Embed Size (px)
Citation preview

Light-Harvesting Bioconjugates asChloroplast Mimics
A thesis submitted in partial fulfilment of the
requirements for admission to the degree of
Doctor of Philosophy
by
David Hvasanov
School of Chemistry
The University of New South Wales
February 2013




Table of Contents
Preface i
Acknowledgements iv
Abstract vi
1 Introduction1.1 Bioconjugation Background 1-4
1.2 Natural Photosystems for Energy Conversion 1-10
1.3 Nanoreactors and Organelles 1-13
1.4 Artificial Cells 1-17
1.5 Artificial Chloroplasts 1-21
1.6 Project Description and Goal 1-25
1.7 References 1-28
2 Synthesis of Ru(II)-Complexes and Quinone Derivatives2.1 Methods For Functionalising 2,2’:6’,2”-Terpyridines 2-2
2.2 Functionalised 4’-Aryl Terpyridines 2-6
2.2.1 Synthesis of nitro-phenyl-terpyridine 3 2-6
2.2.2 Synthesis of amino-phenyl-terpyridine 4 2-7
2.3 Functionalised Ru(II)-Bisterpyridine Complexes 2-8
2.3.1 Synthesis of [Ru(tpy)]Cl3 5 and [Ru(tpy)2](PF6)2 6 2-10
2.3.2 Synthesis of [Ru(tpy)(4’-(4-aminophenyl)-2,2’:6’,2”-tpy)]
(PF6)2 7 2-12
2.3.3 Synthesis of [Ru(tpy)(maleimide-hexylcarboxamido-
phenyl-tpy)](PF6)2 8 2-14
2.3.4 Synthesis of [Ru(4’-(4-aminophenyl)-2,2’:6’2’’-
terpyridine)2](PF6)2 9 2-16
2.3.5 Synthesis of [Ru(4’-(4-maleimide-hexylcarboxyamido-
phenyl)-2,2’:6’2’’-terpyridine)2](PF6)2 10 2-17
2.3.6 Spectroscopic properties of Ru(II)-bistepyridine complexes 2-18
2.3.7 Crystallography 2-20
2.4 Functionalised Anthraquinone-Based Acceptors 2-25

2.4.1 Synthesis of 1-amino-3-azidopropane 11 2-26
2.4.2 Synthesis of anthraquinone-2-azidopropylamide 13 2-26
2.4.3 Attempted synthesis of anthraquinone-2-propylamido-
triazole-maleimide 15 2-29
2.5 Conclusions 2-30
2.6 References 2-31
3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives3.1 Purification of Cytochrome c 3-2
3.1.1 Purification of iso-1 cytochrome c using cation
exchange chromatography 3-3
3.2 Bioconjugation Methods 3-5
3.2.1 Modification of histidine 3-5
3.2.2 Modification of lysine 3-5
3.2.3 Modification of cysteine 3-6
3.3 Bioconjugation of Ru(II)-cyt c (8-cyt c) 3-7
3.4 Synthesis of Dimeric Bioconjugates 3-13
3.4.1 Synthesis of cyt c-10-BSA 3-14
3.4.2 Synthesis of cyt c-10-cyt c 3-19
3.4.3 Synthesis of cyt c-16-GFP 3-22
3.4.4 Synthesis of BSA-10-BSA 3-25
3.4.5 Effect of charge on protein dimer yield 3-27
3.5 Conclusion and Future Work 3-30
3.6 References 3-31
4 Green Fluorescent Protein as a Light-Induced Electron Donor4.1 Background 4-2
4.2 Synthesis of GFP-Acceptor Bioconjugates 4-5
4.2.1 Attempted synthesis of anthraquinone-triazole-GFP
via click chemistry 4-6
4.2.2 Synthesis of anthraquinone-GFP (12-GFP) via amine modification 4-9
4.2.3 Synthesis of viologen-GFP (16-GFP) 4-12
4.3 GFP Donor-Acceptor Studies 4-14

4.3.1 Steady-state spectroscopy studies 4-16
4.3.2 Fluorescence lifetime studies 4-20
4.4 Conclusion 4-25
4.5 References 4-26
5 Supramolecular Aggregates for Protein Encapsulation5.1 Liposomes 5-2
5.1.1 Liposome formation and characterisation 5-4
5.1.2 Enzyme encapsulation 5-11
5.2 Polymersomes 5-16
5.2.1 Aggregate formation using the ‘thermodynamic trapping’
method 5-18
5.2.2 Polymersome formation using the ‘kinetic trapping’ method 5-19
5.3 Conclusion 5-34
5.4 References 5-36
6 Photoinduced Electron Transfer Studies of Cytochrome c6.1 Background 6-2
6.2 Room Temperature Photoinduced Electron Transfer Studies 6-4
6.2.1 Biological activity using cytochrome c oxidase assay 6-12
6.3 Nitrite Reductase Mimics 6-14
6.3.1 Photoinduced nitrite reductase activity of cytochrome c 6-16
6.4 Conclusion and Future Work 6-21
6.5 References 6-22
7 Self-Assembled Light-Driven Proton Pumping Studies7.1 Background 7-2
7.2 Photosynthetic-Respiratory Hybrid System 7-6
7.2.1 Polymersome morphologies and membrane reconstitution 7-9
7.2.2 Photoinduced pH gradient 7-12
7.2.3 Orientation of reconstituted cytochrome c oxidase 7-19
7.2.4 Dependency of proton translocation rates on pH 7-20
7.2.5 Proton pumping quantum efficiencies ( ) 7-21

7.3 Conclusion and Future Work 7-22
7.4 References 7-24
8 Experimental8.1 Chemicals, Equipment and General Methods 8-2
8.2 X-ray Crystallography 8-9
8.3 Synthesis of Terpyridine Chromophores 8-10
8.4 Synthesis of Anthraquinone-Based Acceptors 8-17
8.5 Synthesis and Purification of Bioconjugates 8-20
8.6 Vesicle Formation and Encapsulation 8-27
8.7 Enzyme Activity and Photoreaction Experiments 8-31
8.8 References 8-36
9 Conclusions and Future Work9.1 References 9-9
AppendicesAppendix A – X-ray Crystallography
Appendix B – Bioconjugate Yield Determination
Appendix C – Standard Nitrite Curve
Appendix D – Encapsulation Efficiency
Appendix E – 8-hydroxypyrene-1,3,6-trisulfonic acid (HPTS)
Appendix F – Quantum Yield and Proton Pumping Rates
Appendix G – Abbreviations

i
Preface
The work described in this Thesis was performed in the School of Chemistry at the
University of New South Wales during the candidature. All results discussed herein are
my own work except where specific reference to the work of others is made in the text.
Some of the work in this Thesis has been published in the following journals/books:
"Induced polymersome formation from a diblock PS-b-PAA polymer via encapsulation
of positively charged proteins and peptides", David Hvasanov, Jörg Wiedenmann, Filip
Braet and Pall Thordarson, Chem. Commun., 2011, 6314-6316.
"Light-activated Bioconjugate Complexes", David Hvasanov, Daniel C. Goldstein and
Pall Thordarson, In Molecular Solar Fuels, The Royal Society of Chemistry, 2012, p
426-447.
Some of the work included in this Thesis has also been presented at the following
scientific conferences:
"Membrane-Bound Light Harvesting Bioconjugates as Chloroplast Mimics", David
Hvasanov, Joshua R. Peterson and Pall Thordarson, Light in Life Sciences Conference
(LILS2009), Melbourne, Australia, 24-27 November 2009. (Poster Presentation).

ii
"Membrane-Bound Light Harvesting Bioconjugates as Chloroplast Mimics", David
Hvasanov, Joshua R. Peterson and Pall Thordarson, The Royal Australian Chemical
Institute Annual One-Day Symposium, The University of Sydney, Sydney, Australia, 3
December 2009. (Poster Presentation). Awarded poster prize.
"Membrane-Bound Light Harvesting Bioconjugates as Chloroplast Mimics", David
Hvasanov, Joshua R. Peterson and Pall Thordarson, International Conference on
Nanoscience and Nanotechnology (ICONN2010), Sydney, Australia, 22-26 February
2010. (Poster Presentation).
"Light-harvesting Bioconjugates as Chloroplast Mimics", David Hvasanov, Joshua R.
Peterson, Jörg Wiedenmann and Pall Thordarson, The Royal Australian Chemical
Institute Annual One-Day Symposium, The University of Wollongong, Wollongong,
Australia, 1 December 2010. (Oral Presentation).
"Light-harvesting Bioconjugates as Chloroplast Mimics", David Hvasanov, Joshua R.
Peterson, Jörg Wiedenmann and Pall Thordarson, The 2010 International Chemical
Congress of Pacific Basin Societies (Pacifichem2010), Honolulu, USA, 15-20
December 2010. (Poster Presentation). Awarded poster prize.
"Photoinduced membrane proton pumping via polymersomes as chloroplast mimics",
David Hvasanov, Joshua R. Peterson, Filip Braet, Jörg Wiedenmann and Pall
Thordarson, The Royal Australian Chemical Institute Annual One-Day Symposium, The
University of New South Wales, Sydney, Australia, 30 November 2011. (Poster
Presentation).

iii
"Photoinduced membrane proton pumping via polymersomes as chloroplast mimics",
David Hvasanov, Joshua R. Peterson, Filip Braet, Jörg Wiedenmann and Pall
Thordarson, 2012 International Symposium on Macrocyclic and Supramolecular
Chemistry (ISMSC-7), Dunedin, New Zealand, 29 January-2 February 2012. (Poster
Presentation). Awarded poster prize.
"Photoinduced membrane proton pumping via polymersomes as chloroplast mimics",
David Hvasanov, Joshua R. Peterson, Filip Braet, Jörg Wiedenmann and Pall
Thordarson, 243rd American Chemical Society National Meeting, San Diego, USA, 25-
29 March 2012. (Poster Presentation).

iv
Acknowledgements
Firstly, I would like to thank my supervisor, Dr. Pall Thordarson, for all of his guidance,
support and enthusiasm through my PhD candidacy. I can't thank him enough for giving
me inspiring ideas during difficult periods of my candidature and always willing to look
over a presentation or article. I would also like to thank my co-supervisor, Prof. Martina
Stenzel, who provided guidance in polymer aggregate formation and access to her lab.
A special mention to Dr. Joshua R. Peterson and Dr. Shiva Prasad for their
extensive help and advice during the first year of this project. Thanks to all the members
of the Thordarson group for their helpful contributions, current members - Ethan Howe,
Scott Jamieson, Warren Truong, Ekaterina Nam, Andrew Robinson, James Webb, Alex
Mason, Lev Lewis and Alistair Laos - and past members - Dr. Sabrina Dehn, Dr. Daniel
C. Goldstein, Dr. Joris Meijer, Dr. Katie Tong, Luuk Olijve, Scott Jones, Lip-son Chin,
Katrine Qvortrup, Grace Lim, Haythim Hassanein, Dithepon Pornsaksit, Ben Lewis,
Tommy Pieszko, Dedrick Song and Carol Hua. Thanks also to Dr. Alex Falber for
adding colour to the lab.
Thanks to Ethan Howe, Scott Jamieson, Warren Truong and Dr. Danmar Gloria
(surrogate group member) for letting me release stress when going out to coffee or rock
climbing. William Zhang, it's always interesting getting into discussions about quirky
topics.
Thanks to Dr. Ron Haines for keeping the office in order and Ian Aldred from
the Store. Thanks to the Centre for Macromolecular Design for letting me use their
dynamic light scattering instrument. Thanks to all the staff at the Mark Wainwright
Analytical Centre including, Lewis Alder, Dr. Leanne Stephenson and Dr. Anne Poljak
from the bioanalytical mass spectrometry facility; Dr. Mohan Bhadbhade (X-ray
crystallographer) from the solid state and elemental analysis unit; Katie Levick from the

v
electron microscope unit; Dr. Renee Whan and Dr. Henry Haeberle from the biomedical
imaging facility and all the staff at the NMR facility. I would like to thank Dr. Dylan
Owen for initial fluorescence microscopy studies. I thank A/Prof. Filip Braet and
Delfine Cheng at the Australian Centre for Microscopy and Microanalysis, The
University of Sydney for cryo-TEM analysis. I thank A/Prof. Timothy W. Schmidt, Dr.
Raphaël G. C. R. Clady and Mr. Murad Tayebjee at The University of Sydney for
time-resolved fluorescence lifetime studies and Dr. Jörg Wiedenmann for providing the
GFP and plasmids.
I am grateful to both the University of New South Wales and the Australian
Research Council for financial support of this work. I would also like to thank the
Graduate Research School for a Research Travel Award, which allowed me to travel to
Honolulu to present my work at an international conference.
Finally, I must thank my family and friends for all of their love and support
during my candidature.

vi
Abstract
Cells are highly complex bio-nanoreactors comprised of a complex medium where
multiple multistep reactions occur simultaneously across the cell. To prevent
interference and degradation of these catalytic pathways, cells compartmentalise.
Compartmentalisation achieves control of synthetic pathways at specific sites in the cell
(organelles). In nature, compartmentalisation is demonstrated in plants by the organelle,
chloroplast, which is responsible for photosynthesis. Photosynthesis is one of the most
important biological reactions, responsible for sustaining life.
This Thesis describes the preparation of light-activated donor-acceptor
bioconjugates for use as artificial chloroplasts. The studies were aimed at the
development of synthetic compartments for the reconstitution of light-harvesting
bioconjugates to construct a semi-synthetic electron transport chain capable of
generating a transmembrane proton gradient using light. To achieve that end,
bioconjugates were formed using the redox metalloprotein yeast iso-1 cytochrome c
from Saccharomyces cerevisiae and photoactive maleimide functionalised
ruthenium(II)-bisterpyridine chromophores for attachment to a free single cysteine in
iso-1 cytochrome c (CYS102). The resulting photoactive donor-acceptor bioconjugate is
capable of undergoing electron transfer upon photoactivation using 480 nm light.
The synthesis of a number of terpyridines, ruthenium(II)-bisterpyridine
complexes and maleimide functionalised ruthenium(II)-bisterpyridine complexes for use
as photoactive electron donors were prepared to investigate light-activated electron
transfer. The synthesis of asymmetric maleimide complex was achieved by initially
preparing 4’-(4-aminophenyl)-2,2’:6’,2’’-terpyridine ligand (4) synthetically based on
the modified Kröhnke method. Following from the amine 4'-aryl functionalisation, the
synthesis of asymmetric complex [Ru(tpy)(4’-(4-aminophenyl)-2,2’:6’,2”-tpy)](PF6)2

vii
(7) was achieved using methods adapted from the synthesis of iridium(III)-bisterpyridine
related complexes, followed by maleimide attachment using peptide coupling
chemistry, producing [Ru(tpy)(maleimide-hexylcarboxamido-phenyl-tpy)](PF6)2
complex (8) to allow modification with iso-1 cytochrome c. The bismaleimide complex
equivalent 10, was prepared using a similar approach to allow the preparation of dimeric
bioconjugates. Asymmetric donor-acceptor bioconjugate was prepared from complex 8.
Similarly, bismaleimide complex 10 and bismaleimide viologen 16 were used to
prepare high molecular weight bioconjugates using combinations of single cysteine
residue proteins including iso-1 cytochrome c (cyt c), bovine serum albumin (BSA) and
green fluorescent protein (GFP) to explore factors affecting yield. The resulting
bioconjugates 8-cyt c, cyt c-10-cyt c, cyt c-10-BSA, cyt c-16-GFP and BSA-10-BSA
were purified using chromatographic techniques including immobilised metal affinity
chromatography (IMAC), size exclusion chromatography (SEC) or strong cation
exchange chromatography (CEX). Interestingly, electrostatic interactions played a
significant role in dimer bioconjugate yields. The 80 kDa heterodimer of
complementary charge, cyt c-10-BSA, resulted in a 30% yield. However, like-charged
homo/heterodimer bioconjugates resulted in low yields of less than 1%.
Characterisation of bioconjugates by UV-Vis, MALDI-TOF mass spectroscopy and gel
electrophoresis were exploited to demonstrate attachment and assess purity.
The alternative biological donor-synthetic acceptor system was also explored
using the light-induced electron donor GFP. Synthesis of a GFP bioconjugate as a
potential covalent donor-acceptor system based on the
N-hydroxysuccinimide-anthraquinone 12 acceptor was prepared using non-specific
lysine modification. Additionally, the GFP was site-specifically functionalised with
viologen 16 as an alternative potential donor-acceptor bioconjugate. Light-induced

viii
electron transfer studies between GFP and p-benzoquinone, anthraquinone and viologen
electron acceptors as both non-covalent and bioconjugate mixtures were monitored
using steady-state techniques including UV-Vis and fluorescence spectroscopy and
performed using a Xenon arc lamp or custom built LED array. Time-resolved
fluorescence lifetime measurements were exploited to measure and confirm
electron/energy transfer processes of GFP-acceptor systems. Typically, a ket of the order
109 s-1 for the rate of electron transfer between GFP and the quinone acceptors was
observed.
In order to compartmentalise the ruthenium(II)-cytochrome c bioconjugates to
construct artificial organelles, polymersomes (polymer vesicles) were formed based on
a novel method developed in this project based on the polyelectrolyte,
polystyrene140-b-poly(acrylic acid)48 (PS140-b-PAA48). Polymersomes could be induced
in the presence of positively charged biomolecules including cyt c and GFP,
concomitantly encapsulating the biomolecules in the membrane. In the presence of
simple salts or negatively charged biomolecules such as calmodulin, micelles form.
Liposomes (lipid vesicles) based on egg L- -phosphatidylcholine or synthetic
L-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine were explored, however, their use as
compartments were limited due to poor encapsulation efficiencies.
Photoinduced electron transfer studies between Ru(II)-bisterpyridine complexes
and iso-1 cytochrome c (cyt c) as non-covalent mixtures and asymmetric bioconjugate
8-cyt c were studied. Electron transfer studies were carried out as bulk solution
bioconjugates and membrane encapsulated 8-cyt c. The electron transfer processes
resulting in reduction of the heme of cytochrome c is followed by steady-state UV-Vis
spectroscopy. Electron transfer to the heme was achieved by irradiation using a 465 nm
LED light under anaerobic conditions at room temperature in the presence of sacrificial

ix
electron donor, ethylenediaminetetraacetic acid (EDTA). In addition, light-induced
electron transfer studies between a non-covalent mixture of Ir(III)-bisterpyridine
complex and horse heart cytochrome c was studied to mimic nitrite reductase behaviour.
Excitation and catalysis was performed using a 372 nm LED source under anaerobic
conditions at room temperature in the presence of 100 nm liposomes.
A primitive synthetic chloroplast capable of converting light energy into
chemical energy by generating a proton gradient upon photo-excitation was developed.
An artificial photosynthetic-respiratory hybrid was achieved by replacing chlorophyll
(photosensitiser) with 8-cyt c. This light-harvesting bioconjugate as well as its natural
electron acceptor, cytochrome c oxidase, is encapsulated in the synthetic polymersome
membrane of the diblock copolymer PS140-b-PAA48, reminiscent of a photosynthetic
membrane construct. The successful self-assembly of this complex light-harvesting
enzyme cascade within membranes of polymeric vesicles were characterised using
confocal laser-scanning microscopy (CLSM), TEM and cryo-TEM. Upon
light-activation, cytochrome c oxidase (reduced) is capable of pumping protons across
the membrane to generate an electrochemical gradient which is detected using a
fluorescent pH dye encapsulated within the aqueous core of the polymersome. Upon
irradiation at an initial pH of 7.2, a proton pumping rate of 3.3 × 103 H+/s across the
polymer bilayer generating a gradient up to pH 0.2 was observed.

Chapter 1
Introduction

Chapter 1 Introduction
1-2
Reproduced from New Scientist, 14 April 2012, Turn a new leaf cover story, p 28.

Chapter 1 Introduction
1-3
1 Introduction
This Thesis describes the development of a primitive chloroplast – a system that
converts light energy into chemical energy in the form of an electrochemical gradient
( ). Such a system can in principle be used as a “nanoreactor battery” where the light
induced proton gradient is used to drive chemical reactions or biochemical processes.
In plants, photosynthesis is carried out in organelles known as chloroplasts,
which converts light energy into chemical energy in the form of adenosine triphosphate
(ATP).1 Photosynthesis is highly important as it provides the majority of biomass on
Earth and produces oxygen to sustain life. This process is achieved by photoinduced
generation of an electron-hole pair followed by rapid charge transfer across the electron
transport chain coupled with proton translocation across a membrane, which drives the
production of ATP.2
Biological compartments (organelles) in nature allow chemical reactions to
occur in a confined space which couple reactions in time and space resulting in enzyme
cascade systems.3 This ensures that the product of a chemical conversion is the substrate
or catalyst of the subsequent process. Compartmentalisation achieves well-defined
reaction environments which vary from simple nanometer-sized systems to complex
micrometer-sized assemblies, while isolating catalytic cycles preventing degradation by
proteases and controls the flux of molecules in or out of compartments.4 Synthetic
compartments based on polymer or phospholipid vesicles have been heavily studied in
literature to create nanoreactors.3,5
Inspired by nature, this Thesis aims to develop an artificial photosynthetic
system capable of converting light energy into a stored proton gradient. The system
combines facets of photosynthetic concepts including the construction of a photoactive
hybrid electron transport chain capable of proton translocation which is reconstituted

Chapter 1 Introduction
1-4
using synthetic compartments based on amphiphilic polymer vesicles (polymersomes).6
The key component of the artificial hybrid photosynthetic-respiratory system is a
donor-acceptor bioconjugate linking an electron donating photosensitiser to a redox
protein and electrostatically coupled to mitochondrial cytochrome c oxidase for
vectorial proton translocation.
The development of artificial photosynthetic systems to generate proton
gradients upon photoexcitation have been previously reported using synthesised organic
donor-acceptor triads7 or aryl diimides8 in phospholipid vesicles or reconstitution of a
light-driven transmembrane proton pump (bacteriorhodopsin) in a polymersome
membrane.9 However, issues of these systems still exist including stability,
reproducibility and robustness. This Thesis seeks to address these issues by developing
methods capable of preparing light-activated bioconjugates as a component of a
semi-artificial photosynthetic-respiratory electron transport chain and methods for
concomitant reconstitution of both hydrophilic and hydrophobic enzymes in membrane
domains of polymersomes.
1.1 Bioconjugation Background
Proteins play an important role in nature; they are responsible for the enzymatic
conversions of nearly all processes in living organisms, including DNA replication and
photosynthesis. In addition, they have structural roles, acting as scaffolds in cells to
support and protect tissues and organisms. Furthermore, proteins are involved in cell
signalling within and between cells which regulate the functions of cells via signal
transduction. Commonly, protein functions depend on post-translational modifications

Chapter 1 Introduction
1-5
after expression such as glycosylation which allows for proper folding and increased
stability.†
Scientists have long sought to understand and exploit the properties of proteins
by incorporating synthetic molecules to introduce and enhance novel (unnatural)
functionalities. This has led to the development of biotechnological, biomedical and
pharmaceutical industry related devices such as biosensors10, bioelectronics11, biofuel
cells12 and bioconjugated therapeutic proteins/drugs.13 In this Thesis, this exploitation is
essential towards the development of a primitive chloroplast, as this will allow
researchers to mix and match the best from synthetic and biological light-harvesters
(chromophores) and electron donor/acceptor systems.
Bioconjugation can be defined as the linking of two or more components to form
a novel complex with the combined properties and structural characteristics of the
individual component molecules.14 In general, bioconjugation combines a biological
component (protein, biomolecule) with a synthetic component creating a chimeric
system resulting in a novel class of biomaterials to perform novel functions.
Theoretically, the properties of the bioconjugate is the linear combination of the
properties of the multi-component system.15 This allows bioconjugates to overcome
their intrinsic limitations and to possess properties and functions which otherwise would
not exist in nature.
The advent of bioconjugation methods over the last few decades has led to the
development of different chemoselective conjugation methods.16 Bioconjugation
chemistry has contributed to significant research progress and understanding in the field
of life sciences. Through careful modification of proteins, scientists have been able to
understand protein structure, functions and interactions.17
† Parts of this work have been published: Hvasanov, D.; Goldstein, D. C.; Thordarson, P. In Molecular Solar Fuels; The Royal Society of Chemistry, 2012.

Chapter 1 Introduction
1-6
One of the most extensively studied areas of protein conjugates is PEGylation,
as it allows for enhancement of peptides and proteins for pharmacological and
therapeutic use in vivo and in vitro since it was first demonstrated in 1977.18 Attachment
of the hydrophilic poly(ethylene glycol) (PEG) polymer via PEGylation has led to
improved targeting and tissue penetration for protein therapeutics in the biomedical
industry.19
Biomolecules other than proteins such as low molecular weight co-factors and
ligands are also common targets for bioconjugation and functionalisation. These often
include porphyrins (heme)20, NAD(P)+21, FAD22, lipids23, sugars and oligosaccharides24,
nucleic acids and nucleotides25 and biotin.26 However, these targets are beyond the
scope of this project.
Bioconjugation reactions involving chemical modifications and chemical
cross-linking are often divided into classes of reactions. These can be separated into
three generic types of functionalisation approaches; including direct functionalisation,
indirect functionalisation and functionalisation with cofactors/ligands as shown in
Figure 1.1.15
Direct functionalisation involves chemically modifying a synthetic ligand
possessing a specific functional group with a protein of interest. Indirect
functionalisation requires bioconjugation between the synthetic or biomolecule
components via a spacer containing heterobifunctionality. Finally, indirect
functionalisation is achieved by reaction of a synthetic ligand with a cofactor, which is
then reconstituted into an apoprotein through self-association (ligand binding).

Chapter 1 Introduction
1-7
Figure 1.1. Schematic for the classes of protein functionalisation. (a) direct functionalisation. (b) indirect functionalisation. (c) indirect functionalisation with a cofactor/ligand.27
Proteins and enzymes are most commonly modified by chemical modification
between a synthetic ligand and a targeted amino acid residue. These residues often
include lysine (Lys), cysteine (Cys), histidine (His), tyrosine (Tyr) and glutamine (Gln).
The most common method for chemical modification in bioconjugate chemistry is the
covalent linkage between a ligand and the amines of a protein. Coupling to the -amino
group on a lysine residue is often targeted as they are abundant on the surface of
proteins as well as the -N terminus of a protein. They can be easily modified by
reacting with an N-hydroxysuccimide (NHS) activated ester.15,28 It should be noted,
chemical bond formation between NHS-esters and amines offer poor site-specificity as
the natural occurrence of lysine residues in mammalian proteins is estimated to be
approximately 6%.29

Chapter 1 Introduction
1-8
Another popular target for chemical modification of natural amino acids is
cysteine (cys). Bioconjugation with proteins are generally dependent on nucleophilic
addition or displacement reaction mechanisms of the amino acid residue with an
activated synthetic ligand.17 Based on the theory of nucleophilicity proposed by
Edwards and Pearson30, the thiol group of cysteine is one of the strongest nucleophiles
in proteins. Cysteine residues possess thiol groups with a pKa of approximately 9.14 Due
to these properties, specific chemical modifications of cysteines can proceed selectively
and rapidly under benign conditions.15 Common modification reagents used in the
literature include haloalkyl compounds, thiosulfonates and maleimides. However,
maleimides are ideal as they allow protein modification at neutral pH (6.8-7) coupled
with control of specificity. They can be used to chemically modify cysteines as
maleimide-functionalised ligands are Michael acceptors.31
Cysteine residues are rare in proteins and often only contain a single accessible
residue available for modification. Alternatively, proteins which lack surface cysteine
residues can be introduced via mutagenesis for site-specific modification.32 This is a
major advantage for synthesis of site-specific bioconjugates when targeting natural
amino acids. Furthermore, maleimides are suitable for site-specific bioconjugation as
they specifically react with cysteines at neutral pH and only side-react with other amino
acids above pH 8.15
As a result of the abundance of natural amino acids in proteins, synthetic
biologists and chemists have developed techniques to introduce non-natural amino
acids, allowing for chemoselective bioconjugation reactions. These non-natural amino
acids include azides and alkynes which can be introduced selectively into proteins by
semisynthetic or recombinant methods for [3+2] Huisgen cycloaddition reactions.33

Chapter 1 Introduction
1-9
The methods described above have been used to synthesise light-activated donor
acceptor systems as described in Chapter 3, 6 and 7 for photoinduced electron transfer
measurements. Similarly, electron transfer studies of metal complex modified redox
proteins have been explored over several decades by Millett and Gray.34
One of the approaches to develop a photoactive bioconjugate by Pan and
co-workers is the modification of lysine residues of horse heart cytochrome c with a
Ru(II) bipyridine photoactive chromophore.34e The luminescence decay rates of the
resulting bioconjugates were found to be between (3.5-21) × 106 s-1, indicating possible
electron transfer from the excited triplet state of ruthenium to the ferric heme group of
cytochrome c.
Peterson et al.35 in the Thordarson group have conjugated Ru(II) bisterpyridine
chromophores with yeast iso-1 cytochrome c creating a light-activated donor-acceptor
bioconjugate via a long and short chain spacer as shown in Figure 1.2. Bioconjugates
were modified using maleimide functionalised ligands with the CYS102 residue
through a Michael addition reaction. Electron transfer rates of resulting short and long
chain bioconjugates were 5.95 × 105 and 2.78 × 105 s-1, respectively.
Figure 1.2. Photoinduced electron transfer of Ru(II)-bisterpyridine cytochrome cbioconjugate.27

Chapter 1 Introduction
1-10
1.2 Natural Photosystems for Energy Conversion
Significant breakthroughs have been made in literature to mimic photosynthesis by
extracting natural photosystems from photosynthetic organisms which are covalently
bioconjugated to nanoparticles or semiconductors to produce hydrogen or electricity,
respectively, upon light-activation.
In an elegant approach, Grimme et al.36 have developed a light-harvesting
system resulting in chemical transformation by producing hydrogen as shown in Figure
1.3. They have utilised a biological electron donor, photosystem I (PSI) found in
photosynthetic organisms and conjugated it to a platinum or gold terminal electron
acceptor nanoparticle. Conjugation was initiated by formation of [4Fe-4S] clusters in
solution by reacting sodium sulphide, ferrous ammonium sulphate and mercaptoethanol.
The resulting cluster was then reconstituted into the PSI stromal protein PsaC to yield
holo-C13G/C33S variant PsaC. PSI was then modified by combining reconstituted PsaC
and P700/Fx cores in the presence of PsaD. Finally, 1,6-hexanedithiol was introduced to
displace the mercaptoethanol ligand from the [4Fe-4S] cluster and to covalently link
PSI and Pt/Au nanoparticle. This resulting PSI photoactive bioconjugate evolves
hydrogen upon white light illumination using a Xenon arc lamp. The resulting chemical
transformation is due to the formation of a charge-separated state when PSI absorbs
wavelengths below 700 nm which is then transferred to the nanoparticle with a redox
potential favourable for hydrogen production. In a recent paper, optimisation of this
system has been reported, maximising H2 production in a platinum-based PSI
bioconjugate.37

Chapter 1 Introduction
1-11
Figure 1.3. Photosystem I/nanoparticle bioconjugate evolving hydrogen upon light-activation.36
Iwuchukwu et al.38, have utilised similar concepts by exploiting natural photosystems in
conjunction with nanoparticles to induce hydrogen production after irradiation. The
photosystem I (PSI)/Platinum (Pt) nanoparticle bioconjugate as shown in Figure 1.4 was
prepared by self-assembled platinisation on the stromal surface of PSI. Upon irradiation,
the system allows electron transport from cytochrome c6 to PSI and finally to the Pt
catalyst which produces hydrogen gas with a 25-fold greater yield than current
biomass-to-fuel strategies.

Chapter 1 Introduction
1-12
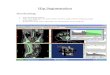
Figure 1.4. Photosystem I/Platinum nanoparticle bioconjugate produce hydrogen gas due to electron transport from cytochrome c6 to photosystem I and finally platinum nanoparticles upon irradiation.38
Other than hydrogen production, photosystem hybrids have been used to generate
electricity for the application of photoelectrochemical cells. In a sophisticated system,
Ham et al.39, have developed a regenerative photoelectrochemical complex capable of
reversible solar energy conversion. As shown in Figure 1.5, the photoelectrochemical
complex involves single-walled carbon tubes which act as a support for self-assembly
of lipid bilayers with membrane scaffold proteins to form nanodiscs, which allow
reconstitution of the photosynthetic reaction centres for electricity generation through
the carbon nanotubes upon irradiation. This system allows self-healing by induced
disassembly upon addition of surfactants and reassembly upon its removal over an
indefinite number of cycles. Similarly, Govorov has studied photosystems for electricity
production using a simpler system by incorporating PSI to a semiconductor nanoparticle
(CdTe) via bioconjugation.40

Chapter 1 Introduction
1-13
Figure 1.5. Reversible self-assembly of photosynthetic reaction centres on single-walled carbon nanotubes via nanodiscs for photoinduced electrical current generation.39
1.3 Nanoreactors and Organelles
Despite efforts to mimic photosynthetic reactions by exploiting photosystems from
natural sources, these systems are not representative of chloroplast organelles as they
are free floating systems in solution. In contrast, living organisms perform reactions and
chemical conversions in a confined environment in order to create a highly organised
reaction space which organises metabolic processes.6 This shows that
compartmentalisation is essential to life and also as proposed by Szotzak et al.41,
compartmentalisation played a crucial role in the origin of life by allowing molecules to
be kept in close proximity and allowing advantageous mutations during replication,
leading to evolution. Cells and organelles perform crucial roles to maintain life
including (1) chemical conversions using enzymes, (2) self-reproducing vesicles, (3)
self-reproducing genetic information and (4) energy storage. In this section, the
developments made towards artificial compartments for chemical reactions as
nanoreactors are discussed.
Since the introduction of liposomes (lipid vesicles) by Bangham et al.42 in 1965
for the preparation of artificial compartments, enzyme encapsulation within the interior
of liposomes have been predominantly employed for protein therapeutic applications43
and drug delivery.44 However, attempts toward nanoreactors using phospholipid

Chapter 1 Introduction
1-14
vesicles have been limited for compartmentalised chemical reactions due to hindered
bilayer permeability preventing diffusion of substrate/product molecules in and out of
the vesicles.45 Walde et al.46 prepared 1-palmitoyl-2-oleoyl-sn-glycero-3-
phosphocholine (POPC) liposomes encapsulating water soluble proteinase
-chymotrypsin in the aqueous interior. This system demonstrated selectivity for small
external substrates including benzoyl-L-Tyr-p-nitroanilide and
acetyl-L-Phe-p-nitroanilide for which the membrane was permeable, allowing enzyme
hydrolysis of the substrate to produce p-nitroaniline. Subsequently, Blocher et al.47
expanded on the system to model the enzyme reactions for encapsulated
-chymotrypsin.
In recent years, there has been growing interest in polymersomes for
compartmentalisation due to their improved stabilities, allowing vesicles to remain
stable for weeks to months rather than hours to days for liposomes.48 Similar to
liposomal systems, polymersomes have been used for drug delivery49 and protein
therapeutic applications for biomedicine.50 However, polymersomes based on block
copolymers are ideal candidates for nanoreactors due to the tunability of monomer
blocks allowing control of permeability51, glass transition temperature (Tg)52 and
membrane thickness.53
In an elegant system, Vriezema et al.54 described an enzyme cascade system
based on polystyrene40-b-poly(L-isocyanoalanine(2-thiophen-3-yl-ethyle)amide)50
(PS-PIAT) which possessed intrinsic membrane permeability due to PIAT forming rigid
rod-like helices during self-assembly, allowing diffusion of small substrates/products in
and out of the water pool. A three enzyme cascade system was prepared by
encapsulation of glucose oxidase (GOX) in the lumen, horse radish peroxidase (HRP) in
the membrane and Candida antarctica Lipase B (CALB) in the bulk solution. Addition

Chapter 1 Introduction
1-15
of substrate 1,2,3,4-tetra-O-acetyl- -glucopyranose (GAc4) resulted in hydrolysis of the
acetate groups by CALB in the bulk solution to produce glucose. Subsequently, glucose
diffused into the water pool following the tandem reaction as shown in Figure 1.6 and
the cascade reaction produced a coloured radical cation product (ABTS +) which was
detected spectroscopically. Extensions of this work have been attempted including
encapsulation of chloroperoxidase55 or covalently linking CALB to polymersomes via
an azide-alkyne click reaction.56
Figure 1.6. Schematic representation of a three-step enzyme cascade system allowing the multistep reaction in PS-PIAT polymersomes.54
Meier et al.57 have investigated nanoreactors by reconstitution of channel proteins with
significant focus on porin OmpF from outer cell walls of Gram-negative bacteria.
Rather than diblock copolymers, the Meier group use triblock copolymers as they
predominantly favour vesicle formation and the polymer used possess low Tg, allowing
extrusion to size polymersomes analogous to liposomal preparation.58 Porin OmpF was
reconstituted in poly(2-methyloxazoline)-b-poly(dimethylsiloxane)-b-poly(2-
methyloxazoline) (PMOXA-PDMS-PMOXA) polymersome membranes which act as a

Chapter 1 Introduction
1-16
size-selective filter allowing passage of molecules below 400 Da while -lactamase was
encapsulated in the interior volume.59 The nanoreactor containing -lactamase
hydrolysed ampicillin (diffused into the interior from the bulk through OmpF) into
ampicillinoic acid which reduces iodine to iodide as shown in Figure 1.7. The reaction
was monitored by micro-iodometry via the decolourisation of a starch-iodine complex.
Variations of this system have been made including the use of channel protein FhuA60,
pH-switchability61, surface immobilisation62 and reduction triggered release4
nanoreactors.
Figure 1.7. Representation of a nanoreactor containing -lactamase and membrane reconstituted porin OmpF from an amphiphilic triblock copolymer.59
Stimuli responsive polymersome nanoreactors have also been explored to control
membrane permeability. Polymersomes were formed in a two component mixture
poly(ethylene glycol)-b-polystyrene:poly(ethylene glycol)-b-poly(styrene boronic acid)
(PEG-b-PS:PEG-b-PSBA) with the minor component PEG-b-PSBA being sensitive to

