Embed Size (px)
Citation preview

Immunobiology 209 (2004) 31–38
ARTICLE IN PRESS
Abbreviations: C
lysoPCacyltransf
acid; PC, phosp
tumour necrosis
*Correspondin
2161.
E-mail addres
0171-2985/$ - see
doi:10.1016/j.imb
www.elsevier.de/imbio
REVIEW
Lysophospholipid acyltransferases in monocyte inflammatory responses
and sepsis
Simon K. Jackson*, Joan Parton
Department of Medical Microbiology, University of Wales College of Medicine, Heath Park, Cardiff CF14 4XN, Wales, UK
Received 1 April 2004; accepted 8 April 2004
Abstract
Acyltransferases are important in the regulation of membrane phospholipid fatty acyl composition and togetherwith phospholipase A2 enzymes control arachidonic acid incorporation and remodelling within phospholipids. Inaddition, monocyte and macrophage acyltransferase activity has been shown to respond to various inflammatorycytokines under conditions that can induce enhanced cellular responses. Work in our laboratory indicates that theenzyme lysophosphatidylcholine acyltransferase may mediate the priming reactions of monocytes to the cytokineinterferon-g: Our recent studies suggest that this enzyme might also affect the responses of monocytes to the bacterialagent lipopolysaccharide that may be important in the development of sepsis. This article summarises the relationshipbetween monocyte lysophosphatidylcholine acyltransferase, lipopolysaccharide and sepsis.r 2004 Elsevier GmbH. All rights reserved.
Keywords: Sepsis, acyltransferase, monocytes
Lipopolysaccharide and sepsis
Sepsis is a consequence of an overwhelming inflam-matory response to infection which has proved excep-tionally difficult to treat despite advances in antibiotictherapy and intensive care. Sepsis occurs in at least oneand a half million people throughout the world eachyear. In the United States alone, sepsis develops in morethan 500,000 patients each year, with a 30–70%mortality rate (Angus et al., 2001). Sepsis develops fromthe systemic inflammatory response to pathogens in theblood. Bacterial pathogens carry surface molecules
oA, coenzyme A; IFN-g, interferon-gamma; LPCAT,
erase; LPS, lipopolysaccharide; PA, phosphatidic
hatidylcholine; PE, phosphatidylethanolamine; TNF,
factor; TLR, toll like receptor
g author. Tel.: +44-29-2074-4725; fax: +44-29-2074-
s: [email protected] (S.K. Jackson).
front matter r 2004 Elsevier GmbH. All rights reserved.
io.2004.04.006
termed pathogen associated molecular patterns (PAMP)which can induce a variety of inflammatory mediators.The resulting inflammatory response can producecardiovascular derangements, hypotension, multipleorgan failure and death (Cohen, 2002). The bestcharacterised of these microbial PAMP is the bacterialmolecule lipopolysaccharide (LPS), the endotoxin pre-sent in the outer membrane of Gram-negative bacteria(Rietschel et al., 1994; Van Amersfoot et al., 2003). Keytarget cells in the pathogenesis of LPS-induced sepsis arethe monocytes and macrophages. These cells canrespond to LPS in the circulation by the production ofinflammatory mediators including cytokines and bioac-tive lipids, and the expression of cell-surface receptorsand adhesion molecules (Bhatia and Moochhala, 2004;Glauser et al., 1991). It is the massive release of theinflammatory mediators that is a primary mechanismfor the initiation of severe sepsis (Hesse et al., 1988;Tracey and Lowry, 1990). Two inflammatory cytokines,

ARTICLE IN PRESSS.K. Jackson, J. Parton / Immunobiology 209 (2004) 31–3832
Tumour necrosis factor (TNF) and interferon-g (IFN-gÞ;have been shown to be particularly important in thedevelopment of septic shock. TNF is central to thepathogenesis as indicated by the relative resistance toLPS-induced toxicity in mice lacking the p55 TNFR,TNF protein or producing high levels of TNFR1 fusionprotein (Pasparakis et al., 1996; Rothe et al., 1993). Thecontrol of TNF production has been a focus for thedevelopment of sepsis therapies. However, the results ofclinical trials of specific anti-TNF therapies for sepsishave been disappointing (Abraham, 1999), reflecting thecomplexity of the syndrome and the involvement ofseveral cytokines with overlapping functions.
IFN-g is another important regulator of LPS-inducedpathology (Doherty et al., 1992; Silva and Cohen, 1992).Administration of IFN-g or neutralizing antibody toIFN-g has been shown to modify the lethal outcomes ofseveral forms of endotoxic shock and Gram-negativebacterial infections and IFN-gR-deficient mice arerelatively resistant to LPS-induced shock (Car et al.,1994; Heinzel, 1990).
The molecular events leading to inflammatory med-iator production by cells in response to LPS arebecoming well understood. LPS, liberated from invad-ing bacteria either spontaneously during growth or as aconsequence of immune-mediated lysis, exists in thecirculation bound to numerous lipid binding proteinsincluding albumin, transferrin, high-density lipoproteinsand LPS-binding protein (LBP), an acute phase proteinsynthesised by the liver in response to infection. LBPcomplexes LPS and acts as a lipid transfer proteinshuttling LPS to the surface of monocytes and macro-phages (Cohen, 2002; Schumann et al., 1990). Thesecells express a receptor for LPS, CD14, a 55 kDaglycosylphosphoinositol (GPI)-anchored membraneprotein, which has been shown to be a receptor forLPS (Pugin et al., 1994; Wright et al., 1990). CD14associates with the signalling receptor TLR4, which inturn initiates the signal cascade resulting in NF-kBtranscription factor activation and inflammatory genetranscription in response to LPS (Hoshino et al., 1999;Van Amersfoot et al., 2003). Regulation of thissignalling pathway for LPS-mediated responses is afocus of research aimed at developing new therapies forsepsis and related inflammatory disease.
