Embed Size (px)
Citation preview

LIST OF PUBLICATIONS
A. RESEARCH PAPERS
1. Sharma, I., Pati, P.K. and Bhardwaj, R. (2010). Regulation of Growth and
Antioxidant Enzyme Activities by 28-Homobrassinolide in Seedlings of Raphanus
sativus L. under Cadmium stress. Indian Journal of Biochemistry and Biophysics. 47:
172-177.
2. Randhawa, N., Sharma, I. and Bhardwaj, R. (2010). Effect of 24-Epibrassinolide and
28-Homobrassinolide on some Biochemical Parameters in Raphanus sativus L.
Plants under Chromium Stress. Seed Science and Biotechnology. 4: 28-32.
3. Sharma, I., Pati, P.K. and Bhardwaj, R. (2011). Effect of 24-epibrassinolide on
oxidative stress markers induced by Nickel metal in Raphanus sativus L. Acta
Physiologae Plantarum. 33: 1723-1735.
4. Sharma, I., Pati, P.K. and Bhardwaj, R. (2011). Effect of 28- homobrassinolide on
antioxidant defence system in Raphanus sativus L. under metal stress. Ecotoxicology.
13: 862-874.
5. Sharma, N., Hundal, G. S., Sharma, I. and Bhardwaj, R. (2011). Effect of 24-
Epibrassinolide on Protein content and activities of Glutathione-S-transferase and
Polyphenol oxidase in Raphanus sativus L. plants under Cadmium and Mercury metal
stress. Terrestrial and Aquatic Environmental Toxicology. (In Press).
6. Sharma, I., Pati, P.K. and Bhardwaj, R. (2011). Induction of Mercury induced
Oxidative Stress Tolerance in Raphanus sativus L. by Brassinosteroids. Journal of
Hazardous Material. (Communicated)
B. REVIEW ARTICLES
1. Bhardwaj, R., Sharma, I., Arora N., Uppal, P., Kanwar, M.K., Sharma, N and
Hundal, G.S. (2009). Regulation of oxidative stress by BRs in plants. In: Oxidative
stress: Role of antioxidants in plants (editor: Parvez Ahmad), Studium Press Pvt.
Ltd., pp. 215-231, New Delhi.
2. Bhardwaj, R., Sharma, I., Arora N., Uppal, P., Kanwar, M.K. (2010). Prospects of
brassinosteroids in medicinal applications. In: Brassinosteroids: A Class of Plant
Hormone (editor: Shamsul Hayat), Springer, pp. 452-473, The Netherlands.

3. Bhardwaj, R., Sharma, I., Kanwar, M. and Handa, N. (2011). Brassinosteroids and
human welfare. In: Practical applications of Brassinosteroids (Eds. Adaucto B.
Pereira-Netto). Bentham Sci. Publishers, UAE. (In press)
4. Bhardwaj, R., Sharma, I., Choudhary, S.P., Uppal P., Kanwar M., Handa N.,
Singh, G., Gautam, V. and Kapoor D. (2011). Cross-talk of Brassinosteroids with
other PGR's in the plants. In: Festschrift book volume to be published in the honor of
Prof. C. P. Malik. (Eds. Pravin C. Trivedi), Jaipur. (In press)
C. ABSTRACTS
1. Sharma, I., Bhardwaj, R., Pati, P.K., Choudhary, S.P. and Sharma, N. (2008).
Brassinosteroids regulated Polyamine content and activities of antioxidative enzymes
in Raphanus sativus L. seedlings under Cr metal stress. In: Golden Jubilee
Conference on “Challenges and Emerging Strategies for Improving Plant
Productivity” organized by Indian Society for Plant Physiology, Indian Agricultural
Research Institute (IARI), New Delhi. pp. 12-14.
2. Sharma, I., Arora, P., Kanwar, M.K., Pati, P.K. and Bhardwaj, R. (2009).
Regulation of Growth and Antioxidant Enzyme activities by 28 – Homobrassinolide
in seedlings of Raphanus sativus L. under cadmium stress. In: Proceedings of
International Conferences on AFRR 2009 & SFRR-India Meeting, Lucknow. pp. 72.
(Paper presentation, received Travel Grant Award)
3. Sharma, I., Pati, P.K. and Bhardwaj, R. (2010). Mitigation of Nickel metal stress in
Raphanus sativus L. seedlings by 24-Epibrassinolide. In: Proceedings of 13th Punjab
Science Congress, Panjab University, Chandigarh.
4. Sharma, I., Pati, P.K. and Bhardwaj, R. (2011). 28-Homobrassinolide mediated
amelioration of stress induced by Mercury in Raphanus sativus L. In: Proceedings of
National Seminar on Environmental Analysis and Remediation, Amritsar. pp. 102-
103.

Effect of 28-homobrassinolide on antioxidant defence system
in Raphanus sativus L. under chromium toxicity
Indu Sharma • Pratap Kumar Pati •
Renu Bhardwaj
Accepted: 14 March 2011 / Published online: 30 March 2011
Ó Springer Science+Business Media, LLC 2011
Abstract Heavy metals have emerged as major envi-
ronmental contaminants due to rapid industrialization and
urbanization. The genotoxic, mutagenic and carcinogenic
effects of heavy metal like chromium (Cr) on man, animals
and plants have been documented. In plants, accumulation
of heavy metals beyond critical levels generates oxidative
stress. This stress is generally overcome by antioxidant
defence system and stress shielding phytohormones. Thus,
the present study has been focused to analyze the effect of
one of imperative group of plant hormones, i.e., brassi-
nosteroids (BRs) which have been reported for its protec-
tive properties for wide array of environmental stresses.
Raphanus sativus L. (Pusa Chetaki) seeds pre-treated with
different concentrations of 28-homobrassinolide (28-HBL)
were raised under various concentrations of Cr(VI). It was
observed that 28-HBL treatment considerably reduced the
impact of Cr-stress on seedlings which was evinced upon
analysis of morphological and biochemical parameters of
7-days old radish seedlings. The toxic effects of Cr in terms
of reduced growth, lowered contents of chlorophyll (Chl),
protein, proline; increased malondialdehyde (MDA) con-
tent and elevated metal uptake were ameliorated by
applications of 28-HBL. Also, the activities of all the
antioxidant enzymes except guaiacol peroxidase (POD),
increased significantly when subjected to Cr stress in
combination with 28-HBL. Overall, seed pre-soaking
treatment of 28-HBL at 10-7 M was most effective in
ameliorating Cr stress. The present work emphasizes the
protective role of 28-HBL on regulation of antioxidant
enzymes and its possible link in amelioration of stress in
plants.
Keywords Brassinosteroids � Antioxidant enzymes �
Radish, metal stress � Lipid peroxidation � Proline content
Introduction
Industrial revolution and anthropogenic activities have
aggravated the metal pollution of biosphere and it has
posed a serious threat to mankind by its incorporation to
the food chain resulting in the degradation of ecosystem
(Chary et al. 2008). Some heavy metals are essential for
most of the redox reactions which are fundamental for
cellular functions. However, their concentrations beyond
tolerable limits lead to production of reactive oxygen
species (ROS). Among heavy metals, Cr is widely dis-
tributed in nature because of their multifarious use in lea-
ther, refractory steel, drilling muds and electroplating
industries. The adverse effects of Cr on the yield, seed
germination, inhibition of some metalloenzymes, growth
and development have been well documented (Shanker
et al. 2005). It occurs in two stable forms which are tri-
valent Cr(III) and hexavalent Cr(VI) species. Cr(VI) usu-
ally occurs associated with oxygen as chromate (CrO42-)
or dichromate (Cr2O72-) oxyanions. Cr(VI) is reported to
be more mobile and toxic compared to Cr(III) species (Yu
et al. 2007). Besides, Cr(VI) is a strong oxidant with high
I. Sharma � R. Bhardwaj (&)
Department of Botanical and Environmental Sciences, Guru
Nanak Dev University, Amritsar, Punjab 143005, India
e-mail: [email protected];
I. Sharma
e-mail: [email protected]
P. K. Pati
Department of Biotechnology, Guru Nanak Dev University,
Amritsar, Punjab 143005, India
e-mail: [email protected]
123
Ecotoxicology (2011) 20:862–874
DOI 10.1007/s10646-011-0650-0

redox potential which accounts for the generation of free
radicals. Thus, Cr toxicity is mediated by the formation of
ROS (Yu et al. 2007) and by catalysis of the Haber–Weiss
reaction (Halliwell and Gutteridge 2007). The ROS are
highly toxic and can oxidize biological macromolecules
such as nucleic acids, proteins and lipids, thereby disturb-
ing the membrane permeability (Schutzendubel and Polle
2002; Sudo et al. 2008). Thus, ROS accumulation leads to
imbalance in pro-oxidative and antioxidative defence sys-
tem resulting in oxidative stress.
Several plant hormones like abscisic acid (ABA), eth-
ylene, jasmonates and BRs play a determinant role in
implicating oxidative stress. However, BRs are unique in
their activities for not only regulating the diverse physio-
logical and morphogenetic responses in plants but also
having a significant role in amelioration of various biotic
and abiotic stresses at nanomolar to micromolar concen-
trations (Krishna 2003; Clouse and Sasse 1998). Recently
28-HBL (10-6–10-11 M) has been reported to combat salt
and heavy metals stress in Brassica juncea, Raphanus
sativus, Triticum aestivum and Zea mays (Bhardwaj et al.
2007; Hayat et al. 2007; Sharma et al. 2007, 2010; Yusuf
et al. 2010). Furthermore, 28-HBL is also observed to
regulate the activities of various enzymes involved in
photosynthesis and plant defence in wheat and Indian
mustard exposed to different abiotic stresses (Hayat et al.
2007; Yusuf et al. 2010). Since BRs control several agro-
nomic traits in plants, therefore their potential in phyto-
remediation ‘‘basically a modified agronomic practice’’ is a
contentious issue from ecological viewpoint (Barbafieri
and Tassi 2011). However, development of efficient phy-
toremediation methods by increasing the efficiency of
uptake of metals/pollutants, species adaptation to particular
ecological niche requires rational scientific study. The
toxicological effects of heavy metals in plants are mediated
at several levels by different factors that also depend on
plant’s genotypic and phenotypic constitution. A number of
plant species, like B. juncea, R. sativus etc. in Brassicaceae
are known as hyperaccumulators, have adapted to envi-
ronmental stresses.
In the seeds of R. sativus, four types of BRs namely cas-
tasterone, brassinolide, 28-homoteasterone and teasterone
have been identified (Schmidt et al. 1991, 1993). In view of
the above as well as the wide occurrence, economic impor-
tance and phytoremediation perspectives of R. sativus, it is
much warranted to explore the possible role of BRs for
ameliorating specific stress in this plant. Thus, the present
study first time reports the effects of 28-HBL on morpho-
logical parameters, chlorophyll content, Cr(VI) metal
uptake, determination of superoxide anion production rate,
protein content, proline content and antioxidant enzymes
like superoxide dismutase (SOD), guaiacol peroxidase
(POD), catalase (CAT), dehydroascorbate reductase
(DHAR), monodehydroascorbate reductase (MDHAR),
glutathione reductase (GR) and ascorbate peroxidase
(APOX) and lipid peroxidation in R. sativus L. (Pusa Che-
taki) seedlings under Cr metal stress.
Materials and methods
Plant material and growth conditions
Seeds of R. sativus L. (Pusa Chetaki) were procured from
Department of Plant Breeding, Punjab Agriculture Univer-
sity, Ludhiana, India. Seeds were surface sterilized with
0.4% sodium hypochlorite for 15 min followed by repeated
rinses in sterile distilled water. Surface sterilized seeds were
given 8-h presoaking treatment in different concentrations of
28-HBL (0, 10-11, 10-9 and 10-7M). These pretreated seeds
were germinated on Whatman No. 1 filter paper lined auto-
claved glass Petri dishes (10 cm diameter, 20 seeds/dish)
each containing 4.0 ml of different concentrations of Cr (0,
0.5, 1.0 and 1.5 mM).TheCr (CrVI)was given in the formof
K2CrO4. The Petri dishes were sealed with parafilm to avoid
the desiccation and evaporation of solutions. However,
2–3 ml test solutions were added on 4th day of the experi-
ment. The experiment was conducted under controlled
conditions of light (16 h photoperiod under fluorescentwhite
light with 175 lmol m-2 s-1 intensity), temperature
(25 ± 5°C) and relative humidity (80–90%). Experiment
was repeated twice with five replicates for each treatment.
Growth analysis
On the last day of the experiment (that is, day 7 after
sowing the seeds including germination time of seedlings),
seedlings were harvested followed by separation of roots
and shoots. Seedling growth in terms of root and shoot
length was recorded. Twenty seedlings per petridish were
used for the determination of morphological parameters
(root/shoot length).
Cr(VI) uptake
Freshly harvested leaves were oven dried at 80°C for 24 h
and then ground to a fine powder. For analysis of metal
uptake dried leaf powder/sample (0.5 g) was acid digested
in H2SO4:HNO3:HClO4 (1:5:1) digestion mixture as
described by Allen et al. (1976). These digests were diluted
with double distilled water, fine filtered through Whatman
No. 1 filter paper and further diluted to make final volume
to 10 ml. The colorimetric method for analyzing the uptake
of Cr(VI) ions spectrophotometrically was followed after
Eaton et al. (1995). From 10 ml solution, 1 ml solution was
taken and diluted 50 times with 2 N H2SO4 (to adjust
Effect of 28-homobrassinolide on antioxidant defence system 863
123

pH = 1.0 ± 0.3). After 20 min, 1 ml Diphenyl carbazide
solution was added and absorbance of pink color end
product was observed at 540 nm. From the standard curve,
the uptake of Cr(VI) was calculated and expressed in terms
of mg g-1 dry biomass of leaf.
Biochemical analysis
To conduct biochemical analysis, the cotyledonary leaves
of 7-days old radish seedlings were harvested. For esti-
mation of the activities of antioxidant enzymes and the
protein content, leaf extracts were prepared by homoge-
nizing 2 g leaves in chilled 6 ml 50 mM phosphate buffer
(pH 7.0), 1 mM ethylenediaminetetraacetic acid (EDTA),
1 mM phenylmethanesulfonylfluoride (PMSF), 0.5% (v/v)
Triton X-100 and 2% (w/v) PVP-30 in a pre-chilled mortar
and pestle. In case of analysis of ascorbate peroxidase and
dehydroascorbate reductase activity, 0.5 mM ascorbate
was added to the extraction buffer. In case of monodehy-
droascorbate reductase activity, 1 g leaves were homoge-
nized in 50 mM Tris–HCl buffer (pH 7.6) containing
2.5 mM Ascorbic acid. The homogenates were centrifuged
at 12,0009g for 20 min at 4°C. The supernatant was fur-
ther used for biochemical analysis of antioxidant enzymes
and proteins.
Chlorophyll content
Chlorophyll (Chl) content was quantified by following the
method of Arnon (1949). Freshly harvested leaves (0.2 g)
were homogenized in 3.0 ml of 80% acetone followed by
its centrifugation at 12,0009g for 15 min. Absorbance of
supernatant was observed at 645 and 663 nm against 80%
acetone as blank. The contents of total Chl, Chl A and Chl
B were calculated in terms of lg ml-1 as described by
Arnon (1949).
Lipid peroxidation
Lipid peroxidation was determined by measuring the con-
tent of Malondialdehyde (MDA), a secondary end product
of the oxidation of polyunsaturated fatty acids, by the
method of Hodges et al. (1999). One gram shoots were
homogenized in 5 ml of 80% ethanol and then centrifuged
at 12,0009g for 5 min. Experiment was conducted using
20.0% (w/v) TCA (trichloroacetic acid) i.e. (-) TBA
(thiobarbituric acid) solution and 0.65% (w/v) TBA in
20.0% (w/v) TCA i.e. (?) TBA solution. One ml aliquot of
supernatant was added to two different test tubes, one
containing 1 ml (-) TBA solution and other test tube
containing (?) TBA solution. Samples were then mixed
vigorously, kept in water bath at 95°C for 30 min and then
cooled quickly on ice bath. Then, samples were centrifuged
at 12,0009g for 5 min. Absorbance of red adduct was
observed at 440, 532, and 600 nm and Malondialdehyde
equivalents g-1 fresh weight (nmol ml-1) were calculated
as described by Hodges et al. (1999).
Proline content
The free proline content was estimated spectrophotomet-
rically following the method of Bates et al. (1973). Fresh
1 g cotyledonary leaves were homogenized in 3.5 ml of
3% sulphosalicylic acid and the homogenates were cen-
trifuged at 12,0009g for 10 min. Then, 2 ml of supernatant
was reacted with 2 ml of acid ninhydrin and 2 ml of glacial
acetic acid in test tubes for 1 h at 100°C. The reaction was
terminated by putting the test tubes in ice bath. The reac-
tion mixture was extracted with 4 ml of toluene and mixed
vigorously by shaking for 15–20 s. Then toluene layer was
separated from aqueous phase and warmed to room tem-
perature. The absorbance of red colored Proline-ninhydrin
product was measured in toluene layer at 520 nm. Proline
concentration was calculated from a standard curve using
0–500 lM concentrations of L-proline.
Determination of superoxide anion
The superoxide anion production rate was determined
spectrophotometrically by monitoring the nitrite formation
from hydroxylamine in the presence of superoxide anion
(O2•-) as described by Wu et al. (2010) with some modi-
fications. Fresh 1 g cotyledonary leaves were homogenized
in liquid N2 in pre-chilled pestle and mortar. Then, 3 ml of
50 mM phosphate buffer (pH 7.8) containing 2% (w/v)
polyvinylpyrrolidone (PVP-30) and 0.5% (v/v) Triton
X-100 was added to the homogenized tissue followed by its
centrifugation at 12,0009g for 15 min at 4°C. The super-
natant (0.5 ml) was mixed with 0.5 ml of 50 mM phos-
phate buffer (pH 7.8) and 0.1 ml of 10 mM hydroxylamine
hydrochloride, and then incubated at 25°C for 30 min. The
incubated solution (1 ml) was added to 1 ml of 58 mM
3-aminobenzenesulphonic acid and 1 ml of 7 mM
1-naphthylamine, and then further kept at 25°C. After
20 min, absorbance was recorded at 530 nm. To calculate
the rate of O2•- production from the reaction of O2
•- with
hydroxylamine a standard curve was used. The O2•- pro-
duction rate was expressed as nmol min-1 g -1 FW.
Protein quantification
Total protein content of different samples of Cr (0, 0.5, 1.0
and 1.5 mM) and 28-HBL (10-7, 10-9 and 10-11 M) alone
or in combinations, was quantified by following the method
of Bradford (1976) using bovine serum albumin as a
standard.
864 I. Sharma et al.
123

