is not significantly affected as most rootlets are
oriented in the same direction.
See also Figure S3.
Developmental Cell
CAs Link Basal Bodies to the Actin Network
Examination of centrin in side projections of morphant cells
(Figure 5F) reveals that, despite the fact that most basal bodies
reach the apical surface, they display variations in positioning
in the z axis, unlike the controls (Figure 5H), suggesting that
many of them do not dock. At higher doses of the FAK MO
(8 ng), basal body transport is impaired and the majority of
basal bodies remain deep in the cytosol (Figure 5G). Imaging
high-dose morphant cells during intercalation reveals that
basal bodies are often dispersed, rather than clustered like in
controls, and some clearly fail to associate with the internal actin
network, previously implicated in their transport (Boisvieux-
Ulrich et al., 1990; Ioannou et al., 2013), suggesting that the
transport defect is due to loss of association between the basal
bodies and the actin network (Figures 5J and 5K). Coinjection of
FAK mRNA also rescues the transport phenotype as shown in
Figure 5I.
In addition to the ciliogenesis and docking defects, FAK
morphants (4 ng) display spacing issues with regions of the
apical surface completely devoid of basal bodies (Figure 6B).
Despite the presence of apical actin enrichment in these
regions, the network appears less dense (Figure 6B0) and the
subapical actin is completely lost (Figure 6B00), unlike control
(Figures 6A–6A00) and rescued cells (Figures 6C–6C00), which pre-
sent uniformly organized apical and subapical actin networks.
In addition, although the apical actin of morphant cells appears
normal at regions with docked basal bodies, the subapical
actin appears less organized. Interestingly, the use of clamp
GFP with centrin2 RFP shows that rotational polarity, which is
76 Developmental Cell 28, 70–80, January 13, 2014 ª2014 Elsevier Inc.
dependent on the microtubule network,
remains relatively unaffected in FAK
morphants, especially when examining
local coordination in basal body clusters
(Figures 6D and 6E). These results suggest that FAK has an
important role in the regulation of the CA complex, and in addi-
tion, they are consistent with a role of this complex as a mecha-
nical link between basal bodies and the actin cytoskeleton.
Since the C terminus of FAK (FRNK) is sufficient for CA local-
ization, we postulated that, in a similar fashion as in FAs, it
would be able to act as a dominant negative by displacing
endogenous FAK (Gilmore and Romer, 1996; Hildebrand
et al., 1993; Schaller et al., 1992; Taylor et al., 2001). We
went on to overexpress FRNK in order to further address the
role of FAK and CAs during ciliogenesis. Expression of FRNK
partially displaces FAK from CAs but fails to induce strong
ciliogenesis defects (Figures S3B and S3C; data not shown).
In FRNK-expressing cells, most basal bodies reach the apical
surface and project cilia; however, in these cells, basal body
spacing is disrupted (Figures S3D–S3F; data not shown). Since
the subapical actin network has been shown to be responsible
for basal body spacing, this result supports a role of the CA
complex linking basal bodies to the actin network (Werner
et al., 2011). The reduced severity of the FRNK phenotype
compared to the FAK MO can be explained by the fact that
FRNK fails to displace all endogenous FAK from CAs. In
addition, FRNK only blocks kinase-dependent functions of
FAK while retaining scaffolding functions that depend on the
C-terminal FAT domain and the two proline-rich regions of
the protein (Gilmore and Romer, 1996; Hildebrand et al.,
1993; Taylor et al., 2001). This result is in agreement with a
role of CA complexes as a mechanical link between actin and

Figure 7. CA Complexes in Multiciliated Cells
(A) A diagram showing the localization of the CA complexes with respect to
the basal bodies, ciliary rootlets, and the actin cytoskeleton in multiciliated
cells. CA complexes interact with the apical and subapical pools of actin.
The upper red circle shows an x-z projection of confocal optical sections of
CA complexes and basal bodies. CAs are marked with GFP FAK (green) and
the basal bodies with centrin2 RFP (red).
(B) Proposed arrangement of the subapical actin network in multiciliated cells
in relation to the basal bodies, rootlets, and CAs.
