Embed Size (px)
Citation preview

Plant Physiol. (1 996) 1 1 1 : 1 O1 1-1 O1 9
lsolation of Mutations Affecting the Development of Freezing Tolerance in Arabidopsis thaliana (L.) Heynh.
Gareth Warren’*, Robert McKown2, Analuisa Marin, and Rita Teutonico3
DNA Plant Technology Corp., 6701 San Pablo Avenue, Oakland, California 94608
We screened for mutations deleterious to the freezing tolerance of Arabidopsis fhaliana (1.) Heynh. ecotype Columbia. lolerance was assayed by the vigor and regrowth of intact plants after cold acclimation and freezing. From a chemically mutagenized popula- tion, we obtained 13 lines of mutants with highly penetrant pheno- types. In 5 of these, freezing sensitivity was attributable to chilling injury sustained during cold acclimation, but in the remaining 8 lines, the absence of injury prior to freezing suggested that they were affected specifically in the development of freezing tolerance. In backcrosses, freezing sensitivity from each line segregated as a single nuclear mutation. Complementation tests indicated that the 8 lines contained mutations in 7 different genes. l h e mutants’ freez- ing sensitivity was also detectable in the leakage of electrolytes from frozen leaves. However, 1 mutant line that displayed a strong phenotype at the whole-plant leve1 showed a relatively weak phe- notype by the electrolyte leakage assay.
What genes are responsible for the development of freez- ing tolerance in hardy plants? A number of laboratories address this question by characterizing genes and proteins induced during cold acclimation (e.g. Houde et al., 1992; Neven et al., 1993; Nordin et al., 1993; Wilhelm and Tho- mashow, 1993; Castonguay et al., 1994; Dunn et al., 1994; Jarillo et al., 1994). Thomashow and co-workers have shown that the product of one such gene can cryoprotect the chloroplast in vivo (Artus et al., 1994); Hincha and colleagues (1990) have shown that cold-induced proteins are cryoprotective to thylakoids. However, it may be diffi- cult to discover whether and how a cold-induced gene contributes to freezing tolerance, e.g. when no function is demonstrable in vitro and suppression of the gene’s trans- lation by antisense RNA (which is usually not absolute) fails to generate a phenotype. Moreover, the usual tech- niques for identifying cold-induced genes may fail to iden- tify cold-induced genes that share homology with consti- tutively expressed genes; also, some genes responsible for freezing tolerance may be constitutive rather than cold induced.
Present address: Department of Biochemistry, Imperial Col- lege of Science Technology and Medicine, London, UK SW7 2AY.
Present address: College of Integrated Science and Technol- ogy, James Madison University, Harrisonburg, VA 22807.
Present address: Technology Forecasters, Inc., 1936 University Avenue, Berkeley, CA 94704.
* Corresponding author; e-mail g.warrenQic.ac.uk; fax 44-171- 225-0960.
The study of QTL offers an alternative method of looking for the genes responsible for freezing tolerance. QTL affect- ing freezing tolerance have been demonstrated in barley, potato, and oilseed Brassica species (Hayes et al., 1993; Stone et al., 1993; Teutonico et al., 1995). A major determi- nant of freezing tolerance has been localized to chromo- some 5 in wheat (Sutka, 1994). It is clear that the loci detected are agronomically significant, but they are not technically accessible to molecular characterization, since it is difficult to clone genes identified only as QTL. Again, there is also the possibility of missing important genes, in this case because they may not show allelic variation be- tween the cultivars tested.
The classical genetic approach would be to identify genes required for freezing tolerance by isolating muta- tions deleterious for this trait. The strengths of this ap- proach are that it is capable of identifying every gene responsible for freezing tolerance and that the mutants’ phenotypes could suggest specific functions for the cognate genes. The weaknesses of the classical approach lie in the difficulty of cloning genes defined only by conventional genetics and the possibility that some mutants may be uninformative. For example, a mutation that exerted its effect by weakening the plant prior to freezing would be uninformative, but mutations having such an effect could probably be screened out. The cloning of genes defined only by conventional genetics has usually required the very laborious process of ”chromosome walking,” but the size of the Arabidopsis tkaliana genome and the molecular tools and information being developed for this species mitigate this difficulty.
There has been one preliminary report of a screen for mutants of Arabidopsis deficient in freezing tolerance (Ar- tus and Thomashow, 1992). A recessive, freezing-sensitive mutation was identified using a screen based on the electrolyte leakage assay (M. Thomashow, personal communication).
We wished to isolate mutations that impaired freezing tolerance in A. tkaliana without using an assay (such as electrolyte leakage) that is conducted on excised leaves. We reasoned that such an assay might fail to detect injury restricted to other tissues, and also might be poorly suited for detecting certain types of injury within the leaf. There- fore, we chose apparent health and regrowth of intact plants after a freezing episode as the criteria by which to
Abbreviations: LT,,, temperature at which 50% injury is in- curred; QTL, quantitative trait loci; SFR, sensitivity to freezing.
1011 www.plantphysiol.orgon May 30, 2018 - Published by Downloaded from
Copyright © 1996 American Society of Plant Biologists. All rights reserved.

