Embed Size (px)
Citation preview

Research Communication
Low Folate Transport Across Intestinal Basolateral Surface isAssociated with Down-regulation of Reduced Folate Carrierin In Vivo Model of Folate Malabsorption
Abid Hamid1, Manjula Kiran1, Satyavati Rana2 and Jyotdeep Kaur11Department of Biochemistry, Postgraduate Institute of Medical Education and Research, Chandigarh, India2Department of Gastroenterology, Postgraduate Institute of Medical Education and Research, Chandigarh, India
Summary
The process of folate transport regulation across biologicalmembranes is of considerable interest because of its ultimaterole in providing one-carbon moieties for key cellular metabolicreactions and exogenous requirement of the vitamin in mam-mals. Although, intestinal folate malabsorption is establishedphenomena in alcoholism; however, there is no knowledgeregarding the mechanism of folate exit across intestinal basolat-eral membrane (BLM) to circulation during alcohol associatedmalabsorption. In the present study, male Wistar rats were fed1 g/kg body weight/day ethanol (20% solution) orally for 3months and regulatory characteristics of folate transport atBLM surface were evaluated. The folate transport was found tobe carrier mediated, saturable, with pH optima at 7.0, besidesexhibiting Na
1independence. The chronic alcohol ingestion
resulted in alteration of transport kinetics, shifting the processto K1 dependent one besides affecting the status of S��S link-age of the transport system. Importantly, chronic ethanol inges-tion reduced the folate exit across the BLM by decreasing theaffinity of transporter (high Km) for substrate and by decreas-ing the number of transporter molecules (low Vmax) on the sur-face. The decreased basolateral transport activity was associ-ated with down-regulation of the reduced folate carrier (RFC)which resulted in decreased RFC protein levels in BLM in ratmodel of alcoholism. The study suggests that during alcoholingestion, RFC mediated deregulated folate transport acrossBLM also attributes to folate malabsorption. � 2009 IUBMB
IUBMB Life, 61(3): 236–243, 2009
Keywords basolateral membrane; folate; alcoholism; reduced folate
carrier; transport.
INTRODUCTION
Folate is a highly lipophobic bivalent anion that can only
minimally traverse biological membranes by simple diffusion. It
plays a critical role in maintaining normal metabolic, differen-
tiation and growth status of mammalian cells (1). Because
humans and other mammals cannot synthesize it, so they must
obtain folate from exogenous source via intestinal absorption.
The epithelium of the small intestine is located at a strategic
interface where the intestinal lumen is in continuity with the
external environment. After intracellular modifications, reduced
and methylated form of folate crosses the basolateral enterocyte
membrane by an active anion exchange mechanism into the
portal circulation via submucosa (2). The most well character-
ized folate transporter, the reduced folate carrier (RFC) is an in-
tegral membrane protein that mediates cellular uptake of
reduced folates and antifolates and is ubiquitously expressed in
tissues (3, 4) to play a central role in tissue folate homeostasis.
However, in contrast to our knowledge of molecular identity,
functional properties and distribution of the RFC uptake system,
little is known about the mechanisms that regulate membrane
folate transport system in absorptive epithelia, particularly dur-
ing pathophysiological conditions involving derangement in its
transport. This is important because the processes involved in
the absorption, transport and intracellular metabolism of folate
are complex and are quite susceptible to cellular microenviron-
ment (5–8).
Folic acid deficiency is associated with chronic alcoholism
worldwide and ethanol is considered as one of the most impor-
tant toxin consumed regularly and in large quantities by humans
(9, 10). Alcohol intake exerts a multifaceted impact on the fo-
late bioavailability and subsequently on one-carbon folate me-
tabolism (11). Regardless of cause, folate deficiency leads to a
variety of clinical abnormalities like megaloblastic anemia and
growth retardation, whereas optimization of folate homeostasis
prevents certain disorders like neural tube defects (12). Alcohol
Present address of Abid Hamid, Division of Pharmacology, Indian
Institute of Integrative Medicine (CSIR), Jammu 180 001, India.
Address correspondence to: Jyotdeep Kaur, Department of Biochem-
istry, Postgraduate Institute of Medical Education and Research,
Chandigarh 160 012, India. Tel: 191-172-2755181. Fax: 191-172-
2744401/2745078. E-mail: [email protected]
Received 3 September 2008; accepted 13 October 2008
ISSN 1521-6543 print/ISSN 1521-6551 online
DOI: 10.1002/iub.153
IUBMB Life, 61(3): 236–243, March 2009

ingestion can be proposed to have role in activity of the RFC
because of its association with intestinal folate malabsorption
and hence folate deficiency. This inturn may influence the folate
delivery via submucosa across basolateral membrane (BLM)
and thereby to the body folate homeostasis, which also may at-
tribute to alcohol induced folate deficiency. Earlier, we reported
the derangement of RFC across intestinal brush border mem-
brane (BBM) surface during chronic alcohol ingestion (13).
However, no studies have been attempted to deduce the activity
of folate transport system in BLM under such conditions and its
role in folate homeostasis. This is important in view of the fact
that the basolateral surface not only regulates absorptive and se-
cretory functions of the folate, but also processes it; thereby
forms an important factor for determining circulatory folate
concentrations. For this, the present work was sought to charac-
terize the folate transport across the intestinal BLM and the role
that expression of the RFC plays in folate transport during alco-
holism. Such a study will help to understand the insights of
folate transport impairment in alcoholism and also in conditions
such as intestinal diseases and congenital disorders in the trans-
port systems that might underlie similar mechanism.
MATERIALS AND METHODS
Animals
Young adult male albino rats (Wistar strain) weighing 100–
150 g were obtained from Institute’s Central Animal House.
