Embed Size (px)
Citation preview

lNF ORMATION Tb USERS
This manusaipt has been nproduced fmm the miaofwm master. UMI films the
text directly from the original or copy submitted. Thus, some thesis and
dissertation copies are in typewriter face, Mile mers may be fmm any type of
cornputer printer.
The qwlity of this nproduction is âepenâont upon the qurlity d the copy
submitîed. Broken or indistinct print, coloced or poor quality illustrations and
photographs, print bkdthmugh, substanâard margins, and improper alignment
can adversely affect reproduction.
In the unlikely event that the auüior ad not send UMI a complete manuscript and
there are missing pages, Cties8 will be noted. AISO. if unauüiorited copyright
material had to be mmoved, a note will indicate the deletion.
Ovenize materials (e.0.. maps, drawings, &arts) are reproduced by monhg
the original, beginning at the upper left-hand corner and continuing frwn left to
fight in equal sections wiîh m a l overlaps.
Photographs induded in the original manusaipt have been reproduœd
xerographicaily in this copy. Higher qwl-Q 6' x 9. black and white photographie
pcints are availirble for any phaographs or illusbations appearing in this copy for
an additional charge. Contact UMI diractly to order.
Bell & Howell Information and Leaming 300 North teeb Road, Ann Amr, MI 481061346 USA


DIETARY PROTEIN DEFICIENCY MODIFIES
SYSTEMIC AND GUT-ASSOCIATED lMMUNE
RESPONSES IN MICE INFECTED WITH
HELJGMOSOMOIDES POLYGYRUS (NEMATODA)
REBECCA YAT LOO ING
Institute of Parasitology & S c h d of Dietetics and Human Nutrition
McGiil University, Montreal Quebec, Canada
A thesis submitted to the Faculty of Graduate Studies and Research in partial fuVilment of the reqoirements for a joint degree of Master of
Science Crom the Institute of Parasitology & S c h d of Dietetics and Human Nutrition
@ Rebecca Y.L. h g October 1998

National Library Bibliothèque nationale du Canada
Acquisitions and Acquisitions et Bibliographie Sewices services bibliographiques
395 Wellington Street 395. rue Wellington Ottawa ON K1A ON4 Ottawa ON K I A ON4 Canada Canada
The author has granted a non- L'auteur a accordé une licence non exclusive licence allowing the exclusive permettant ii la National Library of Canada to Bibliothèque nationale du Canada de reproduce, loan, dismbute or sel1 reproduire, prêter, distribuer ou copies of this thesis in microforni, vendre des copies de cette thèse sous paper or electronic formats. la forme de microfiche/film, de
reproduction sur papier ou sur format électronique.
The author retains ownership of the L'auteur conserve la propriété du copyright in this thesis. Neither the droit d'auteur qui protège cette thèse. thesis nor substantial extracts ffom it Ni la thèse ni des extraits substantiels may be printed or otherwise de celle-ci ne doivent être imprimés reproduced without the author's ou autrement reproduits sans son permission. autorisation.

ABSTRACT
Protein deficiency may increase susceptibility to gastrointestinal (GI) parasitic
infections. possibly as a result of impaired systemic andor intestinal effector responses
induced by downreguhtion of Th2 cytokines andor upreguiation of Th1 cytokines. To test
this hypothesis. femaie B ALBlc mice (n= 1 lldiet) were fed a control(24%), marginal (7%).
or deficient (3%) protein diet and given a challenge infection with the GI nemûtode,
Heligmsomoides polygyrus. The 3% mice had higher wom burdens at 1.2 and 4 weeks
post-challenge uifectio n (pci). 10 wer increases in serum IgE, reduced intestinal eosino philia,
and depressed mucosd mast ceii proliferation and activation at 1 to 2 weeks pci To
determine wkther these suppressed effector responses in the 3% rnice were usociated with
altered spleen and mesenteric lymph node (MLN) cytokine profiles. ceh were restimuiated
in vitro with parasite mtigen and cytokine concentrations were measured. Deficient MLN
ceUs secreted significantly less IL4 and more IFN-y at 1-2 weeks pci thui did control
MLN ceB. Deficient spleen cek &O secreted more I M - y at 2 weeks pci compared with
convol spleen cells. From RT-PCR analyses, the 3% rnice &O had lower IL-4 mRNA
expression in spleen and MLN at 1-2 weeks pci Our study supports the hypothesis that
protein deficiency exacerbates the sudval of a GI nematode parasite by decreasing IL-4
(Th.2) and increasing IFN-y ml) emly in the infection, leading to reduced gut and
systemic Th2 effector responses.

La carence benta i re protidique a démontri! son influence à la prédisposition aux
infections parasitaires gûstrointestinales chez ia souris. Une relation possiblement due à une
d&&ioration de l'action des effecteurs gbnéraw et/ou intestinaux provoqude par une
régdation à la baisse des cytokines Th2 et/ou d'une régulation B la hausse des cytokines '
Thl. Pour tester cette hypotMse, des souris BALBIc (n=l8/di&e) ont tt6 nourri selon trois
types de dietes protidiques diffttrentes: un groupe ternoin (24%)' marghd (7%) et deticient
(3%). De plus, ces souris &aient infestées par le nematode Heligmosomoides polygyms
selon un schéma de premi2re infection suivie d'une réinfection. Le 3 8 groupe avait un
taux d'infection plus elevé à 1,2 et 4 semaines après la réinfection. les augmentations des
taux d'IgE les plus bas dans le serurn, une 6osinopM&nie intestinale, ainsi qu'une
prolifhtion et une activation reduites de mastocytes de h muqueuse à L et 2 semaines
post-réinfection. Pour déterminer si cette diminution de i'activite des effecteurs d m le
3% groupe est le résultat d'un profde dt6rr5 de la rate et des cytokines du GLM, des
cellules ont et6 stirnuEes in vitro avec de l'antigène de parasite. Les cellules du GLM du
groupe déficient ont sécréters beaucoup moins d'IL-4 et d'IFN-y durant les 2 semaines
suivant la réinfection que le groupe de cellules du GLM du groupe ternoin. Les cellules de
la rate ont eues aussi sdcrétées plus d'IFN-y durant les 2 semaines aprés la réinfection en
comparaison aux cellules du groupe tdmoin. D'après des analyses au PCR. le groupe DP
n'exprimait pas aussi bien IL4 ARN messager de k rate et du GLM B 1 ou 2 semaines
post-réinfection. Nos dsultats sou tiennent l'hypo these que la carence protidique exacerbe
la survie d'un dmatode gastrointestiniil en diminuant IL-4 (Th2) et en augmentant CFN-y
(Thl), qui m8nent une diminution de l'activitd des effecteurs Th2 intestinaux et g6n6nux.

ACKNOWLEDGEMENTS
1 thank my supervisors, Dr. Marilyn Scott and Dr. Kristine Koski for their insights,
guidance and expertise which were instrumental to the success of this research project. My
work hm k e n greatly facilitated by their continuous support. enthusiasm and ability to
embrace new concepts and directions. Most irnportantly, I am grateful to them for giving
me the unique opportunity to study both pansitology and nutrition despite my lack of
previous experience in either discipline. 1 also thank Dr. Linda Wykes for her contribution
to my understanding of protein metabolism. I thank McGiU University for the Max Stern
Recruitment Fellowship and the Christine Gagnon Mernorial Scholuship. I also am
gratefûl to rny supervisors for their fmancial support in the last months of my studies.
1 gratefully acknowledge the contributions of my kbmates toward my resevch
project. My deepest appreciation to Fananeh Jalili for her worm counts and assistance
with the infections and mouse harvesting. 1 thank Dr. Zhong Su for performing the
histological work and RT-PCR experiments and for s h h g his immense expertise on
pansite immunology. I will remember fondly the many hugh and beers we shmd
together and especially our late night conversations in the lab. By helping me feed the mice
and enter data, Mike Clark engaged me in lively discussions as weii as spued me many
hours of shvery and monotony. 1 am indebted to Dr. Hai Ning Shi and Sarah Conly for
teaching me the ce1 culture and ELISA procedures. 1 th& Christine Gagnon for her
many words of wisdorn and encouragement and for inspiring in dl of us an appreciation of
science and Ne.
1 th& Gordon Bingham for his tireless and impeccable care of my mice and the
animal facilities. My thousand th& to Shawn Mohammed and Darren Campbell for
helping me troubleshoot problems with the imrnunological assays and for our afternoon
coffee breaks. 1 am gratehil to Christhne Trudeau for her wterfbl French translation of
my absuact. 1 thank Dr. K Chadee, Dr. G. Ruben, Dr. C. Tanner, Dr. J. Smith, Dr. M.
Stevenson. and Dr. S. Mahanty for their ~sistuice and advice during my studies. Also, 1
thYiL dI the students and techniciam at P d t o l o g y and Nutrition for shûring theu
resources and for their fiiendship which Lifted my spirits on the long, tough resevch days.

Thank you Dr. CC. Miny for seeing me through the years it took to me to become
an independent person and competent scientist. Above ail, thank you for showing me the
many wondehl things to iive for beyond acdemics and for your invaluable friendship.
I thank my f d y for their unwavering moral and hancial support which has
sustabled me throughout my academic endeavors. I dedicate this thesis to my parents who
selflessly sacrificed their own professional and personal lives to give their children a better
Me full of endles posibilities. I cm only hope that m y accomplûhments are worthy of
their past hardships. pride and enduring belief in my abilities.
Listly but imrnensely, 1 also dedicate this thesis to J.P. Paiement. Thuik you for
everythmg that you are and more. Elephant shoe.

TABLE OF CONTENTS
TABLE OF CONTENTS . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . v
LISTOFFIGURES ................................................... viii
LIST OF TABLES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ix
THESIS OFFICE STATEMENT . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . x
STATEMENT OF CONTRIBUTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . xi
LIST OF ABBREVIATIONS . . . . . . . . . . . . . . . . . . . . . . . , . . . . . . . . . . . . . . . . . . . . xü
CHAPTER II. LITERATURE REVZEW . . . . . . . . . . . . . . . . . . . . . . . . . , . . . . . . . . . . 5
1. Host Immunity to Nematode Parasites . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5 1.1 Th1 Versus Th2 Response Phenotype as Medhted by Il-4 and IFN-y . . . . . 7 1.2 Th1 Versus Th2 Ceil Differentirition .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9 1.3 Roleofni2Cytokines . . . . . . . . .. .. .. . . .. .. . . .. . . . . . . ... .. . . . . 12 1.4 Roie of Th1 Cytokines . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15 1.5 Overview of Gut-Associated Mucosal Immune Responses . . . . . . . . . . . . 16
2. The Heligmosomoides polygym-mouse Mode1 . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 1 2.1 Host Immunity to Primyy Infection with H. polygyrus . . . . . . . . . . . . . . . 2 1 2.2 Systemic Versus Gut Mucosai Immunity to Primary Mection with H.
polygyrus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26 2.3 Host lmmunity to Chdknge Infection with H. po lygyw . . . . . . . . . . . . . 30
3. Effect of Protein Dekiency on HeImtoth Infections . . . . . . . . . . . . . . . . . . . . . . . . 34 3.1 Effect of Protein Dekiency on Primaxy Infection with H. polygynrr . . . . . 34 3.2 Effect of Protein Defkiency on Challenge Mection with H. polygynrr . . . 36
4. Effect of Protein Deficiency on Systemic CeiI-Mediated and Humorai Immunity . . 38

TABLE OF CONTENTS continued
5 . Effect of Rotein Dekiency on Gut Mucosal Immunity ..................... 42
6 . Effect of Protein Deficiency on Immune Responses to Intestinal Nematodes Including ..................................................... H.polygyms 45
....................................................... . 8 References 49
CHAPTER iII . STIJDY OBJECTIVES ................................... 69
CHAPTER IV . DIETARY PROTEIN DEFICIENCY PROLONGS S U R W A L OF A GASTROINTESTINAL PARASITE IN MTCE BY SUPPRESSING TH2 MMUNITY ......................................................... 70
INTRODUCTION ............................................... 72
.................................... MATERIALS AND METHODS 74 ........................................ Experimental Design 74
Experimental Protocol ..................................... - 7 4 Mice .................................................... 76 Diets ................................................... 77 Parasite and Parasite Antigen ................................. 77 Puasite Wonn Burdens ..................................... 79
....................................... Nutritionai Mesures 79 ELISAs for IgE. parasite-specinc IgGl and MMC protease- 1 ......... 79
..................... Intestinai Mast Cell and Eosinophil Histology 81 Spleen Ceii and Mesenteric Lymph Node CeIl Prepmtion ........... 81 Production of Th2 (IL.4. IL.5. IL- 10) and Th1 (IFN-y) Cytokines ..... 81 Cytokine mRNA Expression by Semi-Quantitative RT-PCR .......... 83
......................................... Statisticd Analyses 84
..................................................... RESULTS 85 ......... ............................ Nutritional Outcornes .. 85
Parasite Outcornes ..... .. .................................. 87 ................................... Senun EEector Responses 87
Intestinal Effector Respoases ................................. 95 Systemic and Local Cytokine Responses to Parasite Antigen in vivo .... 98 Systemic and Local Cytokine mRNA Expression ................. 102

TABLE OF CONTENTS continued
DISCUSSION ................................................. 105
ACKNOWL, EDGEMENTS ....................................... I l 8
REFERENCES ................................................ 119
CHAPTER IV . GENERAL DISCUSSION ................................ 125
APPENDICES ...................................................... 128
APPENDIX A Resuits from Two-Way ANOVA on EEects of Diet. Tirne . & Diet by Time Meraction .............................................. 128
APPENDIX B Results from One-Way ANOVA on Main Effect of Diet ............ 131
APPENDIX C
........... Results from One-Way ANOVA on Main Effect of Time 136
vii

LIST OF FIGURES
CHAPTER 11, LITERATURE REVIEW
Figure 1: Th2 And Th1 Immune Response Phenotypes .......................... 6
Figure 2: DBerentiation of CD4+ T CeUs into Thl or Th2 ........................ 8
Figure 3: Mucosal Immunocpe Tnfnf ..................................... 18
CHAPTER III . DIETARY PROTEIN DEFICIENCY M O D E S TH2 IMMUNE RESPONSES IN MICE TO A GASTROINTESTINAL PARASITIC INFECTION WITH HELIGMOSOMOIDES POLYGYR US (NEMATODA)
Figure 1: Schematic Representation of Experimentai Protocol .................... 75
Figure 2: Effects of Dietary Protein Deficiency and Tirne on W o m Burdens ......... 88
... Figure 3: Effects of Dietary Protein Deficiency and Time on S e m Anu'body Lrvels 90
............ Figure 4: Effects of Dietary Protein Deficiency on Eosinophdh in Blood 93
Figure 5: Effects of Dietary Protein Deficiency and Time on MMC Counts. Intestinal Eosinophil Numbers. and MMCP- 1 Concentration ..................... 96
Figure 6: Effects of Dietary Protein Deficiency on ni2 and Thl Production .......... 99
.... Figure 7: Effects of Dietary Protein Deficiency on Th2 and Thl Cytokine mRNA 103

LIST OF TABLES
CHAPTER II. LITERATURE REVIEW
Tabie 1: Kinetics ofIn2 Cytokine and Effector Production Post-prim;isr and Post-chdlenge .......................... Infection with H. p o l y g y w in BALBk Mice 28
CHAPTER III. DIETARY PROTEIN DEFICIENCY MODIFTES TH2 IMMUNE RESPONSES IN MICE TO A GASTROWSTINAL PARASITIC INFECTION VATH HELJGMOSOMOIDES POLYGYRUS (NEMATODA)
.......................... Table 1: Nutrient Composition of Experirnental Diets 78
Table 2: The Effect of Protein Deficiency on Nutritional Parameters of Challenged Mice KüledonDay28pci . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8 6

THESIS OFFICE STATEMENT
In accordance with the reguiations of the Faculty of Graduate Studies and Research of
McGiIl University, the following statement is included in this thesis:
"Candidates have the option of including, as part of the thesis. the text of a paper(s)
submitted or to be submitted for publication. or the cleuly-duplicated text o h published
paper(s). These texts must be bound as an integral part of the thesis.
' n i e thesis must stiU conforrn to ail other requirements of the "Guidelines for
Thesis Preparation". The thesis must include: A Table of Contents. an abstract in English
and French, an introduction which clevly States the rationale and objectives of the study, a
comprehensive review of the litenture. a final conclusion and surnrnary and a thorough
bibliognphy or referencc üst.
Additional material must be provided where appropriate (e.g. in appendices) and in
sufficient detail to allow a clev and precise judgernent to be made of the importance and
oiginality of the resevch reported in the thesis.
"In case of mmuscripts CO-authored by the candidate and others. the candidate is
required to make an expücit statement in the thesis as to who contributeci to such
wodc and to what extent. Supervison must attest to the accuncy of such statements at
the doctoral oral defense. Since the task of the examiners is made more dinicult in these
cases, it is in the candidate's b a t intrrest to make perkctly clear the responsibilities of dl
the authors of the CO-authored papers. Under no circunirtances can a CO-author of any
component of such a thesis serve as an examiner for that thesis."

STATEMENT OF AUTHORSHIP
AU experimental work in this thesis was performed by Rebecca Y.L. Ing with the
exceptions of the wom counts, histological evaiuations, and RT-PCR assays. Dr. Zhong
Su performed the histology and RT-PCR experiments. This thesis was written by Rebecca
Y.L. hg. Dr. M.E. Scott and Dr. KG. Koski served as research/thesis supenrisors,
critiqued data presentation and andyses, and pre-edited aü written material. The authors
gratefitiiy acknowledge Fananeh .JaU for her work in counting the w o m burdens.

LIST OF ABBREVXATIONS
ADCC
ANOVA
MC
BSA
BUN
CM1
ConA
DNA
DTH
GALT
GM-CSF
HBSS
HEV
HPRT
IE
E L
IL
m-Y r, L I
LP
LPS
mAB
rnRNA
MLN
MHC
MMC
antibody-dependent cellular cytotoxicity
andysis of variance
antigen presenting ceii
bovine serum dbumin
blood urea nitrogen
concanavalin A
deoxyribonucleic acid
dehyed type hypersensitivity
gut-associated lymphoid tissue
granulocyte-macrophage CO lony stimuiating factor
Hank's Balanced Salt Solution
high endothelid venule
hypoxanthine phosphoribosyluansferase
intraepithelium
intraepithelial lymphocytes
interferon-gamma
third-stage W m
fourth-stage lmae
lipo polysaccharide
monoclonal mtibody
messenger riionucleic acid
major histocompatibüity complex
mucosd mut ceU

LIST OF ABBREVIATIONS continued
MMCP- I
NK
PBS
pci
PHA
pi
P P ~ PWM
RNA
RT-PCR
SRBC
Th
Th1
Th2
TCR
TNF-a
mucosd m u t cell protease- l
naturai Mer
phosphate bugered saline
post-challenge section
phyto hemagglu tinin
post-infection
post-prirnary infection
pokeweed mitogen
ribonudeic acid
reverse transcriptase - polyrnerase chah reriction
sheep red blood ceil
T helper
T helper type 1
T helper type 2
T cell receptor
tumour necrosis factor - alpha

CHAPTER 1 INTRODUCTION
Protein malnutrition and helminth infections of the human gastrointestind tract are
among the most cornmon causes of morbidity and mortality worldwide (WHO. 1987). The
coexistence of enteric helminth infections with dietary deficiencies, particularly in
populations of devdoping couniries, has been weU-documented in epidemiological and
clinid studies (Crompton, 1986). Although the prevailing socioeconornic and
environmental conditions (e. g. poor sanitation, inadequate preventive health care) may
predispose the individual to both malnutrition and parasitic infection, the complex
bidirectiond interactions between the two conditions may influence disease patterns and the
efficacy of nutritional repletion or anti-puasitic interventions (Bundy & Golden, 1987).
Malnutrition is known to depress immunocompetence in humans and experimentai animals
(Good & Lorenz, 1992; Kuvibidih et ai., 1993), leadhg to increased suscepuiility CO
parasitic infections. Aiternatively, intestinal parasites rnay influence host defenses ( tg.
villous atrophy and damage to the mucosai epithelial integrity) which in tum disrupt the
uptake and utilization of dietary nutrients (Upadhyay et al., 1985; Solomons, 1993).
Pansitic infections are also implicated in human w u @ and nutrient loss associated with
protein deficiency due to expenditure of resources for imrnunological processes rather thm
for tissue spthesis, to excessive intesthai blood loss and to the lealcage of p h m a proteins
across the damaged gut epitheiial barrier (Lunn & Northrop-CIewes, 1993). Thus, host
immune function may be one mechankm underlying the mutually aggravating interactions
of mdnuuition and helminth infections. Given the possible reiationships in the
malnuuition-uifection-immunity complex, this project was h e d at determining the effect
of protein mainuvition on host immune response to intestinal nematode parasites.
Protein deficiency, in conjunction with inadequate energy intake, h a k e n
associated with profound growth retardation (stunting). metabolic dysfunctions. increased
child mortality, and greater prevaience of intestinal parasitic mfections (WHO, 1987;
Latham, 1990; Kuvii idf et al, 1993; Nesheim, 1993). Epidemiological studies have
suggested a strong association between protein malnutrition and immune function

(Chandra, 1983). however, the evaluation of causal rehtionships in the human setting is
cornpiicated by the presence of CO-existing confounding variables which make it dificult to
establish whether the impaired immune function is a direct result of protein deficiency or
whether it is secondary to the presence of concomitant infection, unrehted diseases, other
nutritional deficiencies, or environmental factors. Despite the i n t ~ s i c interprerative
problerns in epidemiologic data, the conclusions from human studies have k e n supported
by experiments showing that protein deficiency depressed various indices of immune
function in uninfected mimals (e.g. Conzen & Janeway, 1988; Larnont et al., 1988; McGee
& McMurruy, 1988; Woodward et aL, 1995) and increased the transmission d o r
s u ~ v a l of Nippostrongylus brasiliensis. Trichuris muris , and Heligmosornoides polygyrus
in rodents (Duncombe et al.. 1981; Slater, 1988; Keyrner & Tarlton, 1991; Michael &
Bundy. 1992a). However. the majority of these studies either did not involve parvitized
hosts when investigating the rob of protein nutrition in host irnrnunocompetence or did not
examine imrnunologicai rnechanisrns relevant to the control of helminthiasis. Therefore,
rnuch is unknown about the effect of protein malnutrition in host-pansite interactions.
Functional immunity to nematode parasites depends on specific cytokine
production by type 2, T helper cells (Th2) which induce and mediate antibody-dependent
effector responses involving IgE. IgG 1, mucosd mast ce& (MMC) and eosinophils
(MOMO y & E ~ q u e z , 1992; Nredhm & Lillywhite, 1994). Yet. ody a few studies have
investigated the effect of dietary protein deficiency on Th2-type host immune responses to
intestinal nematode parasites (Shter & Keymer, 1988; Boulay et al.. 1998) and therefore it
is currently unclear whether protein deficiency potenthtes intestinal parasitic inf'ections by
impairing the development and maintenance of specific T ceU-dependent humorai immune
responses. In addition. the severe Ieveis of protein restriction (e.g. 0.5%) used in studies on
protein malnutrition and immune function (q. Woodward et al., 1992a & 1992b) limit the
generahbility of the results to human populations in which müd to moderate malnutrition
is much more prevhnt than profound nutrient deficiencies (Kuvibidh et al., 1993). Few
researchen have evaluated comprehensively the relative impact of mild to moderate protein
deficiency on protective immunity to nematode parasites m a dynamic host-parasite system.

Also, there is Utle information regardhg the interactions of host immunity and pro tein
malnutrition during secondary pansitic infections despite accumukted evidence that the
magnitude md efficacy of immune responses are innuenced by the frequency of antigen
exposure. In experiments using repeated or challenge inlection protocols that stimulate
protective immunity in normally immunocompetent hosu, pro tein deficiency (2-346) hu
k e n shown to prolong H. po lygym s u ~ v a l and to impair the production of secondary
total IgG 1 and eosinophil responses (Slater & Keymer. 1986; Keymer & Tarlton, 199 1;
Bouiay et al.. 1998). To date. the imrnunologicd parameters examined in these studies and
in most previous research on host immunity to nematode parasites hm ken confined to the
effector mechanisms in systemic cornputmenu such as the penpherd circuiation and
spleen. As a result, we know less about the nutritional deterrninants of gut mucosal
immunity despite reports that the gut mucosa hm one of the highest protein turnover rates
of ail tissues in the body (Nakshabendi et al., 1995) and is potenMy very sensitive to
changes in host nutritional status (Wykes et al., 1996). Research on gut mucosai immunity
is essential to the understanding of host mûlnutrition-parade interactions because the gut
and its associated lymphoid tissues comprise the fvst site of contact between host immune
de fenses and invasive intestinal nemntodes. Recent studies have s ho wn thrt the expression
of Th2 immunity in intestinal lymphoid tissues may determine the s u ~ v a l of H. polygyrus
during prirnary infection in mice (Behnke et a l , 1993; Fuiklemm et al., 1997; Wahid et ai..
1994). The putative importance of gut mucosal immunity in the resolution of nernatode
infection and its potentiai for nutritiond modulation are persuasive reasons for evaluathg
intestinal as weli as systemic immune processes in ordet to M y ascertain the mech3nisms
underlying the rektionships between protein deficiency and intestinal parasitism
The overall objective of this research project w u to detennine the systemic and
intestinal Th2 immune responses by which protein deficiency may influence the duration
and severity of an intestinai nematode infection. To better understand the cornplex
interrelationships of protein deficiency. host immunity and intesthaî parasitism, this study
integnted ail three conditions using an experimentd H. poIygyncs-mouse system as a mode1
for gastrointestmal helminthiasis and expression of Th2-type immune responses. Mice were

fed varying ievels of dietuy protein and subsequently subjected to a challenge infection
pro toc01 that has k e n shown to stimulate acquired resistance to secondary infections with
polygyms in well-nourished hosts (Behnke & Robinson. 1985). We hypothesized that
pro tein deficiency would reduce the magnitude of intestinal a d o r systemic cytokine and
effector responses duMg challenge infection with H. polygyms. Also. we expected that the
level of diet-induced impairment of host pro tective Unmunity would be proportional to the
severity of protein restriction such that a Iow protein diet (e.g. 3%) would exacerbate
parasite survival by inhiiiting acquired imrnunity to a greater extent th;ui would a
margindy deficient diet (e.g. 7%). We had anticipated that a certain degree of
physiological adaptation to maintain homeostasis and hinctionality might occur with mild to
moderate malnutrition but that there would be a threshold of nutritional deficiency beyond
which imrnunocompetence would be notably impaired. Assuming that gut mucosal
immunity is important for the rapid termination of a nematode infection and for prirning
systemic irnmunity. we postulated that protein deficiency would cause a more profound
impairment of acquired immunity at the intestinal ievel than that at the systemic leveL
Fidy. we speculated that protein deficiency would impact cytokine-medisited effector
mechanisrns to vuyhg intensities at different phases of a nematode infection. To determine
this possible effect of protein deficiency on the kinetics of Th2 immune responses, we
conducted a tirne course study of various effector and cytokine outcomes zt multipk time
points afier prirnafy and challenge kifection with H. polygynrs. Addresshg these
hypotheses would dvance out understanding of the mechaniîrns of parasite survhal in
protein malnourished populations and thereby help d e b e immunoIogicaI or nutritional
targets for therapeutic interventions.

CHAPTER 11 LITERATURE REVIEW
1. HOST IMMUNITY TO NEMATODE PARASITES
In general, immune responses in mice to foreign antigens involve two categories of
T helper (Th) cek: type 1 Th ("M) c e k are responsible for celi-mediated immunity (CMI)
and type 2 Th (TU) cek mediate antibody-dependent immunity. Each Th ceii response
phenotype is characterized by distinct patterns of cytokine and d u n e effector production
that correhte weil with their irnmunologic specincity (see Figure 1). Presentation of
parasite-denved antigens by antigen-presenting celis (APC) to CD4' T c e h induces the
dinerentiation of Th ce& to the Th2 functional phenotype. Th2 ceUs secrete interleukin
(IL,)-4, L-5. IL-9 and IL-10 which promote the prolifention and activation oCTh2-
associated effectors such as IgE- or IgG1-secreting plasma ceiis, eosinophils. and mucosal
m a t ce& (MMC). These efkctors are features of IgE-mediated host defense mechanisms
against many helminth parasites (Cox & Liew, 1992). Altematively, Th1 ceUs produce L-2
and IFN-y and interact with APCs to synthesize IL-12. These Thl-associated cytokines
are important in macrophage activity and isotype selection for IgG2a. IgGZb and IgG3,
which mediate antibody-dependent celluhr cytotoxicity (ADCC) and complement activity
against bacteridor viral infections (Locksley, 1994). BothTh cell types secrete IL-3,
RIF-a, and GM-CSF (Mosmann & Sad, 1996). The pohrked patterns of cytokine and
immune ceil production correspond to activated effector Th phenotypes generated during
immune response to a particuiar antigen but are no t observed in nive CD4+ T ce& or even
long-tem. quiescent memory ce& (Mosmann & Sad, 1996; Bunce & Bell, 1997).
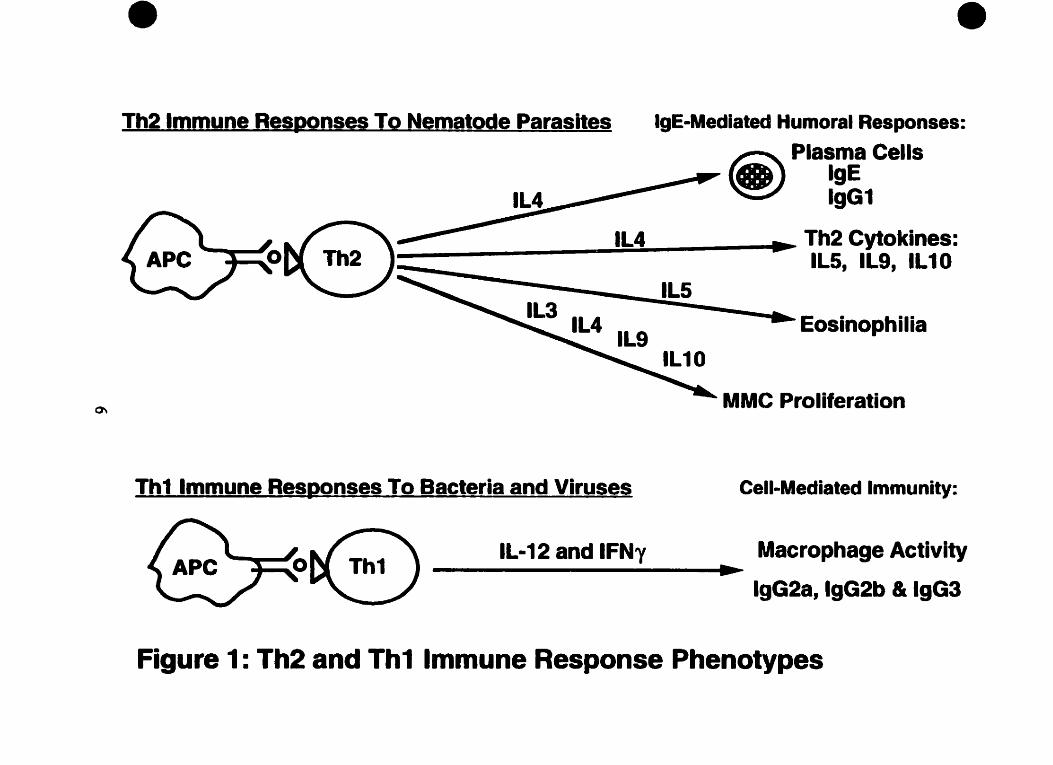
Th2 lmmune Responses To Nematode Parasites IgE-Medieted Humoral Responses:
Plasma Cells IgE lgG1
Th2 Cytokines: ILS, 119, IL10
Eosinophilia
a MMC Proliferation
Th1 Immune Res~onses To Bacteria and Viruses Cell-Mediated tmmunity:
IL-1 2 and IFNy Macrophage Activity
lgG2a, lgG2b & 1963
Figure 1 : Th2 and Th1 lmmune Response Phenotypes

1.1 Th1 Venus Th2 Response Phenotrpe as Mediated by Ii-4 and IFN-y
The characteristic cytokine products of T'hl and Th2 cells are mutudy mtagonistic
for the differentiation and activity of effectors belonging to the reciprocal phenotype (see
Figure 2). Hence, L-4 and in some circumstances 5 1 0 inhibit cytokine synthesis by Th1
cells and macrophage hinction (Kopf et ai., 1993; Lagoo et d., 1994; Mosmann & Sad,
1996) whereas IFN-y suppresses the production of ThL-type effectors and cytokines (King
et aL, 1993; Urban et ai., 1993). This cross-regulation oCTh ceU polarization and immune
responses may explain the svong bias towvds Th2 responses during nematode infections
and why the alteration of Th2 response patterns by IFN-y or other Th1 cytokines (e.g. IL-
12) reverses host resistance to nematode infection (see UrSm et al.. 1993; Fiiemm et aï.,
1994). In humans infected with helminth parasites, serum IgE and IgGl be ls are directly
proportional to parasite-induced IL4 production and inversely related to IFN-y synthesis
(King et al., 1993; King & Nutman, 1993) whereas IgE-medhted eosinophil and mast ce1
activities are associateci with eflective resolution of helminth infections (Men & Maizels.
1996). Experimentai evidence to support the view thrt development of a Th2 cytokine
response stimuiates strong resistance to nematode puasites hm ken denved f?om studies
that measured the cytokine profile of cells isohted from tissues of genetically resistant
animais (Wahid et al., 1994). conducted adoptive tramfer of protection with immune c e h
known to secrete Th2 cpokines in vino (Korenaga et of.. 1989; Ramaswamy et al..
1994a). andyzed knockout animais (Kopf et al., 1993), used neutralizllig antibodies to Th2
cytokines ( C o h a n et al., 1989; Urbm et al.. 199 1 b), and andyzed the effects of different
cytokines on the NI vitro development of Thl and Th2 celi hes (e.g. Maggi et ai-. 1992).
Collectively, observations from these studies suggest that hnctional immunity to helrninths
involves Th2-type cytokines and effectors and that the reciprocal amounu of parasite-
elicited IL4 and LFN-y produced determine the magnitude of Th2 irnrnunity raised against
the nematode pansites.

11-1 2 IL4 O
1FN-y Q t
Humoral CelCMediated lmmunity (viruses, bacteria)
Immunity (Nematodes)
Figure 2: Differentiation of CD4+ T helper cells into Th1 or Th2 type: cross=regulation by 11-4 and IFNy. Adapated from Mosmann & Sad, 1996

1.2 Th1 Versus Th2 Ce11 Differentiation
The source and p ~ c i p a i early events affecting Th 1Khî cell dinerentiation
ultimately influence the type, speed and magnitude of the host immune response to
nematode parasites. Naive CD4' T cells and even some subsets of rnemory Th cells are
uncommitted and have the potentid to secrete cytokines belonging to both phenotypes.
When stimulated by antigen presented on APCs, these Th0 ceh secrete primarily IL-2 as
weU as smdl amounts of other cytokines, including IL-12, IFN-y and IL-4. There is
agreement that IL- 12 and IFN-y direct differentiation to a Th1 phenotype (Seder et al.,
1991; Bradley et al., 1996) and IL-4 drives dif'ferentiation to a Th2 phenotype (Le Gros et
al.. 1990; S wain et al., 1990). Several studies suggest that IL- 12 is denved from
macrophages and IFN-y is secreted by NK cells in response to IL12 or by naive CD4' T
ceik independently of APC or IL- 12 (S wain. 1988). The initial source of IL-4 and factors
required for the genention of IL-4-producing precursors are less weil-dehed. Classical
APCs (e.g. macrophages, dendritic ce&) are not known to produce IL-4 while the
proportion of IL-éproducing T ceUs among naive T ceil populations is very smdi (Swah eet
al., 1988; Le Gros et al., 1990). In fact. naive CD4+ T celis fkeshly isoiated from murine
spleen do not secrete detectabk quantities of IL-4 until afier the cells are primed and
cestimulated with C o d or antibodies to the TCR (Swain et ai., 1988) or ifexogenous IL4
is added to the priming culture (Le Gros et al., 1990; Seder e t al., 1992). Mast cells and
eosinophils cm produce IL-4 (Srder et al.. 199 1; Moqbel et al.. 1995; Ndcajima et aL,
1996) and provide ceil contact signals required for IgE production by B ceils (Gauchat et
al., 1993). but these ceiis are not abundant in lymphoid tissues where T ce1 priming and
daerentiation occur upon fkst exposure to the mtigen. L-4 is ako produced by a minor
subpopulation of T cells. CD3TD4'NKl. l* T cells (Yoshimoto & Paul, 1994) but how
these celis become activated to secrete IL-4 is not known.