1012 Warren et al. Plant Physiol. Vol. 11 1, 1996
judge freezing tolerance. This presented the challenge of recovering mutations that were identified by injury, possi- bly lethal, to the plants that carried them. Also, there were severa1 unknowns that could influence the success of our approach: the frequency with which this type of mutation would occur in a mutagenized population, the frequency with which nonheritable factors would produce a pheno- copy of the desired mutation (the leve1 of "noise" in the screen), and the relative frequency of mutations that would act by causing injury before freezing. It was not even certain that a whole-plant assay could reflect the profi- ciency of Arabidopsis to acclimate; if its roots were to cold acclimate poorly, as has been reported for wheat (Zhou et al., 1994), root injury and consequent plant death would obscure differences of freezing tolerance in the leaves. This report addresses these considerations and demonstrates that it is possible to isolate mutations specifically deleteri- ous to freezing tolerance by means of a whole-plant assay.
MATERIALS AND METHODS
Plant Crowth Conditions for Screening
The pedigreed mutant set of James and Dooner (1990) derives from the Columbia ecotype of Arabidopsis thaliana. Seeds from this set were planted in a grid pattern with the location of each specifying its pedigree. Seedlings were grown for approximately 5 weeks with 8-h photoperiods at 200 to 400 pmol m-* s-' in a shaded greenhouse at 16 to 20"C, then transferred to a growth chamber at 4°C for 2 weeks with 8-h photoperiods at 220 pmol m-* s-'.
Freezing of Whole Plants
For freezes of 60 min, we used a liquid-nitrogen-cooled, controlled-rate freezer (Cryo-Med, Marietta, GA) with a cooling rate of 2"CJmin. For freezes of 24 h, we employed a domestic food freezer modified by the addition of a 7-W fan on each shelf and controlled by an electronic thermo- stat. This arrangement resulted in a regular oscillation in the chamber's air temperature, with a period of approxi- mately 5 min, a magnitude of 0.8"C, and a reproducibility of 0.1"C. In "Results" we discuss varying the freezing temperature; this refers to the nadir (lower limit) temper- ature in these oscillations. Thus, a temperature setting of -6.O"C refers to the condition at which chamber air tem- perature oscillated between -5.2 and -6.O"C. The initial cooling rate was approximately 0.25"C J min. Thawing, which was achieved by removal of the plants to ambient temperature (20"C), was rapid (approximately 5 min) for aerial structures, but longer (approximately 30 min) for soil. Probes of soil temperature indicated that equilibrium was reached approximately 4 h after the start of a freezing cycle, and remova1 of samples at this time showed that all plants and a11 soil divisions were frozen (i.e. ice nucleation had already taken place). Therefore, a11 plant structures, including roots, would remain frozen for at least 20 h of the usual 24-h cycle.
Recovery and Observation of lnjury
After freezing, plants were returned to a shaded green- house at 16 to 20°C. Plants were observed daily for up to 5 d, and those with apparent injury or reduced health relative to wild-type control plants were noted. If still alive, these plants (which represented putative mutants) were transplanted and encouraged to grow to maturity.
Gene and Mutant Designations
The unique gene designation SFR (SFR1, SFR2, etc. for different loci; sfrl , sfr2, etc. for corresponding mutant al- leles) has been registered with Dr. David Meinke, curator of Arabidopsis genetic nomenclature. SFR was chosen to abbreviate sensitivity to freezing. If future work demon- strates the identity of an SFR gene (e.g. SFR5) with that of a previously published gene, the earlier gene designation will take priority and the allele names of cognate mutations ( e g sfr5-1, sfu5-2) will be changed accordingly.
Crosses for Complementation Testing
AI1 crosses were performed using only one of the parents as the pollen donor. The mutant line was the pollen donor in backcrosses. In crosses between mutant lines, the parent to act as the pollen donor was chosen at random. To confirm that the progeny of such crosses were truly F, hybrids (rather than being due to contaminating pollen), we selfed an F, plant that had been scored as freezing tolerant from each cross except FS78 X FS79, and we tested for the expected segregation of freezing sensitivity in a sample of 96 F, plants.
Measurements of Electrolyte Leakage
Six-week-old seedlings were incubated in growth cham- bers with 8 h of light per day at 220 p E mp2, at either 16°C (to provide nonacclimated plants) or 4°C (for cold acclima- tion). After 14 d, young leaves between 40 and 80% of the size of fully expanded leaves were harvested. The leaves were washed and placed in 3-mL aliquots of distilled wa- ter, two leaves to a tube. Tubes were equilibrated to -2"C, nucleated with ice chips, and allowed to remain for 1 h at -2°C to re-equilibrate. The bath temperature was now ramped down to -10°C at a rate of 2°C h-l. Two tubes of each type were withdrawn every half-hour (including two withdrawn at the start) to provide a series of duplicate samples that had experienced nadir temperatures of -2, -3, -4"C, etc. The withdrawn tubes were held at 4°C for 16 h and then warmed to room temperature and shaken for 15 min. Electrical conductivity was measured. The tubes were then heated to 65°C for 30 min to release all electro- lytes, cooled to room temperature, and shaken. A second measurement of conductivity was made to determine the total content of electrolytes in each sample. Division of the first measurement by the second indicated the proportion of total electrolytes that had leaked from the leaves in each sample during the freezing protocol.
Means were taken from duplicate samples and plotted with estimates of SE. Logistic curves were fitted to each set
www.plantphysiol.orgon May 30, 2018 - Published by Downloaded from Copyright © 1996 American Society of Plant Biologists. All rights reserved.

