Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1985 by The American Society of Biological Chernista, Inc.
Vol. 260, No. 17, Issue of August 15, pp. 9713-9719.1985 Printed in U.S.A.
Localization of a Fibrinogen Calcium Binding Site between y-Subunit Positions 3 11 and 336 by Terbium Fluorescence*
(Received for publication, November 30,1984)
Chi V. DangS, Ray F. Ebert, and William R. Bell From the Department of Medicine, Division of Hematology, The Johns Hopkins University School of Medicine, Baltimore, Maryland 21205
Calcium is required for effective fibrin polymeriza- tion. The high affinity Ca” binding capacity of fibrin- ogen was directly localized to the y-chain by autora- diography of nitrocellulose membrane blots of fibrin- ogen subunits incubated with *%a2+. Terbium (Tb”) competitively inhibited “Ca2+ binding to fibrinogen during equilibrium dialysis, accelerated fibrin polym- erization, and limited fibrinogen fragment D digestion by plasmin. The intrinsic fluorescence of Ca’+-depleted fibrinogen was maximally enhanced by Ca” and Tb”, but not by M8+, at about 3 mol of cationlmol of fibrin- ogen. Protein-bound TbS+ fluorescence at 545 nm was maximally enhanced by resonance energy transfer from tryptophan (excitation at 290 nm) at about 2 mol of Tb3+/mol of fibrinogen and about 1 mol of Tb3+/mol of plasmic fragment Der (Mr 94,000). Fibrinogen frag- ments D78 (M. 78,000) and E did not show effective enhancement of Tb3+ fluorescence, suggesting that the Ca2+ site is located within 7303 to 7411, the peptide which is absent in fragment but present in Der. When CNBr fragments of the carboxyamidated y-subunit were assayed for enhancement of Tb3’ fluorescence, peptide CBi (7311-338) bound 1 mol of Tb3+/mol of CBi. Thus, the Caz+ site is located within this peptide. The sequence between 7316 and 7329 is homologous to the calmodulin and parvalbumin Ca” binding sites.
Fibrinogen is composed of three pairs of subunits, Aa, BB, and y (1). Electron microscopic studies of fibrinogen reveal a trinodular rod structure consisting of 2 large outer nodules, the D-domains, connected to a smaller central nodule, the E- domain (2). Calcium is the physiological divalent cation es- sential for the biological functions of fibrinogen and fibrin polymerization (1). Fibrinogen appears to have three high affinity Ca2+ sites (& -1 PM) and many low affinity divalent cation sites (1, 3, 4). Plasmin digestion of fibrinogen (Mr 340,000) yields terminal degradation products of 1 mol of fragment E (Mr -50,000) and 2 of fragment D (Mr -100,000). Fragment D binds 1 Ca2+ ion, which accounts for 2 of the 3 high affinity Caz+ sites (1,4, 5). Plasmic fragment E is devoid of Ca2+ binding capacity; however, the central fibrinogen
* This work was supported in part by Research Grants HL 24898, HL 07377, and HL 29067 from the National Heart, Lung and Blood Institute, National Institutes of Health, The Whitehall Foundation, and Hubert E. and Anne E. Rogers Scholar in Academic Medicine. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$To whom correspondence should be addressed at: Cancer Re- search Institute, University of California, San Francisco, Moffitt 1282, San Francisco, CA 94143.
domain, the amino-terminal disulfide knot, prepared by cyan- ogen bromide degradation appears to contain the third Ca2+ site (1, 5). This site is apparently destroyed by limited prote- olysis of fibrinogen (6).
Ca2+ limits plasmin digestion of fibrinogen fragment D y- subunit (1, 7). In the presence of Caz+, plasmin produces fragment D,, (Mr 93,000) with subunits D@ (Mr 43,000), Dy (MI 38,000), and Da (Mr 12,000) (7). In the presence of EGTA,’ plasmin digestion of Ca2+-depleted fibrinogen yields a smaller fragment, DEGTA (M, 8@,000), which contains a smaller Dy-subunit (MI 25,000) (7). An intermediate fragment Dint (Mr 84,000) containing an intermediate Dy-subunit (Mr 29,000) was prepared recently (8). Both D,, and Dint, but not DEGTA, bind Ca2+, suggesting that the Ca2+ site is located somewhere between positions 7 3 0 3 and y3@ of the y-subunit (8). This segment is present in Dbt and D,.,, but missing in DECTA. Although these facts are highly suggestive of the location of the CaZ+ site, direct evidence for the location of the fibrinogen D-domain Ca2+ site was lacking.