Chapter 1 Introduction
1-17
pH and monosaccharides resulting in disassembly. CALB was encapsulated in the
lumen and enzymatic reactions resulting in hydrolysis of DiFMU octanoate or
p-nitrophenyl acetate substrates was initiated by pore formation after addition of
D-glucose or increasing pH.63
Other than liposomes and polymersomes, there has been growing interest in
nanocapsules64 and capsosomes65 as alternative nanoreactors. Of particular interest are
capsosomes pioneered by the Caruso group due to their capsule:liposome hybrid nature
allowing a microreactor with the possibility of thousands of sub-compartments.66 It
should be noted that specific control of individual sub-compartments still remains a
challenge.
1.4 Artificial Cells
Significant research has been undertaken to investigate the use of compartments for
nanoreactors and enzyme cascades. Another facet of mimicking cells and organelles is
the development of artificial cells using liposomes and polymersomes for genetic
information replication and membrane reproduction. These studies have been conducted
to explore the origin of life using low molecular weight amphiphiles.5b,67
In a landmark article by Szostak et al.41, they proposed three requirements for a
model protocell as (1) bearing an informational substance (DNA or RNA), (2) a catalyst
and (3) a compartment. Although not explicitly mentioned, the requirement for energy
production in a model protocell system is essential to drive these processes. To better
understand the origin of life, two strategies can be employed including the top-down
(minimal cell) and the bottom-up (model protocell) approach.5b Hutchinson et al.68 have
attempted the synthesis of a minimal cell by stripping non-essential cellular components
of a Mycoplasma genitalium bacterium while keeping the cell living. It has been

Chapter 1 Introduction
1-18
predicted that the theoretical minimal number of genes required while maintaining
cellular function is approximately 150.69 However, this system is still complex as
hundreds of genes and thousands of different proteins and molecules remain. In order to
synthesise an artificial cell de novo or better understand the origin of life on Earth,
simpler systems should be studied using the bottom-up approach.41
In order to create a living artificial cell from the bottom-up strategy, it requires a
self-replicating vesicle and a replicase (RNA) with RNA-coded activity to allow
synthesis of amphiphilic lipids for compartment division as shown in Figure 1.8.41 This
allows the 'organism' to be self-sustaining and replicate with evolutionary optimisation
as RNA molecules can be encapsulated spontaneously from the surrounding
environment.
Figure 1.8. Outline of a proposed pathway for the synthesis of a living artificial cell. System composed of RNA replicase and self-replicating vesicle with an RNA-coded linking function for lipid synthesis via a ribozyme.41

Chapter 1 Introduction
1-19
Steps towards the development of a living artificial cell have been attempted. To date,
the focus has been mostly on the replication of the vesicle or the genetic information.
Self-reproducing vesicles were first reported by Walde et al.70 in 1994 using vesicles
composed of fatty oleic and caprylic acids. Growth and reproduction was achieved
when a fatty acid precursor (corresponding anhydride) was added to the solution and
subsequently hydrolysed. Sugawara and co-workers have since reported on
self-reproducing vesicles based on artificially developed amphiphiles.71 Giant vesicles
were composed of amphiphiles with an imine group in its hydrophobic chain which are
formed by a dehydrocondensation reaction between the amphiphilic aldehyde and a
lipophilic aniline derivative as shown in Figure 1.9. When the protected aldehyde
precursor was added to the vesicle suspension containing the lipophilic imine and a
catalyst, dehydrocondensation between the two precursors occurred within the interior
pool producing further amphiphiles, contributing to growth of small vesicles and
eventual exocytosis from the parent giant vesicle.
Figure 1.9. Self-reproducing giant vesicles: (i) protected aldehyde precursor (A') is incorporated into the vesicle composed of V and catalyst C produces reactive precursor A; (ii) A reacts with lipophilic imine (B) within the vesicle to form amphiphilic molecule V via dehydrocondensation reaction; (iii) new vesicles composed of V formed; (iv) exocytosis of new vesicles to the bulk water.71

Chapter 1 Introduction
1-20
In a pioneering investigation, Oberholzer et al.72 replicated a RNA strand using all four
nucleosides and a viral RNA polymerase (Q -replicase) in an oleate/oleic acid small
vesicle (diameter <100 nm) capable of vesicle reproduction upon addition of oleic
anhydride precursor. In an elegant system, Kurihara et al.73 have extended this system
by demonstrating self-reproduction of giant vesicles based on artificial amphiphiles
with concomitant amplification of encapsulated DNA (more robust than RNA) as
shown in Figure 1.10. It was shown that amplification of DNA within a
self-reproducible cationic giant vesicle resulted in distribution of the DNA to the
daughter giant vesicles due to electrostatic interactions between DNA and membrane.
Interestingly, it was reported that the amplification of the encapsulated DNA accelerated
the rate of vesicle division.
Figure 1.10. Schematic representation of the amplification of DNA and self-reproduction of giant vesicles.73
As mentioned earlier, it is essential that an ideal model protocell system can produce
energy to drive processes such as replication of DNA/RNA, catalytic functions or
amphiphile synthesis for vesicle replication. Nearly all organisms on Earth source
energy directly or indirectly from the Sun, with the exception of chemoautotrophs that
live in rocks or deep sea hydrothermal vents.74 In plant based photosynthetic organisms,

Chapter 1 Introduction
1-21
this process occurs in the chloroplast and is therefore also an interesting biomimetic
target for artificial cells.
1.5 Artificial Chloroplasts
To a lesser extent, the biomimicry of chloroplasts have been investigated using
synthetic compartments for photoinduced energy storage as an electrochemical gradient.
In a pioneering investigation by Steinberg-Yfrach et al.7a, a synthetic photosynthetic
reaction centre was prepared by linking a tetraarylporphyrin to a naphthoquinone
moiety fused to a norbornene system bearing a carboxylic acid group and a carotenoid
polyene. This molecular 'triad' was incorporated into the bilayer of a
L- -phosphatidylserine and dioleoylphosphatidylcholine (2:3 molar ratio) liposome.
Upon photoexcitation, the triad undergoes photoinduced electron transfer to generate an
intermediate charge-separated species with a reduction potential near the outer surface
and an oxidation potential near the inner surface of the bilayer membrane. A freely
diffusing quinone molecule alternates between its oxidised and reduced forms due to the
induced redox potential allowing passive transport of protons across the membrane
resulting in acidification of the interior as shown in Figure 1.11. An overall quantum
yield of 0.004 was reported. This work has been extended by incorporating CF0F1-ATP
synthase to catalyse the light-driven synthesis of ATP due to the proton gradient.7b

Chapter 1 Introduction
1-22
Figure 1.11. Schematic representation of photoinduced proton translocation across the liposome bilayer.7a
Bhosale et al.8 have investigated transmembrane proton gradients by incorporating rigid
p-octiphenyl rods to create helical tetrameric -stacks of napthalene diimides that can
span egg yolk lipid bilayer membranes as shown in Figure 1.12. The liposomes used
encapsulated quinone molecules as electron acceptors and the bulk solution contained
ethylenediaminetetraacetic acid as electron donors. Upon irradiation, electron transfer
through the helical stacks internally reduced quinone molecules which subsequently
consumed protons leading to basification of the lumen. Of interest, external ligand
intercalation transformed the photoactive scaffolds into ion channels.

Chapter 1 Introduction
1-23
Figure 1.12. Schematic representation of helical tetrameric -stacks of naphthalene diimides in lipid bilayers that can generate a transmembrane proton gradient upon irradiation. Addition of a ligand transforms the photoactive scaffold into ion channels.8
In another elegant system, biosynthesis of ATP was achieved by reconstitution of a
light-driven proton pump (bacteriorhodopsin) and F0F1-ATP synthase in
PEtOz-PDMS-PEtOz triblock copolymer polymersomes.9 Reconstitution of the proton
pump results in an influx of protons into the vesicle which drives the formation of ATP
as shown in Figure 1.13. Montemagno and co-workers fine-tuned the insertion process
to allow orientational control of bacteriorhodopsin reconstitution.

Chapter 1 Introduction
1-24
Figure 1.13. Schematic representation of proteopolymersomes reconstituted with bacteriorhodopsin (BR) and F0F1-ATP synthase capable of ATP synthesis upon irradiation.9

Chapter 1 Introduction
1-25
1.6 Project Description and Goal
The goal of this project is to develop light-active bioconjugates and incorporate them
into vesicular membranes to create a light-harvesting electron transport chain capable of
generating a transmembrane proton gradient. The light-activation would be a means to
mimic chloroplasts by photoinducing the proton gradient. The design for a
light-activated bioconjugate and the primitive chloroplast is shown in Figure 1.14.
Figure 1.14. Schematic of proposed project goal. (a) a primitive chloroplast capable of generating a transmembrane proton gradient by reconstitution of a light-harvesting bioconjugate with natural electron acceptor and proton pump, cytochrome c oxidase (CcOx). (b) light-harvesting bioconjugate, ruthenium(II)-bisterpyridine cytochrome c(Ru-cyt c).

Chapter 1 Introduction
1-26
The enzyme cytochrome c was chosen for this system due to several advantages it offers
including: (1) a redox metalloprotein, (2) it has been intensively studied and its structure
and photophysical properties are well-known75, (3) it has characteristic UV-Vis
absorption changes corresponding to reduced and oxidised states as well as changes to
protein structure35b, (4) applicable to various bioconjugation techniques because it is
stable in an array of reaction conditions and is highly soluble35b,76, (5) can be purified
readily in large quantities and (6) low cost.
In particular, the iso-1 form of yeast cytochrome c is attractive as a target as it
possesses a single cysteine residue that allows site-specific modification under benign
aqueous conditions using maleimides to form a stable thioester.15 The position of the
cysteine residue is on the opposite side of the active site for cytochrome c oxidase
binding and therefore allows modification without adversely affecting biological and
catalytic function. It has been reported that the cysteine residue is located in a
predominantly hydrophobic pocket which is less accessible to solvents and results in
slower reaction rates, however, reaction conditions have been previously optimised in
the Thordarson group.35b The bioconjugation of iso-1 cytochrome c and a brief review
of bioconjugate techniques and methods have been discussed further in Chapter 3.
Ruthenium(II)-bisterpyridine chromophores have been heavily studied for both
the methods of their synthesis and photophysical properties.77 Ruthenium terpyridine
complexes are often compared to the related bipyridine moieties in terms of their
photophysical properties. Tris(bipyridine)ruthenium(II) complexes are often used for
photophysical studies due to high quantum yields, long-lived luminescence lifetimes
and luminescence can be measured at room temperature.78 Ruthenium bipyridine
complexes have been reported to exhibit room temperature quantum yields and
luminescence lifetimes of 0.04 and 0.6 s, respectively.79 On the other hand, ruthenium

Chapter 1 Introduction
1-27
terpyridine complexes display a lack of room temperature luminescence and low yields
with values of 1 ns and 10-5, respectively, which require quantum yield and lifetime
studies to be performed at 77 K.78
Although ruthenium terpyridine complexes have short-lived room temperature
luminescence lifetimes, the light-harvesting chromophore chosen was based on these
bisterpyridine complexes. The complexes allow (1) assembly of ligands to form linear
complexes without introducing chirality (as observed with bipyridine complexes)77, (2)
the synthesis and photophysical properties of these complexes are heavily studied and
well-known78, (3) photoinduced electron transfer studies involving bisterpyridine
complexes as donor-acceptor systems have been reported80 and (4) electron transfer in
ruthenium(II)-bisterpyridine cytochrome c bioconjugates have been previously
reported.35a A brief review and methods of ruthenium terpyridine complex synthesis is
discussed further in Chapter 2.
As an extension to the synthetic donor-biological acceptor concept used here,
the opposite biological donor-synthetic acceptor motif was also investigated. Following
recent reports by Bogdanov et al.81 that green fluorescent protein (GFP) may act as a
light-induced electron donor, a GFP-synthetic acceptor bioconjugate was constructed to
investigate electron transfer of these donor-acceptor constructs which is discussed
further in Chapter 4.
The use of liposomes and polymersomes has been heavily studied for enzyme
encapsulation.5b,43b In particular, polymersomes based on the diblock copolymer
polystyrene-b-poly(acrylic acid) (PS-b-PAA) was used as the compartments to mimic
chloroplasts in this Thesis. PS-b-PAA offers the following advantages: (1) well studied
polymers for aggregate formation52, (2) improved stabilities and robustness compared to
liposomes48 and (3) morphologies can be fine-tuned by the addition of additives or

Chapter 1 Introduction
1-28
preparation method.82 A brief review of lipid and polymer vesicles and their methods of
preparation are further discussed in Chapter 5.
The photoinduced electron transfer studies of ruthenium(II)-bisterpyridine
cytochrome c bioconjugates and the effect of membrane encapsulation is described in
Chapter 6. Finally, the combination of the light-activated bioconjugate, proton pump
(cytochrome c oxidase) and reconstitution in a polymer membrane to construct a
semi-synthetic electron transport chain for photoinduced transmembrane proton
gradients is described in Chapter 7.
1.7 References
(1) Purves, W. K.; Sadava, D.; Orians, G. H.; Heller, C. H. Life: The Science of Biology; 7th ed.; Sinauer Associates and W. H. Freeman: U.S.A., 2003.
(2) Cordes, M.; Giese, B. Chem. Soc. Rev. 2009, 38, 892.(3) Vriezema, D. M.; Comellas Aragonès, M.; Elemans, J. A. A. W.; Cornelissen, J.
J. L. M.; Rowan, A. E.; Nolte, R. J. M. Chem. Rev. 2005, 105, 1445.(4) Onaca, O.; Sarkar, P.; Roccatano, D.; Friedrich, T.; Hauer, B.; Grzelakowski,
M.; Güven, A.; Fioroni, M.; Schwaneberg, U. Angew. Chem. Int. Ed. 2008, 47,7029.
(5) (a) Lensen, D.; Vriezema, D. M.; van Hest, J. C. M. Macromol. Biosci. 2008, 8,991. (b) Roodbeen, R.; van Hest, J. C. M. BioEssays 2009, 31, 1299.
(6) Peters, R. J. R. W.; Louzao, I.; van Hest, J. C. M. Chem. Sci. 2012, 3, 335.(7) (a) Steinberg-Yfrach, G.; Liddell, P. A.; Hung, S.-C.; Moore, A. L.; Gust, D.;
Moore, T. A. Nature 1997, 385, 239. (b) Steinberg-Yfrach, G.; Rigaud, J.-L.; Durantini, E. N.; Moore, A. L.; Gust, D.; Moore, T. A. Nature 1998, 392, 479.
(8) Bhosale, S.; Sisson, A. L.; Talukdar, P.; Fürstenberg, A.; Banerji, N.; Vauthey, E.; Bollot, G.; Mareda, J.; Röger, C.; Würthner, F.; Sakai, N.; Matile, S. Science2006, 313, 84.
(9) Choi, H.-J.; Montemagno, C. D. Nano Lett. 2005, 5, 2538.(10) Wang, J. Chem. Rev. 2008, 108, 814.(11) Willner, I. Science 2002, 298, 2407.(12) Willner, I.; Yan, Y.-M.; Willner, B.; Tel-Vered, R. Fuel Cells 2009, 9, 7.(13) Ulbrich, K.; Strohalm, J.; Subr, V.; Plocova, D.; Duncan, R.; Rihova, B.
Macromol. Symp. 1996, 103, 177.(14) Hermanson, G. T. Bioconjugate Techniques; 2nd ed.; Elsevier Inc.: San Diego,
CA, 2008.(15) Thordarson, P.; Droumaguet, B. L.; Velonia, K. Appl. Microbiol. Biotechnol.
2006, 73, 243.(16) (a) Hentschel, J.; ten Cate, M. G. J.; Börner, H. G. Macromolecules 2007, 40,
9224. (b) Wang, C.; Stewart, R. J.; KopeCek, J. Nature 1999, 397, 417.

Chapter 1 Introduction
1-29
(17) Lundblad, R. L. Chemical reagents for protein modification; 3rd ed.; CRC Press: Boca Raton, FL, 2005.
(18) Abuchowski, A.; Van Es, T.; Palczuk, N. C.; Davis, F. F. J. Biol. Chem. 1977,252, 3578.
(19) Veronese, F. M.; Morpurgo, M. Il Farmaco 1999, 54, 497.(20) Hamachi, I.; Se; ijlig; Shinkai, i. Eur. J. Org. Chem. 1999, 1999, 539.(21) Bückmann, A. F.; Carrea, G. Adv. Biochem. Eng. Biotechnol. 1989, 39, 97.(22) Bückmann, A. F.; Wray, V.; Stocker, A. Methods Enzymol. 1997, 280, 360.(23) Kung, V. T.; Maxim, P. E.; Veltri, R. W.; Martin, F. J. Biochim. Biophys. Acta
1985, 839, 105.(24) Pozsgay, V.; Vieira, N. E.; Yergey, A. Org. Lett. 2002, 4, 3191.(25) Niemeyer, C. M.; Sano, T.; Smith, C. L.; Cantor, C. R. Nucleic Acids Res. 1994,
22, 5530.(26) Wilbur, D. S.; Chyan, M.-K.; Pathare, P. M.; Hamlin, D. K.; Frownfelter, M. B.;
Kegley, B. B. Bioconjug. Chem. 2000, 11, 569.(27) Hvasanov, D.; Goldstein, D. C.; Thordarson, P. In Molecular Solar Fuels; The
Royal Society of Chemistry, 2012; p 426.(28) Veronese, F. M.; Pasut, G. Drug Discov. Today 2005, 10, 1451.(29) Wu, G.; Ott, T. L.; Knabe, D. A.; Bazer, F. W. J. Nutr. 1999, 129, 1031.(30) Edwards, J. O.; Pearson, R. G. J. Am. Chem. Soc. 1962, 84, 16.(31) Hodgson, D. R. W.; Sanderson, J. M. Chem. Soc. Rev. 2004, 33, 422.(32) Kanaya, S.; Kimura, S.; Katsuda, C.; Ikehara, M. Biochem. J. 1990, 271, 59.(33) (a) van Kasteren, S. I.; Kramer, H. B.; Jensen, H. H.; Campbell, S. J.;
Kirkpatrick, J.; Oldham, N. J.; Anthony, D. C.; Davis, B. G. Nature 2007, 446,1105. (b) de Graaf, A. J.; Kooijman, M.; Hennink, W. E.; Mastrobattista, E. Bioconjug. Chem. 2009, 20, 1281.
(34) (a) Winkler, J. R.; Nocera, D. G.; Yocom, K. M.; Bordignon, E.; Gray, H. B. J. Am. Chem. Soc. 1982, 104, 5798. (b) Meade, T. J.; Gray, H. B.; Winkler, J. R. J. Am. Chem. Soc. 1989, 111, 4353. (c) Mayo, S.; Ellis, W.; Crutchley, R.; Gray, H. Science 1986, 233, 948. (d) Gray, H. B.; Winkler, J. R. Annu. Rev. Biochem.1996, 65, 537. (e) Pan, L. P.; Durham, B.; Wolinska, J.; Millett, F. Biochemistry1988, 27, 7180. (f) Geren, L.; Hahm, S.; Durham, B.; Millett, F. Biochemistry1991, 30, 9450. (g) Millett, F.; Durham, B. Methods Enzymol. 2009, 456, 95. (h) Geren, L.; Durham, B.; Millett, F. Methods Enzymol. 2009, 456, 507.
(35) (a) Peterson, J. R.; Smith, T. A.; Thordarson, P. Chem. Commun. 2007, 1899. (b) Peterson, J. R.; Smith, T. A.; Thordarson, P. Org. Biomol. Chem. 2010, 8, 151.
(36) Grimme, R. A.; Lubner, C. E.; Bryant, D. A.; Golbeck, J. H. J. Am. Chem. Soc.2008, 130, 6308.
(37) Grimme, R. A.; Lubner, C. E.; Golbeck, J. H. Dalton Trans. 2009, 10106.(38) Iwuchukwu, I. J.; Vaughn, M.; Myers, N.; O'Neill, H.; Frymier, P.; Bruce, B. D.
Nat. Nano. 2010, 5, 73.(39) Ham, M.-H.; Choi, J. H.; Boghossian, A. A.; Jeng, E. S.; Graff, R. A.; Heller, D.
A.; Chang, A. C.; Mattis, A.; Bayburt, T. H.; Grinkova, Y. V.; Zeiger, A. S.; Van Vliet, K. J.; Hobbie, E. K.; Sligar, S. G.; Wraight, C. A.; Strano, M. S. Nat. Chem. 2010, 2, 929.
(40) Govorov, A. O. Adv. Mater. 2008, 20, 4330.(41) Szostak, J. W.; Bartel, D. P.; Luisi, P. L. Nature 2001, 409, 387.(42) Bangham, A. D.; Standish, M. M.; Watkins, J. C. J. Mol. Biol. 1965, 13, 238.

Chapter 1 Introduction
1-30
(43) (a) Gaspar, M. M.; Perez-Soler, R.; Cruz, M. E. M. Cancer Chemother. Pharmacol. 1996, 38, 373. (b) Walde, P.; Ichikawa, S. Biomol. Eng. 2001, 18,143.
(44) Soussan, E.; Cassel, S.; Blanzat, M.; Rico-Lattes, I. Angew. Chem. Int. Ed. 2009,48, 274.
(45) (a) Annesini, M. C.; Giorgio, L. D.; Marzio, L. D.; Finazzi-agrò, A.; Serafino, A. L.; Mossa, G. J. Liposome Res. 1993, 3, 639. (b) Cioci, F.; Lavecchia, R. J. Liposome Res. 1993, 3, 725. (c) Walde, P. Curr. Opin. Colloid Interface Sci.1996, 1, 638.
(46) Walde, P.; Marzetta, B. Biotechnol. Bioeng. 1998, 57, 216.(47) Blocher, M.; Walde, P.; Dunn, I. J. Biotechnol. Bioeng. 1999, 62, 36.(48) Yu, Y.; Zhang, L.; Eisenberg, A. Langmuir 1997, 13, 2578.(49) Ranquin, A.; Versées, W.; Meier, W.; Steyaert, J.; Van Gelder, P. Nano Lett.
2005, 5, 2220.(50) O’Neil, C. P.; Suzuki, T.; Demurtas, D.; Finka, A.; Hubbell, J. A. Langmuir
2009, 25, 9025.(51) de Hoog, H.-P. M.; Vriezema, D. M.; Nallani, M.; Kuiper, S.; Cornelissen, J. J.
L. M.; Rowan, A. E.; Nolte, R. J. M. Soft Matter 2008, 4, 1003.(52) Cameron, N. S.; Corbierre, M. K.; Eisenberg, A. Can. J. Chem. 1999, 77, 1311.(53) Ma, L.; Eisenberg, A. Langmuir 2009, 25, 13730.(54) Vriezema, D. M.; Garcia, P. M. L.; Sancho Oltra, N.; Hatzakis, N. S.; Kuiper, S.
M.; Nolte, R. J. M.; Rowan, A. E.; van Hest, J. C. M. Angew. Chem. Int. Ed.2007, 46, 7378.
(55) de Hoog, H. M.; Nallani, M.; Cornelissen, J. J. L. M.; Rowan, A. E.; Nolte, R. J. M.; Arends, I. W. C. E. Org. Biomol. Chem. 2009, 7, 4604.
(56) van Dongen, S. F. M.; Nallani, M.; Schoffelen, S.; Cornelissen, J. J. L. M.; Nolte, R. J. M.; van Hest, J. C. M. Macromol. Rapid Commun. 2008, 29, 321.
(57) Palivan, C. G.; Fischer-Onaca, O.; Delcea, M.; Itel, F.; Meier, W. Chem. Soc. Rev. 2012, 41, 2800.
(58) Nardin, C.; Widmer, J.; Winterhalter, M.; Meier, W. Eur. Phys. J. E 2001, 4,403.
(59) Nardin, C.; Thoeni, S.; Widmer, J.; Winterhalter, M.; Meier, W. Chem. Commun. 2000, 1433.
(60) Nallani, M.; Benito, S.; Onaca, O.; Graff, A.; Lindemann, M.; Winterhalter, M.; Meier, W.; Schwaneberg, U. J. Biotechnol. 2006, 123, 50.
(61) Broz, P.; Driamov, S.; Ziegler, J.; Ben-Haim, N.; Marsch, S.; Meier, W.; Hunziker, P. Nano Lett. 2006, 6, 2349.
(62) Grzelakowski, M.; Onaca, O.; Rigler, P.; Kumar, M.; Meier, W. Small 2009, 5,2545.
(63) Kim, K. T.; Cornelissen, J. J. L. M.; Nolte, R. J. M.; van Hest, J. C. M. Adv. Mater. 2009, 21, 2787.
(64) Kreft, O.; Prevot, M.; Möhwald, H.; Sukhorukov, G. B. Angew. Chem. Int. Ed.2007, 46, 5605.
(65) Städler, B.; Chandrawati, R.; Goldie, K.; Caruso, F. Langmuir 2009, 25, 6725.(66) Städler, B.; Chandrawati, R.; Price, A. D.; Chong, S.-F.; Breheney, K.; Postma,
A.; Connal, L. A.; Zelikin, A. N.; Caruso, F. Angew. Chem. Int. Ed. 2009, 48,4359.
(67) (a) Wick, R.; Luisi, P. L. Chem. Biol. 1996, 3, 277. (b) Blöchliger, E.; Blocher, M.; Walde, P.; Luisi, P. L. J. Phys. Chem. B. 1998, 102, 10383. (c) Walde, P.; Goto, A.; Monnard, P.-A.; Wessicken, M.; Luisi, P. L. J. Am. Chem. Soc. 1994,

Chapter 1 Introduction
1-31
116, 7541. (d) Mansy, S. S.; Schrum, J. P.; Krishnamurthy, M.; Tobe, S.; Treco, D. A.; Szostak, J. W. Nature 2008, 454, 122.
(68) Hutchison, C. A.; Peterson, S. N.; Gill, S. R.; Cline, R. T.; White, O.; Fraser, C. M.; Smith, H. O.; Craig Venter, J. Science 1999, 286, 2165.
(69) Forster, A. C.; Church, G. M. Mol. Syst. Biol. 2006, 2.(70) Walde, P.; Wick, R.; Fresta, M.; Mangone, A.; Luisi, P. L. J. Am. Chem. Soc.
1994, 116, 11649.(71) Takakura, K.; Toyota, T.; Sugawara, T. J. Am. Chem. Soc. 2003, 125, 8134.(72) Oberholzer, T.; Wick, R.; Luisi, P. L.; Biebricher, C. K. Biochem. Biophys. Res.
Commun. 1995, 207, 250.(73) Kurihara, K.; Tamura, M.; Shohda, K.-i.; Toyota, T.; Suzuki, K.; Sugawara, T.
Nat. Chem. 2011, 3, 775.(74) Nakagawa, S.; Takai, K. FEMS Microbiol. Ecol. 2008, 65, 1.(75) (a) Brayer, G. D.; Murphy, M. E. P. Cytochrome c: A Multidisciplinary
Approach; 1st ed.; University Science Books: Sausalito, California, 1996. (b) Louie, G. V.; Brayer, G. D. J. Mol. Biol. 1990, 214, 527.
(76) Peterson, J. R.; Thordarson, P. Chiang Mai J. Sci. 2009, 26, 236.(77) Hofmeier, H.; Schubert, U. S. Chem. Soc. Rev. 2004, 33, 373.(78) Sauvage, J. P.; Collin, J. P.; Chambron, J. C.; Guillerez, S.; Coudret, C.; Balzani,
V.; Barigelletti, F.; De Cola, L.; Flamigni, L. Chem. Rev. 1994, 94, 993.(79) Van Houten, J.; Watts, R. J. J. Am. Chem. Soc. 1976, 98, 4853.(80) Hamachi, I.; Matsugi, T.; Tanaka, S.; Shinkai, S. Bull. Chem. Soc. Jpn. 1996, 69,
1657.(81) Bogdanov, A. M.; Mishin, A. S.; Yampolsky, I. V.; Belousov, V. V.; Chudakov,
D. M.; Subach, F. V.; Verkhusha, V. V.; Lukyanov, S.; Lukyanov, K. A. Nat. Chem. Biol. 2009, 5, 459.
(82) Choucair, A.; Eisenberg, A. Eur. Phys. J. E: Soft Matter Biol. Phys. 2003, 10,37.

Chapter 2
Synthesis of Ru(II)-Complexes and Quinone Derivatives

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-2
2 Synthesis of Ru(II)-Complexes and Quinone Derivatives
In this Chapter, the synthesis of a number of terpyridine, ruthenium(II)-bisterpyridine
complexes and maleimide functionalised ruthenium(II)-bisterpyridine complexes for use
as photoactive electron donors are discussed. The compounds described herein were
synthesised for the attachment to cytochrome c to form light-activated bioconjugates, as
spacers for homo/hetero dimeric bioconjugates or reference compounds for studies
described elsewhere in this thesis. The 4’-aryl functionalised 2,2’:6’,2”-terpyridines
were synthesised using central ring-assembly methodologies based on literature
procedures.1 For this project, methods that lead to the formation of both
homo/heteroleptic complexes which allow asymmetric and dimeric bioconjugation are
desired.
Additionally, quinone derivatives, with focus on anthraquinone analogues were
synthesised as potential electron acceptors. Anthraquinone derivatives were synthesised
as constructs for covalent attachment to green fluorescent proteins (GFP) to form
potential light-activated bioconjugates. Analogues were synthesised using
N-hydroxysuccinimide/N,N’-dicyclohexylcarbodiimide (NHS/DCC) peptide coupling
chemistry2 to introduce azide functionality for attachment to alkyne-tagged GFP via
Huisgen azide-alkyne 1,3-dipolar cycloaddition.3
The procedures for synthesis of terpyridines, complexation and anthraquinone
derivatives are briefly reviewed in this chapter.
2.1 Methods For Functionalising 2,2’:6’,2”-Terpyridines
Terpyridines (tpy) were first discovered by Morgan and Burstall4 who reported its
synthesis as a byproduct in the synthesis of 2,2’-bipyridine (bpy) which was known 44
years earlier (Figure 2.1).5 The synthesis was achieved by heating pyridine with

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-3
anhydrous iron(III) chloride at 340 oC in an autoclave (50 atm) for 36 h. The authors
reported interesting spectroscopic properties of terpyridine with Fe(II) indicating metal
complex formation.
Figure 2.1. Structures of 2,2'-bipyridine (bpy) and 2,2':6',2"-terpyridine (tpy).
Since the first report of terpyridines, tremendous improvements in synthetic strategy to
optimise yields and routes towards site-specific functionalisation have occurred in the
last 80 years. Modern terpyridine synthesis can be categorised into two basic synthetic
routes, which are central ring assembly and cross-coupling procedures.1b,6
Ring assembly strategies toward terpyridine synthesis have been the most
predominant in recent decades. The most common methods developed within the ring
assembly route are the Kröhnke reaction7, Potts methodology8 and Jameson
methodology.9 The most well-known is the Kröhnke reaction (Scheme 2.1) which
involves Michael addition of N-heteropyridinium salts (L-1) with an , -unsaturated
enone (L-2) to yield a 1,5-dione intermediate. The intermediate is subsequently cyclised
using a nitrogen source such as ammonium acetate.10

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-4
Scheme 2.1. Kröhnke synthesis of 4’-functionalised 2,2’:6’,2”-terpyridine. (a) NH4OAc.
An alternative approach to synthesise functionalised terpyridines is the Potts
methodology (Scheme 2.2), also known as the -oxoketene dithioacetal methodology.
In general, the reaction between the enolate of 2-acetylpyridine with carbon disulfide
and methyl iodide gives the resulting -oxoketene dithioacetal (L-3) product.
Subsequent Michael addition with an additional 2-acetylpyridine gives thiol
functionalised 1,5-dione (L-4). Cyclisation is similarly achieved with a nitrogen source
such as ammonium acetate.11
Scheme 2.2. Potts synthesis of 4'-functionalised 2,2':6',2"-terpyridines. (a) i) t-BuOK, THF, ii) CS2, iii) MeI. (b) t-BuOK, 2-acetylpyridine. (c) NH4OAc, HOAc.
Lastly, a less frequently used method to synthesise terpyridines is the Jameson method
(Scheme 2.3), due to the lack of functionalisation ability. Jameson and Guise first

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-5
reacted 2-acetylpyridine with N,N-dimethylformamide dimethyl acetal to form the
enaminone (L-5). The enaminone (L-5) was condensed with a second equivalent of
2-acetylpyridine resulting in a loss of dimethylamine to yield the 1,5-dione (L-6) and
ring closed with ammonium acetate.9
Scheme 2.3. Jameson synthesis of 2,2’:6’,2”-terpyridine. (a) N,N-dimethylformamide dimethyl acetal. (b) 2-acetylpyridine, t-BuOK. (c) NH4OAc, HOAc.
The disadvantages of these ring assembly methods are that the final condensation step
yields tar-like by-products resulting in purification difficulties.1a Over the last decade,
cross-coupling procedures based on modern Pd(0)-catalysed coupling reactions6 have
improved efficiency and synthetic simplicity with substitution control over traditional
methods.12 Pd(0) catalysed cross-coupling procedures include Suzuki13, Negishi14 and
Stille15 couplings.
In this project, a step-wise modified Kröhnke synthesis is employed which
avoids the common drawback of tarry crude by-products and achieves straightforward
synthesis. Chemical modification at the 4’-position of the terpyridine core with aromatic
groups allows the introduction of functional groups while maintaining achirality,

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-6
improved yields16 and increased excited state lifetimes17 making them suitable
photoactive electron donors when coordinated to metal ions such as Ru(II).
2.2 Functionalised 4’-Aryl Terpyridines
2.2.1 Synthesis of nitro-phenyl-terpyridine 3
In order to synthesise functionalised terpyridines with higher yields and ease of
synthesis, nitro-phenyl-terpyridine 3 was synthesised based on a modified classical
Kröhnke synthesis. The modified Kröhnke synthesis is achieved via the azachalcone,
pryidinium iodide route (Scheme 2.4) for terpyridine 3 which prevents the formation of
crude tarry by-products and allows simplified purification procedures based on previous
alternative methods explored by the Thordarson group.18
As shown in Scheme 2.4, the formation of nitro-substituted terpyridine 3 was
carried out in a three-step convergent synthesis with isolation of the intermediates.
Synthesis of 4-nitro-2’-azachalcone 1 via Claisen-Schmidt condensation between one
equivalent of 4-nitrobenzaldehyde and 2-acetylpyridine in a mixture of aqueous sodium
hydroxide and methanol gave a moderate yield of 28%.19 Subsequently, 1-(2-oxo-2-(2-
pyridyl)ethyl)pyridinium iodide 2 was synthesised by addition of 2-acetylpyridine to a
warm solution (60 oC) of iodine in dry pyridine. The mixture was then heated at 100 oC
and gave acylpyridinium iodide 2 in good yield of 79%.20
Finally, reacting the precursors azachalcone 1 with pyridinium iodide 2 and
subsequent ring closure with ammonium acetate in refluxing dry methanol afforded
terpyridine 3 in 75% yield.20

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-7
Scheme 2.4. Synthesis of 4’-(4-nitrophenyl)-terpyridine 3 by the azachalcone/pyridinium iodide route. (a) MeOH, r.t., 29%. (b) dry pyridine, 60 -100 oC, 79%. (c) NH4OAc, dry MeOH, reflux, 75%.
2.2.2 Synthesis of amino-phenyl-terpyridine 4
In order to further functionalise terpyridine 3, an aniline functional group was
introduced to allow further chemical modification with peptide coupling chemistry to
introduce maleimide functionality. Nitro-phenyl-terpyridine 3 was reduced using
previously reported literature procedures according to Scheme 2.5.21

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-8
Scheme 2.5. Synthesis of 4’-(4-aminophenyl)-2,2’:6’,2”-terpyridine by reduction with hydrazine over a Pd/C catalyst. (a) 10% Pd/charcoal, hydrazine monohydrate, EtOH, reflux, 81%.
Terpyridine 3 was activated in refluxing absolute ethanol over a 10% Pd/charcoal
catalyst for 45 min and subsequently hydrazine monohydrate was added to generate the
reduced aniline terpyridine 4 in 81% yield. Upon reduction, the purple terpyridine 3
changed to a yellow crystalline solid 4. It should be noted that in order to maximise
product yield, workup of the crude should be filtered over celite while the mixture is
warm due to formation of microcrystalline solid 4 upon cooling leading to loss of
product.
2.3 Functionalised Ru(II)-Bisterpyridine Complexes
Mononuclear bisterpyridine complexes are well-known and documented in the
literature.1b,17 In general, metal complexes of the type [M(tpy)2]X2, where X = counter
ion (PF6-, Cl- or ClO4
-) are formed from low oxidation state d-block transition metals.
Traditionally, these complexes are prepared from Zn(II), Co(II), Cu(II), Ni(II) transition
metals and the stability of these metal bis-complexes are a result of the strong
metal-ligand (d- *) back-donation, combined with the dynamic chelate effect.22
Complexation with transition metals results in a distorted octahedral geometry23 due to
the hexa-coordinate nature of most d-block metals.

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-9
Synthetic strategies towards the assembly of complexes are dependent on the
type of desired complex (homo/heteroleptic). Homoleptic complexes can be synthesised
in a straightforward one-pot synthesis, where the terpyridine ligand is treated with the
desired metal ion in a 2:1 (ligand:metal) ratio. The resulting complexes can be purified
in a two-step procedure involving the precipitation of symmetric complex with counter
ions (NH4PF6 or TBA-Cl) followed by recrystallisation.24
On the other hand, the preparation of heteroleptic complexes requires a directed
strategy where the desired ligands are introduced in a two-step procedure to prevent
“scrambling” which involves the exchange of ligands. Directed synthesis requires the
formation of the mono-complex which forms a highly insoluble product and is not
characterised by 1H nuclear magnetic resonance (NMR) spectroscopy due to its
paramagnetic nature. The second ligand is then introduced to the mono-complex under
reductive conditions in an alcoholic solvent to yield the asymmetric complex.25 This
technique is generally only applicable to Ru(II), Ir(III) and Os(II) complexes as a result
of the weaker stabilities of other transition metal complexes at elevated temperatures
leading to scrambling.1b In general, heteroleptic complexes require purification over
silica or alumina using a highly polar mobile phase.25-26
Characterisation of metal bisterpyridine complexes can be achieved using a
combination of techniques including 1H NMR and UV-Vis spectroscopy as well as soft
mass spectrometry techniques (MALDI-TOF or Electrospray). Due to terpyridine
ligands possessing anti orientation, 1H NMR spectroscopy can be used to characterise
complexation by monitoring the upfield shift in the 6,6”-proton resonances as a result of
anti to syn ligand orientation upon complexation and the metal-ligand bond.6 UV-Vis
spectroscopy allows confirmation of complexation by the bathochromic shift of the
ligand-centred (LC) band and appearance of the metal-ligand charge transfer (MLCT)

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-10
band giving rise to intense colours of the compounds.27 Soft mass spectrometry
techniques such as MALDI-TOF or electrospray ionisation are ideal to analyse metal
bisterpyridine complexes as it allows the complexes to be detected without
fragmentation while retaining their isotopic patterns. However, when analysing Ir(III)
complexes, MALDI-TOF may cause fragmentation as the MLCT band overlaps the
excitation wavelength (337 nm) of the MALDI-TOF laser.28
In recent decades, metal bis-complexes of Ru(II), Os(II) and Ir(III) metal ions
have been of primary interest, in particular, Ru(II)-complexes due to their interesting
photophysical, electrochemical and photochemical properties. In this project,
functionalised 4’-aryl Ru(II)-complexes have been selected for study due to the MLCT
band taking place in the visible region allowing photo-excitation by visible light,
straightforward synthesis arising from achirality of 4’-functionalisation and their
improved excited state lifetimes after modification with 4’-aryl groups17 make them
ideal photosensitisers for attachment to cytochrome c as light-activated bioconjugates.
2.3.1 Synthesis of [Ru(tpy)]Cl3 5 and [Ru(tpy)2](PF6)2 6
Synthesis of asymmetric mono-complex precursor 5 and reference complex 6 was
achieved following modified literature methods (Scheme 2.6).29 Ru(II) mono-complex 5
was prepared by refluxing 2,2’:6’,2”-terpyridine and two molar equivalent of
ruthenium(III) trichloride hydrate in absolute ethanol overnight yielding precursor 5 as a
dark black solid in 88%. This compound was selected for forming asymmetric
complexes with maleimide functionalisation via the two-step directed methodology.