The animals were housed in clean wire mesh cages with con-
trolled temperature (23 6 1 8C) and humidity (45–55%) and
had 12 h dark light cycle throughout the study. The animals
were acclimatized to laboratory conditions for few days before
the experiment. The rats were randomized into two groups of
six animals each, such that the mean body weights and the
range of body weights for each group of animals were similar.
The rats in group I were given 1 g ethanol (20% solution)/kg
body weight/day and those in group II received isocaloric
amount of sucrose (36% solution) orally by Ryle’s tube daily
for 3 months. The rats were fed commercially available pellet
diet (Ashirwad Industries, India) and water ad libitum. The
body weights of rats were recorded twice weekly. At the end of
the treatment, animals from both the groups were sacrificed
under anesthesia using sodium pentothal and blood was drawn
for alcohol estimations employing alcohol dehydrogenase (14).
Starting from the ligament of Trietz; 2/3rd of small intestine
was removed, flushed with saline and used for various studies.
Prior approval was sought from ‘‘Institutional Animal Ethical
Committee’’ (IAEC) and ‘‘Institutional Biosafety Committee’’
(IBC) on commencement of the study.
Chemicals
Radiolabelled [30, 50, 7, 9-3H]-folic acid, potassium salt with
specific activity 24.0 Ci/mmol was purchased from Amersham
Pharmacia Biotech (Kwai Chung, Hong Kong). D-[U-14C]-glu-
cose with specific activity 140 mCi/mmol was provided by Ra-
dioisotope Division, Bhabha Atomic Research Centre (Mumbai,
India). DL-dithiothreitol (DTT) or Cleland’s reagent were pur-
chased from Sigma Aldrich Co. (St Louis, MO). Cellulose ni-
trate membrane filters (0.45 lm) were obtained from Millipore
Corporation (Bedford, MA) and Percoll was procured from
Fluka (Switzerland). Diaminobenzidine (DAB), HRP-labeled
anti rabbit-IgG secondary antibodies were obtained from Banga-
lore Genei (Bangalore, India). All other chemicals and reagents
used in this study were of analytical grade.
Isolation of Basolateral Membrane Vesicles FromSmall Intestine
Basolateral Membrane Vesicles (BLMV) from intestine
were prepared by the self-generating percoll gradient method
of Scalera et al. (15) with some modifications. Tissue was
flushed with ice cold normal saline and divided into 10 cm
segments. Intestine was then slit open and mucosa scraped
using a metal spatula. The scrapings were homogenized in ice-
cold buffer containing 250 mM mannitol and 12 mM HEPES/
Tris, pH 7.4 using a Waring blender for 3 min and then centri-
fuged at 2,500g for 20 min. The supernatant was then centri-
fuged at 22,000g for 25 min and the resulting fluffy layer of
the pellet resuspended in same buffer followed by homogeni-
zation in glass Teflon homogenizer. The resulting homogenate
was mixed with percoll at a concentration of 15.4% and centri-
fuged at 48,000g for 2 h. A distinct band of basolateral mem-
brane vesicles (BLMV) was seen at the upper one third of the
percoll gradient. The band was aspirated by a syringe and sus-
pended in buffer composing 100 mM mannitol, 100 mM KCl,
12 mM HEPES/Tris, pH 7.4 and centrifuged at 48,000g for 20
min. The pellet obtained was resuspended in loading buffer
containing 280 mM mannitol and 20 mM HEPES/Tris, pH 7.4
and centrifuged at 48,000g for 20 min twice in order to wash
out the residual percoll from membrane preparations. The final
pellet representing purified BLMV was suspended in loading
buffer so as to obtain final protein concentration of 5 mg/mL.
Purity of the membrane preparations was checked by meas-
uring the specific activities of alkaline phosphatase and Na1,
K1-ATPase in BLMV and in original homogenate by the
method of Bergmeyer and Quigley and Gotterer (16, 17)
respectively. The vesicle preparations from both the groups
showed enrichment of 8–10-fold with respect to Na1, K1-
ATPase activity and with negligible activity of alkaline phos-
phatase. Under such conditions, the Na1, K1-ATPase activity
was 3.07 6 0.20 and 3.39 6 0.32 lmol/min/mg protein in
control and ethanol fed group respectively. The observed alka-
line phosphatase activity was 0.102 6 0.002 and 0.123 6
0.010 lmol/min/mg protein in control and ethanol fed group
respectively. It is interesting to mention here that Na1, K1-
ATPase serves as the marker enzyme for the BLM and alka-
line phosphatase to that of brush border membrane. Moreover,
the vesicles were sealed and intact without any contamination
of subcellular organelles, and were similar in the two groups
237INTESTINAL BASOLATERAL FOLATE TRANSPORT IN MALABSORPTION

of rats with right side out orientation. The method of Lowry
et al. (18) was used for the determination of protein concentra-
tion using bovine serum albumin as a standard. The functional
integrity of the BLMV was checked by the [14C]-D-glucose
uptake which didn’t reveal a transient overshoot of the intrave-
sicular glucose concentration over its equilibrium uptake in the
presence of sodium gradient and suggested that the BLMV
preparations were pure and without contamination of brush
border membrane. Under these conditions, the values of [14C]-
D-glucose uptake in control group were 35.29 6 2.20 and
36.20 6 3.20 pmol/30 sec/mg protein and in ethanol fed group
19.06 6 2.81 and 19.69 6 1.90 pmol/30 sec/mg protein,
respectively.
