Embed Size (px)
Citation preview

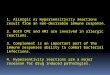
Mast cells are a key cell type in the pathogenesis of im-munoglobulin E (IgE)-dependent hypersensitivity reactions.The binding of multivalent antigens to receptor-bound IgEand the subsequent aggregation of the high-affinity Fc recep-tors for IgE (FceRI) provide the trigger for mast cell activa-tion, as shown in Fig. 1. The FceRI is comprised of four sub-units, an IgE-binding a-subunit, a signal-amplifying and re-ceptor-stabilizing b-subunit, and two disulfide-bound g-sub-units that function as the main signal transducers.2) Both theb- and g-subunits contain two tyrosine residues locatedwithin an immunoreceptor tyrosine-based activation motif(ITAM).3) Engagement of the FceRI leads to activation ofseveral Src family protein tyrosine kinases (PTKs). A Srcfamily PTK, Lyn, phosphorylates the cytosolic tyrosineresidues in the ITAMs in the b- and g-subunits.4) Phosphory-lated b- and g-ITAMs recruit Lyn and Syk, which is anotherPTK with tandem Src homology (SH) 2 domains, respec-tively. Interaction of Syk with the phosphorylated g-ITAMresults in the autophosphorylation and activation of Syk.5) Ithas recently been shown that another Src family PTK, Fyn, isalso activated as a FceRI-proximal event that is independentof Lyn activation.6) Fyn does not effectively phosphorylate b-and g-chains, suggesting that its function is distinct from thatof Lyn. These PTKs lead to the promotion of signaling com-plexes organized by adaptor proteins such as the linker foractivation of T cells (LAT).7) This initiates the downstreamactivation of signal-transducing enzymes including phospho-lipase C (PLC)g , phosphoinositide 3-kinase (PI3K), andphospholipase D (PLD). The lipid-derived messengers, inosi-tol 1,4,5-triphosphate (IP3) that mobilizes Ca2�, diacylglyc-erol that activates protein kinase C (PKC), and phos-phatidylinositol 3,4,5-triphosphate (PIP3) that recruits andactivates signaling proteins, and phosphatidic acid that regu-lates phosphatidylinositol 4,5-bisphosphate (PIP2) produc-tion, act in concert to evoke degranulation and cytokine pro-duction.8) The release of histamine and other potent chemical
mediators that have been implicated in a wide variety of in-flammatory and immunological processes is the primaryevent in acute allergic and inflammatory conditions. Cy-tokine secretion contributes to the leukocyte infiltration asso-ciated with late-phase reactions.9)
In addition to the well-defined IgE-dependent function de-scribed above, mast cells also play a prominent role in non-IgE-mediated hypersensitivity reactions. It has long beendemonstrated that mast cells are closely apposed to nerves ina variety of tissues including the skin, intestine, and duramater.10—12) Some mast cells actually form membrane–mem-brane contact with nerves in vivo.10) Nerves in associationwith mast cells often contain neuropeptides such as sub-stance P and calcitonin gene-related peptide (CGRP). Sincenerves release these neuropeptides upon stimulation andmast cells express receptors for many neuropeptides, it is
September 2005 Biol. Pharm. Bull. 28(9) 1551—1559 (2005) 1551
∗ To whom correspondence should be addressed. e-mail: [email protected] © 2005 Pharmaceutical Society of Japan
Live Cell Imaging to Study Signaling Molecules in Allergic Reactions
Tadahide FURUNO*,1) and Mamoru NAKANISHI
Graduate School of Pharmaceutical Sciences, Nagoya City University; Mizuho-ku, Nagoya 467–8603, Japan.Received May 23, 2005
Mast cells are widely distributed throughout the body, predominantly near blood vessels and nerves, and ex-press effector functions in allergic reactions, inflammatory diseases, and host defense. The activation of mastcells results in secretion of the preformed chemical mediators in their granules by a regulated process of exocyto-sis and leads to synthesis and secretion of lipid mediators and cytokines. Their soluble factors contribute to aller-gic inflammation. Mast cells are associated with hypersensitivity reactions, not only in the classical immunoglob-ulin E (IgE)-dependent mechanism but also in an IgE-independent manner. In particular, investigations of po-tential anatomical and functional interactions between mast cells and the nervous system have recently attractedgreat interest. To understand these molecular mechanisms in mast cell activation, the ability to visualize, track,and quantify molecules and events in living mast cells is an essential and powerful tool. Recent dramatic ad-vances in imaging technology and labeling techniques have enabled us to carry out these tasks with high spa-tiotemporal resolution using confocal laser scanning microscopes, green fluorescent protein and its derivatives,and image analysis systems. Here we review our investigations of the dynamic processes of intracellular signalingmolecules, cellular structure, and interactions with neurons in mast cells to provide basic and valuable informa-tion for allergy and clinical immunology using these new imaging methods.
Key words mast cell; neuroimmunology; confocal laser scanning microscope; green fluorescent protein; degranulation; allergicreaction
Review
Fig. 1. Schematic Representation of Mast Cell Activation

likely that nerve activation results in mast cell activation.Electrical stimulation of neurons induces morphologicalchanges reflecting secretion in mast cells located close to thenerve fibers.13) On the other hand, a variety of molecules aresynthesized by, released from mast cells, and can in turn in-fluence neuronal activity.14,15) Tryptase stored in mast cellgranules directly activates proteinease-activated receptors ex-pressed in neurons.16) Products of arachidonic acid metabo-lism such as cysteinyl leukotrienes and prostaglandins influ-ence the local environment involving nerves.17) Cytokines in-cluding tumor necrosis factor (TNF)-a and growth factorssuch as nerve growth factor (NGF) cause changes in localnerves which lower their threshold to activation.18—20) Thesenerve-mast cell effects are bidirectional and are postulated toparticipate in the promotion and regulation of inflammatorydiseases such as sclerosis, interstitial cystitis, and irritablebowel syndrome.21—23)
Under these situations, we have considered that the analy-sis of intracellular dynamics of signaling molecules providesimportant information from a new aspect to understand themolecular mechanisms of allergic reaction and inflammation.Accordingly, we have studied intracellular movements ofCa2� and signaling molecules with high spatiotemporal reso-lution in a living single mast cell using new microscopictechniques. Further, bioimaging has progressed extraordinar-ily during last 2 decades and has contributed to the study ofcell biology, histology, immunology, and pathology. It hasalso changed the approach to study cell signaling and cellfunction and ushered in a new era in cooperation with techni-cal progress in molecular biology, especially in cell biology.In this review, we describe our imaging analysis of calciumsignals, degranulation, transcriptional activity, and interac-tion with neurons in mast cells at the single-cell level.