In the present report we show that the fibrinogen D-domain CaZ+ site is located between positions y311 and y3=. The results indicate that terbium, a trivalent lanthanide, competitively inhibits Ca2+ binding to fibrinogen, accelerates fibrin polym- erization, and limits plasmin digestion of fragment D. T b 3 +
appears to bind to the D-domain Caz+ site with an enhance- ment of T b 3 + fluorescence by resonance energy transfer from tryptophan. Employing this specific enhancement of Tb3+ fluorescence, we have localized the D-domain Ca2+ site to peptide CBi (7311-336) prepared by cyanogen bromide degra- dation (9) of purified carboxyamidated y-subunit.
EXPERIMENTAL PROCEDURES
Materials-MgCl~. 6Hz0, CdC12. 2Hz0, and CaClz were products of Fisher. Terbium chloride (TbC13. 6HzO) was obtained from Alfa Di- vision, Ventron Corp. (Danvers, MA). %aClZ (19.08 mCi/mg) was obtained from New England Nuclear. Solutions were treated with Chelex 100 (Bio-Rad) to remove metal ions. Contaminating calcium was <I p~ as determined by atomic absorption spectroscopy. Poly- acrylamide and SDS were products of Bio-Rad. Dithiothreitol, EGTA, cyanogen bromide, Tris, and tryptophan were from Sigma. Human fibrinogen, plasmin, and chromogenic substrates S2251 and S2160 were from Kabi-Vitrum (Stockholm, Sweden). Purified human CY-
thrombin (3169 U. S. units/mg) was a gift from Dr. G. Murano (Bureau of Biologics, Bethesda, MD). Nitrocellulose sheets were obtained from Schleicher & Schuell. All other reagents were of highest analytical grade.
Plusmin and Thrombin Assays-Chromogenic substrate assays for plasmin with S2251 and for thrombin with S2160 were performed according to the manufacturer’s instructions.
Protein Determination-The protein concentration was deter-
’ The abbreviations used are: EGTA, ethylene glycol bis(p-amino- ethyl ether)-N,N,N’,iV”tetraacetic acid SDS, sodium dodecyl sul- fate; HPLC, high performance liquid chromatography; PAGE, poly- acrylamide gel electrophoresis.
9713

9714 Fibrinogen Calcium Binding Site mined by the method of Lowry et al. (10). Fibrinogen concentrations were also determined spectrophotometrically a t 280 nm with E;?,,, = 15 (11). The concentration of' peptide CBi was measured at 280 nm with tzRO = 15,300 (Et:,,, = 49), a value which was determined by amino acid analysis.
1 2 3 4
/ Aa
- BP \ Y
FIG. 1. Calcium binding by fibrinogen y-subunit. Carbox- amidated fibrinogen (lanes 2 and 4 ) or purified carboxamidated y- subunit (lanes 1 and 3) were separated in an 8% SDS-polyacrylamide gel under nonreducing conditions. The gel was stained with Coomas- sie Blue G-250 (lanes 3 and 4 ) . Peptides of an identical gel was blotted onto nitrocellulose and stained with "Ca2+. Autoradiography was performed for 24 h (lanes 1 and 2).
c 0
1
X Y
0
8-
6 -
4-
2 - -
O0-
0 I I
\
b \
FIG. 2. Scatchard analysis of calcium binding to fibrinogen in the absence (0) and presence of 20 pM terbium (Tb") (0). The abscissa ( r ) represents mol of bound Ca2+/mol of fibrinogen, and the ordinate ( r / C ) represents r divided by the free Ca2+ concentration (C).
1 I 1 1 I
* I / I I I
n 3 - 2 3 1 4 6 0
Time (minutes) FIG. 3. Clotting of EGTA-treated fibrinogen in the presence
of no added cation ( I ) , 5 mM M g + (2), 20 p~ Ca2+ (3), 3 mM Ca2+ (5), 5 mM Ca2+ (6), or 20 pM Tb3+ (7). Polymerization of citrate-free fibrinogen with no added Ca2' (4 ) . Fibrinogen (1 mg/ml) was polymerized at 25 "C by addition of thrombin (1 U. S. unit/ml) a t time zero.
SDS-PAGE, Protein Blotting, and Detection of "C2+-binding Pep- tides-SDS-PAGE was performed in 0.8-mm slab gels as described (12). Polypeptides were transferred to nitrocellulose sheets (13) in absence of SDS and stained with 45Ca2+ (14) to identify Ca2+-binding polypeptides. Autoradiography was performed with Kodak X-Omat AR film.
