Embed Size (px)
Citation preview

8310892-6638/98/0012-0831/$02.25 Q FASEB
/ 382c 0012 Mp 831 Monday Jul 13 11:49 AM LP–FASEB 0012
Localization, quantification, and activation of platelet-activating factor receptor in human endometriumduring the menstrual cycle: PAF stimulates NO,VEGF, and FAKpp125
ASIF AHMED,*,1 SHARON DEARN,* MUNJIBA SHAMS,* XIAO F. LI,*RAJBANT K. SANGHA,* MAREK ROLA-PLESZCZYNSKI,† JIANQIAO JIANG**Reproductive Physiopathology Group, Departments of Obstetrics and Gynaecology, BirminghamWomen’s Hospital, University of Birmingham, Edgbaston, Birmingham, B15 2TG, U.K.; and†Immunology Division, Faculty of Medicine, Universite de Sherbrooke, Quebec, Canada
ABSTRACT Implantation is characterized by an in-flammatory-like response with expansion of extracel-lular fluid volume, increased vascular permeability,and vasodilatation. These effects are believed to bemediated at the paracrine level by prostaglandin E2
and platelet-activating factor (PAF), but the cellularmechanism (or mechanisms) remains largely un-known. We demonstrate that PAF receptor (PAF-R)immunoreactivity and mRNA are detected in prolif-erative and secretory endometrial glands, however,the responsiveness of endometrium to physiologicalconcentrations of PAF is confined predominantly tothe secretory endometrium. Semiquantitative reversetranscription-polymerase chain reaction revealed thatPAF-R transcript levels were highest in the mid-lateproliferative and late secretory phases of the cycle.Interaction of PAF with its receptor resulted in therapid release of nitric oxide (NO), increased expres-sion of vascular endothelial growth factor (VEGF),and activation of FAKpp125, a focal adhesion kinase,demonstrating that the PAF-R is functionally active.Inhibition of NO synthesis by NG-monomethyl-L-ar-ginine produced dose-dependent attenuation of PAF-evoked NO release, indicating NOS activation; thedependency of PAF-evoked NO release on PKC andextracellular Ca2/ was confirmed by PKC inhibitorRo 31–8220 and by the removal of extracellular Ca2/.PAF up-regulated VEGF gene expression in a concen-tration- and time-dependent fashion in human en-dometrial epithelial cell lysates. Transcription ofVEGF was rapidly followed by secretion of the pro-tein. These data support our premise that this auto-coid acts as an angiogenic mediator in the regenera-tion of the endometrium after menses and as avasodilator to promote blastocyst attachment duringthe implantation process.—Ahmed, A., Dearn, S.,Shams, M., Li, X. F., Sangha, R. K., Rola-Pleszczynski,M., Jiang, J. Localization, quantification, and activa-tion of platelet-activating factor receptor in humanendometrium during the menstrual cycle: PAF stim-
ulates NO, VEGF, and FAKpp125. FASEB J. 12, 831–843 (1998)
Key Words: PAF receptor · uterus · nitric oxide · menstruation· implantation
SUCCESSFUL MAMMALIAN pregnancy requires thepreparation of a receptive endometrium into whichthe embryo attaches and invades. Implantation ischaracterized by an inflammatory-type response withexpansion of extracellular fluid volume, increasedvascular permeability, and vasodilatation (1). Plate-let-activating factor (PAF)2 is a potent inflammatorylipid mediator that increases vascular permeabilityand vasodilatation, processes that accompany humanimplantation (2). PAF is produced by endometrialstromal cells, where its levels are hormonally regu-lated (3). In vitro PAF acts on glandular epitheliumto release prostaglandins (4), and PAF antagonistsinhibit implantation (5).
Actions of PAF are mediated by a specific PAF re-ceptor (PAF-R). Molecular cloning of the PAF-R froma human leukocyte cDNA library has identified aseven-transmembrane spanning, G-protein-linked re-ceptor superfamily that encodes a 342 amino acid
1 Correspondence: Department of Obstetrics and Gynaecol-ogy, Birmingham Women’s Hospital, University of Birming-ham, Edgbaston, Birmingham, B15 2TG U.K. E-mail:[email protected]
2 Abbreviations: PAF, platelet-activating factor; PAF-R, PAFreceptor; PGE2, prostaglandin E2; VEGF, vascular endothelialgrowth factor; NO, nitric oxide; NOS, NO synthase; PBS, phos-phate-buffered saline; HBSS, Hank’s balance salt solution;DMEM, Dulbecco’s modified Eagle’s medium; BSA, bovineserum albumin; FCS, fetal calf serum; HES, human endome-trial epithelial cell line; RT-PCR, reverse transcriptase-poly-merase chain reaction; L-NMMA, NG-monomethyl-L-arginine;SDS, sodium dodecyl sulfate; TBS-T, Tris-buffered saline-Tween; MP, mid-proliferate; MS, mid-secretory; LS, late secre-tory.

832 Vol. 12 July 1998 The FASEB Journal AHMED ET AL.
/ 382c 0012 Mp 832 Monday Jul 13 11:49 AM LP–FASEB 0012
protein with a calculated molecular mass of approx-imately 39 kDa (6). The human PAF-R gene containsno introns in its coding region and maps to chro-mosome 1 (7, 8). Although autoradiographic studiesshow binding of [3H]PAF to rabbit endometrial epi-thelium and to the embryonic disc of day 6 rabbitblastocysts (9), and reverse transcription-polymerasechain reaction (RT-PCR) demonstrated PAF-RmRNA in proliferative and secretory endometrium(10), the exact location of the receptors in humanendometrium remains to be determined. This is es-pecially important since the responsiveness of the en-dometrium to PAF appears to be confined to the se-cretory phase of the menstrual cycle, as demonstratedby PAF-stimulated prostaglandin E2 (PGE2) release(4), PAF-mediated activation of phospholipase C(11), and phospholipase D (12) activity in human se-cretory endometrium. Moreover, there is potentia-tion of PAF-evoked phospholipase D activity by 17b-estradiol in the human endometrial cell line (13).
We provide the first evidence of PAF-R localizationin uterine tissue and quantitative changes in mRNAencoding the PAF-R throughout the menstrual cycle,indicating sex steroid regulation of PAF-R in vivo. Wealso demonstrate that PAF may modulate neovascu-larization through the expression of vascular endo-thelial growth factor (VEGF) in human endome-trium and that the potent vasodilatory properties ofPAF may relate to its ability to stimulate the releaseof nitric oxide (NO), the formation of which isachieved by the conversation of L-arginine to L-cit-rulline by NO synthase (NOS) in the presence of mo-lecular oxygen (14).
MATERIALS AND METHODS
Patient selection
Normal endometrial tissues were obtained from fertile sub-jects undergoing sterilization or from couples complaining ofinfertility with solely tubal damage and/or male infertility. Allfemale subjects had regular menstrual bleeding with a men-strual cycle of 30{5 days for the previous 3 months. Womenwho had received any form of exogenous hormones, had usedan intrauterine device in the previous 3 months, or exhibiteddemonstrable uterine pathology were excluded from thestudy. Endometrial biopsies were collected by routine dilata-tion and curettage. Sections from each endometrial speci-mens were dated from the last menstrual period and wereused in the study only if there was corroboration by indepen-dent histological dating based on the criteria of Fox and Buck-ley (15). Ethical committee approval was obtained from theSouth Birmingham Ethical Committee.
Tissue collection
Endometrial tissue was rinsed in ice-cold phosphate-bufferedsaline (PBS) on ice to remove excess blood. Tissue was eitherimmediately immersed in 4% formaldehyde and processed forparaffin wax embedding, rapidly frozen in liquid N2 for RNA
and protein extraction, or rapidly frozen over dry ice, wrappedin Parafilm to prevent dehydration, and stored at 0807C untilsectioned for in situ hybridization studies. For functional stud-ies, endometrium was collected in sterile Hanks’ balanced saltsolution (HBSS, Gibco Ltd., Uxbridge, U.K.) and transportedon ice to the laboratory.
