Embed Size (px)
Citation preview

An approach to correct the effects of phytoplankton verticalnonuniform distribution on remote sensing reflectance ofcyanobacterial bloom waters
Kun Xue, Yuchao Zhang, Ronghua Ma,* Hongtao DuanKey Laboratory of Watershed Geographic Sciences, Nanjing Institute of Geography and Limnology, Chinese Academy ofSciences, Nanjing 210008, China
Abstract
Cyanobacterial blooms occur frequently in eutrophic lakes and their potentially harmful effects affected
the security of drinking water and food sources, biodiversity, and economic activities, and attracted the
attention of general public worldwide. Cyanobacteria could move vertically in the water column by regulat-
ing their buoyancy, which leads to the assumption of homogeneous water invalid. Ecolight, based on radia-
tive transfer theory, was applied to examine the effects of vertical nonuniform of chlorophyll a
concentrations (Chl a(z)) on remote sensing reflectance spectrum (Rrs(k)) of optically complex inland waters.
Simulations for nonuniform water consisting of three Chl a(z) profile classes, including Gaussian, exponen-
tial, and power, were compared with simulations for a reference homogeneous water whose Chl a was identi-
cal to average value of the nonuniform case. The near-surface aggregation of phytoplankton are shown to
have significant influence on Rrs(k) and Chl a inversion algorithms. Variations of DRrs(k) (relative difference
of Rrs(k) between inhomogeneous and homogeneous waters with same average Chl a concentration) mainly
depended on the Chl a(z) structure parameters and wavelength. A correction scheme was developed based on
the relationships between DRrs(k) and Chl a(z) structure parameters. With knowledge of Chl a(z) profile
parameters, Rrs(k) of inhomogeneous waters can be corrected to the Rrs(k) of uniform waters with same aver-
age Chl a across the water column. Examples of field data from Lake Chaohu illustrated the effects of phyto-
plankton variation on the near infrared-to-red ratio of Rrs and the Rrs correction performance.
Eutrophication of lakes has been a major environmental
and social-economic problem across the world. The
increased intensity of cyanobacterial blooms has affected the
security of drinking water and food sources, biodiversity,
and economic activities. The optical properties of these
waters are determined by the amount of optically active con-
stituents (OACs) like phytoplankton (usually measured as
concentration of chlorophyll a, Chl a), suspended particulate
matter (SPM), and colored dissolved organic matter (CDOM).
In optically complex waters, commonly regard as Case II
waters (Morel and Prieur 1977), large part of the suspended
particulate inorganic matter (SPIM) and CDOM originates
from the inflow rivers and bottom resuspension. As a result,
concentrations of these substances vary a broad range (more
than one order of magnitude), and may not covary with Chl
a as in Case I waters (Gholamalifard et al. 2013).
Remote sensing technology has often been used to estimate
the optical properties, such as the absorption and backscatter-
ing coefficients (Loisel and Stramski 2000; Lee et al. 2002),
and determine key OACs, such as Chl a (Duan et al. 2010;
Song et al. 2013b), SPM (He et al. 2013), and phycocyanin
(Duan et al. 2012; Song et al. 2013a). It is also utilized to
detect the extent and frequency of cyanobacterial blooms
(Kutser et al. 2006; Odermatt et al. 2012; Bresciani et al. 2014;
Zhang et al. 2015). These applications of water color remote
sensing usually assume that the OACs and their correspond-
ing water inherent optical properties (IOPs) are vertically uni-
form. However, OACs of natural waters often show significant
vertical variations, which could result in vertical nonuniform
of IOPs (Andr�e 1992; Nanu and Robertson 1993; Forget et al.
2001). The observed remote sensing reflectance (Rrs(k)) con-
tains information about the optical properties within the pen-
etration depth, at which the downwelling irradiance (Ed) is
reduced to 1=e of its surface value (Gordon and Mccluney
1975). Vertical inhomogeneity of phytoplankton creates a
challenge to the usage of chlorophyll a concentration inver-
sion algorithm and determination of phytoplankton biomass
in the water column (Silulwane et al. 2010).
The vertical profiles of phytoplankton are mainly gov-
erned by meteorological, biological, and hydrological*Correspondence: [email protected]
1
LIMNOLOGYand
OCEANOGRAPHY: METHODS Limnol. Oceanogr.: Methods 00, 2017, 00–00VC 2017 Association for the Sciences of Limnology and Oceanography
doi: 10.1002/lom3.10158

conditions (Cao et al. 2006). Gaussian or shifted Gaussian
models (Mill�an-N�u~nez et al. 1997; Odermatt et al. 2012;
Bresciani et al. 2014) have been used to represent the vertical
profile of Chl a(z) in marine environments. The maximum
Chl a of Gaussian model was regarded as a common feature
of Chl a vertical distribution in these waters (Cullen and
Eppley 1981). In coastal waters or inland lakes, discharge
from rivers or terrestrial runoff may cause the vertical vari-
ability in the upper layer of the water column (Yang et al.
2013). In cyanobacterial bloom waters, cyanobacteria, such
as Microcystis aeruginosa, could regulate their buoyancy, and
move vertically in the water column due to changes of tem-
perature (Tsujimura et al. 2000), light intensity (Walsby and
Booker 1980), and nutrients (Klemer et al. 1982; Kromkamp
et al. 1989). There were few in situ datasets of vertical pro-
files of Chl a concentration in cyanobacterial bloom lakes
(Kutser et al. 2008). Four Chl a vertical profile classes, includ-
ing vertically uniform, Gaussian, exponential, and power
class, were measured in Lake Chaohu, a shallow eutrophic
lake of China (Xue et al. 2015). The Gaussian distribution of
phytoplankton measured in Lake Chaohu had maximum
Chl a value at water surface. In appropriate situations, cya-
nobacteria may float to water surface and form dense sub-
surface accumulations, which could produce extreme high
concentrations of Chl a (Wynne et al. 2010). These situa-
tions usually belong to exponential or power type of Chl a
vertical profiles, whose Chl a concentration decreased dra-
matically with water depth (Xue et al. 2015). In addition,
the typical shape of the chlorophyll a profile is often
assumed to be stable in a given region or season (Hidalgo-
Gonzalez and Alvarez-Borrego 2001). However, some studies
indicated that the vertical profile class of phytoplankton of
lakes change rapidly (D’Alimonte et al. 2014).
Several previous studies have focused on the vertical dis-
tribution of phytoplankton (Morel and Berthon 1989;
Mill�an-N�u~nez et al. 1997) and its effects on remote sensing
reflectance (Tsujimura et al. 2000; Stramska and Stramski
2005; Kutser et al. 2008). It has been demonstrated that the
vertical variations of phytoplankton have significant influ-
ence on the remote sensing reflectance in oceanic waters
(Stramska and Stramski 2005) and cyanobacterial bloom
waters (Kutser et al. 2008). In the Chl a inversion algorithms,
the relationships between Chl a and band ratio or combina-
tion of Rrs(k) were also affected by the vertical profiles of
phytoplankton (Stramska and Stramski 2005). The reality is
that the present empirical Chl a inversion algorithms are
affected to an unknown degree by the nonuniform of Chl
a(z) profiles or IOPs(z) of the water column.
Gordon and Clark (1980) proposed a hypothesis that the
Rrs(k) of vertical inhomogeneous waters is equal to the Rrs(k)
of homogeneous waters in which the Chl a is a weighted
average of the actual Chl a(z) profile within the penetration
depth. This hypothesis was accepted and used widely in
research on vertically nonuniform waters (Mill�an-N�u~nez
et al. 1997; Zaneveld et al. 2005; Bresciani et al. 2014). How-
ever, a common view concerning a reliable model for
weighted mean pigment concentration detectable by remote
sensors has not yet been achieved even for Case 1 waters
(Sokoletsky and Yacobi 2011).
In this study, we explored the effects of vertical profiles of
phytoplankton on remote sensing reflectance Rrs(k) in cyano-
bacterial bloom waters, Lake Chaohu in China. This lake is
characterized by frequent occurrence of algal blooms, and its
SPIM and CDOM are optically important, and do not covary
with Chl a. Radiative transfer simulations for inhomoge-
neous Chl a(z) profiles of three classes, including Gaussian,
exponential, and power distribution, were analyzed com-
pared with reference simulations of a homogeneous water
whose Chl a was the average value of the Chl a(z) profiles.
We aim to: (1) examine the effects of nonuniform Chl a(z)
profiles on spectral remote sensing reflectance Rrs(k) and Chl
a inversion algorithm, using dataset of radiative transfer sim-
ulations; (2) build the relationships between DRrs(k) and
structure parameters of Chl a(z) profiles; (3) convert Rrs(k) of
Chl a(z) inhomogeneous waters to corrected Rrs(k) of uni-
form waters with same average Chl a in the water column.
This is an attempt to estimate to what extent the inhomoge-
neous Chl a(z) profile might affect the remote sensing reflec-
tance and performance of the Chl a inversion algorithms
based on the vertical uniform assumption.
Data and method
Field and laboratory measurements
Lake Chaohu is the 5th largest freshwater lake in China
with a mean depth of 3.0 m and an area of 770 km2 (31825’–
31843’ N, 117817’–117851’ E, Fig. 1) (Chen et al. 2013). The
diversity of phytoplankton, including Chlorophyta, Bacillario-
phyta, Cyanophyta, Cryptophyta, and Euglenuphyta, varies sea-
sonally in Lake Chaohu. Cyanophyta has the highest average
annual density (> 90%) in comparison with other phyto-
plankton species (Yu 2010). Three field surveys were carried
out in 2013 (May 28, July 19–24, and October 10–12) to
measure concentrations of OACs, absorption coefficients,
backscattering coefficients, and remote sensing reflectance.