Transport of [3H]-Folic Acid
Uptake studies were performed at 37 8C using the incubation
buffer of 100 mM NaCl, 80 mM mannitol, 10 mM HEPES, 10
mM 3-[N-morpholino]ethanesulfonic acid (MES), pH 7.5 and
0.5 lM of [3H]-folic acid unless otherwise mentioned. Ten
microliter of vesicles (50 lg protein) were added to incubation
buffer containing [3H]-folic acid for fixed time interval. Reac-
tion was stopped by adding ice-cold stop solution followed by
rapid vacuum filtration. Non-specific binding to the filters was
determined by residual filter counts after filtration of the incuba-
tion buffer and labeled substrate without vesicles. The radioac-
tivity remained on the filters was determined by liquid scintilla-
tion counting (Beckman Coulter LS 6500). To determine the
binding component of folate uptake, vesicles were equilibrated
for 90 min in incubation buffers containing increasing concen-
trations of mannitol. Optimum conditions for maximum trans-
port were chosen for various experiments performed as
described (19). For the determination of the inhibition constants,
[3H]-folic acid at 0.5 and 1.0 lM concentrations were used in
the presence of range of concentrations of either methotrexate
or unlabeled folic acid viz 1, 2, and 3 lM.
RT-PCR Analysis
Total RNA was isolated from the jejunum following the
method of Chomeczynski and Sacchi (20). cDNA synthesis was
carried out from the purified and intact total RNA according to
manufacturer’s instructions. Expression of RFC and b-actin was
evaluated by PCR analysis using sequence specific primers cor-
responding to the sequence in the open reading frame. 20 lLPCR mixture was prepared in 1x PCR buffer consisting of
0.6 U of Taq polymerase, 2 lM of each primer both for b-actinand RFC along with 200 lM of each dNTP. In optimized PCR,
the initial denaturation step was carried out for 2 min @ 958C.The denaturation, annealing and elongation steps were carried
out respectively for 1 min @ 948C, 1 min @ 688C, 1 min @
728C for 35 cycles. The final extension step was carried out for
10 min @ 728C. The primers designed using Primer3 Input
(version 0.3.0) were RFC, Forward50GAACGTCCGGCAAC
CACAG30; Reverse 50GATGGACTTGGAGGCCCAG30 b-actin,
Forward50CACTGTGCCCATCTATGAGGG30; Reverse 50TCC
ACATCTGCTGGAAGGTGG30
Western Blot Analysis
For expression studies, isolated BLMV representing basolat-
eral membrane proteins (100–150 lg) were resolved on 10%
SDS-PAGE following the method of Laemmli (21) and trans-
ferred to nitrocellulose membrane for 4–5 h at 48C and the
transfer was carried out at 25V or 300 mA. Western blotting
was performed using the procedure described by Towbin et al.
(22) using polyclonal primary antibodies as rabbit anti-rat RFC
(1:500 dilution) kindly provided by Dr. Hamid M. Said, Profes-
sor, Physiology & Biophysics, School of Medicine, University
of California Irvine, USA and was raised against specific region
of rat RFC synthetic peptide corresponding to amino acids 495–
512 of the rat RFC. Secondary antibodies used were goat anti-
rabbit IgG HRP-labeled (1:2,000 dilutions).
Statistical Analysis
The data was computed as mean 6 SD. Group means were
compared by using the Student’s t-test, and analysis of variance
was used wherever necessary. The acceptable level of signifi-
cance was P\ 0.05 for each analysis. All RT-PCR and western
blot analyses were performed on 5 separate tissues from differ-
ent animals with comparable results. The densitometric analyses
of the products were determined using scion image software
(Scion Image Corporation, Frederick, MD).
RESULTS
Estimation of Blood Alcohol Levels
In order to reestablish the suitability of rat animal model for
the studies on experimental alcoholism under our experimental
setup, blood was drawn after 24 h of the last dose of ethanol to
determine blood alcohol levels. It was found that considerable
levels of alcohol (15.04 6 1.96 mg/dL) were maintained in
chronic ethanol fed group.
Kinetics of Folate Transport Across Intestinal BLMV
The carrier-mediated folic acid transport across the BLMV
was observed to be 55% less in ethanol fed rats after 3 months
of chronic ethanol ingestion (60.44 6 3.32 vs. 27.19 6 2.60
pmol/30 sec/mg protein). Folic acid uptake was studied at dif-
ferent time intervals from 20 sec to 5 min and uptake attained
maximum value at 60 sec and thereafter decreased abruptly in
control group and attained sharp peak at this time interval.