SimiIarly. the driving force for initial Th2 priming during nematode infection has not
been clevly elucidated. IL-4 mRNA expression duMg prirnary idection in mice with
Heligmosomoides polygyrus is restricted to CD4'. TCRdw ce&. occurs relatively Iate
compared to other Th2 cytokines, and is not preceded by elevations in IL-2 mRNA (Svetic
et al., 1993) as observed in some Th ceil transitions from Th0 to Th2 expression. Also. NK
T ceils with the markers, CD6L-low. CD44-high, CD45RB-low, fiom mice geneticdy
deficient in I L 4 receptor retain their capacity to produce IL4 durhg Nippostrongylus
brasiliensis infection (No ben-Trauth et al., 1997). suggesting diat the induction of Th2-
type responses to nernatode pansites does not require evly IL-4 or signaiing through the
IL-4 receptor. Conside~g that production of IL-4 by mast ceh, eosinophils or CD4' T
cells in response to nernatode parasites appeus to be a Lte event without an initial transient
Th0 phase dominated by IL-2 synthesis. other cytokines or cell types may be hvolved in
initial Th2 dinerentiation. A recent study has s h o w that IL-6 produced by APCs can
vigger the Th2 pathway by inducing naive CD4+ T cells to produce IL-4 which in turn
would act as an autocrine factor to potentiate the upreguktion of IL-4 gene expression
while inhibithg IFN-y production (Rincon et al.. 1997). The CD4' T ceUs are then
polarized to a Th2 phenotype using their own source of IL-4 or IL-4 secreted by
sirnultaneously activoted basophiis and eosinophils. It is possible that these IL-6-dependent
events leading to IL-dproducing Th2 tek occur during responses to nematode parasites
because Svetic et al. (1993) have reponed that non-T ceUs (CD4XD8') are iikely the evly
source for elevated expression of IL-3, IL-5 and IL-9 mRNA and C M + T ce& becorne the
hter source of these cytokines including IL-4. Although IL-6 was not analyzed by Svetic et
al. (1993) or other rehted studies on helminth immunology, it may be a patentid initial
factor iduencing the choice between a Th1 or Th2 response to nematode parasites.
The presence of an appropriate pattern of cytokine production is essentiai for the
development of beneficiai immune responses to intestinal helminths. Even though the
initial sources of IL4 remah uncenain, it is weii recognized that IL-4 is a crucial r e m t o r
of the cornmitment of CD4+ T ceUs to the production of I L 4 The inhibition of early IL4
production ushg antibodies to I L 4 and the IL-4 receptor is sufficient to promote the

dinerentiation of Th1 ceils by increasising IL-2 and IFN-y production by naive CD4+ T cells
(Seder et al., 1992; Tanaka et al., 1993). Consistent with these data. mice deficient in
Stat6 gene (the signal transducer and transcriptionai activator of IL-4) and infected with N.
brarilienris secreted lower amounts of Th2 cytokines (Takeda et aL . 19%) and showed
decreased production of IgE but increased levels of IgG2a. IgG2b and IgG3 which are
dependent on IFN-y (Shmioda et al., 1996). Moreover, blocking of IL4 induced the
development of memory effector Th 1 ceUs (Bradley et al. , 1995). Unlike IL-Cproducing
T cells which require priming and restimulation, naive Th cells secrete IL2 and IFN-y upon
fist antigenic stimulation even in absence of IL- 12 (Bradley et al., 1996). Taken together.
these hdings Mply that the pattern of Th1 cytokines appeus to be the default phenotype
in naive mice as weii as in nematode-infected rnice when IL-4 synthesis or activity is
resuicted. M o . it is clear that IL-4 is an absolute requirement for Th2 cell dinerentiation
and resultant Th2 immune responses because it selectively promotes the expansion of Th2-
committed cells via positive feedback and/or suppresses IFN- y synthesis. Multiple sources
of IL,-4 including CD4' T cek, APC. mast tek. eosinophils and their costirnulatory
molecules (e.g. signaling between B7.1 or B7.2 on B cells and CD28 or C'T'LA-4 on T
ceh; expression of CD40 ligand on activated Th ce&) are involved in induchg the
differentiation of TM cells and subsequent production of IL-4 (Lu et ai., 1994; Lu et al*.
1996; Greenwdd et al*. 1997). A selective impairment of these ceh a d o r of their contact
signais may dom-reguiate ni2 response andor concomitantly up-regulate Th1 responses,
resulting in an inappropriate or inadequate host immune response to nematode parasites.
Because immune effectors are controlled by T ceii-derived cytokines, the foUowing
pangraphs wül review briefly the imrnunologic roles of the major Th2 and Th1 cytokines.
It is apparent that each cytokine has pkiotropic effects on multiple types of lymphoid cek
and that there is mucb duplication of function mongst different cytokines.

1.3 Role of Th2 Cytokines
IL4 is involved in the activation and maintenance of most Th2 effector responses.
IL4 produced by Th2 ceh evly after antigenic stimulation drives ail subsequent immune
responses toward the Th2 phenotype by acting as an autocrine growth factor for
differentiated Th2 ceils (Mosmann 8 Sad, 1996). enhancing the production of other Th2
cytokines (Kopf et al.. 1993), and inhibiting IFN-y production by Th1 cells (Tanaka et al..
1993). Mice deficient in IL-4 receptor displayed markedly diminished IgE, I f i l , IL-5, and
IL- 10 responses to N. brmiliensis Uection (No ben-Trauth et al., 1997). indicating that
induction of Th2 responses to helminths is dependent upon signaling through the IL-4
receptor. iL-4 promotes isotypc: switching of membrane IgA-positive and IgM-positive B
ceiis to IgE- and IgGl-expressing c e b (Finkelman et ai.. 1988 b; Zhang et al.. 199 1). As
IgM4 B ceb are frequently found in peripherd blood, the IgM to IgE switch may account
for much of circukting IgE after nematode infection (Zhang et aL, 1991). Also. IL-4-
mediated switch of IgA to IgE-expressing B c e b may be important for protective mucosai
immunity given the abundance of IgA+ B ceils in gut lyrnphoid tissues such as Peyer's
patches (Cebn & Shroff. 1994) and the association of IgE with anthelrninthic resistance.
Katona et al. (1991) demonstrated that antibodies to the IL-4 protein and recepior
compietely inhiaited secondary IgE responses to H. polygynrr but did not affect the ability
of IgE-expressing B cek to daerentiate into IgE-secreting ce&. Taken together, these
data suggest two important aspects of IL-dinduced imrnunoglobulin responses: 1) IL-4 is
required by virgin B ceh to express the IgE and IgG 1 isotype but may not be necessuy for
the tenninal dinerentiation of these cells into IgE- and IgG1-secreting plasma cek; and 2)
IgE and IgG 1 responses generated in rnice chailenged with H. polygyrus are not derived
from memory B c e h that were primed to express IgE and IgGl during the initiai exposure
but from IgA* or IgM* B cells that were induced by IL4 to switch to IgE and IgG 1
expression during secondw exposure. IgE and IgG 1 in tum, when crossed-iinked by
pansite-denved antigen. trigger the degranuiation of mast cells and eosinophüs (Gounni et
oL, 1994; Chen & Enerback, 1994 & 1996). Aiso, IL-4 has been shown to enhance L-5-

mediated chernotaxis of eosinophils and to induce MMC hyperphia dong with I L 3 and
stem cell factor (Madden et al., 199 1 ; Rennick et al., 1995). IL-4 up-regulates the
expression of endothelial adhesion molecules (e.g.VCAM-1) on intestinal endothelial c e h
and therefore may be important in the reguhtion of effector ceU traficking and horning of
lymphocytes, eosinophils and basophils to intestinal mucosal sites (Chin et al., 199 1;
Haradken et al.. 1996; Schleimer et al., 1992). Similarly, IL-4 treatment in vivo has been
shown to enhance the uptake of injected IgE into the gut mucosa and lumen following
infection with Trichinella spirulis, possibly by upregulating the affuiity of 1% receptors on
MMC and eosinophils present in the infected intestinal tissues (Ramaswarny et al., 1994).
In addition to promoting the expansion and recruitment of Th2 effectors. IL-4 rnay be
cniciril for effector ceil activation and function. IL-4 stirnuiates and promotes IgE binding
to m a t cek and IgE-induced degrmuhion of mut ce& when sensitized by antigen.
possibly via control of mut ceii FcrRI expression (Banks & Coleman. 1996; Tom et al..
1996). Lastly, preliminary experiments by Urban et al. (1996) suggest that IL-4 amplines
smooth muscle contraction in the smail intestine through mast cell and leukoviene D4-
dependent mechvllsrns which could in tum expedite wom expulsion from the intestine.
The main activities of IL-5 are to promote the growth and terminal differentiation of
eosinophils, to attract eosinophils to sites of inflrimmation. and to enhance IgE- or IgG 1-
induced eosinophil degranulation (Co f i a n et al.. 1989; Kita et al.. 1992). These activities
are complemented by L-4 (e.g. via IgE or IgG switching and eosinophil recruitment) and
by IL- 10 (e.g. via IgG 1 switching). IL-5 is secreted by Th2 cek, basophils and eosinophils
afier IgE- and IgG1-induced activation. thus providing an autocrine pathway for locd
eosinophil activation (Dubucquoi et al., 1994). The production of I L 5 in T and NK ceils is
augmented by IL4 and inhîbited by IL- 12 (Warren et al*, 1995).
Preiiminq data suggest that IL-9 potenthtes IL-4induced synthesis of IgE and
IgGl by LPS-prirned or antigen-stimuiated B lymphocytes (Dugas et ul. 1993; Petit-Frere
et al., 1993), optimizes mucosal mastocytosis synergisticany with IL3 and I L 4 (Behnke et
aL, 1993; Huitner et al., 1990). and stimulates the uanscription of genes coding for mast
cell granules such as mouse mast cell protease-1 (MMCP-1) which is secreted specitically

by intestinal MMC (Eklund et ai., 1993). The role of IL-9 in mediating protective
immunity to nematode parasites has not ken studied as extensively as that of other Th2
cytokines, however, a recent paper has demonstrated that IL9 mRNA is expressed early
during Trichuris m r i s infection and its elevation Nt vivo is associated with enhanced
intesthal mastocposis, production of IgE and IgG1, and loss of the parasite from the
intestine (Faulkner et al., 1998). Collectively, these observations indicate thrt IL-9
contribu tes to host resistance to helminths by promoting IgE-dependent MMC responses.
A recognized roh OPIL-IO in mediiiting Th2 responses is its inhibition of Th1 ceil
differentiation via suppression of IFNy production by Thl-cornrnitted celis. IL-10 does
not act directly on Th1 ceils but impairs the capacity of splenic and peritoneal macrophages
to stimulate Th1 cytokine synthesis by Thl cells (Fiorentino et al., 1991). Ding et al.
(1993) showed that this effect is partly due to the ability of IL40 to suppress the activity of
B7.1 (CD80) costimukory molecule on macrophages while de Waal et al. (199 1) found
that IL40 down-regulated the expression of clûss II MHC on APCs that interact with
antigen-activated Thl c e k to induce cytokine synthesis. IL-10 contributes to mucosai
mastocytosis by promoting the growth of mast cell progenitors and expansion of later
colonies in the presence of I L 3 and IL-4 (I3eh.de et al., 1993; Thornpson-Snipes et of.,
199 1). Similar to the effect of IL-9, E- 10 induces the transcription of genes for MMCP- 1
and other serine proteases secreted by MMCs of nematode-infected mice (Ghildyal et al.,
1992 & 1993). IL-10, dong with IL4 and stem cell factor, induces the synthesis of
histamine in MMC (Bissonette et al., 1995). h o , IL40 may promote isotype switching to
IgG4 in humans (Lagoo et al., 1994); although human IgG4 is homologous to muMe
IgG 1, this particular function of IL- 10 ho not been shown with murine B lymphocytes.
IL-3 is a cytokine secreted by Tho. Th1 and ni2 tells (Mosmann & Sad, 1996). Ir
is a multi-colony stimulating factor for severai haematopoietic ceils and thek progenitors in
vitro. Tt-3 is an essentiai cofactor for the growth and maturation of MMC dong with I L 4
and stem ceil factor (Madden et al., 1991; Re-k et al., 1995) and its administration into
nematode-infected mice induces strong mucosal mastocytosis and successful expulsion of
the nematode Snongyloides ratti (Abe & Nawa, 1988; Abe et al., 1993).

1.4 Role of Th1 Cytokines
IFN-y is an important Th1 cytokine that serves primdy to activate natunl killer
ceh and mononuclear phagocytes (e-g. macrophages and neuuophils) which are
responsible for cytotoxic activity againsr tumor celis and microbes (Mosmann & Sad,
1996). Research suggests that IFN-y inhibits or dehys protective immunity to nematodes
by suppressing the growth and expansion of Th2 ce& via a dom-regdation of IL-4
(Maggi et ni., 1992; Urban et al., 1992), inhibithg production of IgGl (Fielman et ai.,
1988a), suppressing the ability of IL4 to elicit IgE switching and dinerentiation (King et
al.. 1993; Uchikawa et al. 1994). blocking eosinophüia (Urban et ai., 1993), decreasing
the TM-a-dependent cytotoxicity of MMC (Bissonette et al.. 1995). and delaying IgE
response and MMC hyperplasia Li N. brudiensis-infected rats (Urban et al.. 1993). These
inhibitory effects of IFN-y on Th2 effector responses have ken associated with prolonged
s u ~ v a l of intestinal nematode parasites such as N. brasiiiensis and T. muris in rodents
(Lirban et al., 1993; Eise et a l , 1994).
L I 2 is secreted by macrophages and B lymphocytes and potenthtes Thl
phenotypic expression by enhancing priming for IFN-y production by naive CD4' T ceUs
(Seder et al., 1993) and naturd killer ceils (Chan et al.. 199 1 ) . IL- 12 also induces Th1
dserentiation &er antigenic stimuhtion by enhncing IFN-y spthesis and inhibiting the
development of IL-4-producing Th c e h (Myietti et al.. 1993; Windhagen et al.. 1996),
even in the absence of the costirnuhtory receptor CD28 on T ceh (Chu et al., 1997). The
presence o f IL- 12 in vivo has k e n shown to reduce the suppressing effecu of IL,-4 on
IFN-y production by naive CD4+ T cells (Seder et al., 1993). These anti-Th2 effects of IL-
1 2 have ais0 been dernonstrated in vivo: IL- 1 2 inhibits IgE S ynthesis and corres ponding IL-
4 production by lymphocytes isolated from helminth-Sected patients (King et al.. 1995)
whüe administration of K- 1 2 to N. brasiliensis-infected rnice prolo nged w o m s u ~ v d ,
stimulated IFN-y production. inhibited IL-4. IL-5 and IL-9 mRNA expression, and
prevented increases in nematode-induced IgE. MMC and eosinophü responses ( F i e h a n
et aL, 1997).

1.5 Overview of Gut-Associatecl Mucosal Immune Responses
Much of Our knowledge about specinc immune responses to nematode infection is
based on analyses of antibody concentrations fiom blood and cytokine production by
splenocytes. The underlying assumptions in these studies are that peripheral levels of
effectors or cytokines refiect intestinal IeveB and that protective T cells exert their principal
hinction by releasing Th2 cytokines in gut mucosa where the parasite resides and
reproduces. The fxst assumption was deemed invalid by hdings of a study showing that
serum IgE concentration in T. spiralis-infected mice represented only 0.2 to 1.0% of total
amount of IgE produced at the peak of intestinal antibody response and that the vast
rnajority of IgE produced in vivo is directed to the small intestine and does not enter the
bloodstream (Negarao-Correa et al.. 1996). Therefore, to understand the bill capacity of
host resistance to intestind parasites, we need to e x d e immune responses occurring in
gut mucosai tissues in addition to those in systemic compartments.
The srnail intestine contains speciaüzed lymphoid tissues that are categorized h t o
two anatornically and hinctionally distinct cornpytments: 1) the affierent hb consists of
aggregated lymphoid ceils (Peyer's potches). isolated lymphatic follicies and mesenteric
lymph nodes (MLN), where APCs present antigen to naive T lymphocytes (Bogen et al.,
199 1; Laissue et al., 1993; Regoli et aL, 1994) and 2) the eKerent h b c o n t h the
inuaepithelium (IE) and lamina pro pria (LP) of the intestinal rnicrovilli, where antigen-
specifk effectors such as IgE, IgG1, MMC and eosinophils mediate their anthelminthic
reactions (Beagley & Elson, 1992). Recently, Saito et al. (1998) desctibed a new intestind
lymphoid cornpartment (cryptopatches) Iocated under the layer of epithelid cells lining the
gut which consisted of immature and dEerentiated intraepithew lymphocytes (EL).
Although progenitors of al l immune cells originate nom bone m m w , Saito et ai. (1998)
found that T ceii precursors in cryptopatches were capable of maturing in the gut without
passing through the thymus and thus provide evidence that the intestinal mucosa contains a
population of thymus-independent mature T cells bat may interact directly wïth enteric
antigem. Cokctively, these immunologicai compartments are kno wn as gut-associated

lymphoid tissues (GALT) which help ensure mucosal integrity and comprise the fkst line of
host protection against ingested antigens and intestinal pathogens. The function of the gut
mucosa includes nonspecitic defenses provided by the epithekl bYrier and mucus as weli
as specinc immune responses medited by gut-associated cytokines and effectors.
The continuous. nonrandom migration of lymphocytes between mucosal lymphoid
tissues and the bloodstream is an essential process in the development of immune responses
at the appropriate sites of infection anywhere in the body. Naive lymphocytes fkst interact
with antigens in the Peyer's patches and isolated lymphoid follicles and then hrther
dinerenthte and mature in the germinal centers of the lymphoid follicles (Guy-Grand et ai.,
1978). Thereaftet. the antigen-primed c e h Ieave these inductive sites of GALT via
mucosai lymphatics. coiIect in the thoracic duct and enter the peripheral circulation (see
Figure 3). From the blood, the lymphocytes migrate to systernic lymphoid organs such 3s
the spleen and peripheral lyrnph nodes where the lymphocytes continue to mature and
proliferate to become effector cells (e.g. secrete cytokines) andor memory ce& (Salmi &
JaIkanen, 1991). Early studies on lymphocyte rectculation proposed a common mucosal
immune system in which lymphocytes mignte freely among the gut, peripheral lymph
nodes. genitd tissues and the lung (McDermott & Bienestock. 1979). However, ment
papers suggest that lymphocytes of lung and gut ongin have distinct recirculation routes
and express different homing adhesion receptors that bind to high endothehl venules
(HEV) lining the mucosai lymphoid tissues (Joel& Chmana, 1985; Abitonbi et al., 1996).
For example. activated and memory gut-drrived lymphocytes preferentidly extravasate
nom the blood to intestinal effector sites by expressing integrin a4P7 and Lselection,
adhesion receptors which are much less expressed by lymphocytes recirculating through the
lung (Schweighoffer et al., 1993; Abitonbi et ai., 1996). niese homing molecules are
receptors for mucosal vascular addressin cell adhesion molecule (MAdCAM)-I whifh is a
vascular addressin found selectively on endothelia of the p t and associated lymphoid
tissues (Nakache et al.. 1989; Berlin et a l . 1993; Roberts 8 Kilshaw. 1993). Thus. most of
the differenthted lymphocytes that enter the effector sites of the GALT (LP, E) are likely
to have had prior contact and specinc activation with antigens located in the gut mucos&

Inductive Sites: Antigen Presentation
Effector Sites: IE and LP
+ Lymphatics Lymphatics
C Systemic MLN -W Thoracic - Circulation
Duct Spleen
PLN
6 Antigen Figure 3: Mucosal lmmunocyte Traff ic @ B ceii
\ IgE or IgGl

In addition to the intestinal migration of lymphocytes, MMC progenitors, basophils,
and circulating eosinophüs are attracted by chernotaxis induced by gut-associated cytokines
to the IE and LP where they mature and degranulate after cross-linkllig of their FceRI
receptors by IgE ruid pansite antigens (Pewce et al.. 1982; Smith & Weis, 1996). MMC
in the gut mucosa or MLN are histochernicdy and functionally dBerent Erom masr cells of
the peritoneal cavity ûnd connective tissues (Shanahan et al., 1986; Befùs et al., 1987) and
are the predomuiant type of mast cells that proliferates in response to parasitic infections of
the gastrointestind tract (Arizono et a l , 1993 & 1994). Infection with N. brusiliemis
induces migration of MMC precursors from peripherd blood to the intestine (Kasugai et
al., 1995), development of MMC cells located Ni situ (Miller & Jarrett, 1971), and release
of serine proteases that may alter intestinai mucosal pemerbility (King & Miller, 1984;
Wasthg et a l , 1997). Mucosal mastocytosis is dependent on T cell-denved IL-3, IL-4 and
IL10 (Haig et ai., 1982; Madden er al., 199 1; ReMick et al., 1995). In tuni. MMC
produce Th2 cytokines (e-g. IL-4) and provide contact signals for IgE synthesis which help
augment local Th2 reactions (PIaut et al., 1989; Seder et al., 1991; Gauchat et al., 1993).
Cytokines such as IL-4, IFN-y and TNF-a rnay enhance the adhesiveness of
endothew ligands on HEV ceils for lymphocytes (ThornhiU et ai., 1990 & 199 1; Chin et
QL, 1991). Previous studies have docurnented the synthesis of Th2 cytokines by
lymphocytes derived fiom Peyer's patches and MLN (Svetic et aL. 1993; Wahid et al.,
1994) as weii as the GALT-specinc production a d o r uptake of Th2 effectors early f ier
nematode iruection (Ramciswmy et al., 1994; UcMcawa et aL, 1994; Wahid et al., 1994).
Cytokine gene expression in GALT and transport of IgE into intestinal lumen occur within
24 hours after primary T. spirulis infection (Ramaswuny et al., 1994; Svetic et ai., 1993)
while cytokine secretion by MLN ceUs, mucosai mastocytosis, and IgE-mediated MMC
degranulation occur by 5 to 10 days of N. braifiensis and H. polygynrs infection (Arizono
et al., 1994; W W et aL, 1994; Chen & Enerback, 1996, 1996). Th2 cytokine leveis in the
intestinal lymph of T. spiralis-infected mice were found to accuntely r e k t the responses
in the gut mucosr. leding to the conclusion that the most important source of Iocd
cytokines duMg early infection is the mucosai ce& residing in the gut and not the draîning

lymph nodes (Ramaswamy et ai.. 1996). Similarly, others have observed that the mjonty
of cytokines produced in MLN, which receive both activated CD4+ T ceUs that were prirned
in Peyer's patches and naive lymphocytes circuhting from systemic cornputmenu, are
produced by cells denved Rom intestinal lyrnphoid tissues (Ho & Massouh, 1997).
Moreover, lymphocytes in the MLN were found to be the most potent source of growth-
stimulating activity for MMC hyperplasia during N. brmiliensis infection compared to
those in spleen or peripheral lymph nodes (Haig et a!. . 1984). These results imply that
cytokine production is inithted locdly at intestinal sites of infection and that these gut-
associated cytokines induce chernotactic recmitment of lymphocyte and effector
progenitors to IE or LP where they mature and dzerentiate into antigen-specific cefl
popuiations. Furthemore. the migrational pattern of mucosd lymphocytes from the gut to
systemic sites suggest that irnmunological mediators produced in GALT are important in
priming and perpetuating host responses in systernic compartments.
A further indication that immune responses in the mucosa are unique is the large
percentage of T c e k with the y 8 receptor. This y 6 popdation, which constitutes only 5%
of circuhting T cells (the majority carry the ap receptor), makes up approxirnately 508 of
the T lymphocytes in the gut mucosa of mice (Kaufmm. 1996). The role of y6 T cek in
Th2- type responses to nematode parasites has not k e n clevly elucidated. Most y6 ceils
are not CD4' T cells which have been shown to be important for protective immunity to
nemûtode parasites (Urbui et al. 199 la; Koyama et ai., 1994) and y 6 ceh do not
recognize MHC-associated peptides. (Raulet. 1989). However, there is evidence that y6
T celis mediate Th2 immune responses by providing 3 contact signal for B cell isotype
switching to IgE production in the presence of IL4 (Gascan et al., 1992; Wen et ai.. 1996)
and by preferentidy secreting Th2 cytokines spontaneously (Hiroi et al., 1995) or during
infection with N. brusiliensis in mice (Femck et ai., 1995). CD4' y8 T ceUs are rare but
may proMerate during infection in the gut and help drive the development of Th2 clones in
vitro (Wen et al., 1998). The selective expression of Th2 cytokines by y6 T cells and the*
substantial accumulation in the gut mucosa of mice suggest that y8 T ceIls play an
important role in the induction of Th2 response to intestinal nematode parasites.

2. THE HEUGMOSOMOIDES POLYGYRUS-MOUSE MODEL
Heligmosomoides polygynrs (= Nematosp iroides dubius) is a trichostrongyloid
pansite of murine rodents and has been proposed as a experirnentd mode1 for human
hookworm infection (B~tlet t & Ball, 1972) and other nematode infections of veterinary
importance (Monro y & E ~ q u e z , 1992). The chronicity of primary infection, the capacity
for acquired immunity to secondary infections, and the similvity of epidemiological
patterns between laboratory-rnaintained and free-living populations (Scott & Tanguay,
1994) make the H. polygyrus-mouse system a suitable mode1 for studying chronic
gastrointestinal helmintbis in humans.
The life cycle of H. polygyrus involves both free-living and parasitic stages and
occurs predomuiately in the small intestine of its murine host. As described by Bryant
(1973), parasite eggs are passed in the feces of the infected host. hatch within
approxirnately 36 hours to become the fûst h a 1 stage (L,), which then undergoes two
moults to form the ensheathed non-feeding infective third lamal stage (LJ. Ingested L-,
exsheath in the lumen of the stomacch and migrate to the srnail intestine where they
penetrate the serosal musculature and mature to fom the fourth larvai stage ( L,). Within
7-8 days post-infection (pi), the pre-adults migrate out of intestinal mucosa and emerge into
the intestinai lumen. Adult worms migrate preferentiaily to the anterior duodenum where
they mature, mate and produce eggs. Signincant levels of egg output are observed within
12- 14 days pi (Bryant, 1973) and an average of 1200- l5OO eggs per fende worm per day
are released in the feces of mice infected once with 100 infective L, (Kerboeuf. 1982).
2.1 Host Immunity to Primary Infection with H. polygyrus
Prirnary infections are chronicdiy mriuitained for 4 to 10 months depending on the
rnouse svain (Lawrence & Pritchard, 1994; Wahid et al., 1989). Apart from nuvitional
factors, parasite establishment, survival and fecundity duMg primary infection are
influenced by multiple externai variables such as host population density. level of exposure,
genetic background. and host immune response phenotype (Behnke & Wahid, 1991;

Kerboeuf, 1985; Keyrner & Tarlton, 1991; Scott, 1990). Even in mouse strains which are
able to reject primary infection with H. polygyms. the process of worrn expulsion is
protracted over several weeks with considerable variation among individual animais
(Behnke & Robinson, 1985). Primary H. p o l y g y m infections in most murine hosts are
long-lived despite the ability for gene expression and production of Th2 cytokines (Svetic et
al. 1993) and the presence of pansitic-specinc effector responses such as IgGl (Wahid &
Behnke, 1993). niis chronic survival of H. polygyms during primvy infection is believed
to be related to the immunosuppressing excretory-secretory proteases of adult w o m
(Monroy et af., 1989). These substances released by the adult worm have been shom to
suppress the activity of host generated immune ce&. to resuict intestinal peristûlsis and
mucus secretion, and to neutralize cytotoxic substances such as free oxygen radicals
produced by mucosal mut cells (MMC) or eosinophils (Dehhwi et al., 1987; Dehlawi &
Wakelin. 1988; Monroy & Enriquez, 1992; Pritchard et al., 1994; Reed et al., 1988; Smith
& Bryant, 1986). Aithough these proteolytic and antioxidmt proteases may moduiate host
immune responses, the overaii contribution of these immunosuppressing activities of the
parasite to the chronicity of primary infection has not been dennitively resolved.
Prirnvy infection with H. polygynrs induces marked enlugement and increased
celiulafity of the spleen and MW (Parker & Inchley, 1990a). Prelllninvy work by Ali and
Behnke (1985) suggested that the intensity and npidity of the prolifentive response in
secondary lymphoid organs are related to the inherent capacity of the mouse strain to expel
the Worms. In support of these hduigs. Parker and Inchley (1990b) reported that an
increase in the proportion of B lymphocytes in MLN (taken as a cmde measure of active
humoral immunity) was greatest and most prolonged in modente to high responder strswis
(BALBk and NIH mice) compared to slow responders (CBA mice). T ceN proüferation
was delayed in cornparison with changes in B cell freguency, while total T ceII numbers and
the ratio of T helper to T suppressor ceb were not ditrerent among the zhm strains.
MaxïmaX hyperplasia of lymphoid organs and peak B lymphocyre cefluiarity occmed within
7-12 days post-infection and coincideci with the appeuance of L, in the gut mucosa (Parker
& Inchley, 1990a & 1990b). These results support other observations that L, stimulate a

strong antibody-dependent inflmmatory and granulomatous response in intestinal tissues of
responder suains (Penttila & Jenkin, 1983; Monro y & EMquez, 1992).
Although primary polygyms infection in mice induces inmtration of innYnmatory
mediators (e.g. neuuophils and eosinophils) and parallel increases in serum IgE and IgG1,
mucosal mastocytosis is limited early d e r infection and noticeably absent once adults have
estabhhed in the anterior duodenurn (Dehlawi & Wakelin. 1988; Reed et al., 1988). The
suppression of mucosal mastocytosis by adult H. polygyms is not due to invinsic defects of
rnast cells because mastocposis can be stimulated by concomitant T. spiralis infection
(Dehhwi & Wakelti, 1988) but is associated with the selective dom-regulation of IL-9
and IL40 production in MLN (Brhnke et al., 1993). Although IL-3 and L-4 are
necessary for helminth-kiduced mucosal mastocytosis (Madden et al., 1991), both IL-9 and
IL40 have k e n shown to be important for optimal MMC proMeration and production of
MMC-derived granule proteases (Ghildyal et al., 1992 & 1993; Hultner et al., 1990;
Rennick et al., 1995). In contrast. other researchers have found high numbers of intestinal
MMC in mice infected with H. polygyruî compmd with that of naive mice (Urban et a l ,
1995) but this nematode-induced mucosal mastocytosis appears only after, and not before,
pronounced worm expulsion (Wahid et al., 1995). It has ken mggested that knpaired IgE-
or IgG1-induced activation of mat cek, and thus the reduced degranulation and release of
granule proteins, may be a more important reason for the prolonged worm s u ~ v a l during
p r i m q infection than the hufficient enlugement of MMC in GALT (Wahid et al., 1994;
Woodbury et al., 1984). Thus, the chronicity of primary infection may result from not only
immunosuppressive secretions from the parasite but also, and perhaps more imponmtiy,
from parasite-induced d ysreguia tion of Th2- type cytokine and effector responses.
It is controvenial as to whether IgGl, IgE, and eosinophil responses are beneficid
to the host infected once with H. polygynrs. Pritchard et al. (1983) reported that puritied
IgGl Crom immune semm of rnice infected with kt polygynrr caused severe stunting of
w o m and promoted adherence of peritoneal exudate ceUs to the surface of L, and adult
w o m in vitro. However, a Iater in vivo study by Wahid et al. (1993) showed t h t
parasite-specific IgGl responses did not correlate with the rate of worm expulsion and

were margindy elevated in rnice with heavier wonn burdens. Moreover, the inhibition of
IL4 and its receptor by neutralizing antibodies increved wom burdens in primary H.
polygyrus idection without concurrent aiterations in IgG 1 responses ( F i e h a n et al..
1988b; Katona et al.. 199 1). Later studies showed that adult woms induced mass
production of nonspecific IgG l (Robinson & Gustad. 1995) that could be bound with
proteins on the surface ofH polygynrs (Robinson et ai.. 1997). This was interpreted by
these authors as evidence of an attempt by the p m i t e to mask specinc antigens from
recognition and interaction with host immune celis. Other researchers have also questioned
the hinctional role of Th2 effectors during primvy H. polygyms infection. The
neutralization of secreted IgE by an anti-IgE antibody fded to promote wonn s u ~ v a l
although injection of anti-IL4 mtibody, which suppressed semm IgE concentrations,
resulted in increased worm burdens and fecundity (Urban et al., 199 Lb). IL4 treatment
elllninated an established primary H. polygyrus infection in immunocompetent mice but was
not associated with changes in eosinophü or serum IgE levek (Urban et al., 1995).
Furthemore, injection of an anti-IL-5 antibody suppressed the development of eosinophilia
but had no effect on worm numbers or egg production (Urban et al.. 1991b). In contnst to
these results, Wahid et al. (1994) found that the ability of mice to expel adult H. polygyrus
d u ~ g primary infection was directly reiated to their potential to secrete Th2 cytokines and
to produce prominent and sustained IgG1, IgE, ûnd MMC responses. Thus, an increase in
parasite-induced production of Th2-type effectors reflects a capacity for specific
immunological responsiveness but the precise functions of these Th2 effectors in mediating
host immunity to H. polygynrs infection remah poorly dehed. The inhiiition of a single
parameter of Th2 defense mechanism without a subsequent detectable eEect on worm
expulsion or fecundity does not n e c e s s ~ y prove that these effectors are not hinctiondly
active or important in the host immune response to nematode parasites. but suggests that
there may be some redundancy of host defenses which has evolved to prevent the selection
of parasites that are resistant to any one of these defenses (as reviewed by Fieiman et a[-,
1997). To this end, the host may have developed a complex set of defense mechanisms that
cokctiveIy, through their interactions and compensatory responses. act to effectivdy resist

infections with nematode puasites. However, for reasons that remain poorly understood.
this putative network of cooperative host effector responses fails to provide adequate
protection against primary infections with H. polygynis. In summuy, low immunologicd
responsiveness and chronic primary infection with H. polygyms appear to be related to
geneticaily detemiined host susceptibility to pansite-induced immunomodul;ition, dthough
the critical protective mechanisrns thnt are suppressed are not clearly defined.
Despite the equivocal evidence for the benefit of Th2-type effector mechitnisms. the
production of Th2-type cytokines is thought to be crucial for the resolution of H. polygyms
Section Injection of anti-IL4 or anti-IL-4 receptor antiiodies into rnice idected with H.
polygyrus increased worm survival and fecundity in mke (Urban et al.. 1991b). The
importance of IL-4 to protection a g h t prirnary infection was confirmed by a later study
which found that treatment of H. polygyms-inocuhted mice with exogenous purûied IL4
signincantly lirnited puasite s u ~ v a l and completely suppressed fecundity (Urban et al..
1995). Interestingly, the effector mech;uiisms by which IL-4 tenninated the primary
infection were uncleu because leveis of MMC. IgE, and eosinophils did not dBer between
IL-4-treated and untrerted rnice. The independent ability of increased IL-4 concentrations
in vivo to selec tively enhance these Th2 e ffec tor responses in the absence of altered
concentrations of other Th2 cytokines is no t known. The observation that IL-4 treatment
inhibited adult worm s u ~ v d to a greater extent than h a 1 establishment led Urbm et al.
(1995) to specuiate that IL-4 rnay activate nonspecitic immune defenses such as intestinal
smooth muscle contraction. mucus secretion, or peristalsis which in tum rnay help facilitate
worm expulsion f?om the s m d intestine. Recently, preliMnary data suggest that IL4 rnay
increase gut spasticity. s m d intestinai permeability, and decrease fluid absorption in the
s m d intestine (Urbui et ai.. 1996). These changes in smooth muscle contractility and
intestinal fluid dynamics may faciltate expulsion of H. polygy~us during primary Section
by decreasing parasite anachment to the intestinal viIli (thus limiting parasite establishment)
ancilor by testricihg access of worms to their food source within mucosai tissues.
Alternatively, IL-4 treatment rnay have initiated immune-mediated responses bat damrged
the lame or young adult Worms and thereby reduced their potentid for chronic suMvaL