In each station, water samples at different depths (surface,
0.1 m, 0.2 m, 0.4 m, 0.7 m, 1.0 m, 1.5 m, 2.0 m, and 3.0 m)
were collected using an ad hoc vertical water collection
device (Xue et al. 2015).
The water samples were filtered with Whatman GF/C
glass-fibre filters (pore size of 1.2 lm), and 90% acetone
extraction was used to extract pigments. Shimadzu (Kyoto,
Japan) UV-2600 spectrophotometer was used to measure
absorbance at 630 nm, 645 nm, 663 nm, and 750 nm, and
then Chl a concentration was calculated (Ma et al. 2006).
For SPM concentrations, Whatman GF/F glass fiber filters
(pore size of 0.7 lm) were pre-combusted at 4508C for 6 h
and pre-weighted. The filters were used to filter water
Xue et al. Phytoplankton vertical nonuniform distribution
2

samples, and then were dried at 1058C for 4–6 h. SPIM (Table
1) was derived gravimetrically by burning organic matter
from the filters at 4508C for 6 h (Duan et al. 2012). The
absorption coefficients of the total particulate matter, phyto-
plankton pigments, and non-algae particulate (NAP) were
determined using the quantitative filter technique (Mitchell
1990) with GF/F glass fiber filters and a Shimadzu UV2600
spectrophotometer from 350 nm to 800 nm with 1 nm inter-
val. The baseline was corrected using a blank filter wetted
with filtered water (Ma et al. 2006). The absorbance spectra
were corrected for path length amplification (Cleveland and
Weidemann 1993) and background by subtracting the aver-
age absorbance at 750 nm from the entire spectra (Ma et al.
2006; Zhang et al. 2007). CDOM absorption (ag(k)) was
determined from filtered water (Millipore filter with 0.22 lm
pore size) using Shimadzu UV2600 spectrophotometer with
Milli-Q water as the reference and ranged from 280 nm to
700 nm with 1 nm interval. Absorbance at each wavelength
was baseline corrected by subtracting the absorbance at
700 nm (Bricaud et al. 1981; Ma et al. 2006; Zhou et al.
2015).
A spectral backscattering sensor, Hydroscat-6 (Washing-
ton, USA), provided backscattering measurements at wave-
lengths of 420 nm, 442 nm, 470 nm, 510 nm, 590 nm, and
700 nm, under the guidance of the manual from Hydro-
Optics, Biology & Instrumentation Laboratories (HOBI Labs
Inc 2010). All Hydroscat-6 data were examined to leave out
exceptional data and the mean of the remaining measure-
ments was calculated, and have been sigma corrected (cor-
rected for attenuation) using of laboratory measurements of
absorption coefficient, and then particulate backscattering
coefficients spectrum (bbp(k), m21) could be derived (Ma
et al. 2006). The backscattering probability of SPM was
derived using the optimization methods provided by Ma
et al. (2008). Following NASA protocols (Mueller et al. 2003),
an analytica spectra devices (ASD) field spectrometer (Field-
Spec Pro Dual VNIR, Boulder, U.S.A.) was used to obtain
remote sensing reflectance. First, total water leaving radiance
(Lsw), radiance of gray panel (Lp), and sky radiance (Lsky)
were measured using the ASD field spectrometer. The water
surface reflectance factor q depended on sky conditions,
wind speed, and solar zenith angle, and was assumed to be
Fig. 1. Study area and observation stations in Lake Chaohu, China. Field measurements of bio-optical parameters made during three cruise surveys in
28 May 2013 (N 5 9), 19–24 July 2013 (N 5 32) and 10–12 October 2013 (N 5 27).
Xue et al. Phytoplankton vertical nonuniform distribution
3

0.028 (Mobley 1999) in terms of the wind speed (< 5 m s21)
and sky conditions (under clear sky or low cloud) in field
measurements of Lake Chaohu. Remote sensing reflectance
(Rrs(k)) was then calculated by the ratio of water-leaving
radiance (Lw(k)) to incident downwelling plane irradiance
(Ed[k, 01]).
RrsðkÞ5LwðkÞ=Edð01; kÞ (1)
Ecolight simulation
Radiative transfer simulations of underwater light field
were carried out with a Hydrolight–Ecolight model (version
5, Sequoia Scientific) to calculate Rrs(k) of the water whose
IOPs are associated with OACs. The Ecolight code solves the
azimuthally-averaged radiative transfer equation (RTE), and
is sufficient for deriving Rrs(k) and spends less time than
solving the azimuthally-dependent RTE in Hydrolight model.
A four-component model of IOPs of water constitutes,
including pure water, phytoplankton, mineral particles, and
CDOM, was used in our computations.
Table 2 lists some common input parameters of Ecolight
simulation. The simulations ranged from 400 nm to 900 nm
with a 5 nm interval. The built-in RADTRAN-X subroutine
of Ecolight was used to model the solar and sky irradiances
incident onto the water surface under clear sky conditions.
The solar zenith angle was assumed to be 308, as it is typical
for the time around midday in the June-September period
in Lake Chaohu. The propagation of light across the air–
water interface was derived by the surface slope statistics
with a wind speed of 2.25 m s21, which is the annual aver-
age wind speed of Lake Chaohu. The field measurements
results indicated that SPIM and CDOM had a lower vertical
variability (coefficient of variation (C.V.)<30%) compared
to Chl a (Xue et al. 2015). In order to estimate the effects of
Chl a(z) profiles on Rrs(k), SPIM and CDOM were assumed
to be vertical independent. The input parameters of SPIM
and CDOM used the mean value of field measurements
(Table 2).
The Fresnel reflectance and transmittance of an air–water
interface was determined by the default refractive index in
Ecolight, n 5 1.34 (Mobley 1994). In addition, Raman scatter-
ing by water molecules was calculated in the simulations.
When the depth of the water is larger than three times of
the Secchi disk depth, the bottom effect on the remote sens-
ing reflectance could be regarded as insignificant (Lodhi and
Table 1. Acronyms, abbreviations, and symbols as appliedhere.
Acronyms and abbreviations
AOPs Apparent optical properties
CDOM Colored dissolved organic matter
IOPs Inherent optical properties
NAP Non-algae particulate
OACs Optically active constituents
RTE Radiative transfer equation
SIOPs Specific inherent optical properties
Symbols
a(k, z) Total absorption coefficient (m21) at wavelength (k)
and depth z m
ag(k) Absorption by CDOM (m21) at wavelength (k)
ad(k) Absorption by NAP (m21) at wavelength (k)
a�d(k) SPIM-specific absorption by NAP (m2 g21) at
wavelength (k)
aph(k) Absorption by phytoplankton (m21) at wavelength (k)
a�ph(k) Chl a-specific absorption by phytoplankton (m2 mg21)
at wavelength (k)
aw(k) Absorption by pure water (m21) at wavelength (k)
b(k, z) Total scattering coefficient (m21) at wavelength (k) and
depth z m, b 5 bp 1 bw
bd(k) Scattering coefficients of NAP (m21) at wavelength (k)
b�d(k) SPIM-specific scattering coefficient of NAP (m2 g21) at
wavelength (k)
bp(k) Scattering coefficients of particulate (m21) at
wavelength (k), bp 5 bph 1 bd
bph(k) Scattering coefficients of phytoplankton (m21) at
wavelength (k)
b�ph(k) Chl a-specific scattering coefficient of phytoplankton
(m2 mg21) at wavelength (k)
bw(k) Scattering coefficients of pure water (m21) at
wavelength (k)
Chl a Chlorophyll a concentration (lg L21)
Chl a(z) Vertical profile of chlorophyll a concentration (lg L21)
Rrs Remote sensing reflectance (sr21)
Rrs-c Corrected remote sensing reflectance (sr21)
Rrs-h Remote sensing reflectance (sr21) of homogeneous waters
Rrs-v Remote sensing reflectance (sr21) of inhomogeneous waters
Sg Spectral slope of ag spectrum (nm21)
DRrs(k) Relative difference of Rrs(k) between inhomogeneous and
homogeneous waters (%)
SPIM Suspended particulate inorganic matter (mg L21)
h Spectral angle (rad)
kc Centroid wavelength of remote sensing reflectance
spectrum (nm)
Table 2. Input parameters used in the Ecolight simulations.
Variable Inputs
Solar zenith angle 308
Wind speed (m s21) 2.25
Particle phase function FF phase function
SPIM (mg L21) 30
ag(440) (m21) 0.85
Sg (nm21) 0.019
k (nm) 400–900, every 5 nm
Xue et al. Phytoplankton vertical nonuniform distribution
4

Rundquist 2001). Thus, water bottom reflection of Lake
Chaohu was ignored (Ma et al. 2006; Ma et al. 2011a; Ma
et al. 2011b).
Inherent optical properties
In the four-component model, total spectral absorption
coefficient of water, a(k, z) (m21), is expressed as:
aðk; zÞ5awðkÞ1aphðk; zÞ1adðkÞ1agðkÞ (2)
where a(k, z) is the spectral absorption coefficient of water at
wavelength k nm and depth z m. aph(k, z) is the spectral
absorption coefficient of phytoplankton at k nm and z m.
aw(k) (Pope and Fry 1997), ad(k), and ag(k) are the spectral
absorption coefficient of pure water, NAP, and CDOM at knm, and were assumed to be independent in depth in this
study. The vertical profile of aph(k, z) is calculated from:
aphðk; zÞ5Chl aðzÞ3a�phðkÞ (3)
where a�ph(k) (m2 mg21) is the spectral Chl a-specific absorp-
tion coefficient of phytoplankton, and is assumed to be con-
stant with depth. The NAP absorption is calculated from:
adðkÞ5SPIM3a�dðkÞ (4)
where a�d(k) (m2 g21) is the spectral SPIM-specific absorption
coefficient of SPIM. The CDOM absorption is modeled as an
exponential function of light wavelength:
agðkÞ5agðk0Þexp ½2Sgðk2k0Þ� (5)
where ag(k0) is the CDOM absorption coefficient estimated
at a reference wavelength k0 (5 440 nm), and Sg (nm21) is
the spectral slope of the CDOM absorption spectrum
between 400 nm and 700 nm. Following the in situ measure-
ments, Sg ranged from 0.012 nm21 to 0.043 nm21 with aver-
age of 0.019 (6 0.004) nm21.