However, in case of ethanol fed group, although the maxima
obtained was at similar time point but the characteristic peak
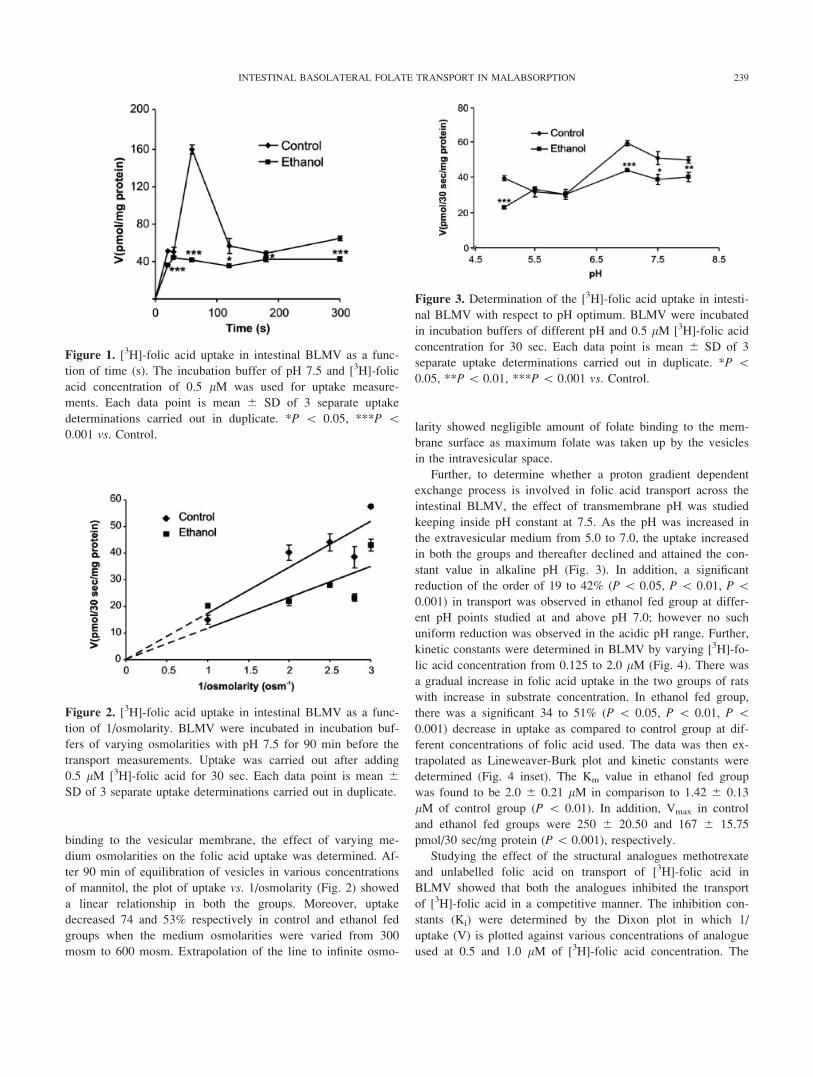
observed in control group was absent (Fig. 1). There was 13 to
73% (P\ 0.05, P\ 0.001) decrease in folate transport in etha-
nol fed group at various time intervals. To examine whether the
folic acid associated with the membranes was due to uptake
into the closed intravesicular space rather than a nonspecific
238 HAMID ET AL.
binding to the vesicular membrane, the effect of varying me-
dium osmolarities on the folic acid uptake was determined. Af-
ter 90 min of equilibration of vesicles in various concentrations
of mannitol, the plot of uptake vs. 1/osmolarity (Fig. 2) showed
a linear relationship in both the groups. Moreover, uptake
decreased 74 and 53% respectively in control and ethanol fed
groups when the medium osmolarities were varied from 300
mosm to 600 mosm. Extrapolation of the line to infinite osmo-
larity showed negligible amount of folate binding to the mem-
brane surface as maximum folate was taken up by the vesicles
in the intravesicular space.
Further, to determine whether a proton gradient dependent
exchange process is involved in folic acid transport across the
intestinal BLMV, the effect of transmembrane pH was studied
keeping inside pH constant at 7.5. As the pH was increased in
the extravesicular medium from 5.0 to 7.0, the uptake increased
in both the groups and thereafter declined and attained the con-
stant value in alkaline pH (Fig. 3). In addition, a significant
reduction of the order of 19 to 42% (P \ 0.05, P \ 0.01, P \0.001) in transport was observed in ethanol fed group at differ-
ent pH points studied at and above pH 7.0; however no such
uniform reduction was observed in the acidic pH range. Further,
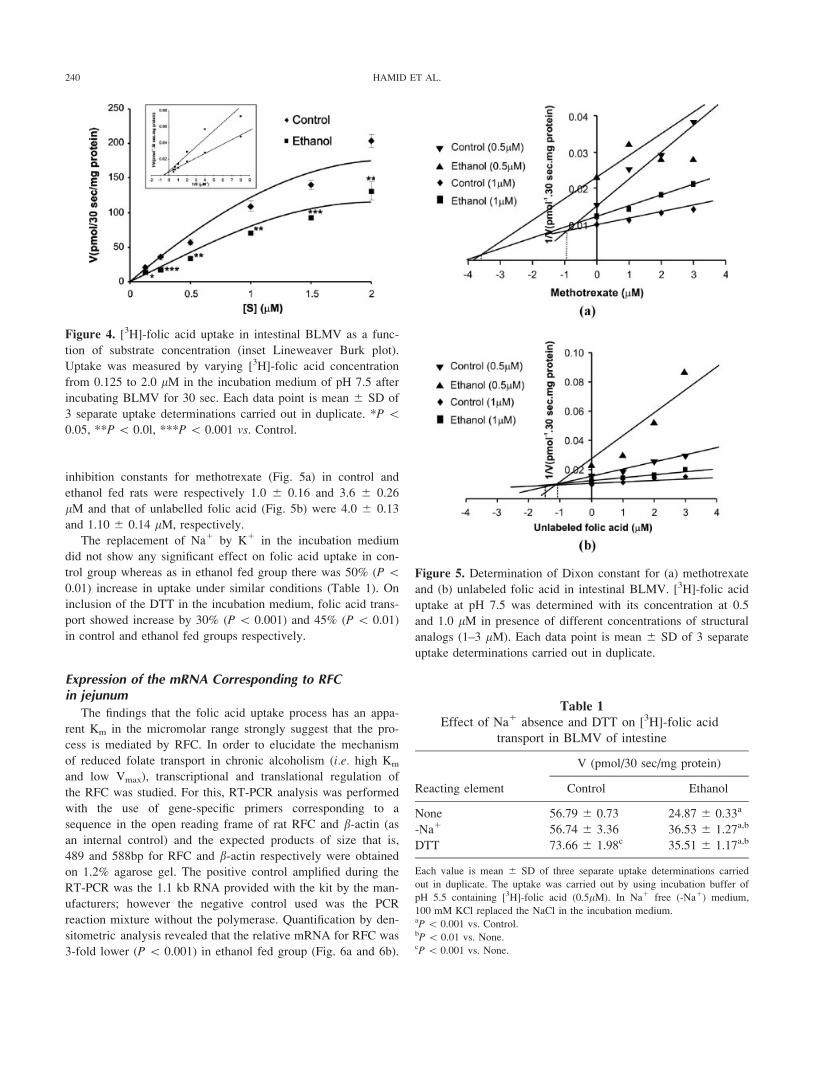
kinetic constants were determined in BLMV by varying [3H]-fo-
lic acid concentration from 0.125 to 2.0 lM (Fig. 4). There was
a gradual increase in folic acid uptake in the two groups of rats
with increase in substrate concentration. In ethanol fed group,
there was a significant 34 to 51% (P \ 0.05, P \ 0.01, P \0.001) decrease in uptake as compared to control group at dif-
ferent concentrations of folic acid used. The data was then ex-
trapolated as Lineweaver-Burk plot and kinetic constants were
determined (Fig. 4 inset). The Km value in ethanol fed group
was found to be 2.0 6 0.21 lM in comparison to 1.42 6 0.13
lM of control group (P \ 0.01). In addition, Vmax in control
and ethanol fed groups were 250 6 20.50 and 167 6 15.75
pmol/30 sec/mg protein (P\ 0.001), respectively.