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-11
Scheme 2.6. Synthesis of [Ru(tpy)]Cl3. (a) EtOH, reflux, 88%.
In order to synthesise the reference complex 6 for photo-reaction control experiments
(Chapter 6), ruthenium(III) trichloride hydrate and two molar equivalent
2,2’:6’,2”-terpyridine was heated at 110 oC in ethylene glycol overnight in one-pot
(Scheme 2.7). The [Ru(tpy)2]2+ chloride salt was diluted in water and precipitated with
ammonium hexafluorophosphate. Complex 6 was recrystallised with acetonitrile/diethyl
ether to afford the red solid in 50% yield. Synthesis of complex 6 was achieved by a
modified approach, substituting N-ethylmorpholine in refluxing absolute ethanol with
neat ethylene glycol which is discussed further in the next section (Chapter 2.3.2).
Scheme 2.7. Synthesis of [Ru(tpy)2](PF6)2. (a) Ethylene glycol, 110 oC, 50%.

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-12
2.3.2 Synthesis of [Ru(tpy)(4’-(4-aminophenyl)-2,2’:6’,2”-tpy)](PF6)2 7
Asymmetric aniline functionalised complex 7 was prepared using a directed synthesis
approach. Initially, asymmetric aniline complex 7 was prepared using mono-complex
precursor 5 (Scheme 2.8). The synthesis of [Ru(tpy)(4’-(4-aminophenyl)-2,2’:6’,2”-
tpy)](PF6)2 7 was achieved by reacting aniline 4 with precursor 5 in refluxing absolute
ethanol overnight. The resulting product was filtered over celite and diluted with water.
Subsequently, complex 7 was precipitated with ammonium hexafluorophosphate and
recrystallised with acetonitrile/diethyl ether to afford red complex in 28% yield. 1H
NMR spectroscopy confirmed complexation due to the upfield shift of the 6,6”-protons
as a result of anti to syn conformation change of the terpyridine ligand as shown in
Figure 2.2.6
Scheme 2.8. Synthesis of [Ru(tpy)(4’-(4-aminophenyl)-2,2’:6’,2”-tpy)](PF6)2. (a) EtOH, reflux, 28%. (b) Ethylene glycol, 110 oC, 76%.

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-13
Figure 2.2. 1H NMR (300 MHz) spectra of aniline terpyridine 4 (top-blue-CDCl3) and asymmetric aniline complex 7 (bottom-black-CD3CN) showing upfield shift of the 6,6”-protons upon complexation due to change in anti to syn orientation of the terpyridine ligands.6 Complex 7 NH2 signal suppressed due to slow proton exchange of labile protons with CD3CN. Symbols represent 6,6”-protons.
Generally, the mono-complex 5 is reacted with the second ligand in ethanol (reductant)
containing N-ethylmorpholine30 (catalyst) and refluxed to afford the bisterpyridine
complex. However, previous studies in the Thordarson group showed that in the
presence of N-ethylmorpholine, symmetric complexation of aniline terpyridines resulted
in N-alkylation of the ligand.18
As a result of the poor yield due to the removal of catalyst in the complexation
reaction of aniline 7, the reaction was repeated using an alternative method based on
Ir(III)-bisterpyridine literature.1b Higher yields after complexation were observed when
performing the reaction in ethylene glycol. The synthesis of complex 7 was achieved by
reacting aniline 4 with mono-complex 5 in ethylene glycol at 110 oC. Subsequently, the
solution was filtered over celite and precipitated with ammonium hexafluorophosphate.

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-14
The resulting precipitate was centrifuged, washed with water and collected with
acetonitrile.
Initially, purification by recrystallisation was attempted; however, purification
by crystallisation for complex 7 was not possible due to scrambling of the ligands as a
result of harsher reaction conditions. Purification of the crude mixture was achieved by
separation over silica (gradient from 90:9:1 to 20:3:1, acetonitrile:water:saturated
potassium nitrate (v/v/v)), followed by a second neutral alumina column (gradient from
acetonitrile to 90:9:1, acetonitrile:water:saturated potassium nitrate (v/v/v)) to afford red
crystalline solid in 76% yield.
It was found that purification by chromatography over silica or alumina (neutral)
required highly polar solvents containing salts. This was a result of binding of the
complex to the stationary phase. Additionally, characterisation of purified fractions
proved difficult at times as thin layer chromatography (TLC) analysis showed
appearance of multiple bands due to counter ion exchange after purification.
2.3.3 Synthesis of [Ru(tpy)(maleimide-hexylcarboxamido-phenyl-tpy)](PF6)2 8
Following the preparation of the amino functionalised complex 7, maleimide modified
complexes were prepared for attachment to cytochrome c and other surface cysteine
containing proteins. Maleimide was introduced to complex 7 by peptide coupling
chemistry using a route similar to that of Hovinen31 due to the unreactive nature of
anilines preventing the use of classical N-hydroxysuccinimide/N,N’-
dicyclohexylcarbodiimide (NHS/DCC) coupling conditions.
Maleimide complex 8 was prepared by reacting a solution of
6-maleimidocaproic acid, O-(7-azabenzotriazole-1-yl)-1,1,3,3-tetramethyluronium
hexafluorophosphate (HATU) and N,N-diisopropylethylamine (DIPEA) in dry

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-15
N,N-dimethylformamide was stirred at room temperature for 1 h (Scheme 2.9). Aniline
7 in dry dimethylformamide was then added to the stirring solution and stirred for a
further 26 h at room temperature. Dichloromethane was added to the solution and the
organic phase was washed with aqueous citric acid (10% w/v), water and dried with
anhydrous sodium sulphate. Dichloromethane was removed in vacuo and the
concentrated dimethylformamide phase was precipitated into dry diethyl ether, filtered
and further washed with diethyl ether. The crude mixture was purified over silica using
a gradient from acetonitrile to 70:29:1 acetonitrile:water:saturated potassium nitrate and
precipitated with ammonium hexafluorophosphate yielding the complex 8 as a red
crystalline solid in 41% yield.
Scheme 2.9. Synthesis of [Ru(tpy)(maleimide-hexylcarboxamido-phenyl-tpy)](PF6)2.(a) O-(7-azabenzotriazole-1-yl)-1,1,3,3-tetramethyluronium hexafluorophosphate (HATU), N,N-diisopropylethylamine, dry N,N-dimethylformamide, r.t., 41%.

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-16
2.3.4 Synthesis of [Ru(4’-(4-aminophenyl)-2,2’:6’2’’-terpyridine)2](PF6)2 9
In order to prepare dimeric bioconjugates, symmetric aniline complex 9 was similarly
prepared using a modified literature method from Ng et al.27 Ruthenium(III) trichloride
hydrate and two molar equivalent of aniline terpyridine 4 in ethylene glycol was heated
at 110 oC overnight (Scheme 2.10). Subsequently, the mixture was diluted with water
and filtered over celite. The product was precipitated using ammonium
hexafluorophosphate and the fine precipitate was collected by centrifugation followed
by washing with water. Purification of the crude mixture was achieved by
recrystallisation in acetonitrile/diethyl ether to yield red crystals in 52% yield.
Scheme 2.10. Synthesis of [Ru(4’-(4-aminophenyl)-2,2’:6’,2”-tpy)2](PF6)2. (a)Ethylene glycol, 110 oC, 52%.

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-17
2.3.5 Synthesis of [Ru(4’-(4-maleimide-hexylcarboxyamido-phenyl)-2,2’:6’2’’-
terpyridine)2](PF6)2 10
Bis-maleimide complex was prepared similarly to compound 8 in order to prepare
dimeric bioconjugates with proteins containing surface exposed cysteines. A solution of
6-maleimidocaproic acid, HATU and DIPEA in dry N,N-dimethylformamide was
stirred at room temperature for 1 h (Scheme 2.11). Aniline 9 in dry dimethylformamide
was then added to the stirring solution and stirred for a further 25 h at room
temperature. Dichloromethane was added to the solution and the organic phase was
washed with aqueous citric acid (10% w/v), water and dried with anhydrous sodium
sulphate. Dichloromethane was removed in vacuo and the concentrated
dimethylformamide phase was precipitated into dry diethyl ether, filtered and further
washed with diethyl ether. The crude mixture was purified over silica using a 20:1:3
acetonitrile:water:saturated potassium nitrate mobile phase and precipitated with
ammonium hexafluorophosphate yielding the complex 10 as a red crystalline solid in
27% yield.

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-18
Scheme 2.11. Synthesis of [Ru(maleimide-hexylcarboxamido-phenyl-tpy)2](PF6)2. (a) O-(7-azabenzotriazole-1-yl)-1,1,3,3-tetramethyluronium hexafluorophosphate (HATU), N,N-diisopropylethylamine, dry N,N-dimethylformamide, r.t., 27%.
2.3.6 Spectroscopic properties of Ru(II)-bistepyridine complexes
Characterisation of metal bisterpyridine complexes can be achieved by complementary
techniques including 1H NMR spectroscopy and mass spectrometry. However, UV-Vis
spectroscopy can be utilised to characterise complex formation in a straightforward
manner. Complexes of [Ru(tpy)2]2+-type display a characteristic spin allowed d- *
metal-ligand charge transfer (MLCT) band as well as ligand centred (LC) absorption
bands (Figure 2.3).22 Successful formation of terpyridine metal complexes is usually
accompanied by a bathochromic (red) shift of the LC absorption band and appearance
of a new MLCT band.

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-19
Figure 2.3. UV-Vis spectra of (a) terpyridines and 4’-aryl terpyridines in dichloromethane, (b) Ru(II)-bisterpyridine and functionalised complexes in acetonitrileshowing bathochromic shifts of ligand centred (LC) bands and appearance of ametal-ligand charge transfer (MLCT) band.
Comparison of [Ru(tpy)2]2+-type complexes of aniline 7 and 9 as well as maleimide
functionalised complexes 8 and 10 shows bathochromic shift of the LC band at ca.
310 nm compared to the ca. 280 nm absorption band of free terpyridine ligand and
functionalised ligands 3 and 4. These bands correspond to - * transitions with
electrons of the aromatic systems (LC). Additionally, appearance of a new absorption
band of the ruthenium(II) complexes is observed centred ca. 480-500 nm corresponding
to the MLCT bands, which relates to the excitation of an electron from the
metal-centred d-orbitals to an unfilled ligand-centred *-orbital.32
From Table 2.1, it can be seen that both asymmetric and symmetric aniline
complexes 7 and 9 exhibit significant red shifting of the MLCT band, 490 and 504 nm
respectively, compared to prototype complex 6 of 476 nm. This is a result of the
introduction of electron donating substituents which induce red shifting of absorption
bands due to decrease of the gap between the LUMO (ligand *) and HOMO (metal t2g,
distorted octahedral) orbitals after modification.17 Additionally, this effect is observed
with the asymmetric and symmetric maleimide functionalised complexes 8 and 10 with
MLCT bands red shifted at 485 and 494 nm, respectively. Finally, Table 2.1 shows a

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-20
general trend of increasing extinction coefficient after introducing phenyl substituents in
the 4’-positions for complexes 7-10 which is as expected based on simple theoretical
predictions.33
Table 2.1. Absorption data for prototype [Ru(tpy)2]2+ complex and 4’-functionalised analogues
Complex Absorption /nma Extinction coefficient /M-1cm-1
[Ru(tpy)2]2+ 6 476 17 70017
[Ru(tpy)(tpyNH2)]2+ 7 490 17 100[Ru(tpyNH2)2]2+ 9 504 31 800[Ru(tpy)(tpymal)]2+ 8 485 26 000[Ru(tpymal)2)]2+ 10 494 41 50018
a Data from 10-6 M solutions in acetonitrile solution
2.3.7 Crystallography
As further evidence of complex formation, single crystal structures of asymmetric
complexes 7 and 8 were obtained providing detailed structural analysis of resulting
complexes and the effects of 4’-aniline and maleimide substitution on geometries and
bond lengths. X-ray crystallography analysis was performed by Dr. Mohan Bhadbhade
(UNSW) with analysis of aniline 7 performed in-house at the UNSW Analytical Centre
and maleimide 8 at the Australian Synchotron facility.
2.3.7.1 [Ru(tpy)(4’-(4-aminophenyl)-2,2’:6’,2”-tpy)](PF6)2 7
The crystal structure of complex 7 dimethylformamide solvate with the composition
[Ru(C15H11N3)(C21H16N4)](PF6)2 2C3H7NO was crystallised by diffusion of diethyl
ether vapour into a solution of 7 in N,N-dimethylformamide in the triclinic space group,
P 1 as red blades. As shown in Figure 2.4, crystal 7 displays an orthorhombic distortion
from octahedral symmetry (N2A-Ru-N3A 78.8(5)o) which is as expected and observed
in [Ru(tpy)2](PF6)2.23

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-21
Figure 2.4. An ORTEP plot of complex 7 2C3H7NO (20% thermal ellipsoids for non-hydrogen atoms). Hydrogen atoms omitted for clarity. Triclinic space group, P1 .
In Table 2.2, the aniline shows a significant twist with a torsional angle of 29.1(2)o
(C7B-C8B-C18B-C17B) about the interannular bond, greater than the corresponding
twist found in the free 4’-phenyl terpyridine ligand.34 Ru-N bond lengths between
ligands A and B are equivalent, 2.033(7) Å and is within reported ranges for
[Ru(tpy)2](PF6)2.23 The C-C and C-N bond lengths within the aromatic rings are normal
and average 1.380(1) and 1.355(6) Å, respectively. The interannular C-C bond distances
of ligands A and B average 1.464(5) and 1.474(7) Å and are consistent with reported
asymmetric complex [Ru(tpy)(4’-5-carboxypentyl-tpy)](PF6)2 of 1.471(1) Å.29
Table 2.2. Selected bond lengths (Å) and angles/torsions (o) for complex 7 2C3H7NO with estimated standard deviations in parentheses.Ru-N1A 2.061(3) Ru-N1B 2.061(3)Ru-N2A 1.980(3) Ru-N2B 1.971(3)Ru-N3A 2.061(3) Ru-N3B 2.065(3)C5A-C6A 1.463(4) C5B-C6B 1.472(4)C10A-C11A 1.466(4) C10B-C11B 1.483(4)
C8B-C18B 1.469(5)
N2A-Ru-N3A 78.8(5) C7B-C8B-C18B-C17B 29.1(2)

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-22
From the packing diagram in Figure 2.5, it shows the complex cation forming
anti-parallel columnar stacks along the c-axis while forming alternating layers with
hexafluorophosphate anions and N,N-dimethylformamide solvent molecules filling the
voids. Intermolecular contact within the columns are face-to-face and edge-to-face
-stacking at a distance of 3.8 Å (N1B and N3B) and 3.6 Å (N1A and N1B),
respectively, in adjacent unit cells.
Figure 2.5. Packing diagram obtained from single-crystal X-ray diffraction analysis of complex 7 2C3H7NO as a capped stick representation. Counter-ions and dimethylformamide omitted for clarity. Representation viewed along b-axis.
2.3.7.2 [Ru(tpy)(maleimide-hexylcarboxamido-phenyl-tpy)](PF6)2 8
The crystal structure of complex 8 dimethylformamide water solvate with the
composition [Ru(C15H11N3)(C31H27N5O3)](PF6)2 2C3H7NO H2O was crystallised by
diffusion of diethyl ether vapour into a solution of 8 in N,N-dimethylformamide in the
monoclinic space group, P21/c as thin red blades. Structural characteristics of complex
8 are similar to complex 7. The main notable difference is the reduction in the torsion
angle between 4’-aryl maleimide group and the central pyridyl ring of ligand B with an
angle of 23.9(8)o as shown in Table 2.3 and Figure 2.6a.

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-23
Table 2.3. Selected bond lengths (Å) and angles/torsions (o) for complex 8 2C3H7NO H2O with estimated standard deviations in parentheses.Ru-N1A 2.076(4) Ru-N1B 2.061(4)Ru-N2A 1.976(4) Ru-N2B 1.985(4)Ru-N3A 2.068(4) Ru-N3B 2.060(4)C5A-C6A 1.458(5) C5B-C6B 1.481(5)C10A-C11A 1.467(5) C10B-C11B 1.462(5)
C8B-C18B 1.486(6)
N2A-Ru-N3A 78.7(9) C7B-C8B-C16B-C17B 23.9(8)
Based on the packing diagram as shown in Figure 2.6b, it can be seen that complex 8 is
stacked in an interlaced anti-parallel motif along the b-axis, possibly to accommodate
the steric hindrance of the flexible maleimide spacer group. Intermolecular contact
within the columns along the a-axis of the crystal structure is face-to-face and
edge-to-face -stacking at a distance of 3.7 Å (N1A and N3A) and 3.3 Å (N1A and
N1B), respectively, coupled with twisted parallel-displaced -stacking between the
outer pyridyl ring (N3B) and 4’-phenyl ring at a distance of 3.8 Å in adjacent unit
cells. Hexafluorophosphate anions form alternating layers with complex 8 along the
a-axis and solvent molecules fill the voids in the packed structure.

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-24
Figure 2.6. Single-crystal X-ray diffraction structures of complex 8 2C3H7NO H2O. (a) an ORTEP plot (20% thermal ellipsoids for non-hydrogen atoms). Hydrogen atoms omitted for clarity. (b) Packing diagram as a capped stick representation. Counter-ions and solvent molecules omitted for clarity. Representation viewed along a-axis.Monoclinic space group, P21/c.

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-25
2.4 Functionalised Anthraquinone-Based Acceptors
In addition to light-activated electron donors based on metal bisterpyrdines, azide
functionalised anthraquinones were prepared as potential ‘clickable’ electron acceptors
to light-activated biological electron donors such as green fluorescent proteins (GFP).
Electron acceptors of the quinone family are one of the most widely studied
acceptors17,26 due to the electron transfer roles of quinones in photosynthesis and
respiration.35 Quinones are highly suitable as electron acceptors due to their electron
shuttling behaviour allowing reversibility between both stable redox states via a two
electron process as shown in Scheme 2.12.36 In these studies, anthraquinones were
selected due to their higher stabilities compared to p-benzoquinones and functionalised
p-benzoquinones which can degrade to form quinhydrones37 when isolated or stored in
solution for prolonged periods.
Scheme 2.12. Stable reversible conversion of oxidised/reduced anthraquinone (AQ) states via a two electron redox process.
Modifying proteins chemically can be difficult due to an array of potential functional
groups including amines, carboxylic acids, histidines and thiols. The Cu(I)-catalysed
[3+2] Huisgen cycloaddition of azides and terminal alkynes, also known as the
azide/alkyne click reaction, is ideal due to benign reaction conditions as well as
chemoselectivity.38 As such, azide functionalised anthraquinones were selected as
synthetic targets in this project to functionalise light-activated biological electron
donors.

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-26
2.4.1 Synthesis of 1-amino-3-azidopropane 11
In order to introduce azide functionality to anthraquinone, a short chain spacer
containing amide and azide functional groups was synthesised to allow attachment to
anthraquinone via peptide coupling chemistry. The 1-amino-3-azidopropane spacer 11
was synthesised based on a previously reported method.39
The azide spacer 11 was prepared by reacting 1-amino-3-bromopropane
hydrobromide with four molar excess of sodium azide in refluxing water via a
displacement reaction for 23 h (Scheme 2.13). Subsequently, the aqueous mixture was
cooled to 0 oC, extracted with diethyl ether and the organic phase was dried with
anhydrous sodium sulphate. The organic phase was removed in vacuo to afford the pure
yellow oil 11 in 43% yield.
Scheme 2.13. Synthesis of 1-amino-3-azidopropane. (a) NaN3, H2O, reflux, 43%.
2.4.2 Synthesis of anthraquinone-2-azidopropylamide 13
Following the synthesis of azide spacer 11, the azide modified anthraquinone 13 was
then prepared using peptide coupling methods based on NHS/DCC chemistry. The
purpose of either maleimide modification using an N-propargyl maleimide spacer to
attach to GFP or direct attachment to an alkyne-tagged GFP using click chemistry is the
formation of a light activated donor-acceptor system based on GFP-anthraquinone,
which is further discussed in Chapter 2.4.3 and Chapter 4.
Azide 13 was prepared by initially preparing the N-hydroxysuccinimide (NHS)
ester of anthraquinone-2-carboxylic acid (Scheme 2.14). Anthraquinone-2-carboxylic
acid and NHS was cooled to 0 oC in dry dichloromethane and
N,N’-dicyclohexylcarbodiimide (DCC) was added to the mixture. The reaction mixture

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-27
was stirred overnight at room temperature and protected from light, the resulting yellow
solution was filtered to remove the insoluble urea by-product. The organic phase was
removed in vacuo to afford the crude NHS-anthraquinone 12 in quantitative yields that
was used in the next step without further purification.
Scheme 2.14. Synthesis of anthraquinone-NHS. (a) N-hydroxysuccinimide, N,N’-dicyclohexylcarbodiimide, CH2Cl2, r.t.
Azide 13 was prepared using a similar method developed by Zhang et al.40 (Scheme
2.15). Crude anthraquinone-NHS 12 was suspended in a mixed acetone/ethanol solvent.
Azide spacer 11 in an aqueous solution of sodium bicarbonate was added to the reaction
mixture and allowed to react for 23 h. The organic phase was removed in vacuo and
product was resuspended in dichloromethane. Product was washed with basic followed
by acidic water and purified on silica using dichloromethane to afford the yellow
product in 35% yield. CAUTION: Small organic azides can be potentially explosive
and handling of the spacer 11 should be treated with care. 1H NMR spectroscopy
confirmed formation of azide 13, as an upfield shift from 2.27 to 1.97 ppm of proton
signals adjacent to the azide group was observed in comparison to bromide 14 as shown
in Figure 2.7.
An alternative approach to prepare 13 was attempted using an indirect route
which is potentially safer due to the formation of a higher molecular weight azide in
comparison to azide 11. Azide 13 was attempted by initially preparing the bromide
precursor 14, followed by azide substitution under mild heating conditions at 60 oC for
3 days. However, it was found that 50% reduction of the anthraquinone moiety was

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-28
observed compared to bromide 14, as well as formation of unidentifiable by-products by
1H NMR spectroscopy and TLC analysis. Oxidation by
2,3-dichloro-5,6-dicyano-1,4-benzoquinone (DDQ) of the reduced species was
attempted with no success.
Scheme 2.15. Synthesis of anthraquinone-2-azidopropane using the direct/indirect methods. (a) Anthraquinone-NHS 12, acetone/EtOH/NaHCO3 (aq), 35%. (b)Anthraquinone-NHS 12, acetone/EtOH/NaHCO3 (aq), 56%. (c) NaN3, CH3CN, 60 oC.
Figure 2.7. 1H NMR (300 MHz, CDCl3) spectra of bromide 14 (top-blue) and azide 13(bottom-black) showing upfield shift of protons after sodium azide displacement reaction indicated by the asterisk.

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-29
2.4.3 Attempted synthesis of anthraquinone-2-propylamido-triazole-maleimide
15
Initially, the synthesis of maleimide modified anthraquinone was attempted using
azide/alkyne click chemistry to allow direct modification of the single cysteine residue
of Acropora millepora green fluorescent protein. Attempted maleimide anthraquinone
15 was prepared by reacting N-propargyl maleimide with azide 13 in a
tetrahydrofuran:water (5:2, v/v) solution containing copper(II) sulphate pentahydrate
and L-ascorbic acid (Scheme 2.16). The solution was reacted overnight and the organic
phase was removed in vacuo and resuspended in dichloromethane, washed with water,
dried with anhydrous sodium sulphate and isolated in vacuo to yield a yellow oil.
Based on 1H NMR spectroscopy, it was observed that the formation of the
triazole was successful with resonances at 8.05 ppm consistent with triazole formation.
However, characteristic maleimide resonance at ca. 6.8 ppm or corresponding
hydrolysed maleimide at ca. 6.3 ppm was not observed.28 Furthermore, mass
spectrometry analysis shows a signal at 596.10 m/z compared to the expected signal for
protonated molecular ion of maleimide 15 at 470.14 m/z. The resulting compound was
not identified; however, it is proposed to be a product of polymerisation.
Scheme 2.16. Attempted synthesis of anthraquinone-triazole-maleimide. (a) CuSO4,L-ascorbic acid, tetrahydrofuran:water (5:2, v/v).

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-30
2.5 Conclusions
Terpyridine ligands bearing amines with 4’-aryl functionalisation were synthesised
based on a modified Kröhnke synthesis with isolation of the azachalcone/pyridinium
iodide precursors. Light-activated chromophores of Ru(II)-bisterpyridine complexes
were prepared based on conditions adapted from Ir(III)-bisterpyridine literature in
ethylene glycol. Homo and heteroleptic aniline complexes 7 and 9 were synthesised to
allow chemical modification by introducing maleimide functionality using peptide
coupling chemistry. Asymmetric and symmetric maleimide complexes 8 and 10 were
synthesised such that these electron donating photosensitisers could be attached to
cysteine bearing proteins/enzymes which are discussed further in Chapter 3. As a
reference compound, complex 6 was prepared for control room temperature
photoreduction studies. Single crystals of complexes 7 and 8 were isolated and analysed
by X-ray crystallography to confirm their structural properties. At the time of writing,
complex 8 is the first reported single crystal X-ray structure of a Ru(II)-bistepyridine
complex with maleimide functionality.
Anthraquinone derivatives were also prepared to function as electron acceptors
after functionalising to a light-activated biological electron donor such as green
fluorescent protein (GFP). Azide bearing anthraquinone 13 was prepared for potential
Cu(I)-catalysed [3+2] Huisgen cycloaddition reactions (click reactions) of azides with
terminal alkynes. Attempts to functionalise azide 13 with maleimides using click
chemistry were unsuccessful and subsequent click chemistry studies of azide 13 with
alkyne-tagged GFP will be discussed further in Chapter 4.

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-31
2.6 References
(1) (a) Heller, M.; Schubert, U. S. Eur. J. Org. Chem. 2003, 947. (b) Hofmeier, H.; Schubert, U. S. Chem. Soc. Rev. 2004, 33, 373.
(2) Anderson, G. W.; Zimmerman, J. E.; Callahan, F. M. J. Am. Chem. Soc. 1964,86, 1839.
(3) (a) Rostovtsev, V. V.; Green, L. G.; Fokin, V. V.; Sharpless, K. B. Angew. Chem. Int. Ed. 2002, 41, 2596. (b) Dirks, A. J.; van Berkel, S. S.; Hatzakis, N. S.; Opsteen, J. A.; van Delft, F. L.; Cornelissen, J. J. L. M.; Rowan, A. E.; van Hest, J. C. M.; Rutjes, F. P. J. T.; Nolte, R. J. M. Chem. Commun. 2005, 4172.
(4) Morgan, G. T.; Burstall, F. H. J. Chem. Soc. 1932, 20.(5) Blau, F. Ber. Dtsch Chem. Ges. 1888, 21, 1077.(6) Schubert, U. S.; Hofmeier, H.; Newkome, G. R. Modern Terpyridine Chemistry;
1st ed.; WILEY-VCH Verlag: Wienheim, Germany, 2006.(7) Kröhnke, F. Synthesis 1976, 1976, 1.(8) Potts, K. T.; Usifer, D. A.; Guadalupe, A.; Abruna, H. D. J. Am. Chem. Soc.
1987, 109, 3961.(9) Jameson, D. L.; Guise, L. E. Tetrahedron Lett. 1991, 32, 1999.(10) Constable, E. C.; Lewis, J. Polyhedron 1982, 1, 303.(11) Cargill Thompson, A. M. W. Coord. Chem. Rev. 1997, 160, 1.(12) (a) Uenishi, J.; Tanaka, T.; Wakabayashi, S.; Oae, S.; Tsukube, H. Tetrahedron
Lett. 1990, 31, 4625. (b) Parks, J. E.; Wagner, B. E.; Holm, R. H. J. Organomet. Chem. 1973, 56, 53.
(13) Miyaura, N.; Suzuki, A. Chem. Rev. 1995, 95, 2457.(14) Negishi, E. Current Trends in Organic Synthesis; 1st ed.; Pergamon, Oxford,
1983.(15) Stille, J. K. Angew. Chem. Int. Ed. 1986, 25, 508.(16) Fallahpour, R.-A. Synthesis 2003, 155.(17) Sauvage, J. P.; Collin, J. P.; Chambron, J. C.; Guillerez, S.; Coudret, C.; Balzani,
V.; Barigelletti, F.; De Cola, L.; Flamigni, L. Chem. Rev. 1994, 94, 993.(18) Peterson, J. R. PhD Thesis, The University of Sydney, 2009.(19) Mukkala, V.-M.; Helenius, M.; Hemmilä, I.; Kankare, J.; Takalo, H. Helv.
Chim. Acta 1993, 76, 1361.(20) Mikel, C.; Potvin, P. G. Polyhedron 2002, 21, 49.(21) Lainé, P.; Bedioui, F.; Ochsenbein, P.; Marvaud, V.; Bonin, M.; Amouyal, E. J.
Am. Chem. Soc. 2002, 124, 1364.(22) Hofmeier, H.; El-ghayoury, A.; Schenning, A. P. H. J.; Schubert, U. S.
Tetrahedron 2004, 60, 6121.(23) Lashgari, K.; Kritikos, M.; Norrestam, R.; Norrby, T. Acta Cryst. C 1999, 55,
64.(24) Goldstein, D. C.; Cheng, Y. Y.; Schmidt, T. W.; Bhadbhade, M.; Thordarson, P.
Dalton Trans. 2011, 40, 2053.(25) Beley, M.; Collin, J. P.; Louis, R.; Metz, B.; Sauvage, J. P. J. Am. Chem. Soc.
1991, 113, 8521.(26) Storrier, G. D.; Colbran, S. B.; Craig, D. C. J. Chem. Soc., Dalton Trans. 1998,
1351.(27) Ng, W. Y.; Gong, X.; Chan, W. K. Chem. Mater. 1999, 11, 1165.(28) Goldstein, D. C. PhD Thesis, The University of New South Wales, 2011.(29) Hofmeier, H.; Andres, P. R.; Hoogenboom, R.; Herdtweck, E.; Schubert, U. S.
Aust. J. Chem. 2004, 57, 419.

Chapter 2 Synthesis of Ru(II)-Complexes and Quinone derivatives
2-32
(30) Maestri, M.; Armaroli, N.; Balzani, V.; Constable, E. C.; Thompson, A. M. W. C. Inorg. Chem. 1995, 34, 2759.
(31) Hovinen, J. Bioconjug. Chem. 2007, 18, 597.(32) Beley, M.; Collin, J.-P.; Sauvage, J.-P.; Sugihara, H.; Heisel, F.; Miehe, A. J.
Chem. Soc., Dalton Trans. 1991, 3157.(33) Phifer, C. C.; McMillin, D. R. Inorg. Chem. 1986, 25, 1329.(34) Constable, E. C.; Lewis, J.; Liptrot, M. C.; Raithby, P. R. Inorg. Chim. Acta
1990, 178, 47.(35) (a) Purves, W. K.; Sadava, D.; Orians, G. H.; Heller, C. H. Life: The Science of
Biology; 7th ed.; Sinauer Associates and W. H. Freeman: U.S.A., 2003. (b) Gust, D.; Moore, T. A.; Moore, A. L. Acc. Chem. Res. 1993, 26, 198.
(36) Luo, W.; Chan, E. W. L.; Yousaf, M. N. J. Am. Chem. Soc. 2010, 132, 2614.(37) (a) Wolfenden, R.; Liang, Y. L.; Matthews, M.; Williams, R. J. Am. Chem. Soc.
1987, 109, 463. (b) Holmes, T. J.; John, V.; Vennerstrom, J.; Choi, K. E. J. Org. Chem. 1984, 49, 4736.
(38) Hvasanov, D.; Goldstein, D. C.; Thordarson, P. In Molecular Solar Fuels; The Royal Society of Chemistry, 2012; p 426.
(39) Lewis, W. G.; Magallon, F. G.; Fokin, V. V.; Finn, M. G. J. Am. Chem. Soc.2004, 126, 9152.
(40) Zhang, Y.; Gu, H.; Yang, Z.; Xu, B. J. Am. Chem. Soc. 2003, 125, 13680.

Chapter 3
Bioconjugate Synthesis of Cytochrome c and RelatedDerivatives

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-2
3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
In this Chapter, the synthesis of a number of iso-1 cytochrome c based bioconjugates
derived from the Saccharomyces cerevisiae (yeast) species for use as either
light-activated bioconjugates or dimeric bioconjugates were prepared. The
bioconjugation reactions described herein are based on the Michael addition reaction
between a single free cysteine of a protein and a maleimide ligand.
An asymmetric Ru(II)-bisterpyridine cytochrome c (8-cyt c) bioconjugate was
prepared as a light-harvesting bioconjugate for use as an electron transfer
photosensitiser. Additionally, to explore the factors affecting dimer formation of
bioconjugates using bismaleimide functionalised spacers, dimers were prepared using
combinations of free single cysteine containing proteins including iso-1 cytochrome c
(cyt c), bovine serum albumin (BSA) and mutated Acropora millepora green
fluorescent protein (GFP).
These bioconjugates were purified using fast protein liquid chromatography
(FPLC) and were characterised by UV-Vis spectroscopy, mass spectrometry and gel
electrophoresis. The procedures for bioconjugate synthesis are briefly reviewed in this
chapter.
3.1 Purification of Cytochrome c
Isolation of yeast cytochrome c was first achieved in 1930 by Keilin1 using only
precipitation techniques involving sulphur dioxide and salts, with residual
non-cytochrome proteins present in the final product. Since the initial report of
purification of cytochrome c, modifications to the methodology including fractionation
with ammonium sulphate, boiling in ammonium sulphate solution or boiling in the
presence of chloroform have been made.2 With the advent of ion-exchange

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-3
chromatography3, isolation of the variants of yeast cytochrome c including iso-1 and
iso-2 forms has been achieved.4 For this project, bioconjugation reactions involving the
iso-1 form as shown in Figure 3.1 are desirable due to the presence of a single available
cysteine residue at CYS102 for modification with maleimides allowing site-specific
modification.
Figure 3.1. Solid ribbon representation of iso-1 cytochrome c with heme group and CYS102 residue indicated as stick representations. The structure was derived from the protein data bank file ‘1YCC’.5
3.1.1 Purification of iso-1 cytochrome c using cation exchange chromatography
The purification of crude yeast cytochrome c (Sigma Aldrich) has been optimised by
Peterson et al.4 in the Thordarson group. Reduced protein (2 mg, reduced with
dithiothreitol) is loaded onto a strong cation exchange column (Supelco,
7.5 cmh × 0.75 cmd, 3.3 mL column volume) and eluted with a 328 to 450 mM sodium
chloride gradient over 14.5 mL in a 20 mM sodium dihydrogen phosphate buffer, pH 7.0
at 1 mL/min as shown in Figure 3.2. Pure iso-1 cytochrome c was collected as the main

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-4
peak, approximately eluting from 15.8 to 18 mL with slight variation across different
batches. The pooled fraction is concentrated and dialysed against water prior to storage
at -20 oC. The initial reduction step prior to column loading improves separation from
iso-2 cytochrome c while also generating the free cysteine residue (CYS102) by
disulfide bridge cleavage allowing for maleimide-cysteine chemical modification.
Figure 3.2. Purification of iso-1 cytochrome c (peak A) on a Tosoh SP-5PW strong cation exchange column (Supelco, 7.5 cmh × 0.75 cmd, 3.3 mL column volume, 10 mparticle size) eluted with a gradient of 328 to 450 mM in 20 mM sodium dihydrogen phosphate buffer, pH 7.0 at 1 mL/min. Peak B corresponds to iso-2 cytochrome c. The gradient (red) is shown for illustration.
A batch purification run (12 mg crude dry weight) using the method shown in Figure
3.2 typically yields 54% (506 nmol) of pure iso-1 cytochrome c based on UV-Vis
absorbance. Throughout this chapter and project, yield is based on UV-Vis absorbance
using the molar absorptivity of iso-1 cytochrome c ( 410 =97.6 mM-1cm-1)4 and the yield
is interchangeably expressed as both dry weight and the moles of protein.