Plasmin Digestion of Fibrinogen and Preparation of Fibrinogen Fragments D and E-Citrate-free fibrinogen was prepared by dialyz- ing human fibrinogen (Kabi, grade L) at 4 "C against 1000 volumes of 0.3 M NaCl (adjusted with NH40H to pH 8) with three 12-h changes, lyophilized, and stored at -20 "C. Citrate-free fibrinogen (2 mg/ml) in 50 mM Tris (pH 7.2) and 150 mM NaCl (final concentra- tion) was incubated with purified plasmin (0.18 IU/ml) and various cations for 36 h at 25 "C. The reaction was stopped by addition of phenylmethylsulfonyl fluoride at a final concentration of 0.2 mM.
Fibrinogen degradation fragments D and E were prepared as de- scribed (15). Human fibrinogen (200 mg) was digested with plasmin (0.0045 IU/mg of fibrinogen) in the presence of 5 mM CaCh or 10 mM EGTA at 37 "C for 15 h. The reactions were stopped with phenylmethylsulfonyl fluoride as above. The digest was applied to a DEAE-cellulose DE52 column (2.5 X 45 cm) and eluted with two linear gradients: 0-0.09 M NaCl and 0.09-0.17 M NaCl in 0.01 M NaHC03 (pH 8.9) buffer. Protein concentration was determined spectrophotometrically a t 280 nm. Pooled fractions were dialyzed at 4 "C against 100 volumes of distilled deionized water with four 6-h changes, lyophilized, and stored at -20 "C. Fragment DS (MI 94,000) was prepared in 5 mM CaC12 and fragment D,B (M, 78,000) in 10 mM EGTA. Fragment E ( M , 52,000) was prepared in CaC12 or EGTA.
Equilibrium Dialysis-Equilibrium dialysis to measure Ca2+ bind- ing was performed as described (6). Duplicate 5O-pl samples of EGTA- treated (6) fibrinogen (1-3 mg/ml) were dialyzed against Ca2+ at concentrations from 1.6 p~ to 0.1 mM in 10 ml of 50 mM Tris (pH 7.0) and 150 mM NaCl at 25 "C for 48 h. Fifteen ml of *Ca (stock solution of 1.0 mCi/ml, 19.08 mCi/mg) were added to each Caz+ solution or to the above buffer. An identical set of samples was dialyzed in Ca" solutions containing 20 p M TbCl3. Samples of 30 pl were removed, diluted in 10 ml of Ready-solvEP (Beckman Instru- ments) and counted for 5 min in a Beckman LS-133 scintillation counter. Counts from duplicate cells did not vary by more than 5%. Scatchard analysis was performed, assuming M, 340,000 for fibrino- gen and employing least squares analysis.
Fibrin Aggregation and Tb3+-induced Protein Aggregation Assays- Fibrin aggregation was monitored by continuously recording turbidity a t 350 nm of solutions containing 1 mg/ml of EGTA-treated or citrate-free fibrinogen, 1.0 U. S. unit/ml of thrombin, and various

Fibrinogen Calcium Binding Site 9715
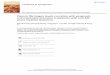
FIG. 4. Protective effects of Ca2+, Tbs+, and Cd2+ in the degradation of fibrinogen by plasmin. Fibrinogen (2 mg/ml) was digested with purified plas- min (0.18 IU/ml) in the presence of 20 pM Ca2+, 20 pM T b 3 + , 20 pM Cd2+, 20 pM Me, 10 mM EGTA, or without addition 094. of any cations in the control (Ct) sample. Standards: phosphorylase b (92 kDa) and bovine serum albumin (66 kDa). Fi- 082. brinogen fragments DM (M, 94,000), De2 D78e ( M , 82,000) and D78 (M, 78,000) are iden- tified.
Ca2+ Tb3+ Cd2+ EGTA Mg2+ Ct
[Tb3'] x M FIG. 5. Terbium-induced aggregation of fibrinogen (10 PM),
fragment E (10 am), and fragment De4 (10 pM) in the absence of thrombin. Protein solutions were incubated (25 "C) with various concentrations of Tb3+ for 5 min and turbidity was measured by absorbance at 400 nm.