Sensitisation to LPS and priming
It is well known from experimental models ofendotoxin-induced sepsis that infectious agents can up-regulate or sensitise responses to subsequent LPSchallenge. Such sensitizing agents include bacterialinfection with P. acnes (Berendt et al., 1980) and theBCG strain of mycobacteria (Peavy et al., 1979) which
can increase the responsiveness of leukocytes to LPSseveral thousand fold. Priming for enhanced responsesto LPS is of considerable interest clinically because it isthought that underlying or sub-clinical infections mayprime patients for exaggerated inflammatory responsesto low concentrations of LPS. However, the molecularmechanisms of this priming are not well understood.
IFN-g has been shown to be an important mediatorof the sensitising actions of infectious agents, such asP. acnes, on macrophages for LPS (Billiau et al., 1987;Katschinski et al., 1992). Indeed, a major contributionof IFN-g to LPS-induced shock may be priming anenhanced activation state in monocytes/macrophages(Adams and Hamilton, 1984; Silva and Cohen, 1992).Upon subsequent exposure to LPS, the primed macro-phages become hyper-activated and produce largeamounts of TNF and IL-1 (Doherty et al., 1992;Heinzel, 1990; Heremans et al., 1990).
Mechanisms of INF-c mediated priming of
macrophages
The precise mechanisms underlying the priming ofmacrophages by INF-g for enhanced responses to LPShave remained elusive. Recent work has suggested thatIFN-g might increase responsiveness to LPS by aug-menting the signal transduction pathway including up-regulating TLR4 expression (Bosisio et al., 2002) orpromoting IL-1 receptor associated kinase expressionand its association to MyD88 (Adib-Conquy andCavaillon, 2002). Our laboratory has been concernedwith elucidating the mechanisms of the primingresponses of monocytes and macrophages to LPS. Wehave previously established that infections, such asBCG, which increase sensitivity to LPS in experimentalmodels of sepsis, alter the membrane phospholipidprofiles of macrophages and monocytes (Stark et al.,1990). Furthermore, these ‘priming’ infections wereshown to induce the production of IFN-g whichmediated the macrophage/monocyte responsiveness.We subsequently showed that IFN-g could producesimilar alterations in macrophage phospholipid compo-sitions that accompany the priming of these cells bothin vivo and in vitro (Jackson et al., 1989, 1993). Inparticular, IFN-g stimulated the increased incorporationof unsaturated fatty acids into phosphatidylcholine (PC)which were then turned over into phosphatidylethano-lamine (PE) (Darmani et al., 1993). The incorporationof unsaturated fatty acids into phospholipids is accom-plished by the deacylation and reacylation of thephopsholipids (the Lands Cycle) mediated by theactivity of the lipid modifying enzymes phospholipasesand acyltransferases (Balsinde, 2002). Thus, we sug-gested that IFN-g might increase responsiveness to LPS

ARTICLE IN PRESSS.K. Jackson, J. Parton / Immunobiology 209 (2004) 31–38 33
in macrophages by up-regulating the activity of theseenzymes. While the activity of phospholipases was notinfluenced by IFN-g; the activity of certain acyltrans-ferases was significantly increased by this priming agent(Schmid et al., 2003).
Acyltransferases
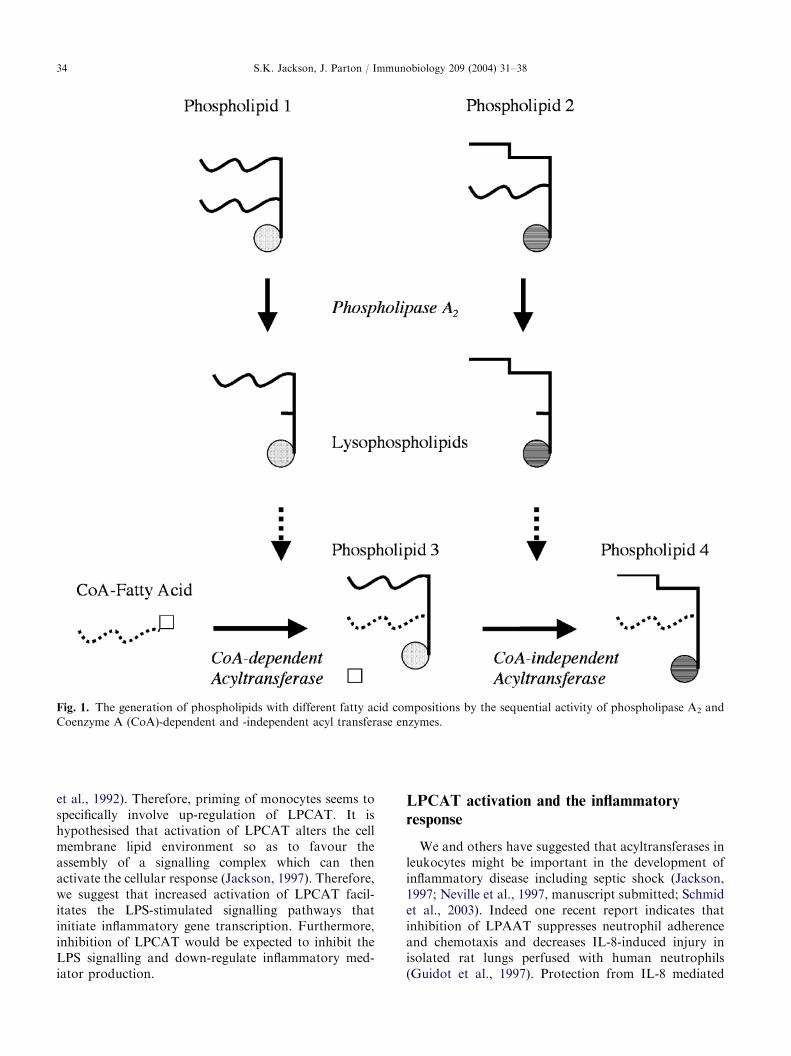
The acylation–deacylation of membrane phospho-lipids (the Lands Cycle) is a mechanism for incorporat-ing unsaturated fatty acids (mainly arachidonic acid)into different phospholipids to provide a range of lipidmediators (Yamashita et al., 1997). The fatty acidsare cleaved from phospholipids by the action ofphospholipase A2 and re-incorporated by acyltrans-ferases. Studies have revealed that arachidonic-acid isincorporated first into phospholipids containing a 1-acyllinkage by Coenzyme A (CoA)-dependent enzymes.The arachidonic acid is then transferred by CoA-independent transacylases from 1-acyl linked phospho-lipids to 1-alkyl and 1-alk-1-enyl lyso phospholipidsto form 1-alkyl and 1-alk-1-enyl-2-arachidonyl phos-pholipids (Chilton et al., 1996), which are importantin the synthesis of platelet activating factor (PAF)(Fig. 1).