Tissue explant procedure and cell line conditions
Endometrial biopsies (1 mm3 size) were placed on square ster-ile capillary matting in a multiwell incubating plate as previ-ously described (11). Tissue was preincubated in Dulbecco’smodified Eagle’s medium (DMEM, Gibco Ltd., Uxbridge,U.K.) supplemented with Hepes buffer (20 mM; Gibco Ltd.,Uxbridge, U.K.), L-glutamine (10 mM; Gibco Ltd., Uxbridge,U.K.), glucose (250 mM; Sigma Chemical Co. Ltd., Poole, Dor-set, U.K.), and 0.1% bovine serum albumin (BSA, SigmaChemical Co. Ltd., Poole, Dorset, U.K.) for 1 h at 377C in 95%O2 and 5% CO2 at 95% humidity prior to agonist stimulation.
A spontaneously transformed human endometrial epithe-lial cell line (HES) generated by repeated passaging of cellsobtained by routine dilatation and curettage was kindly pro-vided by Dr. D. A. Kniss, Department of Obstetrics and Gy-necology, Ohio State University, Columbus, Ohio. HES cellswere maintained in 25 cm2 flasks in medium 199 (M199; GibcoLtd., Uxbridge, U.K.) containing 10% (v/v) fetal calf serum(FCS; Gibco Ltd., Uxbridge, U.K.), 1% L-glutamine (GibcoLtd., Uxbridge, U.K.), and 1% antibiotic mixture (10,000 Upenicillin and 10 mg streptomycin; Gibco Ltd., Uxbridge,U.K.) at 377C in 95% O2 and 5% CO2 at 95% humidity. Priorto agonist stimulation, growth medium was removed and sub-confluent (85%) monolayers were serum starved for 24 h inserum-free M199 supplemented with 0.2% BSA.
Immunocytochemistry
Serial 4 mm sections of formalin-fixed, paraffin-embedded tis-sue were used for immunohistochemistry as previously de-scribed in detail (16). Briefly, sections were deparaffinized,hydrated, and endogenous peroxidase activity was quenchedby the addition of 0.3% (v/v) of hydrogen peroxide for 10min. Nonspecific protein binding sites were blocked with 10%normal goat serum in PBS for 30 min at room temperature ina humidified chamber. Sections were incubated with rabbitpolyclonal anti-PAF-R antibody reported by Muller et al. (17)at 1:200 dilution in 10% goat serum and 0.3% Triton X-100for 60 min at room temperature. Subsequently, the sectionswere incubated with a goat anti-rabbit IgG biotinylated sec-ondary antibody and the localization of PAF-R was detectedusing a streptavidin-biotyn immunoperoxidase kit for rabbitIgG (DAKO Ltd. Bucks, U.K.). Diaminobenzidine was used asthe chromogen. The immunostained sections were conter-stained with Mayer’s hematoxylin. Specificity of the immuno-reactions was determined by comparing the reactivity of theantibodies with that of 10% goat serum in PBS in the absenceof the primary antibody. Hematoxylin and eosin sections weredated, based on the criteria of Fox and Buckley (15).
The specificity of the anti-PAF-R antibody was confirmed bythe fact that PAF-R transfected CHO cells showed labelingwhereas untransfected cells did not. Western blots with anti-PAF-R do not work reliably; the receptor is probably too hy-drophobic to migrate adequately on gels.
Preparation of PAF-R probe
Total RNA from endometrium was prepared using RNAzoland quantified spectrophotometrically at 260 nm. Ethidiumbromide staining confirmed integrity and the quality of the

PAF AND ENDOMETRIAL REMODELING 833
/ 382c 0012 Mp 833 Monday Jul 13 11:49 AM LP–FASEB 0012
RNA was confirmed on 1% agarose-formaldehyde gel electro-phoresis. cDNA was synthesized from total RNA using the Su-perscript Preamplification System (GibcoBRL Life Technolo-gies, Paisley, Scotland). RT-PCR was performed as describedpreviously (10) and the RT-PCR product of 1.3 Kb was sub-cloned into the BamHI and HindIII sites of pBluescript KS.Sequencing analysis was performed on the PCR fragment toconfirm that it encoded the human PAF-R.
In situ hybridization
Frozen tissues was surrounded in embedding medium (Tissue-Teck O. C. T. Compound; Agag Scientific, Stanstead, Essex,U.K.) before 10 mm sections were cut and thaw mounted ontopoly-L-lysine-coated glass slides for in situ hybridization stud-ies. Sections were stored (for less than 2 wk) and dessicatedat 0807C until use. For transcription of the antisense or senseRNA probes, the plasmid-containing RT-PCR product was lin-earized with HindIII or BamHI and the transcripts were gen-erated by [3535S]UTP (1500 Ci mmol/l, Dupont NEN, Boston,Mass.) incorporation using the T7 or T3 RNA polymerases,respectively. The probes thus generated were single-strandedRNA probes and had a specific activity of 1 1 107 dpm/mgplasmid template. In situ hybridization was performed as pre-viously described (18). Pretreated sections were hybridizedwith either sense or antisense probes in hybridization bufferat a final concentration of 11 106 dpm/ml. Hybridization wascarried out at 507C for 16 h. Two adjacent endometrial sec-tions from each sample were hybridized with either sense orantisense probe during the same experiment. After serialwashing, the slides were dipped in photographic emulsion andstored in a lightproof box at 47C for 5 wk. The sections werethen developed and counterstained in Mayer’s hematoxylin.
Semiquantitative RT-PCR for PAF-R
cDNA was synthesized from total RNA. For initial amplifica-tion by PCR, 2 ml of six RT products from throughout themenstrual cycle were amplified with 2.5 units of Taq DNApolymerase and 20 mM of each mouse b-actin primer (5* b-actin: 5* GTTACCAACTGGGACGACA 3*; 3* b-actin: 5*TGGCCATCTCCTGCTCGAA 3*) in 100 ml of reaction mixcontaining 500 mM KCl, 200 mM Tris-HCl, and 2 mM MgCl2
as follows: 947C, 30s; 507C, 40s; 727C, 60s. Reactions were tem-porarily halted after 20–30 cycles by cooling to 47C and 10 mlof PCR product was removed from each tube. Reactions werethen restarted by heating to 947C and amplification was al-lowed to proceed for three additional cycles before removalof a another 10 ml of product. This was continued until eachPCR reaction had been sampled six times. All products (10 ml;six per initial RT product) were then analyzed by 2% agarosegel electrophoresis. Dilutions of RT products were madewhere necessary and the amplification procedure was re-peated until all samples were standardized for b-actin content.After standardization, 2 ml of appropriately diluted RT prod-ucts were amplified with five units of Taq DNA polymeraseand 10 mM of each human PAF-R primer (5* PAF-R: 5*TCTTCTGCAACCTGGTCATCA 3* [608 bp to 628 bp]; 3*PAF-R: 5* GATCTGGTTGAATGGCACAAC 3* [985 bp to 1005 bp])in 100 ml of reaction mix containing 500 mM KCl, 200 mMTris-HCl, and 1.5 mM MgCl2, as follows: 957C, 30 s; 607C, 50s; 727C, 60 s. Sampling was initiated two cycles after the initialsampling cycle for b-actin and reactions were temporarilyhalted every four cycles for sampling purposes. All productswere then analyzed by 2% agarose gel electrophoresis to ob-tain the PAF-R mRNA profile throughout the menstrual cycle.The entire procedure was repeated three more times.