Developmental Cell
CAs Link Basal Bodies to the Actin Network
the basal bodies and, in addition, suggests that FAK has
important kinase-independent roles at the CAs.
DISCUSSION
Overall, this work reveals a function of FA proteins in the
creation of complexes responsible for anchoring basal bodies
to the actin network of multiciliated cells. In addition, our results
show that CAs also form at the ends of ciliary rootlets and
are responsible for connecting the subapical actin network to
the ciliary rootlet. We also show that the subapical network
has two components. One discontinuous network in the front-
to-back direction connects each basal body to the one in front
and the one behind through the ciliary rootlet; and a continuous
network, at a right angle in relation to the first, connects
adjacent basal bodies in the left-to-right direction. This network
is probably responsible for spacing of basal bodies in the
left-to-right orientation while the discontinuous front-to-back
Deve
network determines spacing in the anterior-posterior axis of
the cell. The location of CAs with respect to the basal bodies,
rootlets, and the actin network is summarized in Figure 7.
The localization of the apical CA complex at the cell’s posterior
makes the most sense mechanically, since the maximum
force would be excreted by the cilium on the basal bodies
during the effective stroke rather than the recovery stroke,
pushing the basal bodies in the opposite direction of the stroke.
Linking the basal body to the actin network on this area would
ensure the immobilization of the basal body during the effective
stroke.
The similarities between CAs and FAs are striking, and the
determinants for Paxillin and FAK localization on CAs appear
to be similar to those for FA localization, although a more
detailed analysis will be required to determine if they are, in
fact, identical. FAK has been shown to be a critical regulator
of FA assembly and disassembly. Loss of FAK activity leads
to aberrant ciliogenesis and defects that are dependent on
actin-based processes, like basal body transport, docking,
and spacing, without affecting apical actin enrichment. These
results suggest that FAK, a critical regulator of FAs, is also
an important regulator of CAs, and they are consistent with
a role of this complex in the mechanical linkage of basal
bodies to the actin cytoskeleton. The proximity of the apical
CAs to the basal foot also raises the possibility that CAs are
also connecting basal bodies to the microtubule network,
shown to be responsible for establishing the rotational polarity
within multiciliated cells. However, FAK morphants show no
rotational polarity defects and, in this respect, resemble
cytochalasin-treated cells where spacing is affected but
local rotational polarity is maintained, making it unlikely that
CAs are linking basal bodies to microtubules (Werner et al.,
2011).
The presence of CA complexes in both motile and primary
cilia suggests that these proteins serve a highly conserved
function. The role of the actin cytoskeleton is well documented
both for motile cilia and primary cilia, as well as the flagella
of unicellular eukaryotes (Dentler and Adams, 1992; Engel
et al., 2011; Kim et al., 2010; Paredez et al., 2011; Sharma
et al., 2011). Interestingly, several core FA proteins have
been found in flagellated unicellular eukaryotes, including
Apusozoa and Choanoflagellates, the closest sister group
of Metazoa (Sebe-Pedros et al., 2010). Flagellar motility is
found in every major eukaryotic group (Fritz-Laylin et al.,
2010) and is believed to be an ancestral feature present
in the last common ancestor of all eukaryotic organisms
(Carvalho-Santos et al., 2011; Fritz-Laylin et al., 2010; Mitchell,
2007). This raises the possibility that the core FA proteins
originally evolved to perform functions at flagella and were
later co-opted for use in cell adhesion. Further support for
this notion comes from the fact that FA proteins were lost in
all fungi, with the exception of chytrid fungi, which are the
only members of the kingdom with flagellated gametes,
suggesting an important function for FA proteins in flagellar
function, in agreement with our results (Carvalho-Santos
et al., 2011). Future work will focus on examining the possibility
that CAs also exist in flagellated unicellular eukaryotes and will
also focus on addressing the precise role of individual proteins
within this complex.