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-5
3.2 Bioconjugation Methods
Cytochrome c is one of the most highly studied proteins due to its ease of purification
from multiple species3, use in electron transfer studies as a model redox protein6 and the
important roles they play in the electron transport chain of photosynthesis and
respiration.7 Saccharomyces cerevisiae cytochrome c was the first to be isolated from
the cytochrome c enzyme family by Keilin in 1930.1 Bioconjugation of cytochrome c
has been reported extensively in the literature targeting histidine8, lysine9 and cysteine10
residues and a brief review of these types of reactions will be discussed.
3.2.1 Modification of histidine
Modification of histidine residues of horse heart cytochrome c was achieved by the
formation of coordination complexes via coordinate covalent bonding. This is due to the
lone pair electrons of the histidine nitrogen which can coordinate with transition metal
ions. Winkler et al.8a synthesised a Ru(NH3)5(His-33)3+-ferricytochrome c bioconjugate
to study intramolecular electron-transfer kinetics between the two redox centres by
reacting horse heart cytochrome c (0.2 mM) with fifty-fold excess of
[Ru(NH3)5(H2O)]2+ under argon for 72 h.
3.2.2 Modification of lysine
The most common method for chemical modification in bioconjugate chemistry is the
covalent linkage between a ligand and the amines of a protein. Coupling to the -amino
group on a lysine residue is often targeted as they are abundant on the surface of
proteins as well as the -N terminus of a protein.11 They can be easily modified by
reacting with an N-hydroxysuccimide (NHS) activated ester.12 It should be noted,

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-6
modification of amines offer poor site-specificity as the natural occurrence of lysine
residues in mammalian proteins is estimated to be approximately 6%.13
Pan et al.9a synthesised ten singly labelled horse heart cytochrome c
bioconjugates by reacting with an NHS-activated bipyridine ligand, as well as other
products including unmodified protein as shown in Scheme 3.1.
Scheme 3.1. Reactions of amines with N-hydroxysuccimide activated ester.9a
3.2.3 Modification of cysteine
Another popular target for chemical modification of natural amino acids is cysteine.
Based on the theory of nucleophilicity proposed by Edwards and Pearson14, the thiol
group of cysteine is one of the strongest nucleophiles in proteins. Cysteine residues
possess thiol groups with a pKa of approximately 9.15 Due to these properties, specific
chemical modifications of cysteines can proceed selectively and rapidly under benign
conditions.12a They can be used to chemically modify cysteines as maleimide
functionalised ligands are Michael acceptors.16 In the Thordarson group, Peterson et
al.17 demonstrated site-specific modification of iso-1 cytochrome c from the CYS102
residue and maleimide-functionalised ruthenium-terpyridine complexes to form light-
harvesting donor-acceptor bioconjugates as shown in Scheme 3.2. In this project,
maleimide-cysteine Michael addition reactions with free single cysteine proteins will be
the main focus as it allows site-specific modification under benign conditions
(pH 6.8-7).

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-7
Scheme 3.2. Reactions of cysteine with maleimides.17
3.3 Bioconjugation of Ru(II)-cyt c (8-cyt c)
A light-activated bioconjugate, 8-cyt c was synthesised for electron transfer studies
within a membrane environment and as a component in a photosynthetic-respiratory
system for photoreduction of oxygen as discussed in Chapter 6 and 7, respectively. The
synthesis of bioconjugates followed previously optimised conditions in the Thordarson
group.17b The bioconjugate 8-cyt c was site-specifically reacted by using a single
maleimide group functionalised Ru(II)-bisterpyridine chromophore 8 previously
discussed in Chapter 2.
Iso-1 cytochrome c was added to a solution containing five-fold excess of
complex 8 in a phosphate buffer at pH 7.0 containing ethylenediaminetetraacetic acid
(EDTA) and acetonitrile. The final reaction conditions were iso-1 cytochrome c
(10 M), complex 8 (50 M), phosphate buffer (20 mM), EDTA (20 mM) and acetonitrile
(5% v/v) at pH 7.0 as shown in Scheme 3.3. The mixture was stirred overnight at room
temperature in a plastic reaction vessel in the dark to prevent degradation of maleimide

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-8
and photoreduction of cytochrome c. It should be noted that the addition of EDTA
improves bioconjugation yields by removal of trace copper ions via chelation, as
oxidation of cysteine can occur.1
Scheme 3.3. Synthesis of bioconjugate 8-cyt c. (a) Reduced iso-1 cytochrome c (10 M), phosphate buffer (20 mM), ethylenediaminetetraacetic acid (20 mM), pH 7.0, acetonitrile (5% v/v), r.t., 6%.
It was observed that with the hexafluorophosphate salt of complex 8, significant
precipitation of the complex was observed after stirring for 20 h resulting in extremely
low yields <1%. However, it was noticed that after exchange with a chloride salt,
improved water solubility of the complex 8 was observed with slight formation of
precipitates. Any precipitates that formed during the course of the reaction were
removed by syringe filtration using a 0.2 m filter and the solution was concentrated
and dialysed into water. Dialysis was performed to remove acetonitrile and EDTA from
the protein solution. In particular, removal of EDTA is essential as leaching of

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-9
immobilised nickel (Ni2+) can occur from subsequent purification using immobilised
metal affinity chromatography (IMAC).
Purification of conjugate 8-cyt c was achieved using similar conditions as
previously developed in the Thordarson group by Peterson et al.17a for
Ru(II)-bisterpyridine based bioconjugates. Separation of covalently attached
Ru(II)-bisterpyridine bioconjugates from unreacted proteins and ligands can be achieved
using Ni2+ IMAC chromatography, although the mechanism remains unknown. The
crude product was loaded onto an IMAC (Ni2+) column and purified using an imidazole
gradient from 0 to 125 mM over 11.5 mL in 20 mM sodium dihydrogen phosphate, 0.5 M
sodium chloride at pH 7.0 as shown in Figure 3.3.
Figure 3.3. Purification of bioconjugate 8-cyt c by IMAC (Ni2+) chromatography (HisTrapTM HP, GE Healthcare) using a gradient from 0 to 125 mM imidazole in 20 mMphosphate buffer, 0.5 M sodium chloride, pH 7.0 in 11.5 mL at 0.5 mL/min. Peak A and B – bioconjugate 8-cyt c. The gradient (green) is shown for illustration.

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-10
Prior to the imidazole gradient, elution of unreacted iso-1 cytochrome c and ligand
occurs. Product containing fractions A and B is eluted after the imidazole gradient,
pooled, concentrated and dialysed against water. This afforded the bioconjugate 8-cyt c
with high purity and moderate yield of 6%. UV-Vis spectroscopy is a key method for
determination of protein and bioconjugate mass and yields, but also to monitor protein
stability18 and to determine the effect and extent of bioconjugation.9a,19 UV-Vis
spectroscopy confirmed the purified product as shown in Figure 3.4.
Figure 3.4. UV-Vis spectra of 8-cyt c (green, H2O) and approximated by the linear sum (blue dashed) of oxidised iso-1 cytochrome c (red, H2O) and complex 8 (black, CH3CN).
The UV-Vis absorption spectrum of the Ru(II) bioconjugate 8-cyt c shows the
characteristic absorption peaks of both iso-1 cytochrome c (410 nm) and complex 8
(485 nm). Based on the spectra shown in Figure 3.4 and the molar absorptivities of iso-1
cytochrome c ( 410 = 97.6 mM-1cm-1)4 and complex 8 ( 485 = 26.0 mM-1cm-1), a 1:1: ratio

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-11
of iso-1 cytochrome c and complex is observed. Figure 3.4 shows that bioconjugate
8-cyt c can be approximated as the linear sum of oxidised iso-1 cytochrome c and
[Ru(tpy)(tpymal)](PF6)2 8, indicating that there is negligible ground state
communication between the two species of the bioconjugate.
Bioconjugate 8-cyt c was characterised by MALDI-TOF mass spectrometry
using either a matrix of saturated sinapinic acid/ -cyano-4-hydroxycinnamic acid
mixture or caffeic acid as it is a soft ionisation technique which produces predominantly
singly charged molecular ions with minimal fragmentation. It was found that for
optimal signal-to-noise, high protein and bioconjugate concentrations (>10 M) were
required and caffeic acid produced better spectra. The MALDI-TOF mass spectrum of
8-cyt c is shown in Figure 3.5.
Figure 3.5. MALDI-TOF mass spectra of iso-1 cytochrome c (black) and 8-cyt c (red). Peaks corresponding to the calculated masses 12 706 and 13 559 Da, respectively, were detected. Spectra were baseline corrected.

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-12
The spectra shown in Figure 3.5 shows that the measured mass of bioconjugate 8-cyt c
(13 555 Da) is in good agreement with the expected value (13 559 Da) and correlates
with UV-Vis analysis showing that only a single chromophore is attached to the protein,
which is expected based on maleimide-cysteine coupling at neutral pH. From the
spectra of iso-1 cytochrome c, matrix adducts of caffeic acid can be observed which is
consistent with previous reports by Yang et al.20 which reported observation of adducts
with matrix components via MALDI-TOF.
Finally, in order to confirm the purity of the bioconjugate 8-cyt c, analysis by
gel electrophoresis was performed and stained with SimplyBlueTM Safestain as shown
in Figure 3.6. Gel electrophoresis of conjugate 8-cyt c was prepared by denaturation at
70 oC for 10 min and if samples were to be reduced, dithiothreitol (DTT) was added.
Figure 3.6. Gel electrophoresis of SeeBlue® Plus2 molecular weight marker, iso-1cytochrome c (expected 12 706), 8-cyt c (expected 13 559) and a mixture of iso-1cytochrome c and myoglobin (Mb, expected 16 948). Samples on the left are reduced with dithiothreitol, while samples on the right are non-reduced. Samples stained with SimplyBlueTM Safestain.

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-13
As expected, bioconjugate 8-cyt c as shown in Figure 3.6 migrates as a slightly higher
molecular weight species than unmodified iso-1 cytochrome c. It can be seen that in the
non-reduced samples, a cytochrome c disulfide dimer band can be observed which is
absent after reduction with DTT. However, the bioconjugate is unaffected indicating
that the covalent linkage between maleimide and cysteine cannot be reduced and is
stable under strong reducing conditions.
3.4 Synthesis of Dimeric Bioconjugates
Dimeric bioconjugates were prepared using Ru(II)-bismaleimide complex 10 and 4,4’-
bipyridinium-N,N-di(maleimidopropyl) hexafluorophosphate 16 as small organic
spacers. In order to investigate factors influencing protein dimer formation to improve
yields, protein dimers were prepared using combinations of iso-1 cytochrome c, bovine
serum albumin (BSA, EC 232-936-2) and green fluorescent protein (Acropora
millepora, GFP) possessing varying molecular size, geometry and in particular
isoelectric points (pI). However, the concept of utilising supramolecular ionic
interactions to facilitate synthetic protein dimer formation has been neglected.
Control of protein dimerisation using small molecular ligands allows for
applications in gene expression21, signal transduction22, protein therapeutics23 and tumor
therapy.24 Although dimerisation of proteins has wide applicability, synthetic
dimerisation by covalent modification using small ligands have mostly been limited to
low molecular weight enzymes or peptides (<10 kDa) which primarily target terminal
(N/C) residues in the literature.25 There have been limited examples of isolated high
molecular weight protein synthetic dimers in reasonable yields (>20%); a 52 kDa dimer
of a monoclonal antibody single-chain fragment (di-scFv)26, a 128 kDa haemoglobin
dimer (di-Hb)27, a 34 kDa human interleukin 1 receptor antagonist (di-IL-1ra)28, a

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-14
133 kDa human serum albumin (HAS) dimer29 and a 96 kDa lipase-BSA heterodimer.30
It is worth noting that Hb has a pI around 7.1-7.531, close to neutrality, largely
eliminating any unfavourable ionic interactions between the two proteins. For di-scFv
and di-IL-1ra, the reaction was facilitated by targeting the corresponding protein
terminal residues. The di-IL-1ra was synthesised by native chemical ligation (NCL) of
N-terminal modified IL-1ra while di-scFV was synthesied from scFV modified on the
C-terminus by a cysteine—E-TAG sequence.32 Additionally, the HSA dimer yields
were reported based on the non-isolated product resulting in an overestimate. Finally,
for the lipase-BSA heterodimer, the reaction was achieved by an azide-alkyne click
reaction.
3.4.1 Synthesis of cyt c-10-BSA
The cyt c-10-BSA (oppositely charged heterodimer) was prepared by adding a solution
of bismaleimide complex 10 (as chloride salt) in acetonitrile to ten-fold excess of iso-1
cytochrome c in a final phosphate buffer at pH 7.0 containing EDTA and acetonitrile
(5% v/v) for 2 h as shown in Scheme 3.4. This step ensures formation of
mono-functionalised cyt c-10 conjugate due to the CYS102 residue being buried in the
hydrophobic pocket of the protein, resulting in slower reaction rates ( 1 h for
completion).17b Desalting of the intermediate conjugate did not require desalting to
remove excess ligand 10 due to low yields from unfavourable homodimer formation,
further discussed in Chapter 3.4.5. Subsequently, BSA was added to the stirring solution
and allowed to react for an additional 21 h in the dark at room temperature. BSA
possesses a more reactive cysteine residue (single cysteine residue, CYS34) ( 2 min for
completion)17b as it is exposed in the hydrophilic region of the protein. The final
reaction conditions were iso-1 cytochrome c (100 M), BSA (200 M), complex 10

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-15
(10 M) in acetonitrile (5% v/v) and 20 mM sodium dihydrogen phosphate, 20 mM
ethylenediaminetetraacetic acid, pH 7.0. It should be noted that a twenty-fold excess of
BSA was added as it has been reported that the actual free cysteine available for
functionalisation is 0.5 mol of the protein.33
Scheme 3.4. Synthesis of bioconjugate cyt c-10-BSA. (a) i) Reduced iso-1 cytochrome c(100 M), 2 h, ii) Bovine serum albumin (BSA, 200 M), 21 h. Phosphate buffer (20 mM), ethylenediaminetetraacetic acid (20 mM), pH 7.0, acetonitrile (5% v/v), r.t., 30%. BSA protein structure aligned onto the human serum albumin X-ray crystallographic structure (PDB code: 1AO6).

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-16
Purification of bioconjugate cyt c-10-BSA was achieved by initial dialysis into water to
remove ethylenediaminetetraacetic acid followed by separation of unreacted complex
10 and unreacted proteins using Ni2+ IMAC chromatography. The crude product was
loaded onto an IMAC (Ni2+) column and purified using an imidazole gradient from 0 to
250 mM over 9 mL in 20 mM sodium dihydrogen phosphate, 0.5 M sodium chloride at
pH 7.0 as shown in Figure 3.7a. The product containing fractions (peaks A and B) were
pooled, concentrated and dialysed against water. Unreacted iso-1 cytochrome c, BSA
and complex 10 were eluted prior to the imidazole gradient. Excess unreacted BSA was
present with the cyt c-10-BSA fraction as it is commonly used for enzyme stabilisation
due to its adhesive properties34 and its difficulty of purification has been reported.35 The
product was further purified using a strong cation exchange column using a sodium
chloride gradient from 320 to 450 mM over 14.4 mL in 20 mM sodium dihydrogen
phosphate, pH 7.0 as shown in Figure 3.7b. Excess BSA and mono-functionalised
conjugates (10-cyt c) were removed prior to the sodium chloride gradient and further
residual BSA was removed at a concentration of 1 M sodium chloride (peak E).
Figure 3.7. Purification of bioconjugate cyt c-10-BSA by fast protein liquid chromatography. (a) IMAC (Ni2+) (HisTrapTM HP, GE Healthcare) using a gradientfrom 0 to 250 mM imidazole in 20 mM phosphate buffer, 0.5 M sodium chloride, pH 7.0in 9 mL at 0.5mL/min. Peak A and B – cyt c-10-BSA. (b) Strong cation exchange (CEX) column (SP-5PW, Supelco) using a gradient from 320 to 450 mM sodium chloride in 20 mM phosphate buffer, pH 7.0 in 14.4 mL at 1 mL/min. Peak C and D - cyt c-10-BSA and peak E – residual BSA. The gradient (green) is shown for illustration.

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-17
The product containing fractions C and D is eluted after the sodium chloride gradient,
pooled, concentrated and dialysed against water. This afforded the bioconjugate
cyt c-10-BSA in 30% yield, estimated by gel electrophoresis and UV-Vis spectroscopy
(see Appendix B).
The UV-Vis absorption spectrum of the heterodimer cyt c-10-BSA shows three
primary absorption bands at 280 nm, 410 nm and 495 nm corresponding to BSA,
cytochrome c, and ligand 10, respectively, as shown in Figure 3.8a.
Figure 3.8. UV-Vis spectra of dimeric bioconjugates. (a) cyt c-10-BSA. (b) cyt c-10-cyt c. (c) cyt c-16-GFP. (d) cyt c-10-BSA. All measurements were made in H2O.
Confirmation of cyt c-10-BSA dimer was observed using MALDI-TOF mass
spectrometry with a m/z 80 609 as shown in Figure 3.9.

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-18
Figure 3.9. MALDI-TOF mass spectrum of cyt c-10-BSA. Peaks corresponding to the calculated masses 66 776 and 80 618 Da, respectively, were detected. Spectrum wasbaseline corrected and noise reduced.
The resulting heterodimer was additionally characterised by denaturing gel
electrophoresis. For 12% Bis-Tris SDS PAGE gels, quantitative molecular weight
determination is accurate for proteins less than 30 kDa. Therefore, SDS PAGE
analysis in this study was used qualitatively to confirm formation of product by
observing new bands corresponding to dimer. Hence, the SDS PAGE gel
electrophoresis in Figure 3.10 shows a new dimer band, further confirming conjugate
formation.

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-19
Figure 3.10. Gel electrophoresis of SeeBlue® Plus2 molecular weight marker, iso-1cytochrome c (expected 12 706), green fluorescent protein (GFP, expected 26 807), bovine serum albumin (expected 66 776), cyt c-10-BSA (expected 80 618), cyt c-10-cyt c(expected 26 550), cyt c-16-GFP (expected 40 236) and BSA-10-BSA (expected 134 688). Samples are reduced with dithiothreitol. Samples stained with SimplyBlueTM
Safestain and contrast corrected. Dimer bands highlighted in boxes.
3.4.2 Synthesis of cyt c-10-cyt c
In order to determine the effect of homodimer formation on bioconjugate dimer yields,
the homodimer cyt c-10-cyt c was synthesised. The bioconjugate was prepared by
adding a solution of complex 10 (as a chloride salt) in acetonitrile to ten-fold reduced
iso-1 cytochrome c (five-fold protein per maleimide group) in a final phosphate buffer
solution, pH 7.0, containing EDTA and acetonitrile (5% v/v) for 27 h in the dark at
room temperature as shown in Scheme 3.5. The product was concentrated, dialysed
against water and purified by IMAC (Ni2+) chromatography.

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-20
It should be noted that for all the subsequent protein dimerisation reactions,
conditions used were similar to that of cyt c-10-BSA as cytochrome c is the least
reactive protein, as the free cysteine residue (CYS102) is buried in a hydrophobic
pocket.17b Reaction time varies between different bioconjugation dimer reactions as it is
not a critical factor for dimerisation due to hydrolysis of the maleimide group35-36 and
only requires a minimum overnight incubation to reach completion.17b
Scheme 3.5. Synthesis of bioconjugate cyt c-10-cyt c. (a) Reduced iso-1 cytochrome c(100 M), 27 h. Phosphate buffer (20 mM), ethylenediaminetetraacetic acid (20 mM), pH 7.0, acetonitrile (5% v/v), r.t., 1%.
The crude product was loaded onto an IMAC (Ni2+) column and purified over a gradient
from 0 to 250 mM imidazole over 9 mL in 20 mM sodium dihydrogen phosphate, 0.5 M

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-21
sodium chloride at pH 7.0 as shown in Figure 3.12. Prior to the imidazole gradient,
elution of unreacted iso-1 cytochrome c and ligand was achieved. Product containing
fraction A was eluted after the imidazole gradient, pooled, concentrated and dialysed
against water. This afforded the bioconjugate cyt c-10-cyt c with moderate purity and
low yield of 1%. Due to the low yield of the homodimer, isolation with high purity is
extremely difficult.
Figure 3.11. Purification of bioconjugate cyt c-10-cyt c. IMAC (Ni2+) chromatography (HisTrapTM HP, GE Healthcare) using a gradient from 0 to 250 mM imidazole in 20 mMphosphate buffer, 0.5 M sodium chloride, pH 7.0 in 9 mL at 0.5 mL/min. Peak A –bioconjugate cyt c-10-cyt c. The gradient (green) is shown for illustration.
UV-Vis spectroscopy shows characteristic absorption bands at 410 nm and 495 nm
corresponding to cyt c and complex 10 as shown in Figure 3.8 and dimer was further
confirmed by SDS PAGE gel electrophoresis as shown in Figure 3.10 showing a new
dimer band. The formation of dimer was additionally confirmed by MALDI-TOF mass
spectrometry as shown in Figure 3.12 with a m/z of 26 551.

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-22
Figure 3.12. MALDI-TOF mass spectrum of cyt c-10-cyt c. Peaks corresponding to the calculated masses 25 412 and 26 550 Da, respectively, were detected. Spectrum was baseline corrected.
3.4.3 Synthesis of cyt c-16-GFP
In order to probe the effect of dimer yields based on a heterodimer of similar positive
charge, a cyt c-16-GFP heterodimer was prepared. The spacer chosen for this
bioconjugate was based on a bipyridinium (viologen) moiety 16 which is more water
soluble in comparison to complex 10. The dimer was prepared by addition of a solution
of 4,4’-bipyridinium-N,N-di(maleimidopropyl) hexafluorophosphate 16 (fifteen-fold) in
acetonitrile to a solution of green fluorescent protein (GFP, amFP497) derived from
Acropora millepora in phosphate/EDTA buffer at pH 7.0 for 3 h in the dark as shown in
Scheme 3.6. The functionalisation of recombinant GFP with maleimide could be
achieved due to the single cysteine residue at CYS119 introduced by mutation of the
genetic sequence.37

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-23
Subsequently, the crude product was desalted to remove excess viologen spacer
as shown in Figure 3.13a. Purified reduced iso-1 cytochrome c (four-fold) was added to
the desalted product fraction and reacted for a further 17 h in phosphate/EDTA buffer at
pH 7.0 in the dark.
Scheme 3.6. Synthesis of bioconjugate cyt c-16-GFP. (a) i) Green fluorescent protein (10 M), 3 h, ii) Cyt c-16 (70 M), reduced iso-1 cytochrome c (290 M), 17 h.Phosphate buffer (20 mM), ethylenediaminetetraacetic acid (20 mM), pH 7.0, acetonitrile (5% v/v), r.t., 0.1%. Green fluorescent protein PDB code: 2A4638, aligned with engineered protein sequence for amFP497.
It was proposed that the extremely low yield was a result of CYS102 being buried in the
hydrophobic pocket of cytochrome c.5 Therefore, an alternative approach to synthesise
cyt c-16-GFP was attempted by preparing cyt c-16 prior to addition of green fluorescent
protein containing surface exposed CYS119. However, no improvement in heterodimer
yield was observed. It should be noted that a cytochrome c-GFP heterodimer has been
previously prepared via recombinant methods as a fused product in the mitochondria

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-24
rather than using an organic linker.39 Additionally, bioconjugation variants of GFP for
use as potential donor-acceptor systems are further discussed in Chapter 4.
Recombinant GFP was expressed with a hexahistidine-tag (6×His-tag) allowing
purification using IMAC chromatography.40 The desalted intermediate product was
loaded onto an IMAC column and purified over a gradient from 0 to 250 mM imidazole
in 20 mm sodium dihydrogen phosphate, 0.5 m sodium chloride at pH 7.0. Prior to the
imidazole gradient, elution of unreacted cytochrome c (peak C) occurs. During the
imidazole gradient, further elution of unreacted cytochrome c and unreacted GFP (peak
D and F) was achieved. Product containing fraction E of bioconjugate cyt c-16-GFP is
eluted, pooled, concentrated and dialysed against water. This afforded the bioconjugate
in an extremely low yield of 0.1%. Residual cytochrome c and GFP was present in the
product fraction due to difficulty in purification for low yielding species.
Figure 3.13. Purification of bioconjugate cyt c-16-GFP. (a) Purification of bioconjugate GFP-16 by fast protein liquid chromatography using a desalting column (HiTrapTM Desalting, GE Healthcare) in 20 mM phosphate buffer, pH 7.0 at 0.5 mL/min. Peak A – mixed GFP and GFP-16. Peak B – unreacted 16. (b) Purification of bioconjugate cyt c-16-GFP using an IMAC (Ni2+) column (HisTrapTM HP, GE Healthcare) using a gradient from 0 to 250 mM imidazole in 20 mM phosphate buffer, 0.5 M sodium chloride, pH 7.0 in 6 mL at 0.5mL/min. Peak C – unreacted cyt c, peak D and F - unreacted cyt c and GFP, peak E – bioconjugate cyt c-16-GFP. The gradient (green) is shown for illustration.
UV-Vis spectroscopy shows characteristic absorption bands at 410 nm and 476 nm
corresponding to cytochrome c and GFP as shown in Figure 3.8. The heterodimer was

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-25
also confirmed by reduced SDS PAGE gel electrophoresis showing a new dimer band
indicated in Figure 3.10. Finally, the heterodimer was confirmed by MALDI-TOF mass
spectrometry as shown in Figure 3.14 with a m/z of 40 245.
Figure 3.14. MALDI-TOF mass spectrum of cyt c-16-GFP. Peaks corresponding to calculated mass 40 236 Da was detected. Spectrum was baseline corrected and noise reduced.
3.4.4 Synthesis of BSA-10-BSA
A homodimer of negative charge was prepared by forming a BSA homodimer using
bismaleimide complex 10 as shown in Scheme 3.7. A solution of complex 10 (as
chloride salt) in acetonitrile was added to fourty-fold BSA in a final 20 mM
phosphate/EDTA buffer, acetonitrile (5% v/v) at pH 7.0 for 23 h at room temperature in
the dark. The crude was concentrated, dialysed against water and the crude was purified
by IMAC chromatography.

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-26
Scheme 3.7. Synthesis of bioconjugate BSA-10-BSA. (a) Bovine serum albumin (400 M), 23 h. Phosphate buffer (20 mM), ethylenediaminetetraacetic acid (20 mM), pH 7.0, acetonitrile (5% v/v), r.t., 1%.
The crude was loaded onto an IMAC column and purified over a gradient from 0 to
125 mM imidazole over 6 mL in 20 mM sodium dihydrogen phosphate, 0.5 M sodium
chloride at pH 7.0 as shown in Figure 3.15. Elution of unreacted ligand 10 and BSA
occurred prior to the gradient (peak A). Product containing fraction B eluted during the
imidazole gradient and was pooled, concentrated and dialysed against water to afford
the homodimer in 1% yield.

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-27
Figure 3.15. Purification of bioconjugate BSA-10-BSA by IMAC (Ni2+)chromatography (HisTrapTM HP, GE Healthcare) using a gradient from 0 to 125 mMimidazole in 20 mM phosphate buffer, 0.5 M sodium chloride, pH 7.0 in 6 mL at 0.5 mL/min. Peak A – unreacted 10 and BSA, peak B – BSA-10-BSA. The gradient (green) is shown for illustration.
UV-Vis spectroscopy shows characteristics absorption bands at 280 nm and 495 nm due
to BSA and spacer 10, respectively, as shown in Figure 3.8. Additionally, reduced gel
electrophoresis shows a new band corresponding to dimer formation as shown in Figure
3.10. Due to the high molecular weight (>130 kDa), the BSA homodimer could not be
detected by MALDI-TOF mass spectrometry.
3.4.5 Effect of charge on protein dimer yield
The synthesis of bioconjugate dimers discussed above showed that yields may be
dependent of protein charge. The role and importance of protein charge to probe protein
function after amino acid modification has been utilised in the literature, such as the use
of protein charge ladders.41 However, the exploitation of global protein charge to

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-28
facilitate dimer bioconjugate synthesis via supramolecular interactions has been
neglected.
In this study it was found that a heterodimer of complementary charge and high
molecular weight (>80 kDa), cyt c-10-BSA, could be prepared in up to 30% yield
overcoming steric hindrance as shown in Table 3.1. Cytochrome c is a small protein
(12.7 kDa) with positive charge at physiological pH 7.4 as it possesses a high positive
pI of 10.63 and BSA is a negatively charged large protein (66.7 kDa) with a negative pI
of 4.7.34b
In addition, a positively charged homodimer cyt c-10-cyt c was synthesised and
resulted in a low yield of 1%. Similarly, the positively charged heterodimer
cyt c-16-GFP, which used green fluorescent protein as a like charged protein with a
molecular weight of 26.8 kDa and a high positive pI of 8.342 resulted in extremely low
yields of less than 1%. Additionally, the negatively charged homodimer, BSA-10-BSA
also resulted in an extremely low yield of 1%. Based on these like-charged homo and
heterodimers, it appears the low yields are due to unfavourable dimer formation due to
electrostatic repulsion.
Table 3.1. Summary of conjugate dimer pairings.
Entry Conjugate Protein 1 (pI) Protein 2 (pI) Charge Yielda
1 cyt c-10-BSA cyt c (10.6) BSA (4.7) +/- 30%2 cyt c-10-cyt c cyt c (10.6) cyt c (10.6) +/+ 1%3 cyt c-16-GFP cyt c (10.6) GFP (8.3)b +/+ 0.1%4 BSA-10-BSA BSA (4.7) BSA (4.7) -/- 1%
a Estimated by gel electrophoresis and UV-Vis spectroscopy. b pI for GFP estimated by theoretical calculations of peptide sequence.
To exclude the possibility that the higher yielding complementary charged heterodimer
cyt c-10-BSA was not induced by localised charge effects, but rather global charge, the
protein electrostatic surface of cytochrome c, green fluorescent protein and bovine

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-29
serum albumin was modelled by PyMol (version 1.3). The model assumes that the
positively charged cytochrome c and green fluorescent protein possess 2-8 positive
charges at physiological pH 7.4 and negatively charged bovine serum albumin has
approximately 13 negatively charged residues. From Figure 3.16, the solvent accessible
single cysteine residues of the proteins do not appear to be in particularly negatively or
positively charged regions on the calculated electrostatic surface.43
Figure 3.16. The three proteins used in this model and their key properties. For bovine serum albumin (BSA), the sequence of BSA (ExPASy code: P02769) has been aligned onto the X-ray crystallographic structure of human serum albumin (PDB code: 1AO6). Similarly, the modified Acropora millepora green fluorescent protein (GFP) sequence37
has been aligned with another Anemonia majano GFP structure (PDB code: 2A46). The sequence alignment was performed using ClustalW2 (http:www.ebi.ac.uk/Tools/clustalw2/). The structure for iso-1 cytochrome c (cyt c) was used without further modification (PDB code: 1YCC). Negative (red), neutral (white)and positive (blue) electrostatic surface features are presented according to the inserted polarised scale. Images were generated with PyMol (Version 1.3, Schrödinger, LLC) using the APBS plug-in to calculate the electrostatic surface potentical.43 The pI values for BSA34b and cyt c3 were obtained from the literature while for GFP, it has been estimated by theoretical calculations based on its sequence.42 The number of + or –charges refers to the net whole charges at pH =7.4 assuming only Lys (+), Arg(+), Glu (–) and Asn (–) are charged at that pH (the N- and C-protein terminus cancel each other out). The target cysteine residues for bioconjugation are coloured in green and indicated by an arrow.

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-30
3.5 Conclusion and Future Work
In this Chapter, the successful synthesis of an asymmetric terpyridine cytochrome c
bioconjugate 8-cyt c was prepared for electron transfer studies in membranes and for
use as a component in a polymer membrane reconstituted electron transfer chain as a
photosynthetic-respiratory hybrid which is further discussed in Chapter 6 and 7,
respectively. The purification of ruthenium(II)-bisterpyridine based bioconjugate could
be achieved using immobilised metal affinity chromatography (IMAC, Ni2+) in high
purity with 6% yield based on procedures previously developed in the Thordarson
group by Peterson et al.17a although the exact mechanism remains unknown. The
limitation of low yielding asymmetric bioconjugate 8-cyt c is a result of the poor water
solubility, and this issue could be addressed further in the future by preparing more
water soluble ligands based on polyethylene glycol modified
ruthenium(II)-bisterpyridine complexes.
Several bioconjugate dimers have been prepared based on combinations of
cytochrome c, green fluorescent protein (Acropora millepora) and bovine serum
albumin to probe factors affecting dimerisation yield. It was found that a high molecular
weight heterodimer of complementary charge cyt c-10-BSA (>80 kDa) was able to be
prepared in up to 30% yield. It is noteworthy that like-charged homo and heterodimers
were prepared in extremely low yields of less than 1%. Based on these studies, it is
proposed that global protein charge can be utilised to induce higher dimer yields by
electrostatic supramolecular ionic attraction. Additionally, the absolute purification of
dimers proved to be difficult due to the low yields of the dimers. In order to confirm
that electrostatic attraction can affect bioconjugate yields, further experiments including
pH titrations on dimer formation (pI effect), Curtin-Hammett regimes44 and

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-31
involvement of other proteins (site directed mutagenesis to introduce CYS) should be
conducted in the future.
The conjugates were characterised by MALDI-TOF mass spectrometry, UV-Vis
spectroscopy and gel electrophoresis.
3.6 References
(1) Keilin, D. Proc. R. Soc. Lond. B 1930, 106, 418.(2) Keilin, D.; Hartree, E. F. Proc. R. Soc. Lond. B 1937, 122, 298.(3) Minakami, S. J. Biochem. 1955, 42, 749.(4) Peterson, J. R.; Thordarson, P. Chiang Mai J. Sci. 2009, 26, 236.(5) Louie, G. V.; Brayer, G. D. J. Mol. Biol. 1990, 214, 527.(6) (a) Millett, F.; Durham, B. Methods Enzymol. 2009, 456, 95. (b) Geren, L.;
Durham, B.; Millett, F. Methods Enzymol. 2009, 456, 507.(7) (a) Cordes, M.; Giese, B. Chem. Soc. Rev. 2009, 38, 892. (b) Gray, H. B.;
Winkler, J. R. Annu. Rev. Biochem. 1996, 65, 537.(8) (a) Winkler, J. R.; Nocera, D. G.; Yocom, K. M.; Bordignon, E.; Gray, H. B. J.
Am. Chem. Soc. 1982, 104, 5798. (b) Yang, X.-J.; Drepper, F.; Wu, B.; Sun, W.-H.; Haehnel, W.; Janiak, C. Dalton Trans. 2005, 256.
(9) (a) Pan, L. P.; Durham, B.; Wolinska, J.; Millett, F. Biochemistry 1988, 27,7180. (b) Hahm, S.; Durham, B.; Millett, F. Biochemistry 1992, 31, 3472.
(10) Wang, K.; Mei, H.; Geren, L.; Miller, M. A.; Saunders, A.; Wang, X.; Waldner, J. L.; Pielak, G. J.; Durham, B.; Millett, F. Biochemistry 1996, 35, 15107.
(11) Hvasanov, D.; Goldstein, D. C.; Thordarson, P. In Molecular Solar Fuels; The Royal Society of Chemistry, 2012; p 426.
(12) (a) Thordarson, P.; Droumaguet, B. L.; Velonia, K. Appl. Microbiol. Biotechnol.2006, 73, 243. (b) Veronese, F. M.; Pasut, G. Drug Discov. Today 2005, 10,1451.
(13) Wu, G.; Ott, T. L.; Knabe, D. A.; Bazer, F. W. J. Nutr. 1999, 129, 1031.(14) Edwards, J. O.; Pearson, R. G. J. Am. Chem. Soc. 1962, 84, 16.(15) Hermanson, G. T. Bioconjugate Techniques; 2nd ed.; Elsevier Inc.: San Diego,
CA, 2008.(16) Hodgson, D. R. W.; Sanderson, J. M. Chem. Soc. Rev. 2004, 33, 422.(17) (a) Peterson, J. R.; Smith, T. A.; Thordarson, P. Chem. Commun. 2007, 1899. (b)
Peterson, J. R.; Smith, T. A.; Thordarson, P. Org. Biomol. Chem. 2010, 8, 151.(18) Kaminsky, L. S.; Davison, A. J. Biochemistry 1969, 8, 4631.(19) Geren, L.; Hahm, S.; Durham, B.; Millett, F. Biochemistry 1991, 30, 9450.(20) Yang, H.; Liu, N.; Qiu, X.; Liu, S. J. Am. Soc. Mass Spectrom. 2009, 20, 2284.(21) Carlotti, F.; Zaldumbide, A.; Martin, P.; Boulukos, K. E.; Hoeben, R. C.;
Pognonec, P. Cancer Gene Ther. 2005, 12, 627.(22) Spencer, D. M.; Belshaw, P. J.; Chen, L.; Ho, S. N.; Randazzo, F.; Crabtree, G.
R.; Schreiber, S. L. Curr. Biol. 1996, 6, 839.(23) Press, O. W.; Corcoran, M.; Subbiah, K.; Hamlin, D. K.; Wilbur, D. S.; Johnson,
T.; Theodore, L.; Yau, E.; Mallett, R.; Meyer, D. L.; Axworthy, D. Blood 2001,98, 2535.

