Embed Size (px)
Citation preview

lable at ScienceDirect
Soil Biology & Biochemistry 43 (2011) 206e211
Contents lists avai
Soil Biology & Biochemistry
journal homepage: www.elsevier .com/locate/soi lbio
Macro-invertebrates accelerate litter decomposition and nutrient releasein a Hawaiian rainforest
Wallace M. Meyer III a,b,*, Rebecca Ostertag c, Robert H. Cowie a
aCenter for Conservation Research and Training, Pacific Biosciences Research Center, University of Hawaii at Manoa, 3050 Maile Way, Gilmore 408, Honolulu, HI 96822, USAbDepartment of Zoology, University of Hawaii at Manoa, 2538 McCarthy Mall, Edmonson 152, Honolulu, HI 96822, USAcDepartment of Biology, University of Hawaii at Hilo, 200 W. Kawili Street, Hilo, HI 96720, USA
a r t i c l e i n f o
Article history:Received 25 June 2010Received in revised form15 September 2010Accepted 4 October 2010Available online 16 October 2010
Keywords:Ecosystem processesLitter bagNitrogenManganeseEarthwormssnails/slugsArthropods
* Corresponding author. Present address: Departmeof Arizona, 1140 E. South Campus Drive, Forbes 410, Tu520 621 2847; fax: þ1 520 621 1150.
E-mail address: [email protected] (W.M. M
0038-0717/$ e see front matter � 2010 Elsevier Ltd.doi:10.1016/j.soilbio.2010.10.005
a b s t r a c t
Macro-invertebrates (>2 mm in size) can play a key role in litter decomposition by influencing litterchemistry and other components of the decomposer community, thus affecting rates of decomposition,nutrient release, and primary production. However, in many ecosystems the influences of macro-invertebrates on key ecosystem processes have not been adequately addressed. We investigated theinfluence of the macro-invertebrate community in litter decomposition and the cycling of nutrients ina young rainforest site on the island of Hawaii by using litter bags with and without 2.5 cm holes to allowor prevent access by macro-invertebrates. Presence of macro-invertebrates increased rates of litterdecomposition by 16.9% and rates of nutrient release for N and Mn by 33.2% and 30.3%, respectively.Macro-invertebrate activity thus has a major impact on N release accounting for 3.32 kg/ha/yr. Thisinternal ecosystem transfer of N from the litter is greater than estimates of nitrogen inputs from rainwater, dry deposition, volcanic sources, atmospheric dust, and nitrogen fixation for this ecosystem. Thesefindings demonstrate that improved knowledge of the ecosystem effects of macro-invertebrates isnecessary to understand how ecosystems function.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Litter decomposition is an important ecosystem process thatreleases carbon to the atmosphere and nutrients in forms that canbe used for plant and microbial production (Chapin et al., 2002;Duffy, 2002). Macro-invertebrates (invertebrates >2 mm in size)can play a key role in litter decomposition by influencing litterchemistry and other components of the decomposer community,thus affecting rates of decomposition, nutrient release, and primaryproduction (Swift et al., 1979; Jones et al., 1994; Belovsky and Slade,2000; Holdsworth et al., 2008). Depending on species compositionand abundances they can contribute directly (via metabolism) andindirectly (by modification of the habitat to enhance either micro-arthropod or microbial activity) to the decomposition of leaf litterand the cycling of nutrients (Jennings and Barkham, 1979; Newell,1967; Shachak et al., 1987; Schaefer, 1991; Theenhaus and Scheu,1996). Because soil animals only account for x5% of soil
nt of Entomology, Universitycson, AZ 85721, USA. Tel.: þ1
eyer III).
All rights reserved.
respiration, their major effect on decomposition has beenpresumed to be enhancement of microbial activity (Wall et al.,2001). As such, macro-invertebrates are considered ecosystemengineers, species that modify the soil or litter in ways that eitherpromote or constrain the activities of soil microbes and other soilanimals (Jones et al., 1994; Chapin et al., 2002; Groffman et al.,2004; Eisenhauer et al., 2007).
Despite the established importance of the macro-invertebrates,their influences on key processes in many ecosystems have notbeen adequately addressed. This is especially true in the HawaiianIslands, which are renowned as a model system for ecosystemstudies (Vitousek, 2004). Many studies have examined howecosystem dynamics change along substrate, temperature, andprecipitation gradients throughout the archipelago, with much ofthe research focused on the island of Hawaii (Vitousek et al., 1994;Crews et al., 1995; Austin and Vitousek, 2000; Austin, 2002). Specialattention has been paid to the effects of nutrient availability andquality on litter decomposition rates (Russell and Vitousek, 1997;Hobbie, 2000; Mack and D’Antonio, 2003). Although interactionsamong the flora and microbial communities have been assessedand discussed (Norman, 2008), the impact of animals on theseprocesses has rarely been mentioned. In the few studies involving

W.M. Meyer III et al. / Soil Biology & Biochemistry 43 (2011) 206e211 207
animals (e.g., Gruner, 2004; Sin et al., 2008), litter dwelling macro-invertebrates such as terrestrial molluscs, annelids, millipedes, andisopods were not included.
We investigated the influence of the macro-invertebratecommunity in litter decomposition and the cycling of nutrients ina young rainforest on the island of Hawaii by using litter bags withand without 2.5 cm holes to allow or prevent access by macro-invertebrates. Because young Hawaiian ecosystems tend to benitrogen limited (Vitousek, 2004), we were also interested inunderstanding if and to what extent macro-invertebrates influencenitrogen cycling so we estimated the amount of nitrogen releasedby the decomposition process (kg/ha/yr) in the two treatments. Wethen compared the amount of N released via this internalecosystem transfer to estimates of external ecosystem nitrogeninputs (e.g., rain water, dry deposition, volcanic sources, cloudwater, atmospheric dust, and nitrogen fixation). Within thiscontext, our results provide a better understanding of the role ofmacro-invertebrates in decomposition and N cycling processes inHawaiian ecosystems.
