Embed Size (px)
Citation preview

Proc. Nat!. Acad. Sci. USAVol. 88, pp. 7106-7110, August 1991Cell Biology
The low-affinity p75 nerve growth factor (NGF) receptor mediatesNGF-induced tyrosine phosphorylation
(signal transduction)
MARGARET M. BERG*, DAVID W. STERNBERGt, BARBARA L. HEMPSTEADt, AND MOSES V. CHAO*t*Department of Cell Biology and Anatomy, and tDivision of Hematology/Oncology, Department of Medicine, Cornell University Medical College, New York,NY 10021; and tLaboratory of Molecular Oncology, Rockefeller University, New York, NY 10021
Communicated by Bernard L. Horecker, May 6, 1991
ABSTRACT Protein tyrosine phosphorylation is a potentialmechanism for initial sinaling in PC12 cells during dfferenti-ation in response to nerve growth factor (NGF). NGF-inducedtyrosine phosphorylation has been found to be initiated by the trkprotooncogene, which participates in the formation of high-affinity NGF binding sites. In contrast to transfection of wild-type low-affinity p75 NGF receptors, transfection of p75NGmwith mutations in the cytoplasmic domain resulted in an inabilityof NGF to elicit tyrosine phosphorylation of intracellular sub-strates, indicating that p75NG is involved in initiating pbos-phorylation events by NGF. Even though the p75NGFR receptordoes not possess any inherent tyrosne kinase activity, theseexperiments demonstrate that the p75NGFR has a potential rolein NGF-induced tyrosine phosphorylation.
Protein tyrosine phosphorylation is initiated by growth factorbinding to receptors for insulin, epidermal growth factor(EGF), platelet-derived growth factor (PDGF), and fibroblastgrowth factor (FGF), all of which possess intrinsic tyrosinekinase activity (1). Nerve growth factor (NGF) interacts witha 75-kDa receptor (2) that lacks a cytoplasmic tyrosine kinasedomain (3, 4), yet rapid and transient protein tyrosine phos-phorylation has been observed upon treatment of PC12 cellswith NGF (5-8). The kinase responsible for this activity hasbeen identified as the trk protooncogene, a receptor tyrosinekinase that binds NGF and is activated in PC12 cells,neuroblastoma cells, and embryonic sensory neurons (8, 9,38).NGF initiates the differentiation and survival of selective
neuronal populations by binding to cell surface receptors. Inresponsive cells, NGF receptors can be divided into twoclasses differing 100-fold in their rates of dissociation ofNGF. The high-affinity form of the receptor, representinglop0 ofthe total number ofbinding sites on responsive cells,binds NGF with a Kd of 10-11 M and mediates the biologicaleffects of NGF on target cells. Gene transfer experimentswith the cloned human cDNA (10-12) demonstrated that thelow-affinity receptor (Kd = 10-9 M), represented by thep75NGFR receptor protein, is involved in the formation of thehigh-affinity sites. Affinity crosslinking studies using "'I-labeled NGF (1251I-NGF) binding in responsive cells sug-gested that an additional protein was involved in the forma-tion of the high-afflinity NGF receptor complex (13-17). Theproduct of the trk tyrosine kinase has been identified as alow-affinity NGF receptor that participates with p75NGFR toform high-affinity NGF receptors (9, 17).
Since p75NGF has been shown to be required for high-affinity NGF binding (10, 17), we have tested whetherexpression of the p75 low-affinity NGF receptor is requiredfor activation ofprotein tyrosine phosphorylation in responseto NGF. Our results indicate that appropriate NGF-induced
protein tyrosine phosphorylation requires expression ofp75NGFR and that a highly conserved cytoplasmic domain ofthis receptor is crucial for this response.
MATERIALS AND METHODSCell Culture. PC12 and NR18 cells were maintained in
Dulbecco modified Eagle medium (DMEM) with 10%o fetalbovine serum and 5% horse serum (GIBCO). A875 humanmelanoma cells were maintained in DMEM with 10%6 fetalbovine serum.
Constructs. Construction of the deletion mutants of theNGF receptor has been described (15). All cDNAs weresubcdoned into the EcoRI site of the pMV7 expression vectorand recombinant retroviruses produced as described (18).