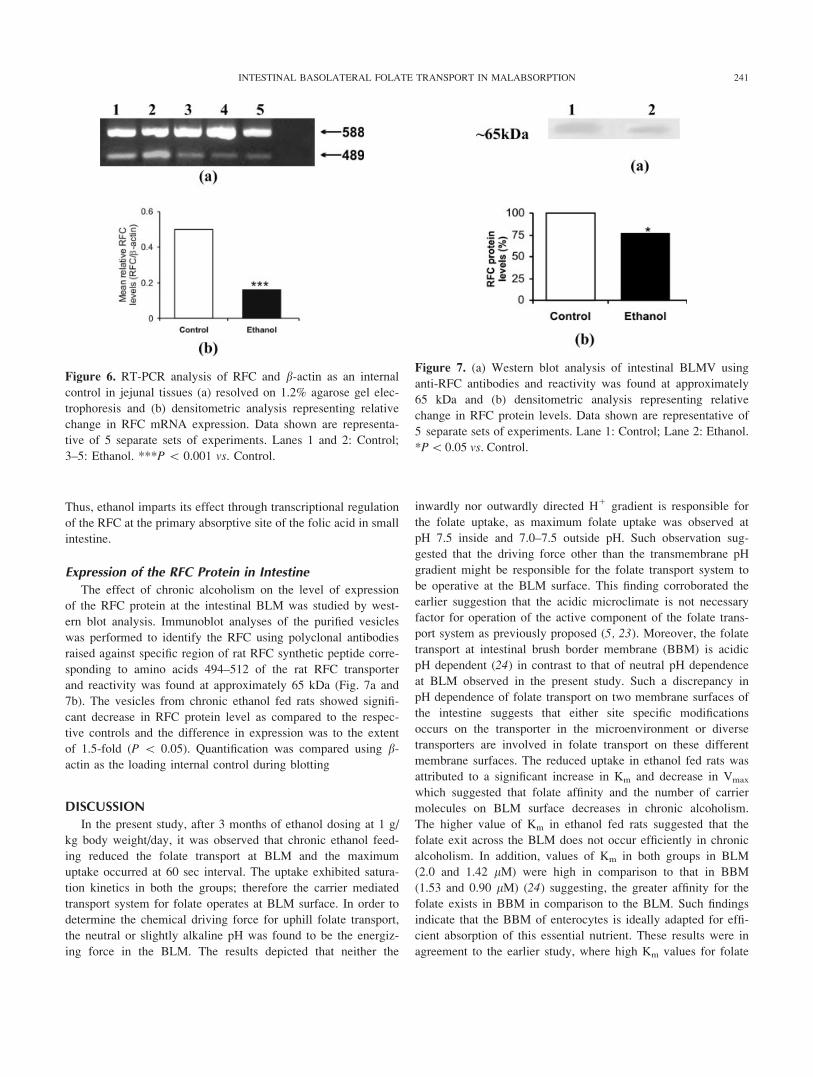
Studying the effect of the structural analogues methotrexate
and unlabelled folic acid on transport of [3H]-folic acid in
BLMV showed that both the analogues inhibited the transport
of [3H]-folic acid in a competitive manner. The inhibition con-
stants (Ki) were determined by the Dixon plot in which 1/
uptake (V) is plotted against various concentrations of analogue
used at 0.5 and 1.0 lM of [3H]-folic acid concentration. The
Figure 2. [3H]-folic acid uptake in intestinal BLMV as a func-
tion of 1/osmolarity. BLMV were incubated in incubation buf-
fers of varying osmolarities with pH 7.5 for 90 min before the
transport measurements. Uptake was carried out after adding
0.5 lM [3H]-folic acid for 30 sec. Each data point is mean 6
SD of 3 separate uptake determinations carried out in duplicate.
Figure 3. Determination of the [3H]-folic acid uptake in intesti-
nal BLMV with respect to pH optimum. BLMV were incubated
in incubation buffers of different pH and 0.5 lM [3H]-folic acid
concentration for 30 sec. Each data point is mean 6 SD of 3
separate uptake determinations carried out in duplicate. *P \0.05, **P\ 0.01, ***P\ 0.001 vs. Control.
Figure 1. [3H]-folic acid uptake in intestinal BLMV as a func-
tion of time (s). The incubation buffer of pH 7.5 and [3H]-folic
acid concentration of 0.5 lM was used for uptake measure-
ments. Each data point is mean 6 SD of 3 separate uptake
determinations carried out in duplicate. *P \ 0.05, ***P \0.001 vs. Control.
239INTESTINAL BASOLATERAL FOLATE TRANSPORT IN MALABSORPTION
inhibition constants for methotrexate (Fig. 5a) in control and
ethanol fed rats were respectively 1.0 6 0.16 and 3.6 6 0.26
lM and that of unlabelled folic acid (Fig. 5b) were 4.0 6 0.13
and 1.10 6 0.14 lM, respectively.