RNAse protection assay
A shorter PAF-R cDNA template of 420 base pairs was obtainedby restriction digestion of the original plasmid containing theinsert with Blp1. The transcript was generated using T7 poly-merase, which hybridizes with human total RNA to yield a 420base pair protected fragment of human PAF-R mRNA. ThepTRI-b-actin control template (Ambion, Whitney, Oxon) con-tained a 247 base pair fragment of human cytoplasmic actin(b-actin) gene. The TRIPLE-script vector was transcribed to a298 base antisense RNA probe using T3 polymerase, whichgenerated a 247 base pair protected fragment of human b-actin mRNA. This was used as a constitutively expressed geneproduct to control for possible differences in RNA amountsbetween samples in order to allow accurate quantitation ofPAF-R mRNA abundance between RNA samples.
RNAse protection assay was carried out on an endometrialsample throughout the cycle as described previously (19). An-tisense RNA was transcribed from each plasmid using a[32P]-CTP (Dupont NEN) to specific activities of 0.4–1.2 1 109
cpm/mg. Briefly, sample RNA (20 mg) was mixed witha[32P]-labeled riboprobe (0.5–1.0 1 105 cpm), denatured at85 { 57C for 5 min, and incubated overnight at 457C. Non-hybridized RNA was removed by the addition of RNAse diges-tion; thereafter, RNAses were inactivated, and RNA was ex-tracted and precipitated. RNA pellets were resuspended inloading buffer; samples were denatured and electrophoresedon 4% polyacrylamide/8 mol/l urea gels. The gels werewashed in buffer containing 10% methanol and 10% aceticacid before transfer to Whatman 3 mm paper. The gels werecovered with saran wrap and dried under vacuum on a geldrier (BioRad Gel Dryer 583; BioRad Laboratories, Inc. Hert-fordshire, U.K.) at 807C for 30–60 min. Autoradiography wasperformed using X-ray film (Kodak Biomax MR) with inten-sifying screens at 0707C for 2 to 6 days. Quantitation of au-toradiographic signals was performed by densitometric anal-ysis, using densitometry computer software (Gelbase).Transcripts detected were assigned relative densitometricunits and the ratio of PAF-R/actin was calculated for eachsample. Negative controls included 20 mg of yeast tRNA in tworeaction tubes. Control reaction 1 included radiolabeledprobes in the absence of RNAse treatment, which should yieldthe full-length probes. Control reaction 2 was treated in thesame manner as the RNA samples, thereby testing for non-specific hybridization of radiolabeled probes.
The VEGF antisense control template was formulated froma 600 base pair cDNA (EcoR1) and cloned into pBluescriptSK/. Transcripts were generated using T3 polymerase, whichhybridizes with human total RNA to yield a 570 base pair pro-tected fragment of human VEGF mRNA. b-Actin was used asa constitutively expressed gene product for comparison ofVEGF mRNA abundance between RNA samples. Ribonucleaseprotection analyses were performed as described above.
Measurement of NO release by chemiluminesence
For concentration dependence experiments, increasing con-centrations of PAF-acether (De-hydro-PAF [C18]; Sigma Chem-ical Co. Ltd., Poole, Dorset, U.K.) in serum-free M199 buffercontaining 0.2% BSA were added to endometrial tissue ex-plants from both proliferative and secretory phases of themenstrual cycle. For experiments with PAF or vechicle in thepresence of the PAF-R antagonist, WEB 2170 (20), or the NOsynthesis inhibitor NG-monomethyl-L-arginine (L-NMMA), tis-sues were pretreated with the antagonists for 1 h and thenstimulated in a final volume of 1.0 ml at 377C for a another 2h. A similar procedure was adopted for experiments with lowCa2/ (150 nM) HBSS and protein kinase C inhibitor Ro 31–8220. Reactions were terminated by removal of the superna-

834 Vol. 12 July 1998 The FASEB Journal AHMED ET AL.
/ 382c 0012 Mp 834 Monday Jul 13 11:49 AM LP–FASEB 0012
tant, which was subsequently stored at 0807C for NO analysis.Levels of NO were measured in the gas phase using a SieversNOA 270B chemiluminescence analyzer as described previ-ously (21). The total amount of NO present in conditionedmedia was expressed as picomoles of NO/mg protein. Proteinwas extracted from endometrial tissue explants in PBS with 5M sodium hydroxide and 0.1% sodium dodecyl sulfate (SDS)at 857C for 1 h.
Western blot analysis
After exposure of cells to either PAF (0.1 nM–1 mM) or vehi-cle, the conditioned medium was removed and stored at0707C. The cells were washed with ice-cold PBS and trans-ferred to prefrozen Eppendorfs. After centrifugation (13,000rpm, 47C, 10 min), the supernatant was discarded and thepellet was frozen in liquid nitrogen for 5 min.
Samples were then allowed to thaw on ice in 100 ml ice-coldhigh salt buffer (KCl 0.4 M, Hepes pH 7.4 20 mM, DTT 1 mM,glycerol 20%, bacitracin 0.5 mg/ml, PMSF 40 mg/ml, pepsta-tin 5 mg/ml, leupeptin 5 mg/ml) prior to protein estimation.Protein extraction from the medium was performed by etha-nol precipitation. Media was mixed with three volumes (15ml) of ice-cold ethanol and stored at 0707C for 24 h. Aftercentrifugation (6000 rpm, 47C, 10 min), the supernatant wasdiscarded and pellets were resuspended in PBS. Protein con-centration was determined using the Bio-Rad protein assay(Bio-Rad Laboratories Inc., Herts, U.K.), with BSA as stan-dard.
The nonradioactive enhanced chemuluminascence West-ern blotting system (Amersham, Bucks, U.K.) was used to de-tect VEGF in cell lysates and conditioned medium as describedpreviously (19). Briefly, 50 mg total protein was separated byelectrophoresis on a polyacrylamide gel consisting of a stack-ing gel overlaying the separating gel. The samples were loadedin a final volume of 25 ml with sample buffer containing0.002% bromophenol blue and electrophoresed at 50 V forapproximately 2 h. A kaleidoscope protein marker (Bio-RadLaboratories, Richmond, Calif.) was run alongside the sam-ples. This protein marker is a protein standard calibrated mo-lecular weight and prestained, which allows accurate molec-ular weight determination of unknown protein. Afterelectrophoresis, the protein was transferred onto a nitrocel-lulose membrane in a cooling system (107C) overnight at 36V. The filter was blocked to reduce nonspecific binding of theantibody using Tris-buffered saline-Tween (TBS-T) containing10% Marvel and 2.5% BSA for 4 h. After washing in TBS-T,the membrane was incubated overnight at 47C with the VEGFantibody. The primary rabbit polyclonal anti-VEGF antibodyraised against a 20 amino acid peptide corresponding to res-idues 1 to 20, mapping at the amino terminus of humanVEGF, was used at a dilution of 1:1000. The filter was washedand incubated with the secondary anti-rabbit antibody for 1.5h at room temperature. After a final wash in TBS-T, the filterwas incubated for 1 min at room temperature in detectionreagent, immediately wrapped in saran wrap, and exposed forperiods of 30 s, 1 min, 5 min, and 10 min to X-ray film.
Tyrosine phosphorylation
Confluent monolayers of endometrial glandual epithelial cellsin 25 cm2 flasks were incubated with either PAF (1 mM), VEGF(1 ng/ml), or vehicle in a final volume of 5 ml of serum-freeM199 supplemented with 0.2% BSA. Reactions were termi-nated at the appropriate time (1, 5, 15, or 30 min after stim-ulation) by rapid aspiration and the cells were washed twicewith ice-cold PBS. Modified radioimmunoprecipitation assaybuffer (0.5 ml; RIPA: 50 mM Tris-HCL, pH 7.4, 1% NP-40,
0.25% sodium deoxycholate, 150 mM NaCl, 1 mM EGTA, 1mM PMSF, 1 mg/ml aprotinin, leupeptin, and pepstatin, 1 mMactivated Na3VO4, and 1 mM NaF) was added and the flaskswere rocked on an orbital shaker at 47C for 15 min. Cell lysateswere collected and centrifuged (13,000 rpm, 47C, 10 min).Total protein (25 mg) was electrophoresed in a 10% SDS-poly-acrylamide minigel at 50 V and proteins were electroblottedonto nitrocellulose for 3 h at 50 V. Blots were blocked for 2 hat room temperature in TBS-T containing 1% rabbit serum.After washing, the membranes were incubated overnight at47C with an anti-pTyr mAb (PY20: Affiniti, Exeter, U.K.) di-luted 1:2000. Blots were washed and incubated with a second-ary peroxidase-labeled anti-mouse antibody diluted 1:3000 for1 h at room temperature. After a final wash in TBS-T, blotswere incubated for 1 min at room temperature in detectionreagent, immediately wrapped in saran wrap, and exposed toX-ray film.