CALCIUM SIGNALS
A number of biochemical events occur subsequently to theaggregation of FceRI. Activation of PLCg is an importantprocess in the signaling pathway. PLCg catalyzes the hydrol-ysis of membrane-associated PIP2, thereby generating diacyl-glycerol and IP3. IP3 diffuses rapidly from the plasma mem-brane to bind with IP3 receptors on the endoplasmic reticu-lum (ER) which serve as Ca2� channels to release luminalCa2� and generate the initial Ca2� signal phase.24) The Ca2�
depletion in the ER activates store-operated Ca2� channels(SOCs) mediating Ca2� entry and generates the second phaseof Ca2� signals.25) We showed using a confocal laser scan-ning microscope (CLSM) that single-cell imaging of Ca2�
signals provides interesting and important information in ratbasophilic leukemia (mast cell line, RBL-2H3) cells. CLSMfluorescence images of RBL-2H3 cells loaded with Fluo-3 (aCa2�-specific fluorescent probe) were dim in the restingstate. They were excited by an argon ion laser (488 nm).After stimulation with antigen, Fluo-3 fluorescence intensi-ties in individual cells increased with different lag times, andfluorescence images became more heterogeneous. The areasof bright fluorescence appeared to belong to the nucleus inRBL-2H3 cells based on the morphological pattern. We con-firmed this using a DNA-specific fluorescence probe (Hoech-est 33342) excited by a He-Cd laser (325 nm) and the probewas coloaded with RBL-2H3 cells with Fluo-3. The brighter
part of Fluo-3 fluorescence intensity was identical to the nu-cleus, suggesting that the FceRI-mediated Ca2� signals weretransferred not only to the cytoplasm but also to the nu-cleus.26) Based on ratio-imaging using Indo-1 (a ratiometricCa2�-specific fluorescent probe), it was estimated that theCa2� concentration was 200—300 nM higher in the nucleusthan in the cytoplasm. The receptor-mediated nuclearCa2�signals were also found in B lymphocytes and T lym-phocytes.27—29) These findings suggested the possibility thatCa2� may act as a nuclear messenger in mast cells and lym-phocytes. Further, to study the mechanism of the intranuclearCa2� increase, thapsigargin which is an inhibitor of Ca2�-ATPase on the ER was added to mast cells and lymphocytes.We found that thapsigargin elcited an increase in cytosolicand nuclear Ca2� concentration by depleting Ca2� from theER, and that the subsequent Ca2� influx via SOCs by the de-pletion of Ca2� in the ER also elicited an increase.30—32)
These results suggest that increased intranuclear Ca2� wasmainly derived from the cytoplasm.
DEGRANULATION
The increase in intracellular Ca2� concentration ([Ca2�]i)and the activation of PKC, induced by IP3 and diacylglycerol,respectively, lead to degranulation, causing the release of his-tamine and other potent inflammatory mediators from mastcells. In assessing mast cell function, a degranulation re-sponse has typically been defined by measuring histamine,arachidonic acid metabolism, tryptase, and b-hexos-aminidase released from abundant mast cells or by observingthe density of granules in mast cells using electron mi-croscopy (EM). However, since it is difficult to study thesteps of the degranulation pathway in mast cells using con-ventional microscopy, new microscopic strategies to visual-ize the degranulation process at the living single-cell levelhave been anticipated to study the molecular mechanisms ofdegranulation and perform functional analysis in mast cells.Here we introduce our microscopic approaches to visualizedegranulation in individual mast cells.
CD63 CD63 is a glycoprotein with a molecular mass of50—60 kDa known as a member of the transmembrane 4 su-perfamily (TM4SF) and is located not only on the plasmamembrane but also on the basophilic granule membranes inresting basophils, mast cells, and platelets.33,34) Activation ofbasophils or mast cells induces fusion of cytoplasmic granulemembrane with the plasma membrane and the successive re-lease of inflammatory mediators such as histamine. Upon ac-tivation of human basophilic granulocytes with anti-IgE, theexpression of CD63 on the cell surface is increased, as de-tected by flow cytometry, together with anti-CD63 mono-clonal antibody.35) Therefore the surface expression of CD63in individual mast cells was examined under a CLSM usingmouse monoclonal anti-CD63 antibody (AD1).33) The sur-face expression of CD63 was found to occur almost simulta-neously with histamine release after antigen stimulation inRBL-2H3 cells.36) The surface expression of CD63 was alsomeasured in three types of P815 mastocytoma cells trans-fected with mutant FceRI.36) In P815 cells with wild-typeFceRI (P815wt), the surface expression of CD63 increasedupon the stimulation with antigen. However, in P815bct andP815gct cells with mutant FceRI in which the C-terminal do-
1552 Vol. 28, No. 9

mains including ITAM of the b- and g-chains had been trun-cated, respectively, surface expression did not increase afterantigen stimulation. This indicates that the surface expres-sion of CD63 reflects degranulation in real-time in livingmast cells. However, the dynamic movements of CD63 onthe intracellular secretory granule membranes remain to beelucidated. To determine the dynamics of secretory granules,we prepared a plasmid of CD63-green fluorescent protein(GFP) and introduced it into RBL-2H3 cells to obtain stabletransfectant cells.37) CD63-GFP chimera proteins were lo-cated on both the granule membranes and the plasma mem-branes. The diameter of the fluorescent granules in the cyto-plasm varied from 0.5 to 1.5 mm. The major population offluorescent granules did not move at all in resting cells, al-though a few granules apparently moved straight along thelongitudinal axis of a pseudopodium. In contrast, with anti-gen stimulation, the plasma membrane ruffled violently andthe granules moved dramatically. One-third of fluorescentgranules moved toward to the plasma membrane, and the av-erage velocity of the granules was calculated to be 0.10�0.02 mm/s. They reached the plasma membrane in a few min-utes and fused with it instantaneously. Thus real-time obser-vation of plasma membrane ruffling and intracellular granulemovements utilizing CD63-GFP chimera proteins provided anew insight into the elementary process of degranulation inmast cells.