lopmental Cell 28, 70–80, January 13, 2014 ª2014 Elsevier Inc. 77

Developmental Cell
CAs Link Basal Bodies to the Actin Network
EXPERIMENTAL PROCEDURES
Embryo Manipulations and Microinjections
Female adult Xenopus laevis were ovulated by injection of human chorionic
gonadotropin. Eggs were fertilized in vitro, dejellied in 2% cysteine (pH 7.8),
and subsequently reared in 0.1 3 Marc’s modified ringers (MMR) and staged
according to Neiuwkoop and Faber (Nieuwkoop and Faber, 1994). For micro-
injections, embryos were placed in a solution of 4% Ficoll in 0.33 3MMR and
injected using a glass capillary pulled needle, forceps, a Singer Instruments
MK1 micromanipulator, and a Harvard Apparatus pressure injector. After
injections, embryos were reared for 2 hr or until stage 8 in 4% Ficoll in
0.33 3 MMR and then washed and maintained in 0.1 3 MMR. For most
experiments, injections were made into the ventral blastomeres to target the
epidermis at the 4-cell, 8-cell, or 16-cell stage. For all experiments, we injected
morpholinos at 4 to 8 ng per blastomere and mRNAs at various amounts.
Embryos were allowed to develop to the appropriate stage and then imaged
live or fixed in MEMFA (Sive et al., 2010) for 1–2 hr at room temperature
(RT). Fixed embryos were used immediately. For live imaging, embryos were
anesthetized in 0.01% benzocaine in 0.1 3 MMR.
Immunostaining
Indirect immunofluorescence assays were carried out as described elsewhere
(Demetriou et al., 2008; Skourides et al., 1999) with modifications. NIH 3T3
cells were seeded on glass coverslips (charged with HCl), washed three
times with PBS, preextracted with 0.2% Triton X-100 for 20 s, and then
fixed for 10 min in 4% paraformaldehyde solution in PBS. Fixation was
followed by addition of 50 mM glycine solution in PBS, and then the cells
were blocked using 10% normal goat serum (Jackson Immunoresearch) for
30 min. Cells were incubated with primary antibodies diluted in 10% normal
goat serum solution in PBS for 1.5 hr. The primary antibodies used were
Paxillin mouse monoclonal (BD Transduction Laboratories) used in com-
bination with Acetyl-a-Tubulin (Lys40) rabbit polyclonal (Cell Signaling) and
g Tubulin goat polyclonal (Santa Cruz Biotechnology); and FAK [pS732]
Phosphospecific rabbit polyclonal (Invitrogen) used in combination with
Acetylated tubulin, mouse monoclonal (Santa Cruz Biotechnology) and g
Tubulin goat polyclonal (Santa Cruz Biotechnology). Cells were then washed
five times in PBS. Secondary antibodies used were Cy3 anti-mouse and
anti-rabbit (Jackson Immunoresearch), Alexa 488 anti-goat (Molecular
Probes Invitrogen), and IgG-CFL 647 anti-mouse and anti-rabbit (Santa Cruz
Biotechnology).
For whole-mount immunostaining, fixed embryos were permeabilized in
PBDT (1 3 PBS + 0.5% Triton X-100 + 1% DMSO) for several hours at
RT and blocked in PBDT + 1% normal goat serum for 1 hr at RT. Primary
antibodies were added in block solution, and embryos were incubated for
4 hr at RT or overnight at 4�C. The primary antibodies used were: Acetylated
tubulin mouse monoclonal (Santa Cruz Biotechnology), Vinculin mouse
monoclonal (Developmental Studies Hybridoma Bank), and FAK pS732 rabbit
polyclonal (Invitrogen). The next day, embryos were washed 4 3 10 min in
PBDT. Embryos were then incubated in secondary antibodies—anti-mouse
Alexa 488 (Invitrogen) or anti-mouse IgG-CFL 647 (Santa Cruz Biotech-
nology)—at RT for 1–2 hr. Actin labeling was performed using Alexa Fluor
Phalloidin 488 (Invitrogen) and Phalloidin CruzFluor 647 (Santa Cruz Biotech-
nology) at RT for 1–2 hr. Embryos were then washed 4 3 10 min in PBDT.
Embryos were postfixed in MEMFA for 10-15 min at RT, washed in 1 3 PBS
and imaged immediately.
