Embed Size (px)
Citation preview

THE JOUKNAL OF COMPARATIVE NEUKOLOGY 367:264-273 (1 996)
Localization of Enkephalin Immunoreactivity in the Spinal Cord
of the Long-Tailed Ray Himantura fai
PETER J. SNOW, GlLLlAN M.C. RENSHAW, AND KATHERIYE E. HAMLIN Cerebral and Sensory Function Unit, Department of Anatomical Sciences,
University of Queensland, St Lucia, Queensland, Australia 4072
ABSTRACT Enkephalin-like immunoreactivity (ENK-LI) was found throughout the spinal cord of the
long-tailed ray Himantura f a i The densest ENK-LI was in the superficial portion of lamina A of the dorsal horn. Lamina B and the deeper parts of laminaA contained radially oriented, labelled fibres. Laminae C, D, and E contained many longitudinally orientated fascicles which were surrounded by a reticulum of transversely orientated, labelled fibres, some of which projected into the ventral and lateral funiculi. Labelled fibres were found in the dorsal commissure and around the central canal, but the later did not cross the midline. One-third of all enkephaliner- gic cells were found throughout laminae A and B, while two-thirds were located in the medial half of C, D, and E. Occasionally a labelled cell was located in the lateral funiculus. The ventral horn (laminae F and G) contained many enkephalinergic fibres but no labelled nuclei.
A few dorsal column axons contained ENK-LI. In the lateral funiculus there were two groups of labelled axons, a superficial, dorsolateral group, and a deeper group, occupying a crescent-shaped region. The ventral funiculus also contained many labelled axons. The central projection of the dorsal root passed through the substantia gelatinosa and divided into rostrally and caudally projecting fascicles within lamina C. The root, and these fascicles, both lacked ENK-LI. In contrast, the fascicles in laminae D and E did contain enkephalinergic fibres. The origin of the various fibre systems and the role of enkephalin in the regulation of sensory processing and motor output are discussed.
Indexing terms: fish, pain, opiates, evolution, immunocytochemistry
cj 1996 Wiley-Liss, Inc.
The endogenous opiates are generally regarded as inhibi- tory transmitters. In the spinal cord of mammals, enkepha- linergic terminals are found throughout the spinal grey matter, but are most concentrated in Rexed's laminae I and I1 of the superficial dorsal horn. In this region, enkephalin is found in synapses on the cells of origin of the lateral spinothalamic tract -a system which, in mammals, is highly responsive to a variety of noxious stimuli and is known to receive monosynaptic input from unmyelinated (C-fibre) and fine, myelinated (AG-fibre) nociceptive affer- ents (Dubner et al., 1984; Willis and Coggeshall, 1991). Laminae I and I1 in mammals are therefore regarded as important relay stations in the transmission of nociceptive information to ascending tract neurons. The Atlantic stin- gray lacks unmyelinated somatic afferents (Coggeshall et al., 1978) but in this animal, the field potential studies of Leonard et al. (1978) suggest that the smaller, A6, myelin- ated primary afferents project to the superficial regions of the dorsal horn, while the larger myelinated afferents project to deeper parts of the dorsal horn.
The cartilaginous fish, the chondrichthyes, include the elasmobranchs (sharks, skates, and rays) and the holocepha- lons (ratfish). They are considered to have evolved from the earliest jawed vertebrates, the gnathostomes slightly before the bony fish (osteichthyes). The gnathostomes themselves are considered to have evolved from an early group of jawless vertebrates, the agnathans, which today are repre- sented only by the lamprey and the hagfish (Northcutt, 1981). In many ways, we might therefore expect living representatives of these groups to bear the characteristics of their fossil ancestors. However, as Northcutt (1981) is careful to point out, the survival of these lifeforms for many hundreds of millions of years has most likely only been possible by their undergoing considerable adaptation to an ever changing environment.
Accepted October 26, 1995. Addrcss reprint requests to Associate Professor Peter J. Snow, Cerebral
and Sensory Functions Unit, Department ofhatomica1 Sciences, University of Quecnsland, St. Lucia, Australia 4072. E-mail:p.snowtflmailbox.uq.oz.au
G 1996 WILEY-LlSS, INC.

ENKEPHALIN IN ELASMOBRANCH SPINAL CORD
The endogenous opiate enkephalin is not found in the spinal cord of the lamprey (Buchanan et al., 1987). How- ever, its occurrence has been reported in the dorsal and ventral horn of the spinal cord of a teleost, Salmo gairdneri (Vecino et al., 1992) and in the dorsal horn of an elasmo- branch, Rhinobatus batillurn (Cameron et al., 1990). The presence of enkephalin in the dorsal horn of the elasmo- branch spinal cord is of special interest because elasmo- branchs lack the unmyelinated sensory axom which in mammals constitute a large percentage of the nociceptive afferent fibres (Coggeshall et al, 1978; Snow and Plender- leith, 1992; Snow et al., 1993). In addition these animals lack a cytoarchitectonic area equivalent to lamina I of the mammalian dorsal horn (Cameron et a]., 1990)-a region where nociceptive ascending tract neurons are located (Willis and Coggeshall, 1991). Thus, while Leonard (1985) has reported that a few primary afferents in the stingray do respond to high-intensity mechanical stimuli, it would seem that elasmobranchs have, at best, a very rudimentary system for encoding potentially damaging somatic stimuli (Snow et al., 1993). I t is, therefore, interesting to know more of the extent and detail of the enkephalinergic innervation of the spinal grey of elasmobranchs. The present study was undertaken to examine details of the morphology and distribution of enkephalin-like immunore- active (ENK-LI) profiles in the spinal cord of the long-tailed ray, Himantura fai.
265
MATERIALS AND METHODS Tissue was taken from four long-tailed rays collected
from designated collecting sites at Heron Island, Queens- land, Australia. Previously we have referred to these ani- mals as Dasyatis fluuiorum (Cameron et al., 1990) and Himaritura uranak (Kitchener, Wilson and Snow et al., 1993; Snow, 19941, but a recent taxonomic revision has reclassified them as being Himantura fai (Whittington and Last, 1994).