Antioxidant enzyme activities
Ascorbate peroxidase assay The ascorbate peroxidase
(APOX, EC 1.11.1.11) activity was determined spectro-
photometrically as described by Nakano and Asada (1981).
The 3.0 ml reaction mixture contained 50 mM Potassium
phosphate buffer (pH 7.0), 0.5 mM ascorbate, 1.0 mM
H2O2 and 100 ll enzyme extract. The H2O2 dependent
oxidation of ascorbate was followed by monitoring the
decrease in absorbance at 290 nm using the extinction
coefficient 2.8 mM-1 cm-1. The reaction was carried out
for 3 min at 25°C. One unit of APOX activity is defined as
the amount of enzyme that can oxidize 1 lmol of ascorbate
per minute.
Catalase assay Catalase (CAT, EC 1.11.1.6) activity was
assayed by measuring the initial rate of H2O2 disappear-
ance using the method of Aebi (1984). The 3.0 ml reaction
mixture contained 50 mM Potassium phosphate buffer (pH
7.0), 15 mM H2O2 and 100 ll enzyme extract The
decrease in hydrogen peroxide was followed as decline in
optical density at 240 nm for 30 s at 25°C. The enzyme
activity was calculated using an extinction coefficient
39.4 mM-1 cm-1 for H2O2.
Dehydroascorbate reductase assay Dehydroascorbate
reductase (DHAR, EC 1.8.5.1) activity was measured fol-
lowing the method given by Dalton et al. (1986). The
3.0 ml reaction mixture contained 50 mM Potassium
phosphate buffer (pH 7.0), 0.2 mM dehydroascorbate,
0.1 mM EDTA, 2.5 mM reduced glutathione (GSH) and
100 ll enzyme extract. The DHAR activity was measured
by following the increase in absorbance at 265 nm due to
ascorbate formation at 265 nm using extinction coefficient
of 14 mM-1 cm-1.
Glutathione reductase assay Glutathione reductase (GR,
EC 1.6.4.2) activity was determined by using the method of
Carlberg and Mannervik (1975). The 3.0 ml reaction
mixture contained 50 mM potassium phosphate buffer (pH
7.6), 1 mM oxidized glutathione (GSSG), 0.5 mM EDTA,
0.1 mM reduced nicotinamideadenine dinucleotidephos-
phate (NADPH) and 100 ll enzymes extract. The reaction
was initiated by addition of 0.1 mM NADPH at 25°C. The
GR activity was determined by the oxidation of NADPH at
340 nm with extinction coefficient of 6.22 mM-1 cm-1.
Guaiacol peroxidase assay Guaiacol peroxidase (POD,
EC 1.11.1.7) activity was assayed using the method of
Sanchez et al. (1995) with some modifications. The 3.0 ml
reaction mixture contained 50 mM Potassium phosphate
buffer (pH 7.0), 20 mM guaiacol, 12.3 mM H2O2 and
100 ll enzyme extract. The POD activity was determined
by measuring the absorbance at 436 nm and using an
extinction coefficient of 26.6 mM-1 cm-1. One unit of
POD activity represents the amount of enzyme catalyzing
the oxidation of 1 lmol of guaiacol in 1 min.
Monodehydroascorbate reductase assay Monodehydro-
ascorbate reductase (MDHAR, EC 1.6.5.4) activity was
assayed using the method of Hossain et al. (1984). The
3.0 ml reaction mixture contained 50 mM Tris–HCl (pH
7.6) containing 2.5 mM Ascorbic acid, 0.1 mM reduced
nicotinamideadenine dinucleotide (NADH), 0.14 units of
ascorbic acid oxidase and 100 ll of enzyme extract.
Reaction was started by adding ascorbic acid oxidase and
the enzyme activity is measured by following the decrease
in absorbance due to the oxidation of NADH at 340 nm.
This decrease in absorbance was measured for 1 min and
enzyme activity was determined using extinction coeffi-
cient of 6.2 mM-1 cm-1.
Superoxide dismutase assay Superoxide dismutase (SOD,
EC 1.15.1.1) activity was assayed by measuring the ability
of the enzyme extract to inhibit the photochemical reduc-
tion of nitrobluetetrazolium (NBT) (Kono 1978). For total
SOD assay, 3.0 ml reaction mixture contained 50 mM
sodium carbonate (pH 10.2), 24 lM NBT, 0.1 mM EDTA,
1 mM hydroxylamine, 0.03% (v/v) Triton X-100 and 70 ll
enzyme extract. The absorbance was recorded at 560 nm
for 2 min. One unit of SOD activity was defined as the
amount of enzyme required that caused 50% of NBT
reduction at 25°C.
Statistical analysis
Five replicates (each containing 20 seedlings) for each
treatment were maintained. All data were subjected to two-
way analysis of variance (ANOVA) for scrutinizing inter-
actions of Cr and 28-HBL and expressed as the
mean ± standard error of five replicates. The Holm–Sidak
post hoc test was applied for the multiple comparisons
versus control using SigmaStat Version 3.5 and signifi-
cance of difference between the Cr-stress and 28-HBL
treatments was set at p B 0.05.
Results
Morphological parameters
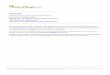
Chromium stress imposition resulted in decreased root and
shoot length of radish seedlings (Table 1). About 4.9 times
(11.3–2.3 cm) and 2.82 times (5.967–2.117 cm) decrease
was observed at 1.5 mM Cr stress in root length and shoot
length respectively. Application of 28-HBL (10-11, 10-9
Effect of 28-homobrassinolide on antioxidant defence system 865
123
and 10-7 M) alone had significantly improved their root
and shoot lengths (Table 1). Further, supplementation of Cr
solutions with 28-HBL considerably reduced the inhibitory
effect of Cr on seedling length. Root length was observed
to be improved significantly by the application of 10-7 M
(16.22 cm), 10-9 M (15.37 cm) and 10-11 M (15.2 cm)
28-HBL alone (Table 1). When 10-7 M 28-HBL supple-
mented with 0.5 mM Cr solution, root length (15.65 cm)
was enhanced as compared to control (seedlings treated
with 0.5 mM Cr alone) (6.64 cm) (Table 1). A similar
trend was found when effect of 28-HBL was studied on
shoot length of the seedlings (Table 1). It was observed
that irrespective of the concentrations of 28-HBL, it has
promoting effect on root and shoot length of radish seed-
lings under Cr stress. Overall, 10-7 M 28-HBL application
was most effective in enhancing growth of seedling treated
with Cr(VI).
Uptake of Cr(VI) ions
To understand effect of 28-HBL on Cr accumulation in
radish shoots, the uptake of Cr(VI) ions was analyzed.
Observations on uptake of Cr(VI) metal ions in Cr-treated
radish seedlings revealed a significant increase in leaves
(Table 1). The content of Cr(VI) ions was highest
(0.398 mg g-1 dry matter) in 1.0 mM Cr stressed radish
leaves. But, seed-presoaking treatment of 28-HBL alone
had no significant effect on Cr(VI) uptake in comparison to
control. However, these treatments significantly lowered
uptake of Cr ions in stressed seedlings (Table 1). About
sixfolds (0.286 to 0.045 mg g-1 dry matter) decrease in
Cr(VI) uptake was observed at 0.5 mM Cr by 10-7 M
28-HBL than 0.5 mM Cr alone. Similarly, same treatment
was effective in lowering Cr(VI) ions uptake to approxi-
mately 2 times in 1.0 and 1.5 mM Cr stressed radish
seedlings. Overall among different treatments of 28-HBL,
10-7 M concentration showed maximum decrease in
Cr(VI) ions uptake in Cr treated radish leaves.
Biochemical parameters
The studies carried out on the biochemical parameters
(Tables 2, 4, 5, 6) in radish shoots revealed the consider-
able effects of 28-HBL treatments. The analyses of rate of
superoxide radical formation, activities of antioxidant
enzymes as well as contents of chlorophyll, protein and
MDA, revealed that total protein content, chlorophyll
contents and activities of antioxidant enzymes significantly
increased with increasing concentrations of 28-HBL except
for the activity of POD under Cr metal stress. Although
treatment of 28-HBL alone could not increase free proline
content significantly but 28-HBL under Cr-stress signifi-
cantly enhanced free proline content (Table 3). However, aTable
1Effectof28-H
BLonrootlength,shootlength
andCr(VI)
ionsuptakein
7-daysold
R.sativusseedlingsunder
Crstress
Rootlength
(cm)at
varioustreatm
ents
of28-H
BL
Shootlength
(cm)at
varioustreatm
ents
of28-H
BL
Cr(VI)
uptake(m
g-1g-1dry
weight)at
varioustreatm
ents
of28-H
BL
0M
10-11M
10-9M
10-7M
0M
10-11M
10-9M
10-7M
0M
10-11M
10-9M
10-7M
0mM
Cr
11.3
±0.601a,p
15.37
±0.426a,q
15.2
±0.295a,q
16.22
±0.590a,q
5.967
±0.219a,p
6.28
±0.099a,p
6.443
±0.170a,p
6.873
±0.233a,p
0.000
±0.000a,p
0.0001
±0.0021a,p
0.0007
±0.009a,p
0.0001
±0.007a,p
0.5
mM
Cr
6.64
±0.322b,p
9.68
±0.180b,q
11.87
±0.560b,r
15.65
±0.381b,s
4.5 ±0.153b,p
5.6 ±0.115b,q
4.933
±0.203b,r
6.467
±0.067b,s
0.045
±0.002b,p
0.151
±0.0018b,qr
0.129
±0.006b,r
0.286
±0.008b,r
1.0
mM
Cr
6.333
±0.580b,p
8.79
±0.120bc,q
10.24
±0.373c,r
14.31
±0.960b,s
3.233
±0.088c,p
5.533
±0.012b,q
4.567
±0.004b,r
5.467
±0.600b,s
0.221
±0.008c,p
0.215
±0.012c,q
0.289
±0.0073c,s
0.398
±0.0096c,s
1.5
mM
Cr
2.3
±0.270c,p
6.78
±0.203c,q
9.73
±0.279c,r
13.42
±0.219c,s
2.117
±0.113d,p
3.6 ±0.058c,q
4.067
±0.016b,q
5.367
±0.007c,r
0.175
±0.0027d,p
0.136
±0.003d,qs
0.213
±0.007c,s
0.204
±0.0019c,rs
Datapresentedis
mean
±SE.Differentletters(a,b,c,d)within
acolumnaresignificantlydifferent(H
olm
–Sidakposthoctest,pB
0.05)whereasdifferentletters(p,q,r,s)within
arowforasingleparam
eter
aresignificantlydifferent(H
olm
–
Sidakpost
hoctest,pB
0.05)andsignifyinteractionsofdifferentconcentrations28-H
BLwithCronrootlength,shootlength
andCr(VI)
ionsuptakerespectively
866 I. Sharma et al.
123
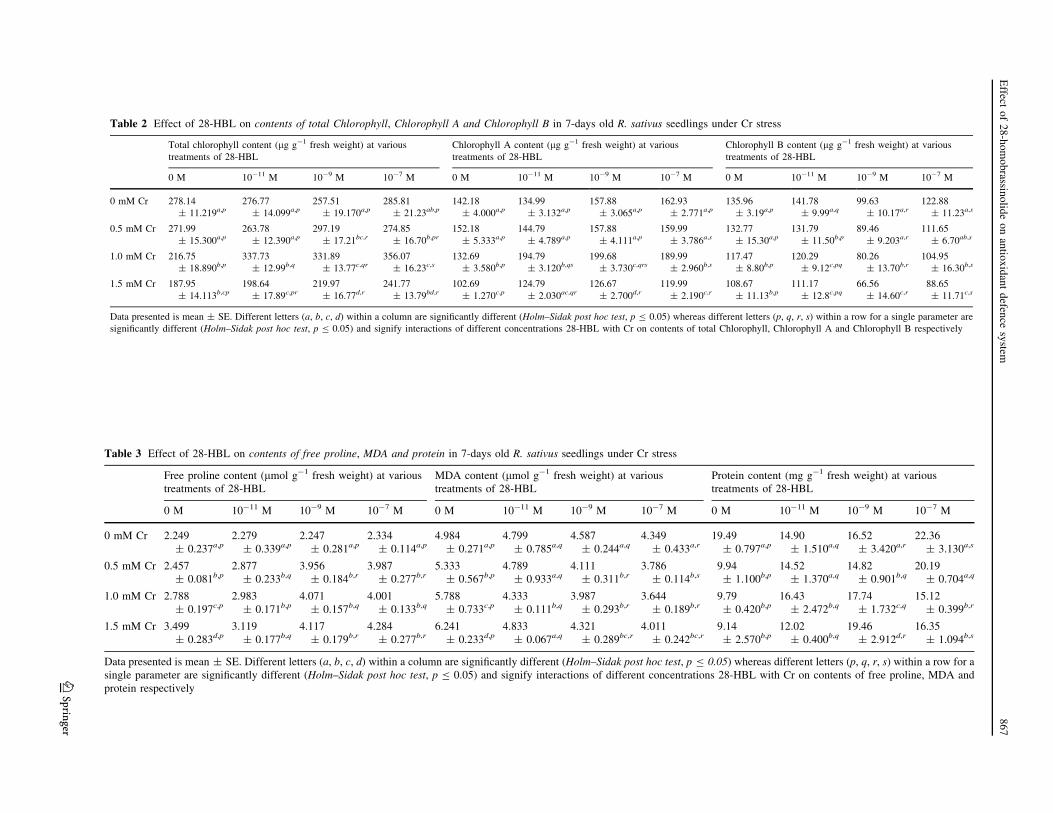
Table 2 Effect of 28-HBL on contents of total Chlorophyll, Chlorophyll A and Chlorophyll B in 7-days old R. sativus seedlings under Cr stress
Total chlorophyll content (lg g-1 fresh weight) at various
treatments of 28-HBL
Chlorophyll A content (lg g-1 fresh weight) at various
treatments of 28-HBL
Chlorophyll B content (lg g-1 fresh weight) at various
treatments of 28-HBL
0 M 10-11 M 10-9 M 10-7 M 0 M 10-11 M 10-9 M 10-7 M 0 M 10-11 M 10-9 M 10-7 M
0 mM Cr 278.14
± 11.219a,p276.77
± 14.099a,p257.51
± 19.170a,p285.81
± 21.23ab,p142.18
± 4.000a,p134.99
± 3.132a,p157.88
± 3.065a,p162.93
± 2.771a,p135.96
± 3.19a,p141.78
± 9.99a,q99.63
± 10.17a,r122.88
± 11.23a,s
0.5 mM Cr 271.99
± 15.300a,p263.78
± 12.390a,p297.19
± 17.21bc,r274.85
± 16.70b,pr152.18
± 5.333a,p144.79
± 4.789a,p157.88
± 4.111a,p159.99
± 3.786a,s132.77
± 15.30a,p131.79
± 11.50b,p89.46
± 9.203a,r111.65
± 6.70ab,s
1.0 mM Cr 216.75
± 18.890b,p337.73
± 12.99b,q331.89
± 13.77c,qr356.07
± 16.23c,s132.69
± 3.580b,p194.79
± 3.120b,qs199.68
± 3.730c,qrs189.99
± 2.960b,s117.47
± 8.80b,p120.29
± 9.12c,pq80.26
± 13.70b,r104.95
± 16.30b,s
1.5 mM Cr 187.95
± 14.113b,cp198.64
± 17.89c,pr219.97
± 16.77d,r241.77
± 13.79bd,r102.69
± 1.270c,p124.79
± 2.030ac,qr126.67
± 2.700d,r119.99
± 2.190c,r108.67
± 11.13b,p111.17
± 12.8c,pq66.56
± 14.60c,r88.65
± 11.71c,s
Data presented is mean ± SE. Different letters (a, b, c, d) within a column are significantly different (Holm–Sidak post hoc test, p B 0.05) whereas different letters (p, q, r, s) within a row for a single parameter are
significantly different (Holm–Sidak post hoc test, p B 0.05) and signify interactions of different concentrations 28-HBL with Cr on contents of total Chlorophyll, Chlorophyll A and Chlorophyll B respectively
Table 3 Effect of 28-HBL on contents of free proline, MDA and protein in 7-days old R. sativus seedlings under Cr stress
Free proline content (lmol g-1 fresh weight) at various
treatments of 28-HBL
MDA content (lmol g-1 fresh weight) at various
treatments of 28-HBL
Protein content (mg g-1 fresh weight) at various
treatments of 28-HBL
0 M 10-11 M 10-9 M 10-7 M 0 M 10-11 M 10-9 M 10-7 M 0 M 10-11 M 10-9 M 10-7 M
0 mM Cr 2.249
± 0.237a,p2.279
± 0.339a,p2.247
± 0.281a,p2.334
± 0.114a,p4.984
± 0.271a,p4.799
± 0.785a,q4.587
± 0.244a,q4.349
± 0.433a,r19.49
± 0.797a,p14.90
± 1.510a,q16.52
± 3.420a,r22.36
± 3.130a,s
0.5 mM Cr 2.457
± 0.081b,p2.877
± 0.233b,q3.956
± 0.184b,r3.987
± 0.277b,r5.333
± 0.567b,p4.789
± 0.933a,q4.111
± 0.311b,r3.786
± 0.114b,s9.94
± 1.100b,p14.52
± 1.370a,q14.82
± 0.901b,q20.19
± 0.704a,q
1.0 mM Cr 2.788
± 0.197c,p2.983
± 0.171b,p4.071
± 0.157b,q4.001
± 0.133b,q5.788
± 0.733c,p4.333
± 0.111b,q3.987
± 0.293b,r3.644
± 0.189b,r9.79
± 0.420b,p16.43
± 2.472b,q17.74
± 1.732c,q15.12
± 0.399b,r
1.5 mM Cr 3.499
± 0.283d,p3.119
± 0.177b,q4.117
± 0.179b,r4.284
± 0.277b,r6.241
± 0.233d,p4.833
± 0.067a,q4.321
± 0.289bc,r4.011
± 0.242bc,r9.14
± 2.570b,p12.02
± 0.400b,q19.46
± 2.912d,r16.35
± 1.094b,s
Data presented is mean ± SE. Different letters (a, b, c, d) within a column are significantly different (Holm–Sidak post hoc test, p B 0.05) whereas different letters (p, q, r, s) within a row for a
single parameter are significantly different (Holm–Sidak post hoc test, p B 0.05) and signify interactions of different concentrations 28-HBL with Cr on contents of free proline, MDA and
protein respectively
Effect
of28-homobrassin
olid
eonantio
xidantdefen
cesystem
867
123
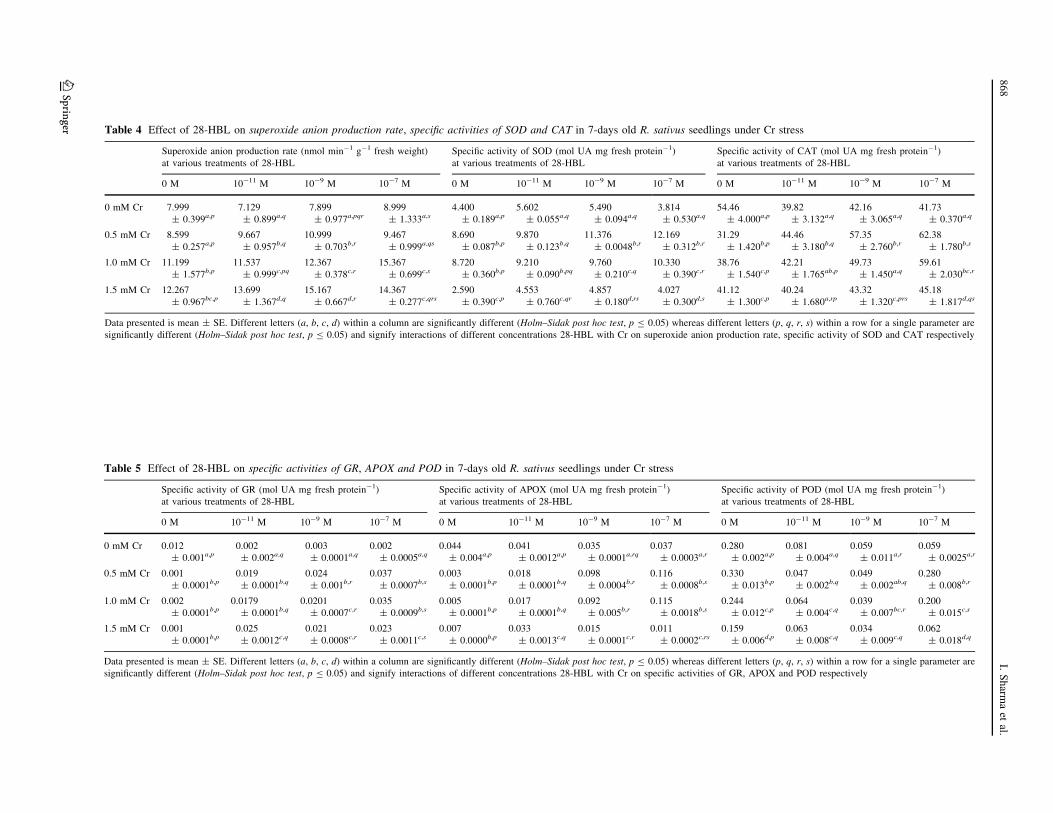
Table 4 Effect of 28-HBL on superoxide anion production rate, specific activities of SOD and CAT in 7-days old R. sativus seedlings under Cr stress
Superoxide anion production rate (nmol min-1 g-1 fresh weight)
at various treatments of 28-HBL
Specific activity of SOD (mol UA mg fresh protein-1)
at various treatments of 28-HBL
Specific activity of CAT (mol UA mg fresh protein-1)
at various treatments of 28-HBL
0 M 10-11 M 10-9 M 10-7 M 0 M 10-11 M 10-9 M 10-7 M 0 M 10-11 M 10-9 M 10-7 M
0 mM Cr 7.999
± 0.399a,p7.129
± 0.899a,q7.899
± 0.977a,pqr8.999
± 1.333a,s4.400
± 0.189a,p5.602
± 0.055a,q5.490
± 0.094a,q3.814
± 0.530a,q54.46
± 4.000a,p39.82
± 3.132a,q42.16
± 3.065a,q41.73
± 0.370a,q
0.5 mM Cr 8.599
± 0.257a,p9.667
± 0.957b,q10.999
± 0.703b,r9.467
± 0.999a,qs8.690
± 0.087b,p9.870
± 0.123b,q11.376
± 0.0048b,r12.169
± 0.312b,r31.29
± 1.420b,p44.46
± 3.180b,q57.35
± 2.760b,r62.38
± 1.780b,s
1.0 mM Cr 11.199
± 1.577b,p11.537
± 0.999c,pq12.367
± 0.378c,r15.367
± 0.699c,s8.720
± 0.360b,p9.210
± 0.090b,pq9.760
± 0.210c,q10.330
± 0.390c,r38.76
± 1.540c,p42.21
± 1.765ab,p49.73
± 1.450a,q59.61
± 2.030bc,r
1.5 mM Cr 12.267
± 0.967bc,p13.699
± 1.367d,q15.167
± 0.667d,r14.367
± 0.277c,qrs2.590
± 0.390c,p4.553
± 0.760c,qr4.857
± 0.180d,rs4.027
± 0.300d,s41.12
± 1.300c,p40.24
± 1.680a,rp43.32
± 1.320c,prs45.18
± 1.817d,qs
Data presented is mean ± SE. Different letters (a, b, c, d) within a column are significantly different (Holm–Sidak post hoc test, p B 0.05) whereas different letters (p, q, r, s) within a row for a single parameter are
significantly different (Holm–Sidak post hoc test, p B 0.05) and signify interactions of different concentrations 28-HBL with Cr on superoxide anion production rate, specific activity of SOD and CAT respectively
Table 5 Effect of 28-HBL on specific activities of GR, APOX and POD in 7-days old R. sativus seedlings under Cr stress
Specific activity of GR (mol UA mg fresh protein-1)
at various treatments of 28-HBL
Specific activity of APOX (mol UA mg fresh protein-1)
at various treatments of 28-HBL
Specific activity of POD (mol UA mg fresh protein-1)
at various treatments of 28-HBL
0 M 10-11 M 10-9 M 10-7 M 0 M 10-11 M 10-9 M 10-7 M 0 M 10-11 M 10-9 M 10-7 M
0 mM Cr 0.012
± 0.001a,p0.002
± 0.002a,q0.003
± 0.0001a,q0.002
± 0.0005a,q0.044
± 0.004a,p0.041
± 0.0012a,p0.035
± 0.0001a,rq0.037
± 0.0003a,r0.280
± 0.002a,p0.081
± 0.004a,q0.059
± 0.011a,r0.059
± 0.0025a,r
0.5 mM Cr 0.001
± 0.0001b,p0.019
± 0.0001b,q0.024
± 0.001b,r0.037
± 0.0007b,s0.003
± 0.0001b,p0.018
± 0.0001b,q0.098
± 0.0004b,r0.116
± 0.0008b,s0.330
± 0.013b,p0.047
± 0.002b,q0.049
± 0.002ab,q0.280
± 0.008b,r
1.0 mM Cr 0.002
± 0.0001b,p0.0179
± 0.0001b,q0.0201
± 0.0007c,r0.035
± 0.0009b,s0.005
± 0.0001b,p0.017
± 0.0001b,q0.092
± 0.005b,r0.115
± 0.0018b,s0.244
± 0.012c,p0.064
± 0.004c,q0.039
± 0.007bc,r0.200
± 0.015c,s
1.5 mM Cr 0.001
± 0.0001b,p0.025
± 0.0012c,q0.021
± 0.0008c,r0.023
± 0.0011c,s0.007
± 0.0000b,p0.033
± 0.0013c,q0.015
± 0.0001c,r0.011
± 0.0002c,rs0.159
± 0.006d,p0.063
± 0.008c,q0.034
± 0.009c,q0.062
± 0.018d,q
Data presented is mean ± SE. Different letters (a, b, c, d) within a column are significantly different (Holm–Sidak post hoc test, p B 0.05) whereas different letters (p, q, r, s) within a row for a single parameter are
significantly different (Holm–Sidak post hoc test, p B 0.05) and signify interactions of different concentrations 28-HBL with Cr on specific activities of GR, APOX and POD respectively
868
I.Sharm
aet
al.
123