Atomic Force Microscopy Compared with light mi-croscopy, atomic force microscopy (AFM) has advantagesfor spatial resolution close to those of EM.38) AFM has beenused for many biological specimens such as DNA, proteins,membranes, and cells. Exocytosis-related events at the cellsurface in mast cells have been visualized with AFM.39,40)
The surfaces of RBL-2H3 cells were observed under tap-ping-mode AFM under air pressure after fixation with glu-taraldehyde. None of the crucial treatments such as metalcoating for preparing EM samples are needed for AFM.AFM images showed that resting RBL-2H3 cells had a roundand thick cell body with long pseudopodia, with a nearlysmooth surface. Antigen stimulation for 15 min induced astrong spreading response, accompanied by a transformationof the cell surface to a lamellar topography. It was also foundthat there were various pits on the surface of cell membranes.The diameters of these pits varied from 0.05 to 1.0 mm.These pits were classified roughly into two groups, larger andsmaller. The diameters of the larger and smaller pits wereconsistent at about 0.4—1.0 mm and 0.05—0.2 mm, respec-tively. The larger pits were assumed to be discharged gran-ules because the time-course of their appearance on the cellsurface coincided well with that of b-hexosaminidase secre-tion and they were not observed in the absence of extracellu-lar Ca2�. On the other hand, the smaller pits might be due tocoated pit-like structures related to FceRI-mediated endocy-tosis because their diameters correlated with those of coatedpits observed under EM and they disappeared following pre-treatment with wortmannin, an inhibitor of PI3K. These re-sults indicate the value of AFM measurements in the detec-tion and analysis of degranulation in mast cells.
TRANSCRIPTIONAL REGULATION
In addition to the release of inflammatory mediators from
cytoplasmic granules, there are significant nuclear compo-nents to the activation of mast cells after the ligation ofFceRI. Signaling molecules that translocated from the cyto-plasm to the nucleus upon the aggregation of FceRI play im-portant roles in the induction of the expression of variousgenes and the synthesis of cytokines such as interleukin (IL)-3, IL-4, IL-5, IL-6, IL-10, granulocyte macrophage-colonystimulating factor (GM-CSF), and TNF-a , as shown in Fig.1. Transcriptional factors such as nuclear factor of activatedT cells (NF-AT) and nuclear factor kB (NF-kB) are bound topromoters in cytokine genes.41—44) Nuclear location and acti-vation of NF-AT are regulated by calcineurin, a Ca2�-depen-dent phosphatase. NF-kB is activated and translocated to thenucleus via the pathway including NF-kB-inducing kinase(NIK) and IkB kinase (IKK) that induce the phosphorylationand degradation of IkB.45) Furthermore, three major subfam-ilies of mitogen-activated protein kinases (MAPKs), whichare extracellular signal-regulated kinases (ERKs), c-JunNH2-terminal kinases (JNKs), and p38, respectively, are acti-vated through sequential phosphorylation of cytosolic proteinkinases to lead signals from the plasma membrane to theircytosolic and nuclear targets, regulating nuclear transcriptionfactors.46,47) The activation of these different MAPKs in-volves their phosphorylation by MAPK kinase (MAPKK)family proteins including MAPK/ERK kinase (MEKs). Theactivation of MAPKKs involves their phosphorylation byMAPKK kinase family proteins including Raf-1. The MAP-KKs phosphorylate specific MAPK family members on botha tyrosine and a threonine, resulting in MAPK activation.Moreover, each pathway is downregulated by a variety ofphosphatases that function specifically in different pathways.
It is important to study the process of nuclear translocationin those signaling proteins intimately involved in transcrip-tional activity to understand how cellular signaling is trans-duced to the nucleus from the cytoplasm. GFP and its deriva-tives are considered to be potent and powerful tools to ob-serve nucleocytoplasmic mobilization and to study the mole-cular mechanisms of nuclear translocation. Here we discussthe intracellular dynamics of ERK2, one of the MAPK fam-ily proteins, after the ligation of FceRI in mast cells.
To monitor nucleocytoplasmic movements of ERK2 in realtime, the chimera protein of ERK2 with yellow fluorescentprotein (YFP), a GFP derivative, was prepared and intro-duced into RBL-2H3 cells. YFP-ERK2 was mainly presentin the cytoplasm in resting RBL-2H3 cells, and in somecases the expression of excess amounts of ERK2 resulted inits nuclear accumulation, as shown in Fig. 2a. Upon stimula-tion with antigen, translocation of YFP-ERK2 into the nu-cleus occurred mostly in the RBL-2H3 cells following the in-crease in [Ca2�]i.
48) The fluorescence intensity of YFP-ERK2increased first, and it decreased again in the nucleus of theRBL-2H3 cells. On the contrary, the fluorescence intensity ofYFP-ERK2 decreased first in the cytoplasm with a similarlag time and then increased again. The average fluorescenceintensities in the nucleus and the cytoplasm did not changeduring 30 min, as shown in Fig. 2b. These results indicatethat the import of ERK2 reached the maximum at 6—7 minand that the export to the cytoplasm was almost completedca. 15 min after the ligation of FceRI. The results of kineticstudy of ERK2 translocation were consistent with the phos-phorylation patterns of transfected and endogenous ERKs, as
September 2005 1553

shown in Fig. 2c. These demonstrated that ERK2 was shut-tling between the cytoplasm and the nucleus dependent on itsphosphorylation state after stimulation with antigen.