2.2 Systemic Versus Gut Mucosal Immunity to Rimary Infection 16th H. poiygyms
Aside from the analysis of mucosai rnastocytosis, much of our knowledge on
immune responses to H. polygyrus ha been based on serum levels oETh2-associated
antibodies (e.g. IgE and IgG 1) or cytokine production (TL-4, IL-5. IL9 or IL- 10) in the
spleen. Studies examining the specifc advantages of Th2 effector mechanisms such as IgE-
mediated immune reactions have produced inconsistent results perhaps because they have
not examined these Th2 immune responses at the usual site of infection in GALT. It
appears logical that induction of Th2 cytokine and effector ceU production also occurs in
intestinal mucosal tissues where the nematode piirasite first interacts with the host and
spends the majority of its life cycle. However. ody recently bas there k e n concentnted
research on the role and relative importance of gut mucosd immunity in the resolution of
intestinal nematode infections.
Parker & Inchley (1990a) reponed that the pcak cellular proliferative response to
H. polygyms infection occurred eulier in MLN than in spleen (Parker & Inchley 19903,
thus suggesting that the k s t Ipphocytic response to an intestinal nematode is restricted to
infected enteric regions. Unfortunately, the authors did not determine whether this eulier
cellular proliferative response in MLN resulted in a more rapid omet of functionai immunity
at the gut kveL To mess tirne-dependent cornpartmentalization of Th2 immune responses
to H. polygyrus, Svetic et al. (1993) studied Th2 cytokine gene expression in spleen and
GALT (MLN and Peyer's patch) within 12 days after primary Section The ruthors found
that IL-3. IL-4, IL-5, and IL9 mRNA expression as measured by a RT-PCR assay was
markedly elevated in the Peyer's patch and MLN ceils within 2 days pi and remained above
u ~ e c i e d levels for up to 12 days afier infection. In contrasr. the gene expression for
these Th2 cytokines remained unchmged in splenic c e h diroughout the experirnentd
period. These results suggested that the early hinctional immune response to H. polygynrs
as measured by mRNA activîty w u locaiized at intestinai sites of infection CurrentIy, no
study has c o n h e d whether the higher Th2 cytokine mRNA expression in the gut during
EL polygym infection is accornpanied b y equivalent increases in pro tein secretion although

gene expression usuaiiy correlates well with protein synthesis. Table 1 shows a compilation
of data on the kinetics and site-specifcity of Th2 cytokine and effector responses following
prllnary Section with H. polygynrs. The principal conclusion to note from this table and
the above studies is that Th2 cytokine gene expression and secretion in GALT usually
precede the production of Th2 cytokines in the spleen as weU as the onset and peak of Th2
effector responses in the peripheral circulation.
Because wom outcomes were not assessed by Parker & Inchley (1990a) or by
Svetic et al. (1993), it is unclear whether this earlier omet of ceiiuiar and cytokine
prouerative responses in GALT i s critical for protective irnmunity to the nematode
parasite. In one study that examined the correlation between Th2 immune responses in
GALT and worm burden of a primary response, Wahid et al. (1994) detemiined whether
Th2 cytokine secretion in MLN as weil as MMC responses difZered between fast and slow
responder mouse s trah. The fast responder suain (SAR mice) expelled over 961 of the
Worms by 8 weeks pi compared to the slow responder strain (CBA mice) which
experienced no signincuit worm Ioss throughout the experimental penod. The MLN of
SAR mice secreted consistently higher amounts of L-3, IL-4, and IL-9 over the course of
infection compared to MLN of CBA rnice. Moreover, significantly higher MMC counts
and senirn concentrations of MMCP-I (used as a systernic marker of mucosal activation
ancUor matocytosis) were found in SAR rnice versus the CI3 A mice. Thus, suMvaI of the
parasite and ultimately the chronicity of infection may depend on the intensity and
persistence of Th2 cytokines and effector responses ar the intestinal kveL The relative
importance of gut mucosal versus systemic immunity to infection outcomes such as worm
s u ~ v a i has not been studied in Our experimental H. polygynrs-mouse modeL

Table 1: Kinedcs of Th2 Cytokine and Effetor Production Post-primary (PH) and Post-challenge (PCI) Infection with H. pofygyms in BALBlc mice (unless othenvise indicated).
The Post-infectio n PPI PCI CytokineEffector Response Tissue Location
Onset of IL-3, a-5, IL-9 gene expression PP1 Marginal increase in IL4 gene expression MLNL
1-2 days Sustained L 3 , IL-9 gene expression PP & MLN'
2-4 days
4-6 days
6-8 days
Omet of IL4 gene expression PP & MLN' Onset of I L 3 & IL-4 secretion (SAR mice) MLN'
3-5 days Omet of eosino philia (NIH mice) Whole Blood3
Peak IL-5 gene expression Peak IL-3 gene expression
PP' PP & MLN'
4-6 days Onset of total IgE response Serum4* ' Onsei of total I Q response (NIH mice) Semrn3
5-7 days Peak eosinophilia Whole Blood3* Onset of parasite-specifk IgG 1 (NIH mice) Senimg
Sustained IL-5 gene expression MLN ' Peak IL-9 secretior (SAR mice) MLF Muginai increase in IL- 10 secretion MLN2 Onset of MMCP- I increase Semm2 Onset of total IgG 1 response (NIH mice) Serurn3
7 days Peak total IgG l response Serum6
8-9 days Peak total IgE response Sem4* ' Deche of eosinoph (NIH mice) Whole B100d3
7-14 days Onset of to tai IgE and to ta1 IgG 1 response Serum4*
continued PP = Peyer's patches MLN = Mesenteric lymph nodes

Table 1 : continued
T h e Post-infection PP PC CytokindEffector Response Tissue Location
12 days Peak MMC numbers (SAR. CBA mice) Smdl IntestineZ Peak IL-5 gene expression MLNL
10-15 days Peak pansite-specific IgG l response serurns
14-15 days
16 days
20-21 days
24-30 days
Peak eosinophilia Whok Blood6 Peak total & specitic IgGl response serumb
14-2 1 days Peak total & specitic IgGl response Serum6* ' Peak serum IgE response Serum63
Peak MMCP-1 leveis (SAR. CBA rnice) Semmz
Second peak of IL-9 secretion M N Decline of IL-4 secretion (SAR/CB A rnice) MLN2 Peak total IgE response Senim4
Dccline of MMC numbers (SAR mice) S m d Intes the' Decline of MMCP- 1 leyels (CBA mice) Serum'
MLN = Mesenteric lyrnph nodes T
' Svetic et ai., 1993 * W;ihid et al., 1994 ' Shter & Keymer, 1988 Urban et al., 199 1 a ' Wahid & Behnke, 1993 Boulay et al., 1998
'Urban et al., 1991b ' Katona et al.. 1991
Wahid & Behnke. 1992

2.3 Host Imminity to Challenge Infeftion with H. polygyrus
The hast is capable of developing effective resistance against secondary exposures
to H. polygyrus following the removal of adult worms from a primary infection by an
anthelmintic-abbreviated regirne (Behnke & Wakeh, 1977; Behnke & Robinson. 1985).
oral inocuiation with irndiated b a e (Slater & Keymer. 1988). or other immunizing
methods (Urban et al., 199 la). The rapidity and intensity of acquired immunity achieved
upon chdenge infection depend on the dose of the immunking inoculum, the genetic
susceptibility of the host, and the infectivity of the larvae ( Jacobson et al., 1982; Ali &
Behnke, 1985; Behnke & Robinson. 1985; Dobson et al.. 1985; Enriquez et al., 1988).
Protective immunity to challenge infection with H. polygyms is demonstrated by reduced
won site, depressed fecundity. and more rapid expulsion of adult wonns (as reviewed by
Monroy & E ~ q u e z , 1992). The L, and L, IYvae of H. polygyms are believed to be potent
inducers of protective immunity in responder strains of mice (Monroy & Enriquez. 1992;
Wahid & Behnke, 1992). In most murine straiils, p r i m q H. polygyms infections, which
are terminated before the parasites mature to adulthood by drug treatment or irradiation.
induce high levels of acquired resistance to iarval challenge infections (Behnke et al.. 1983;
Behnke & Robinson. 1985; Wahid & Behnke, 1992). The strong and npid acquired
imrnunity to secondary lyval exposures fo flowing an inmuniMg regime is thought to
overcome any inherent immunomoduIatory stntegies employed by the nematode puasites
(Monro y & Enriquez, 1992). However. develo pment of ihis pro tective immunity depends
on the stage of the parasite used in the irnmuniPng pmcedures because no protection wûs
evident in mice when adult w o m were used for the initial sensitization. when the primyy
infection was terminated after the appearance of adults worms in the intestinal lumen, and
when the mice were chatiienged with adult worms (Behnke et al.. 1983; Enriquez et al..
1988; Jacobson et al., 1982; Pleass & Bianco, 1994). The appevance of increased effector
responses upon h a 1 challenge of previously immunized hosts suggests that immun0 Io gical
processes primed during primuy exposure are important in the effective resolution of
secondary infections (as reviewed by Monroy & Enriquez, 1992).

Immunological memory is characterized by accelerated speed, increased size and
persistence, dl of which are observed during secondary immune responses to challenge or
repeated idections with nematode parasites. Immunocornpetent hosts are able to develop
strong resistance ta challenge infection with H. polygyrw as evidenced by the dramatic
reduction in worm survival, wonn size, and fecundity shortly after secondvy exposure
(Behnke & Wakelin, 1977; Behnke & Robinson, 1985). This rapid and intense temination
of secondary H. polygynrs infections is thought to result from the mobilization of protective
Th2 immune responses that had been primed during previous mtigenic exposure. Acquired
immunity to challenge infection following an anthelminthic-abbreviiited primary infection is
CD4' T celi dependent and involves Th2 effector responses (Urban et ai., 1991a). Indeed,
senun IgGl rose ten-fold from resting levels in mice repeatedly immunized with immune
mouse serurn (Williams & Behnke, 1983) and the-fold in mice challenged with the
pansite compared to mice given only a single inocuiation (Urban et al., 199 la). Passive
tramfer of pr imvy infection senirn containhg pansite-specinc IgGl did not protect
unprimed mice from H. polygyrw (Williams & Behnke, 1982) whereas multiple Section
sera were capable of transferting immunity to naive recipients (Behnke & Parish. 1979).
Also. concentrations of semm IgE and numbers of MMC were significantly higher in
chaiienged mice relative to primary infection controls (Dehlawi & Wakelin, 1988; Urban et
al., 199 la). Consistent with the concept that secondary exposwe to polygyncs in
previously immunized mice stimuhtes npid activation of memory Th2 immunity, Table 1
shows that the onset and peak of IgE, IgG 1 and eosinophil responses in the peripheral
circulation of challenged mice generdy occurs 2 days to a week earlier than that foUowing
primary infection. Unfortunately, there are no studies evduating specitic cytokine profles
during challenge infection and therefore it is u n c o ~ e d as to whether an ekvated
secretion of Th2 cytokines is responsibk for the apparent potent activation of Th2-type
effector responses in mice chdienged with nematode parasites. However, this cytokine and
eEector interaction in the gut is highly probable considering that cytokines are the primary
factors that induce and reguiate effector responses. A cytokine inhriition study by Urbm et
al. (199 1 b) provided some indirect evidence that Th2 cytokines are important for the

resolution of secondary infections. Injection of an anti-IL4 antibody suppressed
secondary IgE responses and was associated with increasised worm counts and fecundity on
day 48 pci Interestingly, the role of IL4 and associated eosinophil response in acquired
imrnunity to H. p o l y g y m is not clear because neutrdimtion of IL-5 blocked nematode-
induced eosinophilia but did not affect worm burden or fecundity in chaiienged mice.
The relative contribution of naive and rnemory effector c e h to immune responses
observed during challenge infection is not cleu. Memory Th ceUs are not as stringent in
their rcquirements for IL-4 or costirnulatory signais (CD28/CTLA4 and B7.1tB7.2
interactions) during activation and effector activity compared to naive Th ceUs (Gause et
aL. 1995; Huang et aL. 1997). It is possible that the priming of Th2 cytokines during
prirnary infection rnay be a more criticai immunologicai event than secondvy responses for
the development of protective imrnunity to nematode parasites. However, some studies
have suggested that memory CD4+ T ceh are unstable, depend on the persistence of
antigen, and may reven to a resting, quiescent form that is phenotypicdy indistinguishable
from naive CD4' T c e h (Bunce & Bell. 1997; Bell et al.. 1998). IL-4 was found to be
crucial for maximal capacity for Th2 cytokine secretion by memory Th effector c e h that
came from a heterogeneous pool of resting memory CD4* T ceUs (Bradley et al.. 1995)
and therefore may be important in potentiating previousiy comrnitted active T ce& as weii
as in reactivating memory revenant T cek. In addition. Katona et al. (1991) and
F i e h a n et al. (1 997) reported that IgE responses in secondary infections with H.
polygynrs were derived mostly from naive or uncommitted B cells rather thm fiom IgE-
committed memory B ceils produceci during initiai nematode infection. These preliminary
results must be verined by experirnents on other Th2 effectors to provide precise
description of the immune popuktions and reguliitory factors that are activated during
challenge infection.

In sumrnuy, functional immunity to H. polygyms appears to be a Th2-dependent
process because it involves specific cytokines such as IL-4 and effectors such as IgE-
dependent eosinophils and MMC. The magnitude and eficacy of Th2-mediated immune
mechanisrns may d s e r according to the infection protocol used with the greatest level of
protection seen d e r secondary exposure. Preliminary data point to the importance of Th2
immunity in GALT which may play a d e in promothg the O bserved increases in serurn
Th2 effector responses. Given that immunocompetent hosts are able to mount a strong and
effective acquired immunity upon subsequent infections, it would be important to determine
the magnitude. kinetics and companmentdhtion of Th2 immune responses during
challenge infection and to mess whether these characteristics are altered by nutritionai
deficiencies such as protein malnutrition.

3. EFFECT OF PROTEIN DEFICIENCY ON HELMINTH INFECTIONS
Pro tein deficiency is generdy synergistic with bo th naturd and experimental
helminth infections and thereby results in an interaction that is more seriously harmhl than
would be expected Erom the occurrence of either condition alone (Scrimshaw et al., 1968).
Protein malnutrition and intestinal parasitism share similar geogrrphicd distribution patterns
with the sarne individuais experiencing bo th diseae States simultaneously throug hout their
lifetime. For exampie, a study of rurd Nigerim children found that protein intakes were
inversely proportional to the number of CO-existing intestinai parasitic infections
(Rosenberg & Bowmm, 1984). Additiondy. Trichuris egg counts in children iiving in an
urbm slum region of Indonesia were correlated with stunting. a physiological indicator of
chronic protein calorie malnutrition (Hadju et ai., 1995. These data supports the concept
that malnourished children living in helminth endernic areas are more heavily parasitized
than theY webnourished counterparts. However. given the multi-dimensionai nature of
malnutrition-parasite interactions and the interpretative pro blems of epidemiological data, it
is difncult to ascertain which condition was the preceding or causative factor.
3.1 Effect of Pmtein Deficiency on Pnmrry Infection with H. polygyms
ControUed experirnents are more conducive to studying the nature of putative
casual rehtionships between protein mdnutrition and intestinal parasitic Uuection. These
systems atternpt to reconsvuct the natunl transmission of nematode parasites o c c u ~ g in
human settings by establishg a cohon of susceptible animals inhabithg an arer of endemic
helminth infection. Shter (1988) housed N c e in large, unrestricted cages which provided
free access to a 2% or 16% protein diet. An initialcohort of mke w u infected with H.
polygyrus, and at 5 week intervals, subsequent cohorts of young uninfected rnice were
introduced into the enclosures whüe previous cohorts were removed. The rate at which the
uninf'ted mice acquired the parasites was higher in the low protein colony, thus îndicaiing
th t protein deficiency may enhance the rate of pansite transmission within a mobile
population of hosts. The prevalence of infection arnong protein-deficient mice was higher

compared with that of the welI-nourished mice and may be related to the observed increase
in s u ~ v a i of aduit womis in these deficient animals over the experhental period. These
results show that, under naturd infection conditions. protein deficiency may potentiate the
prevalence and intensity of a nematode parasitic infection.
Evidence to support these fkdings using controiled p h a r y infection protocois is
inconclusive. An euly study by Bawden (1969) showed that a marginal protein diet (white
bread with 12.3% crude protein) produced greater establishment of H. polygym in mice.
signincant stunting of female adult w o m . and more widespread distribution of nematodes
dong the srnall intestine relative to the nomal protein diet (pelleted concentrate with
25.2% protein). Interestingly, protein restriction had no effect on the long-term expulsion
rate of nematodes but w u associated with decreased fecundity at 28 days post-infection.
Bawden suggested that the retarded growth and dtered distribution of femde w o m
recovered in the malnourished hosu adversely affected their reproductive capacity. Given
that H. polygym w o m tend to mate and produce eggs in the antenor duodenum, the
10 nger migration route induced by pro tein restriction rnay have subsequently impeded their
usud reproductive processes. It is dficult to interpret these results due to the intermediate
level of protein restriction and the use of diets with difTerent macronutrient constituents.
These uncontrolled d i e t q factors may have been responsible for the absence of nutritional
impact on chronic worm survivd Using purifed synthetic diets, Bnilsford and Mapes
(1987) reported that a more severe restriction of dietvy protein (2% casein) did not
significmtiy increase wom establishment or s u ~ v a l despite inhibiting body weight gain
and producing overt hypoalbuminaemiû. during primuy infection. These authors found that
worm burdens in control(204b protein) and protein-deficient (2% pro tein) animais
remained equivalent at 3-4 weeks post infection. Previous studies in our laboratory found
that marginal (7%) and low (3%) protein diets resulted in greater worm burdens at day 29
post-primary infection (Boulay et al.. 1998). These results dEer fiom those of Bnilsford
& Mapes (1987) perhps because Boulay et al. (1998) found unusudly low parasite
numbers in the control diet group (rather than larger wonn burdens in the deficient mice)
which may have kad to signûicant differences in worm sumival between dietxy groups.

It is possible that a primary infection protocol in which laboratory anima are
exposed only once to the nematode parasite may not be a sensitive experimental system to
evduate the influence of dietary protein on outcornes of H. polygyn<s infection. P N n q
infections with H. p o l y g y m are usually chronic in othenvise immunocompetent murine
hosts. suggesting that the parasite has the inherent capacity to down-regulate or prevent
protective host immune responses (Behnke et al., 1992; Dehhwi & Wakelin, 1988; Smith
& Bryant, 1986). Assuming that secondary infections nonnally stimulate development of
protective immunity in weU-nourished immunocompetent hosts (Monroy & Enriquez
1992) ami that protein deficiency impairs irnmunological processes, a repeated or challenge
infection protocol mny help to maximize the chances of detecting any possible effects of
dietary pro tein deficiency on intestinal helminth popuiation dynamics.
3.2 E f k t of Protein Deficiency on Challenge Infection with H. polygym
Experimental infection studies show that the level of dietvy protein influences the
extent to which the host cm develop protective irnrnunity to repeated or secondvy
infections. Slater and Keyrner (1986) fed mice 2% or 8% protein diets and repeatedly
infected animais with vvying doses of infective k a e of H. polygyms every 2 weeks for a
period of 12 weeks. Mice fed the 2% protein diet accumulated worms in direct proportion
to the intensity of exposure throughout the experiment whereas mice fed the 896 protein
diet showed partial resistance to repeated infection at higher doses. Also. net egg
production was increased in rnice fed the Iow protein diet due to both higher per capita egg
production of femde w o m and hewier worm burdens. The authors attributed the
increased s u ~ v a i and fecundity of w o m in the mainourished mice to either a diet-induced
inhibition of acquired immunity to reinfection or to increued sensitivîty to parasite-induced
irnmunosuppression. Both explanations rely on the assumption that protein deficiency
impairs optimal irnmunocompetence. Consistent with the results obtained by Skiter and
Keymer (1986). Keymer and TuIton (1991) observed higher wonn burdens in repeatedly
infected mice fed 3% protein diet compared with those fed 16% protein diet. These
authors also used a challenge infection protocol in which the primary infection was

abbreMated by an anthelmintic dmg and the mice were then reinfected once with 100
infective bac. At two weeks post-challenge infection, mice fed the low protein diet had
more w o m thm their weU-nourished counterparts, suggesting that pro tein deficiency
impairs development of irnrnunological memory normally primed by previous exposure to
infectious pathogens. Using a chailenge infection protocol. Shter & Keymer (1988)
recovered significantly more w o m in &male NIH mice fed diets containhg 2% protein
compared with 16% protein at 21 days post-challenge infection. In contrat to restlts of
their previous study (Shter & Keyrner, 1986) which had ernployed a repeated infection
protocol, Slater & Keymer (1988) found that per capita fecundity of a femde worrn in mice
challenged with larval H. pofygyms was not altered signüicantly by the low protein diet. It
is unknown whether repeated infection protocois elicit different immunological responses
than challenge infection protocols and whether this rnay be responsible for these confiicting
effecu of protein deficiency on worm fecundity. Bouhy et al., (1998) reported that o 3%
protein diet increased total egg output compared to a 24% protein diet throughout four
weeks of challenge infection with H. polygyrus. Taken together, protein restriction
impaired the capacity for acquired host resistmce to secondary infection as indicated by the
increased worm burdens in previously immunized protein-deficient mice.
In sumrnary, Gndings from controiled labo ratory and experimentai epiderniology
studies demonstrate that the overd protein nutriture of the host affects the population
dynamics of a nematode parasite. Although many researchen have attributed the increased
survivai and fecundity of nematodes in protein-deficient hosu to impaired host imrnunity,
only a few studies directly examined Th2 immune responses after both nematode infection
and protein deprivation (e.g. Boulay et al., 1998). Inasmuch as host immune defenses are
important in mediating susceptibility to parasitic infection, the immunologic mechanisms by
which protein deficiency rnay exacerbate intestinal parasitism deserve in-depth
investigation. To this end, the foilowing sections discuss in d e t d some of the literature on
protein malnutrition and host immunity.

4. EFFECT OF PROTEIN DEFICIENCY ON SYSTEMIC CELL-MEDIA'IED AND HUMORAL IMMUNITY
The predorninant approach of previous literature on interactions of malnutrition
and immunity has k e n to examine diet-induced structural and hnctional changes in
systemic lymphoid tissues and theu associated immune ce1 populations. The thymus and
bone rnarrow are essential tissues for the production of lymphocyte, monocyte and mast
celi pro genito rs, ho wever, the y are no t specitically designed to facilitate immune responses
upon antigenic stimulation. Initiation of rnost primary immune responses to blood-borne
pathogens including antigen presentation, activation of B and T lymphocytes, and the
production of antibodies and T ceU-derived cytokines occurs mainly in the spleen. Antigen-
specific immune effectors (e.g. imrnunoglobulins and differenthted T cells) produced in the
spleen enter the systernic circulation to migrate to sites of infection and are detectable in the
peripheral blood. Thus, the term systemic immunity used in this paper wili apply to
nonspecific and specitic immune responses in penpherai sites such as the bloodstreun,
thymus. or spleen.
Dietary protein deficiency has been shown to adversely affect the morphology of
systemic lymphoid orgus, particukly in the irnrnediate postweaning period which
represents a crucial stage of development and maturation of the lymphomyeloid system
(Speu & Edehan, 1974). Thymic and splenic atrophy is chmcteristic of protein-energy
malnutrition in children and young kboratory animais (Chuidra, 1983). Bell et al. (1976)
found significant organ weight loss, decreved ceil yields, and stnicturally disorganized cells
in the thymus and spleen of w e d h g mice fed a 4% protein diet for four weeks cornpmd
to mice fed a 2 8 4 protein diet. Given the importance of the thymus and spleen in the
production and maturation of T lymphocytes, respectively, a likely consequence of
lymphoid organ involution is decreased numbers or altered proportions of T lymphocyte
subpopulations. Pro tein-deficient rats and patients with kwashiorkor have lower counts of
total cucuiating or splenic T lymphocytes (Abbott et al., 1986; Bises et aL, 1987;
SIo bodivlik et al., 1984) whiIe pro tein-rnalnourished adults also had lower percentages of
CD3+. CD4+, and CD8' T cells in blood (Abbott et aL , 1986). Contradicting these fjndings,

Woodward and Miller (1991) and Woodward et al. (1995) reported rhat a very low protein
diet (0.5% protein) increased the percentages of CD4' and CD8+ ce& reiative to totd ceU
numbers in spleens of weanluig mice. The biological significance of these proportional
increases is unclear considering that the total cell population was drastically reduced (96-
998) by the severe protein deficiency. Some researchers found greater proportion of
cytotoxic/suppressor (CD8+) T cells relative to that of T helperfinducer (CD4+) c e h in
pro tein-deficient anirnals (Nimmanwudipong et aL. 1992) while others reported that the
ratio oFCD4+ to CD8+ celis did not v u y from control values in protein mainourished
animals or humans (Abbott et al., 1986; Bises et al.. 1987; Woodward & Miller. 1991).
Therefore, it is equivocal as to whether an altered ratio of helper to suppressor T ceUs
contributes to the Th cell-dependent immunodepression observed in protein malmurished
animals (Woodward & M e r , 1991). A recent paper examining the effects of protein
deficiency in experimental pulmonary tuberculosis in guiner pigs reponed that pmtein
deprivation (10% ovalbumin versus 30%) did not affect the proportions of ap T c e h
(CD4' and CD$+) in the spleens of naive animais (Mainali & McMurray, 1998). When
mycobacterial infection was superimposed on the protein deficiency, there was a d m &
Ioss of ap T ce& Erom the spleen ûnd a large decrease in the CD4+/CD8+ ratio in lymph
nodes due to a disproportionate increase of the CD8' subset at the expense of CD4+ T ceiis.
These results imply that analysis of immune cells in uninfected animais may not necessdy
reflect induced responses to antigenic stimuhtion. Lower numbers of CD4' ceh would
have O bvious implications for the capacity of the host io respond effectively to nematode
parasites because CD4+ T cells control the development and activation of B ceUs and
effectors via secretion of cytokines or direct cellular interactions. Interestingly, Woodward
et of. (1995) observed higher numbers of CD4+ and CDS* cells expressing the CD45RA'
surface market of quiescent or hyporesponsive T cek relative to ceils with the CD45RA'
marker of memory-phenotype T ceIlse These results provide the first indication that protein
deficiency decreases the number of responsive T ceîl subsets in the absence of exposure to
an extemal antigen The exact mechuiisms by which protein d e e n c y contnïlutes to
depressed activation and maturation of T ceiis remain to be elucidated.

Recent data point to the involution and dysfunction of specinc thymic
compartments, namely the thymic epithelium which promotes functional dserentiation of T
lymphocytes through the release of thymulin and other hormonal factors, as contribuhg
factors in the decrease in T lymphocyte nurnbers seen in protein depleted subpcts (Wade et
aL. 1988; Woodward et al., 1992a). Administration of thymuh by Parent et al. (1994) to
malnourished children with marasmus andor kwashiorkor increased the percentages of
CD3'. CD4+ and CD8' and decreased the proportion of immature T lymphocytes (CD 1 a) in
their penpherai blood, Also, thyrnulin is reputed to exert a positive influence on
prolifentive capacity of peripheral T ce&. Thus. the thymic and splenic atrophy induced by
protein malnutrition in we&gs may result in depressed maturation and prolifierative
capacity of specific T lymphocytes. Unfortunately, none of these studies e x d e d the
hinctional hpücations ofdecreased or altered T ceii subpopuktions during infection in
mainourished mixnds or humans.
In addition to dtered nurnbers of specific types of T lymphocyte. protein deficiency
may have detrimental effects on the efiïcacy of CMI responses to Mtogenic or antigenic
stimulation. Deitch et al. (1992) found that splenic lymphocytes from mice fed a nearly
protein-kee diet (0.03% protein) for 7, 14 or 21 days had progressively lower biastogenic
responses to two T ceil mitogens (PHA and ConA) compmd to basehe levels. The lower
mitogen responsiveness of T lymphocytes from mice fed a low protein diet is a crude
indicator of depressed nonspecitic CM1 but does not necessarily refiect altered T cell
responses to speciiïc antigens. Lamont es al. (1988) showed that the vansfer of ovdbumin-
primed lymphocytes fiom donor mice fed a 24% protein diet to the footpads of naive mice
fed 4% protein die< resulted in a depressed cutaneous delayed type hypersensitivity @TH)
response (a characteristic feature of CMI) whereas a positive DTH response w u elicited in
footpads of naive mice fed 24% protein. Using a graft rejection assay. Larnont et al.
(1988) found that splenic T cells derived Crom rnice fed 4% protein induced significant
splenornegaly in webnourished recipients. The authors interpreted these resuits as evidence
that protein deficiency did not impair the functional capacity oCT cells perse but rather it
altered other ceU-mediated immune responses (e.g. innltratioa of leukocytes to site of

antigen challenge or T cekdependent activation of macrophages) which are necessary for a
positive DTH response. Thus. protein deficiency may decrease not only the proliferative
capacity of T lymphocytes but dso impaircd T cell-dependent immune processes and
interfered with celluiar interactions required for effective CM1 responses.
Although thymic involution and su bsequent alterations in T lymphocyte numbers are
frequent rndestations of protein deficiency, evidence is inconclusive as to whether CMI is
more seriously impaired by malnutrition than is thymus-dependent humord imrnunity.
Severe protein restriction (0.5% protein) induced a greater reduction in splenic B
lymphocyte numbers than in T lymphocyte numbers (Woodward & Miller, 1991).
However, changes in lymphocyte numbers do not necessarily correlate with altered
hnctiond lymphocyte responsiveness. Woodward et al. (1 W b ) found that protein
denciency impaired DTH to sherp red blood cells (SRBC) but had no effect on the serum
titres of anti-SRBC antibodies. Human studies consistently report depressed DTH
responses, lower mitogen-stimulated proliferative capacity of T lymphocytes, and lower
numbers of circulating T lymphocytes but presemed B cell function and
hyperimmunoglobuiinemia in chiidren with protein malnutrition (Chandra. 1983; Ozkm et
aL, 1993; Punilo et al.. 1976). In support of these fmdings, Deitch et al. (1992) reported
that severe protein deficiency (0.03% protein) decreased biastogenesis of spknic
lymphocytes to T cell mitogens (PHA and C o d ) to a greater extent than that to a B celi
mitogen (PWM). Although these mitogens do not elicit exclusive proliferation of either T
or B lymphocytes, these results do imply that competence of CM1 rather than of humorai
immunity is more sensitive to changes in dietary protein IeveL In sumrnary, protein
deficiency causes structural and functional defects in systemic lymphoid organs and
peripheral lymphocytes which are required for the induction and propagation of CM1
ac tivities and T cell-de pendent hum0 rd respo mes.

5. EFFECT OF PROTELN DEFICIENCY ON GUT MUCOSAL IMMUNITY
The functional relevance of systemic immune respoases to intestinal heiminths is not
clear because infections with these parasites are predominantly localized in enteric tissues.
Given that the gut c o n t a speciaüzed secondary lymphoid tissues that can react directly
with antigens present in intestinal mucosa or lumen. it is of cruchi importance to examine
the consequences of protein deficiency on gut mucosal immunity to nematode parasites.
The impact of protein deficiency on gut mucosd immunity cm be tirst observed by changes
in the gross morphological chancteristics of GALT. Bell et al. (1976) reported that dietvy
protein restriction (4% protein) induced atrophy of MLN of we;uiling mice as seen by the
reduction in organ s k and loss of mature B cells over the four week feeding penod.
However. the authors found chat the tissue loss and structurai defects in the protein
deficient mice were less pronounced in MLN than those in the thymus and spleen.
Simiiiirly, Lochmilier et al. ( 1992) reported no signifcant differences in the number and
average total weight of Peyer's patches in the srnail intestine among weanling rats fed 4%
and 16% protein diets for 6 weeks. When expressed relative to body weight. Peyer's
patches were 84 to 95% larger in the protein-deficient rats compared to the wehourished
rats. indicûting that the low protein diet crused a greater decrease in total body weight than
in the weight of Peyer's patches. In contrast. Woodward and Miller (1991) and Deitch et
al. (1992) found that severe protein rndnuuition equaily depressed splenic and M W orgm
weight and lymphocyte pools. The CD4/CD8 ratio and percentage of CD4' or CDS' ceh
in MLN were not affected by feeding a very low protein for 3 weeks (Lee & Woodward.
1996). Taken together, these studies indicate bat gut lymphoid tissues mîy be more
resistant to gross organ stunting induced by moderate protein restriction than peripheral
lymphoid organs or body mus in growing animais. However, the very severe ievels of
protein restriction used in bo th studies (0.5% and 0.03% pro tein, respectively) and the use
of nYve animais preclude my applicable conclusions to be drawn Erom these 6ndings
toward nematode infections in protein-depkted humans. Regardless of the relative extent
of gross structural changes to intestinal lymphoid tissues. there is considerable evidence that

protein deficiency induces morphological abnomalities in the intestinal villi (e.g. crypt
hypoplasia) md increves pemeability of the gut mucosal bvrier (Curnmins et al.. 1987a &
1987b; Deitch et al., 1992). Diet-induced villus atrophy and a disrupted mucosal barrier
integrity may enable parasites to penctnte and establish in gut mucosal tissues and thus
could be more dethentai to host resistance thm gross changes in lyrnphoid organs and
lymphocyte numbers. Moreover, an increasingly penneable gut barrier may ailow leakage
of p h m a proteins and nutrients which may further compromise gut-associated Unmunity.
In addition io the O bserved morpho lo gical defects, reseuch has documented
irnpaired functional capacity of gut-associated immunity in pro tein-restricted animals.
B lastogenic responsiveness of lymphocytes from M W and Peyer's patches to T cell
mitogens (PHA and ConA) was depressed over t h e in mice fed 0.03% protein diet (Deitch
et al., 1992). Although B cell responsiveness to mitogen is rehtively retained in protein-
deficient mice (Deitch et al.. 1992). protein restricted diets (2 to 4% protein) have k e n
associated with reduced intestinal IgA concentrations (Lim et al., 198 1; McGee &
McMurray, 1988; Sullivan et al.. 1993). impriired dinerentiation of IgA' precursors in
Peyer's patches ta a mature IgA-expressing B ceil phenotype (Lopez & Roux . 1989). and
dimùiished ability of gut lymphocytes to mediate mtibody-dependent ceil-mediated
cytotoxicity ngainst SRBC (Lim et al.. 1981). Recent reports show that the reduced
intestinal I ~ A concentrations during protein-energy malnuvition may not result fiom
impaired IgA synthesis but more Likely fiom decreased transport of IgA precursors to the
intestinal epithelium (Ha et aL. 1996; Ha & Woodward, 1997). The devance of these
defecu in gut-associated IgA and rnitogen-stimulated proliferative responses to helminth
section is not cleu. dthough decreases in the intestinal IgA+ B cell pool mriy lower the
potential for nemntode-elicited production of IgE- and IgGl-expressing B cek in GALT.

