Embed Size (px)
Citation preview

Board Meeting: September 2015 Agenda Number: 12 (i) File Ref: PU0024
LORD HOWE ISLAND BOARD
Business Paper ITEM
Lord Howe Island Rodent Eradication Program Update
BACKGROUND
On 18 May 2015, after the community consultation process over late 2014 and early 2015 ending with the community survey, the Board decided to proceed with the planning and approvals stage of the Program leading towards implementation of the rodent eradication plan, if the required approvals were received. The decision-making process was captured in a diagram, which assisted the Board on the day and which has been amended to include additional information and is attached. The rodent eradication program has now been divided into three stages: Stage One: Preliminary planning and community consultation
This stage has already been completed. It involved undertaking required initial trials including captive management and toxin resistance trials as well as initial operational planning. It included the biosecurity review and progressing of biodiversity outcome monitoring. Finally it included the community consultation and engagement process and the community survey.
Stage Two: Planning and Approvals
This stage is now underway. The key tasks during this stage are:
• Assemble personnel to undertake the work on the next stages • Review the Rodent Eradication Plan to ensure that it takes into consideration all new
information since it was drafted in 2009 • Develop individual property and livestock management plans, which will inform the
eradication plan and the approval process. This will involve a detailed property by property consultation with individual leaseholders and residents.
• Undertake any necessary studies required for the approval process, including independent health assessment
• Continue the relevant baseline outcome monitoring • Finalise detailed planning and all necessary risk assessments; • Obtain required permits and approvals, • Update and finalise operational details; • Prepare tender documentation

Stage Three: Implementation and evaluation of the eradication plan
This Stage will not happen until Stage Two is completed.
After all the necessary approvals are obtained and the required planning is undertaken, the decision-makers, that is the Commonwealth and State funding bodies and the Board will consider all the information and make the decision about proceeding to Stage Three.
Stage Three will involve the eradication plan being implemented in winter 2017 over a three month period. In summary the key tasks leading up to and during this stage are:
• Establish monitoring programs • Procurement of bait and helicopter operators • Recruitment of temporary field staff for hand baiting • Captive management of selected non target species • Carrying out the baiting operation • Environmental monitoring • Evaluation of the outcome of the eradication
CURRENT POSITION
1. Assembling the personnel and resources The Project Manager’s position was advertised because the existing Project Manager, Pete McClelland, was not able to return to the Island for personal reasons. However Pete will continue to be involved with the Program from off the Island in reviewing strategies and plans until the Project Manager position is filled, and then be available for advice and consultation after that. Interviews for the Project Manager position will be held in Sydney on 15 September. One of the Assistant Project Manager positions was advertised and recruitment has now been completed. The successful candidate is Anthony Wilson, currently Catchment Coordinator, Northeast Catchment Management Authority, Victoria. Anthony will be moving to the Island to take up the position with his wife and young son in early October. There is another Assistant Project Manager position in the Program structure. However recruitment to this position will not take place until the planning and approvals have been further progressed.
2. Project timelines
The Planning and Approvals Stage will take up to two years to complete. There is significant work to be undertaken in preparing and gathering information from property management and making applications for various approvals. The approvals also take time to be processed and determined.

Following is a broad outline of the steps in Stage 2: Planning and Approvals. It is subject to change depending on the time taken for each step.
Step Description Start Date End Date Status
1 Assemble resources 1 July 2015 30 June 2016
Engage staff to undertake the Planning and Approvals work:
- Project Manager - Asst Project Manager (Community) - Asst Project Manager (Operations)
Develop role descriptions for field staff
3 /8/15 1/715 1/3/16
31/10/15 30/9/15 30/6/16
2 Communications and consultation 1 July 2015 30 June 2017
Communications Plan developed Communications and consultation undertaken
3 Property Management Plans 1 Oct 2015 31 Mar 2016
Develop 130 Property Management Plans Develop 30 Livestock Management Plans
4 Captive Management arrangements 1 Nov 2015 30 June 16
Species to be managed – agreed Contract with Taronga Zoo drafted
1/11/15 1/1/16
31/12/15 30/6/16
5 Operations Plan update 1 Oct 2015 31 Mar 2016
Update to include operational constraints arising from Prop & Livestock Mgt Plans.
1/10/15 31/03/16
6 Permits and approvals 31 Mar 2016 30 Nov 2016
Prepare applications for approvals: • Species impact statement • APVMA approval • EPCB approval • Captive holding permit • Animal ethics approval • NSW Pesticides Act approval • POEO Act Flight Ops approval • CASA Flight Ops approval
7 Human health assessment review 31 Mar 2016 30 Nov 2016

Identify appropriate reviewer Request for review to be undertaken
8 Biodiversity outcome monitoring 1 Nov 2015 30 June 2017
Monitoring Plan confirmed Monitoring undertaken Report completed
9 Biosecurity arrangements 1 Nov 2015 30 June 2017
Finalise Biosecurity Plan Develop implementation plan Implement plan (subject to funding)
10 Preparation of contracts 30 June 2016 28 Feb 2017
Tender documentation prepared for: • Bait procurement • Helicopter operations • Captive management
11 Technical and financial feasibility assessment
31 Dec 2016 30 Mar 2017
The feasibility assessment will take into account any changes to the methodology resulting from the planning and approvals process and cost estimates for the implementation.
11 Stage 3 Implementation Plan 31 Dec 2016 30 June 17
Decision made by LHIB, CfOC and ET to proceed to Stage 3 Stage 3 Implementation Plan developed
3. Outcome Monitoring report
The monitoring project is being undertaken by the Office and Environment and Heritage’s Science Division under the project leadership of Dr Nicholas Carlile. It is an ongoing process taking place over the duration of the Rodent Eradication Program. For 2014/2015, there were numerous trips to the Island by project staff looking at different animal and plant species.
The following studies were undertaken during the 2014/2015 year:
• Terrestrial bird study • Seabird study of Black Noddy • Reptile study of the LHI Skink.

• Seabird study of Black-winged Petrel • Flora studies of Mountain Palms • Flora studies of other flower plants
A Final Report (attached) of this part of the Outcome Monitoring Project has been produced: Monitoring, evaluating and reporting the biodiversity benefits of eradicating exotic rodents from Lord Howe Island.
RECOMMENDATION
That the Board note the LHI Rodent Eradication Program update
Prepared __________________ Penny Holloway Chief Executive Officer

Rodent EradicationProcess for resolution
•2015 Community consultation outcome•Majority view to proceed•Level of support likely to impact success
•APVMA Approval process•EPBC Approval process•NSW approval processes•Independent Health assessment incorporating international evidence and advice
•Board Go/No go decision•Informed by:•Status of approvals•Additional advice from chief scientist•Ongoing community consultation
Principle 1:Board needs to respond to majority view
Principle 2: Board committed to attempting to resolve outstanding community concerns
Principle 3:Board will act to keep our people safe, and to protect the Island’s environment, World Heritage status and unique tourism assets
Ongoing Community Consultation

i
OFFICE OF ENVIRONMENT AND HERITAGE
Monitoring, evaluating and reporting the biodiversity benefits of eradicating exotic rodents from Lord Howe Island.
Final report:15 June 2015
Nicholas Carlile Science Manager, Ecosystem Management Science

ii
Contents
Summary iv
Background 1
Flora Studies Tony Auld, Andrew Denham, Mark Tozer, Ian Hutton 4
Introduction 4
Methods Impact of Black rats on fruits and seeds 6
Impacts of Black rats on the Little Mountain Palm on Mt Gower 8
Extinction risk for the Gnarled Mossy Cloud Forest EC 9
Results Impact of Black rats on fruits and seeds 9
Impacts of Black rats on the Little Mountain Palm on Mt Gower 12
Extinction risk for the Gnarled Mossy Cloud Forest EC 14
Discussion 15
Reptile Studies Robert Wheeler, George Madani 18
Introduction 18
Methods 19
November 2014 19
April 2015 22
Results and Discussion 22
North Bay November 2014 22
Salmon Beach November 2014 26
North Bay / Salmon Beach April 2015 27
Recommendations 29
Bird Studies 30
Seabird studies for identifying candidates for biodiversity benefits monitoring
on Lord Howe Island Nicholas Carlile, Lisa O’Neill, Ben Hope 30
Introduction 30
White Tern 31
Methods 31
Results and Discussion 32
Grey Ternlet 32

iii
Methods 32
Results and Discussion 34
Black Noddy 34
Methods 35
Results and Discussion 35
Black-winged Petrel 36
Methods 36
Results and Discussion 37
Seabird Recommendations 38
Land birds on Lord Howe Island Peter Fullagar, Chris Davey, A O Nicholls and
Ian Hutton SUMMARY 39
Land Bird Recommendations 39
Acknowledgements 40
References 41
Appendix 1 44
Appendix 2 (PDF) Assessing impacts of introduced rats on Lord Howe Island
plants including the Little Mountain Palms and its habitat on Mt Gower
(with images) to be attached 45
Appendix 3 (PDF) Auld TD & Leishman MR (2015). Ecosystem risk assessment for
Gnarled Mossy Cloud Forest, Lord Howe Island, Australia. Austral Ecology 40,
364–372 to be attached

iv
Summary
The Biodiversity Benefits Program associated with the rodent eradication project has
been established to assess and document the biodiversity benefits of removing rats
and mice from the World Heritage Lord Howe Island. The program provides a
measure of the return on investment. It also allows an evaluation of current status of
species and so any impacts of the eradication of rodents on key non-target species
can be tracked during their recovery. Over time, results from the various monitoring
components could be integrated to identify and explore changes to ecosystem
processes.
The following key species and taxa were targeted specifically during this revised two-
year project (2013-14 and 2014-15): Flora: Big Mountain Palm Hedyscepe
canterburyana; Little Mountain Palm Lepidorrhachis mooreana and fruiting plants
(including Blue Plum Chionanthus quadristamineus, Berrywood Ochrosia elliptica,
Green Plum Atractocarpus stipularis, Pandanus Pandanus forsteri and Tamana
Elaeodendron curtipendulum); Reptiles: LHI Skink Oligosoma lichenigera ; Seabirds:
White Tern Gygis alba; Grey Ternlet Procelsterna cerulean, Black Noddy Anous
minutus, Black-winged Petrel Pterodroma nigripennis; land birds.
Flora studies used selective caging and cafeteria trials to determine removal and
loss of fruits and seeds to Black Rats Rattus rattus. Of the 16 species examined,
losses to Black Rats were very high for six species (Howea forsteriana, Olea
paniculata, Baloghia inophylla, Jasminium simplicifolium, Smilax australis and
Geitonoplesium cymosum); potentially very high but variable for Berrywood;
moderate for three species (Syzygium fullagarii, Dietes robinsoniana and Blue Plum)
and low-moderate in the remaining 6 species. No species tested was entirely free of
seed or fruit losses to Black Rats. This suggests Black Rats are likely to impact on
many species across LHI and further testing of species is warranted to examine the
number of species likely to be at risk due to impacts on seed production, seed
survival and plant recruitment. In addition, further work on what impact the loss of
seeds or fruits may have on the affected plant populations is also necessary.
Additionally the high impact of Black Rats on the Little Mountain Palm on Mt Gower
that had been identified in 2007 was confirmed by repeat sampling in 2014. While

v
there is some successful recruitment in this species where rodent baiting occurs on
Mt Gower, recruitment failure is evident at locations over 100 m or so from bait
stations. This suggests that to successfully maintain this critically endangered palm
on Mt Gower rat control or eradication will be required across the Mt Gower summit.
The habitat of the Little Mountain Palm, the Gnarled Mossy Cloud Forest, was
assessed as critically endangered using the IUCN Red List for Ecosystems criteria.
Control of Black Rats on the summit of Mt Gower is the most practical means to
reduce the threat to this ecological community in the short term.
The LHI Skink is endemic to Lord Howe Island, and is listed as Vulnerable in both
NSW and national environmental legislation. Surveys for this species were
undertaken at various locations on the island in November 2014 and April 2015 to
obtain baseline data on the skink before an eradication takes place. Population
structure, relative abundance and distribution were attempted measurements for
comparative studies following the rodent eradication.
It was found that the skink is in low numbers, if present at all, in the southern and
central areas of Lord Howe Island. However, the population in North Bay appears to
be abundant although largely restricted to the sedge-grass plant community found on
sand dunes.
Surveillance camera studies on seabirds had mixed results. White Tern showed no
impact from rodents in incubation and chick provisioning periods but were present on
nesting branches when sites were inactive. Accessible Grey Ternlets nests were
scarce, reflecting the potential impacts of rodents on this species on the main island.
From two monitored nests, one was unsuccessful with subsequent rodent predation
of an unguarded egg. Black Noddy nesting was unaffected by rodents and none
were recorded at nest sites during the study. Black-winged Petrel were monitored
monthly during the breeding season and those nesting areas where rats were
uncontrolled had poor success in incubation and total failure of fledglings. In areas
where rats were controlled, breeding success was mixed but the results were also
confounded by heavy Masked Owl Tyto novaehollandiae predation of adults. If the
Biodiversity Benefits Program was to continue in its revised form future studies of
seabirds should concentrate on Grey Ternlet and Black-winged Petrel breeding sites

vi
using surveillance cameras. Consideration should also be given to monitoring Little
Shearwater Puffinus assimilis breeding sites using similar methods.
Comparing only two years of land bird surveys showed that most common bird was
the Golden Whistler Pachycephala pectoralis followed by the Silvereye Zosterops
lateralis. A noticeable reduction in all indices occurred for the next four most
frequently reported species – Blackbird Turdus merula, Pied Currawong Strepera
graculina, Magpie-lark Grallina cyanoleuca and Welcome Swallow Hirundo nigricans.
The suggested potential differences between 2013 and 2014 for some species (Buff-
banded Rail Gallirallus philippensis, Magpie-lark, Welcome Swallow and Blackbird)
need further exploration and additional observations before it can be determined that
the detected differences between years are truly significant.

1
Monitoring, evaluating and reporting the biodiversity benefits of eradicating exotic rodents from Lord Howe Island.
Final report: 15 June 2015
Background:
The Lord Howe Island group (LHIG; 31°31'S, 159°03’E) is located 760 kilometres
north east of Sydney. The group comprises the main island (Lord Howe Island; 1455
ha) and 28 smaller islets and rocks (Figure 1). The most significant of these are the
Admiralty Group (1 km to the north of LHI) and Balls Pyramid (23 km to the south
east). The island group is a World Heritage Area with global environmental
significance, and Australia has an international obligation to effectively manage the
island’s substantial, world-renowned natural values.
In common with a number of oceanic islands, populations of invasive rodents have
been accidentally introduced to Lord Howe Island (LHI). The house mouse Mus
musculus probably arrived before 1860, and the ship rat Rattus rattus in 1918. Both
species of introduced rodent have significant impacts on the biodiversity of Lord
Howe Island. Rats are implicated in the extinction of at least five endemic bird taxa
(species or subspecies), and the local extirpation of the endangered LHI Phasmid
Dryococelus australis, now found only on Balls Pyramid, and Wood-feeding
Cockroach Panesthia lata on offshore islands including the Admiralty Group. The
rodents are recognised as an ongoing threat to at least another 13 bird species, two
reptiles, 49 flowering plants, 12 vegetation communities, and numerous threatened
invertebrates, many of which are endemic to LHI. Rats also cause significant
damage to the Kentia Palm Howea forsteriana, with resulting economic losses to the
local palm industry. Predation by ship rats on LHI is listed as a Key Threatening
Process under both New South Wales (NSW) state and Australian Government
legislation.
The eradication of rodents from LHI will have enormous benefits for a range of
species currently adversely affected by rats and mice, including a suite of threatened
endemic species. In particular, we expect increases in the abundance of seeds and

2
seedlings, thereby enhancing the process of forest regeneration as well as increases
in the abundance and distribution of invertebrates, reptiles, seabirds, and land birds.
The monitoring program associated with the rodent eradication project is currently
assessing and documenting the biodiversity benefits of removing rats and mice. This
provides a measure of the return on investment. It also evaluates any impacts of the
eradication on key non-target species and will track their recovery in the absence of
rodents. Over time, results from the various monitoring components could be
integrated to identify and explore changes to ecosystem processes.
The initial project plan (April 2013 to June 2018) intended to research and monitor
key taxa identified in the Rodent Eradication Plan to determine the changes in the
distribution and abundance of those species. The work commenced prior to the
actual eradication operation planned at the time, to allow collection of baseline, pre-
eradication data, and was to continue for at least three years after the eradication to
assess the benefits of the operation. The following taxa were targeted specifically:
Kentia Palm; Big Mountain Palm; Little Mountain Palm and other fruiting plants; land
snails; the LHI Wood-feeding Cockroach; LHI Gecko and LHI Skink; LH Pied
Currawong; land birds and seabirds. In addition, data from the annual surveys of LH
Woodhen conducted by the LHI Board was to be incorporated into the integrated
analysis. By December 2013 only surveys on targeted flora, land birds and selected
seabirds had occurred as the progress of the eradication plan altered due to
changing circumstances on LHI.
A revised project plan was amended in March 2014 in response to the decision
taken in early 2014 to move away from immediate implementation of the eradication
program to full community engagement and consultation. In response and to further
minimise costs, the scope of the monitoring program was reduced from the original
eradication plan. Reviewing existing data, the project refocused on those species or
taxa for which existing monitoring data was absent or inadequate. It is envisaged
that full monitoring could commence once implementation of the eradication program
resumes, with 1-2 years of data collectable before baiting occurs.

3
The following key species and taxa being targeted specifically during this revised
two-year project (2014-15 and 2015-16): Flora: Big Mountain Palm Hedyscepe
canterburyana; Little Mountain Palm Lepidorrhachis mooreana and fruiting plants
(including Blue Plum Chionanthus quadristamineus, Berrywood Ochrosia elliptica,
Green Plum Atractocarpus stipularis, Pandanus Pandanus forsteri and Tamana
Elaeodendron curtipendulum). Reptiles: LHI Skink Oligosoma lichenigera; Seabirds:
Black-winged Petrel Pterodroma nigripennis, Black Noddy Anous minutus; and land
birds.
This final report covers the period August 2013 to May 2015 (Appendix 1) as agreed
to in the LHIB Revised Project Plan, March 2014.

4
Flora Studies
Assessing impacts of introduced rats on Lord Howe Island plants including the Little Mountain Palms and its habitat on Mt Gower
Tony D Auld1, Andrew Denham1, Mark Tozer1 and Ian Hutton2
1, NSW Office of Environment and Heritage, PO Box 1967 Hurstville NSW 2220
2, PO Box 157, Lord Howe Island, NSW 2898
Email: [email protected]
Introduction
The flora of Lord Howe Island is recognised as globally significant due to the high
level of endemism and unique vegetation communities. Auld and Hutton (2004)
detail this as: “The flora of Lord Howe Island has a high level of endemism and many
of the floristic assemblages are also unique to the island group (Pickard 1983, Green
1994). There are five plant genera endemic to Lord Howe Island — Negria,
Lordhowea and three palms, Howea, Hedyscepe and Lepidorrachis. For the vascular
plants, Green (1994) lists 459 species, 241 indigenous (53%), of which 105 are
endemic (44%) and 218 naturalised (48%). Of the indigenous vascular plants, 58
species are ferns and 183 are flowering plants. Several more species of both
indigenous and naturalised plants have been found since Green’s (1994) work. The
high level of endemism is typical of islands and comparable with megadiverse
regional areas of continents (Lowry 1998).”
It is widely recognised that invasive species may have significant negative impacts
on oceanic islands, particularly for many narrow range endemics and ecological
communities. While Lord Howe Island (LHI) was originally free of mammals (except
for small insectivorous bats) when first encountered by humans in 1788 and first
settled in 1834 (Hutton 1986), Black Rats Rattus rattus were accidentally introduced
when a steamship ran aground in 1918 (Billing and Harden 2000). A number of plant

5
species and vegetation communities have been identified as being at risk from the
impacts of Black Rats and House Mice Mus musclus on LHI. These impacts include:
• Loss of fruits and seeds to rats, e.g. Baloghia inophylla, Chionanthus
quadristamineus, Drypetes deplanchei, Elaeodendron curtipendulum,
Hedyscape canterburyana, Howea forsteriana, Howea belmoreana,
Lepidorrhachis mooreana, Pandanus forsteri, Ochrosia elliptica (Auld &
Hutton 2004, Auld et al. 2010);
• loss of seedlings and stem damage (the four palm species, Hedyscape
canterburyana, Howea forsteriana, Howea belmoreana, Lepidorrhachis
mooreana, Dietes robinsoniana (Wedding Lily) and at least two fern species,
Asplenium milnei and Adiantum hispidulum) (Auld & Hutton 2004);
• impacts on the critically endangered Gnarled Mossy Cloud Forest Ecological
Community confined to the summits of the southern mountains (NSW
Scientific Committee 2011);
As well, the extinctions of two LHI plant species, Sicyos australis and Solanum
bauerianum, were most likely influenced by consumption of seeds and fruits by
introduced animals (Auld & Hutton 2004; NSW Scientific Committee 2010). Solanum
bauerianum is now considered to be globally extinct.
In this work, we undertook to quantify the impact of introduced rats on 2 aspects of
the vegetation of LHI. Firstly, we wanted to understand the breadth of plants species
that may be impacted by rats. To do this, in the field we tested the magnitude of fruit
and seed loss to rats across a range of species, with the species tested being
dictated by fruit and seed availability. For some species we were able to replicate
the trials in different locations or at different times of the year. Secondly, we re-
examined the size distribution of plants of the Little Mountain Palm Lepidorrhachis
mooreana on Mt Gower. Previously, Auld et al. (2010) had shown that rats were
preventing the establishment of new plants into the parts of the population that were
not baited. We re-measured these sites to determine if the impact of rats was
ongoing and to try and see if rat baiting was leading to sufficient protection for these
palms.

6
Finally, through funding support from an Australian Research Council Grant to TA
and others we assessed the extinction risk for the Gnarled Mossy Cloud Forest
Ecological Community using the recently developed IUCN Red List for Ecosystems
criteria (Keith et al. 2013).
Methods Impact of Black Rats on fruits and seeds
At one or more sites (See Table 1) we established 5 plots beneath mature plants of
several species (Kentia Palm Howea forsteriana, Berrywood Ochrosia elliptica,
Maulwood Olea paniculata, Coprosma putida) (Table 1). Each plot contained three
treatments: uncaged; caged to exclude only birds (rodents could enter in gaps at
each end (see Fig. 1 Appendix 2); and caged to exclude both rodents and birds. At
each plot, in each treatment we placed 5 mature fruits. The density of fruits so
placed was not dissimilar to that found naturally below plants with mature fruits. We
recorded the fate of seeds over 3-7 days. We also established an infared camera at
one plot for each species to record what animal was responsible for seed removal or
loss (these cameras were set up to activate and take 10 photos when a warm
blooded animals comes into view of the camera (i.e. a rat , a mouse, a bird, a
human).
Table 1 Seed and fruit losses across species and sites. Family Scientific
Name Common Name
Life form
Experiment Type
Site Date of trial
Seeds lost to rats (%)
Fleshy Fruit species Arecaceae Howea
forsteriana Kentia Palm
Palm Single Neds Beach
Nov-13
100
Howea forsteriana
Single Research Station
Nov-13
100
Howea forsteriana
Single Windy Ridge W
Nov-13
100
Howea forsteriana
Single Windy Ridge
Nov-13
100
Howea forsteriana
Single Windy Ridge E
Nov-13
80
Howea forsteriana
Single Neds Beach
Jan-14
100
Oleaceae Olea paniculata Maulwood Tree Single Research Station
Nov-13
100
Olea paniculata Single Middle Beach
Nov-13
100

7
Olea paniculata Single Windy Ridge W
Nov-13
100
Olea paniculata Single Windy Ridge E
Nov-13
100
Apocynaceae Ochrosia elliptica
Red Berrywood
Shrub Single Neds Beach
Nov-13
96
Ochrosia elliptica
Single Middle Beach
Nov-13
96
Ochrosia elliptica
Single Windy Ridge W
Nov-13
12
Ochrosia elliptica
Single Windy Ridge E
Nov-13
4
Myrtaceae Syzygium fullagarii
Scalybark Tree Single Research Station
Jun-14
44
Syzygium fullagarii
Cafeteria Erskine Valley
Jun-14
40
Rubiaceae Coprosma putida
Stinkwood Shrub Single Research Station
Jun-14
0
Coprosma putida
Single Erskine Valley
Jun-14
16
Oleaceae Jasminium simplicifolium
Jasmine Vine Cafeteria Research Station
Nov-13
64
Jasminium simplicifolium
Cafeteria Middle Beach
Nov-13
100
Jasminium simplicifolium
Cafeteria Windy Ridge E
Nov-13
100
Jasminium simplicifolium
Cafeteria Windy Ridge
Nov-13
100
Smilacaceae Smilax australis Vine Cafeteria Research Station
Nov-13
92
Smilax australis Cafeteria Middle Beach
Nov-13
100
Smilax australis Cafeteria Windy Ridge E
Nov-13
92
Smilax australis Cafeteria Windy Ridge
Nov-13
96
Smilacaceae Geitonoplesium cymosum
Vine Cafeteria Research Station
Nov-13
68
Geitonoplesium cymosum
Cafeteria Middle Beach
Nov-13
100
Rutaceae Sarcomelicope simplicifolia
Yellow Wood
Tree Cafeteria Research Station
Nov-13
12
Rubiaceae Psychotria carronis
Black Grape
Tree Cafeteria Erskine Valley
Jun-14
20
Iridaceae Dietes robinsoniana
Wedding lily
Herb Cafeteria Erskine Valley
Jun-14
36
Oleaceae Chionanthus quadristamineus
Blue Plum Tree Cafeteria Erskine Valley
Jun-14
32
Meliacaea Dysoxylon pachyphyllum
Island Apple
Tree Cafeteria Erskine Valley
Jun-04
4
Old Fruit with no pulp Euphorbiaceae Drypetes
deplanchei Greybark Tree Cafeteria Research
Station Nov-13
8
Cafeteria Middle Beach
Nov-13
32
Dry seeds Fabaceae Sophora Lignum Tree Cafeteria Research Nov- 32