Chapter 3 Bioconjugate Synthesis of Cytochrome c and Related Derivatives
3-32
(24) Quintarelli, C.; Vera, J. F.; Savoldo, B.; Giordano Attianese, G. M. P.; Pule, M.; Foster, A. E.; Heslop, H. E.; Rooney, C. M.; Brenner, M. K.; Dotti, G. Blood2007, 110, 2793.
(25) (a) Ziaco, B.; Pensato, S.; D’Andrea, L. D.; Benedetti, E.; Romanelli, A. Org. Lett. 2008, 10, 1955. (b) Xiao, J.; Tolbert, T. J. Org. Lett. 2009, 11, 4144. (c) Eger, S.; Scheffner, M.; Marx, A.; Rubini, M. J. Am. Chem. Soc. 2010, 132,16337. (d) Li, X. Chem. Asian. J. 2011, 6, 2606.
(26) Natarajan, A.; Du, W.; Xiong, C.-Y.; DeNardo, G. L.; DeNardo, S. J.; Gervay-Hague, J. Chem. Commun. 2007, 695.
(27) (a) Foot, J. S.; Lui, F. E.; Kluger, R. Chem. Commun. 2009, 7315. (b) Yang, Y.; Kluger, R. Chem. Commun. 2010, 46, 7557.
(28) Xiao, J.; Hamilton, B. S.; Tolbert, T. J. Bioconjug. Chem. 2010, 21, 1943.(29) Komatsu, T.; Oguro, Y.; Teramura, Y.; Takeoka, S.; Okai, J.; Anraku, M.;
Otagiri, M.; Tsuchida, E. Biochim. Biophys. Acta, Gen. Subj. 2004, 1675, 21.(30) Hatzakis, N. S.; Engelkamp, H.; Velonia, K.; Hofkens, J.; Christianen, P. C. M.;
Svendsen, A.; Patkar, S. A.; Vind, J.; Maan, J. C.; Rowan, A. E.; Nolte, R. J. M. Chem. Commun. 2006, 2012.
(31) Loughran, M.; Tsai, S.-W.; Yokoyama, K.; Karube, I. Curr. Appl. Phys. 2003, 3,495.
(32) Natarajan, A.; Xiong, C.-Y.; Albrecht, H.; DeNardo, G. L.; DeNardo, S. J. Bioconjug. Chem. 2004, 16, 113.
(33) Riener, C.; Kada, G.; Gruber, H. Anal. Bioanal. Chem. 2002, 373, 266.(34) (a) Kilty, C. G.; John, B. R.; John, M. D.; Biotrin Intellectual Properties Limited,
Ire., 2005. (b) Ang, W. S.; Elimelech, M. J. Membr. Sci. 2007, 296, 83.(35) Zöphel, L.; Eisele, K.; Gropeanu, R.; Rouhanipour, A.; Koynov, K.;
Lieberwirth, I.; Müllen, K.; Weil, T. Macromol. Chem. Phys. 2010, 211, 146.(36) Gregory, J. D. J. Am. Chem. Soc. 1955, 77, 3922.(37) Angelo, C.; Denzel, A.; Vogt, A.; Matz, M. V.; Oswald, F.; Salih, A.; Nienhaus,
G. U.; Wiedenmann, J. Mar. Ecol. Prog. Ser. 2008, 364, 97.(38) Henderson, J. N.; Remington, S. J. Proc. Natl. Acad. Sci. U. S. A. 2005, 102,
12712.(39) Goldstein, J. C.; Munoz-Pinedo, C.; Ricci, J. E.; Adams, S. R.; Kelekar, A.;
Schuler, M.; Tsien, R. Y.; Green, D. R. Cell Death Differ. 2005, 12, 453.(40) Hochuli, E.; Bannwarth, W.; Dobeli, H.; Gentz, R.; Stuber, D. Nat. Biotech.
1988, 6, 1321.(41) (a) Gitlin, I.; Carbeck, J. D.; Whitesides, G. M. Angew. Chem. Int. Ed. 2006, 45,
3022. (b) Gitlin, I.; Mayer, M.; Whitesides, G. M. J. Phys. Chem. B 2003, 107,1466.
(42) Hvasanov, D.; Wiedenmann, J.; Braet, F.; Thordarson, P. Chem. Commun. 2011,47, 6314.
(43) Baker, N. A.; Sept, D.; Joseph, S.; Holst, M. J.; McCammon, J. A. Proc. Natl. Acad. Soc. U. S. A. 2001, 98, 10037.
(44) Seeman, J. I. Chem. Rev. 1983, 83, 83.

Chapter 4
Green Fluorescent Protein as a Light-Induced Electron Donor

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-2
4 Green Fluorescent Protein as a Light-Induced Electron Donor
In this Chapter, the synthesis of a mutant green fluorescent protein (GFP) bioconjugate
as a potential covalent donor-acceptor system based on the N-hydroxysuccinimide
anthraquinone 12 acceptor was prepared using non-specific lysine modification.
Additionally, green fluorescent protein has been mutated to contain a single cysteine
residue (CYS119)1 using recombinant methods and is derived from the Acropora
millepora species. The synthesis of site-specific functionalised GFP as potential
donor-acceptor systems using a 4,4’-bipyridinium (viologen) derivative 16 was
achieved and a Cu(I)-catalysed [3+2] Huisgen cycloaddition using azide functionalised
anthraquinone 13 attempted.
Light-induced electron transfer studies between GFP (electron donor) and
p-benzoquinone, anthraquinone and viologen electron acceptors as both non-covalent
and bioconjugate mixtures were monitored using steady-state techniques including
UV-Vis and fluorescence spectroscopy and performed using a Xenon arc lamp or
custom built LED array. The fluorescence lifetime of = 1.65±0.06 ns was determined
for GFP. Additionally, quenched lifetimes due to energy/electron transfer of 0.14±0.01
and 0.23±0.04 ns with corresponding electron transfer rates of ket = 6.5±0.5 × 109 and
3.7±0.7 × 109 s-1 were determined for a non-covalent mixture of GFP with
p-benzoquinone or anthraquinone-2-carboxylic acid, respectively. The fluorescence
lifetime for bioconjugate 12-GFP indicated electron transfer with a quenched lifetime of
0.28±0.04 ns (ket = 2.9±0.4 × 109 s-1).
4.1 Background
Over the last two decades, the green fluorescent protein (GFP) has become one of the
most utilised fluorescent probes in cell biology and molecular biology.2 This is due to

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-3
the ability to fuse GFP to proteins of interest3 and express the resulting gene in vivo to
function as a visual indicator using standard fluorescence microscopy techniques. GFP
has been used for protein localisation4 in cells or organisms or in protein-protein
interaction studies5 via fragment reassembly or Förster resonance energy transfer.
The prevalent role of GFP as a fluorescent probe in cell biology eventually led
to the award of the 2008 Nobel Prize in Chemistry shared by Professors Osamu
Shimomura, Martin Chalfie and Roger Y. Tsien. Wild type GFP (wtGFP) from the
jellyfish Aequorea victoria was first isolated by Shimomura et al.6 in 1962. Chalfie and
co-workers4 demonstrated the versatility of GFP as a visual probe in organisms and
Tsien et al.7 expanded their applications by introducing chromophore mutations to
create spectroscopic variants.
GFPs can be found in natural sources of jellyfish (Aequorea victoria), sea corals
and sea anemones.2c GFP is an approximately 230 amino acid residue fluorescent
protein with monomer molecular weight of 26.8 kDa as shown in Figure 4.1.8 The high
quantum yield and unique fluorescent properties of the protein is a result of sequestering
of the chromophore from the bulk solvent in an -helix that runs down the centre of an
11-stranded -barrel and prevents eventual nonradiative cis to trans isomerisation of the
central chromophore.9

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-4
Figure 4.1. Solid ribbon representation of green fluorescent protein (Acropora millepora) with tripeptide chromophore (GLN68-TYR69-GLY70) and CYS119 residue indicated as stick representations. The modified protein sequence was aligned against another Anemonia majano GFP structure from the protein data bank file ‘2A46’.10
The wild type GFP chromophore is formed via an intramolecular autocatalytic
generation from the tripeptide SER65-TYR66-GLY67 as a
p-hydroxybenzylidene-imidazolidinone chromophore11 with coplanar cis
conformation.12 The chromophore in wild type GFP and variants from different species
and mutations can exist in two tautomeric forms as shown in Figure 4.2, including the
neutral and anionic form responsible for absorption centred near 395 and 475 nm,
respectively.13 The tautomeric forms of the chromophore result in the same emission
band due to ultrafast excited state proton transfer from the neutral to anionic form upon
photoexcitation.14

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-5
Figure 4.2. Tautomeric forms of the green fluorescent (GFP) chromophore. Neutral form is the protonated phenolic form of the chromophore with a keto oxygen on the imidazole ring responsible for the absorption band centred near 395 nm. Anionic form is responsible for the absorption band centred near 475 nm. The equilibrium ratio is dependent on electrostatic interactions between the chromophore and surroundingresidues and are dependent on GFP variants and mutations.9a
The biological purpose and function of bioluminescence of GFPs in jellyfish and sea
corals remain unknown. However, GFP may participate in photochemical reactions with
oxidants as reported by Lukyanov and co-workers who have shown that GFPs from
different species can act as light-induced electron donors.15 In this chapter, mutant GFP
based bioconjugates derived from Acropora millepora (amFP497) with viologen or
quinones as potential electron acceptors were prepared with a GLN68-TYR69-GLY70
chromophore to explore potential light-induced electron transfer reactions in GFP.1
4.2 Synthesis of GFP-Acceptor Bioconjugates
Viologen and quinone derivatives are the most widely studied electron acceptors.16 In
this project, covalent modification of GFP viologen 16 and NHS-anthraquinone 12 was
achieved using cysteine-maleimide coupling17 or amine (lysine) modification,
respectively.18

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-6
4.2.1 Attempted synthesis of anthraquinone-triazole-GFP via click chemistry
The synthesis of a 13-triazole-GFP bioconjugate as a light-activated donor-acceptor
system was attempted using Cu(I)-catalysed [3+2] Huisgen cycloaddition between a
terminal alkyne and azide, also known as the click reaction. Since the initial report of
the Cu(I)-catalysed click reaction between alkyne and azides in 2002 by Sharpless et
al.19, this approach has been a popular strategy for bioconjugation reactions.20 The click
reaction offers advantages over other bioconjugation techniques such as high coupling
efficiency and high selectivity which prevents side reactions with other functional
groups in a reaction mixture, even allowing reactions within cells.21 Furthermore, due to
the weak acid-base properties of azides and alkynes, it is suitable for protein
modification due to benign reaction conditions as well as its chemoselectivity.20b
In order to perform click chemistry on GFP, the alkyne precursor bioconjugate
was prepared as shown in Scheme 4.1. This bioconjugation was performed by reacting
alkyne-maleimide 17 with mutant GFP containing a single cysteine residue (CYS119)
as discussed in Chapter 3. This strategy using the same alkyne-maleimide has been
reported previously to modify bovine serum albumin (BSA).22 GFP was added to a
solution containing ten-fold excess of alkyne-maleimide 17 in a phosphate buffer at
pH 7.0 containing ethylenediaminetetraacetic acid (EDTA) and
N,N-dimethylformamide. The final reaction conditions were GFP (10 M),
alkyne-malemide 17 (100 M), phosphate buffer (20 mM), EDTA (20 mM), and
N,N-dimethylformamide (5% v/v) at pH 7.0. The mixture was stirred for 16.5 h at room
temperature in the dark. The crude mixture was concentrated to ca. 1 mL and desalted
to remove excess alkyne-maleimide 17. Throughout this chapter, yield is based on
UV-Vis absorbance using the molar absorptivity of GFP (amFP497,

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-7
476 = 31.42 mM-1cm-1) and the yield is interchangeably expressed as both dry weight
and the moles of protein.
Scheme 4.1. Synthesis of bioconjugate 17-GFP. (a) Green fluorescent protein (10 M), phosphate buffer (20 mM), ethylenediaminetetraacetic acid (20 mM), pH 7.0,N,N-dimethylformamide (5% v/v), r.t., 50%.
The characterisation of alkyne-tagged GFP, 17-GFP by MALDI-TOF mass
spectrometry showed complete conversion into the modified product after desalting as
shown in Figure 4.3 with a product signal of m/z 26 967. Due to the small change in
molecular weight, gel electrophoresis did not show any significant change in band
migration. It should be noted that the direct bioconjugation approach could not be used
as the synthesis of anthraquinone-maleimide 15 was unsuccessful which was discussed
previously in Chapter 2.

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-8
Figure 4.3. MALDI-TOF mass spectrum of alkyne-tagged GFP (17-GFP). Peak corresponding to the calculated mass of 26 965 Da was detected. Spectrum was baseline corrected and noise reduced.
The attempted synthesis of anthraquinone functionalised GFP was attempted using
anthraquinone-azide 13 via click chemistry. Anthraquinone was selected due to
improved stabilities compared to p-benzoquinone which can degrade into quinhydrones
when isolated or stored in solution.23 The conditions used were based on conditions
optimised previously in the Thordarson group by Goldstein et al.24 A solution of a
preformed Cu(I) source, tetrakis(acetonitrile)copper(I) hexafluorophosphate and azide
13 in N,N-dimethylformamide was added to alkyne functionalised GFP (17-GFP) in
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) buffer at pH 7.0 for 19 h
in the dark at 30 oC with a nitrogen overlay. Significant precipitation was observed after
incubation. The filtrate was characterised after syringe filtration using a 0.2 m
membrane, however, UV-Vis spectroscopy and size exclusion chromatography (HiTrap

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-9
DesaltingTM, GE Healthcare) showed no trace of protein, indicating complete
denaturation of protein.
It was proposed that protein denaturation may have been a result of the
formation of a histidine-Cu(II) chelation complex25 (recombinant GFP expressed with
hexahistidine-tag) or Cu(I)-acetylide intermediate.26 In order to address these issues, a
control experiment under identical reaction conditions containing all components was
repeated using unmodified GFP. Precipitation was observed within 30 min of stirring
indicating protein denaturation due to presence of Cu(II) from oxidation. Based on this
control experiment, the primary factor causing denaturation is a result of the
histidine-Cu(II) chelate as GFP has been previously modified using click chemistry with
polymersomes which did not possess a hexahistidine-tag.27
4.2.2 Synthesis of anthraquinone-GFP (12-GFP) via amine modification
The anthraquinone-GFP bioconjugate (12-GFP) was prepared by non-specific lysine
modification using an NHS-functionalised anthraquinone 12 as shown in Scheme 4.2. A
solution of GFP was added to crude anthraquinone 12 in aqueous sodium bicarbonate
and N,N-dimethylformamide at pH 8.3. The final reaction conditions were GFP
(10 M), anthraquinone 12 ( 100 M), aqueous sodium bicarbonate (50 mM),
N,N-dimethylformamide (5% v/v) at pH 8.3. The mixture was reacted for 16.5 h,
concentrated to ca. 1 mL and desalted by size exclusion chromatography affording the
bioconjugate in 27% yield, based on total protein concentration.

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-10
Scheme 4.2. Synthesis of anthraquinone-GFP (12-GFP). (a) Green fluorescent protein (10 M), aqueous sodium bicarbonate (50 mM), pH 8.3, N,N-dimethylformamide (5% v/v), r.t., 27%.
Characterisation of the desalted product by MALDI-TOF mass spectrometry shows a
mixture of unmodified GFP and singly and doubly labelled 12-GFP in an approximate
ratio of 1:1:1 based on peak intensity as shown in Figure 4.4.
Figure 4.4. MALDI-TOF mass spectrum of anthraquinone-GFP (12-GFP). Peaks corresponding to the calculated masses of 26 807, 27 041 and 27 275 Da, respectively, were detected. Spectrum was baseline corrected and noise reduced.

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-11
The UV-Vis absorption spectrum of the anthraquinone bioconjugate 12-GFP shows the
characteristic absorption peaks of both GFP (476 nm) and the anthraquinone moiety 12
(258 nm). Based on the spectra shown in Figure 4.5 and the molar absorptivities of GFP
(amFP497, 476 = 31.42 mM-1cm-1) and anthraquinone 14 ( 258 = 29.62 mM-1cm-1),
deconvolution of the spectra indicated that the desalted product was composed of 25%
bioconjugate 14-GFP relative to unmodified GFP. UV-Vis spectroscopy provides a
more accurate approach to determine composition in contrast to MALDI-TOF mass
spectrometry as the ionisation efficiency of different molecular weight species varies. It
should be noted that anthraquinone-bromide 14 was chosen as the reference compound
as anthraquinone-2-carboxylic acid displays poor solubility in organic solvents and
water.
Figure 4.5. UV-Vis spectra of 14-GFP (green, H2O) and approximated by the linear sum (blue dashed) of green fluorescent protein (red, H2O) and anthraquinone 14(black, CH2Cl2).

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-12
4.2.3 Synthesis of viologen-GFP (16-GFP)
The potential donor-acceptor system based on viologen (16-GFP) was prepared using
cysteine-maleimide chemistry previously discussed in Chapter 3. GFP was added to a
solution containing ten-fold excess of bismaleimide viologen 16 in a phosphate buffer at
pH 7.0 containing EDTA and acetonitrile. The final reaction conditions were GFP
(10 M), viologen 16 (100 M), phosphate buffer (20 mM), EDTA (20 mM) and
acetonitrile (5% v/v) at pH 7.0 as shown in Scheme 4.3. The mixture was stirred
overnight at room temperature in the dark and quenched with 2-mercaptoethanol.
Scheme 4.3. Synthesis of viologen-GFP (16-GFP). (a) Green fluorescent protein (10 M), phosphate buffer (20 mM), ethylenediaminetetraacetic acid (20 mM), pH 7.0,acetonitrile (5% v/v), r.t., 11%.
The crude was concentrated to ca. 1 mL and purified by Ni2+ immobilised metal affinity
chromatography using an imidazole gradient from 0 to 500 mM over 16 mL in 20 mM
sodium dihydrogen phosphate, 0.5 M sodium chloride at pH 7.0 as shown in Figure 4.6.

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-13
Prior to the imidazole gradient, elution of unreacted ligand 16 (peak A) occurred. Both
unreacted GFP (peak B) and product containing fraction (peak C) as a mixture of
unmodified protein and bioconjugate 16-GFP were eluted after the imidazole gradient.
The product containing fraction was concentrated and dialysed against water affording
the product in 11% yield.
Figure 4.6. Purification of bioconjugate 16-GFP by IMAC (Ni2+) chromatography (HisTrapTM HP, GE Healthcare) using a gradient from 0 to 500 mM imidazole in 20 mMphosphate buffer, 0.5 M sodium chloride, pH 7.0 in 16 mL at 0.5 mL/min. Peak A –unreacted ligand 16, peak B – unmodified GFP and peak C – bioconjugate 16-GFP (mixture of unmodified GFP and bioconjugate).
Characterisation of bioconjugate 16-GFP by MALDI-TOF mass spectrometry spiked
with unmodified GFP showed the single site-specific attachment of ligand 16 with a
measured mass of 27 246 Da and is in agreement with the expected value of 27 239 Da
as shown in Figure 4.7.

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-14
Figure 4.7. MALDI-TOF mass spectrum of bioconjugate 16-GFP spiked with unmodified green fluorescent protein. Peaks corresponding to the calculated masses 26 807 and 27 239 Da, respectively, were detected. Spectrum was baseline correctedand noise reduced.
4.3 GFP Donor-Acceptor Studies
Electron transfer reactions of redox proteins including the electron transport chain for
photosynthesis and respiration are critical to the biological function of organisms.
Electron transfer reactions have been primarily focused on redox proteins including
cytochrome c28, however, it has been reported recently that a possible role of GFP is a
potential light-induced electron donor in the presence of suitable acceptors.15
GFP undergoes photoconversion into a red fluorescent state under anaerobic
conditions29 where the oxygen concentration is below 1% without external agents.30 On
the other hand, light-induced red photoconversion of GFP in the presence of electron
acceptors from a green to red fluorescent state under aerobic conditions has been
reported by Bogdanov et. al.15, known as oxidative redding. The proposed red

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-15
photoconversion mechanism is a one-photon and step-wise two-electron oxidation
process as shown in Scheme 4.1, however, the exact nature of the red chromophore is
unknown. In order for oxidative redding to occur, a tyrosine residue is required in the
protein tripeptide chromophore.
Scheme 4.4. Proposed photoinduced aerobic red photoconversion mechanism. An excited green chromophore (Chrg) transfers one electron to an acceptor molecule (A) resulting in a short-lived intermediate (Chr +). If the intermediate can transfer a second electron in its lifetime, a red chromophore (Chrr) is formed; otherwise, a permanent bleached chromophore (Chrbl) is formed.15
Quinone and viologen based electron acceptors are suitable as potential acceptor
molecules of GFP as quinone can be reduced via a two-electron process31 as shown in
Scheme 4.5a and viologen can be reduced to form a mono-reduced species which can be
detected spectroscopically as shown in Scheme 4.5b.32
Scheme 4.5. Reversible conversion of oxidised/reduced states of electron acceptors. (a) p-benzoquinone. (b) viologen.

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-16
4.3.1 Steady-state spectroscopy studies
4.3.1.1 Viologen 16 acceptor studies
To monitor whether GFP (amFP497) could be a potential light-induced electron donor,
initial studies were performed on the viologen-GFP bioconjugate, 16-GFP. Electron
transfer studies were monitored using UV-Vis spectroscopy by following the
characteristic mono-reduced viologen species with a characteristic absorption band at
600 nm.32 Photoexcitation was performed at room temperature on the bioconjugate
16-GFP ( 13 M) in a phosphate buffer, pH 7.0 (20 mM) in the presence of EDTA
(present to stabilise the mono-reduced viologen species).33 Due to the instability of the
mono-reduced viologen species in the presence of oxygen, samples were degassed
(120 mbar) in a specialised small volume cuvette at 0 oC for 30 min and overlayed with
nitrogen. Photoexcitation was performed using a high powered xenon lamp (450 W)
fitted with a 2 mm iris to restrict the amount of excitation light and a UV filter equipped
for 30 min at a distance of 10 cm. The irradiated sample was monitored using UV-Vis
spectroscopy and followed the absorbance from 250 to 800 nm as shown in Figure 4.8a.
The characteristic reduced viologen species was not observed indicating no electron
transfer processes taking place and is consistent with time-resolved fluorescence studies
discussed in the next section (Chapter 4.3.2). In order to eliminate the possibility that
the formation of the mono-reduced species is hindered as a result of bioconjugation, the
degassed bioconjugate 16-GFP was chemically reduced using a strong reductant,
sodium dithionite and the characteristic 600 nm absorption band could be observed as
shown in Figure 4.8b. Similarly, in order to discern whether the lack of electron transfer
was a result of donor-acceptor distance ( 26 Å estimated from chromophore imidazole
ring to central bipyridinium ring via CYS119)10 or viologen is an unsuitable electron
acceptor, a non-covalent mixture of GFP (20 M) and viologen 16 (1 mM) was

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-17
photoexcited using a custom built LED source (3 × 490 nm, 30 W) for 3 h and no shifts
in absorbance or fluorescence emission bands were observed indicating no
photoconversion.
Figure 4.8. Photoexcitation studies of degassed viologen-GFP bioconjugate (16-GFP). (a) Irradiation for 0 min (black) and 30 min (red) with Xenon arc lamp (450 W) showing no change in absorbance spectra corresponding to photoreduction. (b) Chemical reduction of bioconjugate with sodium dithionite (black) showing mono-reduced species with characteristic absorption band at 600 nm; re-oxidised (red) with vigorous shaking showing reversibility of viologen oxidation states.
4.3.1.2 p-benzoquinone acceptor studies
To determine whether the lack of light-induced electron transfer was a result of the GFP
used in this study or viologen is a poor electron acceptor for GFP, p-benzoquinone (BQ)
was employed as an alternative acceptor molecule.16,34 As a result of the lack of
photoinduced electron transfer for viologen-GFP (16-GFP), non-covalent studies using
quinones were performed to determine if a donor-acceptor bioconjugate system is
viable. A non-covalent mixture of GFP (20 M) with fifty-fold excess of
p-benzoquinone (1 mM) in a phosphate buffer, pH 7.0 (20 mM) was irradiated using the
LED source for 3 h at a distance of 2 cm and monitored by UV-Vis and fluorescence
spectroscopy. After irradiation, the appearance of a new absorbance band was observed
at ca. 540 nm as shown in Figure 4.9a. No shift in the fluorescence emission spectra
(ca. 1.6 M) was observed as shown in Figure 4.9b, however, electron/energy transfer

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-18
was confirmed by fluorescence lifetime measurements which is discussed in the next
section.
Figure 4.9. Light-induced photoconversion of a non-covalent mixture containing GFP (20 M) and p-benzoquinone (BQ, 1 mM) in a 20 mM phosphate buffer, pH 7.0. (a) UV-Vis spectra (20 M) showing formation of new absorption band at 540 nm after 3 hirradiation (red) due to electron transfer compared to GFP (black). (b) Fluorescence emission spectra ( 1.6 M, ex = 476 nm) showing no red shift after 3 h irradiation (red) compared to GFP (black).
4.3.1.3 Anthraquinone acceptor studies
Steady state measurements of a non-covalent mixture of p-benzoquinone and GFP
demonstrated light-induced electron transfer showing quinone family acceptors are
viable for covalent donor-acceptor systems. However, due to their inherent long-term
instability23 and lack of functionalisation possibilities, anthraquinone was also screened.
A non-covalent mixture of GFP (20 M) and fifty-fold excess of
anthraquinone-2-carboxylic acid (AQ) in phosphate buffer (20 mM), pH 7.0 was
irradiated using the LED source for 3 h at a distance of 2 cm and photoconversion was
monitored by UV-Vis and fluorescence spectroscopy. After irradiation, bleaching of the
anionic chromophore absorbance band (476 nm) occurred as shown in Figure 4.10a.
Based on Figure 4.10b, analysis of the diluted mixture (ca. 1.6 M) by fluorescence
spectroscopy, a 15 nm stoke shift of the fluorescence emission band from 497 to

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-19
512 nm after irradiation was observed, indicating red photoconversion due to
photo-induced electron transfer.
Figure 4.10. Light-induced photoconversion of a non-covalent mixture containing GFP (20 M) and anthraquinone-2-carboxylic acid (AQ, 1 mM) in a 20 mM phosphate buffer, pH 7.0. (a) UV-Vis spectra (20 M) showing bleaching of the anionic chromophore absorption band at 476 nm after 3 h irradiation (red) due to electron transfer compared to GFP (black). (b) Fluorescence emission spectra ( 1.6 M, ex = 476 nm) showing red shifting of the fluorescence emission band from 497 to 512 nm after 3 h irradiation (red) compared to GFP (black).
It should be noted that the photophysical properties of GFP are different after irradiation
between p-benzoquinone and anthraquinone-2-carboxylic acid, either showing the
appearance of a new absorbance band or red shift of the fluorescence emission band,
respectively. It is proposed that this may be due to the different redox potentials
between the quinone species of p-benzoquinone and anthraquinone-2-carboxylic acid
(estimated using 9,10-anthraquinone) with redox potentials of +0.283 and -0.28 V (vs
SHE), respectively, affecting the chromophore differently.35
It was determined that anthraquinone-2-carboxylic acid is a viable electron
acceptor for light-induced electron transfer with GFP based on non-covalent studies.
Therefore, further photoexcitation experiments were performed on the non-specific
functionalised donor-acceptor bioconjugate, 12-GFP. The bioconjugate 12-GFP (18 M)
in 20 mM phosphate buffer, pH 7.0 was irradiated with the LED source for 3 h at a
distance of 2 cm. Irradiation showed conversion of the chromophore from the anionic to

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-20
neutral chromophore and photobleaching, however, no stoke shift of the fluorescence
emission band was observed indicating red photoconversion of 12-GFP did not occur as
shown in Figure 4.11. The lack of red photoconversion may be due to the
donor-acceptor distance being greater than distance required for electron transfer
(assuming maximum tunnelling distance of 20 Å via superexchange).36 The exact
residues functionalised (21 possible lysines available)1 is unknown due to the
non-specific nature of amine modification. In order to confirm whether energy/electron
transfer processes are taking place, time-resolved fluorescence spectroscopy were
performed on both the non-covalent mixture of GFP and anthraquinone-2-carboxylic
acid as well as the bioconjugate 12-GFP.
Figure 4.11. Photoexcitation of 12-GFP (18 M) in a 20 mM phosphate buffer, pH 7.0.(a) UV-Vis spectra (18 M) showing bleaching of the anionic chromophore absorption band at 476 nm and partial shifting to the neutral state after 3 h irradiation (red) compared to GFP (black). (b) Fluorescence emission spectra ( 1.6 M, ex = 476 nm) showing no shift of the fluorescence emission band (497 nm) after 3 h irradiation (red) compared to GFP (black).
4.3.2 Fluorescence lifetime studies
Fluorescence lifetime studies of GFP and electron transfer measurements were
performed in collaboration with A/Prof. Timothy W. Schmidt, Dr. Raphaël G. C. R.
Clady and Mr. Murad Tayebjee at the University of Sydney, Australia. Time-resolved
fluorescence spectroscopy was performed on GFP, a non-covalent mixture of

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-21
p-benzoquinone/anthraquinone-2-carboxylic acid and GFP, bioconjugates 12-GFP and
16-GFP.
Lifetime measurements of all samples were performed at a concentration of
1.6 M in a 5 mM phosphate buffer, pH 7.0 containing 5 mM
ethylenediaminetetraacetic acid at room temperature. Prior to measurements, samples
were irradiated with the polarised laser source at an excitation wavelength of 476 nm for
10 min to allow stabilisation of the fluorescence emission intensity. Additionally, it
should be noted that fluorescence emission was probed at a wavelength of either 515 or
540 nm as detection at the GFP emission maximum (497 nm) resulted in saturation of
the detector. Fluorescence lifetime measurements were performed in a specialised low
volume quartz cuvette with a 2-3 mm pathlength to minimise background fluorescence
due to delayed fluorescence of sample across the entire pathlength of the cell.
Based on Figure 4.12, it was found that GFP had a fluorescence lifetime of
1.65±0.06 ns. This is not dissimilar to wild type jellyfish Aequorea GFP, with a 2.8 ns
fluorescence lifetime due to photochromicity between the protonated and deprotonated
species.37 The fluorescence data was fitted assuming a mono-exponential decay (single
fluorescent species with no quencher) and estimated using the relative amplitudes:9b
/0)( teFtF (4.1)
where F(t) is the fluorescence intensity, F0 is the initial fluorescence intensity, t is time
and is the fluorescence lifetime. Additionally, fluorescence decay of bioconjugate
16-GFP showed a similar decay profile and modelled according to Equation 4.1. The
fluorescence lifetime of 16-GFP was estimated to be 1.66±0.05 ns, indicating no
electron or energy transfer processes occurring.
In contrast, quenching of the fluorescence decay of the non-covalent mixture of
GFP and p-benzoquinone was observed as shown in Figure 4.12, indicative of

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-22
electron/energy transfer processes.38 The fluorescence showed double exponential decay
behaviour with components having lifetimes of 0.76±0.04 and 0.14±0.01 ns, which
appears to correspond to two different types of static quenching which was modelled
according to Equation 4.2:9b,39
21 /0
/0 ')( tt eFeFtF (4.2)
where F(t) is the fluorescence intensity, F0 and F0’ is the initial fluorescence intensity, t
is time and is the fluorescence lifetime. Similarly, the initial fluorescence intensity
coefficients were found to be 0.47±0.03 and 0.95±0.05 for the two quenching modes,
respectively, indicating approximately 67% of the GFP population with the shorter
lifetime component is likely participating in electron/energy transfer processes.
Assuming that electron transfer takes place, from the lifetime value of the shorter
quenched lifetime, the rate constant of the forward electron transfer (ket) can be
calculated according to Equation 4.3:40
0
11etk (4.3)
where and 0 are the respective lifetimes in the presence and absence of the electron
acceptor. The calculated value was ket = 6.5±0.5 × 109 s-1. The corresponding ket for the
longer quenching lifetime would be 7.1±0.5 × 108 s-1.

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-23
Figure 4.12. Time-resolved fluorescence (ultrafast time-resolved photoluminescence) of 1.6 M green fluorescent protein ( = 1.65±0.06 ns, black), 16-GFP (red,= 1.66±0.05 ns) and a non-covalent mixture of green fluorescent protein and 1 mM
p-benzoquinone (blue, = 0.76±0.04 and 0.14±0.01 ns) in 5 mM phosphate buffer, 5 mMethylenediamine tetraacetic acid, pH 7.0. Excitation with 476 nm laser source and detected at 540 nm at room temperature.
Initial lifetime studies were based on ultrafast time-resolved photoluminescence
(UFTRPL). Subsequent lifetime measurements for both a non-covalent mixture of
anthraquinone-2-carboxylic acid and GFP as well as 12-GFP were performed using
time-correlated single photon counting (TCSPC) which sacrifices temporal resolution in
favour of a greater range and sensitivity compared to UFTRPL. Quenching of
fluorescence decay of anthraquinone samples were observed as shown in Figure 4.13.

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-24
Figure 4.13. Time-resolved fluorescence (time-correlated single photon counting) of a non-covalent mixture of 1.6 M green fluorescent protein and 1 mManthraquinone-2-carboxylic acid ( = 2.04±0.02 and 0.23±0.04 ns, black) and bioconjugate 12-GFP (red, = 1.87±0.02 and 0.28±0.04 ns) in 5 mM phosphate buffer, 5 mM ethylenediamine tetraacetic acid, pH 7.0. Excitation with 476 nm laser source and detected at 515 nm at room temperature.
The non-covalent mixture of GFP and anthraquinone-2-carboxylic acid was modelled as
a double exponential decay using Equation 4.2. Hence, the fluorescence lifetimes of the
non-covalent mixture was estimated to be =2.04±0.02 and 0.23±0.04 ns,
corresponding to native GFP fluorescence and quenching, possibly due to electron
transfer, respectively. The electron transfer rate was determined to be
ket = 3.7±0.7 × 109 s-1 using Equation 4.3. The larger value of the GFP fluorescence
lifetime component of =2.04±0.02 ns compared to the GFP only control
(1.65±0.06 ns) is attributed to the limitation of TCSPC, where the temporal resolution is

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-25
200 ps. Based on the initial fluorescence intensity coefficients, approximately 76%
of the GFP population participated in electron/energy transfer processes.
Similarly, quenching of the bioconjugate 12-GFP fluorescence decay was
observed in Figure 4.13 and modelled according to Equation 4.2 which gave
fluorescence lifetimes of =1.87±0.02 and 0.28±0.04 ns (ket = 2.9±0.4 × 109 s-1),
corresponding to GFP fluorescence and electron transfer, respectively. Based on these
time-resolved fluorescence spectroscopy measurements, electron transfer of the
bioconjugate was observed in contrast to steady-state fluorescence measurements which
did not exhibit red photoconversion. This may be attributed to the high power of the
laser source inducing intramolecular electron transfer of the bioconjugate 12-GFP.
Based on the initial fluorescence intensity coefficients, approximately 62% of the
12-GFP population participated in electron/energy transfer processes. From lifetime
studies, the possible electron transfer pathway is summarised in Scheme 4.6
Scheme 4.6. Schematic representation of the photo-induced electron transfer of green fluorescent protein (GFP, amFP497) at room temperature. (a) Non-covalent mixture of GFP and p-benzoquinone (BQ) or anthraquinone-2-carboxylic acid (AQ) with GFP fluorescent lifetime of = 1.65 ns and electron transfer rates of 6.5 109 and 3.7 109 s-1, respectively. (b) Bioconjugate 12-GFP with electron transfer rate of 2.9 109 s-1.
4.4 Conclusion
In this Chapter, the synthesis of covalent donor-acceptor systems based on green
fluorescent protein (GFP) as a light-induced electron donor were prepared for electron

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-26
transfer studies. For this purpose, viologen-GFP (16-GFP) and anthraquinone-GFP
(12-GFP) bioconjugates were prepared via cysteine-maleimide coupling and amine
modification, respectively. Based on steady-state UV-Vis and fluorescence
spectroscopy studies, evidence of electron transfer was observed by red
photoconversion of the chromophore with p-benzoquinone and anthraquinone acceptor
molecules. Time-resolved fluorescence spectroscopy measurements showed a GFP
fluorescence lifetime of 1.65±0.06 ns. Furthermore, quenching of GFP lifetimes to
0.14±0.01 and 0.23±0.04 ns was observed with non-covalent mixtures of GFP and
p-benzoquinone and anthraquinone-2-carboxylic acid, respectively, corresponding to a
rate constant of forward electron transfer of ket = 6.5±0.5 × 109 and 3.7±0.7 × 109 s-1.
The investigation of photoinduced electron transfer rates between GFP and electron
acceptors have never been reported. However, the donor-acceptor viologen
bioconjugate 16-GFP showed no evidence of photoinduced electron transfer as viologen
acts as a poor electron acceptor in 16-GFP. On the other hand, fluorescence lifetime
studies of the anthraquinone bioconjugate 12-GFP showed possible electron transfer
with a quenched GFP fluorescence lifetime of 0.28±0.04 ns and an electron transfer rate
of ket = 2.9±0.4 × 109 s-1.
4.5 References
(1) Angelo, C.; Denzel, A.; Vogt, A.; Matz, M. V.; Oswald, F.; Salih, A.; Nienhaus, G. U.; Wiedenmann, J. Mar. Ecol. Prog. Ser. 2008, 364, 97.
(2) (a) Tsien, R. Y. Annu. Rev. Biochem. 1998, 67, 509. (b) Lippincott-Schwartz, J.; Patterson, G. H. Science 2003, 300, 87. (c) Alieva, N. O.; Konzen, K. A.; Field, S. F.; Meleshkevitch, E. A.; Hunt, M. E.; Beltran-Ramirez, V.; Miller, D. J.; Wiedenmann, J.; Salih, A.; Matz, M. V. PLoS One 2008, 3, e2680.
(3) Goldstein, J. C.; Munoz-Pinedo, C.; Ricci, J. E.; Adams, S. R.; Kelekar, A.; Schuler, M.; Tsien, R. Y.; Green, D. R. Cell Death Differ. 2005, 12, 453.
(4) Chalfie, M.; Tu, Y.; Euskirchen, G.; Ward, W.; Prasher, D. Science 1994, 263,802.
(5) (a) Wilson, C. G. M.; Magliery, T. J.; Regan, L. Nat. Meth. 2004, 1, 255. (b) Wiedenmann, J.; Oswald, F.; Nienhaus, G. U. IUBMB Life 2009, 61, 1029.