Retroviral Gene Transfer. Cell lines expressing the wild-type and mutant NGF receptor cDNAs were generated afterretroviral infection of helper-free virus stocks. NR18 trans-fectants were isolated after neomycin (0.5 mg ofG418 per ml;GIBCO) selection and purification by rosetting with themonoclonal antibody ME20.4 and rabbit anti-mouse IgG-coupled human erythrocytes (2, 3, 15).
Immunoblotting. Cells were grown to near confluence on60-mm tissue culture dishes and were serum starved for 1 hrby incubation in serum-free DMEM. The medium was thenreplaced with DMEM containing 50 ng of NGF per ml or 20ng ofEGF per ml (Bioproducts for Science, Indianapolis). Atthe indicated times the cells were rapidly rinsed with ice-coldTris-buffered saline, solubilized in 120 ,l of sol buffer [10mMTris hydrochloride, pH 7.4/1% sodium dodecyl sulfate(SDS)/1 mM sodium orthovanadate/0.1 mM sodium molyb-date/1 mM phenylmethylsulfonyl fluoride/5 ,ug of aprotininper ml/5 ,ug of leupeptin per ml], and immediately frozen indry ice and ethanol. The samples were then boiled for 5 min.Protein concentrations were determined with a protein assaykit (Bio-Rad). Proteins (150 ,ug) were separated by SDS/PAGE on 6-13% linear gradient polyacrylamide gels (unlessotherwise noted) and electrophoretically transferred to ni-trocellulose filters as described (19). The filters were incu-bated with rabbit anti-phosphotyrosine antisera preparedagainst bacteria expressing v-abl and purified by affinitychromatography as described (19). Labeled proteins werevisualized by incubating the filters with 125I-labeled protein A(Amersham) followed by autoradiography on x-ray film withan intensifying screen for 3-5 days, unless otherwise noted.
RESULTSHigh-Affinity NGF Receptors Are Required for Induction of
Protein Tyrosine Phosphorylation. Treatment of PC12 cellswith NGF induces the rapid and transient phosphorylation ofseveral distinct proteins on tyrosine residues (5). Since all
Abbreviations: NGF, nerve growth factor; EGF, epidermal growthfactor.
7106
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 88 (1991) 7107
known biological responses to NGF require interaction withhigh-affinity NGF receptors (20, 21), we compared the pat-tern of induced protein phosphorylation in cell lines thatdisplay different receptor affinities. Two cell lines that ex-press only low-affinity receptors, A875 human melanomacells and the PC12 mutant line PC12nnr5 (21), were comparedto native PC12 cells, which express low- and high-affinityreceptors.Each cell line was treated with 50 ng of NGF per ml for
varying times, and lysates were prepared and probed with ananti-phosphotyrosine antibody by Western blotting. Within afew minutes ofNGF treatment ofPC12 cells, a rapid increasein phosphotyrosine content in proteins with apparent molec-ular masses of 114, 99, 42, and 39 kDa was observed. Inunstimulated cells, a major tyrosine-phosphorylated proteinof 130 kDa was present, and levels of phosphorylation of thisprotein did not change dramatically with NGF treatment (Fig.1).The time course of phosphorylation for each protein was
distinctive and reproducible. Phosphorylation of a 99-kDaprotein occurred within 30 sec after NGF treatment (data notshown), peaked at 1-5 min, and was undetectable at 60 min.Phosphorylation of the 114-, 42-, and 39-kDa proteins wasfirst detected at 1 min after stimulation and was found atreduced levels 60 min after the addition of NGF. After 4 hrofNGF treatment, levels of protein tyrosine phosphorylationreturned to the level observed in untreated cells (data notshown). The differences in molecular masses of the phos-photyrosine proteins induced by NGF described here andthose reported by Maher (5, 6) may be due to differences inspecificity ofthe anti-phosphotyrosine antibodies or to clonalvariance in the PC12 cells used. As a positive control, lysatesfrom rat 3Y1 cells transformed with the v-src oncogene ofRous sarcoma virus (SR3Y1) were included in Fig. 1 (19).When 2.5 mM phosphotyrosine was added to the antiserumbefore probing blots of NGF-treated PC12 cell lysates, nolabeled proteins were detected (Fig. 1A). When phospho-seine or phosphothreonine was incubated, no competitionwas observed (data not shown), demonstrating that theantibodies specifically recognize phosphotyrosine.When A875 human melanoma cells (22, 23), which express
only the p75 NGF receptor, were treated with NGF in asimilar manner, no changes in protein tyrosine phosphory-lation were observed (Fig. 1A). Similarly, as shown in Fig.lB, NGF does not induce any change in levels of tyrosinephosphorylation in the PC12nnr5 cell line (NNR) (21) or in the
APC12
PC12 A875 C +ptyr_ _ 0 I al: r F -i
0 1 5 60 0 1 5 60 () 0 1 5220-
109-
72-
46-
-
p75NGFR-negative cell line, NR18 (24). The PC12nnr5 and theNR18 cell lines have been shown to be unresponsive to NGF(21, 24). The trk tyrosine kinase is expressed in extremely lowlevels in NR18 cells (17) and PC12nnr5 cells (L. Greene,personal communication) but is not present in A875 cells (9).These data are consistent with a model in which NGFinteraction with the high-affinity NGF receptor results in therapid activation of a protein tyrosine kinase and mere expres-sion of p75NGFR low-affinity receptors alone in most celltypes is not sufficient to stimulate protein tyrosine phosphor-ylation by NGF.p75NGFR Receptor Mutants and Tyrosine Phosphorylation.
