-
8/22/2019 Long-Term Enhancement of Synaptic Transmission
Between
1/16
J Physiol591.1 (2013) pp 287302 287
e
Jou
a
o
ysoo
gy
Neuros
cience Long-term enhancement of synaptic transmission
between
antennal lobe and mushroom body in cultured Drosophilabrain
Kohei Ueno1, Shintaro Naganos1, Yukinori Hirano1,2, Junjiro
Horiuchi3 and Minoru Saitoe1
1Tokyo Metropolitan Institute of Medical Science, 2-1-6
Kamikitazawa, Setagaya-ku, Tokyo, 1568506, Japan2PRESTO, Japan
Science and Technology Agency, 4-1-8 Honcho, Kawaguchi, Saitama
3320012, Japan3Tokyo Metropolitan University, 1-1 Minami-osawa,
Hachiouji, Tokyo, 1920397, Japan
Key points
During olfactory aversive conditioning in Drosophila, odour and
shock information aredelivered to the mushroom bodies (MBs) through
projection neurons in the antennal lobes(ALs) and ascending fibres
of the ventral nerve cord (AFV), respectively.
Using an isolated cultured brain expressing a Ca2+
indicator in the MBs, we demonstrated thatthe simultaneous
stimulation of the ALs and AFV establishes long-term enhancement
(LTE)in AL-induced Ca2+ responses.
The physiological properties of LTE, including associativity,
input specificity and persistence,are highly reminiscent of those
of olfactory memory.
Similar to olfactory aversive memory, LTE requires the
activation of nicotinic acetylcholinereceptors that mediate the
AL-evoked Ca2+ response, NMDA receptors that mediate theAFV-induced
Ca2+ response, and D1 dopamine receptors during the simultaneous
stimulationof the ALs and AFV.
Considering thephysiologicalandgenetic analogies,we
proposethatLTE at theALMB synapsecan be a relevant cellular model
for olfactory memory.
Abstract In Drosophila, the mushroom body (MB) is a critical
brain structure for olfactoryassociative learning. During aversive
conditioning,the MBs are thought to associateodour signals,conveyed
by projection neurons (PNs) from the antennal lobe (AL), with shock
signals conveyedthrough ascending fibres of the ventral nerve cord
(AFV). Although synaptic transmissionbetween AL and MB might play a
crucial role for olfactory associative learning, its
physiologicalproperties have not been examined directly. Using a
cultured Drosophilabrain expressing a Ca2+
indicator in the MBs, we investigated synaptic transmission and
plasticity at the ALMB synapse.Following stimulation with a glass
micro-electrode, AL-induced Ca2+ responses in the MBs weremediated
through Drosophilanicotinic acetylcholine receptors (dnAChRs),
while AFV-inducedCa2+ responses were mediated through
DrosophilaNMDA receptors (dNRs). ALMB synaptic
transmission was enhanced more than 2 h after the simultaneous
associative-stimulation of ALand AFV, and such long-term
enhancement (LTE) was specifically formed at the ALMB synapsesbut
not at the AFVMB synapses. ALMB LTE was not induced by intense
stimulation of theAL alone, and the LTE decays within 60 min after
subsequent repetitive AL stimulation. Thesephenotypesof
associativity, inputspecificityand persistence of ALMB LTE are
highly reminiscentof olfactory memory. Furthermore, similar to
olfactory aversive memory, ALMB LTE formationrequired activation of
the DrosophilaD1 dopamine receptor, DopR, along with dnAChR
anddNR
C2012 The Authors. The Journal of Physiology
C2012 The Physiological Society DOI:
10.1113/jphysiol.2012.242909
) by guest on August 3, 2013jp.physoc.orgDownloaded from J
Physiol (
http://jp.physoc.org/http://jp.physoc.org/http://jp.physoc.org/
-
8/22/2019 Long-Term Enhancement of Synaptic Transmission
Between
2/16
288 K. Ueno and others J Physiol591.1
during associative stimulations. These physiological and genetic
analogies indicate that ALMBLTE might be a relevant cellular model
for olfactory memory.
(Received 12 August 2012; accepted after revision 28 September
2012; first published online 1 October 2012)
Corresponding authors K. Ueno and M. Saitoe: Tokyo Metropolitan
Institute of Medical Science, 2-1-6 Kamikitazawa,
Setagaya-ku, Tokyo, 1568506, Japan. Emails:
[email protected], [email protected]
Abbreviations AFV, ascending fibres of the ventral nerve cord;
AL, antennal lobe; AN, antennae; CS, conditioned
stimulus;DA, dopamine; dnAChR, Drosophilanicotinicacetylcholine
receptor;dNR, DrosophilaNMDA receptor; DopR,
DrosophilaD1 dopamine receptor; GPCR, G-protein coupled
receptor; LH, lateral horn; LTE, long-term enhancement;
LTP, long-term potentiation; MB, mushroom body;MP, maxillary
palp;ORN, olfactory receptorneuron;PN, projection
neuron; US, unconditioned stimulus; VNC, ventral nerve cord.
Introduction
After olfactory aversive conditioning, Drosophilaselectively
increases avoidance toward the conditionedodour (conditioned
stimulus, CS), which was previouslypresented with foot-shock
(unconditioned stimulus,US). Because chemical ablation of the
mushroom bodies
(MBs) completely prevents olfactory learning (de Belle&
Heisenberg, 1994) and blocking of the synapticoutput from MBs
disrupts retrieval (Dubnau et al. 2001;McGuire et al. 2001), the
MBs are considered as a criticalneuronal structure for integrating
odour and foot-shockinformation. Consistent with this model, recent
in vivoimaging studies have demonstrated the formation ofmemory
traces in the MBs after conditioning; Ca2+
responses in the MBs to an odour, which were previouslypaired
with foot-shock, are increased for more than 1 h(Wang et al. 2008;
Tan et al. 2010). However, the synapticmechanisms involved in
neural plasticity in the MBsremain unknown.
In Drosophila, odour information is delivered to theMBs through
projection neurons (PNs) in the antennallobe (AL) (Marin et al.
2002; Wong et al. 2002), whereasfoot-shock information from the
body is delivered to theMBs via the ascending fibres of the ventral
nerve cord(AFV) (Fig. 1A). Although in vivoimaging has
suggestedthat the physiological properties of the ALMB
synapsesarepotentiallyimportantforneuronalplasticityintheMB,current
in vivoimaging methods are unsuited to analysingALMB synaptic
transmission by stimulating AL directly.Furthermore, considerable
brainmovement duringinvivorecording reduces the signal-to-noise
ratio, thereby hind-
ering the kinetic analysis of MB responses.To overcome these
difficulties and analyse ALMBsynaptic transmission and plasticity,
we employed invitro imaging (Wang et al. 2008; Tomchik &
Davis,2009) using an isolated cultured Drosophila brain todirectly
stimulate the AL and AFV without brainmovement during recording of
the Ca2+ responsesin the MBs. Using this in vitro imaging system
inconjunction with high-speed scanning confocal micro-scopy, we
observed that cholinergic ALMB synaptictransmission was enhanced
for more than 2 h after the
simultaneous stimulation of the AL and AFV. Strikingly,the
physiological properties and genetic requirementsof this long-term
enhancement (LTE) at the ALMBsynapse are highly reminiscent of the
characteristics ofolfactory memory. We propose that ALMB LTE
mightbe a reasonable cellular model for learning and memorysimilar
to long-term potentiation(LTP) at the mammalian
hippocampal synapses.