Another mechanism by which protein deficiency may contribute to impaired host
immunity to intestinal parasites is altention of the homing patterns of mucosal lymphocytes.
McDermott et al. (1982) transferred radiohbded MLN lymphobhts prepared from well-
nourished donor rats to recipients with or without protein-calorie malnutrition and found
that radioactivity in small and h g e intestines did not differ among normal or mahourished
rats. In conuast, radiolabekd MLN lymphobhu fiom malnourished donors inje!cted into
well-fed recipients resulted in decreased radioactivity recovered in s m d and large
intestines. The authors concluded h3t protein-calorie mdnutrition did not alter the
capacity of the gut to take up mucosd lymphocytes but did impair the selective intestinal
locûliz;ition of GALT-derived lymphobiasts. Although these preliminq data do not
provide any information regarding molecuIar or ceiiular mechmkms, the authors specuiated
that protein deficiency may have modified the function mdlor structure of lymphocyte
horning receptors that mediate lymphocyte binding to the endothelium of HEV in gut
mucosd tissues. Composition of the diets and the degree of deprivation induced in the
recipient rats were not specifed and therefore these results are diffcult to hterpret and to
generalize to systems involving both rnûlnutrition and intesthai parasitic infection.

6. EFFECT OF PROTEIN DEFICIENCY ON IMMUNE RESPONSES TO INTESTINAL NEMATODES INCLUDmG H. POLYGYRUS
Given that T helper lymphocytes are cnicial for antibody responses. depressed T
lymphocyte numbers, impaired functiond capacity of CM1 responses. and defects in homing
of gut-derived T lymphocytes have important implications for Th2-mediated humorai
immunity to intestinal nematode pansites. It is premature. however, to conclude that these
changes o bserved in blood and lymphoid organs of naive subjects interfere with host
immunologicd responsiveness to helminths, especidly considering that the impact of
protein deficiency on host protection depend on the proliferative and functional capacity of
Th2 cells. As discussed previously, changes in humoral responses are not chuacteristic of
protein malnutrition in uninfected humans. Malnourished children have k e n reported to
have elevated antibody titres in blood (Ozkan et al.. 1993) and elevated totd IgE ievek
(Abbassy et al.. 1974) while other studies found no relationship between malnutrition and
IgE levels (Punïio et al., 1976). However. these results on total antibody levels do not
reflect antigen-specific responses during protein malnutrition. Hagel et al. (1995) reponed
that Ascaris-infected cliildren h d increased tord serum IgE but also lower anti-Ascaris IgE
concenuations. This stud y sho wed that mainu trition may enhance polyclonal IgE synthesis
induced by the parasite and in tum dirninish the ability to mount specifk anti-parasite IgE
responses that are important in host resistmce to helminthic infections.
In another study thet is relevant to anthelmirithic immunity, Rickard and Lagunoff
(1989) found that a 0.5% protein diet reduced the nurnber of IgE receptors on peritoned
mat ceb of naive rats but did not affect the nurnber of mut c e k nor the histamine content
per ceL The authors suggested two possibüities for these results: 1) protein deficiency is
more detrimental to IgE receptor synthesis than to synthesis of pro teins necessary for m u t
ceiî or histamine production andor 2) decreased levels of circulating IgE down-regulated
the expression of mast ceiî receptors for IgE. The kst explmation requires ihat pro tein
malnutrition does not cause generalized hypoimmunoglobuluiemia. however, the htter is
eqiially plausible in individu& whose puasite-specinc IgE synthesis is impaired by protein
restriction (Hagel et aL, 1995). Interestingly. Chen & Enerback (1994 & 1996) found that

the density of FccRI receptors and binding of IgE to FceRI on peritoneal m u t ce& of
naive or N. brasiliensis-uifected rats were independent of T lymphocytes as demonstrated
by the lack of differences in these panmeters between athymic and euthymic rats. In
athymic rats which lack T lymphocytes. IgE synthesis may be induced by Th2 cytokines and
cell contact signals rebased from mut ceUs and basophüs (Gauchat et al., 1993; Huek et
a l , 1995). However, the IgE-mediated histamine reiease by mut c e k in athyrnic rats was
signincantly lower compared with that of the normal rats despite similar leveis of semm 1%
and IgE receptors. Thus, interaction with T ceils and their medktors may be necessary for
activation and degranulation of mast cells. The potential for protein deficiency to impair
the synthesis of IgE receptor and the secretory capacity of MMC by depressing Th2
proiif'ntion and cytokine production rem& to be investigated.
In one study that exiunined gut-associated immunological outcomes of protein
deficiency during nematode infection, Cummins et al. (1987a & 1987b) found that a 66
protein diet venus a 208 protein diet impaired worm expulsion and decreased the number
of intestinal MMC but did not affect semm leveh of rat mucosal mu t ceIî protease II
(released upon MMC degranulation) in rats infected once with N. brasiliensis. Thus.
increased worm s u ~ v a l in the protein-deficient rats rnay be attributed to depressed MMC
proliferation rather than to defective mast ceil activation. It is possible that the rehtively
rmld level of pro tein deficiency (6% protein ) was no t conducive to determining the impact
of protein malnutrition on histamine releasibility of mast ceh. Also. protease
concentrations are an indirect meuure of MMC activation and may be infiuenced by the
rate of protease turnover in blood. Given that cytokine production w u not measured in
these studies, it is unknown whether protein deficiency down-reguiates the growth and
maturation of MMC by decreasing the production and release of cytokines from Th2 and
other immune ceils. These hdings of diet-relnted impairment of intestinal immunity have
obvious implications for the capacity for tesistance to nematodes in mahourished hosu.

Surprisingly, there are few studies that have examined whether protein d e f i n c y
potentiates chronicity of infection with nematode puasites by impairing Th2 immune
responses. A low pro tek diet (4%) increased the establishment. survival and egg output of
T. muris in mice (Michael & Bundy. 1991) compared with a 16% protein diet. but this was
not associated with defects in humoral immunity as indicated by the normal to elevated
serurn titres of pansite-specific IgGl and IgA in the protein deficient group (Michael &
Bundy, 1992a & 1992b). These fhdings were interpreted by the researchers as evidence
that protein deficiency is detrimentai to irnmunological processes such as CMI or intestinal
barrier function but has little impact on systernic antibody-mediated responses. Studies
using the mouse-H. polygyms system have produced more conclusive data showing the
detrimental effecr of protein malnutrition on Th2 irnmunity during nematode infection.
Slater and Keymer (1988) fed mice a 2% or 16% protein diet and then chalienged the
animals with H. polygym. The worm burdens at day 21 post-challenge infection were
significantly higher in mice fed the low protein diet cornpared to those recovered in the
protein sufficient mice. The blood eosinophil response was signincantly deiayed and
reduced in the protein deficient rnice over the chdlenge infection period. The rate of
increase in parasite-specific IgG response was signüicantly reduced in the protein deficient
mice while absolute concentrations in IgG did not dSer between the diet groups. It is
unciear whether this slower omet of IgG response may account for the lower level of
acquired protection observed in these animais because changes in total IgG response may
not necessarily correspond with that of subset IgG 1 response which is known to be mediate
protective immunity against H. polygynrr. Consistent with these hdings by Shter and
Keymer (1988). p u t experirnents in Our Lbontory showed that mice fed a low (3%)
protein diet had significantly higher w o m burdens and higher net egg production at 4
weeks post-chalIenge infection compued to mice fed 24% protein (Boulay et al., 1998).
The hck of functional resistance to challenge infection in these pro tein-deficient rnice was
related to their lower periphenl eosinophiüa and lower senun total IgGl concentrations
observed throughout the c W n g e infection process. Considering that Shter and Keymer
(1988) and Bouiay et aL (1998) did not memure production of other Th2 immune effectors

such as IgE, intestinal eosinophils. MMC and specific Th2 cytokines. there are still major
gaps in out understanding of the s pecific immunologicai mechanisms underlying the
interrelationship between protein deficiency and challenge infection with H. polygyms. In
particular, we know very Little about how pro tein deficiency aEects the development of
protective irnmunity to secondary exposures to H. polygyncr in GALT. It is obvious from
the limited scope of current ïiterature that much more research on protein deficiency and
specific Th2 immunity to helminth infections is needed. Our study integrated dl t h e
conditions (ie. protein deficiency, intestinal nematode infection, and host irnmunity) in a
single experirnentai host-pansite system with particular focus on them systernic and
intestinal immun010 gical mechIlliiSms mediating the interac rions between pro tein
malnutrition and intestinal parasitism.

REFERENCES
Abbott, W.C., Tayek, J.A., Bistrian, B A , Maki, T., Ainsley, B.M., Reid, L.A., & Blackburn, G.L. (1986). The effect of nutritional support on T-lymphocyte subpopulations in protein calorie malnutrition. i. Am. Cokge Nutr. 5: 577-584.
Abe, Te & Nawa, Y. (1988). Wonn expulsion and mucosal mst ceil response induced by repetitive iL-3 administration in Strongyloides rani-infected nude rnice. Immunology 63: 181-185.
Abe, Te, Sugaya, He, Ishida, K., Khan, W.L, Tasàemir, I., & Yoshimura, K. (1993). Intestinal protection against Strongyloides roM' and mastocytosis induced by administration of interkukin-3 in mice. Immunology 80: 1 16- 12 1.
Abitorabi, M.A., Mackay, CR., Jerome, E.H., Osorio, O., Butcher, E.C., & Erie, D.J. (1996). DSerential expression of horning molecules on recùcuhting lymphocytes from
sheep gut. peripheral. and lung lymph. J. ImmunoL 156: 3 1 1 1-3 1 17.
Allen, J.E. & Maizels, RM. (1996). Immunology of human helminth infection. Int. Arch AUergy & IrnmunoL 109: 3- 10.
Ali, N.M.H. & Behnke, J.M. (1985). Observations on the gross changes in the secondary lymphoid organs of mice Uifected with Nematospiroides dubius. J . Helminth. 59: 167-174.
Arizono, Na, Kasugai, Ta, Yamada, M., Okada, M., Morimoto, M., Tei, H., Newlands, G.F.J., Miller, H.R.P., & Kitamura, Y. (1993). Infection of Nipposnongylus brusiliemis induces development of mucosal-type but not comective tissue-type m a t cells in gene tically mat ceii-deficient Ws/Ws nts. Blood 81:2572-257 8.
Arizono, Ne, Yamada, M., Tegoshi, Ta, Okada, M., Uchikawa, R, & Matsuda, S. (1994). Mucosal m u t ceîi proliferation folIowing normal and heterotopic îdections of the nernatode Nippostrongyius brasiliensis in nu. APMIS 102: 589-596.
Abbassy, AS., Badd ECDin, M.K., Hassan, A L , Aref, G.H., Hammad, S.A., El- Araby, LI., & Badd ECDin, A.A. (1974). S tudies of ceU-mediated immunity and akrgy in protein energy malnutrition. II. Immedkte hypersensitivity. J. Trop. Med. & Hyg. 77: 18-21.
Banks, E.M.S., & Coleman, LW. (1996). A comparative study of peritoneal mut cells fiom mutant IL4 deficient and normal mice: evidence that IL4 is no t essential for mast ce1 development but enhances secretion via control of IgE binding and passive sensitization. Cytokine 8: 190-196.

Bartiett, A. & Bdl, P.A. J. (1972). Nematospiroides dubius in the mouse as a possible model of endemic human hookworm infection. Am. Trop. Med. P~asitoL 66: 129- 134.
Bawden, RJ. (1969). Some effects of the diet of mice on Nematospiroides dubius (Nematoda). Parasitology 59: 203-2 13.
Beagley, &W. & Elson, C O . (1992). Ceh and cytokines in mucosal immunity and inflammation. GastroenteroL Clin. N.A 21 : 347-365.
Befus, A.D., Dyck, No, Goodacre, R., & Bienenstock, J. (1987). Mast ceUs fiom the human intestinal lamina propria. Isoiation. hisrochernical subtypes and functional characterization. J. Imrnunol. 138: 2604-26 10.
Behnke, J.M. & Wakeün, D. (1977). Nematospiroides dubius: stimulation of acquked imrnunity in inbted strains of mice. J. Helminthol 51: 167-176.
Behnke, J.M. & Parish, HA. (1979). Expuision of Nemotospiroides dubius nom the intestine of mice treated with immune serum. Parasite Imrnunol 1: 13.
Behnke, J.M.9 Hannah, J., & Pritchard, D.I. (1983). Nemurospiroides dubius in the mouse: evidence that adult w o m depress the expression of homologous immunity. Parasite Immuno 1. 5: 397-408.
Behnke, J.M. & Robinson, M. (1985). Genetic control of imrnunity to Nematospiroides dubius: a 9-day anthelminthic abbreviated inununiMg regime which separates weak md strong responder strains of mice. Parasite Immun01 7: 235-253.
Behnke, J.M. & Wahid, RN. (1991). Immunological relationships during primary infection with Heligmsomoides polygynis (Nematospiroides dubius): H-2 linked genes determine w o m survival. Parasitolo gy 103: 157- 164.
Behnke, J.M., Cnbaj, W., & Wakeün, D. (1992). Susceptibility of adult Heligmsomoides polygyms to intestinal infiammatory responses induced heterologous infection. Int. J. PmsitoL 22: 75-86.
Blnke, JeMo, W W , F.N.9 Gren&, RK., Else, KJo, Ben-Smith, A.W., & Goyd, P.K. (1993). Immunological relationships during primary infection with Heligmosomoides polygyrics (Nema~ospiroides drtbius) : do wnreg ulation of speciûc cytokine secretio n (IL-9 and IL-IO) correlates with poor mastocytosis and chronic swvival of adult w o m . Parasite ImmunoL 15: 415-421.
Bell, E.R, Sparshott, SoM., & Bunce, C. (1998). CD4+ T-cell memory, CD45R subsets and the penistence of antigen - r unifjing concept. ImmunoL Today 19: 60-64.

Bell, RG.9 Hazell, L.A., & Mce, P. (1976). Muence of dietary protein restriction on immune cornpetence II. Effect on lymphoid tissue. C h Exp. ImmunoL 26: 3 14-326.
Berlin, Cm, Berg, ILLw, Briskin, M. J., Andrew, D.P., Kilîhaw, P.J., Holzmann, B., Weisman, LL., Hamann, A., & Butcher, E.C. (1993). a4P7 integrin mediates Iymp hoc yte binding to the mucosal vascular addressin MadCAM- 1. Cell74: 1 8 5.
Bises, Ge, Mengheri, E., & Goetad, S. (1987). T-lymphocyte subsets in zinc deficient and in protein mahourished rats. Nutr. Rep. Int. 36: 1371-1378.
Bissonette, E.Y., Chin, B., & Befw, A.D. (1995). Interferons dinerentially regdate histamine and TNF-cr in rat intestinal mucosal mast ceiis. Immunology 86: 12-17.
Bogen, S.A., Weinberg, DaS., & Abbas, AwKo (1991). Histologic analysis of T lymphocyte activation in reactive lymph nodes. J. Immunol. 147: 1537- 154 1.
Boulay, M., Scott, M.E.9 Conly, S.& Stevenson, M.M., & Koski, K.G. (1998). Dietvy pro tein and zinc restrictions independently modw a Helig msomoides polygyrus (Nematoda) infection in mice. Puasitology 116: 449-462.
Bradley, LM*, Yoshimoto, K., & Swoin, S.L. (1995). The cytokines IL-4. EN-y, and L 1 2 regulate the development of subsets of mernory effector helper T celis in vitro. J. ImmunoL 155: 1713-1724.
Bradley, Lw, M., Dalton, DeK., & Croft, M. (1996). A direct role for IFN-y in reguhtion of Th1 cell development. J. IrnmunoL 157: 1350-1358.
Brailslord, T J. & Mapes, C. Je (1987). Cornparisons of Heligmosomoides polygyrus primary infiection in protein-deficient and well-nourished mice. P~asitology 95: 3 1 1-32 1.
Bryant, V. (1973). The life cycle of Nematuspiroides dubius. Baylis 1926 (Nematoda: Heligmosomidae). J. Helminthol 47: 263-268.
Bunce, C. & Bell, E.B. (1997). CD45RC isoforms dehe two types of CD4 memory T cek. one of which depends on persisting antigen. J. Exp. Med. 185(4): 767-776.
Bundy, DeAmPm and Golden, M.H.N. (1987). The impact of host nutrition on gastrointestinal helminth populations. Pansitology 95: 623-635.
Cebm, JJ. & S h d , K.E. (1994). Peyefs patches as inductive sites for IgA cornmitment. In: (Eds. P.L. Ogn, W. Stroker, J. Mesteckey. et al.), Handbook of Mucosd Immunology. New York: Academic Press Inc.

Chan, S.H., Penissia, B., Gupta, J.W.9 Kobyashi, M., Pospisil, M., Young, H.A., Wolf, S.F., Young, D., Clark, S.C., & Tdnichieri, G. (1991). Induction of interferon y production by natural killer ceil stimulatory factor: characterhtion of the responder celis and synergy with other inducers. J. Exp. Med. 173: 869.
Chandra, R.K. (1983). Nutrition. irnmunity. and infection: present knowledge and hture directions. The Lmcet 1: 688-69 1.
Chen, XJ. & Enerback, L. (1994). IgE receptors, IgE content and secretory response of mast cells in athyrnic rats. Immunology 83: 595-600.
Chen, X J. & Enerback, L. (1996). Immune response to a Nippostrongylus brasiliensis Lifection in athymic and euthymic rats: sudace expression of IgE receptors. IgE occupancy and secretory ability of mast cells. Int. Arch. Allergy & IrnmunoL 109: 250-257.
Chin, Y.H., Cai, J.-P., & Xu, Xe-M. (199 1). Tissue-specific homing receptor rnediates lymphocyte adhesion to cytokine-stimulated lymph node high endothehl venule ceb. Immunology 74: 478-483.
Chu, NA, DeBenedette, M.A., Stiernholm, J.N., Barber, B.H., & Watts, T.H. (1997). Role of IL,- 12 and 41BB ligand in cytokine production by CD28+ and CD28- T ceils. I. ImmunoL 158: 3081-3089.
Coffman, R.L., Seymour, B.W., Hudak, S., Jackson, J., & Rennidr, D. (1989). Antibody to IL-5 inhibits heiminth-induced eosinophilia in rnice. Science 245: 308-3 10.
Conzen, S.D. & Janeway, C.A. (1988). Defective mtigen presentation in chronically protein-deprived mice. Irnmunology 63: 683-689.
Cox, F.E.G. & Liew, E.Y. (1992). T-ceU subsets and cytokines in pyasitic infiections. ParasitoL Today 8: 371-375.
Crompton, D.W.T. (1986). Nutritional aspects of infection. Trans. R Soc. Trop. Med. & Hyg. 80: 697-705.
Cumniins, A.G., Kenny, A.L., Duncombe, V.M., Boün, T.D., & Davis, A.E. (1987a). The effect of protein deficiency on systemic reiease of rat mucosd mast ceii protease II during Nipposnongylus brusiliensis infection and foilowing systemic anaphyW. IrnmunoL CeU BioL 65( Pt 4): 357-363.
Cummins, A.G., Duncombe, V.M., Boiin, T.D., Davis, A.E., & Yong, J. (1987b). The respoase of the s d intestine of the pro tein-deficient rat to infection with Nippostrongylus brmiIiensis. fnt. J. ParasitoL 17: 1445- 1450.

Dehlawi, M.S., Wakelin, Dm, & Behnke. (1987). Suppression of mucosal mastocposis by infection with the intestinal nematode Nemutospiroides dubius. Pansite Immun01 9: 187- 194.
Dehlawi, M.S. & Wakeün, D. (1988). Suppression of mucosal mastocposis by Nematospiroides dubiur results from an adult wom-mediated effect upon host lymphocytes. Parasite ImmunoL 16: 85-95.
Mtch, E.A., Xu, Dm, Spdan, RD., & Berg, RD. (1992). Protein malnutrition done and in combination with endotolcin impairs systemic and gut-associated imrnunity. P E N 16: 25-3 1.
Ding, Lm, Linsley, P.S., Huang, Lm-Y., Germain, R.N., & Shevach, E.M. (1993). IL10 inhibits macrophage costimulatory activity by selectively inhibithg the up-regulation of B7 expression. J. IrnmunoL 151: 1224.
Dobson, C., Sitepu, P., & Brindley, P.J. (1985). Muence of primary infection on the population dynamis of Nematospiroides dubius after challenge infections in rnice. Int. J. ParasitoL 15: 353-359.
Dubucquoi, S., Desreumaux, P., Janin, A., Klein, O., Coldman, M., Tavernier, Je, Capron, A., & Capron, M. (1994). Interleukin 5 synthesis by eosinophüs: association with granules and immunoglobulin-dependent secretion. J. Exp. Med. 179: 703-709.
Dugas, B., Renauld, J.C., Pene, J., Bonnefoy, J.Y., Peti-Frere, C., Braquet, P., Bousquet, J., Van Snick, J., & Menaa-Huerta, J.M. (1993). Interleukin-9 potenthtes the interleukin-4-induced irnmunoglo bulin (QG, IgM and IgE) production by nomai human B lymphocytes. Eur. J. ImmunoL 23: 1687- 1692.
Duncombe, V.M., Boün, T.D., Davis, A.E., & Kelly, J.D. (198 1). Dehyed expulsion of the nematode Nippmtrongylus brusiliensis fiom rats on a low protein diet: the role of a bone marrow derived component. Am. J. Clin. Nutr. 34: 400-403.
Eklund, K.K., Ghildyal, N., Austen, K.F., & Stevens, R*L. (1993). Induction by L-9 and suppression by L-3 and IL-4 of the be l s of chromosome lbderived transcripts that encode late-expressed mouse mast ce1 proteases. J. ImmunoL 151: 4266-4274.
Else, KJ., Finkelman, F.D., Maliszewski, CR, & Grencis, RK. (1994). Cytokine- mediated regulation of chronic intestinal helminth infec tioa J. Exp. Med. 179: 347-35 1.
Enriqucz, FJ., Cypess, RH*, & W-nt, D.L. (1988). Influence of mimunizing dose and presence or absence of adult wonns on the development of resistance to Nematospiroides dubius challenge infections of mice. J. ParasitoL 74: 409-4 14.

Faulkner, Hm, Renauld, J.-Ca, Van Snlck, j., & Grenas, RK. (1998). Interieukin-9 enhances resistance to the intestinal nematode Tn'churis muris. fnfect. Immun. 66: 3832- 3840.
Femck, D.A., Schrenzel., M.D., Mulvania, T., Hsieh, B., Ferlin, W.G., & Lepper, Ha (1995). Differential production of interferon-y and interleukin-4 in response to Thl- and Th2-stimulating pathogens by y6 T cells in vivo. Nature 373: 255-257.
Finkelman, F.D., Katona, LM., Momann, Tek , & Coflman, R*L. (1988a). IFN-y regdates the iso types of Ig secreted during in vivo humoral immune responses. J. Immun01 lm: 1022- 1027.
Finkelman, F.D., Katona, EM., Urban, J.F., Holmes, J., Ohara, J., Tung, AS., vG Sample, J., & Paul, W.E. (1988b). IL4 is required to generate and sustain in vivo IgE responses. J. ImrnunoL 141: 233XU41.
Finkleman, F.D., Madden, K.B., Cheever, A.W., Katona, LM., Morris, S.C., Gately, M.K., Hubbard, B.R, Gause, W.C., & Urban, J.F. (1994). EfTects of interkukin 12 on immune responses and host protection in mice infected with intestinal nematode parasites. J. Exp. Med. 179: 1563-1572.
Finkelman, F.D., Shea-Donohue, Tm, Goldhill, J., Sullivan, C.A., Morris, S.C., Madden, K.B., Cause, W.C., & Urban, J.F. (1997). Cytokine regulation of host defense against pansitic gastrointestinal nematodes. Annu. Rev. IrnmunoL 15: 505-533.
Fiorentino, D.F., Zlothnik, A., Vieira, P., Mosmmn, T.R., Howard, M., Moore, K.W., & OpGama, A. (1991). IL40 acts on the mtigen-presenting ceii to inhibit cytokine production by Th1 ce&. J. ImmunoL 146: 3444-345 1.
Flo, J. & Massouh, E. (1997). Age-rehted changes of naive and memory CD4 rat lymphocyte subsets in mucosal and systernic lymphoid organs. Dev. & Comp. IrnmunoL 21: 443-53
Gascan, H., Aveisa, G.G., Gauchat, J.F., Van Vlasselae, P., Ronurolo, M.G., Yssel, Hm, Kehry, M., Spits, Ha, & De Vries, JE. (1992). Membranes of activated CD4+ T cells expressing T c d receptor (TCR) alpha beta or TCR gamma delta induce IgE synthesis by human B ceb m the presence of interleukin-4. Eur. J. ImmunoL 22: 1 133- 1141.
Gauchat, J.-Fa, Aenchoz, S., Mazzel, Ga, Aubry, &P., Brumer, T., Blasey, H., Life, P., Talabot, Dm, Fiores-Romo, Lm, Thompson, J., Kishi, K., Butterfield, J., Dahinden, C., & Bonnefoy, &Y. (1993). Induction of human IgE synthesis in B cells by mast cells and basophihi. Nature 365: 340-343.

Gaw, W.C., Urban, J.F., Linsley, P., & Liu, P. (1995). Role of B7 signalling in the differentiation of naive CD4+ T cells to effector interleukin-4-producing T helper cells. ImmunoL Res. 14: 176-188.
Ghidyal, N., McNeil, H.P., Stechschulte, S., Austen, K.F., Silberstein, D., Gurish, M.F., Somerville, L.L., & Stevens, EL. (1992). IL-10 induces transcription of the gene for mouse mast cell protease-l, a serine protease preferentially expressed in rnucosal mast cells of Trichinella spiralis-infected mice. J. ImmunoL 149: 2123-2129.
Ghidyal, N., Friend, D.S., Nicodemus, C.F., Austen, K.F., & Stevens, RL. (1993). Reverslfile expression of mouse mast cell protease 2 mRNA and protein in cultured mast cells exposed to IL-10. J. ImmunoL 151: 3206-3214.
Good, RA. & Lorenz, E. (1 992). Nutrition and cellulv immunity. Int. J. ImmunopharmacoL 14: 36 1-366.
Gounni, A.S., Larnkhloued, B., Ochalal, K., Tanaka, T., Delaporte, E., Capmn, A., Kinet, J.-P., & Capmn, M. (1994). High-affinity IgE receptor on eosinop hils is involved in defence against parasites. Nature 367: 183- 186.
Greenwald, R J., Lu, P., Hdvomn, M. J., Zhou, X., Chen, S.-J., Madden, K.B., Pemn, PJ., Moms, S.C., Finkelman, F.D., Peach, R., Linsley, P.S., Urban, J.F., & Gaw, W.C. (1997). Effects of blocking B7-1 and B7-2 interactions during a type 2 in vivo immune responses. J. ImmunoL 158: 4088-4096.
Guy-Grand, D., Griscelll, C., & Vassalli, P. (1978). The mouse gut T lymphocyte: 3
novel type of T celL Nature, origin, and traffic in mice in normal and graft-versus-host conditions. J. Exp. Med. 148: 1661- 1677.
Ha, C.-L., Paulino-Radne, L.E., & Woodward, B.D. (1996). Expylsion of the humord effkctor cell compartment of both systemic and mucosal immune systems in a weanling murine model which duplicates critical features of human proteinenergy malnutrition. Br. I. NUU. 75: 445-460.
Ha, C.-L. & Woodward, B. ( 1997). Reduction in the quantity of the polymeric immunoglobulin receptor is sufficient to account for the low concentration of intestinal secretory immunoglobulin A in a weanling mouse model of wasting pro tein-energy malnutrition. J. Nutr. 127: 427-435,
Hagel, I., Lynch, N.R., Di Prisco, M.C., Sanchez, J., 8 Perez, M. (1995). Nutritional status and the IgE response against Ascuris lumbricoides in children from a tropical slum. Trans. R Soc. Trop. Med. & Hyg. 89: 562-565.

Haig, D.M., McKee, T.A., Jarrett, E.E.E., Woodbury, R, & Miller, HoRP. (1982). Generation of mucosd mast ceUs is stirnulated in vitro by factors derived from T cells of helminth-infected rats, Nature 300: 188- 190.
Haig, D.M., Jarrett, E.E.E., & Tas, J. (1984). In vitro studies on mast ceU prolifention in Nipposrrongylus brasiliensis infection. Immunology 51: 643-65 1.
Haradlsen, G., Kvale, De, Lien, B., Farstad, LN., & Brandbaeg, P. (1996). Cytokine- regulated expression of E-selectin, interceiiulv adhesion molecule- 1 (ICAM-1). and vascular ceU adhesion molecule- 1 (VCAM- 1) in human intestinal microvascuiar endo thelial ceh. J. IrnmunoL 156: 258-2565,
Hiroi, Ta, Fujhashi, K., McGhee, J.R., & Kiyono, H. (1995). Polarued Th2 cytokine expression by both mucosal y6 and ap T ceb. Eur. J. ImmunoL 25: 2743-2751.
Huang, H., Hu-Li, Je, Chen, H., Ben-Sasson, S.Z., & Paul, W.E. (1997). IL,-4 md L- 13 production in dinerentiated T helper type 2 ce& is not IL,-4 dependent. J. ImmunoL 159: 373 1-3738.
Hueh, C., Germann, Tm, Goedert, S., Hoehn, P., Koelsch, S., Hultner, L., Palm, Na, Rude, E., & Schmitt, E. (1995). Co-activation of naive CD4' T cells and bone marrow- derived mast results in the development of Th2 ceh. Int. h u n o L 4: 525-532.
Hultner, L., Druez, & C., Mwller, J., (1990). Mast cell growth enhancing activity (MEA) is stnicturdly rekted and functionûlly identicai to the novel mouse T ceIl growth factor P40/TCGFIII (Interleukin 9). Eur. J. ImmunoL 20: 14 13- 14 16.
Jacobsen, RH., Brooks, B.O., & Cypess, RH. (1982). Irnmunity to Nematospiroides dubius: parasite stages responsible for md subject to resistmce in high responder (LAFl/J) mice. J. ParasitoL 68: 1053-1058.
Joe!, DoD.9 & Chanana, A.D. (1985). Cornparison of pulmonary and intestinal lymphocyte migrationid patterns in sheep. Ann. NY Acad. Sci. 459: 56.
Kasugai, Te, Tei, H., Oknda, M., Hirota, S., Morimoto, M., Yamada, M., Nakama, A., Arizono, N., & Etmura, Y. (1995). Infection with Nippostrongylus brcrsiliensis induces invasion of mast cefl precursors from peripherai blood to small intestine. Blood 85: 1334-1340.
Katona, LM., Urban, J.Frn, Kmg, S.S., Paul, W.& & Finkelmnn, F.D. (1991). IL4 requirements for the generation of secondq in vivo IgE responses. J. h u n o L 146: 42154221.

Kaubann, S.H. (1996). Gamrnddelta and other unconventional T lymphocytes: what do they see and what do they do? Proc. NatL Acad. Sci. USA 93: 2272.
Kerboeuf, Ds (1 982). Egg output of Heligmosomoides polygyrus (Nematospiroides dubius) in mice infected once ody. Ann. Recherches Veterinaires 13: 69-78.
Kerboeuf, Dm (1985). The effect of host density on the establishment and fecundity of Heligmosomoides polygynrs. Parasitology 91 : 177- 183.
Keymer, A.E. & Tariton, A.E. (1991). The population dynvnics of acquired immunity to Heligmosomuides polygyrur in the laboratory mouse: strain. diet and exposure. Parasitology 103: 121-126.
King, CL., Low, C C , & Nutman, T.B. (1993). IgE production in human helminth infection: ceciprocal interrelationship between IL-4 and IFN-y. J. ImrnunoL 150: 1873- 1880.
King, C.L. & Nutman, T.B. (1993). IgE and IgG subclass reguhtion by IL4 and IFN-y in human helminth infections: assessrnent by B ceil precursor frequencies. J. ImmunoL 151: 458-465.
King, CL., Hakirni, J., Shab, M.T., & Medhat, A. (1995). IL-12 reguiation of parasite antigen-dnven IgE production in human helminth infections. J. ImmunoL 155: 454-46 1.
King, S. J., & Miller, H.RP. (1984). Anaphylactic release of mucosai m u t ceU pro tease and its relationship to gut permeability in Nipposnongylus-primed nu. Immunology 51: 653-660.
Kita, Hm, Weiler, D.A., Abu-Ghazaleh, R., Sandemn, CJ., & Gleich, G.J. (1992). Release of granule proteins from eosinophils cultured with IL-5. J. Imrnunol. 149: 629- 635.
Kopf, M., Le Gros, G., Bachmmn, M., Lamen, M.C., Bluethmann, H., & Kahler, G. (1993). Disruption of the murine ïL-4 gene blocks Th2 cytokine responses. Nature 362: 245-248.
Kocenaga, M., Wang, CH., Bell, RG., Zhu, Dm, & Ahmad, A. (1989). Intestinal immunity to Trichinella spiralis is a propeny of 0x8' 0x22 T helper ceh that are generated in the intestine. Immunology 66: 588-594.
Koyama, K., Tamauchi, Hm, & Ito, Y. (1994). The role ofCD4+ and CD8+ T ceils in protective immunity to the murine nematode parasite Tkhuris muris. Parasite ImmunoL 17: 161-165.

Kuvibidila, S., Yu, L., Ode, Dm, & W d e r , RP. (1993). The immune response in protein-energy malnutrition and single nutrient deficiencies. In: (Ed. D.M. Klurfeld), Human Nutrition - A Comprehensive Treatise. Volume 8: Nutrition and Immunology. New York: Plenum Press.
Lagoo, AS., Eldridge, J.H., Lagoo-Deenadaylan, S., Black, C.A., Ridwan, B.U., Hardy, K. J., McGhee, J.R, & Beagley, K. W. (1994). Peyer's patch CD8+ memory T ce& secrete T helper type 1 and type 2 cytokines and provide help for immunog10buli.n secretion, Eur. J. Immun01 24: 3087-3092.
Laissue, J.A., Chappuis, B.B., Miller, C., Reubi, J.C., & Gebbers, JO-0. (1993). The intestinal immune system and its relation to disease. Digestive Diseases 11: 298-3 12.
Lamont., A.G., Gordon, M., & Ferguson, A. (1988). T lymphocyte fùnction in protein deprived mice. Clin. Exp. Imrnunol. 72: 1 13- 1 17.
Latham, M.C. (1990). Protein-energy rnahutrition - its epidemiology and conuol. J. Environ PathoL ToxicoL & OncoL (JEPTO) 10: 168-180.
Lawrence, C.E. & Pritchard, D.L. (1994). Immune response pro f i s in responsive and nonresponsive mouse strains infected with Heiigmosomoides polygyrus. Int. J. PansitoL 24: 487-494.
Lee, W.-H. & Woodward, B.DO (1996). The CD4KD8 ratio in the blood does not reflect the response of this index in secondvy lymphoid organs of weanling mice in models of protein-energy malnutrition known to depress thymus-dependent irnmunity. J. Nutr. 126: 849-859.
Le Gros, R, Ben-Sasson, S.Z, Seder, R., Finkelman, F.D. & Paul, W.E. (1990). Generation of interleub 4 (IL-4)-producing cek in vivo and in vitro: IL2 and IL-4 are required for in vitro genention of IL-4 producing cells. J. Exp. Med. 172: 921-929.
Lim, T.S., Messiha, N., & Watson, RR. (198 1). Immune components of the intestinal mucosae of ageing and protein deficient mice. Immunology 41: 401-407,
Locksley, RM. (1994). Th2 ce&: help for helminths. J. Exp. Med. 179: 1405-1407.
Lochrniller, RL., Vestey, M.R., & Nash, Da (1992). Gut-associated lymphoid tissue in the Cotton rat (Sigrnodon hispidus) and its response to protein restriction. J. Wiidlife
Lopez, M. del C. & Roug M.E. (1989). Irnpaired dinerentiation of IgA-B ce1 precursors in the Peyer's patches of protein depieted rats. Dev. & Cornp. ImmunoL 253-262.