2. Methods
2.1. Study site
The study was conducted in the Upper Waiakea Forest Reserve(N 19.33479, W 155.15023) on the eastern (windward) side of theisland of Hawaii. The site is a wet forest with dense canopy coverthat typically receives an annual rainfall of 2500 to 5000 mm (Juvikand Juvik, 1998). It is a young Hawaiian forest (the last lava flow tocover the area was w2100 y ago) and as such is probably nitrogenlimited (Vitousek, 2004). The plant community is characterized bynative tree species (Metrosideros polymorpha, Cheirodendron trigy-num, Melicope spp.), mid-canopy tree ferns (Cibotium spp.), andunderstory plants (e.g., Broussaisia arguta and Peperomia sp.), andby the absence or extremely low density of the invasive tree speciesPsidium cattleianum and Morella faya, both of which can have largeecosystem effects (Vitousek et al., 1987; Mascaro et al., 2008). Sevensites were chosen within a w1000e1250 m elevational range.
2.2. Leaf litter decomposition experiment
Leaves were collected just after abscission between June 2007and December 2007 at all seven sites by placing plastic sheeting ateach site and collecting all fresh leaves every two weeks. Leaveswere then air dried for at least two weeks at room temperature.M.polymorpha and Cibotium spp. were the most abundant speciespresent, but all litter species collected were used and probablyencompassed a broad range of decomposability (Vitousek, 2004).The initial C:N ratio (�1 SE) of the mixed species litter was 49.2(�7.3) with carbon and nitrogen constituting 45.1 (�0.9) and 0.93(�0.11) percent of the litter, respectively.
To evaluate litter decomposition, litter was placed in bags,which were then deployed in the experimental treatments. Litterbags consisted of 4.0 � 0.02 g (mean air-dry mass � range) of leaflitter enclosed in a 10 � 10 � 8 cm plastic mesh food carton (toprovide a 3-dimensional framework) covered with plastic screen(2.0 mm mesh). Leaves from all plant species and sites were mixedthoroughly to ensure that a randommixture of leaf litter was addedto each bag. For Cibotium spp., the main rachis was removed andonly the compound leaves were placed in the litter bags.
The experiments consisted of two treatments applied at each ofthe seven sites to determine the influence of macro-invertebrateson litter decomposition and nutrient release: 1) macro-inverte-brates excluded, the litter decomposition bags having no holes, and2) macro-invertebrates allowed access via holes (2.5 cm diameter)
cut in the mesh on opposite sides of the bags. In total for these twotreatments, 350 litter bags were deployed (25 per treatment at eachsite). The bags were placed on top of the natural litter layer in pairsconsisting of one experimental and one control bag w0.1 m apart.These treatments ran from January 20, 2008 to January 9, 2009. Fivelitter bags were collected from each treatment at each site atmonths 1, 2, 4, 6, and 12.
Prior to processing, leaf litter was sorted to remove inverte-brates, roots, and large pieces of soil. All macro-invertebratescollected during sorting were identified to group: Mollusca,Annelida, Platyheminthes, and Arthropoda. Arthropods werefurther divided into Diplura, Dermaptera, Coleoptera, Isopoda, andDiplopoda. Litter samples were then dried at 50 �C for 48 h, andweighed to the nearest 0.001 g. Ash-free dry mass (AFDM) wasdetermined after grinding each sample in a Wiley mill (20 mesh)and then ashing one subsample (x0.5 g) at 500 �C for 4 h. Thisaccounted for any soil attached to the litter samples and permittedcalculation of the mass of litter remaining in each bag. Onesubsample from three litter bags from each treatment at each sitefor months 2, 4, 6, and 12 was analyzed for C, N, Ca, Mg, Mn, P, andK concentrations. Concentrations of C and N were determined ona Costech ECS 4010 Elemental Analyzer (Valencia, CA). For theother nutrients, 0.25 g dried sample was first ashed at 500 �C for5 h, cooled and then resuspended in 5 ml of 1 N HCl and allowedto digest for 0.5 h. Then 20 ml of reagent-grade water was addedto each sample and the samples were run on a Varian Vista MPXICP-OES Spectrometer (Palo Alto, CA). Peach leaves (NIST SRM1547) were used as an international standard and recovery aver-aged 92%.
2.3. Statistical analyses
Some macro-invertebrate groups may be able to access bothtreatments and some may not be willing to enter the litter bags atall. We therefore used c2 analyses to determine if abundances ofvarious macro-invertebrate groups differed in the two treatmenttypes.
To examine differences in litter decomposition and nutrientrelease rates, we first tested whether a linear or exponential modelof decay (Wieder and Lang, 1982) best fit decay of litter and eachnutrient based on R2 values for each treatment at each site. A linearmodel was superior for decomposition rates of litter and allnutrient types except K, for which an exponential model wassuperior. Decomposition rate constants (k, y�1) were calculated forthe two treatments at each site. Paired t-tests were used to deter-mine if there were differences in litter decomposition and nutrientdecay rates (k) between the two treatments, with the seven sites asreplicates. To further understand when in the decompositionprocess rates of litter decomposition and nitrogen release differedamong treatments, we used paired t-tests to compare differences indecomposition constants (k-values) between treatments for thefollowing time intervals 0e2, 2e4, 4e6, and 6e12 months with theseven sites used as replicates.