Statistical analysis
Statistical analysis was performed using Student’s paired t test.
RESULTS
Localization of PAF receptor protein
PAF-R-like immunoreactivity was detected in endo-metrium throughout the menstrual cycle in all tissuesexamined and showed marked differences in the pat-tern of staining between different phases of the men-strual cycle. PAF-R-like-immunoreactivity was de-tected in glandular epithelial cells throughout themenstrual cycle. Stromal cells, however, stainedstrongly in the early proliferative and the late secre-tory endometrium, whereas in the mid-late prolifer-ative and the mid-secretory endometrium they ap-peared relatively negative to the PAF-R antibody.During the early proliferative phase (EP, nÅ4), in-tense cytoplasmic immunostaining was seen in theglandular epithelium and stroma (Fig. 1A). In mid-proliferative endometrium (MP, nÅ6), however,staining was confined largely to endometrial glandswith relatively no staining of stromal cells (Fig. 1B).The endometrial blood vessels also showed little orno staining in the MP phase (Fig. 1B). During thelate proliferative phase (LP, nÅ4), the intensity ofPAF-R immunostaining varied from light to moderatein the endometrial glands whereas the endometrialstroma appeared negative (Fig. 1C). In mid-secretory(MS, nÅ5) endometrium, strong to moderate stain-ing was observed in the endometrial glands and lightstaining was also detected in the cytoplasm of vascularendothelial cells in endometrial blood vessels. Neg-ligible immunostaining was seen in the endometrialstromal cells (Fig. 1D). In the late secretory (LS, nÅ6)endometrium, strong to moderate staining was seenin the glandular epithelium whereas discrete cellswithin the stroma showed weak staining (Fig. 1E). Noimmunostaining was observed in secretory endome-

PAF AND ENDOMETRIAL REMODELING 835
/ 382c 0012 Mp 835 Monday Jul 13 11:49 AM LP–FASEB 0012
Figure 1. Immunohistochemical localization of PAF-R in human endometrium during the menstrual cycle. Strong PAF-R im-munoreactivity was seen in endometrial glandular (G) epithelial cells in early (A), mid- (B), and late (C) proliferative endo-metrium whereas no staining (arrows) was seen in the endometrial blood vessels (Bv) in these sections. In mid-secretory (D),staining was strong in glands and blood vessels, whereas glandular immunostaining was moderate to strong in late secretory (E)endometrium. Endometrial stroma showed strong PAF-R immunoreactivity in early proliferative (A) and thereafter was weak tononexistent throughout the cycle. Plate F is stained with nonimmune control serum. 1280.
trium (Fig. 1F) when using nonimmune serum in-stead of the primary antibody.
Localization of PAF-R mRNA
In situ hybridization using 35S-labeled antisense ri-boprobe demonstrated cyclic changes in the expres-sion of PAF-R mRNA in endometrial tissue during the
menstrual cycle, with the highest level in the LS(nÅ4) phase of the cycle. Comparison of the hybrid-ization signal obtained with sections of EP (nÅ3) en-dometrium with either control sense (Fig. 2A, B) orthe antisense (Fig. 2C, D) probe demonstrated noexpression of PAF-R mRNA in the early proliferativeendometrium. In contrast, the level of hybridizationsignal was high in the late proliferative (nÅ4) endo-

836 Vol. 12 July 1998 The FASEB Journal AHMED ET AL.
/ 382c 0012 Mp 836 Monday Jul 13 11:49 AM LP–FASEB 0012
Figure 2. In situ localization of PAF-R mRNA expression in proliferative endometrium. Bright-field (A, C, E) and dark-field (B,D, F) photomicrographs of sections of endometrium after in situ hybridization with 35S-labeled sense (A, B) and antisense (CF)PAF-R RNA probe. Early proliferative phase (AD) and late proliferative phase endometrium (E, F). Endometrial glands (G).Blood vessels (Bv). Bar Å 50 mm.
metrium and localized over the glands (Fig. 2E, F).Similarly, PAF-R mRNA expression was high in theearly secretory (nÅ3) endometrium, but was uni-formly distributed over the glandular epithelium andstroma (Fig. 3C, D). In the late secretory endome-trium, the hybridization signal persisted over theglands and stromal cells and increased in intensity(Fig. 3E, F). The degree of hybridization was consis-tently lower in the stroma compared with the glandsthroughout the menstrual cycle. A strong hybridiza-tion signal was seen around the blood vessels. Thespecificity of the signal was confirmed by the use ofa sense probe (Fig. 3A, B).
Semiquantitation of PAF-R mRNA throughout themenstrual cycle
To quantify changes in PAF-R gene expression in en-dometrium throughout the menstrual cycle, varia-
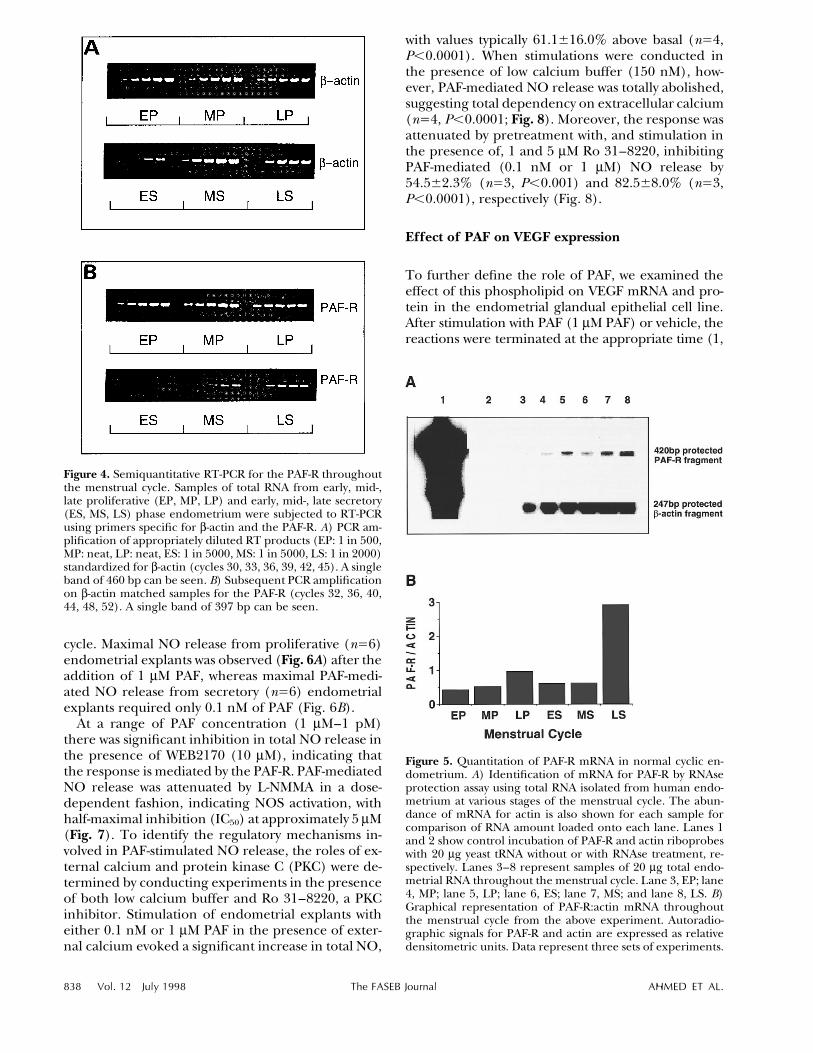
tions in transcript levels were determined using a se-miquantitative RT-PCR by normalization of theamount of PCR product for the PAF-R against thatfor b-actin. The overall trend in PAF-R mRNA ex-pression throughout the menstrual cycle, despiteconsiderable patient variation, showed that the tran-script levels were highest in endometrium from themid- to late proliferative and in the late secretoryphases of the menstrual cycle. Amplification of cDNAwith primers specific for mouse b-actin produced asingle band of the predicted size (460 bp). Figure 4Ashows amplification of appropriately diluted RTproducts from throughout the menstrual cycle stan-dardized for b-actin and on which amplification wassubsequently performed, using primers specific forthe PAF-R. Subsequent amplification of cDNA appro-priately diluted with primers specific for the PAF-Rsimilarly produced a single band of the predicted size(397 bp). Figure 4B demonstrates the cyclic changes

PAF AND ENDOMETRIAL REMODELING 837
/ 382c 0012 Mp 837 Monday Jul 13 11:49 AM LP–FASEB 0012
Figure 3. In situ localization of PAF-R mRNA expression in secretory endometrium. Bright-field (A, C, E) and dark-field (B, D,F) photomicrographs of sections of endometrium after in situ hybridization with 35S-labeled sense (A, B) and antisense (C–F)PAF-R RNA probe. Early secretory phase (A–D) and late secretory phase endometrium (E, F). Endometrial glands (G). Bloodvessels (Bv). Bar Å 50 mm.