Lu, P., di Zhou, Xe, Chen, S.& Mwnnan, M., Morris, S.C., Finkelman, RD., Linsley, P., Urban, J.F., & Gause, W.C. (1994). CTLA-4 ligands are required to induce an in vivo interleukin 4 response to a gastrointesthd nematode parasite. J. Exp. Med. 180: 693-698.
Lu, P., Urban, J.F., di Zhou, X., Chen, S.- J., Madden, K., Mooman, M., Npyen, Ha, Morris, S C , Finkelman, F.D., & Gause, W.C. (1996). CD40-mediated stimulation contributes to lymphocyte proliferation. antibody production, e o s i n o p ~ and mastocytosis during an in vivo type 2 response. but is not required for T cell IL-4 production. J. ImmunoL 156: 3327-3333.
Lunn, PG. & Northrop-Clewes, C.A. (1993). Symposium on 'Parasitism and protein and energy metabolism in man and animais. Proc. Nutr. Soc. 52: 101-1 11.
Madden, K.B., Urban, J.F., Ziltener, HmJ*, Schrader, J.W., Finkelman, F.D., & Katona, LM. (199 1). Antibodies to L-3 and IL-4 suppress helminth-indoced intestinal mastocytosis. J. Irnmunol. 147: 1367-139 1.
Maggi, E., Parronchi, P., Manetti, Ra, Simonelli, CD, Piccinni, M..P., Rugiu, ES., De Cadi, M., Rica, M., & Romagnani, S. (1992). Reciprocal reguhtory effects of IFN-y and IL-4 on the in vitro development of human Th 1 and Th2 clones. J. ImmunoL 148: 2142-2147.
Maindi, ES. & McMurray, D.N. (1998). Protein deficiency induces aiterations in the distribution of T-ce1 su bsets in experimental puhonary tu bercuiosis. Infect. Immun. 66: 927-93 1.
Manetti, R, Parronchi, P., GiudiP, MG., Picrinni, M., Ma&, ED, Trinchieri, G., & Romagnani, S. (1993). Naturd killer ceii stimuhtory factor (interleukin 12 (IL-12) induces T helper type (Th1)-specific immune responses and inhibits the development of L- 4-produchg Th ce&. J. Exp. Med. 177: 1199.
McDermott, M.R & Bienenstock, J. (1979). Evidence for a cornmon mucosd immunologie system. Migration of B immunoblasts into intestinal. respiratory, and genitd tissues. J. ImmunoL 122: 1892.
McDemott, MeRa, Mark, DA., Beliis, A.D., Baîiga, B.S., & Suskhd, RM. (1982). Irnpakd intestinal locaiization of mesenteric lymphobhts associated with vitamin A deficiency and pro teindorie malnutrition. Immun010 gy 45: 1-5.
McGee, D.W. & McMumy, D.N. (1988). The effect of protein malnutrition on the IgA immune nsponse in rnice. Immunology 63: 25-29.

Michael, E. & Bundy, D.A.P. (1991). The effect of the protein content of CBNCa mouse diet on the population dynamics of Trichuris m r i s (Nematoda) in primary infection. Parasitology 103: 403-4 1 1.
Michael, E. & Bundy, D.A.P. (19923). Nutrition, immunity and helminth infection: effect of dietary pro tein on the dynamics of the primary antibody response to Trichuris m r i s (Nematoda) in CB AICa mice. Parasite ImmunoL 14: 169- 183.
Michael, E., & Bundy, D.A.P. (199%). Protein content of CBA/Ca mouse diet: relationship with host antibody responses and the population dynamics of Tnchuris muris (Nematoda) in repeated infection. Parasitology 105: 139- 150.
Miller, H.R.P. & Jarrett, W.F.H. (197 1). Immune reactions in mucous membranes. 1. Intestinal mat ceil response during heiminth expulsion in the rat. Imrnunology 20: 277- 288.
Monroy, FaGa, Adam, J.H., East, L J., & Dobson, C. (1989). Excretmy-secretory antigens from adult Nematospiroides dubius. Immunol. Ceil Bi01 67: 1 15- 120.
Monroy, F.G. & Enriquez, FJ. (1992). Heligmsomoides polygyrus: a mode1 for chronic gastrointestind helminthiasis. PansitoL Today 8: 49-54
Moqbel, Ra, Ying, S., Barkans, J., Newman, T.M., Kirnmitt, P., Wakeün, M., Taborda-Barata, Lm, Meng, Q., Comgrn, C a J., Durham, SR, & Kay, A.B. (1995). Identification of messenger RNA for IL-4 in human eosinophils with granule localkation and releue of the translated producr. J. Immunol. 155: 4939-4947.
Mosmann, T.R & Sad, S. (1996). The expanding universe of T-ceil subsets: TM, Th2 and more. Immunol. Today 17: 138- 146.
Nakache, M., Berg, EL., Streeter, P.Ra, & Butcher, E.C. (1989). The mucosal vascuiar addressin is a tissue-specific endothelid cell adhesion molecule for circuhting lymphocytes. Nature 337: 179.
Nakajima, Ha, Gleich, G.J., & Kits, He (1996). Constitutive production of IL-4 and IL- 10 and siimuked production of L-8 by normal peripherd blood eosinophils. J. IrnmunoL 156: 4859-4866.
NaMabendi, LM., Obeidat, W., Russell, RL, Downie, S., Smith, K., & Rennie, MJ* (1995). Gut mucosal protein synthesis meuured using intravenous and intragastric delivery of stable tracer amho acids. Am. J. PhysioL 269: E9960E999.

Needham, C.S. & Lillywhite, J.E. (1994). Immunoepidemiology of intestinal h e h t h i c infections. 2. Immunological correlates with pattern of Trichuris uifection Trans. R. Soc. Trop. Med. & Hyg. 88: 262-264.
Negrao-Coma, D., Adams, L.S., & Bell, R.G. (1996). intestinal transport and catabolism of IgE. J. ImmunoL 157: 4037-4044.
Nesheim, M.C. (1993). Human nutrition needs and p d t i c infections. P~asitology 107 (Suppl): S7-S 1 8.
Nimmanwudipong, T., Cheadle, W.G., Appel, S.H., & Polk, H.C. (1992). Effect of protein malnutrition and irnmunomodulation on immune ceil populations. I. Surg. Res. 52: 233-238.
Noben-Trauth, N., Shultz, L.D., Brombacher, F., Urban, J.F., Gu, H., & Paul, W.E. (1997). An interleukin 4 (IL-4)-independent pathwûy for CD4+ T cell IL-4 production is revealed in IL-4 receptor-deficient mice. Proc. NatL Acad. Sci USA 94: 10838-10843.
Ozkan, Hm, Logun, N., Sasmaz, E., Abacroglu, H., Okuyan, M., & Cevik, N. (1993). Nutrition. immunity and Uifections: T lymphocyte subpopuhtions in protein-energy malnutrition. J. Trop. Ped. 39: 257-260.
Parent, G., Chevaiier, P., Zalles, L., Sevilla, IL, Bustos, M., Dhenin, J.M., & Jambon, B. ( N94). In vitro lymphocyte-differentiating effects of thyrnulin (Zn-FTS) on lymphocyte subpopulations of severely malnourished children. Am. J. Clin. Nutr. 60: 274-8.
Parker, SJ. & Inchley, C. J. (1990a). E d y lymphocytic responses to Heligmosomuides polygym infections in rnice. J. HelminthoL 64: 35-45.
Parker, SJ. & Inchley, C. J. (1990b). Heligmosomoides polygyncs: infiuence of infection on lymphocyte subpopulations in mouse mesenteric lymph nodes. Exp. ParasitoL 71: 249-258.
Pearce, F.L., Befus, A.D., Gauldie, J., & Bienenstock, Je (1982). Mucosal mast cea. II. Effects of anti-alergic cornpounds on histamine secretion by isohted intestinal mast celis. J. ImmunoL 12%: 248 1-2486.
Penttila, I.A., Ey, P.L., & Jenkin, C.R (1983). Adherence of murine peripheral blood eosinophils and neutrophils to the different parasitic stages of Nematospiroides dubiur. Aust. J. Exp. BioL Med. Sci. 61: 617-627.
Petit-Frem, Cm, Dugas, B., Braquet, P., & Mena-Huerta, J.M. (1993). Interkukin-9 potentiates the interleukin+hduced IgE and IgGl rekase fiom murine B lymphocytes. Immunology 79: 146-151.

Plaut, M., Pierce, J.H., Watson, Cm J., Hanley-Hyde, J., Nordan, R.P., & Paul, W.E. (1989). Mast cell lines produce lymphokines in response to cross-linkage of FcERI or to calcium ionophores. Nature 339: 64-67.
Pleass, R. J. & Bianco, A.E. (1994). The role of adult w o m in suppressing function protective imrnunity to Heligmosomoides po lygym bakeri challenge infections. Parasite ImmunoL 16: 619-628.
Pritchard, D.I., William, D.J.L., Behnke, J.M., & Lee, T.D.G. (1983). The role of IgG l hyperg;unmaglobulinaemia in immunity to the gastrointestinal nematode Nema tospiroides dubius. The immunochemical purification. antigen-s pecincit y and in vivo anti-parasite effect of IgG 1 from immune semm. Immunology 49: 353-365.
Pritchard, D.L, Lawrence, C.E., Appleby, P., Gibb, LA., & Glover, K. (1994). Immunosuppressive proteins secreted by the gastrointestinal nematode parasite Heligmosomoides polygyrus. Int. J. ParasitoL 24: 495-500.
Purtilo, D.T., Riggs, RS., Evans, R., & Nedie, RCw (1976). Humoral immunity of parasitized. mainourished chüdren. Am. J. Trop. Med. & Hyg. 25: 229-232.
Ramaswamy, K., Goodman, RE., & Bell, R.G. (1994a). Cytokine profile of protective yiti -Trichinella spiralis CD4' 0x22' and non-protective CD4+ OX22+ thoracic duct ceils in rats: secretion of IL-4 done does not determine protective capacity. Parasite Imrnunol. 16: 435-445.
Rarnaswamy, K., Hakirni, J., & Bell, R.G. (1994b). Evidence for an interleukin-4- inducible immunoglobulin E uptake and transport mechuiism in the intestine. J. Exp. Mcd. TM: 1793-1803.
Ramaswamy, Kw, Negrao-Coma, D., & Bell, R (1996). Local intestinal immune responses to infections with Trichinella spiralis. J. ImmunoL 156: 4328-4337.
Raulet, D.H. (1989). The structure, Function. and molecul;ir genetics of the gammddelta T ce1 receptor. Annu. Rev. ImmunoL 7: 175.
Reed, N.D., Dehlawi, M.S., & Wakelin, D. (1988). Medium conditioned by spleen cek of Nematospiroides dubius-infected mice does not support development of cultured m a t ceUs. Int. Arch. AUergy & AppL Immun01 85: 1 13- 1 15.
Regoii, M., Borghesi, C., BerteIli, E., & Nicoletti, C. (1994). A morphological study of the lymphocyte trattic in Peyer's patches d'ter an in vivo antigen stimulation. Anat. Rec. 239: 47-54.

Rennidc, De, Hunte, B., Hollmd, Ge, & Thompson-Snipes, L. (1995). Cofactors are essential for stem ceil factor-dependent growth and maturation of mast ceil progenitors: comparative effects of interleukin-3 (IL-3). IL-4. IL-10. and fibroblasts. Blood 85: 57-65.
Rickard, ADLm & Lapnotr, Dm (1989). Protein malnutrition: effect on rat peritoned mast cell number, histamine content. and IgE receptors. Am. J. C h . Nutr. 49: 641-645.
Rincon, M., Anguita, J., Nakamura, Te, Fikrig, E., & Fiavell, RA. (1997). Interleukin a ) - 6 directs the differentiation of IL-4-producing CD4+ T cells. J. Exp. Med. 185: 46 1 - 469.
Roberts, K. & Kilshaw, P. JD (1993). The mucosal T cell integrin aM290P7 recognizes a ligand on mucosai epitheiid ce1 lines. Eur. J. ImmunoL 23: 1630.
Robinson, M. & Gustrid, T+R. (1995). In vitro stimulation of naive mouse lymphocytes by H e l i g m s o m i d e s polygyms adult worm antigens induces the production of IgG 1. Parasite IrnmunoL 18: 87-93.
Robinson, M., Gustad, TDRD, & Meinhamit, S. (1997). Non-specific binding of mouse IgGl to Heligmsomoides polygyrus: parasite homogenate cm riffinity purify mouse monoclonal antibodies. Parasitology 114: 70-84.
Rosenberg, I.H. & Bownman, B*B. (1984). Impact of intestinal parasites on digestive fiinction in humans. Fed. Proc. 43: 246-250.
Saito, Ho, Kanamori, Y., Takemori, T., Nariuchi, Hm, Kubota, E.9 Takahashi- Iwanaga, Iwanaga, Tm, & Ishikawa, Ha (1998). Genention of intestinal ceh f?om progenitors residing in gut cryptoprtches. Science 280: 275-278.
Salmi, M. & Jalkanen, S. (1992). Regdation of lymphocyte tmftïc to mucosa-associated lymphatic tissues. GastroenteroL Clin. N.A 20: 495-510.
Schleimer, R.P, Sterbinsky, S.A., Kaiser, J., Bickel, C.A., Klunk, D.A., Tomioka, K., Newman, W., Lusrinskas, F.W., Gimbmne, MrnA.9 Mcïntyre, B.W., & Bochner, B.S. (1992). IL4 induces adherence of human eosinophils and basophüs but not neutrophils to endo theliurn. Association with expression of VCAM- 1. J. ImrnunoL 148: 1086- lO92.
Schweighoffer, Ta, Tanaka, Y., Tidswell, M., Ede, DJa, Horgan, KJ., Luce, G.E., L m v i t s , A o L , Buck, Dm, & Shaw, S. (1993). Selective expression of integrin a4P7 on a subset of human CD4' memory T cek with hallmarks of gut-uophism. J. Immun01 151:717.
Scott, M.E. (1990). An experimental theoreticai study of the dyndcs of a mouse- nematode (Heligmsomoides polygynis) interaction Parasitology 101: 75-92.

Scott, M.E. & Tanpay, G.V. (1994). Heligmosomoides polygyrus: a iaboratory mode1 for direct Me cycle nematodes of humans and livestock. In: (Eds. M.E. Scott & G. Smith), Pansitic and Xnfectious Diseases. pp. 279-300. New York: Academic Press hc.
Scrimshaw, N.S., Taylor, C.E., & Gordon, J.E. (1968). Interactions of Nuvition and Infection. WHO Monognph Series No. 57: Geneva,
Seder, RA., Plaut, M., Barvieri, S., Urban, J., Finkelmm, F.D., & Paul, W.E. (1991). Mouse splenic and bone marrow celi populations hat express high aEfÏnity Fc epsilon receptors and produce interleukin 4 are highly enriched in basophils. Proc. NatL Acad. Sci USA 88: 2835-2839.
Seder, RA., Paul, W.E., Davis, M.M., Fazekas, D.S., & Groth, B. (1992). The presence of interleukin-4 duMg in vitro priming determines the lymphokine producing potential of CD4+ T cells from T ceii receptor transgenic mice. J. Exp. Med. 176: 109 1.
Seder, RA., Gdnel l i , R, Sher, A., & Paul, W.E. (1993). IL- 12 acts directîy on CD4+ T ceils to enhance priming for IFN-y production and diminish IL-4 inhibition of such priming. Proc. Nati. Acad. Sci USA 90: 1018840192.
Shanahan, F., Lee, T.D.G., Denburg, J.A., Bienenstock, J., & Befus, A.D. (1986). Functional chuacterization of mast cells generated in vivo fiom the rnesentenc lymph node of rats infected with Nippostrongylus brasiliensis. Immunology 57: 455-459.
Shirnoda, K., van Deursen, J., Snagster, M.Y., Sarawar, S.R., C~rson, RT., Tripp, RA., Chu, C., Quelle, RW., Nosaka, T., Vignali, D.A.A., Doherty, P.C., Grosveld, DG., Paul, W.& & Dile, J.N. (1996). Lack of IL-4-induced Th2 response and IgE clas switching in rnice with disrupted Stst6 gene. Nature 380: 630-633.
Slater, A.F.G. (1988). The influence of dietvy protein on the experimental epiderniology of Heiigmosomoides polygyrus (Nematoda) in the iabontory mouse. Proc. R. Soc. Lond. B 234: 239-254.
Slater, A.F.G. & Keymer, A.E. (1986). Heligmosomides polygyw (Nematoda): the influence of dietvy protein on the dynamics of repeated infection. Proc. R Soc. Lond. B 229: 69-83.
Slater, A.F.G. & Keymer, A.E. (1988). The influence of protein deficiency on immunity to Heligmosomoides polygyrur (Nematoda) in Mce. Parasite ImmunoL 10: 507-522.
Slobodianik,N.H.,Cos~nsky,RC., Langini,S.H., Roux, M.E., Rio,M.E.,& Sanahuja, J.C. (1984). Effect of severe protein deficiency on surface and intracelluiar markers of growing rats' lymphoid organs. Nuu. Rep. Int 29: 957-964.

Smith, N.C. & Bryant, C. (1986). The role of host generated free radicals in helminih infections: Nippostrongylus brdiensis and Nemutospiroides dubius compared. Int. J. ParasitoL 16: 6 17-622.
Smith, T J. & Weis, J.H. (1996). Mucosai T cells and mast cells share cornmon adhesion receptors. ImmunoL Today 17: 60-63.
Solomons, N.W. (1993). Pathways to the impairment of human nutritional status by gastrointestind pathogens. Parasitology 107 (Suppl): S 19433.
Spear, P. & Edelman, G.M. (1974). Maturation of the humoral immune response in mice. i. Exp. Med. 139: 249.
Sulüvan, D.A., Vaerman, J.-P., & Soo, C. (1993). Influence of severe protein malnutrition on rat lacrimal. salivary and gastrointestind immune expression duMg development. adulthood and ageing. Immunology 78: 308-317.
Svetic, A., Madden, K.B., di Zhou, X., Lu, P., Katona, LM., Finkelman, F.D.9 Urban, J.F., & Cause, W.C. (1 993). A prirnary intestinal helrninthic infection rapidly induces a gut-associated elevation of Th2-associated cytokines and IL-3. J. Imrnunoi. 150: 3434.
Swain, S.L., McKenPe, D.T., Weinberg, A.D., & Hancock, W. (1988). Characterization of T helper 1 and 2 ceU subsets in normal mice: Helper T cells responsible for IL-4 and IL-5 production are present as precursors that require priming before they develop into lymphokine-secreting ceils. J. ImmunoL 141: 344503455.
Swain, S.L., Weinberg, A.D., Engüsh, M., & Huston, G. (1990). IL-4 directs the development o f Th2-W<e helper effectors. J. IrnmunoL 145: 3796-3806.
Takeda, K., Tmaka, Tm, Shi, W., Matsumoto, M., Minami, M., Kashiwamura, S.L, Nakanishi, K., Yoshida, N., Kishimotom, Tq & Akira, S. (1996). Essential role of Stat6 in IL-4 s ignahg Nature 380: 627-630.
Tanaka, T., Hu-Li, J., Seer, R.A., de, S., Groth, B.F., & Paul, W.E. (1993). Interleukin 4 suppresses interleukinZ and interferon gamma production by naive T ceUs stimuhted by accessory cell-dependent receptor engagement. Proc. Nat. Acad. Sci USA 90: 5914.
Thompson-Snipes, L., Mar, V., Bond, M.W., Mosrnann, T.R., Moore, K.W., & Rennick, D. (1991). Interleukm 10: a novel stimuhtory factor for mast ceb and their progenitors. J. Exp. Med. 173: 507.
Thomhill, M.&, Kyan-Aung, U., & Haskard, D.0. (1990). IL4 increases human endotheiiai ceïï adhesiveness for T ceiis but not for neutrophils. J. ImmunoL 144: 3060.

Thomhill, M.H., Welücome, S.M., Mahiouz, D.L., Lanchbury, J.S.S., Kyan-Aung, U., & Haskard, D.O. (1991). Tumor necrosis factor combines with IL-4 and IFN-y to selectively enhance endothelid ceU adhesiveness for T cells. J. Immunol 146: 592-598.
TON, Ha, Ra, C., Nonyama, S., Suniki, K., Yata, Je-Ia, & Nakahata, T. (1996). Induction of the high-dfhity IgE receptor (FcrRJ) on human mast cells by IL-4. Int. ImmunoL 8: 1367-1373.
Uchikawa, Ra, Yamada, M., Matsuda, S., K u d a , A., & Anzono, N. (1994). IgE antibody production is associated with suppressed interferon-y levels in mesentenc lymph nodes of rats infected with the nematode Nipposnongylus brmiliensis. hrnunology 82: 427-432,
Upadhyay, P., Ganply, N.K., Mahajan, R.C., & Walia, BeNaSm (1985). Intesthai uptake of nutrients in normal and mahourished animais infected with Giardia lamblia. Digestion 32: 243-248.
Urban, J.F., Katona, LM., & Finkelman, F.D. (1991a). Heligrnusomides p o l y g y w : CD4+ but not CDS+ T ceb regulate the IgE response and protective imrnunity in mice. Exp. ParasitoL 73: 500-5 1 1.
Urban, J.F., Katona, LM., Paul, W.E., & Finkelman, F.D. (199 1 b). Interleukin 4 is important in protective immunity to a gastrointestinal nematode infection in mice. Proc. Natl. Acad, Sci USA 88: 35 13-34 1%
Urban, J.F., Madden, K.B., Svetic, A., Cheever, A., Trotta, P.P., Cause, W.C., Katona, LM., & Finkelman, F.D. (1992). The importance of Th2 cytokines in pro tective immunity to nematodes. ImmunoL Rev. 127: 205-2 19.
Urban, J.F., Madden, KWBa, Cheever, A.W., Trotta, P.P., Katona, LM., & Finkelman, F.D. (1993). IFN inhibits inflanmatory responses and protective immunity in mice infected with the nernatode parasite, Nipposnongylus brdiensis . I. Immunol. 151: 7086- 7094.
Urban, J.F., Maliszewski, C.R., Madden, K.B., Katona, LM., & Finkelman, FaDe (1995). L-4 treatment can cure established gastrointestinal nematode infections in imrnunocompetent and Unmunodeficient mice. J. ImrnunoL 154: 4675-4684.
Urban, J.F., Fayer, R a , Sulüvan, C., Gofdhill, J., Shea-Donohue, T., Madden, K., Morris, S.C., Katona, L, Cause, W., Ruff, MM., Mansfield, L.S., & Finkelman, F.D. (1996). Local Th1 and Th2 responses to parasitic infection in the intestine: reguhtion by IFN-gamma and 1 . 4 . Vet. ImmunoL & ImmunopathoL 54: 337-344.

de Waal Malefijt, R a , Haanen, Je, Spits, H., Roncarolo, M.-Ga, teVelde, A., Figddor, C., Johnston, K., Kaastelein, Ra, Yssel, He, & deVries, J.E. (1991). Seiection of a humm T helper type 1-lüce T cell subset by mycobacterh. J. Exp. Med. 174: 9 15.
Wade, S., Parent, Ge, Bleiberg-Daniel, F., Maire, B., Fall, M., Schneider, Dm, Le Moullac, B., & Dardenne, M. (1 988). Thymulin (Zn-FïS) activiîy in pro tein-energy malnutrition: new evidence for interaction between malnutrition and infection on thymic function. Am. J. Clin. Nutr. 47: 305- t 1.
Wahid, F.N., Robinson, M., Behnke, J.M. (1989). Irnmunological relationships during primary infection with Heligmosomides polygynrs (Nemutospiroides dubius): expulsion of adult worms from fast-responder syngenetic and inbred strains of mice. P&tology 98: 459-469.
Wahid, F.N. & Behnke, J.M. (1992). Stimuli for acquired resistmce to Heligmsomides polygyms from intestinal tissue resident L3 and LA larvae. Int. J. P~ûsitoL 22: 699-7 10.
Wahid, F.N. & Behnke, J.M. (1993). Immunologicd relationships during primary infection with Heligmusomoides polygyrus (Nemutospiroides dubius): parasite-specifc IgGl antibody responses and primvy response phenotype. Parasite Irnrnunol 15: 401- 413.
Wahid, F.N., Behnke, J.M., Grenas, R.K., Else, KJ., & Ben-Smith, A.W. (1994). Immun010 gical relatio nships during p h y y infection with Heligmosomoides polygyms: Th2 cytokines and primary response phenotype. P~asitology 108: 46 1-47 1.
Warren, &S., Kinnear, B.F., Philüps, J.H., & Lanier, L.L. (1995). Production of IL-5 by human NK ceils and regulation of IL-5 secretion by IL-4. IL- 10. and IL- 12. J. ImrnunoL 154: 51445152.
Wastllng, J.M., Scudamore, C.L., Thomton, E.M., Newlands, G.F.J., & Miller, H.RP. (1997). Constitutive expression of mouse m u t ceil protese-1 in normai BALBk mice and its up-regulation duMg intestinal nematode infection. Immunology 90: 308-3 13.
Wen, L., Pao, W., Wong, ES., Peng, Q., C d , J., Zheng, B., Kelsoe, G., Owen, J. Je, & Hayday, A.C. (1996). Germinal center formation immunogIobulin ciass switching and autoantibody production driven by "non ap" T ce&. J. Exp. Med. 183: 227 1.
Wen, L., Barber, D.F., Pao, W., Song, F.S., Owen, MJ., B Hayday, A. (1998). Primary y6 ce1 clones can be dehed phenotypicdy and functionally as T h l m tek and illustrate the association of CD4 with Th2 differentiation. J. ImmunoL 160: 1965-1974.

Wodd Health Organization (WHO). (1987). Prevention and Control of Intestinal Parasitic Infections. Geneva: World Health O r g h t i o n , Technical Report Series, no. 749.
Wilüams, D J.L. & Behnke, J.M. (1982). Host protective antibodies and semm immunoglobulin isotypes in mice chronically infected or repeatedly immunized with the nematode parasite Nemutospiroides dubiiis. Immunology 48: 37-47.
Windhagen, A., Anderson, D.E., Carrizosa, A., Williams, RE., & Hafler, D.A. (1996). IL- 12 induces human T ce& secreting IL- IO with IFNy. J. ImmunoL 157: 1 12%
W d b u y , RG., Miller, H.R.P., Huntley, J.F., Newlands, G.F.J., Palliser, A.C., & Wakeün, Dm (1984). Mucosal mast ceh are hinctiondy active during spontaneous expulsion of intestinal nemntode infections in nt. Nature 312: 450-452.
Woodward, B.D. & Miller, R.G. (1991). Depression of thymus-dependent immunity in wasting protein-energy miilnutrition does not depend on an altered ratio of helper (CD47 to suppressor (CDg*) T ceh or on a disproportionately iarge atrophy of the T-ceii relative to the B-ceil pool Am. J. Clin. Nutr. 53: 329-335.
Woodward, B., Dwivedi, A., & Nmgpd, A. (1992a). Mechanisms of thymic epitheliai involution in weanüng pro tein-energy mainutrition. Nutr. Res. 12: 1253- 1264.
Woodward, B.D., Woods, J.W., & Crouch, D.A. (1992b). Direct evidence that primvy acquired ceil-mediated immunity is Iess resistant thm is primary thymus-dependent humoral immunity to the depressive Suence of wasting protein-energy malnutrition in weaniing mice, Am. J. Clin, Nutr. 55: 1 180- I 185.
Woodward, B.D., Bezuison, K.D., Hillyer, L.M., & Lee, W.-H. (1995). The CD45RA' (quiescent) celluiar phenotype is overabunduit reiative to the CD45W phenotype within the involuted splenic T ceil popuhtion of weanling mice subjected tow wasting protein-energy malnutrition. J. Nutr. 125: 247 1-2482.
Wykes, L.J., Fiorotto, M., Bumn, DG., Del Rosario, M., Frazer, ME., Pond, W.C., & Jaboor, F. (1996). Chronic low protein intake reduces tissue protein synthesis in a pig mode1 of pro tein malnutrition. J. Nutr. 126: 148 1- 1488.
Yoshimoto, T. & Paul, W.E. (1 994). CD4*M<l. 1+ T ceUs prornp tly produce interleukin- 4 in response to in vivo ch;illenge with mti-CD3. J. Exp. Med. 179: 1285-1295.
Zhang, X., Werner-Fom, Ce, Tang, H., Bmuswers, Ne, Bonnefoy, J.Y., & Zubkr, RH* (1991). IL4dependent IgE switch in membrane IgA-positive humaa B cells. J. IinmunoL 147: 3001-3004.

CHAPTER III STUDY OBJECTIVES
Previous studies have reported independentiy that protein deficiency increases the
prevalence and intensity of experimental nemtode infections and impairs systemic and gut
mucosal immune huiction in naive animais. However, very few studies have examkied the
interactions between all three conditions (Le. protein deficiency, nematode infection and
host irnmuniiy) in the s m e host-parasite system. Inasmuch as host resistance to challenge
H. polygyrus infection depends on intense. npid and sustained Th2 immune responses. it is
important to examine whether the capacity for acquired immunity to intestinal nematodes is
adversely modified by protein malnutrition. The overd purpose of this study was to
detennine whether dietvy protein deficiency potentiates a chdenge Uifection in rnice with
the gastrointestinal nematode, H. polygyrus, by impairing the magnitude. site-specificity
and kinetics of secondary cytokine and effector responses. A the course study of serum
IgE. parasitic-specific IgG1, peripheral and intestinal eosinophils, MMC, MMCP-1, and
cytokine production in spleen and M W was performed from days 3 to 28 pci These
immune responses were &O measured in naive rnice yidlor afker primary section to
compare the ievel of protective immunity acquired during challenge infection.
The k s t specinc objective of this study was to determine the effects of dietary
protein deficiency on systemic md intestinal Th2 immune responses to polygyms
Mection as indicated by the level of serum effecton (eosinophils, IgE, parasite-specinc
IgG l), the production of spleen cytokines (lL-4, IL-5. IL- 10, IFN-y ), the proliferation of
gut mucosd effectors (MMC, intesthai eosinophüs, MMCP- I), and the synthesis of gut-
associated cytokines in MLN. Secondly, we wanted to ascertain whether the seventy of
dietary protein restriction (7% versus 3%) would afTect the extent of impaired Th2
immunity and parasite survivd. The third objective was to compare the impact of protein
deficiency on systemic versus intestinal Th2 immune responses to validate our assumption
that gut mucosal immunity may be more suscepuile to protein mdnutritioa Fiidy, we
intended to characterize and contrast the kinetics of systemic and local Th2 immune
responses during graded leveis of protein dekiency in mice challenged with polygynrs.

PROTEIN DEFICIENCY PROLONGS SURVIVAL OF A GASTROINTESTINAL
PARASITE IN MICE BY SUPPRESSING TH2 IMMUMTY
Rebecca Ing12, Zhong Su', Marilyn E. Scott', Kristine G. Kosk?
'Institute of Parasitology and 'School of Dietetics and Humm Nuuition.
McGill University (Macdonald Campus)
21.1 11 Lakeshore Road, Ste-Anne de Bellevue, Quebec H9X 3V9. Canada
Corresponding Author:
Dr. KG. Koski
School of Dietetics and Human Nutrition
Macdonald Campus, McGilI University
21.1 11 Lakeshore Road
Ste-Anne de Bellevue, Quebec H9X 3V9, Canada
Telephone: 5 14-398-7845
Fsix: 5 14-398-7739
Email: koski@agndiahmcgülca

ABSTRACT
Protein deficiency has been shown to increase susceptibüity to gastrointestind (GI)
parasitic infections in mice, possibly as a result of impaired systemic and intestinal effiector
responses induced by the dom-regulation of Th2 cytokine d o r up-regdation of Th l
cytokine. To test this hypothesis, B ALBlc mice (n= 18/diet/time point) were fed a control
(24%), marginal (7%). or deficient (3%) protein diet and given a challenge infection with
the nematode, Heligmosomides polygyrus. The protein deficient (PD) rnice had higher
food intake per gram of body weight, lower body weight gain, reduced BUN concentration,
and depressed reiative spleen and mesentenc lymph node (MLN) weights compared to the
control and marginal mice. The PD mice had higher worm burdens at 1,2 and 4 weeks
post-change infection (pci), suggesting chronic absence of a fbnctional immune response.
Indeed, the PD rnice h d lower increues in serum IgE, reduced penpheral and intestinal
eosinophilia, and depressed MMC pmliferation and activation at 1 to 2 weeks pci
Parasitic-specinc IgG 1 was not affected by protein deficiency. To determine whether these
suppressed effector responses in the PD rnice may have resulted in piut from aitered spleen
and MLN cytokine pronles, c eh were stimulated Ni vino with parasite antigen and
cytokine concentration in the culture supematmts was measured by sandwich ELISA. The
deficient MLN ceh secreted signincantly las IL4 and more EN-y within 2 weeks pci
than did control MLN cek. The deficient spleen ce& &O secreted more IFNy at 2 weeks
pci compared with control spleen cells. There were no significuit dserences in IL-5 or IL-
10 secretion by MW or spleen c e h among the dietuy groups. Frorn RT-PCR mûlyses,
the PD mice also had lower IL-4 rnRNA expression in spleen and MLN at 1-2 weeks pci
In summary, protein deficiency has detrimental type- and site-specifk effects on cytokine
and effector responses at euly pci time points critical for expulsion of H. polygyrur. Our
data support the hypothesis that protein deficiency exacerbates the suniival of a GI
nernatode parasite by decreasing IL-4 (TM) and increasing IFN-y ml), leading to
reduced gut and systemic Th2 effecto r respo mes.