To help evaluate the magnitude of the effect of macro-inverte-brates on litter decomposition, we compared our decompositionrate estimates to those measured for various plant species acrossthe entire archipelago (Crews et al., 1995; Thompson and Vitousek,1997; Vitousek et al., 1997a; Ostertag and Hobbie, 1999; Hobbie andVitousek, 2000). To determine the influence of macro-invertebrateson nitrogen cycling, we calculated the average amount of nitrogenreleased from the litter (converting mg/100 cm2/yr to kg/ha/yr) foreach treatment type and compared these values to estimates fromthe literature of ecosystem inputs of nitrogen from rain water, drydeposition, volcanic sources, cloud water, atmospheric dust, andnitrogen fixation (Heath and Huebert, 1999; Coeppicus, 1999;
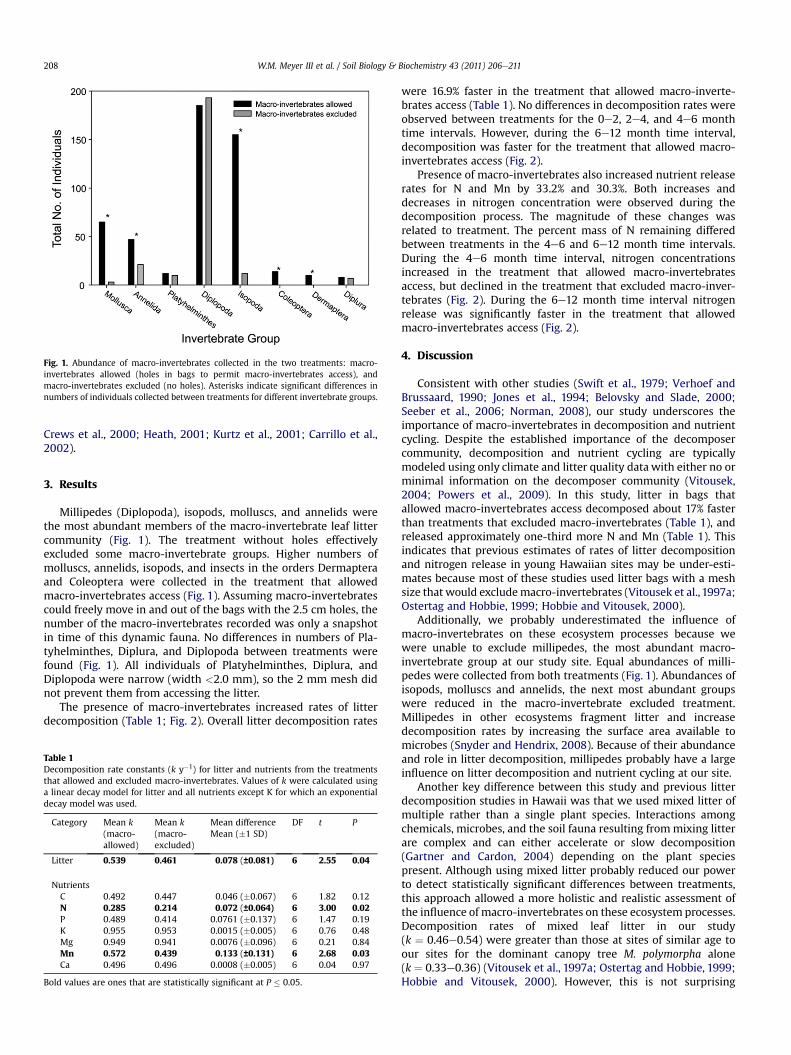
Fig. 1. Abundance of macro-invertebrates collected in the two treatments: macro-invertebrates allowed (holes in bags to permit macro-invertebrates access), andmacro-invertebrates excluded (no holes). Asterisks indicate significant differences innumbers of individuals collected between treatments for different invertebrate groups.
W.M. Meyer III et al. / Soil Biology & Biochemistry 43 (2011) 206e211208
Crews et al., 2000; Heath, 2001; Kurtz et al., 2001; Carrillo et al.,2002).
3. Results
Millipedes (Diplopoda), isopods, molluscs, and annelids werethe most abundant members of the macro-invertebrate leaf littercommunity (Fig. 1). The treatment without holes effectivelyexcluded some macro-invertebrate groups. Higher numbers ofmolluscs, annelids, isopods, and insects in the orders Dermapteraand Coleoptera were collected in the treatment that allowedmacro-invertebrates access (Fig. 1). Assuming macro-invertebratescould freely move in and out of the bags with the 2.5 cm holes, thenumber of the macro-invertebrates recorded was only a snapshotin time of this dynamic fauna. No differences in numbers of Pla-tyhelminthes, Diplura, and Diplopoda between treatments werefound (Fig. 1). All individuals of Platyhelminthes, Diplura, andDiplopoda were narrow (width <2.0 mm), so the 2 mm mesh didnot prevent them from accessing the litter.
The presence of macro-invertebrates increased rates of litterdecomposition (Table 1; Fig. 2). Overall litter decomposition rates
Table 1Decomposition rate constants (k y�1) for litter and nutrients from the treatmentsthat allowed and excluded macro-invertebrates. Values of k were calculated usinga linear decay model for litter and all nutrients except K for which an exponentialdecay model was used.