Methods
Fly stocks
All fly stocks were maintained at 25 2C and 60 10%humidity under
a 12/12 h lightdark cycle. All transgenicflies and mutants were
outcrossed to ourwild-type controlline w(CS10) (Dura et al. 1993;
Tamura et al. 2003). Weused female flies for imaging analyses, and
both male andfemale flies for behavioural tests.
Imaging analysis
Brains with attached ventral nerve cords (VNCs) (Fig. 1B)were
dissected in 0 mM Ca2+ HL3 medium (in mM, NaCl,70; sucrose,115;
KCl,5; MgCl2, 20; NaHCO3,10;trehalose,5; Hepes, 5; pH 7.3) (Stewart
et al. 1994). The isolatedbrains were immobilized by placing their
optic lobesbetween two nylon fibre bundles attached to the
platinumgrid and were placed in a bath chamber (Fig. 1C). We
cuttheVNC at thecervical connective. The ALs andAFVwereelectrically
stimulated using glass micro-electrodes with
a stimulator (SEN-7103; Nihon Kohden, Tokyo, Japan)and an
isolator (SS-104J for AL, SS202J for AFV, NihonKohden). During the
experiments, fresh HL3 medium (inmM, NaCl, 70; sucrose, 115; KCl,
5; MgCl2, 20; CaCl2, 1.8;NaHCO3, 10; trehalose, 5; Hepes, 5; pH
7.3) was infusedinto the chamber using a peristaltic pump (2 ml
min1,MiniPuls3, Gilson, Inc., Middleton, WI, USA).
Images were captured using a high-speed scanningconfocal
microscope system (A1R, Nikon Corp., Tokyo,Japan) with a 20
water-immersion lens (numericalaperture 0.5; Nikon Corp.). In this
imaging system,
C2012 The Authors. The Journal of Physiology
C2012 The Physiological Society
) by guest on August 3, 2013jp.physoc.orgDownloaded from J
Physiol (
http://jp.physoc.org/http://jp.physoc.org/http://jp.physoc.org/
-
8/22/2019 Long-Term Enhancement of Synaptic Transmission
Between
3/16
J Physiol591.1 Synaptic plasticity in cultured Drosophila brain
289
512 512 pixel images canbe capturedat 30 Hzfrequency.Before the
experiments, the offset value was set toobtain a background
intensity of approximately 0. TheF value was calculated for each
pixel in the region ofinterest using NIS-elements software
(NIS-Elements Ar;Nikon Corp.).
To record AL- and AFV-induced Ca2+ responses, we
stimulated the AL or AFV with three trains of 30 pulses(100 Hz,
1.0 ms pulse duration, intensity 12 thresholdcurrent) with an
inter-train interval of 10 s. We obtainedinitial fluorescence
values (F0) by averaging the F valuesobtained in the five
sequential frames before stimulationonset. We averaged the three
fluorescent traces obtainedafter stimulation and calculated (F
F0)/F0 to obtainF/F0. To evaluate the relative Ca
2+ responses inducedthrough the associative stimulation of the
AL andAFV or intensive AL stimulation, the F/F0 obtainedafter
associative or intensive stimulation was dividedby the F/F0
obtained prior to associative or intensive
stimulation.
Electrical stimulation protocols
For associative stimulation, the AL and AFV weresimultaneously
stimulated with 12 trains at an inter-val of 5 s, and the average
of the first three responseswas considered as the relative Ca2+
response duringthe associative stimulation. For unpaired
stimulation,AL stimuli were induced at 3 min after the end ofthe
AFV stimuli. In intensive AL-alone stimulation,the pulse duration
for AL stimulation was extended
from 1.0 to 1.5 ms to obtain a Ca2+
response duringintensive stimulation comparable with that
elicited duringassociative stimulation.
Behavioural tests
The procedure for measuring olfactory memory hasbeen previously
described (Tully & Quinn, 1985). Briefly,two aversive odours
(OCT or MCH) were sequentiallydelivered to approximately 100 flies
for 1 min at an inter-val of 45 s between each odour exposure. When
the flieswere exposed to the first treatment, CS odour (either
OCT
or MCH), they were also subjected to 1.5 s pulses of 60 VDC
electric shocks every 5 s. To test olfactory memory, theflies were
placed at the choice point of a T-maze whereboth odours were
delivered and were allowed to choosebetween theodours. After 1.5
min, memory wascalculatedas a performance index, such that a 50:50
distribution (nomemory) yielded a performance index of zero and a
0:100distributionawayfromtheCSyieldedaperformanceindexof 100.
For extinction, the flies were sequentially exposed tothe CS
odours for 1 min at 5 min intervals in the training
chamber after conditioning to expose the conditioned fliesto the
CS odours 11 times before examining 1 h memory.
LexA/LexAop system in DopR rescue flies
To construct the pMBp-LexA construct, which contains
the LexA gene under the control of an MBpromoter/enhancer, a DNA
fragment containing 247 bpof the 5 flanking sequence from the Mef2
gene (whichis expressed in the MBs) and the hsp70Bb minimalpromoter
was subcloned into pCasper-W-LexA::GAD(Diegelmann et al. 2008)
using the Gateway system (LifeTechnologies Corp., Carlbad, CA, USA)
according to themanufacturers instructions. To obtain
LexAop-G-CaMP2for expression of G-CaMP2 from a LexA driver,
G-CaMP2cDNA amplified from pN1-G-CaMP2 was subclonedinto
pCasper-lexAop-W (Diegelmann et al. 2008). Thetwo constructs were
injected into embryos from ourstandard w(CS10) strain to obtain
w;MBp-LexA andw;LexAop-G-CaMP2transgenic flies.
In the DopRf02676 mutant, DopRexpression is inhibitedby
insertion of a piggyBac construct in the first intronof the DopR
gene (Kim et al. 2007). Because thepiggyBac construct carries a UAS
sequence (Thibaultet al. 2004), DopR expression can be induced from
anin-frame ATG in the second exon using a GAL4 driver(Kim et al.
2007). Using this system, we
generatedw;MBp-LexA,LexAop-G-CaMP2/+;DopRf02676/DopRf02676
and w;MBp-LexA/c309,LexAop-G-CaMP2;DopRf02676/DopRf02676
flies.
Statistics
All data are expressed as mean SEM. Students t test forpaired
data was used to evaluate the statistical significancebetween two
data sets. Formultiple comparisons, one-wayANOVA was used, followed
by Bonferroni post hocanalyses. The time constant for decay
kinetics (see Fig. 3I)was calculated by fitting a single
exponential curve, usingPrism (GraphPad Software, Inc., La Jolla,
CA, USA).
Results
Imaging analyses of ALMB synaptic transmission
We isolated Drosophila brains expressing the Ca2+
indicator G-CaMP in the MBs using a GAL4/UAS binarysystem (Brand
& Perrimon, 1993) and immobilized theisolated brains in the
recording bath chamber (Fig. 1Band C). To measure synaptic
transmission between PNsand MB neurons, we stimulated the AL using
a glassmicro-electrode (Fig. 1C).