To investigate the nucleocytoplasmic movements of ERK2and MEK simultaneously, YFP-ERK2 and MEK-cyan fluo-rescent protein (CFP) were coexpressed in RBL-2H3 cells.YFP-ERK2 and MEK-CFP were present mainly in the cyto-plasm in resting cells. After the addition of antigen, YFP-ERK2 was imported into the nucleus, but MEK-CFP wasnot. While the import of ERK2 reached the maximum at 6—7 min and then the imported ERK2 was exported from thenucleus, no significant change in MEK distribution was de-tected after the ligation of FceRI. MEK has a nuclear exportsignal (NES) sequence in the N-terminal region (residues33—44), while ERK2 has neither a nuclear localization sig-nal (NLS) nor NES sequences.49) The present results suggestthat the NES sequence in MEK might play a role in the rapidnuclear shuttling of ERKs after the ligation of FceRI. WhenRBL-2H3 cells were pretreated with leptomycin B, an in-hibitor of Crm1-dependent nuclear export,50) MEK was par-tially distributed in the nucleus. After the addition of antigen,
the nuclear import of ERK2 was unaffected, but the nuclearexport became slower by pretreatment with leptomycin B.These results suggest that the rapid nuclear shuttling ofERK2 was dynamically regulated by MEK after antigenstimulation in RBL-2H3 cells.48) Nuclear shuttling of ERK2was also likely to be controlled by other signaling molecules.Differences in [Ca2�]i levels were found to affect the phos-phorylation and nuclear import of ERK2. The transient[Ca2�]i increase that was induced either by the FceRI aggre-gation in the absence of extracellular Ca2� or by the coclus-tering of FceRI and FcgRIIB, a low-affinity IgG receptorwith the immunoreceptor tyrosine-based inhibition motif(ITIM), did not trigger the optimal translocation of ERK2into the nucleus and suppressed the subsequent TNF-a pro-duction in RBL-2H3 cells.48,51)
There is other evidence that Ca2� regulates transcriptionalactivities. It was reported that the amplitude and the durationof Ca2� signals controlled differential activation of the pro-inflammatory transcriptional regulators NF-kB, JNK, andNF-AT in B lymphocytes.52) It appears, therefore, that thespecificity of cellular responses encoded by calcium signal-ing patterns results from differences in the Ca2� sensitivitiesand kinetic behavior of these transcriptional regulators. Nu-clear Ca2� as well as cytosolic Ca2� controls gene expres-sion. It was found that an increase in nuclear Ca2� concentra-tion controlled Ca2�-activated gene expression mediated bythe cyclic AMP-response element (CRE) in the c-fos pro-moter and that the CRE-binding protein (CREB) can func-tion as a nuclear Ca2�-responsive transcription factor.53)
CREB activity is regulated by its phosphorylation on serine133, which can be catalyzed by a number of protein kinasesincluding cAMP-dependent protein kinase, calcium/calmod-ulin-dependent kinase (CaMK) IV, ribosomal S6 kinase(RSK) 2, and MAP kinase-activated protein (MAPKAP) ki-nase 2. These indicate that Ca2� is intimately involved ingene expression via the activation of transcriptional regula-tors and contributes to the efficacy and specificity of cellularsignaling to mediate a diverse array of cell functions.
INTERACTION WITH NEURONS
There has been an exponential increase in data illustratingthat the immune and nervous systems are not disparate enti-ties.54,55) The mast cell–nerve relationship has served as aprototypic association and has provided substantial evidencefor bidirectional communication between nerves and immunecells.56) Many studies have shown that messenger molecules(e.g., neuropeptides) from nerves can elicit mast cell degran-ulation and that mast cell-derived mediators (e.g., histamine,serotonin, and platelet-activating factor) modulate neuro-transmission.57,58) Early studies elegantly described the non-random spatial association of mast cells and nerves in a vari-ety of tissues in which membrane–membrane contacts couldbe observed.10) However, while the tissue studies showed afunctional nerve–mast cell interplay, they did not dismiss thepossibility that an intermediate cell transduces or modulatesthe nerve–mast cell communication. We utilized an in vitrococulture model comprising RBL-2H3 cells and neurite-sprouting murine superior cervical ganglia (SCG) to examinedirect neurite–mast cell communication. A selective associa-tion was seen between SCG and RBL-2H3 cells in cocul-
1554 Vol. 28, No. 9
Fig. 2. Nuclear Shuttling of ERK2 in RBL-2H3 Cells after Antigen Stim-ulation
(a) CLSM images of YFP-ERK2 in RBL-2H3 cells after the addition of DNP-conju-gated BSA (DNP-BSA; 500 ng/ml). The times indicated are after addition of DNP-BSA. (b) Time-course of fluorescence intensity changes in YFP-ERK2 in the nucleus(�), in the cytoplasm (�), and the average in the nucleus and the cytoplasm (�). (c)Western blotting analysis comparing phosphorylation of YFP-ERK2 and endogenousERK2.

tures.59) EM images showed that the contacts between theirmembranes were less than 20 nm and dense-core vesicleswere found in the neurites proximate to RBL-2H3 cells.60)
Changes in the electrical properties of RBL-2H3 cell mem-branes upon contact were also noted, and RBL-2H3 cellsstopped dividing upon contact and increased their mediatorcontent and number of granules.61,62)
Interactions between Nerves and RBL Cells We haveshown direct communication between RBL-2H3 cells andSCG in cocultures using Ca2� imaging to indicate evidencefor activation and signaling. The addition of bradykinin orscorpion venom to SCG cultures only resulted in a dose-de-pendent neurite activation but did not evoke an increase in[Ca2�]i when added directly to RBL-2H3 cells in the absenceof SCG neurites. In coculture studies, bradykinin or scorpionvenom-induced neurite activation was invariably followed byactivation in RBL-2H3 cells in contact with the activatedneurites, as shown in Fig. 3.63) The number of RBL-2H3 cellsresponding increased in accordance with increasing concen-trations of bradykinin or scorpion venom. These results showthat activation of a nerve fiber directly elicits an activationevent in an RBL-2H3 cell attached to the activated fiber. Onthe other hand, when RBL-2H3 cells were stimulated withanti-FceRIa antibody in cocultures to determine a signaltransduction from RBL-2H3 cells to SCG neurites, neuriteactivation occurred in response to RBL activation.63,64) Neu-rites were affected at considerable distance (�160 mm) fromactivated RBL-2H3 cells. These data clearly show that SCGneurite or RBL-2H3 cell activation can result in activation of
the reciprocal cell type in the absence of any modulating ortransducing effects of an intermediary cell.
Substance P can induce the release of histamine from cer-tain types of mast cells such as peritoneal and mucosal mastcells, human mast cells, and human pulmonary mast cells,further supporting the functional interaction between mastcells and substance P-containing nerves in vivo.57,65—67) Sincesubstance P in isolated SCG neurons maintained in cultureshowed a 25-fold increase within 48 h in vitro,68) we pro-ceeded to assess substance P as a putative mediator responsi-ble for the direct activation from SCG neurites to RBL-2H3cells. Measurements in which cocultures were stimulatedwith bradykinin in the presence of neutralizing anti-sub-stance P antibodies revealed that the inclusion of the antibod-ies dose-dependently prevented the activation in RBL-2H3cells. It has been reported that substance P induces mast cellactivation via both a receptor-independent and a receptor-de-pendent pathway. Higher concentrations of substance P causedirect activation of G protein. The positively charged N-ter-minal amino acid residues of substance P are responsible forthis receptor-independent pathway.69) On contrary, lower con-centrations of substance P cause mast cell activation by inter-action with the neurokinin (NK)-1 receptor, which is atachykinin receptor preferentially bound to substance P.65)
Additionally, RBL-2H3 cells were shown to express NK-1receptors on their surface.70) Next, the ability of tachykininreceptor antagonists to abrogate the activation in RBL-2H3cells after neurite activation was examined. Inclusion of theNK-1 receptor antagonist (CP99,994-01) in cocultures did
September 2005 1555
Fig. 3. Time-Courses of Neurite-Induced RBL-2H3 Cell Activation
(a) CLSM images of Fluo-3-loaded SCG neurites and an associated RBL-2H3 cell in coculture after bradykinin stimulation (10 nM). The times indicated are after addition ofbradykinin. (b) Representative Fluo-3 fluorescence changes in neurites and associated RBL-2H3 cells. Arrow shows the time when bradykinin was added.