Category Mean k(macro-allowed)
Mean k(macro-excluded)
Mean differenceMean (�1 SD)
DF t P
Litter 0.539 0.461 0.078 (±0.081) 6 2.55 0.04
NutrientsC 0.492 0.447 0.046 (�0.067) 6 1.82 0.12N 0.285 0.214 0.072 (±0.064) 6 3.00 0.02P 0.489 0.414 0.0761 (�0.137) 6 1.47 0.19K 0.955 0.953 0.0015 (�0.005) 6 0.76 0.48Mg 0.949 0.941 0.0076 (�0.096) 6 0.21 0.84Mn 0.572 0.439 0.133 (±0.131) 6 2.68 0.03Ca 0.496 0.496 0.0008 (�0.005) 6 0.04 0.97
Bold values are ones that are statistically significant at P � 0.05.
were 16.9% faster in the treatment that allowed macro-inverte-brates access (Table 1). No differences in decomposition rates wereobserved between treatments for the 0e2, 2e4, and 4e6 monthtime intervals. However, during the 6e12 month time interval,decomposition was faster for the treatment that allowed macro-invertebrates access (Fig. 2).
Presence of macro-invertebrates also increased nutrient releaserates for N and Mn by 33.2% and 30.3%. Both increases anddecreases in nitrogen concentration were observed during thedecomposition process. The magnitude of these changes wasrelated to treatment. The percent mass of N remaining differedbetween treatments in the 4e6 and 6e12 month time intervals.During the 4e6 month time interval, nitrogen concentrationsincreased in the treatment that allowed macro-invertebratesaccess, but declined in the treatment that excluded macro-inver-tebrates (Fig. 2). During the 6e12 month time interval nitrogenrelease was significantly faster in the treatment that allowedmacro-invertebrates access (Fig. 2).
4. Discussion
Consistent with other studies (Swift et al., 1979; Verhoef andBrussaard, 1990; Jones et al., 1994; Belovsky and Slade, 2000;Seeber et al., 2006; Norman, 2008), our study underscores theimportance of macro-invertebrates in decomposition and nutrientcycling. Despite the established importance of the decomposercommunity, decomposition and nutrient cycling are typicallymodeled using only climate and litter quality data with either no orminimal information on the decomposer community (Vitousek,2004; Powers et al., 2009). In this study, litter in bags thatallowed macro-invertebrates access decomposed about 17% fasterthan treatments that excluded macro-invertebrates (Table 1), andreleased approximately one-third more N and Mn (Table 1). Thisindicates that previous estimates of rates of litter decompositionand nitrogen release in young Hawaiian sites may be under-esti-mates because most of these studies used litter bags with a meshsize that would excludemacro-invertebrates (Vitousek et al., 1997a;Ostertag and Hobbie, 1999; Hobbie and Vitousek, 2000).
Additionally, we probably underestimated the influence ofmacro-invertebrates on these ecosystem processes because wewere unable to exclude millipedes, the most abundant macro-invertebrate group at our study site. Equal abundances of milli-pedes were collected from both treatments (Fig. 1). Abundances ofisopods, molluscs and annelids, the next most abundant groupswere reduced in the macro-invertebrate excluded treatment.Millipedes in other ecosystems fragment litter and increasedecomposition rates by increasing the surface area available tomicrobes (Snyder and Hendrix, 2008). Because of their abundanceand role in litter decomposition, millipedes probably have a largeinfluence on litter decomposition and nutrient cycling at our site.
Another key difference between this study and previous litterdecomposition studies in Hawaii was that we used mixed litter ofmultiple rather than a single plant species. Interactions amongchemicals, microbes, and the soil fauna resulting frommixing litterare complex and can either accelerate or slow decomposition(Gartner and Cardon, 2004) depending on the plant speciespresent. Although using mixed litter probably reduced our powerto detect statistically significant differences between treatments,this approach allowed a more holistic and realistic assessment ofthe influence of macro-invertebrates on these ecosystem processes.Decomposition rates of mixed leaf litter in our study(k ¼ 0.46e0.54) were greater than those at sites of similar age toour sites for the dominant canopy tree M. polymorpha alone(k ¼ 0.33e0.36) (Vitousek et al., 1997a; Ostertag and Hobbie, 1999;Hobbie and Vitousek, 2000). However, this is not surprising
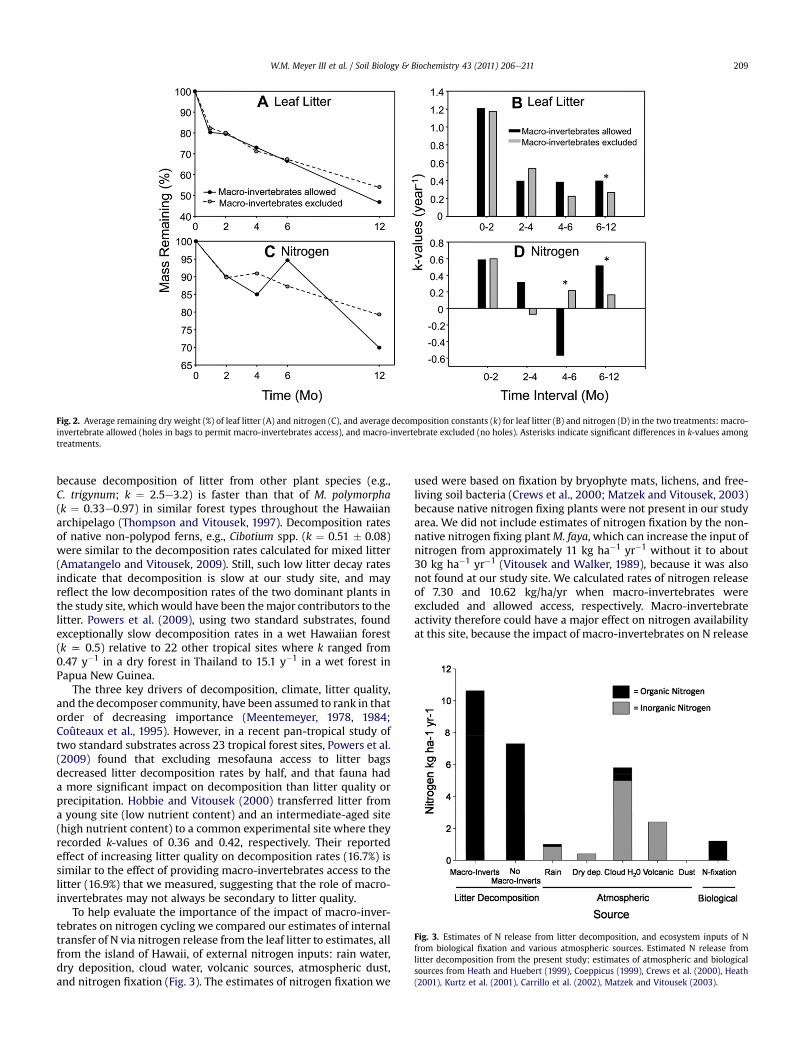
Fig. 2. Average remaining dry weight (%) of leaf litter (A) and nitrogen (C), and average decomposition constants (k) for leaf litter (B) and nitrogen (D) in the two treatments: macro-invertebrate allowed (holes in bags to permit macro-invertebrates access), and macro-invertebrate excluded (no holes). Asterisks indicate significant differences in k-values amongtreatments.