Prior to fixation, animals were anaesthetised with MS222 (60 mg/kg), and 1 ml of heparin and 1 ml of 1% sodium nitrite were administered intracardially. Animals were then perfused through the conus arteriosus at a pressure of 60 mm Hg with 1 litre of 0.1M phosphate buffered saline (PBS) containing 0.4M urea and 1% sodium nitrite then 1 litre of 4% paraformaldehyde in 0.1M phosphate buffer (pH 7.4) containing 0.4M urea, followed by a second litre of the same fixative perfused over 1 hour. The spinal cord directly posterior to the brachial chamber was removed, postfixed in 4% paraformaldehyde overnight and stored in 1% parafor- maldehyde.
Selected blocks were placed in 30% sucrose overnight and sectioned on the freezing microtome in the transverse (25, 50, and 100 pm), parasagittal (50 pm), and horizontal (50 pm) planes. Free-floating sections were rinsed for 5 min- utes in each of 3 changes of 0.1M PBS with 0.5% Triton-X. To block endogenous peroxidases sections were placed for 15 minutes in each of 2 changes of 50% alcohol followed by 30 minutes in 50% ethanol with 1% hydrogen peroxide. Sections were again rinsed for 5 minutes in each of 3 changes of O.1M PBS with 0.5% Triton-X and then trans- ferred for 60 minutes to 3% bovine serum albumin (BSA) in 0.1M PBS with 0.5% Triton-X to reduce nonspecific bind- ing. Sections were incubated in rabbit antimethionine- enkephalin (ENK, Incstar lot number 105753 at 1: 16,000)
\ Fig. 1. Illustration of the lamination of the spinal grey matter and
the distribution of 92 enkephalinergic cells in the spinal cord of the long-tailed ray, Hirnunturu fai. Cell distribution was plottcd from labelled cells located on the left side of ten, 50-pm serial sections through a 500-pm length of postbrachial spinal cord. Scale bar = 1 mm.
in 0.1M PBS with 1% BSA and 0.5% Triton-X, for 16-36 hours at 4°C. Sections were then rinsed for 5 minutes in each of 3 changes of O.1M PBS with 0.5% Triton-X, incubated with biotinylated donkey antirabbit (Jacksons, lot number 23159 at 1:600) in O.1M PBS with 1% BSA and 0.5% Triton-X, for 60 minutes and rinsed again in 3 washes of PBS prior to incubation in the avidin-biotin-HRP com- plex (Vector Elite B C kit) for 60 minutes. Sections were then washed again in PBS, and preincubated for 5 minutes in diaminobenzidine prior to the addition of hydrogen peroxide and further incubation for 10 minutes. Sections were mounted on subbed slides, cleared, and dehydrated; and the distribution of diaminobenzidine reaction product was examined and photographed on the light microscope. The position of labelled spinal cells in 25- and 50-pm trans- verse sections was plotted using a camera lucida (Fig. 1).
To test for specificity of labelling some sections were processed using antimethionine-enkephalin (1:16,000) which had been preabsorbed by exposure to 100 mg/ml methionine enkephalin (Auspep, Batch No. D542111) for 24 hours, whereas other sections were taken through the above process omitting only the exposure to the primary antisera. No labelling was found in either of these two sets of sections.
RESULTS General observations
In describing the distribution of enkephalin-like imrnuno- reactivity (ENK-LI), we have used the nomenclature and cytoarchitectonic divisions of the spinal grey described by Cameron et al. (1990). These cytoarchitectural divisions are illustrated in Figure 1. Within the elasmobranch cord three components could be easily recognised: a) grey matter of the dorsal (laminae A and B) and ventral horn (laminae F and G), w-hich was devoid of fascicles and tracts of axons; b) a series of longitudinally oriented fascicles which composed much of the cross-sectional area of laminae C, D, and E; and c) the white matter of the dorsal, lateral and ventral funiculi (Fig. 1).
Enkephalin-like immunoreactivity was found in cells, axons, and preterminal arborizations (Figs. 1 and 2). In

Fig. 2. Enkephalin-like IENK-LI) irnmunoreactivity in a transverse section of the spinal cord of the long-tailed ray. The highest concentra- tion of ENK-LI was in the most superficial part of the dorsal horn (superficial part of lamina A). Note that the longitudinal fascicles of lamina C (small solid arrows) lack ENK-LI. Note the ENK-LI in the ventral horn, in the longitudinal fascicles of laminae D and E, in the
fibres forming a reticulum around longitudinal fascicles of laminae C-E, and in the dorsal commissure (medium open arrows). In the white matter ENK-LI was found in a group of axons near the pial surface in the dorsolateral part of the lateral funiculus (large open arrow), in axons in located within a cresent-shaped region deeper in the lateral funiculus (medium solid arrows), and in the ventral funiculus. Scalebar = 200 km.

ENKEPHALIN IN ELASMOBRANCH SPINAL CORD 267
axons and terminals, we observed two types of ENK-LI: a) In punctate ENK-LI, the reaction product was located in dense arrays of particles (Figs. 3a,D. When only a few process were within a field of view, these particles could be seen to form a line, indicating that they were within a main axon or preterminal process (Fig. 3a). b) In soZid ENK-LI the axoplasm or cytoplasm appeared to be completely filled with reaction product, giving a Golgi-like appearance to the stained process (Figs. 3c, 5b). This form of labelling was also seen in cells which contained ENK-LI (Figs. 3f, Fja,d). Within the grey matter punctate ENK-LI was by far the most common form, but in the white matter many longitu- dinally oriented axons contained solid ENK-LI.