Western Blot Analysis
Protein lysates were prepared by homogenizing embryos (by pipetting up and
down) in ice-cold MK’s modified lysis buffer (50 mM Tris, pH 8.0, 150 mM
NaCl, 0.5% NP-40, 0.5% Triton X-100, 100 mM EGTA, 5 mM NaF) supple-
mented with protease inhibitors (1 mM phenylmethylsulfonyl fluoride and
protease cocktail; Sigma). Homogenates were cleared by centrifugation at
15,000 3 g for 30 min at 4�C (Kragtorp and Miller, 2006). Proteins (3/4–1 em-
bryo equivalent) were run on 7.5% SDS-polyacrylamide gels with the West-
ernC ladder (Bio-Rad) and blotted on to nitrocellulose membranes. Blots
were blocked in 5% milk (in TBSTw: 13 TBS buffer and 0.1% Tween 20)
and then incubated with the primary antibodies in 5% milk overnight at 4�C.The primary antibodies used were: FAK mouse monoclonal (Millipore), Paxillin
78 Developmental Cell 28, 70–80, January 13, 2014 ª2014 Elsevier In
mouse monoclonal (BD Transduction Laboratories), and Vinculin mouse
monoclonal (Developmental Studies Hybridoma Bank). Visualization was
performed using horseradish peroxidase-conjugated antibodies (anti-rabbit
and anti-mouse; Santa Cruz Biotechnology), after 1 hr incubation at RT, and
signal was detected using LumiSensor (GeneScript) on a UVP BioSpectrum
Imaging System. For loading control, actin rabbit polyclonal antibody (Santa
Cruz Biotechnologies) was used.
GRP Assay
Embryos were fixed in MEMFA at stage 17. The GRP was manually dissected
and postfixed for a further 15 min. GRP tissue was then used for immunofluo-
rescence as described earlier.
Imaging and Quantification
Imaging was performed on a Zeiss LSM 710 laser scanning confocal micro-
scope with Zen 2010 software.
Quantification of the ciliogenesis defects was conducted based on the
number of cilia projecting from the apical surface of each cell. Cells have
been denoted as normal, with mild phenotype (having more than 50% of their
cilia but less than control cells), with moderate phenotype (having 50% or
lower reduction of their cilia), and with severe phenotype (having near com-
plete loss of their cilia).
Quantification of the basal body distribution was conducted by overlaying
a grid over each cell and counting the number of basal bodies included in
each cell of the grid, using Adobe Photoshop CS2 software.
Acceptor Photobleaching FRET
FRET experiments were accomplished using a laser scanning confocal micro-
scope (Zeiss LSM 710) with a Plan-Apochromat 633/1.40 Oil DIC M27 objec-
tive lens (Zeiss). Stage 33–35 embryos expressing GFP FAK and mKate2 actin
or centrin GFP and mKate2 actin were anesthetized in 0.01% benzocaine in
MMR and immobilized in silicone grease wells on glass slides. A 543 nm laser
was used for acceptor (mKate2 actin) photobleaching (100% power) within a
region of interest (ROI). A 488 nm laser (0.8% power) was used for acquisition
of GFP FAK, and emission was detected between 493 and 538 nm. A 543 nm
laser (1.2% power) was used for acquisition of mKate2 actin, and emission
was detected between 599 and 758 nm. One frame was acquired as a pre-
bleaching control, and the ROI (rectangular region) was bleached within one
frame. Zeiss Zen 2010 software was used for FRET analysis. FRET efficiency
was calculated using the following equation: FRETEfficiency = (Donor Post �Donor Pre)/(Donor Post) 3 100 (Donor Post: donor emission intensity after
photobleaching; Donor Pre: donor emission intensity before photobleaching).
SUPPLEMENTAL INFORMATION
Supplemental Information includes Supplemental Experimental Procedures
and three figures and can be found with this article online at http://dx.doi.
org/10.1016/j.devcel.2013.12.003.
ACKNOWLEDGMENTS
We thank Drs. JohnWallingford, Brian Mitchell, and Reinhard Koster for kindly
providing plasmids. We also thank Dr. Niovi Santama for providing reagents
for the immunofluorescence experiments on NIH 3T3 cells. Finally, we thank
Charalambos Kourouklaris for generating the 3D reconstruction of the pro-
posed model. Funding was provided by the Cyprus Research Promotion
Foundation (YGEIA/BIOS/0609(BE)/14, TEXNOLOGIA/YLIKA/0311(BIE)/10).