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-27
(6) Shimomura, O.; Johnson, F. H.; Saiga, Y. J. Cell. Comp. Physiol. 1962, 59, 223.(7) Heim, R.; Cubitt, A. B.; Tsien, R. Y. Nature 1995, 373, 663.(8) Yang, F.; Moss, L. G.; Phillips, G. N. Nat. Biotech. 1996, 14, 1246.(9) (a) van Thor, J. J.; Sage, J. T. Photochem. Photobiol. Sci. 2006, 5, 597. (b)
Cotlet, M.; Hofkens, J.; Maus, M.; Gensch, T.; Van der Auweraer, M.; Michiels, J.; Dirix, G.; Van Guyse, M.; Vanderleyden, J.; Visser, A. J. W. G.; De Schryver, F. C. J. Phys. Chem. B 2001, 105, 4999.
(10) Henderson, J. N.; Remington, S. J. Proc. Natl. Acad. Sci. U. S. A. 2005, 102,12712.
(11) Ormö, M.; Cubitt, A. B.; Kallio, K.; Gross, L. A.; Tsien, R. Y.; Remington, S. J. Science 1996, 273, 1392.
(12) Shu, X.; Shaner, N. C.; Yarbrough, C. A.; Tsien, R. Y.; Remington, S. J. Biochemistry 2006, 45, 9639.
(13) Ward, W. W. In Green Fluorescent Protein: Properties, Applications, and Protocols; 2nd ed.; Chalfie, M., Kain, S. R., Eds.; John Wiley & Sons, Inc., 2006; Vol. 47.
(14) Chattoraj, M.; King, B. A.; Bublitz, G. U.; Boxer, S. G. Proc. Natl. Acad. Soc. U. S. A. 1996, 93, 8362.
(15) Bogdanov, A. M.; Mishin, A. S.; Yampolsky, I. V.; Belousov, V. V.; Chudakov, D. M.; Subach, F. V.; Verkhusha, V. V.; Lukyanov, S.; Lukyanov, K. A. Nat. Chem. Biol. 2009, 5, 459.
(16) Sauvage, J. P.; Collin, J. P.; Chambron, J. C.; Guillerez, S.; Coudret, C.; Balzani, V.; Barigelletti, F.; De Cola, L.; Flamigni, L. Chem. Rev. 1994, 94, 993.
(17) Hodgson, D. R. W.; Sanderson, J. M. Chem. Soc. Rev. 2004, 33, 422.(18) (a) Thordarson, P.; Droumaguet, B. L.; Velonia, K. Appl. Microbiol. Biotechnol.
2006, 73, 243. (b) Veronese, F. M.; Pasut, G. Drug Discov. Today 2005, 10,1451.
(19) Rostovtsev, V. V.; Green, L. G.; Fokin, V. V.; Sharpless, K. B. Angew. Chem. Int. Ed. 2002, 41, 2596.
(20) (a) van Kasteren, S. I.; Kramer, H. B.; Jensen, H. H.; Campbell, S. J.; Kirkpatrick, J.; Oldham, N. J.; Anthony, D. C.; Davis, B. G. Nature 2007, 446,1105. (b) Wang, Q.; Chan, T. R.; Hilgraf, R.; Fokin, V. V.; Sharpless, K. B.; Finn, M. G. J. Am. Chem. Soc. 2003, 125, 3192.
(21) (a) Hvasanov, D.; Goldstein, D. C.; Thordarson, P. In Molecular Solar Fuels;The Royal Society of Chemistry, 2012; p 426. (b) Bertozzi, C. R. Acc. Chem. Res. 2011, 44, 651.
(22) Dirks, A. J.; van Berkel, S. S.; Hatzakis, N. S.; Opsteen, J. A.; van Delft, F. L.; Cornelissen, J. J. L. M.; Rowan, A. E.; van Hest, J. C. M.; Rutjes, F. P. J. T.; Nolte, R. J. M. Chem. Commun. 2005, 4172.
(23) Holmes, T. J.; John, V.; Vennerstrom, J.; Choi, K. E. J. Org. Chem. 1984, 49,4736.
(24) Goldstein, D. C. PhD Thesis, The University of New South Wales, 2011.(25) Sarkar, B.; Wigfield, Y. J. Biol. Chem. 1967, 242, 5572.(26) Himo, F.; Lovell, T.; Hilgraf, R.; Rostovtsev, V. V.; Noodleman, L.; Sharpless,
K. B.; Fokin, V. V. J. Am. Chem. Soc. 2004, 127, 210.(27) Opsteen, J. A.; Brinkhuis, R. P.; Teeuwen, R. L. M.; Lowik, D.; van Hest, J. C.
M. Chem. Commun. 2007, 3136.(28) Gray, H. B.; Winkler, J. R. Annu. Rev. Biochem. 1996, 65, 537.(29) Sawin, K. E.; Nurse, P. Curr. Biol. 1997, 7, R606.

Chapter 4 Green Fluorescent Protein as a Light-Induced Electron Donor
4-28
(30) Takahashi, E.; Takano, T.; Nomura, Y.; Okano, S.; Nakajima, O.; Sato, M. Am. J. Physiol. Cell Physiol. 2006, 291, C781.
(31) Bailey, S. I.; Ritchie, I. M. Electrochim. Acta 1985, 30, 3.(32) Prasad, D. R.; Mandal, K.; Hoffman, M. Z. Coord. Chem. Rev. 1985, 64, 175.(33) (a) Abdul-Ghani, A. J.; Abdul-Kareem, S. J. Photochem. Photobiol., A 1990, 51,
391. (b) Mandal, K.; Hoffman, M. Z. J. Phys. Chem. 1984, 88, 185.(34) Hay, S.; Wallace, B. B.; Smith, T. A.; Ghiggino, K. P.; Wydrzynski, T. Proc.
Natl. Acad. Sci. U. S. A. 2004, 101, 17675.(35) Clark, W. M. Oxidation-reduction potentials of organic systems; Robert E.
Krieger Publishing Company, 1972.(36) (a) Gray, H. B.; Winkler, J. R. Q. Rev. Biophys. 2003, 36, 341. (b) Cordes, M.;
Giese, B. Chem. Soc. Rev. 2009, 38, 892.(37) Striker, G.; Subramaniam, V.; Seidel, C. A. M.; Volkmer, A. J. Phys. Chem. B.
1999, 103, 8612.(38) Peterson, J. R.; Smith, T. A.; Thordarson, P. Chem. Commun. 2007, 1899.(39) (a) Andersson, A.; Danielsson, J.; Gräslund, A.; Mäler, L. Eur. Biophys. J. 2007,
36, 621. (b) Valeur, B. Molecular Fluorescence: Principles and Applications;WILEY-VCH Verlag: Wienheim, Germany, 2002.
(40) Nelissen, H. F. M.; Kercher, M.; De Cola, L.; Feiters, M. C.; Nolte, R. J. M. Chem. Eur. J. 2002, 8, 5407.

Chapter 5
Supramolecular Aggregates for Protein Encapsulation

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-2
5 Supramolecular Aggregates for Protein Encapsulation
In this Chapter, the formation of vesicular compartments based on phospholipids and a
diblock copolymer for enzyme encapsulation and light-induced proton pumping studies
is described. Liposomes were prepared using either egg L- -phosphatidylcholine or
synthetic L-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine based on variations of the
film hydration method allowing internal encapsulation of fluorescently labelled iso-1
cytochrome c.
Induced formation of polymersomes based on the polyelectrolyte diblock
copolymer, polystyrene140-b-poly(acrylic acid)48 (PS140-b-PAA48) in the presence of
positively charged proteins/peptides allowed concomitant encapsulation of highly
charged hydrophilic proteins with high encapsulation efficiencies.
Vesicles were purified by size exclusion chromatography or membrane dialysis
to remove non-encapsulated material and were characterised by dynamic light
scattering, transmission electron microscopy and confocal laser-scanning microscopy. A
brief review of the vesicle formation methods of liposomes and polymersomes is
outlined in this chapter.
5.1 Liposomes
The term liposome (also termed lipid vesicles) was first introduced by Bangham et al.1
in 1965 which was used to describe aqueous dispersions of multilamellar vesicle (MLV)
systems produced by physical agitation of an aqueous medium in the presence of a dry
lipid film. In modern literature, it is a generic term used to describe polymolecular
aggregates formed in aqueous solution on the dispersion of certain bilayer forming
amphiphilic lipids which may be large or small and may be unilamellar or multilamellar
in nature.2

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-3
Liposomes are spherical shells consisting of one or more (concentric) lamellae
composed of the amphiphiles under osmotically balanced conditions as shown in Figure
5.1.2b The lamellae are curved and self-closed molecular bilayers in which the
hydrophobic portion of the amphiphiles forms the hydrophobic interior of the bilayer
and the polar head group is in contact with the aqueous phase.3 The interior of the lipid
vesicle consists of an aqueous core with an approximate composition of the bulk phase.
The properties of vesicles including mean diameter, lamellarity and physical stability
are generally dependent on the method of vesicle preparation rather than the structure of
natural phospholipid amphiphile used.4
Figure 5.1. Schematic representation of a liposome (lipid vesicle).2b
Liposomes can be prepared based on four primary routes including: film hydration5,
emulsion6, micelle forming detergents7 and organic solvent injection.8 The most
common approach for forming liposomes for enzyme encapsulation is the film
hydration method due to the use of biologically friendly conditions requiring only an
enzyme containing buffered aqueous solution.9 Additionally, variations of the film
hydration method allows customisation of vesicle properties including lamellelarity and
mean size via dispersion (large multilamellar vesicles (LMV), diameters typically

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-4
1 m)10, sonication (small unilamellar vesicles (SUV), diameters typically <50 nm)11,
extrusion (large unilamellar vesicles (LUV), diameters typical of pore size used)12 or
electroformation (LMV, diameters typically 30 m).13 In general, vesicles for enzyme
encapsulation are typically prepared by hydration of a dry lipid film using a buffered
aqueous solution containing the (bio)molecule to be encapsulated resulting in LMV.
The mixture is then subjected to freeze-thaw or lyophilisation steps to increase
encapsulation efficiency and finally sized to form a homogenous mixture by either
sonication or extrusion. Subsequently, the liposome mixture is passed through a size
exclusion column or dialysed to remove non-encapsulated material.4b,c It should be
noted that mechanical treatments during preparation have to be carried out at 5-10 oC or
more above the main lamellar chain-melting phase transition temperature (Tm) where
the saturated hydrophobic chains change from a crystalline state (trans conformation) to
a fluid state (gauche conformation).14
Commonly, liposomes in literature are employed as drug delivery agents,15 gene
vectors16, nanoreactors17 and artificial cells.18 In this project, the ultimate goal is the
development of a photosynthetic-respiratory artificial organelle which requires
compartmentisation via an enclosed membrane which is discussed further in Chapter 7.
Liposomes are ideal for this purpose as LUV ( 100 nm) closely resemble biological
membranes based on surface curvature and membrane fluidity.2b,7
5.1.1 Liposome formation and characterisation
Liposomes based on the phospholipid egg L- -phosphatidylcholine (egg PC) derived
from egg yolk were selected for enzyme encapsulation purposes. Egg PC as shown in
Figure 5.2 was chosen as it is one of the most highly studied amphiphiles in lipid vesicle

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-5
literature2b, commercially available, present in eukaryotic cells19 and possesses a low Tm
(-5.8±6.5 oC)20 leading to a fluid membrane for potential enzyme encapsulation studies.
Figure 5.2. Chemical structure of egg L- -phosphatidylcholine (egg PC) or synthetically derived 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC).
In this project, large unilamellar vesicles (LUV) of ca. 100 nm in diameter are required
as potential compartments for enzyme encapsulation of light-harvesting bioconjugates
discussed in Chapter 2 and the reconstitution of transmembrane cytochrome c oxidase
for charge transfer and proton translocation studies. In order to achieve this, liposomes
from egg PC were prepared by formation of a thin film of dry lipid from a stock
solution in chloroform as shown in Scheme 5.1. The organic solvent was removed under
a stream on nitrogen and residual chloroform was removed under high vacuum for at
least 4 h as presence of trace organic solvent can affect vesicle morphology.3 The lipids
were hydrated in a phosphate buffer (pH 7.0) over 1 h with regular agitation.
Subsequently, the liposome preparation was extruded by passing the sample through a
100 nm polycarbonate membrane to size the vesicles into a homogenous mixture as it
has been reported that extrusion of vesicles through <200 nm pore membranes produce
unilamellar vesicles.12

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-6
Scheme 5.1. Schematic summary of the preparation of liposomes via the film hydration method.
The extruded liposomes were characterised by dynamic light scattering (DLS) to
determine the average hydrodynamic diameter (Dh) of the particles as it is an optimal
technique to determine particle size in dispersions and colloids.21 Particles (liposomes)
cause scattering of the light in all directions when the light passes through the
dispersion. With small particles (compared to the wavelength of the light), the intensity
of scattered light is uniform in all directions due to Rayleigh scattering. In contrast, with
larger particles (above 250 nm diameter), the scattering intensity is angle dependent
(Mie scattering). DLS allows detection of time-dependent fluctuations in the scattered
intensity due to Brownian motion of the particles causing constructive and destructive
interference of light scattered by neighbouring particles using a coherent
monochromatic (laser) source. The diffusion coefficient can be determined based on
analysis of the time dependence of the intensity fluctuation. As a result, the
hydrodynamic diameter of the particles can be elucidated via the Stokes-Einstein
equation, knowing the viscosity of the medium.22
DLS experiments carried out on 1 mg/mL egg PC extruded (100 nm) liposomes
showed a monodisperse sample with an average hydrodynamic radius of 105±12 nm as
shown in Figure 5.3.

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-7
Figure 5.3. Dynamic light scattering (DLS) experiments on 100 nm extruded egg phosphatidylcholine (egg PC) liposomes (1 mg/mL) in 20 mM sodium dihydrogen phosphate, pH 7.0. The average diameter of the aggregates is 105±12 nm.
Freeze-thaw and lyophilisation treatment prior to extrusion has been shown to increase
encapsulation efficiencies.2b In order to confirm that the treatment does not adversely
affect mean size, DLS experiments were also performed on the extruded freeze-thaw
and lyophilised samples as shown in Table 5.1. It was found that lyophilisation prior to
extrusion did not significantly affect mean diameter of the liposomes, however,
freeze-thaw cycles led to small average diameters (75±7 nm). This may be a result of
fracture or rupture of the vesicles leading to a change in size distribution.

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-8
Table 5.1. Mean size of different treatment methods of egg phosphatidylcholine liposomes.Treatment Mean hydrodynamic diameter (Dh) /nma
Extrusion 105±12Freeze-thaw/Extrusion 75±7Lyophilisation/Extrusion 114±8
a Experiments carried out on 1 mg/mL liposomes in 20 mM sodium dihydrogen phosphate, pH 7.0 at 25 oC. Errors are standard deviation.
Additionally, liposomes were characterised by transmission electron microscopy (TEM)
to confirm structural morphology and lamellarity. Due to the poor electron scattering
properties of phospholipids for TEM imaging, samples were negatively stained using
2% phosphotungstic acid. Negative stains are composed of salt solutions of strongly
electron scattering heavy metal compounds which are added to the drying specimen
which forms a mold of the specimen.23 TEM studies showed vesicular morphologies for
extruded liposomes as shown in Figure 5.4a. However, multilamellar vesicles were
observed even though it has been reported in literature that extrusion produces
unilamellar vesicles.12 To ascertain whether further mechanical treatment of the
liposomes produces unilamellar vesicles, TEM micrographs of freeze-thawed and
lyophilised specimens prior to extrusion were imaged; Figure 5.4b and c, respectively,
which showed multilamellar structural features.

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-9
Figure 5.4. TEM micrographs of egg phosphatidyl (egg PC) liposomes (0.5 mg/mL). (a) 100 nm pore extrusion. (b) Freeze-thawed and extruded. (c) Lyophilised and extruded. Specimens negatively stained with 2% phosphotungstic acid. Scale bars: 50 nm. Arrows indicate multilamellar structures.
It has been reported that naturally isolated egg PC contains negatively charged
impurities.24 Instead, synthetically isolated L-palmitoyl-2-oleoyl-sn-glycero-3-
phosphocholine (POPC) which is chemically identical to the natural product was used
for liposome formation to eliminate the possibility that impurities were causing
multilamellar features. To determine whether the multilamellar features are a result of
the use of 2% phosphotungstic acid, an alternative negative stain, 2% uranyl acetate was
used for POPC liposomes as shown in Figure 5.5a and b, respectively. Based on TEM
micrographs of Figure 5.5, identical multilamellar features were observed compared to
2% phosphotungstic acid stains of egg PC liposomes (Figure 5.4).

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-10
Figure 5.5. TEM micrographs of negatively stained L-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC) liposomes (0.5 mg/mL). (a) 2% phosphotungstic acid. (b) 2% uranyl acetate. Scale bars: 100 nm. Arrows indicate multilamellar structures.
To eliminate the use of negative stains and drying of sample during TEM preparation,
cryogenic TEM (cryo-TEM) was employed in collaboration with A/Prof. Filip Braet at
the Australian Centre for Microscopy & Microanalysis (The University of Sydney).
Cryo-TEM allows the imaging of suspended organic material in aqueous solution.25
Thin films (ca. 100 nm) of suspended sample on a TEM grid is plunged into liquid
ethane at -183 oC using an automated robot (vitrobot) which allows the nanostructures
to become instantaneously embedded in an electron transparent film of vitrified
amorphous ice.26 This allows samples to be analysed in their near native hydrated
state.27 As shown in Figure 5.6, it was found that 70% of the extruded (100 nm) POPC
liposomes were unilamellar and the morphology were generally typically ‘filled-cups’.
This demonstrates that staining of liposomes and the drying process was responsible for
multilamellar features resulting in electron micrograph artifacts.

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-11
Figure 5.6. Cryo-TEM micrograph of L-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC) liposomes. Typically ‘filled-cup’ morphologies with 70% unilamellarity. Scale bars: 50 nm. Arrows indicate unilamellar vesicle.
5.1.2 Enzyme encapsulation
In order to evaluate liposomes as potential compartments for light-induced electron
transfer studies of light-activated bioconjugates (discussed further in Chapter 6),
enzyme encapsulation studies of iso-1 cytochrome c as a model enzyme were
performed. Initially, to determine if cytochrome c could be successfully encapsulated in
the interior volume of POPC liposomes, fluorescently labelled iso-1 cytochrome c
(cyt c-Oregon Green 488) was encapsulated and monitored optically via confocal
laser-scanning microscopy (CLSM).
Encapsulation was achieved by adding cyt c-Oregon Green 488 (5 M) in 20 mM
sodium dihydrogen phosphate buffer, pH 7.0, to a dry lipid film resulting in a lipid
concentration of 15 mg/mL. Subsequently, the dispersion was lyophilised to increase
encapsulation efficiency (method with highest encapsulation efficiencies)2b, rehydrated
and extruded. It should be noted that extrusion was performed using 400 nm
polycarbonate membranes rather than 100 nm pores to allow visualisation of aggregates
as 100 nm particles are below the Abbe diffraction limit ( 250 nm, assuming numerical

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-12
aperture NA of 1.4).28 Non-encapsulated enzymes were removed by size exclusion
chromatography (Superdex 75) and imaged by CLSM as shown in Figure 5.7,
demonstrating successful encapsulation.
Figure 5.7. Transmission light (a) and confocal laser-scanning micrograph (b)-excitation 488 nm-of enzyme encapsulated fluorescently labelled cytochrome c(cyt c-Oregon Green 488) in L-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC) liposomes. Scale bars: 50 m.
Interestingly, it was found that purification of liposomes by size exclusion
chromatography (approximately three purification cycles) resulted in gradual blockage
of the column and required cleaning using an aqueous 70% v/v ethanol solution as
shown in Figure 5.8.

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-13
Figure 5.8. Size exclusion chromatogram showing separation of fluorescently labelled cytochrome c-containing vesicles (peak A) from non-entrapped enzyme (peak B) using Superdex 75 (GE Healthcare) in 20 mM sodium dihydrogen phosphate buffer, pH 7.0 at 0.5 mL/min. Red trace indicates blockage of column preventing elution of liposomes. Black trace indicates elution of liposomes after 70% ethanol (v/v) wash. Elution was monitored at optical density 280 nm.
Evaluation of POPC as a potential compartment for enzyme encapsulation was screened
by quantifying the encapsulation efficiency of unmodified iso-1 cytochrome c (cyt c).
The encapsulation efficiency (EE(%)) was quantified based on Equation 5.1 and is
defined as the percentage amount of enzyme entrapped in the vesicles in relation to the
total amount of enzyme present during the vesicle formation and entrapment
procedure.2b,3 The EE(%) is dependent on a number of factors including method of
preparation, concentration of amphiphile used, chemical nature of amphiphile and
concentration and nature of (bio)molecule to be encapsulated.
100(%)usedenzymeofamounttotal
vesicleslipidtheinentrappedenzymeofamountEE (5.1)

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-14
Encapsulation was achieved by adding cytochrome c (100 M) in 20 mM sodium
dihydrogen phosphate buffer, pH 7.0, to a dry lipid film resulting in a lipid
concentration of 15 mg/mL. The dispersion was lyophilised, rehydrated and extruded
through a 100 nm membrane. Subsequently, the enzyme containing liposomes were
purified using size exclusion chromatography (Superdex 75) in 20 mM sodium
dihydrogen phosphate buffer, pH 7.0, as shown in Figure 5.9. To ensure maximum
recovery of encapsulated enzyme, the column was first pre-cleaned with 70% v/v
ethanol solution before loading liposomes onto column.
Figure 5.9. Size exclusion chromatogram showing separation of iso-1cytochrome c-containing vesicles (peak A) from non-entrapped enzyme (peak B) using Superdex 75 (GE Healthcare) in 20 mM sodium dihydrogen phosphate buffer, pH 7.0 at 0.5 mL/min.

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-15
The EE(%) was determined by collecting the purified liposomal fraction (peak A) and
estimating the concentration of encapsulated cyt c by UV-Vis spectroscopy using the
heme absorption band ( 410 = 97.6 mM-1cm-1).29 Due to scattering of the UV-Vis
baseline caused by liposome aggregates, the vesicles were disrupted by addition of
surfactant Triton X-100 (2%, v/v) in a ratio of 1:1 (v/v)30 and a low EE(%) of 1.5±0.1%
was determined. The maximum theoretical EE(%) under identical conditions and
assuming a random statistical distribution with an average POPC lipid head group area
of 69 31 was 7.0% (see Appendix D). A random statistical distribution was assumed as
there are no charge interactions between lipid and cytochrome c due to the zwitterionic
nature of POPC. Interestingly, from DLS studies, it was found that the average diameter
of the enzyme encapsulated vesicles was greater than enzyme free POPC liposomes
with diameters of 96±1 nm and 100±3 nm, respectively, as shown in Figure 5.10. This
is most likely due to accommodation of enzyme within the interior volume of the
compartment.

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-16
Figure 5.10. Dynamic light scattering (DLS) experiments on lyophilised-100 nm extruded L-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC) liposomes (1 mg/mL) in 20 mM sodium dihydrogen phosphate, pH 7.0. The average diameter of the aggregates is 96±1 nm and 100±3 nm for enzyme free (black) and enzyme encapsulated (red) vesicles, respectively.
5.2 Polymersomes
One of the major disadvantages of liposomes is their poor thermodynamic and
mechanical properties resulting in morphology changes, degradation and leakage of
internal contents mainly due lipid peroxidation or hydrolysis when stored in aqueous
solution.32 In contrast, polymersomes overcome this inherent instability due to their
higher molecular weight, lower critical aggregation concentration, slower chain
mobility and polymer entanglement.33 Polymersomes are generally based on diblock or
triblock copolymer amphiphiles.34 Polymersomes have been used for the encapsulation
of drugs35, biomolecules as nanoreactors17, protein therapeutic applications for
biomedicine36 or artificial organelles.37

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-17
Polymersomes are part of a continuum of supramolecular structures consisting
of micelles, rods, lamellae and vesicles.38 Multiple factors can be tailored to influence
resulting structures including temperature, solvent, block length, ratio of the blocks, pH
and ionic strength which affects the critical packing parameter (p = v/aolc), where v is
the volume of the hydrocarbon chains, ao is the optimal area of the hydrophilic (corona)
group and lc is the critical chain length of the hydrophobic (core) group as shown in
Figure 5.11.39 In this project, polymersome aggregate studies used the well-known
diblock copolymer polystyrene-b-poly(acrylic acid) (PS-b-PAA).40 It has been reported
that the major factor influencing PS-b-PAA morphologies was the length of the
hydrophilic block.41 Diblock copolymers such as PS-b-PAA self-assemble when they
are mixed with a solvent selective for one of the blocks, resulting in aggregation and
phase separation.42
Figure 5.11. Different morphologies predicted by the packing parameter (p). It is predicted that micelles form when p<1/2, vesicles when 1/2<p<1 and inverted structures are expected when p>1.17
Polymersomes are traditionally prepared using the ‘thermodynamic trapping’ method
with the block copolymer dissolved in an organic common solvent and water then
slowly added to induce aggregate morphologies, which is a non-biologically friendly
technique.43 Alternatively, they can be prepared using a biologically friendly ‘kinetic

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-18
trapping’ method44, which involves injecting the dissolved block copolymer in organic
solution directly into the aqueous buffer. In this Chapter, both methodologies are
studied using commercially available PS140-b-PAA48 with a polydispersity index (PDI)
of 1.10 to determine which preparation method is ideal and robust for enzyme
encapsulation.
5.2.1 Aggregate formation using the ‘thermodynamic trapping’ method
Polymersome formation using PS140-b-PAA48, as shown in Figure 5.12, was attempted
using the ‘thermodynamic trapping’ method, also known as the slow addition method.
The role of solvent is crucial to the free energy of aggregation and polymer-solvent
interactions determine coil dimensions for each block of the diblock copolymer, which
affects the resulting aggregate dimensions and morphologies.39a Tetrahydrofuran (THF)
and dioxane were used as solvents for aggregation studies as they are a common solvent
for both blocks of the copolymer. Additionally, the solubility parameter of the solvents
is similar to the polymer.45
Figure 5.12. Chemical structure of polystyrene140-b-poly(acrylic acid)48(PS140-b-PAA48) with a polydispersity index (PDI) of 1.10.
Polymeric aggregates were prepared by adding water dropwise over a 3 h period
into a dissolved polymer solution (10 mg/mL) in THF or dioxane until a turbid solution
formed, indicating formation of polymeric aggregates. Finally, the turbid solution was
dialysed against water extensively to remove the organic solvent. The aggregates were
characterised by TEM microscopy without the use of negative stains due to the electron
scattering properties of the PS block.46 It should be noted that polymer aggregate TEM

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-19
studies required a lower accelerating voltage (80 kV) compared to liposomes as higher
energy electron beams caused deformation and damage to the specimen. Only micelle
and large compound micelle structures were observed as shown in Figure 5.13. Due to
the unsuccessful formation of polymersome structures via the ‘thermodynamic trapping’
method and the biologically unfriendly conditions required, further aggregate studies
using this method was not pursued. As a result, the ‘kinetic trapping’ method was
investigated for enzyme encapsulation which is discussed in the next section (Chapter
5.2.2) and THF was exclusively used in further polymersome aggregate studies due to
the possible carcinogenic effects of dioxane.47
Figure 5.13. TEM micrographs of polysytrene140-b-poly(acrylic acid)48(PS140-b-PAA48) aggregates. (a) Tetrahydrofuran (THF). (b) Dioxane. Scale bars: 200 nm.
5.2.2 Polymersome formation using the ‘kinetic trapping’ method
It was found that polymersome formation could be induced from negatively charged
PS140-b-PAA48 using proteins and peptides with a complementary positive charge to
drive the formation of vesicles over micelles in this system under ‘kinetic trapping’
conditions.† This method of membrane encapsulation for biomolecules overcomes the
issues of traditional polymersome encapsulation, as discussed in the previous section
† Parts of this work have been published: Hvasanov, D.; Wiedenmann, J.; Braet, F.; Thordarson, P. Chem. Commun. 2011, 47, 6314.

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-20
(Chapter 5.2.1), including denaturation of enzymes by harsh preparation conditions and
offers functionalities that polyelectrolyte capsules lack.48 For example, large internal
volumes could encapsulate a second functional (biological) system within the cavity for
cascade reactions as organelle mimics.34b The proteins and peptides employed in this
study induced polymersome formation and concurrently efficiently encapsulated in this
polymer vesicle system.
Aggregates were prepared by injection of a dissolved PS140-b-PAA48 in THF
into an enzyme containing phosphate buffer with the syringe submerged in solution at a
THF:water ratio of 1:6 (v/v). The solution was equilibriated for 24 h and dialysed
(50 kDa molecular weight cut-off) against water extensively to remove
non-encapsulated enzyme and THF. Due to the robust nature of polymer aggregates,
purification of enzyme-containing polymersomes via size exclusion chromatography is
not possible.
In this work, polymeric vesicles are defined as morphologies displaying distinct
contrast within individual particles indicating the presence of bilayer and cavity
formation, whereas micelles show uniform contrast. The addition of 5 M of a
positively charged biomolecule such as iso-1 cytochrome c to 8 M of PS140-b-PAA48
is very effective in promoting polymersome formation. Evidence of the dependence of
polymersome formation on positively charged proteins or peptides has been confirmed
by TEM microscopy studies as shown in Figure 5.14.49

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-21
Figure 5.14. TEM micrographs of polysytrene140-b-poly(acrylic acid)48(PS140-b-PAA48) aggregates in the presence of 5 M additives in PBS (25 oC). (a) PBS control (no enzyme). (b) Iso-1 cytochrome c. (c) Green fluorescent protein. (d) Poly-L-lysine. (e) Myoglobin. (f) Bovine serum albumin. (g) Calmodulin. Scale bars: 200 nm.
The polymer aggregates were further characterised by cryo-TEM as shown in Figure
5.15 to conclusively verify aggregate morphologies. The predominant observed
morphology were micelles in the presence of buffer control (Figure 5.15a), consistent
with classical TEM analysis (Figure 5.14). Additionally, the electron micrographs
confirm the formation of vesicles when formed with positively charged additives such
as iso-1 cytochrome c (cyt c) (Figure 5.15c), green fluorescent protein (GFP) (Figure
5.15d) and poly-L-lysine (Figure 5.15e).

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-22
Figure 5.15. Cryo-TEM micrographs of polysytrene140-b-poly(acrylic acid)48(PS140-b-PAA48) aggregates in the presence of 5 M additives in PBS (25 oC). (a) PBS control (no enzyme). (b) Calmodulin. (c) Iso-1 cytochrome c. (d) Green fluorescent protein. (e) Poly-L-lysine. Scale bars: 50 nm.
Interestingly, using negatively charged biomolecules such as bovine serum albumin
(BSA) and calmodulin (Figure 5.15b) in PBS resulted in micelle structures while with
the neutral myoglobin (Mb) inducer, an approximate 1:1 mixture of polymersomes and
micelles is obtained (Table 5.2).

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-23
Table 5.2. Effect of polymersome formation with different biomolecules at physiological pH.Proteina M.W. /kDa pI Charge Morphologyb
None (H2O) N/A N/A N/A MicellesNone (PBS) N/A N/A +/- Micellesc
Cyt c50 12.7 10.6 + VesiclesCyt cd (30 min) 12.7 10.6 + VesiclesCyt cd (24 h) 12.7 10.6 + MicellesGFP49 26.8e 8.3f + VesiclesPoly-L-lysine 1-5 N/A + VesiclesMb51 17.6 7.3 Neutral Micelles/Vesiclesg
BSA52 66 4.7 - MicellesCalmodulin53 16.7 3.9-4.3 - Micelles
a Conditions: enzyme in buffered PBS, pH 7.2. b Determined by TEM (see also Figure 5.14). c Asmall amount of vesicles observed. d Post-addition to mixture. e GFP can form non-covalent dimers and tetramers. f pI for GFP estimated by theoretical calculations of peptide sequence. g
Mb displayed 1:1 ratio of micelles:vesicles.
The combined addition of salts and positively charged biomolecules (cyt c), induces
electrostatic shielding along the partially ionised corona block which decreases the coil
dimension and reduces steric hindrance.39a,54 This allows more polymer chains to
aggregate, leading to increased core-chain stretching, resulting in structural changes
from micelles to rods to vesicles due to unfavourable thermodynamics associated with
excessive stretching. Interestingly, post-addition of cytochrome c to a polymer micelle
mixture after 30 minutes produces a mixed population of polymersomes and amorphous
aggregates compared to micelles and delayed post-addition after 24 hours has no effect
on resulting micelle morphologies (Table 5.2 and Figure 5.16).

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-24
Figure 5.16. TEM micrographs of polystyrene140-b-poly(acrylic acid)48(PS140-b-PAA48) aggregates in the presence of 5 M cytochrome c after post-addition to polymer micelle mixture in PBS (25 oC). (a) 30 min. (b) 24 h. Scale bars: 200 nm.
Only micelles are observed with PS140-b-PAA48 in the presence of BSA and
calmodulin. Further, there is no indication of BSA or calmodulin interacting with the
micelles in contrast to positively charged biomolecules. This suggests for negatively
charged proteins, there is electrostatic repulsion between the corona chains in
PS140-b-PAA48. The net result is that the aggregation of polymer chains and eventual
formation of polymersomes due to increased core-chain stretching is inhibited, leading
to micelle morphologies.
Given the variation in molecular weight and 3-dimensional structure of the
biomolecules screened, the primary factor causing vesicle formation are electrostatic
interactions between the biological and synthetic polymers rather than templation of the
surface curvature of the proteins. This is consistent with the myoglobin (Mb) results,
whereby, the aggregates form an approximate 1:1 ratio between micelles and
polymersomes. This is a result of the near neutrality of Mb with only a few positively
charged residues at pH 7.2 (pI = 7.3)51 even though it is fairly similar in size and shape
to cytochrome c (pI = 10.6).50 The induced polymersome formation can be generally
summarised as shown in Scheme 5.2.

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-25
Scheme 5.2. Schematic showing effect of charged biomolecule additives on polysytrene140-b-poly(acrylic acid)48 (PS140-b-PAA48) aggregate morphologies. Negatively charged biomolecules lead to micelles and positively charged additives induce polymersome structures.
5.2.2.1 Optimisation of polymersome formation
In order to determine optimum conditions for polymersome formation, such as minimal
structural defects and larger encapsulation volumes in a sample population, different salt
buffer conditions and temperatures were selected as shown in Table 5.3. It can be seen
that the optimum parameters required for polymersome preparation was room
temperature incubation in the presence of phosphate buffered saline (PBS). It is
interesting to observe that higher incubation temperatures and lower salt concentration
results in poorly formed polymersomes with smaller vesicle diameters. Lower salt
concentration buffers (20 mM NaH2PO4, pH 7.0) reduces the degree of electrostatic
repulsion causing smaller vesicles to form compared to PBS (pH 7.2) with sizes
162±61 nm and 376±259 nm at room temperature, respectively. In addition, the increase
in buffer temperature increases the rate of evaporation of the organic co-solvent
plasticiser (THF). As a result, the vesicles are ‘frozen’ over a shorter time scale
compared to ambient temperature due to decreasing core-chain mobilities leading to
smaller vesicles being formed.46 It should be noted that the size determination of
polymersomes were analysed using TEM microscopy. DLS was unsuitable for average

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-26
diameter determination of polymersomes as the aggregates were highly polydisperse,
resulting in poor data quality.
Table 5.3. Summary of salt buffer and temperature effects on polymersome formation.Enzymea Buffer T /oC Size /nm EE(%)b
Cyt c 20 mM NaH2PO4, pH 7.0 25 162±61 55±1Cyt c PBS, pH 7.2 25 376±259 66±7Cyt c 20 mM NaH2PO4, pH 7.0 40 96±25 -Cyt c PBS, pH 7.2 40 267±193 -GFP PBS, pH 7.2 25 308±136 35±1
a Enzymes prepared at 5 M concentration. b Determined for optimally formed polymersomes. EE(%) estimated using fluorescently labelled cyt c. Errors are standard deviation.
5.2.2.2 Concentration dependence of positively charged biomolecules
In order to probe the mechanism of polymersome formation, the percentage ratio of
polymersome-to-micelle aggregates were determined relative to the ratios of the
positive charge composition F+:55
]charges[]charges[charges][1 FF (5.2)
assuming 70% partial ionisation of the poly(acrylic acid) block ( 8 M) at pH 7.2.56
Additionally, yeast iso-1 cytochrome c has +14 charges (14 lysine residues)57 and green
fluorescent protein (Acropora millepora) has +17 charges (17 lysine residues).58 From
Figure 5.17, it can be seen that there is rapid binding leading to significant
polymersome formation with up to 60% formation with less than F+ 0.05 ( 1 M) for
both cytochrome c and green fluorescent protein due to the strong charge interactions
between corona and biomolecule.59 This is followed by saturation after addition of
greater than F+ 0.15 for both additives.§ At the saturation point, this is equivalent to an
approximate molar ratio of 2:1 polymer chain-to-enzyme.
§ Each data point is performed as independent experiments.

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-27
Figure 5.17. Influence of different enzyme inducers on relative polymersome/micelle (p/m) formation as a function of positive charge composition (F+) for cytochrome c ( )and green fluorescent protein ( ). Error bars are standard deviation.
Additionally, box plot analysis of the effect of charge ratio F+ on the size of
polymersomes for cytochrome c (Figure 5.18a) and green fluorescent protein (Figure
5.18b) shows that the polymersome size is independent of F+.

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-28
Figure 5.18. Influence of different enzyme inducers on dependence of polymersome size. (a) Cytochrome c. (b) Green fluorescent protein. Boxes extend to 25th and 75th and whiskers to 5th and 95th percentile, respectively. Red horizontal line marks the median.
5.2.2.3 Model of polymersome formation
Based on these observations, it is proposed that the positively charged cytochrome c and
green fluorescent protein are bound electrostatically to the coronal PAA block as shown
in Figure 5.19, causing aggregation of polymer chains and its vesicle structure. The 2:1
ion-pair supramolecular complex induces contraction of the effective PAA block length,
due to neutralisation of the PAA coronal chain with the multiply charged
cytochrome c57 and green fluorescent protein58 leading to micelle to vesicle
morphological transitions.39a This ion-pair interaction is reminiscent of gemini
surfactants, shown to form vesicles for long spacer groups between individual
amphiphiles consistent with the model.60

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-29
Figure 5.19. Proposed model for polymersome formation (PS140-b-PAA48) as a 2:1 ion-pair supramolecular complex. (a) Cytochrome c. (b) Green fluorescent protein.
Owing to the highly polydisperse nature of the polymersomes, diameters as large as
3 m have been observed as shown in Figure 5.20, allowing optical visualisation of
coronal encapsulation via fluorescence.
Figure 5.20. Representative histogram of PS140-b-PAA48 polymersomes for 5 Mcytochrome c in PBS at 25 oC.

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-30
The proposed model is consistent with confocal laser-scanning microscopy (CLSM)
images which show fluorescently labelled cyt c (cyt c-Oregon Green 488) and GFP
encapsulated in the membrane as shown in Figure 5.21.
Figure 5.21. Transmission light (a) and (c) and confocal laser scanning microscopy images (b) and (d) – excitation 488 nm – of enzymes encapsulated in the membrane of PS140-b-PAA48 polymersomes for fluorescently labelled cytochrome c (a) and (b) and green fluorescent protein (c) and (d). Scale bars: 50 m and inset 5 m. Contrast enhanced.
5.2.2.4 Biological stability
The membrane encapsulation of proteins and biomolecules into the polymersomes via
the injection method requires the presence of co-solvent, THF. In order to ensure that
the encapsulated enzymes are not denatured and remain biologically active for
encapsulation of light-harvesting bioconjugates and photo-induced charge transfer
studies further discussed in Chapter 6 and 7, biological stability studies were performed.

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-31
The photophysical properties of the proteins studied including green fluorescent
protein and cytochrome c were monitored via fluorescence or UV-Vis spectroscopy,
respectively, to ensure that the emission band or absorption bands were not quenched or
shifted, indicating correct folding of the protein tertiary structure after polymersome
formation. It was found that the encapsulation of GFP maintained its native folded state
as evident by its preserved fluorescence as shown in Figure 5.21d and Figure 5.22.
Figure 5.22. Fluorescence emission profiles of green fluorescent protein (GFP) excited at 476 nm with emission maxima 497 nm. Solution GFP (solid red line), encapsulated GFP (dashed black line).
The stability of cytochrome c conformation was monitored using the Soret band at
410 nm indicating that the tertiary structure was preserved as shown in Figure 5.23.61
Significant blue shifting (>10 nm) of the Soret band indicating protein denaturation was
not observed.