INTRODUCTION
Protein malnutrition and gastrointestind nematode infections are chronic diseases
that frequently CO-exist in human populations of developing countries (Crompton, 1986).
Acquired resistance to intestinal parasitisrn depends on the magnitude and rapidity of Th2
cytokine (e.g. E-4, IL-5. IL-10) and effector responses ( e o s h o p ~ . IgE, wtocytosis) as
well as the concomitant absence of Th1 responses such as IFN-y and macrophage activity
(Urban et al., 1993; Monroy & E ~ q u e z . 1992; Urbm et ai., 199 1). Although protein
deficiency increases susceptibility to nematode parasites during repeated and challenge
infections (Shter & Keymer, 1988; Keymer & Tarlton. 1991), it rernains unclev whether
protein deficiency potenthtes intestinai parasitic infections by hpaVing the development
and maintenance of specific Th2-dependent immune responses in penpheral andior
intestinal tissues. Earlier studies showed that higher worm burdens in mice fed low protein
dieu were associated with depressed blood eosinophilia and lower titres of cuculating
parasite-speciflc IgG (Slater & Keymer, 1988; Bouhy et al.. 1998). However, it is
unknown whether milder protein restriction has the same effect on Th2 immunity and
whether the irnpaired effector responses resulted from diet-induced perturbations in
cytokine synthesis. Additionally, lit tle Uifomation exists on the CO nsequences of pro tein
deficiency on host protective immunity in gut-associated Iymphoid tissues (GALT) even
though immune responses in GALT comprise the k s t line of defense ag;iinst intestinal
parasites and may influence the appropriateness, proiiferative magnitude and traffïcking of
systemic responses. The rate of protein turnover in gut mucosal tissues is 25-30 times
greater than that observed for muscle (Nakshabendi et al.. 1995) and is potentisilly very
sensitive to changes in host protein status (Wykes et al., 1996).
Our labontory has been involved in studies on the interactions between acquired
immunity, parasitic infection, and nutritional deficiencies in mice infected with a
gastrointestinal mouse nematode, Heiigmosomoides polygynrr. P r i m q infections with H.
polygyncs are usually chronic but immunocompetent hosts are capable of developing
resistance to challenge infections. The capacity to resist secondary infections and the
intesthal site of infection make the polygyrus-mouse system a suitable mode1 for

studying the Muence of nutritionai factors on acquired immunity and gut mucosal immune
responses during gastrointestinal heùninthiasis. Previous experimenu in our labontory
have shown that protein deficiency uicreased worm burdens, decreased serum IgGl and
lowered eosinophilia in mice chdienged with H. p o f y g y w (Boulay et al.. 1998). Gut-
associated Th2 effector responses and cytokine production in a protein deficient host
chaknged with polygyncs have no t k e n chuacterized. The overd objective of this
study was to determine the effects of graded levels of protein deficiency on the magnitude,
site-specincity and time course of Th2 immune responses during challenge infection with H.
pofygynrs. We postuhted that pro tein deficiency prolongs parasite s u ~ v a l by impairhg
host cytokine respo mes in gut-associated ando r systemic lymphoid tissues that normaily
help to resolve a secondary H. polygyms infection. The reduced Th2 cytokine responses
andlor increased T'hl cytokine synthesis may subsequently lead to decreased concentrations
of Th2 effectors in intestinal and systemic sites. Assuming that gut mucosal immunity is
more susceptible thui systemic immunity to protein malnutrition and given its putative role
in priming and directing host resistance, we expected protein deficiency to cause a more
profound impairment of intestinai irnmunity than of systemic responses. Also, we expected
that a low protein diet (3%) would inhibit acquired immunity, and in tum increase
parasitisrn, to a greater extent than would a marginally deficient diet (7%). FimaDy, we
speculated that these effects would diner in intensity at dinerent phases of infection given
that the most intense stimulus for acquired immunity is elicited by tissue-resident Wae at
1-2 weeks post-challenge infection (Wahid & Behnke, 1992).
Our results show that protein deficiency promoted suxvival oEH. p o l y g y w by
altering cytokine and effector responses in a selective mamer depending on the tissue
cornpartment, the level of restriction and the time point of infection. In pyticulu. the host
responses that normaily help to resolve challenge Uifections with H. pofygynrs (increased
IL-4, decreased IFN-y, eievated IgE, mucosal rnastocytosis, MMC activation) were
pro foundiy impaired by pro tein deficienc y. These data support the hypo thesis that pro tein
denciency prolongs sunRvaï of n nematode parasite by decreasing gut-associated I L 4
production and Th2 effector responses in the gut and peripherd circulation.

MATERIALS & METHODS
Experimental Design
Mice were fed a diet that was sufficient (24%). marginal (7%). or deficient (3%) in
protein and given prirnary or chûllenge uifections with H. polygynrs. Penpheral
eosinophih was measured in tail veh blood using the Unopette Test (#5877 Fiher
Scientinc, Montreal. PQ). Worms in smali intestines of infected mice were counted at 6
and 28 days post-primuy infection (ppi) and at 6, 14.2 1 and 28 days post-challenge
infection (pci). Cardiac blood w u separated into serum and plasma sampks and stored at
-80°C in Microtaher tubes (Becton Dickinson, Rutherford, NJ. USA) for iater antibody and
mucosal mast ceU pro tease- 1 (MMCP- 1) analyses. Intestinal tissues were O btained for
histological counting of mucosal mast cells (MMC) and intestinai eosinophils. At 3. 6, 14
and 28 days pci, spleen and rnesentenc lymph nodes (MW) ceii suspensions were
prepared, and their cytokine production and gene expression were determined in vitro.
Expenmental Protocol
The experiment was conducted using an aithelmintic-abbreviated immunizing
protocol that hm been shown to stimulate protective imrnunity against a challenge infection
with H. polygynis (Behnke & Robinson, 1985). niree weeks after feeding with
experimentd diets, mice (6/diet group/time point) were intubated ordy with 100 third-
stage Iarvae (L,, of H. polygynrs (day O ppi); 12 rnice per diet group remained uninfected
(Figure 1). Six days &er primary infection, 6 mice per diet group were iciIIed to connmi the
Uifectivity of L, (P6 rnice). On days 9 and 14 ppi, the remaining infected rnice were trerted
oraily with pyruitel pamoate (Pfker Canada Inc.. Kirkland, Quebec, Canada, dose of 172
mgkg body weight) to remove the adult parasites fkom the intestine. Six mice per dietary
group were LiUed CO ensure efficac y of the dmg ueatment which required an absence of
w o m in the intestine. One week foilowing the second anthelmintic treatment ( d g 2 1
ppi), mice (6ldiet group/time point) were reinfected with 100 L, of H. poiygyn*r (challenge
cohort).

Challenge Cobort - -21 D DO D6 D 9 D l 4 D 21 1 Posi-primary Infection (ppi)
DO D 3 D 6 D 9 D 14 D 21 D 28 Post-challenge Infection (M)
Feeûing differenr diets 100 L, P6 Drug h g 100 L, C3 C6 0 Cl4 C21 C28 Primary Challenge Infeciion In feciion
-21 r) DO D 28 - Posi-primary Infection (ppi)
Feeding different diets 100 L, Primary Infection
Feeding different die& Nai ve mirols
Figure 1: Schematic representation of the experimental protocol and various infection groups. The numbers on top of each line represent tirne course (in days) of the infection protocol. The values below the iine represent the procedures perfomed ai the correspondhg day and infection groups killed at each t h e point.

Two groups of control mice were included in the experiment. One group of previously
uninfected mice (6 per diet group) were given a primary infection with 100 L, at the tirne of
challenge infection and killed at 28 days ppi (P28 mice). These primary infection mice were
used to compare the degree of protection acquired duMg the challenge infection protocoL
A second cohon of mice (6 per diet gmup) remained uninfected throughout the
experimental penod to serve as naive infection controls and were kiüed on day 28 pci
(naive controls). Mice given the challenge infection were H e d at 3, 6.9, 14, 2 1 and 28
days pci and are referred to as C3, C6, Cg, C 14. C2 1 and C28 Mce. respectively.
Mice
Three week old femaie B ALBlc rnice (Charles River, St. Constant, PQ, Canada)
were acchatized to a 14h light-l0h duk cycle in a temperature-controlled (22-25'C)
animai room for 3 days while fed the protein-sufficient diet. Mice were then weighed and
randomly docated to one of the 3 dietary groups. AU experimental diets were offered ad
libitum in plastic Mouse Powder Feeders (Lab Products Inc., Montreal, PQ) specifically
designed to minimize food spihge. Distilled water was rvaiiable to ail animais ad libinim.
Mice were housed individudly in Nalgene cages (Flher Sckntific, Montreai, PQ) with
stainless steel covers, filter tops, and heat-treated hardwood bedding. AU procedures were
approved by the McGill Animai Care Committee according to the Canadian Council on
Animal Care (1984).

Diets
Mice were fed a semi-pudied, biotin-fortified diet with spray-dried egg-white soiids
as the sole source of protein (Bouhy et al., 1998). The 2446 (control) protein level is
considered adequate and not excessive for the hboratory mouse (NRC, 1995). The
protein-restncted diets contained either 7% (marginal) or 3% (deficient) protein and were
made isoenergetic to the 24% protein diet by substitution of protein with equivalent
amounts of comtatch on a weight buis (Table 1). AU other nuvients were identical
among the three diets and were included at levels sufflciently above the NRC (1995) mouse
requirernents to ensure that a 30% reduction in food intake as a result of protein restriction
or infection would not generate any other nuvient deficiencies in the marginal or deficient
protein groups. Biotin was added at 18 times the NRC recommended level to counteract
the biotin-chehting effects of avidin in the egg-white solids.
Parasite and parasite an Agen
Infective third-stage Irirvae (b) of H. polygyms were obtained by culturing the
feces of stock CD1 mice (Charles River) on moist fdter paper for 7 days. The cultured
l ame were suspended in distilled water (approlamately la0 Y20 pl) and adrninistered by
o n l gavage to mice using a Gilson pipetmm (Mandel ScientSc, Guelph. Ontario). The
accuracy of the dose and viabiîity of the iarvae was estimated by counting the number of
live b a e per 20 p1 after every 15-20 intubations.
Parasite antigen was prepared using fourth stage lyvae (LJ of H. p o l y g y m
obtained fiom s m d intestines of mice infected with L, 6-7 days previously. The coiîected
L, were washed with PBS and sonicnted on ice for 10-15 min. The homogenate was
centrifuged at 1500 g at 4°C for 1 hr. The recovered supernatant was sterjlized using a
0.22 pm Acrodisc (Gehui Sciences, AM Arbor, MI, USA) and stored at -20°C. Pro tein
concentration of the antigen homogenate was determined ushg a dye-binding protein assay
(Bio-Rad, Mississauga, Ontario, Canada) with BSA as the standard.

Table 1: Nutrient composition of experimental diets.'
1ngredient2 Pro tein sufficient Pro tein marginal Protein deficient (24%) (7%) (346)
Egg white 240
Corn oil 80
Corn starch 296
Glucose 296
Alphacel 30
Vitiunin mix3 12
Mineral mix' 46
l Ingredients are g/kg diet
ICN Biochemicals. Division of ICN Biocfiemiuis, Inc.
V i m i n mix (mgkg diet) as formulnted with X N Biahemicals & Anachernia Science (Anachemin
Canadû, ïnc.); niacin 50.0; d-calcium panthothenate 32.0: riboflcivin 24.0; pyridoxine bydrochloride 24;
thivnin hydrocbloride 16.0; folic acid 2.0; d-biotin 3.6; cymocobalunin 0.05; a-tocopheryi aceote (1 .O
iU/mg) 96.0; menaquinone 9.0: vitamin 4 (400.W iUlg) 7.5; viiamin A acetate ( 5 0 0 . 0 i ü g ) 8.0;
butylated hydroxytoluene 100.0; choline chIoride 4000.0; alphacel3748.5.
' Minerai mix (gkg diet) as farmulated with ICN Biochemicals. Anachemia Science, Sigma Chernicals
(Sigma Canada), & Fisha Chemids (Fisher Scientific Canada); diuiaum phosphate 27.16; potassium
bicarbaiate 10.25; soâium chloride 2.54; magnesium sulphate 4.95; chromium potassium sulphnte
0.0384; cupric carbonate 0.0157; potassium iodate 0.01; ferrous sulpbate 0.2489; manganous carbonate
0.1883; zinc carbonate 0.115; sodium selenite 0.0004; sodium moiybdate 0.00045; potassium fluoride
0.0099; citric acid 0.48;

Parasite wonn burdens
The numbers of L, parasites were counted at 6 days after primary or challenge
uifection. The nurnbers of adult worrns in chdienged rnice were determined at days 14.2 1
and 28 pci and in the primary group at day 28 post-infection. L, pansites embedded in the
intestinal mucosa were counted from the smaii intestine that was pressed between two g h s
slides. Parasite s u ~ v a l was indicated by the number of w o m recovered from the entire
srnail intestine at necropsy.
Nutritional measures
Mouse body weight and food intake were measured every 3-4 days. Body weight
gain was calculated by subuacting the initiai body weight taken prior to randornkition to
the experirnental dietary groups from the hal body weight obtained at necropsy. The
weighu of spleen, thymus and MLN are expressed relative to the h a 1 body weight. The
concentration of albumin in plasma simples was osayed by dye-binding of bromcresol
green to the protein (No. 63 1. Sigma. St-Louis. MO) as rneasured with the Abbott VP
Sumper S ystem Discrete Analyzer (Abbo tt Diagnostic, Mississauga, Ontario). Blood urea
nitrogen @UN) concentration was detemhed by coupled enzyme reactions (No. 67-UV.
Sigma) as measured with the Abbott Analyzer.
ELISAs for IgE, parasite-specific Ig GI and MMC proteme-I
Total semm IgE and MMCP-1 were rneasured by two-site ELISA Immulon II
phtes (Dynatech Laboratories Inc., Chantilly, VA) were coated with purined rat mti-mouse
IgE mAb (021 11D clone R35-72, PharMingen, San Diego, CA) at 2 pgrd and incubated
ovemight at 4°C. Plates were blocked with 1% BSA in PBS for 2 hrs at room tempenture
to imnmiize nonspecinc binâing in weUs. Serinl dilutions of purified mouse IgE standard
mAb (03 121D, PharMingen) and diluted sera were added to the welk and incubated
oveniight at 4°C. Biotinylated, streptavidin-horseradish peroxidase conjugated rat anti-IgE
mAb (02122D clone R35-92, PharMingen) at 2 pglml was used as a detecting antibody-
Avidk- pro xidase conjug ate (Cedariane Labontories Limited, Homb y, ON) was used as

the secondary hyer and the reaction w as visualized b y addition of 2.2'-azino-(3-ethyl-
bennhiazoline-6-sulphonic acid) (ABTS; Sigma) and read at 405 nrn after 25 min with an
ELISA microplate reader (Dynatech MR5000. Dynatech Laboratories). IgE concentrations
were deterrnined 6rom standard curves and are expressed as mean p g / d î SE.
For measurement of semm MMCP-1. Immulon ï f plates (Dynatech Laboratories)
were coated with sheep mti-MMCP-1 capture antibody (Moredun Scientif'c Ltd.. Peniciuk.
Scotland) at 2 pglml and incubated for 24 hr at 4°C. Purified mouse MMCP-1 isolated
fkom mouse srnaii intestine was used as the standard (Moredun Scientinc). M e r overnight
incubation at 4°C with standards and diiuted serurn sampks. rabbit anti-MMCP- l mAb
(Moredun Scientinc) conjugated to sueptavidin-horseradish peroxidase was added as the
detecting antibody. The reaction was visuaijzed by the substrate. orthophenylene diamine
(OPD. Sigma) and read at 450 m with ELISA reader after 25 min incubation. MMCP-1
concentrations are derived Erom MMCP-1 standard curves and expressed as nglml I SE.
Parasite-speciûc IgGl in sera was deterrnined by direct ELISA Immulon II phtes
(Dynatech Laboratories) were coated with L, antigen homogenate (2.5 pg/rnl). prepared as
above. in PBS and incubated ovemight at 4OC. Piates were blocked with 1% BSA in PBS
for 2-3 hr at room temperature. Serum samples and interna1 convols from uninfected and
infected mice diluted 1:100 in PBS with 1% BSA were incubated in weUs at 4°C overnight.
Biotinyhted, streptavidin-horsendish peroxidase conjugnted rat anti-mouse IgGl mAb
(PharMingen) was used as the detecting antibody. Avidh-peroxidase conjugate (Ceduhe
Limited) was dded and plates were developed with ABTS substrate solution for 12 min.
The reaction was read at 405 nm with an ELISA reder. The results are reported as mean
optical density (OD) I SE.

Intestinal mus? cell and eo~hophil histology
Segments of small intestine (approximately 0.5 cm long, 10 cm fkom pyloms) were
h e d in Camoy's fixative (for MMC) or in 4% paraformaldehyde at 4°C overnight (for
intestinai eosinophils). The h e d tissue was dehydrated and embedded in paranin following
standard histological procedure. To count mast cek, sections (5-6 prn thick) were stained
with Aician Blue, counterstained with Sakanine 0, and the aumber is expressed as mem
per 20 villus crypt units (v.c.u.). Eosinophils in intestinal mucosal sections (5-6 pm thick)
were stained with 0.5% Chromotrope 2R and are expressed as mem counts per V.C.U.
Spleen ceil and mesenteric iymph node ceil preparation
Spleens and MLN were weighed. teased apart with needles. and then passed
aseptically through a 60 mesh stainless steel screen (Sigma). Red blood ceiIs were lysed by
osmotic shock with cold W C 1 lysing bufEer solution. Membrane debris was removed
through repeated washings with RPMI 1640 supplemented with 1096 FBS and
centrihigation at 469 g for 10 min at 4°C. Ceil viability was deterrnined by trypan blue
exclusion (0.1% Trypan Blue in PBS; Gibco) and was dways greater thm 90%. The ceh
were resuspended in complete medium consisting of RPMI 1640 (Gibco, Burlington,
Ontario), 10% heat-inactivated FBS (Gibco), 20rnM HEPES buffer (Sigma), 200 Ulm1
penicillin (Sigma), 200 p ghd streptomycin sulfate (Sigma), 25 p g/d gentamich (Gibco),
and 50 pM 2-mercaptoethanol diluted in Hank's balanced salt solution (Gibco). The stock
concentration was adjusted to 10 x 106 ceWrnL
Production of Th2 (IL-4, IL-S and IL- I O) and ni I (IFN- y) cytokines
Spleen and MLN ceil suspensions were plated in complete medium at a hai
concentration of 5 x 106 ceWmL Aliquots of 1.0 ml were incubated in 24-weîI Hat-bottom
ceil cuhure pktes (Costar, Charlotte, NC) with L, antigen (7.5 pg proteWml) prepared as
above. This concentration of parasite antigen was determined in a separate experiment to
be the lowest concentration required to stimulate maximal cytokine secretion by infected
s p k n cek. CeIl cultures were incubated at 37OC in 5% CO2 aunosphere with 90%

humidity. Supernatants were recovered afler 48 hr incubation by centrifugation of the
suspensions at 300 x g for 10 min. The supematants were stored at -80°C until they were
assayed for IL-4, IL-5, IL- 10 and IM-y. A subset of the cultured ce& (3 per diet, second
block) were lysed with Trizol (Gibco) for mRNA analysis.
Cytokine concentrations in the supematants were measured by two-site sandwich
ELISA that involved the following paired antibodies: purified rat anti-mouse IL4 mAb
1 1B 11 (18 MD. PharMingen. San Diego, CA) and biotinylated rat anti-mouse IL-4 mAb
BVD6-24G2 (180429, PhYMingen); purified rat anti-mouse IL-5 mAb TRFK-5 (18051D,
PharMingen) and biotinylated rat anti-mouse IL-5 rnAb TRFK-4 (18062D, PharMingen);
purined rat anti-mouse IL- 10 mAb JES5-2A5 (1 8 141D, PhYMingen) and biothyhted rat
anti-mouse IL-10 mAb SxC-1 (18 152D, PharMingen); and purified rat anti-mouse IFN-y
mAb W6A2 (1 8 18 1D. PharMingen) and biotinylated rat anti-mouse IFN-y mAb XMG 1.2
(18 112D, PhrirMingen). Irnmulon II piates (Dynatech Laboratories) were coated with the
capture antibody ai 2 p g h l in O. 1 M Na&PO, (pH 9.0) and incubated ovemight at 4°C.
Seriai dilutions of cytokine standards and supernatants were added and incubated ovemight
at 4°C. Biotinylated anti-cytokine detection antibodies were added at 1 pg/d and
s~ptavidin-horseradish peroladase conjugate (Cedariane Laboratories Limited, Homby,
ON, Canada) was used as the secondvy hyer. The reaction was visudized with the ABTS
(Sigma). The concentration of cytokine was cdcuhted fiom standard curves generated
fiom known concentrations of recombinant (r) murine I L 4 (1 923 1 V, PharMingen), rIL-5
(19241V, PhitfMingen), rIL-10 (19281V, PhluMingen), and rIM-y (19301T.
PhuMingen).

Cytokine mUNA expression by semi-quantitative RT-PCR
Cytokine mRNA Ievels in cultured spleen and MLN cells were quantitated by RT-
PCR assay. RNase free plastics and water were used throughout the assay. Concentration
of the extracted RNA was determined spectrophotometrically. The RNA was suspended in
water, heated to 65°C for 10 min and reverse tramcriid in a 65 PI final volume containing
6.5 pl of a 10 mM mU of four deoxynucleotide triphosphates (m), 13 pl of 5X RT
buffer, 22. pl of O.1M dithiothreital, 0.26 p1 of RNase inhibitor (40 UIpI), 0.52 p1 Oligo dT
(0.5 pg/pI), 0.65 p l of Moloney Murine Leukemh Virus (MMLV) reverse transcriptase
(200 Ulpl) (Gibco), 36.27 pl distilled water, and 5.2 pl total RNA (0.3 pgfpl). The mix
was incubated at 37'C for 1 hr and then heated to 95°C for 5 min. A 10-pl aliquot of the
RT reaction was used in a 100 p1 PCR reaction which contained 0.5 p1 of Taq DNA
polymeme (Gibco), 10 pl of 10x PCR buffer, 0.2 pl of 10 m M d m . 4 11 of 50 rnM
MgCl, 1 pl of each appropriate forwvd and reverse primers (50 pdpl). The sequence
primers for IL-4, IL-5, IL10 and IFN-y and the ceference gene, hypoxanthine phosphori-
bosyltransferase (HPRT) were selected as descnid by Svetic et al. (1993). The samples
were subjected to 29 cycles as foilows: 1) 94OC for 0.5 min; 2) 56°C for 1 min; and 3) 72°C
for 2 min. A final extension step was performed by heating to 72°C for 8 min. The nurnber
of PCR cycles and the total RNA input in the RT reaction were determined in a preliminary
experiment to ensure that the amplification was well below the saturation under this assg
condition. After PCR amplitication. a 10 p1 diquoi w u mixed with 2 pl loading buffer and
ekctrophoresed on a 1.2% agrose gel containing ethidium b r o d e (0.25 pg/ml). The
resulting DNA bands in the gel were photographed using a Polaroid camera Quantitation
of cytokine mRNA was accomplished by densitomeuic scanning of the photograph ushg
Macintosh Photoshop and NIH Image 160 software. HPRT mRNA levels did not dZer
signincantly among dietary groups (FI, 4 4 7 ; p=0.49), infection times (F,, e l .77;
p=û.17), or tissues (F,,,,=û.O6; p=0.81). The mRNA Ievel for each cytokine w u
nomialued to HPRT and expressed as ratio to HPRT mRNA for each sample.

Statistical analyses
nie experimental protocol was repeated 3 times over a penod of 8 months. The
results for nutritional, parasite, antibody and histologicd outcornes were obtained from 3
blocks. The cytokine concentration data were from the second and third blocks while the
mRNA expression data were obtained from a subset (n=3/diet/time point) in the second
block. Results are reported as means SE. Bmktt's test was used to detennine
homogeneity of variance among groups to be compmd. When the variances of the
cornpuison groups were non-homogenous (PcO.OS), analyses of eosinophils, IgE, MMCP-
1, histological counts, cytokine were perfomed with logarithmic transfonned data.
To assess the overall effects of diet and tirne, data on wom burdens. mtibody titres.
histological counts, MMCP- 1 concentration, cytokine production and cytokine mRNA
expression were pooled by infection protocol (primary or challenge) and each was anaiyzed
for the effects of diet, tirne and diet by time using a two-way analysis of variance
(ANOVA). Interactions between diet and tirne were reponed only if signincant. Repeated
mesures ANOVA was used to assess effects of diet and iime on peripheral eosinophils.
One-way ANOVA was used to detect the effects of diet on nutritional parmeters of
chdenged mice killed on day 28 and on parasite and irnmunological data of naive mice.
Although our study was designed to examine the overail effects of diet and tirne
during the entire course of the prirnary or challenge infection (as assessed using two-way
ANOVA), we ako studied the efTect of diet at each post-infection tirne point because we
expected dietuy modulation of host immune response to vary in magnitude at different
stages of the infection. To thiS end, the w o m burdens, antibody titres, histologicd counts,
MMCP-1 concentration. and cytokine data were analyzed for the effect of diet at each post-
~ e c t i o n time point with a one-way ANOVA foliowed bypost hoc p-e cornpuisons
(Tuicey) when diet w u signiticant. Only results from pusr-hoc cornparisons are reported.
The analyses were perfomed using SAS (SAS Institute I~c., Cary, NC) and considered
sipifkant at P c 0.05.

Num*tional Outcornes
Table 2 summarizes the nutritional outcomes of a representative group. challenged
mice killed on day 28 pci Although challenged rnice fed the 3% protein diet ate the same
amount of food curnulatively, and hence calories, as mice fed the 24% protein diet, they
gzined significantly less body weight. Thus. the 3% protein diet induced protein but not
energy deficiency that Frequentiy accompanies dietary protein restriction. In fact. the 3%
protein group ate significmtly more food per gram body weight compared to the other 2
dietvy groups. However, ihis increased relûtive energy intake did not support normal
growth as s h o w by the 71% lower body weight gain of protein deficient mice compared to
the control mice. In contrast to the 3% group, the marginal protein (7%) group ate
signincantly more food overd and as a result gained comparable weight as the 24% group.
Protein restriction at both the 3% and 7% level signincantly decreased rehtive
weight of mesentenc lymph nodes (MLN). The 3% protein diet also signincantiy lowered
relative spleen weight. Thymus weight was not affected by protein restriction. Dietary
protein deficiency modified biochemical indices of protein status: plasma albumin
concentration was decreased by the 3% protein diet only when compared with the 7%
group, but neither groups differed from the 24% mice. However, blood urei nivogen
(BUN) concentration showed a dose-dependent response to protein level in the diet.


Parasite Outcornes
The hinctional significance of protein deficiency on immunity is shown in Figure 2
which presents the worm burdens of mice fed the experirnental dieu and given either a
primq or challenge Section wiih H. polygyrus. Previously irnmunized mice fed the 24%
and 7% protein diets were capable of expeiling over 95% of their parasites by day 28 pci
compared to their primary infection counterparts who eliminated only 56-60s of the
infective dose. In contrut. worm burdens were signincmtly higher in mice fed the 3 8
pro tein diet throughout the primary and chdenge infections, thus supporting the prernise
that protein deficiency impairs the development of protective immunity to H. po1ygyrusS
During challenge infection. protein deficiency increased both the establishment of H.
p o l y g y w at day 6 pci and the s u ~ v a l of aduit wonns at days 14-28 pci The wom
burdens of the 3% mice were also signincuitly higher cornpared to the 24% and 7% mice
during prirnvy infection. however, these were not as dnmatic as that observed during
challenge infection.
Serum Effector Responses
To assess the effects of protein deficîency on serum IgE responses, we measured
IgE concentration in nive mice, during primary infection and throughout the challenge
infection 3A). Serum levek of IgE were neady undetectable in uninfected rnice
and low in prirnary mice but increased dnmatically after chdenge infection. Serum IgE
levels in ail dietvy groups changed over the course of challenge infection (time effect,
Pcû.05): mice fed 24% and 7% protein diets h d noticeable peaks in serum IgE at days 6 to
9 pci whüe this elevation was blunted in rnice fed 3% protein. In al chaiienged mice. serum
IgE levels gradudy deched over tirne and by 28 days pci were simi7u to ievels observed
during primary Section. Mice fed 3% protein had signiacantly lower serum IgE levels
cornpared to mice fed 24% pro tein throughout the chdknge infection and on day 28 pp i
Serum IgE ievels in the 71 mice were simily to the 24% mice on days 3.6 and 14 pci.
were intermediate between the 24% and 3% groups on days 9 and 2 1 pci, and were as low
as the 3% group by day 28 pci, thus suggesting a possible dinerential response over time to
graded levels of dietary protein

Infection Groups
Figure 2

Figure 2. Effects of d i e t q protein deficiency and time on worm burdens of mice given r primary or challenge infection with H. polygynrs. Values are mean + SE. During primvy infection. the number of wonns was significantly higher in 3% mice compared to 24% and 7% mice (F,=13.35. Pd.0001) and was signXcmtly higher on day 6 ppi cornpared to day 28 ppi (F,*,=5.29, P=0.024). During challenge infection, the worm burdens were signincantly higher in 3% mice compared to 24% and 7% rnice (F,,,,=163.37, P=O.Oûû 1) and were signiticmtly ai3ected by time (F,,,,,=47.63, P=0.000 1). There were signincant diet and time interactions in primary infection (F,,=4.22. P=û.018) and in challenge infection (F6,1,=9.15, P=O.ûûûl), indicating that 24% and 7% rnice but not 3% mice eiiminated progressively more worms over thne. Dinerent letters at each time point represent significant dEerence between dietary groups ( P d . 0 5 ) based on posr-hoc pairwise cornparisons (Tukey).

ParasiteSpecif ic lgGl O.D. O O a --L
O in ô in P O
Serum IgE (uglml)

Figure 3. Effects of dietary protein deficiency and time on semm antibody bels. Values are + SE. Total semm IgE (A) and parasite-specitïc IgGl (B) were determllied in naive mice, primary irifection (days 6 and 28 ppi). and challenge infection (days 3.6.9. 14, 21. and 28 pci). A: In naive mice, there were no signiricant clifferences in serum IgE detected between the dietary groups (F,,,=O. 89, P=O.43). During primary infection. semm IgE was not affected by diet overaii (F,=2.43, Pd.094) but was signüicuitly higher on day 28 ppi compmd to day 6 ppi (F,,=254.5, P=0.0001). During challenge infection. serum IgE was signülcantly different arnong ail three diets (F,,,=44.3, W.0 1) and signincantiy dtered over t h e (F,,,,=17.67, P=0.0001). There were no signincant interactions of diet and t h e in serum IgE during primiuy or challenge Uifection. B: Parasite-specific IgG 1 concentration wu no t signincantiy affected by diet overd in naive rnice, primvy groups or challenge gro ups. Tirne significantiy afTected parasite-specinc IgG 1 levels among die tvy groups d u ~ g primary infection (F,,8,=583.18, P4.000 1) and during challenge infection (F5,,,=150.62, P=0.0001). For IgE, different letters represent signincant merence between dktq groups (Pd .05) based on post-hoc painvise cornparisons (Tukey) perfbrmed following signincant effect of diet in one-way ANOVAs at each time point.

Parasite-specinc IgGt responses were also measured in the different infection and
dietary groups (Figure 3B). Sirniliir to IgE responses. parasite-specific IgGl was not
detected in naive mice or evly during primary section but w u signincuitly elevated after
challenge infection. However, protein deficiency did not impair the synthesis of parasite-
specific IgGI at any tirne during the prirnary or challenge infection period. In ail dietary
groups, parasite-specific IgGl rose rapidly evly after chaUenge infection and peaked by day
9 pci (tirne effect. P<0.05) to levels greater thm that observed during primvy Uifection.
Higher parasite-specinc IgG 1 levels persisted in aii dietary groups after 4 weeks pci despite
the near absence of worms in the rnice fed 24% or 7% diets.
The patterns of peripheral eosinophil responses were deterrnined duMg p r i m ~ y and
challenge infection with H. polygyrus (Figure 4). Pro tein deficiency signincantly Io wered
the numbers of peripheral eosinophils throughout the primary infection (Figure 4A).
Although eosinophil counts appeued to be higher in the 24% mice compared to the 3%
mice on day 5 pci, the overd effect of diet on peripheral eosinophilia throughout challenge
infection was no t signincant (Figure 4B). At day O pci (on which the challenge mife
received theû second infection and the p r i m q mice received their fïrst infection), blood
eosinophil counts in previously immunized mice of the challenge group were higher than in
mice of the primary group. indicating the more intense secondary response characteristic of
acquired irnmunity. In the f h t 2 days afier challenge infection. circuiating eosinophil
numbers in t d vein blood decreased in di dietary groups. suggesting a preferenthi
recniitment of blood eosinophils to the parasite-infected intestine. This was followed, in
mice fed 24% or 7% protein. with a shvp increase in blood eosinophh during the fbst
week of infection which was double die eosinophilüi observed in prirnary mice fed either
diets. Blood eosinophil responses declined rapidly after day 5 pci in the chaknged mice
fed 24% or 7% protein possibly because of the decreasing number of w o m in the gut. In
contrast. elevated blood eosinophilia persisted in the protein sufficient mice throughout the
primary infection.

B Challenge Infection
Figure 4
2 5 13
Days Post-Infection

Figun 4. EfTects of dietary protein deficiency on eosinophüÿt in tail vein whoie blood during both primary (A) and challenge infection (B) with H. po lygyw . Values are î SE. A: During primary infec tien, perip henl eosinophili;i was signitfcuitly aEected by die t (F,=8.7 1, P = W û 1) and tirne (F6,,,4=47.99, W.000 1). B: During challenge infection, peripheral eosinophdia was significantiy aected by rime oniy (F,,,,,=6.87. Pd.000 1 ). There were no significmt interactions of diet and t h e during primtuy or challenge infection.