The replacement of Na1 by K1 in the incubation medium
did not show any significant effect on folic acid uptake in con-
trol group whereas as in ethanol fed group there was 50% (P \0.01) increase in uptake under similar conditions (Table 1). On
inclusion of the DTT in the incubation medium, folic acid trans-
port showed increase by 30% (P \ 0.001) and 45% (P \ 0.01)
in control and ethanol fed groups respectively.
Expression of the mRNA Corresponding to RFCin jejunum
The findings that the folic acid uptake process has an appa-
rent Km in the micromolar range strongly suggest that the pro-
cess is mediated by RFC. In order to elucidate the mechanism
of reduced folate transport in chronic alcoholism (i.e. high Km
and low Vmax), transcriptional and translational regulation of
the RFC was studied. For this, RT-PCR analysis was performed
with the use of gene-specific primers corresponding to a
sequence in the open reading frame of rat RFC and b-actin (as
an internal control) and the expected products of size that is,
489 and 588bp for RFC and b-actin respectively were obtained
on 1.2% agarose gel. The positive control amplified during the
RT-PCR was the 1.1 kb RNA provided with the kit by the man-
ufacturers; however the negative control used was the PCR
reaction mixture without the polymerase. Quantification by den-
sitometric analysis revealed that the relative mRNA for RFC was
3-fold lower (P \ 0.001) in ethanol fed group (Fig. 6a and 6b).
Figure 5. Determination of Dixon constant for (a) methotrexate
and (b) unlabeled folic acid in intestinal BLMV. [3H]-folic acid
uptake at pH 7.5 was determined with its concentration at 0.5
and 1.0 lM in presence of different concentrations of structural
analogs (1–3 lM). Each data point is mean 6 SD of 3 separate
uptake determinations carried out in duplicate.
Table 1
Effect of Na1 absence and DTT on [3H]-folic acid
transport in BLMV of intestine
Reacting element
V (pmol/30 sec/mg protein)
Control Ethanol
None 56.79 6 0.73 24.87 6 0.33a
-Na1 56.74 6 3.36 36.53 6 1.27a,b
DTT 73.66 6 1.98c 35.51 6 1.17a,b
Each value is mean 6 SD of three separate uptake determinations carried
out in duplicate. The uptake was carried out by using incubation buffer of
pH 5.5 containing [3H]-folic acid (0.5lM). In Na1 free (-Na1) medium,
100 mM KCl replaced the NaCl in the incubation medium.aP\ 0.001 vs. Control.bP\ 0.01 vs. None.cP\ 0.001 vs. None.
Figure 4. [3H]-folic acid uptake in intestinal BLMV as a func-
tion of substrate concentration (inset Lineweaver Burk plot).
Uptake was measured by varying [3H]-folic acid concentration
from 0.125 to 2.0 lM in the incubation medium of pH 7.5 after
incubating BLMV for 30 sec. Each data point is mean 6 SD of
3 separate uptake determinations carried out in duplicate. *P \0.05, **P\ 0.0l, ***P\ 0.001 vs. Control.
240 HAMID ET AL.
Thus, ethanol imparts its effect through transcriptional regulation
of the RFC at the primary absorptive site of the folic acid in small
intestine.
Expression of the RFC Protein in Intestine
The effect of chronic alcoholism on the level of expression
of the RFC protein at the intestinal BLM was studied by west-
ern blot analysis. Immunoblot analyses of the purified vesicles
was performed to identify the RFC using polyclonal antibodies
raised against specific region of rat RFC synthetic peptide corre-
sponding to amino acids 494–512 of the rat RFC transporter
and reactivity was found at approximately 65 kDa (Fig. 7a and
7b). The vesicles from chronic ethanol fed rats showed signifi-
cant decrease in RFC protein level as compared to the respec-
tive controls and the difference in expression was to the extent
of 1.5-fold (P \ 0.05). Quantification was compared using b-actin as the loading internal control during blotting
DISCUSSION
In the present study, after 3 months of ethanol dosing at 1 g/
kg body weight/day, it was observed that chronic ethanol feed-
ing reduced the folate transport at BLM and the maximum
uptake occurred at 60 sec interval. The uptake exhibited satura-
tion kinetics in both the groups; therefore the carrier mediated
transport system for folate operates at BLM surface. In order to
determine the chemical driving force for uphill folate transport,
the neutral or slightly alkaline pH was found to be the energiz-
ing force in the BLM. The results depicted that neither the
inwardly nor outwardly directed H1 gradient is responsible for
the folate uptake, as maximum folate uptake was observed at
pH 7.5 inside and 7.0–7.5 outside pH. Such observation sug-
gested that the driving force other than the transmembrane pH
gradient might be responsible for the folate transport system to
be operative at the BLM surface. This finding corroborated the
earlier suggestion that the acidic microclimate is not necessary
factor for operation of the active component of the folate trans-
port system as previously proposed (5, 23). Moreover, the folate
transport at intestinal brush border membrane (BBM) is acidic
pH dependent (24) in contrast to that of neutral pH dependence
at BLM observed in the present study. Such a discrepancy in
pH dependence of folate transport on two membrane surfaces of
the intestine suggests that either site specific modifications
occurs on the transporter in the microenvironment or diverse
transporters are involved in folate transport on these different
membrane surfaces. The reduced uptake in ethanol fed rats was
attributed to a significant increase in Km and decrease in Vmax
which suggested that folate affinity and the number of carrier
molecules on BLM surface decreases in chronic alcoholism.