Mutations Affecting Freezing Tolerance in Arabidopsis 1013
of data. The logistic function (Causton and Venus, 1981) may be written:
f ( t ) = c + A(l + e(KP--Kt))-l
with asymptotes at f(t) = c and f(t) = c + A, and inflection at t = p. We adjusted parameters c, A, K , and p to minimize the sum of squares of differences between f(t) and the sample means for each data set. We did not follow the recommendation of Von Fircks and Verwijst (1993) to use a derivative of the logistic function (the Richards function) because the small improvements in curve fit do not appear to justify the introduction of another parameter, unless data with very small SE values are available.
Mapping
The sfr2, sfr4, and sfr5-1 lines (FS61, FS67, and FS68, respectively), which have a Columbia background, were crossed with Landsberg evecta. The F, and F, generations were allowed to self, and the progeny of individual F, plants were freeze-tested to reveal (by the segregation of phenotypes) the sfr genotypes of their respective parents. DNA was isolated from families (pooling DNA from 18 or more F, plants per family to assure complete representa- tion of each family's parenta1 genome), digested with EcoRI, and subjected to a Southern blot. Probes were non- radioactively labeled by the digoxigenin method using the Genius System V2.0 kit from Boehringer Mannheim. Hy- bridization and detection methods followed the manufac- turer's recommendations.
Recombination was estimated by the maximum likeli- hood method (Mather, 1951) and converted to map dis- tances by the mapping function of Kosambi (1944).
RESULTS
Defining the Conditions for Screening
It was first desirable to find conditions for freezing intact specimens of A. thaliana (L.) Heynh. that would distinguish cold-acclimated from nonacclimated plants. Our tests uti- lized wild-type, vegetatively growing seedlings of ecotype Columbia, the ecotype that would be screened subse- quently for mutants. Initially, we cooled plants to various temperatures at a rate of 2°C min-' and then held temper- ature constant for 60 min before rapid thawing. A holding temperature of -8.O"C gave the clearest distinction (by differences in visible damage, observed 2 d later) between cold-acclimated and nonacclimated plants. However, dam- age to nonacclimated plants increased considerably from the center to the outside of a tray; we ascribed this to slower temperature equilibration at the center. To circum- vent this effect, we extended the plants' low-temperature exposure to 24 h and reduced the cooling rate to 0.25"C min-'.
We varied the holding temperature under this new re- gime and found that temperatures between -4.9 and -6.O"C clearly differentiated cold-acclimated from nonac- climated plants. Below -6.0°C, partia1 damage to cold- acclimated plants reduced the clarity of the distinction.
Above -4.9"C, the incompleteness of the damage to non- acclimated plants made their sensitivity less obvious. We tried reducing the holding period to 12 h, but this reintro- duced variability according to position within a tray, and so we abandoned this modification. For screening we chose the lowest temperature compatible with clear detection of the cold-acclimated state (-6.O"C) because we wanted to be able to detect mutants only slightly less hardy than the wild type.
lsolating Mutants
Because the testing process might be lethal for the de- sired mutants, we screened a collection of mutagenized germ plasm that was organized so that siblings of a puta- tive mutant could be found. The collection of James and Dooner (1990) consists of M, seed pools derived from 1804 M, plants from ethyl methanesulfonate mutagenesis of the ecotype Columbia. We screened 2 plants from each M, pool. In 30 of the 1804 pools, just 1 of the 2 screened plants appeared sensitive to freezing, whereas in 2 pools, both plants were injured. (Thus, we observed a total of 34 plants injured out of approximately 3600 tested.) Injury was not always lethal: 14 of the injured plants survived and grew to maturity. Their M, progeny were subsequently tested for freezing sensitivity. However, this left 18 pools from which the putative mutation had to be sought among siblings of a killed plant. Eight siblings from each such pool were grown to maturity, and their M, progeny were tested for freezing sensitivity.
By testing the M, families we could identify M, parents that had bred true for freezing sensitivity, because all M, members of such families would be freezing sensitive. This was true in 13 pedigrees; we were thus left with 13 true- breeding lines, discounting duplicate M, lines from the same pedigree. The other 19 pedigrees failed to yield true- breeding M, plants. Some gave no freezing sensitives at a11 among the M, plants; this is attributable to purely coinci- dental injury (from some other cause) to the pedigree's representative in the original screen. However, some ped- igrees gave apparent segregation of freezing sensitivity among M, families without yielding any M, families in which all members displayed the trait. Severa1 explana- tions for this are plausible: (a) among a pedigree's M, plants, mutant homozygotes were inviable or infertile, so that only heterozygotes gave rise to progeny; (b) the mu- tation had incomplete penetrance, therefore giving the ap- pearance of segregation even among progeny of homozy- gotes; or (c) by chance, none of the M, plants that had grown out were homozygous. Because of the last possibil- ity, we repeated our sampling of M, siblings, grew them out, and tested their M, families. However, no more true- breeding lines were obtained. We decided not to pursue mutations likely to be (a) homozygous inviable or sterile or (b) incompletely penetrant, and therefore abandoned all pedigrees that had failed to yield a true-breeding M, plant.
Since our goal was to isolate mutations causing a specific deficiency in the process of cold acclimation, we wished to identify and eliminate mutant lines in which freezing sen- sitivity was due to injury incurred before freezing (during
www.plantphysiol.orgon May 30, 2018 - Published by Downloaded from Copyright © 1996 American Society of Plant Biologists. All rights reserved.