8
howinsula Vitae Station 13 Sophora
howinsula Cafeteria Middle
Beach Nov-13
36
Sophora howinsula
Cafeteria Windy Ridge E
Nov-13
0
Sophora howinsula
Cafeteria Windy Ridge
Nov-13
0
Euphorbiaceae Baloghia
inophylla Bloodwood Tree Cafeteria Neds
beach Nov-13
100
Baloghia inophylla
Cafeteria Middle Beach
Nov-13
100
Baloghia inophylla
Cafeteria Windy Ridge E
Nov-13
100
Baloghia inophylla
Cafeteria Windy Ridge
Nov-13
100
We also established plots for multiple species in a ‘cafeteria’ trial within the habitat
where they generally occur. We did this for 12 species at 5 sites (See Table 1). In
this trial 5 fruits or seeds of 4-6 species were placed together in small clumps (see
Fig. 2 Appendix 2). At each plot all seeds were uncaged. Again, like above, the fate
of the fruits/seeds was followed over 3-7 days if possible.
Impact of Black Rats on the Little Mountain Palm (Lepidorrhachis mooreana) on Mt
Gower
We repeated the sampling of Auld et al. (2010) (see end of Appendix 2) and sampled
the size structure of stands of Little Mountain Palms in cloud forest on the Mount
Gower plateau. We stratified the sampling between areas that have been baited by
the Lord Howe Island Board to control rat numbers and those that have never been
baited. In each of the baited and unbaited areas we sampled three separate plots for
the Little Mountain Palm (6 plots). We also sampled an additional 4 plots in the
gradient between baited and unbaited areas. We estimated the distance for each of
our 10 sampled plots from an existing bait station to determine if there was any
additional benefit of baiting into surrounding unbaited areas. At each plot, we
established a 5 m wide transect across the site and sampled all individual palms
within the transect. The transects were sampled until we had encountered a
minimum of 30-50 Little Mountain Palm individuals with an emergent trunk. We

9
measured the trunk height to the base of the leaves, the number of leaf scars for
individuals with a trunk and the height of individuals for individuals without a trunk.
Extinction risk for the Gnarled Mossy Cloud Forest Ecological Community
To assess the Gnarled Mossy Cloud Forest against the IUCN Red List for
Ecosystems criteria (Keith et al. 2013), we searched the available literature for
evidence of any threats to the ecosystem that may result in ecosystem decline. We
assessed the Gnarled Mossy Cloud Forest against four of the IUCN criteria (see
Appendix 2, Auld and Leishman 2015 for more details):
• Decline was assessed using historical vegetation reports (Maiden 1898, 1914;
Oliver 1916; Pickard 1983; Green 1994; Mueller-Dombois & Fosberg 1998;
Harris et al. 2005). For more recent changes, we inspected satellite imagery
(Google Earth and 2011 air photo imagery).
• For Restricted geographic distribution we used available Geographical
Information System data layers for the distribution of Gnarled Mossy Cloud
Forest (mapped from Pickard 1983) to estimate both the extent of occurrence
and area of occupancy of the ecosystem.
• To assess changes in abiotic features that directly impact on Gnarled Mossy
Cloud Forest, we used two metrics based on collated data from the Australian
Bureau of Meteorology from mid 1940s onwards (Rainfall and cloud cover).
• For biotic degradation, we reviewed the impact of exotic rats on limiting
recruitment of key species in the ecosystem and in changing the structure and
composition of the ecosystem.
Results Impact of Black Rats on fruits and seeds
In total, we were able to test the removal of fruits or seeds in 16 species, comprising
a mixture of plant families, life forms (trees, shrubs, vines) and habitats. Most
species examined had a fleshy fruit. In summary, seed or fruit losses were apparent
in all study species, at least at some sites.
Virtually all seed losses occurred at night and there was little removal of fleshy fruits
by birds from our ground based trials. It is likely that most bird removal of fruits

10
occurs on the plants and not the ground. Occasionally a shearwater returning to its
nest disturbed one of our plots and scattered fruits, but this was not recorded as fruit
loss in our calculations. Evidence of seed or fruit losses to rats was indicated by:
• Camera images of rats removing or consuming fruits/seeds (see Figs 3-6
Appendix 2)
• In situ seed or fruit remains indicating rats (eg, incisor marks etc) (see Figs 7-
11 Appendix 2)
We found no clear pattern of greater loss in areas without rat baiting stations present
versus those with baiting stations.
Trees
Howea forsteriana
There was high levels of seed removal in this species in all trials (5 sites, 2 time
periods, Table 1), with 5/6 trials with 100% fruit loss and 1/6 with 80% loss.
Olea paniculata
All four trails across 4 different sites showed 100% seed loss.
Syzygium fullagarii
Fruit loss was around 40-44% at 2 study sites. For both sites, the trials only ran for 2-
3 days so further fruit losses could be expected.
Sarcomelicope simplicifolia
12% fruit loss at one site (insufficient fruits available for further study).
Psychotria carronis
20% fruit loss at one site (trial only ran for 2 nights so further losses may occur).
Chionanthus quadristamineus
32% fruit loss at one site (trial only ran for 2 nights so further losses may occur).
Dysoxylon pachyphyllum
4% fruit loss at one site (trial only ran for 2 nights so further losses may occur).

11
Drypetes deplanchei
Seed loss ranged from 8-32% across 2 sites. Only fruits with no flesh available for
trials.
Sophora howinsula
Seed loss ranged from 0-36% across four sites. When eaten seed, seeds were
consumed in situ (see Fig. 8).
Baloghia inophylla
100% seed loss in all trials across 4 sites.
Shrubs
Ochrosia elliptica
Fruit loss varied from 4-96% across sites (Table 1). One of the sites with low losses
was in a location with no adult plants. The cameras showed clear evidence of rats
removing fruits (Fig. 4 Appendix 2). On other occasions fruits were eaten in situ and
fruit remains where present.
Coprosma putida
Fruit loss varied from 0-16% across 2 study sites.
Dietes robinsoniana
36% fruit loss at one site (trial only ran for 2 nights so further losses may occur).
Vines
Jasminium simplicifolium
Total fruit loss (100%) at three sites and 64% at the fourth
Smilax australis
Fruit loss varied from 92-100% across four sites.
Geitonoplesium cymosum
68-100% fruit loss across 2 sites.

12
Impact of Black Rats on the Little Mountain Palm (Lepidorrhachis mooreana) on Mt
Gower
Based on sampling in 2006, Auld et al. (2010) showed that not only were the fruits of
Little Mountain Palm consumed by rats, but the recruitment and establishment of a
juvenile seedling bank only occurred in the baited habitats on Mt Gower. Our re-
sampling of these and other sites in 2014 found the pattern remains unchanged. Two
of the 3 sites that were unbaited lacked new seedlings and small juveniles (eg. Fig.
12) and one of these showed a decline in larger juveniles over time, suggesting
ongoing recruitment failure. The unbaited site nearest the baiting stations showed a
small pulse of juveniles less than 50 cm in height, suggesting that that there may
have been some decline in rat impacts for a period in this area. The three sites in the
baited area, all showed evidence of some small juveniles being present, but the
abundance of small juveniles was greatly reduced in 2014 compared to 2006 (eg see
Fig. 13). For the 4 additional sites sampled, one had no juvenile plants at all, two had
no seedlings (one of these sites had only 1 juvenile less than 50 cm high), while only
one site had a few seedlings.
Fig. 12. A comparison of juvenile plants sizes between 2006 and 2014 at an unbaited site of Little Mountain Palm on Mt Gower. Note the lack of small juveniles and seedlings.
seedl
<50cm
<100cm
<150cm
<200cm
>200cm
Juvenile plant size (cm)
0
2
4
6
8
10
12
Freq
uenc
y
20142006
YEAR

13
Fig. 13. A comparison of juvenile plants sizes between 2006 and 2014 at a baited site of Little Mountain Palm on Mt Gower. Note the presence of small juveniles and seedlings (particularly in 2006).
There was a pattern of greater recruitment of juvenile plants closer to bait stations
(Fig. 14) with recruitment of small plants declining to zero at plots sampled around
250 m from bait stations.
Fig. 14. Abundance of seedlings (black circles) and small juveniles (black triangles) of Little Mountain Palms in relation to distance from nearest bait station on Mt Gower.
seedl
<50cm
<100cm
<150cm
<200cm
>200cm
Juvenile plant size (cm)
0
100
200
300
400
500
Freq
uenc
y
20142006
YEAR
0 100 200 300 400 500Distance to nearest bait station (m)
0
10
20
30
40
50
60
70
Abun
danc
e
0 100 200 300 400 500Distance to nearest bait station (m)
0
50
100
150
Abun
danc
e

14
Extinction risk for the Gnarled Mossy Cloud Forest Ecological Community
Overall the Gnarled Mossy Cloud Forest is considered to be Critically Endangered
based on a restricted geographic distribution combined with continuing decline.
Historical Decline: There have been no observed changes in the extent of this
ecosystem in the last 50 years (Pickard 1983). No changes are indicated since
Pickard’s 1983 vegetation classification based on inspection of recent satellite
imagery (Google Earth and 2011 air photo imagery) and field traverses. The status
of the ecosystem is therefore Least Concern under criterion A1.
Geographical Extent and Continuing Decline: The geographic distribution of the
ecosystem is below the IUCN threshold for Critically Endangered (Auld and
Leishman 2015). Decline was inferred from: a loss of moisture from declining rainfall
and cloud cover due to climate change (affecting disturbance regimes, gap formation
and species survival and recruitment); ongoing exotic rat predation on seeds and
seedlings of several sensitive species that are structural components of the
ecosystem (affecting survival and recruitment); and the fact that the ecosystem is
considered to exist at only one location (Auld and Leishman 2015).
Decline in abiotic processes and features: There was a trend for decline in both
rainfall and the frequency of cloud cover although there is still a great deal of
variation in the degree of decline predicted (Auld and Leishman 2015, see end of
Appendix 3).
Decline in biotic processes and interactions: Rats damage key functional plant
species in the ecosystem via predation on seeds and seedlings (Auld & Hutton 2004;
Auld et al. 2010). Most of the area (93%) of Gnarled Mossy Cloud Forest is subject
to impact by rats (7% is baited to reduce impact). The extent and severity of seed
predation by rats is high for two key species (mountain palms, but the severity of
seed predation by rats on other taxa in the ecosystem is uncertain. More work on
this aspect is required.

15
Discussion Impact of Black Rats on fruits and seeds
There was evidence that rats were consuming fruits or seeds in all 16 species
examined. In summary, seed or fruit losses were apparent in all study species, at
least at some sites. Losses were very high for six study species (Howea forsteriana,
Olea paniculata, Baloghia inophylla, Jasminium simplicifolium, Smilax australis and
Geitonoplesium cymosum); potentially very high but variable for one species
(Ochrosia elliptica); moderate for three species (Syzygium fullagarii, Chionanthus
quadristamineus, Dietes robinsoniana) (the actual losses may be higher as the trials
only ran for a short period); generally low in 4 species (Sarcomelicope simplicifolia,
Psychotria carronis, Dysoxylon pachyphyllum, Coprosma putida) (but the actual
losses may be higher where the trials only ran for a short period); and low-moderate
in two species (Sophora howinsula, Drypetes deplanchei).
Further work on examining the impact of fruit losses on the ecology of the study
species would assist interpretation of these data. Does the impact of these losses
result in reduced potential for recruitment of new plants in the study species as has
been previously demonstrated for the Little Mountain Palm and Big Mountain Palm
Hedyscepe canterburyana (Auld et al. 2010, Simmons et al. 2012)? Given that
losses are occurring in all tested species, it would also be worthwhile to extend the
sampling to additional species as many other species are also likely to be impacted
by rats.
Impact of Black Rats on the Little Mountain Palm (Lepidorrhachis mooreana) on Mt
Gower
Repeat sampling in 2014 confirmed the earlier findings of Auld et al. (2010) that
Black Rats are significantly impacting on Little Mountain Palms on Mt Gower. Baiting
appears to be effective in allowing the development of a juvenile bank of palms,
although there was evidence that the size of this juvenile bank was greatly reduced
in 2014 compared to 2006. Sites within about 100m of bait stations had some
seedlings and small juveniles (indicating recent recruitment), but sites 250m or
further away mostly had zero recent recruitment. This suggests that to successfully

16
maintain this critically endangered palm on Mt Gower rat control or eradication
across the Mt Gower summit will be required.
Extinction risk for the Gnarled Mossy Cloud Forest Ecological Community
Assessment of the Gnarled Mossy Cloud Forest against the IUCN Red List for
Ecosystems criteria indicated that the ecosystem is critically endangered due to its
very highly restricted geographic distribution (confined to summits of Mt Gower and
Mt Lidgbird) combined with continuing decline due to a loss of moisture from
declining rainfall and cloud cover as a result of climate change (affecting disturbance
regimes, gap formation and species survival and recruitment); ongoing exotic rat
predation on seeds and seedlings of several sensitive species that are structural
components of the ecosystem (affecting survival and recruitment); and the fact that
the ecosystem is considered to exist at only one location (see Auld and Leishman
2015, Appendix 3 for details. This supports the current listing of this ecological
community as critically endangered under the NSW Threatened Species
Conservation Act (NSW Scientific Committee 2011).
The recognition that the introduced Black Rat is a threat to the Gnarled Mossy Cloud
Forest mirrors similar threats from exotic species and climate change to other Pacific
island cloud forests (Meyer et al. 2010). Eradication of rats from Lord Howe Island
will reduce the immediate risk to this ecosystem, however, only global mitigation of
greenhouse gases could alleviate risk from declining cloud cover and moisture
availability. If rat eradication does not occur or is unsuccessful, regular rodent baiting
across much of the distribution of the ecosystem, particularly on Mt Gower would be
needed to alleviate the threat. This may not be a long-term solution as the rodents
may gain resistance to the poisons. A reduction in rat impact is needed to both allow
successful seed production and recruitment of new plants in the Gnarled Mossy
Cloud Forest.

17
Reptile Studies
Lord Howe Island Skink Survey: November 2014 and April 2015
Robert Wheeler1, George Madani2
1, NSW Office of Environment and Heritage, PO Box 1967 Hurstville NSW 2220
Email: [email protected]
Introduction
Funding has been obtained from the New South Wales and Australian governments
to undertake the eradication of introduced rodents from Lord Howe Island.
Monitoring key non-target species is crucial to evaluating the biodiversity benefits of
such an eradication. One such species is the Lord Howe Island Skink Oligosoma
lichenigera which is listed as Vulnerable under both the NSW Threatened Species
Conservation Act 1995 and the Commonwealth’s Environment Protection and
Biodiversity Conservation Act 1999.
This species is restricted to Norfolk Island and Lord Howe Island (DECC 2007)
although Cogger et al. (2005) suggest that the two island populations are genetically
distinct, and should not be regarded as the same species. On the Lord Howe group
of islands the skink is present on the main island, Balls Pyramid, Blackburn Island
and Roach Island (DECC 2007) as well as many smaller islets (Carlile & Priddel
2013a; 2013b; 2013c; 2013d). A wide range of vegetation communities, ranging from
lowland rainforest to montane forest as well as grasslands on the islets appear to be
acceptable to the skink (DECC 2007). On the main island, the skink seems to be
confined to sedge-grass habitat (Bray personal communication), the dense structure
of which may protect the skink from predators.
The skink feeds on beetles, spiders, ants and other invertebrates amongst the leaf
litter (DECC 2007) and fruit (Bray pers. com.).

18
Rats prey upon this species and are probably the main reason for its decline on the
main island (DECC 2007). The introduced Delicate Skink Lampropholis delicata,
which arrived in the early 1990’s, has spread from the settlement to the Northern
Hills and Intermediate Hill, and may compete for food with this species (DECC
2007).
To obtain baseline information on the status of the Lord Howe Island Skink (LHI
Skink) prior to rodent eradication, this survey of the lizard was undertaken to
determine:
• its relative abundance and distribution;
• population demography; and
• the extent of dispersal into habitats fringing sedge-grass habitat, currently the
stronghold of the species.
In addition, the feasibility of using tracking tunnels to assess broader distribution
patterns was evaluated.
Method
November 2014
Two potential study sites, North Bay and Windy Point, were inspected as to
suitability following information supplied by Rebecca Bray, a PhD candidate studying
Lord Howe Island’s two endemic lizard species, the LHI Skink and the Lord Howe
Island Gecko Christinus guentheri. The sedge-grass community located on the
foredune of North Bay is known to contain the largest population of the LHI Skink
(Bray, unpublished field-trip report September 2010). It may be no coincidence that
North Bay is also regularly baited for rats to protect the Lord Howe Island Flax Snail
Placostylus bivaricosus.
In November 2014, Sooty Terns Onychoprion fuscata were nesting along the dune,
particularly in the northern and central sections. The southern section had a
relatively small number of tern nests so it was selected as one study site as this
would result in little disturbance to these birds. This area corresponds to Rebecca
Bray’s North Bay Dune site. The sedge-grass community at Windy Point, which is

19
approximately 3 km south-east of the North Bay site, was known to be the next-most
populated site for the skink (Bray, unpublished field-trip report September 2010).
Unfortunately, approximately half of this site has since been washed away. The
sedge-grass community at Salmon Beach at the southern end of Lord Howe Island
was chosen as a substitute for Windy Point.
At each site, two lines of pitfall traps (20 traps at each site) were set in the sedge-
grass community on the sand dunes, and ran approximately parallel to the nearby
seashore. Distance between traps in each line was 5 metres. At North Bay the lines
were identified as Green (traps 1-12) and Blue (traps 1-8). The sedge-grass
community trap lines at Salmon Beach were separated by 7 metres, and both
contain ten traps each, with 5 metres between sequential traps. Additionally at each
site, pitfall traps were set in the adjacent habitats. For North Bay, two lines (the North
and Middle lines) ran at 3250 to the sedge-grass community lines, and passed
through a shrub community dominated by hopbush Dodonaea viscosa burmanniana
then into a Kentia Palm Howea forsteriana forest. The North and Middle lines each
had one trap in the sedge-grass community, one on the boundary of sedge-grass
community and the adjacent habitat and eight traps in the adjacent communities. A
South Line (one trap in the sedge-grass, one on the boundary and four in adjacent
habitat) ran through a Kentia Palm forest growing on shallow soil over limestone with
rock outcrops. At Salmon Beach a line of ten traps ran perpendicular to the sedge-
grass community through lowland rainforest towards Mt Lidgbird. Traps in this
Lidgbird Line were left closed for two days after being set as a precaution in case the
smell of newly turned soil attracted rodents or the Lord Howe Woodhen Gallirallus
sylvestris to them.
Nearest-neighbour distance for traps 1, 2, 3 and 4 in the lines into adjacent habitat
(i.e., the North, Middle, South and Lidgbird lines) was 5 metres. Except for the South
Line, distance between traps after trap 4 was 10 metres. For the South Line traps,
the nearest-neighbour distance for all the traps was 5 metres, and, unlike the straight
line followed in the North, Middle and Lidgbird lines, the course of the South Line
was somewhat zig-zagged because of the scarcity of soil deep enough to fully burry
the pit-trap.

20
Each pit-trap line consisted of a series of 5-litre buckets (190 mm deep, 210 mm
across, MaxiPailR) buried into the soil up to the rim. A quarter of a pear (for bait)
(Goulburn ValleyR tinned pears), several leaves (for shelter) and pieces of pumice or
polystyrene (for floatation in case of flooding) were placed into each bucket. A
minimum of eight holes, each 8 mm in diameter, was also drilled into the bottom of
each bucket for drainage. However two traps on the South Line (and situated on a
slope) filled with water as a consequence of moderate rainfall during one morning.
Traps were set for four consecutive days and nights at the dune sites (North Bay and
Salmon Beach) and the adjoining habitat sites at North Bay, in the period 18 – 26
November 2014. Traps were left open overnight with the exception of those traps
along the Lidgbird line, which were closed at night to safeguard against being raided
by rats. The pears used to lure skinks into the pit-traps were missing from the
majority of traps at the Lidgbird site by late afternoon on the second day of trapping
which suggested that this line was also being visited by currawongs or woodhen,
hence the Lidgbird line was closed after only two days.
Traps were cleared between 7:30 and 10 a.m., and again around mid-day and late
afternoon/dusk. Skinks caught were weighed using a Pesola spring balance,
measured (snout-vent) on a ruler, and individually numbered using a non-toxic
marker pen (Sharpie Fine-point Permanent MarkerR). Sex determination was partly
based on colouration. A yellowish or copper-colour underside was taken as evidence
of the skink being male; gravid females had pronounced bulges to their sides.
Differentiation between adults and sub adults was based on the amount of colouring
(bright colouration is indicative of breeding status) and animal size. Only data
collected from those skinks caught in the sedge-grass community, the known
stronghold of the species, has been used to examine demography.
To evaluate the practicality of using tracker tunnels for surveying skink distribution,
two captured skinks were released at one end of a tunnel, and encouraged to leave
at the other end, and in so doing, pass over an area painted with printer’s ink. The
aim was to determine if the resulting foot prints were sufficiently clear to distinguish
tracks made by the LHI Skink from that of the Delicate Skink.

21
April 2015
Surveys for the skink were conducted on Lord Howe Island between the 16 - 26 April
2015 at the two sites examined in November 2014 as well as an additional site at the
playground where a survey had been undertaken by Rebecca Bray in April-May
2011. The additional site was included due to the lack of capture of the LHI Skink at
Salmon Beach the previous spring.
Trap lines opened at North Bay (along the dune, and into the adjacent habitat) and at
Salmon Beach (along the dune) were the same as for November but the line leading
towards Mount Lidgbird from Salmon Beach was not used due to the significant
presence of predators. Two lines, each of 10 traps, were set out approximately
parallel to the beach near the Playground, and another line of 10 traps ran
perpendicular to these, and passed through a Norfolk Pine-Kentia Palm community.
Nearest-neighbour spacing of traps at this site was 5 metres.
Traps were baited with pieces of pear, which was replenished after three days.
Trapping was conducted at Salmon Beach and the playground from 16 – 26 April,
and at North Bay from 20 – 26 April. Trapping was not conducted on the 21st and
22nd April when over 230 mm of rain was recorded falling on Lord Howe Island.
Results and Discussion
North Bay November 2014
In the equivalent of 92 pit-trap days at the sedge-grass dune site at North Bay (12
traps in the Green Line, eight in the Blue Line, plus the first trap of the North, Middle
and South lines, each line opened for four days and nights) 123 LHI Skinks were
caught, with daily totals of 34, 25, 37, and 27. There were only three recaptures. This
contrasts markedly with the trapping result for September 2010 when 32 LHI Skinks
were caught in 20 traps over five days (100 pit-trap days) in the same area (Bray,
unpublished field-trip report September 2010). In September 2010, recaptures
numbered four, a 12.5% recapture rate compared to less than 3% for November
2014. It is possible that the ink marking used to number the skinks was lost due to
abrasive contact between the marked lizard and the coarse sedge vegetation or
sand, and this resulted in recaptured individuals being recorded as new captures by

22
mistake. When descriptions of lizards of the same length (+ 1mm) are compared in
regard to noted features (such as sex, weight, tail characteristics, when and where
caught) there were at least 99 individuals caught at North Bay in November 2014 so
it is possible that there may have been up to 24 other recaptures that were recorded
as new captures. Although, the time between the initial and subsequent capture with
the three known recaptures was no more than two days, the marking was still
distinctive so it is likely that a substantial proportion of these 24 captures were of
new animals. Even if it is accepted that there were only 99 individuals caught, and
not 123, this still represents a three-fold increase in the September 2010 total with
approximately the same capture effort. The difference may be due to a rise in the
skink population or, perhaps, to an increase in skink activity (and, therefore,
probability of capture) as spring progressed.
Sixty-three of the dune skinks (25 females, 24 males, 14 sex unknown; 34 adults and
29 sub-adults) were noted to have wounds to some degree, ranging from scratch
marks on the head and body to a regenerated tail or part thereof. Such wounds may
be a product of robust mating activity or possibly antagonistic behaviour as a result
of overcrowding.
Of the 67 captured LHI Skinks from North Bay that were judged to be adult, at least
42 are female but only 19 to 25 are believed to be male (Table 1) although at least
40% of the sub-adult population is comprised of males. It is unlikely that the unequal
sex ratio for the adults is a product of trapping bias with adult males being somehow
less prone to falling into traps. The approximately two-to-one bias in favour of
females may represent the adult population structure of the dune colony. More than
half the number of skinks caught in, or at the border with, the adjacent habitats were
believed to be male (6 males, 5 unknown sex; Table 2). More than 81% of the skinks
caught in the nearby habitats or at the very edge of the sedge-grass community were
judged to be sub-adults. These findings suggest that males, particularly young
males, are being disproportionably excluded from the better habitat.
Four skinks were caught in the Dodonaea shrub community, at 5 metres (n = 2, one
of which was an adult), 10 metres (n = 1) and 20 metres (n = 1) from the sedge
grass area. Three, including one adult, were caught in the palm forest up to 70

23
metres from the dune, and one skink was caught in the rock-outcrop area (i.e., along
the South Line) 15 metres from the sedge-grass community.
No Delicate Skinks were caught at North Bay although they were commonly seen
along the path on the eastern side of the Dawson Point Ridge.
Table 1: Size and gender information for those Lord Howe Island Skinks caught in the sedge-grass
dune community at North Bay in November 2014 (Table 1a) and April 2015 (Table 1b). For the
determination of sex and age, see text. Length, in millimetres, the distance between the tip of the
snout and the vent; weight in grams.
Table 1a: Capture data from November 2014
All
captured
skinks
Sub-
adults
sex
unknown
Adults
sex
unknown
Sub-
adults
female
Adults
female
Sub-
adults
male
Adults
male
Number
caught
123 33 7 1 41 22 19
Mean
length
(SD; n)
63.0
(8.1; 122)
55.4
(3.7; 32)
63.9
(6.6; 7)
58 69.2
(4.9; 41)
56.6
(3.3; 22)
70
(5.8; 19)
Length
range
47-79 47-63 57-77 - 60-78 49-63 60-79
Mean
weight
(SD; n)
5.4
(2.2; 121)
3.6
(0.8; 33)
5.2
(1.2; 7)
4.2 7.1
(1.8; 40)
3.7
(0.5; 21)
6.9
(1.8; 19)
Weight
range
2.0-11.5 2.0-6.0 4.0-7.5 - 4.5-11.5 2.5-4.7 4.0-11