Labelled axons and terminals were found in the grey matter of all laminae, whereas labelled cells were found only in the dorsal, and intermediate regions (laminae A-E) (Figs. 1, 2). The longitudinal fascicles in lamina C were virtually devoid of ENK-LI, whereas those of laminae D and E were weakly labelled. In the white matter longitudinally oriented, labelled axons were found in the lateral, ventral, and even the dorsal funiculus, as well as in transversely oriented fibres in the lateral funiculus and ventrolateral parts of the ventral funiculus (Fig. 2).
Enkephalin-like immunoreactivity in axons and terminals
Lateral, ventral, and dorsal funiculi. Axons and termi- nals containing ENK-LI were found running in the longitu- dinal and transverse axis of the spinal cord. Longitudinally oriented, labelled axons were found throughout the lateral and ventral funiculi (Fig. 2). A few labelled axons were also found in the dorsal funiculus (Fig. 3e). In the lateral and ventral funiculi there were regions where there was a high concentration of such fibres. Thus in the lateral funiculus, fibres containing punctate ENK-LI were found concen- trated superficially in the dorsolateral part while in a more deeply located, cresent-shaped area which arcs around the lateral border of the grey matter were many fibres which contained solid ENK-LI (Fig. 2). In the ventral funiculus labelled fibres were most concentrated in the dorsal portion and usually contained solid ENK-LI (Fig. 2). Transversely oriented labelled axons were seen penetrating well into the lateral funiculi and ventrolateral part of the ventral fu- niculi (Fig. 2). Generally these axons contained punctate ENK-LI, but some clearly contained solid labelling, and many of the latter had a clearly beaded appearance (Fig. 3c).
The dorsal roots projected into the dorsal horn along the mediolateral axis of the cord passing through the substantia gelatinosa (laminae A and B) (Figs. 4a,b) and giving rise to rostrally and caudally projecting fascicles in lamina C (Fig. 4c). The central projections of the roots were clearly devoid of any ENK-LI (Figs. 4, 5). Within the dorsal horn the projection of the root was often divided into a rostral and caudal portion by dorsoventrally oriented fibres which contained ENK-LI (Fig. 4b).
The grey matter in the ventral portions of the dorsal horn (lamina C) and the intermediate zone, opposite the central canal (laminae D and E) con- tained many longitudinal fascicles (Figs. 1, 2) (Cameron et al., 1990). Fibres running along the fascicles of lamina C were devoid of ENK-LI (Fig. 21, presumably because in horizontal section these fascicles could be seen to be composed of the rostral and caudal projections of single dorsal roots which themselves lack ENK-LI (Fig. 412). Close examination of parasagittal sections showed that the fas-
Dorsal roots.
Ihgitudinal fascicles.
cicles of laminae D and E did contain a small number of fibres with punctate ENK-LI, indicating that these fascicles contained fibres which did not arise from the dorsal roots (Fig. 5a).
All the longitudinal fascicles were surrounded by trans- versely oriented labelled fibres most of which contained punctate ENK-LI (Figs. 2, 30 . Enkephalinergic fibres, therefore, participated in a network, or reticulum, around all the longitudinal fascicles. Occasionally, beaded fibres containing solid ENK-LI were observed crossing the longi- tudinal fascicles of laminae C, D, or E, suggesting that some transversely oriented, enkephalinergic s o n s synapsed with axons in the longitudinally oriented fascicles.
In transverse section, the lat- eral lobe of the substantia gelatinosa often appeared to be divided by the central projection of a dorsal root (Fig. 3b). Medially, much of the substantia gelatinosa curved ven- trally to terminate immediately above the dorsal commis- sure so that much of the medial lobe lay in the parasagittal plane (Fig. 2). Labelled axons within the substantia gelati- nosa mostly contained punctate ENK-LI.
The highest level of ENK-LI in the cord was found in the superficial part of lamina A, where even in 50-pm sections it was impossible to distinguish individual axons which contained punctate ENK-LI (Fig. 3b). Within lamina B and the deeper parts of lamina A, labelling was found in radially oriented fibres (Fig. 3b). In both laminae there were some transversely oriented, beaded axons containing solid ENK- LI, indicating that enkephalinergic axons made en passant synapses across the substantia gelatinosa (Fig. 3d).
Most of the transversely oriented fibres that form a reticulum around the longitudinally projecting fascicles of laminae C, D, and E contain punctate EIVK-LI (Fig. 30, while a few were beaded in appearance and contained solid ENK-LI. Later- ally, both solidly labelled and punctately labelled fibres projected well into the white matter, even, on occasion approaching the surface of the lateral funiculus (Figs. 2, 34. A few fibres of both types were also found within the ventral funiculus. Occasionally, beaded, solidly labelled fibres were also seen crossing the longitudinal fascicles. Trans- versely oriented, beaded, labelled fibres within the longitudinal fascicles and the lateral (Fig. 3c) and ventral funidi , indicated the presence of en passant enkephalinergic synapses within these major longitudinal fibre systems of the ray cord.
The dorsal commis- sure lay directly below the ventromedial tip of the substan- tia gelatinosa (Cameron et al., 1990). The fibres in the dorsal commissure ran dorsolaterally along the boundary between lamina C and lamina D, and appeared to form a tract of communication between the dorsal horn of either side. Weak, punctate ENK-LI could be seen in the fibres of the dorsal commissure (Fig. 2), but no fibres containing solid ENK-LI were observed.
Ventral to the dorsal commissure were a number of large longitudinal fascicles. Ventral to these fascicles was the ventral commissure which surrounded the central canal (Fig. 2). The ventral commissure appeared to form a tract that between lamina F of the left and right ventral horns and contained some weak punctate ENK-LI. Some fibre tracts (the commissura accessoria, Nieuwenhuys, 1964) could also be seen projecting across the ventral funiculus between lamina G of the each ventral horn (Fig. 2).
Transversely oriented, beaded fibres containing solid ENK-LI could be seen running across the ventral comrnis-
Substantia gelatinosa.
Reticulated zone and the white matter.