The MBs contain approximately 5000 intrinsic neuronscalled
Kenyon cells. Kenyon cells receive the presynaptic
C2012 The Authors. The Journal of Physiology
C2012 The Physiological Society
) by guest on August 3, 2013jp.physoc.orgDownloaded from J
Physiol (
http://jp.physoc.org/http://jp.physoc.org/http://jp.physoc.org/
-
8/22/2019 Long-Term Enhancement of Synaptic Transmission
Between
4/16
290 K. Ueno and others J Physiol591.1
terminals of the PNs at the calyx, dendritic regions ofthe MBs,
and project terminals to vertical and
horizontallobes.Kenyoncellsareclassifiedas/,/ andneuronsusing
functional and anatomical criteria (Ito et al. 1997;Crittenden et
al. 1998). While AL-induced Ca2+ responseswere consistently
observed in the distal ends of the verticallobes (/), these
responses were weak and infrequent in
the horizontal lobes (//) (Fig. 2A and B) and in thecalyx (Fig.
2C and D).
In contrast, previous imaging studies havedemonstrated that
odour and electrical stimulationto antennae (AN) induces
significant Ca2+ responsesnot only in vertical lobes but also in
horizontal lobesand the calyx (Martin et al. 2007; Wang et al.
2008).Notably, while our external recording solution contains20 mM
Mg2+, these previous studies employed lowerconcentrations of Mg2+
(4 mM). Therefore, we suspectthat the robust Ca2+ responses in the
horizontal lobeobserved in other studies are due to the low
concentration
of Mg2+
.To test this possibility, we examined AL-induced
responses in 4 mM Mg2+ recording solution. As suspected,we
observed a significant increase in responses in thehorizontal lobes
and in the calyx in recording solutioncontaining 4 mM Mg2+ (Fig.
2E). This result suggests that
Ca2+ responses in the horizontal lobe and in the calyx aremore
sensitive to external Mg2+ than Ca2+ responses inthe vertical
lobes.
Consistent with anatomical studies indicating that PNsof each AL
send their axons to the ipsilateral MB (Marinet al. 2002; Wong et
al. 2002), stimulation of an ALinduced Ca2+ responses in the
ipsilateral, but not the
contralateral, MB (Fig. 2F and G). In contrast to ALstimuli, AFV
stimulation induced Ca2+ responses in theMBs of both brain
hemispheres (Fig. 2H and I). Weemployed 20 mM Mg2+ in the recording
solution in thisstudy, as this concentration lies within the
physiologicallyrelevant range (see Discussion), and we primarily
focusedon Ca2+ responses in the vertical lobes in
subsequentexperiments.
LTE of ALMB synaptic transmission
In our in vitro imaging system, we observed the LTEof AL-induced
Ca2+ responses in the MB after thesimultaneous stimulation of AL
and AFV for 1 min(Fig. 3AC). In contrast to associative AL and
AFVstimulation, unpaired AL and AFV stimulation did notenhance the
Ca2+ responses (Fig. 3A, B and D). In this
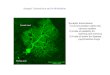
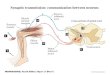
Figure 1. Preparation of isolated cultured Drosophila brains for
imaging
A, odour signalling pathway in Drosophila. An odour signal
perceived through the dendrites of the ORNs in the AN
is transmitted to PNs in the AL and subsequently to MB or LH
neurons that project inhibitory GABAergic terminals
onto MB neurons. The foot-shock signal from the body is
delivered through the AFV. B, a c309;UAS-G-CaMP
fly brain with the VNC. The VNC was cut at the cervical
connection (arrowhead) for stimulating AFV. G-CaMP
expression was observed throughout the , , , and lobes of the
MBs. OL, optic lobe; SEG, sub-oesophageal
ganglion. C, schematic diagram of the recording setup. Glass
micro-electrodes were placed on the AL, and the
cut ends of the AFV were suctioned for stimulation.
C2012 The Authors. The Journal of Physiology
C2012 The Physiological Society
) by guest on August 3, 2013jp.physoc.orgDownloaded from J
Physiol (
http://jp.physoc.org/http://jp.physoc.org/http://jp.physoc.org/
-
8/22/2019 Long-Term Enhancement of Synaptic Transmission
Between
5/16
J Physiol591.1 Synaptic plasticity in cultured Drosophila brain
291
Figure 2. Ca2+ responses in the MBs
A, Ca2+ responses induced by AL stimulation in the vertical
(arrow) and horizontal (arrowhead) lobes of the
MB. While robust Ca2+ responses were observed in the distal end
of the vertical lobe, the significant responses
were not observed in the horizontal lobe (33 of 38 brains). B,
nine traces of AL-induced Ca2+ responses in the
vertical (upper) and horizontal (lower) lobes of the MB. C, the
calyx exhibited weak AL-induced Ca2+ responses. D,
summary of Ca2+ responses in the vertical lobes (black line) and
the calyx (grey line) of the MB (P< 0.05 between
the vertical lobe and the calyx; n = 6). E, AL-induced Ca2+
responses were increased in the horizontal lobes (upper)
and in the calyx (middle) with decreasing extracellular Mg2+
concentration from 20 to 4 mM (P< 0.05, n = 5).
F and G, AL stimulation produced Ca2+ responses in the
ipsilateral but not in the contralateral MB lobe. Hand I,
AFV stimulation produced Ca2+ responses in both sides of MB
lobes (n = 5). All data in this study are shown as
mean SEM.
C2012 The Authors. The Journal of Physiology
C2012 The Physiological Society
) by guest on August 3, 2013jp.physoc.orgDownloaded from J
Physiol (
http://jp.physoc.org/http://jp.physoc.org/http://jp.physoc.org/
-
8/22/2019 Long-Term Enhancement of Synaptic Transmission
Between
6/16
292 K. Ueno and others J Physiol591.1
study, we primarily used the c309 Gal4 driver to measurethe Ca2+
response in the MBs because we observed themost significant LTE
with this MB driver. For example,although the OK107 MB driver
generated higher UAStransgene expression than c309 (Aso et al.
2010), theLTE observed with OK107 was lower than that observed
with c309; the relative Ca2+ responses at 15 min after
theassociativestimuli were 1.48 with OK107 (see Fig. 4H)and2.40
with c309 (Fig. 3C).
These results suggest that the associative AL and AFVstimulation
produces the LTE of ALMB synaptic trans-mission. However, PNs in
the AL also convey olfactory
Figure 3. LTE of AL-induced Ca2+
responses in the MBA, stimulation protocol. In associative
stimulation, AL and AFV were stimulated simultaneously with 12
stimulus
trains (AL + AFV). In unpaired stimulation, AL stimulation was
induced after the end of AFV stimulation. B,
typical changes in G-CaMP fluorescence at the distal end of the
MB vertical lobes (/ lobes), and the traces
of AL-induced Ca2+ responses in the MBs before and after
associated or unpaired stimulation. The arrowhead
indicates stimulation onset. C and D, summary of AL-induced Ca2+
responses in the MBs measured before (pre)
and after associative (C) and unpaired (D) stimulation. The
shaded column in C indicates Ca2+ responses during
associative stimulation (AL + AFV) (P< 0.05 compared with pre
responses). E, averaged traces of AL-induced
Ca2+ responses before and after treatment with 50 M picrotoxin
for 15 min. F, averaged traces of AL-induced
Ca2+ responses before and 5 min after associative AL + AFV
stimulation. G and H, peak Ca2+ responses were
normalized to 1.0 to evaluate the effects of picrotoxin (G) and
associative stimulations (H) on decay kinetics. I,
picrotoxin treatment reduced the decay kinetics (1/, where
represents the time constant) of the Ca2+ response,
while associative stimulations did not (P< 0.05 by t test). n
= 68 for all data.