1014 Warren et al. Plant Physiol. Vol. 111, 1996
the period of cold acclimation). Mutations of the latter type(i.e. chilling-sensitive mutants) have been previously ob-served by Hugly et al. (1990) and Schneider et al. (1995).
Seedlings of the 13 lines were cold acclimated and thenexamined for chilling injury by two criteria: visible damageduring cold acclimation and stunted growth upon a returnto normal temperature. Five mutant lines showed injuryduring acclimation, in every case by both criteria. Thesefive lines were therefore eliminated from further consider-ation. We tentatively concluded that the remaining eightlines contained mutations causing more specific deficien-cies in the processes that confer freezing tolerance on thewild-type plant. The freezing-sensitive phenotypes bywhich these lines were recognized are illustrated in Figure1. The controls shown in this figure, which are plants of themutant lines grown and cold acclimated in parallel but notfrozen, illustrate the absence of chilling injury in theselines.
Penetrance, Dominance, and Mendelian Inheritance
We assessed the penetrance of each mutant line's phe-notype with a minimum of three separate tests. We mea-sured not only the frequency with which mutant pheno-types were distinct from the typical wild type, but also thefrequency with which wild-type controls, tested in parallel,were distinct from the typical mutant phenotype. The ef-fective penetrance is given as the product of both frequen-cies (Table I). Penetrance was high enough in each case topermit genetic analysis.
To assess dominance, backcrosses were made to wildtypes of the same genetic background (Columbia). TwelveF] plants from each cross, divided between two separatelyconducted tests, were frozen alongside wild-type and ho-mozygous mutant control plants, and the F5 plants' freez-ing tolerance was compared with both. As reported inTable I, F, phenotypes were in most cases similar to wildtype, indicating that the respective mutations are recessive.The Fj plants derived from backcrosses of lines FS68 andFS79 were exceptional, having intermediate levels of freez-ing tolerance, and thus showing codominance.
Figure 1. Phenotypes of mutant lines in com-parison to wild type (wt). Three plants of eachline (Tests) are shown 9 d after freezing understandard conditions (24 h at -6.0°C). Twoplants of each line (Controls), with a parallelhistory except for freezing, are shown ascontrols for any injurious effects of the cold-acclimation period.
F, plants from the backcrosses were allowed to self andthe F2 plants were screened to observe segregation of themutant phenotype. In all cases the segregation ratios dif-fered significantly from 15:1 but not from 3:1. This is con-sistent with single-gene segregation and refutes the hy-pothesis that freezing sensitivity is caused by the combinedeffects of two unlinked recessive mutations (Table II). An-other two-gene hypothesis is that freezing sensitivity iscaused by the combined action of a recessive mutation anda dominant mutation at unlinked loci. This predicts an F2segregation ratio of 13:3, which is experimentally difficultto distinguish from 3:1. However, according to this hypoth-esis (or any hypothesis requiring two unlinked genes), only1 in 16 F2 progeny will breed true for the mutant pheno-type. Therefore, we examined the genotypes of F2 individ-uals by freezing families of their F3 progeny. For everymutant line, the ratio of F2 individuals breeding true forfreezing sensitivity to all other F2 individuals differed sig-nificantly from 1:15 but not from 1:3 (Table II). This indi-cates that in each of the 7 mutant lines tested (FS79 wasomitted), freezing sensitivity was caused by mutation in asingle nuclear gene.
Complementation
The eight mutant lines were crossed together in all pair-wise combinations, except for some involving FS79 (seebelow). Approximately 12 F, progeny from each of thesecrosses were frozen alongside control plants of both paren-tal lines and wild type. In the majority of such tests (TableIII), the F, progeny appeared less freezing sensitive thaneither parent and usually no more sensitive than the wildtype (an example is shown in Fig. 2a). Such a result indi-cates complementation and therefore suggests that the re-spective mutations affect different genes. The only excep-tion involved the F, progeny from the cross FS68 x FS79,which were as freezing sensitive as either parent andclearly distinct from wild type (Fig. 2b). The lack ofcomplementation here suggests that the mutations in theFS68 and FS79 lines are allelic. To confirm their allelism, weallowed the F] progeny from FS68 X FS79 to self, and froze
www.plantphysiol.orgon May 30, 2018 - Published by Downloaded from Copyright © 1996 American Society of Plant Biologists. All rights reserved.

Mutations Affecting Freezing Tolerance in Arabidopsis 1015
Table 1. Penetrance and dominance of mutations
Mutants Distinct Wild Type Distinct Effective Phenotypes of Fla from Wild Type from Mutant lnference
FS59 59/61 69/72 92% O O 12 Recessive FS61 6 816 8 73/73 100% O O 9 Recessive FS65 69/69 68/69 98% O O 12 Recessive FS67 5 615 6 5 715 7 100% O 4 20 Recessive FS68 56/57 56/56 98% 3 8 1 Codominant FS69 44/44 44/44 100% O O 12 Recessive FS78 5 615 6 5 715 7 100% O O 13 Recessive FS79 5 815 8 63/63 100% 1 6 6 Codominant
Mutant Line Penetrance Mutant lntermediate Wild type
a F, plants derived from backcrosses to ecotype Columbia.
the F, plants. A11 108 of the F, plants examined were freezing sensitive. The absence of segregants with wild- type freezing tolerance refutes other possible explanations for the lack of complementation in the F, progeny and confirms allelism. After reaching this conclusion, we per- formed no additional crosses with FS79.