The total spectral scattering coefficient, b(k, z)(m21), is
the sum of scattering coefficient of pure water, bw(k) (m21)
(Morel 1974), and SPM, bp(k, z) (m21):
bðk; zÞ5bwðk; zÞ1bpðk; zÞ (6)
bp(k, z) is the sum of scattering coefficient of phytoplankton,
bph(k) (m21), and NAP, bd(k) (m21):
bpðk; zÞ5bphðk; zÞ1bdðk; zÞ (7)
b�ph(k) (m2 mg21) and b�d(k) (m2 g21) are the spectral specific
scattering coefficient of phytoplankton and NAP. b�ph(k) and
b�d(k) were determined by partitioning the bbp into scattering
coefficient of phytoplankton and NAP (Eq. 7) using multiple
linear regression with Chl a and SPIM concentration (Eq. 8)
(Snyder et al. 2008):
bpðk; zÞ5Chl aðzÞ3b�phðkÞ1SPIM3b�dðkÞ (8)
Similarly to mass specific absorption coefficient, the values
of b�ph(k) and b�d(k) were assumed to be independent of
depth.
The specific inherent optical properties (SIOPs) of Lake
Chaohu were derived from the field measurements, and then
IOPs could be derived based on the above Eqs. 2–-8. Optical
measurements of the water samples ranging from 350 nm to
800 nm were extrapolate to 900 nm in the near infrared
(NIR) part of the spectrum. Typical examples of specific
absorption and backscattering coefficient spectra of Lake
Chaohu were set as the input files of Ecolight simulation
(Fig. 2).
The scattering phase functions for pure water, phyto-
plankton, and mineral particles were also key input IOPs.
The scattering phase function of pure water was provided by
the file (pureh2o.dpf) in the Ecolight model. The Fournier-
Fig. 2. Average SIOPs for Lake Chaohu: (a) the spectral absorptioncoefficient of pure water (aw, m21) and the specific absorption coeffi-cient spectra of Chl a (aph*, m2 mg21), NAP (ad*, m2 g21) and CDOM
(ag*, dimensionless); (b) the spectral backscattering coefficient of purewater (bbw, m21) and the specific backscattering coefficient spectra of
Chl a (bbph*, m2 mg21), NAP (bbd*, m2 g21). aw(k) and bw(k) are readfrom file HE5\data\H20abDefaults.txt in Hydrolight-Ecolight model.
Xue et al. Phytoplankton vertical nonuniform distribution
5
Forand (FF) phase function (Fournier and Forand 1994) was
specified as the scattering phase function of phytoplankton
and NAP (Mobley et al. 2002). FF phase function can be
determined mainly by the backscattering fraction, bb/b,
which was derived by optimization method. Ecolight simu-
lation was performed by changing the backscattering frac-
tion of phytoplankton and NAP, and the simulation with
lowest errors between simulated Rrs(k) and field measured
Rrs(k) had optimum value of backscattering fraction. Back-
scattering fraction was 0.02 and 0.05 for the phytoplankton
and NAP, respectively. Some studies demonstrated that the
backscattering ratio is wavelength dependent and not a con-
stant value (Aas et al. 2005). The particulate backscattering
fractions and the particulate scattering phase functions were
assumed to be independent of light wavelength and depth
in this study.
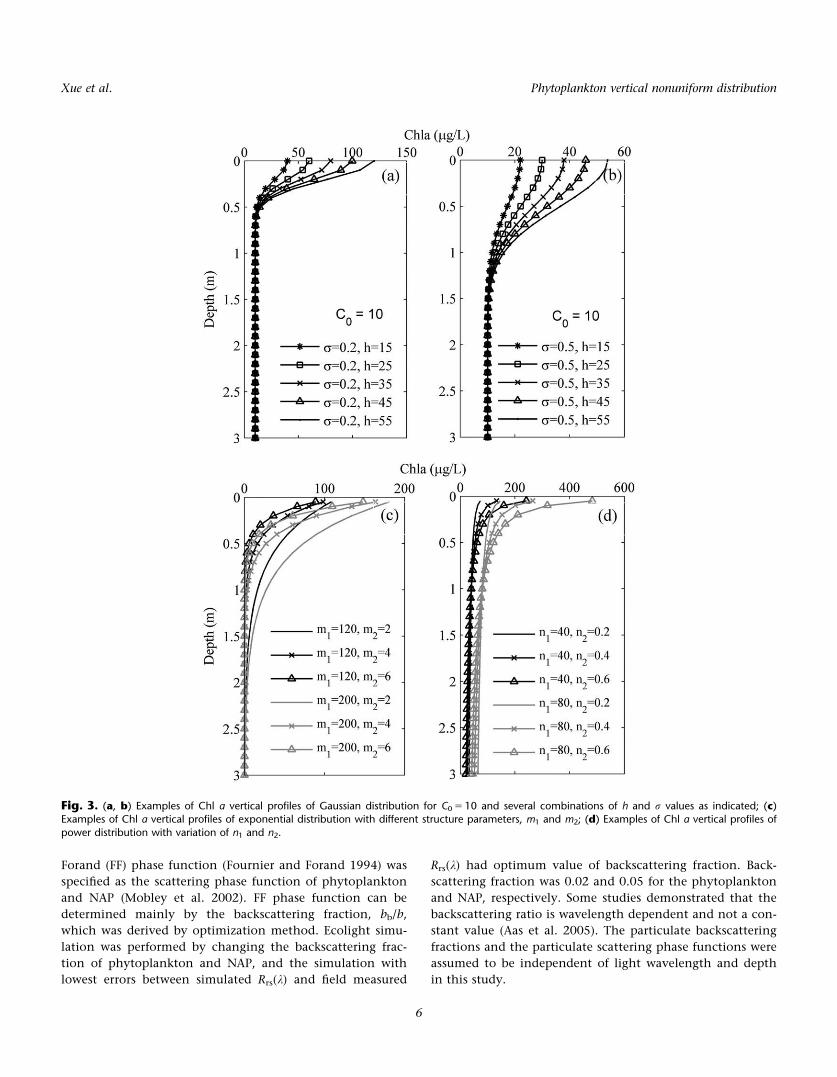
Fig. 3. (a, b) Examples of Chl a vertical profiles of Gaussian distribution for C0 5 10 and several combinations of h and r values as indicated; (c)Examples of Chl a vertical profiles of exponential distribution with different structure parameters, m1 and m2; (d) Examples of Chl a vertical profiles of
power distribution with variation of n1 and n2.
Xue et al. Phytoplankton vertical nonuniform distribution
6

Chl a(z) profiles
Previous study found that the Chl a(z) profiles of Lake
Chaohu exhibited four vertical classes, including vertical
uniform, Gaussian, exponential, and power distribution
(Xue et al. 2015). These inhomogeneous vertical Chl a(z)
profiles usually had maximum concentration at the water
surface in cyanobacterial bloom waters. Specifically, Chl
a(z) profiles of the three inhomogeneous profile classes
were derived according to the range and step of vertical
structure parameters of each class, which is derived from
field measurements (Eqs. 9–-11; Fig. 3). Based on the range
and step of structure parameters as presented in Table 3,
1008, 2400, and 576 profiles of Chl a(z) were generated for
Gaussian, exponential, and power class, respectively.
Hence, total of 3984 radiative transfer simulations were
made with different sets of input IOPs representing nonuni-
form water column.
Chl aGðzÞ5C01h
rffiffiffiffiffiffi2pp exp
�2
1
2ðzrÞ2�
(9)
Chl aEðzÞ5m13exp ðm23zÞ (10)
Chl aPðzÞ5n13zn2 (11)
In addition to the simulations of vertical inhomogeneous
water, reference simulations for vertically homogeneous
water were also performed. In the reference simulation, the
average Chl a of the corresponding inhomogeneous case was
used to determine the independent IOPs. These reference
simulations examined the effects of vertical Chl a(z) profiles
on Rrs(k) relative to the vertically homogeneous water. For
each of 3984 simulations of nonuniform water, the relative
difference (DRrs(k)) between Rrs(k) for the nonuniform case
and Rrs(k) for the corresponding uniform case was calculated
as:
DRrsðkÞ5Rrs2vðkÞ2Rrs2hðkÞ
Rrs2hðkÞ3100% (12)
where Rrs-v(k) and Rrs-h(k) was the spectral Rrs(k) of the verti-
cally nonuniform and uniform water, respectively. DRrs(k)
was used to analyze the sensitivity of Rrs(k) to Chl a(z) pro-
files within the water column.
Method to correct Rrs(k) of nonuniform waters
Rrs-v(k) contains information of Chl a(z) profiles, but it is
difficult to decide to what extent Rrs(k) is depended on verti-
cal effects. A Rrs(k) correction scheme was developed to solve
this problem and decreased the influence of vertical distribu-
tion of Chl a(z) on Rrs(k) based on the relationships between
DRrs(k) and Chl a(z) structure parameters. According to Eq.
12, Rrs-v(k) can be converted to Rrs-h(k), if DRrs(k) could be
estimated. First, as DRrs(k) varies with structure parameters of
Chl a(z) (S), empirical relationships between DRrs(k) and Chl
a(z) profile parameters of three Chl a(z) profile classes were
build. Then, corrected Rrs(k) (Rrs-c(k)) could be derived
according to Eq. 13:
Rrs2ciðkÞ5Rrs2viðkÞ
DRrs2iðkÞ=10011(13)
Hence, the corrected Rrs(k) of each Chl a profile class, Rrs-
ci(k), would be derived using the Rrs-vi(k) and structure
parameters of Chl a(z), Si. i 5 1, 2, 3, represents Gaussian,
exponential, and power class, respectively.