Dorsal and ventral commissures.

268 P.J. SNOW ET AL.
Fig. 3. a: Axons containing punctate ENK-LI (arrows) in a trans- verse section through lamina C of the dorsal horn. b: Radially oriented fibres containing ENK-LI coursing through lamina B. c: Beaded axon containing solid ENK-LI in a transverse section of the lateral funiculus (arrow). d Solid ENK-LI in a beaded axon terminal (arrow) in lamina A. e: Axon containing ENK-LI (arrow) in a longitudinal section of the
dorsal column and underlying, densely immunoreactive, laminae A (asterisk). f Cell containing ENK-LI (arrow) amongst the reticular fibres of lamina D. Note the alignment of the dendrites of this cell with immunoreactive reticular fibres. Scale bars: a = 50 pm; b = 500 pm; c = 50 pm; d = 50 km; e = 100 pm; f = 50 pm.
sure towards the central canal (Fig. 5b). These fibres were not, however, commissural for on reaching the central canal they either terminated below the canal, or more often curved to run dorsally in the midsagittal plane between the large medial longitudinal fascicles (Fig. 5b). Their beaded
appearance suggested that they formed en passant syn- apses along their course.
In the ventral horn (laminae F and G) ENK-LI, though less dense than in the dorsal horn, was conspicuous and uniform (Fig. 2). Fibres which contained
Ventral horn.
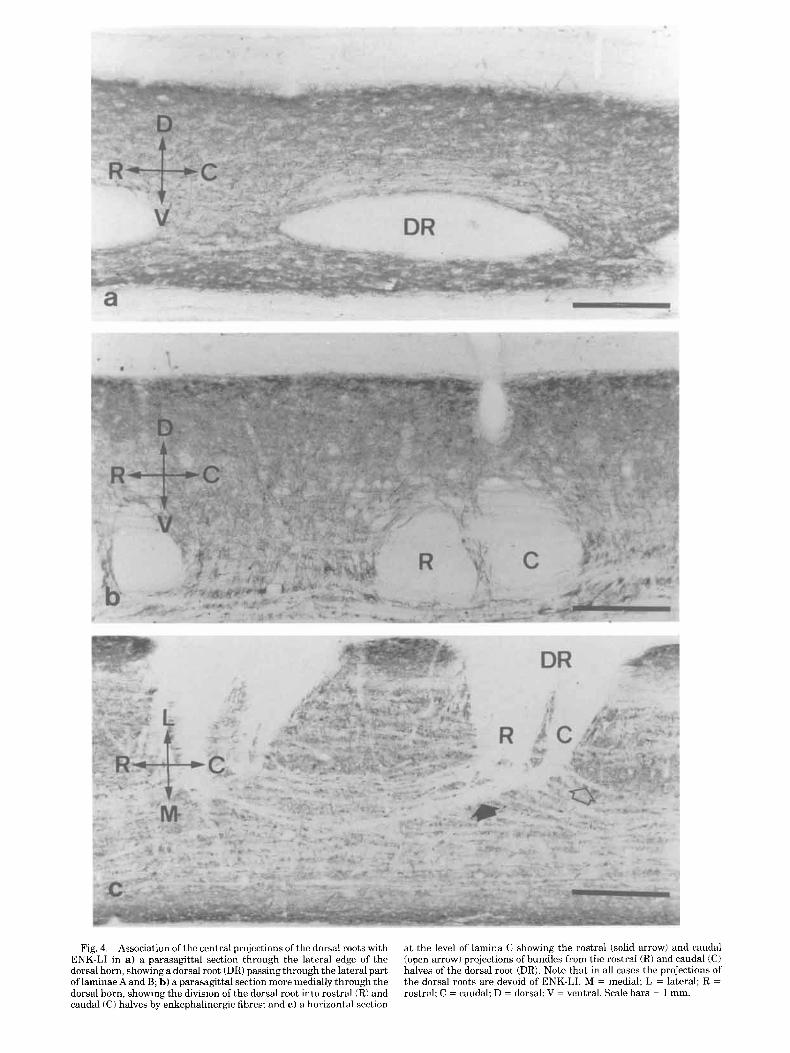
Fig. 4. Association of the central projections of the dorsal roots with ENK-LI in a) a parasagittal section through the lateral edge of the dorsal horn, showing a dorsal root (DR) passing through the lateral part of laminae A and B; b) a parasagittal section more medially through the dorsal horn, showing the division of the dorsal root into rostral (R) and caudal (C) halves by enkephalincrgic fibres; and c ) a horizont,al section
at the level of lamina C showing the rostral (solid arrow1 and caudal (open arrow) projections ofbundles from the rostral (Ri and caudal (CI halves of the dorsal root (DR). Note that in all cases the projections of the dorsal roots are devoid of ENK-LI. M = medial; L = lateral; R = rostral; C = caudal; D = dorsal; V = ventral. Scale bars = 1 mm.

270 P.J. SNOW ET AL.
Fig. 5. Enkephalin-like immunoreactivity a) in a parasagittal sec- tion medial part of the grey matter, b) in a transverse section through the central canal, c ) in axons in the ventral horn, and d) in axons and a cell in a parasagittal section through the lateral funiculus. Panel a shows the punctate ENK-LI in longitudinal fascicle of lamina D and a spindle-shaped enkephalinergic cell (arrow). Pancl b shows beaded, enkephalinergic axons projecting towards the central canal and turning
to run dorsally (arrows) in the midsagittal plane. Panel c shows solid (arrows) ENK-LI and punctate ENK-LI in axons in the ventral horn. Panel d shows ENK-LI in axons coursing along the lateral funiculus, in axons crossing the lateral funiculus in the transverse plane (arrows), and in a cell within the lateral funiculus (open arrow). Scale bars: a = 100 pm; b = 50 pm; c = 100 pm; d = 200 pm.