Description of Freezing Sensitivity and Pleiotropic Effects
Since only two of the eight mutant lines contained mu- tually allelic mutations, the eight lines defined seven genes that were required for freezing tolerance after cold accli- mation. In accord with convention, we named these genes for the mutant phenotype (sensitivity to freezing): they are SFRl to SFR7. The correspondence of mutant alleles (sfvl , etc.) to mutant lines is given in Table IV.
As is apparent from Figure 1, the mutant lines do not manifest freezing sensitivity in identical ways. The differ- ences we observed are summarized in Table IV (third column). In three of the lines we also observed other her- itable characteristics that distinguished them from wild type (Table IV, fourth column). A11 such characteristics were recessive. To determine whether these characteristics were pleiotropic effects of the sfr mutations, we examined their cosegregation with freezing sensitivity. The F, popu- lations described in Table I1 were used. There was complete cosegregation in each case (Table IV, fifth column), indi- cating that the other phenotypes were indeed pleiotropic effects of the sfr mutations.
Electrolyte Leakage Assays
The plasmalemma may be the primary site of freezing injury in nonacclimated plants (Steponkus, 1984). The leak- age of electrolytes from frozen and thawed tissues is a sensitive indicator of loss of integrity by the plasmalemma, and it has been commonly used to assay freezing injury (reviewed by Calkins and Swanson, 1990). We applied this assay to the mutants to determine whether their freezing sensitivity was manifested by increased sensitivity of the plasmalemma in leaf tissues. We also wished to assess the applicability of the electrolyte leakage assay as a screening method for mutants.
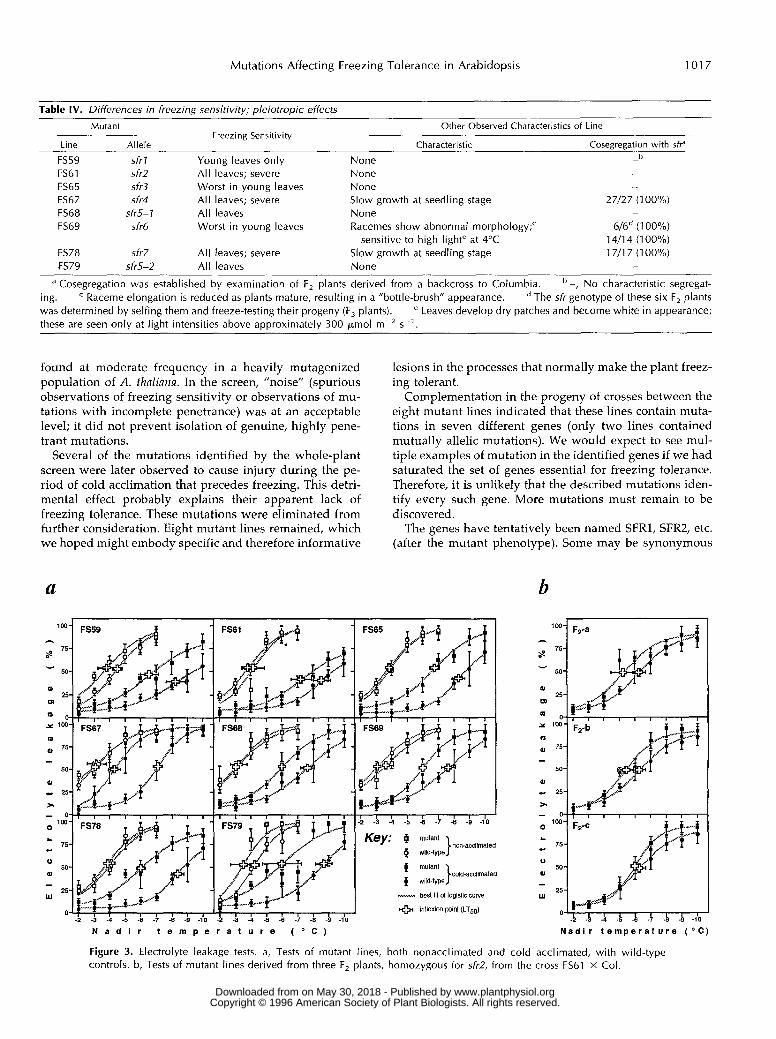
The electrolyte leakage assay was applied both to non- acclimated and cold-acclimated leaf tissue of each geno- type. It was not technically feasible to conduct a11 the tests simultaneously; therefore, leaf tissue from wild-type plants, grown and treated in parallel, was included as a control each time the assay was conducted. For each data set, a best fit to the sigmoid logistic function was calcu- lated. The inflection point of each fitted curve was taken as the best estimate of LT,,, and confidence limits for this temperature were calculated based on the goodness of fit of the curve. The data and estimates of LT,, are presented in Figure 3a.