Fig. 3. Estimates of N release from litter decomposition, and ecosystem inputs of Nfrom biological fixation and various atmospheric sources. Estimated N release fromlitter decomposition from the present study; estimates of atmospheric and biologicalsources from Heath and Huebert (1999), Coeppicus (1999), Crews et al. (2000), Heath(2001), Kurtz et al. (2001), Carrillo et al. (2002), Matzek and Vitousek (2003).
W.M. Meyer III et al. / Soil Biology & Biochemistry 43 (2011) 206e211 209
because decomposition of litter from other plant species (e.g.,C. trigynum; k ¼ 2.5e3.2) is faster than that of M. polymorpha(k ¼ 0.33e0.97) in similar forest types throughout the Hawaiianarchipelago (Thompson and Vitousek, 1997). Decomposition ratesof native non-polypod ferns, e.g., Cibotium spp. (k ¼ 0.51 � 0.08)were similar to the decomposition rates calculated for mixed litter(Amatangelo and Vitousek, 2009). Still, such low litter decay ratesindicate that decomposition is slow at our study site, and mayreflect the low decomposition rates of the two dominant plants inthe study site, whichwould have been themajor contributors to thelitter. Powers et al. (2009), using two standard substrates, foundexceptionally slow decomposition rates in a wet Hawaiian forest(k x 0.5) relative to 22 other tropical sites where k ranged from0.47 y�1 in a dry forest in Thailand to 15.1 y�1 in a wet forest inPapua New Guinea.
The three key drivers of decomposition, climate, litter quality,and the decomposer community, have been assumed to rank in thatorder of decreasing importance (Meentemeyer, 1978, 1984;Coûteaux et al., 1995). However, in a recent pan-tropical study oftwo standard substrates across 23 tropical forest sites, Powers et al.(2009) found that excluding mesofauna access to litter bagsdecreased litter decomposition rates by half, and that fauna hada more significant impact on decomposition than litter quality orprecipitation. Hobbie and Vitousek (2000) transferred litter froma young site (low nutrient content) and an intermediate-aged site(high nutrient content) to a common experimental site where theyrecorded k-values of 0.36 and 0.42, respectively. Their reportedeffect of increasing litter quality on decomposition rates (16.7%) issimilar to the effect of providing macro-invertebrates access to thelitter (16.9%) that we measured, suggesting that the role of macro-invertebrates may not always be secondary to litter quality.
To help evaluate the importance of the impact of macro-inver-tebrates on nitrogen cycling we compared our estimates of internaltransfer of N via nitrogen release from the leaf litter to estimates, allfrom the island of Hawaii, of external nitrogen inputs: rain water,dry deposition, cloud water, volcanic sources, atmospheric dust,and nitrogen fixation (Fig. 3). The estimates of nitrogen fixation we
used were based on fixation by bryophyte mats, lichens, and free-living soil bacteria (Crews et al., 2000; Matzek and Vitousek, 2003)because native nitrogen fixing plants were not present in our studyarea. We did not include estimates of nitrogen fixation by the non-native nitrogen fixing plantM. faya, which can increase the input ofnitrogen from approximately 11 kg ha�1 yr�1 without it to about30 kg ha�1 yr�1 (Vitousek and Walker, 1989), because it was alsonot found at our study site. We calculated rates of nitrogen releaseof 7.30 and 10.62 kg/ha/yr when macro-invertebrates wereexcluded and allowed access, respectively. Macro-invertebrateactivity therefore could have a major effect on nitrogen availabilityat this site, because the impact of macro-invertebrates on N release

W.M. Meyer III et al. / Soil Biology & Biochemistry 43 (2011) 206e211210
(the difference between our two treatments: 3.32 kg/ha) is greaterthan estimates of nitrogen inputs from rain water, dry deposition,volcanic sources, atmospheric dust, and nitrogen fixation (Fig. 3).However, N is a mobile element and might have high potential forleaching or denitrification in tropical wet forests (Vitousek et al.,1997b; Hall and Matson, 2003). Therefore, future studies shouldexamine if the amount and timing of N released due to macro-invertebrate activity is actually influencing microbial and plant Nuptake and growth.