remarkable reduction in content of MDA (Table 3) and rate
of superoxide radical formation (Table 4) was observed in
28-HBL treated seedlings grown under Cr toxicity.
Chlorophyll content
Phytotoxicity of Cr(VI) ions is due to its high mobility
across biological membranes and its interference with
electron transport in respiration and photosynthesis (Yu
et al. 2007). Thus, to determine toxicity of Cr(VI) ions,
chlorophyll (Chl) content was measured. The toxicity
induced by Cr resulted in significant decrease in contents of
total chlorophyll (Chl), Chl A and B (Table 2). About 1.5
times (from 278.18 to 187.95 lg g-1 FW) decrease in total
Chl content was decreased in 1.5 mM Cr stress than control
(Table 2). Similarly, 1.4 and 1.25 folds decline in contents
of Chl A (Table 2) and B (Table 2) was observed under
1.5 mM Cr treated seedlings than untreated. Seed presoa-
king treatments of 28-HBL alone had no significant effect
on total Chl and Chl A content. However, treatments of
28-HBL alone had significant enhancing effect on content
of Chl B (Table 2). Furthermore, 28-HBL treatment had
significantly increased Chl A and total Chl content in Cr
metal treated seedlings. Seed presoaking treatments at 10-7
M 28-HBL had resulted in 1.7 times increase in total Chl
content in 1.0 mM Cr stressed seedlings in comparison to
1.0 mM Cr alone (Table 2). A similar increasing effect of
28-HBL treatment was followed in Chl A content. While
28-HBL treatments significantly decreased the contents of
Chl B as compared to control.
Free proline content
The free proline content (Table 3) was observed to increase
significantly in radish seedlings grown under Cr toxicity
(2.457–3.499 lmol g-1 FW) when compared to control
(2.249 lmol g-1 FW) (Table 3). However, applications of
28-HBL alone could vaguely enhance free proline contents
and maximum enhancement at 10-7 M concentration
(2.344 lmol g-1 FW) as compared to untreated seedlings
(2.249 lmol g-1 FW) (Table 3). But seeds pretreated with
28-HBL under Cr metal stress, significantly enhanced free
proline levels than only Cr metal treated radish seedlings
(Table 3). 10-7 M of 28-HBL significantly increased pro-
line levels under Cr stress, with 1.224-fold increase at
1.5 mM Cr in conjunction with 10-7 M 28-HBL
(4.284 lmol g-1 FW) as compared to 1.5 mM Cr treat-
ment alone (3.499 lmol g-1 FW).
Lipid peroxidation
Chromium metal stress significantly enhanced the MDA
content (5.333–6.421 lmol g-1 FW) as compared toTable
6Effectof28-H
BLonspecificactivitiesofDHARandMDHARin
7-daysold
R.sativusseedlingsunder
Crstress
SpecificactivityofDHAR
(molUA
mgfreshprotein-1)at
varioustreatm
ents
of28-H
BL
SpecificactivityofMDHAR
(molUA
mgfreshprotein-1)at
varioustreatm
ents
of28-H
BL
0M
10-11M
10-9M
10-7M
0M
10-11M
10-9M
10-7M
0mM
Cr
0.0387±
0.003a,p
0.0321±
0.002a,q
0.0397±
0.0027a,r
0.041±
0.0029a,r
0.377±
0.033a,p
0.372±
0.043a,p
0.367±
0.013a,p
0.383±
0.019a,p
0.5
mM
Cr
0.0344±
0.0013b,p
0.0256±
0.0015b,q
0.0323±
0.002b,r
0.0392±
0.0028a,s
0.327±
0.017b,p
0.357±
0.019ba,q
0.379±
0.025ab,r
0.407±
0.018ab,s
1.0
mM
Cr
0.0367±
0.0012b,p
0.0287±
0.004b,q
0.0396±
0.0047c,r
0.0421±
0.0015a,s
0.254±
0.028c,p
0.334±
0.037bc,q
0.389±
0.041a,r
0.423±
0.026b,s
1.5
mM
Cr
0.0247±
0.006c,p
0.0264±
0.0048b,q
0.0347±
0.0064b,r
0.0407±
0.0018a,s
0.237±
0.019c,p
0.317±
0.037c,q
0.351±
0.029b,r
0.389±
0.029a,s
Datapresentedismean±
SE.Differentletters(a,b,c,d)within
acolumnaresignificantlydifferent(H
olm
–Sidakposthoctest,pB
0.05)whereasdifferentletters(p,q,r,s)within
arowfora
single
param
eter
aresignificantlydifferent(H
olm
–Sidakpost
hoctest,pB
0.05)andsignifyinteractionsofdifferentconcentrations28-H
BLwithCronspecificactivitiesofDHAR
and
MDHAR
respectively
Effect of 28-homobrassinolide on antioxidant defence system 869
123

control (4.984 lmol g-1 FW) in radish seedlings
(Table 3). The maximum increase in MDA content was
observed at 1.5 mM Cr (6.421 lmol g-1 FW) and this
increase in MDA content was further reduced significantly
by 28-HBL presoaking treatments (Table 3). The maxi-
mum reduction in MDA content was recorded in seedlings
treated with 10-7 M 28-HBL under 1.0 mM Cr metal stress
(3.644 lmol g-1 FW) (Table 3). It was also observed that
seedlings treated with 28-HBL alone could lower MDA
levels than control (Table 3). The most effective 28-HBL
concentration which reduced MDA level to the lowest level
was 10-7 M (4.349 lmol g-1 FW) (Table 3).
Total protein content
The observations on Cr stressed shoots revealed that pro-
tein content decreased with increased concentrations of Cr
metal (Table 3). Minimal protein content was observed in
case of shoots treated with 1.5 mM Cr (9.14 mg g-1 FW)
as compared to control (19.49 mg g-1 FW), where only
distilled water was given (Table 3). Shoots treated with
28-HBL alone showed significant increase in soluble pro-
tein content (Table 3) in comparison to untreated shoots.
The treatment of shoots with 10-7 M of 28-HBL resulted in
significantly enhanced protein content (22.36 mg g-1 FW)
when compared to the control (19.49 mg g-1 FW). The
protein content (Table 3) was significantly higher in the
shoots treated with Cr along with 28-HBL than Cr alone.
Superoxide (O2•-) anion production
Cr-induced oxidative stress is mediated via generation,
accumulation and imbalance of a steady-state level of ROS
in stressed plants. To analyze effect of 28-HBL in amelio-
ration of oxidative stress, the generation of O2•- anion was
determined (Table 4). In radish shoots, the production of
O2•- anion increased about 1.53 and 1.4 times respectively
under 1.5 and 1.0 mM Cr stress than control. Moreover,
28-HBL treatments alone had significantly increased O2•-
content as compared to control. Furthermore, 28-HBL had
also remarkably enhanced O2•- anion production under Cr-
metal toxicity. Seed presoaking 28-HBL treatment at 10-7M
had resulted in 1.37 folds (11.199–15.367 nmol min-1 g-1
FW) and 1.92 folds (7.999–15.367 nmol min-1 g-1 FW)
increase in O2•- production in 1.0 mM Cr when compared to
1.0 mM Cr alone and control respectively. Overall, 10-7 M
28-HBL treatment was most effective in enhancing the
generation of O2•- anion during Cr metal stress (Table 4).
Activities of antioxidant enzymes
Activity of SOD (Table 4) increased significantly under
0.5 mM (8.69 mol Unit Activity mg protein-1) and
1.0 mM (8.72 mol UA mg protein-1) Cr metal stress in
radish shoots as compared to control (4.4 mol UA mg
protein-1). Seed presoaking treatments of 28-HBL alone
was not able to alleviate the decreased levels of SOD
activity. However, when Cr metal solutions were supple-
mented alongwith 28-HBL, it resulted in increased levels
of SOD activity (Table 4). Maximum activity of SOD was
reported in case of radish shoots treated with 10-7 M
28-HBL supplemented with 0.5 mM Cr (12.169 mol UA
mg protein-1) when compared to 0.5 mM Cr alone
(8.69 mol UA mg protein-1) (Table 4). Activities of CAT
(Table 4) and GR (Table 5) were decreased under Cr stress
than control. There activities were significantly enhanced
in Cr treated shoots supplemented with 28-HBL in com-
parison to Cr metal alone. Activity of CAT (Table 4) was
maximum (62.38 mol UA mg protein-1) in shoots treated
with 10-7 M 28-HBL supplemented with 0.5 mM of Cr
solution (31.29 mol UA mg protein-1). Similarly, GR
activity (Table 5) was also increased significantly by
applications of different concentrations of 28-HBL under
Cr stress. Maximum enhancement in the activity of GR
(0.037 mol UA mg protein-1) was observed in case of
shoots, treated with 0.5 mM of Cr solution supplemented
with 10-7 M 28-HBL (Table 5). Activity of APOX was
increased with increased concentrations of Cr as compared
to control (Table 5). It was further enhanced by applica-
tions of different concentrations of 28-HBL under Cr stress
(Table 5). The maximum APOX activity (Table 5) was
observed in case of shoots, treated with 0.5 mM of Cr
solution supplemented with 10-7 M 28-HBL (0.116 mol
UA mg protein-1).
Application of 28-HBL significantly decreased the POD
activity (Table 5) and it was also decreased under metal
stress except 0.5 mM Cr stress. Supplementation of 10-9
and 10-11 M 28-HBL along with Cr metal significantly
decreased POD activity in the shoots (Table 5). The
activities of DHAR (Table 6) and MDHAR (Table 6) got
decreased from 0.0387 mol UA mg protein-1 and
0.377 mol UA mg protein-1 respectively in control to
0.0247 mol UA mg protein-1 (DHAR) and 0.237 mol UA
mg protein-1 (MDHAR) in Cr metal treated seedlings.
Pretreatments of 28-HBL alone significantly enhanced
DHAR activity in radish seedlings than control
(0.0387 mol UA mg protein-1), with maximum increase at
10-7 M 28-HBL alone (0.041 mol UA mg protein-1)
(Table 6). However, there was insignificant change in
MDHAR activity under the seed presoaking treatments of
28-HBL alone (Table 6). Yet, 28-HBL treatments under
metal stress could restore the activity of MDHAR and the
maximum enhancement was recorded at 10-7 M 28-HBL
along with 1.0 mM Cr (0.423 mol UA mg protein-1)
(Table 6). Also, in Cr stressed seedlings maximum activity
of DHAR (0.0421 mol UA mg-1 protein) was observed at
870 I. Sharma et al.
123