The freezing tolerance of nonacclimated leaf tissue is depicted in the data identified by open symbols. In most cases, the inferred LT,, value for mutant tissue was very close to that for the wild-type control. The inferred LT,,
Table II. Segregation of phenotypes and genotypes
Phenotypes of F,a lndividuals Cenotypes of F, lndividuals
Probability of chance Observed Probability of chance deviation from segregation deviation from Mutant Line Observed segregation
Wild type/intermediate Mutant 15:l 3:l Other m/m 15:l 3: l
FS59 80 22 <0.001 0.499 ndh nd nd nd FS61 54 22 <0.001 0.500 8 5 0.002 0.41 2 FS65 88 39 <0.001 0.1 71 13 4 0.038 1.000 FS67 67 27 <0.001 0.469 21 6 0.01 2 0.942 FS68 88 39 <0.001 0.1 71 8 4 0.01 o 0.702 FS69 74 20 <0.001 0.482 15 6 0.003 0.867 FS78 75 17 <0.001 0.1 80 26 6 0.026 0.556
a F, plants derived from backcrosses to ecotype Columbia. nd, Not determined.
www.plantphysiol.orgon May 30, 2018 - Published by Downloaded from Copyright © 1996 American Society of Plant Biologists. All rights reserved.

1016 Warren et al. Plant Physiol. Vol. 111, 1996
Table III. Complementation resultsPhenotypes of F, plants, resulting from pairwise crosses of mutant lines, are reported as follows: wt, freezing tolerance
indistinguishable from wild type; MUT, freezing tolerance indistinguishable from one or both of the parental mutant lines;int, freezing tolerance intermediate between mutant and wild type; nd, not determined.
Crossed with
FS59FS61FS65FS67FS68FS69FS78
FS79
ndintintint
MUTndnd
FS78
wtwtwtwtintwt
FS69
wtwtwtwtint
FS68
intintintint
FS67 FS65
wt wtwt wtwt
FS61
wt
values for FS59 and FS79 differed the most from theirrespective controls, but even here the differences were notstatistically significant. Therefore, we failed to observe anyincreased freezing sensitivity of the mutants in the nonac-climated state.
The freezing tolerance of cold-acclimated leaf tissue isdepicted in the data identified by filled symbols. Everymutant yielded an inferred LT50 value that was signifi-cantly higher than that of the wild-type control. The dif-ference was greatest for FS67 and FS78, which is consistentwith their severely freezing-sensitive phenotypes, deter-mined by visual observation. However, the inferred LT50value for FS61 was closest to that of the cognate control.Since we know by the whole-plant assay that FS61 also hasa severely freezing-sensitive phenotype, this result seemedodd, and perhaps due to an interfering phenotype fromanother mutation. Therefore, we repeated both types ofassay on three lines selected from the F2 of the FS61 back-cross; these lines are homozygous for the sfr2 mutation, butit is unlikely that they would all be homozygous for asecond mutation. Nevertheless, the electrolyte leakage as-say again showed a relatively small effect (1.1°C) on LT50(Fig. 3b), whereas the whole-plant assay reproduced the
severe freezing sensitivity seen in the original sfr2 line,FS61 (data not shown). Therefore, we concluded that cold-acclimated sfr2 leaf tissue behaves anomalously betweenthe two assays.
Preliminary Mapping
By examining recombination between three sfr mutationsand a set of markers designed for preliminary mapping(Fabri and Schaffner, 1994), we were able to map the mu-tations to their respective chromosomes. sfr2, sfr4, andsfr5-l mapped to chromosomes 3, 2, and 1, respectively(Table V). It was to be expected from the complementationdata that they would map to distinct loci.
DISCUSSION
After freezing intact plants, including their root systems,it proved possible to reliably distinguish cold-acclimatedfrom nonacclimated plants. This permitted identification ofmutants impaired in freezing tolerance after cold acclima-tion without prejudicing the screen by looking for partic-ular symptoms of injury. Highly penetrant mutations were
Figure 2. Examples of positive and negativecomplementation results, a, F, from the crossFS61 x FS65. Lack of freezing injury showscomplementation and demonstrates that thecognate mutations are not allelic. b, F, from thecross FS68 X FS79. Freezing injury shows anabsence of complementation and demonstratesthat the cognate mutations are allelic. In eachtest, six plants of each parental line and of thewild type were grown and frozen in parallel.Photographs were taken 4 d after freezing.
aFS61
bFS61 x FS65 FS68 FS68 x FS79
wild type FS65 wild type FS79 www.plantphysiol.orgon May 30, 2018 - Published by Downloaded from
Copyright © 1996 American Society of Plant Biologists. All rights reserved.
Muta t ions Affecting Freezing Tolerance in Arabidopsis 1 0 1 7
Table IV. Differences in freezina sensitivitv; oleiotrooic effects
Mutant
Line
FS59 FS61 FS65 FS67 FS68 FS69
FS78 FS79
Allele sfr 1 sfr2 sfr3 sfr4
sfr5- 1 sfr6
sfr7 sfr5-2
Freezing Sensitivity
Young leaves only All leaves; severe Worst in young leaves All leaves; severe All leaves Worst in young leaves
All leaves; severe Ali leaves
Other Observed Characteristics of Line
Characteristic
None None None Slow growth at seedling stage None Racemes show abnormal morphology;'
Slow growth at seedling stage None
sensitive to high light' at 4°C
Cosegregation with sf? b -
- 27/27 (1 00%)
6/6d (1 009'0) 1 4/14 (1 00%) 17/17 (1 00%)
-
-
a Cosegregation was established by examination of F, plants derived from a backcross to Columbia. Raceme elongation is reduced as plants mature, resulting in a "bottle-brush" appearance.