Statistical analyses
To exhibit the variability of Chl a(z) vertical profiles, the
C.V. was derived with the ratio of standard deviation (SD)
and mean value:
CV5SD
mean3100% (14)
Centroid wavelength of remote sensing reflectance spectrum
(kc) was used to present the difference of centroid wave-
length as Chl a(z) profiles changed. It was calculated as (Lin
et al. 2012):
kc5
X900
k5400
k3RrsðkÞ
X900
k5400
RrsðkÞ(15)
Spectral angle (h) measures differences between Rrs(k) of ver-
tical uniform and nonuniform waters in spectral shape, and
is calculated as (Kruse et al. 1993; Dennison et al. 2004):
h5cos 21
X900
k5400
Rrs2hðkÞRrs2vðkÞffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiX900
k5400
Rrs2h2ðkÞ
vuutffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiX900
k5400
Rrs2v2ðkÞ
vuut
0BBBBBB@
1CCCCCCA
(16)
h ranges from 0 to p/2, h 5 0 represents that the two spec-
trum are similar, and large value illustrates large difference.
Table 3. Range and step of Chl a(z) structure parameters con-sidered in radiative transfer simulations of underwater light field.
Class Parameters Range Step Numbers
Gaussian C0 0–40 5 1008
h 1–76 5
r 0.2–1.4 0.2
Exponential m1 120–600 20 2400
m2 0.5–10 0.1
Power n1 10–80 2 576
n2 0.2–0.95 0.05
Xue et al. Phytoplankton vertical nonuniform distribution
7

R2 and root mean square error (RMSE) in log space were
used to assess algorithm performance:
R2512
XN
i51ðyi2xiÞ2XN
i51ðyi2�xiÞ2
(17)
RMSEðlog Þ5
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi1
N
XNi51
ðlog 10ðyiÞ2log 10ðxiÞÞ2vuut (18)
where xi and yi refer to the referenced and modeled values of
the ith sample, N is the number of samples.
Results
Effects of Chl a vertical nonuniform on Rrs(k)
There exist many Chl a(z) vertical profiles with different
structure parameters of each vertical profile class (Gaussian,
exponential, and power class), even the average Chl a con-
centration of the water column is constant. Centroid wave-
length (kc) and spectral angle (h) reflect the general variation
of remote sensing reflectance spectrum (Fig. 4). kc represents
the centroid position of Rrs(k) spectra between 400 nm and
900 nm. h could be used to evaluate the similarity of Rrs(k)
spectra between homogeneous water and inhomogeneous
water with same average Chl a concentration in the water
column.
The relationships between kc and C.V. of Chl a(z) profiles
of different vertical profile classes showed that kc increased
with increase of C.V. among the three classes. Compared to
homogeneous waters, kc of Gaussian class moved from near
612 nm to the longer wavelength, about 660 nm (Fig. 4a).
Spectral angle (h) between homogeneous water and
inhomogeneous water had similar tendency with kc (Fig. 4b).
In addition, same C.V. was corresponding to different kc and
h in different Chl a(z) profile classes, and extent of the
effects of Chl a vertical nonuniform on Rrs(k) was different
among three classes, in which the Gaussian class was more
sensitive than other two classes. Thus, we analyzed the
effects of Chl a vertical nonuniform on Rrs(k) of each class
separately in the following content.
When the average Chl a concentration of the water col-
umn is unchangeable, Rrs(k) spectra of inhomogeneous Chl
a(z) profiles can depart apparently from the spectra of the
homogeneous ones. Example results of Rrs(k) and DRrs(k) sim-
ulated by Ecolight for different pairs of Chl a vertical struc-
ture parameters were provided (Fig. 5). In each class of Chl
a(z) profile, the multiple plots of Rrs(k) spectra corresponded
to different values of structure parameters.
For Gaussian class, the presence of near-surface high Chl
a value led to decrease in Rrs(k) from 400 nm to about
700 nm. Under these conditions, DRrs(k) in the green and
red wavelengths reaches the values of 240% to 260% (Fig.
5a,b). Water column stratification had reverse effect on Rrs(k)
and enhanced the Rrs(k) at wavelength longer than 700 nm.
These departures were most obvious for high contrast in Chl
a(z) profiles. For example, DRrs(k) reached 260% in the red
and 70% in the NIR spectrum when the top layer was very
thin, i.e., r 5 0.2 (Fig. 5a). The increase in C0 naturally
resulted in reduction of the observed effects due to weak
stratification. For large C0 values (e.g., C0 5 40 in Fig. 5c,d),
the magnitude of Rrs-v(k) was close to that of Rrs-h(k) from
400 nm to 900 nm.
The simulations for exponential class also showed that
the largest difference was observed at about 600–700 nm
when the top layer is thin as m2 increase (Fig. 5e,f). Howev-
er, Rrs(k) had highest values in red wavelength when most of
Fig. 4. Variation of (a) centroid wavelength (kc) and (b) spectral angle (h) with C.V. of Chl a(z) profiles in Gaussian (“G”), exponential (“E”), andpower (“P”) class.
Xue et al. Phytoplankton vertical nonuniform distribution
8

Fig. 5. Example results of radiative transfer simulations of Gaussian (a–d), exponential (e, f) and power (g, h) profile classes, showing the differences inRrs(k) and DRrs(k) between the uniform water (black curves) and the inhomogeneous water (color curves). The average Chl a (Chl a_ave) in the nonuni-
form case is identical to that in the uniform case. (a, b) C0 5 10, Chl a_ave 5 45 lg L21; (c, d) r 5 0.4, Chl a_ave 5 45 lg L21; (e, f) Chl a_ave 5 60 lgL21; (g, h) Chl a_ave 5 60 lg L21. Each graph for DRrs(k) (right-hand panels) is corresponding to the data presented in the left-hand panels.
Xue et al. Phytoplankton vertical nonuniform distribution
9

the cyanobacteria were close to the water surface (Fig. 5g,h).
For instance, as the decrease of n1 for power class, the phyto-
plankton aggregated near the water surface, and Rrs(k) of Chl
a vertical nonuniform was larger than the vertical uniform
case in the visible region and lower in the NIR band. The
Rrs(k) spectra of uniformly water were significantly lower
than those of nonuniform vertical distributions with average
Chl a 60 lg L21 in the water column. This had similar results
as Kutser et al. (2008), and showed opposite tendency with
the other two profile classes.
The Chl a(z) inhomogeneous profiles not only introduced
the differences of shape and magnitude in remote sensing
reflectance spectra, but also affected remote sensing esti-
mates of Chl a concentration. Standard Chl a inversion algo-
rithms of Case I waters are based on the blue-green ratio,
and fail badly in complex inland waters. First, we made a
test using the band ratio (Rrs(709)/Rrs(675)) of simulated data
to illustrate the influence of Chl a vertical profiles on Chl a
inversion algorithm. The results indicated that correlation
between Rrs-h(709)/Rrs-h(675) and Chl a of vertical uniform
waters was good (R2 5 0.99):
x5Rrsð709Þ=Rrsð675Þ
Chl a546:29x2137:52x246:563(19)
However, the Chl a estimated by the above relationship
using Rrs-v(709)/Rrs-v(675) of nonuniform waters did not
match the homogeneous ones (Fig. 6a–c). The scatter plots
between Chl a of homogeneous water and modeled Chl a of
inhomogeneous waters based on the above equation illus-
trated that vertical inhomogeneous of Chl a(z) profiles
severely influenced the accuracy of Chl a algorithm. General-
ly, overestimation of Chl a inversion algorithm was intro-
duced, and the average relative error between Chl a-model
and Chl a–h was 79.8%, 549.9%, and 99.6% for Gaussian,
exponential, and power class, respectively. For example, sur-
face Chl a of nonuniform case ranged about 50–113 lg L21,
while the average concentration in the uniform case was
Fig. 6. Comparison between modeled Chl a and Chl a of homogeneous waters simulated by Ecolight of Gaussian (a, d), exponential (b, e), andpower (c, f) class. Modeled Chl a of (a–c) are derived by band ratio algorithm (Eq. 19), modeled Chl a of (d–f) are derived by FLH inversion algo-rithm (Eq. 20). The points close to the 1:1 line represent simulations with negligible effect of inhomogeneous Chl a(z) profiles on Chl a inversion
algorithm.
Xue et al. Phytoplankton vertical nonuniform distribution
10

45 lg L21. The relative error of DRrs-v(709)/Rrs-v(675) was 14
times of DRrs(443), that is to say, when the relative error of
DRrs(443) was 10%, the error of Rrs(709)/Rrs(675) would be
140%, and the relative error of Chl a estimation raised up to
153%.
As the sensitivity of Chl a algorithm to Rrs variation was
different, another Chl a algorithm developed based on fluo-
rescence line height (FLH) index (Letelier and Abbott 1996)
was used to estimate the effect of Chl a vertical nonuniform
on Chl a inversion:
Fig. 7. Dependence of relative difference between inhomogeneous and homogeneous waters of Rrs (DRrs) on Chl a vertical structure parameters at
443 nm, 550 nm, 675 nm, and 709 nm. C0 is given to be 10 in the 1st column, and h is 40 in the 2nd column.