10-7 M 28-HBL in combination with 1.0 mM Cr metal
(Table 6). It was observed that 28-HBL helped in con-
quering the stress in radish plants by regulating the activ-
ities of antioxidant enzymes. The Present study thus
indicates a possible regulation of antioxidant enzymes by
BRs in amelioration of heavy metals stress in plants
(Fig. 1).
Discussion
Phytotoxicity induced by Cr have been explored in many
important crop plants and also in lower plants (Panda and
Choudhury 2005; Shanker et al. 2005; Yu et al. 2007).
Chromium causes wilting and plasmolysis in root cells and
thus affects root growth of plants (Panda and Choudhury
2005). Total root length of wheat was affected by 20 mg
Cr(VI) kg-1 soil (Chen et al. 2001). Rout et al. (1997)
reported adverse effects of Cr on plant height and shoot
growth. Growth inhibition by Cr can be due to chromo-
somal aberrations which lead to inhibition of cell division
(Liu et al. 1993). In present study, decrease in root length
(Table 1); shoot length (Table 1); Chl A, Chl B and total
Chl contents (Table 2) has been observed under Cr stress.
A similar declining trend for Chl content was observed in
Glycine max under water deficits which was improved by
brassinolide treatments (Zhang et al. 2008). The present
work reveals that seed-presoaking treatments of 28-HBL
improved the seedling growth and contents of total Chl
(Table 2) and Chl A (Table 2) under Cr stress. Besides,
increased uptake of Cr(VI) ions was remarkably lowered
by 28-HBL treatments thereby resulted in mitigation of Cr
metal toxicity in radish seedlings (Table 1). These results
are in coherence with the reports of Sharma et al. (2007). In
B. juncea, Zn metal ions uptake was significantly lowered
by 28-HBL treatments. Thus, applications of 28-HBL may
increase cell division, reduces chromosomal aberrations
and alters the membrane permeability (Fig. 1). Besides,
stress protective effects of 28-HBL on plant growth, pho-
tosynthesis, activities of carbonic anhydrase, nitate reduc-
tase and antioxidant enzymes in B. juncea plants under Cd
stress were studied by Hayat et al. (2007). They observed
that the toxic effects generated by Cd were reduced with
28-HBL spraying treatment due to increase in plant growth
and enhanced enzyme activities. Also, BRs have been
reported to increase the ATPase activity in azuki bean
epicotyls and maize roots, leading to proton extrusion, and
induced cell wall relaxation (Cerana et al. 1984; Haubrick
and Assmann 2006). Further, Bajguz (2000) stated that
BRs increased DNA, RNA and protein contents of Chlo-
rella vulgaris. Similarly, in the current study protein con-
tent was found to be significantly improved in Cr stressed
shoots treated with 28-HBL (Table 3).
Heavy-metal detoxification and tolerance is mediated
via metal-binding cystein-rich compounds like phytochel-
atins (PC) which facilitates the chelation of Cr ions in
cytosol or their compartmentalization in vacuoles (Fig. 1)
(Shanker et al. 2005). However, in higher concentrations
Fig. 1 Possible mechanism of brassinosteroids regulated heavy metal
stress tolerance in plants. Dotted line represents the proposed possible
mechanisms, which remains to be explored. ROS reactive oxygen
species, PC phytochelatins, BR brassinosteroid, BRI1 brassinosteroid
insensitive 1, BAK1 BRI1-associated kinase 1, P phosphate, BIN2
brassinosteroid insensitive 2, BZR1 brassinozole-resistant 1, BES1
BRI1-ems-suppressor 1, V-ATPase vacuolar H?-ATPase, APOX
ascorbate peroxidase, CAT catalase, POD guaiacol peroxidase, DHAR
dehydroascorbate reductase, MDHAR monodehydroascorbate redu-
catse, GR glutathione reductase
Effect of 28-homobrassinolide on antioxidant defence system 871
123

Cr(VI) generates ROS like superoxide radicals (O2•-),
hydrogen peroxide (H2O2), hydroxyl radicals (HO-), per-
oxy radicals (ROO•), alkoxy radicals (RO•), singlet oxygen
(O21), etc. (Bhattacharjee 2005; Shanker et al. 2005). Cr
induced toxicity leads to over-production of ROS and thus
causes oxidative damage leading to membrane destruction
which in turn affects the levels of antioxidants and metal-
lozymes or antioxidant enzymes (Pandey et al. 2005;
Shanker et al. 2005). Similarly, in present study, generation
of O2•- anion had significantly increased under Cr stress
(Table 4). Thus, Cr stress may enhance accumulation of
ROS resulting in disturbance in its detoxification. The
balance between ROS production and detoxification is
maintained by ROS-scavenging antioxidant enzymes such
as SOD, CAT, APOX, GR, DHAR, MDHAR and POD etc.
(Fig. 1) (Mittler 2002). Superoxide dismutase is a family of
metallozymes which catalyze the disproportionation of
superoxide radicals (O2•-) to O2 and H2O2 (Fig. 1). The
H2O2 is further removed by CAT in the peroxisomes or by
APOX in the chloroplast or by membrane bounded POD
(Foyer et al. 1997). Glutathione reductase is a member of
flavoenzyme family which catalyzes the NADPH depen-
dent reduction of glutathione disulphide (GSSG) to gluta-
thione (GSH) and it maintains glutathione in the reduced
state (Fig. 1), which in turn reduces dehydroascorbate to
ascorbate (Noctor and Foyer 1998). Besides, ascorbic acid
in its reduced form is essential for ROS scavenging and its
cellular pool is maintained by DHAR and MDHAR using
NADPH as reducing power (Mittler 2002). This may lead
to altered redox potential (Fig. 1) of the cells ensuing
membrane destabilization under stress (Bhattacharjee
2005; Scandalios 2005; Skorzynska-Polit et al. 2010).
Recently, Liu et al. (2010) reported that two mitogen-
activated protein kinases (MAPKs), MPK3 and MPK6, are
triggered in response to Cd via the accumulation of ROS in
Arabidopsis. Also, in response to both abiotic and biotic
stresses, Calcium-dependent protein kinases (CDPKs),
calmodulins, calmodulin-like proteins or calcineurin B-like
proteins are activated as important sensors of Ca2? in
plants (Ludwig et al. 2004). The role of stress on antioxi-
dant enzymes has been overemphasized in literature (Mit-
tler 2002; Scandalios 2005). However, the effect of BRs
and its implications against stress is not well understood.
McCord (2000) reported that 28-HBL treated seedlings
might be scavenging ROS more effectively than the seed-
lings treated with metal alone. It may be possible that
28-HBL maintained the altered redox status of cells by
regulating activities of antioxidant enzymes viz. SOD,
CAT, APOX, GR, DHAR, MDHAR etc. (Fig. 1). Further,
redox potential may be re-established by reducing lipid
peroxidation of phosho-lipids in cell membranes and
accumulation of certain osmoprotectants like proline or
betaine (Fig. 1). Previous reports also showed that
exogenous application of BRs modified antioxidant
enzyme activity in stressed maize, mustard, radish and
wheat plants (Bhardwaj et al. 2007; Sharma et al. 2007,
2010, 2011; Yusuf et al. 2010). BRs (modified Sterols) can
modulate the activity of proteins and other enzymes within
the membrane by affecting either protein conformation
(functionality) or protein activity by direct protein-sterol
interactions (Lindsey et al. 2003). Li et al. (2002) reported
that BRs are perceived by a protein complex that includes
the leucine-rich repeat receptor-like protein kinase (LRR-
RLK) brassinosteroid-insensitive 1 (BRI1) (Fig. 1). Wang
and He (2004) suggested the possibility that in plants BRI1
plays a role in defence by perceiving peptide signals.
Under stress these generated signals may regulate defence
responses. Recently, Dhaubhadel and Krishna (2008)
identified the six differentially expressed genes in 24-epi-
brassinolide treated heat stressed Brassica napus seedlings
which signify that BRs regulated genes are involved in
diverse physiological responses in stress conditions. Vari-
ous stress responsive genes regulated by BRs may code for
the phytochelatins, organic acids, osmolytes, metallothio-
nines and stress protective proteins (Late Embryogenesis
Abundant i.e., LEA proteins and Heat Shock Proteins, i.e.
HSPs) (Gendron and Wang 2007; McSteen and Zhao 2008)
(Fig. 1).
Over-production of ROS during Cr-stress might increase
oxidative degradation of phospholipids, ensuing in dis-
ruption of the permeability of cell membranes which can
not be re-stored by antioxidant enzymes (Sudo et al. 2008;
Skorzynska-Polit et al. 2010). However, it’s feasible that
treatments of BRs contribute in inhibition of lipid degra-
dation as well as stimulation of activities of antioxidant
enzymes under stress (Fig. 1). In present study, similar
increased expression of GR (Table 5), DHAR (Table 6)
and MDHAR (Table 6) in presence of 28-HBL enhances
tolerance to oxidative stress generated by Cr has been
observed. This information is in accordance to the previous
reports where exogenous application of BRs modifies
antioxidant enzyme activity. Here, POD activity (Table 5)
is decreased in Cr treated seedlings (except 0.5 mM Cr)
and further decreased by 28-HBL treatments, suggesting no
protective role for 28-HBL on POD activity. However,
28-HBL enhanced APOX activities in seedlings grown
under Cr metal stress (Table 5). Both POD and APOX are
involved in removal of H2O2. Thus, it can be concluded
that 28-HBL enhanced the activities of antioxidant
enzymes which might be owed to the up-regulation of the
Brassinosteroids-mediated gene expression during metal
stress (Fig. 1). This aligns with the prior reports of
Bhardwaj et al. (2007) and Hayat et al. (2007) in B. juncea
and Z. mays respectively. So, the alteration of antioxidant
enzyme activities may suggest that 28-HBL treated seed-
lings were less affected by Cr metal than the untreated
872 I. Sharma et al.
123

seedlings. Therefore, improvement in seedling growth,
contents of proline, protein, total Chl and Chl A, lowered
Cr(VI) ions uptake, lowered MDA content and activities of
antioxidant enzymes recommends the stress protective
effects of BRs.
Intensifying environmental pollution and shrinking
habitats has obligated to trail new, scientific and eco-
friendly techniques for rehabilitating and restoring polluted/
toxic sites for social use. Being economic, environment-
friendly and plant-based natural technique, phytoremedia-
tion offers a promising prospect for addressing ecological
concerns of a range of pollutants. Since plants with high and
rapid rate of biomass production and hyperaccumulation
potential are appropriate for phytoremediation, subse-
quently recent studies are focused to improve their effi-
ciency by use of additives for their enhanced applicability in
phytoremediation. Assisted-phytoremediation exploits the
use of chemical additives like chelators, organic acids etc.
to soil for increasing the bioavailability of heavy metals for
plant uptake. Recently, a few studies have addressed the use
of phytohormones as additives to plants for phytoremedia-
tion purpose (Barbafieri and Tassi 2011). Since, BRs con-
trol several agronomic traits in plants; therefore these are of
great concern for its implications in a modified agronomic
practice i.e., phytoremediation. Thus, studies on effects of
BRs could provide positive interactions with plants for
more efficient assisted-phytoremediation techniques to
reduce the risks associated with the exposure of human and
environment to sites contaminated with heavy metals.
Conclusions
In recent years, BRs have been reported for increasing
plant productivity and plant protection against biotic as
well as abiotic stress. The stress-amelioration might be
mediating at different levels of metal toxicity viz., metal
binding and its translocation, accumulation and detoxifi-
cation of ROS. Besides metal toxicity in plants also
depends on its uptake and bioaccumulation which is
influenced by the bioavailability, route of uptake, storage,
degradation, immobilization, excretion, and avoidance/
tolerance mechanisms. These characteristics are important
for enhancing the potential of assisted-phytoremediation in
hyperaccumulator plants to decontaminate the polluted
sites. Also, the role of phytohormones in assisted-phytor-
emediation technologies may be exploited to decontami-
nate the habitats contaminated with heavy metals.
However, the acquisition of knowledge at the molecular
level and its advancement will further give an insight into
understanding the anti-stress effect of 28-HBL in greater
detail and facilitate in developing suitable strategies for
plant protection against heavy metals stresses. Further
investigation on interactive effects of BRs with other
phytohormones can be focused to sort out the intricacies of
phytohormones signaling involved in various defence
mechanisms.
Acknowledgments Financial assistance from Department of Sci-
ence and Technology (DST), Ministry of Science & Technology,
Government of India, New Delhi, India is duly acknowledged.
References
Aebi H (1984) Catalase in vitro. Methods Enzymol 105:121–126
Allen SE, Grimshaw HM, Parkinson JA, Quarmby C, Roberts JD
(1976) Chemical analysis. In: Chapman SB (ed) Methods in plant
ecology. Blackwell Scientific Publications, London, pp 424–426
Arnon DI (1949) Copper enzymes in isolated chloroplasts. Polyphe-
nol oxidase in Beta vulgaris. Plant Physiol 24:1–15
Bajguz A (2000) Effect of brassinosteroids on nucleic acids and
protein content in cultured cells of Chlorella vulgaris. Plant
Physiol Biochem 38(3):209–215
Barbafieri M, Tassi E (2011) Brassinosteroids for phytoremediation
application. In: Hayat S, Ahmad A (eds) Brassinosteroids: a class
of plant hormone. Springer, Dordrecht, pp 403–438
Bates LS, Waldern RP, Teare ID (1973) Rapid determination of free
proline for water stress studies. Plant Soil 39:205–207
Bhardwaj R, Arora N, Sharma P, Arora HK (2007) Effects of
28-homobrassinolide on seedling growth, lipid peroxidation and
antioxidative enzyme activities under nickel stress in seedlings
of Zea mays L. Asian J Plant Sci 6(5):765–772
Bhattacharjee S (2005) Reactive oxygen species and oxidative burst:
roles in stress, senescence and signal transduction in plants. Curr
Sci 89(7):1113–1121
Bradford MM (1976) A rapid and sensitive method for the quantitation
of microgram quantities of protein utilizing the principle of
protein-dye binding. Anal Biochem 72(1–2):248–254
Carlberg I, Mannervik B (1975) Purification of the flavoenzyme
glutathione reductase from rat liver. J Biol Chem 250:5475–5480
Cerana R, Lado P, Anastasia M, Ciuffreda P, Allevi P (1984)
Regulating effects of brassinosteroids and of sterols on growth
and H? secretion in maize roots. J Plant Physiol 114:221–225
Chary NS, Kamala CT, Raj DS (2008) Assessing risk of heavy metals
from consuming food grown on sewage irrigated soils and food
chain transfer. Ecotoxicol Environ Saf 69(3):513–524
Chen N-C, Kanazawa S, Horiguchi T (2001) Effect of chromium on
some enzyme activities in the wheat rhizosphere. Soil Microorg
55(1):3–10
Clouse SD, Sasse JM (1998) Brassinosteroids: essential regulators of
plant growth and development. Annu Rev Plant Physiol Plant
Mol Biol 49:427–451
Dalton DA, Russell SA, Hanus FJ, Pascoe GA, Evans HJ (1986)
Enzymatic reactions of ascorbate and glutathione that prevent
peroxide damage in soybean root nodules. Proc Natl Acad Sci
USA 83:3811–3815
Dhaubhadel S, Krishna P (2008) Identification of differentially
expressed genes in brassinosteroid-treated Brassica napus seed-
lings. Plant Growth Regul 27:297–308
Eaton AD, Clesceri LS, Greenberg AE (1995) Coloriometric method-
chromium. In: Standard methods for the examination of water
and wastewater, 19th edn. American Public Health Association,
Washington, pp 3 59–3 60
Foyer CH, Lopez-Delgado H, Dat JF, Scott IM (1997) Hydrogen
peroxide and glutathione associated mechanisms of acclamatory
stress tolerance and signaling. Physiol Plant 100:241–254
Effect of 28-homobrassinolide on antioxidant defence system 873
123