C2012 The Authors. The Journal of Physiology
C2012 The Physiological Society
) by guest on August 3, 2013jp.physoc.orgDownloaded from J
Physiol (
http://jp.physoc.org/http://jp.physoc.org/http://jp.physoc.org/
-
8/22/2019 Long-Term Enhancement of Synaptic Transmission
Between
7/16
J Physiol591.1 Synaptic plasticity in cultured Drosophila brain
293
Figure 4. LTE occurred in both and lobes but not in the
calyx
A, G-CaMP was expressed in each and lobe by the c305a and
c739 GAL4 drivers, respectively. B and C, typical traces of
AL-induced Ca2+ responses in the (B) and lobes (C) before
(pre)
and 5 min after associative AL and AFV stimulation. D and E,
AL-induced Ca2+ responses were enhanced after associative
stimulation in both (D), and (E) lobes (P< 0.05 compared
with
pre responses, and P< 0.05 comparison between 5 and 30
min).
signalling to lateral horn (LH) neurons (Marin et al.2002; Wong
et al. 2002), which in turn project theirGABAergic terminals onto
MB neurons (Perez-Oriveet al. 2002; Yasuyama et al. 2002; Fig. 1A).
Therefore,it is possible that the enhanced Ca2+ responses in theMBs
reflect the inhibition of LH neurons rather thanincreases in
excitatory transmission between the AL and
the MB.To test this possibility, we recorded AL-induced
Ca2+ responses in the brain following treatment withpicrotoxin,
a GABA receptor blocker (Su & ODowd,2003). Picrotoxin treatment
increased the amplitude ofthe AL-induced Ca2+ response (Fig. 3E)
but significantlydecreased the decay kinetics of the Ca2+
response(Fig. 3G and I). However, after associative AL and
AFVstimulation, the AL-induced Ca2+ response amplitudesincreased
(Fig. 3F) without any changes in the decaykinetics (Fig. 3H and I).
These results support theidea that LTE in the MBs results from an
increase in
excitatory synaptic transmission between PNs and MBneurons.
The vertical lobes in Drosophila consist of and
lobes (Fig. 4A; Ito et al. 1997). Although previous
studiesdemonstrate that the associative stimulation of the ANand
VNC enhances AN-induced Ca2+ responses in
but not lobes (Wang et al. 2008), we observed that
theassociative AL and AFV stimulation enhances AL-inducedCa2+
responsesin bothlobes. To confirm the enhancementin both the and
the lobes, we employed the and
lobe-specific GAL4 drivers c739 and c305a, respectively(Yang et
al. 1995; Krashes et al. 2007).
As shown in Fig. 4A and B, LTE was significantlyinduced in both
lobes after associative AL and AFVstimulations. Notably, the PNs in
the AL received olfactoryinformation from the olfactory receptor
neurons (ORNs)in the maxillary palps (MPs) in addition to the
AN(Rajashekar & Shamprasad, 2004), and AN stimuli evokedmuch
higher Ca2+ responses in the lobe than in the lobe (Wang et al.
2008). Because we stimulated the entireAL, AL-induced Ca2+
responses and LTE in the lobemight reflect the activation of PNs
that receive input fromORNs in the MPs. Consistent withthe results
of a previousstudy (Wang et al. 2008), we did not observe
substantialLTE at the calyx (Fig. 4F and G), although
pre-synaptic
PNs form synapses with post-synaptic MB neurons at thecalyx
(Fig. 1A).
F and G, typical traces of AL-induced Ca2+ responses in the
vertical
lobe (F) and the calyx (G) before (pre) and after associative AL
and
AFV stimulation. Both traces were obtained from identical
OK107-GAL4/UAS-G-CaMPbrains. Hand I, LTE was observed after
associative stimulation in the vertical lobe (H) but not in the
calyx (I)
(P< 0.05 compared with pre responses). n = 67 for all
data.
C2012 The Authors. The Journal of Physiology
C2012 The Physiological Society
) by guest on August 3, 2013jp.physoc.orgDownloaded from J
Physiol (
http://jp.physoc.org/http://jp.physoc.org/http://jp.physoc.org/
-
8/22/2019 Long-Term Enhancement of Synaptic Transmission
Between
8/16
294 K. Ueno and others J Physiol591.1
Associativity, input specificity and persistence of LTE
LTP in the mammalian hippocampus and post-tetanicpotentiation in
the Drosophila neuromuscular junction(Zhong & Wu, 1991) can be
induced through theintensive stimulation of fibres of a single
input pathway.Therefore, we examined whether the association of
AL
and AFV stimuli is essential for LTE formation or whetherintense
AL stimulation alone is sufficient(Fig. 5A). Duringassociative
stimulation, robust increases in Ca2+ responseswere observed in MB
neurons (Fig. 5B and C). Toincrease Ca2+ responses in MB neurons
during intense
ALstimulationalone,weextendedthepulsedurationfrom1.0to 1.5 ms.
Although this intense AL stimulation producedincreases in Ca2+
responses comparable with associativestimulation (Fig. 5BD), no
subsequent enhancement ofAL-induced Ca2+ responses was observed
(Fig. 5Band D).Thus, LTE at the ALMB synapse probably requires
theassociation of AL and AFV stimuli.
The association of an odour with foot-shock selectivelyincreases
avoidance to the odour but not to the shock(Fig. 5E). This
observation prompted us to test the inputspecificity of LTE in the
MBs. As shown in Fig. 5F,the simultaneous stimulation of AL and AFV
enhancedAL-induced responses but not AFV-induced responses.This
indicates that although stimulation of the AFV isrequired for LTE
induction, LTE is selectively induced atALMB synapses but not at
AFVMB synapses.
To further assess the physiological properties of LTEat the ALMB
synapse, we examined the persistenceof this LTE. When we stimulated
AL at 120 min afterthe induction of LTE, we still observed a
significant
enhancement in the Ca2+
response (Fig. 6A, filled circles).However, when we stimulated
AL repeatedly every 5 minafter the associative stimulations, LTE
decayed rapidly anddisappeared within 1 h (Fig. 6B, filled
circles). This decaywas not theresult of synaptic transmission
fatigue betweenthe AL and MB or the desensitization of
postsynapticresponses, as repeated AL stimulation every 5 min
didnot decrease AL-induced Ca2+ responses (Fig. 6B,
opencircles).