I I 1 I 1 I I
A 8
d
I I I I I
-0 2 4 6 O U Y " I
Mol Cation/Mol Fibrinogen FIG. 6. Enhancement of EGTA-treated fibrinogen (10 PM)
intrinsic fluorescence at 345 nm (excitation at 280 nm) by the addition of Mg2+ (0), TbS+ (A), or Ca2+ (0). Precipitation of fibrinogen occurred at Tb3+ concentrations greater than 40 p ~ .
concentrations of cations in 50 mM Tris (pH 7.4) and 150 mM NaCl at 25 "C.
Solutions (1 ml) of EGTA-treated fibrinogen or fragments Dm or E in 50 mM Tris (pH 7.0) and 150 mM NaCl were incubated with varying amounts of T b 3 + for 5 min at 25 "C. Turbidity was recorded at 400 nm and was stable over 30 min.
- 92kDa
c 66kDa
Fluorescence Spectroscopy-Fluorescence measurements were made at 25 "C on a Perkin-Elmer 650-10s fluorescence spectropho- tometer a t a spectral band width of 2 nm for excitation and 10 nm for emission. Samples were diluted in 50 mM Tris (pH 7.0) and 150 mM NaCl in a 3-ml cuvette with 1-cm path length. Titrations with TbC13, CaC12, or MgCI2 were made by sequential addition of 5 pl of a 0.4 M stock solution of cation to a 1-ml solution of EGTA-treated fibrinogen, fragment D, or fragment E. Binding of cations to fibrin- ogen was followed by an increase in intrinsic fluorescence at 345 nm on excitation at 280 nm. Binding of T b 3 + to fibrinogen, fragment D, or fragment E was measured by an increase in fluorescence at 545 nm by resonance energy transfer from Trp to T b 3 + on exciting the former at 290 nm (16). Binding of Tb3+ to peptide CBi (y311-~) was performed in water adjusted to pH 7.0 with 0.1 M NH,OH. Titrations were made by sequential addition of 5 pl of a 40 p~ stock T b 3 +
solution. The total volume change was 15% or less. Fluorescence intensities were corrected for dilution. Inner filter effect may occur a t the high protein concentrations required to detect T b 3 + fluores- cence. However, since relative fluorescence was measured, this poten- tial source of error is minimized. Solutions of CNBr peptides had absorbances below 0.10 at 280 nm.
Preparation of Carboxyamidated Fibrinogen and Purification of Carboxyamidated y-Subunit-Citrate-free fibrinogen (300 mg) in 0.3 M NaCl was reduced with 50 mg of dithiothreitol and carboxyami- dated with 0.3 g of iodoacetamide according to Murano et al. (171, except that iodoacetamide was used instead of iodoacetic acid. Car- boxyamidated y-subunit was purified to >95% from carboxyamidated fibrinogen by CM-cellulose column (2.5 X 20 cm) chromatography (17).
Cyanogen Bromide Degradation of Carboxyamidated Fibrinogen y- Subunit-Carboxyamidated y-subunit (55 mg) was dissolved in 2 ml of 70% formic acid and incubated with 0.5 g of CNBr for 12 h at 25 "C according to Henschen and Warbinek (9). The sample was applied to a Sephadex G-50 column (2.5 X 100 cm) and eluted with 5% formic acid at 70 ml/h. Three-ml fractions were collected, and protein was determined spectrophotometrically at 280 nm. The three peaks of CNBr peptides were collected, lyophilized, and stored at -20 "C.
Isolation and Amino Acid Analysis of CNBr Peptides by HPLC-A Varian 5500 liquid chromatograph equipped with a UV-200 detector and CDS-402 data station was used for HPLC. Solvents used were: A, 0.1% aqueous trifluoroacetic acid; and B, acetonitrile. Peak 111 material from the Sephadex G-50 purification step was lyophilized and resuspended in water (adjusted to pH 8.0 with 0.1 M NH,OH). Aliquots of 200 pl were injected onto a Vydac T P C-4 column (1 x 25 cm, 214TP510) at room temperature. Starting conditions (10% sol- vent B, 2 ml/min) were held for 5 min, then a linear gradient of 10- 40% solvent B was executed over a 40-min period. All peptides with UV absorption at 280 or 210 nm were eluted under these conditions. The column was washed with 65% solvent B between runs.
Amino acids after hydrolysis (in 6 N HCI at 150 "C for 90 min) were analyzed as the phenylthiocarbamyl derivatives by the method of Cohen et al. (18).