Accepted: December 6, 2013
Published: January 13, 2014
REFERENCES
Afzelius, B.A. (1976). A human syndrome caused by immotile cilia. Science
193, 317–319.
Afzelius, B.A. (2004). Cilia-related diseases. J. Pathol. 204, 470–477.
c.

Developmental Cell
CAs Link Basal Bodies to the Actin Network
Baker, K., and Beales, P.L. (2009). Making sense of cilia in disease: the human
ciliopathies. Am. J. Med. Genet. C. Semin. Med. Genet. 151C, 281–295.
Basto, R., Lau, J., Vinogradova, T., Gardiol, A., Woods, C.G., Khodjakov, A.,
and Raff, J.W. (2006). Flies without centrioles. Cell 125, 1375–1386.
Bershteyn, M., Atwood, S.X., Woo, W.M., Li, M., and Oro, A.E. (2010). MIM
and cortactin antagonism regulates ciliogenesis and hedgehog signaling.
Dev. Cell 19, 270–283.
Bisgrove, B.W., and Yost, H.J. (2006). The roles of cilia in developmental
disorders and disease. Development 133, 4131–4143.
Boisvieux-Ulrich, E., Laine, M.C., and Sandoz, D. (1990). Cytochalasin D
inhibits basal body migration and ciliary elongation in quail oviduct epithelium.
Cell Tissue Res. 259, 443–454.
Brown, M.C., Perrotta, J.A., and Turner, C.E. (1996). Identification of LIM3
as the principal determinant of paxillin focal adhesion localization and charac-
terization of a novel motif on paxillin directing vinculin and focal adhesion
kinase binding. J. Cell Biol. 135, 1109–1123.
Carvalho-Santos, Z., Azimzadeh, J., Pereira-Leal, J.B., and Bettencourt-Dias,
M. (2011). Evolution: Tracing the origins of centrioles, cilia, and flagella. J. Cell
Biol. 194, 165–175.
Chailley, B., Nicolas, G., and Laine, M.C. (1989). Organization of actin micro-
filaments in the apical border of oviduct ciliated cells. Biol. Cell 67, 81–90.
Cooper, L.A., Shen, T.L., and Guan, J.L. (2003). Regulation of focal adhesion
kinase by its amino-terminal domain through an autoinhibitory interaction.
Mol. Cell. Biol. 23, 8030–8041.
Dawe, H.R., Farr, H., and Gull, K. (2007). Centriole/basal body morphogenesis
and migration during ciliogenesis in animal cells. J. Cell Sci. 120, 7–15.
Demetriou, M.C., Stylianou, P., Andreou, M., Yiannikouri, O., Tsaprailis, G.,
Cress, A.E., and Skourides, P. (2008). Spatially and temporally regulated
alpha6 integrin cleavage during Xenopus laevis development. Biochem.
Biophys. Res. Commun. 366, 779–785.
Dentler, W.L., and Adams, C. (1992). Flagellar microtubule dynamics in
Chlamydomonas: cytochalasin D induces periods of microtubule shortening
and elongation; and colchicine induces disassembly of the distal, but not
proximal, half of the flagellum. J. Cell Biol. 117, 1289–1298.
Engel, B.D., Ishikawa, H., Feldman, J.L., Wilson, C.W., Chuang, P.T.,
Snedecor, J., Williams, J., Sun, Z., and Marshall, W.F. (2011). A cell-based
screen for inhibitors of flagella-driven motility in Chlamydomonas reveals a
novel modulator of ciliary length and retrograde actin flow. Cytoskeleton
(Hoboken) 68, 188–203.
Fonar, Y., Gutkovich, Y.E., Root, H., Malyarova, A., Aamar, E., Golubovskaya,
V.M., Elias, S., Elkouby, Y.M., and Frank, D. (2011). Focal adhesion kinase pro-
tein regulates Wnt3a gene expression to control cell fate specification in the
developing neural plate. Mol. Biol. Cell 22, 2409–2421.