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-32
Figure 5.23. UV-Vis spectra of soret band (410 nm) of cytochrome c (cyt c). Solution cyt c (solid red line) and encapsulated cyt c (dashed black line). Sloping baseline for encapsulated cyt c due to scattering by polymersome aggregates.
The catalytic activity of cytochrome c has been widely reported with specific attention
on the oxidation of 2,2’-azino-bis[ethyl-benzothiazoline-(6)-sulfonic acid] (ABTS).62
Samples were prepared to 0.5 M solution (bulk) cytochrome c or encapsulated
cytochrome c, 200 M ABTS, 20 mM phosphate buffer at pH 7.0 and initiated with the
addition of hydrogen peroxide to a final concentration of 10 mM. Cytochrome c was
catalytically active after encapsulation by monitoring oxidation of ABTS at 415 nm as
shown in Figure 5.24.61 Interestingly, the encapsulated cytochrome c had an initial
increase in catalytic activity compared to native enzyme in solution due to sequestering
of substrate towards the enzyme.63

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-33
Figure 5.24. Catalytic activity of cytochrome c using the ABTS assay. (a) Chemical structure of 2,2’-azino-bis[ethyl-benzothiazoline-(6)-sulfonic acid] (ABTS). (b) Schematic of catalytic oxidation of ABTS using cytochrome c (cyt c). (c) Catalytic activity of solution cyt c (solid red line) and encapsulated cyt c (dashed black line) by monitoring oxidation of ABTS at 415 nm.
5.2.2.5 Enzyme encapsulation efficiencies
It is usually difficult to achieve high encapsulation efficiencies with polymersomes, due
to thicker membranes (d 8-21 nm) compared to liposomes (d 3-5 nm).36 Highly
polydisperse membrane thicknesses of 120±117 nm for PS140-b-PAA48 were observed
and thicker than previously reported for PS-b-PAA systems.38 This may be due to the

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-34
positively charged biomolecules acting as electrostatic bridges resulting in multilamellar
vesicles. A bilayer of PS140-b-PAA48 in the fully extended state is 92 nm.46
Furthermore, the bridging ability of these inducers is supported by observations of
polymersome aggregation and cluster formation in electron (Figure 5.14) and confocal
microscopy (Figure 5.21).
The method of encapsulation of biomolecules within the coronal block by
induction of positively charged additives overcomes the inherent disadvantage of
conventional polymersome encapsulation methods due to their thicker membranes.
Encapsulation efficiencies as high as 55±1% and 66±7% in 20 mM NaH2PO4 and PBS,
respectively, for cytochrome c and 35±1% for green fluorescent protein were observed
for optimised polymersomes (Table 5.3). It should be noted that the EE(%) of enzymes
in polymersomes was performed using steady-steady fluorescence of fluorescently
labelled cytochrome c (cyt c-Oregon Green 488) and native fluorescence of green
fluorescent protein to estimate loading (see Appendix D). This is due to the fact that
unlike liposomes, polymersomes are robust and treatment with a surfactant such as
Triton X-100 does not disassemble the aggregates which contributes to scattering of the
UV-Vis spectra. Hence, fluorescence emission provides a more accurate EE(%)
estimate.
5.3 Conclusion
In this Chapter, the formation of vesicles as compartments for enzyme encapsulation
and potential light-induced charge transfer studies were achieved. Lipid vesicles were
formed using the film hydration method and sized via extrusion to produce
monodisperse unilamellar vesicles of ca. 100 nm in diameter. Confirmation of
monodispersity and average diameter were achieved using dynamic light scattering

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-35
techniques. Additionally, unilamellar vesicles were characterised using traditional TEM
microscopy and cryo-TEM techniques and it was found that structural characterisation
of lamellar features using conventional TEM microscopy introduced artifacts as a result
of the necessity of negative stains and drying of specimen. The possibility of liposomes
based on egg PC and POPC as potential compartments were evaluated by the
encapsulation of model protein cytochrome c, which is used as an electron transfer
metalloprotein for photo-active bioconjugates. Encapsulation was confirmed using
confocal laser-scanning microscopy (CLSM) of fluorescently labelled cytochrome c.
However, low encapsulation efficiencies of 1.5±0.1% precluded their use for further
bioconjugate encapsulation studies.
Alternatively, polymersomes were formed as potential compartments. A novel
method of polymersome preparation by the injection method which induces
polymersome aggregates with the polyelectrolyte-based diblock copolymer,
PS140-b-PAA48, in a facile and biologically friendly manner was demonstrated.
Concomitantly, encapsulation of positively charged biomolecules occurs within the
coronal block. Additionally, this induced formation is versatile and can be applied to
various positively charged biomolecules with high encapsulation efficiencies allowing
for potential applications as nanoreactors64 (further discussed in Chapter 6 and 7) or
protein therapeutic vessels including use of polycationic copolymers as non-viral
vectors for DNA/RNA gene therapy65 and simplified preparation compared to
polyelectrolyte microcapsules.66 Similar to liposomes, polymersomes were
characterised by conventional and cryo-TEM microscopy as well as CLSM. The
biological activities of encapsulated enzymes were demonstrated to remain active using
spectroscopy studies and the ABTS assay. Due to the high encapsulation efficiencies of
55±1% and 66±7% in 20 mM NaH2PO4 and PBS, respectively, for cytochrome c and

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-36
35±1% for green fluorescent protein, polymersomes were selected as the most suitable
compartments for enzyme encapsulation.
5.4 References
(1) Bangham, A. D.; Standish, M. M.; Watkins, J. C. J. Mol. Biol. 1965, 13, 238.(2) (a) Mayer, L. D.; Hope, M. J.; Cullis, P. R. Biochim. Biophys. Acta, Biomembr.
1986, 858, 161. (b) Walde, P.; Ichikawa, S. Biomol. Eng. 2001, 18, 143.(3) Ostro, M. J. Liposomes; Marcel Dekker, Inc.: New York, 1983.(4) (a) Haluska, C. K.; Riske, K. A.; Marchi-Artzner, V.; Lehn, J.-M.; Lipowsky,
R.; Dimova, R. Proc. Natl. Acad. Soc. U. S. A. 2006, 103, 15841. (b) Blocher, M.; Walde, P.; Dunn, I. J. Biotechnol. Bioeng. 1999, 62, 36. (c) Gaspar, M. M.; Perez-Soler, R.; Cruz, M. E. M. Cancer Chemother. Pharmacol. 1996, 38, 373.
(5) Walde, P.; Marzetta, B. Biotechnol. Bioeng. 1998, 57, 216.(6) Szoka, F.; Papahadjopoulos, D. Proc. Natl. Acad. Soc. U. S. A. 1978, 75, 4194.(7) Enoch, H. G.; Strittmatter, P. Proc. Natl. Acad. Soc. U. S. A. 1979, 76, 145.(8) Schenning, A. P. H. J.; Escuder, B.; van Nunen, J. L. M.; de Bruin, B.; Löwik,
D. W. P. M.; Rowan, A. E.; van der Gaast, S. J.; Feiters, M. C.; Nolte, R. J. M. J. Org. Chem. 2000, 66, 1538.
(9) Das, N.; Gupta, S.; Mazumdar, S. Biochem. Biophys. Res. Commun. 2001, 286,311.
(10) Lichtenberg, D.; Markello, T. J. Pharm. Sci. 1984, 73, 122.(11) Fuhrhop, J.-H.; Mathieu, J. Angew. Chem. Int. Ed. 1984, 23, 100.(12) Mui, B.; Chow, L.; Hope, M. J. Methods Enzymol. 2003, 367, 3.(13) Angelova, M. I.; Dimitrov, D. S. Faraday Discuss. Chem. Soc. 1986, 81, 303.(14) Yeagle, P. The Structure of Biological Membranes; CRC Press, 2011.(15) Touitou, E.; Junginger, H. E.; Weiner, N. D.; Nagai, T.; Mezei, M. J. Pharm.
Sci. 1994, 83, 1189.(16) Ewert, K.; Slack Nelle, L.; Ahmad, A.; Evans Heather, M.; Lin Alison, J.;
Samuel Charles, E.; Safinya Cyrus, R. Curr. Med. Chem. 2004, 11, 133.(17) Vriezema, D. M.; Comellas Aragonès, M.; Elemans, J. A. A. W.; Cornelissen, J.
J. L. M.; Rowan, A. E.; Nolte, R. J. M. Chem. Rev. 2005, 105, 1445.(18) (a) Steinberg-Yfrach, G.; Liddell, P. A.; Hung, S.-C.; Moore, A. L.; Gust, D.;
Moore, T. A. Nature 1997, 385, 239. (b) Steinberg-Yfrach, G.; Rigaud, J.-L.; Durantini, E. N.; Moore, A. L.; Gust, D.; Moore, T. A. Nature 1998, 392, 479. (c) Kurihara, K.; Tamura, M.; Shohda, K.-i.; Toyota, T.; Suzuki, K.; Sugawara, T. Nat. Chem. 2011, 3, 775.
(19) Kent, C.; Carman, G. M. Trends Biochem. Sci. 1999, 24, 146.(20) Koynova, R.; Caffrey, M. Biochim. Biophys. Acta Rev. Biomembr. 1998, 1376,
91.(21) Ottaviani, M. F.; Matteini, P.; Brustolon, M.; Turro, N. J.; Jockusch, S.;
Tomalia, D. A. J. Phys. Chem. B 1998, 102, 6029.(22) Burchard, W. Adv. Polym. Sci. 1983, 48, 1.(23) (a) Kunitake, T.; Okahata, Y. J. Am. Chem. Soc. 1980, 102, 549. (b) Ottaviani,
M. F.; Favuzza, P.; Bigazzi, M.; Turro, N. J.; Jockusch, S.; Tomalia, D. A. Langmuir 2000, 16, 7368.

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-37
(24) Dimitrova, M. N.; Tsekov, R.; Matsumura, H.; Furusawa, K. J. Colloid Interface Sci. 2000, 226, 44.
(25) Taylor, K. A.; Glaeser, R. M. Science 1974, 186, 1036.(26) Dubochet, J.; Chang, J. J.; Freeman, R.; Lepault, J.; McDowall, A. W.
Ultramicroscopy 1982, 10, 55.(27) Frederik, P. M.; Hubert, D. H. W. Methods Enzymol. 2005, 391, 431.(28) Lipson, A.; Lipson, S. G.; Lipson, H. Optical Physics; 4th ed.; Cambridge
University Press: New York, 2011.(29) Peterson, J. R.; Thordarson, P. Chiang Mai J. Sci. 2009, 26, 236.(30) Kato, T.; Higuchi, M.; Endo, R.; Maruyama, T.; Haginoya, K.; Shitomi, Y.;
Hayakawa, T.; Mitsui, T.; Sato, R.; Hori, H. Pestic. Biochem. Physiol. 2006, 84,1.
(31) Huang, C.; Mason, J. T. Proc. Natl. Acad. Soc. U. S. A. 1978, 75, 308.(32) Klein, R. A. Biochim. Biophys. Acta 1970, 210, 486.(33) Yu, Y.; Zhang, L.; Eisenberg, A. Langmuir 1997, 13, 2578.(34) (a) Nardin, C.; Thoeni, S.; Widmer, J.; Winterhalter, M.; Meier, W. Chem.
Commun. 2000, 1433. (b) Vriezema, D. M.; Garcia, P. M. L.; Sancho Oltra, N.;Hatzakis, N. S.; Kuiper, S. M.; Nolte, R. J. M.; Rowan, A. E.; van Hest, J. C. M. Angew. Chem. Int. Ed. 2007, 46, 7378.
(35) Ranquin, A.; Versées, W.; Meier, W.; Steyaert, J.; Van Gelder, P. Nano Lett.2005, 5, 2220.
(36) O’Neil, C. P.; Suzuki, T.; Demurtas, D.; Finka, A.; Hubbell, J. A. Langmuir2009, 25, 9025.
(37) Roodbeen, R.; van Hest, J. C. M. BioEssays 2009, 31, 1299.(38) Ma, L.; Eisenberg, A. Langmuir 2009, 25, 13730.(39) (a) Cameron, N. S.; Corbierre, M. K.; Eisenberg, A. Can. J. Chem. 1999, 77,
1311. (b) Blanazs, A.; Armes, S. P.; Ryan, A. J. Macromol. Rapid Commun.2009, 30, 267.
(40) Shen, H.; Eisenberg, A. J. Phys. Chem. B. 1999, 103, 9473.(41) Zhang, L.; Eisenberg, A. Science 1995, 268, 1728.(42) Lensen, D.; Vriezema, D. M.; van Hest, J. C. M. Macromol. Biosci. 2008, 8,
991.(43) Lim Soo, P.; Eisenberg, A. J. Polym. Sci. Part B: Polym. Phys. 2004, 42, 923.(44) Vriezema, D. M.; Hoogboom, J.; Velonia, K.; Takazawa, K.; Christianen, P. C.
M.; Maan, J. C.; Rowan, A. E.; Nolte, R. J. M. Angew. Chem. Int. Ed. 2003, 42,772.
(45) (a) Brandrup, J.; Immergut, E. H. Polymer handbook; 3rd ed.; Wiley-Interscience: New York, 1989. (b) Lide, D. R. CRC handbook of chemistry and physics; 78th ed.; CRC Press: New York, 1997-1998.
(46) Zhang, L.; Eisenberg, A. J. Am. Chem. Soc. 1996, 118, 3168.(47) Report on Carcinogens, 11th Ed.; U.S. Department of Health and Human
Services, Public Health Service, National Toxicology Program.(48) He, Q.; Cui, Y.; Li, J. Chem. Soc. Rev. 2009, 38, 2292.(49) Hvasanov, D.; Wiedenmann, J.; Braet, F.; Thordarson, P. Chem. Commun. 2011,
47, 6314.(50) Minakami, S. J. Biochem. 1955, 42, 749.(51) (a) Darbre, P. D.; Romero-Herrera, A. E.; Lehmann, H. Biochim. Biophys. Acta
1975, 393, 201. (b) Radola, B. J. Biochim. Biophys. Acta 1973, 295, 412.(52) Ang, W. S.; Elimelech, M. J. Membr. Sci. 2007, 296, 83.

Chapter 5 Supramolecular Aggregates for Protein Encapsulation
5-38
(53) (a) Lin, Y. M.; Liu, Y. P.; Cheung, W. Y. J. Biol. Chem. 1974, 249, 4943. (b) Crouch, T. H.; Klee, C. B. Biochemistry 1980, 19, 3692. (c) Watterson, D. M.; Sharief, F.; Vanaman, T. C. J. Biol. Chem. 1980, 255, 962.
(54) Discher, D. E.; Eisenberg, A. Science 2002, 297, 967.(55) Lindhoud, S.; de Vries, R.; Norde, W.; Stuart, M. A. C. Biomacromolecules
2007, 8, 2219.(56) Choi, J.; Rubner, M. F. Macromolecules 2005, 38, 116.(57) Louie, G. V.; Brayer, G. D. J. Mol. Biol. 1990, 214, 527.(58) Angelo, C.; Denzel, A.; Vogt, A.; Matz, M. V.; Oswald, F.; Salih, A.; Nienhaus,
G. U.; Wiedenmann, J. Mar. Ecol. Prog. Ser. 2008, 364, 97.(59) Choucair, A.; Eisenberg, A. Eur. Phys. J. E: Soft Matter Biol. Phys. 2003, 10,
37.(60) Prévost, S.; Wattebled, L.; Laschewsky, A.; Gradzielski, M. Langmuir 2010, 27,
582.(61) Peterson, J. R.; Smith, T. A.; Thordarson, P. Chem. Commun. 2007, 1899.(62) Valderrama, B.; García-Arellano, H.; Giansanti, S.; Baratto, M. C.; Pogni, R.;
Vazquez-Duhalt, R. FASEB J. 2006, 20, 1233.(63) Gauthier, M. A.; Klok, H.-A. Polym. Chem. 2010, 1, 1352.(64) Minten, I. J.; Claessen, V. I.; Blank, K.; Rowan, A. E.; Nolte, R. J. M.;
Cornelissen, J. J. L. M. Chem. Sci. 2011, 2, 358.(65) Ropert, C. Braz. J. Med. Biol. Res. 1999, 32, 163.(66) Zelikin, A. N.; Becker, A. L.; Johnston, A. P. R.; Wark, K. L.; Turatti, F.;
Caruso, F. ACS Nano 2007, 1, 63.

Chapter 6
Photoinduced Electron Transfer Studies of Cytochrome c

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-2
6 Photoinduced Electron Transfer Studies of Cytochrome c
In this Chapter, photoinduced electron transfer studies between Ru(II)-bisterpyridine
complexes and iso-1 cytochrome c (cyt c) as non-covalent mixtures and asymmetric
bioconjugate 8-cyt c were explored. Electron transfer studies were carried out as bulk
solution bioconjugates and membrane encapsulated 8-cyt c. The electron transfer
processes resulting in reduction of the heme of cytochrome c is followed by steady-state
UV-Vis spectroscopy. Electron transfer to the heme was achieved by irradiation using a
465 nm LED light under anaerobic conditions at room temperature in the presence of
sacrificial electron donor, ethylenediaminetetraacetic acid (EDTA). Quantum
efficiencies for bulk and polystyrene140-b-poly(acrylic acid)48 (PS140-b-PAA48)
encapsulated 8-cyt c, were estimated to be 5.9±1.5 10-4% and 1.1±0.3 × 10-3%,
respectively.
In addition, light-induced electron transfer studies between a non-covalent
mixture of Ir(III)-bisterpyridine complex and horse heart cytochrome c were studied to
mimic nitrite reductase behaviour. Excitation and catalysis were performed using a
372 nm LED source under anaerobic conditions at room temperature. Nitrite reductase
behaviour was observed in the presence of 100 nm L—
palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC) liposomes and detected using
the Griess assay.
6.1 Background
Electron transfer processes in a direct and controlled fashion are critical for all
functioning living organisms.1 The most notable electron transfer reactions in nature are
photosynthesis and respiration which is responsible for the generation of adenosine

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-3
triphosphate (ATP). ATP is used to drive biochemical reactions and to store energy by
converting carbon dioxide into carbohydrates.2
Cytochrome c is a naturally occurring metalloprotein with iron (cytochroe) heme
cofactor and is part of the electron transport chain in photosynthesis and mitochondria
(respiration) where quinones are its natural electron donors.1 Cytochrome c is heavily
studied for electron transfer mechanisms as it is a robust small protein with an
accessible redox centre, highly stable and soluble in aqueous solvents and is well
characterised with crystal structures.
To improve the understanding of electron transfer processes in complex
biological systems, electron transfer models have been developed and have been
experimentally studied using the model protein cytochrome c with notable contributions
by Gray3 and Millet4 over the last three decades. Electron transfer studies involving
cytochrome c has generally been achieved by modification of either wild type or mutant
species with ruthenium bipyridine complexes at different positions via functionalisation
of histidine3a,5, lysine4a,6 or cysteine residues.7 Electron transfer rates of the
bioconjugates can be elucidated using molecular modelling by combining crystal
structure data as shown in Figure 6.1. These studies have been used to measure electron
pathways in proteins, such as electron tunnelling electron transfer velocities are
dependent on the peptide matrix3d as well as distance dependence between donor and
acceptor components.8 Moreover, the electron transfer process of cytochrome c with its
biological redox partners cytochrome c oxidase9, cytochrome c peroxidase4b and
plastocyanin10 have been studied.

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-4
Figure 6.1. X-ray crystal structure of cytochrome c peroxidase:cytochrome c complex. Mutant yeast iso-1 cytochrome c was functionalised, where the histidine residue 39 has been substituted with cysteine and attached to a tris(bipyridyl)ruthenium(II) complex by molecular modelling.7
The Thordarson group have reported light-harvesting bioconjugates capable of electron
transfer based on Ru(II)-bisterpyridine iso-1 cytochrome c bioconjugates.11
Ru(II)-bisterpyridine complexes have shorter fluorescent lifetimes and lower quantum
yields compared to bipyridine complexes and therefore less desirable for room
temperature electron transfer studies.12 However, they are employed in this project due
to ease of preparation due to their achirality and ease of 4’-functionalisation.
6.2 Room Temperature Photoinduced Electron Transfer Studies
The room temperature light-activated electron transfer between ruthenium bipyridine
complexes (donor) and variants of cytochrome c (acceptor) using sacrificial electron

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-5
donors such as aniline13 and ethylenediaminetetraacetic acid (EDTA)4b,14 have been well
established in the literature. Peterson et al.11a in the Thordarson group have previously
demonstrated room temperature photoinduced electron transfer in Ru(II)-bisterpyridine
cytochrome c bioconjugates using a short and long chain spacer. Photoreduction was
monitored using steady state UV-Vis spectroscopy and low temperature time-resolved
fluorescence lifetime measurements. It was found that long chain spacers resulted in
optimum electron transfer as the use of short chain spacers caused inactivation of the
protein.
Based on the findings of Peterson et al.11a, photoreduction studies of the novel
long chain spacer bioconjugate 8-cyt c and non-covalent mixtures with reference
complex 6 and iso-1 cytochrome c as shown in Figure 6.2, discussed in Chapter 2 and 3
were used to determine the effect of membrane encapsulation for use as light-activated
photosynthesis-respiratory hybrids (further discussed in Chapter 7).
Figure 6.2. Bioconjugate 8-cyt c and reference compounds [Ru(tpy)2]2+ 6 and iso-1cytochrome c (cyt c) used for room temperature photoreduction studies.

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-6
Previous photoreduction studies by the Thordarson group were based on
literature procedures by Hamachi and co-workers, who measured room temperature
electron transfer in apomyoglobin reconstituted with ruthenium bisterpyridine complex
appended heme as the enzyme cofactor.15 Reported photoexcitation conditions were
performed at room temperature with a high pressure mercury lamp (equipped with
<450 nm filter) in the presence of sacrificial electron donor EDTA under argon
atmosphere. It was found that complete photoreduction was observed after 5 h in the
presence of 20 mM EDTA, pH 6.3 at 25 oC.
In contrast to Hamachi et al.15 or Peterson et al.11b which used either a high
pressure mercury lamp or xenon arc lamp that are broad spectrum light sources,
photoreduction studies in this project were based on well-defined narrow wavelength
LED sources (465 nm). In order to demonstrate light-activated electron transfer in
bioconjugate 8-cyt c, samples were prepared in a specialised small volume cuvette,
degassed and irradiated (2.5 cm) with a constant area at a bioconjugate concentration of
2.3±0.1 M and an equivalent 1:1 non-covalent mixture of complex 6 and iso-1 cyt c in
a 5 mM phosphate buffer, 5 mM EDTA at pH 7.0 in either bulk solution or encapsulated
in the PS140-b-PAA48 membrane. This experiment was also conducted under different
conditions to determine the effect on heme reduction rate, including the presence of
oxygen or the absence of sacrificial electron donor EDTA. The concentrations were
estimated by UV-Vis absorption spectroscopy with molar absorption coefficients: iso-1
cyt c/8-cyt c ( 410 = 97.6 mM-1cm-1)16 and reference complex 6 ( 476 = 17.7 mM-1cm-1).12
It should be noted that concentrations used for photoreduction studies were low to
prevent intermolecular electron transfer in bioconjugate samples.

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-7
The reduction rate of the heme group in cytochrome c bioconjugate 8-cyt c was
monitored using UV-Vis spectroscopy from 350 nm to 650 nm as shown in Figure 6.3
and the degree of reduction was calculated based on absorbance changes at 550 nm
according to Equation 6.1:
100%)Reduction(oxred
oxt
AAAA (6.1)
where At is the absorbance after reduction time t, Aox is the original absorbance of
oxidised cytochrome c or bioconjugate and Ared is the final absorbance after complete
reduction.
Figure 6.3. UV-Vis absorbance spectra showing an increase in 550 nm absorbance band over time corresponding to photoreduction of Ru(II) bioconjugate 8-cyt c(2.3±0.1 M) in 5 mM phosphate buffer, 5 mM ethylenediaminetetraacetic acid (EDTA), pH 7.0. Samples irradiated with 20±2.3 mW/cm2 of 465 nm light.

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-8
Bioconjugate 8-cyt c appeared to be fully reduced in 5 mM phosphate buffer, 5 mM
ethylenediaminetetraacetic acid (EDTA), pH 7.0 under degassed conditions in a period
of 50 minutes as shown in Figure 6.4.
Figure 6.4. Room temperature photoreduction of ruthenium(II)-bisterpyridine based iso-1 cytochrome c (cyt c) samples irradiated with 20±2.3 mW/cm2 of 465 nm light. All sample measurements were made at a concentration of 2.3±0.1 M in 5 mM sodium dihydrogen phosphate, 5 mM ethylenediaminetetraacetic acid (EDTA), pH 7.0. Ru(II)bioconjugate 8-cyt c ( ), 8-cyt c:polymersome ( ), 8-cyt c non-degassed ( ), 8-cyt c no EDTA ( ), non-covalent (1:1) Ru(II) 6:cyt c mixture ( ) and non-covalent (1:1) Ru(II)6:cyt c:polymersome mixture ( ).† Error bars indicate standard deviation.
To interpret changes in the 550 nm absorbance band in terms of quantum yield ( ) for
photoreduction, the rate of photon emission of the 465 nm light source was determined
to be 1.40±0.02 × 1016 photons/s at a distance of 2.5 cm between sample and light
source. In the context of these studies, quantum yield is expressed as the ratio of initial
† Different photoreduction conditions were scaled relative to bulk 8-cyt c experiment as a ratio of molar percentage of reduced protein.

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-9
rate of heme reduction vs. absorbed photons. The number of photons absorbed could be
determined by correcting for the optical density of the Ru(II) complex. A quantum
efficiency ( ) of bioconjugate 8-cyt c was estimated to be 5.9±1.5 10-4% (see
Appendix F). It is most likely that the low quantum efficiency is attributed to the short
lifetimes of Ru(II)-bisterpyridine complex at room temperature and the relatively long
distance between the donor and acceptor.11b,12
The Ru(II) bioconjugate 8-cyt c was analysed via molecular modelling to
estimate the long range electron transfer distance from Ru(II) complex to heme as
shown in Figure 6.5. The donor-acceptor distance was estimated by summation of the
linear bond distance between the heme group and cysteine (CYS102) in the crystal
structure of iso-1 cytochrome c (Fe-S: 11 Å) and the maximum distance between the
ruthenium to thiol (CYS102) in a protein attached complex 8 (Ru-S: 21 Å), resulting in
an estimated maximum distance between ruthenium and heme centre of 32 Å.
Figure 6.5. A model of 8-cyt c generated from the X-ray crystal structure of yeast iso-1cytochrome c (cyt c) attached to the X-ray structure of ruthenium(II)-bisterpyridine complex 8 via the CYS102 residue. Maximum distance between Ru-Fe estimated as
32 Å. Cytochrome c structure derived from the protein data bank file ‘1YCC’.17

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-10
To determine the effect of electron transfer after encapsulation within PS140-b-PAA48
polymersome membranes, the polymer aggregates were characterised by transmission
electron microscopy (TEM) prior to room temperature photoreduction measurements to
ensure that bioconjugate 8-cyt c induced polymersome formation as discussed in
Chapter 5. As shown in Figure 6.6, polymersome aggregates were observed with an
average diameter of 290±132 nm.
Figure 6.6. TEM micrograph of polystyrene140-b-poly(acrylic acid)48 (PS140-b-PAA48)polymersome aggregates in the presence of 14 M Ru(II) bioconjugate 8-cyt c in PBS (25 oC). Average polymersome diameter of 290±132 nm. Scale bar: 200 nm.
As shown in Figure 6.4, encapsulation of Ru(II) 8-cyt c resulted in an increase in initial
rate of reduction of heme and fully reduced within 50 minutes. Encapsulation was
estimated to be double the of bulk solution 8-cyt c photoreduction with a of
1.1±0.3 × 10-3%. The increase in could be explained using semi-classical theory
(Marcus-Hush theory of electron transfer) which describes electron tunnelling in
proteins as shown in Equation 6.1:3d,18

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-11
TkGH
Tkhk
B
o
ADB
ET 4)(exp)4(
22
2
3
(6.1)
where the electron transfer rate (kET) from a donor (D) to an acceptor (A) at fixed
separation and orientation depends on the reaction driving force (- Go), a nuclear
reorganisation parameter ( ) and the electronic-coupling strength representing the
probability that an electron tunnels through the potential barrier between A and D
(HAD). Also, kB is the Boltzmann constant, T is temperature and h is Planck’s constant.
It has been reported that embedding reactants in a membrane (low dielectric medium)
dramatically reduces reorganisational energies ( ), hence, the encapsulation of 8-cyt c
in the polymersome membrane leads to an increase in electron transfer rate (kET).3d
Another possibility leading to increased may be due to the flexible linker used in
complex 8 for attachment to cytochrome c allowing the complex to lie flat on the
protein surface when embedded in the polyelectrolyte membrane as shown in Figure
6.5. This decrease in distance between the donor and acceptor increases the coupling
strength HAD by reducing the potential barrier.1
To determine the effect of covalent linkage of Ru(II)-bisterpyridine donor to
cytochrome c, non-covalent studies using a 1:1 mixture of reference complex 6 and
cytochrome c in bulk solution or encapsulated in membrane were performed in the
presence of EDTA and degassed. Complex 6 was chosen as a control complex
compared to complex 8 as the lack of maleimide functional group prevents reaction
with protein. It was observed in Figure 6.4 that a dramatic decrease in photoreduction
yield resulted in non-covalent mixtures. This indicates that covalent attachment to
protein is essential to ensure proximity between donor and acceptor to increase
photoreduction efficiency. However, the behaviour of increasing was observed after
encapsulation, consistent with covalent bioconjugate 8-cyt c studies. The reduction

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-12
efficiencies and quantum efficiencies of bioconjugate 8-cyt c and non-covalent mixtures
in bulk solution and membrane encapsulation is summarised in Table 6.1.
Table 6.1. Estimated rates of heme reduction, reduction efficiency and quantum efficiency ( ) for the photoinduced reduction of Ru(II)-cytochrome c systems.Samplea Heme reduction
(electrons/s)bEfficiency(%) (electrons/photons)c
(%) (electrons/absorbed photons)d
8-cyt ce 7.9±1.8 109 5.6±0.1 10-5 5.9±1.5 10-4
8-cyt cf 1.5±1.8 1010 1.1±0.3 10-4 1.1±0.3 10-3
6::cyt ce,g 4.6±0.6 109 3.3±0.6 10-5 3.4±0.6 10-4
6::cyt cf,g 5.1±1.3 109 3.6±1.0 10-5 3.8±1.1 10-4
a Sample concentration of 2.3±0.1 M in 5 mM sodium dihydrogen phosphate, 5 mMethylenediaminetetraacetic acid (EDTA), pH 7.0 at 25 oC with 80 L volume. Irradiated with 20±2.3 mW/cm2 of 465 nm light. b Initial rate of heme reduction was estimated using amount of protein reduced in the first 1860 s interval. c Efficiency(%) was determined by dividing initial rate of heme reduction by incident photons (1.40±0.2 1016 photons/s). Incident photons calculated from power ouput over an irradiation area of 1.0 0.3 cm for a 465 nm photon. d Quantum efficiency was determined by correcting for the optical density of the solutions (see Appendix F). e Bulk solution measurement. f Encapsulated measurement in PS140-b-PAA48membrane. g Non-covalent mixture. Errors are standard deviation.
Photoexcitation of Ru(II)-bisterpyridine to the triplet metal-to-ligand charge-transfer
(3MLCT) state is short lived and non-luminescent at room temperature with an
excitated-state lifetime estimated to be 250 ps.19 To investigate if the presence of
oxygen significantly quenches the excited chromophore Ru(II)*, photoreduction studies
were performed without degassing. A quenching of photoreduction by 50% was
observed in non-degassed experiments as shown in Figure 6.4. Additionally, the
presence of sacrificial electron donor, EDTA, is essential as bioconjugate 8-cyt c in
phosphate buffer shows negligible photoreduction (Figure 6.4).
6.2.1 Biological activity using cytochrome c oxidase assay
The biological activity of bioconjugate 8-cyt c in bulk solution and encapsulated in
PS140-b-PAA48 membranes were measured by cytochrome c oxidase after
photoreduction. The cytochrome c oxidase assay is used to demonstrate that the proteins

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-13
remain biologically active after bioconjugation and encapsulation in the presence of
organic co-solvents, acetonitrile and tetrahydrofuran, respectively. Additionally,
unmodified iso-1 cytochrome c was oxidised as a reference. The oxidation of reduced
cytochrome c by cytochrome c oxidase can be followed by UV-Vis absorbance
following the characteristic decrease in the 550 nm absorbance band as shown in Figure
6.7. Biological activity was measured using horse heart cytochrome c oxidase. The
samples were prepared at 2.3 M reduced cytochrome c or bioconjugate 8-cyt c (bulk or
encapsulated) in 5 mM phosphate buffer, 5 mM EDTA, pH 7.0 and oxidation initiated by
addition of a catalytic amount of cytochrome c oxidase (0.25 M).
Figure 6.7. Biological activity of iso-1 cytochrome c ( ), Ru(II) bioconjugate 8-cyt cand ( ) and 8-cyt c:polymersome ( ). Samples are at concentration of 2.3±0.1 M in 5 mM sodium dihydrogen phosphate, 5 mM ethylenediaminetetraacetic acid (EDTA), pH 7.0 and oxidation achieved using a catalytic amount of horse heart cytochrome coxidase (0.25 M). Error bars indicate standard deviation.

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-14
As shown in Figure 6.7, bioconjugates 8-cyt c in bulk solution and encapsulated in
membranes demonstrate comparable biological activity to that of native iso-1
cytochrome c indicating attachment of complex 8 to the CYS102 residue does not
influence cytochrome c oxidase binding.4d Furthermore, the ability of cytochrome c
oxidase to oxidise bioconjugate 8-cyt c after encapsulation also supports the
polymersome model discussed previously in Chapter 5, where bioconjugates are
encapsulated in the coronal block of the PS-b-PAA diblock copolymer.20
6.3 Nitrite Reductase Mimics
Nitric oxide (NO) is a biologically relevant cellular signalling molecule responsible for
mediating biological responses such as hypoxic vasodilation, regulation of gene and
protein expression and cytoprotection after ischemia-reperfusion.21 Nitrite anions (NO2-
) comprise the largest vascular storage pool of nitric oxide under physiological
conditions and regulates nitric oxide levels via reduction reactions with
heme-containing proteins.22 It was originally proposed that nitrite was biologically
inert23, however, it has been demonstrated recently that nitrite is an intrinsic vasodilator
and signalling molecule at physiological concentrations in vivo.24 Similar to nitric oxide,
nitrite has been shown to inhibit cytotoxicity and apoptosis after ischemia-reperfusion
injury of the heart, liver and brain.25

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-15
Two distinct types of nitrite reductases are capable of catalysing the reduction of
nitrite anions into nitric oxide, heme cd1 containing chromophores and copper based
enzymes.26 The reaction catalysed by both types of enzymes can be represented by the
simple Equation 6.2:
OHNO2HeNO 22 (6.2)
It should be noted however, that other nitrite reductases reduce nitrite to ammonia such
as cytochrome c nitrite reductase (Escherichia coli) which catalyses the six electron
reduction of nitrite to ammonia.27
One of the most common enzymes responsible for nitric oxide synthesis in
biological systems is nitric oxide synthase (EC 1.14.13.39). Nitric oxide synthase is a
complex enzymatic system which acts on molecular oxygen, arginine and reduced
nicotinamide adenine dinucleotide phosphate (NADPH) to produce nitric oxide,
citrulline and NADP+ as shown in Figure 6.8.28 Five additional cofactors including
flavin mononucleotide (FMN), flavin adenine dinucleotide (FAD), heme, calmodulin
and tetrahydrobiopterin as well as two divalent cations (calcium and heme iron) are
required for catalytic function.
It has been demonstrated that heme containing proteins are capable of additional
roles as nitrite reductase mimics including hemoglobin24, myoglobin29, xanthine
oxidoreductase30, cytochrome c21 and cytochrome c oxidase31 under anoxic conditions.
In this project, nitrite reductase activity of cytochrome c was induced by
light-activation.