in PAF-R mRNA expression in endometrial samplesstandardized for b-actin, showing transcript levels in-creasing from the early to mid-proliferative, persist-ing in late proliferative endometrium, only to de-crease in early secretory and then to increaseprogressively in the mid- and late secretory endo-metrium.
Quantitation of PAF-R mRNA throughout themenstrual cycle
Finally, levels of PAF-R mRNA were determinedthroughout the normal menstrual cycle using RNAseprotection assay. Autoradiographic signals were ap-parent in all phases of the cycle; however, variationin signal intensity was observed between differentphases (Fig. 5A). Quantitation of autoradiographicsignals by densitometric analysis showed relatively lowabundance of PAF-R mRNA throughout the cycle in
EP, MP, and LP phases of the cycle, as shown in Fig.5B. The expression of PAF-R remained low in the ESand MS phases, giving densitometric PAF-R/b-actinratios similar to those found for proliferative phaseendometrium. In the LS phase, the abundance ofPAF-R mRNA increased dramatically by sixfold rela-tive to all other cycle phases.
Effect of PAF on NO release
Addtion of PAF to endometrial explants resulted inthe rapid generation of NO consistent with activationof a constitutive isoform of NOS, with values exceed-ing basal release by 47.7{9.7% within 2 h of stimu-lation. No demonstrable difference was observed inthe basal values of NO released by proliferative andsecretory endometrium. The concentration of PAFrequired to evoke maximal NO release, however, ap-pears to be governed by the phase of the menstrual
838 Vol. 12 July 1998 The FASEB Journal AHMED ET AL.
/ 382c 0012 Mp 838 Monday Jul 13 11:49 AM LP–FASEB 0012
Figure 4. Semiquantitative RT-PCR for the PAF-R throughoutthe menstrual cycle. Samples of total RNA from early, mid-,late proliferative (EP, MP, LP) and early, mid-, late secretory(ES, MS, LS) phase endometrium were subjected to RT-PCRusing primers specific for b-actin and the PAF-R. A) PCR am-plification of appropriately diluted RT products (EP: 1 in 500,MP: neat, LP: neat, ES: 1 in 5000, MS: 1 in 5000, LS: 1 in 2000)standardized for b-actin (cycles 30, 33, 36, 39, 42, 45). A singleband of 460 bp can be seen. B) Subsequent PCR amplificationon b-actin matched samples for the PAF-R (cycles 32, 36, 40,44, 48, 52). A single band of 397 bp can be seen.
Figure 5. Quantitation of PAF-R mRNA in normal cyclic en-dometrium. A) Identification of mRNA for PAF-R by RNAseprotection assay using total RNA isolated from human endo-metrium at various stages of the menstrual cycle. The abun-dance of mRNA for actin is also shown for each sample forcomparison of RNA amount loaded onto each lane. Lanes 1and 2 show control incubation of PAF-R and actin riboprobeswith 20 mg yeast tRNA without or with RNAse treatment, re-spectively. Lanes 3–8 represent samples of 20 mg total endo-metrial RNA throughout the menstrual cycle. Lane 3, EP; lane4, MP; lane 5, LP; lane 6, ES; lane 7, MS; and lane 8, LS. B)Graphical representation of PAF-R:actin mRNA throughoutthe menstrual cycle from the above experiment. Autoradio-graphic signals for PAF-R and actin are expressed as relativedensitometric units. Data represent three sets of experiments.
cycle. Maximal NO release from proliferative (nÅ6)endometrial explants was observed (Fig. 6A) after theaddition of 1 mM PAF, whereas maximal PAF-medi-ated NO release from secretory (nÅ6) endometrialexplants required only 0.1 nM of PAF (Fig. 6B).
At a range of PAF concentration (1 mM–1 pM)there was significant inhibition in total NO release inthe presence of WEB2170 (10 mM), indicating thatthe response is mediated by the PAF-R. PAF-mediatedNO release was attenuated by L-NMMA in a dose-dependent fashion, indicating NOS activation, withhalf-maximal inhibition (IC50) at approximately 5 mM(Fig. 7). To identify the regulatory mechanisms in-volved in PAF-stimulated NO release, the roles of ex-ternal calcium and protein kinase C (PKC) were de-termined by conducting experiments in the presenceof both low calcium buffer and Ro 31–8220, a PKCinhibitor. Stimulation of endometrial explants witheither 0.1 nM or 1 mM PAF in the presence of exter-nal calcium evoked a significant increase in total NO,
with values typically 61.1{16.0% above basal (nÅ4,Põ0.0001). When stimulations were conducted inthe presence of low calcium buffer (150 nM), how-ever, PAF-mediated NO release was totally abolished,suggesting total dependency on extracellular calcium(nÅ4, Põ0.0001; Fig. 8). Moreover, the response wasattenuated by pretreatment with, and stimulation inthe presence of, 1 and 5 mM Ro 31–8220, inhibitingPAF-mediated (0.1 nM or 1 mM) NO release by54.5{2.3% (nÅ3, Põ0.001) and 82.5{8.0% (nÅ3,Põ0.0001), respectively (Fig. 8).
Effect of PAF on VEGF expression
To further define the role of PAF, we examined theeffect of this phospholipid on VEGF mRNA and pro-tein in the endometrial glandual epithelial cell line.After stimulation with PAF (1 mM PAF) or vehicle, thereactions were terminated at the appropriate time (1,

PAF AND ENDOMETRIAL REMODELING 839
/ 382c 0012 Mp 839 Monday Jul 13 11:49 AM LP–FASEB 0012
Figure 6. Dose dependency of PAF-mediated NO release fromexplants of proliferative (A) and secretory (B) phase endo-metrium. After equilibration, explants were stimulated for 2h with PAF (0.1 nM–1 mM). Media was removed and NO levelswere determined using a Sievers NOA 270B chemilumines-cent analyzer. Results are expressed as mean{SEM of a typicalexperiment (duplicate determinations per experiment; sixsimilar experiments).