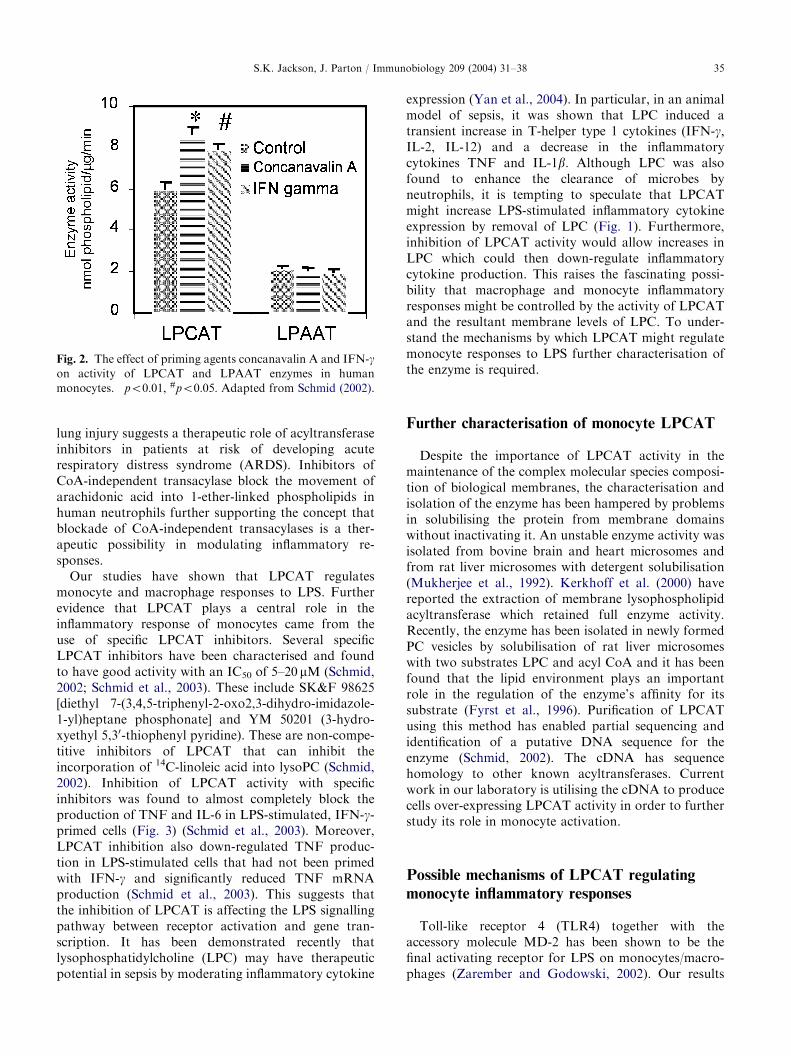
Both CoA-dependent acyltransferases and CoA-in-dependent transacylases (CoAIT) in addition to theirimportant role in providing substrates for lipid media-tors of inflammation, have been found to be involved inlipid signalling pathways (Jackson, 1997; Prokazovaet al., 1998) and leukocyte activation. They may alsomodulate the activities of other membrane-localisedenzymes (Prokazova et al., 1998). However, little isknown about the regulation of these enzymes duringinflammation, although a study using human neutro-phils described a PAF-induced increase in the arachi-donoyl-CoA-specific lysophospholipid acyltransferaseactivity (Tou, 1987). Similarly, TNF was found toincrease the CoAIT activity in human neutrophils(Winkler et al., 1994) and IL-1a was shown to increasethe incorporation of arachidonate into phosphatidyli-nositol and phosphatidylserine in rat mesangial cells,implying an enhanced rate of arachidonate-selectivelysophospholipid acyltransferase(s) (Winkler et al.,1995). Studies from our laboratory recently demon-strated that TNF can modify phospholipid composi-tions in monocytes via activation of CoA-independenttransacylases (Neville et al., manuscript submission).Furthermore, we showed that IFN-g and concanavalinA, another priming agent, could selectively activatelysophosphatidyl choline acyltransferase (LPCAT) butnot the lysophosphatidic acid acyltransferase (LPAAT)(Schmid et al., 2003) (Fig. 2). Thus, LPCAT activity isup-regulated under conditions of priming monocytes/macrophages for increased responses to LPS.