RESULTS
Calcium Binding by Fibrinogen ?-Subunit The relative Ca2+ binding affinities of the carboxyamidated
fibrinogen subunits were determined by blotting of the sub- units separated by SDS-PAGE onto a nitrocellulose mem- brane and probing with 4sCa2+. Only the y-subunit bound

Fibrinogen Calcium Binding Site
1 I
Fibrinogen A
-Ca2+ or
0 0 0+Mg 2t- 0
0 0
0 0
0
0 0
FIG. 8. CNBr peptides (2-ml sam- ple) from cyanogen bromide- cleaved carboxyamidated y-subunit (55 mg) were applied to a Sephadex G-50 column (2.5 X 100 cm) and eluted with 5% formic acid at 70 ml/ h. Fractions (3 ml) were assayed by ab- sorbance at 280 nm. Pooled fractions (horizontal bars) were lyophilized and tested for Tb3+ fluorescence (545 nm) enhancement by resonance energy trans- fer from Trp excited at 290 nm. Vertical open bars represent the relative enhance- ment of Tb'+ fluorescence by pools I , 11, and 111.
4sCa2+ (1 p M ) (Fig. 1) in the presence of 5 mM M&l2, 60 mM KCl, and 10 mM imidazole (pH 6.8). When blotted, reduced human fibrinogen subunits were separated by SDS-PAGE, only the y-subunit bound 4sCa2+ (data not shown).
Terbium Substitution for Ca" in Fibrinogen To ascertain whether T b 3 + can effectively substitute for
Caz+ in fibrinogen, we examined the following: (i) inhibition of Ca2+ binding to fibrinogen by T b 3 + as determined by equilibrium dialysis; (ii) acceleration of fibrin polymerization by Ca2+ and T b 3 + ; (iii) limitation of plasmin digestion of fibrinogen fragment D by Ca2+ and T b 3 + ; and (iv) enhance- ment of intrinsic fibrinogen fluorescence by Ca2+ and T b 3 + .
(i) Inhibition of Ca2+ Binding to Fibrinogen by Tb3+-Ter- bium was found to inhibit Ca" binding to fibrinogen as
U
Fraction Number
determined by equilibrium dialysis (Fig. 2). The dissociation constant (&) for Ca2+ is 3.7 p~ in the absence of T b 3 + , and an apparent K d = 19 p M was found in the presence of 20 pM Tb3+. At 50 p~ T b 3 + , "CaZ+ binding to fibrinogen was very low and not reliably detectable. In the presence of 50 p~ Ba2+, Ca2+ binding was inhibited with an apparent K d of 5.7 p M (not shown). Thus, T b 3 + is a relatively potent inhibitor of Ca2+ binding to fibrinogen.
(ii) Acceleration of Fibrin Polymerization by Ca2+ and T b 3 + -
The ability of Caz+ and T b 3 + to accelerate fibrin polymeriza- tion was assessed by continuous turbidity measurement at 350 nm of solutions containing 1 mg/ml of EGTA-treated fibrinogen and 1 U. S. unit/ml of thrombin. Thrombin was added at time zero. Thrombin activity as determined by the hydrolysis of chromogenic substrate S2160 was not altered by

Fibrimgen Calcium Binding Site 9717
E t
4 N
FIG. 9. HPLC of pool I11 CNBr peptides from the Sephadex G-SO E o.2
- column (see legend to Fig. 8). Con- 8 ditions were as described under "Exper- c imental Procedures." For this chromat- 0.' - ogram, 0.5 mg of peptides injected in 200 2 d. u)
6
2
2 O . 3
0
Wavelength (nm) FIG. 10. Fluorescence spectra of peptide CBi (5 @M) in the pres-
ence (-) and absence (- - -) of saturating T b 3 + are shown in the rightpanel. Excitation was at 290 nm. First order Rayleigh scattering at 290 nm is noted. The left panel shows enhanced T b 3 + fluorescence (545 nm) which was not detectable in the absence of peptide CBi.
T b 3 + when compared with Ca2+ or M F at cation concentra- tions used in the fibrin polymerization assay.
In the absence of added cation (Fig. 3, curve 1) there was minimal change in turbidity. M F (5 mM) did not enhance aggregation (Fig. 3, curve 2). At a concentration of Ca2+ (20 pM) adequate to limit plasmin digestion of fibrinogen frag- ment D and to saturate the high affinity binding sites, slight enhancement of turbidity resulted (Fig. 3, curve 3). Near maximal increase in turbidity was attained at Ca2+ concentra- tions of 2 and 5 mM (Fig. 3, curves 5 and 6). Citrate-free fibrinogen exhibited an increase in turbidity in the absence of added Ca2+ (Fig. 3, curve 4). In contrast, at 20 p~ Tb3+ there was a large thrombin-dependent increase in turbidity (Fig. 3, curve 7). The molar ratio of T b 3 + to fibrinogen was 7:l at the concentrations of 20 p~ T b 3 + and 1 mg/ml of fibrinogen.