Intestinal Effector Responses
We counted the number of MMC and intestinai eosinophils in sections of intestinal
tissues obtained from nive rnice, primary mice. and chaIIenged rnice (Figure 5). In aii
challengecl mice. the numbers of intestinal MMC increased 5-fold by day 3 pci compared
with the uninfected levels and were consistently higher at ai l pci t h e points cornpared with
early p h v y mice (Figure 5A). Overali, the 3% protein diet significantly impaired MMC
proliferation during challenge infection but no t during primary infection. High numbers of
MMC in chailenged mice fed 24% or 7% protein die6 persisted throughout the infection
period (tirne effect. PsO.05) but dropped in the 396 group after day 9 pci (time effect,
PeO.05). Therefore, significant dserences in MMC numbers between 245 and 3% groups
were detected on day 9 pci and successively thereafier. Numbers of intestinal MMC were
lower in the 7% protein diet compared with the 24% group ody on day 9 pci and at other
pci time points were either sunilar to 244 mice or intermediate between the 24% and 3%
groups. Hence, the marginai protein mice maintained intermedite but adequate leveis of
MMC d u ~ g challenge Section. MMC counts were not statisticdly different among the
dietary groups in d e c t e d mice or duMg prirnary infection.
Proiein de ficienc y also significuitly prevented the increase of intestinal eosino phils
during challenge infection but did not af5ect numbers in naive or primary mice (Figure SB).
In spite of the fluctuation in the numbers of intestinal eosinophils after challenge inlection.
they showed m overall dose-response to the dietary protein leveL Compared to the
uwected level. a doubiing of intestinal eosinophil numbers was apparent within 2 weeks
pci only in mice fed 24% protein. Marked increases in intestinai eosinophilia were not
obserwd in the chaîienged rnice fed 7% or 3% protein compared to their naive
counterpans, and therefore. gut eosinophils fded to prouerate after challenge infection in
these protein restricted groups. Due to the considenble variability in counts obtained at
each pci time point (e-g. dûys 3 and 9 pci), intestinai eosinophil bels did not change
signincantiy over tirne in any of the dietary groups (ticne effect, A0.05). It is important to
note that high numbers of gut eosinophiis were found in 24% mice on day 3 pci despite Iow
numbers of circulating eosinoplds obsemed very eariy afier challenge infection (Figure 4).
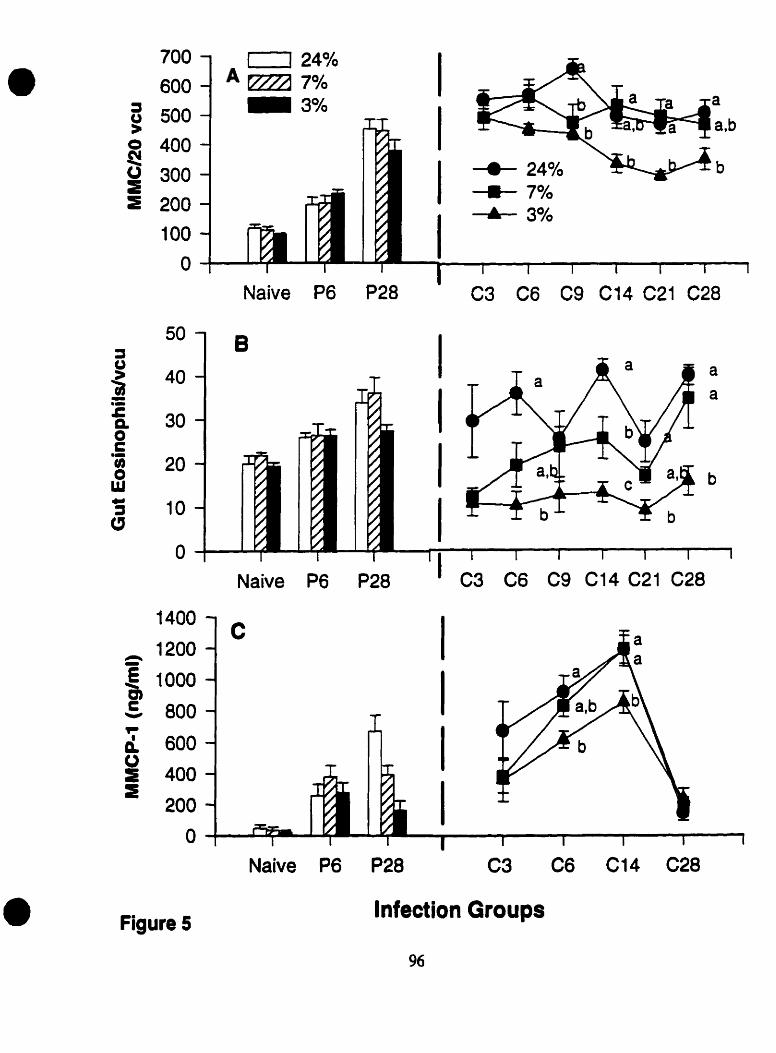
Naive
Naive P6 P28 C3 C6 Cl4 C28
Infection Groups Figure 5

Figure 5. Effects of dietvy protein deficiency and t h e on the numbers of MMC per 20 vcu (A), numbers of intestinal eosinophils per vcu (B), and the secretory capacity of MMC as measured by MMCP- 1 concentration (C) in naive mice, p M i q mice (6 and 28 days ppi), and challenge mice (3.6.9, 14.21 and 28 days pci). Values are * SE. A: In naive mice, the numbers of MMC were not signiticantly atrected by diet. During primary infection. the numbers of MMC were not significantly different among dietary groups but were signincmrly altered over the (F,e7O. 19, P=0.000 1). DuMg challenge infection, the nuben of MMC were signüifantly lower in 3% mice compared to 24% and 7% rnice (F,,=21.69, Ps0.0001) and signincantly affected by tirne (F,,,=4.09, P d . 0 0 3 ) . B: Numbers of intestinal eosinophiis were no t significantly affected by diet overail in naive or prirnary rnice. During primary infection, intestinai eosinophih was signincantly highet on day 28 ppi compared to day 6 ppi (F,,=7.02, P4.014). During challenge infection, the numbers of intestinal eosinophüs were signincantly dinerent among di dietary groups (F,,=29.6 1, P=ûAlûû 1) and signincuitly different over time (F,,=3.67, P=0.0059). C: S e m MMCP-I concentration was not significantly affected by diet in naive mice. During primary infection. semm MMCP-1 levels were not significantiy affected by diet or time but there was a signincant diet and time interaction (F,,=3.84, P4.028). During challenge infection, MMCP-1 concentration was not significmtly aected by diet overall but w u significantly affected by t h e (F,,,=44.59, P=O.ûûû 1). For MMC, gut eosinophils and MMCP-1, the different ietters represent signincuit difference between dietary groups (P4.05) based on post-hoc pairwise cornparisons (Tukey) foIiowing significant effect of diet in one-way ANOVAs at each t h e point.

To determine the secretory capacity of MMC duMg nematode infection in mice
with v-g degrees of protein deficiency, we measured the concentration of s e m
MMCP-1, a protease expressed predominantly in intestinal MMC (Figure SC). Protein
deficiency did not alter MMCP-1 ievek in uninfected rnice, prirnary mice or in challenged
mice. Although there were no overd diet effects during chailenge infection, we found that
protein deficiency significantly inhibited the increase of MMCP- 1 secretion at days 6 and 14
pci ( P d . 0 5 ) . The time course of MMCP- 1 response parallels that of semm IgE such that
MMCP-1 concentration in the protein suficient mice were higher after challenge infection
compared with unllifected or prirnary mice. In ail dietary groups, peak MMCP-1
concentration occurred on day 14 pci (tirne effect, P<0.05) which occurred &ter peak
MMC numbers in the gut mucosa (in 244 mice on day 9 pci) and peak bels of serum IgE
and parasite-specinc IgGl (day 9 pci).
Systemic and Local Cytokine Responses tu Parasite Antigen in vitro
To determine whether the impaired effector responses observed in the protein
deficient mice were reiated to diet-induced perturbations in cytokine production in local
a d o r systemic lymphoid tissues, we measured post-challenge Th2 cytokine (a-4, IL-5,
IL- IO) and Th 1 cytokine (IM-y) production by spleen and MLN ceik restimulated with
parasite antigen in vitro mgure 6). In general, Th2 cytokine secretion was higher in the
fnst week of primuy infection compared with the challenge infection (with the exception of
IL-5 production in spleen), indicating a strong priming of CD4* c e h to secrete Th2-type
cytokines. Also, the Th2 cytokine proNe durhg challenge infection was similu among the
3 dietuy groups in that the response was greater in MW cells than in spleen cells, peaked
by one week pci, and declined npidly thereder. Effecu of protein deficiency on Th2
cytokine production were detected only in IL,-4 secretion by MLN cells early after
challenge infection @gure 6A & 6B). The MLN ceik of the 7% protein group secreted
significantly Iess IL-4 at day 3 pci compared to their 2446 protein counterparts. S W l y at
day 6 pci, IL-4 production was significantly lower in MLN of mice fed 3% protein
compmd to mice fed 24% protein We also observed differences in mucosal IL-4

SPLEEN MLN
Figure 6 Infection Groups

Figure 6. Effects of dietary protein deficiency on Th2 cytokine (IL-4. IL-5. IL-10) and Th1 cytokine (EN-y) production in spleen (A, C. E. G) or MLN (B. D, F, H) ceUs in mice with either primary or challenge infection of H. polgynrs. Values are I SE. Spleen and MLN ceh from P6. C3, C6, Cl4 and C28 mice were cuitured, restimuhted with parasite antigen in vitro, and assayed for cytokine secretion using sandwich ELISA Values are mean nglml I SE. A and B: At day 6 ppi, IL-4 production was signincantly lower in spleen 01 3% mice compared to 24% mice (F,,=3.87, P=0.03 1) but not in MLN. During chdknge infection, IL-4 production in spleen was signincantly affected by tirne only (F, ,=6.55, P4.0004) and in MLN was signincuitly &ted by both diet overdi (~;,=6.08, W . 0 0 3 3 ) and tirne CF,,=14.06, P=0.0001). C and D: IL-5 secretion by spleen or MLN cells was not signiticantly dinerent among dietary groups at day 6 ppi and during c W n g e infection. Time signincantly aEected IL-5 production oniy in MLN ceiis duMg chdenge infection (F3,,,,=16.74, Pd.0001). E and F: IL-IO production by spleen or MLN ceUs wu not significantly affected by diet overd at day 6 ppi and throughout the challenge infec tioa During challenge infection. tirne significantly affected IL- 10 production in spleen (F, p l 3.4 1, Pa000 1) and in M W (F3,8,=9.97, P=û.0001). G and H: IFN- y production kas not signincantly atrected by diet overall at day 6 ppi or throughout challenge infection. During chailenge infection, time significantly affected IFN- y production in spleen (F3,,,=35.06. P=0.0001) and in MLN (F,,,=30.89. P=0.0001). For a i l cytokines produced in spleen or MLN, different letters represent significant differences between dietary groups (PcO.05) based on post-hoc p M e cornparison (Tukey) following significant enect of diet in one-way ANOVAs at each post-challenge time point.

production between the dietary groups at a t h e immediately prior to challenge section
(basal challenge levels). In these primed mice. the MLN cells of the 2 4 4 group produced
more IL4 (8.66 ng/d * 0.71. n=2) compared to 3% group (3.98 nglml I 0.96, n=2) while
the MLN of mice fed 7% pro tein secreted an intermediate level (5.16 nglml* 2.43). The
7% mice were capable of increasing IL-4 production by 6 days pc5, suggesting a capacity to
up-regdate IL-4 production at a post-infection tirne critical for maximal stimulation of
effector responses. By cornparison, the decreased production of IL-4 in MLN cells of m k e
fed 3% protein preceded the impairment of intestinal and systemic Th2 effector responses.
As shown on Fgure 6C to 6F, the protiles of IL5 and IL40 production in spleen and
MLN were s h h r to IL-4 responses but. unlike IL-4, were not signifïcantly affected by
protein deficiency overaii duMg chaüenge infection. Despite an overail nonsignifïcant
effect of diet, we found that L-5 secretion in MLN was higher in mice fed 3 4 protein
compared with either 24% or 7% groups at day 3 pci but this was not associated with
m k e d elevation of gut eosinophils.
As shown in Figure 6G & 6H. IFN-y secretion by both spleen and MLN ceh
restimuhted with parasite antigen in vitro increued signifïcuidy throughout the challenge
infection in a.li d i e t q groups (tirne effect, P d . 0 5 ) . except in spleens of mice fed 3%
protein (tirne effect, P>0.05) This higher IFN-y response in spleen and MLN over time
coincided with decreasing ieveis of IL-4 and IL10 in MLN and increasing worm loss in the
protein sficient mice. We did not detect an overall significant effect of diet on IFN-y
production during challenge infection period. However, at day 14 pci, mice fed the 3 2
protein secreted significantly more IFN-y compared to rnice fed 24% protein in spleen
(P=0.0003) and in M'LN ( P 4 . 0 2 3 ) . Taken together, protein deficiency did not alter the
inherent capacity of spleen or MLN ce& to synthesize ?FN-y but increased IFN-y
production at the onset of adult worm expulsion (day 14 pci) in a c h a n g e infection.

Systemic and Lucal Cytokine m W A Expression
To address the question of whether altered IL-4 and IFN-y profiles in protein
deficient mice resulted from diet-induced changes in cytokine mRNA expression, we
analyreci be l s of cytokine mRNA using semi-quantitative RT-PCR (Figure 7). The spleen
and MLN ceh of mice fed either 24% or 3% protein (n=3) were restimdated with parasite
antigen in vitro and then assayed for E-4, IL-5, IL10 and IFN-y mRNA In mice fed 3%
protein. IL4 mRNA expression in spleen and MLN cek fded to increase and remained
low throughout the chalienge infection (Figure 7A & 78). The dinerences between dietary
groups were statisticdy signincant on d g 14 pci in spleen celis and on days 6 and 14 pci in
MLN ce&. Also, IL40 mRNA expression in spleen and MLN was significantly lower in
mice fed 3% protein compared with their 24% protein counterpûru at day 14 pci (Figure
7E & 7F). The ievels of IL-5 (Figure 7C & 7D) and IFN-y (Figure 7G & 7H) rnRNA
expression were not significuitly affected by protein deficiency at any t h e point during
c hdenge infection.


Figure 7 Effecu of dietary protein deficiency and tirne on Th2 cytokine mRNA (IL-4, IL- 5, IL-10) a d ni1 cytokine mRNA (IFN-y) expression in spleen (A,C. E, G) or MLN (B, D. F, H) ceh in mice with challenge infection of H. p o l y g y m . Spleen and MLN cells from C3, C6. Cl4 and C28 mice were cultured and restimulated with parasite antigen in vitro. Cells fiom the supematants were lysed for total mRNA and mRNA expression of each cytokine was determined with semi-quantitative RT-PCR Values are expressed as the ratio of cytokine mRNA to HPRT mRNA for individual simples (mean î SE, n=3). A and B: IL4 mRNA levels in spleen and MLN were signincantly affected by diet (F,,=12.28. P=û.ûû 15) and the (F,,3,=6.43. P4.M) 17). C and D: No significant dinerences in IL-5 mRNA expression in spleen and MW were detected between the dietary groups or time points. E and F: IL,- 10 rnRNA leveis in spleen and MLN were signtncantly aEected by diet (F ,,,, =6.64, Pr0.015) and tirne (F,,p17.18, P4.0001). G and H: IFNy mRNA expression in spleen and MLN was no t significantly affected by diet or time. For ail cytokine mRNA levek in spleen or MLN, dserent letters represent signincant dinerences between dietvy groups (P4.05) based on post-hoc pYwise cornparisons (Tukey) foiiowing signincant effect of diet in one-way ANOVAs at each tirne point.

DISCUSSION
This study is the fist comprehensive investigation on the effects of dietary protein
deficiency on the magnitude and kinetics of Th2 immune responses in mice to a challenge
infection with a gastrointestinal nematode parasite, Heligmosomoides p o l y g y w . Contrary
to conventional belief, dietriry protein deficiency did not suppress aU immune processes but
instead exened dinerential site- and type-specific effects on cytokine and effector responses
to H. polygym infection To iuustrate this point, four important hdings are noted: 1)
Protein deficiency impaired the proliferation of gut eosinophils and mucosal mast cells
(MMC) but only duMg the evly phase of infection; 2) Pro tein deficiency decreased the
levels of circulating IgE but not of parasite-specifc IgGI; 3) Protein deficiency down-
regulated IL-4 production only in a gut Iymphoid tissue and did not ûffect secondary
responses of other Th2 cytokines; and 4) Protein deficiency up-reguhted levek of IFN-y at
2 weeks pci which is a cnticai period for expression of acquired immunity. Additiody,
the Ievel of d i e t q protein influenced the degree of protective immunity and parasite
s u ~ v a l duMg a challenge infection with H. polygynrs. Our results support the hypothesis
that protein deficiency prolongs sunival of a gastrointestinal nematode parasite by
decresing gut-associated IL4 (TM) and increuing IFN-y (Thi) early in the infection,
leading to reduced intestinal and sysremic Th2 enector responses.
Acquired immunity to secondary infections with H. polygyms is assochted with
strong and persistent Th2 effector responses (Monroy & Enriquez, 1992), although the
precise mechuiisms by which these Th2 effectors exen their protective effect have not ken
ciearly elucidated. Eosinophiiia is a typicai Th2 infiamatory response observed during
nematode parasitic infections but some researchers have suggested that host resistance to
H. polygyrus or Trichinella spirulis infections in mice is not dependant on eosiaophilis
Prevention of e o s i n o p h induced by the neutraüzation of IL4 antibody or deletion of the
IL-5 gene did not afliect worm survival or fecundity (Urban et al.. 199 1; Herndon & Kayes,
1992; Takamoto et aL, 1997). However, these authors measured eosiwphiiia in circulating
blood, an indicator bat does not necessvily refiect eosinophil responses in intesiinal sites of

Section which are more relevant to host immunity against enieric parasites (e.g. Svetic et
ai.. 1993; Negno-Coma et al., 1996). An example of the svong bias towards gut mucosal
localization of eosinophils during nematode infection is our hding of a transient drop in
blood eosinophils at day 2 pci that coincided with elevated numbers of intestinal eosinophils
in mice fed 24% protein. We interpret this pattern as evidence that eosinophüs are
trmported preferentiaiiy to the infec ted intestine hmedkely folIo wing c haiienge infection
until gut-associated lyrnphoid tissues are capable of producing high quantities of local
eosinophils. Similx to this reputed migrational pattern of eosinophils during polygy~us
challenge infection, others have observed that the uptake of IgE by intestinal ceUs is greatly
increased within 24 hours after T. spiralis Uifection (Rarnaswamy et al., 1994) and that the
vast rnajority of intestinal IgE (>99%) detected by 4 days post-infection is produced locdy
in the gut (Negrao-Correa et al.. 1996). These results suggest that high levels of immune
effectors in the gut are hinctiondy more important than peripherai quantities during
intestinal nematodiasis. Thus, Our fkding that intestinal but not peripherd eosinophilio was
inhibited by protein deficiency during challenge infection may be one factor contnbuting to
the prolonged worm s u ~ v a l in the protein deficient mice. However, the roie of
eosinophils in pro tective immunity to helminth infections has been chaUenged by studies
showing that murine eosinophils, unWte human eosinophüs, hck IgE receptors (the high-
S i t y FcdU and the low-afnnity FceRWCD23) but do express IgG receptors (de Andres
et al., 1997). Accordingly, the cross-Mage of IgG receptoa by pansite antigen may be
the main activation pathway of eosinophil degmuiation and oxidative burst. Thus, the
functional significance of reduced gut eosinophilia in mice fed 3% protein to parasite
expulsion remains uncenain because the synthesis of puasite-specinc IgGl (the IgG isotype
associated with Th2 responses to nematodes) after chdenge infection with H. p o l y g y m
was no t impaired by pro tein dekiency.

This lack of responsiveness of parasite-specifi~ IgGl to the detrimental effecu of
protein deficiency during primary or challenge infection with H. polygyrus corresponds
with previous results from Our laboratory (Bouhy et aL, 1998) and rnay be explained by
literature suggesting that the regdation of IgGl response does not require IL-4. Eatly
studies demonstmted that the synthesis and expansion of IgG1-secreting B cells is
dependent on CD4+ T cells and that IL-4 promo tes isotype switching of IgM+ B ce& to
membrane expression and antibody secretion of IgGl (Vitetta et al.. 1985; Coffmui et al..
1986). However. hter animai studies showed that IgGl responses can be induced duMg
H. polygynrs or Nippos~ongylus brasiliensis infections even in the absence of IL-4 activity
in vivo and the disruption of the murine IL-4 gene (FYike1rna.n et al.. 1988; Katona et al.,
1991; Kopf et al.. 1993). Furthemore. treatment with exogenous IL-4 increased the
transport and uptake of IgE but not of IgG1 into the intestines of T. spiralis-infected rats
(Ramaswany et al., 1994). Therefore, any disturbances in IL-4 production caused by
protein mainutrition as observed in Our study may not necessdy lead to subsequent
changes in IgGl responses during H. polygyms Section. Also, the observation that
protein deficiency signincantiy prolonged parasite s u ~ v d without impairing concomitant
IgGl responses indicate that we should reûppraise the immunologie d e of IgGl in
protective immunity to nematode parasites. Pritchard et ai. (1983) reported that purified
IgGl from immune sera of mice infected with H. polygyrus caused severe stunting of
woms and promoted adherence of pentoneal exudate celis to the surface of immature and
adult woms in vino. However. Wahid et aL (1993) later showed that parasite-specific
IgGl responses did not correlate with the rate of wonn expulsion and were rnargindy
elevated in mice with heavier worm burdens. Moreover, the inhibition of IL4 and its
receptor by neutraiizing antibodies prolonged worm survivaî in primary H. poiygyrus
infection without concunent impairment of IgG 1 responses (Finkelman et al.. 1988;
Katona et al., 199 1). The undtered parasite-specific IgGl levels observed in the heavüy
pivasitized protein deficient mice fûnher support the contention that IgGl may contribute
iess to host protection a g d t intestinal parasites than other Th2 effectoa in a c W n g e
infection.

MMC hyperpiasia is another prominent feature of Th2 effector responses to
intestinal helrninths including T. spiralis and N. brasiliensis (Woodbury et ai., 1984;
Dehlawi et al., 1987). The presence of adult polygyrur in the intestinal lumen is
associated with poor mastocytosis, chronic parasite s u ~ v a l , and inhibition of MMC
responses stimuhted by concomitant T. spirulis infections, indicating that adult H.
p o l y g y w exerts an immunosuppressive effect on MMC development and proliferation
(Dehlawi & Wakelin. 1988; Behnke et al., 1993). Moreover, spontaneous expulsion of H.
polygyrus in some resistmt strains of mice and elimination of N. brasiliensis in nu were
not accompanied by concurrent mucosal mastocytosis which had peaked afier maximal
worm loss (Woodbury et al., 1985; Dehhwi et al*, 1988; Wahid et al., 1994). However,
increased size and accelerated development of MMC responses have been observed during
challenge infections with FL polygyrus in which there was &O a reduction in the number of
penetnting b a e and expedited elhination of adult w o m (Dehhwi et al., 1987). thus
implying that MMC may contribute to acquired resistance by arresting h a 1 development
or darnaging young adult worms following b e n a l emergence. Indeed, we found that a
challenge infection with If. polygyms induced prominent and persistent MMC responses
that peaked (day 9 pci) prior to mvked eltnination of adult parasites (beginning on day 14
pci). The concept that secondq MMC responses are a critical mechanism of acquired
resistance is supported by Our finding that protein deficiency irnpûired the intestinai
prolûeration of MMC within 2 weeks pci but did not affect MMC numbers during prirnvy
Uifection in which more w o m were recovered thm after chdienge infection Funhermore.
the onset of depressed mucosai mastoc~tosis immediately preceded the loss of adult w o m
in the pro tein sutficient mice, thereby implying that MMC responses were elicited by larvae
tesident in the gut mucosa and that local MMC induced early after chdienge infection
directly phcipated in pansite expulsion in conjunction with IgE or IgG1.

IgE response is an essential component of acquired immunity to nematode pansites
and any increases in this antibody represent an induced response in helminth infections. We
found that serum IgE levels were low in uninfected mice and duMg primary infection but
hcreased substmtially after challenge infection- However, this nematode-induced eievation
in IgE was absent in the protein deficient mice which ais0 had lower MMC levels in the gut
mucosa. nie importance of IgE in mediating protective immunity to helminth parasites is
infened from studies showing that the neutraiization of IL-4 in vivo or inhibition of IL-4
receptor ameliorated semm IgE responses and increased worm numbers (Fieman et a l ,
1990; Urban et al.. 199 1). Moreover, mice with a mutation of the IgE gene have increased
burdens of Schistosoma monroni despite prominent elevations of antigen-specific IL-4-
secreting ceils (King et al., 1997). Binding of IgE to high atfinty receptors on mast ceh
and cross-linking by parasite antigen cause the degranulation of MMC and reiease of
inflanmatory mediators. Accordingly. an effective IgE-medhted immune response requires
not only sufficient Ieveis of MMC md associated IgE receptors but also abundant IgE
antibody at the appropriate sites of infection in order to enhance m a t ceil FceRI expression
and secretory capacity (Chen et al., 1994; Yamaguchi et ni., 1997). In our study, the t h e
courses of serum IgE response, MMC proliferation and parasite expulsion were tightly
correlated such that the onset of parasite expulsion in the protein sufficient mice (day 14
pci) occurred immediately following pedc serum IgE and intestinal MMC numbers (day 9
pni. Funhermore, the protein deficient mice h d higher w o n burdens throughout 4 weeks
of challenge infection as wel as consistently decreased serum IgE titres and lower numbers
of MMC in the gut mucosa Taken together, these results provide additionai evidence that
protective immunity to nematode parasites requires maximal induction of IgE production
and concurrent proliferation of MMC in situ or intestinai trafkking of mast ceii precursors.

Aithough semm IgE concentration was found to correlate poorly with intestinal IgE
ievels (Negrao-Coma et al., 1996) and with IgE occupancy or density of IgE receptors on
mast cells (Chen & Enerback, 1994 & 1996). Our results on the secretory capacity of MMC
iikely implicate a functional consequerice of lower serum IgE bels detected in the protein
deficient mice. Inasmuch as MMC activation md degranulation depend on expression of
IgE receptors and acquisition of IgE by the mast cell. the lower MMCP- 1 concentrations in
protein deficient mice at days 6 and 14 pci may be due to both decreased MMC numbers
and reduced levels of IgE in gut-associated lymphoid tissues. Lower serurn MMCP-I
levels likely represent the true extent of the immunosuppressive efEects of protein deficiency
on MMC secretory capacity given that the intestine is the major source of systernicdIy
secreted MMCP-1 and that semm levels correlate positively with concentrations in the
intestinal lumen of nematode-infected mice (Wastling et al.. 1997). There are &O
preliminary indications that MMCP-1 has a physiological role in puasite expulsion by
affecthg epithelial permeabüity and translocation of antibodies from blood to the gut lumen
(Wastling et a l , 1997). Lower IgE responses, depressed MMC numbers and impaired
MMC secretory activity were apparent by day 9 pci in mice fed 346 protein at a t h e point
that preceded the onset of marked parasite expulsion in immunocompetent mice fed 24%
protein. Although we did not measure MMCP-1 on day 9 pci, other studies have shown
hat MMC protease concentration in blood peaks slightly iater than maximal intestinal
mastocytosis in nematode-parasitized mice and rats (Woodbury et al., 1984; Wastiing et
aL, 1997). Thus, prolonged w o n survivd in rnice fed 3% protein may be attributed in part
to the impaired Ig E-dependent MMC pro Werative and secre tory responses O bserved evly
after challenge infection with H. polygynrs.
We investigated whether these impaired enector responses in protein deficient rnice
were reLzted to perturbations in Th2 cytokine production. It is currently accepted that IL-
4 is the critical cytokine directing ni2 differentiation and respowe phenotype d u ~ g
nematode iiifections (Mosmann & Sad, 1996). IL-4 promotes the dinerentiation of Th ceh
to the Th2 phenotype (Rincon et al., 1997). the generation of memory Th2 and B cells
(Bradley et ai.. 1995). isotype switching of B cells from IgA+ and IgW to I g F (Zhang et

al., 1991), the proliferation of IgE-comrnitted cells and differenthted MMC (Fïnkleman et
al., 1988; Katona et al., 1991), mast ceii FceRI expression (Tom et al., 1996). binding of
plasma IgE to intestinal ce& of T. spirulis-infected mice (Ranaswany et al.. 1994). and the
expulsion of established H. p o l y g y w in mice (Urban et ai.. 199 1). Our kdings b a t
protein deficiency decreased the capacity of MLN celis of H. polygynrs-infected mice to
secrete IL,-4 and to express IL-4 mRNA, depressed Th2 effector responses, and prolonged
parasite s u ~ v a l support previous reports (e.g. Urban et al.. 1991 & 1995) that IL-4 is a
crucial cytokine required for strong and persistent Th2 effector responses and npid
elhination of nematode parasites. In contrast to IL-4. protein deficiency did not impair IL-
S and IL10 production even though the 3% protein diet reduced IL40 mRNA ievels.
(Yamaguchi et al.. 1988; Coffman et al.. 1989; Behnke et al., 1993). This indicaies that
relaiively smdi amounts of IL-4 may be suEcknt for production of other Th2 cytokines
and that protein deficiency may a e c t gene expression without irnpairing translation of
mRNA to protein. In fact, IL-5 secretion by MLN was relatively higher in mice fed 3%
protein at day 3 pci, but the functiond signif~cance of this event is unclear because the up-
regulated IL5 response at this early tirne point was not associated with concomitant
elevation in gut eosinophilia, It is possible that IL-5 and IL- 10 are constitutively expressed
during nemat ode infection and their regdation is less responsive to exogenous factors.
Other researchers have questioned the hnctional impact of IL-5 and IL40 during
nematode infection The expression of E-10 mRNA does not correlate with the mRNA
levels of other Th2 cytokines in H. polygyms Mection (Svetic et al.. 1993) and IL12 cm
enhance the production of IL-10 by humm T ce& (Widhagen et al.. 1996). thus implying
that IL- LO may no t be a s tric tly Th2-resuic ted cytokine. h o , IL- IO gene expression is
stirnulated by IL42 trertment in mice during brusiliensis infection in which there was
l o prolonged w o m survival and reduced IgE, MMC and eosinophil responses (Fihicehan
et al., 1994). SimilarIy, Th2-type mechanisms of resistance to nematode parasites rnay be
independent of I L 4 as shown in experiments in which the neutraiization of endogenous IL-
5 fyled to S e c t parasite loads in T. spiral& H. polygynrs or brailiensis Sections even
though anti-IL4 antiiody prevented eosinophüia in blood and lung mucosa (Herndon &

Kayes, 1992; F i e m a n et al.. 1997). This selective impact of dietary protein deficiency on
host immune function has also been reported in studies on cytokine production m murine
tuberculosis (Chan et al.. 1996) and on neutrophü mobïiization during staphyIococcal
infection (Nwankwo et al., 1985). Taken together, the dinerential effects on ni2 cytokines
suggest protein deficiency impairs specinc components of Th2 cytokine response (a-4
production in intesthi lymphoid tissues) that are important for develo pment of acquired
immunity to H. polygyrus.
The significant effect of t h e on Th2 cytokine response profiles suggests that
kinetics of cytokine production during chdenge infection are important to the regulation of
host protection. Cytokines are likely most effective in promoting effector responses during
the first week of infection when H. polygynrs is in its tissue-dwehg b a l stage which is
h o wn to elicit the most intense expression of acquired immunity in mice (Wahid &
Behnke, 1992). Indeed. we found that Th2 cytokine concentrations in MW were higher
than in spleen and bat levels graduaily decline after 14 days pci Consistent with these
tirne-dependent effects of protein deficiency, Chan et al., (1996) detected depressed
cytokine mRNA expression in lungs of protein deficient mice only at early tirnes (cl4 days)
afier tuberculosis infection. In our study, the early intense response in local lymphoid tissue
refkcts the strong antigenic stimuli provided by the invasive and tissue-resideni I;trvae.
Once the worm matures and enters the pt lumen, there is much less antigenic stimulation
to gut-assochted lymphoid ceils for cytokine synthesis. Also, the presence of adult worms
in the anterior duodenum has been associated with suppression of mucosal mastocytosis
(Dehwi & Wakeh, 1988) and selective down-reguktion of IL-9 and IL-10 production
(Behnke et al., 1993), leding to the conclusion thot adult parasites are capable of
compromishg host immune responses. Taken together, these results suggest that acquired
mistance to H. po lygym requires elevated Th2 cytokine production in intestinal lymphoid
tissues within one week pci before adult parasites emerge into the gut lumen. During
challenge mfection with polygyrics, protein deficiency modified IL4 production by MLN
only during the first week pci in a selective manner depending on the seventy of restriction.
The MLN ceUs of marginal protein mice were able to rebound nom initiai impaired I L 4

synthesis (day 3 pci) whereas the MLN ce& of the protein deficient mice were unable to
secrete sufficient quantities of IL-4 at a slightly hter t h e point (day 6 pci). Tbis hter time
point rnay have been critical for stimuhtion of subsequent Th2 effector responses because
peak serum IgE and MMC responses occurred on day 9 pci Moreover, we found that
basal IL4 production by MLN of mice fed 24% protein was high prior to challenge
infection and remained elevated during the first week pci but that IL-4 secretion in MLN of
mice fed 3% protein was low at baseline and had f i d to increase after challenge infection.
Therefore, susthed elevations in IL-4 production in the early phase of challenge infection
rnay be equally or more important for priming of effector rnechuiisrns thui transient
increases at any specific pci tirne point.
It is imponant to note that Th2 cytokine production by spleen and MLN w u
prominent early after primvy infection as suniluly reported by Svetic et al. (1993) and Shi
et al. (1997). However, this intense primary cytokine response was no t associated with
subsequent mvked increases in IgE, eosinophila or mucosal mastocytosis which were
significantly higher afier chdenge infection. Secondary IL-4 and L- 10 cytokine responses
to H. polygym were relatively low CO mpared to primuy levels and elevated levels were
limited to the first week of chailenge infection. It is possibie that the priming or initiation of
Th2 cytokine synthesis rnay be a more important immunological event dian secondary
responses for the developrnent of B ceii or Th2 memory and phenotypic expression.
However, IL-4 produced during chaenge but not primary infection with H. polygynrr is
required to generate and sustain secondary IgE responses which denve mostly from naive
or uncomrnitted B ce& rather than from IgE-committed memory B cells produced during
prirnary infection (Fiinkelrnan et ai., 1990; Katona et ai.. 1991). Therefore, the impaired
production of gut-associated IL4 in protein deficient mice observed early in challenge
infection rnay be the determinhg factor responsible for the subsequent impaired IgE, MMC
and intesbd eosino p hi1 responses, and in tum for the prolonged wonn s u ~ v a i

The reciprocal amounts of IL-4 and IFN-y produced duMg challenge infection may
be an additional factor affecthg the magnitude of Th2 Unmunity raised against nematode
parasites. IL-4 and IFN-y are mutuaily antagonistic for Th ceii differentiation and effector
activity belonging to reciprocal Th ceil phenotypes. IL-4 inhibits cytokine spthesis by Th1
cek and impairs macrophage function (Kopf et al., 1993; Tan& et al., 1993; Lagoo et
al., 1994) whereas IFN-y suppresses the production of Th2 effectors and cytokine and
reverses host resistance to nematode parasites (Maggi et al., 1992; King et al., 1993; Urban
et ai.. 1993). Collectively, these data indicate that a high level of IL.-4 and the concomitant
Iow quantity of IFN-y ultimately influence the appropriateness. speed and magnitude of
Th2 immune responses raised against nemtode parasites. We found that protein deficiency
decreased gut-associated IL4 response (protein level and mRNA expression) and increased
both gut-assochted and systemic IFN-y production (protein level on day 14 pci). Although
we did not detect effects of diet on IFN-y gene expression. it is possible that protein
defkiency aitered other steps in the IFN-y synthesis pathway such as mRNA tnnsiation or
post-translational modification. The up-reguhted IM-y production duMg chaîienge
infection coincided with decreasing K-4 leveis in MLN and thus may be a by-product of
decreasing antigenic stimulation of Thtassochted responses within gut mucosal tissues.
Despite an overall nonsignincant effec t of diet on IFN-y throughout challenge Section. we
detected signincmtiy higher IFN-y production in spleen and MLN of mice fed 3% protein
compared to controk at day 14 pci During the initial 2 weeks pci, a criticai the for
developrnent of appropriate and sufficient Thî responses, the increased IFN-y coupled
with decreased IL-4 rnay have contnbuted to the lower Th2 enector responses in the
protein deficient mice. The mech;uiisrns by which protein deficiency could up-regdate
nonpro tective IFN-y and simultaneously dom-regulate protective IL4 are unknown but
these results suggest that malnutrition aiters the balance of cytokine responses that
r e m t e IgE-dependent effector mechanisms during challenge infection with H. polygyw.

The selective impact of protein malnutrition on host imrnunity was further
evidenced in the graded responses to varying levels of dietary proteh We found that 3%
proteh deficiency signincantly decreased body weight gain, rehtive spleen and MLN
weights and BUN concentration but h d no effect on piasrna aibumin concentration despite
a feeding period of 10 weeks. The absence of hypoalburninaemia in Our mice fed 7% or 3 1
protein is consistent with previous experiments in our labontory using similar diet and
infection protocols (Boulay et al., 1998) and with protein restriction in rabbits (Weidel et
of., 1994). Although biochemical indicators of pro tein deficiency typicdy have included
levels of transport proteins. urea nitrogen and hematocrit (Golden, 1982), plasma albumin
concentration has k e n regarded classically as an index of long-terni protein status
(McFuhe et al.. 1969). However, recent studies on pro tein deficiencies in pigs and
rabbits show that phma albumin concentration does not refiect albumin turnover nor
provide information on the mechanisms of how the phma albumin pool is maintained
(Jahoor et al., 1996). Several studies have reported that protein deficiency significmtly
reduces the fiactional rates of albumin synthesis and catabolism (Kirsch et al., 1968; Jahoor
et al., 1996), but the rate of dburnin turnover was sirniiar between control and protein
deficient animais (Weidel et al., 1994). These metabolic adaptations in aibumin turnover
rate m a y e x p h the unchanged plasma albumin concentration duMg protein deficiency
despite obvious impairment of growth and other physiological functions. Immune indices
are P sensitive parameter of nutritional status and help define the bc t iond consequences
of malnutrition by predicting morbidity and diseme outcornes (Chandra, 199 1). To this end,
Our results show ihat impaired immune fùnction occurs prior to detectable changes in
plasma aibumin concentration. Also, we cm suggest that the smakr body weight in the
protein deficient mice may have ennbled these mice to maintain plasma albumin homeostasis
at the expense of pro tein synthesis for other hinctionai capacities such as host
immunocompetence. Indeed, the suppressed Th2 immunity observed in H. polygytus-
inlected mice fed 3% protein confhned that we had created a functionaî protein deficiency
without concunent reductions in plasma albumin concentration.