Deletion of the cytoplasmic domain of p75NGFR resulted inthe loss of high-affinity '25I-NGF binding in NR18 cells (15).These mutant receptors lacked portions of the highly con-served region surrounding the membrane spanning domainand were introduced into NR18 by retrovirally mediated genetransfer. All mutant receptors were found to be expressedappropriately at the cell surface offibroblasts and NR18 cells(10, 15) and bound 125I-NGF in both cell types with a similarKd of 10-9 M. In contrast, the wild-type p75NGFR intro-duced into NR18 cells displayed high- and low-affinity bind-ing, but displayed only low-affinity binding when introducedinto fibroblast cells (10).NGF-induced protein tyrosine phosphorylation was tested
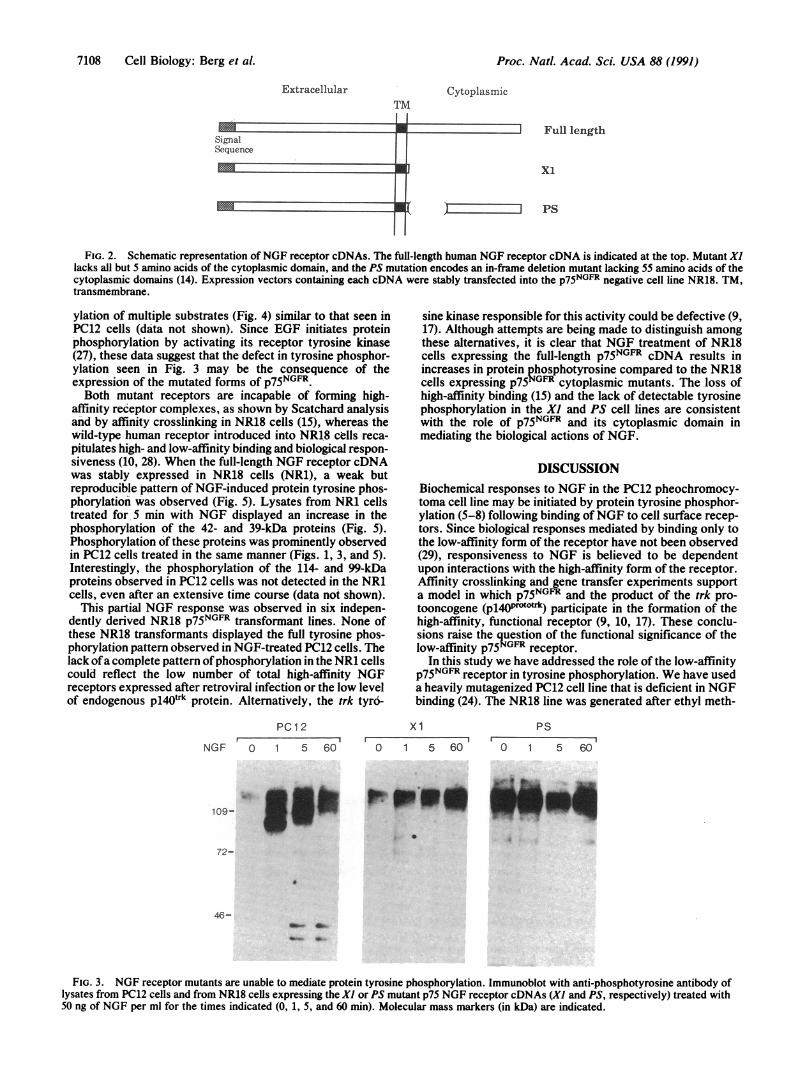
with the two different receptor mutants. Mutant Xl encodesaNGF receptor lacking the entire cytoplasmic domain exceptfor 5 amino acids. Mutation PS encodes an in-frame deletionof 55 amino acids in the highly conserved juxtamembraneregion of the cytoplasmic domain (Fig. 2). The mutantreceptor cDNAs were subcloned in pMV7 (25), transfectedinto the PA317 recipient cell line (26), and used to generaterecombinant retrovirus. After infection of NR18 cells andselection in neomycin, individual cell lines were isolated andanalyzed for phosphotyrosine responses.No changes in protein tyrosine phosphorylation were de-
tected upon NGF treatment in NR18 cells expressing eitherreceptor containing mutant cytoplasmic domains (Fig. 3).Slight changes in intensity of the high molecular mass pro-teins were observed but were not reproducibly increased byNGF treatment. These cells were assayed during the sametime period as in Fig. 1. The lack of phosphotyrosine re-sponse in these cells was not the consequence of G418selection or a defect in the ability of the cell to displaytyrosine phosphorylation. EGF treatment ofthese cells underthe sane conditions rapidly induced the tyrosine phosphor-
PC 12 N N R NR18 >O
0 1 5 60 0 1 5 60 0 1 5 0CIu
_R
46-
28-