Figure 7. Effect of inhibition of NO synthesis by L-NMMA onPAF-mediated NO release. Explants of human endometriumwere stimulated for 2 h with either 0.1 nM PAF (secretoryphase endometrium) or 1 mM PAF (proliferative phase en-dometrium) in the presence of increasing concentrations ofL-NMMA. Reactions were terminated by aspiration of theconditioned media and NO levels were determined using aSievers NOA 270B chemiluminescent analyzer. The results areexpressed as means{SEM of a typical experiment performedon secretory phase endometrium (duplicate determinationsper experiment; six similar experiments). Basal NO releasewas 19,687.5 pmol/well and PAF-mediated NO release25,983.1 pmol/well.
Figure 8. Effect of low external calcium and inhibition of pro-tein kinase C on platelet-activating factor (PAF) mediated NOrelease from explants of human endometrium. After equili-bration, explants were pretreated at 377C with low calciumbuffer or Ro 31–8220 (1 and 5 mM) for 1 h. Explants werethen stimulated with either 0.1 nM PAF (secretory phase en-dometrium) or 1 mM PAF (proliferative phase endometrium)in the presence of 150 nM external calcium, 1 mM Ro 31–8220, 5 mM Ro 31–8220, or vehicle alone (control). The re-sults are expressed as means { SEM of a typical experimentperformed on secretory phase endometrium (duplicate de-terminations per experiment; four similar experiments). Sta-tistical analysis was performed using Student’s unpaired t test.*P õ 0.01; **P õ 0.001; ***P õ 0.0001, when compared withPAF-stimulated NO release.
2, 4, 8, 12, 24, or 48 h) by removal of the conditionedmedium from confluent cell monolayers, and totalRNA or protein was extracted. Stimulation with PAFresulted in at least a twofold increase in VEGF mRNAat 24 h compared to control (Fig. 9). Prior to 24 h,mRNA levels were consistently lower in all samples;however, at 48 h VEGF mRNA was increased in bothcontrol and PAF-stimulated cells. PAF similarly evokeda dose-dependent up-regulation of VEGF protein incell lysates after 24 h. The rabbit polyclonal anti-VEGFantibody raised against a 20 amino acid synthetic pep-tide, corresponding to the first 20 residues at theamino terminus of human VEGF, recognized one ma-jor band at 46 kDa (Fig. 10). This effect of PAF onVEGF protein expression was inhibited, in a dose-de-pendent manner, by the specific PAF-R antagonist
WEB 2170 (0.1 mM–10 mM; data not shown). No VEGFwas detected in the conditioned media of either con-trol or PAF-stimulated samples by Western blotting.

840 Vol. 12 July 1998 The FASEB Journal AHMED ET AL.
/ 382c 0012 Mp 840 Monday Jul 13 11:49 AM LP–FASEB 0012
Figure 9. Effect of PAF on VEGF mRNA expression in humanendometrial epithelial cells. Cells were stimulated for varyingtimes in the presence and absence of 1 mM PAF. Identificationof VEGF mRNA was by RNAse protection assay. The abun-dance of mRNA for b-actin is also shown for each sample forcomparison of VEGF mRNA abundance between RNA sam-ples. Lanes C1 and C2 show control incubation of VEGF andactin probes in the absence and presence of RNAse treatment,respectively. Lanes 1 to 14 represent control and PAF-stimu-lated cells, respectively, at 1 h (1/2), 2 h (3/4), 4 h (5/6), 8h (7/8), 12 h (9/10), 24 h (11/12), and 48 h (13/14). Tran-scripts detected were assigned relative densitometric units andthe ratio of VEGF/actin RNA was calculated for each sample.
Figure 10. Effect of PAF on VEGF protein expression in humanendometrial epithelial cells. After serum starvation, confluentmonolayers were stimulated with PAF (0.1 nM–1 mM) for 24 h.Protein was then extracted from cells using high-salt buffer priorto protein estimation; Western blot analysis was performed todetect VEGF in endometrial epithelial cell lysates.
Figure 11. Agonist-evoked protein tyrosine phosphorylation inhuman endometrial epithelial cells. Cells were stimulated ina time-dependent fashion with 1 mM PAF for 1, 5, 15, and 30min (A), with 1 ng/ml VEGF for 1, 5, 15, and 30 min (B), orvehicle. Cell lysates were assayed for phosphotyrosine contentby SDS-polyacrylamide gel electrophoresis. The identity of the125 kDa band was confirmed by reprobing the anti-phospho-tyrosine blot with anti-focal adhesion kinase. This is a typicalrepresentation of three separate experiments.
Effect of PAF and VEGF on protein tyrosinephosphorylation
In unstimulated human endometrial epithelial cells,a number of tyrosine phosphorylated proteins wereobserved (Fig. 11). PAF (1 mM) stimulated a time-dependent increase in the tyrosine phosphorylationof two major proteins of approximately 125 and 42–44 kDa (Fig. 11A). The phosphorylation state of pp125
was similarly increased by VEGF (1 ng/ml), but theresponse was more rapid than that observed withPAF; a significant increase in the phosphorylationstate of this protein was apparent within 1 min ratherthan after 5 min of stimulation (Fig 11B). Reprobingof the anti-phosphotyrosine blot with anti-focal ad-hesion kinase anti-FLKpp125 antibody confirmed theidentity of this band (data not shown).
DISCUSSION
The major findings of this study are the cellular lo-calization of mRNA encoding PAF-R and its trans-

PAF AND ENDOMETRIAL REMODELING 841
/ 382c 0012 Mp 841 Monday Jul 13 11:49 AM LP–FASEB 0012
lated protein in human endometrium, and the cyclicchanges in PAF-R mRNA levels throughout the men-strual cycle. Moreover, it shows that PAF may mediateendometrial function by NO and VEGF release andthat the ability of endometrium to respond to physi-ological concentrations of PAF varies markedly dur-ing the menstrual cycle. This does not correlate withthe cellular localization of PAF-R mRNA or its trans-lated product, supporting our original propositionthat ovarian steroids prime the PAF-R for PAF signalin human uteri (13). Indeed, recent studies by Satoand colleagues (22) showing the up-regulation by es-tradiol of PAF-evoked intracellular Ca2/ in culturedendometrial cells confirm our earlier findings.
Cyclic changes in the expression of PAF-R mRNAdemonstrated when using semiquantitative RT-PCRreflect similar changes observed both in the patternof immunostaining and the intensity of hybridizationsignal between different phases of the menstrual cy-cle. This demonstrates that expression of the PAF-Ris modulated by sex steroids in vivo. The discrepancybetween no hybridization signal in contrast to PAF-R-like immunoreactivity in the early proliferative en-dometrium most likely reflects increased turnover ofsteady-state PAF-R mRNA in this phase of the cycle.An increase in PAF-R mRNA expression from theearly proliferative to the late proliferative endome-trium suggests that the receptor mRNA may be up-regulated by estrogen. Indeed, positive transcrip-tional regulation of human PAF-R transcript 2 (tissuetype) by estrogen has been demonstrated in JR-stcells, a human stomach cancer cell line that expressesboth functional endogenous PAF-R transcripts (23),and, more recently, in endometrial cells in culture(22). Furthermore, PAF-R mRNA expression in thelate proliferative endometrium was confined to in-dividual endometrial glands, suggesting that the PAF-R gene is likely to be transcriptionally activated inthese cells.
PAF production has been shown to be regulated byovarian steroids in endometrium (3). Moreover, theobservation that PAF-R mRNA expression is high inthe secretory phase of the cycle suggests that proges-terone does not antagonize PAF-R mRNA expressionstimulated by estrogen, but most likely enhances pref-erential expression in the glands and endothelialcells of the blood vessels. PAF is known to stimulatesynthesis of PGE2 by enriched glandular fractions ofhuman secretory endometrium (4). Functional PAF-R are confined to the glandular epithelium, as dem-onstrated by us (for review, see ref 2) and confirmedby Sato and co-workers (22). A similar pattern of sexsteroid regulation of expression was reported forVEGF (24, 25) and bradykinin (26) in endometrium.Moreover, PAF-evoked phospholipase D activity (13)and intracellular Ca2/ (22) was up-regulated by 17b-estradiol. These results strongly suggest that PAF isinvolved in the physiological process of reproduction
and in the preparation of a receptive endometriumfor implantation.