Findy, this study is important because it suggests that a certain degree of
physiological adaptation occurs with marginal malnutrition but there is a threshold of
nutritional deficiency below which immune function is signincantly compromised. Mice fed
a marginal protein diet (78) were capable of eliminating rnost of their parasites by day 28
pci and showed intermediate but adequate b e l s of IgE, MMC nurnbers. and eosinophilia.
In contrast, a further decrease in dietary protein level caused obvious defects in host
protective immunity at both intestinal and peripheral b e l s which lead to increased parasite
s u ~ v a l . The74b protein group adopted to their marginal protein intake by eating more
food which resulted in these mice ingesting more energy than the control and deficient
groups. This was confmed by the higher plasma albumin concentration in the 7% mice
compared with the 3% group. Increased energy intake rnay spare endogenous protein for
utilkation in protein synthesis and allow nitrogen balaace to be achieved duMg Wied
protein intake (Waterlow, 1986). Indeed, mmy of the nutritional and irnmuno1ogical
outcornes of the 7% mice were not statisticdy different h m rhose of the 24% rnice,
suggesting that the higher energy consurnption compensated for the margind protein intake
and in tum help protect these 7% mice against reidection with H. polygynrs.
A comparison of Mmunological responses between the 7% and 3% protein groups
rnay heIp to clarify the signiacance of Th2 effector responses in protective imrnunity against
nematode parasites. Previous studies in which a single pumeter of Th2 defense
mechuiisms was inhibited by neuualipng antibodies (e.g. Urbm et al., 1991) or deletion o f
a cytokine-specific gene (e.g. Takmoto et d, 1997) were inconclusive as to whether Th2
eEectors such as IgE, MMC and eosinophüs were hinctionaiiy important in host resistance
to nematode parasites, but nised the possibility that there are some redundancies in host
immune hinction. Our study examining multiple parameters and tissue compuments of
host immunity suggests that Th2 effector and cytokine defenses act collectively andor
synergisticdy to promote acquired resistance to H. polygym. The rnarginaily resvicted
mice were capable of eliciting intermediate bels of Th2 effector responses b t were
apparently effective for rapid parasite expulsion In conuast, the 3% mice had lower gut-
associated IL-4, higher gut-associated and spkntc IFN-y. and consistently decreased IgE,

MMC and intesiind eosinophil responses during challenge infection which cumuiatively
may have resulted in theY higher parasite burdens.
In summary. Our study demonstrated that pro tein deficiency promo tes parasite
s u ~ v a l by dtering cytokine and effector responses in a seiective manner depending on the
specifc tissue cornpartment, the level of protein restriction and the time point of infection.
We found that protein deficiency w u more detrimental to IL-4 production in MLN than in
spleen during the early phase of a chdenge infection which m q have k e n responsible for
the subsequent decreased IgE, MMC and gut eosinophü responses. Based on Our results,
we suggest that protein deficiency exerts its adverse effecu at the following steps dong the
cascade of immunological responses to a Th2-induchg parasite. Adequate intake of
protein is necessary for mRNA expression and protein synthesis of TL-4 in GALT but does
not S e c t IL-5 or IL-10 protein production which appeared to be normal even in mice with
limited capacity to produce IL-4. This decreased IL-4 combined with increased IFN-y led
to reduced levels of IgE, MMC and gut eosinophils and in tum to prolonged parasite
survivai in the protein deficient mice. That intestinal MMC and eosinophil responses were
irnpaired in the 3 8 mice despite adequate production of IL-5 and IL40 suggests that
dietary protein affects cytokine-independent pathways in the cellular prolifention or
traficking of immune effectors. F m d y , the integration of protein deficiency into the Hp-
mouse system provided evidence thût, among the many cytokines and effectors elicited
during m Hp infection, the set of IL-Mependent, IgE-mediated MMC and eosinophil
responses in the gut mucosa is more cntical to host protection than IL-5, L- 10, peripherd
eosinophils or IgGI. Broadly, we conclude thrt dietvy protein deficiency predisposes
individu& to acquire higher levels of nematode uifection by suppressing specinc Th2
immunity at the intestinal kveL By contributhg to the understanding of the immunologicai
mechanisms of nematode parasite s u ~ v a l in protein-depleted popuiations, Our results
reved immunological targets and nutritional strategies that may help improve the naturd
abilities of the host immune system to protect against giutrointestinal nematode parasites.

This research was supported by the Naturd Sciences and Engineering Research Council of
Canada (NSERC) Collaborative Project Grant (0163762) and NSERC Research Grants
(A3624 and A3585). Research at the Institute oEParasitobgy is hnded in part by the
Fonds FCAR pour l'aide et le soutien à ia recherche, a Quebec provincial hnding agency.
The authors thank Famineh Jalili for her expert technical assistance.

REFERENCES
Behnke, J.M & Robinson, M. (198 5). Genetic control of immunity to Nematospiroides dubius: A 9-day anthelmintic abbrevhted immunizing regime which separates weak and strong responder strains of rnice. Parasite IrnmunoL 7: 235-253.
Behnke, J.M., Wahid, F.N., Grenas, R.K., Else, KJ., Ben-Smit, A.W. & Goyai, P.K. (1993). Immun010 gical relationships during primary infection with Heligmsomoides polygyrus (Nematospiroides dubius): downregulation of specinc cytokine secretion (n-9 and IL-IO) correhtes with poor mastocytosis and chronic s u ~ v a l of adult w o m . Parasite ImmunoL 15: 415-421,
Boulay, M., Scott, M.E., Conly, S.L., Stevenson, M.M. & Koski, K.G. (1998). Dietary protein and zinc restrictions independently rnodify a Heligmsomoides polygyrur (Nematoda) infection in mice. Parasitology 116: 449-462.
Bradley, L.M., Yoshimoto, K., & Swain, S.L. (1995). nie cytokines IL-4. IFN-y, and IL,-12 regulate the development of subsets of memory effector helper T cells in vitro. J. Immunol. 155: 17 13-1724.
Chan, J., Tian, Y., Tanaka, K.E., Tsang, M.S., Yu, K., Salgame, P., Carroll, Da, Kross, Y., Teitelbaum, R., & Bloom, B.R. (1996). Effects of protein cdorie malnutrition on tuberculosis in mice. Proc. Natl. Acad. Sci USA 93: 14857- 1486 1.
Chandra, R.K. (1991). Immunocompetence is a sensitive and hinctional barorneter of nutritional statu. Acta Pediatr. Scand. SuppL 374: 129-132.
Chen, X.J. & Enerback, L. (1994). IgE receptors, IgE content and secretory response of mast cells in athymic nts. Imrnunology 83: 595-600.
Chen, X.J. & Enerback, L. (1996). Immune response to a Nippostrongylus brariliensis infection in athymic and euthymic rau: surface expression of IgE recepton, IgE occupancy and secretory abiiity of mast cells. Int. Arch. Ailergy & Imrnunol. 109: 250- 257.
Cof'finan, RL., Ohara, J., Bond, M.W., Carty, J., Wotnik, A., & Paul, W.E. (1986). B ceii stimulatory factor4 enhances the IgE response of lipopolysacchuide-activated B ceUs. J. ImmunoL 136: 4538-4541.
Codlman, RL., Seymour, B.W.P., Hudak, S., Jackson, J., & Rennick, D. (1989). Antibody to interleukin-5 inhibits hekninth-induced eosinophilia in mice. Science 245: 308-3 10.

Crampton, D.W.T. (1986). Nutritional aspects of infection. Trans. R Soc. Trop. Med. & Hyg. 80: 697-705.
de Andres, B., Rakasz, E., Hagen, M., McCormik, ML., Mueller, A.L., Elliot, D., Metwail, A., Sandor, Britigan, B.E., Weinstock, J.V., & Lynch, RG. (1997). Lack of Fc-E receptors on murine eosinophils: implications for the functional significance of elevated IgE and eosinophils in pansitic infections. Blood 89: 382693836.
Dehfawi, M.S., Wakeün, Dm, & Behnke, J.M. (1987). Suppression of rnucosd mastocytosis by infection with the intestinal nematode Nematospiroides dubius. Parasite ImrnunoL 9: 187-194,
Dehlawi, M.S. & Wakelin, D. (1988). Suppression of mucosal mastocposis by Nemutospiroides dubius results from an adult wo rm-mediated effect upon hos t lymphocytes. Parasite ImmunoL 10: 85-95.
Finkelman, F.D., Katona, LM., Urban, J.F., Holmes, J., Ohara, J., Tung, A.S., vG Sample, J., & Paul, W.E. (1988). IL4 is required to generate ruid sustain in vivo IgE responses. J. Immunol. 141: 2335-234 1.
Finkelman, F.D., Holmes, J., Katona, LM., Urban, J.F., Beckmann, M.F., Park, L.S., Schooley, K.A., Cof'fman, R.L., Mosmann, T.R., & Paul, W.E. (1990). Lymphokine convol of in vivo immunoglobulin isotype selection. Annu. Rev. ImmunoL 8: 303-33.
Finkelman, F.D., Madden, K.B., Cheever, A.W., Katona, LM., Morris, S.C., Gately, M.K., Hubbard, B.R., Gause, W.C., & Urban, J.F. (1994). Effects of interleukin 12 on immune responses and host protection in M c e ~ e c t e d with intestinal nematode parasites. J. Exp. Med. 179: 1563-1572.
Finkelman, FaDe, Sheo-Donohue, T., GoldhilI, J., Sullivan, CA., Morris, S.C., Madden, &B., Gause, W.C., & Urbon, J.F. (1997). Cytokine regulation of host defense against parasitic g~uointestinal nematodes. Annu. Rev. Immunol. 15: 505-533.
Golden, MaHoNo (1982). Transport proteins as indices of protein status. Am. J. Clin. Nutr. 35: 1 159- 1 165.
Hemdon, F.J., & Kayes, S.G. (1992). Depletion of eosinophüs by anti-IL5 monoclond antibody treatment of mice infected with Trichinella spiralis does not alter parasite burden or irnmunologic resistance to reinfection J. ImmunoL 149: 364203647.
Jahoor, F., Bhattipmlu, S., Del Rosario, M., Bumn, D., Wykes, L., & Frazer, M. (1996). Chronic protein deficiency dEerentialIy atrects the kinetics of plasma proteins Pi young pigs. J. Nutr. 126: 1489-1495.

Katona, LM., Urban, J.F., Kang? S.S., Paul, W.E., & Finkelman, RD. (1991). IL-4 requirements for the generation of secondary in vivo IgE responses. J. Imrnunol. 146: 4215-4221.
Keymer, AwEm & Tarlton, A.E. (1991). The population dynmics of acquired immunity to Heligmosomoides polygyms in the laboratory mouse: suain, diet and exposure. P~asitology 103: 12 1- 126.
King, CL, Low, CC., & Nutman, T.B. (1993). IgE production in humm helminth infection: reciprocai intenelationship between IL-4 and IFN-y. J. Imrnunol. 150: 1873- 1880.
King, CmLm, Xianli, J., Malhoba, I., Liu, S., Mahmoud, A.A.F., & Oettgen, HwC. (1997). Mice with a targeted deletion of the IgE gene have increased worm burdens and reduced grnulornatous infiammation following primvy infection with Schistosoma mansoni. J. Immunot. 158: 294-300.
Kirsch, Rw, Frith, L., Black, E., & Hoffenberg, R. (1968). Regulation of albumin synthesis and catabolism by alteration of dietary protein. Nature 217: 578-579.
Kopf, M., Le Gros, G., Bachmann, M., Lamers, M.C., Bluethmnnn, H., & Kohler, Gw (1993). Disniption of the munne IL-4 gene blocks Th2 cytokine responses. Nature 362: 245-248.
Lagoo, A.S., Eldridge, J.H., Lagoo-Deenadaylan, S., Black, C.A., Ridwan, B.U., Hardy, K. J., McGhee, J.R., & Beagley, K. W. (1994). Peyer's patch CD8' memory T ceh secrete T helper type 1 and type 2 cytokines and provide help for irnrnunoglobuin secretion. Eur. J. IrnmunoL 24: 3087-3092.
Maggi, E., Paronchi, P., Manetti, R., Simonelü, C., Piceinni, Mrn-Pw, Rugiu, F.S., de Carli, M., Ricci, M., & Rornagnani, S. (1992). Reciprocal reguliitory effects of IFN-y a d IL-4 on the in vitro development of hurnan Th 1 and Th2 clones. J. IrnmunoL 148: 214292147.
McFarlane, H., Ogbeide, ML, Reddy, S., Adock, K. J., Adeshina, Hm, Gurney , J.M., Cwke, A., Taylor, G.O., & M o d e , J.A. (1969). Biochemical assessrnent of protein- calorie malnutrition. The Lancet 1: 392-394.
Monroy, F.G. & Endquez, F J. (1992). Heligmsomuides polygynrs: A mode1 for c hro nic gasuointestinai helrninthiasis. Parasito L Today 8: 49-53.
Mosmann, T.R 8 Sad, S. and more. ImmunoL Today
(1996). The expanding universe of T-cell subseu: Thl, Th2, 17: 138-146.

Nakshabendi, LM., Obeidat, W., Russell, RmL, Downle, S., Smith, K., & Rennie, M.J. (1995). Gut mucosai protein synthesis measured using htravenous and intragastric deiivery of stable tracer unino acids. Am. J. PhysioL 269: E996-E999.
Negrao-Correa, Dm, Adams, L.S., & Bell, RG. (1996). Intestinai transport and catabolism of IgE. J. Immunol. 157: 4037-4044.
National Research Council (NRC). (1995). Nutrient requirements of the mouse. In: Nuvient Requirements of Labontory Animals. Fourth Revised Edition. National Academy of Sciences, Washington, D.C.
Nwankwo, M.U., Schuit, K.E., & Glew, R.H. (1985). Effects oEmrtemd protein deprivation on the nutritional status and neutrophil function of suckling neonatai rats. J. Infect. Dis. 5 1 : 23-32.
Pawlowski, ZmSa (1984). Implications of parasite-nutrition interactions from a world perspective. Fed. Proc. 43: 256-260.
Pritchard, DaL, Williams, DDJ.Le, Behnke, J.M.9 & Lee, T.D.G. (1983). The role of IgGl hypergammaglobulinaemin in immunity to the gastrointestinal nematode Nematospiroides dubius. The immunochemicd purification. antigen-specificity and in vivo anti-parasite effect of IgGl from immune serum. Immunology 49: 353-365.
Ramaswarny, K., Hakimi, J., & Bell, R.G. (1994). Evidence for an interleukin-4- inducible immunoglobulin E uptake and transport mechanism in the intestine. J. Exp. Med. 18û: 1793-1803.
Rincon, M., Anguita, J., Nakamura, T., Fikrig, E., & Flavell, R.A. (1997). Interleukin (a)-6 directs the differentiation of Ldproducing CD4+ T cells. J. Exp. Med. 185: 46 1-469.
Shi, H.N., Koski, K.G., Stevenson, M.M., & Scott, M.E. (1 997). Zuic deficiency impairs T ceil function in mice during both primuy and challenge infection widi Heligmosomoides polygyrus (Nematoda). Parasite Imrnunol. 19: 363-373.
Slater, A.F.G. & Keyner, A.E. (1988). The influence of protein deficiency on immunity to Heligmosomoides polygyrur (Nematoda) in mice. Parasite ImmunoL IO: 507-522.
Svetic, A., Madden, K.B., di Zhou, X., Lu, P., Katona, LM., Finkelman, F.D., Urban, J.F., & Gause, W.C. (1993). A primary intestinai helminthic infection rapidly induces a gut-associated elevation of Th2-associated cytokines and IL-3. 1. Immunol. 150: 3434.

Takamoto, M., Ovington, K.S., Behm, C.A., Sugane, K., Young, LG., & Matthae, K.1. (1997). Eosinophilia, parasite burden and lung damage in Toxocara cmis infection in C57BY6 mice geneticdy deficient in IL-5. Immunology 90: 5 1 1-5 17.
Tanaka, T., Hu-Li, J., Seder, R.A., Fazekas de St. Groth, B., & Paul, W.E. (1993). 1nterleuk.i.n 4 suppresses interleukin 2 and interfemn y production by naive T ceils stimulated by accessory celi-dependent receptor engagement Roc. Natl. Acd. Sci. USA 90: 5914-5918.
Toru, Hm, Ra, C., Nonyama, S., Suzuki, K., Yata, J.-I., & Nakahata, T. (1996). Induction of the high-affinity IgE receptor (FceRI) on human mûst ceUs by IL4 Int. Immunol. 8: 1367- t 373.
Urban, J.F., Katona, LM., Paul, W.E., & Finkelman, FD. (199 1). Interleukin 4 is important in protective immunity to a gastrointestinal nematode infection in mice. Froc. Natl. Acad. Sci, USA 8%: 35 13-35 1%
Urban, J.F., Madden, &B., Cheever, A.W., Trotta, P.P., Katona, LM., & Finkelman, F.D. (1993). IFN inhibits infiunmûtory responses and protective immunity in mice infected with the nematode parasite, Nipposmnglyzu brusiliensis. J. Immunol. 151: 7086-7094.
Urban, J.F., Maliszewski, Cor., Madden, K.B., Katona, LM., & Finkelman, F.D. (1995). IL-4 treatment cm cure established gastrointestind nematode infections in irnrnunocompetent and immunodeficient rnice. J. ImrnunoL 154: 4675-4684.
Vitetta, ES., Ohara, J., Myea, C., Layton, J., Krammer, P.H., & Paul, W.E. (1985). Serological, biochemicai and functionai identity of B ceiI-stimulatory factor 1 and B ceii differentiation factor for IgG1. J. Exp. Med. 161 : 1726-3 1.
Wahid, FaNa & Behnke, J.M. (1992). Stimuli for acquired resistance to Heligmosomides polygyrus from intestinal tissue resident L3 and LA b a e . IntL J. Parasitoi. 22 (6): 699-7 10.
Wahid, F.N. & Behnke, J.M. (1993). Immunological relationships during primary infection with Heligmosomoides polygynrr (Nematospiroides dubius): parasite-specific IgG 1 antibody responses and primary response phenotype. Parasite Immunol. 15: 40 1 - 413.
Wahid, F.N., Behnke, J.M., Gnncis, RK., Else, KJ., & Ben-Smith, A.W. (1994). Immunological niationships during primary infection with Heligmosomoides polygyrus: Th2 cytokines and primary response phenotype. Parasitology 108: 46 1-471.

Wastüng, J.M., Scudamore, CL., Thornton, E.M., Newlands, G.F. J., & Miller, HmRPw (1997). Constitutive expression of mouse mast cell protease- l in nomal BALBlc mice and its up-regulation during intestinai nernatode infection. Immunoiogy 90: 308-3 13.
Waterlow, J.C. (1986). Metabolic adaptation to Iow intakes of energy and protein. Annu. Rev, Nutr. 6: 495-526.
Weidel, S.E., Smith, G., & Fleck, A. (1994). The effects of experimental malnutrition on albumin metabolism and distribution in mbbits. Br. J. Nutr. 72: 369-384.
Windhagen, A., Anderson, D.E., Camzosa, A., Wilüams, R.E., & Haflet, D.A. (1996). IL- 12 induces human T cells secreting IL- 10 with IFN-y . J. ImmunoL 157: 1 127- 1131.
Woodbury, R.G.9 Miller, H.R.P., Huntiey, J.F., Newlands, GwFwJa, Palüser, A.C. & Wakeiin, D. (1984). Mucosal mast ceils are hinctionally active during spontmeous expulsion of intestinal nematode infections in rat. Nature 312: 450-452.
Wykes, LJ., Fiorotto, M., Bumn, D.G., Del Rosario, M., Frazer, M.E., Pond, W.G., & Jahoor, F. (1996). Chronic low protein intake reduces tissue protein synthesis in a pig mode1 of protein malnutrition J. Nutr. 126: 148 1- 1488.
Yamaguchi, Y., Suda, T., Suda, J., Eguchi, M., Miura, Y., Harada, N., Tominaga, A., & Takatus, K. (1988). Pudied interleukin 5 supports the terminai differenthtion and proliferation of muMe eosinophilic precursors. J. Exp. Med. 167: 43-56.
Zhang, Xe, Werner-Favre, Cm, Tang, Ha, Brounvers, N., Bonnefoy, J.Y., & Zubler, RwHw (1991). IL-Uependent IgE switch in membrane IgA-positive human B celIs. J. Immunol, 147: 300 1-3004.

CHAPTER IV
GENERAL DISCUSSION
Protein malnutrition and gastrointestinai nematode infections are chronic diseases
thar are frequentiy endemic in populations of the developing world. Protein deficiency i s
associated with irnpaited host irnmunocompetence; however, the specific mechanisms by
which protein deficiency may influence the duration and severity of an intestinal nematode
infection are cornplex and poorly undentood. Inasmuch as host resistance to intestinai
nematode infections depends on the development and maintenance of Th2-type cytokine
and effector responses. we investigated whether protein deficiency exacerbates suMval of
a murine nematode puasite. Heligmsomoides polygym. by impairing parasite-induced
Th2 immunity in systemic a d o r intestinal cornpartments. Previous studies on the
interactions of protein deficiency, nematode infection and host immunity showed that
increased parasite s u ~ v a l was associated with depressed antibody (IgG, t o d IgGl) and
eosinophil responses. The effects of pmtein deficiency on secondary effector responses in
the GALT (e.g. MMC, intestinal eosinophilia) and on gut-associated or splenic cytokine
production have not been characterized. Our study w u designed to address these current
gaps in knowfedge as well as to asscss the temponl and comparmientalization aspects of
protein malnutrition and host immunity to nematode parasites. The major finding of rhis
study was that protein deficiency at the 3% level promoted parasite suMval in previously
immunized mice in association with altered cytokine responses to H. polygyrus (Le. up-
regulated Th l and down-regulated Th2) and impaired proliferation and activation of Th2
effectors within 2 weeks pci. Specifcaily. protein deficiency decreased gut-associated IL-
4 production, increased gut-associnted and systemic IFN-y synthesis, and inhibited
increases in intestinal and serum Th2 effector responses that were observed in weil-
nourished hosts during challenge infection. Interestingly, protein deficiency appeved to
exert differential site- and type-specîf?c effects on cytokine and effec tor responses to H.
po lygym challenge infection. For exarnple, protein deficiency inhibited the parasite-
elicited responses of intestinal eosinophils, IgE, MMC and I L 4 but did not signincmtly
depress peripherai eosinophilia, parasite-specifc IgGl synthesis, or production of other

Th2 cytokines (IL4 and IL-IO). Also, the intestinal cornpartment (gut eosinophils. IL-4
production in MLN) w u more dversely affected by protein resmction than systemic
responses in periphed circulation or spleen. The responses that were profoundly
irnpaired during protein malnutrition (n-41IFN-y balance, elevated IgE titres, mucosai
mastocytosis) are those known to be the most important markers associated with host
protective immunity to H. polygyrus. These impaired responses observed in the protein
deficient mice were detected by 3 days pci (e.g. IgE) until2 weeks pci (e-g. up-regulated
IFN-y), suggesting that functional Th2 immunity is most effective at terminating the
infection when activated at the e d y stage of challenge infection before the parasite
matures and migntes to the intestinal lumen. Once a worm reaches the gut lumen it is less
in contact with immune effectors in the gut mucosa and thus is capable of avoiding the
potentially dvnaging mediators of host immunity. Also. the level of protein restriction
Uifluenced the magnitude of host protection and parasite survival such that mice fed a
marginal protein diet were able to mobilize adaptive rnechanisms (e.g. increased total food
intake) which may have enabled them to mount an effective immune response and in tum
eliminate a challenge infection. However, a further decrease in dietary protein level from
7% to 3% caused profound defects in secondary Th2 immune responses as well as
significuitly prolonged parasite survival.
This study is the fint cornprehensive investigation of the interactions between
protein deficiency, Th2 immunity and nematode pansitic infection. and the results reveal
sevenl points for further investigation. Cytokine production in MLN likely reflects the
activities of lymphocytes in the Peyer's patches and cryptopatches of the gut mucosa that
are in more direct contact with parasite antigens. Considering that MLN receives
immunological components from both intestinal tissues and the periphenl bloodstream, a
definitive study of malnutrition-associated changes in cytokine production would require
measurement of cytokine production by lymphocytes in Peyer's patches or other epithelial
lymphoid tissues. Also, it would be important to determine the kinetics of cytokine
production dunng primary infection in order to ascertain whether primary cytokine
responses direct the magnitude and direction of secondary cytokine Rsponses and
resultant effector mechanisms. Moreover, the potential for imrnunornodulation by protein

malnutrition early after challenge infection (Le. 2 weeks pci) suggesu that nutritional
interventions (e.g. refeeding) or immunological therapy (e.g. I L 4 treaûnent) may be most
successful when implemented before or immediately following reinfection with nematode
parasites. Lady , the differentiai outcornes of the marginal protein group (7%) and the
protein deficient group (3%) indicate that total energy or cdoric intake may have an
important impact on host immunocompetence irrespective of, or in conjunction with, the
level of dietuy protein. The greater food intake by the marginal protein group which had
successfully terminated a challenge infection suggesu that one revonably cost-effective
and feasible stntegy to improve host resistmce to nematode parasites is to increase the
amount of energy consumption without a concomitant supplementation of dietvy protein.
Broadly, our study supports the premise that protein deficiency exacerbates the
suMvd of a gastrointestinal nematode pmsite by decreasing gut-associated L-4 (Th2)
and increasing systemic and gut-associated IFN-y (Thl) early in the challenge infection.
leading to reduced gut and systemic Th2 effector responses. These data contribute to Our
understanding of the immunologicd mechmisms of parasite survivai in proteindepleted
populations. Nso, the results reveal immunological mgeu and tissue compartments for
thenpeutic modulation in order to improve the nauiral abilities of the host immune system
to respond to intestinal nematode pansites. Cleuly. a consideration of the nutritionai
status in the population is essentid to the success of vaccination programs Ymed at
enabling npid and strong acquired immunity to secondary parasitic infections.

APPENDICES
APPENDIX A: RESULTS FROM TWO-WAY ANOVA ON EFFECTS OF DIET, TIME, & DIET BY TIME INTERACTION*
APPENDIX B: RESULTS FROM ONE-WAY ANOVA ON MAIN EFFECT OF DIET.
APPENDIX C: RESULTS FROM ONE-WAY ANOVA ON MAIN EFFECT OF TIME*

APPENDIX A: Results from two-way ANOVA analyses of diet, tirne and diet*time interaction main effects on panmeters measured postprimary infection BPI) or postchallenge infection (PCI). Numbea in parentheses are degrees of kedom.
Variable I Tinte
Worms - PPI
Worms - PCI
BI& Eosinophils - PPI BI& Eosinophils - PCI
S e m IgE - PPI
Semm IgE - PCI
Semm IgG1- PPI F (2.8 1) d .74 1 Pd.48 17
MMC - PPI
F (2.24) = 1 .O6 PPI GutEosinophiIs~ 1 M.3615

APPENDLX A continued. --
1 Variable 1 Die t Time
I IL-5 Spleui - PCI F (2, 109) = 2.41 I P=0.0948
I IL-5 MLN - PCI F (2, 101) = 0.02 1 P4.9839 1 P=0.0001 F (3, 101) = 16.74
1 iFN-y MLN - PCI F (2.92) = 0.35 I ~=0.7031

APPENDM B. Results nom one-way ANOVA analyses on the main effect of diet on parameters meaured at each postprirnary (PPI) and postchaknge (PCI) time point and in
1 Variable
1 Worm Burdens - P6
1 Worm Burdeos - C6
1 Worm Burdcns - Cl4
1 Worm Burdens - C28
1 Puasite-spedfic IgGl - P6

APPENDIX B continued.
1 Variable Die t
1 MMC - Cl4
1 MMC - C28
1 Gut Eosimphils - Naive
1 Gut Eosinaphils - P6
1 Gut Eosinophils - P28
1 Gut Eosinophils - C9 1 Gut Eosinophils - Cl4
I Gut Eosinophils - C21

Variable I
MMCP-1 - Naive
1 I L 4 Spleen - P6
MMCP- 1 - C6
MMCP- 1 - C 14
. - - . - - -
IL4 Spleen - C6 IF(2.29) = 1.91 id. 1665 1 - 1
Diet
F(2,6)=1.64 Pd.2705
Posthoc cornparisons J
F (2.47) = 4.37 P=O.O182
F (2.47) = 5.41 P=0.0077
1 IL4 Spleen - C6 1 F (2.30) = 0.39 P=0.6820 1 1
3% < 24%; 7%=345 & S M
1
3% c (24% = 7%)
IL4 Spleen - C 14 IL4 Spleen - C28
L
1 1L-5 Spleen - Cl4 1 F (2.27) = 1.59 Pd.2229 ( 1
F (2 .3 ) = 2.39 P=0.1126
F (2.20) = 0.18 P=0.8334
1 L i 0 Spleen - C6 1 F (2.30) = 3.14 P4.0576 1 1
1
IL-10 Spleen - P6 I
IL40 Spleen - C3
( ILI0 Spleen - Cl4 1 F (2.28) = 0.76 M.4765 1 1
F (2. 32) = 2.62 M.0880
F(2.25)=0.12 P=û.8856 -
I

APPENDIX B continued.
1 Variable 1 Diet 1 Posthoccomparisons
( m - y Spleen - P6 1 F (2. 32) = 1.29 P4.2897 ( 1 IFN-y Spleen - C3 IF(225)=1.15 Pd.3319 1
IL-4 MLN - C28
iFN-y Spleen - C 14
IFN-y Spleen - CS8
1 IL40 MLN - C3 1 F (2.22) = 0.70 M.5075 1
F (2.25) = 1 1.19 P=0.0003
F (2. 24) = 3.10 P=0.0633
IL-5 MLN - Cl4
IL-5 MLN - C28
1 IL40 MLN - C6 1 F (2.26) = 1.07 M.3564 1
(24% = 7%) < 3%
F ( 2.29) = 0.72 P4.4933
F (2, 22) = 2.1 1 Pa. 1452
1 L I 0 MW - C28 1 F (2. 10) = 3.22 M.0835 1

APPENDIX B continued.
1 Variable 1 Diet 1 PosthoccompPriso~ 1
1 IFN-y MLN - C3 1 F (2, 21) = 1.48 PG.2496 1 1 EN-y MLN - P6 F (2, 11) = 0.67 Pd.5330
i'
IFN-Y MLN - C6 v
IFN-y MLN - Cl4
1 Rdative Food Intake - C28 1 F (2.47) = 77.03 P=0.0001 1 1
Body Wei@ Gain - C28 Total Food btake - C28
1 Relative S p l m Weight - C28 ( F (2.39) = 7.27 P=û.0018 1 1
F (2, 22) = 0.29 P=0.7506
F(2,29)=4.31 P=0.0230
F (2,47) = 85.35 P=O.ûûûl
F (2,47) = 16.63 P=O.OUûl
1 Relative MLN Weight - C28 1 F (2, 32) = 8.57 k0.0007 1 1
24%<3%; 7% = 3% lk 24%
RelativeThymusWeight C28
Plasma Albumin Concentration - C28
- -
F(232)=0.21 P=û.8125
F (2,37) = 4.16 P4.234û 1

APPENDM C. Remlts from one-way N O V A analyses of thne on puameters measured within each dietary group (24%, 7%. 3%) postprimary infection (PPI) or postchdenge infection (PCI).
1 Variable 1 Time 1 1 Worm Burdens - 24% PPI 1 F (1.28) = 3.93 Pd.0573 1 1 Worm Burdens - 7% PPI 1 F (1,31) = 5.25 Pd.0289 1 1 Wonn Burdens - 3% PPI 1 F (1.33) = 2.37 Pd.1333 1 1 Worm Burdens - 24% PCI 1 F (3.49) = 22.29
1 W o m Burdens - 3% PCI I F ( ~ , s ~ ) = 1.31 P=û.2815 1
hem IgE - 24% PPI
( Senun IgE - 3 1 PPI 1 F (1, 31) = 78.53
1 Serurn IgE - 24% PCI
1 Se- IgE - 7% PCI 1 F (5.69) = 9.46 P~.OOOl 1
1 Parasite-speciâc IgG1-24% PPI 1 F (1,27) = 163.41 P4.0001 1

1 MMC - 7% PPI 1 F (1, 10) = 1.59 Pd.2392 1
Variable L
MMC - 24% PPI
1 MMC - 3% PPI 1 F(1,9) =OS1 Pd.4935 1
Time
F (1.9) = 5.09 P=0.0504
( Gut Eosinophils - 24% P H IF(^, 7) =0.00 P4.9645 1
I
MMC - 24% K I 1
1 Gut Eosinophils - 7% PPI
J
F (5.21) = 2.41 P4.07 1 1
1 Gut Eosinophils - 3% PPI 1 F (1,8) = 0.35 P4.5686 1 1 Gut EosinophiIs - 24% PCI 1 F (5.21) = 3.45 P4.1960 1 Gut Eosinophils - 7% PCI
-
IF&8)=1.40 P4.2708 1 1 Gut Eosinophils - 3 2 PCI 1 F (5, 19) = 0.54 P4.7471 1
1 MMCPl - 24% PPI 1 F (1. 19) = 13.47
1 MMCPl - 7% PPI 1 F (1, 18) = 0.14 P4.7136 1 1 MMCPl - 3% PPI 1 F(1, 13) ~ 0 . 2 4 W.6307 1
1 MMCPl - 7% PCI 1 F (3,58) = 19.85
I MMCPl - 3% PCI I F (3.66) = 10.23

APPENDIX C continued.
1 Variable 1 Time 1 1 I L 4 Spleen - 2 4 4 PCI ( F (3.35) = 6.85 P=O.OO09 1
-
1 IL4 Spleen - 7% PCI
1 IL-4 spleen - 3% PCI 1 F (3,34) = 1.35 P4.2747 1 1 IL-5 Spleen - 2446 PCI 1 F (3.37) = 0.18 P4.9073 1 1 IL5 Spleen - 7% PCI 1 F (3.35) = 0.67 Pd.5761 1 1 IL-5 spleen - 3% PCI 1 F (3.37) = 0.15 P4.9308 1 1 IL10 Spleen - 245 PCI 1 F (3.32) = 2.09 P=û.1205 1 1 IL-IO Spleen - 7% PCI
1 IL-10 spleen - 3% PCI 1 F (3.3 1) = 4.56 P==O.0093 1 1 IFN-y Spleen - 2446 PCI 1 F (3.35) = 22.97 P=O.ooOl 1 ( IFN-y spleen - 796 PCI 1 F (3,37) = 31.52 P=O.OOO~ 1
- . -. ..
1 IL-4 MLN - 24% PCI 1 F (3, 33) = 19.32 P=û.ûûûl 1 1 IL-4 MLN - 7 4 PCI 1 F (3,35) = 4.56
1 IL-5 MLN - 7% PCI 1 F (3. 36) = 6.03 P=û.0019 1
1 IL- 10 MLN - 24% PCI 1 F (3,31) = 5.50 P=0.0038 1
1 IFN-y MLN - 24% PCI 1 F (3,31) = 10.18





