FIG. 1. NGF-induced protein tyrosine phosphorylation requires expression of the high-affinity NGF receptor. (A) Immunoblot withanti-phosphotyrosine antibody of lysates from PC12 cells and A875 human melanoma cells treated with 50 ng of NGF per ml for 0, 1, 5, and60 min. Lysates from rat 3Y1 cells transformed with v-src oncogene (SR3Y1) were used as a positive control. The specificity of the antibodiesis shown at right in which the blot was incubated with anti-phosphotyrosine antibodies in the presence of 2.5 mM phosphotyrosine. Molecularmass markers (in kDa) are indicated. (B) Immunoblot as in A with lysates from the PC12 cell variant cell lines NNR and 9NR. Whole cell lysateswere run on 8% polyacrylamide gels and transferred to nitrocellulose as described in text.
Cell Biology: Berg et al.
Proc. Natl. Acad. Sci. USA 88 (1991)
ExtracellularTM
Cytoplasmic
Full length
xi
PS
FIG. 2. Schematic representation of NGF receptor cDNAs. The full-length human NGF receptor cDNA is indicated at the top. Mutant Xllacks all but 5 amino acids of the cytoplasmic domain, and the PS mutation encodes an in-frame deletion mutant lacking 55 amino acids of thecytoplasmic domains (14). Expression vectors containing each cDNA were stably transfected into the p75NGFR negative cell line NR18. TM,transmembrane.
ylation of multiple substrates (Fig. 4) similar to that seen inPC12 cells (data not shown). Since EGF initiates proteinphosphorylation by activating its receptor tyrosine kinase(27), these data suggest that the defect in tyrosine phosphor-ylation seen in Fig. 3 may be the consequence of theexpression of the mutated forms of p75NGFR*Both mutant receptors are incapable of forming high-
affinity receptor complexes, as shown by Scatchard analysisand by affinity crosslinking in NR18 cells (15), whereas thewild-type human receptor introduced into NR18 cells reca-pitulates high- and low-affinity binding and biological respon-siveness (10, 28). When the full-length NGF receptor cDNAwas stably expressed in NR18 cells (NR1), a weak butreproducible pattern of NGF-induced protein tyrosine phos-phorylation was observed (Fig. 5). Lysates from NR1 cellstreated for 5 min with NGF displayed an increase in thephosphorylation of the 42- and 39-kDa proteins (Fig. 5).Phosphorylation of these proteins was prominently observedin PC12 cells treated in the same manner (Figs. 1, 3, and 5).Interestingly, the phosphorylation of the 114- and 99-kDaproteins observed in PC12 cells was not detected in the NR1cells, even after an extensive time course (data not shown).