The higher value of Km in ethanol fed rats suggested that the
folate exit across the BLM does not occur efficiently in chronic
alcoholism. In addition, values of Km in both groups in BLM
(2.0 and 1.42 lM) were high in comparison to that in BBM
(1.53 and 0.90 lM) (24) suggesting, the greater affinity for the
folate exists in BBM in comparison to the BLM. Such findings
indicate that the BBM of enterocytes is ideally adapted for effi-
cient absorption of this essential nutrient. These results were in
agreement to the earlier study, where high Km values for folate
Figure 6. RT-PCR analysis of RFC and b-actin as an internal
control in jejunal tissues (a) resolved on 1.2% agarose gel elec-
trophoresis and (b) densitometric analysis representing relative
change in RFC mRNA expression. Data shown are representa-
tive of 5 separate sets of experiments. Lanes 1 and 2: Control;
3–5: Ethanol. ***P\ 0.001 vs. Control.
Figure 7. (a) Western blot analysis of intestinal BLMV using
anti-RFC antibodies and reactivity was found at approximately
65 kDa and (b) densitometric analysis representing relative
change in RFC protein levels. Data shown are representative of
5 separate sets of experiments. Lane 1: Control; Lane 2: Ethanol.
*P\0.05 vs. Control.
241INTESTINAL BASOLATERAL FOLATE TRANSPORT IN MALABSORPTION

transport were observed in colonic BLM as compared to that in
luminal membrane (25). The osmolarity manipulation study
demonstrated that the folic acid uptake by intestinal BLM was
directed into the intravesicular space rather than towards extra-
vesicular binding. Such data demonstrated that the binding com-
ponent is not associated with the folate transport in BLM in
contrast to renal BBM where a distinctive binding process
involving folate binding protein has been suggested (19). The
values of the inhibition constants (Ki) in presence of the struc-
tural analogues under physiological conditions suggest the com-
mon uptake route for folic acid and its analogues in intestinal
BLM as Ki values were similar to the Km values of the folate.
A similar suggestion has also been made by earlier investigators
(26). However, in chronic alcoholism, the greater Ki values
observed in the present study, suggested the folate transport sys-
tem operates less efficiently for the analogs too upon chronic
alcohol ingestion and may propose the deranged antifolate bioa-
vailability may prevail more in alcoholics. Moreover, such find-
ings further corroborated the involvement of a carrier mediated
system for the transport of folic acid in BLM.
In line with the previous evidence (2), the folate uptake was
found to be Na1 and K1 independent under the physiological
conditions; however under chronic alcoholic conditions, the pro-
cess seemed to be dependent on the presence of K1 at the
absorptive surface. Such an observation suggests that the alco-
hol ingestion may alter the presence of these ions at the folate
exit site. Additionally, it remains to be determined whether, the
carrier mediated folate transport is facilitated at BLM surface,
because maximum transport was observed at neutral or slightly
alkaline pH and the physiological system was found to be Na1
and K1 independent. The folate uptake system at the basolateral
surface showed enhanced transport in the presence of DTT
which might be attributed to increase in –SH group (s) on the
transporter. The more increase in uptake in ethanol fed group in
presence of DTT suggests that the higher number of accessible
–SH group(s) on the transporter which inturn proposes that the
conformation of the carrier at the folate exit site on BLM sur-
face might be altered during alcoholism.
Importantly, the decreased Vmax in ethanol fed rats could
relate to the reduced number of RFC molecules on BLM sur-
face. In this context, the decrease in RFC mRNA and protein
expression during alcoholism observed in the present study can
be explained by the lesser stability of RFC mRNA or enhanced
degradation of the RFC protein molecules at BLM surface dur-
ing alcoholism. In addition, substantial evidence from studies
in micropigs suggests that chronic exposure to ethanol at
intoxicating serum levels alters RFC expression in jejunum
(27). Assuming that the RFC message level is a reflection of
functional uptake across intestinal BLM surface, our results
suggest that decreased RFC expression concomitantly with
observed decrease in transport efficiency in BLM surface as a
possible reason of lower blood folate levels commonly found
in chronic alcoholics. Importantly, after chronic ethanol feed-
ing, the decreased expression level of the RFC seems to be
regulated at BLM surface by transcriptional and/or transla-
tional machinery. Future work should focus on the mechanism
that underlies down-regulation of RFC during alcoholic condi-
tions. In addition, recently identified proton coupled folate
transporter (PCFT) has also been found important for folate
transport in diverse tissues including intestine. However, role
of PCFT as folate transporter at intestinal basolateral surface
is questioning because of its acidic pH dependence in contrast
to neutral or slightly alkaline pH dependence of folate trans-
port at BLM.
Taken together, these results suggests that the folate trans-
port at the intestinal BLM to be carrier mediated, saturable, pH
dependent but Na1 independent process. Chronic ethanol inges-
tion reduces the folate exit activity across the BLM by a mecha-
nism involving decreased affinity of the RFC to the folate and
reduced number of its molecules on membrane surface as
revealed by the kinetic and expression studies respectively. In
addition, alcohol ingestion shifted the process to K1 dependent
besides affecting the –SH status of the transport system.
ACKNOWLEDGEMENTS
This work was supported by the financial grant sanctioned to Dr.
Jyotdeep Kaur from Indian Council of Medical Research, New
Delhi, India. Abid Hamid was recipient of fellowship from the
Council of Scientific and Industrial Research, New Delhi, India.
REFERENCES1. Sabharanjak, S. and Mayor, S. (2004) Folate receptor endocytosis and
trafficking. Adv. Drug. Deliv. Rev. 56, 1099–1109.
2. Said H. M. and Redha, R. (1987) A carrier-mediated transport for folate
in basolateral membrane vesicles of rat small intestine. Biochem. J. 247,
141–146.
3. Sirotnak, F. M. and Tolner, B. (1999) Carrier-mediated membrane trans-
port of folates in mammalian cells. Annu. Rev. Nutr. 19, 91–122.