24
Table 1b: Capture data from April 2015
All
captured
skinks
Sub-
adults
sex
unknown
Adults
sex
unknown
Sub-
adults
female
Adults
female
Sub-
adults
male
Adults
male
Number
caught
32 2 2 9 4 15
Mean
length
(SD; n)
63.2
(9.1; 32)
50
(2.8; 2)
55
(1.4; 2)
67.7
(7.7; 9)
50
(3.8; 4)
66.3
(7.2; 15)
Length
range
47-76 48-52 54-56 58-76 47-56 56-76
Mean
weight
(SD; n)
5.63
(2.5; 26)
2.9
(0; 2)
3.45
(0.2; 2)
7.0
(2.4; 9)
2.6
(1.0; 3)
6.1
(2.5; 10)
Weight
range
2.2-9.7 2.9-2.9 3.3-3.6 4.4-9.7 2.2-4.2 3.2-9.1

25
Table 2: Size and gender information for those Lord Howe Island Skinks caught in habitats in the
vicinity of the sedge-grass dune community at North Bay in November 2014. For the determination of
sex and age, see text. Length, in millimetres, the distance between the tip of the snout and the vent;
weight in grams.
All
captured
skinks
Sub-
adults
sex
unknown
Adults
sex
unknown
Sub-
adults
female
Adults
female
Sub-
adults
male
Adults
male
Number
caught
11 5 0 0 0 4 2
Mean
length
(SD; n)
52.1
(8.2; 11)
49.6
(7.7; 5)
- - - 50.3
(6.9; 4)
62
(1.0; 2)
Length
range
39-63 39-59 - - - 39-58 61-63
Mean
weight
(SD; n)
3.2
(1.1; 11)
2.7
(1.0; 5)
- - - 3.1
(0.9; 4)
4.4
(0.4; 2)
Weight
range
1.5-4.8 1.5-4.0 - - - 2.0-4.5 4.0-4.8
Salmon Beach November 2014
The only skink species caught at Salmon Beach was the introduced Delicate Skink;
ten were caught in the sedge-grass community in 80 trap days, and one 70 metres
into the forest towards Mount Lidgbird during 20 trap days. In the sedge-grass
community, four skinks were caught on the first day, three on the second, two on the
third and one was caught on the last day; there were no recaptures.
Suitability of Tracking Tunnels

26
Two LHI Skinks were put through a tracking tunnel to evaluate the usefulness of
these tunnels for monitoring lizard distribution. The results were not encouraging.
The prints of a large skink (snout-vent length of 72 mm and weight of 8 g) were not
discernible being nothing more than smudges of ink on the running board, while a
smaller skink (snout-vent 52 mm, weight 3 g) got stuck on the ink board although it
eventually managed to free itself.
North Bay April 2015
Unlike in November, the greater majority of adult LHI Skinks caught at North Bay in
April are believed to be male (at least 53% compared to 28% for November) (Table
1). Overall, there were 35 individual skinks caught at this site, 32 in the dune traps
and three (an adult female, an adult male and a sub-adult of indeterminate sex) in
the adjacent habitat.
Sixty-seven LHI Skinks were trapped at North Bay over seven days/nights in autumn
2011 (Bray, unpublished field-trip report April-May 2011). The difference in the
numbers caught in 2011 compared to 2015 (67 and 32 respectively) may simply be
due to a difference in trapping effort, and not represent a decline in the population at
North Bay. Traps were closed for several days in April 2015 when recorded rainfall
exceeded 230 mm. Traps were also only opened during the day because of the
potential threat of overnight rain, therefore the trapping effort in 2015 was less than
that for 2011.
Three LHI Skinks were caught in the Dodonaea community up to 20 metres from the
sedge habitat. One Delicate Skink was also captured in this area. No LHI Skink was
caught in the palm forest in contrast to the capture result in November 2014.
Salmon Beach April 2015
The only skink found at Salmon Beach was the Delicate Skink; six captures being
recorded.

27
The playground April 2015
Low numbers of the LHI Skink were recorded at the playground in 2010 (e.g., three,
Bray unpublished fieldtrip report: Lord Howe Island Lizards - Nov-Dec 2010). No LHI
Skink was caught at the sedge-grass community or in the adjacent Norfolk Pine-
Kentia Palm habitat although five Delicate Skinks were trapped in the sedge-grass
community. One pit-trap was found to be without its pear bait on the sixth morning of
the survey which may indicate that the trap had been visited recently by a rodent or
currawong.
Current Status of the Lord Howe Island Skink
Trapping results from 2010 and 2011 (Bray, unpublished reports) and this survey
(2014 and 2015) indicate that the LHI Skink is, at best, present in very low numbers
in the southern and central areas of Lord Howe Island. The skink is, however,
abundant in the sedge-grass community growing on the dunes at North Bay. The
population also appears to be healthy; at least 44% of the adult females caught in
North Bay were carrying young in November. In addition, over 20% of the LHI Skinks
caught in 2014, and 18% of those caught in 2015 had been marked with translucent
tags by Rebecca Bray in 2010 and 2011 which may suggest a significant level of
survival for individuals as well as a high reproductive output.
Large numbers of LHI Skinks at North Bay may be a consequence of the rat baiting
that takes place in the area to protect the LHI Snail. However, the carcass of one rat
and the odour from several other dead rats were detected at the site in November
just after the vicinity was baited, so rats are still in the area. Baiting also takes place
at the playground (one bait station was located within two metres of a pit-trap in the
Norfolk Pine-Kentia Palm community) yet no LHI Skinks were found there, and a rat
was seen scurrying through the sedge-grass community at dusk. Mice also prey on
reptiles and their eggs, and can severely reduce reptile populations (Towns and
Broome 2003). If mice are not being controlled by the current baiting programme for
rats then it is likely that mice are also impacting the the populations of the LHI Skink.
Although the population at North Bay appears to be substantial and healthy, its long-
term future is not necessarily guaranteed. The species was once found in a variety of
habitats on the main island (DECC 2007) but now it is largely restricted to the sedge-

28
grass community found on sand dunes, and even here its distribution has shrunk to
the northern part of the island.
Recommendations
It is recommended that:
• rodent baiting continues at North Bay;
• the artificial wall built to stabilise the area at Windy Point is searched for the
LHI Skink;
• tracker tunnels are set in the sedge-grass community at the playground and
North Bay to determine the degree that rats or mice use these areas;
• reptile surveys are undertaken in the other habitats at North Bay if rodents are
eradicated from the main island; and
• the LHI Board considers translocating LHI Skinks to other parts of the main
island if a rodent-eradication programme proves successful.

29
Bird Studies
Seabird studies for identifying candidates for biodiversity benefits monitoring on Lord Howe Island
Nicholas Carlile, Lisa O’Neill, Ben Hope
NSW Office of Environment and Heritage, PO Box 1967 Hurstville NSW 2220
Email: [email protected]
Introduction
The Biodiversity Benefits Program associated with the rodent eradication project was
established to assess and document the likely positive outcomes from the removal of
rats and mice. With costly programs for the eradication of rodents, it is desirable to
provide a meaningful measure of the return on investment. Despite the presence of
rodents on Lord Howe Island (LHI) for a century there has been little direct evidence
collected of their impact on seabirds. This group of animals makes up the most
significant component of the islands fauna group through the variety of species and
the sheer numbers of individuals. The potential extent of their occurrence over the
island, their annual breeding and foraging habits significantly add to, and possibly
‘drive’, the islands productivity through the transfer of oceanic nutrients to the
terrestrial system. The loss of species from the main island ecosystem and the
suppression of populations that breed here due to rodent impacts has the potential to
be catastrophic for the long-term viability of the current ecosystems that have
evolved here when they were more populous.
Two species of seabirds that once bred on the main island no longer occur here.
Populations of White-bellied Storm-petrel Fregetta grallaria thrive on the outer
islands (i.e. Carlile et al. 2013; Carlile & Priddel 2013a; 2013b;) and Kermadec Petrel
Pterodroma neglecta also occurs on the outer islands (Hutton 1991; Carlile & Priddel
2013a) but both were originally known from the southern mountains as well as other
locations on the main island (Hindwood 1940). It is likely, with the removal of rodents
that these species will return to breed on the main island.

30
Evaluation of any impacts of the eradication on key non-target species would be
most profitable monitoring changes in breeding response during recovery of species
currently breeding on the main island.
The revised Biodiversity Benefits Program specifically targeted spring-summer
breeding White Tern Gygis alba; Grey Ternlet Procelsterna cerulean, Black Noddy
Anous minutus and Black-winged Petrel Pterodroma nigripennis. It was considered
that these species were most likely to have suffer suppression of breeding success
(all species) and extent of occurrence (Grey Ternlet) since rodents, and specifically
Black Rats Rattus rattus became established on the island.
White tern
The White Tern breeds only within the settled lowlands, and colonies are highly
visible. The first substantiated breeding of this species on LHI was in 1968 (McAllen
et. al 2004). The preferred habitat is large, mature Norfolk Island Pines Araucaria
heterophylla that line Lagoon Road along the western shoreline of the island. These
trees were planted during the 1900s and have historical significance. Younger
stands of Norfolk Island Pines have since established within areas of native
vegetation. Concerned about the invasive nature of this species, the LHI Board has
declared Norfolk Island Pines an exotic weed (Lord Howe Island Board 2006), and
self-seeded stands that do not have historical value are being systematically
removed.
White Terns do not make a nest but lay their egg directly on a tree branch, usually
using a dimple or depression in the branch to help hold the egg on the branch. A
single egg is laid, though the terns do have capacity to lay a second and sometimes
even a third egg if prior eggs are lost.
Methods
Motion sensor cameras were set to monitor nests within the settlement area,
principally in the private garden of Wilsons Bike Hire business. Cameras were set
between 1.5 and 5.5 metres above the ground, attached to tree branches. In total 26
cameras were used to monitor 26 nests from egg laying (or just prior to laying) to

31
time of chick fledging or prior loss of the chick or egg. The majority of cameras were
set prior to nesting commencing. Over 629,000 images were captured at these nests
from 15th of September 2013 to 5th of April 2014 for analysis. Cameras were serviced
monthly to remove and download data cards, replace batteries and alter viewing
area or re-set camera positions depending on the outcome of the breeding attempt
at the monitored site.
Results and Discussion
The monitoring showed an extremely low survival rate from egg to fledgling chick.
Only three monitored chicks fledged and these chicks were not hatched in these
nests but were put into these nests as a young chick by ‘Macca’ of Wilsons Bike Hire
after being rescued from having fallen out of a nest elsewhere.
Of 25 first eggs laid, nine hatched (Hatching Success (HS) = 36%). There were 14
nests where a second egg was laid, ten of these were laid after loss of the previous
egg and four after loss of the previous chick. Of these 14 nests, eight eggs hatched
(HS = 57%). There were five nests where a third egg was laid. Three of these were
nests where the second egg was lost and two where the chick was lost. Of the five
third laid eggs, two hatched (HS = 40%). One nest was monitored from part way
through the season and it is not known if it was a first or subsequent nest, so is not
included in the analysis above. This nest successfully hatched its chick which was
later lost to an unknown cause.
In total, from 26 nests, 45 eggs were laid, 20 chicks hatched from those eggs, and
none survived to fledge. The causes of egg lost were varied, including falling from
the nest (4), predation by LH Currawong Strepera graculina crissalis (5) and egg
infertility (4). Of the remaining eggs (12 or 48%) that were lost, a cause of loss could
not be determined.
The major cause of chick loss was predation by LH Currawong, but one third of chick
disappearances could not be attributed to any cause. Chicks ranging in age from one
day old to 26 days old were predated by currawongs. Breeding Success (BS) at this
site was 0% although limited numbers of near-fledged young and flying juveniles
were observed at other locations (Carlile unpub data).

32
The lack of complete image capture from nest sites to determine egg and chick
losses highlights the failure of the infrared trigger system in the cameras to pick up
all bird movements. Birds are covered in feathers, which provide excellent insulation,
and do not allow indication of heat differentials between their body heat and the
ambient temperature. Only when the areas of high temperature differential, the front
of the head (eyes and open beak) or the underwings (when exposed while landing or
taking off) are presented to the infrared sensor does the camera trigger. Conversely,
mammals have an excellent thermal signal and would have been detected by the
cameras, so they can be excluded as the cause of the unknown nest failures.
White Tern showed no impact from rodents in incubation and chick provisioning
periods but Black Rats were present on nesting branches when sites were inactive
either before nesting or directly after depredation events. This indicates that the
presence of adult White Tern on nesting branches is a sufficient deterrent for rat
investigation of potential prey. Far from being threatened by either rodents or
predatory birds, the White Tern population continues to expand and is currently
estimated at 600 pairs in 2014, up from approximately 500 pairs in 2006 (Carlile &
Priddel 2015). Additional bird species observed during surveillance camera
monitoring of White Terns included two sightings of Long-tailed Cuckoo Eudynamys
taitensis 25/11/2013 and 14/12/2013.
Grey Ternlet
Grey Ternlets are a cavity nesting seabird, one of only two such species on LHI. The
White-bellied Storm-petrel Fregetta grallaria now only persists in nesting cavities on
the outer islands (i.e. Carlile et al. 2013) since the arrival of the Black Rat in 1918.
The Grey Ternlet, present on LHI since European occupation, is now confined to
occasional nesting sites within the North Hills and on the main islands east coast. It
is far more prevalent on the outer islands (Hutton pers comm), possibly as a result of
rodent interactions (McAllen et al. 2004).
Methods
Between the 16th and 21st of October 2013, surveys were made of the cliffs in the
North Hills in search of nesting Grey Ternlets which were expected to be incubating

33
or with freshly hatched young (Hutton 1991). Ian Hutton assisted in surveys around
Old Gulch and Dean Hiscox directed surveyors to sites at New Gulch. Surveillance
cameras were installed on accessible nests and monitored during the breeding
period until the fledging period in January 2014.
Results and Discussion
Surveys of Old Gulch recorded only two nests in total. The only accessible nest
already had a 10-day old chick in it and was deemed too advanced for camera
monitoring. Surveys at New Gulch recorded a total of six active Grey Ternlet nests
sites. Only two of these were accessible without the need for rope assistance, for
camera monitoring. One of the two nests had adults visiting at the time of camera
installation but the egg was cold, possibly indicating an abandoned nesting attempt.
Subsequent monitoring here showed that the egg was not being incubated and it
was removed by a Black Rat 10 days after the camera was installed. The second
nest was successful in raising a chick to fledging, and there were no images of rat
visitation. The chick was not left alone, even after initial brooding as adults remained
in attendance at the nest entrance at night for the duration of the in-nest provisioning
period as indicated by regular incursion to the nesting chamber during the hours of
darkness.
It is possible that by mid-October many nesting attempts had failed and the adult
Grey Ternlets had already abandoned their breeding sites. Future surveys for this
species should commence in early September and include Hells Gate and cliff lines
along the east coast of the main island as far as Red Point. Boat surveys in early
September would determine the most productive areas to search from the land for
camera monitoring.
Black Noddy
The Black Noddy is a tree nesting seabird, one of only two such species on LHI. It
has been breeding on LHI Island since 1989 (Hutton 1991). The population has, until
recently, been confined to North Bay where it has been expanding slowly in the
introduced Norfolk Island Pines behind the fore dune. Recently breeding had
commenced in a few Norfolk Island Pines along Lagoon Road in the settlement area

34
(Hank Bower pers comm). It was suspected that, if the population was breeding
successfully, a more rapid expansion would have been expected over the last 45
years. It is possible that the species is suffering from some form of reduced breeding
success that has hindered its expansion.
The Black Noddy lays a single egg in a shaped nest, usually on the branch of a large
established tree. There is some evidence to suggest that if the egg is lost a pair will
re-lay (this study).
Methods
A pilot study into the breeding success and causes of breeding failure for Black
Noddy was initiated at the start of October 2014. Initially, nine motion sensor
cameras were set up to monitor Black Noddy nests in introduced Norfolk Island
Pines at North Bay. Cameras were set at between 2.5 and 3.5 metres above the
ground. At this time a small number of nests were already with incubating eggs but
nest-building was still underway. An additional camera was added to make ten
cameras in November 2014. From these 10 cameras it was possible to monitor 12
nests.
Results and Discussion
Over 380,000 images of breeding activities were captured for analysis. Egg survival
to hatching was high, with only 2 eggs being lost, one fallen from the nest and one
cracked. A second egg was laid in the nest from which an egg fell, giving a total of
13 eggs able to be monitored from 12 nests. The replacement egg was successfully
incubated providing an overall nest HS of 91%. All pairs that successfully hatched
their chick (11), also raised it to fledging giving an overall BS of 91%.
There was no evidence on camera that rats were visiting or preying upon Black
Noddy at their nests. There was no evidence of any predation at any monitored nest.
The identical hatch and fledging success would indicate that the adult attendance
was high, preventing rodent access to young chicks. This was corroborated by the
surveillance cameras which showed adults on or at the nest during hours of
darkness. The study area at North Bay is included in the baiting program, principally
to protect the Lord Howe Island Flax Snail Placostylus bivaricosus. The low rodent
population may assist the Black Noddy in their high breeding success. The current

35
population at North Bay is 510 nesting pairs. If the effectiveness of baiting for
controlling rodents at this site declines due to rodent tolerance to current available
baits, or if baiting ceased to be carried out, Black Noddy breeding may be impacted.
This would warrant further investigation as to rodent impacts in any future
Biodiversity Benefits Program.
Black-winged Petrel
Black-winged Petrels are a burrow-nesting seabird, one of only two small burrowing
seabirds present on the main island of LHI. The Black-winged Petrel was first
observed breeding on LHI in 1971 (Hutton & Priddel 2002). The population has
established in several locations close to the coast at the northern end of the island
and along the eastern coast of LHI (McAllen et al. 2004), expanding slowly to an
approximate estimate of 500 pairs (Carlile unpub. data). It is suspected that if the
population was breeding successfully that a more rapid expansion would have been
expected over the last 45 years. On Phillip Island, off Norfolk Island, a similar
population established at the same time as the LHI birds (Priddel et al. 2003). Here,
they have expanded to between 5,000 and 10,000 pairs on an island without
introduced rodents (Carlile unpub. data). It is possible that the species on LHI is
suffering from some form of reduced breeding success that has hindered its
expansion.
Methods
Two sites were selected for a survey of breeding success of Black-winged Petrel,
with 21 burrows monitored on North Head and 35 burrows monitored just north of
Blinkys Beach on the main islands east coast. The North Head site is isolated from
the nearest rodent baiting grid by 200 m and is within lowland mixed forest. The
Blinkys Beach site is within a regularly maintained baiting grid of coastal forest
dominated by Melaleuca howeana and intersected by a fishers track to the eastern
shore of the island. The two sites were monitored roughly monthly (6-7 January, 10-
11 February, 9-10 March, 28-29 April) and their contents recorded. The burrows
were initially inspected to coincide with when the majority of eggs would have been
laid (Hutton & Priddel 2002) using an infrared burrow scope (Peep-A-Roo: Sandpiper

36
Technologies, Manteca, CA, USA). Following observations were timed to hatching
and early brooding of young by parents, half-grown pullus without parental protection
during daylight hours and near fledged young when abandoned by adults prior to
their fledging period (Hutton & Priddel 2002).
Results and Discussion
Success differed at the two sites. At North Head, two of 21 eggs (10%) were
observed to have hatched young in February 2015 and a single egg was still being
incubated. A further three adults were within their nesting chamber, possibly
incubating eggs (total: 4 potential eggs or maximum HS of 29% if all eggs survived to
this point). The sitting adults could not be shifted to emphatically indicate an egg
beneath them but their behaviour during disturbance by the burrowscope indicated
they were likely incubating an egg. There were six scatters of egg shell fragments
found on the surface with signs of rat chewing. It was not possible to determine
whether rats had destroyed the eggs while being incubated or after the eggs were
abandoned from failed nesting attempts. By March there was no sign of any eggs or
chicks and most nests were already deserted. In April, there were no adults in any of
the previously active nests indicating a BS of 0%.
At the Blinkys Beach site, 23 of 35 eggs (66%) were being incubated and a single
chick had hatched in February. By March there were seven chicks (HS = 20%)
observed and by April, five large chicks remained (BS = 14%). One of these chicks
was malformed and appeared unlikely ever to fly, reducing potential BS to 11%. The
nest site that had contained the freshly hatched chick in February was empty
indicating an additional chick may have fledged successfully but this could not be
confirmed. There were five adult bodies found in the lower, more open, section of the
Blinkys Beach colony near the fishers track that appeared to have been killed by
Masked Owl Tyto novaehollandiae.
The two sites showed differences in timing of laying, with proportionally more chicks
hatching early at North Head, but with much poorer egg survival at North Head. A
more thorough monitoring of nests, using surveillance cameras, may be able to
determine whether predation is a factor in the poor egg survival and hatching to
fledging survival once adults no longer attend the chicks (from about 3-5 days old) at

37
North Head. Cameras at the Blinkys Beach site may help determine the extent of owl
and rat depredation.
Seabird Recommendations:
• If the Biodiversity Benefits Program was to continue in its revised form future studies of seabirds should concentrate on Grey Ternlet and Black-winged Petrel breeding sites using surveillance cameras.
• Consideration should also be given to monitoring Little Shearwater Puffinus assimilis breeding sites using similar methods. This species is known to breed in both areas of rodent control (Priddel et al. 2003) as well as at other locations around LHI (Carlile pers comm). It has been established that this species shares burrows with summer-breeding Black-winged Petrel (Hutton & Priddel 2002) and, due to its similar breeding demography, would be a candidate for rodent predation, similar to that concluded from this Black-winged Petrel study.

38
Landbirds on Lord Howe Island, NSW
Survey report 2 (summary)
Note: Final reports have been provided elsewhere to the LHI Board
Peter Fullagar1, Chris Davey1, A O Nicholls1 and Ian Hutton2
1, Canberra Ornithological Group PO Box 301 CIVIC SQUARE ACT 2608
2, PO Box 157, Lord Howe Island, NSW 2898
Email: [email protected]
The purpose of the terrestrial bird study was to provide baseline information on the
numbers and distribution of land birds on Lord Howe Island. Point counts within
natural vegetation were chosen as the preferred method for the island survey. The
data collecting protocol used was one that best fitted the limitations for time and
effort available in the field. Additional transect counts within the settlement were
used to track the terrestrial bird presence in highly disturbed environments.
The 2013 and the 2014 survey covered 96 sites and were sampled four times within
the survey periods, distributed across the lowlands, by the Canberra Ornithologists
Group (Fullagar et al. 2014; Fullagar et al. 2015). Full results and site selection
methods are available in reports already with the Lord Howe Island Board.
In essence, when comparing only two years of land bird surveys, the most common
bird was the Golden Whistler Pachycephala pectoralis followed by the Silvereye
Zosterops lateralis. A noticeable reduction in all indices occurred for the next four
most frequently reported species – Blackbird Turdus merula, Pied Currawong
Strepera graculina, Magpie-lark Grallina cyanoleuca and Welcome Swallow Hirundo
nigricans. The suggested potential differences between 2013 and 2014 for some
species (Buff-banded Rail Gallirallus philippensis, Magpie-lark, Welcome Swallow
and Blackbird) need further exploration.
Land Bird Recommendations:
• Additional observations are required before it can be determined that the detected differences between years are truly significant.

39
Acknowledgements
Many field programs are reliant on the efforts of volunteers and this Biodiversity
Benefits Program is no different. We would like to acknowledge the efforts of Mark
Allen, Lia Battisson, David Cook, Kathy Cook, John Goldie, Christine Ledger, Martyn
Moffat, Gail Newmann, Michael Robbins and Kathy Walter from the Canberra
Ornithologists Group who assisted in the land bird surveys. Juliet Dingle, Margie
Grant, Ian Knight, Anicee Lombal, Melinda Norton and George Madani are thanked
for their assistance in seabird and reptile data collection. Lord Howe Island Board
assisted with transport, logistics and advice, particularly Christo Haselden, Hank
Bower and Michael ‘Fitzy’ Fitzgerald. Lord Howe Island residents Ian Hutton
provided support in the flora, land bird and seabird studies; Dean Hiscox provided
direction for seabird survey sites; Craig ‘Macca’ Campbell provided logistical support
and access to breeding seabirds on his property and we thank him and his family for
their patience, the paper is coming. The residents of the island shared their stories of
seabird experiences as well as their general knowledge of their habits and for that
we are grateful.
The assessment of extinction risk for the Gnarled Mossy Cloud Forest was
supported by an Australian Research Council Linkage Grant to TA and others (LP
LP130100435) and we thank Prof. David Keith and all the participants of the
Australian Centre for Ecological Analysis and Synthesis workshop for their thoughtful
contributions to the development of the risk assessment for this ecosystem
The project was funded by the Lord Howe Island Board under the Rodent
Eradication Program. Ethics approval was provided through the OEH animal ethics
committee numbers 021028/02 and 021028/01.