ENKEPHALIN IN ELASMOBRANCH SPINAL CORD 271
both punctate ENK-LI and solid ENK-LI were present (Fig. 5cj. The latter type of labelling was often in beaded fibres (Fig. 5c), suggesting the presence of en passant, enkephalin- ergic synapses throughout this region.
Enkephalin-like immunoreactivity in spinal cells
Cells containing ENK-LI were found only in the dorsal and intermediate regions of the spinal grey matter (laminae A-E) (Figs. 1, 3f, 5a) and very occasionally in the lateral funiculus (Fig. 5d). Figure 1 shows the distribution of 92 labelled cells located in the left side of a set of serial sections through a 500-pm length of postbrachial spinal cord. The other side of this length of cord contained 72 labelled cells, suggesting that there was on each side of the postbrachial cord about 164 enkephalinergic cells/mm.
In a total sample of 223 labelled cells 15.7% were located in lamina A, 18.4% in lamina B, 25.6%> in lamina C, 37.6% in lamina D, and 2.7% in lamina E. Thus. about one-third (34%) of labelled cells were located in the substantia gelatinosa (laminae A and B), whereas two-thirds (65%) were found within the reticulated zone (laminae C-E). No enkephalinergic cells were found in the ventral horn (lami- nae F and G) .
In laminae A and B labelled cells were bipolar and were found scattered across the entire width of the dorsal horn. When viewed in the transverse plain their proximal den- drites were radially oriented such that they were lined up with the enkephalinergic fibres found in this region of the spinal grey. In laminae C-E, labelled cells were multipolar and were generally located in the medial half of these laminae. In laminae C and E the dendrites of these cells often followed the course taken by the bundles of trans- versely oriented, enkephalinergic fibres that surrounded the longitudinal fascicles (Fig. 3f). In parasagittal sections, enkephalinergic cells in lamina E were often spindle shaped with their long axis oriented in the dorsoventral axis of the cord (Fig. 5a), whereas within the lateral funiculus spindle- shaped cells had their long axis in the rostrocaudal axis of the cord (Fig. 5d).
DISCUSSION Comparison with other species
Although Northcutt (1981) has emphasized that living representatives of the gnathosomes and the agnathans may represent specializations as much as they do ancestral forms, it is nevertheless interesting to note that there does seem to be some correlation between phylogenetic origin and the presence of the endogenous opioid, enkephalin. In the elasmobranchs we first reported the presence of ENK-LI in the spinal cord of the shovel-nose ray, Rhinobatus battilum (Cameron et al., 1990). The present work has extended this by revealing enkephalinergic axons and termi- nals throughout all parts of the cord of the long-tailed ray, Hirnantura fai, and enkephalinergic cells within the dorsal horn and intermediate region of the grey matter. In the osteichthye, Salmo gairdneri, ENK-LI has been reported in axons and terminals of the dorsal horn (Vecino et al., 1992), although no immunoreactive cells were found.
In other nonmammalian groups ENK-LI has been re- ported in both cells and axons in the spinal cord of the frog, Rana esculenta (Lozez and Kemli, 1981); the lizard, Anolis carolinensis (Naik et al., 1981); the turtles, Chrysemys picta and Pseudemys scripta (Reiner, 1987); and the domes-
tic fowl, Gallus domesticus (LaValley and Ho, 1983). How- ever, in the cord of the lizard, Varanus exanthematicus, ENK-LI was detected only in axons and terminals (Wolters et al., 1986). In contrast to the gnathosomes, no ENK-LI has been found in the spinal cord of the lamprey, Ichthyomy- zon unicuspis (Buchanan et al., 19871, suggesting that at least in relation to spinal circuitry, involvement of the endogenous opioids might be a characteristic which has evolved with the jawed vertebrates.
In the long-tailed ray, cells con- taining ENK-Ll were found in all laminae except the ventral horn (laminae F and G) (Fig. 1). A few labelled cells were also found in the lateral funiculus. Enkephalinergic cells have also been reported in the dorsal horn, the intermediate region, and around the central canal in turtles (Reiner, 1987), the lizard (Naik et al., 19811, and the frog (Lorez and Kemali, 1981) but not in the white matter of these species. In mammals, no enkephalinergic cells have been reported in the ventral horn, but such cells have been found throughout the dorsal horn and the intermediate region (Sar et al., 1978; Uhl et al., 1979; Glazer and Basbaum, 1981).
Within both the grey and the white matter, the dendritic morphology of labelled cells often seemed to be determined by the orientation of fibre systems present within the area of cord occupied by their somata. Thus the morphology of these neurons was related to their position within the spinal grey matter.
Enkephalinergic axons and terminals in the spinal grey mutter. In the dorsal horn of the long-tailed ray, ENK-LI was clearly most intense across the most superficial part of lamina A of the dorsal horn and strong labelling was also apparent in the deeper parts of lamina Aand in lamina B. A similarly superficial concentration of enkepahlinergic termi- nals has been reported in spinal cord of other nonmamma- lian species, including the shovel-nose ray (Cameron et al., 1990), the rainbow trout (Vecino et al., 1992), the turtle (Reiner, 1987), lizard (Naik et al, 1981; Wolters et al., 1986), the domestic chicken (LaValley and Ho, 19831, and the frog (Lorez and Kemali, 1981). In mammals, enkepha- linergic terminals are also concentrated in Rexed’s laminae I and I1 of the superficial dorsal horn, where they have been shown to synapse with the cells of origin of the lateral spinothalamic tract (Willis and Coggeshall, 1991).
From the cytoarchitecture of the elasmobranch cord, Cameron et al. (1990) concluded that the most superficial part of the dorsal horn (lamina A) was not structurally homologous to Rexed’s lamina I of the mammalian dorsal horn. Although, in the ray, both laminae A and B each have a distinctive cytoarchitecture (Cameron et al., 1990), in unstained sections they have together the clear appearance which in mammals characterizes the substantia gelatinosa as defined originally by Clarke (1859). Thus, the concentra- tion of ENK-LI in lamina A of the ray supports the cytoarchitectural division of the substantia gelatinosa of the ray into lamina A and lamina B.