LPCAT and priming in macrophages and
monocytes
Cytokines are typical physiological priming agentsand priming by several cytokines has been shown to up-regulate PAF and eicosanoid generation in different celltypes (Glaser et al., 1990). CoA-dependent acyltrans-ferases and CoA-independent transacylases would alsobe expected to be key enzymes involved in the primingprocess for increased lipid mediator production. Inter-leukin-1a can increase the activity of arachidonic acid-lysophospholipid acyltransferase and stimulates arachi-donic acid incorporation into phospholipids in rat(Nakazato and Sedor, 1992) and stimulates lysoPAacyltransferase in human mesangial cells (Bursten et al.,1991). Increased acyltransferase activity would protectthe mesangial cell membranes from the potentiallydamaging effects of PLA2-generated lysophospholipids.A CoA-independent transacylase was up-regulated inhuman neutrophils after treatment with TNF which wasshown to prime these cells for enhanced arachidonicacid metabolism following activation with the agonistformyl peptide, f-MetLeuPhe (fMLP) (Gegner et al.,1995). Acyltransferases have been characterised inmacrophages and evidence from our work suggests thatpriming cytokines such as IFN-g can increase theactivity of both CoA-dependent and CoA-independenttransacylases in a human monocyte cell line (Nevilleet al., 1997).
We demonstrated that IFN-g could induce the alteredphospholipid profiles seen in monocytes and macro-phages both in vivo and in vitro which accompaniedtheir increased inflammatory responses to LPS (Darma-ni et al., 1993; Jackson et al., 1993). In particular, wefound that IFN-g could directly up-regulate the activityof lysophosphatidylcholine acyltransferase (LPCAT)which reacylates lysoPC with unsaturated fatty acids(Schmid et al., 2003). Such studies suggest that thatLPCAT may regulate the priming of monocytes by IFN-g and stimulate increased inflammatory cytokine pro-duction in response to LPS. In studies of T-lymphocyteactivation, Szamel et al. (1993, 1998) have demonstratedthat activation of the T-cell antigen receptor/CD3complex leads to increased incorporation of polyunsa-turated fatty acids into phosphatidylcholine, alsomediated by an LPCAT. Thus, LPCAT may play acrucial role in the early phase of T-cell activation byelevated incorporation of polyunsaturated fatty acidsinto plasma membranes phospholipids.
Results from our work (Schmid et al., 2003) suggestthat LPCAT plays a similar role in the activation ofmonocytes by priming agents such as IFN-g: Interest-ingly, our results showed that neither of the primingagents IFN-g or concanavalin A, affected the activity ofLPAAT, which has been reported to be up-regulated bythe lipid A portion of LPS in mesangial cells (Bursten
ARTICLE IN PRESS
Fig. 1. The generation of phospholipids with different fatty acid compositions by the sequential activity of phospholipase A2 and
Coenzyme A (CoA)-dependent and -independent acyl transferase enzymes.
S.K. Jackson, J. Parton / Immunobiology 209 (2004) 31–3834
et al., 1992). Therefore, priming of monocytes seems tospecifically involve up-regulation of LPCAT. It ishypothesised that activation of LPCAT alters the cellmembrane lipid environment so as to favour theassembly of a signalling complex which can thenactivate the cellular response (Jackson, 1997). Therefore,we suggest that increased activation of LPCAT facil-itates the LPS-stimulated signalling pathways thatinitiate inflammatory gene transcription. Furthermore,inhibition of LPCAT would be expected to inhibit theLPS signalling and down-regulate inflammatory med-iator production.
LPCAT activation and the inflammatory
response
We and others have suggested that acyltransferases inleukocytes might be important in the development ofinflammatory disease including septic shock (Jackson,1997; Neville et al., 1997, manuscript submitted; Schmidet al., 2003). Indeed one recent report indicates thatinhibition of LPAAT suppresses neutrophil adherenceand chemotaxis and decreases IL-8-induced injury inisolated rat lungs perfused with human neutrophils(Guidot et al., 1997). Protection from IL-8 mediated
ARTICLE IN PRESS
Fig. 2. The effect of priming agents concanavalin A and IFN-gon activity of LPCAT and LPAAT enzymes in human
monocytes. �po0:01; #po0:05: Adapted from Schmid (2002).
S.K. Jackson, J. Parton / Immunobiology 209 (2004) 31–38 35
lung injury suggests a therapeutic role of acyltransferaseinhibitors in patients at risk of developing acuterespiratory distress syndrome (ARDS). Inhibitors ofCoA-independent transacylase block the movement ofarachidonic acid into 1-ether-linked phospholipids inhuman neutrophils further supporting the concept thatblockade of CoA-independent transacylases is a ther-apeutic possibility in modulating inflammatory re-sponses.