40
References
Auld TD & Hutton I (2004). Conservation issues for the vascular flora of Lord Howe Island. Cunninghamia 8, 490–500.
Auld T. D., Hutton I., Ooi M. K. J. & Denham A. J. (2010). Disruption of recruitment in
two endemic palms on Lord Howe Island by invasive rats. Biological Invasions 12, 3351-3361.
Auld TD & Leishman MR (2015). Ecosystem risk assessment for Gnarled Mossy
Cloud Forest, Lord Howe Island, Australia. Austral Ecology 40, 364–372. Billing J, & Harden B (2000). Control of introduced Rattus rattus L. on Lord Howe
Island. I. The response of mouse populations to warfarin bait used to control rats. Wildl Res 27, 655–658.
Carlile, N., Priddel, D. (2013a). Seabird islands No. 261: Mutton Bird Island, Lord
Howe Group, New South Wales. Corella 37 (4) 94-96. Carlile, N., Priddel, D. (2013b). Seabird islands No. 258: South Island, Lord Howe
Group, New South Wales. Corella 37 (4) 88-89. Carlile, N., Priddel, D. (2013c). Seabird islands No. 259: Sugarloaf Island, Lord
Howe Group, New South Wales. Corella 37 (4) 90-91. Carlile, N., Priddel, D. (2013d). Seabird islands No. 257: Tenth of June Island, Lord
Howe Group, New South Wales. Corella 37 (4) 86-87. Carlile, N. & Priddel, D. (2015) Establishment and growth of the White Tern Gygis
alba population on Lord Howe Island, Australia. Marine Ornithology 43: 113–118.
Carlile, N., Priddel, D., & Bower, H. (2013). Seabird islands No. 256: Roach Island,
Lord Howe Group, New South Wales. Corella 37 (4) 82-85. Cogger, H., Cameron, E. & Sadlier, R. (1979). The terrestrial reptiles of islands in the
Norfolk Island complex. Australian Museum, Sydney. pp.1—122. DECC (2007). Lord Howe Island Biodiversity Management Plan. Department of
Environment and Climate Change. Sydney. http://www.environment.gov.au/resources/parks/LHI_bmp.pdf
Fullagar, P., Davey, C., Nicholls, A. O. & Hutton I. Landbirds on Lord Howe Island,
NSW: Survey report 1, September 2013. Report prepared for the Lord Howe Island Board by the Canberra Ornithologists Group, February 2014.
Fullagar, P., Davey, C., Nicholls, A. O. & Hutton I. Landbirds on Lord Howe Island,
NSW: Survey report 2, September 2014. Report prepared for the Lord Howe Island Board by the Canberra Ornithologists Group, March 2015.

41
Green PS (1994). Flora of Australia Vol. 49 Oceanic Islands 1. Australian Government Printing Service, Canberra.
Harris R, Cassis G, Auld T & Hutton I (2005). Floristics and structure of the mossy
cloud forest of Mt Gower summit, Lord Howe Island. Pac. Conserv. Biol. 11, 246–56.
Hindwood, K. A. (1940). The birds of Lord Howe Island. Emu 40 1-86. Hutton, I (1986). Lord Howe Island (Conservation Press: Canberra ACT). Hutton, I. (1991). ‘Birds of Lord Howe Island: Past and Present’. (Ian Hutton: Coffs
Harbour). Hutton, I. & Priddel, D. (2002). Breeding biology of the Black-winged Petrel,
Pterodroma nigripennis, on Lord Howe Island. Emu 102 361-365. Keith DA, Rodríguez JP, Rodríguez-Clark KM et al. (2013). Scientific Foundations for
an IUCN Red List of Ecosystems. PLoS ONE 8, e62111. Lord Howe Island Board. 2006. Weed management strategy for Lord Howe Island. Lowry, PP (1998). Diversity, endemism, and extinction in the flora of New Caledonia:
a review. In Rare, threatened and endangered floras of Asia and the Pacific Rim (eds. C.-I Peng & P.P. Lowry II), (Institute of Botany, Academia Sinicia Monograph Series No. 16. Taipei). pp. 181–206.
Maiden JH (1898). Observations of the vegetation of Lord Howe Island. Proc. Linn.
Soc. N. S.W. 23, 112–58. Maiden JH (1914). Further notes on the botany of Lord Howe Island (fifth paper).
Proc. Linn. Soc. N. S.W. 39, 377–84. Meyer J-Y (2010). Montane cloud forests on remote islands of Oceania: the example
of French Polynesia (South Pacific Ocean). In: Tropical Montane Cloud Forests: Science for Conservation and Management (eds L. A. Bruijnzeel, F. N. Scatena & L. S. Hamilton) pp. 121–9. Cambridge University Press, Cambridge.
McAllan, I.A.W., Curtis, B.R., Hutton, I & Cooper R.M. (2004). The birds of the Lord
Howe Island Group: A review of records. Australian Field Ornithology supplement. 21, 1–82.
Mueller-Dombois D & Fosberg FR (1998). Vegetation of the Tropical Pacific Islands.
Springer-Verlag, NewYork. NSW Scientific Committee (2010). Final determination. Solanum bauerianum -
species presumed extinct listing [Cited 9th June 2015] Available from URL: http://www.environment.nsw.gov.au/determinations/solanumbauerianumFD.htm

42
NSW Scientific Committee (2011). Final determination. Gnarled Mossy Cloud Forest
on Lord Howe Island – critically endangered ecological community listing. [Cited 23September 2014.] Available from URL: http://www.environment.nsw.gov.au/determinations/gnarledmossycloudforestlhifd.htm
Oliver WRB (1916). The vegetation and flora of Lord Howe Island. Trans. Proc. N. Z.
Inst. 49, 94–161. Pickard J (1983). Vegetation of Lord Howe Island. Cunninghamia 1, 133–265. Priddel, D., Carlile, N., Evans, O., Evans, B & McCoy H. (2010). A review of the
seabirds of Phillip Island in the Norfolk Island Group. Notornis 57, 113-137. Priddel, D., Hutton, I., Carlile, N & Bester, A. (2003). Little Shearwaters, Puffinus
assimilis assimilis, breeding on Lord Howe Island. Emu 103 67-70. Simmons L, Auld TD, Hutton I, Baker WJ, & Shapcott A (2012). Will climate change,
genetic and demographic variation or rat predation pose the greatest risk for persistence of an altitudinally distributed island endemic. Biology 1(3), 736-765.
Towns, D.R. & Broome, K. G. (2003). From small Maria to massive Campbell: forty
years of rat eradications from New Zealand islands. New Zealand Journal of Zoology 30, 377-398.
Wilkinson IS & Priddel D (2011). Rodent eradication on Lord Howe Island:
challenges posed by people, livestock, and threatened endemics. In: Island Invasives: Eradication and Management (eds CR Veitch, MN Clout & DR Towns) pp. 508–14. IUCN, Gland.
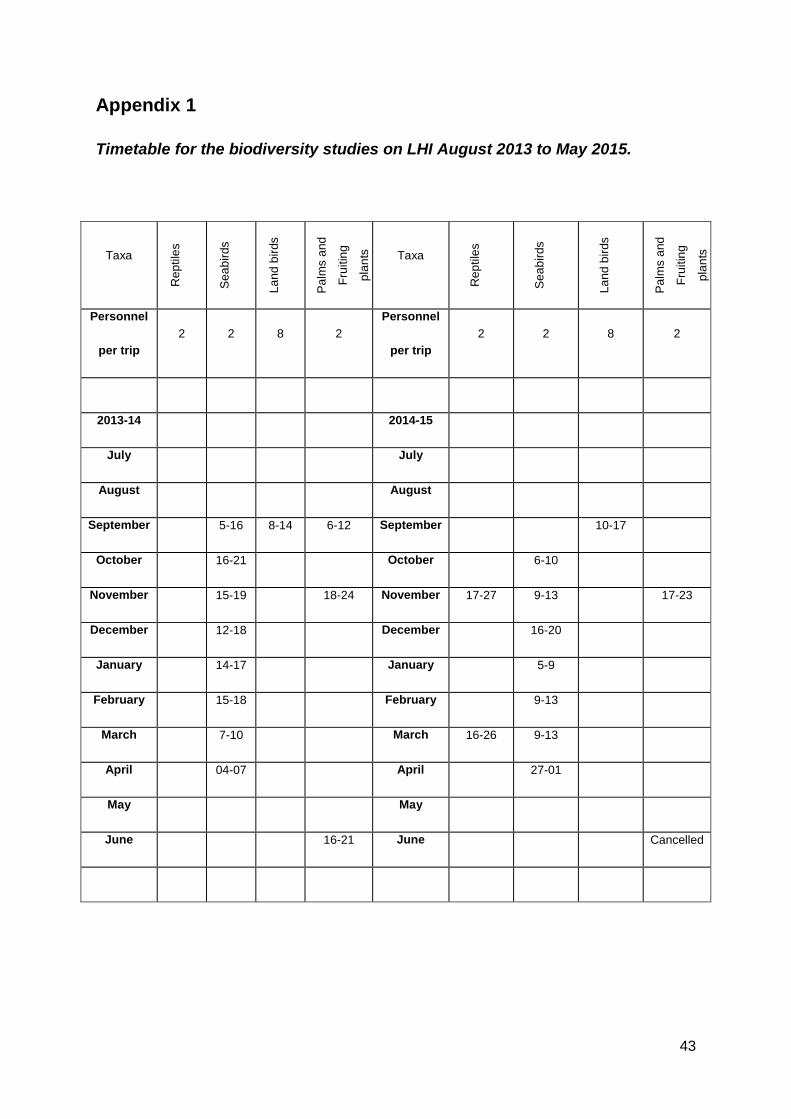
43
Appendix 1
Timetable for the biodiversity studies on LHI August 2013 to May 2015.
Taxa
Rep
tiles
Sea
bird
s
Land
bird
s
Pal
ms
and
Frui
ting
plan
ts
Taxa
Rep
tiles
Sea
bird
s
Land
bird
s
Pal
ms
and
Frui
ting
plan
ts
Personnel
per trip 2 2 8 2
Personnel
per trip 2 2 8 2
2013-14 2014-15
July July
August August
September 5-16 8-14 6-12 September 10-17
October 16-21 October 6-10
November 15-19 18-24 November 17-27 9-13 17-23
December 12-18 December 16-20
January 14-17 January 5-9
February 15-18 February 9-13
March 7-10 March 16-26 9-13
April 04-07 April 27-01
May May
June 16-21 June Cancelled

1
Assessing impacts of introduced rats on Lord Howe Island plants including
the Little Mountain Palms and its habitat on Mt Gower
June 2015
Tony D Auld1, Andrew Denham1, Mark Tozer1 and Ian Hutton2
1, NSW Office of Environment and Heritage, PO Box 1967 Hurstville NSW 2220
2, PO Box 157, Lord Howe Island, NSW 2898
Email: [email protected]
Summary
We used selective caging and cafeteria trials to determine removal and loss of fruits
and seeds to Black Rats (Rattus rattus) on Lord Howe Island. Of the 16 species
examined, losses to Black Rats were very high for six species (Howea forsteriana,
Olea paniculata, Baloghia inophylla, Jasminium simplicifolium, Smilax australis and
Geitonoplesium cymosum); potentially very high but variable for one species
(Ochrosia elliptica); moderate for three species (Syzygium fullagarii, Chionanthus
quadristamineus, Dietes robinsoniana) (the actual losses may be higher as the trials
only ran for a short period) and low-moderate in the remaining 6 species. No species
tested was entirely free of seed or fruit losses to Black Rats. This suggests Black
Rats are likely to impact on many species across LHI and further testing of species is
warranted to examine the number of species likely to be at risk due to impacts on
seed production, seed survival and plant recruitment. In addition, further work on
what impact the loss of seeds or fruits may have on the affected plant populations is
also necessary.
The high impact of Black Rats on the Little Mountain Palm (Lepidorrhachis
mooreana) on Mt Gower that had been identified by Auld et al. (2010) was confirmed
by repeat sampling 8 years after the original sampling was done. While there is
some successful recruitment in this species where rodent baiting occurs on Mt
Gower, recruitment failure is evident at locations over 100 m or so from bait stations.
This suggests that to successfully maintain this critically endangered palm on Mt
Gower rat control or eradication will be required across the Mt Gower summit. The
habitat of the Little Mountain Palm, the Gnarled Mossy Cloud Forest, was assessed
as critically endangered using the IUCN Red List for Ecosystems criteria. Control of
Black Rats on the summit of Mt Gower is the most practical means to reduce the
threat to this ecological community in the short term.

2
Introduction
The flora of Lord Howe Island is recognised as globally significant due to the high level of endemism and unique vegetation communities. Auld and Hutton (2004) detail this as: “The flora of Lord Howe Island has a high level of endemism and many of the floristic assemblages are also unique to the island group (Pickard 1983, Green 1994). There are five plant genera endemic to Lord Howe Island — Negria, Lordhowea and three palms, Howea, Hedyscepe and Lepidorrachis. For the vascular plants, Green (1994) lists 459 species, 241 indigenous (53%), of which 105 are endemic (44%) and 218 naturalised (48%). Of the indigenous vascular plants, 58 species are ferns and 183 are flowering plants. Several more species of both indigenous and naturalised plants have been found since Green’s (1994) work. The high level of endemism is typical of islands and comparable with megadiverse regional areas of continents (Lowry 1998).” It is widely recognised that invasive species may have significant negative impacts on oceanic islands, particularly for many narrow range endemics and ecological communities. While Lord Howe Island was originally free of mammals (except for small insectivorous bats) when first encountered by humans in 1788 and first settled in 1834 (Hutton 1986), Black rats (Rattus rattus) were accidentally introduced when a steamship ran aground in 1918 (Billing and Harden 2000). A number of plant species and vegetation communities have been identified as being at risk from the impacts of Black Rats and house mice on LHI. These impacts include:
• Loss of fruits and seeds to rats, e.g. Baloghia inophylla, Chionanthus quadristamineus, Drypetes deplanchei, Elaeodendron curtipendulum, Hedyscape canterburyana, Howea forsteriana, Howea belmoreana, Lepidorrhachis mooreana, Pandanus forsteri, Ochrosia elliptica (Auld & Hutton 2004, Auld et al. 2010);
• loss of seedlings and stem damage (the four palm species, Hedyscape canterburyana, Howea forsteriana, Howea belmoreana, Lepidorrhachis mooreana, Dietes robinsoniana (Wedding Lily) and at least two fern species, Asplenium milnei and Adiantum hispidulum) (Auld & Hutton 2004);
• impacts on the critically endangered Gnarled Mossy Cloud Forest Ecological Community confined to the summits of the southern mountains (NSW Scientific Committee 2011);
As well, the extinctions of two Lord Howe Island plant species, Sicyos australis and Solanum bauerianum, were most likely influenced by consumption of seeds and fruits by introduced animals (Auld & Hutton 2004; NSW Scientific Committee 2010). Solanum bauerianum is now considered to be globally extinct. In this work, we undertook to quantify the impact of introduced rats on 2 aspects of the vegetation of LHI. Firstly, we wanted to understand the breadth of plants species that may be impacted by rats. To do this, in the field we tested the magnitude of fruit and seed loss to rats across a range of species, with the species tested being dictated by fruit and seed availability. For some species we were able to replicate the trials in different locations or at different times of the year. Secondly, we re-examined the size distribution of plants of the little mountain palm (Lepidorrhachis mooreana) on Mt Gower. Previously, Auld et al. (2010) had shown that rats were preventing the establishment of new plants into the parts of the population that were

3
not baited. We re-measured these sites to determine if the impact of rats was ongoing and to try and see if rat baiting was leading to sufficient protection for these palms. Finally, through funding support from an Australian Research Council Grant to TA and others we assessed the extinction risk for the Gnarled Mossy Cloud Forest Ecological Community using the recently developed IUCN Red List for Ecosystems criteria (Keith et al. 2013). Methods Impact of Black Rats on fruits and seeds At one or more sites (See Table 1) we established 5 plots beneath mature plants of several species (Howea forsteriana (Kentia Palm), Ochrosia elliptica (Berrywood), Olea paniculata (Maulwood), Coprosma putida) (Table 1). Each plot contained three treatments: uncaged; caged to exclude only birds (rodents could enter in gaps at each end (see Fig. 1); and caged to exclude both rodents and birds. At each plot, in each treatment we placed 5 mature fruits. The density of fruits so placed was not dissimilar to that found naturally below plants with mature fruits. We recorded the fate of seeds over 3-7 days. We also established an infared camera at one plot for each species to record what animal was responsible for seed removal or loss (these cameras were set up to activate and take 10 photos when a warm blooded animals comes into view of the camera (i.e. a rat , a mouse, a bird, a human). We also established plots for multiple species in a ‘cafeteria’ trial within the habitat where they generally occur. We did this for 12 species at 5 sites (See Table 1). In this trial 5 fruits or seeds of 4-6 species were placed together in small clumps (see Fig. 2). At each plot all seeds were uncaged. Again, like above, the fate of the fruits/seeds was followed over 3-7 days if possible. Impact of Black Rats on the Little Mountain Palm (Lepidorrhachis mooreana) on Mt Gower We repeated the sampling of Auld et al. (2010) (see Appendix 1) and sampled the size structure of stands of Little Mountain Palms in cloud forest on the Mount Gower plateau. We stratified the sampling between areas that have been baited by the Lord Howe Island Board to control rat numbers and those that have never been baited. In each of the baited and unbaited areas we sampled three separate plots for the Little Mountain Palm (Lepidorrhachis mooreana) (6 plots). We also sampled an additional 4 plots in the gradient between baited and unbaited areas. We estimated the distance for each of our 10 sampled plots from an existing bait station to determine if there was any additional benefit of baiting into surrounding unbaited areas. At each plot, we established a 5 m wide transect across the site and sampled all individual palms within the transect. The transects were sampled until we had encountered a minimum of 30-50 Lepidorrhachis individuals with an emergent trunk. We measured the trunk height to the base of the leaves, the number of leaf scars for individuals with a trunk and the height of individuals for individuals without a trunk. Extinction risk for the Gnarled Mossy Cloud Forest Ecological Community To assess the Gnarled Mossy Cloud Forest against the IUCN Red List for Ecosystems criteria (Keith et al. 2013), we searched the available literature for

4
evidence of any threats to the ecosystem that may result in ecosystem decline. We assessed the Gnarled Mossy Cloud Forest against four of the IUCN criteria (see Appendix 2, Auld and Leishman 2015 for more details):
• Decline was assessed using historical vegetation reports (Maiden 1898, 1914; Oliver 1916; Pickard 1983; Green 1994; Mueller-Dombois & Fosberg 1998; Harris et al. 2005). For more recent changes, we inspected satellite imagery (Google Earth and 2011 air photo imagery).
• For Restricted geographic distribution we used available Geographical Information System data layers for the distribution of Gnarled Mossy Cloud Forest (mapped from Pickard 1983) to estimate both the extent of occurrence and area of occupancy of the ecosystem.
• To assess changes in abiotic features that directly impact on Gnarled Mossy Cloud Forest, we used two metrics based on collated data from the Australian Bureau of Meteorology from mid 1940s onwards (Rainfall and cloud cover).
• For biotic degradation, we reviewed the impact of exotic rats on limiting recruitment of key species in the ecosystem and in changing the structure and composition of the ecosystem.
Results Impact of Black Rats on fruits and seeds In total, we were able to test the removal of fruits or seeds in 16 species, comprising a mixture of plant families, life forms (trees, shrubs, vines) and habitats. Most species examined had a fleshy fruit. In summary, seed or fruit losses were apparent in all study species, at least at some sites. Virtually all seed losses occurred at night and there was little removal of fleshy fruits by birds from our ground based trials. It is likely that most bird removal of fruits occurs on the plants and not the ground. Occasionally a shearwater returning to its nest disturbed one of our plots and scattered fruits, but this was not recorded as fruit loss in our calculations. Evidence of seed or fruit losses to rats was indicated by:
• Camera images of rats removing or consuming fruits/seeds (see Figs 3-6) • In situ seed or fruit remains indicating rats (eg, incisor marks etc) (see Figs 7-
11) We found no clear pattern of greater loss in areas without rat baiting stations present versus those with baiting stations. Trees Howea forsteriana There was high levels of seed removal in this species in all trials (5 sites, 2 time periods, Table 1), with 5/6 trials with 100% fruit loss and 1/6 with 80% loss. Olea paniculata All four trails across 4 different sites showed 100% seed loss. Syzygium fullagarii Fruit loss was around 40-44% at 2 study sites. For both sites, the trials only ran for 2-3 days so further fruit losses could be expected. Sarcomelicope simplicifolia

5
12% fruit loss at one site (insufficient fruits available for further study). Psychotria carronis 20% fruit loss at one site (trial only ran for 2 nights so further losses may occur). Chionanthus quadristamineus 32% fruit loss at one site (trial only ran for 2 nights so further losses may occur). Dysoxylon pachyphyllum 4% fruit loss at one site (trial only ran for 2 nights so further losses may occur). Drypetes deplanchei Seed loss ranged from 8-32% across 2 sites. Only fruits with no flesh available for trials. Sophora howinsula Seed loss ranged from 0-36% across four sites. When eaten seed, seeds were consumed in situ (see Fig. 8). Baloghia inophylla 100% seed loss in all trials across 4 sites. Shrubs Ochrosia elliptica Fruit loss varied from 4-96% across sites (Table 1). One of the sites with low losses was in a location with no adult plants. The cameras showed clear evidence of rats removing fruits (Fig. 4). On other occasions fruits were eaten in situ and fruit remains where present. Coprosma putida Fruit loss varied from 0-16% across 2 study sites. Dietes robinsoniana 36% fruit loss at one site (trial only ran for 2 nights so further losses may occur). Vines Jasminium simplicifolium Total fruit loss (100%) at three sites and 64% at the fourth Smilax australis Fruit loss varied from 92-100% across four sites. Geitonoplesium cymosum 68-100% fruit loss across 2 sites.

6
Impact of Black Rats on the Little Mountain Palm (Lepidorrhachis mooreana) on Mt Gower Based on sampling in 2006, Auld et al. (2010) showed that not only were the fruits of Lepidorrhachis mooreana consumed by rats, but the recruitment and establishment of a juvenile seedling bank only occurred in the baited habitats on Mt Gower. Our re-sampling of these and other sites in 2014 found the pattern remains unchanged. Two of the 3 sites that were unbaited lacked new seedlings and small juveniles (eg. Fig. 12) and one of these showed a decline in larger juveniles over time, suggesting ongoing recruitment failure. The unbaited site nearest the baiting stations showed a small pulse of juveniles less than 50 cm in height, suggesting that that there may have been some decline in rat impacts for a period in this area. The three sites in the baited area, all showed evidence of some small juveniles being present, but the abundance of small juveniles was greatly reduced in 2014 compared to 2006 (eg see Fig. 13). For the 4 additional sites sampled, one had no juvenile plants at all, two had no seedlings (one of these sites had only 1 juvenile less than 50 cm high), while only one site had a few seedlings. There was a pattern of greater recruitment of juvenile plants closer to bait stations (Fig. 14) with recruitment of small plants declining to zero at plots sampled around 250 m from bait stations. Extinction risk for the Gnarled Mossy Cloud Forest Ecological Community Overall the Gnarled Mossy Cloud Forest is considered to be Critically Endangered based on a restricted geographic distribution combined with continuing decline. Historical Decline: There have been no observed changes in the extent of this ecosystem in the last 50 years (Pickard 1983). No changes are indicated since Pickard’s 1983 vegetation classification based on inspection of recent satellite imagery (Google Earth and 2011 air photo imagery) and field traverses. The status of the ecosystem is therefore Least Concern under criterion A1. Geographical Extent and Continuing Decline: The geographic distribution of the ecosystem is below the IUCN threshold for Critically Endangered (Auld and Leishman 2015). Decline was inferred from: a loss of moisture from declining rainfall and cloud cover due to climate change (affecting disturbance regimes, gap formation and species survival and recruitment); ongoing exotic rat predation on seeds and seedlings of several sensitive species that are structural components of the ecosystem (affecting survival and recruitment); and the fact that the ecosystem is considered to exist at only one location (Auld and Leishman 2015). Decline in abiotic processes and features: There was a trend for decline in both rainfall and the frequency of cloud cover although there is still a great deal of variation in the degree of decline predicted (Auld and Leishman 2015, see Appendix 2). Decline in biotic processes and interactions: Rats damage key functional plant species in the ecosystem via predation on seeds and seedlings (Auld & Hutton 2004; Auld et al. 2010). Most of the area (93%) of Gnarled Mossy Cloud Forest is subject to impact by rats (7% is baited to reduce impact). The

7
extent and severity of seed predation by rats is high for two key species (mountain palms, but the severity of seed predation by rats on other taxa in the ecosystem is uncertain. More work on this aspect is required. Discussion Impact of Black Rats on fruits and seeds There was evidence that rats were consuming fruits or seeds in all 16 species examined. In summary, seed or fruit losses were apparent in all study species, at least at some sites. Losses were very high for six study species (Howea forsteriana, Olea paniculata, Baloghia inophylla, Jasminium simplicifolium, Smilax australis and Geitonoplesium cymosum); potentially very high but variable for one species (Ochrosia elliptica); moderate for three species (Syzygium fullagarii, Chionanthus quadristamineus, Dietes robinsoniana) (the actual losses may be higher as the trials only ran for a short period); generally low in 4 species (Sarcomelicope simplicifolia, Psychotria carronis, Dysoxylon pachyphyllum, Coprosma putida) (but the actual losses may be higher where the trials only ran for a short period); and low-moderate in two species (Sophora howinsula, Drypetes deplanchei). Further work on examining the impact of fruit losses on the ecology of the study species would assist interpretation of these data. Does the impact of these losses result in reduced potential for recruitment of new plants in the study species as has been previously demonstrated for the Little Mountain Palm (Lepidorrhachis mooreana) and Big Mountain Palm (Hedyscepe canterburyana) (Auld et al. 2010, Simmons et al. 2012)? Given that losses are occurring in all tested species, it would also be worthwhile to extend the sampling to additional species as many other species are also likely to be impacted by rats. Impact of Black Rats on the Little Mountain Palm (Lepidorrhachis mooreana) on Mt Gower Repeat sampling in 2014 confirmed the earlier findings of Auld et al. (2010) that Black Rats are significantly impacting on Little Mountain Palms on Mt Gower. Baiting appears to be effective in allowing the development of a juvenile bank of palms, although there was evidence that the size of this juvenile bank was greatly reduced in 2014 compared to 2006. Sites within about 100m of bait stations had some seedlings and small juveniles (indicating recent recruitment), but sites 250m or further away mostly had zero recent recruitment. This suggests that to successfully maintain this critically endangered palm on Mt Gower rat control or eradication across the Mt Gower summit will be required. Extinction risk for the Gnarled Mossy Cloud Forest Ecological Community Assessment of the Gnarled Mossy Cloud Forest against the IUCN Red List for Ecosystems criteria indicated that the ecosystem is critically endangered due to its very highly restricted geographic distribution (confined to summits of Mt Gower and Mt Lidgbird) combined with continuing decline due to a loss of moisture from declining rainfall and cloud cover as a result of climate change (affecting disturbance regimes, gap formation and species survival and recruitment); ongoing exotic rat predation on seeds and seedlings of several sensitive species that are structural components of the ecosystem (affecting survival and recruitment); and the fact that

8
the ecosystem is considered to exist at only one location (see Auld and Leishman 2015, Appendix 2 for details. This supports the current listing of this ecological community as critically endangered under the NSW Threatened Species Conservation Act (NSW Scientific Committee 2011). The recognition that the introduced Black Rat is a threat to the Gnarled Mossy Cloud Forest mirrors similar threats from exotic species and climate change to other Pacific island cloud forests (Meyer et al. 2010). Eradication of rats from Lord Howe Island will reduce the immediate risk to this ecosystem, however, only global mitigation of greenhouse gases could alleviate risk from declining cloud cover and moisture availability. If rat eradication does not occur or is unsuccessful, regular rodent baiting across much of the distribution of the ecosystem, particularly on Mt Gower would be needed to alleviate the threat. This may not be a long-term solution as the rodents may gain resistance to the poisons. A reduction in rat impact is needed to both allow successful seed production and recruitment of new plants in the Gnarled Mossy Cloud Forest. Acknowledgements We thank the Lord Howe Island Board for providing support to this project along with use of the Research Facility on the island (for TA, AD and MT). The assessment of extinction risk for the Gnarled Mossy Cloud Forest was supported by an Australian Research Council Linkage Grant to TA and others (LP LP130100435) and we thank Prof. David Keith and all the participants of the Australian Centre for Ecological Analysis and Synthesis workshop for their thoughtful contributions to the development of the risk assessment for this ecosystem. References Auld TD & Hutton I (2004) Conservation issues for the vascular flora of Lord Howe Island. Cunninghamia 8, 490–500. Auld TD, Hutton I, Ooi MKJ & Denham AJ (2010) Invasive species on oceanic islands: disruption of recruitment in narrow endemic palms. Biol. Invasions 12, 3351–61. Auld TD & Leishman MR (2015) Ecosystem risk assessment for Gnarled Mossy Cloud Forest, Lord Howe Island, Australia. Austral Ecology 40, 364–372. Billing J, Harden B (2000) Control of introduced Rattus rattus L. on Lord Howe Island. I. The response of mouse populations to warfarin bait used to control rats. Wildl Res 27, 655–658. Green PS (1994) Flora of Australia Vol. 49 Oceanic Islands 1. Australian Government Printing Service, Canberra. Harris R, Cassis G, Auld T & Hutton I (2005) Floristics and structure of the mossy cloud forest of Mt Gower summit, Lord Howe Island. Pac. Conserv. Biol. 11, 246–56. Hutton, I (1986) Lord Howe Island (Conservation Press: Canberra ACT).