Understanding how and when macro-invertebrates influencelitter decomposition rates and nutrient concentrations in the leaflitter can provide insights into the mechanisms of how theseorganisms influence these ecosystem processes. Soil animals onlyaccount for w5% of soil respiration, so their major effect ondecomposition is presumed to be enhancement of microbialactivity (Wall et al., 2001). Because differences in litter decompo-sition between treatments were not observed until 6e12 monthsinto the litter decomposition process, ingestion by soil animals isprobably less important than indirect effects on microbes. Macro-invertebrates can promote microbial abundance, diversity, andactivity by the following mechanisms: 1) fracturing litter, therebyincreasing the surface area available to microbes (Swift et al., 1979;Lavelle et al., 1997); 2) increasing available nutrients through wasteproduction (Belovsky and Slade, 2000); 3) consuming microbes,which can lead to vigorous microbial growth (Seastedt andCrossley, 1980; Hedlund and Augustsson, 1995; Lavelle, 1997); 4)modifying the habitat in ways that facilitate microbial growth/activity (Jones et al., 1994; Eisenhauer et al., 2007); and 5) modi-fying the habitat in ways that increase recruitment of other smallerinvertebrates (i.e., <2 mm) that can influence litter decompositionby the four previous mechanisms. Large increases in N concentra-tions indicate N immobilization by microbes and thus increasedmicrobial abundance or activity (Chapin et al., 2002). As such, thedramatic increase in nitrogen concentrations during the 4e6month time interval in the treatment that allowed macro-inverte-brates access indicates that presence of macro-invertebrates facil-itated microbial recruitment/activity. This increase in Nconcentrationwas followed by a period (6e12month time interval)of increased N release in the macro-invertebrate allowed treatment(Fig. 2). Although we are not able to identify specifically howmacro-invertebrates promote microbes, our results demonstratethat commensalisms among macro-invertebrates and microbes areprobably driving increased rates of decomposition and nutrientcycling. The results also demonstrate that the duration of theexperiment can influence how the role of macro-invertebrates isinterpreted. For example, if the experiment had been conducted foronly 6 months we may have concluded that macro-invertebratesincrease N in the litter (Fig. 2), whereas our 12month study showedthat the presence of macro-invertebrates leads to increased Nrelease (Table 1). Also, we followed decomposition dynamics fora short period (1 year). If followed for longer, the results mightdiffer due to more recalcitrant litter, but if commensalisms amongmacro-invertebrates and microbes persisted, increased rates ofdecomposition and nutrient release would probably continue inlitter exposed to macro-invertebrates.
Our results contribute to a small but growing literature on theinfluence of macro-invertebrates on key ecosystem processes. Wedemonstrated that the macro-invertebrate community can facili-tate litter decomposition and release of N and Mn. While wefocused on the macro-invertebrates on one Hawaiian island, ourfindings may be pertinent throughout the Hawaiian Islands andelsewhere, given the wide distributions of many of the invasivemacro-invertebrates discussed here (e.g., snails/slugs: Cowie, 2001;Cowie et al., 2008), and the significance of macro-invertebrates inmany ecosystem processes (Swift et al., 1979; Verhoef and
Brussaard, 1990; Jones et al., 1994; Belovsky and Slade, 2000).However, the influence of macro-invertebrates may be different insites that are not N-limited (Vitousek, 2004). Improved knowledgeof the ecosystem effects of soil macro-invertebrates is a necessarypart of understanding how these ecosystems function.
Acknowledgements
We thank the National Science Foundation (NSF DEB0710435 toR. H. Cowie and W. M. Meyer), the Ecology, Evolution and Conser-vation Biology program of the University of Hawaii (NSFDGE0538550 to K. Y. Kaneshiro), Conchologists of America, UnitasMalacologica, the University of Hawaii Jessie Kay MemorialFellowship, and the Hawaiian Malacological Society for funding. R.Ostertag was facilitated by NSF CAREER0546868. Randi Schneiderran the elemental analyses at the UH Hilo Analytical Laboratory,which was supported by NSF EPSCoR 0237065 and 0554657 (J.Gaines PI, and D. Price and K. Kaneshiro co-PIs). Any opinions,findings, and conclusions or recommendations expressed herein arethose of the authors and do not necessarily reflect the views of theNational Science Foundation. We also thank Pat Aldrich, JeremyClaisse, Jeff Eble, Creighton Litton, Jodie Schulten, Derek Skillingsand Barbara Rowe for help in the field and lab, and Brian Bowen, TadFukami, and Andy Taylor for comments on a draft of themanuscript.The Hawaii Department of Land and Natural Resources (permitFHM08-147) allowed access to our study sites.
References
Amatangelo, K.L., Vitousek, P.M., 2009. Contrasting predictors of fern versusangiosperm decomposition in a common garden. Biotropica 41, 154e161.
Austin, A.T., Vitousek, P.M., 2000. Precipitation, decomposition and litter decom-posability of Metrosideros polymorpha in native forests on Hawai’i. Journal ofEcology 88, 129e138.
Austin, A.T., 2002. Differential effects of precipitation on production and decom-position along a rainfall gradient in Hawaii. Ecology 83, 328e338.
Belovsky, G.E., Slade, J.B., 2000. Insect herbivory accelerates nutrient cycling andincreases plant production. Proceedings of the National Academy of Sciences 97,14412e14417.
Carrillo, J.H., Galanter-Hastings, M., Sigman, D.M., Huebert, B.J., 2002. Atmosphericdeposition of inorganic and organic nitrogen and base cations in Hawaii. GlobalBiogeochemical Cycles 16 (4), 1076, doi:10.1029/2002GB001892.