Gendron JM, Wang ZY (2007) Multiple mechanisms modulate
brassinosteroid signaling. Curr Opin Plant Biol 10:436–441
Halliwell B, Gutteridge JMC (2007) Free radicals in biology and
medicine, 4th edn. Clarendon Press, Oxford
Haubrick LL, Assmann SM (2006) Brassinosteroids and plant
function: some clues, more puzzles. Plant Cell Environ
29:446–457
Hayat S, Ali B, Hassan SA, Ahmad A (2007) Brassinosteroids
enhanced antioxidants under cadmium stress in Brassica juncea.
Environ Exp Bot 60(1):33–41
Hodges DM, DeLong JM, Forney CF, Prange RK (1999) Improving
the thiobarbituric acid-reactive-substances assay for estimating
lipid peroxidation in plant tissues containing anthrocyanin and
other interfering compounds. Planta 207:604–611
Hossain MA, Nakano Y, Asada K (1984) Monodehydroascorbate
reductase in spinach chloroplasts and its participation in
regeneration of ascorbate for scavenging hydrogen peroxide.
Plant Cell Physiol 25(3):385–395
Kono Y (1978) Generation of superoxide radical during autooxidation
of hydroxylamine and an assay for superoxide dismutase. Arch
Biochem Biophys 186:189–195
Krishna P (2003) Brassinosteroids-mediated stress responses. J Plant
Growth Regul 22:289–297
Li J, Wen J, Lease KA, Doke JT, Tax FE, Walker JC (2002) BAK1, an
ArabidopsisLRR receptor-like protein kinase, interacts with BRI1
and modulates brassinosteroid signaling. Cell 110(2):213–222
Lindsey K, Pullen ML, Topping JF (2003) Importance of plant sterols
in pattern formation and hormone signaling. Trends Plant Sci
8(11):521–525
Liu DH, Jaing WS, Li MX (1993) Effect of chromium on root growth
and cell division of Allium cepa. Isr J Plant Sci 42:235–243
Liu X-M, Kim KE, Kim K-C, Nguyen XC, Han HJ, Jung MS, Kim
HS, Kim SH, Park HC, Yun D-J, Chung WS (2010) Cadmium
activates Arabidopsis MPK3 and MPK6 via accumulation of
reactive oxygen species. Phytochemistry 71:614–618
Ludwig AA, Romeis T, Jones JDG (2004) CDPK-mediated signalling
pathways: specificity and cross-talk. J Exp Bot 55(395):181–188
McCord JM (2000) The evolution of free radicals and oxidative
stress. Am J Med 108:652–659
McSteen P, Zhao Y (2008) Plant hormones and signaling: common
themes and new developments. Dev Cell 14:467–473
Mittler R (2002) Oxidative stress, antioxidants and stress tolerance.
Trends Plant Sci 7:405–410
Nakano Y, Asada K (1981) Hydrogen peroxide is scavenged by
ascorbate specific peroxidase in spinach chloroplasts. Plant Cell
Physiol 22(5):867–880
Noctor G, Foyer CH (1998) Ascorbate and glutathione: keeping
active oxygen under control. Annu Rev Plant Physiol Plant Mol
Biol 49:249–279
Panda SK, Choudhury S (2005) Chromium stress in plants. Braz J
Plant Physiol 17(1):95–102
Pandey V, Dixit V, Shyam R (2005) Antioxidative responses in
relation to growth of mustard (Brassica juncea c.v. Pusa
Jaikisan) plants exposed to hexavalent chromium. Chemosphere
61:40–47
Rout GR, Samantaray S, Das P (1997) Differential chromium
tolerance among eight mungbean cultivars grown in nutrient
culture. J Plant Nutr 20:473–483
Sanchez M, Revilla G, Zara I (1995) Changes in peroxidase activity
associated with cell walls during pine hypocotyl growth. Ann
Bot 75:415–419
Scandalios JG (2005) Oxidative stress: molecular perception and
transduction of signals triggering antioxidant gene defences.
Braz J Med Biol Res 38:995–1014
Schmidt J, Yokota T, Adam G, Takahashi N (1991) Castasterone and
brassinolide in Raphanus sativus seeds. Phytochemistry
30:364–365
Schmidt J, Yokota T, Spengler B, Adam G (1993) 28-Homoteaster-
one, a naturally occurring brassinosteroid from seeds of Raph-
anus sativus. Phytochemistry 34:391–392
Schutzendubel A, Polle A (2002) Plant responses to abiotic stresses:
heavy metal induced oxidative stress and protection by myco-
rrhization. J Exp Bot 53(372):1351–1365
Shanker AK, Cervantes C, Tavera-Loza H, Avudainayagam S (2005)
Chromium toxicity in plants. Environ Int 31:739–753
Sharma P, Bhardwaj R, Arora N, Arora HK (2007) Effect of
28-homobrassinolide on growth, zinc metal uptake and antiox-
idative enzyme activities in Brassica juncea L. seedlings. Braz J
Plant Physiol 19(3):203–210
Sharma I, Pati PK, Bhardwaj R (2010) Regulation of growth and
antioxidant enzyme activities by 28-homobrassinolide in seed-
lings of Raphanus sativus L. under cadmium stress. Indian J
Biochem Biophys 47(3):172–177
Sharma I, Pati PK, Bhardwaj R (2011) Effect of 24-epibrassinolide on
oxidative stress markers induced by nickel-ion in Raphanus
sativus L. Acta Physiol Plant. doi:10.1007/s11738-010-0709-1
Skorzynska-Polit E, Drazkiewicz M, Krupa Z (2010) Lipid peroxi-
dation and antioxidative response in Arabidopsis thaliana
exposed to cadmium and copper. Acta Physiol Plant 32:169–175
Sudo E, Itouga M, Yoshida-Hatanaka K, Ono Y, Sakakibara H (2008)
Gene expression and sensitivity in response to copper stress in
rice leaves. J Exp Bot 59(12):3465–3474
Wang Z-Y, He J-X (2004) Brassinosteroid signal transduction—
choices of signals and receptors. Trends Plant Sci 9(2):91–96
Wu GL, Cui J, Tao L, Yang H (2010) Fluroxypyr triggers oxidative
damage by producing superoxide and hydrogen peroxide in rice
(Oryza sativa). Ecotoxicology 19:124–132
Yu X-Z, Gu J-D, Huang S-Z (2007) Hexavalent chromium induced
stress and metabolic responses in hybrid willows. Ecotoxicology
16:299–309
Yusuf M, Fariduddin Q, Hayat S, Hasan SA, Ahmad A (2010)
Protective response of 28-homobrassinolide in cultivars of
Triticum aestivum with different levels of nickel. Arch Environ
Contam Toxicol. doi:10.1007/s00244-010-9535-0
Zhang M, Zhai Z, Tian X, Liusheng D, Li Z (2008) Brassinolide
alleviated the adverse effect of water deficits on photosynthesis
and the antioxidant of soybean (Glycine max L.). Plant Growth
Regul 56:257–264
874 I. Sharma et al.
123

1 23
Acta Physiologiae Plantarum
ISSN 0137-5881
Volume 33
Number 5
Acta Physiol Plant (2011)
33:1723-1735
DOI 10.1007/s11738-010-0709-1
Effect of 24-epibrassinolide on oxidativestress markers induced by nickel-ion inRaphanus sativus L.Indu Sharma, Pratap Kumar Pati & RenuBhardwaj

1 23
Your article is protected by copyright and
all rights are held exclusively by Franciszek
Górski Institute of Plant Physiology, Polish
Academy of Sciences, Kraków. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you
wish to self-archive your work, please use the
accepted author’s version for posting to your
own website or your institution’s repository.
You may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

ORIGINAL PAPER
Effect of 24-epibrassinolide on oxidative stress markers induced
by nickel-ion in Raphanus sativus L.
Indu Sharma • Pratap Kumar Pati •
Renu Bhardwaj
Received: 20 October 2010 / Revised: 23 December 2010 / Accepted: 30 December 2010 / Published online: 21 January 2011
Ó Franciszek Gorski Institute of Plant Physiology, Polish Academy of Sciences, Krakow 2011
Abstract The present study illustrates the effect of
24-epibrassinolide (24-EBL) on morphological and bio-
chemical parameters in radish (Raphanus sativus L.) seed-
lings grown under nickel (Ni) ion stress. The radish seeds
pre-soaked in different concentrations of 24-EBLwere sown
in petridishes containing various concentrations of heavy
metal (Ni).Observations were made on root/shoot length,
fresh biomass, activities of antioxidant enzymes (ascorbate
peroxidase, superoxide dismutase, catalase, monodehydro-
ascorbate reductase, dehydroascorbate reductase, guaiacol
peroxidase and glutathione reductase), lipid peroxidation,
proline and protein content in 7-day-old Ni-stressed radish
seedlings. Results indicate that seeds presoaked with
24-EBL reduced the impact of Ni-stress which was evident
by assessing the morphological parameters, protein content
and antioxidant enzyme activities. It was also observed that
24-EBL reduced the toxicity of heavy metal by influencing
proline and malondialdehyde (MDA) content. The present
study lays a foundation for understanding the role of 24-EBL
in heavy metal stress amelioration, particularly in food crop.
Analysis of behaviour of antioxidant enzymes will play a
critical role in understanding the stress networking, further
filling the knowledge gap on the subject.
Keywords Antioxidant enzymes � Radish � Nickel ion
stress � 24-epibrassinolide � Lipid peroxidation
Introduction
Brassinosteroids (BRs) are an emerging group of steroidal
phytohormones which are essential for plant growth and
development (Pinol and Simon 2009). Recently, several
BR-regulated genes associated with diverse physiological
responses, such as cell division and expansion, differenti-
ation, programmed cell death, stomatal development and
functions, homeostasis and gene expression have been
isolated by genome-wide microarray analysis (Divi and
Krishna 2009; Tanaka et al. 2009). In addition to their
growth regulatory activities, BRs have also been reported
to play pivotal potential for their implication in both stress-
protection and stress-amelioration (Krishna 2003; Bajguz
and Hayat 2009). Furthermore, BRs are tested for anti-
genotoxicity by employing Allium cepa chromosomal
aberration bioassay, to ensure their safe use in agricultural
practices (Sondhi et al. 2008). In Arabidopsis, Li et al.
(2007) isolated a gene HSD1 encoding a protein with
homology to animal 11-b-hydroxysteroid dehydrogenase
(HSD). Divi and Krishna (2009) observed that overex-
pression of AtHSD1 in Arabidopsis led to BR-responsive
gene expression and in Brassica napus enhanced stress
tolerance. Divi and Krishna (2009) suggested that crop
yields and stress tolerance in plants could be achieved by
manipulating the genes involved in the BRs biosynthetic
and signalling pathways.
Communicated by S. Lewak.
I. Sharma � R. Bhardwaj (&)
Department of Botanical and Environmental Sciences,
Guru Nanak Dev University, Amritsar 143005, Punjab, India
e-mail: [email protected]
I. Sharma
e-mail: [email protected]
P. K. Pati
Department of Biotechnology, Guru Nanak Dev University,
Amritsar 143005, Punjab, India
e-mail: [email protected]
123
Acta Physiol Plant (2011) 33:1723–1735
DOI 10.1007/s11738-010-0709-1
Author's personal copy

Heavy metal stress is one major stress faced by the
agricultural crops. Some metals like Fe, Se, Mn, Co, Zn,
Mo and Ni, are essential micronutrient, but when ‘‘certain
trace levels’’ exceed, they are highly toxic to plants (Eskew
et al. 1983; Hall and Williams 2003). Also, being persistent
in nature, these heavy metals get accumulated in soils and
plants. Nickel (Ni) is an essential micronutrient and func-
tions as an active centre of the enzyme urease required for
the hydrolysis of urea and nitrogen metabolism in higher
plants (Brown et al. 1987; Gerendas et al. 1999). But at
higher levels, Ni produces toxic symptoms like stunting
growth, leaf chlorosis, mitotic inhibition, vein necrosis, etc.
in plants (Seregin and Kozhevnikova 2006; Llamas et al.
2008). Anthropogenic activities, for example effluent dis-
posal, metal mining, smelting, electroplating, sewage
sludge and fertilizer application result in Ni pollution
(Chen et al. 2009). From contaminated soils, Ni is easily
absorbed by the plants and its excessive uptake leads to
altered plant growth, fruit quality and quantity (Chen et al.
2009). Also, Ni ion leads to the alteration of biochemical
parameters including accumulation of reactive oxygen
species (ROS) and enhancement of lipid peroxidation in
plant tissues (Gajewska and Skłodowska 2008). The
imbalance between ROS production and amelioration leads
to oxidative stress causing phytotoxicity (Sudo et al. 2008).
To overcome this stress, the plants possess enzymatic and
non-enzymatic antioxidant defence system (Mittler 2002;
Skorzynska-Polit et al. 2010). Also, the antioxidant defence
system of plants is regulated by certain phytohormones like
Abscisic acid, Jasmonates and BRs under stress (Bari and
Jones 2009). But, the role of BRs in this direction is yet to
be studied in Raphanus sativus plants under heavy metal
stress.
In view of the above as well as the wide occurrence and
economic importance of R. sativus (radish), the present
study was carried out to explore the possible role of BRs in
ameliorating specific stress. The effects of 24-EBL have
been focused on the present piece of work to study the
morphological parameters, activities of antioxidant
enzymes, lipid peroxidation, proline and protein content in
Raphanus sativus L. (Pusa Chetaki) seedlings under Nickel
(Ni) ion stress.
Materials and methods
Plant material and growth conditions
Seeds of Raphanus sativus L. (Pusa Chetaki) were pro-
cured from Department of Plant Breeding, Punjab Agri-
culture University, Ludhiana, India. Seeds were surface
sterilized with 0.4% sodium hypochlorite for 15 min fol-
lowed by repeated rinses in sterile distilled water. Surface-
sterilized seeds were given 8-h presoaking treatment in
different concentrations of 24-EBL (0, 10-11, 10-9 and
10-7 M). These pretreated seeds were germinated on
Whatman No. 1 filter paper lined autoclaved glass Petri
dishes, each containing various concentrations of Ni (0,
0.5, 1.0 and 1.5 mM). The Ni (II) ion stress was given in
the form of NiSO4�7H2O. The experiment was conducted
under controlled conditions of light (16 h photoperiod
under fluorescent white light with 175 lmol m-2 s-1
intensity), temperature (25 ± 5°C) and relative humidity
(80–90%). The experiment was repeated twice with five
replications (each containing 20 seedlings) for each
treatment.
Growth analysis
Seven-day-old seedlings were harvested and roots and
shoots were separated. Seedling growth in terms of root
and shoot length was recorded. Twenty seedlings per pet-
ridish were used for the determination of morphological
parameters (root/shoot length), fresh biomass and per-
centage germination. The seedlings were oven-dried at
80°C for 24 h to determine their dry weights.
Biochemical analysis
Lipid peroxidation
Lipid peroxidation was determined by measuring the con-
tent of Malondialdehyde (MDA), a secondary end product
of the oxidation of polyunsaturated fatty acids, by the
method of Hodges et al. (1999). One gramme of shoots was
homogenized in 5 ml of 80% ethanol and then centrifuged
at 12,0009g for 5 min. Experiment was conducted using
20.0% (w/v) TCA (trichloroacetic acid), i.e. (-) TBA
(thiobarbituric acid) solution and 0.65% (w/v) TBA in
20.0% (w/v) TCA, i.e. (?) TBA solution. One ml aliquot of
supernatant was added to two different test tubes, one
containing 1 ml (–) TBA solution and other test tube
containing (?) TBA solution. Samples were then mixed
vigorously, kept in water bath at 95°C for 30 min and then
cooled quickly on ice bath. Then, samples were centrifuged
at 12,0009g for 5 min. Absorbance of red adduct was
observed at 440, 532, and 600 nm and Malondialdehyde
equivalents/g fresh weight (nmol ml-1) were calculated as
described by Hodges et al. (1999).
Proline content
The free proline content was estimated spectrophotomet-
rically following the method of Bates et al. (1973). Fresh
1 g of cotyledonary leaves was homogenized in 3.5 ml of
3% sulphosalicylic acid and the homogenates were
1724 Acta Physiol Plant (2011) 33:1723–1735
123
personal copy