9
Keith DA, Rodríguez JP, Rodríguez-Clark KM et al. (2013) Scientific Foundations for an IUCN Red List of Ecosystems. PLoS ONE 8, e62111. Lowry, PP (1998) Diversity, endemism, and extinction in the flora of New Caledonia: a review. In Rare, threatened and endangered floras of Asia and the Pacific Rim (eds. C.-I Peng & P.P. Lowry II), (Institute of Botany, Academia Sinicia Monograph Series No. 16. Taipei). pp. 181–206. Maiden JH (1898) Observations of the vegetation of Lord Howe Island. Proc. Linn. Soc. N. S.W. 23, 112–58. Maiden JH (1914) Further notes on the botany of Lord Howe Island (fifth paper). Proc. Linn. Soc. N. S.W. 39, 377–84. Meyer J-Y (2010) Montane cloud forests on remote islands of Oceania: the example of French Polynesia (South Pacific Ocean). In: Tropical Montane Cloud Forests: Science for Conservation and Management (eds L. A. Bruijnzeel, F. N. Scatena & L. S. Hamilton) pp. 121–9. Cambridge University Press, Cambridge. Mueller-Dombois D & Fosberg FR (1998) Vegetation of the Tropical Pacific Islands. Springer-Verlag, NewYork. NSW Scientific Committee (2010) Final determination. Solanum bauerianum - species presumed extinct listing [Cited 9th June 2015] Available from URL: http://www.environment.nsw.gov.au/determinations/solanumbauerianumFD.htm NSW Scientific Committee (2011) Final determination. Gnarled Mossy Cloud Forest on Lord Howe Island – critically endangered ecological community listing. [Cited 23 September 2014.] Available from URL: http://www.environment.nsw.gov.au/determinations/gnarledmossycloudforestlhifd.htm Oliver WRB (1916) The vegetation and flora of Lord Howe Island. Trans. Proc. N. Z. Inst. 49, 94–161. Pickard J (1983) Vegetation of Lord Howe Island. Cunninghamia 1, 133–265. Simmons L, Auld TD, Hutton I, Baker WJ, Shapcott A (2012) Will climate change, genetic and demographic variation or rat predation pose the greatest risk for persistence of an altitudinally distributed island endemic. Biology 1(3), 736-765. Wilkinson IS & Priddel D (2011) Rodent eradication on Lord Howe Island: challenges posed by people, livestock, and threatened endemics. In: Island Invasives: Eradication and Management (eds CR Veitch, MN Clout & DR Towns) pp. 508–14. IUCN, Gland.

10
Table 1 Seed and fruit losses across species and sites. Family Scientific
Name Common Name
Life form
Experiment Type
Site Date of trial
Seeds lost to rats (%)
Fleshy Fruit species Arecaceae Howea
forsteriana Kentia Palm
Palm Single Neds Beach
Nov-13
100
Howea forsteriana
Single Research Station
Nov-13
100
Howea forsteriana
Single Windy Ridge W
Nov-13
100
Howea forsteriana
Single Windy Ridge
Nov-13
100
Howea forsteriana
Single Windy Ridge E
Nov-13
80
Howea forsteriana
Single Neds Beach
Jan-14
100
Oleaceae Olea paniculata Maulwood Tree Single Research Station
Nov-13
100
Olea paniculata Single Middle Beach
Nov-13
100
Olea paniculata Single Windy Ridge W
Nov-13
100
Olea paniculata Single Windy Ridge E
Nov-13
100
Apocynaceae Ochrosia elliptica
Red Berrywood
Shrub Single Neds Beach
Nov-13
96
Ochrosia elliptica
Single Middle Beach
Nov-13
96
Ochrosia elliptica
Single Windy Ridge W
Nov-13
12
Ochrosia elliptica
Single Windy Ridge E
Nov-13
4
Myrtaceae Syzygium fullagarii
Scalybark Tree Single Research Station
Jun-14
44
Syzygium fullagarii
Cafeteria Erskine Valley
Jun-14
40
Rubiaceae Coprosma putida
Stinkwood Shrub Single Research Station
Jun-14
0
Coprosma putida
Single Erskine Valley
Jun-14
16
Oleaceae Jasminium simplicifolium
Jasmine Vine Cafeteria Research Station
Nov-13
64
Jasminium simplicifolium
Cafeteria Middle Beach
Nov-13
100
Jasminium simplicifolium
Cafeteria Windy Ridge E
Nov-13
100
Jasminium simplicifolium
Cafeteria Windy Ridge
Nov-13
100
Smilacaceae Smilax australis Vine Cafeteria Research Station
Nov-13
92
Smilax australis Cafeteria Middle Beach
Nov-13
100
Smilax australis Cafeteria Windy Ridge E
Nov-13
92
Smilax australis Cafeteria Windy Nov- 96

11
Ridge 13
Smilacaceae Geitonoplesium cymosum
Vine Cafeteria Research Station
Nov-13
68
Geitonoplesium cymosum
Cafeteria Middle Beach
Nov-13
100
Rutaceae Sarcomelicope simplicifolia
Yellow Wood
Tree Cafeteria Research Station
Nov-13
12
Rubiaceae Psychotria carronis
Black Grape
Tree Cafeteria Erskine Valley
Jun-14
20
Iridaceae Dietes robinsoniana
Wedding lily
Herb Cafeteria Erskine Valley
Jun-14
36
Oleaceae Chionanthus quadristamineus
Blue Plum Tree Cafeteria Erskine Valley
Jun-14
32
Meliacaea Dysoxylon pachyphyllum
Island Apple
Tree Cafeteria Erskine Valley
Jun-04
4
Old Fruit with no pulp Euphorbiaceae Drypetes
deplanchei Greybark Tree Cafeteria Research
Station Nov-13
8
Cafeteria Middle Beach
Nov-13
32
Dry seeds Fabaceae Sophora
howinsula Lignum Vitae
Tree Cafeteria Research Station
Nov-13
32
Sophora howinsula
Cafeteria Middle Beach
Nov-13
36
Sophora howinsula
Cafeteria Windy Ridge E
Nov-13
0
Sophora howinsula
Cafeteria Windy Ridge
Nov-13
0
Euphorbiaceae Baloghia
inophylla Bloodwood Tree Cafeteria Neds
beach Nov-13
100
Baloghia inophylla
Cafeteria Middle Beach
Nov-13
100
Baloghia inophylla
Cafeteria Windy Ridge E
Nov-13
100
Baloghia inophylla
Cafeteria Windy Ridge
Nov-13
100

12
Fig. 1 Layout of single treatment with fully enclosed cages, uncaged fruits and cage open at each end. Ochrosia elliptica fruits at Windy Pt west.
Fig. 2. Layout of cafeteria treatment. Species visible are (L to R): Smilax, Sarcomelicope, Baloghia.

13
Fig. 3. Black Rat removing mature fruit of Howea forsteriana near Ned’s Beach (Photo Ian Hutton)
Fig. 4. Black Rat removing fruit of Ochrosia elliptica from open ended cage.

14
Fig. 5. Black Rat removing fruit of Chionanthus quadristamineus fruit in Erskine Valley.
Fig 6. Black Rat fruit damage on Howea belmoreana (photo Ian Hutton)

15
Fig.7 Black Rat damage on Chionanthus quadristamineus in Erskine Valley
Fig. 8. Black Rat damage to Sophora howinsula seeds

16
Fig. 9 Black Rat damage to Olea paniculata fruits.
Fig. 10. Black Rat damage to Baloghia seeds.
Fig. 11. Black Rat damage to Smilax fruits and seeds
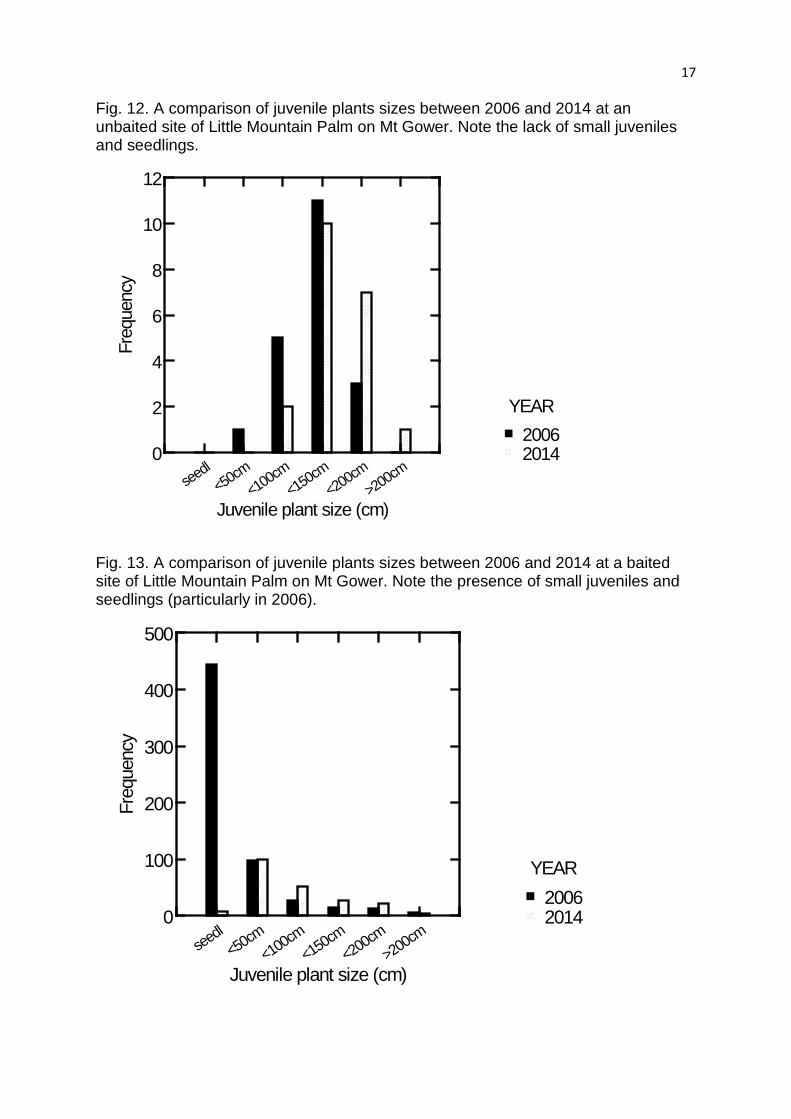
17
Fig. 12. A comparison of juvenile plants sizes between 2006 and 2014 at an unbaited site of Little Mountain Palm on Mt Gower. Note the lack of small juveniles and seedlings.
Fig. 13. A comparison of juvenile plants sizes between 2006 and 2014 at a baited site of Little Mountain Palm on Mt Gower. Note the presence of small juveniles and seedlings (particularly in 2006).
seedl
<50cm
<100cm
<150cm
<200cm
>200cm
Juvenile plant size (cm)
0
2
4
6
8
10
12
Freq
uenc
y
20142006
YEAR
seedl
<50cm
<100cm
<150cm
<200cm
>200cm
Juvenile plant size (cm)
0
100
200
300
400
500
Freq
uenc
y
20142006
YEAR

18
Fig. 14. Abundance of seedlings (black circles) and small juveniles (black triangles) of Little Mountain Palms in relation to distance from nearest bait station on Mt Gower.
0 100 200 300 400 500Distance to nearest bait station (m)
0
10
20
30
40
50
60
70
Abun
danc
e
0 100 200 300 400 500Distance to nearest bait station (m)
0
50
100
150
Abun
danc
e

19
Appendix 1 Auld TD, Hutton I, Ooi MKJ and Denham AJ (2010) Disruption of recruitment in two endemic palms on Lord Howe Island by invasive rats. Biological Invasions 12, 3351-61.

ORIGINAL PAPER
Disruption of recruitment in two endemic palmson Lord Howe Island by invasive rats
Tony D. Auld • Ian Hutton • Mark K. J. Ooi •
Andrew J. Denham
Received: 12 November 2009 / Accepted: 15 February 2010 / Published online: 6 March 2010
� Springer Science+Business Media B.V. 2010
Abstract Invasive species may have negative
impacts on many narrow range endemics and species
restricted to oceanic islands. Predicting recent
impacts of invasive species on long-lived trees is
difficult because the presence of adult plants may
mask population changes. We examined the impact
of introduced black rats (Rattus rattus) on two
palm species restricted to cloud forests and endemic
to Lord Howe Island, a small oceanic island in
the southern Pacific. We combined estimates of the
standing size distribution of these palms with the
proximal impacts of rats on fruit survival in areas
baited to control rats and in unbaited areas. The size
distribution of palms with trunks was comparable
across baited and unbaited sites. Small juvenile palms
lacking a trunk (\50 cm tall) were abundant in baited
areas, but rare in unbaited sites for Lepidorrhachis
mooreana, and rare or absent in 3 out of 4 unbaited
Hedyscepe canterburyana sites. All ripe fruits were
lost to rats in the small fruited L. mooreana. Fruit
removal was widespread but less (20–54%) in
H. canterburyana. Both palms showed evidence of
a reduced capacity to maintain a juvenile bank of
palms through regular recruitment as a consequence
of over 90 years of rat impact. This will limit the
ability of these species to take advantage of episodic
canopy gaps. Baiting for rat control reduced fruit
losses and resulted in the re-establishment of a
juvenile palm bank. Conservation of both endemic
palms necessitates control (or eradication) of rat
populations on the unique cloud forest summits of the
island.
Keywords Seed predation � Lord Howe Island �Australia � Palms � Black rat Rattus rattus
Introduction
Invasive species disrupt a range of ecological habitats
via direct impacts on individuals (either as adults or
juveniles) or by altering habitats, ecological pro-
cesses (Soule 1990; Lonsdale 1999) or disturbance
regimes (Mack and D’Antonio 1998). Invasive spe-
cies are generally competitors, predators, herbivores
or pathogens. Endemic populations of plants and
animals on oceanic islands often comprise few
individuals, occupy highly restricted areas or specia-
lised habitats and cannot recolonise from other
areas. These populations are particularly sensitive to
T. D. Auld (&) � A. J. Denham
Climate Change Science, Department of Environment,
Climate Change and Water (NSW), PO Box 1967,
Hurstville, NSW 2220, Australia
e-mail: [email protected]
I. Hutton
PO Box 157, Lord Howe Island, NSW 2898, Australia
M. K. J. Ooi
Department of Animal and Plant Sciences, University
of Sheffield, Sheffield S10 2TN, UK
123
Biol Invasions (2010) 12:3351–3361
DOI 10.1007/s10530-010-9728-5

declines or extinction caused by invasive species
(Sakai et al. 2002; Coote and Loeve 2003; Wiles et al.
2003; Blackburn et al. 2004; Towns et al. 2006;
Trevino et al. 2007), often in combination with loss
of habitat (Hunt 2007; Athens 2009).
In parts of the Pacific, much of the flora and fauna
have evolved in the absence of mammalian predators
and grazers. The subsequent introduction of such
predators, including humans, has led to a number of
serious declines or extinctions of species, including
birds (Hindwood 1940; Holdaway 1989; Blackburn
et al. 2004; Cheke 2006), invertebrates (Priddel et al.
2003; Coote and Loeve 2003; Towns et al. 2006) and
plants (Duncan and Young 2000; Campbell and
Atkinson 2002). For long-lived plants, extinctions
caused by introduced species have rarely been
recorded (Prebble and Dowe 2008), and in some
cases the life span of individual plants may be longer
than the period since the introduction of particular
weeds, predators or pathogens. For such plants,
continuing impacts may result in population declines,
even if these declines are hard to detect. Janzen
(1986) described such situations as blurry catastro-
phes and such species may be candidates for the
‘‘living dead’’ (sensu Janzen 1986) where a key part
of the life history has been disrupted and successful
reproduction has ceased. To identify recent impacts
of invasive species on long-lived plants we need to
determine if reproduction is sufficient to maintain a
stable population.
Past changes to population dynamics in plant
species have been inferred using paeleoenvironmen-
tal records (Hunt 2007; Athens 2009) or standing size
distributions across a range of plant communities
from arid shrublands (Crisp and Lange 1976; Walker
et al. 1986) to rainforests (Ogden 1985), including
palms (Enright 1985) and other species on oceanic
islands (Allen et al. 1994). To successfully apply
these techniques, an understanding of proximal
population dynamics is also needed. Assumptions
about growth rates or turnover in size or stage classes
must be made or estimated from direct observation
(Enright 1985; Ogden 1985). Proximate factors
limiting fecundity and survival, and the life or size
stages at which they act, need to be understood to
permit effective conservation management.
In this paper, we assess the impact of introduced
black rats (Rattus rattus) on two highly restricted
endemic palm species (Arecaceae) from Lord Howe
Island in the southern Pacific Ocean. Palms are a
useful study group as they are readily counted
and age classes may be inferred from leaf scars
(Tomlinson 1979). We inferred past impacts on
populations by examining standing size structures in
combination with some limited data on leaf produc-
tion. Concurrently, we estimated the proximal impact
of seed predation by rats across habitats either baited
(to reduce rat numbers) or unbaited. We applied
selective caging experiments to exclude vertebrates
from gaining access to seeds on the ground. Finally,
we combined these two approaches to infer the
conservation implication of the invasion of rats on the
endemic palm populations and the likely response of
these species to rat removal.
Methods
Study site
Lord Howe Island (31�300S, 159�050E) is a small
oceanic island (0.3–2.8 km wide and 11 km long,
1,455 ha) formed from volcanic activity 6.3–7 million
years ago (Hutton et al. 2007). It is situated *570 km
from the east coast of Australia, 1,350 km from New
Zealand and 1,250 km from New Caledonia (Pickard
1983a). Almost 80% of the island is protected in a
permanent park preserve (Davey 1986). It has been a
UNESCO World Heritage Area since 1982 (Depart-
ment of Environment and Climate Change NSW
2007). Topography on the island is dominated by its
two southern mountains (Mt Lidgbird at 777 m and
Mt Gower at 875 m). Both mountains are basaltic in
origin and Mt Gower is considered to be an eroded
caldera (McDougall et al. 1981).
The Lord Howe Island climate is humid-subtrop-
ical. It has a mean annual temperature of 19.2�C,
ranging from 17 to 25�C in December–February to
14–18�C in June–August (Mueller-Dumbois and
Fosberg 1998). Average annual rainfall in the low-
lands is 1,645 mm (Australian Bureau of Meterology),
although rainfall in the southern mountains is likely
to be much higher (Department of Environment and
Climate Change NSW 2007). Rainfall is unevenly
distributed throughout the year, with the driest period
in late summer (February, average of 100 mm)
and the wettest in winter (June/July average of
200 mm).
3352 T. D. Auld et al.
123

The island was originally free of mammals (except
for small insectivorous bats) when first encountered by
humans in 1788 and first settled in 1834 (Hutton 1986).
It now has a resident population of about 300 people
(Hutton et al. 2007). There is no archaeological
evidence of Polynesian occupation of the island prior
to 1788 (Anderson 2003) and the Pacific rat (Rattus
exulans) is not considered to have ever reached the
island. Black rats (Rattus rattus) were accidentally
introduced when a steamship ran aground in 1918
(Billing and Harden 2000). Rat control measures have
been sporadic until 1986 when a number of baited
areas were established and ongoing baiting was
maintained (Billing and Harden 2000). Approximately
8% of the island is currently baited (Billing 2000). This
rat control program is now part of an integrated
management plan for biodiversity conservation on
Lord Howe Island (Department of Environment and
Climate Change NSW 2007). The primary aim of the
baiting has been to reduce seed predation by rats upon
the two species of the endemic lowland palm genus
Howea, especially H. forsteriana which forms the
basis of a commercial seedling export enterprise on the
island for horticultural trade (Pickard 1980). The
baiting program also benefits other species, including
seabirds (McAllan et al. 2004), invertebrates (Depart-
ment of Environment and Climate Change NSW 2007)
and plants (Auld and Hutton 2004).
Study species and habitat
We sampled the two endemic mountain palm species
from Lord Howe Island. Lepidorrhachis mooreana
(F. Muell.) O.F. Cook (Arecoideae: Areceae;
Dransfield et al. 2008) is a small palm (trunks
generally up to 3 m) restricted to the cloud forest
vegetation (Pickard 1983a; Harris et al. 2005; Baker
and Hutton 2006) on the 27 Ha summit plateau of Mt
Gower (about 700–875 m in elevation). It has also
been reported from the very small summit of Mt
Lidgbird, but this requires confirmation. The species
is considered to be rare on Lord Howe Island (Pickard
1983b), with introduced rats and climate change
considered to threaten the cloud forest habitat of the
species (Baker and Hutton 2006). Hedyscepe canter-
buryana (C. Moore and F. Muell.) H Wend. & Drude
has a trunk of up to 6 m and is generally confined to
the higher elevations (600–875 m) of the southern
mountains, although on sheltered slopes it can be
found down to 300 m. It is listed as vulnerable under
the IUCN red list of threatened trees. It co-occurs
with L. mooreana on the summit of Mt Gower, but
also occurs at lower elevations in forests on several
extensive ledges below the summits of both Mt
Gower and Mt Lidgbird (Pickard 1983a). Since the
two palm species belong to monospecific genera we
hereafter refer to them as Hedyscepe and
Lepidorrhachis.
The size and dry weight (air dried at room
temperature) of fruits of the species are very differ-
ent. Lepidorrhachis fruits are globular drupes
*1.3 cm in diameter, weighing an average of
1.4 ± 0.04 g (n = 20 fruits). Fruits of Hedyscepe
are large and ovoid, *3 9 4.4 cm, weighing 24.8
± 0.5 g (n = 30 fruits).
Standing size structure
We sampled the size structure of stands of the two
species in cloud forest on the Mount Gower plateau.
We stratified the sampling between areas that have
been baited by the Lord Howe Island Board to control
rat numbers and those that have never been baited.
Baiting currently involves placement of poison every
3 months at fixed stations throughout the baited area.
Baiting has been conducted on Mt Gower for the last
20 years (Billing 2000). Only a small proportion
(*7%) of the eastern area of the summit has been
baited (1.9 ha of a total 27 ha) and not all the baited
or unbaited habitat is occupied by the study species.
Although both mountain palms occur across the
summit plateau and Hedyscepe and Lepidorrhachis
co-occur in places, there is a tendency for Hedyscepe
to occupy more sheltered sites and gully slopes while
Lepidorrhachis is more common on small ridge tops.
In each of the baited and unbaited areas we sampled
three separate plots for each palm species (12 plots
overall). At each plot, we established a 5 m wide
transect across the site and sampled all individual
palms within the transect. The transects were sampled
until we had encountered a minimum of 30 Hedy-
scepe or 50 Lepidorrhachis individuals with an
emergent trunk. Hence, the number of juvenile plants
without a trunk that were sampled varied between
plots. Transects ran for a minimum of 20 m up to
35 m, but occasionally to obtain sufficient numbers
of plants with trunks we had to sample additional
trunked plants beyond this length. These latter plants
Disruption of recruitment in two endemic palms on Lord Howe Island 3353
123