(iii) Protective Effect of Tb3+ and Ca2+ in the P h m i n Deg- radation of Fibrinogen-Tb3+ and Cd", but not M$+, limited
IO 20 30 40 Elution Time (minutes)
plasmin digestion of citrate-free fibrinogen in a manner sim- ilar to that of Ca2+ (Fig. 4). The ability of plasmin to hydrolyze the chromogenic substrate ,92251 was not altered at compa- rable concentrations of Ca2+ and T b 3 + (not shown). In the presence of EGTA, Ca2+-depleted fibrinogen was degraded to fragment D78. A reduced SDS-polyacrylamide gel of fragments De4 and D78 showed differences only in the length of the y- chain (not shown).
(iu) Enhancement of Fibrinogen Intrinsic Fluorescence by Ca2+ and Tb3+-EGTA-treated fibrinogen was titrated with Tb3+, Ca", and M$+ to determine whether intrinsic fluores- cence would change upon binding of these cations. Before proceeding to fluorescence spectroscopy, the effect of T b 3 +
concentration on the turbidity (due to aggregation) of solu- tions of fibrinogen (3.4 mg/ml), fragment Dm (1 mg/ml), and fragment E (0.5 mg/ml) was determined (Fig 5). Thrombin- independent increases in turbidity were reversible with 5 mM EGTA. The results indicate that fluorescence experiments must be performed at Tb3+ final concentrations well below 0.1 mM.
Fig. 6 shows the enhancement of fibrinogen intrinsic fluo- rescence (excitation at 280 nm and emission at 345 nm) by Ca2+ and T b 3 + . Maximal enhancement occurs at 3 mol of Ca2+ or Tb3+/mol of EGTA-treated fibrinogen. At Tb3+ concentra- tions greater than 40 p~ turbidity prevented accurate mea- surements of fluorescence. M e showed slight enhancement of fluorescence without obvious saturability. The increase in fluorescence was reversed by the addition of EGTA (5 mM final concentration). Thus, T b 3 + and Ca2+ have similar effects on the intrinsic fluorescence of fibrinogen.
Fluorescence Resonance Energy Transfer from Tryptophan to Tb3+ in Fibrinogen and Fragment Dg4-Tb3+ fluorescence at 545 nm was enhanced by resonance energy transfer from Trp excited at 290 nm when fibrinogen bound T b 3 + . Maximal enhancement occurred at 2 mol of Tb3+/mol of EGTA-treated fibrinogen (Fig. 7A). T b 3 + fluorescence was inhibited by Ca2+ but not by M$+. An uncorrected excitation spectrum of protein-bound Tb3+ (emission at 545 nm) exhibited a maxi- mum at 285 nm (not shown), indicating that Trp is the donor group. Excitation at 260 nm did not enhance T b 3 + fluores- cence at 545 nm. The results suggest that 2 fibrinogen Ca2+ sites have Trp in their immediate environment.
When plasmic fragments Dg4, D78, and E were titrated with Tb3+ (Fig. 7B), fragment E showed only weak enhancement of T b 3 + fluorescence. For fragment D78, a moderate enhance- ment of T b 3 + fluorescence which appeared maximal at 10 mol of Tb3+/mol of D78 was observed. Fragment Dg4 exhibited two classes of sites: one which binds 1 mol of Tb3+/mol of Dg4, and a second which binds 10 mol of Tb3+/mol of Dg4. Similar

9718 Fibrinogen Calcium Binding Site
TABLE I Sequences of the calmodulin, parvalbumin, and proposed fibrinogen Ca2+ sites
The sequences are aligned to show homologous regions. Dashes (between -X and -Z) represent an insertion in the y-subunit. X, Y, and 2 indicate the octahedral vertices of Ca2+ ligands. The parvalbumin EF-loop -X vertex is HzO.