Fritz-Laylin, L.K., Prochnik, S.E., Ginger, M.L., Dacks, J.B., Carpenter, M.L.,
Field, M.C., Kuo, A., Paredez, A., Chapman, J., Pham, J., et al. (2010). The
genome of Naegleria gruberi illuminates early eukaryotic versatility. Cell 140,
631–642.
Gilmore, A.P., and Romer, L.H. (1996). Inhibition of focal adhesion kinase (FAK)
signaling in focal adhesions decreases cell motility and proliferation. Mol. Biol.
Cell 7, 1209–1224.
Goetz, S.C., and Anderson, K.V. (2010). The primary cilium: a signalling centre
during vertebrate development. Nat. Rev. Genet. 11, 331–344.
Hayashi, I., Vuori, K., and Liddington, R.C. (2002). The focal adhesion targeting
(FAT) region of focal adhesion kinase is a four-helix bundle that binds paxillin.
Nat. Struct. Biol. 9, 101–106.
Herreros, L., Rodrıguez-Fernandez, J.L., Brown, M.C., Alonso-Lebrero, J.L.,
Cabanas, C., Sanchez-Madrid, F., Longo, N., Turner, C.E., and Sanchez-
Mateos, P. (2000). Paxillin localizes to the lymphocyte microtubule organizing
center and associates with the microtubule cytoskeleton. J. Biol. Chem. 275,
26436–26440.
Hildebrand, J.D., Schaller, M.D., and Parsons, J.T. (1993). Identification of
sequences required for the efficient localization of the focal adhesion kinase,
pp125FAK, to cellular focal adhesions. J. Cell Biol. 123, 993–1005.
Deve
Hildebrand, J.D., Schaller, M.D., and Parsons, J.T. (1995). Paxillin, a tyrosine
phosphorylated focal adhesion-associated protein binds to the carboxyl
terminal domain of focal adhesion kinase. Mol. Biol. Cell 6, 637–647.
Hildebrandt, F., Benzing, T., and Katsanis, N. (2011). Ciliopathies. N. Engl. J.
Med. 364, 1533–1543.
Ioannou, A., Santama, N., and Skourides, P.A. (2013). Xenopus laevis
nucleotide binding protein 1 (xNubp1) is important for convergent extension
movements and controls ciliogenesis via regulation of the actin cytoskeleton.
Dev. Biol. 380, 243–258.
Kim, J., Lee, J.E., Heynen-Genel, S., Suyama, E., Ono, K., Lee, K., Ideker, T.,
Aza-Blanc, P., and Gleeson, J.G. (2010). Functional genomic screen for
modulators of ciliogenesis and cilium length. Nature 464, 1048–1051.
Klotz, C., Bordes, N., Laine, M.C., Sandoz, D., and Bornens, M. (1986). Myosin
at the apical pole of ciliated epithelial cells as revealed by a monoclonal
antibody. J. Cell Biol. 103, 613–619.
Knowles, M.R., and Boucher, R.C. (2002). Mucus clearance as a primary
innate defense mechanism for mammalian airways. J. Clin. Invest. 109,
571–577.
Kragtorp, K.A., and Miller, J.R. (2006). Regulation of somitogenesis by
Ena/VASP proteins and FAK during Xenopus development. Development
133, 685–695.
Lemullois, M., Klotz, C., and Sandoz, D. (1987). Immunocytochemical locali-
zation of myosin during ciliogenesis of quail oviduct. Eur. J. Cell Biol. 43,
429–437.
Lemullois, M., Boisvieux-Ulrich, E., Laine, M.C., Chailley, B., and Sandoz, D.
(1988). Development and functions of the cytoskeleton during ciliogenesis
in metazoa. Biol. Cell 63, 195–208.
Mitchell, D.R. (2007). The evolution of eukaryotic cilia and flagella as motile
and sensory organelles. Adv. Exp. Med. Biol. 607, 130–140.
Mitchell, B., Jacobs, R., Li, J., Chien, S., and Kintner, C. (2007). A positive
feedback mechanism governs the polarity and motion of motile cilia. Nature
447, 97–101.
Mitchell, B., Stubbs, J.L., Huisman, F., Taborek, P., Yu, C., and Kintner, C.
(2009). The PCP pathway instructs the planar orientation of ciliated cells in
the Xenopus larval skin. Curr. Biol. 19, 924–929.