This partial NGF response was observed in six indepen-dently derived NR18 p75NGIF transformant lines. None ofthese NR18 transformants displayed the full tyrosine phos-phorylation pattern observed in NGF-treated PC12 cells. Thelack ofa complete pattern ofphosphorylation in the NR1 cellscould reflect the low number of total high-affinity NGFreceptors expressed after retroviral infection or the low levelof endogenous p140trk protein. Alternatively, the trk tyro-
sine kinase responsible for this activity could be defective (9,17). Although attempts are being made to distinguish amongthese alternatives, it is clear that NGF treatment of NR18cells expressing the full-length p75NGFR cDNA results inincreases in protein phosphotyrosine compared to the NR18cells expressing p75NGFR cytoplasmic mutants. The loss ofhigh-affinity binding (15) and the lack of detectable tyrosinephosphorylation in the Xl and PS cell lines are consistentwith the role of p75NGFR and its cytoplasmic domain inmediating the biological actions of NGF.
DISCUSSIONBiochemical responses to NGF in the PC12 pheochromocy-toma cell line may be initiated by protein tyrosine phosphor-ylation (5-8) following binding ofNGF to cell surface recep-tors. Since biological responses mediated by binding only tothe low-affinity form of the receptor have not been observed(29), responsiveness to NGF is believed to be dependentupon interactions with the high-affinity form of the receptor.Affinity crosslinking and gene transfer experiments supporta model in which p75NGFR and the product of the trk pro-tooncogene (pl40Pr°'0'rk) participate in the formation of thehigh-affinity, functional receptor (9, 10, 17). These conclu-sions raise the question of the functional significance of thelow-affinity p75NGFR receptor.
In this study we have addressed the role of the low-affinityp75NGFR receptor in tyrosine phosphorylation. We have useda heavily mutagenized PC12 cell line that is deficient in NGFbinding (24). The NR18 line was generated after ethyl meth-
PC12
NGF 0 1 5 60
xi1 5 60
0 1 5 60
PS
0 1 5 60
109-
72-
46-
FIG. 3. NGF receptor mutants are unable to mediate protein tyrosine phosphorylation. Immunoblot with anti-phosphotyrosine antibody oflysates from PC12 cells and from NR18 cells expressing the Xl or PS mutant p75 NGF receptor cDNAs (Xl and PS, respectively) treated with50 ng of NGF per ml for the times indicated (0, 1, 5, and 60 min). Molecular mass markers (in kDa) are indicated.
7108 Cell Biology: Berg et al.
Proc. Natl. Acad. Sci. USA 88 (1991) 7109
xi PS
EGF '0 1 5 60 0 1 5 60
220-
109 -
72-
46-
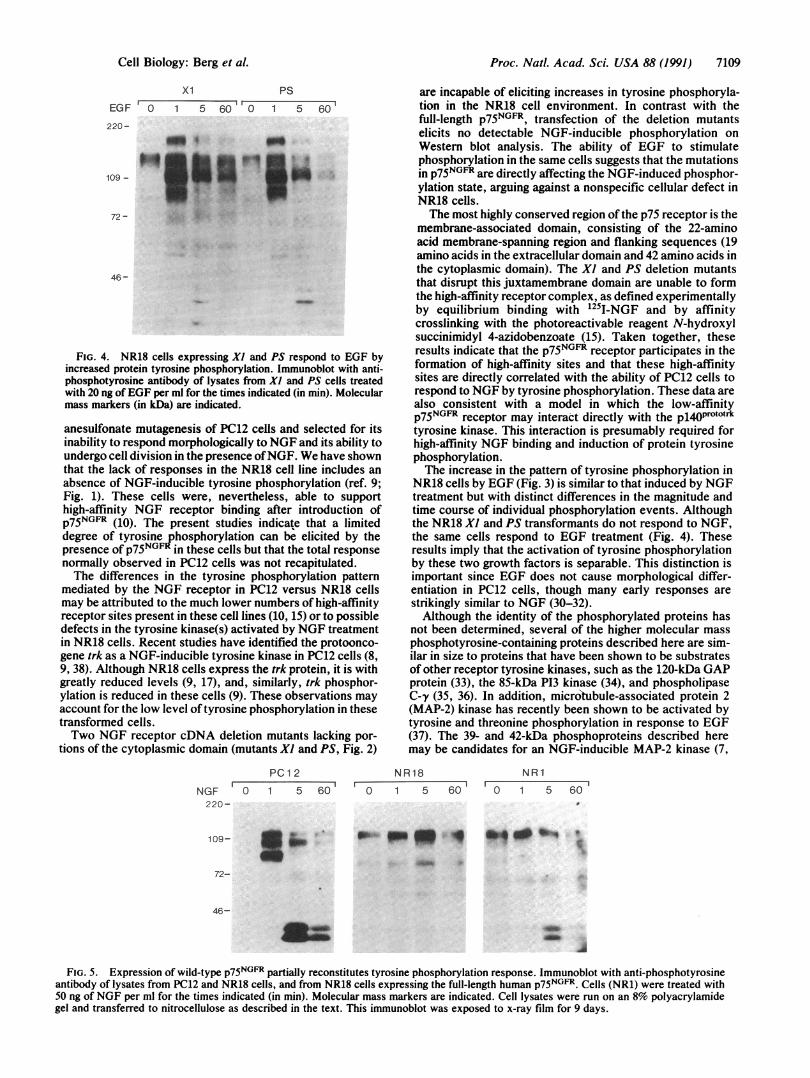
FIG. 4. NR18 cells expressing Xl and PS respond to EGF byincreased protein tyrosine phosphorylation. Immunoblot with anti-phosphotyrosine antibody of lysates from Xl and PS cells treatedwith 20 ng of EGF per ml for the times indicated (in min). Molecularmass markers (in kDa) are indicated.