+X Fibrinogen
+Y +Z -Y -X -Z 3 1 5 T r p - ~ - A s n - A s p - A s n - ~ - L y s - ~ - G l u - G l y - A s n - C y s - A l a - ~
ParvalbuminEF CD
89Gly-Asp-Ser-Asp-Gly-Asp-Gly-Lys-Ile-Gly-Val - Asp-% 50Ile-Asp-Glu-Asp-Lys-~-Gly-P&-Ile-~-Glu - Asp-%
Calmodulin I lSPhe-Asp-Lys-Asp-Gly-&-Gly-=-Ile-Th-Thr - Lys-Q I1
I11 55Val-Asp-Ala-Asp-Gly-@-Gly-=-Ile-&-Phe - Pro-=
I V 92Phe-Asp-Lys-Asp-Gly-&-Gly-~-Ile-Ser-Ala - Ala-% 128Ala-&-Ile-Asp-Gly-Asp-Gly-~-Val-&-Tyr - Glu-%
stoichiometries were obtained at 5 and 10 PM Dgr. However, since formation of low order oligomers cannot be ruled out, the stoichiometries observed may include the effect of oligo- merization.
Localization of a Ca2+ Site within the y-Subunit-Cyanogen bromide degradation products of the carboxyamidated y-sub- unit were separated by chromatography on a Sephadex G-50 column according to Henschen and Warbinek (9) (Fig. 8). The three peptide peaks were pooled and lyophilized. Pools I1 and I11 dissolved in distilled deionized water (pH 7.0, adjusted with 0.1 M NH,OH). Pool I was soluble at pH 6.0 or below. Pools I and I1 showed insignificant T b 3 + fluorescence enhancement when compared with pool I11 at the same pH.
Pool I11 has been reported (9) to contain two peptides: peptide CBi (7311-Glu-Phe-Ser-Thr-Trp-Asp-Asn-Asp-Asn- Asp-Lys-Phe-Glu-Gly-Asn-Cys-Ala-Glu-Gln-Asp-Gly-Ser- Gly-Trp-Trp-Hse) and peptide CBh (790-Leu-Glu-Glu-Ile- Hse).
The components of pool I11 were resolved by HPLC (Fig. 9). Peaks 1 and 2 were unidentifiable by amino acid compo- sition. The amino acid composition of peak 3 corresponds to peptide CBh. Peaks 4, 5, 6, and 7 (91% of total material absorbing at 280 nm) had identical amino acid compositions corresponding to that of peptide CBi. The microheterogeneity of peptide CBi is not understood, but it may be due to homoserine-/homoserine-lactone interconversion, partial deamination, or ion-pairing upon HPLC analysis. The yield of peptide CBi in pool I11 was 60% of the starting carboxy- amidated y-subunit.
Titration of pool I11 with T b 3 + showed maximal enhance- ment of fluorescence at 1.2 mol of Tb3+/mol of peptide CBi (Fig. 10). Tb3+ fluorescence was inhibited 70% by a 50-fold molar excess of Ca2+ or a 200-fold excess of M%+, and it was abolished by 5 mM EGTA. The peptide intrinsic fluorescence was quenched identically by Tb3+ and Ca2+. After HPLC (Fig. 9), Tb3+ fluorescence was enhanced by peak 4, but enhance- ment was markedly diminished in peaks 5, 6, and 7. The reason for this is unknown.
The Trp-Tb3+ distance was estimated to be greater than 6- 8 A (calculated to be 10.7 A) according to the Forster theory (16). This estimate was based on the determination of Trp quantum yield (a = 0.06) for peptide CBi using aqueous Trp (a = 0.14) as a standard (19). The ratio of the integrated emission intensities of T b 3 + (Am) and Trp AT^) in peptide CBi was 3.31 X (three determinations). The critical distance for 50% energy transfer, R,, was estimated to be 3.17 A and the Tb3+ fluorescence quantum yield was assumed to be 0.30 (16).
DISCUSSION
The binding of Caz+ to the fibrinogen D-domain Ca2+ site is necessary for the proper function of fibrinogen. Occupancy
of the high-affinity D-domain Ca2+ site, identified in this report, limits plasmin digestion of the y-subunit carboxyl- terminal segment. This segment is necessary for fibrinogen binding to the platelet membrane (20), clumping of staphy- lococci (21), fibrin polymerization (22), and cross-linking of fibrin by Factor XI11 (23).