Nieuwkoop, P.D.F., and Faber, J. (1994). Normal Table of Xenopus laevis
(Daudin): A Systematical and Chronological Survey of the Development from
the Fertilized Egg till the End of Metamorphosis, First Edition. (New York:
Garland).
Pan, J., You, Y., Huang, T., and Brody, S.L. (2007). RhoA-mediated apical
actin enrichment is required for ciliogenesis and promoted by Foxj1.
J. Cell Sci. 120, 1868–1876.
Paredez, A.R., Assaf, Z.J., Sept, D., Timofejeva, L., Dawson, S.C., Wang, C.J.,
and Cande, W.Z. (2011). An actin cytoskeleton with evolutionarily conserved
functions in the absence of canonical actin-binding proteins. Proc. Natl.
Acad. Sci. USA 108, 6151–6156.
Park, T.J., Haigo, S.L., and Wallingford, J.B. (2006). Ciliogenesis defects in
embryos lacking inturned or fuzzy function are associated with failure of
planar cell polarity and Hedgehog signaling. Nat. Genet. 38, 303–311.
Park, T.J., Mitchell, B.J., Abitua, P.B., Kintner, C., andWallingford, J.B. (2008).
Dishevelled controls apical docking and planar polarization of basal bodies in
ciliated epithelial cells. Nat. Genet. 40, 871–879.
Park, A.Y., Shen, T.L., Chien, S., and Guan, J.L. (2009). Role of focal adhesion
kinase Ser-732 phosphorylation in centrosome function during mitosis. J. Biol.
Chem. 284, 9418–9425.
Pazour, G.J., and Witman, G.B. (2003). The vertebrate primary cilium is a
sensory organelle. Curr. Opin. Cell Biol. 15, 105–110.
Pedersen, L.B., Veland, I.R., Schrøder, J.M., and Christensen, S.T. (2008).
Assembly of primary cilia. Dev. Dyn. 237, 1993–2006.
Petridou, N.I., Stylianou, P., Christodoulou, N., Rhoads, D., Guan, J.L., and
Skourides, P.A. (2012). Activation of endogenous FAK via expression of its
amino terminal domain in Xenopus embryos. PLoS ONE 7, e42577.
lopmental Cell 28, 70–80, January 13, 2014 ª2014 Elsevier Inc. 79

Developmental Cell
CAs Link Basal Bodies to the Actin Network
Petridou, N.I., Stylianou, P., and Skourides, P.A. (2013). A dominant-negative
provides new insights into FAK regulation and function in early embryonic
morphogenesis. Development 140, 4266–4276.
Ravanelli, A.M., and Klingensmith, J. (2011). The actin nucleator Cordon-bleu
is required for development of motile cilia in zebrafish. Dev. Biol. 350, 101–111.
Roy, S. (2009). The motile cilium in development and disease: emerging new
insights. Bioessays 31, 694–699.
Sawamoto, K., Wichterle, H., Gonzalez-Perez, O., Cholfin, J.A., Yamada, M.,
Spassky, N., Murcia, N.S., Garcia-Verdugo, J.M., Marin, O., Rubenstein,
J.L., et al. (2006). New neurons follow the flow of cerebrospinal fluid in the
adult brain. Science 311, 629–632.
Schaller, M.D., Borgman, C.A., Cobb, B.S., Vines, R.R., Reynolds, A.B., and
Parsons, J.T. (1992). pp125FAK a structurally distinctive protein-tyrosine
kinase associated with focal adhesions. Proc. Natl. Acad. Sci. USA 89,
5192–5196.
Sebe-Pedros, A., Roger, A.J., Lang, F.B., King, N., and Ruiz-Trillo, I. (2010).
Ancient origin of the integrin-mediated adhesion and signaling machinery.
Proc. Natl. Acad. Sci. USA 107, 10142–10147.
Shah, A.S., Ben-Shahar, Y., Moninger, T.O., Kline, J.N., and Welsh, M.J.
(2009). Motile cilia of human airway epithelia are chemosensory. Science
325, 1131–1134.
Sharma, N., Kosan, Z.A., Stallworth, J.E., Berbari, N.F., and Yoder, B.K. (2011).