Xue et al. Phytoplankton vertical nonuniform distribution
11
FLH5Rrs k2ð Þ2 Rrs k1ð Þ1 Rrs k3ð Þ2Rrs k1ð Þð Þ k22k1
k32k1
� �
Chl a538:89EXPð2359:9FLHÞ(20)
where, k1 5 665 nm, k2 5 681 nm, k3 5 709 nm, R2 5 0.99. In
this model, the average relative error between Chl a-model
and Chl a–h was 140%, 508%, and 92% for Gaussian, expo-
nential, and power class, respectively. Most data points of
the nonuniform Chl a(z) profiles lie on the left-hand side of
1:1 line or 1:2 line (Fig. 6d–f). It indicated that application
of the band ratio or FLH algorithm can lead to substantial
overestimation of surface Chl a, if the vertical variation of
phytoplankton in cyanobacterial bloom waters was ignored.
Relationships of DRrs(k) and Chl a structure parameters
The relationships between relative difference of remote
sensing reflectance (DRrs(k)) and Chl a vertical structure
parameters were illustrated for the three inhomogeneous Chl
a vertical profile classes (Fig. 7). For Gaussian class, when h
increased or r decreased, the variation of Chl a(z) profile
within the Chl a peak increased, and then DRrs(k) increased
as well. The effect of Chl a(z) profile on Rrs(k) weakened with
the increase of r. For r 5 0.2 and h 5 40, Rrs-v(443) was lower
by � 20%, Rrs-v(550) was lower by 30% than the uniform
ones. If the Chl a peak was wide (r 5 0.8, h 5 40), DRrs(550)
was about 210%. For exponential profile class, DRrs increased
with increase of m1 and decrease of m2. Variation of m2 had
little influence on DRrs, especially when m2 was large. For
power profile class, n2 had significant effect on DRrs, and var-
iation of DRrs was less than 10% with the increase of n1 from
10 to 80.
Structure parameters of different Chl a profile classes have
important influence on DRrs(k). Figures 5 and 7 showed sev-
eral examples of the effects caused by changing structure
parameters of the Chl a(z) profile. Based on about 3/4 of the
simulated data (N 5 2874), we established a relationship
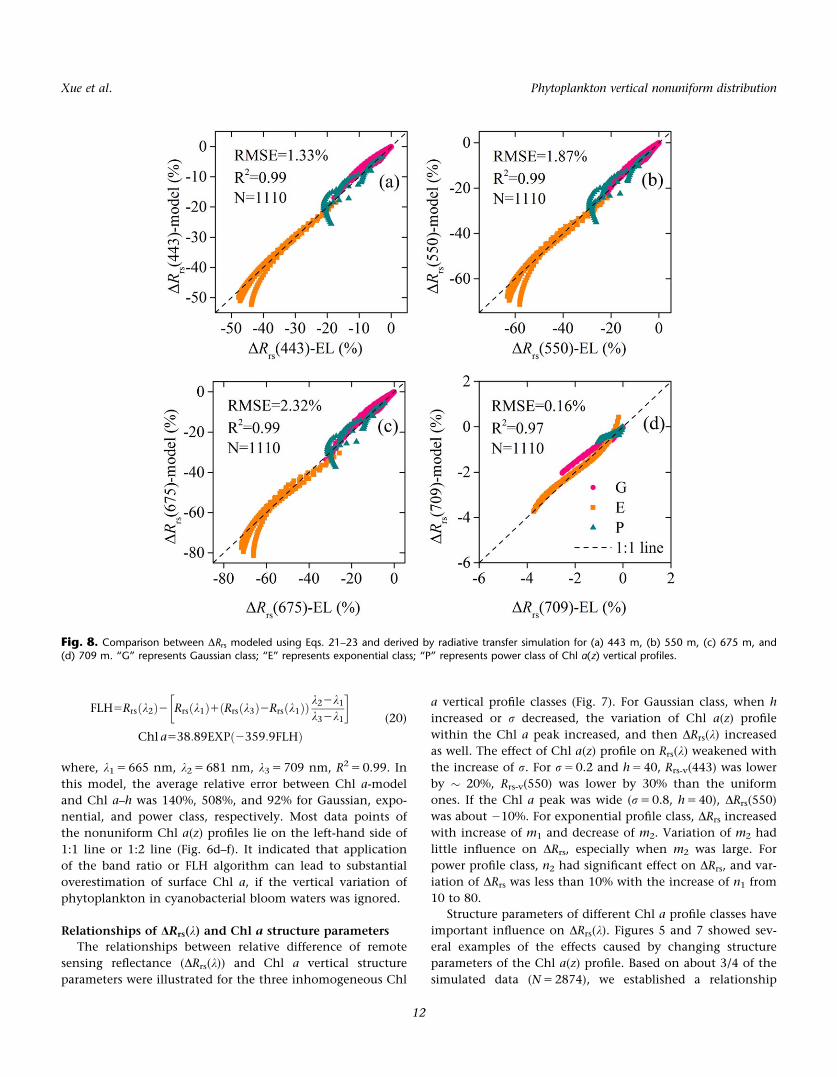
Fig. 8. Comparison between DRrs modeled using Eqs. 21–23 and derived by radiative transfer simulation for (a) 443 m, (b) 550 m, (c) 675 m, and(d) 709 m. “G” represents Gaussian class; “E” represents exponential class; “P” represents power class of Chl a(z) vertical profiles.
Xue et al. Phytoplankton vertical nonuniform distribution
12
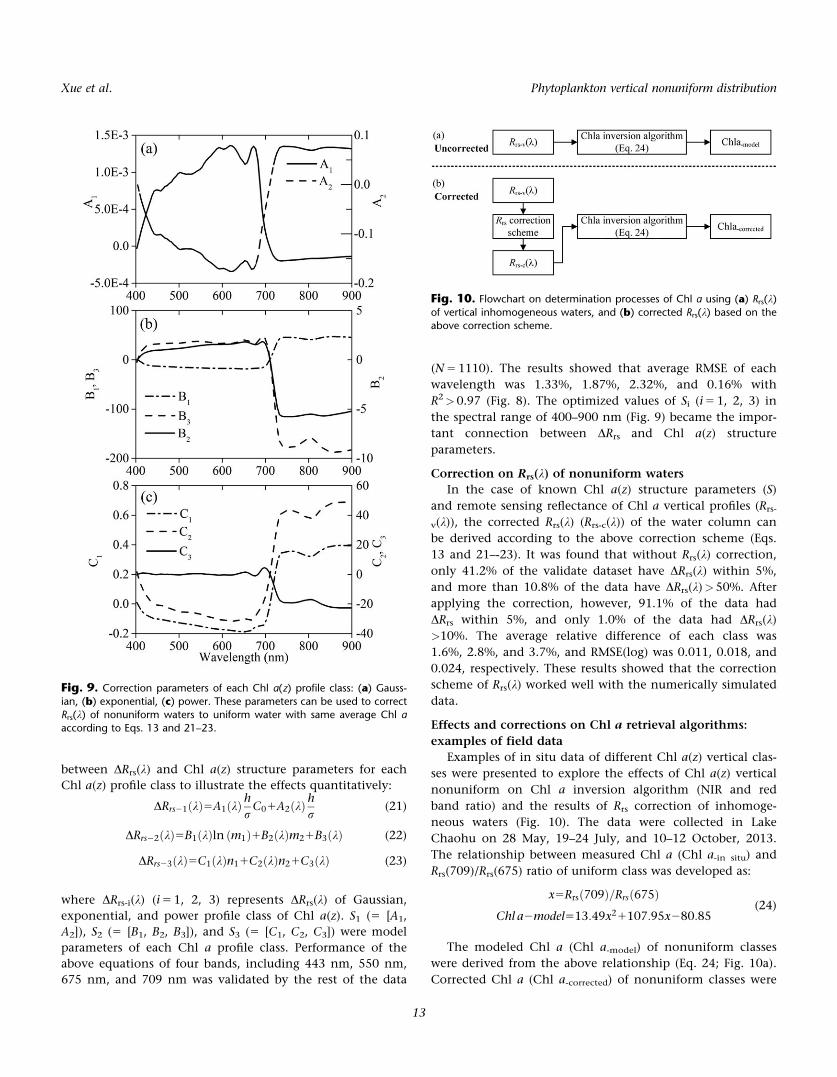
between DRrs(k) and Chl a(z) structure parameters for each
Chl a(z) profile class to illustrate the effects quantitatively:
DRrs21ðkÞ5A1ðkÞh
rC01A2ðkÞ
h
r(21)
DRrs22ðkÞ5B1ðkÞln ðm1Þ1B2ðkÞm21B3ðkÞ (22)
DRrs23ðkÞ5C1ðkÞn11C2ðkÞn21C3ðkÞ (23)
where DRrs-i(k) (i 5 1, 2, 3) represents DRrs(k) of Gaussian,
exponential, and power profile class of Chl a(z). S1 (5 [A1,
A2]), S2 (5 [B1, B2, B3]), and S3 (5 [C1, C2, C3]) were model
parameters of each Chl a profile class. Performance of the
above equations of four bands, including 443 nm, 550 nm,
675 nm, and 709 nm was validated by the rest of the data
(N 5 1110). The results showed that average RMSE of each
wavelength was 1.33%, 1.87%, 2.32%, and 0.16% with
R2>0.97 (Fig. 8). The optimized values of Si (i 5 1, 2, 3) in
the spectral range of 400–900 nm (Fig. 9) became the impor-
tant connection between DRrs and Chl a(z) structure
parameters.
Correction on Rrs(k) of nonuniform waters
In the case of known Chl a(z) structure parameters (S)
and remote sensing reflectance of Chl a vertical profiles (Rrs-
v(k)), the corrected Rrs(k) (Rrs-c(k)) of the water column can
be derived according to the above correction scheme (Eqs.