PAF is a potent vasodilator, a response dependenton intact endothelium-derived NO synthesis (27, 28).PAF was reported to stimulate NO release in a humanendometrial epithelial cell line, HEC-1B, suggestingthat NO might augment the vasodilatory effects ofPAF (2). The current study demonstrates not only theproduction of NO in human endometrium, but alsoshows that the vasodilatory action of PAF is via NOrelease. Implantation is characterized by vasodilation(1). PAF-evoked NO generation in human endome-trial explants was rapid and calcium dependent, con-sistent with the activation of a constitutive NOS (14).PAF-evoked NO was blocked by PAF-R antagonistWEB2170, indicating a receptor-mediated response.The maximal endometrial NO release was governednot only by the concentration of PAF, but also by thephase of the menstrual cycle. At 1.0 mM, PAF evokedmaximal NO release in the proliferative endome-trium, whereas only 0.1 nM PAF was required to gen-erate maximal NO release in the secretory endome-trium. The different responses exhibited by theproliferative and secretory endometrium may reflectcyclic variation in PAF-R density and/or the activityof PAF acetylhydrolase, the enzyme responsible fordegradation of PAF during the menstrual cycle. It isequally possible, however, that estrogen primes theuterus to become more responsive to PAF. Estradiol-17b up-regulates PAF-evoked phospholipase D activ-ity (13) and PAF-induced intracellular Ca2/ release(22). The observation that PAF-mediated NO releaseis attenuated by Ro 31–8220 suggests that PKC maybe an important point of modulation in the releaseof NO from endometrial explants. Ro 31–8220 inhib-its members of the conventional PKC family (PKCisoenzymes PKCs-a, -b, and -g) that are Ca2/ andphospholipid dependent (29). Use of Ro 31–8220suggests a role for a member of the conventional PKCfamily in regulating PAF-mediated NO release. In-deed, PAF stimulates NO release from HEC-1B cellsvia PKCa (30).
Increased formation of NO could lead to increasedendometrial bleeding (31). PAF-R is localized aroundthe endometrial blood vessels during the secretoryphase of the menstrual cycle. Since PAF-R expressionincreases dramatically duning the late secretory en-dometrium and PAF stimulates NO release, it is pos-sible that one physiological function of PAF may beto promote menstruation. After menstruation, repairof the endometrium involves profound angiogenesis.During the secretory phase of the menstrual cycle,the spiral artery development occurs under the influ-ence of progesterone, a process during which spiralarterioles develop from straight arteries in the myo-metrium and basal endometrium into the thickenedsuperficial zone of the secretory endometrium (32).Thus, the increase in PAF-R gene expression and pro-

842 Vol. 12 July 1998 The FASEB Journal AHMED ET AL.
/ 382c 0012 Mp 842 Monday Jul 13 11:49 AM LP–FASEB 0012
tein observed in the late proliferative and late secre-tory endometrium suggests that PAF may modulatecell growth and neovascularization. Recent studieshave confirmed that PAFs possess angiogenic activity(33). PAF stimulates directional migration of endo-thelial cells in vitro, consistent with the morphologicalterations and redistribution of cytoskeleton seen inprevious studies (34, 35), by acting on specific PAF-Rs without affecting cell proliferation (33). Further-more, PAF has been implicated as a mediator of theangiogenesis induced by tumor necrosis factor (36)and to cooperate with other angiogenic moleculesand chemokines, including VEGF, acidic and basicFGF, and hepatocyte growth factor (HGF), in induc-ing vascular development in Kaposi’s sarcoma (37).The fact that PAF can stimulate VEGF mRNA andprotein in endometrium supports our notion thatPAF may modulate endometrial remodeling aftermenses, probably by up-regulating VEGF expression.Increases in both VEGF gene expression (38) andprotein production (25) have been demonstrated inthe late proliferative and late secretory phases of themenstrual cycle.
PAF and VEGF both stimulate the tyrosine phos-phorylation of a number of proteins, including focaladhesion kinase, which raises the possibility that PAFdirectly or via VEGF may regulate endometrial cellmotility and adhesion by activating this pathway. PAFantagonists have previously been shown to inhibit tro-phoblast outgrowth in vitro (5), suggesting that PAFmay be involved not only in the activation of blasto-cyst attachment, but also in the differentiation of thetrophectoderm into invasive trophoblast. Together,these results suggests that functional PAF receptorsmay play a role in the regulation of growth and divi-sion in endometrial cell by coupling to a tyrosine ki-nase pathway. In summary, the cyclic changes in PAF-R-like immunoreactivity and PAF-R mRNAexpression in human endometrium, together withthe observed stimulatory effects of PAF on NO re-lease, VEGF mRNA, and protein expression, suggestroles for PAF in the control of the uterine vascularbed as well as in regeneration of the endometriumin preparation for implantation after endometrialshedding.
This work was funded by Medical Research Council grant#G94–15051.
REFERENCES
1. McRae, A. C., and Heap, R. B. (1988) Uterine vascular perme-ability, blood flow and extra cellular fluid space during implan-tation in rats. J. Reprod. Fertil. 82, 617–625
2. Ahmed, A., and Dearn, S. (1997) The role of platelet-activatingfactor and its receptor in endometrial receptivity. Adv. Exp. Med.Biol. 416, 277–290
3. Alecozay, A. A., Harper, M. J. K., Schenken, R. S., and Hanahan,D. J. (1991) Paracrine interaction between platelet-activating fac-
tor and prostaglandins in hormonally-treated human lutealphase endometrium in vitro. J. Reprod. Fertil. 91, 301–312
4. Smith, S. K., and Kelly, R. W. (1988) Effect of platelet-activatingfactor on the release of PGF2a and PGE2 by separated cells ofhuman endometrium. J. Reprod. Fertil. 82, 271–276
5. Spinks, N. R., Ryan, J. P., and O’Neill, C. (1990) Antagonists ofembryo-derived platelet-activating factor act by inhibiting theability of the mouse embryo to implant. J. Reprod. Fertil. 88, 241–248
6. Nakamura, M., Honda, Z., Izumi, T., Sakanaka, C., Mutoh, H.,Minami, M., Bito, H., Seyama, Y., Matsumoto, T., Noma, M., andShimizu, T. (1991) Molecular cloning and expression of platelet-activating factor receptor from human leukocytes. J. Biol. Chem.266, 20400–20405
7. Seyfried, C. E., Schweickart, V. L., Godiska, V. L. R., and Gray,P. W. (1992) The human platelet-activating factor receptor gene(PTAFR) contains no introns and maps to chrosome 1. Genomics13, 832–834
8. Mutoh, H., Bito, H., Minami, M., Nakamura, M., Honda, A.,Izumi, T., Nakata, R., Kurachi, Y., Terano, A., and Shimizu, T.(1993) Two different promoters direct expression of two distinctforms of mRNA of human receptor from human leukocytes.FEBS Lett. 322, 129–134