not affect the neurite activation evoked bradykinin, but sig-nificantly and dose-dependently inhibited the subsequent in-crease in RBL activation. In contrast, the NK-2 receptor an-tagonist (SR48,968) did not affect either neurite activation orthe subsequent Ca2� mobilization in RBL-2H3 cells in cocu-tures. These results indicate that the neuronal-derived sub-stance P activated the RBL-2H3 cells in contact with the ac-tivated neurites via NK-1 receptors.63)
Degranulation in RBL Cells by Nerve ActivationNext, to assess mast cell function in nerve-mast cell commu-nication, we studied the degranulation process in RBL-2H3cells by measuring the cell surface structure in AFM andCD63-GFP granule movements in CLSM.71,72) AFM imagesof neurite-RBL coculture showed that a thin neurite with agrowth cone extended over a fiber-like pseudopodium ofRBL-2H3 cells and that contact between the growth cone andthe RBL-2H3 cells occurred over about 7 mm, as shown inFig. 4a. Whereas RBL-2H3 cells attached to neurites had aspherical and nearly smooth cell body in cocultures, they
spread and flattened 20 min after the addition of bradykinin,as shown in Fig. 4b.71) Several discharged granules with di-ameters of 0.5—1.0 mm were also found on the surface ofRBL-2H3 cells. Such morphological changes in RBL-2H3cells induced by nerve activation were similar to those uponantigen stimulation. However, the number of dischargedgranules after nerve stimulation was much fewer than thatafter antigen stimulation. This may be due to the local activa-tion of RBL-2H3 cells through the contact site with neurites.These morphological changes were not found in RBL-2H3cells not in contact with neurites. In addition, the NK-1 re-ceptor antagonist mostly inhibited the activated neurite-in-duced degranulation in RBL-2H3 cells. AFM images showedthat activation of the SCG neurites was able to elicit degranu-lation in RBL-2H3 cells attached to neurites.
To investigate further the degranulation process in RBL-2H3 cells induced by neurite activation, we used a coculturesystem composed of SCG neurites and RBL-2H3 cells stablyexpressing CD63-GFP, as shown in Figs. 4c and d. The
1556 Vol. 28, No. 9
Fig. 4. Degranulation in RBL-2H3 Cells Attached to Neurites
(a) AFM image of SCG neurites attached to a pseudopodium and the cell body of an RBL-2H3 cell. A thin neurite with a growth cone extends over a fiber-like pseudopodium ofan RBL-2H3 cell. (b) Typical example of AFM image of neurite and RBL-2H3 cell unit 20 min after bradykinin stimulation (100 nM). Several discharged granules shown by whitearrowheads are observed in the area close to neurites. (c) and (d) Phase-contrast image and CLSM image of neurites and RBL-2H3 cells expressing CD63-GFP, respectively. (e)and (f) Enlarged sequential images of boxes e and f in Fig. 4(d), respectively. The times indicated are after addition of bradykinin (10 nM).

plasma membrane ruffling was examined in RBL-2H3 cells in contact with bradykinin-stimulated neurites. Afterbradykinin stimulation of neurites, there was significant andobvious RBL cell membrane ruffling, but this was restrictedto the point of contact with the neurite. The addition of scor-pion venom to the neurite-RBL cell cocultures also induced alocal ruffling response in the pseudopodium of RBL cells incontact with neurites but not in the pseudopodia that weredistinct from neurites, as shown in Figs. 4e and f. The per-centage of RBL cells responding to neurite activation bymembrane ruffling was dose-dependently inhibited in thepresence of the NK-1 receptor antagonist, but not in the pres-ence of the NK-2 receptor antagonist. RBL-2H3 cells ex-pressing CD63-GFP allowed tracking of granule movementsfollowing activation. In the absence of neurite stimulation,the RBL cell granules were largely inactive. In contrast, afterneurite activation, we found increased granule movements inthe contacting RBL cells, with the most dramatic granulemovements being predominantly located adjacent to the neu-rite contact point. Those granules located close to the neurite(within 10 mm of the contact point) became highly motileand moved toward the plasma membrane at an average veloc-ity of 0.13�0.04 mm/s. Granules distant from the site of con-tact were considerably less motile (0.05�0.01 mm/s), andtheir movement patterns were, on average, largely reminis-cent of the granules in RBL-2H3 cells in unstimulated cocul-tures.72)
These results support an important role for substance P innerve-to-mast cell communication and define the movementsof locally sited granules toward the plasma membrane as aconsequence of excitation and a possible precursor to de-granulation. The demonstration that neurite activation notonly causes a Ca2� response in the RBL-2H3 cells associatedwith the neurites but also is followed by dramatic plasmamembrane ruffling and morphological change at cell–cellcontact sites highlights a very precise, confined, and specificmeans of cross-talk between nerves and mast cells.