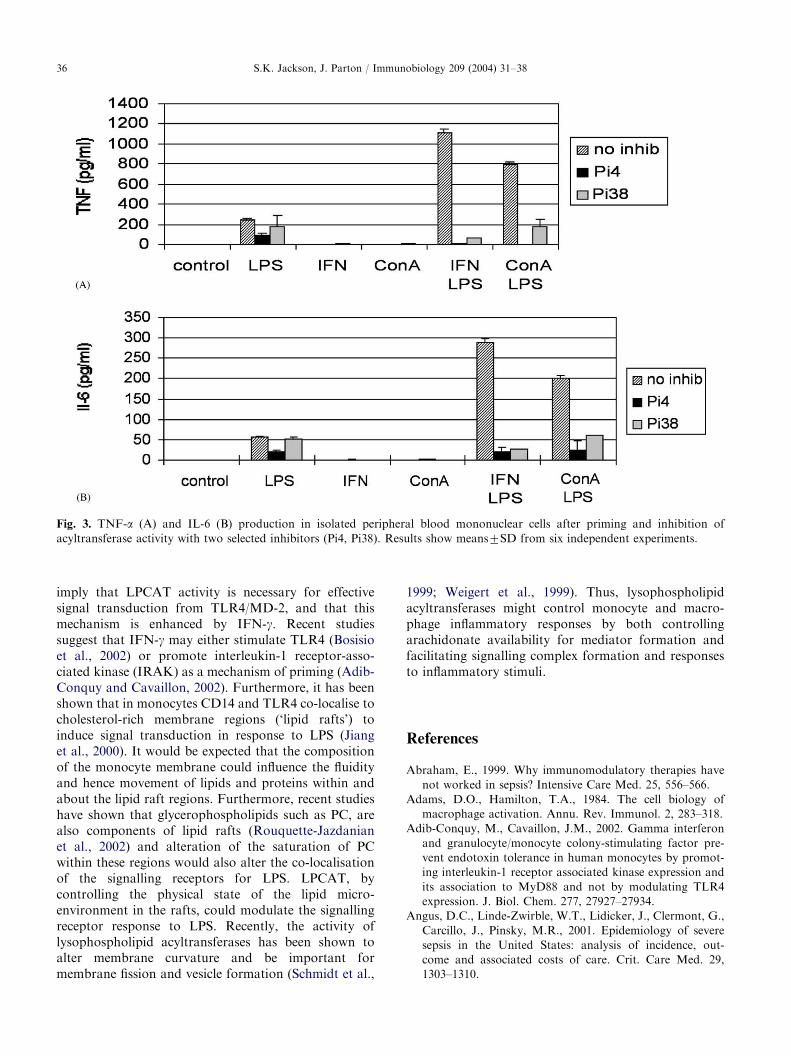
Our studies have shown that LPCAT regulatesmonocyte and macrophage responses to LPS. Furtherevidence that LPCAT plays a central role in theinflammatory response of monocytes came from theuse of specific LPCAT inhibitors. Several specificLPCAT inhibitors have been characterised and foundto have good activity with an IC50 of 5–20 mM (Schmid,2002; Schmid et al., 2003). These include SK&F 98625[diethyl 7-(3,4,5-triphenyl-2-oxo2,3-dihydro-imidazole-1-yl)heptane phosphonate] and YM 50201 (3-hydro-xyethyl 5,30-thiophenyl pyridine). These are non-compe-titive inhibitors of LPCAT that can inhibit theincorporation of 14C-linoleic acid into lysoPC (Schmid,2002). Inhibition of LPCAT activity with specificinhibitors was found to almost completely block theproduction of TNF and IL-6 in LPS-stimulated, IFN-g-primed cells (Fig. 3) (Schmid et al., 2003). Moreover,LPCAT inhibition also down-regulated TNF produc-tion in LPS-stimulated cells that had not been primedwith IFN-g and significantly reduced TNF mRNAproduction (Schmid et al., 2003). This suggests thatthe inhibition of LPCAT is affecting the LPS signallingpathway between receptor activation and gene tran-scription. It has been demonstrated recently thatlysophosphatidylcholine (LPC) may have therapeuticpotential in sepsis by moderating inflammatory cytokine
expression (Yan et al., 2004). In particular, in an animalmodel of sepsis, it was shown that LPC induced atransient increase in T-helper type 1 cytokines (IFN-g;IL-2, IL-12) and a decrease in the inflammatorycytokines TNF and IL-1b. Although LPC was alsofound to enhance the clearance of microbes byneutrophils, it is tempting to speculate that LPCATmight increase LPS-stimulated inflammatory cytokineexpression by removal of LPC (Fig. 1). Furthermore,inhibition of LPCAT activity would allow increases inLPC which could then down-regulate inflammatorycytokine production. This raises the fascinating possi-bility that macrophage and monocyte inflammatoryresponses might be controlled by the activity of LPCATand the resultant membrane levels of LPC. To under-stand the mechanisms by which LPCAT might regulatemonocyte responses to LPS further characterisation ofthe enzyme is required.
Further characterisation of monocyte LPCAT
Despite the importance of LPCAT activity in themaintenance of the complex molecular species composi-tion of biological membranes, the characterisation andisolation of the enzyme has been hampered by problemsin solubilising the protein from membrane domainswithout inactivating it. An unstable enzyme activity wasisolated from bovine brain and heart microsomes andfrom rat liver microsomes with detergent solubilisation(Mukherjee et al., 1992). Kerkhoff et al. (2000) havereported the extraction of membrane lysophospholipidacyltransferase which retained full enzyme activity.Recently, the enzyme has been isolated in newly formedPC vesicles by solubilisation of rat liver microsomeswith two substrates LPC and acyl CoA and it has beenfound that the lipid environment plays an importantrole in the regulation of the enzyme’s affinity for itssubstrate (Fyrst et al., 1996). Purification of LPCATusing this method has enabled partial sequencing andidentification of a putative DNA sequence for theenzyme (Schmid, 2002). The cDNA has sequencehomology to other known acyltransferases. Currentwork in our laboratory is utilising the cDNA to producecells over-expressing LPCAT activity in order to furtherstudy its role in monocyte activation.
Possible mechanisms of LPCAT regulating
monocyte inflammatory responses
Toll-like receptor 4 (TLR4) together with theaccessory molecule MD-2 has been shown to be thefinal activating receptor for LPS on monocytes/macro-phages (Zarember and Godowski, 2002). Our results
ARTICLE IN PRESS
Fig. 3. TNF-a (A) and IL-6 (B) production in isolated peripheral blood mononuclear cells after priming and inhibition of
acyltransferase activity with two selected inhibitors (Pi4, Pi38). Results show means7SD from six independent experiments.