were not included in our estimates of plant densities.
We measured canopy height (=leaf height in plants
lacking a trunk) for all individuals. For plants with
emergent trunks, we also scored flowering or fruiting
and measured two additional plant size attributes:
1. trunk height to the base of the leaf sheaths; and
2. the number of leaf scars on the trunk.
Occasionally, trunks were covered in mosses and
lichens and it was not possible to count the number of
leaf scars on the entire trunk. Where this occurred,
mainly in Hedyscepe, we counted a section of the
trunk and extrapolated to the length of the trunk. This
assumed even rates of growth between leaf scars. In
Hedyscepe and Lepidorrhachis, there was a pattern of
larger intervals between leaf scars at the base of
trunks and smaller intervals at the top (after the plants
had matured). Consequently, where we had to
subsample a trunk, we chose a section that included
the transition between smaller and larger intervals
between leaf scars. We also calculated the average
leaf scar interval per trunked plant from the stem
length divided by number of leaf scars.
We additionally sampled one population of Hedy-
scepe at the summit of Mt Lidgbird. The summit of
this mountain is very narrow and there is little
available habitat. The habitat of Hedyscepe palms on
this mountain top has not been subject to rat baiting.
Sufficient habitat was available to sample one plot,
where we ran a 5 m transect along the undulating
summit and sampled as per above. It was not possible
to sample Lepidorrhachis on Mt Lidgbird as no plants
could be located.
Hedyscepe also occurs on large ledges below the
summits of Mt Lidgbird and Mt Gower. However, to
ensure that all our samples were confined to compa-
rable habitat on the mountain summits in cloud forest
vegetation, we did not sample in these areas.
Inferring plant age or growth rates
For Lepidorrhachis, we tagged leaf bases on 10
mature trunked plants to determine the rate of
production of leaf scars over a 31 month period.
The length of time a palm had a trunk was then
inferred from the mean number of leaves produced
per year times the number of leaf scars. All plants
sampled were on ridges on the summit of Mt Gower
across the gradient in elevation. Loss of tags from
Hedyscepe plants prevented a similar estimation in
that species.
Impact of rats on fruit survival
To examine the direct proximal impacts of rats on the
palms we established seed predation exclusion exper-
iments on Mt Gower. We ran the experiments on
2 occasions in 2006 (winter–July; and late spring–
November). This replication allowed us to examine
temporal variation in seed predation rates across the
long period of fruit maturation. For both replicate
trials, there were developing and ripe fruits available
on mature plants of both species. For Lepidorrhachis,
there were considerably more ripe fruits in July than
November, while for Hedyscepe, slightly more ripe
fruits were available in November. At each time, we
sampled 4 sites per species, 2 in rat baited and 2 in
unbaited areas. At each site, we established 10
randomised plots of treatments, with the location of
each plot constrained by being under a palm with a
fruiting infructescence, but such plants were chosen
randomly from those available at the site. For Hedy-
scepe, at each chosen palm, we set up 2 batches of 10
mature fruits in a cache, one caged and one uncaged.
The caged fruits were covered with a 1 cm2 wire mesh
cage that was held into place by wire stakes. This
excluded rodents and birds. We followed the fates of
fruits over a 2 week period. For Lepidorrhachis, we
observed that green fruits were eaten by rats. In some
baited areas, fruits, in which the red fleshy outer layer
(the mesocarp and exocarp) had been removed or worn
off, persisted on the forest floor (leaving only the seed
and endocarp). So for Lepidorrhachis, we used caged
and uncaged ripe red fruits along with two additional
uncaged fruit categories at each plot. These categories
were large green fruit and endocarp-only fruit. We
observed Lord Howe currawongs (Strepera graculina
crissalis, a diurnal corvid) remove ripe fruits from
infructescences of Lepidorrhachis and regurgitate
endocarps. As this bird may have removed fruits in
our exclusion trials, the inclusion of endocarp-only
fruits in the experiment allowed us to compare
removal rates for fruits that were attractive to these
birds (ripe red fruits) and fruits that were not (endo-
carp-only). The Lord Howe currawongs may occa-
sionally handle the much larger fruits of Hedyscepe
(Hutton pers observ.), but no endocarp-only fruits
were available on the summit plateau.
3354 T. D. Auld et al.
123

In the July trial, the experiments were checked
overnight and again at *2 weeks after establishment,
with the final data point used in the analyses. In the
November trial, we checked removal and fruit loss at
plots overnight, at 3–3.5 days and after 2 weeks. We
used these temporal sampling data to examine the early
rates of fruit loss.
To infer the fates of fruits we explored several
possibilities. We spent time observing fruit caches in
the day and night to monitor removal agents. We also
examined the pattern of fruit mesocarp removal and
fruit consumption in situ to infer likely agents.
Data analysis
We compared the population size structure in stands of
each palm across baited and unbaited areas. For each
species, we used nested 2 Factor ANOVAs (fixed
factor rat baiting, with sites nested in rat baiting) to
compare each size attribute in trunked plants. Species
were not included as a factor as Hedyscepe is much
larger than Lepidorrhachis and as we were interested in
the nested site effect within a species. Cochran’s Test
was used to test for homogeneity of variances. Where
heterogeneous variances were detected, the data were
log transformed (Underwood 1997). Where ANOVAs
were significant, individual means were compared
using Student–Newman–Keuls tests (Zar 1974).
For the predator exclusion experiment, for each
species, we compared the magnitude of fruit removal/
loss across the treatment plots. We used a GLIM with
four factors—time of experiment, rat baiting, caging
treatment and sites (nested) with a binomial error
structure. Removal from a plot was scored if there
had been any seed removal as the plot was the unit of
replication. In the November trial, we used a failure-
time analysis (Fox 2001) to compare the rates of seed
removal between baited and unbaited areas and
different caging treatments, pooling data across plots
and sites.
Results
Standing size structure
In trunked palms of both Hedyscepe and Lepidorrhachis,
there was no significant difference between baited and
unbaited areas for all size attributes, but there was
variation among study sites (Table 1). In Hedyscepe,
there was a significant difference in crown sizes
(F4,135 = 3.3, P = 0.01), trunk lengths (F4,177 = 5.4,
P \ 0.001) and the number of leaf scars per trunk
(F4,170 = 2.7, P = 0.03) across sites (Table 1). In
Lepidorrhachis, there was significant variation among
sites in crown sizes (F4,211 = 3.5, P = 0.001) and
trunk lengths (F4,294 = 2.8, P = 0.025).
The size distribution of trunked plants was broadly
similar across sites, although in Hedyscepe there were
relatively more individuals in the largest size classes
than in Lepidorrhachis (Figs. 1, 2). When plants
without trunks were compared, there were clear
differences between the population structures in the
rat baited versus unbaited areas (Figs. 1, 2). Small
juvenile palms (\25 cm) were only present in areas
that were baited, with the exception of one Hedyscepe
site (H2rat; Fig. 1). Large juveniles that did not yet
have an aerial trunk and plants with a trunk that had
clearly not yet flowered were present at all sites,
although they were very scarce at one Hedyscepe site
where rats were not baited (H1rat, Table 2). The
Hedyscepe site from the summit of Mt Lidgbird was
comparable to the unbaited Mt Gower sites as there
were few plants without a trunk (Table 2). Densities
of mature (0.06–0.4 m-2) and juvenile plants (0.05–
8.3 m-2) varied across sites (Table 2).
Trunked Lepidorrhachis plants produced an aver-
age of 2.6 (±0.8) leaves each year (range 2.3–3.1).
To infer an estimate of the time spent as a trunked
Lepidorrhachis palm, we assumed plants produced
between 2 and 3 leaf scars per year. This would
suggest that each site contains a few very old
individuals that have had trunks for 100–200 years
(cf. Fig. 2). Most of the plants are estimated to have
had trunks for less than 70 years.
Impact of rats on fruit survival
Fruit loss varied markedly between baited and
unbaited areas. For Hedyscepe, there was no signif-
icant difference between the replicate experiment
times and no significant interactions involving this
term. No fruits were consumed in situ in this species.
There were significant differences between baited and
unbaited areas and between caging treatments, along
with significant site variation. Some fruits were
removed from all plots at one of the unbaited sites
Disruption of recruitment in two endemic palms on Lord Howe Island 3355
123

(a total 54% of individual fruits removed) and from
85% of plots at the other unbaited site (a total 20% of
individual fruits removed). One of the baited sites
also had some fruit removed from 85% of plots
(a total 22% of individual fruits removed), while the
second baited site had removal from 45% of plots
(a total 7% of individual fruits removed). There was
evidence of rats scraping away the thin mesocarp and
trying to remove the fibrous endocarp. Nocturnal
observations revealed that rats dragged fruits away
from the experimental plots. It is likely that fruits
were consumed in caches and such caches with eaten
fruits were observed in the habitat.
In Lepidorrhachis, there was a significant two way
interaction between caging treatment and date of
sampling (July or November), along with a signifi-
cant difference between baited and unbaited areas
and a significant nested site effect. In the unbaited
areas, uncaged fruits were removed in all plots,
except for 2 plots of green fruits. All red fruits and
those with the flesh removed (endocarp-only) were
taken in both July and November sampling. The
removal of green fruits varied from 98% in July to
17% in November. Evidence that rats consumed
fruits was indicated by the presence of chewed pieces
of pericarps in situ, the presence of fresh rat scats in
the plots and our nocturnal observations of rats
feeding on fruits. While some ripe red fruits may
have been taken and dispersed by Lord Howe
currawongs, it is likely that rats are consuming fruits
since there was a loss of both ripe red fruits and
endocarp-only fruits in all the unbaited sites. Fruits
with the mesocarp removed should not be attractive
to currawongs. At these unbaited sites, no adult plants
had ripe fruits on infructescences and no fruits with
the mesocarp removed (endocarp-only) were present
on the ground. In contrast, ripe fruits were common
both on infructescences and on the ground in the
baited areas. In baited areas, we found that there was
some removal of ripe red fruits in all plots in
November (a total 64–94% of individual fruits
removed across sites) and in 85% of plots in July
(a total 35–40% of individual fruits removed across
sites). For fruits with only an endocarp, there was
more variation between the two sampling seasons,
with removal in 95% of plots in November (a total
24–96% of individual fruits removed across sites)
compared to only 30% of plots in July (a total 6% of
individual fruits removed across sites). Removal of
green fruits was comparable across sampling seasons,
65% of plots in November (a total 14–25% of
individual fruits removed across sites) and 55% of
plots in July (a total 12–13% of individual fruits
removed across sites). The losses in November at one
baited site were directly attributed to rats as evi-
denced by large amounts of fruit remains in situ. This
Table 1 Summary size measurements for trunked palms sampled on Lord Howe Island
Species Site Baited for rats Trunk Length (m) Mean plant (±SE)
Crown size (m) Leaf scars Leaf scar
interval (cm)
Hedyscepe H1rat No 4.01(0.34) 2.08(0.19) 193(26) 2.7(0.3)
H2rat No 2.73(0.33) 1.98(0.10) 138(32) 3.4(0.3)
H3rat No 2.05(0.27) 2.38(0.10) 87(22) 4.2(0.5)
H4 Yes 2.63(0.33) 2.43(0.10) 115(25) 4.1(0.4)
H5 Yes 2.39(0.35) 2.38(0.13) 148(34) 4.5(0.5)
H6 Yes 3.03(0.32) 2.03(0.09) 208(39) 3.1(0.4)
HLrat No 2.79(0.21) 1.89(0.07) 170(14) 2.3(0.2)
Lepidorrhachis L1rat No 1.37(0.14) 1.73(0.05) 75(9) 2.1(0.2)
L2rat No 1.67(0.15) 1.85(0.08) 68(9) 2.5(0.2)
L3rat No 1.59(0.12) 1.63(0.07) 70(8) 2.4(0.1)
L4 Yes 1.73(0.12) 2.02(0.06) 76(9) 2.8(0.2)
L5 Yes 1.21(0.13) 1.95(0.08) 53(9) 2.9(0.3)
L6 Yes 1.46(0.11) 1.80(0.05) 75(9) 2.3(0.1)
For each size measurement 30 Hedyscpe or 50 Lepidorrhachis trunked plants were sampled
3356 T. D. Auld et al.
123

may reflect re-colonisation of this site by rats at the
time.
In the November trial, there was some initial
(overnight) rapid removal of ripe fruits of Hedyscepe
in unbaited areas and then a slow rate of removal in
both baited and unbaited areas (Fig. 3). In Lep-
idorrhachis, loss of ripe red fruits was rapid where
rats were not baited (Fig. 4), with some 90% fruits
lost within a day and all fruits gone within 3 days.
Most of these fruits were consumed in situ by rats
over the first night of the experiment, again support-
ing the conclusion that consumption by rats rather
than fruit removal and dispersal by Lord Howe
currawongs explains fruit losses. The loss of all
endocarp-only fruits within 3 days was also likely
due to rat predation since these fruits are not
attractive to currawongs (Fig. 4).
Discussion
The introduction of rats (Rattus rattus) to the oceanic
Lord Howe Island is likely to have increased the risk
of extinction for the two endemic mountain palms.
This is a consequence of rat predation of fruits which
has the potential to limit recruitment in both palm
species. Past observations highlight the lack of ripe
fruits on Lepidorrhachis plants unless mesh caging
was applied to exclude rats from developing fruits
(Moore 1966; Pickard 1980). The paucity of small
0 100 200 3000
10
20
30
0 100 200 300 400 500 600 7000
10
20
30Fr
eque
ncy
Leaf Height (cm) Number of Leaf scars
0 100 200 3000
100
200
300
0 100 200 300 400 500 600 7000
10
20
30
Freq
uenc
y
Leaf Height (cm) Number of Leaf scars
0 100 200 3000
100
200
300
0 100 200 300 400 500 600 7000
10
20
30
Freq
uenc
y
Leaf Height (cm) Number of Leaf scars0 100 200 300
0
100
200
300
0 100 200 300 400 500 600 7000
10
20
30
Freq
uenc
yLeaf Height (cm) Number of Leaf scars
0 100 200 3000
10
20
30
0 100 200 300 400 500 600 7000
10
20
30
Freq
uenc
y
Leaf Height (cm) Number of Leaf scars0 100 200 300
0
100
200
300
0 100 200 300 400 500 600 7000
10
20
30
Freq
uenc
y
Leaf Height (cm) Number of Leaf scars
H1rat
H2rat
H3rat
H4
H5
H6
Fig. 1 Size distributions for Hedyscepe canterburyana of plant height for juvenile plants lacking a trunk (open bars) and the number
of leaf scars in trunked individuals (filled bars). rat No rat baiting conducted at the site. Note: Y axis scale varies across sites
Disruption of recruitment in two endemic palms on Lord Howe Island 3357
123

juvenile plants in areas which have not been baited
for rat control, both in our study plots and more
generally throughout this unbaited area, supports the
more general contention that the introduction of rats
affects plant recruitment in a range of island ecosys-
tems (Campbell and Atkinson 2002; Delgado Garcıa
2002; Towns et al. 2006; Meyer and Butaud 2009;
Traveset et al. 2009). It is possible that rats also
reduce seedling and juvenile growth as they have
been observed eating palm leaf sheaths and causing
leaf fall in both juvenile and adult palms consistent
with similar damage observed on plants elsewhere
(Traveset et al. 2009). The impact of rats is greatest in
Lepidorrhachis, where fruit losses reached 100% and
small juvenile plants (\50 cm) were extremely rare
in the presence of rats. Any possible seed escape in
Lepidorrhachis via dispersal by the Lord Howe
currawong is negated by the loss of endocarp-only
fruits (matching regurgitated fruits) in unbaited areas.
For Hedyscepe, observed fruit losses were less (total
of 20–54%), and while some seed escape may be
occurring, only one out of four sites sampled on the
mountain tops has an appreciable number of small
juveniles (\50 cm; Table 2). Differences between
the two palm species may relate to the fact that the
fruits of Hedyscepe are up to 18 times bigger than
Lepidorrhachis and rats may have more difficulty
eating into Hedyscepe fruits as they have a thicker
and very fibrous endocarp wall. Neither palm species
can be considered the ‘‘living dead’’ (sensu Janzen
0 100 200 3000
10
20
30
0 100 200 300 400 5000
10
20
30Fr
eque
ncy
Leaf Height (cm) Number of Leaf scars0 100 200 300
0
100
200
300
400
500
600
700
0 100 200 300 400 5000
10
20
30
Freq
uenc
y
Leaf Height (cm) Number of Leaf scars
0 100 200 3000
10
20
30
0 100 200 300 400 5000
10
20
30
Freq
uenc
y
Leaf Height (cm) Number of Leaf scars0 100 200 300
0
100
200
300
400
0 100 200 300 400 5000
10
20
30
Freq
uenc
y
Leaf Height (cm) Number of Leaf scars
0 100 200 3000
10
20
30
0 100 200 300 400 5000
10
20
30
Freq
uenc
y
Leaf Height (cm) Number of Leaf scars0 100 200 300
0
100
200
300
400
0 100 200 300 400 5000
10
20
30
Freq
uenc
y
Leaf Height (cm) Number of Leaf scars
L1rat
L2rat
L3rat
L4
L5
L6
Fig. 2 Size distributions for Lepidorrhachis mooreana of plant height for juvenile plants lacking a trunk (open bars) and the number
of leaf scars in trunked individuals (filled bars). rat No rat baiting conducted at the site. Note: Y axis scale varies across sites
3358 T. D. Auld et al.
123

1986) as there are juvenile plants over 50 cm in
height at all sites, independent of the current rat
baiting program on the island. Some episodic
recruitment may occur as a result of sporadic seed
escape from rats, possibly in relation to natural
fluctuations of abundance within the rat population.
Rats reached Lord Howe Island some 90 years
prior to this study and are reported to have spread
rapidly (Billing and Harden 2000). Consequently, rat
predation could have affected palm recruitment for a
number of decades. Is this reflected in the estimated
Table 2 Plant densities across sampled plots for Lepidorrhachis mooreana and Hedyscepe canterburyana
Species Site Baited for rats Density (m-2)
Trunked plants Juveniles with no trunks
Mature Juv Height (m)
\0.5 0.5–1 [1
Hedyscepe H1rat No 0.10 0.02 0 0.01 0.01
H2rat No 0.13 0.02 0.85 0.01 0.03
H3rat No 0.11 0.14 0.03 0.03 0.18
H4 Yes 0.14 0.07 4.02 0.56 0.19
H5 Yes 0.06 0.09 1.52 0.07 0.06
H6 Yes 0.08 0.03 2.91 0.11 0.03
HLrat No 0.40 0.11 0.01 0.02 0.04
Lepidorrhachis L1rat No 0.34 0.06 0.04 0.09 0.10
L2rat No 0.24 0.06 0.01 0.09 0.15
L3rat No 0.35 0.04 0.01 0.04 0.11
L4 Yes 0.19 0 8.13 0.13 0.08
L5 Yes 0.12 0.10 2.38 0.19 0.11
L6 Yes 0.25 0.04 5.41 0.28 0.14
All sites on Mt Gower, except HLrat from Mt Lidgbird
0 5 10 15 20DAY
0.0
0.1
0.2
0.3
0.4
0.5
Frui
t rem
oval
(pr
op.)
Fig. 3 Rate of fruit loss in Hedyscepe canterburyana, pooled
across sites. Data are 1-(Kaplan–Meier estimates) with
standard errors. Filled symbols are sites with no rat baiting,
symbols are sites with rat baiting. Vertical bars are standard
errors. Treatments are: Cage (circle); uncaged ripe fruit
(triangle). No fruits were removed in the caged treatment
0 5 10 15 20DAY
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Frui
t rem
oval
(pr
op.)
Fig. 4 Rate of fruit loss in Lepidorrhachis mooreana, pooled
across sites. Data are 1-(Kaplan–Meier estimates) with
standard errors. Filled symbols are sites with no rat baiting,
open symbols are sites with rat baiting. Vertical bars are
standard errors. Treatments are: Cage (circle); uncaged ripe
fruit (triangle); uncaged green fruit (diamond) uncaged
endocarp-only fruit (square)
Disruption of recruitment in two endemic palms on Lord Howe Island 3359
123

age structure of Lepidorrhachis? Plant age in palms
has been inferred by the number of leaf scars
(Sarukhan 1980; Enright and Watson 1992). In
Lepidorrhachis, there were few plants (*6% of the
population) with more than 120 leaf scars (Fig. 2).
This represents some 40–60 years of trunk growth.
The remainder of the trunked palms are more or less
evenly distributed up to 100 or 120 leaf scars (Fig. 2).
If individual mortality was constant across these size
classes then this would suggest a declining population
and may reflect a long term impact of rats on
recruitment. Estimating plant age requires additional
data on the time required for juvenile plants to grow
large enough to produce a trunk, and this may vary
with habitat quality and disturbance regime. This pre-
trunk period typically takes decades in some palm
species (e.g. 35–60 years, Enright 1985; Enright and
Watson 1992), but is currently unknown for Lep-
idorrhachis. Further data on juvenile survival and
growth are needed to clarify the age structure of the
population.
Recovery of vegetation following removal of rats
is not always straightforward (Mulder et al. 2009;
Towns 2009). Rats may impact on nutrient cycling
(through impacts on seabirds, Mulder et al. 2009) and
the impacts of mice may increase if rats alone are
removed (Angel et al. 2009). In our study area on Mt
Gower, there has only been limited rat control in the
last 20 years. There is a lack of small juveniles in
unbaited areas at all sampled Lepidorrhachis sites
and at 75% of the Hedyscepe sites. Baited areas are
distinguished by the abundance of young juvenile
palms, implying a restoration of plant recruitment
and a re-establishment of the juvenile bank of
palms (sensu Silvertown 1982). Successful growth
to maturity of juvenile palms will probably be
influenced by local canopy gap formation after tree
death or tree fall in storms.
Long-term conservation of the endemic palm
genera in this study may be compromised by a
reduced capacity to establish a juvenile bank and to
adapt to a changing climate. Both species occur in the
unique cloud forests on the island (Harris et al. 2005)
and changes to moisture regimes (e.g. through a
reduction in cloud cover) with a changing climate
may limit establishment opportunities in the future.
Targeted control, or eradication of rats is essential to
maximise the potential for long-term conservation of
these species.
Acknowledgments We thank the Lord Howe Island Board
for permission to carry out this study and for support in the use
of a research facility for TA, MO and AD. William Baker made
helpful comments on the manuscript.
References
Allen RB, Lee WG, Rance BD (1994) Regeneration in indig-
enous forest after eradication of Norway rats, Breaksea
Island, New Zealand. NZ J Bot 32:429–439
Anderson A (2003) Investigating early settlement on Lord
Howe Island. Aust Archaeol 57:98–102
Angel A, Wanless RM, Cooper J (2009) Review of impacts of the
introduced house mouse on islands in the Southern Ocean:
are mice equivalent to rats? Biol Invas 11:1743–1754
Athens JS (2009) Rattus exulans and the catastrophic disap-
pearance of Hawai’i’s native lowland forest. Biol Invas
11:1489–1501
Auld TD, Hutton I (2004) Conservation issues for the vascular
flora of Lord Howe Island. Cunninghamia 8:490–500
Baker WJ, Hutton I (2006) Lepidorrhachis. Palms 50:33–38
Billing J (2000) The control of introduced Rattus rattus L. on
Lord Howe Island. II. The status of warfarin resistance in
rats and mice. Wildl Res 27:659–661
Billing J, Harden B (2000) Control of introduced Rattus rattusL. on Lord Howe Island. I. The response of mouse pop-
ulations to warfarin bait used to control rats. Wildl Res
27:655–658
Blackburn TM, Cassey P, Duncan RP, Evans KL, Gaston KJ
(2004) Avian extinction and mammalian introductions on
oceanic islands. Science 305:1955–1958
Campbell DJ, Atkinson IAE (2002) Depression of tree
recruitment by the Pacific rat (Rattus exulans Peale) on
New Zealand’s northern offshore islands. Biol Conserv
107:19–35
Cheke AS (2006) Establishing extinction dates—the curious
case of the Dodo Raphus cucullatus and the Red Hen
Aphanapteryx bonasia. Ibis 148:155–158
Coote T, Loeve E (2003) From 61 species to five: endemic tree
snails of the Society Islands fall prey to an ill-judged
biological control programme. Oryx 37:91–96
Crisp MD, Lange RT (1976) Age structure, distribution and
survival under grazing of the arid-zone shrub Acaciaburkittii. Oikos 27:86–92
Davey A (1986) Plan of management. Lord Howe Island per-
manent park preserve. NSW National Parks and Wildlife
Service, Sydney
Delgado Garcıa JD (2002) Interaction between introduced rats
and a frugivore bird–plant system in a relict island forest.
J Nat Hist 36:1247–1258
Department of Environment and Climate Change NSW (2007)
Lord Howe Island Biodiversity Management Plan.
Department of Environment and Climate Change (NSW),
Sydney
Dransfield J, Uhl NW, Asmussen CB, Baker WJ, Harley MM,
Lewis CE (2008) Genera Palmarum. The evolution and
classification of palms. Kew Publishing, UK
Duncan RP, Young JR (2000) Determinants of plant extinction
and rarity 145 years after European settlement of Auck-
land, New Zealand. Ecology 81:3048–3061
3360 T. D. Auld et al.
123