-, No characteristic segregat- The sfr genotype of these six F, plants
e Leaves develop dry patches and become white in appearance; ing. was determined by selfing them and freeze-testing their progeny (F, plants). these are seen only at light intensities above approximately 300 pmol m-, s-'.
found at moderate frequency in a heavily mutagenized population of A. thaliana. In the screen, "noise" (spurious observations of freezing sensitivity or observations of mu- tations with incomplete penetrance) was at an acceptable level; it did not prevent isolation of genuine, highly pene- trant mutations.
Severa1 of the mutations identified by the whole-plant screen were later observed to cause injury during the pe- riod of cold acclimation that precedes freezing. This detri- mental effect probably explains their apparent lack of freezing tolerance. These mutations were eliminated from further consideration. Eight mutant lines remained, which we hoped might embody specific and therefore informative
a
lesions in the processes that normally make the plant freez- ing tolerant.
Complementation in the progeny of crosses between the eight mutant lines indicated that these lines contain muta- tions in seven different genes (only two lines contained mutually allelic mutations). We would expect to see mul- tiple examples of mutation in the identified genes if we had saturated the set of genes essential for freezing tolerance. Therefore, it is unlikely that the described mutations iden- tify every such gene. More mutations must remain to be discovered.
The genes have tentatively been named SFR1, SFR2, etc. (after the mutant phenotype). Some may be synonymous
b
N a d i r t e m p e r a t u r e ( " C ) N a d i r t e m p e r a t u r e ( " C )
Figure 3. Electrolyte leakage tests. a, Tests of mutant lines, both nonacclimated and cold acclimated, with wild-type controls. b, Tests of mutant lines derived from three F, plants, homozygous for sfi-2, from the cross FS61 X Col.
www.plantphysiol.orgon May 30, 2018 - Published by Downloaded from Copyright © 1996 American Society of Plant Biologists. All rights reserved.

1 0 1 8 Warren et al. Plant Physiol. Vol. 1 1 1 , 1996
Table V. Preliminarv mappinn data SeparatiorP (centiMorgan) Marker Observed
Recombination Estimate 95% confidence limits Allele nb
sfr2 560B1 3:21 .5 112 0.210 22 12-31
sfr4 497A 2:18.1 48 0.333 40 24-80 sfr5- I 241 A 1:32.0 92 0.1 85 19 12-29
Name Location"
249A 3:64.6 134 0.436 67 42- unl in ked
254A 1:56.3 92 0.385 50 34-85
a Map location is given in the form of (chromosome:position). n is the number of gametes that were informative for the measurement of recombination. Separation is derived from recomhination data hy the mapping function of Kosambi (1 944).
with genes already described on the basis of other pheno- types or on the basis of their cold-inducible expression. The aba mutation is known to have a strong effect on freezing tolerance (Heino et al., 1990). None of our mutants matches the usual aba phenotype (yellow-brown color, withering), but it remains possible that one of them may represent an allele of aba. The abil mutation has a weak effect on cold- induced freezing tolerance (Mantyla et al., 1995), and like- wise, one of the sfr mutations may be allelic with abil . The question of possible identity with these and many other previously characterized genes will best be resolved by mapping. We have presented preliminary mapping data for three of the mutations.
By examining the freezing sensitivity of leaves of the mutants in a different assay, we can say whether they could have been isolated had we based our screen on such an assay. The electrolyte leakage assay commonly used to quantify variations in the level of freezing tolerance, de- tected freezing sensitivity to various degrees in a11 of the mutants. Therefore, in theory, a11 could have been detected in a screen based on that assay. Because of the variability inherent in the electrolyte leakage assay, we suspect that in practice such a screen would not be completely efficient in detecting mutant lines such as FS61 or FS68 and might also be subject to considerable noise (spurious indications of sensitivity from plants without a heritable deficiency in freezing tolerance).
The electrolyte leakage assays, because they are per- formed on isolated leaves, show that each mutation is directly deleterious to the freezing tolerance of leaf tissue. Moreover, because the assay depends on injury to the plasma membrane, the mutants must sustain such injury to a greater extent than the wild type within 16 h of thawing. This leaves open the possibility that the plasmalemma is the mutants' primary site of injury. However, a mutation with a strong phenotype at the level of the whole plant, sfr2, has one of the weakest phenotypes by the electrolyte leakage assay. The source of this discrepancy, among the many differences between the two types of assays, may be interesting.
We hope that our investigation of genetic lesions in freezing tolerance will have synergy with research else- where on cold-induced genes. If an sfr mutation is found to affect a cold-induced gene, the mutation's phenotype will provide evidence of that gene's role in freezing tolerance. Moreover, the mutant line could be used to test the activity
of the gene after manipulations designed to probe its function.
ACKNOWLEDCMENTS
We are indebted to Drs. Hugo Dooner and Douglas James for their generous help and encouragement. Reba Brindley and Chris- tine Chung provided technical assistance.
Received December 11, 1995; accepted April 29, 1996. Copyright Clearance Center: 0032-0889/96/111/ 1011 /09.