13 and 21–-23). It was found that without Rrs(k) correction,
only 41.2% of the validate dataset have DRrs(k) within 5%,
and more than 10.8% of the data have DRrs(k)>50%. After
applying the correction, however, 91.1% of the data had
DRrs within 5%, and only 1.0% of the data had DRrs(k)
>10%. The average relative difference of each class was
1.6%, 2.8%, and 3.7%, and RMSE(log) was 0.011, 0.018, and
0.024, respectively. These results showed that the correction
scheme of Rrs(k) worked well with the numerically simulated
data.
Effects and corrections on Chl a retrieval algorithms:
examples of field data
Examples of in situ data of different Chl a(z) vertical clas-
ses were presented to explore the effects of Chl a(z) vertical
nonuniform on Chl a inversion algorithm (NIR and red
band ratio) and the results of Rrs correction of inhomoge-
neous waters (Fig. 10). The data were collected in Lake
Chaohu on 28 May, 19–24 July, and 10–12 October, 2013.
The relationship between measured Chl a (Chl a-in situ) and
Rrs(709)/Rrs(675) ratio of uniform class was developed as:
x5Rrsð709Þ=Rrsð675Þ
Chl a2model513:49x21107:95x280:85(24)
The modeled Chl a (Chl a-model) of nonuniform classes
were derived from the above relationship (Eq. 24; Fig. 10a).
Corrected Chl a (Chl a-corrected) of nonuniform classes were
Fig. 9. Correction parameters of each Chl a(z) profile class: (a) Gauss-
ian, (b) exponential, (c) power. These parameters can be used to correctRrs(k) of nonuniform waters to uniform water with same average Chl a
according to Eqs. 13 and 21–23.
Fig. 10. Flowchart on determination processes of Chl a using (a) Rrs(k)
of vertical inhomogeneous waters, and (b) corrected Rrs(k) based on theabove correction scheme.
Xue et al. Phytoplankton vertical nonuniform distribution
13
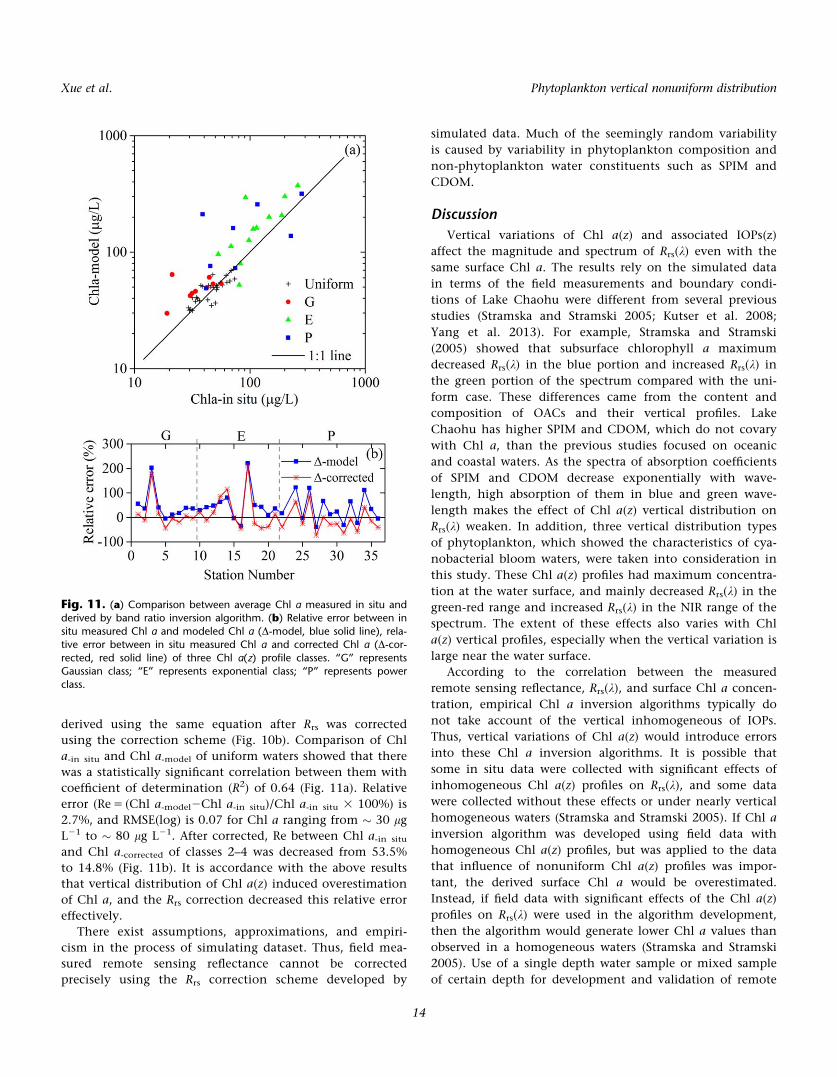
derived using the same equation after Rrs was corrected
using the correction scheme (Fig. 10b). Comparison of Chl
a-in situ and Chl a-model of uniform waters showed that there
was a statistically significant correlation between them with
coefficient of determination (R2) of 0.64 (Fig. 11a). Relative
error (Re 5 (Chl a-model2Chl a-in situ)/Chl a-in situ 3 100%) is
2.7%, and RMSE(log) is 0.07 for Chl a ranging from � 30 lg
L21 to � 80 lg L21. After corrected, Re between Chl a-in situ
and Chl a-corrected of classes 2–4 was decreased from 53.5%
to 14.8% (Fig. 11b). It is accordance with the above results
that vertical distribution of Chl a(z) induced overestimation
of Chl a, and the Rrs correction decreased this relative error
effectively.
There exist assumptions, approximations, and empiri-
cism in the process of simulating dataset. Thus, field mea-
sured remote sensing reflectance cannot be corrected
precisely using the Rrs correction scheme developed by
simulated data. Much of the seemingly random variability
is caused by variability in phytoplankton composition and
non-phytoplankton water constituents such as SPIM and
CDOM.
Discussion
Vertical variations of Chl a(z) and associated IOPs(z)
affect the magnitude and spectrum of Rrs(k) even with the
same surface Chl a. The results rely on the simulated data
in terms of the field measurements and boundary condi-
tions of Lake Chaohu were different from several previous
studies (Stramska and Stramski 2005; Kutser et al. 2008;
Yang et al. 2013). For example, Stramska and Stramski
(2005) showed that subsurface chlorophyll a maximum
decreased Rrs(k) in the blue portion and increased Rrs(k) in
the green portion of the spectrum compared with the uni-
form case. These differences came from the content and
composition of OACs and their vertical profiles. Lake
Chaohu has higher SPIM and CDOM, which do not covary
with Chl a, than the previous studies focused on oceanic
and coastal waters. As the spectra of absorption coefficients
of SPIM and CDOM decrease exponentially with wave-
length, high absorption of them in blue and green wave-
length makes the effect of Chl a(z) vertical distribution on
Rrs(k) weaken. In addition, three vertical distribution types
of phytoplankton, which showed the characteristics of cya-
nobacterial bloom waters, were taken into consideration in
this study. These Chl a(z) profiles had maximum concentra-
tion at the water surface, and mainly decreased Rrs(k) in the
green-red range and increased Rrs(k) in the NIR range of the
spectrum. The extent of these effects also varies with Chl
a(z) vertical profiles, especially when the vertical variation is
large near the water surface.
According to the correlation between the measured
remote sensing reflectance, Rrs(k), and surface Chl a concen-
tration, empirical Chl a inversion algorithms typically do
not take account of the vertical inhomogeneous of IOPs.
Thus, vertical variations of Chl a(z) would introduce errors
into these Chl a inversion algorithms. It is possible that
some in situ data were collected with significant effects of
inhomogeneous Chl a(z) profiles on Rrs(k), and some data
were collected without these effects or under nearly vertical
homogeneous waters (Stramska and Stramski 2005). If Chl a
inversion algorithm was developed using field data with
homogeneous Chl a(z) profiles, but was applied to the data
that influence of nonuniform Chl a(z) profiles was impor-
tant, the derived surface Chl a would be overestimated.
Instead, if field data with significant effects of the Chl a(z)
profiles on Rrs(k) were used in the algorithm development,
then the algorithm would generate lower Chl a values than
observed in a homogeneous waters (Stramska and Stramski
2005). Use of a single depth water sample or mixed sample
of certain depth for development and validation of remote
Fig. 11. (a) Comparison between average Chl a measured in situ andderived by band ratio inversion algorithm. (b) Relative error between in
situ measured Chl a and modeled Chl a (D-model, blue solid line), rela-tive error between in situ measured Chl a and corrected Chl a (D-cor-
rected, red solid line) of three Chl a(z) profile classes. “G” representsGaussian class; “E” represents exponential class; “P” represents powerclass.
Xue et al. Phytoplankton vertical nonuniform distribution
14

sensing algorithm is inadequate in many situations (Kutser
et al. 2008).
The internal inconsistency correlating the surface Chl a
with Rrs(k) may generate errors in surface Chl a estimation
from the present remote sensing inversion algorithms.
Development of regional or seasonal algorithms with careful-
ly chosen field data known vertical profiles may minimize
this problem, but it is difficult and could not overcome the
problem fundamentally. The correction scheme of Rrs(k) on
nonuniform waters eliminated the errors caused by vertical
variation of phytoplankton, and could be used to estimate to
what extent the inhomogeneous Chl a(z) profile might affect
remote sensing reflectance and the performance of the cur-
rent empirical algorithm.
The Rrs(k) correction scheme indicates the potential of
decreasing the effects of vertical inhomogeneous of phyto-
plankton on remote sensing reflectance. This approach is
established based on field measured data and radiative trans-
fer simulated data, and it is suitable for shallow cyanobacte-
rial bloom waters. With the knowledge of Chl a(z) profile
parameters, Rrs(k) of nonuniform waters can be corrected to
the Rrs(k) of uniform case. The application of this approach
to satellite water color data remains to be determined.