centrifuged at 12,0009g for 10 min. Then, 2 ml of super-
natant was reacted with 2 ml of acid ninhydrin and 2 ml of
glacial acetic acid in test tubes for 1 h at 100°C. The
reaction was terminated by putting the test tubes in ice
bath. The reaction mixture was extracted with 4 ml of
toluene and mixed vigorously by shaking for 15–20 s.
Then toluene layer was separated from aqueous phase and
warmed to room temperature. The absorbance of red col-
oured Proline-ninhydrin product was measured in toluene
layer at 520 nm. Proline concentration was calculated from
a standard curve using 0–500 lM concentrations of
L-proline.
Preparation of leaf extracts
Leaf extracts were prepared to estimate the activities of
antioxidant enzymes and the protein content by homoge-
nizing 2 g cotyledonary leaves of 7-day-old seedlings in
chilled 6 ml 50 mM phosphate buffer (pH 7.0), 1 mM
ethylenediaminetetraacetic acid (EDTA), 1 mM phe-
nylmethanesulfonylfluoride (PMSF), 0.5% (v/v) Triton
X-100 and 2% (w/v) polyvinylpyrrolidone (PVP-30) in a
pre-chilled mortar and pestle. For estimation of ascorbate
peroxidase and dehydroascorbate reductase 0.5 mM
ascorbate was added to the extraction buffer. In case of
monodehydroascorbate reductase activity, 1 g of leaves
was homogenized in 3 ml of 50 mM Tris–HCl buffer (pH-
7.6) containing 2.5 mM Ascorbic acid. The homogenates
were centrifuged at 12,0009g for 20 min at 4°C. The
supernatant was further used for biochemical analysis.
Protein quantification
Total protein content of different samples of Ni (0, 0.5, 1.0
and 1.5 mM) and 24-EBL (0, 10-11, 10-9 and 10-7 M)
alone or in combinations, was quantified by following the
method of Bradford (1976) using bovine serum albumin as
a standard.
Ascorbate peroxidase assay
The ascorbate peroxidase (APOX, EC 1.11.1.11) activity
was determined spectrophotometrically as described by
Nakano and Asada (1981). The 3.0 ml reaction mixture
contained 50 mM Potassium phosphate buffer (pH 7.0),
0.5 mM ascorbate, 1.0 mM H2O2 and 100 ll enzyme
extract. The H2O2-dependent oxidation of ascorbate was
followed by monitoring the decrease in absorbance at
290 nm using the extinction coefficient 2.8 mM-1 cm-1.
The reaction was carried out for 3 min at 25°C. One unit of
APOX activity is defined as the amount of enzyme that can
oxidize 1 lmol of ascorbate per minute.
Catalase assay
Catalase (CAT, EC 1.11.1.6) activity was assayed by
measuring the initial rate of H2O2 disappearance using the
method of Aebi (1984). The 3.0 ml reaction mixture con-
tained 50 mM Potassium phosphate buffer (pH 7.0),
15 mM H2O2 and 100 ll enzyme extract The decrease in
hydrogen peroxide was followed as decline in optical den-
sity at 240 nm for 30 s at 25°C. The enzyme activity was
calculated using an extinction coefficient 39.4 mM-1 cm-1
for H2O2. One unit of enzyme activity is defined as the
decomposition of 1 mmol H2O2 per minute/g fresh
biomass.
Dehydroascorbate reductase assay
Dehydroascorbate reductase (DHAR, EC 1.8.5.1) activity
was measured following the method given by Dalton et al.
(1986). The 3.0 ml reaction mixture contained 50 mM
Potassium phosphate buffer (pH 7.0), 0.2 mM dehydro-
ascorbate, 0.1 mM EDTA, 2.5 mM reduced glutathione
and 100 ll enzyme extract. DHAR activity was measured
by following the increase in absorbance at 265 nm due to
ascorbate formation at 265 nm using extinction coefficient
of 14 mM-1 cm-1.
Glutathione reductase assay
Glutathione reductase (GR, EC 1.6.4.2) activity was
determined by using the method of Carlberg and Manner-
vik (1975). Three ml of reaction mixture contained 50 mM
Potassium phosphate buffer (pH 7.6), 1 mM oxidized
glutathione (GSSG), 0.5 mM EDTA, 0.1 mM reduced ni-
cotinamideadenine dinucleotidephosphate (NADPH) and
100 ll enzymes extract. The reaction was initiated by
addition of 0.1 mM NADPH at 25°C. The GR activity was
determined by the oxidation of NADPH at 340 nm with
extinction coefficient of 6.22 mM-1 cm-1.
Guaiacol peroxidase assay
Guaiacol peroxidase (POD, EC 1.11.1.7) activity was
assayed using the method of Sanchez et al. (1995) with
some modifications. The 3.0 ml reaction mixture contained
50 mM Potassium phosphate buffer (pH 7.0), 20 mM
guaiacol, 12.3 mM H2O2 and 100 ll enzyme extract.
Activity of POD was determined by measuring the absor-
bance at 436 nm and using an extinction coefficient of
26.6 mM-1 cm-1. One unit of POD activity represents the
amount of enzyme catalysing the oxidation of 1 lmol of
guaiacol in 1 min.
Acta Physiol Plant (2011) 33:1723–1735 1725
123
personal copy
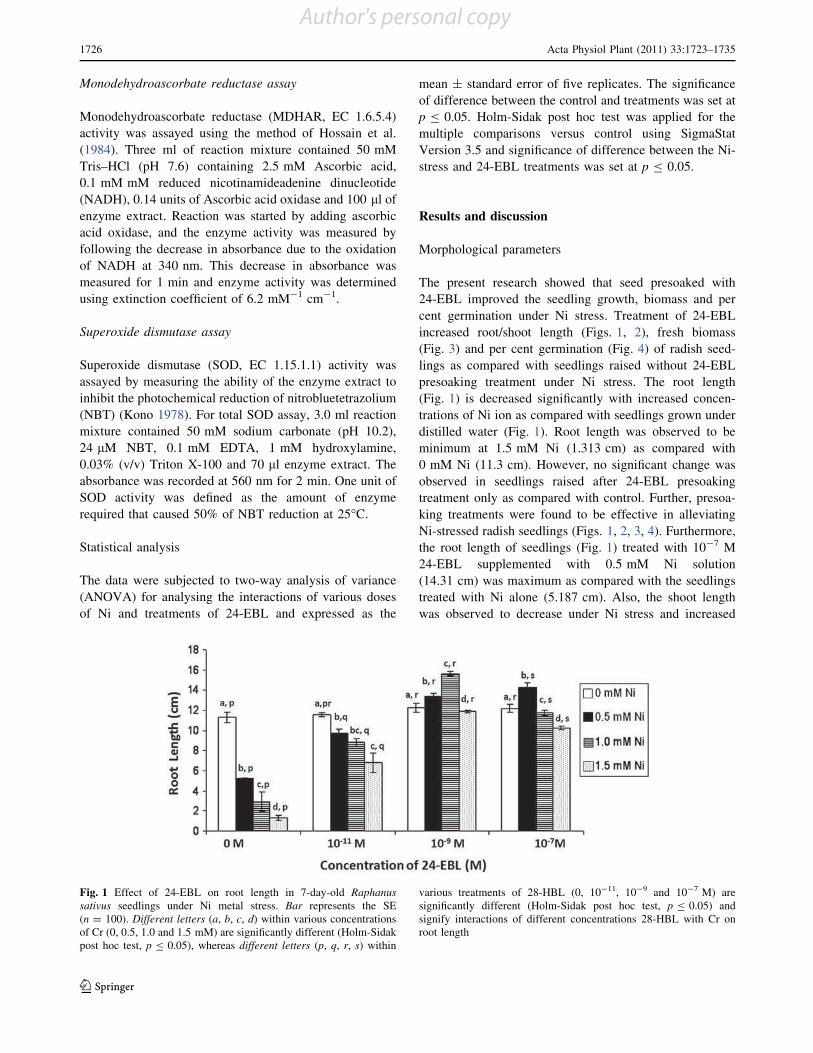
Monodehydroascorbate reductase assay
Monodehydroascorbate reductase (MDHAR, EC 1.6.5.4)
activity was assayed using the method of Hossain et al.
(1984). Three ml of reaction mixture contained 50 mM
Tris–HCl (pH 7.6) containing 2.5 mM Ascorbic acid,
0.1 mM mM reduced nicotinamideadenine dinucleotide
(NADH), 0.14 units of Ascorbic acid oxidase and 100 ll of
enzyme extract. Reaction was started by adding ascorbic
acid oxidase, and the enzyme activity was measured by
following the decrease in absorbance due to the oxidation
of NADH at 340 nm. This decrease in absorbance was
measured for 1 min and enzyme activity was determined
using extinction coefficient of 6.2 mM-1 cm-1.
Superoxide dismutase assay
Superoxide dismutase (SOD, EC 1.15.1.1) activity was
assayed by measuring the ability of the enzyme extract to
inhibit the photochemical reduction of nitrobluetetrazolium
(NBT) (Kono 1978). For total SOD assay, 3.0 ml reaction
mixture contained 50 mM sodium carbonate (pH 10.2),
24 lM NBT, 0.1 mM EDTA, 1 mM hydroxylamine,
0.03% (v/v) Triton X-100 and 70 ll enzyme extract. The
absorbance was recorded at 560 nm for 2 min. One unit of
SOD activity was defined as the amount of enzyme
required that caused 50% of NBT reduction at 25°C.
Statistical analysis
The data were subjected to two-way analysis of variance
(ANOVA) for analysing the interactions of various doses
of Ni and treatments of 24-EBL and expressed as the
mean ± standard error of five replicates. The significance
of difference between the control and treatments was set at
p B 0.05. Holm-Sidak post hoc test was applied for the
multiple comparisons versus control using SigmaStat
Version 3.5 and significance of difference between the Ni-
stress and 24-EBL treatments was set at p B 0.05.
Results and discussion
Morphological parameters
The present research showed that seed presoaked with
24-EBL improved the seedling growth, biomass and per
cent germination under Ni stress. Treatment of 24-EBL
increased root/shoot length (Figs. 1, 2), fresh biomass
(Fig. 3) and per cent germination (Fig. 4) of radish seed-
lings as compared with seedlings raised without 24-EBL
presoaking treatment under Ni stress. The root length
(Fig. 1) is decreased significantly with increased concen-
trations of Ni ion as compared with seedlings grown under
distilled water (Fig. 1). Root length was observed to be
minimum at 1.5 mM Ni (1.313 cm) as compared with
0 mM Ni (11.3 cm). However, no significant change was
observed in seedlings raised after 24-EBL presoaking
treatment only as compared with control. Further, presoa-
king treatments were found to be effective in alleviating
Ni-stressed radish seedlings (Figs. 1, 2, 3, 4). Furthermore,
the root length of seedlings (Fig. 1) treated with 10-7 M
24-EBL supplemented with 0.5 mM Ni solution
(14.31 cm) was maximum as compared with the seedlings
treated with Ni alone (5.187 cm). Also, the shoot length
was observed to decrease under Ni stress and increased
Fig. 1 Effect of 24-EBL on root length in 7-day-old Raphanus
sativus seedlings under Ni metal stress. Bar represents the SE
(n = 100). Different letters (a, b, c, d) within various concentrations
of Cr (0, 0.5, 1.0 and 1.5 mM) are significantly different (Holm-Sidak
post hoc test, p B 0.05), whereas different letters (p, q, r, s) within
various treatments of 28-HBL (0, 10-11, 10-9 and 10-7 M) are
significantly different (Holm-Sidak post hoc test, p B 0.05) and
signify interactions of different concentrations 28-HBL with Cr on
root length
1726 Acta Physiol Plant (2011) 33:1723–1735
123
Author's personal copy
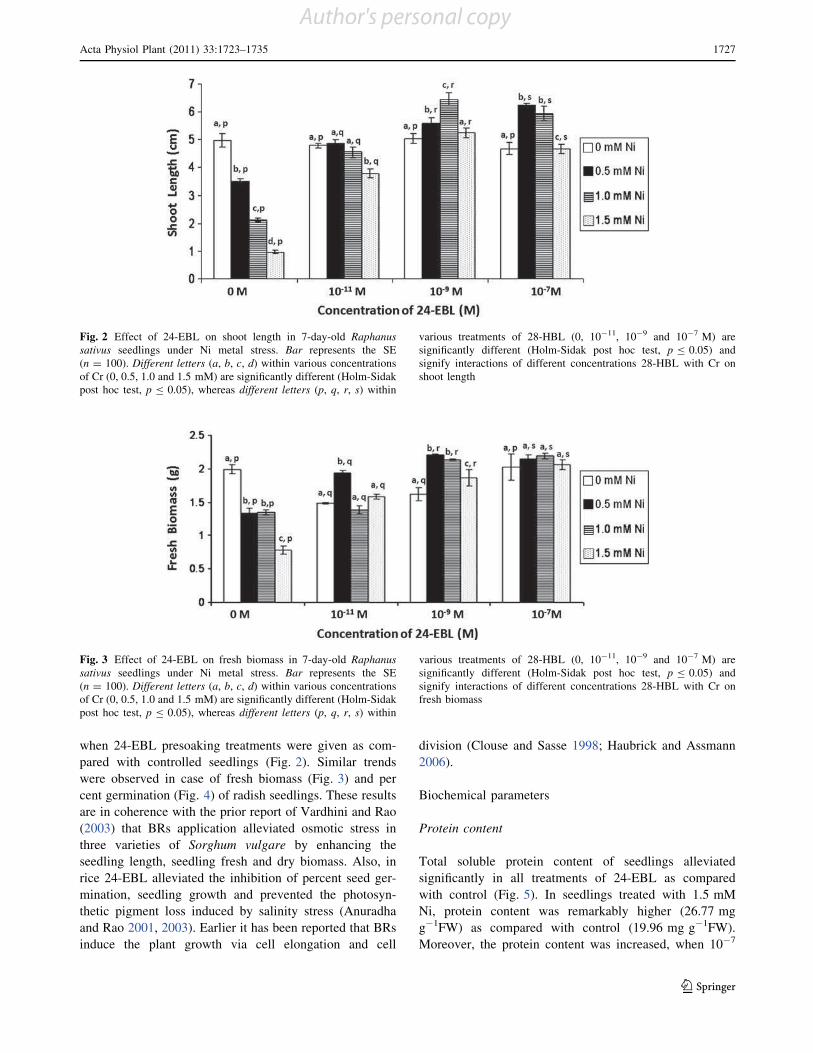
when 24-EBL presoaking treatments were given as com-
pared with controlled seedlings (Fig. 2). Similar trends
were observed in case of fresh biomass (Fig. 3) and per
cent germination (Fig. 4) of radish seedlings. These results
are in coherence with the prior report of Vardhini and Rao
(2003) that BRs application alleviated osmotic stress in
three varieties of Sorghum vulgare by enhancing the
seedling length, seedling fresh and dry biomass. Also, in
rice 24-EBL alleviated the inhibition of percent seed ger-
mination, seedling growth and prevented the photosyn-
thetic pigment loss induced by salinity stress (Anuradha
and Rao 2001, 2003). Earlier it has been reported that BRs
induce the plant growth via cell elongation and cell
division (Clouse and Sasse 1998; Haubrick and Assmann
2006).
Biochemical parameters
Protein content
Total soluble protein content of seedlings alleviated
significantly in all treatments of 24-EBL as compared
with control (Fig. 5). In seedlings treated with 1.5 mM
Ni, protein content was remarkably higher (26.77 mg
g-1FW) as compared with control (19.96 mg g-1FW).
Moreover, the protein content was increased, when 10-7
Fig. 2 Effect of 24-EBL on shoot length in 7-day-old Raphanus
sativus seedlings under Ni metal stress. Bar represents the SE
(n = 100). Different letters (a, b, c, d) within various concentrations
of Cr (0, 0.5, 1.0 and 1.5 mM) are significantly different (Holm-Sidak
post hoc test, p B 0.05), whereas different letters (p, q, r, s) within
various treatments of 28-HBL (0, 10-11, 10-9 and 10-7 M) are
significantly different (Holm-Sidak post hoc test, p B 0.05) and
signify interactions of different concentrations 28-HBL with Cr on
shoot length
Fig. 3 Effect of 24-EBL on fresh biomass in 7-day-old Raphanus
sativus seedlings under Ni metal stress. Bar represents the SE
(n = 100). Different letters (a, b, c, d) within various concentrations
of Cr (0, 0.5, 1.0 and 1.5 mM) are significantly different (Holm-Sidak
post hoc test, p B 0.05), whereas different letters (p, q, r, s) within
various treatments of 28-HBL (0, 10-11, 10-9 and 10-7 M) are
significantly different (Holm-Sidak post hoc test, p B 0.05) and
signify interactions of different concentrations 28-HBL with Cr on
fresh biomass
Acta Physiol Plant (2011) 33:1723–1735 1727
123
Author's personal copy
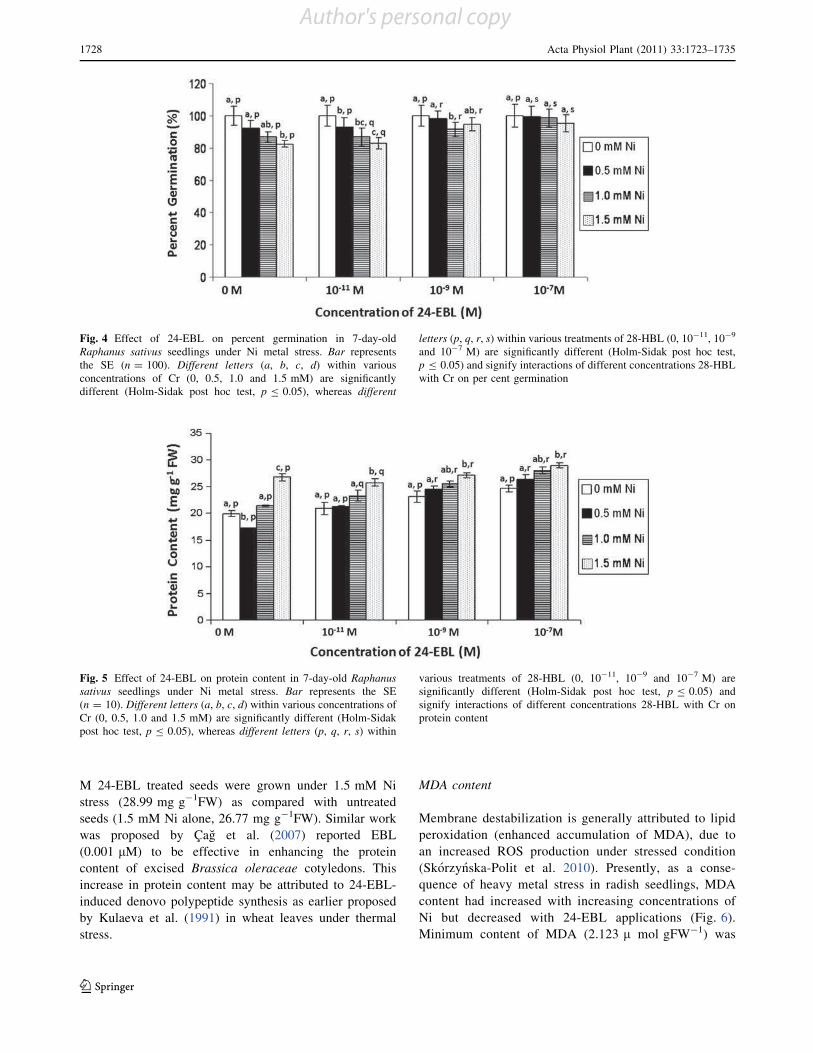
M 24-EBL treated seeds were grown under 1.5 mM Ni
stress (28.99 mg g-1FW) as compared with untreated
seeds (1.5 mM Ni alone, 26.77 mg g-1FW). Similar work
was proposed by Cag et al. (2007) reported EBL
(0.001 lM) to be effective in enhancing the protein
content of excised Brassica oleraceae cotyledons. This
increase in protein content may be attributed to 24-EBL-
induced denovo polypeptide synthesis as earlier proposed
by Kulaeva et al. (1991) in wheat leaves under thermal
stress.
MDA content
Membrane destabilization is generally attributed to lipid
peroxidation (enhanced accumulation of MDA), due to
an increased ROS production under stressed condition
(Skorzynska-Polit et al. 2010). Presently, as a conse-
quence of heavy metal stress in radish seedlings, MDA
content had increased with increasing concentrations of
Ni but decreased with 24-EBL applications (Fig. 6).
Minimum content of MDA (2.123 l mol gFW-1) was
Fig. 4 Effect of 24-EBL on percent germination in 7-day-old
Raphanus sativus seedlings under Ni metal stress. Bar represents
the SE (n = 100). Different letters (a, b, c, d) within various
concentrations of Cr (0, 0.5, 1.0 and 1.5 mM) are significantly
different (Holm-Sidak post hoc test, p B 0.05), whereas different
letters (p, q, r, s) within various treatments of 28-HBL (0, 10-11, 10-9
and 10-7 M) are significantly different (Holm-Sidak post hoc test,
p B 0.05) and signify interactions of different concentrations 28-HBL
with Cr on per cent germination
Fig. 5 Effect of 24-EBL on protein content in 7-day-old Raphanus
sativus seedlings under Ni metal stress. Bar represents the SE
(n = 10). Different letters (a, b, c, d) within various concentrations of
Cr (0, 0.5, 1.0 and 1.5 mM) are significantly different (Holm-Sidak
post hoc test, p B 0.05), whereas different letters (p, q, r, s) within
various treatments of 28-HBL (0, 10-11, 10-9 and 10-7 M) are
significantly different (Holm-Sidak post hoc test, p B 0.05) and
signify interactions of different concentrations 28-HBL with Cr on
protein content
1728 Acta Physiol Plant (2011) 33:1723–1735
123
Author's personal copy
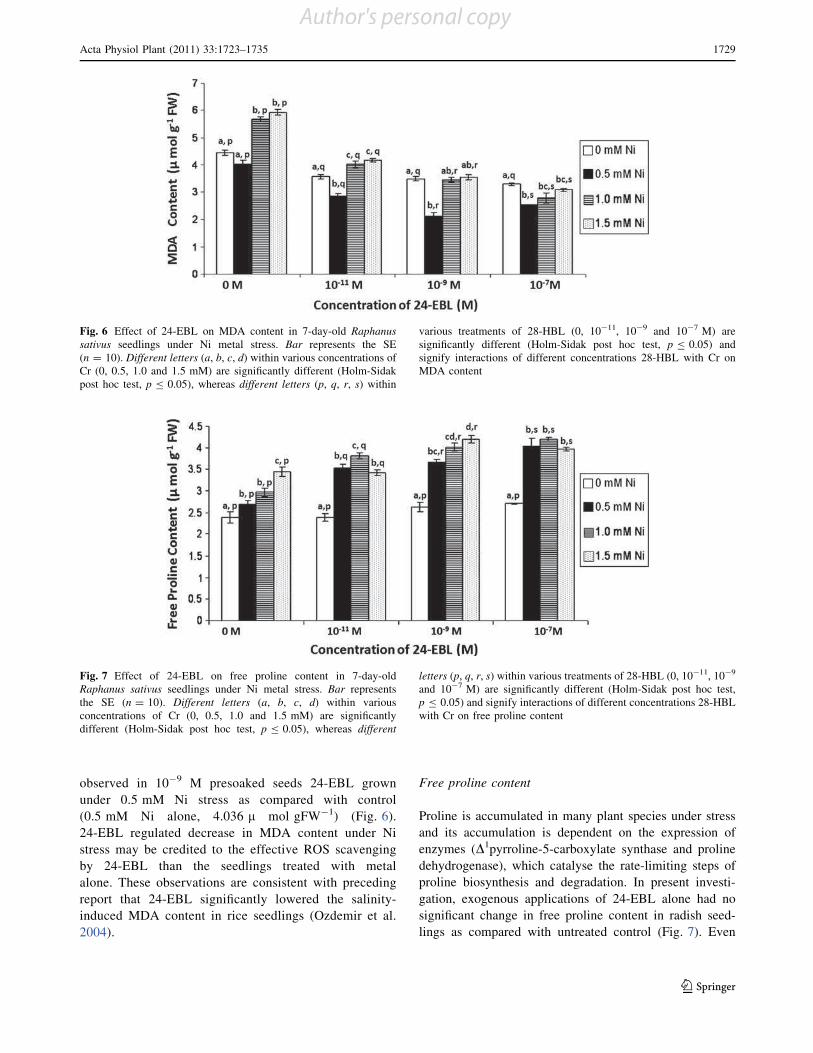
observed in 10-9 M presoaked seeds 24-EBL grown
under 0.5 mM Ni stress as compared with control
(0.5 mM Ni alone, 4.036 l mol gFW-1) (Fig. 6).
24-EBL regulated decrease in MDA content under Ni
stress may be credited to the effective ROS scavenging
by 24-EBL than the seedlings treated with metal
alone. These observations are consistent with preceding
report that 24-EBL significantly lowered the salinity-
induced MDA content in rice seedlings (Ozdemir et al.
2004).
Free proline content
Proline is accumulated in many plant species under stress
and its accumulation is dependent on the expression of
enzymes (D1pyrroline-5-carboxylate synthase and proline
dehydrogenase), which catalyse the rate-limiting steps of
proline biosynthesis and degradation. In present investi-
gation, exogenous applications of 24-EBL alone had no
significant change in free proline content in radish seed-
lings as compared with untreated control (Fig. 7). Even
Fig. 6 Effect of 24-EBL on MDA content in 7-day-old Raphanus
sativus seedlings under Ni metal stress. Bar represents the SE
(n = 10). Different letters (a, b, c, d) within various concentrations of
Cr (0, 0.5, 1.0 and 1.5 mM) are significantly different (Holm-Sidak
post hoc test, p B 0.05), whereas different letters (p, q, r, s) within
various treatments of 28-HBL (0, 10-11, 10-9 and 10-7 M) are
significantly different (Holm-Sidak post hoc test, p B 0.05) and
signify interactions of different concentrations 28-HBL with Cr on
MDA content
Fig. 7 Effect of 24-EBL on free proline content in 7-day-old
Raphanus sativus seedlings under Ni metal stress. Bar represents
the SE (n = 10). Different letters (a, b, c, d) within various
concentrations of Cr (0, 0.5, 1.0 and 1.5 mM) are significantly
different (Holm-Sidak post hoc test, p B 0.05), whereas different
letters (p, q, r, s) within various treatments of 28-HBL (0, 10-11, 10-9
and 10-7 M) are significantly different (Holm-Sidak post hoc test,
p B 0.05) and signify interactions of different concentrations 28-HBL
with Cr on free proline content
Acta Physiol Plant (2011) 33:1723–1735 1729
123
Author's personal copy
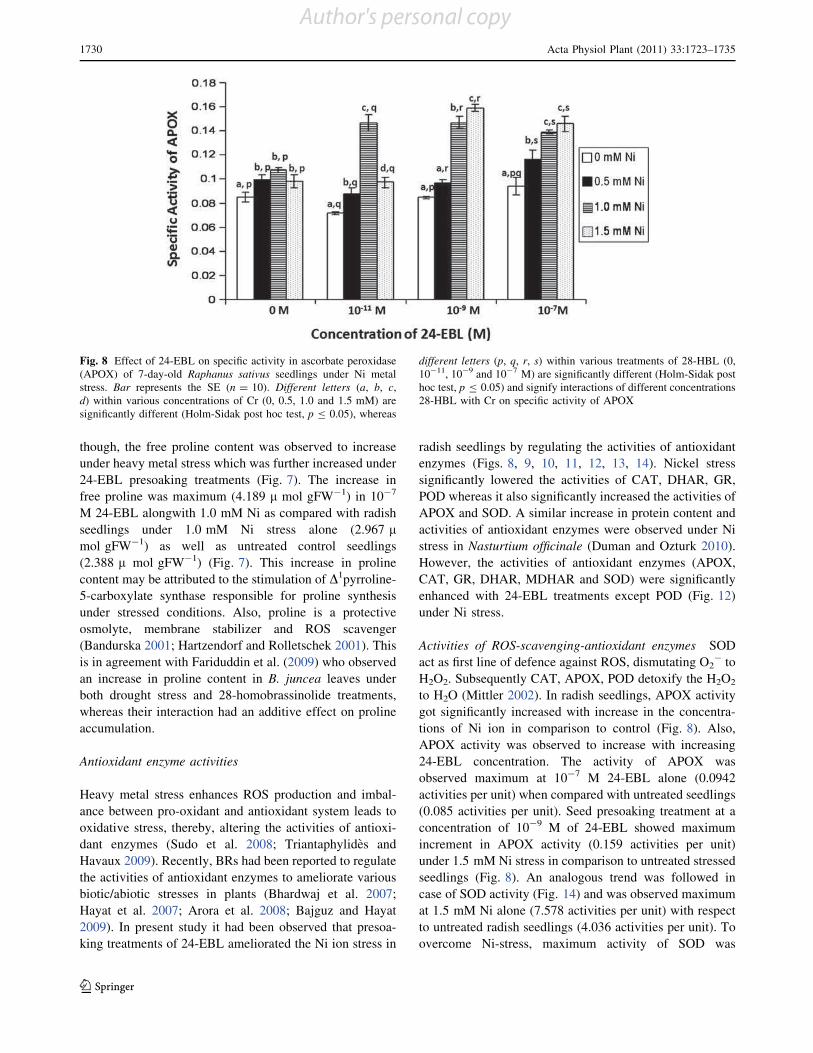
though, the free proline content was observed to increase
under heavy metal stress which was further increased under
24-EBL presoaking treatments (Fig. 7). The increase in
free proline was maximum (4.189 l mol gFW-1) in 10-7
M 24-EBL alongwith 1.0 mM Ni as compared with radish
seedlings under 1.0 mM Ni stress alone (2.967 l
mol gFW-1) as well as untreated control seedlings
(2.388 l mol gFW-1) (Fig. 7). This increase in proline
content may be attributed to the stimulation of D1pyrroline-
5-carboxylate synthase responsible for proline synthesis
under stressed conditions. Also, proline is a protective
osmolyte, membrane stabilizer and ROS scavenger
(Bandurska 2001; Hartzendorf and Rolletschek 2001). This
is in agreement with Fariduddin et al. (2009) who observed
an increase in proline content in B. juncea leaves under
both drought stress and 28-homobrassinolide treatments,
whereas their interaction had an additive effect on proline
accumulation.
Antioxidant enzyme activities
Heavy metal stress enhances ROS production and imbal-
ance between pro-oxidant and antioxidant system leads to
oxidative stress, thereby, altering the activities of antioxi-
dant enzymes (Sudo et al. 2008; Triantaphylides and
Havaux 2009). Recently, BRs had been reported to regulate
the activities of antioxidant enzymes to ameliorate various
biotic/abiotic stresses in plants (Bhardwaj et al. 2007;
Hayat et al. 2007; Arora et al. 2008; Bajguz and Hayat
2009). In present study it had been observed that presoa-
king treatments of 24-EBL ameliorated the Ni ion stress in
radish seedlings by regulating the activities of antioxidant
enzymes (Figs. 8, 9, 10, 11, 12, 13, 14). Nickel stress
significantly lowered the activities of CAT, DHAR, GR,
POD whereas it also significantly increased the activities of
APOX and SOD. A similar increase in protein content and
activities of antioxidant enzymes were observed under Ni
stress in Nasturtium officinale (Duman and Ozturk 2010).
However, the activities of antioxidant enzymes (APOX,
CAT, GR, DHAR, MDHAR and SOD) were significantly
enhanced with 24-EBL treatments except POD (Fig. 12)
under Ni stress.
Activities of ROS-scavenging-antioxidant enzymes SOD
act as first line of defence against ROS, dismutating O2- to
H2O2. Subsequently CAT, APOX, POD detoxify the H2O2
to H2O (Mittler 2002). In radish seedlings, APOX activity
got significantly increased with increase in the concentra-
tions of Ni ion in comparison to control (Fig. 8). Also,
APOX activity was observed to increase with increasing
24-EBL concentration. The activity of APOX was
observed maximum at 10-7 M 24-EBL alone (0.0942
activities per unit) when compared with untreated seedlings
(0.085 activities per unit). Seed presoaking treatment at a
concentration of 10-9 M of 24-EBL showed maximum
increment in APOX activity (0.159 activities per unit)
under 1.5 mM Ni stress in comparison to untreated stressed
seedlings (Fig. 8). An analogous trend was followed in
case of SOD activity (Fig. 14) and was observed maximum
at 1.5 mM Ni alone (7.578 activities per unit) with respect
to untreated radish seedlings (4.036 activities per unit). To
overcome Ni-stress, maximum activity of SOD was
Fig. 8 Effect of 24-EBL on specific activity in ascorbate peroxidase
(APOX) of 7-day-old Raphanus sativus seedlings under Ni metal
stress. Bar represents the SE (n = 10). Different letters (a, b, c,
d) within various concentrations of Cr (0, 0.5, 1.0 and 1.5 mM) are
significantly different (Holm-Sidak post hoc test, p B 0.05), whereas
different letters (p, q, r, s) within various treatments of 28-HBL (0,
10-11, 10-9 and 10-7 M) are significantly different (Holm-Sidak post
hoc test, p B 0.05) and signify interactions of different concentrations
28-HBL with Cr on specific activity of APOX
1730 Acta Physiol Plant (2011) 33:1723–1735
123
Author's personal copy
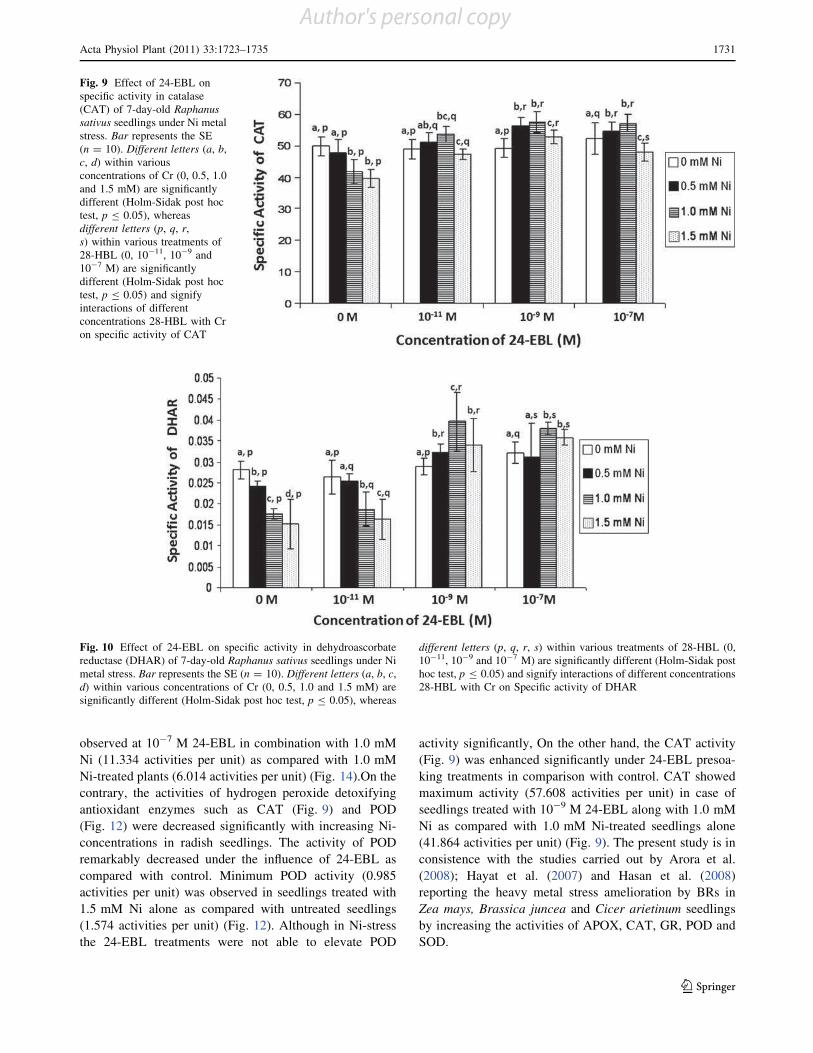
observed at 10-7 M 24-EBL in combination with 1.0 mM
Ni (11.334 activities per unit) as compared with 1.0 mM
Ni-treated plants (6.014 activities per unit) (Fig. 14).On the
contrary, the activities of hydrogen peroxide detoxifying
antioxidant enzymes such as CAT (Fig. 9) and POD
(Fig. 12) were decreased significantly with increasing Ni-
concentrations in radish seedlings. The activity of POD
remarkably decreased under the influence of 24-EBL as
compared with control. Minimum POD activity (0.985
activities per unit) was observed in seedlings treated with
1.5 mM Ni alone as compared with untreated seedlings
(1.574 activities per unit) (Fig. 12). Although in Ni-stress
the 24-EBL treatments were not able to elevate POD
activity significantly, On the other hand, the CAT activity
(Fig. 9) was enhanced significantly under 24-EBL presoa-
king treatments in comparison with control. CAT showed
maximum activity (57.608 activities per unit) in case of
seedlings treated with 10-9 M 24-EBL along with 1.0 mM
Ni as compared with 1.0 mM Ni-treated seedlings alone
(41.864 activities per unit) (Fig. 9). The present study is in
consistence with the studies carried out by Arora et al.
(2008); Hayat et al. (2007) and Hasan et al. (2008)
reporting the heavy metal stress amelioration by BRs in
Zea mays, Brassica juncea and Cicer arietinum seedlings
by increasing the activities of APOX, CAT, GR, POD and
SOD.
Fig. 9 Effect of 24-EBL on
specific activity in catalase
(CAT) of 7-day-old Raphanus
sativus seedlings under Ni metal
stress. Bar represents the SE
(n = 10). Different letters (a, b,
c, d) within various
concentrations of Cr (0, 0.5, 1.0
and 1.5 mM) are significantly
different (Holm-Sidak post hoc
test, p B 0.05), whereas
different letters (p, q, r,
s) within various treatments of
28-HBL (0, 10-11, 10-9 and
10-7 M) are significantly
different (Holm-Sidak post hoc
test, p B 0.05) and signify
interactions of different
concentrations 28-HBL with Cr
on specific activity of CAT
Fig. 10 Effect of 24-EBL on specific activity in dehydroascorbate
reductase (DHAR) of 7-day-old Raphanus sativus seedlings under Ni
metal stress. Bar represents the SE (n = 10). Different letters (a, b, c,
d) within various concentrations of Cr (0, 0.5, 1.0 and 1.5 mM) are
significantly different (Holm-Sidak post hoc test, p B 0.05), whereas
different letters (p, q, r, s) within various treatments of 28-HBL (0,
10-11, 10-9 and 10-7 M) are significantly different (Holm-Sidak post
hoc test, p B 0.05) and signify interactions of different concentrations
28-HBL with Cr on Specific activity of DHAR
Acta Physiol Plant (2011) 33:1723–1735 1731
123
Author's personal copy
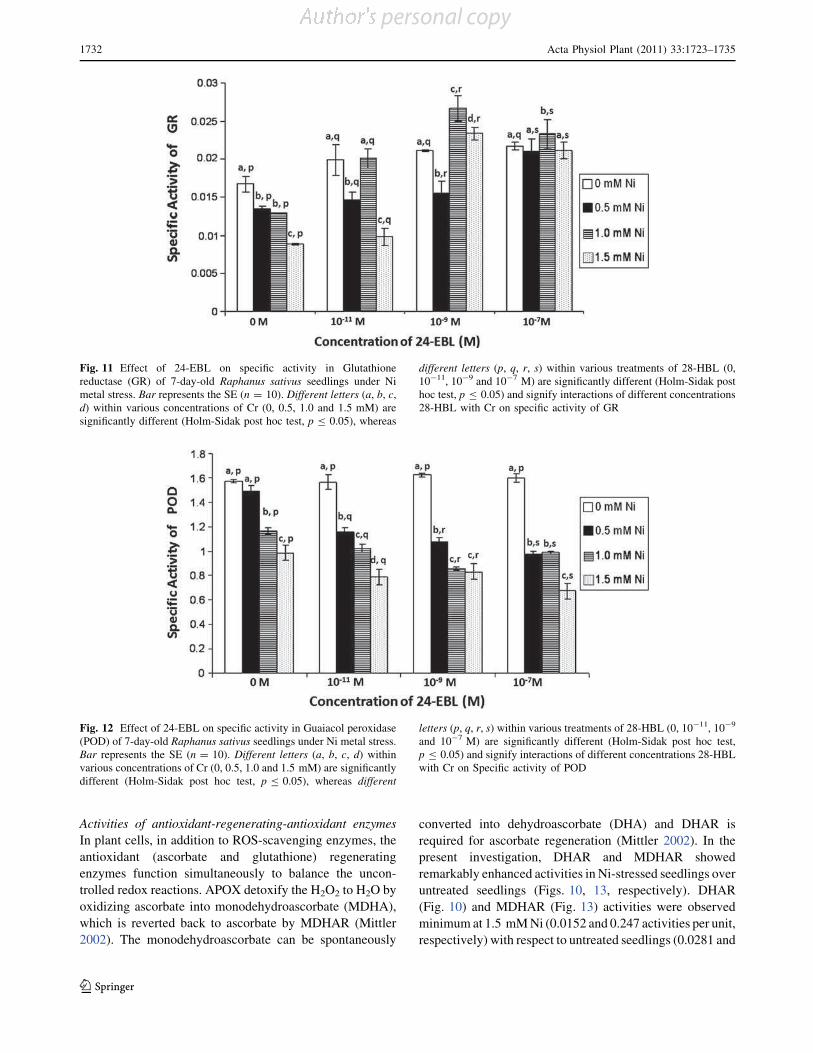
Activities of antioxidant-regenerating-antioxidant enzymes
In plant cells, in addition to ROS-scavenging enzymes, the
antioxidant (ascorbate and glutathione) regenerating
enzymes function simultaneously to balance the uncon-
trolled redox reactions. APOX detoxify the H2O2 to H2O by
oxidizing ascorbate into monodehydroascorbate (MDHA),
which is reverted back to ascorbate by MDHAR (Mittler
2002). The monodehydroascorbate can be spontaneously
converted into dehydroascorbate (DHA) and DHAR is
required for ascorbate regeneration (Mittler 2002). In the
present investigation, DHAR and MDHAR showed
remarkably enhanced activities in Ni-stressed seedlings over
untreated seedlings (Figs. 10, 13, respectively). DHAR
(Fig. 10) and MDHAR (Fig. 13) activities were observed
minimum at 1.5 mMNi (0.0152 and 0.247 activities per unit,
respectively) with respect to untreated seedlings (0.0281 and
Fig. 11 Effect of 24-EBL on specific activity in Glutathione
reductase (GR) of 7-day-old Raphanus sativus seedlings under Ni
metal stress. Bar represents the SE (n = 10). Different letters (a, b, c,
d) within various concentrations of Cr (0, 0.5, 1.0 and 1.5 mM) are
significantly different (Holm-Sidak post hoc test, p B 0.05), whereas
different letters (p, q, r, s) within various treatments of 28-HBL (0,
10-11, 10-9 and 10-7 M) are significantly different (Holm-Sidak post
hoc test, p B 0.05) and signify interactions of different concentrations
28-HBL with Cr on specific activity of GR
Fig. 12 Effect of 24-EBL on specific activity in Guaiacol peroxidase
(POD) of 7-day-old Raphanus sativus seedlings under Ni metal stress.
Bar represents the SE (n = 10). Different letters (a, b, c, d) within
various concentrations of Cr (0, 0.5, 1.0 and 1.5 mM) are significantly
different (Holm-Sidak post hoc test, p B 0.05), whereas different
letters (p, q, r, s) within various treatments of 28-HBL (0, 10-11, 10-9
and 10-7 M) are significantly different (Holm-Sidak post hoc test,
p B 0.05) and signify interactions of different concentrations 28-HBL
with Cr on Specific activity of POD
1732 Acta Physiol Plant (2011) 33:1723–1735
123
personal copy
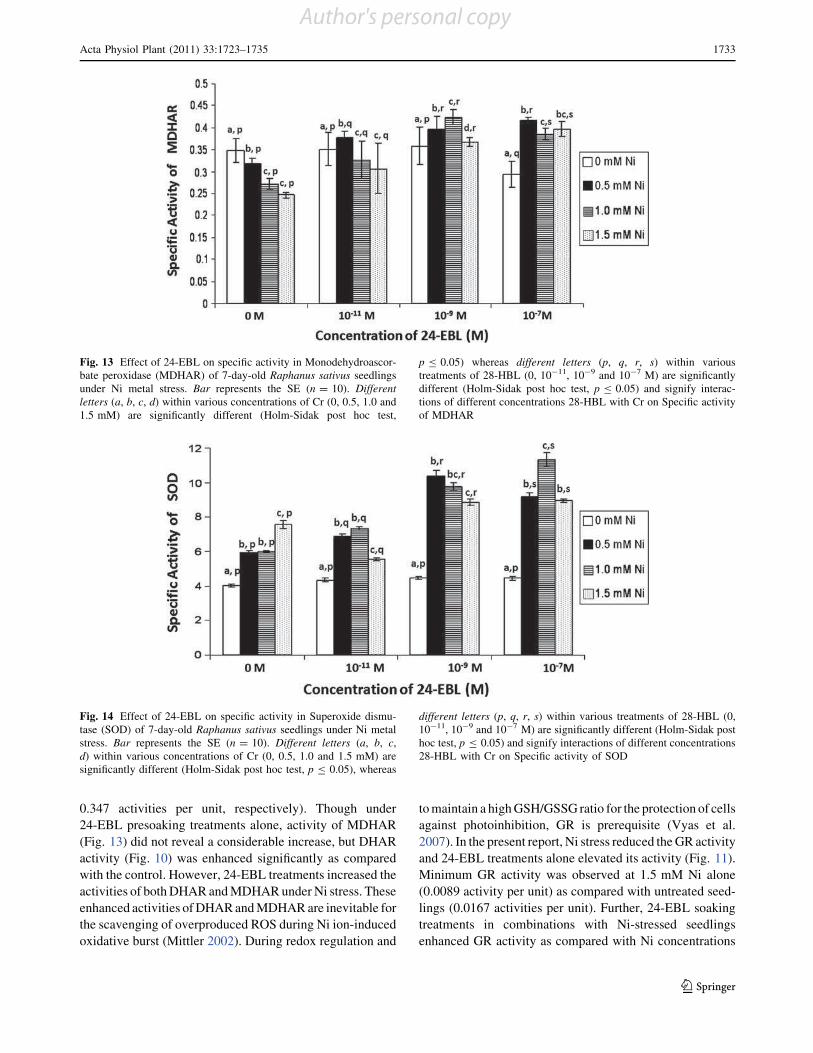
0.347 activities per unit, respectively). Though under
24-EBL presoaking treatments alone, activity of MDHAR
(Fig. 13) did not reveal a considerable increase, but DHAR
activity (Fig. 10) was enhanced significantly as compared
with the control. However, 24-EBL treatments increased the
activities of bothDHARandMDHARunder Ni stress. These
enhanced activities ofDHARandMDHARare inevitable for
the scavenging of overproduced ROS during Ni ion-induced
oxidative burst (Mittler 2002). During redox regulation and
tomaintain a highGSH/GSSG ratio for the protection of cells
against photoinhibition, GR is prerequisite (Vyas et al.
2007). In the present report, Ni stress reduced theGR activity
and 24-EBL treatments alone elevated its activity (Fig. 11).
Minimum GR activity was observed at 1.5 mM Ni alone
(0.0089 activity per unit) as compared with untreated seed-
lings (0.0167 activities per unit). Further, 24-EBL soaking
treatments in combinations with Ni-stressed seedlings
enhanced GR activity as compared with Ni concentrations
Fig. 13 Effect of 24-EBL on specific activity in Monodehydroascor-
bate peroxidase (MDHAR) of 7-day-old Raphanus sativus seedlings
under Ni metal stress. Bar represents the SE (n = 10). Different
letters (a, b, c, d) within various concentrations of Cr (0, 0.5, 1.0 and
1.5 mM) are significantly different (Holm-Sidak post hoc test,
p B 0.05) whereas different letters (p, q, r, s) within various
treatments of 28-HBL (0, 10-11, 10-9 and 10-7 M) are significantly
different (Holm-Sidak post hoc test, p B 0.05) and signify interac-
tions of different concentrations 28-HBL with Cr on Specific activity
of MDHAR
Fig. 14 Effect of 24-EBL on specific activity in Superoxide dismu-
tase (SOD) of 7-day-old Raphanus sativus seedlings under Ni metal
stress. Bar represents the SE (n = 10). Different letters (a, b, c,
d) within various concentrations of Cr (0, 0.5, 1.0 and 1.5 mM) are
significantly different (Holm-Sidak post hoc test, p B 0.05), whereas
different letters (p, q, r, s) within various treatments of 28-HBL (0,
10-11, 10-9 and 10-7 M) are significantly different (Holm-Sidak post
hoc test, p B 0.05) and signify interactions of different concentrations
28-HBL with Cr on Specific activity of SOD
Acta Physiol Plant (2011) 33:1723–1735 1733
123
Author's personal copy