anesulfonate mutagenesis of PC12 cells and selected for itsinability to respond morphologically to NGF and its ability toundergo cell division in the presence ofNGF. We have shownthat the lack of responses in the NR18 cell line includes anabsence of NGF-inducible tyrosine phosphorylation (ref. 9;Fig. 1). These cells were, nevertheless, able to supporthigh-affinity NGF receptor binding after introduction ofp75NGFR (10). The present studies indicate that a limiteddegree of tyrosine phosphorylation can be elicited by thepresence of p75NGFR in these cells but that the total responsenormally observed in PC12 cells was not recapitulated.The differences in the tyrosine phosphorylation pattern
mediated by the NGF receptor in PC12 versus NR18 cellsmay be attributed to the much lower numbers of high-affinityreceptor sites present in these cell lines (10, 15) or to possibledefects in the tyrosine kinase(s) activated by NGF treatmentin NR18 cells. Recent studies have identified the protoonco-gene trk as a NGF-inducible tyrosine kinase in PC12 cells (8,9, 38). Although NR18 cells express the trk protein, it is withgreatly reduced levels (9, 17), and, similarly, trk phosphor-ylation is reduced in these cells (9). These observations mayaccount for the low level of tyrosine phosphorylation in thesetransformed cells.Two NGF receptor cDNA deletion mutants lacking por-
tions of the cytoplasmic domain (mutants Xl and PS, Fig. 2)
PC1 2
NGF 0 1 5 60220-
are incapable of eliciting increases in tyrosine phosphoryla-tion in the NR18 cell environment. In contrast with thefull-length p75NGFR, transfection of the deletion mutantselicits no detectable NGF-inducible phosphorylation onWestern blot analysis. The ability of EGF to stimulatephosphorylation in the same cells suggests that the mutationsin p75NGFR are directly affecting the NGF-induced phosphor-ylation state, arguing against a nonspecific cellular defect inNR18 cells.The most highly conserved region of the p75 receptor is the
membrane-associated domain, consisting of the 22-aminoacid membrane-spanning region and flanking sequences (19amino acids in the extracellular domain and 42 amino acids inthe cytoplasmic domain). The Xl and PS deletion mutantsthat disrupt this juxtamembrane domain are unable to formthe high-affinity receptor complex, as defined experimentallyby equilibrium binding with 125I-NGF and by affinitycrosslinking with the photoreactivable reagent N-hydroxylsuccinimidyl 4-azidobenzoate (15). Taken together, theseresults indicate that the p75NGFR receptor participates in theformation of high-affinity sites and that these high-affinitysites are directly correlated with the ability of PC12 cells torespond to NGF by tyrosine phosphorylation. These data arealso consistent with a model in which the low-affinityp75NGFR receptor may interact directly with the pl4OPrototrktyrosine kinase. This interaction is presumably required forhigh-affinity NGF binding and induction of protein tyrosinephosphorylation.The increase in the pattern of tyrosine phosphorylation in
NR18 cells by EGF (Fig. 3) is similar to that induced by NGFtreatment but with distinct differences in the magnitude andtime course of individual phosphorylation events. Althoughthe NR18 Xl and PS transformants do not respond to NGF,the same cells respond to EGF treatment (Fig. 4). Theseresults imply that the activation of tyrosine phosphorylationby these two growth factors is separable. This distinction isimportant since EGF does not cause morphological differ-entiation in PC12 cells, though many early responses arestrikingly similar to NGF (30-32).Although the identity of the phosphorylated proteins has