Chapin III, F.S., Matson, P.A., Mooney, H.A., 2002. Principles of Terrestrial EcosystemEcology. Springer, New York.
Coeppicus, S., 1999. Atmospheric deposition of fixed nitrogen and base cations andthe volcanic source of fixed nitrogen at the Thurston lava tube, Hawai‘i. M.S.thesis, University of Hawai‘i, Manoa, Honolulu.
Coûteaux, M.M., Bottner, P., Berg, B., 1995. Litter decomposition, climate and litterquality. Trends in Ecology and Evolution 10, 63e66.
Cowie, R.H., Hayes, K.A., Tran, C.T., Meyer III, W.M., 2008. The horticultural industryas a vector of alien snails and slugs: widespread invasions in Hawaii. Interna-tional Journal of Pest Management 54, 267e276.
Cowie, R.H., 2001. Invertebrate invasions on Pacific islands and the replacement ofunique native faunas: a synthesis of the land and freshwater snails. BiologicalInvasions 3, 119e136.
Crews, T.E., Kitayama, K., Fownes, J.H., Riley, R.H., Herbert, D.A., Mueller-Dombois, D., Vitousek, P.M., 1995. Changes in soil phosphorus fractions andecosystem dynamics across a long chronosequence in Hawaii. Ecology 76,1407e1424.
Crews, T.E., Farrington, H., Vitousek, P.M., 2000. Changes in asymbiotic, heterotro-phic nitrogen fixation on leaf litter of Metrosideros polymorpha with long-termecosystem development in Hawaii. Ecosystems 3, 386e395.
Duffy, J.E., 2002. Biodiversity and ecosystem function: the consumer connection.Oikos 99, 201e219.
Eisenhauer, N., Partsch, S., Parkinson, D., Scheu, S., 2007. Invasion of a deciduousforest by earthworms: changes in soil chemistry, microflora, microarthropodsand vegetation. Soil Biology and Biochemistry 39, 1099e1110.
Gartner, T.B., Cardon, Z.G., 2004. Decomposition dynamics in mixed-species leaflitter. Oikos 104, 230e246.
Groffman, P.M., Bohlen, P.J., Fisk, M.C., Fahey, T.J., 2004. Exotic earthworm invasionand microbial biomass in temperate forest soils. Ecosystems 7, 45e54.
Gruner, D.S., 2004. Attenuation of top-down and bottom-up forces in a complexterrestrial community. Ecology 85, 3010e3022.
Hall, S.J., Matson, P.A., 2003. Nutrient status of tropical rain forests influences soil Ndynamics after N additions. Ecological Monographs 73, 107e129.

W.M. Meyer III et al. / Soil Biology & Biochemistry 43 (2011) 206e211 211
Heath, J.A., Huebert, B.J., 1999. Cloudwater deposition as a source of fixed nitrogenin a Hawaiian montane forest. Biogeochemistry 44, 119e134.
Heath, J.A., 2001. Atmospheric nutrient deposition in Hawai‘i: methods, rates andsources. PhD Dissertation, University of Hawai‘i, Manoa, Honolulu.
Hedlund, K., Augustsson, A., 1995. Effects of enchytraeid grazing on fungal growthand respiration. Soil Biology and Biochemistry 27, 905e909.
Hobbie, S.E., Vitousek, P.M., 2000. Nutrient limitation of decomposition in Hawaiianforests. Ecology 81, 1867e1877.
Hobbie, S.E., 2000. Interactions between litter lignin and soil nitrogen availabilityduring leaf litter decomposition in a Hawaiian montane rainforest. Ecosystems3, 484e494.
Holdsworth, A.R., Frelich, L.E., Reich, P.B., 2008. Litter decomposition in earthworm-invaded northern hardwood forests: role of invasion degree and litterchemistry. Écoscience 15, 536e544.
Jennings, T.J., Barkham, J.B., 1979. Litter decomposition by slugs in mixed deciduouswoodland. Holarctic Ecology 2, 21e29.
Jones, C.G., Lawton, J.H., Shachak, M., 1994. Organisms as ecosystem engineers.Oikos 69, 373e386.
Juvik, S.P., Juvik, J.O., 1998. Atlas of Hawai‘i, third ed. University of Hawai‘i Press,Honolulu.
Kurtz, A.C., Derry, L.A., Chadwick, O.A., 2001. Accretion of Asian dust to Hawaiiansoils: isotopic, elemental, and mineral mass balances. Geochimica et Cosmo-chimica Acta 65, 1971e1983.
Lavelle, P., Bignell, D., Lepage, M., 1997. Soil function in a changing world: the role ofinvertebrate engineers. European Journal of Soil Biology 33, 159e193.
Lavelle, P., 1997. Faunal activities and soil processes: adaptive strategies thatdetermine ecosystem function. Advances in Ecological Research 27, 93e132.
Mack, M.C., D’Antonio, C.M., 2003. The effects of exotic grasses on litter decom-position in a Hawaiian woodland: the importance of indirect effects. Ecosys-tems 6, 723e738.
Mascaro, J., Becklund, K.K., Hughes, R.F., Schnitzer, S.A., 2008. Limited native plantregeneration in novel, exotic-dominated forests on Hawai’i. Forest Ecology andManagement 256, 593e606.
Matzek, V.A., Vitousek, P.M., 2003. Nitrogen fixation in bryophytes, lichens, anddecaying wood along a soil age gradient in Hawaiian montane rainforests.Biotropica 35, 12e19.