alone. Furthermore, 10-7 M 24-EBL treatments in combi-
nation with 1.0 mM Ni (0.0267 activities per unit) showed
maximum GR activity as compared with 1.0 mMNi-treated
plants (0.0129 activities per unit) in radish seedlings. Hence,
24-EBL pretreatments might help in maintaining the novel
pool of glutathione in the reduced state by enhancing GR
activity under metal-stressed conditions.
Recently, Dhaubhadel and Krishna (2008) identified six
differentially expressed genes in 24-EBL-treated heat-
stressed Brassica napus seedlings. BRs have been specu-
lated to act via receptor/ligand complex that binds to nuclear
or cytoplasmic sites to regulate the expression of specific
stress-related genes. Stress-responsive genes activated by
BRs may code for the PCs, organic acids, osmolytes and
stress-protective proteins (LEA proteins and HSPs) (Gen-
dron and Wang 2007; McSteen and Zhao 2008). Also,
Bajguz (2002) reported that BRs acted as phytochelatin
stimulator in Chlorella vulgaris to ameliorate lead metal
toxicity. Thus, the present study specifies the effect of
24-EBL by analysing the behaviour of antioxidant enzymes
in heavy metal stress amelioration in a food crop (radish).
Further, detailed studies in validating the synergistic role of
BRs as well as other phytohormones under stress will play a
critical role in understanding the stress networking in plants.
Conclusions
The seed presoaking treatments of 24-EBL improved radish
seedling growth, germination, antioxidant enzyme activi-
ties, proline and MDA content under both stressed and non-
stressed conditions. Under Ni-stress, 24-EBL enhanced
antioxidant enzyme activities more significantly, suggesting
the amelioration of metal-induced ROS. Thus, 24-EBL
mediated Ni-stress amelioration might be mediated through
the modification of antioxidant defence system of plant.
Acknowledgments Financial assistance from Department of Sci-
ence and Technology (DST), Ministry of Science & Technology,
Government of India, New Delhi, India, is duly acknowledged.
References
Aebi M (1984) Catalase in vitro. Methods Enzymol 105:121–126
Anuradha S, Rao SSR (2001) Effect of brassinosteroids on salinity
stress induced inhibition of germination and seedling growth of
rice (Oryza sativa L.). Plant Growth Regul 33:151–153
Anuradha S, Rao SSR (2003) Application of brassinosteroids to rice
seeds (Oryza sativa L.) reduced the impact of salt stress on
growth, prevented photosynthetic pigment loss and increased
nitrate reductase activity. Plant Growth Regul 40:29–32
Arora N, Bhardwaj R, Sharma P, Arora HK (2008) Effects of
28-homobrassinolide on growth, lipid peroxidation and antiox-
idative enzyme activities in seedlings of Zea mays L. under
salinity stress. Acta Physiol Plant 30:833–839
Bajguz A (2002) Brassinosteroids and lead as stimulators of
phytochelatins synthesis in Chlorella vulgaris cells. J Plant
Physiol 159:321–324
Bajguz A, Hayat S (2009) Effects of brassinosteroids on the plant
responses to environmental stresses. Plant Physiol Biochem
47:1–8
Bandurska H (2001) Does proline accumulated in leaves of water
deficit stressed barley plants confine cell membrane injuries? II.
Proline accumulation during hardening and its involvement in
reducing membrane injuries in leaves subjected to severe
osmotic stress. Acta Physiol Plant 23:483–490
Bari R, Jones JDG (2009) Role of plant hormones in plant defence
responses. Plant Mol Biol 69:473–488
Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free
proline for water-stress studies. Plant Soil 39:205–207
Bhardwaj R, Arora N, Sharma P, Arora HK (2007) Effects of
28-homobrassinolide on seedling growth, lipid peroxidation and
antioxidative enzyme activities under nickel stress in seedlings
of Zea mays L. Asian J Plant Sci 6:765–772
Bradford MM (1976) A rapid and sensitive method for the
quantification of microgram quantities of protein utilizing the
principle of protein-dye binding. Anal Biochem 72:248–254
Brown PH, Welch RM, Cary EE (1987) Nickel: a micronutrient
essential for higher plants. Plant Physiol 85:801–803
Cag S, Goren-Saglam N, Cingil-Baris C, Kaplan E (2007) The effect
of different concentration of epibrassinolide on chlorophyll,
protein and anthocyanini content and peroxidase activity in
excised red cabbage (Brassica oleracea L.) cotyledons. Biotech-
nol Biotechnol Equip 21:422–425
Carlberg I, Mannervik B (1975) Purification of the flavoenzyme
glutathione reductase from rat liver. J Biol Chem 250:5475–5480
ChenC,HuangD,Liu J (2009) Functions and toxicity of nickel in plants:
recent advances and future prospects. Clean 37(4–5):304–313
Clouse S, Sasse J (1998) Brassinosteroids: essentials regulators of
plant growth and development. Annu Rev Plant Physiol Plant
Mol Biol 49:427–451
Dalton DA, Russell SA, Hanus FJ, Pascoe GA, Evans HJ (1986)
Enzymatic reactions of ascorbate and glutathione that prevent
peroxide damage in soybean root nodules. Proc Natl Acad Sci
USA 83:3811–3815
Dhaubhadel S, Krishna P (2008) Identification of differentially
expressed genes in brassinosteroid-treated Brassica napus seed-
lings. Plant Growth Regul 27:297–308
Divi UK, Krishna P (2009) Brassinosteroid: a biotechnological target
for enhancing crop yield and stress tolerance. New Biotechnol.
doi:10.1016/j.nbt.2009.07.006
Duman F, Ozturk F (2010) Nickel accumulation and its effect on
biomass, protein content and antioxidative enzymes in roots and
leaves of watercress (Nasturtium officinale R. Br.). J Environ Sci
(China) 22(4):526–532
Eskew DL, Welch RM, Cary EE (1983) Nickel: an essential
micronutrient for legumes and possibly all higher plants. Science
222:621–623
Fariduddin Q, Yusuf M, Hayat S, Ahmad A (2009) Effect of
28-homobrassinolide on antioxidant capacity and photosynthesis
in Brassica juncea plants exposed to different levels of copper.
Environ Exp Bot 6:418–424
Gajewska E, Skłodowska M (2008) Differential biochemical
responses of wheat shoots and roots to nickel stress: antioxida-
tive reactions and proline accumulation. Plant Growth Regul
54:179–188
Gendron JM, Wang ZY (2007) Multiple mechanisms modulate
brassinosteroid signaling. Curr Opin Plant Biol 10:436–441
Gerendas J, Polacco JC, Freyermuth SK, Sattelmacher B (1999)
Significance of nickel for plant growth and metabolism. J Plant
Nutr Soil Sci 162:241–256
1734 Acta Physiol Plant (2011) 33:1723–1735
123
personal copy