In the long-tailed ray the terminals of enkephalinergic fibres coursed radially through lamina R and the deeper parts of lamina A. This form of organization has a parallel in the somatotopic organization of the mammalian dorsal horn, where spinal cells which lie along any line which describes the radius of the dorsal horn were activated by afferents from the same area of skin (Wilson et al., 1986; Woolf and Fitzgerald, 1986). If, in the ray, the skin was represented in laminae A and B, in the same manner as it
Enkephalinergic cells.

2 72
was in laminae I-V of the mammalian dorsal horn, then the radial orientation of enkephalinergic terminals would indi- cate that these fibres might provide selective inhibition of afferent information from particular parts along the proxi- modistal axis of each dermatome (Kitchener et al., 1994).
Laminae C-E of the ray have numerous longitudinal fascicles which were surrounded by a reticulum of fibres, many of which contained ENK-LI. A similar network of enkephalinergic fibres has been reported in the dorsal horn of the shovel-nose ray (Cameron et al., 1990). In elasmo- branchs, laminae E and D constitute what is generally known as the intermediate region of the spinal grey, a region which in turtles (Reiner, 19871, lizards (Naik et al, 1981; Wolters et al, 19861, the domestic fowl (LaValley and Ho, 19831, and the frog (Lorez and Kemali, 1981) has also been shown to contain ENK-LI. Similarly ENK-LI has been found in the region around the central canal of the long- tailed ray, the lizard, the frog, domestic fowl, and the rat (Lorez and Kemali, 1981; Gibson et al., 1981; Naik et al., 1981; LaValley and Ho, 1983; Wolters et al., 1986; Reiner, 1987; Lamotte, 1988).
The ventral horn, including the motor nucleus (lamina F) of the long-tailed ray, also contained many enkephalinergic s o n s and terminals such as have been reported in the ventral horn of turtles (Reiner, 19871, the lizard (Wolters et al., 19861, the domestic fowl (LaValley and Ho, 1983), the frog (Lorez and Kemali, 19811, the rainbow trout (Vecino et al., 1992), and in mammals, including man (Schoenen et al., 1985; Tashiro et al., 1987).
The grey matter of the elasmobranch spinal cord was characterized by a series of longitudinal fascicles which occupy much of the volume of laminae C-E (Nieuwenhuys, 1964; Cameron et al., 1990). According to Nieuwenhuys (19641, some dorsal root fibres enter these longitudinal fascicles (intracor- nual bundles), whereas others contribute to the much reduced dorsal columns. In the long-tailed ray only the longitudinal fascicles in lamina C were, like thc dorsal roots, devoid of ENK-LI whereas the dorsal columns and the longitudinal fascicles in laminae D and E contained many enkephalin-positive fibres.
Nieuwenhuys (1964) has suggested that the dorsal funiculi of elasmobranchs are composed of a subpopulation of dorsal root fibres. Although this may be partially correct, our finding that some axons in the dorsal funiculi of the long-tailed ray contain ENK-LI suggests that at least some fibres are not primary afferents.
Enkephalinergic fibres have been described in the dorsal part of the lateral funiculi of turtles (Reiner, 1987) and in the lateral and ventral funiculi of the frog (Lorez and Kemali, 1981). In the long-tailed ray enkephalinergic axons were found in the dorsolateral portion of the lateral funicu- lus near the pial surface and in a deeper crescent-shaped band in the lateral funiculus, as well as throughout the ventral funiculus. Chronic transection of the spinal cord in the long-tailed ray resulted in a disappearance of enkepha- linergic axons in the lateral and ventral funiculus caudal to the lesion, but no loss of ENK-LI in the dorsolateral, superficial part of the lateral funiculus (Snow, Renshaw, Wise, and Hamlin, pers obs). Enkephalinergic axons in the ventral funiculus and deep in the lateral funiculus, there- fore, probably belong to a descending spinal tract(s), whereas those in the superficial part of the dorsolateral region belong to an as yet unidentified ascending system. It is relevant to mention that in mammals some enkephalinergic
Longitudinal fascicles and the dorsal roots.
Funiculi o f cord.
P.J. SNOW ET AL.
cells in laminae VI and VII and around the central canal are actually spinothalamic or spinoreticular tract neurons (Na- hin and Micevych, 1986; Coffield and Miletic, 1987; Nahin, 1988; Willis and Coggeshall, 1991).
Enkephalinergic fibres have been seen in commissural fibres of turtle (Reiner, 1987) and in the ventral commissures of the primate (Lamotte, 1988). In the long-tailed ray both the dorsal and ventral commis- sures contained punctate ENK-LI, suggesting that enkepha- lin may be involved in the integration of spinal centers on different sides of the cord.
In the long-tailed ray many enkephalinergic fibres could also be seen projecting into the lateral and ventral funiculi, although none were seen entering the dorsal funiculus. Similar enkephalinergic processes have been reported in lateral and ventral funiculi of the spinal cord of turtles (Reiner, 19871, suggesting that in elasmobranchs and turtles there may be opiate-mediated control of transmission along long ascending and descending spinal pathways.