Notably, the persistence of LTE is highly reminiscentof the
persistence of olfactory memory. Similar to LTE,the olfactory
memory formed by olfactory conditioningis extinguished through
subsequent repeated exposure tothe CS odours (Quinn et al. 1974;
Qin & Dubnau, 2010).Indeed, the 1 h memory after olfactory
conditioningwas extinguished through repetitive CS
presentationsapplied every 5 min (Fig. 6C). This effect was not due
todesensitization, as repeated exposure to the CS odoursevery 5 min
for 1 h prior to olfactory conditioningdid not affect the 3 min
memory, which representslearning ability and is lowered by
desensitization tothe odour used in subsequent olfactory
conditioning(Fig. 6D).
Figure 5. Associative stimulation of AL and AFV induces LTE
in
the ALMB synaptic transmission
A, stimulation protocols for associative stimulations
(associative) and
intense AL stimulations (AL alone). B, typical traces of
AL-induced
Ca2+ responses before, during and after associative or
AL-alone
stimulation. C and D, summary of AL-induced Ca2+ responses in
the
MB before and after associative (C) or AL-alone (D) stimulation.
The
shaded columns indicate Ca2+ responses during associative
and
intense AL-alone stimulation (P< 0.05 compared with pre
responses). E, avoidance of flies to 30 and 60 V electric
shocks. Flies
that had been previously trained to associate 3-octanol (OCT)
or
4-methylcyclohexanol (MCH) with foot-shocks did not exhibit
altered
shock avoidance. Avoidance tests were performed at 15 min
after
conditioning. F, AL- and AFV-induced Ca2+ responses before
(pre)
and 15 min after the associative stimulation (P< 0.05). NS,
not
significant; n = 68 for all data.
C2012 The Authors. The Journal of Physiology
C2012 The Physiological Society
) by guest on August 3, 2013jp.physoc.orgDownloaded from J
Physiol (
http://jp.physoc.org/http://jp.physoc.org/http://jp.physoc.org/
-
8/22/2019 Long-Term Enhancement of Synaptic Transmission
Between
9/16
J Physiol591.1 Synaptic plasticity in cultured Drosophila brain
295
nAChR and NMDA receptors mediate Ca2+ responses
and LTE in the MBs
In previous studies, it was demonstrated that thecholinergic
synaptic currents in MB neurons are mediatedthrough Drosophila
nicotinic acetylcholine receptors(dnAChRs), which are widely
expressed in Drosophila
brain including MBs (Jonas et al. 1994; Su & ODowd,2003).
Consistent with this result, we observed thatAL-induced Ca2+
responses were suppressed in the pre-sence of mecamylamine, an
nAChR antagonist (Kazama& Wilson, 2008; Fig. 7AC). However, the
AFV-inducedCa2+ responses were not suppressed by mecamylamine(Fig.
7Aand B).
In olfactory aversive conditioning, foot-shockinformation, which
is conveyed through the AFV, canbe substituted by artificial
stimulation of dopaminergic
(DAergic) neurons (Claridge-Chang et al. 2009; Aso et al.2010).
DA neurons innervate the MBs (Mao & Davis,2009), and the
dopamine D1 receptor, DopR (Sugamoriet al. 1995), is required in
the MBs for olfactory aversivememory formation (Kim et al. 2007).
Therefore, we werecurious to test whether AFV-induced Ca2+
responsesare mediated by DopR, although it is classified as
G-protein coupled receptor (GPCR) (Sugamori et al.1995).
However, AFV-induced Ca2+ responses were notsuppressed
byDopRIn(3LR)234 mutations (Kim et al. 2007)or exposure to
butaclamol, a DopR antagonist (Sugamoriet al. 1995; Fig. 7D and E).
AL-induced Ca2+ responseswere also unaffected by DopRIn(3LR)234
mutations andbutaclamol (Fig. 7F and G). These results suggest
thatneither AL- nor AFV-induced Ca2+ responses in the MBsare
mediated through DopR.
Figure 6. LTE is extinguished through
repetitive post-associative AL stimulation
A, AL-induced Ca2+ responses in the MB
measured at 60 and 120 min after associative
stimulation (filled circles) were still significantly
higher than the control responses (open
circles), indicating that LTE persists for at least120 min. B,
the enhanced Ca2+ responses
declined within 60 min through repetitive AL
stimuli, induced every 5 min, after associative
stimulation (filled circles). Ca2+ responses were
not diminished by repetitive AL stimuli in the
control MBs (open circles). C, extinction of
olfactory memory in trained flies through
repetitive presentation of the conditioned
odour. D, the extinction protocol administered
prior to olfactory conditioning did not affect
olfactory learning (P< 0.05 compared with
pre-associative responses). n = 68 for all data.
C2012 The Authors. The Journal of Physiology
C2012 The Physiological Society
) by guest on August 3, 2013jp.physoc.orgDownloaded from J
Physiol (
http://jp.physoc.org/http://jp.physoc.org/http://jp.physoc.org/
-
8/22/2019 Long-Term Enhancement of Synaptic Transmission
Between
10/16
296 K. Ueno and others J Physiol591.1
C2012 The Authors. The Journal of Physiology
C2012 The Physiological Society
) by guest on August 3, 2013jp.physoc.orgDownloaded from J
Physiol (
http://jp.physoc.org/http://jp.physoc.org/http://jp.physoc.org/
-
8/22/2019 Long-Term Enhancement of Synaptic Transmission
Between
11/16
J Physiol591.1 Synaptic plasticity in cultured Drosophila brain
297
Drosophila NMDA receptors (dNRs) are expressed inthe MBs and are
involved in olfactory memory formation(Xia et al. 2005; Wu et al.
2007; Sinakevitch et al. 2010;Miyashita et al. 2012); therefore, we
hypothesized thatdNRs might mediate the AFV-induced Ca2+
response.Consistent with this hypothesis, AFV-induced Ca2+
responses were suppressed in response to exposure to the
NR antagonist MK801 (Fig. 7HJ). These results indicatethat while
AL-induced Ca2+ responses are mediatedthrough dnAChRs, AFV-induced
Ca2+ responses aremediated through dNRs.
To test whether dnAChRs and dNRs are required forLTE formation,
each receptor antagonist was appliedduring associative stimulations
(Fig. 7K). As shown inFig. 7LQ, each antagonist suppressed LTE
formation.Because AL-induced Ca2+ responses are mediatedthrough
dnAChRs and the effects of mecamylaminelast approximately 10 min
after washout (Fig. 7C),mecamylamine treatment reduced AL-induced
Ca2+
responses at 5 min after associative stimulation (Fig. 7P).
DopR in the MBs is required for PAE formation
As reported previously (Kim et al. 2003, 2007), DopRis
preferentially expressed in MB and required forolfactory aversive
memory formation (Fig. 8A), althoughit mediates neither AL nor
AFV-induced Ca2+ responses.To address whether DopRs are necessary
for inducing LTE,we examined LTE in DopR mutants. Consistent with
thebehavioural data, LTE was not produced in DopRmutants(Fig. 8B
and C). Furthermore, LTE formation in
controlbrainswassuppressedinresponsetobutaclamoltreatment
(Fig. 8D and E) during associative stimulations (seeFig. 7K),
indicating that DopR is physiologically requiredfor LTE formation.