Mast Cell Activation According to Level of NK-1 Re-ceptor Expression While mucosal and skin mast cells canbe activated by substance P, evidence for this communicationoccurring directly through NK-1 receptors has so far beenlimited. It was demonstrated that RBL-2H3 cells expressNK-1 receptors on their surface,70) but it was not widely ac-cepted that mast cells express NK receptors under normalphysiological conditions. Since RBL-2H3 cells are a cell lineand bone marrow-derived mast cells (BMMCs) are thoughtto be mucosal mast cell counterparts, we conducted experi-ments to establish the determinants of response in cocultureand correlate this with the degree of expression of NK-1 re-ceptors on BMMCs. It has recently been found that sub-stance P causes dose-dependent degranulation in BMMCscultured with IL-4 and SCF, and that BMMCs express func-tional NK-1 receptors under the influence of IL-4 or SCF butexpress the greatest numbers when IL-4 and SCF are bothpresent.73—75) Therefore we cocultured BMMCs, which hadbeen cultured under various conditions to obtain differentpercentages of NK-1 receptor expression, with SCG to inves-tigate the population of mature BMMCs positive for the NK-1 receptor (c-kit�/NK-1�) followed by the probability ofCa2� response after nerve activation.76) When scorpionvenom was added to the coculture, the [Ca2�]i increase in
BMMCs invariably occurred after the [Ca2�]i increase inneurites. BMMCs did not respond to scorpion venom in theabsence of neurites. At a high level of NK-1 receptor expres-sion, about 70% of BMMCs attached to neurites respondedafter scorpion venom stimulation, as shown in Fig. 5. TheCa2� response in BMMCs was inhibited by pretreatmentwith the NK-1 receptor antagonist. The response rates de-clined in direct correlation with the decrease in NK-1 recep-tor expression (see Fig. 5). When no NK-1 receptors wereexpressed, even though the majority of BMMCs were c-kitpositive, the response was negligible. These results indicatethat BMMCs respond in association with activated nervefibers according to the level of NK-1 receptor expression,suggesting that the presence of NK-1 receptors on BMMCsconfers increased sensitivity to substance P derived fromneurites.76)
Adhesion Proteins Involved in Interactions betweenNerves and Mast Cells We attempted to identify the adhe-sion proteins involved in the contact between neurites andmast cells. Once formed, the contact between SCG neuritesand RBL-2H3 cells was maintained for up to 120 h and onaverage there was �20-nm distance between apposed mem-branes. Dense-cored neurosecretory vesicles accumulated inthe neurite endings apposed to the RBL-2H3 cell membrane,while no specialized structures were seen either in neurites orRBL-2H3 cells.60) We focused on cadherins, which are afamily of Ca2�-dependent and homophilic cell adhesion pro-teins playing a role in the formation of synaptic plasticity.Therefore the involvement of cadherins in contact betweennerves and mast cells has been examined using in vitro co-culture of SCG neurites with BMMCs instead of RBL-2H3cells, since RBL-2H3 cells are much more adhesive thanBMMCs and adhere non-specifically to culture dishes. West-ern blotting and immunostaining analysis showed that bothSCG and BMMCs expressed N- and E-cadherins, and that N-and E-cadherins are present mainly in the cytoplasm ofBMMCs in the absence of SCG neurites. With the associa-tion of neurites, however, N-cadherin was localized on theplasma membrane of BMMCs, but E-cadherin was not. b-Catenin, which is a protein associated with cadherins in thecytoplasm, was also accumulated at the periphery of theplasma membrane in BMMCs attached to neurites, while itwas distributed in the cytoplasm without neurites. The pre-
September 2005 1557
Fig. 5. Correlation of NK-1 Receptor Expression and Responding Rate toActivated Neurites in BMMCs
BMMCs responded to activated neurites after scorpion venom (1 mg/ml) stimulationaccording to level of NK-1 receptor expression.

sent results suggest that N-cadherin, but not E-cadherin, isinvolved in the contact formation between nerves and mastcells in collaboration with b-catenin.77)
In addition, Biederer et al.78) and Ito et al.79) have recentlyisolated the same molecule, designated the synaptic cell ad-hesion molecule (SynCAM) or spermatogenic immunoglob-ulin superfamily (SgIGSF) in different processes. Biederer etal. showed that SgIGSF/SynCAM is localized preferentiallyon both sides of most synapses in the brain and functions asa homophilic adhesion molecule that spans the synaptic cleftin the nervous system.78) Ito et al. found that SgIGSF/Syn-CAM mediates the adhesion of BMMCs to fibroblasts andthat the expression of SgIGSF/SynCAM in BMMCs is regu-lated by the microphthalmia transcriptional factor (MITF).79)
Since SgIGSF/SynCAM is strongly considered to be one ofthe candidates involved in the attachment and communica-tion between mast cells and nerves, we examined this possi-bility by coculturing SCG neurons and mast cells with orwithout SgIGSF/SynCAM. SgIGSF/SynCAM was localizedintensively at the contact site between mast cells and neu-rites. Mast cells lacking SgIGSF/SynCAM attached poorly toneurites, and ectopic expression of this adhesion moleculesignificantly improved their attachment. The attachment lev-els of mast cells expressing SgIGSF/SynCAM were dose-de-pendently reduced in the presence of an anti-SgIGSF/Syn-CAM blocking antibody. These results suggest thatSgIGSF/SynCAM is the major adhesion molecule that medi-ates in vitro attachment between mast cells and neurites.80)
We also studied the involvement of SgIGSF/SynCAM inthe communication between mast cells and nerves. Our re-sults showed that there is a significant difference in the pro-portion responding to neurite activation between mast cellswith and without SgIGSF/SynCAM. Only one-quarter ofmast cells without SgIGSF/SynCAM responded to neuriteactivation, whereas more than one-half of mast cells with theadhesion molecule responded. The responding rate of mastcells with SgIGSF/SynCAM to neurite activation dose-de-pendently decreased in the presence of an anti-SgIGSF/Syn-CAM blocking antibody. In addition, the NK-1 receptor an-tagonist (CP99,994-01) blocked the responding rate to neu-rite activation in mast cells without SgIGSF/SynCAM atmuch lower concentrations than in mast cells with the adhe-sion molecule. Thus SgIGSF/SynCAM is likely not only tofunction as simple glue in nerve-mast cell interaction, butalso to promote the development of a microenvironmentwhere mast cells have an enhanced susceptibility to nerve ac-tivation, as shown in Fig. 6.
CONCLUSION
In this review, we mainly presented our work on the imag-ing analysis of cellular signaling in mast cells. The micro-scopic technology to study the dynamic molecular organiza-tion and signaling events in individual living cells has re-cently advanced and played an essential role in many fieldsof life science such as cell biology, immunology, physiology,and neurology. Our approach utilizing the advantages of mi-croscopic analysis provided new aspects of information onthe mobilization of intracellular Ca2�, secretory granules,and signaling proteins in FceRI-mediated mast cell activa-tion. On the basis of techniques to study signaling events in
single cells, the microscopic methods could be further devel-oped for investigating unequivocally the bidirectional com-munication between mast cells and neurons which may be in-volved in physiologic or pathologic processes in inflamma-tion, disease, and reactions to stress.
Mast cells are considered to function as ancillary aidsagainst infection, promoters of tissue repair, regulators ofblood vessel tone, permeability, and angiogenesis, importantmediators of stress and allergy, regulators of the neuroen-docrine system, purveyors of sensory information to the cen-tral nervous system, and effector targets for efferent nervouspathways.81) Therefore it is important to study molecularmechanisms through which mast cells function. The role ofmicroscopic techniques is becoming more important sincethey yield information on specific molecular components ofcells with high spatiotemporal resolution. We hope that ourstrategy and results can contribute to an understanding of theimmune response and complicated regulation in our body.