not been determined, several of the higher molecular massphosphotyrosine-containing proteins described here are sim-ilar in size to proteins that have been shown to be substratesof other receptor tyrosine kinases, such as the 120-kDa GAPprotein (33), the 85-kDa P13 kinase (34), and phospholipaseC-y (35, 36). In addition, microtubule-associated protein 2(MAP-2) kinase has recently been shown to be activated bytyrosine and threonine phosphorylation in response to EGF(37). The 39- and 42-kDa phosphoproteins described heremay be candidates for an NGF-inducible MAP-2 kinase (7,
NR18
0 1 5
NR1
wY1 ,0 1 5 60'
-w q a"" W4109- 3b
72-
46-
9
FIG. 5. Expression of wild-type p75NGFR partially reconstitutes tyrosine phosphorylation response. Immunoblot with anti-phosphotyrosineantibody of lysates from PC12 and NR18 cells, and from NR18 cells expressing the full-length human p75NGFR. Cells (NR1) were treated with50 ng of NGF per ml for the times indicated (in min). Molecular mass markers are indicated. Cell lysates were run on an 8% polyacrylamidegel and transferred to nitrocellulose as described in the text. This immunoblot was exposed to x-ray film for 9 days.
Cell Biology: Berg et al.

Proc. Nati. Acad. Sci. USA 88 (1991)
39, 40). The availability of specific antibodies will greatlyfacilitate the identification of these protein substrates. De-termination of the functional significance of tyrosine phos-phorylation of these proteins and the generality of thisresponse in vivo will aid in elucidating the signaling pathwaysused by NGF in carrying out its ultimate biological effects.
We are grateful to H. Hanafusa, T. Miyasaka, and A. Saltiel forhelpful advice, L. A. Greene for the PC12nnrS cell line, and Nila Patilfor the deletion constructs. This work was supported by grants fromthe National Institutes of Health (NS21072), the Hirschl/CaulierTrust Fund, and the Dorothy Rodbell Cohen Foundation to M.V.C.and the National Cancer Institute (K08CA01367) and Andrew MellonFoundation to B.L.H.
1. Ullrich, A. & Schlessinger, J. (1990) Cell 61, 203-212.2. Chao, M. V., Bothwell, M. A., Ross, A. H., Koprowski, H.,
Lanahan, A., Buck, C. R. & Sehgal, A. (1986) Science 232,518-521.
3. Johnson, D., Lanahan, A., Buck, C. R., Sehgal, A., Morgan,C., Mercer, E., Bothwell, M. & Chao, M. V. (1986) Cell 47,545-554.
4. Radeke, M. J., Misko, T. P., Hsu, C., Herzenberg, L. A. &Shooter, E. M. (1987) Nature (London) 325, 593-597.
5. Maher, P. A. (1988) Proc. Natl. Acad. Sci. USA 85,6788-6791.6. Maher, P. A. (1989) J. Neurosci. Res. 21, 29-37.7. Miyasaka, T., Sternberg, D., Miyasaka, J., Sherline, P. &
Saltiel, A. (1991) Proc. Natl. Acad. Sci. USA 88, 2653-2657.8. Kaplan, D. R., Martin-Zanca, D. & Parada, L. F. (1991) Na-
ture (London) 350, 158-160.9. Kaplan, D. R., Hempstead, L. S., Martin-Zanca, D., Chao,
M. V. & Parada, L. F. (1991) Science 252, 554-557.10. Hempstead, B., Schleifer, L. S. & Chao, M. V. (1989) Science
243, 373-375.11. Pleasure, S. J., Reddy, U. R., Venkatakrishnan, G., Roy,
A. K., Chen, J., Ross, A. H., Trojanowski, J. Q., Pleasure,D. E. & Lee, V.-Y. (1990) Proc. Natl. Acad. Sci. USA 81,8496-8500.