LITERATURE ClTED
Artus NN, Gilmour SJ, Lin C, Thomashow MF (1994) Use of transgenic Arabidopsis plants to evaluate the roles of cold- regulated genes in freezing tolerance (abstract no. 939). Plant Physiol 105: S-169
Artus NN, Thomashow MG (1992) A cold acclimation mutant of Arabidopsis (abstract no. 735). Plant Physiol 99: S-123
Calkins JB, Swanson BT (1990) The distinction between living and dead plant tissue-viability tests in cold hardiness research. Cryobiology 27: 194-211
Castonguay Y, Laberge S, Nadeau P, Vezina L-P (1994) A cold- induced gene from Medicago sativa encodes a bimodular protein similar to developmentally regulated proteins. Plant Mo1 Biol24
Causton DR, Venus JC (1981) The Biometry of Plant Growth. Edward Arnold, London
Dunn MA, Goddard NJ, Zhang L, Pearce RS, Hughes MA (1994) Low-temperature-responsive barley genes have different control mechanisms. Plant Mo1 Biol 24: 879-888
Fabri CO, Schaffner AR (1994) An Arabidopsis thaliana RFLP map- ping set to localize mutations to chromosomal regions. Plant J 5:
Hayes PM, Blake T, Chen THH, Tragoonrung S, Chen F, Pan A, Liu B (1993) Quantitative trait loci on barley (Hordeum vulgare L.) chromosome 7 associated with components of winterhardiness. Genome 36: 66-71
Heino P, Sandman G, Lang V, Nordin K, Palva ET (1990) Abscisic acid deficiency prevents development of freezing tolerance in Arabidopsis thaliana L. Heynh. Theor Appl Genet 79: 801-806
Hincha DK, Heber U, Schmitt JM (1990) Proteins from frost- hardy leaves protect thylakoids against mechanical freeze-thaw damage in vitro. Planta 180: 416-419
Houde M, Danyluk J, Laliberte JF, Rassart E, Dhindsa RS, Sar- han F (1992) Cloning, characterization, and expression of a cDNA encoding a 50-kilodalton protein specifically induced by cold acclimation in wheat. Plant Physiol 99: 1381-1387
Hugly S, McCourt P, Browse J, Patterson GW, Somerville C (1990) A chilling sensitive mutant of Arabidopsis with altered steryl-ester metabolism. Plant Physiol 93: 1053-1062
James DWJ, Dooner HK (1990) Isolation of EMS-induced mutants in Arabidopsis altered in seed fatty acid composition. Theor Appl Genet 8 0 241-245
799-804
149-156
www.plantphysiol.orgon May 30, 2018 - Published by Downloaded from Copyright © 1996 American Society of Plant Biologists. All rights reserved.

Mutations Affecting Freezing Tolerance in Arabidopsis 1019
Jarillo JA, Cape1 J, Leyva A, Martinez-Zapater JM, Salinas J (1994) Two related low-temperature-inducible genes of Arabidopsis en- code proteins showing high homology to 14-3-3 proteins, a family of putative kinase regulators. Plant Mo1 Biol 2 5 693-704
Kosambi DD (1944) The estimation of map distance from recom- bination values. Annals of Eugenics 1 2 172-175
Mantyll E, Ling V, Palva ET (1995) Role of abscisic acid in drought-induced freezing tolerance, cold acclimation, and accu- mulation of LT178 and RAB18 proteins in Arabidopsis thaliana. Plant PhysiollO7 141-148
Mather K (1951) The Measurement of Linkage in Heredity. Methuen/ John Wiley, London
Neven LG, Haskell DW, Hofig A, Li QB, Guy CL (1993) Charac- terization of a spinach gene responsive to low temperature and water stress. Plant Mo1 Biol 21: 291-305
Nordin K, Vahala T, Palva ET (1993) Differential expression of two related low-temperature-induced genes in Arabidopsis thali- una L. Heynh. Plant Mo1 Biol 21: 641-653
Schneider JC, Hugly S, Somerville CR (1995) Chilling-sensitive mutants of Arabidopsis. Plant Mo1 Biol Rep 13: 11-17
Steponkus PL (1984) Role of the plasma membrane in freez- ing injury and cold acclimation. Annu Rev Plant Physiol 35: 543-584
Stone JM, Palta JP, Bamberg JB, Weiss LS, Harbage JF (1993) Inheritance of freezing resistance in tuber-bearing Solanum spe- cies: evidence for independent genetic control of nonacclimated freezing tolerance and cold acclimation capacity. Proc Natl Acad Sci USA 90: 7869-7873
Sutka J (1994) Genetic control of frost tolerance in wheat (Triticum aestivum L.). Euphytica 77: 277-282
Teutonico RA, Yandell B, Satagopan JM, Ferreira ME, Palta JP, Osborn TC (1995) Genetic analysis and mapping of genes con- trolling freezing tolerance in oilseed Brassica. Molecular Breed- ing 1: 329-339
Von Fircks HA, Verwijst T (1993) Plant viability as a function of temperature stress: the Richards function applied to data from freezing tests of growing shoots. Plant Physiol 103: 125-130
Wilhelm KS, Thomashow MF (1993) Arabidopsis thaliana corl5b, an apparent homologue of corl5a, is strongly respon- sive to cold and ABA, but not drought. Plant Mo1 Biol 23:
Zhou BL, Arakawa K, Fujikawa S, Yoshida S (1994) Cold-induced alterations in plasma membrane proteins that are specifically related to the development of freezing tolerance in cold-hardy winter wheat. Plant Cell Physiol 35: 175-182
1073-1077
www.plantphysiol.orgon May 30, 2018 - Published by Downloaded from Copyright © 1996 American Society of Plant Biologists. All rights reserved.