S.K. Jackson, J. Parton / Immunobiology 209 (2004) 31–3836
imply that LPCAT activity is necessary for effectivesignal transduction from TLR4/MD-2, and that thismechanism is enhanced by IFN-g: Recent studiessuggest that IFN-g may either stimulate TLR4 (Bosisioet al., 2002) or promote interleukin-1 receptor-asso-ciated kinase (IRAK) as a mechanism of priming (Adib-Conquy and Cavaillon, 2002). Furthermore, it has beenshown that in monocytes CD14 and TLR4 co-localise tocholesterol-rich membrane regions (‘lipid rafts’) toinduce signal transduction in response to LPS (Jianget al., 2000). It would be expected that the compositionof the monocyte membrane could influence the fluidityand hence movement of lipids and proteins within andabout the lipid raft regions. Furthermore, recent studieshave shown that glycerophospholipids such as PC, arealso components of lipid rafts (Rouquette-Jazdanianet al., 2002) and alteration of the saturation of PCwithin these regions would also alter the co-localisationof the signalling receptors for LPS. LPCAT, bycontrolling the physical state of the lipid micro-environment in the rafts, could modulate the signallingreceptor response to LPS. Recently, the activity oflysophospholipid acyltransferases has been shown toalter membrane curvature and be important formembrane fission and vesicle formation (Schmidt et al.,
1999; Weigert et al., 1999). Thus, lysophospholipidacyltransferases might control monocyte and macro-phage inflammatory responses by both controllingarachidonate availability for mediator formation andfacilitating signalling complex formation and responsesto inflammatory stimuli.
References
Abraham, E., 1999. Why immunomodulatory therapies have
not worked in sepsis? Intensive Care Med. 25, 556–566.
Adams, D.O., Hamilton, T.A., 1984. The cell biology of
macrophage activation. Annu. Rev. Immunol. 2, 283–318.
Adib-Conquy, M., Cavaillon, J.M., 2002. Gamma interferon
and granulocyte/monocyte colony-stimulating factor pre-
vent endotoxin tolerance in human monocytes by promot-
ing interleukin-1 receptor associated kinase expression and
its association to MyD88 and not by modulating TLR4
expression. J. Biol. Chem. 277, 27927–27934.
Angus, D.C., Linde-Zwirble, W.T., Lidicker, J., Clermont, G.,
Carcillo, J., Pinsky, M.R., 2001. Epidemiology of severe
sepsis in the United States: analysis of incidence, out-
come and associated costs of care. Crit. Care Med. 29,
1303–1310.

ARTICLE IN PRESSS.K. Jackson, J. Parton / Immunobiology 209 (2004) 31–38 37
Balsinde, J., 2002. Roles of various phospholipases A2 in
providing lysophospholipid acceptors for fatty acid phos-
pholipids incorporation and remodelling. Biochem. J. 364,
695–702.
Berendt, M.J., Newborg, M.F., North, R.J., 1980. Increased
toxicity of endotoxin for tumor-bearing mice and mice
responding to bacterial pathogens: macrophage activation
as a common denominator. Infect. Immun. 28, 645–647.
Bhatia, M., Moochhala, S., 2004. Role of inflammatory
mediators in the pathophysiology of acute respiratory
distress syndrome. J. Pathol. 202, 145–156.
Billiau, A., Heremans, H., Vandekerckhove, F., Dillen, C.,
1987. Anti-interferon-g antibody protects mice against the
generalised Shwartzman reaction. Eur. J. Immunol. 17,
1851–1854.
Bosisio, D., Polentarutti, N., Sironi, M., Bernasconi, S.,
Miyake, K., Webb, G.R., Marin, M.U., Mantovani, A.,
Muzio, M., 2002. Stimulation of toll-like receptor 4
expression in human mononuclear phagocytes by interfer-
on-g: a molecular basis for priming and synergism with
bacterial lipopolysaccharide. Blood 99, 3427–3431.
Bursten, S.L., Harris, W.E., Bomsztyk, K., Lovett, D.,
1991. Interleukin-1 rapidly stimulates lysophosphatidate
acyltransferase and phosphatidate phosphohydrolase
activities in human mesangial cells. J. Biol. Chem. 266,
20732–20743.
Bursten, S.L., Harris, W.E., Resch, K., Lovett, D., 1992. Lipid
A activation of glomerular mesangial cells: mimicry of the
bioactive lipid, phosphatidic acid. Am. J. Physiol. 262,
C328–C338.
Car, B.D., Eng, V.M., Schnyder, B., Ozmen, L., Huang, S.,
Gallay, P., Heumann, D., Auget, M., Ryffel, B., 1994. IFN-
g receptor deficient mice are resistant to endotoxic shock.
J. Exp. Med. 179, 1437–1444.
Chilton, F.H., Fontech, A.N., Surette, M.E., Triggiani, M.,
Winkler, J.D., 1996. Control of arachidonate levels within
inflammatory cells. Biochim. Biophys. Acta 1299, 1–15.
Cohen, J., 2002. The immunopathogenesis of sepsis. Nature
420, 885–891.
Darmani, H., Harwood, J., Jackson, S.K., 1993. Interferon-gstimulated uptake and turnover of linoleate and arachido-
nate in macrophages: a possible pathway for hypersensi-
tivity to endotoxin. Cell. Immunol. 152, 59–71.
Doherty, G.M., Lange, J.R., Langstein, H.N., Alexander,
H.R., Buresch, C.M., Norton, J.A., 1992. Evidence for
IFN-g as a mediator of lethality of endotoxin and TNF-a.J. Immunol. 149, 1666–1670.
Fyrst, H., Pham, D.V., Lubin, B.H., Kuypers, F.A., 1996.
Formation of vesicles by the action of acyl-CoA:1-
acyllysophosphatidylcholine acyltransferase from rat liver
microsomes: optimal solubilization conditions and analysis
of lipid composition and enzyme activity. Biochemistry 35,
2644–2650.
Gegner, J.A., Ulevitch, R.J., Tobias, P.S., 1995. Lipopolysac-
charide (LPS) signal transduction and clearance. Dual roles
for LPS binding protein and membrane CD14. J. Biol.