Meentemeyer, V., 1978. Micro-climate and lignin control of litter decompositionrates. Ecology 59, 465e472.
Meentemeyer, V., 1984. The geography of organic decomposition rates. Annals ofthe Association of American Geographers 74, 551e560.
Newell, P.F., 1967. Mollusca. In: Burges, A., Raw, F. (Eds.), Soil Biology. AcademicPress, London, pp. 413e433.
Norman, M.N., 2008. An invasive producer (Albizia) and a predator (coqui frogs)alter decomposition processes in a Hawaiian tropical forest. M.S. thesis,University of Hawai‘i at Hilo, Hilo, HI.
Ostertag, R., Hobbie, S.E., 1999. Early stages of root and leaf decomposition inHawaiian forests: effects of nutrient availability. Oecologia 121, 564e573.
Powers, J.S., Montgomery, R.A., Adair, E.C., Brearley, F.Q., DeWalt, S.J.,Castanho, C.T., Chave, J., Deinert, E., Ganzhorn, J.U., Gilbert, M.E., González-Iturbe, J.A., Bunyavejchewin, S., Grau, H.R., Harms, K.E., Hiremath, A., Iriate-Vivar, S., Manzane, E., de Oliveira, A.A., Poorter, L., Ramanamanjato, J., Stalk, C.,Varela, A., Weiblen, G.D., Lerdau, M.T., 2009. Decomposition in tropical forests:a pan-tropical study of the effects of litter type, litter placement and
mesofaunal exclusion across a precipitation gradient. Journal of Ecology 97,801e811.
Russell, A.E., Vitousek, P.M., 1997. Decomposition and potential nitrogen fixation inDicranopteris linearis litter on Mauna Loa, Hawaii. Journal of Tropical Ecology 13,579e594.
Schaefer, M., 1991. Secondary production and decomposition. In: Rohrig, E.,Ulrich, M. (Eds.), Temperate Deciduous Forests. Ecosystems of the World, vol. 7.Elsevier, Amsterdam, pp. 175e218.
Seastedt, T.R., Crossley, D.A., 1980. Effects of microarthropods on the seasonaldynamics of nutrients in forest litter. Soil Biology and Biochemistry 12, 337e342.
Seeber, J., Scheu, S., Meyer, E., 2006. Effects of macro-decomposers on litterdecomposition and soil properties in alpine pastureland: a mesocosm experi-ment. Applied Soil Ecology 34, 168e175.
Shachak, M., Jones, C.G., Granot, Y., 1987. Herbivory in rocks and the weathering ofa desert. Science 236, 1098e1099.
Sin, H., Beard, K.H., Pitt, W.C., 2008. An invasive frog, Eleutherodactylus coqui,increases new leaf production and leaf litter decomposition rates throughnutrient cycling in Hawaii. Biological Invasions 10, 335e345.
Snyder, B.A., Hendrix, P.F., 2008. Current and potential roles of soil macro-invertebrates (earthworms, millipedes, and isopods) in ecological restoration.Restoration Ecology 16, 629e636.
Swift, M.J., Heal, O.W., Anderson, J.M., 1979. Decomposition in Terrestrial Environ-ments. Blackwell Scientific, Oxford.
Theenhaus, A., Scheu, S., 1996. The influence of slug (Arion rufus) mucus and castmaterial addition on microbial biomass, respiration, and nutrient cycling inbeech leaf litter. Biological Fertility of Soils 23, 80e85.
Thompson, M.V., Vitousek, P.M., 1997. Asymbiotic nitrogen fixation and decompo-sition during long-term soil development in a Hawaiian montane rain forest.Biotropica 29, 134e144.
Verhoef, H.A., Brussaard, L., 1990. Decomposition and nutrient mineralization innatural and agro-ecosystems: the contribution of soil animals. Biogeochemistry11, 175e211.
Vitousek, P.M., Walker, L.R., 1989. Biological invasion by Myrica faya in Hawai’i:plant demography nitrogen fixation, and ecosystem effects. Ecological Mono-graphs 59, 247e265.
Vitousek, P.M., Walker, L.R., Whiteaker, L.D., Mueller-Dombois, D., Matson, P.A., 1987.Biological invasion by Myrica faya alters ecosystem development in Hawaii.Science 238, 802e804.
Vitousek, P.M., Turner, D.R., Parton, W.J., Sanford, R.L., 1994. Litter decomposition onthe Mauna Loa environmental matrix, Hawai‘i: patterns, mechanisms, andmodels. Ecology 75, 418e429.
Vitousek, P.M., Chadwick, O.A., Crews, T., Fownes, J., Hendricks, D., Herbert, D., 1997a.Soil and ecosystem development across the Hawaiian Islands. GSA Today 7, 1e8.
Vitousek, P.M., Aber, J.D., Howarth, R.W., Likens, G.E., Matson, P.A., Schindler, D.W.,Schlesinger, W.H., Tilman, D.G., 1997b. Human alteration of the global nitrogencycle: sources and consequences. Ecological Applications 7, 737e750.
Vitousek, P.M., 2004. Nutrient Cycling and Limitation: Hawaii as a Model Ecosystem.Princeton University Press, Princeton.
Wall, D.H., Adams, G., Parsons, A.N., 2001. Soil biodiversity. In: Chapin III, F.S.,Sala, O.E., Huber-Sannwald, E. (Eds.), Global Biodiversity in a Changing Envi-ronment: Scenarios for the 21st Century. Springer-Verlag, New York, pp. 47e82.
Wieder, R.K., Lang, G.E., 1982. A critique of the analytical methods used in exam-ining decomposition data obtained from litter bags. Ecology 63, 1636e1642.