Interestingly, Ca2+ responses duringthe associative stimulation
were enhanced above controllevels as a result of the DopR mutation
(Fig. 8F) and inresponse to butaclamol treatment (Fig. 8G),
although AL-and AFV-induced Ca2+ responses were not different
fromthose in the control (Fig. 7DG).
Considering that DopR mutations do not affectAFV-inducedCa2+
responsesintheMBs,DopRsexpressedin areas outside of MB neurons
might be required forLTE in contrast to the DopR requirement for
olfactoryaversive memory formation. To test this possibility,
weused rescue experiments with DopRf02676 mutants (Kimet al. 2007).
DopRf02676 is a null allele as a result of inter-
ference of transcription from its endogenous promoterby the
insertion of piggyBac in the first intron. ThispiggyBac insertion
acts as a carrier of rescue constructin the presence of GAL4, as
the inserted piggyBac containsUAS, whichfunctions as an exogenous
enhancer/promoterwhen bound by GAL4 to initiate transcription of
thedownstream gene (Thibault et al. 2004). We used boththe GAL4/UAS
(Brand & Perrimon, 1993) and theLexA/LexAop binary systems (Lai
& Lee, 2006; Fig. 9Aand B) to generate transgenic flies
expressing both theDopR transgene and G-CaMP2 (Tallini et al.
2006)independently in the MBs ofDopRf02676. Similar to hypo-
morphic DopRIn(3LR)234
mutants, LTE did not occur in theDopRf02676-null mutants (Fig.
9C and D). However, in arescue line expressing DopRsin the MBs, LTE
was restoredto the control level (Fig. 9E). Notably, while LTE
wasrestored in this transgenic line, the hyper-enhancementof Ca2+
responses during associative stimulation was notreduced to normal
levels (Fig. 9F). These results suggestthat while DopR in the MBs
is essential for LTE formation,the larger Ca2+ response during
associative stimulationresults from defects in DopRexpressed in
areas outside ofthe MBs.
Discussion
Using in vitro imaging, we directly measured ALMBsynaptic
transmission and successively observed theinduction, persistence
and decline of LTE at the ALMBsynapse. Previous studies have shown
that odour-inducedCa2+ responses in the MBs were increased more
than 1 hafter single-cycle olfactory aversive conditioning
(Wang
Figure 7. dnAChR and dNR activities during associative
stimulation are required for LTE
A, typical tracesof AL-and AFV-induced Ca2+ responses before and
afterthe application of 100 M mecamylamine.
B, summary of the effects of mecamylamine on AL- and AFV-induced
Ca 2+ responses (P< 0.05). C, after bath
application of 100 M mecamylamine, AL-induced Ca2+
responses were decreased and abolished within 9
min.Subsequently, the responses were restored at 10 min after
washout (18 min). D and E, neither DopR mutations nor
the DopR antagonist (50 M butaclamol) altered AFV-induced Ca2+
responses. Typical traces of AFV-induced Ca2+
responses (D) and the summary ofthe effects of DopR inhibitions
(E). Fand G, neither DopR mutationsnor theDopR
antagonist altered AL-induced Ca2+ responses. H, AFV-induced
Ca2+ responses were suppressed using 100 M
MK801, while mecamylamine and the AMPA receptor antagonist CNQX
(6-cyano-7-nitroquinoxaline-2,3-dione)
had no effect (P< 0.05 compared with the response at 0 min).
I and J, MK801 suppressed AFV-induced Ca2+
responses but not AL-induced Ca2+ responses. Typical AL- and
AFV-induced Ca2+ traces (I), and averaged peak
responses in saline (filled columns) and after (open columns)
application of 100 M MK801 (J) (P< 0.05). KQ,
both dnAChR and dNR activities during associative stimulation
are required for LTE formation. K, experimental
design. While LTE was induced in control conditions (L, O), the
application of mecamylamine (M, P) and MK801
(N, Q) during associative stimulation prevented LTE induction
(P< 0.05 compared with pre-associative responses
(pre). n = 68 for all data.
C2012 The Authors. The Journal of Physiology
C2012 The Physiological Society
) by guest on August 3, 2013jp.physoc.orgDownloaded from J
Physiol (
http://jp.physoc.org/http://jp.physoc.org/http://jp.physoc.org/
-
8/22/2019 Long-Term Enhancement of Synaptic Transmission
Between
12/16
298 K. Ueno and others J Physiol591.1
et al. 2008; Tan et al. 2010). However, the synaptic basisof
such neural plasticity in the MBs is not understood.Odour
information received through ORNs in the ANand MPs is transmitted
to PNs in the AL, which in turnconvey signals to MB neurons.
Although synaptic trans-mission from ORNs to PNs is also increased
after olfactoryconditioning, this increase disappears within 10 min
(Yu
et al. 2004). Thus, it has been suggested that memory tracemight
be formed through the enhancement of ALMB
synaptic transmission. Consistent with this hypothesis,LTE at
the ALMB synapse lasts more than 2 h.
ALMB LTE exhibits characteristic physiologicalproperties,
including associativity, input specificity andpersistence.
Importantly, these properties are alsoobserved in olfactory
aversive memory. While ALMBLTE formation requires the correlated
activity of AL
and AFV inputs, olfactory aversive memory formationrequires the
correlated presentation of CS odour and
Figure 8. DopR activity during associative stimulation is
required for LTE
A, 5 min memory was impairedin the DopR mutant, DopRIn(3LR)234 .
B and C, DopR mutants are defectivefor ALMB
LTE formation. D and E, application of 50 M butaclamol during
associative stimulation blocked LTE induction. F
and G, DopR mutations and butaclamol treatment further increased
Ca2+ responses during associative stimulation
compared with the wild-type control and DMSO treatment (P<
0.05). n = 58 for all data.
C2012 The Authors. The Journal of Physiology
C2012 The Physiological Society
) by guest on August 3, 2013jp.physoc.orgDownloaded from J
Physiol (
http://jp.physoc.org/http://jp.physoc.org/http://jp.physoc.org/
-
8/22/2019 Long-Term Enhancement of Synaptic Transmission
Between
13/16
J Physiol591.1 Synaptic plasticity in cultured Drosophila brain
299
US foot-shock. In olfactory aversive conditioning, odourand
foot-shock signals are associated in the MBs.In this study, we
demonstrated that signals deliveredfrom the AL and AFV are
associated in the MBs andproduce LTE at ALMB synapses. Trained
flies increaseavoidance to CS odour but not to US
foot-shock.Similarly, LTE is specifically formed at ALMB but
not
AFVMB synapses. This input specificity of LTE mightexplain why
avoidance to foot-shock is not increasedfollowing olfactory
aversive conditioning. Furthermore,both ALMB LTE and olfactory
memory were reducedafter subsequent repetitive AL stimulations and
CS odourpresentations, respectively. This suggests that
attenuatedALMB synaptic transmission might be involved in
Figure 9. DopRs expressed in the MBs are required for LTE
formation
A, G-CaMP2 fluorescence observed in the MBs of an
MBp-LexA;LexAop-G-CaMP2 fly. B, G-CaMP2 fluorescence
is observed in all , , , and lobes of an MBp-Lex;LexAop-G-CaMP2
brain. CE, ALMB LTE
(C) disrupted in DopR mutants (D) was restored through
expression of the DopR+ transgene in the
MBs (E). AL-induced Ca2+ responses before (pre) and after
associative stimulation were recorded in the
MBs from control MBp-Lex,LexAop-G-CaMP2/+ (C),
MBp-Lex,LexAop-GCaMP2/+;DopRf02676/DopRf02676 (D)
and MBp-Lex,LexAop-GCaMP2/c309;DopRf02676/DopRf02676 flies (E)
(P< 0.05 compared with pre-associative
stimulation). F, the larger increase in Ca2+ responses during
associative stimulation in DopRf02676 (open column)
were not decreased in the rescue line (shaded column) (P<
0.05 compared with control brains). n = 67 for all
data.