Chem. 270, 5320–5325.
Glaser, K.B., Asmis, R., Dennis, E.A., 1990. Bacterial
lipopolysaccharide priming of P388D1 macrophage-like
cells for enhanced arachidonic acid metabolism. Platelet-
activating factor receptor activation and regulation of
phospholipase A2. J. Biol. Chem. 265, 8658–8664.
Glauser, M.P., Zanetti, G., Baumgartner, J.D., Cohen, J.,
1991. Septic shock: pathogenesis. Lancet 338, 732–736.
Guidot, D.M., Bursten, S.L., Rice, G.C., Chaney, R.B.,
Singer, J.W., Repine, A.J., Hybertson, B.M., Repine, J.E.,
1997. Modulating phosphatidic acid metabolism de-
creases oxidative injury in rat lungs. Am. J. Physiol. 273,
L957–966.
Heinzel, F.P., 1990. The role of IFN-g in the pathology of
experimental endotoxemia. J. Immunol. 145, 2920–2925.
Heremans, H., Van Damme, J., Dillen, C., Dijkmans, R.,
Billiau, A., 1990. Interferon gamma, a mediator of lethal
LPS-induced Shwartzman-like reactions in mice. J. Exp.
Med. 171, 1853–1869.
Hesse, D.G., Tracey, K.J., Fong, Y., Manogue, K.R.,
Palladino Jr., M.A., Cerami, A., Shires, G.T., Lowry,
S.F., 1988. Cytokine appearance in human endotoxemia
and primate bacteremia. Surg. Gynecol. Obstet. 166,
147–153.
Hoshino, K., Takeuchi, O., Kawai, T., Sanjo, H., Ogawa, T.,
Takeda, Y., Takeda, K., Akira, S., 1999. Cutting edge: toll-
like receptor 4 (TLR4)-deficient mice are hyporesponsive to
LPS: evidence for TLR4 as the Lps gene product.
J. Immunol. 162, 3749–3752.
Jackson, S.K., 1997. The role of lipid metabolites in the
signalling and activation of macrophages by lipopolysac-
charide. Prog. Lipid Res. 36, 227–244.
Jackson, S.K., Stark, J.M., Taylor, S., Harwood, J.L., 1989.
Changes in phospholipid fatty acid composition and
triacylglycerol content in mouse tissues after infection with
bacille Calmette-Guerin. Br. J. Exp. Path. 70, 435–441.
Jackson, S.K., Darmani, H., Harwood, J.L., 1993. Interferon-gincreases membrane fluidity and stimulates the uptake of
linoleic acid in murine macrophages. J. Interferon Res. 13,
427–431.
Jiang, Q., Akashi, S., Miyake, K., Petty, H.R., 2000.
Lipopolysaccharide induces physical proximity between
CD14 and toll-like receptor 4 (TLR-4) prior to nuclear
translocation of NF-kappaB. J. Immunol. 165, 3541–3544.
Katschinski, T., Galanos, C., Coumbos, A., Freudenberg,
M.A., 1992. Gamma interferon mediates P. acnes –induced
hypersensitivity to LPS in mice. Infect. Immun. 60,
1994–2001.
Kerkhoff, C., Trumbach, B., Gehring, L., Habben, K.,
Schmitz, G., Kaever, V., 2000. Solubilization, partial
purification and photolabelling of the integral membrane
protein lysophospholipid: acyl-coa acyltransferase (LAT).
Eur. J. Biochem. 267, 6339–6345.
Mukherjee, J.J., Tardi, P.G., Choy, P.C., 1992. Solubilization
and modulation of acyl-CoA: 1-acyl-glycerophosphocho-
line acyltransferase activity in rat liver microsomes.
Biochim. Biophys. Acta 1123, 27–32.
Nakazato, Y., Sedor, J.R., 1992. IL-1 alpha increases
arachidonyl-CoA: lysophospholipid acyltransferase activity
and stimulates [3H]arachidonate incorporation into phos-
pholipids in rat mesangial cells. Life Sci. 50, 2075–2082.
Neville, N.T., Jackson, S.K., Harwood, J.L., 1997. The effects
of inflammatory cytokines on acyl-CoA-dependent acyl-
transferase. Biochem. Soc. Trans. 25, 496S.

ARTICLE IN PRESSS.K. Jackson, J. Parton / Immunobiology 209 (2004) 31–3838
Neville, N.T., Parton, J., Harwood, J.L., Jackson, S.K.,
submitted. The activity of monocyte lysophosphatidylcho-
line acyltransferase and coenzyme A-independent transa-
cylase is changed by the inflammatory cytokines tumor
necrosis factor alpha and interferon gamma.
Pasparakis, M., Alexopoulou, L., Episkopou, V., Kollias, G.,
1996. Immune and inflammatory responses in TNFadeficient mice: a critical requirement for TNFa in the
formation of primary b cell follicles, follicular dendritic cell
networks and germinal centres, and in the maturation
of the humoral immune response. J. Exp. Med. 184,
1397–1411.
Peavy, D.L., Baughn, R.E., Musher, D.M., 1979. Effects of
BCG infection on the susceptibility of mouse macrophages
to endotoxin. Infect. Immun. 24, 59–64.
Prokazova, N.V., Zvezdina, N.D., Korotaeva, A.A., 1998.
Effect of lysophosphatidylcholine on transmembrane signal
transduction. Biochemistry 63, 31–37.
Pugin, J., Heumann, I.D., Tomasz, A., Kravchenko, V.V.,
Akamatsu, Y., Nishijima, M., Glauser, M.P., Tobias, P.S.,
Ulevitch, R.J., 1994. CD14 is a pattern recognition
receptor. Immunity 1, 509–516.