Soluble levels of cytosolic tubulin regulate ciliary length control. Mol. Biol. Cell
22, 806–816.
Sive, H.L., Grainger, R.M., and Richard, M.H. (2010). Early Development of
Xenopus laevis: A Laboratory Manual. (Cold Spring Harbor, NY: Cold Spring
Harbor Laboratory Press).
Skourides, P.A., Perera, S.A., and Ren, R. (1999). Polarized distribution of
Bcr-Abl in migrating myeloid cells and co-localization of Bcr-Abl and its target
proteins. Oncogene 18, 1165–1176.
80 Developmental Cell 28, 70–80, January 13, 2014 ª2014 Elsevier In
Sorokin, S.P. (1968). Reconstructions of centriole formation and ciliogenesis
in mammalian lungs. J. Cell Sci. 3, 207–230.
Tamm, S., and Tamm, S.L. (1988). Development of macrociliary cells in Beroe.
I. Actin bundles and centriole migration. J. Cell Sci. 89, 67–80.
Taylor, J.M., Mack, C.P., Nolan, K., Regan, C.P., Owens, G.K., and Parsons,
J.T. (2001). Selective expression of an endogenous inhibitor of FAK regulates
proliferation and migration of vascular smooth muscle cells. Mol. Cell. Biol. 21,
1565–1572.
Wallingford, J.B. (2010). Planar cell polarity signaling, cilia and polarized ciliary
beating. Curr. Opin. Cell Biol. 22, 597–604.
Wallingford, J.B., and Mitchell, B. (2011). Strange as it may seem: the many
links between Wnt signaling, planar cell polarity, and cilia. Genes Dev. 25,
201–213.
Werner, M.E., Hwang, P., Huisman, F., Taborek, P., Yu, C.C., andMitchell, B.J.
(2011). Actin and microtubules drive differential aspects of planar cell polarity
in multiciliated cells. J. Cell Biol. 195, 19–26.
Worthington, W.C., Jr., and Cathcart, R.S., 3rd. (1963). Ependymal cilia: distri-
bution and activity in the adult human brain. Science 139, 221–222.
Yan, X., and Zhu, X. (2013). Branched F-actin as a negative regulator of cilia
formation. Exp. Cell Res. 319, 147–151.
Yin, Y., Bangs, F., Paton, I.R., Prescott, A., James, J., Davey, M.G.,Whitley, P.,
Genikhovich, G., Technau, U., Burt, D.W., and Tickle, C. (2009). The Talpid3
gene (KIAA0586) encodes a centrosomal protein that is essential for primary
cilia formation. Development 136, 655–664.
Zariwala, M.A., Knowles, M.R., and Omran, H. (2007). Genetic defects in
ciliary structure and function. Annu. Rev. Physiol. 69, 423–450.
c.


![The Actin Cytoskeleton: Functional Arrays forUpdate on the Actin Cytoskeleton The Actin Cytoskeleton: Functional Arrays for Cytoplasmic Organization and Cell Shape Control1[OPEN] Dan](https://img.pdfslide.us/doc/110x75/5f0830197e708231d420c69d/the-actin-cytoskeleton-functional-arrays-update-on-the-actin-cytoskeleton-the-actin.jpg)








![Actin cytoskeleton and cell motility - Indico [Home]indico.ictp.it/event/a10138/session/33/contribution/22/material/0/... · Actin cytoskeleton and cell motility Julie Plastino, UMR](https://img.pdfslide.us/doc/110x75/5bcc339f09d3f232618dcbfd/actin-cytoskeleton-and-cell-motility-indico-home-actin-cytoskeleton-and.jpg)









![CYTOSKELETON NEWS - fnkprddata.blob.core.windows.net · Dynamic remodeling of the actin cytoskeleton [i.e., rapid cycling between filamentous actin (F-actin) and monomer actin (G-actin)]](https://img.pdfslide.us/doc/110x75/609edd2b88630103265d18ee/cytoskeleton-news-dynamic-remodeling-of-the-actin-cytoskeleton-ie-rapid-cycling.jpg)