Hall JL, Williams LE (2003) Transition metal transporters in plants.
J Exp Bot 54:2601–2613
Hartzendorf T, Rolletschek H (2001) Effect of NaCl-salinity on
amino acid and carbohydrate contents of Phragmites australis.
Aquat Bot 69:195–208
Hasan SA, Hayat S, Ali B, Ahmad A (2008) 28-Homobrassinolide
protects chickpea (Cicer arietinum) from cadmium toxicity by
stimulating antioxidants. Environ Pollut 151:60–66
Haubrick LL, Assmann SM (2006) Brassinosteriods and plant
function: some clues, more puzzles. Plant Cell Environ
29:446–457
Hayat S, Ali B, Hasan SA, Ahmad A (2007) Brassinosteriod enhance
the level of antioxidants under cadmium stress in Brassica
juncea. Environ Exp Bot 60:33–41
Hodges DM, DeLong JM, Forney CF, Prange RK (1999) Improving
the thiobarbituric acid reactive-substances assay for estimating
lipid peroxidation in plant tissues containing anthrocyanin and
other interfering compounds. Planta 207:604–611
Hossain MA, Nakano Y, Asada K (1984) Monodehydroascorbate
reductase in spinach chloroplasts and its participation in
regeneration of ascorbate for scavenging hydrogen peroxide.
Plant Cell Physiol 25(3):385–395
Kono Y (1978) Generation of superoxide radical during autoxidation
of hydroxylamine and an assay for superoxide dismutase. Arch
Biochem Biophys 186:189–195
Krishna P (2003) Brassinosteoids-mediated stress responses. J Plant
Growth Regul 22:289–297
Kulaeva ON, Burkhanova EA, Fedina AB et al (1991) Effect of
brassinosterods on protein synthesis and plant-cell ultrastructure
under stress conditions. In: Culter HG, Yokota T, Adam G (eds)
Brassinosteroids––chemistry, bioactivity and applications. ACS
Symp. Ser., Washington, pp 141–157
Li F, Asami T, Wu X, Tsang EWT, Cutler AJ (2007) A putative
hydroxysteroid dehydrogenase involved in regulating plant
growth and development. Plant Physiol 145:87–97
Llamas AC, Ullrich I, Sanz A (2008) Ni2? toxicity in rice: effect on
membrane functionality and plant water content. Plant Physiol
Biochem 46:905–910
McSteen P, Zhao Y (2008) Plant hormones and signaling: common
themes and new developments. Dev Cell 14:467–473
Mittler R (2002) Oxidative stress, antioxidants and stress tolerance.
Trends Plant Sci 7:405–410
Nakano Y, Asada K (1981) Hydrogen peroxide is scavenged by
ascorbate specific peroxidase in spinach chloroplasts. Plant Cell
Physiol 22:867–880
Ozdemir F, Bor M, Demiral T, Turkan I (2004) Effects of
24-epibrassinolide on seed germination, seedling growth, lipid
peroxidation, proline content and antioxidative system of rice
(Oryza sativa L.) under salinity stress. Plant Growth Regul
42:203–211
Pinol R, Simon E (2009) Effect of 24-Epibrassinolide on chlorophyll
fluorescence and photosynthetic CO2 assimilation in Vicia faba
plants treated with the photosynthesis-inhibiting herbicide
terbutryn. J Plant Growth Regul. doi:10.1007/s00344-008-
9077-0
Sanchez M, Revilla G, Zara I (1995) Changes in peroxidase activity
associated with cell walls during pine hypocotyl growth. Ann
Bot 75:415–419
Seregin IV, Kozhevnikova AD (2006) Physiological role of nickel
and its toxic effects on higher plants. Russ J Plant Physiol
53:257–277
Skorzynska-Polit E, Drazkiewicz M, Krupa Z (2010) Lipid peroxi-
dation and antioxidative response in Arabidopsis thaliana
exposed to cadmium and copper. Acta Physiol Plant 32:169–175
Sondhi N, Bhardwaj R, Kaur S, Kumar N, Singh B (2008) Isolation of
24-epibrassinolide from leaves of Aegle marmelos and evalua-
tion of its antigenotoxicity employing Allium cepa chro-
mos€omal aberration assay. Plant Growth Regul 54:217–224
Sudo E, Itouga M, Yoshida-Hatanaka K, Ono Y, Sakakibara H (2008)
Gene expression and sensitivity in response to copper stress in
rice leaves. J Exp Bot 59(12):3465–3474
Tanaka A, Nakagawa H, Tomita C et al (2009) BRASSINOSTEROID
UPREGULATED 1, Encoding a helix-loop-helix protein, is a
novel gene involved in brassinosteroid signaling and controls
bending of the lamina joint in rice. Plant Physiol 151:669–680
Triantaphylides C, Havaux M (2009) Singlet oxygen in plants:
production, detoxification and signaling. Trends Plant Sci
14(4):219–228
Vardhini BV, Rao SSR (2003) Acceleration of ripening of tomato
pericarp discs by brassinosteroids. Phytochemistry 61:843–847
Vyas D, Kumar S, Ahuja PS (2007) Tea (Camellia sinensis) clones
with shorter periods of winter dormancy exhibit lower accumu-
lation of reactive oxygen species. Tree Physiol 27:1253–1259
Acta Physiol Plant (2011) 33:1723–1735 1735
123
personal copy


![How Auxin May Contribute to the Regulation of Plant ...acid (SA), jasmonates, brassinosteroids (BR) and strigolactones [5-7]. Regarding their function, auxins, BR, CK, GA and strigolactones](https://img.pdfslide.us/doc/110x75/5f8b1647dd500c0935619b8c/how-auxin-may-contribute-to-the-regulation-of-plant-acid-sa-jasmonates-brassinosteroids.jpg)