12. Matsushima, H. & Bogenmann, E. (1990) Mol. Cell. Biol. 10,5015-5020.
13. Hosang, M. & Shooter, E. M. (1985) J. Biol. Chem. 260,655-662.
14. Green, S. H. & Greene, L. A. (1986) J. Biol. Chem. 261,15316-15326.
15. Hempstead, B. L., Patil, N., Thiel, B. & Chao, M. V. (1990) J.Biol. Chem. 265, 9595-9598.
16. Meakin, S. 0. & Shooter, E. M. (1991) Neuron 6, 153-163.17. Hempstead, B. L., Martin-Zanca, D., Kaplan, D. R., Parada,
L. F. & Chao, M. V. (1991) Nature (London) 350, 678-683.
18. Brown, A. M. C. & Scott, M. R. D. (1987) in DNA Cloning,ed. Glover, D. M. (IRL, New York), Vol. 3, pp. 189-212.
19. Hamaguchi, M., Grandori, C. & Hanafusa, H. (1988) Mol. Cell.Biol. 8, 3035-3042.
20. Bernd, P. & Greene, L. A. (1984) J. Biol. Chem. 259, 15509-15516.
21. Green, S. H., Rydel, R. E., Connolly, J. L. & Greene, L. A.(1986) J. Cell Biol. 102, 830-843.
22. Grob, P. M., Berlot, C. H. & Bothwell, M. A. (1983) Proc.Natl. Acad. Sci. USA 80, 6819-6823.
23. Buxser, S., Puma, P. & Johnson, G. L. (1985) J. Biol. Chem.260, 1917-1926.
24. Bothwell, M. A., Schechter, A. L. & Vaughn, K. M. (1980)Cell 21, 857-866.
25. Kirschmeier, P. T., Housey, G. M., Johnson, M. D., Perkins,A. S. & Weinstein, I. B. (1988) DNA 7, 219-225.
26. Miller, A. D. & Buttimore, C. (1986) Mol. Cell. Biol. 6,2895-2902.
27. Schlessinger, J. (1986) J. Cell Biol. 103, 2067-2072.28. Volonte, C. & Greene, L. A. (1990) Growth Factors 2, 321-331.29. Hempstead, B. L., Patil, N., Olson, K. & Chao, M. (1988) Cold
Spring Harbor Symp. Quant. Biol. 53, 477-485.30. Greenberg, M. E., Greene, L. A. & Ziff, E. B. (1985) J. Biol.
Chem. 260, 14101-14110.31. Bartel, D. B., Sheng, M., Lau, L. F. & Greenberg, M. E.
(1989) Genes Dev. 3, 304-313.32. Pollock, D., Krempin, M. & Rudy, B. (1990) J. Neurosci. 10,
2626-2637.33. Kaplan, D. R., Morrison, D. K., Wong, G., McCormick, F. &
Williams, L. T. (1990) Cell 61, 125-133.34. Kaplan, D. R., Whitman, M., Schaffhausen, B., Pallas, D. C.,
White, M., Cantley, L. & Roberts, T. M. (1987) Cell 50,1021-1029.
35. Kim, U.-H., Fink, D., Kim, H. S., Park, D. J., Contreras,M. L., Guroff, G. & Rhee, S. G. (1991) J. Biol. Chem. 266,1359-1362.
36. Vetter, M., Martin-Zanca, D., Parada, L. F., Bishop, J. M. &Kaplan, D. R. (1991) Proc. Natl. Acad. Sci. USA 88, 5650-5654.
37. Anderson, N. G., Maller, J. L., Tonks, N. K. & Sturgill, T. W.(1990) Nature (London) 343, 651-653.
38. Klein, R., Jing, S., Nanduri, V., O'Rourke, E. & Barbacid, M.(1991) Cell 65, 189-197.
39. Boulton, T. G., Nye, S. H., Robbins, D. J., Ip, N. Y., Rad-ziejewska, E., Morgenbesser, S. D., DePinho, R. A., Panay-otatos, N., Cobb, M. H. & Yancopoulos, G. D. (1991) Cell 65,663-676.
40. Schanen-King, C., Nel, A., Williams, L. K. & Landreth, G.(1991) Neuron 6, 915-922.
7110 Cell Biology: Berg et al.













![RESEARCHARTICLE …...prototypes and low-and high-BMI maleprototypes. Theprototypeswerecreated bydigitally averagingten facesofindividualswith thedesired BMIs[25].Thelow-BMIfemale](https://img.pdfslide.us/doc/110x75/5ec130bf6e6df93a5b2f5959/researcharticle-prototypes-and-low-and-high-bmi-maleprototypes-theprototypeswerecreated.jpg)