Conclusion
Vertical variation of IOPs characterized by Chl a(z) verti-
cal profiles with maximum value at water surface, including
Gaussian, exponential, and power class, often occurs in
eutrophic shallow waters. Radiative transfer simulations of
different vertical profiles of Chl a(z) indicated that such ver-
tical variations can significantly affect the remote sensing
reflectance, compared with vertically uniform water (with
the average Chl a identical to the nonuniform case across
the water column). The differences in Rrs(k) between the
vertical nonuniform and uniform water can be as high
as>70% when the near-surface Chl a decreases dramatically
with depth, i.e., r 5 0.2. The magnitude of these differences
depends strongly on Chl a(z) structure parameters of each
profile class and wavelength of light. Large near-surface var-
iations in Chl a(z) are more likely to produce significant
errors in band ratio of Rrs and present Chl a inversion algo-
rithms. The overestimation of Chl a estimation based on
the NIR-to-red reflectance ratio was 79.8%, 549.9% and
99.6% for Gaussian, exponential and power class,
respectively.
The good relationships between DRrs(k) and the struc-
ture parameters in the spectral region from 400 nm to
900 nm was established in each profile class. With the
knowledge of Chl a(z) profile parameters, Rrs(k) of nonuni-
form waters can be corrected to the Rrs(k) of uniform
waters with same average Chl a across the water column.
This Rrs correction scheme was validated by both Ecolight
simulated data and field measured data. Some inversion
uncertainties of bio-optical parameters can be removed if
the effects of Chl a(z) nonuniform on Rrs corrected.
Although the implication of this correction scheme to oth-
er waters should know about the SIOPs, the demonstrated
potential for correcting the relative difference of Rrs(k)
caused by Chl a(z) vertical variation is encouraging. The
Rrs correction performed just rely on several bands of Rrs(k)
in the case of not knowing Chl a(z) profile is necessary in
further research.
Author contributions
Ronghua Ma proposed the research question and helped
to interpret the results. Kun Xue assembled and analyzed the
data, and drafted the manuscript. Yuchao Zhang and Hon-
gtao Duan contributed to the data simulation, and analysis
of the results. All co-authors revised the manuscript.
References
Aas, E., J. Høkedal, and K. Sørensen. 2005. Spectral backscat-
tering coefficient in coastal waters. Int. J. Remote Sens.
26: 331–343. doi:10.1080/01431160410001720324
Andr�e, J. M. 1992. Ocean color remote-sensing and the sub-
surface vertical structure of phytoplankton pigments.
Deep-Sea Res. Part A Oceanogr. Res. Pap. 39: 763–779.
doi:10.1016/0198-0149(92)90119-E
Bresciani, M., M. Adamo, G. De Carolis, E. Matta, G. Pas-
quariello, D. Vaici�ut _e, and C. Giardino. 2014. Monitoring
blooms and surface accumulation of cyanobacteria in the
Curonian Lagoon by combining MERIS and ASAR data.
Remote Sens. Environ. 146: 124–135. doi:10.1016/
j.rse.2013.07.040
Bricaud, A., A. Morel, and L. Prieur. 1981. Absorption by dis-
solved organic matter of the sea (yellow substance) in the
UV and visible domains 1. Limnol. Oceanogr. 26: 43–53.
doi:10.4319/lo.1981.26.1.0043
Cao, H. S., F. X. Kong, L. C. Luo, X. L. Shi, Z. Yang, X. F.
Zhang, and Y. Tao. 2006. Effects of wind and wind-induced
waves on vertical phytoplankton distribution and surface
blooms of Microcystis aeruginosa in Lake Taihu. J. Freshw.
Ecol. 21: 231–238. doi:10.1080/02705060.2006.9664991
Chen, X., X. Yang, X. Dong, and E. Liu. 2013. Environmen-
tal changes in Chaohu Lake (southeast, China) since the
mid 20th century: The interactive impacts of nutrients,
hydrology and climate. Limnol. Ecol. Manage. Inland
Waters 43: 10–17. doi:10.1016/j.limno.2012.03.002
Cleveland, J. S., and A. D. Weidemann. 1993. Quantifying
absorption by aquatic particles: A multiple scattering cor-
rection for glass-fiber filters. Limnol. Oceanogr. 38: 1321–
1327. doi:10.4319/lo.1993.38.6.1321
Cullen, J., and R. Eppley. 1981. Chlorophyll maximum layers
of the Southern-California Bight and possible mechanisms
of their formation and maintenance. Oceanol. Acta 4: 23–
32. http://archimer.ifremer.fr/doc/00121/23207/
Xue et al. Phytoplankton vertical nonuniform distribution
15

D’Alimonte, D., G. Zibordi, T. Kajiyama, and J. F. Berthon.
2014. Comparison between MERIS and regional high-level
products in European seas. Remote Sens. Environ. 140:
378–395. doi:10.1016/j.rse.2013.07.029
Dennison, P. E., K. Q. Halligan, and D. A. Roberts. 2004. A
comparison of error metrics and constraints for multiple
endmember spectral mixture analysis and spectral angle
mapper. Remote Sens. Environ. 93: 359–367. doi:10.1016/
j.rse.2004.07.013
Duan, H., R. Ma, Y. Zhang, S. A. Loiselle, J. Xu, C. Zhao, L.
Zhou, and L. Shang. 2010. A new three-band algorithm
for estimating chlorophyll concentrations in turbid inland
lakes. Environ. Res. Lett. 5: 044009. doi:10.1088/1748-
9326/5/4/044009
Duan, H., R. Ma, and C. Hu. 2012. Evaluation of remote
sensing algorithms for cyanobacterial pigment retrievals
during spring bloom formation in several lakes of East
China. Remote Sens. Environ. 126: 126–135. doi:10.1016/
j.rse.2012.08.011
Forget, P., P. Broche, and J. J. Naudin. 2001. Reflectance sen-
sitivity to solid suspended sediment stratification in coast-
al water and inversion: A case study. Remote Sens.
Environ. 77: 92–103. doi:10.1016/S0034-4257(01)00197-3
Fournier, G. R., and J. L. Forand. 1994. Analytic phase func-
tion for ocean water. Proc. SPIE Int. Soc. Opt. Eng. 2258:
194–201. doi:10.1117/12.190063
Gholamalifard, M., A. Esmaili-Sari, A. Abkar, B. Naimi, and
T. Kutser. 2013. Influence of vertical distribution of phy-
toplankton on remote sensing signal of case II waters:
Southern Caspian Sea case study. J. Appl. Remote Sens. 7:
073550–073550. doi:10.1117/1.JRS.7.073550 doi:10.1117/
1.JRS.7.073550
Gordon, H. R., and W. Mccluney. 1975. Estimation of the
depth of sunlight penetration in the sea for remote sens-
ing. Appl. Opt. 14: 413–416. doi:10.1364/AO.14.000413
Gordon, H. R., and D. K. Clark. 1980. Remote sensing optical
properties of a stratified ocean: An improved interpreta-
tion. Appl. Opt. 19: 3428–3430. doi:10.1364/AO.19.003428
He, X., Y. Bai, D. Pan, N. Huang, X. Dong, J. Chen, C. T. A.
Chen, and Q. Cui. 2013. Using geostationary satellite
ocean color data to map the diurnal dynamics of sus-
pended particulate matter in coastal waters. Remote Sens.
Environ. 133: 225–239. doi:10.1016/j.rse.2013.01.023
Hidalgo-Gonzalez, R. M., and S. Alvarez-Borrego. 2001. Chlo-
rophyll profiles and the water column structure in the
Gulf of California. Oceanol. Acta 24: 19–28. doi:10.1016/
S0399-1784(00)01126-9
HydroScat-6 spectral backscattering sensor & fluorometer
user’s manual (revision J). 2010. Hydro-Optics, Biology &
Instrumentation Laboratories: Tucson, AZ, USA.
Klemer, A., J. Feuillade, and M. Feuillade. 1982. Cyanobacte-
rial blooms: Carbon and nitrogen limitation have oppo-
site effects on the buoyancy of Oscillatoria. Science 215:
1629–1631. doi:10.1126/science.215.4540.1629
Kromkamp, J., A. Van Den Heuvel, and L. R. Mur. 1989. For-
mation of gas vesicles in phosphorus-limited cultures of
Microcystis aeruginosa. J. Gen. Microbiol. 135: 1933–1939.
doi:10.1099/00221287-135-7-1933
Kruse, F. A., A. B. Lefkoff, J. W. Boardman, K. B.
Heidebrecht, A. T. Shapiro, P. J. Barloon, and A. F. H.
Goetz. 1993. The spectral image processing system
(SIPS)—interactive visualization and analysis of imaging
spectrometer data. Remote Sens. Environ. 44: 145–163.
doi:10.1016/0034-4257(93)90013-N
Kutser, T., L. Metsamaa, N. Str€ombeck, and E. Vahtm€ae.
2006. Monitoring cyanobacterial blooms by satellite
remote sensing. Estuar. Coast. Shelf Sci. 67: 303–312. doi:
10.1016/j.ecss.2005.11.024
Kutser, T., L. Metsamaa, and A. G. Dekker. 2008. Influence
of the vertical distribution of cyanobacteria in the water
column on the remote sensing signal. Estuar. Coast. Shelf
Sci. 78: 649–654. doi:10.1016/j.ecss.2008.02.024
Lee, Z., K. L. Carder, and R. A. Arnone. 2002. Deriving inher-
ent optical properties from water color: A multiband
quasi-analytical algorithm for optically deep waters. Appl.
Opt. 41: 5755–5772. doi:10.1364/AO.41.005755
Letelier, R. M., and M. R. Abbott. 1996. An analysis of
chlorophyll fluorescence algorithms for the moderate
resolution imaging spectrometer (MODIS). Remote
Sens. Environ. 58: 215–223. doi:10.1016/S0034-
4257(96)00073-9
Lin, Y., Y. Gao, Y. Lu, L. Zhu, Y. Zhang, and Z. Chen. 2012.