In this report we describe observations indicating that T b 3 +
and Ca2+ bind to the same or equivalent sites on the y-subunit. These results provided a rational basis for localizing the Ca2+ site by monitoring T b 3 + fluorescence. The enhancement of Tb3+ fluorescence was localized to a peptide derived by CNBr degradation of purified carboxyamidated y-subunit. This pep- tide corresponds to peptide CBi (9) which contains residues
The sequence between positions 7315 to 7329 of peptide CBi is homologous to the calmodulin. (24) and to the parvalbumin Ca" binding sequences (Table I) (25). We provide evidence that this sequence is most likely the D-domain Ca2+ site which is composed of many ligand groups containing oxygen. A model of the plausible conformation of the fibrinogen Ca" site based upon homology with the parvalbumin EF-loop may be constructed (25); however, elucidation of the conformation of this fibrinogen Ca2+ site awaits detailed NMR studies using purified or synthetic peptide CBi or segments thereof. It is noteworthy that peptide CBi has a relatively low ion specific- ity compared with that of other EF-hand Ca2+ binding sites.
7311 to 7336.
Acknowledgments-We thank Dr. L. M. Amzel, Department of Biophysics, The Johns Hopkins University School of Medicine and Dr. D. C. H. Yang, Department of Chemistry, Georgetown University, for helpful suggestions and discussions and Kimberly Pruitt for preparation of the manuscript.
REFERENCES 1. Doolittle, R. F. (1984) Annu. Rev. Biochem. 53,195-229 2. Fowler, W. E., and Erickson, H. P. (1977) J. MOL Biol. 134,241-
249 3. Marguerie, G., Chagniel, G., and Suscillon, M. (1977) Biochim.
Biophys. Acta 490,94-103 4. Nieuwenhuizen, W., Van Ruijven-Vermeer, I. A. M., Nooijen, W.
J., Vermond, A., and Haverkate, F. (1981) Thromb. Res. 22,
5. Nieuwenhuizen, W., Vermond, A., and Hermans, J. (1983)
6. Marguerie, G., and Ardaillou, N. (1982) Biochim. Biophys. 701,
7. Haverkate, F., and Timan, G. (1977) Thromb. Res. 10,803-812 8. Nieuwenhuizen, W., Voskuilen, M., Vermond, A., Haverkate, F.,
and Hermans, J. (1982) Biochim. Biophys. Acta 707,190-192 9. Henschen, A., and Warbinek, R. (1975) Hoppe-Seyler's Z . Physiol.
Chem. 356,1981-1984 10. Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J.,
(1951) J. Bwl. Chem. 193,265-275 11. Doolittle, R. F. (1975) in Plasma Proteins (Putnam, F. W., ed)
Vol. 2, pp. 109-161, Academic Press, New York
653-657
Thromb. Res. 31,81-86
410-412

Fibrinogen Calcium Binding Site 9719 12. Ebert, R. F., and Bell, W. R. (1983) Proc. Natl. Acad. Sci U. S. 19. Anderson, J. S., Bowitch, G. S., and Brewster, R. L. (1983)
13. Towbin, H. T., Staehelin, T., and Gordon, J. (1970) P m . N d . 20. Klocswiak, M., Timmons, S., Lukw, T. J., and Hawiger, J.
14. Mamyma* K** Mikawa* T.* and s. (lga) J . Biochem. 21. Hawiger, J., Timmons, S., Strong, D. D., Cottrell, B. A., Riley,
A. 80,7318-7322
Acad. Sci. U. S. A. 76,4350-4354
Bwplymers 22,2459-2476
(1984) Biochemistry 23,1767-1774
M., and Doolittle, R. F. (1982) Biochemistry 21, 1407-1413
3544-3549
(Tokyo) 95,511-519 15. Doolittle, R. F.9 Cwsmen, K. G., COttRll, B. A*, Friezner, s. J-9 22. Olexa, s. A., and Budzynski, A. Z. (1981) J, BWL chm. 256,
and Takagi, T. (1977) Biochemistry 16, 1710-1715 16. Horrocks, W. D., Jr., and Collier, W. E. (1981) J. Am. Chem. SOC.
17. Murano, G., Wiman, B., Blomback, M., and Blomback, B. (1971) 24- Henwhen, A*, LottaWich, F-7 Kehl, M-9 and Southan, c. (1983) 18. Cohen, S. A., Tarvin, T. L., and Bidlingmeyer, B. A. (1984) Am. 25. Kretainger, R. H., andNockolda, C. E. (1973) J. BWL Chem. 248,
103,2856-2862 23. Chen, R., and Doolittle, R. F. (1971) Biochemistry 10,4486-4491
FEBS h t t . 14,3741 Ann. N. Y. Acad. Sci. 408, 28-43
Lab. 16.49-59 3313-3326