Functional considerations Dorsal horn. The endogenous opiates are generally
considered to mediate neuronal inhibition (Willis and Cog- geshall, 1991). In the mammalian dorsal horn enkephalin is thought to specifically inhibit ascending tract cells which receive input from nociceptive afferents (Willis and Cogge- shall, 1991). One might therefore expect a correlation between the presence of enkephalinergic terminals and cells in the dorsal horn and presence of terminals of nociceptive afferent fibres. To date, of all vertebrates where the immunocytochemical detection of spinal enkephalin has been attempted, only the lamprey appears devoid of ENK-LI (Buchanan et al., 19871, and it is interesting that electrophysiological studies of primary afferents in this animal have not revealed any nociceptive afferent fibres (Christenson et al., 1988). Like other elasmobranchs, the long-tailed ray lacks the unmyelinated primary afferent fibres, which in mammals constitutes a large proportion of the nociceptive fierents (Coggeshall et al.: 1978; Snow et al., 1993). In the Atlantic stingray, Dasyatis sabina, Leo- nard (1985) has reported recording from a few high- threshold mechanoreceptors, but there is no evidence for the existence of afferent fibres that respond to heat or irritant chemicals, or for the polymodal nociceptors, such as have been well documented in mammals (Willis and Cogge- shall, 1991). For these reasons we have suggested that elasmobranchs have no more than a very rudimentary system for registering tissue damaging stimuli (Snow et al., 1993). It was, therefore, curious to find in the superficial dorsal horn of the long-tailed ray such high levels of ENK-LI in s o n s and terminals and 34% of all the enkepha- linergic spinal cells. In rays the enkephalinergic system within the substantia gelatinosa might inhibit the transmis- sion of information from the AS-afferent fibres which have been shown to project to this region (Leonard et al., 1978).
The ventral horn, including the motor nucleus (lamina F), of the long-tailed ray was rich in enkephalinergic terminals. There exists a large literature documenting the occurrence of enkephalin in the ventral horn of' vertebrates where clearly it is unlikely to be involved in the regulation of the perception of pain. In mammals physiological studies have demonstrated the involvement of endogenous opiates in the suppression of spinal reflexes (Clarke et al., 1992; Zhuo et al., 1993). However, enkephalinergic cells have not been reported in
Transverse fibre systems.
Ventral horn.

ENKEPHALIN IN ELASMOBRANCH SPINAL CORD 273
Leonard, K.B. (1985) Primary afferent receptive field properties and neuro- transmitter candidates in a vertebrate lacking unmyelinated fibres. Prog. Clin. Biol. Res. 176:135-145.
Leonard, H.U., P. Rudomin, and W.D. Willis (1978) Central effects of volleys in sensory and motor componcnts of peripheral nerve in the stingray, Dasyatcs sahina. *J. Neiirophysiol. 41:108-125.
Lorez, H P . , and M. Kemali !1981) Substance P, met-enkephalin and somatostatin like immunoreactivity distribution in the frog spinal cord. Neurosci. Lett. 26:119-124.
Nahin, R.L. (1988) Immunocytochemical identification of long ascending, peptidergic lumbar spinal neuronl; terminating in either the medial or lateral thalamus in the rat. Brain Res. 443345-349,
Nahin, R.L., and P.E. Micevych (1986) A long ascending pathway o f enkephalin-like immunoreactive spinoreticular neurons in the rat. Neu- rosci. Lett. 65:271-276.
Naik, D.R., M. Sar, and W.E. Stiimf (1981) Immunohistochemical localiza- tion of enkcphalin in the central nervous system and pituitary of the lizard, Anolzs carolinensis. J. Comp. Neurol. 198: 583-601.
Niewenhuys, R. (1964) Comparative anatomy of the spinal cord. In J.C. Eccles and J .P. Schade (eds): Progress in Brain Res. New York: Elsevier, vol. 11, pp. 1-57.
Northcutt, R.G. (1981) Evolution of the trlcncephalon in nonmammals. Annu. Rev. Neurosci. 4:301-350.
Reiner, A. (1987) The distribution of proenkephalin-derived peptides in the central nervous system of turtles. J . Comp. Neurol25.9:65-91.
Sar, M., W.E. Stumpf, R.J. Miller, K. Chang, and P. Cuatrecasas (19781 lmmunohistochemical localization of enkephahn in rat brain and spinal cord. J . Comp. Neurol. 182:17-38.
Sasek, C.A., and R.P. Elde (19861 Cooexistence of enkephalin and dynorphin immunoreactivities in neurons in the dorsal gray commissure of the sixth lumbar and first lumbar spinal cord segments in rat. Brain Res. 38%-14.
Schoenen, J . , F. Lutstra, G. Vierendeels, M. Reznik, and J.J. Vanderhae- ghen. (1985) Substance P, enkephalins. somatostatin, cholecystokinin, oxytocin and vasopressin in human spinal cord. Neurology 35881-890.
Snow, P.J , M.U. Plenderleith, and L.L. Wright. (19931 Quantitative study of primary sensory neuronc populations of three species of elasmobranch fish. J. Comp. Neurol. 334,97-103.
Snow, P. ?J., and M.B. Plenderleith 11992) Sensory neurons, peptides and pain-functional, phylogenetic and developmental considerations. In S.K. Manchanda, W. Selvanurthy, and V. Mohan Kumar icds): Advances in Physiological Science. New Dehli: Macmiillan India Ltd., pp. 705-716
Tashiro, T., 0. Takahashi, T. Satoda, H. Matsushima, and N. Mizuno. (1987) Immunohistochemical demonstration of coexistence of enkephalin- and substance P-like immunoreactivities in axonal components in the lum- bar segments ofcat spinal cord. Brain Res. 424:391-395.
Uhl: G. R., R. R. Goodman, M.J. Kuhar, S.K. Childers, and S.H. Snydidrr 11979) Immunohistochemical mapping of enkephalin cell bodies, fibres and nerve terminals in the brainstem of the rat. Brain Res. 166.75-94.
Vecino. E,, C. Pinuela, K. Arevalo, J. Lara, J.R. Alonso, and J. Aijon. (1992) Distribution of enkephalin-like immunoreactivity in the central nervous system of the rainbow trout: An immunocytochemicztl study. J. Anat. 180:436-453.
Whittington, I.D., and P.H. Last (19Y4) Himanluro fai Jordan and Seale (Myliobatiformes: Dasyatididae), from Heron Island and its monogenean parasite fauna. Memoirs of the Queensland Museum 35: 285-289.