Study of temperature sensitive optical parameters and
junction temperature determination of light-emitting
diodes. Appl. Phys. Lett. 100: 202108. doi:10.1063/
1.4718612 doi:10.1063/1.4718612
Lodhi, M., and D. Rundquist. 2001. A spectral analysis of
bottom-induced variation in the colour of Sand Hills
lakes, Nebraska, USA. Int. J. Remote Sens. 22: 1665–1682.
doi:10.1080/01431160117495
Loisel, H., and D. Stramski. 2000. Estimation of the inherent
optical properties of natural waters from the irradiance
attenuation coefficient and reflectance in the presence of
Raman scattering. Appl. Opt. 39: 3001–3011. doi:10.1364/
AO.39.003001
Ma, R., J. Tang, and J. Dai. 2006. Bio-optical model with
optimal parameter suitable for Taihu Lake in water colour
remote sensing. Int. J. Remote Sens. 27: 4305–4328. doi:
10.1080/01431160600857428
Ma, R., Q. Song, G. Li, and D. Pan. 2008. Estimation of back-
scattering probability of Lake Taihu waters. J. Lake Sci.
20: 375–379. doi:10.18307/2008.0318
Ma, R., H. Duan, Q. Liu, and S. A. Loiselle. 2011a. Approxi-
mate bottom contribution to remote sensing reflectance
in Taihu Lake, China. J. Great Lakes Res. 37: 18–25. doi:
10.1016/j.jglr.2010.12.002
Ma, R., G. Jiang, H. Duan, L. Bracchini, and S. Loiselle.
2011b. Effective upwelling irradiance depths in turbid
Xue et al. Phytoplankton vertical nonuniform distribution
16

waters: A spectral analysis of origins and fate. Opt. Express
19: 7127–7138. doi:10.1364/oe.19.007127
Mill�an-N�u~nez, R., S. Alvarez-Borrego, and C. C. Trees. 1997.
Modeling the vertical distribution of chlorophyll in the
California Current System. J. Geophys. Res. 102: 8587.
doi:10.1029/97JC00079 doi:10.1029/97JC00079
Mitchell, B. G. 1990. Algorithms for determining the
absorption-coefficient of aquatic particulates using the
quantitative filter technique (QFT). Proc. SPIE Int. Soc.
Opt. Eng. 1302: 137–148. doi:10.1117/12.21440
Mobley, C. D. 1994. Light and water: Radiative transfer in
natural waters. Academic Press.
Mobley, C. D. 1999. Estimation of the remote-sensing reflec-
tance from above-surface measurements. Appl. Opt. 38:
7442–7455. doi:10.1364/AO.38.007442
Mobley, C. D., L. K. Sundman, and B. Emmanuel. 2002.
Phase function effects on oceanic light fields. Appl. Opt.
41: 1035–1050. doi:10.1364/AO.41.001035
Morel, A. 1974. Optical properties of pure water and pure
seawater, p. 1–24. In N. G. Jerlov, E. Steemann Nielsen,
[eds.], Optical Aspects of Oceanogrpahy. Academic, San
Diego, California.
Morel, A., and L. Prieur. 1977. Analysis of variations in
ocean color1. Limnol. Oceanogr. 22: 709–722. doi:
10.4319/lo.1977.22.4.0709
Morel, A., and J. F. Berthon. 1989. Surface pigments, algal
biomass profiles, and potential production of the eupho-
tic layer: Relationships reinvestigated in view of remote-
sensing applications. Limnol. Oceanogr. 34: 1545–1562.
doi:10.4319/lo.1989.34.8.1545
Mueller, J. L., and others. 2003. Ocean optics protocols for
satellite ocean color sensor validation, revision 5, volume
V: Biogeochemical and bio-optical measurements and
data analysis protocols, p. 36. NASA Tech. Memo.
211621.
Nanu, L., and C. Robertson. 1993. The effect of suspended
sediment depth distribution on coastal water spectral
reflectance: Theoretical simulation. Int. J. Remote Sens.
14: 225–239. doi:10.1080/01431169308904334
Odermatt, D, F. Pomati, J. Pitarch, J. Carpenter, M. Kawka,
M. Schaepman, and A. W€uest. 2012. MERIS observations
of phytoplankton blooms in a stratified eutrophic lake.
Remote Sens. Environ. 126: 232–239. doi:10.1016/
j.rse.2012.08.031
Pope, R. M., and E. S. Fry. 1997. Absorption spectrum
(380–700 nm) of pure water. II. Integrating cavity measure-
ments. Appl. Opt. 36: 8710–8723. doi:10.1364/
AO.36.008710
Silulwane, N. F., A. J. Richardson, F. A. Shillington, and B. A.
Mitchell-Innes. 2010. Identification and classification of
vertical chlorophyll patterns in the Benguela upwelling
system and Angola-Benguela front using an artificial neu-
ral network. S. Afr. J. Mar. Sci. 23: 37–51. doi:10.2989/
025776101784528872
Snyder, W. A., and others. 2008. Optical scattering and back-
scattering by organic and inorganic particulates in US coastal
waters. Appl. Opt. 47: 666–677. doi:10.1364/AO.47.000666
Sokoletsky, L. G., and Y. Z. Yacobi. 2011. Comparison of chlo-
rophyll a concentration detected by remote sensors and
other chlorophyll indices in inhomogeneous turbid waters.
Appl. Opt. 50: 5770–5779. doi:10.1364/AO.50.005770
Song, K., L. Li, Z. Li, L. Tedesco, B. Hall, and K. Shi. 2013a.
Remote detection of cyanobacteria through phycocyanin
for water supply source using three-band model. Ecol.
Inform. 15: 22–33. doi:10.1016/j.ecoinf.2013.02.006
Song, K., and others. 2013b. Remote estimation of
chlorophyll-a in turbid inland waters: Three-band model
versus GA-PLS model. Remote Sens. Environ. 136: 342–
357. doi:10.1016/j.rse.2013.05.017
Stramska, M., and D. Stramski. 2005. Effects of a nonuniform
vertical profile of chlorophyll concentration on remote-
sensing reflectance of the ocean. Appl. Opt. 44: 1735–
1747. doi:10.1364/AO.44.001735
Tsujimura, S., H. Tsukada, H. Nakahara, T. Nakajima, and M.
Nishino. 2000. Seasonal variations of Microcystis popula-
tions in sediments of Lake Biwa, Japan. Hydrobiologia
434: 183–192. doi:10.1023/A:1004077225916
Walsby, A., and M. Booker. 1980. Changes in buoyancy
of a planktonic blue-green alga in response to light
intensity. Brit. Phycol. J. 15: 311–319. doi:10.1080/
00071618000650321
Wynne, T. T., R. P. Stumpf, M. C. Tomlinson, and J. Dyble.
2010. Characterizing a cyanobacterial bloom in western
Lake Erie using satellite imagery and meteorological data.
Limnol. Oceanogr. 55: 2025–2036. doi:10.4319/
lo.2010.55.5.2025
Xue, K., Y. Zhang, H. Duan, R. Ma, S. Loiselle, and M.
Zhang. 2015. A remote sensing approach to estimate ver-
tical profile classes of phytoplankton in a eutrophic lake.
Remote Sens. 7: 14403–14427. doi:10.3390/rs71114403
Yang, Q., D. Stramski, and M. X. He. 2013. Modeling the
effects of near-surface plumes of suspended particulate
matter on remote-sensing reflectance of coastal waters.
Appl. Opt. 52: 359–374. doi:10.1364/AO.52.000359
Yu, T. 2010. Phytoplankton community structure in Chaohu
Lake. Master’s thesis. Anhui Univ.
Zaneveld, J. R., A. Barnard, and E. Boss. 2005. Theoretical
derivation of the depth average of remotely sensed optical
parameters. Opt. Express 13: 9052–9061. doi:10.1364/
OPEX.13.009052
Zhang, Y., B. Zhang, X. Wang, J. Li, S. Feng, Q. Zhao, M.
Liu, and B. Qin. 2007. A study of absorption characteris-
tics of chromophoric dissolved organic matter and par-
ticles in Lake Taihu, China. Hydrobiologia 592: 105–120.
doi:10.1007/s10750-007-0724-4
Zhang, Y., R. Ma, M. Zhang, H. Duan, S. Loiselle, and J. Xu.
2015. Fourteen-year record (2000–2013) of the spatial and
temporal dynamics of floating algae blooms in Lake
Xue et al. Phytoplankton vertical nonuniform distribution
17

Chaohu, observed from time series of MODIS images.
Remote Sens. 7: 10523–10542. doi:10.3390/rs70810523
Zhou, Y., Y. Zhang, K. Shi, C. Niu, X. Liu, and H. Duan.
2015. Lake Taihu, a large, shallow and eutrophic aquatic
ecosystem in China serves as a sink for chromophoric dis-
solved organic matter. J. Great Lakes Res. 41: 597–606.
doi:10.1016/j.jglr.2015.03.027
Acknowledgments
The authors would like to thank the colleagues from NIGLAS (Jing Li,Min Tao, Jinghui Wu, Zhaojun Nai, Zhigang Cao, Qichun Liang, MingShen, and Yixuan Zhang) for their help with field measurements and
sample collections. Financial support was provided by State Key Program
of National Natural Science of China (41431176), National Natural Sci-ence Foundation of China (41471287), and National High Technology
Research and Development Program of China (2014AA06A509).
Conflict of Interest
None declared.
Submitted 01 June 2016
Revised 23 August 2016
Accepted 17 November 2016
Associate editor: Xiao Hua Wang
Xue et al. Phytoplankton vertical nonuniform distribution
18