Acknowledgments We are grateful to Prof. J. Bienen-stock, Prof. Y. Kitamura, and Drs. R. Teshima, A. Ito, N. Hi-rashima, and R. Suzuki for their advice, creative work, anddiscussions. This study was supported in part by Grants-in-Aid for Scientific Research from the Japan Society for thePromotion of Science and for Young Scientists from the Min-istry of Education, Culture, Sports, Science and Technologyof Japan.
REFERENCES AND NOTES
1) Present address: School of Pharmacy, Aichi-Gakuin University, 1–100Kusumoto-cho, Chikusa-ku, Nagoya 464–8650, Japan.
2) Blank U., Ra C., Miller L., White K., Metzger H., Kinet J. P., Nature(London), 337, 187—189 (1989).
3) Reth M., Nature (London), 338, 383—384 (1989).4) Yamashita T., Mao S. Y., Metzger H., Proc. Natl. Acad. Sci. U.S.A., 91,
11251—11255 (1994).5) Jouvin M. H., Adamczewski R., Numerof R., Letourneur O., Valle A.,
Kinet J. P., J. Biol. Chem., 269, 5918—5925 (1994).6) Parravicini V., Gadina M., Kovarova M., Odom S., Gonzalez-Espinosa
C., Furumoto Y., Saitoh S., Samelson L. E., O’Shea J. J., Rivera J.,Nat. Immunol., 3, 741—748 (2002).
1558 Vol. 28, No. 9
Fig. 6. Schematic Representation of Mast Cell Activation by ActivatedNeurites

7) Saitoh S., Arudchandran R., Manetz T. S., Zhang W., Sommers C. L.,Love P. E., Rivera J., Samelson L. E., Immunity, 12, 525—535 (2000).
8) Blank U., Rivera J., Trends Immunol., 25, 266—273 (2004).9) Galli S. J., Costa J. J., Allergy, 50, 851—862 (1995).
10) Stead R. H., Tomioka M., Quinonez G., Simon G. T., Felten S. Y., Bi-enenstock J., Proc. Natl. Acad. Sci. U.S.A., 84, 2975—2979 (1987).
11) Arizono N., Matsuda S., Hattori T., Kojima K., Maeda T., Galli S. J.,Lab. Invest., 62, 626—634 (1990).
12) Rozniecki J. J., Dimitriadou V., Lambracht-Hall M., Pang X., Theo-harides T. C., Brain Res., 849, 1—15 (1999).
13) Dimitriadou V., Buzzi M. G., Moskowitz M. A., Theoharides T. C.,Neuroscience, 44, 97—112 (1991).
14) Frieling T., Cooke H. J., Wood J. D., Am. J. Physiol., 261, G1017—G1023 (1991).
15) Frieling T., Cooke H. J., Wood J. D., Am. J. Physiol., 264, G74—G80(1994).
16) Corvera C. U., Dery O., McConalogue K., Gamp P., Thoma B., Al-AniB., Caughey G. H., Hollenberg M. D., Bunnett N. W., J. Physiol., 517,741—756 (1999).
17) Marone G., Galli S. J., Kitamura Y., Trends Immunol., 23, 425—427(2002).
18) van Houwelingen A. H., Kool M., de Jager S. C., Redegeld F. A., vanHeuven-Nolsen D., Kraneveld A. D., Nijkamp F. P., J. Immunol., 168,5297—5302 (2002).
19) Leon A., Buriani A., Dal Toso R., Fabris M., Romanello S., Aloe L.,Levi-Montalcini R., Proc. Natl. Acad. Sci. U.S.A., 91, 3739—3743(1995).
20) Suzuki M., Furuno T., Teshima R., Sawada J., Nakanishi M., Biol.Pharm. Bull., 21, 1267—1270 (1998).
21) Theoharides T. C., Int. J. Tissue React., 18, 1—21 (1996).22) Theoharides T. C., Cochrane D. E., J. Neuroimmunol., 146, 1—12
(2004).23) Barbara G., Stanghellini V., De Giorgio R., Cremon C., Cottrell G. S.,
Santini D., Pasquinelli G., Morselli-Labate A. M., Grady E. F., Bun-nett N. W., Collins S. M., Corinaldesi R., Gastroenterology, 126,693—702 (2004).
24) Berridge M. J., Bootman M. D., Lipp P., Nature (London), 395, 645—648 (1998).
25) Putney J. W., Bird G. S., Cell, 75, 199—201 (1993).26) Nakato K., Furuno T., Inagaki K., Teshima R., Terao T., Nakanishi M.,
Eur. J. Biochem., 209, 745—749 (1992).27) Yamada H., Mizuguchi J., Nakanishi M., FEBS Lett., 284, 249—251
(1991).28) Furuno T., Hamano T., Nakanishi M., Biophys. J., 65, 665—669
(1993).29) Nagao Y., Yamada H., Nakanishi M., Immunol. Lett., 34, 151—153
(1992).30) Furuno T., Hamano T., Nakanishi M., Bioimages, 1, 9—12 (1993).31) Horikoshi Y., Furuno T., Teshima R., Sawada J., Nakanishi M.,
Biochem. J., 304, 57—60 (1994).32) Okamoto Y., Furuno T., Hamano T., Nakanishi M., Biochem. J., 305,
1011—1015 (1995).33) Kitani S., Berenstein E., Mergenhagen S., Tempst P., Siraganian R. P.,
J. Biol. Chem., 266, 1903—1909 (1991).34) Metzelaar M. J., Wijngaard P. L., Peters P. J., Sixma J. J., Nieuwenhuis
H. K., Clevers H. C., J. Biol. Chem., 266, 3239—3245 (1991).35) Knol E. F., Mul F. P., Jansen H., Calafat J., Roos D., J. Allergy Clin.
Immunol., 88, 328—338 (1991).36) Furuno T., Teshima R., Kitani S., Sawada J., Nakanishi M., Biochem.
Biophys. Res. Commun., 219, 740—744 (1996).37) Amano T., Furuno T., Hirashima N., Ohyama N., Nakanishi M., J.