Wilson, P., D.E.R. Meyers, and P.J. Snow (1986) The detailed somatotopic organization of the dorsal horn in the lumbosacral enlargement of the cat spinal curd. J. Neuroph.ysio1. 5Ii:604-617.
Willis, W.D., and R.E. Coggeshall(1991’ Sensory Mechanisms of the Spinal Cord. New York: Plenum Press.
Woolf, C.J., and M. Fitzgerald 11986) Sornatotnpic organization of cutaneous afferent terminals and dorsal horn neuronal receptive fields in the superficial and deep laminar nf the rat lumbar spinal cord. J. Comp. Neurol. 2513 17-53 1.
Wnlters, J G . , H.J. ten Donkelaar, and A.A.J. Verhofstad (19861 Distribution of some peplides (substance P, leu-enkephalin, met-enkephalin) in the brain stem and spinal cord or a lizard, Varanus p ~ a r ~ t h ~ . n ~ ~ t i c ~ ~ . Neuro- science 18:917-946.
Zhuo, H., S.J. Fung. and C.D. Barnes (1993) Opioid action on spinal cord reflexes due to dorsolateral pontine tegmentum stimulation. Neurophar- macol. 32621-631.
the ventral horn of any vertebrate. Thus the source of enkephalinergic terminals in the ventral horn must be cells in either the dorsal or intermediate portions of the spinal grey or in descending tracts such as the pontospinal path- way in the cat (Zhuo et al., 1993). The involvement of enkephalin in the regulation of motor outflow may not he restricted to the regulation of reflexes. For instance, it may be that these animals utilize enkephalinergic inhibition of moroneurons to enable them to attain the complete immo- bilization which they so often adopt when they lie half- buried in the sand.
AKIVOWLEDGMENTS We thank the Director of the Heron Island Research
Station, Dr. Ian Lawn, for providing the facilities which has made this research possible. We are also indebted to Drs. P. Wilson and P. D. Kitchener for critically reading this manuscript and Ms. L. Banh and Mr. A. Hamlin for their skilled technical assistance.
LITERATURE CITED Buchanan, J.Y., L. Brodin, T. Hokfelt, P.A.M. van Dongen, anrl 6. Grillncr
(1987) Survey of neuropeptide-like immunoreactivity in the lamprey spinal cord. Brain Res. 408.299-302.
Cameron, A.A.? M.B. Plenderleith, and P.J. Snow (1990) Organisation of the spinal cord in four species of elasmohranch fish: cytoarchitecture anrl distribution nf serutonin and selected neuropeptides. J. Comp. Neurol.
Christenson, J.. A. Boman, P.A. Lagerback, and S. Grillner i1988) The dorsal cell, one class of primary sensory neurons in the lamprey spinal cord. I. Touch, pressure but no nociception - a physiological study. Brain Res. 44O:l-8.
Cuggeshall, R.E., R.B. Leonard, MI,. Appelbaum, and W.D. Willis (1978) Organization of peripheral nerves and spinal roots of the Atlantic stingray, Dasyatis sabina. J. Neurophysiol4 1:97-107.
Clarke, J.L. (1859) Further researches on the gray substance of the spinal cord. Phil. Trans. B 149:437-468.
Clarke, R.W., F.J. Galloway, J. Harris, J.S. Taylor, and T.W. Ford (19921 Opioidergic inhibition of flexor and extensor reflexes in the rabbit.. .I. Physiol. (Lond.) 449: 493-501.
Coffield, J.A., and V. hliletic (1987) Immunoreactlveenkephalin in contained within some trigeminal and spinal neurons projecting to the medial thalamus. Brain Res. 425:380-383.
Dubner, R.. M.A. Ruda, V. Miltex, M.J. Hofferet, G.J. Bennett, N. Nishi- kawa, and J. Coffield (1984) Neural circuitry mediating nociception in the medullaryand spinal dorsal horns. In: L. Kruger and J. C. Liebeskind (cds): Advances in Pain Research and Therapy. New York: Raven Press 6:15 1- 166.
Gibson, S.J.: J.M. Polak, S.K. Bloom, and P.D. Wall (1981) The distribution of nine peptides in rat spinal cord with special emphasis on the substantia gelatinosa and on the area around the central canal (Lamina XI. J. Comp. Neurol. 201r65-79.
Glaser, E.d.: and A.I. Basbaum (1981) Immunnhistnchemical localisation of lcucinr-cnk<*phali n in the spinal cnrd o f the cat: enkephalin containing marginal neurnns and pain modulation. J. Comp. Keurol. 196: 377-389.
Kitchener, P.D., P. Wilson, and P.J. Snow (1994) Sensory dcrmatomes in the pectoral and pelvic fins of the stingray Hirnanturu uamak. Brain Behaviour and Evolution (in Press).
Kitrhmer, P.D.: G. Wise, and P.J. Snow (1994) Inter- and intrasegment.al spinal reflexes in the long-tailed stingray, Hirnanturu fa;. Proc. Aust. Physinl. Pharmacol. Soc. 2512):168p.
Lamotte, C.C. (1988) Lamina X of primate spinal cord: Distribution of five neuropeptides and serotonin. Neuroscience 25:639-658.
LaValley, A.L., and R.H. Ho (1983) Substance P , somatostatin and methio- nine-enkephalin immunoreactive elements in the sponal cord of the domestic fowl, Gallus domesticus. J. Comp. Neurol. 21.3: 406-413.
z 9 7 : ~ 1 i -218.




![BMC Biology BioMed Central - Springer[D-ala2,D-leU5]enkephalin (DADLE) reduFigure 2ces necrotic and apoptotic (panel B) cell death associated with ischemia [D-ala2,D-leU5]enkephalin](https://img.pdfslide.us/doc/110x75/5e396225173d974deb7f955c/bmc-biology-biomed-central-springer-d-ala2d-leu5enkephalin-dadle-redufigure.jpg)
