Enright NJ (1985) Age, reproduction and biomass allocation in
Rhopalostylis sapida (Nikau palm). Aust J Ecol 10:461–
467
Enright NJ, Watson AD (1992) Population dynamics of the
Nikau palm, Rhopalostylis sapida (Wendl et Drude), in a
temperate forest remnant near Auckland, New Zealand.
NZ J Bot 30:29–43
Fox GA (2001) Failure-time analysis: Studying times to events
and rates at which events occur. In: Scheiner SM, Gu-
revitch J (eds) Design and analysis of ecological experi-
ments. Oxford University Press, Oxford, pp 235–266
Harris R, Cassis G, Auld TD, Hutton I (2005) Patterns in flo-
ristic composition and structure in the mossy cloud forest
of Mt Gower summit, Lord Howe Island. Pac Conserv
Biol 11:246–256
Hindwood KA (1940) The birds of Lord Howe Island. Emu
40:1–86
Holdaway RN (1989) New Zealand’s pre-human avifauna and
its vulnerability. NZ J Ecol 12:11–25
Hunt TL (2007) Rethinking Easter Island’s ecological catas-
trophe. J Archaeol Sci 34:485–502
Hutton I (1986) Lord Howe Island. Conservation Press, Can-
berra, ACT
Hutton I, Parkes JP, Sinclair ARE (2007) Reassembling island
ecosystems: the case of Lord Howe Island. Anim Cons
10:22–29
Janzen DH (1986) Blurry catastrophes. Oikos 47:1–2
Lonsdale WM (1999) Global patterns of plant invasions and
the concept of invasibility. Ecology 80:1522–1536
Mack MC, D’Antonio CM (1998) Impacts of biological inva-
sions on disturbance regimes. Trends Ecol Evol 13:195–
198
McAllan IAW, Curtis BR, Hutton I, Cooper RM (2004) The
birds of Lord Howe Island Group: a review of records.
Aust Field Ornithologist 21(Suppl):1–82
McDougall I, Embleton BJ, Stone DB (1981) Origin and
evolution of Lord Howe Island, southwest Pacific Ocean.
J Geol Soc Aust 28:155–176
Meyer JY, Butaud JF (2009) The impacts of rats on the
endangered native flora of French Polynesia (Pacific
Islands): drivers of plant extinction or coup de grace
species? Biol Invas 11:1569–1585
Moore HE (1966) Palm hunting around the world. IV Lord
Howe Island. Principes 10:13–21
Mueller-Dumbois D, Fosberg FR (1998) Vegetation of the
tropical Pacific islands. Springer, New York
Mulder CPH, Grant-Hoffman MN, Towns DR, Bellingham PJ,
Wardle DA, Durrett MS, Fukami T, Bonner KI (2009)
Direct and indirect effects of rats: does rat eradication
restore ecosystem functioning of New Zealand seabird
islands? Biol Invas 11:1671–1688
Ogden J (1985) An introduction to plant demography with
special reference to New Zealand trees. NZ J Bot 23:751–
772
Pickard J (1980) The palm seed industry of Lord Howe Island.
Principes 24:3–13
Pickard J (1983a) Vegetation of Lord Howe Island. Cun-
ninghamia 1:133–266
Pickard J (1983b) Rare or threatened vascular plants of Lord
Howe Island. Biol Conserv 27:125–139
Prebble M, Dowe JL (2008) The late Quaternary decline and
extinction of palms on oceanic Pacific islands. Quatern
Sci Rev 27:2546–2567
Priddel D, Carlile N, Humphrey M, Fellenberg S, Hiscox D
(2003) Rediscovery of the ‘extinct’ Lord Howe Island
stick-insect (Dryococelus australis (Montrouzier)) (Phas-
matodea) and recommendations for its conservation.
Biodiversity Conserv 12:1391–1403
Sakai AK, Wagner WL, Mehrhoff LA (2002) Patterns of
endangerment in the Hawaiian flora. Syst Biol 51:276–
302
Sarukhan J (1980) Demographic problems in tropical systems.
In: Solbrig O (ed) Demography and evolution in plant
populations. Blackwell Scientific Publications, Oxford, pp
161–188
Silvertown JW (1982) Introduction to plant population ecol-
ogy. Longman Press, London
Soule ME (1990) The onslaught of alien species, and other
challenges in the coming decades. Conserv Biol 4:233–
239
Tomlinson PB (1979) Systematics and ecology of the Palmae.
Annu Rev Ecol Syst 10:85–107
Towns DR (2009) Eradications as reverse invasions: lessons
from Pacific rat (Rattus exulans) removals on New Zealand
islands. Biol Invas 11:1719–1733
Towns DR, Atkinson IAE, Daugherty CH (2006) Have the
harmful effects of introduced rats on islands been exag-
gerated? Biol Invas 8:863–891
Traveset A, Nogales M, Alcover JA, Delgado JD, Lopez-Da-
rias M, Godoy D, Igual JM, Bover P (2009) A review on
the effects of alien rodents in the Balearic (Western
Mediterranean Sea) and Canary Islands (Eastern Atlantic
Ocean). Biol Invas 11:1653–1670
Trevino HS, Skibiel AL, Karels TJ, Dobson FS (2007) Threats
to avifauna on oceanic islands. Conserv Biol 21:125–132
Underwood AJ (1997) Experiments in ecology. Their logical
design and interpretation using analysis of variance.
Cambridge University Press, Cambridge
Walker BH, Stone L, Hendersen L, Verede M (1986) Size
structure analysis of the dominant trees in a South African
savannah. S Afr J Bot 52:397–402
Wiles GJ, Bart J, Beck RE, Aguon CF (2003) Impacts of the
brown tree snake: patterns of decline and species persis-
tence in Guam’s avifauna. Conserv Biol 17:1350–1360
Zar JH (1974) Biostatistical analysis. Prentice-Hall Inc, NJ
Disruption of recruitment in two endemic palms on Lord Howe Island 3361
123

20
Appendix 2 Auld TD, Leishman MR (2015) Ecosystem risk assessment for Gnarled Mossy Cloud Forest, Lord Howe Island, Australia. Austral Ecology 40, 364–372.

Ecosystem risk assessment for Gnarled Mossy CloudForest, Lord Howe Island, Australia
TONY D. AULD1,2* AND MICHELLE R. LEISHMAN3
1Science Division, NSW Office of Environment and Heritage, PO Box 1967, Hurstville, 2220 (Email:[email protected]), 2Centre for Ecosystem Science, University of New SouthWales,Sydney, and 3Department of Biological Sciences, Macquarie University, North Ryde, New SouthWales,Australia
Abstract Gnarled Mossy Cloud Forest is a globally unique ecosystem, combining floristic elements fromAustralia, New Zealand and New Caledonia. It is restricted to a very small area (28 ha) at elevations above 750 mon the summits of two mountains on Lord Howe Island in the Pacific Ocean, approximately 570 km off the eastcoast of Australia. Moisture derived from clouds is a key feature of the ecosystem. We assessed the conservationstatus of this ecosystem using the International Union for the Conservation of Nature Red List criteria forecosystems. There has been no historical clearing of the ecosystem, but declines (with large uncertainty bounds)were estimated for two abiotic variables that are important in maintaining the component species (cloud cover andrainfall). Overall, we found the ecosystem to be Critically Endangered based on a restricted geographic distributioncombined with continuing decline (criterion B1aii, iii, B1b, B1c and B2aii, iii, B2b, B2c). Decline was inferredfrom: a loss of moisture from declining rainfall and cloud cover due to climate change (affecting disturbanceregimes, gap formation and species survival and recruitment); ongoing exotic rat predation on seeds and seedlingsof several sensitive species that are structural components of the ecosystem (affecting survival and recruitment); andthe fact that the ecosystem is considered to exist at only one location. This mirrors similar threats from exoticspecies and climate change to other Pacific island cloud forests. Eradication of rats from Lord Howe Island willreduce the immediate risk to this ecosystem; however, only global mitigation of greenhouse gases could alleviate riskfrom declining cloud cover and moisture availability.
Key words: moisture loss, rat predation, Red List, restricted distribution.
INTRODUCTION
Risk assessment criteria for ecosystems have beendeveloped by the International Union for the Conser-vation of Nature (IUCN) (Keith et al. 2013). Thesecriteria can be used to both identify those ecosystemsmost in danger of being lost and prioritize actions forprotection and conservation management.The criteriahave elements comparable with those used in red listcriteria for species (Keith et al. 2013) and reflectbroadly similar types of criteria used in some nationallisting criteria for ecological communities (e.g. in Aus-tralia, Nicholson et al. 2014).
Ecosystems of oceanic islands comprise manyunique combinations of both flora and fauna(Mueller-Dombois & Fosberg 1998), making thempriorities for conservation management. Isolated cloudforests are scattered across remote islands of thePacific where they form highly diverse plant commu-nities with high levels of endemism (Meyer 2010).While globally, cloud forest is thought to comprise
some 12% of tropical forests (Mulligan & Burke2005), in Australia and on oceanic islands of thePacific, it is very restricted (Mueller-Dombois &Fosberg 1998; Meyer 2010; Scatena et al. 2010). Herewe assess the conservation status of a cloud forestecosystem endemic to Lord Howe Island. Lord HoweIsland is a small (11 km by 0.6–2.8 km) oceanic islandfound approximately 570 km off the east coast of Aus-tralia, 1350 km from New Zealand and 1250 km fromNew Caledonia (Auld & Hutton 2004). The islandvaries in topography because of two steep rising south-ern mountains (up to 895 m a.s.l.) and is consideredto be a remnant of a large shield volcano (McDougallet al. 1981). The climate of Lord Howe Island ishumid-subtropical.
Lord Howe Island has a mix of floristic elements frommainland Australia, New Zealand and New Caledonia(Pickard 1983; Green 1994), and its entire flora andfauna are considered to have reached the island bylong-distance dispersal (Auld & Hutton 2004). Thisdispersal trait, combined with the large topographicrelief, has led to many of the species assemblages on theisland being globally unique. A large proportion of theplant and invertebrate taxa on the island are endemic
*Corresponding author.Accepted for publication September 2014.
Austral Ecology (2015) 40, 364–372
bs_bs_banner
© 2014 The Authors doi:10.1111/aec.12202Austral Ecology © 2014 Ecological Society of Australia

(105 vascular plant species (44% of the total flora ofLord Howe Island), Green 1994; 600 invertebratespecies (approx. 37% of the total invertebrate fauna ofLord Howe Island), Cassis et al. 2003).
There have been a number of studies on the vegeta-tion of Lord Howe Island (Maiden 1898; Oliver 1916;Pickard 1983; Mueller-Dombois & Fosberg 1998).Given that plants provide a significant component ofthe species diversity as well as a habitat resource forother species, we have here interpreted Pickard’s(1983) vegetation communities as ecosystems. The27-ha summit of Mt Gower and the narrow summit ofMt Lidgbird support an endemic cloud forest knownas Gnarled Mossy Cloud Forest (Harris et al. 2005).In this paper, we assess the conservation status of theGnarled Mossy Cloud Forest ecosystem using theIUCN Red List criteria for ecosystems (Keith et al.2013) by addressing reduction in geographic distribu-tion, restricted geographic distribution, environmentaldegradation and disruption of biotic processes. Wedevelop a conceptual model of ecosystem dynamicsthat would lead to ecosystem collapse (sensu Keithet al. 2013) applicable to cloud forests globally butwith special reference to the disruption to biotic orabiotic processes most relevant to Lord Howe Island.We then used this model to quantify measures ofabiotic degradation and disruption of biotic processes.
ECOSYSTEM DESCRIPTION
Classification
Regionally, this ecosystem is classified as a CriticallyEndangered Ecological Community ‘Gnarled MossyCloud Forest on Lord Howe Island’under the New SouthWales Threatened Species Conservation Act 1995. Theecosystem is currently recognized following the vegetationclassification on Lord Howe Island by Pickard (1983) whodescribed it as Gnarled Mossy Forest. Other studiesdescribe the same ecosystem as Moss Forest (Oliver1916), Cloud Forest (Mueller-Dombois & Fosberg 1998)and Mossy Cloud Forest (Harris et al. 2005).Recent work(Harris et al. 2005) has detailed the plant species compo-sition and internal variation within the ecosystem, alongwith its conservation significance.Under the IUCN Habi-tats Classification Scheme (Version 3.1), the ecosystem isclassified as ‘1. Forest/1.9 Subtropical/Tropical MoistMontane Forest’.
Characteristic native biota
Gnarled Mossy Cloud Forest on Lord Howe Island isa closed-canopy forest of 2–8 m height (Figs 1, 2),with its composition depending on aspect and whether
it occurs on ridges or in drainage lines (Harriset al. 2005). The ecosystem occurs on the summitsof Mt Gower and Mt Lidgbird (Fig. 3). On thesummit of Mt Gower, the dominant plant species areZygogynum howeanum and Dracophyllum fitzgeraldii(Pickard 1983; Harris et al. 2005). Associated treesinclude Cryptocarya gregsonii, Elaeocarpus costatus,Leptospermum polygalifolium ssp. howense, Negriarhabdothamnoides, Pittosporum erioloma, Symplocus can-delabrum, and the palms Hedyscepe canterburyanaand Lepidorrhachis mooreana. Tree ferns (Cyatheaspp.), large tussock sedges (Machaerina insularis andGahnia howeana), ferns Blechnum fullagarii, Blechnumcontiguum, Blechnum howeanum, Grammitis wattsiand other ferns, mosses (e.g. Echinodium hispidum,Spiridens vieillardii; Brotherus & Watts 1915) andlichens (e.g. Strigula spp., Byssoloma subdiscorans,Echinoplaca spp., Gyalectidium filicinum, Porina rufula
Fig. 1. Gnarled Mossy Cloud Forest ecosystem (photo T.Auld).
Fig. 2. Summit plateau of Mt Gower with Gnarled MossyCloud Forest. The very narrow ridgetop of Mt Lidgbird canbe seen at the back, left-hand side (photo T. Auld).
CLOUD FOREST LHI 365
© 2014 The Authors doi:10.1111/aec.12202Austral Ecology © 2014 Ecological Society of Australia

and Trichothelium javanicum; McCarthy et al. 1996)are abundant. Gnarled Mossy Cloud Forest on thesummit ridgetop of Mt Lidgbird above 750 m eleva-tion is much more exposed and restricted in area com-pared with the ecosystem on Mt Gower (Pickard1983). A vegetation plot (10 m by 40 m) on thesummit of Mt Lidgbird in Gnarled Mossy CloudForest had a dominant canopy of H. canterburyana,C. gregsonii, Dysoxylum pachyphyllum, Negria rabdo-thamnoides, P. erioloma and Cyathea macarthurii, alongwith Grammitis diminuta, Carex sp., Olearia mooneyi,Rapanea myrtillina, Z. howeanum, Lordhowea insularis,Gahnia howeana, Coprosma lanceolaris, Dendrobiummoorei, Coprosma putida, Macropiper hooglandii,Microsorium scandens, Asplenium milnei, Aspleniumsurragatum, Elatostema grande, Hymenophyllum sp.(Hutton & Auld, unpubl. data, 2007).
Gnarled Mossy Cloud Forest on Lord Howe Island onlyoccurs on the summit plateau and ridgetops of Mt Gowerand Mt Lidgbird, and is readily distinguished from adja-cent ecosystems that lack the extensive development ofnon-vascular epiphytes and several endemic mountain
vascular plant species.Overall, some 86% of vascular plantspecies in theecosystemareendemic toLordHoweIsland,and some 17% are endemic to the mountain summits onthe island (Harris et al. 2005). Dracophyllum-Metrosiderosscrub (of Pickard 1983 and Mueller-Dombois & Fosberg1998) occurs on benches below the summits. Pickard(1983) notes that Gnarled Mossy Cloud Forest on LordHowe Island is floristically distinct but close toDracophyllum-Metrosideros scrub and H. canterburyanaclosed forest, neither of which are considered to be part ofthe Gnarled Mossy Cloud Forest ecosystem.
Gnarled Mossy Cloud Forest on Lord Howe Island isa key component contributing to the southern moun-tains biodiversity hotspot on Lord Howe Island(Department of Environment and Climate Change2007), particularly for plants and invertebrates. Cassiset al. (2003) found that the assemblage of terrestrialinvertebrates in the Gnarled Mossy Cloud Forestexhibits high species richness, high levels of endemismto Lord Howe Island and many species that arerestricted to the Gnarled Mossy Cloud Forest ecosys-tem. It also forms important habitat for several verte-brates (DECC 2007) including the main breedinghabitat for providence petrels (Pterodroma solandri) andrefugial breeding habitat for the Lord Howe Islandwoodhen (Tricholimnas sylverstis).
Abiotic environment
Lord Howe Island as a remnant volcanic shield isgeologically and topographically diverse (McDougallet al. 1981). The climate is temperate, and sea levelparts of the island have a mean annual temperatureof 19.2°C, ranging from 17°C–25°C in summer to14°C–18°C in winter (Mueller-Dombois & Fosberg1998). At sea level, average annual rainfall is1717 mm, with a maximum of 2886 mm and aminimum of 998 mm (Mueller-Dombois & Fosberg1998). Temperature decreases with altitude in thesouthern mountains (0.9°C for every 100 m rise inaltitude; Simmons et al. 2012). Cloud forests onPacific islands typically occur between 800 and 900 ma.s.l (Meyer 2010), and on Lord Howe Island, theGnarled Mossy Cloud Forest ecosystem occurs from750 to 875 m a.s.l. The annual rainfall in GnarledMossy Cloud Forest is thought to be much higher thanat sea level (although this has not been quantified) andspread throughout the year (DECC 2007). The twosouthern mountains (Mts Gower and Lidgbird) obtainsignificant moisture from both rainfall and directcanopy interception of cloud water (horizontal precipi-tation or cloud stripping), and their peaks are oftenshrouded in cloud (Auld & Hutton 2004). Cloudforests are characterized by increased rainfall andcooler temperatures than forest with no cloud (Jarvis &Mulligan 2011), and this is thought to also apply to theGnarled Mossy Cloud Forest ecosystem.
0 2 4 Kilometres
N
Fig. 3. Map of the two occurrences (shown as the shadedareas) of Gnarled Mossy Cloud Forest on Lord Howe Island.As existing threats are known to occur across both occur-rences, it is considered that these occurrences constitute onelocation under the definitions of the International Union forthe Conservation of Nature (IUCN) guidelines (Keith et al.2013). Source of map data: Pickard (1983).
366 T. D. AULD AND M. R. LEISHMAN
© 2014 The Authorsdoi:10.1111/aec.12202Austral Ecology © 2014 Ecological Society of Australia

Distribution
Gnarled Mossy Cloud Forest is confined to LordHowe Island in New South Wales, Australia. On theisland, it is restricted to the summit plateau of MtGower (some 27 ha) and in a greatly reduced formand extent on the narrow summit ridge of Mt Lidgbird(estimated to be less than 1 ha) (Pickard 1983)(Figs 2, 3). Gnarled Mossy Cloud Forest occurs above750 m in elevation (Pickard 1983).
Key processes, interactions and threats
The ecosystem is sustained by a combination of cloudcover allowing moisture stripping by component species,relatively cool temperatures at >750 m elevation and anocean-moderated climate. This cloud layer provides asource of precipitation (occult precipitation) and main-tains the humidity required by about 86% of the island’sendemic plant species including the profusion of epiphytesin the Gnarled Mossy Cloud Forest (Australian NationalUniversity 2009). Ecosystem dynamics that could lead tocollapse of the Gnarled Mossy Cloud Forest depend ontwo key ecological processes.These are likely increases infrequencyandextentofdisturbance regimesunderclimatechange (e.g. through increased storm damage) and areduction of survival and recruitment of componentspecies in the ecosystem (e.g. through warming, desicca-tion and exotic species) (Fig. 4). Both factors will operateglobally for cloud forest ecosystems (along with the majorissue of loss of habitat), while local factors may also play arole (Fig. 4). As an example, increased desiccation isexpected to reduce species survival and limit opportunities
for recruitment of new individuals.This may result fromcombinations of reduced cloud cover or precipitation,increased warming or increased extreme storm activitythat opens up the existing canopy. In particular, epiphytesmay be very sensitive to changes in microclimate resultingfrom predicted global climate changes (Benzing 1998;Nadkarni 2010), yet these plants play a key role in light,hydrological and nutrient regimes in the forests in whichthey occur (Foster 2001).
Survival and recruitment of species will also beimpacted by local factors, including for Lord HoweIsland, the impact of exotic species such as rats, patho-gens or weeds (Auld & Hutton 2004). Impacts of exoticspecies will also be a global concern for cloud forestsand have been shown to be a major threat to other cloudforests of the Pacific (Meyer 2010). Exotics can selec-tively alter species composition, modify habitats and, incombination with changing moisture availability andwarming temperatures,modify both the structure of theecosystem and the species that can occur in the area.Increased storm damage, declining cloud cover,warming temperatures and impacts of exotic specieswill reduce survival and recruitment in componentspecies in the ecosystem.This is likely to result in botha changed ecosystem composition and ultimately eco-system collapse (Fig. 4).
Globally cloud forest ecosystems are threatened bychanges to land use through clearing, alien species andclimate change (Scatena et al. 2010). On Lord HoweIsland, there are a number of threats to the GnarledMossy Cloud Forest ecosystem (NSW ScientificCommittee 2011) (Fig. 4). Climate change is consid-ered to be a major threat to the ecosystem. As theGnarled Mossy Cloud Forest is confined to mountainsummits, there is no bioclimatic zone for componentspecies to move into with warming temperatures(Australian National University 2009; NSW ScientificCommittee 2011). Decreases in cloud formation(timing, duration and frequency) on the mountains willimpact on the moisture regime and species survival inthe cloud forest (Auld & Hutton 2004) and may favourspecies from lower down the mountains that can bettertolerate desiccation. The ‘lift-cloud-base hypothesis’states that the climate of tropical mountains will gradu-ally change because of an elevation in sea surface tem-peratures (Pounds et al. 1997, 1999; Still et al. 1999).Still et al. (1999) suggest that under a changing climatethere are likely to be upward altitudinal shifts in theareas that experience cloud formations. Pounds et al.(1999) suggest that such a change may have contrib-uted to frog and toad decline in a Costa Rican CloudForest. If this prediction holds for Lord Howe Island,then there may be reduced formation of clouds on thesouthern mountains and reduced moisture availability.The impacts of a rise of the cloud layer, caused by risingsea surface temperatures, constitute a major climate-related threat to the island’s ecosystems. Simmons et al.
Fig. 4. Conceptual model of ecosystem dynamics leadingto collapse for Gnarled Mossy Cloud Forest. Grey boxesrepresent global threats and outcomes for cloud forests,excluding loss of habitat. Unfilled boxes are local threats forLord Howe Island (but may also be applicable to other cloudforests). Arrows indicate direction of transition betweenstates.
CLOUD FOREST LHI 367
© 2014 The Authors doi:10.1111/aec.12202Austral Ecology © 2014 Ecological Society of Australia

(2012) estimated that sea level temperatures aroundLord Howe Island have risen by some 0.6°C since1940. Predicted changes in global mean air tempera-tures from global circulation models forecast a changeof 1.5–6°C by 2100 (Intergovernmental Panel onClimate Change 2007). Australian National University(2009) suggests that average annual air temperature onLord Howe Island is expected to rise (compared with1990 levels) by 1.3 ± 0.6°C by 2030, although there ismuch uncertainty around such estimates and attempt-ing to apply global projections to Lord Howe Island.
The exotic ship rat Rattus rattus has been on LordHowe Island for some 90 years. It is known to havesevere impacts on the two endemic palm genera(Hedyscepe and Lepidorrhachis) that occur in theGnarled Mossy Cloud Forest (Baker & Hutton 2006;Auld et al. 2010), and results in a reduction in groundcover across areas impacted by rats. Rats limit recruit-ment in the palms and consume a large proportion ofseeds and leaves of a number of other taxa (Auld &Hutton 2004; unpubl.). Hence, rats are likely to impacton recruitment in many plant species on Lord HoweIsland. Some 7% of the summit plateau of Mt Gower isbaited to reduce rat impacts, and there is a plan to try toeradicate rats across the whole island (Lord HoweIsland Board 2009). There have been other introduc-tions of exotic animals to Lord Howe Island. Pigs andgoats were naturalized since the mid-1800s. Goats andpossibly pigs had locally significant impacts on GnarledMossy Cloud Forest (Pickard 1983), but the long-termimpacts are unknown. Both have now been eradicatedfrom the island (DECC 2007).
Weeds are only a minor threat on the summit of MtGower, where a small number of weeds have beenrecorded within the Gnarled Mossy Cloud Forest eco-system. Most are facilitated by disturbance and takeadvantage of cliff edges or areas of tree fall, lightningstrike, tree death and other minor disturbances.Theseweeds may compete with native plants for recruitmentin canopy gaps and cliff edges.The species compositionand structure of the ecosystem will be modified if weedsare able to exclude natives.Weeds identified in GnarledMossy Cloud Forest on Mt Gower include: Agrostisavenaceus, Arenaria serpyllifolia, Avena barbata, Bromuscarthartius, Ehrharta erecta, Lolium perenne, Luzulalongiflora, Paspalum sp., Poa annua, Polycarpontetraphyllum, Potentilla indica, Pseudognaphalium luteo-album, Rumex sp., Solanum nigrum, Sonchus oleraceus,Sporobolus africanus, Stellaria media and Vulpiabromoides. On Mt Lidgbird, the more limited extent ofGnarled Mossy Cloud Forest along a narrow ridge(Figs. 2, 3) has allowed more aggressive weedsAgeratina adenophora, Lilium formosanum and S. nigrumto invade (NSW Scientific Committee 2011).
Several other potential threats could impact on speciescomposition and structure in the Gnarled Mossy CloudForest. The exotic pathogen Phytophthora cinnamomi
occurs on Lord Howe Island and could spread to theGnarled Mossy Cloud Forest via walkers or birds,although education measures have been undertaken tominimize this risk. Currently, there are few data on thesusceptibility of key Lord Howe Island endemic plants tothis pathogen (see Auld & Hutton 2004). A number ofpotentially susceptible species are major structural domi-nants in the Gnarled Mossy Cloud Forest. Species occur-ring as part of the Gnarled Mossy Cloud Forest on LordHowe Island and belonging to the Family Myrtaceae(Metrosideros nervulosa and Leptospermum polygalifoliumssp. howense) may be susceptible to exotic rust pathogensincluding myrtle rust, which is present on mainland NSW(NSW Scientific Committee 2011).
METHODS
To assess the Gnarled Mossy Cloud Forest Ecosystem againstthe IUCN Red List for ecosystems criteria (Keith et al. 2013),we searched the available literature for evidence of any threatsto the ecosystem that may result in ecosystem decline.We alsoexamined geographic distribution and both biotic and abioticfactors associated with the ecosystem in the light of theconceptual model of ecosystem dynamics that could lead tocollapse for the Gnarled Mossy Cloud Forest (Fig. 4).
For decline, we examined historical vegetation reports(Maiden 1898, 1914; Oliver 1916; Pickard 1983; Green1994; Mueller-Dombois & Fosberg 1998; Harris et al. 2005).For more recent changes, we inspected satellite imagery(Google Earth and 2011 air photo imagery). We used avail-able Geographical Information System data layers for thedistribution of Gnarled Mossy Cloud Forest (mapped fromPickard 1983) to estimate both the extent of occurrence andarea of occupancy of the ecosystem. Extent of occurrencewas estimated using a minimum convex polygon enclosingthis distributional layer, while area of occupancy was esti-mated by determining the number of 10 × 10 km grids thatthe ecosystem covers.
To assess changes in abiotic features that directly impacton Gnarled Mossy Cloud Forest, we used two metrics basedon collated data from the Australian Bureau of Meteorologyfrom mid 1940s onwards. These metrics relate to how cli-matic moisture conditions may have changed (and the poten-tial for future change). The two metrics were:1. Frequency of cloud cover on the summits of the south-
ern mountains where the Gnarled Mossy Cloud Forestecosystem occurs. We estimated the number of days perannum where there was cloud cover on the summit ofthe two southern mountains of Lord Howe Island usingdata from 1946 to 2013. These peaks are observablefrom the Lord Howe Island meteorological station andcloud cover is recorded several times per day. As theGnarled Mossy Cloud Forest ecosystem is restricted toelevations above 750 m, this metric allowed us to esti-mate the long-term changes in cloud cover directly rel-evant to this ecosystem. These data were used tocalculate changes in the number of days of cloud cover(with plausible bounds) over the last 50 years and for thenext 50 years. In cloud forests, the degree of cloud covervaries with elevation and distance from the coast (Jarvis
368 T. D. AULD AND M. R. LEISHMAN
© 2014 The Authorsdoi:10.1111/aec.12202Austral Ecology © 2014 Ecological Society of Australia