4. Zhao, R. and Goldman, I. D. (2003) Resistance to antifolates. Onco-
gene. 22, 7431–7457.5. Kumar, C. K., Nguyen, T. T., Gonzales, F. B., and Said, H. M. (1998)
Comparison of intestinal folate carrier clone expressed in IEC- 6 cells
and in Xenopus oocytes. Am. J. Physiol. Cell. Physiol. 274, C289–
C294.
6. Mason, J. B. and Choi, S. W. (2005) Effects of alcohol on folate metab-
olism: implications for carcinogenesis. Alcohol. 35, 235–241.
7. Nguyen, T. T., Dyer, D. L., Dunning, D. D., Rubin, S. A., Grant, K. E.,
and Said, H. M. (1997) Human intestinal folate transport: cloning,
expression, and distribution of complementary RNA. Gastroenterol.
112, 783–791.
8. Rajgopal, A., Sierra, E. E., Zhao, R., and Goldman, I. D. (2001) Expres-
sion of the reduced folate carrier SLC19A1 in IEC-6 cells results in two
distinct transport activities. Am. J Physiol. Cell. Physiol. 281, C1579–
C1586.
9. Poschl, G. and Seitz, H. K. (2004) Alcohol and cancer. Alcohol. Alco-
holism. 39, 155–165.
10. Salaspuro, M. P. (2003) Alcohol consumption and cancer of the gastro-
intestinal tract. Best. Prac. Res. Clin. Gastroenterol. 17, 679–694.11. Halsted, C. H., Villanueva, J. A., Devlin, A. M., Niemela, O., Parkkila,
S., Garrow, T. A., Wallock, L. M., Shigenaga., M. K., Melnyk, S., and
James, S. J. (2002) Folate deficiency disturbs hepatic methionine
242 HAMID ET AL.

metabolism and promotes liver injury in the ethanol-fed micropig. Proc.Natl. Acad. Sci. USA 99, 10072–10077.
12. de Marco, P., Calevo, M. G., Moroni, A., Merello, E., Raso, A., Finnell,
R. H., Zhu, H., Andreussi L., Cama A., and Capra V. (2003) Reduced
folate carrier polymorphism (80A?G) and neural tube defects. Eur. J
Hum. Genet. 11, 245–252.
13. Hamid, A., Wani, N. A., Rana, S., Vaiphei, K., Mahmood, A., and
Kaur, J. (2007). Down-regulation of reduced folate carrier may result in
folate malabsorption across intestinal brush border membrane during ex-
perimental alcoholism. FEBS J. 274, 6317–6328.
14. Burtis, C. A. and Ashwood, E. R. (1994) Tietz Textbook of Clinical
Chemistry, 2nd edn. p. 1170, WB Saunders, Philadelphia.
15. Scalera, V., Storelli, C., Storelli-Joss, C., Haase, W., and Murer, H. (1980)
A simple and fast method for the isolation of basolateral membrane from rat
small-intestinal epithelial cells. Biochem. J. 186, 177–181.16. Bergmeyer, H. U. (1974) Alkaline phosphatase from calf intestine. In
Methods of Enzymatic Analysis, 2nd edn. (Bergmeyer, H. U. ed.) pp.
496–498, Academic Press, New York.
17. Quigley, J. P. and Gotterer, G. S. (1969) Distribution of Na1-K1 stimulated
ATPase in rat intestinal mucosa. Biochim. Biophys. Acta. 173, 456–468.
18. Lowry, O. H., Rosenbergh, N. J., Farr, A. L., and Randall, R. J. (1951)
Protein measurement with the folin phenol reagent. J. Biol. Chem. 193,
265–275.
19. Hamid, A. and Kaur, J. (2006) Chronic alcoholism alters the transport
characteristics of folate in rat renal brush border membrane. Alcohol.
38, 59–66.
20. Chomeczynski, P. and Sacchi, N. (1987) Single-step method of RNA
isolation by acid guanidinium thiocyanate-phenol-chloroform extraction.
Anal. Biochem. 162, 156–159.
21. Laemmli, U. K. (1970) Cleavage of structural proteins during the as-
sembly of the head of bacteriophage T4. Nature. 227, 680–685.
22. Towbin, H., Staehelin, T., and Gordon, J. (1979) Electrophoretic trans-
fer of proteins from polyacrylamide gels to nitrocellulose sheets: proce-
dure and some applications. Proc. Natl. Acad. Sci. USA 76, 4350–4354.
23. Mason, J. B. and Rosenberg, I. H. (1994) Intestinal absorption of folate
In Physiology of the Gastrointestinal Tract, 3rd edn. (Johnson, L. R.).
pp. 979–995, Raven Press, New York.
24. Hamid, A., Kaur, J., and Mahmood, A. (2007). Evaluation of the kinetic
properties of the folate transport system in intestinal absorptive epithe-
lium during experimental ethanol ingestion. Mol. Cell. Biochem. 304,
265–271.
25. Dudeja, P. K., Kode, A., Alnounou, M., Tyagi, S., Torania, S., Subra-
manian, V. S., and Said, H. M. (2001) Mechanism of folate transport
across the human colonic basolateral membrane. Am. J Physiol. Gastro-
intest. Liver. Physiol. 281, G54–G60.26. Montalar, M., Nalda-Molina, R., Rodriguez-Ibanez, M., Garcia-Valcar-
cel, I., Garrigues, T. M., Merino, V., and Bermejo, M. (2003) Kinetic
modeling of triamterene intestinal absorption and its inhibition by folic
acid and methotrexate. J. Drug. Target. 11, 215–223.27. Villanueva, J. A., Devlin, A. M., and Halsted, C. H. (2001) Reduced fo-
late carrier: tissue distribution and effects of chronic ethanol intake in
the micropig. Alcohol. Clin. Exp. Res. 25, 415–420.
243INTESTINAL BASOLATERAL FOLATE TRANSPORT IN MALABSORPTION