C2012 The Authors. The Journal of Physiology
C2012 The Physiological Society
) by guest on August 3, 2013jp.physoc.orgDownloaded from J
Physiol (
http://jp.physoc.org/http://jp.physoc.org/http://jp.physoc.org/
-
8/22/2019 Long-Term Enhancement of Synaptic Transmission
Between
14/16
300 K. Ueno and others J Physiol591.1
the memory extinction process. In addition to thesephysiological
similarities, both ALMB LTE and olfactoryaversive memory require
the activity of dNRs and DopRsduring association. These results
demonstrate that LTE atthe ALMB synapse can be an appropriate
cellular modelfor olfactory memory.
In this study, we employed 20mM Mg2+ in the
external recording solution because various groups haveshown
that the Mg2+ concentration in the Drosophilahaemolymph ranges
between 20 and 33 mM (Croghan &Lockwood, 1960; Begg &
Cruickshank, 1963; Stewart et al.1994). In mammals, the Mg2+
concentration is higher inthe cerebrospinal fluid than in the
plasma (McKee et al.2005). Furthermore, while 10 mM Mg2+
concentrationis not sufficient for the Mg2+ blockade of
DrosophilaNMDA receptors, 20 mM Mg2+ is sufficient to producethis
effect (Miyashita et al. 2012). We thus propose that the20 mM Mg2+
concentration used in our study lies withinthe physiologically
relevant range. We employed a much
higher Mg2+
concentration (20 mM) than those used inprevious in vivo and in
vitro imaging studies (
-
8/22/2019 Long-Term Enhancement of Synaptic Transmission
Between
15/16
J Physiol591.1 Synaptic plasticity in cultured Drosophila brain
301
Aso Y, Siwanowicz I, Bracker L, Ito K, Kitamoto T &
TanimotoH (2010). Specific dopaminergic neurons for theformation of
labile aversive memory. Curr Biol20,14451451.
Begg M & Cruickshank WJ (1963). A partial analysis
ofDrosophilalarval haemolymp. Proc Roy Soc Edinburgh68,215236.
Brand AH & Perrimon N (1993). Targeted gene expression as
ameans of altering cell fates and generating dominantphenotypes.
Development 118, 401415.
Claridge-Chang A, Roorda RD, Vrontou E, Sjulson L, Li H,Hirsh J
& Miesenbock G (2009). Writing memories withlight-addressable
reinforcement circuitry. Cell 139,405415.
Crittenden JR, Skoulakis EM, Han KA, Kalderon D & Davis
RL(1998). Tripartite mushroom body architecture revealed
byantigenic markers. Learn Mem 5, 3851.
Croghan PC & Lockwood APM (1960). The composition of
thehaemolyph of the larva ofDrosophila melanogaster. J ExpBiol 37,
339343.
Daw NW, Stein PS & Fox K (1993). The role of NMDA
receptors in information processing. Annu Rev
Neurosci16,207222.
de Belle JS & Heisenberg M (1994). Associative odour
learningin Drosophilaabolished by chemical ablation of
mushroombodies. Science263, 692695.
Diegelmann S, Bate M & Landgraf M (2008). Gateway
cloningvectors for the LexA-based binary expression system
inDrosophila. Fly (Austin) 2, 236239.
Dubnau J, Grady L, Kitamoto T & Tully T (2001). Disruptionof
neurotransmission in Drosophilamushroom body blocksretrieval but
not acquisition of memory. Nature411,476480.
Dura JM, Preat T & Tully T (1993). Identification oflinotte,
anew gene affecting learning and memory in Drosophilamelanogaster.
J Neurogenet 9, 114.
Gruart A & Delgado-Garcia JM (2007).
Activity-dependentchanges of the hippocampal CA3-CA1 synapse during
theacquisition of associative learning in conscious mice.
GenesBrain Behav 6(Suppl 1), 2431.
Ito K, Awano W, Suzuki K, Hiromi Y & Yamamoto D (1997).The
Drosophilamushroom body is a quadruple structure ofclonal units
each of which contains a virtually identical set ofneurones and
glial cells. Development 124, 761771.
Jonas PE, Phannavong B, Schuster R, Schroder C &Gundelfinger
ED (1994). Expression of the ligand-bindingnicotinic acetylcholine
receptor subunit D alpha 2 in theDrosophilacentral nervous system.
J Neurobiol 25,
14941508.Kazama H & Wilson RI (2008). Homeostatic matching
and
nonlinear amplification at identified central synapses.Neuron58,
401413.
Kim YC, Lee HG & Han KA (2007). D1 dopamine receptordDA1 is
required in the mushroom body neurons foraversive and appetitive
learning in Drosophila. J Neurosci27,76407647.
Kim YC, Lee HG, Seong CS & Han KA (2003). Expression of aD1
dopamine receptor dDA1/DmDOP1 in the centralnervous system
ofDrosophila melanogaster. Gene ExprPatterns 3, 237245.
Krashes MJ, Keene AC, Leung B, Armstrong JD & Waddell
S(2007). Sequential use of mushroom body neuron subsetsduring
Drosophilaodour memory processing. Neuron53,103115.
Lai SL & Lee T (2006). Genetic mosaic with dual
binarytranscriptional systems in Drosophila. Nat
Neurosci9,703709.
Mao Z & Davis RL (2009). Eight different types
ofdopaminergic neurons innervate the Drosophilamushroombody
neuropil: anatomical and physiological heterogeneity.Front Neural
Circuits3, 5.
Marin EC, Jefferis GS, Komiyama T, Zhu H & Luo L
(2002).Representation of the glomerular olfactory map in
theDrosophilabrain. Cell 109, 243255.
Martin JR, Rogers KL, Chagneau C & Brulet P (2007). In
vivobioluminescence imaging of Ca signalling in the brain
ofDrosophila. PLoS One2, e275.
McGuire SE, Le PT & Davis RL (2001). The role of
Drosophilamushroom body signalling in olfactory memory. Science293,
13301333.
McKee JA, Brewer RP, Macy GE, Phillips-Bute B, Campbell KA,
Borel CO, Reynolds JD & Warner DS (2005). Analysis of
thebrain bioavailability of peripherally administeredmagnesium
sulfate: a study in humans with acute braininjury undergoing
prolonged induced hypermagnesemia.Crit Care Med 33, 661666.
Miyashita T, Oda Y, Horiuchi J, Yin Jerry CP, Morimoto T
&Saitoe M (2012). Mg2+ block ofDrosophilaNMDA receptorsis
required for long-term memory formation andCREB-dependent gene
expression. Neuron74, 887898.