9. Kudolo, G. B., Kasamo, M., and Harper, M. J. K. (1991) Auto-radiographic localisation of platelet-activating factor (PAF) bind-ing sites in the rabbit endometrium during the peri-implantationperiod. Cell Tissue Res. 265, 231–241
10. Ahmed, A., Sage, S. O., Plevin, R., Shoaibi, M. A., Sharkey,A. M., and Smith, S. K. (1994) Functional platelet-activating fac-tor receptors linked to inositol lipid hydrolysis, calcium mobili-sation and tyrosine kinase activity in the human endometrialHEC-1B cell line. J. Reprod. Fertil. 101, 459–466
11. Ahmed, A., and Smith, S. K. (1992) Platelet activating factor stim-ulates phospholipase C activity in human endometrium. J. Cell.Physiol. 152, 207–214
12. Ahmed, A., Ferriani, R. A., Plevin, R., and Smith, S. K. (1992)Platelet-activating factor mediates phosphatidylcholine hydroly-sis by phospholipase D in human endometrium. Biol. Reprod. 47,59–65
13. Ahmed, A., Plevin, R., and Smith, S. K. (1995) Oestradiol-17bmodulates PAF-evoked phospholipase D activity but not inositollipid hydrolysis in human endometrial cell line, HEC-1B. Cell.Signal. 7, 1–7
14. Lowenstein, C. J., Dinerman, J. L., and Snyder, S. H. (1994) Ni-tric oxide: a physiologic messenger. Ann. Int. Med.. 120, 227–233
15. Fox, H., and Buckley, C. H. (1983) Endometrium. In Atlas ofGynaecological Pathology (Gresham, G. A., ed) pp. 61–63, MTPPress Ltd., London
16. Ahmed, A., Li, X. F., Shams, M., Gregory, J., Rollason, T., Barnes,N. M., and Newton, J. R. (1995) Localization of the angiotensinII and its receptor subtype expression in human endometriumand identification of a novel high-affinity angiotensin II bindingsite. J. Clin. Invest. 96, 848–857
17. Muller, E., Dagenais, P., Alami, N., and Rola-Pleszczynski, M.(1993) Identification and functional characterization of platelet-activating factor receptors in human leukocyte populations us-ing polyclonal anti-peptide antibody. Proc. Natl. Acad. Sci. USA90, 5818–5822
18. Shams, M., and Ahmed, A. (1994) Localisation of mRNA forbasic fibroblast growth factor in human placenta. Growth Factors11, 105–111
19. Sangha, R. K., Li, X.-F., Shams, M., and Ahmed, A. (1997) FGFReceptor-1 (flg) is a critical component for endometrial remod-elling: Localisation and expression of FGF-2 and FGF-R1 in hu-man endometrium during the menstrual cycle and decreasedFGF-R1 expression in menorrhagia. Lab. Invest. 77, 389–402
20. Heuer, H. O., Casals-Stenzel, J., Muacevic, G., and Weber, K. H.(1990. Pharmacologic activity of bepafant (WEB 2170), a newand selective hetrazepinoic antagonist of platelet-activating fac-tor. J. Pharmacol. Exp. Ther. 255, 962
21. Ahmed, A., Dunk, C., Kniss, D., and Wilkes, M. (1997) Role ofVEGF receptor (Flt-1) in mediating calcium-dependent nitric ox-ide release and limiting DNA synthesis in human trophoblastcells. Lab. Invest. 76, 779–791
22. Sato, S., Kume, K., Takan, T., Mutoh, H., Taketani, Y., and Shi-mizu, T. (1997) Up-regulation of the intracellulkar Ca2/ signal-ing and mRNA expression of platelet-activating factor receptor

PAF AND ENDOMETRIAL REMODELING 843
/ 382c 0012 Mp 843 Monday Jul 13 11:49 AM LP–FASEB 0012
by estradiol in human uterine endometrial cells. Adv. Exp. Med.Biol. 416, 95–100
23. Mutoh, H., Kume, K., Sato, S., Kato, S., and Shimizu, T. (1994)Positive and negative regulations of human platelet-activatingfactor receptor transcript 2 (tissue-type) by estrogen and TGF-beta 1. Biochem. Biophys. Res. Commun. 205, 1130–1136
24. Charnock-Jones, D. S., Sharkey, A., Rajput-Williams, L., Burch,D., Schofield, J. P., Fountain, S. A., Boocock, C. A., and Smith,S. K. (1993) Identification and localisation of alternately splicedmRNAs for vascular endothelial growth factor in human uterusand estrogen regulation in endometrial carcinoma cell lines.Biol. Reprod. 48, 1120–1128
25. Li, X. F., Gregory, J., and Ahmed, A. (1994) Immunolocalisationof vascular endothelial growth factor in human endometrium.Growth Factors 11, 277–282
26. Shams, M., Sisi, aP., and Ahmed, A. (1996) Localisation of bra-dykinin type II receptor mRMA in human endometrium. Mol.Human Reprod. 2, 35–39
27. Kamata, K., Mori, T., Shigenobu, K., and Kasuya, Y. (1989) En-dothelium-dependent vasodilator effects of platelet-activatingfactor on rat resistance vessels. Br. J. Pharmacol. 98, 360–364
28. Juncos, L. A., Ren, Y., Arima, S., and Ito, S. (1993) Vasodilatorand constrictor actions of platelet-activating factor in the isolatedmicroperfused afferent arteriole of the rabbit kidney. Role ofendothelium-derived relaxing factor/NO and cyclooxygenaseproducts. J. Clin. Invest. 91, 1374–1379
29. Wilkinson S. E., Parker, P. J., and Nixon, J. S. (1993) Isoenzymespecificity of bisindolylmaleimides, selective inhibitors of proteinkinase C. Biochem J. 294, 335–337
30. Dearn, S., and Ahmed, A. (1998) Activation of platelet-activatingfactor receptor stimulates nitric oxide release in human endo-metrial epithelial cell line (HEC-1B) via protein kinase C alpha.Lab. Invest. In press
31. Chaudhuri, G., and Nathan, L. (1992) Nitric oxide and endo-metrial blood flow. In Steroid Hormones and Uterine Bleeding (Al-
exander, N. J., and d’Arcangues, C., eds) pp. 241–244, AAASPublication, Washington, D.C.
32. Guyton, A. C. (1981) Textbook of Medical Physiology, pp. 1005–1020, W. B. Saunders, Philadelphia
33. Camussi, C., Montrucchio, G., Lupia, E., De Martino, A., Perona,L., Arese, M., Vercellone, A., Toniolo, A., and Bussolino, F.(1995) Platelet-activating factor directly stimulates in vitro mi-gration of endothelial cells and promotes in vivo angiogenesisby a heparin-dependent mechanism. J. Immunol. 154, 6492–501
34. Bussolino, F., Camussi, G., Aglietta, M., Braquet, M., Bosia, P.,Pescarmona, G., Sanavio, F., D’Urso, M., and Marchisio, M.(1987) Human endothelial cells are targets for platelet-activatingfactor. I. Platelet-activating factor induces changes in cytoskeletalstructure and promotes albumin diffusion across endothelialmonolayer. J. Immunol. 139, 2439–2446
35. Camussi, G., Turello, E., Bussolino, F., and Baglioni, C. (1991)Tumour necrosis factor alters cytoskeletal organization and bar-rier function of endothelial cells. Int. Arch. Allergy Appl. Immunol.96, 84
36. Montrucchio, G., Lupia, E., Battaglia, E., Passerini, G., Bussolino,F., Emanuelli, G., and Camussi, G. (1994) Tumour necrosis fac-tor alpha-induced angiogenesis depends on in situ platelet-acti-vating factor biosynthesis. J. Exp. Med. 180, 377–382
37. Bussolino, F., Arese, M., Montrucchio, G., Primo, L., Benelli, R.,Sanavio, F., Aglietta, M., Ghigo, D., Rola-Pleszczynski, M., Bosia,A., Albini, A., and Camussi, G. (1995) Platelet-activating factorproduces in vitro by Kaposi’s sarcoma cells induces and sustainsin vivo angiogenesis. J. Clin. Invest. 96, 940–952
38. Shweiki, D., Itan, A., Soffer, D., and Keshet, E. (1992) Vascularendothelial growth factor induced by hypoxia may mediate hy-poxias-initiated angiogenesis. Nature (London) 359, 843–845
Received for publication June 23, 1997.Revised for publication January 20, 1998.





