& Mulligan 2011). As cloud forests are confined to habi-tats with generally >60% cloudy days per annum(Mulligan & Burke 2005) and data from Lord HoweIsland suggested cloud cover was historically in thisrange (average over 78 years of 69%, annual range of55–83%), we chose a threshold of collapse of 50%cloudy days per year for this index.
2. Changes to rainfall. The meteorological station, locatedjust above sea level, can be used to estimate if there hasbeen any decline in rainfall. We used data on annualrainfall for Lord Howe Island that had been analysed bySimmons et al. (2012) to make inferences about chang-ing rainfall on the island that would also reflect changesto moisture availability in the Gnarled Mossy CloudForest ecosystem. Use of this index assumes thatchanges in rainfall at sea level would indicate a compara-tive level of change at the mountain top elevations ofGnarled Mossy Cloud Forest and that any decreases willreduce the persistence of species within the ecosystem.Again, comparisons were made between annual rainfallmeasures in the last 50 years and for the next 50 years.For rainfall, Pickard (1983) reported dying plant frondsand leaf fall on lowland trees and desiccation of herbs onLord Howe Island during a dry period in 1977. Annualrainfall on Lord Howe Island in 1977 was 1071 mm,compared with the average of 1526 mm. 1977 was thesecond lowest annual rainfall recorded in the last 73years (the lowest being in 1997 at 990 mm). Hence, weused a collapse threshold of 900 mm annual rainfall(below the lowest ever recorded).
Temperature increases have been identified for near sea levelon Lord Howe Island (Simmons et al. 2012),and any increases intemperatures observed at sea level are likely to be reflected intemperatures on the southern mountains as Simmons et al.(2012) found an adiabatic lapse rate of some 0.9°C per 100 mincrease in altitude on Mt Gower on Lord Howe Island.We didnot use temperature as an abiotic variable as we had no data toinform the threshold of collapse of Gnarled Mossy Cloud Forestin relation to temperature. Instead, we examined the strength ofthe correlations between annual average minimum andmaximum temperatures with both cloud cover frequency andannual rainfall to determine if temperature might be a usefulpredictor of these abiotic variables.
There is further potential abiotic degradation from anyincreases in storm damage from extreme events underclimate change. Remote sensing data could also be used toestimate changes in storm frequency; however, no index ofthis is currently available.
For biotic degradation, we reviewed the impact of exotic ratson limiting recruitment of key species in the ecosystem and inchanging the structure and composition of the ecosystem.
For assessment of criteria A and B, collapse was assumedto occur when the mapped distribution of the ecosystemdeclines to zero. For criterion C, ecosystem collapse wasdefined in relation to changes in cloud days (collapse at 50%)or precipitation (collapse at annual rainfall of 900 mm).Decreases in cloud days or precipitation lead to a reductionin the persistence of species within the ecosystem until thethresholds of collapse are reached. Under criterion D, eco-system collapse was assessed based on changes in communitycomposition, particularly loss of canopy dominants and char-acteristic epiphytes.We could not undertake an estimation of
the probability of ecosystem collapse (criterion E), and thiscriterion was considered to be Data Deficient.
RESULTS
Criterion A – decline in distribution
There have been no observed changes in the extent of thisecosystem in the last 50 years (Pickard 1983).The remotelocation, steep terrain and conservation value of thesouthern mountains has ensured no clearing of theecosystem. No changes are indicated since Pickard’s 1983vegetation classification based on inspection of recent sat-ellite imagery (Google Earth and 2011 air photo imagery)and field traverses.The status of the ecosystem is thereforeLeast Concern under criterion A1.
No projections are available for the future distribu-tion, although it is possible the current bioclimaticenvelope for the ecosystem may shift sufficiently tocause ecosystem collapse. Further data on climaticconditions within the ecosystem are needed toexamine this possibility.The status of the ecosystem istherefore Data Deficient under criterion A2.
There has been no observed change in the extent ofthis ecosystem since 1750 (Pickard 1983). The statusof the ecosystem is therefore Least Concern undercriterion A3.
Criterion B – distribution size
The extent of occurrence was estimated to be 20 km2
based on a minimum convex polygon enclosing alloccurrences mapped by Pickard (1983).
The area of occupancy of Gnarled Mossy CloudForest is estimated to be one 10 × 10 km grid cell.
Long-term monitoring data on both rainfall andcloud cover provide evidence for a continuing declinein abiotic components of the ecosystem (AustralianNational University 2009, Simmons et al. 2012, seebelow and Fig. 5). Additionally, rat predation on seedsand seedlings of several sensitive species (e.g. moun-tain palms) that are structural components of the eco-system is inferred to result in continuing decline of thestanding vegetation and seed banks (Auld & Hutton2004; Auld et al. 2010). This rat predation on seedsand seedlings is likely to cause continuing reduction/elimination of recruitment of several key canopyspecies in the next 20 years. The distribution of thisecosystem (Mt Gower and Mt Lidgbird) is consideredto be a single location, as the existing known threatsoccur across both remaining patches.This renders theecosystem susceptible to stochastic processes such astropical cyclones and wildfire within a short timeperiod in an uncertain future.
CLOUD FOREST LHI 369
© 2014 The Authors doi:10.1111/aec.12202Austral Ecology © 2014 Ecological Society of Australia
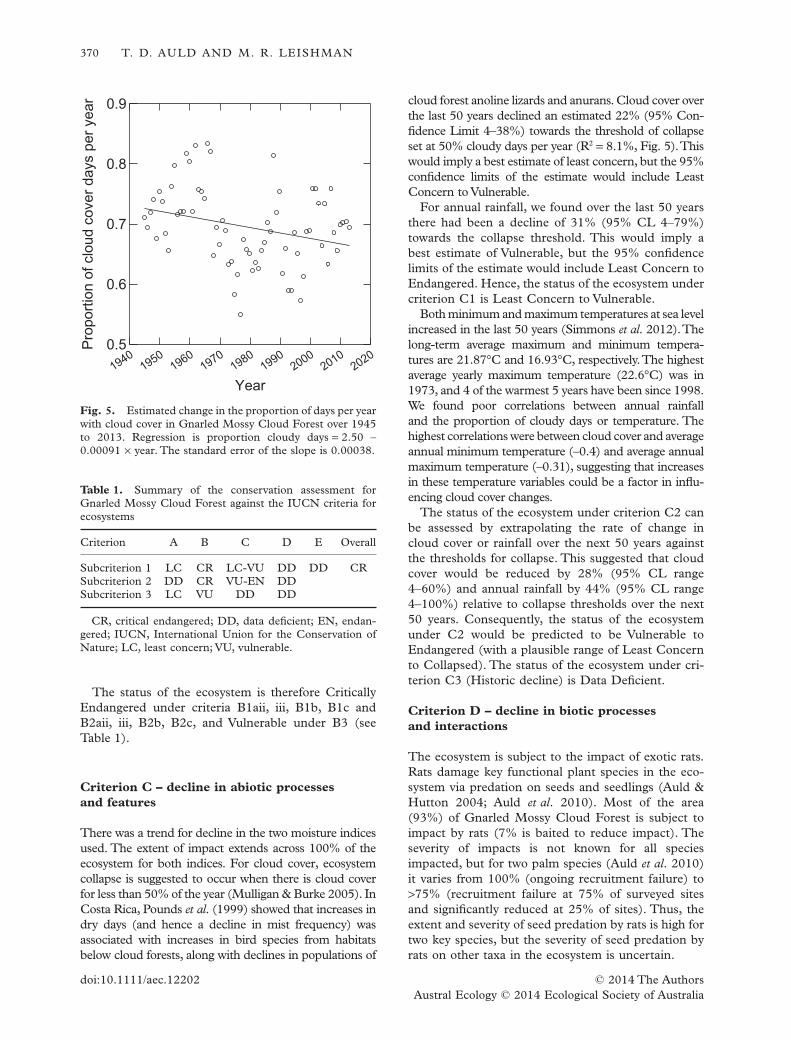
The status of the ecosystem is therefore CriticallyEndangered under criteria B1aii, iii, B1b, B1c andB2aii, iii, B2b, B2c, and Vulnerable under B3 (seeTable 1).
Criterion C – decline in abiotic processesand features
There was a trend for decline in the two moisture indicesused. The extent of impact extends across 100% of theecosystem for both indices. For cloud cover, ecosystemcollapse is suggested to occur when there is cloud coverfor less than 50% of the year (Mulligan & Burke 2005). InCosta Rica, Pounds et al. (1999) showed that increases indry days (and hence a decline in mist frequency) wasassociated with increases in bird species from habitatsbelow cloud forests, along with declines in populations of
cloud forest anoline lizards and anurans. Cloud cover overthe last 50 years declined an estimated 22% (95% Con-fidence Limit 4–38%) towards the threshold of collapseset at 50% cloudy days per year (R2 = 8.1%, Fig. 5).Thiswould imply a best estimate of least concern, but the 95%confidence limits of the estimate would include LeastConcern to Vulnerable.
For annual rainfall, we found over the last 50 yearsthere had been a decline of 31% (95% CL 4–79%)towards the collapse threshold. This would imply abest estimate of Vulnerable, but the 95% confidencelimits of the estimate would include Least Concern toEndangered. Hence, the status of the ecosystem undercriterion C1 is Least Concern to Vulnerable.
Both minimum and maximum temperatures at sea levelincreased in the last 50 years (Simmons et al. 2012).Thelong-term average maximum and minimum tempera-tures are 21.87°C and 16.93°C, respectively.The highestaverage yearly maximum temperature (22.6°C) was in1973, and 4 of the warmest 5 years have been since 1998.We found poor correlations between annual rainfalland the proportion of cloudy days or temperature. Thehighest correlations were between cloud cover and averageannual minimum temperature (–0.4) and average annualmaximum temperature (–0.31), suggesting that increasesin these temperature variables could be a factor in influ-encing cloud cover changes.
The status of the ecosystem under criterion C2 canbe assessed by extrapolating the rate of change incloud cover or rainfall over the next 50 years againstthe thresholds for collapse. This suggested that cloudcover would be reduced by 28% (95% CL range4–60%) and annual rainfall by 44% (95% CL range4–100%) relative to collapse thresholds over the next50 years. Consequently, the status of the ecosystemunder C2 would be predicted to be Vulnerable toEndangered (with a plausible range of Least Concernto Collapsed). The status of the ecosystem under cri-terion C3 (Historic decline) is Data Deficient.
Criterion D – decline in biotic processesand interactions
The ecosystem is subject to the impact of exotic rats.Rats damage key functional plant species in the eco-system via predation on seeds and seedlings (Auld &Hutton 2004; Auld et al. 2010). Most of the area(93%) of Gnarled Mossy Cloud Forest is subject toimpact by rats (7% is baited to reduce impact). Theseverity of impacts is not known for all speciesimpacted, but for two palm species (Auld et al. 2010)it varies from 100% (ongoing recruitment failure) to>75% (recruitment failure at 75% of surveyed sitesand significantly reduced at 25% of sites). Thus, theextent and severity of seed predation by rats is high fortwo key species, but the severity of seed predation byrats on other taxa in the ecosystem is uncertain.
19401950
19601970
19801990
20002010
2020
Year
0.5
0.6
0.7
0.8
0.9P
ropo
rtio
n of
clo
ud c
over
day
s pe
r ye
ar
Fig. 5. Estimated change in the proportion of days per yearwith cloud cover in Gnarled Mossy Cloud Forest over 1945to 2013. Regression is proportion cloudy days = 2.50 –0.00091 × year. The standard error of the slope is 0.00038.
Table 1. Summary of the conservation assessment forGnarled Mossy Cloud Forest against the IUCN criteria forecosystems
Criterion A B C D E Overall
Subcriterion 1 LC CR LC-VU DD DD CRSubcriterion 2 DD CR VU-EN DDSubcriterion 3 LC VU DD DD
CR, critical endangered; DD, data deficient; EN, endan-gered; IUCN, International Union for the Conservation ofNature; LC, least concern; VU, vulnerable.
370 T. D. AULD AND M. R. LEISHMAN
© 2014 The Authorsdoi:10.1111/aec.12202Austral Ecology © 2014 Ecological Society of Australia

The ecosystem is also susceptible to P. cinnamomi andmyrtle rust as several dominants in the canopy are likely tobe impactedby thesepathogens.Phytophthora cinnamomi ispresent on Lord Howe Island (Auld & Hutton 2004). Noestimate of severity or impact is available.
The status of the ecosystem under criteria D1, D2and D3 are all Data Deficient.
DISCUSSION
Unlike other global cloud forest ecosystems (Scatenaet al. 2010), Gnarled Mossy Cloud Forest has notbeen subject to recent clearing or fragmentation. Wefound Gnarled Mossy Cloud Forest to be CriticallyEndangered based on a restricted geographic distribu-tion combined with continuing decline in abiotic andbiotic processes inferred from: a loss of moisture fromdeclining rainfall and cloud cover due to climatechange; ongoing exotic rat predation on seeds andseedlings of several sensitive species that are structuralcomponents of the ecosystem; and the single locationof the ecosystem. These are likely to lead to reducedsurvival and recruitment in species in the ecosystem(Fig. 4). Hence, this ecosystem is impacted by similarthreats to other Pacific island cloud forests (Meyer2010).
We successfully applied several related indices ofchange in the climate that may affect the GnarledMossy Cloud Forest ecosystem. However, the sensitiv-ity of relationships between the degree of decline ineither cloud cover or rainfall, and species and ecosys-tem decline remains somewhat uncertain. Our esti-mates of thresholds for collapse are based on limitedinformation and the estimates of decline in cloud coverand rainfall had large plausible bounds (e.g. resultingin 95% confidence limit estimates of Least Concern toCollapsed in C2). While cloud cover data have themost direct relevance to species survival in the ecosys-tem, average annual temperature changes were nega-tively correlated with cloud cover suggesting the ‘liftcloud base hypothesis’ (Pounds et al. 1997, 1999; Stillet al. 1999) may be applicable to Lord Howe Island. Itis possible that increases to temperature near sea levelmay be a suitable surrogate for changes in cloud coverin the Gnarled Mossy Cloud Forest ecosystem. Dataon biotic processes and interactions were available foronly two of the canopy palm species in the ecosystem.While we had data on both severity (75–100%) andextent (93%) for these species, uncertainty remainsabout the severity across other key structural taxa inthe ecosystem (just how many species need to beimpacted to drive the ecosystem to decline). As well, asmature plants may be long-lived, recruitment failurewould need to consistently occur across decades (as isthe case in the two palms; Auld et al. 2010). Factorsinfluencing adult tree survival (such as reduced mois-
ture availability, increased extreme weather and light-ning strikes) will also be important estimating theseverity of rat impacts.
Future prognosis for the ecosystem
A rodent eradication programme has been developedfor Lord Howe Island (Lord Howe Island Board 2009;Wilkinson & Priddel 2011) and is planned to occur inthe coming years. If successful, this has the potential toeliminate the major immediate threat to thisecosystem. If unsuccessful, regular rodent baitingacross much of the distribution of the ecosystemwould be needed, although this is not likely to be along-term solution as the rodents gain resistance to thepoisons. A reduction in rat impact is needed to bothallow successful seed production and recruitment ofnew plants. As rats also consume leaves of developingplants, a period of a decade or more of minimal ratimpacts would be needed to promote regeneration.
Phytosanitory guidelines have been developed forLord Howe Island in an effort to reduce any risk ofspread of the existing P. cinnamomi (DECC 2007) andlimit the introduction of novel pathogens such asmyrtle rust. An ongoing programme of early detectionis needed to complement this. Weed impacts are cur-rently minor except for the ecosystem occurrence onMt Ligdbird, where large habitat edges favouringcrofton weed are impacting on the ecosystem. Unfor-tunately, the remote and rugged nature of the terraincurrently prevents any control efforts.
Any prevention of climate warming and reduction incloud-derived precipitation will depend on global effortsto reduce greenhouse gas emissions.The current progno-sis for this is not favourable (Peters et al. 2013).
Collapse of the Gnarled Mossy Cloud Forest Eco-system would have cascade effects for many of thecomponent species and others dependent upon thisecosystem. Visiting the ecosystem is also part of thepopular tourist walk to Mt Gower. Decline and loss ofthis unique ecosystem would likely impact on thepopularity of this walk and the tourism values of theisland.
ACKNOWLEDGEMENT
This work was supported by an Australian ResearchCouncil Linkage Grant (LP LP130100435). JudyScott and Chris Simpson developed the distributionmap of Gnarled Mossy Cloud Forest.We thank DavidKeith for his helpful comments on earlier versionsof this manuscript and all the participants of the Aus-tralian Centre for Ecological Analysis and Synthesisworkshop for their thoughtful contributions to thedevelopment of the risk assessment for this ecosystem.
CLOUD FOREST LHI 371
© 2014 The Authors doi:10.1111/aec.12202Austral Ecology © 2014 Ecological Society of Australia

REFERENCES
Auld T. D. & Hutton I. (2004) Conservation issues for thevascular flora of Lord Howe Island. Cunninghamia 8, 490–500.
Auld T. D., Hutton I., Ooi M. K. J. & Denham A. J. (2010)Invasive species on oceanic islands: disruption of recruit-ment in narrow endemic palms. Biol. Invasions 12, 3351–61.
Australian National University (2009) Implications of climatechange for Australia’s World Heritage properties: a prelimi-nary assessment. A report to the Department of ClimateChange and the Department of the Environment, Water,Heritage and the Arts by the Fenner School of Environmentand Society, the Australian National University.
Baker W. J. & Hutton I. (2006) Lepidorrhachis. Palms 50, 33–8.Benzing D. H. (1998) Vulnerabilities of tropical forests to climate
change: the significance of resident epiphytes. Clim. Change39, 519–40.
Brotherus V. F. & Watts W.W. (1915) The mosses of Lord HoweIsland. Proc. Linn. Soc. N. S.W. 40, 363–85.
Cassis G., Meades L., Harris R. et al. (2003) Lord Howe Islandterrestrial invertebrate biodiversity and conservation.Report to the NSW National Parks and Wildlife Service bythe Australian Museum Centre for Biodiversity and Con-servation Research.
Department of Environment and Climate Change (2007) LordHowe Island biodiversity management plan. Department ofEnvironment and Climate Change (NSW), Sydney.
Foster P. (2001) The potential negative impacts of global climatechange on tropical montane cloud forests. Earth Sci. Rev.55, 73–106.
Green P. S. (1994) Flora of Australia Vol. 49 Oceanic Islands 1.Australian Government Printing Service, Canberra.
Harris R., Cassis G., Auld T. & Hutton I. (2005) Floristics andstructure of the mossy cloud forest of Mt Gower summit,Lord Howe Island. Pac. Conserv. Biol. 11, 246–56.
Intergovernmental Panel on Climate Change (2007) ClimateChange 2007: The Physical Science Basis. Contributions ofWorking Group 1 to the Fourth Assessment Report of the Inter-governmental Panel on Climate Change. Cambridge Univer-sity Press, Cambridge.
Jarvis A. & Mulligan M. (2011) The climate of cloud forests.Hydrol. Process. 25, 327–43.
Keith D. A., Rodríguez J. P., Rodríguez-Clark K. M. et al. (2013)Scientific Foundations for an IUCN Red List of Eco-systems. PLoS ONE 8, e62111.
Lord Howe Island Board (2009) Draft Lord Howe Island rodenteradication plan. Lord Howe Island Board, Lord HoweIsland.
McCarthy P. M., Streimann H. & Elix J. A. (1996) Newfoliicolorus species of Strigula from Lord Howe Island,Australia. Lichenologist 28, 239–44.
McDougall I., Embleton B. J. J. & Stone D. B. (1981) Origin andevolution of Lord Howe Island, southwest Pacific Ocean.J. Geol. Soc. Austr. 28, 155–76.
Maiden J. H. (1898) Observations of the vegetation of LordHowe Island. Proc. Linn. Soc. N. S.W. 23, 112–58.
Maiden J. H. (1914) Further notes on the botany of Lord HoweIsland (fifth paper). Proc. Linn. Soc. N. S.W. 39, 377–84.
Meyer J.-Y. (2010) Montane cloud forests on remote islands ofOceania: the example of French Polynesia (South Pacific
Ocean). In: Tropical Montane Cloud Forests: Science for Con-servation and Management (eds L. A. Bruijnzeel, F. N.Scatena & L. S. Hamilton) pp. 121–9. Cambridge Univer-sity Press, Cambridge.
Mueller-Dombois D. & Fosberg F. R. (1998) Vegetation of theTropical Pacific Islands. Springer-Verlag, New York.
Mulligan M. & Burke S. (2005) Global cloud forests and envi-ronmental change in a hydrological context. DFID FRPProject ZF0216. Final Report. December 2005. [Cited 23September 2014.] Available from URL: http://www.ambiotek.com/cloudforests/cloudforest_finalrep.pdf
Nadkarni N. M. (2010) Potential effects of global climate changeon epiphytes in a tropical montane cloud forest: an experi-mental study from Monteverde, Costa Rica. In: TropicalMontane Cloud Forests: Science for Conservation and Manage-ment (eds L. A. Bruijnzeel, F. N. Scatena & L. S. Hamilton)pp. 557–65. Cambridge University Press, Cambridge.
Nicholson E., Regan T. J., Auld T. D. et al. (2014) Towardsconsistency, rigour and compatibility of risk assessments forecosystems and ecological communities. Austral Ecol. onlineearly.
NSW Scientific Committee (2011) Final determination.Gnarled Mossy Cloud Forest on Lord Howe Island – criti-cally endangered ecological community listing. [Cited 23September 2014.] Available from URL: http://www.environment.nsw.gov.au/determinations/gnarledmossycloudforestlhifd.htm
Oliver W. R. B. (1916) The vegetation and flora of Lord HoweIsland. Trans. Proc. N. Z. Inst. 49, 94–161.
Peters G. P., Andrew R. M., Boden T. et al. (2013) The challengeto keep global warming below 2°C. Nat. Clim. Change 3,4–6.
Pickard J. (1983) Vegetation of Lord Howe Island.Cunninghamia 1, 133–265.
Pounds J. A., Fogden M. P. L. & Campbell J. H. (1999) Biologi-cal response to climate change on a tropical mountain.Nature 398, 691–715.
Pounds J. A., Fogden M. P. L., Savage J. M. & Gorman G. C.(1997) Tests of null models for amphibian declines on atropical mountain. Conserv. Biol. 11, 1307–22.
Scatena F. N., Bruijnzeel L. A., Bubb P. & Das S. (2010) Settingthe stage. In: Tropical Montane Cloud Forests: Science for Con-servation and Management (eds L. A. Bruijnzeel, F. N.Scatena & L. S. Hamilton) pp. 3–13. Cambridge UniversityPress, Cambridge.
Simmons L., Auld T. D., Hutton I., Baker W. J. & Shapcott A.(2012) Will climate change, genetic and demographic vari-ation or rat predation pose the greatest risk for persistence ofan altitudinally distributed island endemic? Biology 1, 736–65.
Still C. J., Foster P. N. & Schneider S. H. (1999) Simulating theeffects of climate change on tropical montane cloud forests.Nature 398, 608–10.
Wilkinson I. S. & Priddel D. (2011) Rodent eradication on LordHowe Island: challenges posed by people, livestock, andthreatened endemics. In: Island Invasives: Eradication andManagement (eds C. R. Veitch, M. N. Clout & D. R. Towns)pp. 508–14. IUCN, Gland.
372 T. D. AULD AND M. R. LEISHMAN
© 2014 The Authorsdoi:10.1111/aec.12202Austral Ecology © 2014 Ecological Society of Australia





















![Issue #: [Date] Dolor Sit Amet Lord Howe Island Board](https://img.pdfslide.us/doc/110x75/61a16916f2fcc72d8717e0db/issue-date-dolor-sit-amet-lord-howe-island-board.jpg)