Rietschel, E.T., Kirikae, T., Schade, F.U., Mamat, U.,
Schmidt, G., Loppnow, H., Ulmer, A.J., Zahringer, U.,
Seydel, U., Di Padova, F., et al., 1994. Bacterial endotoxin:
molecular relationships of structure to activity and func-
tion. FASEB J. 8, 217–225.
Rothe, J., Lesslauer, W., Lotscher, H., Lang, Y., Koebel, P.,
Kontgen, F., Althage, A., Zinkernagel, R., Steinmetz, M.,
Bluethmann, H., 1993. Mice lacking the TNF receptor 1 are
resistant to TNF-mediated toxicity but highly susceptible to
infection by Listeria monocytogenes. Nature 364, 798–802.
Rouquette-Jazdanian, A.K., Pelassy, C., Breittmayer, J-P.,
Cousin, J-L., Aussel, C., 2002. Metabolic labeling of
membrane microdomains/rafts in Jurkat cells indicates the
presence of glycerophospholipids implicated in signal
transduction by the CD3 T-cell receptor. Biochem. J. 363,
645–655.
Schmid, B., 2002. Cell and molecular biology of a monocyte
lysophosphatidylcholine acyltransferase. Ph.D. Thesis,
University of Wales.
Schmid, B., Finnen, M.J., Harwood, J.L., Jackson, S.K., 2003.
Acylation of lysophosphatidylcholine plays a key role in
the response of monocytes to lipopolysaccharide. Eur.
J. Biochem. 270, 2728–2788.
Schmidt, A., Wolde, M., Thiele, C., Fest, W., Kratzin, H.,
Podtelejnikov, A.V., Witke, W., Huttner, W.B., Soling,
H.D., 1999. Endophilin 1 mediates synaptic vesicle forma-
tion by transfer of arachidonate to lysophosphatidic acid.
Nature 401, 133–141.
Schumann, R.R., Leong, S.R., Flaggs, G.W., Gray, P.W.,
Wright, S.D., Mathison, J.C., Tobias, P.S., Ulevitch, R.J.,
1990. Structure and function of lipopolysaccharide binding
protein. Science 249, 1429–1431.
Silva, A.T., Cohen, J., 1992. Role of IFN-g in experimental
Gram-negative sepsis. J. Infect. Dis. 166, 331–335.
Stark, J.M., Jackson, S.K., Taylor, S., Davies, I., Harwood, J.,
1990. The effect of endotoxin on membrane fatty acid
composition in BCG-sensitized mice. Experientia 46,
472–474.
Szamel, M., Bartels, F., Resch, K., 1993. Cyclosporin A
inhibits T cell receptor-induced interleukin-2 synthesis of
human T lymphocytes by selectively preventing a trans-
membrane signal transduction pathway leading to sus-
tained activation of a protein kinase C isoenzyme, protein
kinase C-beta. Eur. J. Immunol. 23, 3072–3081.
Szamel, M., Kaever, V., Leufgen, H., Resch, K., 1998. The
role of lysophosphatide acyltransferases and protein kinase
C isoforms in the regulation of lymphocyte responses.
Biochem. Soc. Trans. 26, 370–374.
Tou, J., 1987. Platelet-activating factor modulates phospholi-
pid acylation in human neutrophils. Lipids 22, 333–337.
Tracey, K.J., Lowry, S.F., 1990. The role of cytokine
mediators in septic shock. Adv. Surg. 23, 21–25.
Van Amersfoot, E.S., Van Berkel, T.J.C., Kuiper, J., 2003.
Receptors, mediators and mechanisms involved in bac-
terial sepsis and septic shock. Clin. Microbiol. Rev. 16,
379–414.
Weigert, R., Silletta, M.G., Spano, S., Turacchio, G., Cericola,
C., Colanzi, A., Senatore, S., Mancini, R., Polishchuk,
E.V., Salmona, M., Facchiano, F., Burger, K.N., Mironov,
A., Luini, A., Corda, D., 1999. CtBP/BARS induces fission
of Golgi membranes by acylating lysophosphatidic acid.
Nature 402, 429–433.
Winkler, J.D., Sung, C-M., Huang, L., Chilton, F.H., 1994.
CoA-independent transacylase activity is increased in
human neutrophils after treatment with tumor necrosis
factor alpha. Biochim. Biophys. Acta 1215, 133–140.
Winkler, J.D., Fonteh, A.N., Sung, C.M., Heravi, J.D., Nixon,
A.B., Chabot-Fletcher, M., Griswold, D., Marshall, L.A.,
Chilton, F.H., 1995. Effects of CoA-independent transacy-
lase inhibitors on the production of lipid inflammatory
mediators. J. Pharmacol. Exp. Ther. 274, 1338–1347.
Wright, S.D., Ramos, R.A., Tobias, P.S., Ulevitch, R.J.,
Mathison, J.C., 1990. CD14, a receptor for complexes of
LPS and LPS binding protein. Science 249, 1431–1433.
Yamashita, A., Sugiura, T., Waku, K., 1997. Acyltransferases
and transacylases involved in fatty acid remodelling of
phospholipids and metabolism of bioactive lipids in
mammalian cells. J. Biochem. 122, 1–16.
Yan, J.J., Jung, J.S., Lee, J.E., Lee, J., Huh, S.O., Kim, H.S.,
Jung, K.C., Cho, J.Y., Nam, J.S., Suh, H.W., Kim, Y.H.,
Song, D.K., 2004. Therapeutic effects of lysophosphatidyl-
choline in experimental sepsis. Nat. Med. 10, 161–167.
Zarember, K.A., Godowski, P.J., 2002. Tissue expression of
human toll-like receptors and differential regulation of toll-
like receptor mRNAs in leukocytes in response to microbes,
their products and cytokines. J. Immunol. 168, 554–561.





