Perez-Orive J, Mazor O, Turner GC, Cassenaer S, Wilson RI
&Laurent G (2002). Oscillations and sparsening of
odourrepresentations in the mushroom body. Science297,359365.
Qin H & Dubnau J (2010). Genetic disruptions
ofDrosophilaPavlovian learning leave extinction learning intact.
GenesBrain Behav9, 203212.
Quinn WG, Harris WA & Benzer S (1974). Conditionedbehaviour
in Drosophila melanogaster. Proc Natl Acad Sci U SA71, 708712.
Rajashekar KP & Shamprasad VR (2004). Maxillary
palpglomeruli and ipsilateral projections in the antennal lobe
ofDrosophila melanogaster. J Biosci29, 423429.
Sinakevitch I, Grau Y, Strausfeld NJ & Birman S
(2010).Dynamics of glutamatergic signalling in the mushroom bodyof
young adult Drosophila. Neural Dev5, 10.
Stewart BA, Atwood HL, Renger JJ, Wang J & Wu CF
(1994).Improved stability ofDrosophilalarval neuromuscular
preparations in haemolymph-like physiological solutions.J Comp
Physiol A175, 179191.
Su H & ODowd DK (2003). Fast synaptic currents
inDrosophilamushroom body Kenyon cells are mediated
by-bungarotoxin-sensitive nicotinic acetylcholine receptorsand
picrotoxin-sensitive GABA receptors. J Neurosci23,92469253.
Sugamori KS, Demchyshyn LL, McConkey F, Forte MA &Niznik HB
(1995). A primordial dopamine D1-like adenylylcyclase-linked
receptor from Drosophila melanogasterdisplaying poor affinity for
benzazepines. FEBS Lett 362,131138.
C2012 The Authors. The Journal of Physiology
C2012 The Physiological Society
) by guest on August 3, 2013jp.physoc.orgDownloaded from J
Physiol (
http://jp.physoc.org/http://jp.physoc.org/http://jp.physoc.org/
-
8/22/2019 Long-Term Enhancement of Synaptic Transmission
Between
16/16
302 K. Ueno and others J Physiol591.1
Tallini YN, Ohkura M, Choi BR, Ji G, Imoto K, Doran R, Lee
J,Plan P, Wilson J, Xin HB, Sanbe A, Gulick J, Mathai J,Robbins J,
Salama G, Nakai J & Kotlikoff MI (2006). Imagingcellular
signals in the heart in vivo: cardiac expression of thehigh-signal
Ca2+ indicator GCaMP2. Proc Natl Acad Sci U SA103, 47534758.
Tamura T, Chiang AS, Ito N, Liu HP, Horiuchi J, Tully T
&
Saitoe M (2003). Aging specifically impairsamnesiac-dependent
memory in Drosophila. Neuron40,10031011.
Tan Y, Yu D, Pletting J & Davis RL (2010).
Gilgameshisrequired for rutabaga-independent olfactory learning
inDrosophila. Neuron67, 810820.
Thibault ST, Singer MA, Miyazaki WY, Milash B, Dompe NA,Singh
CM, Buchholz R, Demsky M, Fawcett R, Francis-LangHL, Ryner L,
Cheung LM, Chong A, Erickson C, Fisher WW,Greer K, Hartouni SR,
Howie E, Jakkula L, Joo D, Killpack K,Laufer A, Mazzotta J, Smith
RD, Stevens LM, Stuber C, TanLR, Ventura R, Woo A, Zakrajsek I,
Zhao L, Chen F,Swimmer C, Kopczynski C, Duyk G, Winberg ML
&Margolis J (2004). A complementary transposon tool kit for
Drosophila melanogasterusing P and piggyBac. Nat Genet36,
283287.
Tomchik SM & Davis RL (2009). Dynamics oflearning-related
cAMP signalling and stimulus integrationin the Drosophilaolfactory
pathway. Neuron64,510521.
Tsydzik V & Wright NJ (2009). Dopamine modulation of the
invivoacetylcholine response in the Drosophilamushroombody. Dev
Neurobiol69, 705714.
Tully T & Quinn WG (1985). Classical conditioning
andretention in normal and mutant Drosophila melanogaster.
J Comp Physiol A157, 263277.Wang Y, Mamiya A, Chiang AS &
Zhong Y (2008). Imaging of
an early memory trace in the Drosophilamushroom body.J
Neurosci28, 43684376.
Wong AM, Wang JW & Axel R (2002). Spatial representation
ofthe glomerular map in the Drosophilaprotocerebrum. Cell109,
229241.
Wu CL, Xia S, Fu TF, Wang H, Chen YH, Leong D, Chiang AS&
Tully T (2007). Specific requirement of NMDA receptorsfor long-term
memory consolidation in Drosophilaellipsoidbody. Nat Neurosci10,
15781586.
Xia S, Miyashita T, Fu TF, Lin WY, Wu CL, Pyzocha L, Lin
IR,Saitoe M, Tully T & Chiang AS (2005). NMDA receptorsmediate
olfactory learning and memory in Drosophila. CurrBiol 15,
603615.
Yang MY, Armstrong JD, Vilinsky I, Strausfeld NJ & Kaiser
K(1995). Subdivision of the Drosophilamushroom bodies
byenhancer-trap expression patterns. Neuron15, 4554.
Yasuyama K, Meinertzhagen IA & Schurmann FW (2002).Synaptic
organization of the mushroom body calyx inDrosophila melanogaster.
J Comp Neurol 445, 211226.
Yu D, Akalal DB & Davis RL (2006). Drosophila/ mushroombody
neurons form a branch-specific, long-term cellularmemory trace
after spaced olfactory conditioning. Neuron52, 845855.
Yu D, Ponomarev A & Davis RL (2004). Altered
representationof the spatial code for odors after olfactory
classicalconditioning; memory trace formation by
synapticrecruitment. Neuron42, 437449.
Yuan N & Lee D (2007). Suppression of excitatory
cholinergicsynaptic transmission byDrosophiladopamine
D1-likereceptors. Eur J Neurosci26, 24172427.
Zhong Y & Wu CF (1991). Altered synaptic plasticity
inDrosophilamemory mutants with a defective cyclic AMPcascade.
Science251, 198201.
Author contributions
K.U. designed and performed most of the experiments. S.N.
contributed to the imaging study, and Y.H. contributed to
thegenetics and behaviour studies. M.S. supervised and wrote
the
manuscript with K.U. and J.H.
Acknowledgements
We thank J. Dubnau (Cold Spring Harbor Laboratory)for DopR
mutants, K. Scott (University of California,
Berkeley) for UAS-G-CaMP transgenic flies, S. Diegelmann
(University of Cambridge, UK) for pCasper-W-LexA::GAD and
pCasper-lexAop-W vectors and J. Nakai (Saitama University,
Japan) for G-CaMP2. We also thank Ms Fukuda and MsOfusa for
stock maintenance and rearing flies. This research
was supported by a Grant-in-Aid for Scientific Research in
Innovative Areas Systems Molecular Ethology to M.S., and
MEXT/JSPS KAKENHI Grant (21700376 and 23700405) to K.U.
C2012 The Authors The Journal of Physiology
C2012 The Physiological Society