Biochem. (Tokyo), 129, 739—744 (2001).38) Hansma P. K., Eling V. B., Marti O., Bracker C. E., Science, 242,
209—216 (1988).39) Nakamura R., Furuno T., Nakanishi M., Bioimages, 6, 69—75 (1998).40) Teshima R., Nakamura R., Furuno T., Nakanishi M., Bioimeges, 7,
97—103 (1999).41) Shaw J. P., Utz P. J., Durand B. B., Toole J. J., Emmel E. A., Crabtree
G. R., Science, 241, 202—205 (1988).42) Turner H., Cantrell D. A., J. Exp. Med., 185, 43—53 (1997).43) Turner H., Gomez M., McKenzie E., Kirchem A., Lennard A., Cantrell
D. A., J. Exp. Med., 188, 527—537 (1998).44) Kitaura J., Asai K., Maeda-Yamamoto M., Kawakami Y., Kikkawa U.,
Kawakami T., J. Exp. Med., 192, 729—739 (2000).
45) Ghosh S., Karin M., Cell, 109, S81—S96 (2002).46) Pearson G., Robinson F., Beers Gibson T., Xu B. E., Karandikar M.,
Berman K., Cobb M. H., Endocr. Rev., 22, 153—183 (2001).47) Hirasawa N., Sato Y., Fujita Y., Mue S., Ohuchi K., J. Immunol., 161,
4939—4943 (1988).48) Furuno T., Hirashima N., Onizawa S., Sagiya N., Nakanishi M., J. Im-
munol., 166, 4416—4421 (2001).49) Fukuda M., Gotoh I., Gotoh Y., Nishida E., J. Biol. Chem., 271,
20024—20028 (1996).50) Fukuda M., Asano S., Nakamura T., Adachi M., Yoshida M., Yanagida
M., Nishida E., Nature (London), 390, 308—311 (1997).51) Ohyama N., Furuno T., Hirashima N., Nakanishi M., Immunol. Lett.,
90, 173—176 (2003).52) Dolmetsch R. E., Lewis R. S., Goodnow C. C., Healy J. I., Nature
(London), 386, 855—858 (1997).53) Hardingham G. E., Chawla S., Johnson C. M., Bading H., Nature
(London), 385, 260—265 (1997).54) Ottaway C. A., Gastroenterol. Clin. North Am., 20, 511—529 (1991).55) Cohen N., Moynihan J. A., Ader R., Int. Arch. Allergy Immunol., 105,
101—106 (1994).56) McKay D. M., Bienenstock J., Immunol. Today, 15, 533—538 (1994).57) Shanahan F., Denburg J. A., Fox J., Bienenstock J., Befus D., J. Im-
munol., 135, 1331—1337 (1985).58) Weinreich D., Undem B. J., Leal-Cardoso J. H., Ann. N.Y. Acad. Sci.,
664, 293—308 (1992).59) Blennerhassett M. G., Bienenstock J., Neurosci. Lett., 120, 50—54
(1991).60) Blennerhassett M. G., Tomioka M., Bienenstock J., Cell Tissue Res.,
265, 121—128 (1991).61) Blennerhassett M. G., Janiszewski J., Bienenstock J., Am. J. Respir.
Cell Mol. Biol., 6, 54—59 (1992).62) Blennerhassett M. G., Bienenstock J., J. Neurobiol., 35, 173—182
(1998).63) Suzuki R., Furuno T., McKay D. M., Wolvers D., Teshima R., Naka-
nishi M., Bienenstock J., J. Immunol., 163, 2410—2415 (1999).64) Suzuki R., Furuno T., Teshima R., Nakanishi M., Biol. Pharm. Bull.,
24, 291—294 (2001).65) Ogawa K., Nabe T., Yamamura H., Kohno S., Eur. J. Pharmacol., 374,
285—291 (1999).66) Gibbs B. F., Wierecky J., Welker P., Henz B. M., Wolf H. H., Grabbe J.,
Exp. Dermatol., 10, 312—320 (2001).67) Heaney L. G., Cross L. J., Stanford C. F., Ennis M., Clin. Exp. Allergy,
25, 179—186 (1995).68) Kessler J. A., Adler J. E., Bell W. O., Black I. B., Neuroscience, 9,
309—317 (1983).69) Mousli M., Bueb J. L., Bronner C., Rouot B., Landry Y., Trends Phar-
macol. Sci., 11, 358—362 (1992).70) Cooke H. J., Fox P., Alferes L., Fox C. C., Wolfe S. A., Jr., Can. J.
Physiol., Pharmacol., 76, 188—193 (1998).71) Ohshiro H., Suzuki R., Furuno T., Nakanishi M., Immunol. Lett., 74,
211—214 (2000).72) Mori N., Suzuki R., Furuno T., McKay D. M., Wada M., Teshima R.,
Bienenstock J., Nakanishi M., Am. J. Physiol. Cell Physiol., 283,C1738—C1744 (2002).
73) Karimi K., Redegeld F. A. M., Heijdra B., Nijkamp F. P., Exp. Hema-tol., 27, 654—662 (1999).
74) Karimi K., Redegeld F. A. M., Heijdra B., Nijkamp F. P., Exp. Hema-tol., 28, 626—634 (2000).
75) van der Kleij H. P. M., Ma D., Redegeld F. A., Kraneveld A. D.,Nijkamp F. P., Bienenstock J., J. Immunol., 171, 2074—2079 (2003).
76) Furuno T., Ma D., van der Kleij H. P. M., Nakanishi M., BienenstockJ., Neurosci. Lett., 372, 185—189 (2004).
77) Suzuki A., Suzuki R., Furuno T., Teshima R., Nakanishi M., Biol.Pharm. Bull., 27, 1891—1894 (2004).
78) Biederer T., Sara Y., Mozhayeva M., Atasoy D., Liu X., Kavalali E. T.,Sudhof T. C., Science, 297, 1525—1531 (2002).
79) Ito A., Jippo T., Wakayama E., Morii E., Koma Y., Onda H., NojimaH., Iseki S., Kitamura Y., Blood, 101, 2601—2608 (2003).
80) Furuno T., Ito A., Koma Y., Watabe K., Yokozaki H., Bienenstock J.,Nakanishi M., Kitamura Y., J. Immunol., 174, 6934—6942 (2005).
81) Maurer M., Theoharides T., Granstein R. D., Bishoff S. C., Bienen-stock J., Henz B., Kovanen P., Piliponsky A. M., Kambe N., VliagoftisH., Levi-Schaffer F., Metz M., Miyachi Y., Befus D., Forsythe P., Kita-mura Y., Galli S., Exp. Dermatol., 12, 886—910 (2003).
September 2005 1559