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-16
Figure 6.8. Schematic of nitric oxide synthase (EC 1.14.13.39). (a) Representation showing nitric oxide synthesis by acting on molecular oxygen, arginine and reduced nicotinamide adenine dinucleotide phosphate (NADPH) to produce nitric oxide, citrulline and NADP+.32 (b) Solid ribbon representation of human inducible nitric oxide synthase with heme group indicated as stick representations. The structure was derived from the protein data bank file ‘1NSI’.33
6.3.1 Photoinduced nitrite reductase activity of cytochrome c
Basu et al.21 have shown that chemically reduced cytochrome c is capable of reducing
nitrite anions under anaerobic conditions. In order for heme proteins to react with
nitrite, a pentacoordinate state is required. Cytochrome c is a six-coordinate heme iron
which hinder reactions which nitrite, however, in the presence of anionic phospholipids
the weakening or breaking of the iron-methionine bond can occur resulting in a

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-17
pentacoordinate state to activate nitrite reductase activity.34 In addition, the
iron-methionine bond in cytochrome c has been proposed to play a role in apoptosis.35
In this study, an iridium(III)-bistepyridine complex 18 was used to photo-reduce
cytochrome c in the presence of POPC liposomes using a UV LED light source
(372 nm) as shown in Scheme 6.1.
Scheme 6.1. Photoinduced nitrite reductase activity of horse heart cytochrome c (cyt c)using complex 18 irradiated with UV light (372 nm) to reduce cyt c under anaerobic conditions in the presence of L—palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC) liposomes which induces pentacoordination of heme state. Solid ribbon representation of horse heart cytochrome c with heme group, MET80 and HIS80 indicated as stick representations. The structure was derived from the protein data bank file ‘1HRC’.36
Photoreduction studies inducing nitrite reductase activity were performed using a 1:1
non-covalent mixture between Ir(III) complex 18 and cytochrome c as work in the
Thordarson group have previously shown that photoreduction efficiency is similar to the
bioconjugate equivalent unlike its ruthenium(II) counterpart.37 Due to experimental
limitations preventing large scale synthesis of Ru(II) bioconjugate 8-cyt c, photoinduced

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-18
nitrite reductase activity studies could not be performed. Additionally, horse heart
cytochrome c was used in this study rather than yeast iso-1 cytochrome c due to its low
cost and does not require further purification.
Nitrite reductase measurements were conducted using conditions adapted from
Basu and coworkers.21 Samples were prepared in plastic eppendorf vials contained in a
vacuum flask and irradiated (2.0 cm) using a 1:1 non-covalent mixture of horse heart
cyt c and complex 18 at a concentration of 400 M, 10 M sodium nitrite, 3 mg/mL
POPC liposomes (4 mM) in a 25 mM phosphate buffer, 5 mM EDTA at pH 5.4 under
anoxic conditions. The concentration of horse heart cytochrome c was estimated by
UV-Vis absorption spectroscopy with molar absorption coefficient
410 = 106.1 mM-1cm-1.38 After irradiation for 3 h followed by incubation for a further
14 h, the final nitrite concentration was determined via the Griess assay as shown in
Scheme 6.2 which converts nitrite into a deep purple azo compound and monitored at
540 nm using UV-Vis absorbance to estimate nitric oxide conversion by difference (see
Appendix C).39
Scheme 6.2. Detection of nitrite via the Griess assay by converting nitrite into a deep purple azo compound and concentration measured spectroscopically at an absorbance of 540 nm.39

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-19
It should be noted that in order to prevent spectroscopic interference by cytochrome c,
the solution was centrifuged using a 3 000 molecular weight cut-off concentrator to
remove protein and filtrate was analysed using the Griess assay. Additionally, studies
were performed at pH 5.4 as Equation 6.2 predicts faster nitrite reduction under acidic
conditions.
Irradiation with 372 nm light of the non-covalent mixture of Ir(III) 18 and
cytochrome c in the presence of POPC liposomes at an initial concentration of 10 M
sodium nitrite under anoxic conditions resulted in photo-activated nitrite reductase
activity leading to an 82% conversion of nitrite anions into nitric oxide (1.8±0.2 M
nitrite remaining) as shown in Figure 6.9. It should be noted that although the liposomes
induce a pentacoordinate heme, unconverted nitrite was present when a forty-fold
excess of cytochrome c relative to nitrite was used due to auto-inhibition of
cytochrome c caused by binding of nitric oxide preventing further reactions with nitrite
and there is always substantial hexacoordinate cytochrome c remaining.21
To eliminate the possibility of interference during the centrifugation step such as
nitrite-membrane binding, a nitrite only control was studied and no effect on nitrite
concentration was observed after centrifugation (NO2- = 10.5±1.0 M). Additionally,
chemically reduced cytochrome c (no liposomes) exhibited neglible nitrite reductase
behaviour as no nitrite was converted during the time-scale of the study within
experimental error, NO2- = 9.3±1.3 M. Nitrite reductase activity was observed only
after reduced cytochrome c was incubated in the presence of POPC liposomes with
50% NO conversion (4.9±1.6 M nitrite remaining). Reduced cytochrome c in the
presence of liposomes is essential to induce nitrite reductase activity as the
cytochrome c (oxidised):liposome mixture exhibited no observable nitrite conversion
Pre-reduced chemically with sodium dithionite, desalted and dialysed against water for 2 h.

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-20
(NO2- = 11.5±1.7 M) as shown in Figure 6.9. This also indicates that zwitterionic
phospholipid liposomes can induce pentacoordination in addition to anionic
phospholipids.34
Figure 6.9. Room temperature photoinduced nitrite reductase activity of horse heart cytochrome c (cyt c) under an anaerobic environment using a 1:1 non-covalent mixture of cyt c and Ir(III) complex 18 (400 M), 3 mg/mL 100 nm POPC liposomes (4 mM), sodium nitrite (10 M) and control conditions. Irradiated with a 0.04±0.02 mW/cm2 UV light source (372 nm) at a distance of 2.0 cm for 3 h and incubated for a further 13 hand 45 min. All sample measurements were made in 25 mM sodium dihydrogen phosphate, 5 mM ethylenediaminetetraacetic acid (EDTA), pH 5.4. Error bars arestandard deviation. Pre-reduced chemically with sodium dithionite, desalted and dialysed for 2 h.

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-21
To confirm that nitrite reductase activity was photoinduced, a cytochrome c
(oxidised):liposome mixture was irradiated with 372 nm in the absence of Ir(III) 18 and
no observable nitrite reduction was detected (NO2- = 9.5±0.3 M). Furthermore, a dark
control sample composed of Ir(III) 18:cytochrome c (oxidised):liposomes was
investigated, however, approximately 57% NO conversion was observed (4.3±0.8 M
nitrite remaining) as shown in Figure 6.9. This is attributed to sample exposure to
ambient light during sample preparation resulting in photoreduction of cytochrome c. It
has been shown in the Thordarson group that Ir(III)-bisterpyridine complexes are
extremely effective in heme photoreduction even in aerobic conditions in contrast to
Ru(II) complexes allowing cytochrome c to be fully reduced in under 10 min.37 Also,
the long-lived photoreduction EDTA by-products (amine radicals) are capable of
continued protein reduction once transferred to a dark environment.
6.4 Conclusion and Future Work
In this Chapter, the photophysical properties of a Ru(II)-bistepyridine cytochrome c
bioconjugate 8-cyt c was investigated. Using steady-state UV-Vis absorption
spectroscopy studies, the photoreduction of 8-cyt c could be followed by monitoring the
increase in the 550 nm absorption band corresponding to heme reduction. It was found
that Ru(II) 8-cyt c could be fully reduced in 50 min via irradiation with a 465 nm LED
light source. The quantum efficiency ( ) of bioconjugate 8-cyt c in bulk solution was
estimated to be 5.9±1.5 10-4%. In order to determine the effect of membrane
encapsulation for use as a component in a semi-synthetic electron transport chain,
bioconjugate 8-cyt c was encapsulated in diblock copolymer
polystyrene140-b-poly(acrylic acid)48 (PS140-b-PAA48) as discussed in Chapter 5.
Polymersome formation was confirmed by transmission electron microscopy with an

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-22
average diameter of 290±132 nm. The encapsulated conjugate 8-cyt c, exhibited a
two-fold enhancement of the rate of heme reduction with an estimated of
1.1±0.3 × 10-3%. Molecular modelling of 8-cyt c showed that electron transfer is
occurring over a maximum distance between the ruthenium and heme centre of 32 Å.
Expanding on photoreduction of cytochrome c, electron transfer studies between
a non-covalent mixture of Ir(III)-bisterpyridine complex 18 and horse heart
cytochrome c using a 372 nm UV light source was investigated to induce nitrite
reductase mimicry behaviour. It was shown that in the presence of zwitterionic
phospholipid (POPC) liposomes (100 nm), a pentacoordinate heme state could be
induced allowing reactions with nitrite anions to form nitric oxide. The photo-activated
nitrite reductase activity of cytochrome c exhibited an 82% conversion of nitrite anions
into nitric oxide after irradiation under anoxic conditions in an acidic phosphate
buffered environment (pH 5.4). Due to experimental limitations, the exact irradiation
power could not be determined due scattering and absorption by the vacuum flask and
nitrite levels could only measured at a single time point. For future work, a custom
experimental set-up could be developed to allow controlled sample irradiation under an
anaerobic environment and an amperometric nitric oxide probe could be incorporated to
allow kinetic measurements of nitric oxide formation.
6.5 References
(1) Cordes, M.; Giese, B. Chem. Soc. Rev. 2009, 38, 892.(2) Purves, W. K.; Sadava, D.; Orians, G. H.; Heller, C. H. Life: The Science of
Biology; 7th ed.; Sinauer Associates and W. H. Freeman: U.S.A., 2003.(3) (a) Winkler, J. R.; Nocera, D. G.; Yocom, K. M.; Bordignon, E.; Gray, H. B. J.
Am. Chem. Soc. 1982, 104, 5798. (b) Meade, T. J.; Gray, H. B.; Winkler, J. R. J. Am. Chem. Soc. 1989, 111, 4353. (c) Mayo, S.; Ellis, W.; Crutchley, R.; Gray, H. Science 1986, 233, 948. (d) Gray, H. B.; Winkler, J. R. Annu. Rev. Biochem.1996, 65, 537.
(4) (a) Pan, L. P.; Durham, B.; Wolinska, J.; Millett, F. Biochemistry 1988, 27,7180. (b) Geren, L.; Hahm, S.; Durham, B.; Millett, F. Biochemistry 1991, 30,

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-23
9450. (c) Millett, F.; Durham, B. Methods Enzymol. 2009, 456, 95. (d) Geren, L.; Durham, B.; Millett, F. Methods Enzymol. 2009, 456, 507.
(5) Yang, X.-J.; Drepper, F.; Wu, B.; Sun, W.-H.; Haehnel, W.; Janiak, C. Dalton Trans. 2005, 256.
(6) Hahm, S.; Durham, B.; Millett, F. Biochemistry 1992, 31, 3472.(7) Wang, K.; Mei, H.; Geren, L.; Miller, M. A.; Saunders, A.; Wang, X.; Waldner,
J. L.; Pielak, G. J.; Durham, B.; Millett, F. Biochemistry 1996, 35, 15107.(8) Gray, H. B.; Winkler, J. R. Q. Rev. Biophys. 2003, 36, 341.(9) Geren, L. M.; Beasley, J. R.; Fine, B. R.; Saunders, A. J.; Hibdon, S.; Pielak, G.
J.; Durham, B.; Millett, F. J. Biol. Chem. 1995, 270, 2466.(10) Pan, L. P.; Frame, M.; Durham, B.; Davis, D.; Millett, F. Biochemistry 1990, 29,
3231.(11) (a) Peterson, J. R.; Smith, T. A.; Thordarson, P. Chem. Commun. 2007, 1899. (b)
Peterson, J. R.; Smith, T. A.; Thordarson, P. Org. Biomol. Chem. 2010, 8, 151.(12) Sauvage, J. P.; Collin, J. P.; Chambron, J. C.; Guillerez, S.; Coudret, C.; Balzani,
V.; Barigelletti, F.; De Cola, L.; Flamigni, L. Chem. Rev. 1994, 94, 993.(13) Nilsson, T. Proc. Natl. Acad. Soc. U. S. A. 1992, 89, 6497.(14) Mandal, K.; Hoffman, M. Z. J. Phys. Chem. 1984, 88, 185.(15) Hamachi, I.; Matsugi, T.; Tanaka, S.; Shinkai, S. Bull. Chem. Soc. Jpn. 1996, 69,
1657.(16) Peterson, J. R.; Thordarson, P. Chiang Mai J. Sci. 2009, 26, 236.(17) Louie, G. V.; Brayer, G. D. J. Mol. Biol. 1990, 214, 527.(18) Marcus, R. A.; Sutin, N. Biochim. Biophys. Acta 1985, 811, 265.(19) Winkler, J. R.; Netzel, T. L.; Creutz, C.; Sutin, N. J. Am. Chem. Soc. 1987, 109,
2381.(20) Hvasanov, D.; Wiedenmann, J.; Braet, F.; Thordarson, P. Chem. Commun. 2011,
47, 6314.(21) Basu, S.; Azarova, N. A.; Font, M. D.; King, S. B.; Hogg, N.; Gladwin, M. T.;
Shiva, S.; Kim-Shapiro, D. B. J. Biol. Chem. 2008, 283, 32590.(22) Dejam, A.; Hunter, C. J.; Tremonti, C.; Pluta, R. M.; Hon, Y. Y.; Grimes, G.;
Partovi, K.; Pelletier, M. M.; Oldfield, E. H.; Cannon, R. O.; Schechter, A. N.; Gladwin, M. T. Circulation 2007, 116, 1821.
(23) Lauer, T.; Preik, M.; Rassaf, T.; Strauer, B. E.; Deussen, A.; Feelisch, M.; Kelm, M. Proc. Natl. Acad. Soc. U. S. A. 2001, 98, 12814.
(24) Cosby, K.; Partovi, K. S.; Crawford, J. H.; Patel, R. P.; Reiter, C. D.; Martyr, S.; Yang, B. K.; Waclawiw, M. A.; Zalos, G.; Xu, X.; Huang, K. T.; Shields, H.; Kim-Shapiro, D. B.; Schechter, A. N.; Cannon, R. O.; Gladwin, M. T. Nat. Med.2003, 9, 1498.
(25) (a) Webb, A.; Bond, R.; McLean, P.; Uppal, R.; Benjamin, N.; Ahluwalia, A. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 13683. (b) Gladwin, M. T. Nat. Chem. Biol. 2005, 1, 245. (c) Duranski, M. R.; Greer, J. J. M.; Dejam, A.; Jaganmohan, S.; Hogg, N.; Langston, W.; Patel, R. P.; Yet, S.-F.; Wang, X.; Kevil, C. G.; Gladwin, M. T.; Lefer, D. J. J. Clin. Invest. 2005, 115, 1232. (d) Jung, K.-H.; Chu, K.; Ko, S.-Y.; Lee, S.-T.; Sinn, D.-I.; Park, D.-K.; Kim, J.-M.; Song, E.-C.; Kim, M.; Roh, J.-K. Stroke 2006, 37, 2744.
(26) Averill, B. A. Chem. Rev. 1996, 96, 2951.(27) Clarke, T. A.; Mills, P. C.; Poock, S. R.; Butt, J. N.; Cheesman, M. R.; Cole, J.
A.; Hinton, J. C. D.; Hemmings, A. M.; Kemp, G.; Söderberg, C. A. G.; Spiro, S.; Van Wonderen, J.; Richardson, D. J. Methods Enzymol. 2008, 437, 63.
(28) Liu, Q.; Gross, S. S. Methods Enzymol. 1996, 268, 311.

Chapter 6 Photoinduced Electron Transfer Studies of Cytochrome c
6-24
(29) Shiva, S.; Huang, Z.; Grubina, R.; Sun, J.; Ringwood, L. A.; MacArthur, P. H.; Xu, X.; Murphy, E.; Darley-Usmar, V. M.; Gladwin, M. T. Circ. Res. 2007, 100,654.
(30) Li, H.; Samouilov, A.; Liu, X.; Zweier, J. L. J. Biol. Chem. 2004, 279, 16939.(31) Castello, P. R.; Woo, D. K.; Ball, K.; Wojcik, J.; Liu, L.; Poyton, R. O. Proc.
Natl. Acad. Soc. U. S. A. 2008, 105, 8203.(32) Cayman Chemical Company: Ann Arbor, MI, 2009.(33) Li, H.; Raman, C. S.; Glaser, C. B.; Blasko, E.; Young, T. A.; Parkinson, J. F.;
Whitlow, M.; Poulos, T. L. J. Biol. Chem. 1999, 274, 21276.(34) Tuominen, E. K. J.; Wallace, C. J. A.; Kinnunen, P. K. J. J. Biol. Chem. 2002,
277, 8822.(35) Kagan, V. E.; Tyurin, V. A.; Jiang, J.; Tyurina, Y. Y.; Ritov, V. B.; Amoscato,
A. A.; Osipov, A. N.; Belikova, N. A.; Kapralov, A. A.; Kini, V.; Vlasova, I. I.; Zhao, Q.; Zou, M.; Di, P.; Svistunenko, D. A.; Kurnikov, I. V.; Borisenko, G. G. Nat. Chem. Biol. 2005, 1, 223.
(36) Bushnell, G. W.; Louie, G. V.; Brayer, G. D. J. Mol. Biol. 1990, 214, 585.(37) Goldstein, D. C. PhD Thesis, The University of New South Wales, 2011.(38) Margoliash, E.; Frohwirt, N. Biochem. J. 1959, 71, 570.(39) Titheradge, M. A. In Methods in Molecular Biology: Nitric Oxide Protocols;
Titheradge, M. A., Ed.; Humana Press Inc.: Totowa, NJ, 1998; Vol. 100.

Chapter 7
Self-Assembled Light-Driven Proton Pumping Studies

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-2
7 Self-Assembled Light-Driven Proton Pumping Studies
In this Chapter, the construction of a self-assembled artificial hybrid
photosynthetic-respiratory electron transport chain for light-induced proton pumping is
described. The synthetic hybrid photosynthetic-respiratory chain is based on
ruthenium(II)-bistepyridine linked to cytochrome c (8-cyt c) coupled with the terminal
electron acceptor of the mitochondrial electron-transport chain, cytochrome c oxidase
(CcOx) and membrane encapsulated in diblock copolymer polystyrene140-b-poly(acrylic
acid)48 based on the work discussed in previous chapters of this Thesis.
Upon irradiation at an initial pH of 7.2, a proton pumping rate of 3.3 ×
103 H+/s across the polymer bilayer generating a gradient up to pH 0.2 was observed
by steady-state fluorescence spectroscopy studies using fluorescent pH dye, 8-hydroxy-
1,3,6-pyrenetrisulfonate (HPTS). Reconstituted cytochrome c oxidase exhibited a
greater than 50% native mitochondrial orientation in the polymersome membrane. This
chapter demonstrates that the reconstituted photoactive hybrid electron-proton chain
generates a proton gradient which can store chemical energy and is a step towards the
development of a model artificial protocell.
7.1 Background
A high order of organisation of assembly of discrete components to allow direct and
controlled electron transfer processes are critical for all functioning living organisms.1
Electron transfer across the photosynthetic electron transport chain is an ideal example
of this phenomenon which is involved in the production of adenosine triphosphate
(ATP) to drive biological processes.2
Photosynthesis in plants is one of the most important reactions in maintaining
life by providing the source of oxygen and biomaterial on Earth. It converts light energy

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-3
into stored chemical energy by driving the reaction between carbon dioxide and water to
produce sugar and the release of oxygen.3 In plants, photosynthesis occurs in the
organelle chloroplast, which comprises the electron transport chain in the thylakoid
membrane. Upon absorption of light energy (photons), charge separation events are
initiated by reaction centres allowing rapid electron transfer through cofactor chains
which stabilises the charge separation and prevents recombination. Light induced
charge separation generates a strong oxidant (water oxidising complex) in photosystem
II (PSII) which extracts electrons from water producing oxygen and subsequent electron
transfer to photosystem I (PSI) generates a strong reducing agent (oxidised chlorophyll
P700) catalysing nicotinamide adenine dinucleotide phosphate (NADP+) reduction as
shown in Figure 7.1.4 The electron flow between PSII and PSI through the electron
transport chain is coupled to proton translocation in a redox loop mechanism to produce
a proton gradient which generates ATP by ATP synthase.2,4
Figure 7.1. Light-induced photosynthetic electron transport chain catalysing oxidation of water generating oxygen and the subsequent proton gradient produces adenosine triphosphate (ATP) to drive biological processes.3

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-4
The respiratory electron transport chain of the mitochondria is analogous to
photosynthesis where electrons are translocated through protein complexes that span
cell membranes consisting of chains of organic and inorganic cofactors that act as
stepping stones for electron transfer, which is coupled to proton translocation. The
proton gradient is used to drive ATP production by ATP synthase.5
In the process of respiration, the proton gradient is contributed by the terminal
oxidase of the respiratory chain of mitochondria, cytochrome c oxidase (complex IV,
EC 1.9.3.1). Since the discovery by Wikström in 1977 that cytochrome c oxidase redox
activity is coupled to membrane translocation of protons, it has become one of the most
well characterised and studied integral membrane proteins.6 Cytochrome c oxidase
catalyses the reduction of oxygen to form water consuming four electrons (from
cytochrome c) and protons while concomitantly translocating four protons across a
membrane as shown in Figure 7.2.7 It is a 200 kDa Y-shaped multisubunit enzyme
where the two arms of the Y span the inner membrane and the stalk extends into the
cytoplasmic side allowing binding to cytochrome c.8 In this project, work is primarily
focused on cytochrome c oxidase, however, it should be noted that complex I (NADH
dehydrogenase) and complex III (cytochrome bc1 complex) also contribute to the
generation of a proton gradient across the inner mitochondrial membrane during
respiration.2

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-5
Figure 7.2. Solid ribbon representation of bovine heart cytochrome c oxidase (complex IV, EC 1.9.3.1). The structure was derived from the protein data bank file ‘2OCC’.9
Natural electron transport systems (photosynthetic or respiratory) are highly
sophisticated and require control of components in space, energy and time while being
organised in a supramolecular system.1 Moreover, the storage of chemical energy in a
compartmentalised space is also one of the key requisites for any model artificial
protocell; the other key attributes being the ability to self-replicate the vesicle
compartment and genetic information.10 Scientists have attempted to develop artificial
photosynthetic systems to produce chemical energy as an electrochemical proton
gradient across a membrane in a confined space using self-assembly.
One approach to construct a photosynthetic hybrid is to use a light-harvesting
photosensitiser linked to a donor and acceptor capable of generating a redox potential
gradient across a membrane. This gradient can then translocate protons across a

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-6
phospholipid membrane vesicle (liposome) using a quinone relay as first reported by
Steinberg-Yfach et al.11 Additionally, Bhosale et al.12 have developed an elegant mimic
using aryl diimides which undergo - stacking, and upon photoexcitation achieves
directional electron transfer in liposomes coupled with proton transfer due to quinone
reduction which acts as an electron acceptor. In both these cases, the proton gradient is
generated passively by the virtue of quinone-redox chemistry that either shuttles protons
into or depletes protons from the interior of the vesicle. Finally, another approach
reported to produce a photosynthetic mimic was to encapsulate an active light-driven
transmembrane proton pump, bacteriorhodopsin in a polymer vesicle (polymersome) to
generate a proton gradient.13 Here, the proton pumping action is based on the natural
ability of bacteriorhodopsin to unidirectionally gate protons through an ion channel
upon light-induced transient conformational changes in the protein chromophore.
Attempts have been made by Meier et al.14 as well as Rowan and van Hest et
al.15 to form proteo-polymersomes by reconstitution of channel proteins or catalytic
enzymes to form a catalytic cascade pathway, respectively. However, the reconstitution
of enzymes in polymersome membranes to form a semi-synthetic electron transport
chain has never been attempted.
7.2 Photosynthetic-Respiratory Hybrid System
The construction of a chloroplast mimic that links membrane-bound photoinduced
electron transfer across an enzyme cascade with concomitant proton translocation to
generate an electrochemical potential ( ) based on synthetic polymersomes is
described. In order to develop the chloroplast hybrid using a synthetic polymer
membrane, ruthenium(II)-bisterpyridine complex 8 was selected as the light-harvesting
component and linked to iso-1 cytochrome c (8-cyt c) which functions as an electron

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-7
transport redox protein as shown in Figure 7.3. Ru(II) 8-cyt c was prepared by
covalently linking the synthetic maleimide functionalised ruthenium(II)-bisterpyridine
complex 8 with CYS102 of cytochrome c via Michael addition as discussed in Chapter
3.16
Figure 7.3. Bioconjugate 8-cyt c used as the light-harvesting electron transport redox protein.
To complete the photosynthesis-electron transport chain hybrid, mitochondrial
cytochrome c oxidase was selected as the natural acceptor which mediates electron
transfer across the enzyme cascade and simultaneously vectorially translocates protons
upon reduction.17 The primitive chloroplast is a modified synthetic hybrid of the
electron transport chain and is capable of converting photon energy into a by direct
electron transport from the light-harvesting centre of the hybrid 8-cyt c to the active
proton pump cytochrome c oxidase as shown in Figure 7.4.

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-8
Figure 7.4. Schematic representation of light-harvesting enzyme cascade generating a proton gradient upon irradiation. Encapsulation of 8-cyt c within hydrophilic poly(acrylic acid) region of diblock copolymer membrane PS140-b-PAA48 coupled electrostatically with transmembrane cytochrome c oxidase in the hydrophobic polystyrene block. Irradiation in the presence of sacrificial electron donor ethylenediaminetetraacetic acid (EDTA) leads to generation of pH which is detected by fluorescent pH dye, 8-hydroxy-1,3,6-pyrenetrisulfonate (HPTS). Proton translocation from the interior negative (N-) side to the positive (P-) side illustrated.
The resulting semi-synthetic enzyme cascade was membrane encapsulated in
polymersomes based on the polystyrene140-b-poly(acrylic acid)48 (PS140-b-PAA48)
diblock copolymer. Polymeric vesicles were prepared in this study using the ‘kinetic
trapping’ method previously discussed in Chapter 5 by injecting a tetrahydrofuran
solution of dissolved PS140-b-PAA48 into a solution of bioconjugate 8-cyt c and
cytochrome c oxidase. It was found that this preparation method is highly robust against
changes in salt concentration and pH and that the predominant factor controlling

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-9
formation of polymersomes based on PS140-b-PAA48 is the addition of positively
charged proteins which induce vesicle formation (Chapter 5).18 This achieves
concomitant encapsulation of both enzymes within the membrane. Additionally, Ru(II)
8-cyt c and cytochrome c oxidase were selectively encapsulated in separate membrane
domains, with the positively charged 8-cyt c entrapped in the hydrophilic poly(acrylic
acid) block and lipophilic cytochrome c oxidase in the hydrophobic polystyrene block.
7.2.1 Polymersome morphologies and membrane reconstitution
To eliminate the possibility that the induced pH gradient was a result of morphological
changes of polyelectrolyte PS140-b-PAA48 during irradiation, transmission electron
microscopy studies (TEM) were performed on enzyme cascade (8-cyt c:CcOx)
polymersome membrane reconstituted samples upon irradiation with 465 nm light over
56 min. TEM studies demonstrated that there were no significant changes in
polymersome morphologies and diameters are identical within experimental
uncertainty, indicating that the pH gradient is a result of proton translocation (inducing
a pH gradient at an initial pH of 7.2) and excluding the possibility that the pH gradient
is a result of changing morphologies of polymer aggregates as shown in Figure 7.5a-d.
These polymersome morphologies have been previously confirmed by cryo-TEM in
Chapter 518 and vesicles are defined as morphologies displaying a distinct contrast
within individual particles indicating the presence of bilayer and cavity formation.

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-10
Figure 7.5. Structural characterisation of hybrid proteo-polymersomes (8-cyt c:CcOx:HPTS) by transmission electron microscopy. (a) Low magnification electron micrograph of proteo-polymersomes at time 0 min and initial pH 7.2. Scale bar: 400 nm. (b) Electron micrograph of proteo-polymersomes at time 0 min and initial pH 7.2. (367±185 nm). Scale bar: 200 nm. (c) Electron micrograph after irradiation for 24 min using 465 nm light. (273±83 nm). Scale bar: 200 nm. (d) Electron micrograph after irradiation for 56 min using 465 nm light to a final light induced pH gradient of
pH 7.4. (291±74 nm). Scale bar: 200 nm. All sample measurements were made in a 5 mM sodium dihydrogen phosphate, 5 mM ethylenediaminetetraacetic acid buffer at an initial pH 7.2. Errors are standard deviation.
In order to confirm reconstitution of cytochrome c oxidase within the hydrophobic
membrane of PS140-b-PAA48, confocal laser scanning microscopy was performed on
reconstituted fluorescently labelled cytochrome c oxidase excited using a 488 nm laser
as shown in Figure 7.6a-b.

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-11
Figure 7.6. Structural characterisation of hybrid proteo-polymersomes (8-cyt c:CcOx:HPTS) by confocal laser scanning microscopy of fluorescently labelled cytochrome c oxidase encapsulated in PS140-b-PAA48 polymersome membrane.(a) Transmission light micrograph of fluorescently labelled cytochrome c oxidaseencapsulated proteo-polymersomes. (b) Confocal laser scanning micrograph excited at 488 nm (contrast enhanced). Scale bars: 10 m and inset 1 m. All sample measurements were made in a 5 mM sodium dihydrogen phosphate, 5 mM ethylenediaminetetraacetic acid buffer at an initial pH 7.2.
Polymersomes containing cytochrome c:cytochrome c oxidase-Dye could be visualised
optically via fluorescence due to aggregate structures with diameters up to 1 m as
shown in Figure 7.7. This indicates that cytochrome c oxidase is successfully
reconstituted in the polymer membrane rather than remaining as a homogenous mixture
in the bulk or internal polymersome aqueous phase due to the localised fluorescence in
the polymersome aggregates.18

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-12
Figure 7.7. Representative histogram of PS140-b-PAA48 polymersome diameters for 7.5 M cytochrome c:0.75 M cytochrome c oxidase in PBS at 25 oC.
7.2.2 Photoinduced pH gradient
Water soluble fluorescent pH dye, 8-hydroxy-1,3,6-pyrenetrisulfonate11-13,19 (HPTS,
Figure 7.8a) was entrapped in the internal volume of polymersomes to monitor pH
correlating to proton translocation from the negative (N-) to the postive (P-) side of the
membrane. Figure 7.8b shows the pH dependent changes in the fluorescence properties.

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-13
Figure 7.8. pH dependent photophysical properties of 8-hydroxy-1,3,6-pyrenetrisulfonate (HPTS). (a) Chemical structure of HPTS. (b) pH titration curve of HPTS fluorescence intensity emission ( ex = 460 nm, em = 510and encapsulated pyranine ( ) in PS140-b-PAA48 in 20 mM sodium dihydrogen phosphate. Bulk pKa = 7.56 and encapsulated pKa = 7.62. Inset: excitation spectra ( em = 510 nm) of internally encapsulated HPTS in dialysed polymersomes and incubated at pH 5.1 (black solid line) and pH 9.2 (black dashed line). Error bars are standard deviation.
Internal pH changes in hybrid proteo-polymersomes were determined based on the
fluorescence intensity of enclosed HPTS as well as the ratiometric ratio of the two
peaks 405 and 460 nm as shown in Figure 7.9. Irradiation with a 465 nm light source

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-14
results in initiation of electron transport across the 8-cyt c:CcOx electron transport
chain, leading to an overall proton translocation in the presence of sacrificial electron
donor ethylenediaminetetraacetic acid (EDTA).
Figure 7.9. Light-induced transmembrane proton translocation in hybrid proteo-polymersomes. Fluorescence excitation (left, em 510 nm) and emission spectra (right, ex = 460 nm; em = 510 nm) of encapsulated HPTS after irradiation at an initial pH 6.5. The polymersomes were irradiated for 0 ( ), 12 ( ), 24 ( ), 36 ( )and 56 ( ) min with 20.0±2.3 mW/cm2 of 465 nm light. Measurement was made in a 5 mM sodium dihydrogen phosphate, 5 mM ethylenediaminetetraacetic acid buffer, pH 6.5.
Determination of internal pH could be achieved by monitoring fluorescence intensity
( ex = 460 nm; em = 510 nm) and calibrating to a pH titration curve (Figure 7.8b).19
The proteo-polymersome ensemble (Figure 7.4) shows an increase in fluorescence
intensity after irradiation with actinic light at 465 nm over a 1 h period. An increase in
fluorescence intensity corresponds to basification of the internal aqueous compartment

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-15
indicating overall vectorial proton translocation from the N- to P-side. Proton pumping
rate at an initial pH of 7.2 was estimated to be 3.3±1.0 × 103 H+/s (see Appendix F).
Figure 7.10. Light-induced transmembrane proton translocation in hybrid proteo-polymersomes. Relative fluorescence intensity of internal HPTS as a function of irradiation time at an initial pH of 7.2. 8-cyt c:CcOx:polymersome
[H+] = 1.6 × 10-8 M 8-cyt c:CcOx bulk [H+] = 3.8 × 10-9 M ( ), 8-cyt c:polymersome [H+] = 6.8 × 10-9 M ( ), internal EDTA [H+] = 4.3 × 10-9 M( ), cyt c:CcOx:polymersome [H+] = 4.4 × 10-10 M ( ), dark control
[H+] = 1.3 × 10-9 M ( ), polymer micelle [H+] = -1.8 × 10-9 M ( ) and poly-L-lysine induced:polymersome [H+] = -1.1 × 10-9 M (×). Error bars indicate standard deviation. All sample measurements were made in a 5 mM sodium dihydrogen phosphate, 5 mM ethylenediaminetetraacetic acid buffer at a desired pH.
Additionally, in order to exclude artefacts, the excitation spectra as shown in Figure 7.9
displays a ratiometric change of the two maxima peaks at 405 and 460 nm ensuring that
the fluorescence intensity increase is a result of pH change. Based on Figure 7.11, it can
be observed that there is a decrease in the ratiometric intensity of the two peaks
indicating basification of the interior at an initial pH of 6.5 in which the proton pumping

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-16
activity is greater (see Chapter 7.2.4). This correlates to the fluorescence intensity
increase in the emission spectra indicating basification of the interior compartment.
Interestingly, the excitation spectra showed unusual behaviour with concomitant
increase in the excitation peaks as shown in Figure 7.9. It is proposed that this
behaviour is attributed to scattering caused by EDTA by-products which is formed by
photoreduction of oxygen with the Ru(II)-bisterpyridine complex (see Appendix E). It
was found that due to scattering and the small overall pH changes in the hybrid
proteo-polymersome system, the fluorescence intensity emission method was more
reliable and sensitive to pH changes and the proton pumping rates were determined
using this approach which has been previously reported for monitoring internal pH
changes in cells.19-20
Figure 7.11. Intensity ratio of the two peaks (405 and 460 nm) of the excitation spectra of internal HPTS after irradiation at an initial pH 6.5. Error bars are standard deviation. Measurements were made in a 5 mM sodium dihydrogen phosphate, 5 mMethylenediaminetetraacetic acid buffer, pH 6.5.

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-17
To demonstrate that internal pH was a result of proton translocation within the
polymersome, pumping studies consisting of the ensemble components in a bulk
solution were performed without encapsulation. Fluorescence studies indicated a minor
increase in internal pH. However, it can be seen that the change in intensity is
significantly smaller than the encapsulated 8-cyt c:CcOx:polymersome light-harvesting
hybrid. It is proposed that this background change in internal pH may be due to the
electron transfer between sacrificial electron donor EDTA and 8-cyt c producing
reaction intermediates consuming internal protons. Another possibility may be a result
of light-induced or structurally enzyme modified loss of peroxidase-like activity which
leads to radical generation and propagation mechanisms.16,21 Additionally, this
background pH change is similarly observed for the 8-cyt c:polymersome control (i.e.,
no CcOx). A pH of 0.2 units can be observed for the 8-cyt c:CcOx:polymersome
construct, whereas, the control samples generally exhibit a pH <0.07 units.
The block copolymer membranes of the polyelectrolyte system PS140-b-PAA48
are considerably thicker (98±35 nm) than phospholipid bilayers ( 5 nm).22 This
corresponds to a bilayer of PS140-b-PAA48 in the fully extended state. Cytochrome c
oxidase (CcOx) spans 12 nm9 and can be reconstituted to span the polymer membrane
due to the high flexibility and the conformational freedom of the polymer molecules,
allowing the block copolymer to adapt to the dimensions or conformation of the
transmembrane protein without loss of free energy while remaining functional. This
effect has been reported by Meier et al.14a for reconstitution of small channel proteins
(bacterial porin OmpF) within a triblock copolymer. In addition, van Hest et al.23 have
shown that in mixed solvent mixtures of water and tetrahydrofuran, where
tetrahydrofuran is a good solvent and plasticiser of polystyrene (PS), the flexibility of
hydrophobic block (PS) is preserved despite the high glass transition temperature (Tg).

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-18
This property of PS allows the rearrangement of the diblock copolymer to conform to
the dimensions of CcOx as the self-assembly process of polymersomes occurs in the
presence of the transmembrane protein in a co-solvent mixture of water and
tetrahydrofuran over at least 12 h. This is supported by previous work as discussed in
Chapter 5 which showed that PS140-b-PAA48 polymersomes could be induced from
micelle aggregates after addition cytochrome c within 30 min in a THF:PBS (1:6 v/v)
co-solvent mixture at room temperature.18
The above shows that the measured fluorescence intensity changes of
encapsulated HPTS were light-induced effects. Both the dark incubation control and
cyt c:CcOx:polymersome which lacks the light-harvesting chromophore as shown in
Figure 7.10 resulted in neglible intensity change. It should be noted that EDTA
by-products are highly effective at reducing cytochrome c as discussed in Chapter 6. To
minimise background effects caused by EDTA and its by-products, EDTA solution was
incubated in a dark environment for at least 24 h and the hybrid proteo-polymersome
system was incubated in the dark for 1 h prior to dark control measurements. Moreover,
the dark control sample was measured only at time 0 min and 56 min as the excitation
wavelength for HPTS fluorescence can cause photoexcitation of the
Ru(II)-bisterpyridine complex.
Additionally, to demonstrate that the polyelectrolyte PS-b-PAA does not
contribute to the basification of the interior compartment, a polymer only control
(micelle) and a biologically inert positively charged polypeptide (poly-L-lysine) control
was used to induce polymersome formation encapsulating HPTS were chosen to
eliminate the possibility that poly(acrylic acid) was acting as a proton transporter upon
irradiation. Interestingly, over the period of illumination, slight acidification of the
interior was observed in contrast to basification exhibited by the enzyme ensemble

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-19
system. This slight acidification in the absence of the proton pumping assembly may be
attributed to the weak acid nature of poly(acrylic acid).24 This shows that our system
undergoes light-induced vectorial proton translocation.
7.2.3 Orientation of reconstituted cytochrome c oxidase
Determination of the average orientation of reconstituted cytochrome c oxidase can be
deduced based on the relative increase or decrease of HPTS emission intensity. HPTS
undergoes an increase in fluorescence luminescence from acidic to a basic environment.
Fluorescence measurements as shown in Figure 7.9 and Figure 7.11 indicate an increase
in HPTS emission after irradiation with actinic light indicating basification of the
interior volume of the polymersome. This indicates favoured orientation of the average
reconstituted cytochrome c oxidase in PS140-b-PAA48 in the native mitochondrial
configuration (Figure 7.4).2,5 It should be noted that the orientation of the reconstituted
cytochrome c oxidase indicates a bias in the average orientation (>50% in the native
mitochondrial configuration) and not exclusively uniform orientation in the
cytochrome c oxidase population. This indicates that proton pumping from the exterior
into the interior compartment (acidification) can occur simultaneously and the
fluorescence measurements indicate an average proton translocation from the N- to P-
side (basification).
The bias of the average orientation (>50% native orientation) during
reconstitution may be due to the polar region of cytochrome c oxidase on the native
cytoplasmic side of the mitochondrial membrane being significantly larger than the
polar region on the matrix side.25 This may cause the orientation to be biased during the
self-assembly process, whereby the larger radii of curvature of the outer polymersome
(PAA block) accommodates this polar surface. The observation of favoured native

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-20
oriented cytochrome c oxidase was further confirmed based on the internally entrapped
ethylenediaminetetraacetic acid (EDTA) control as shown in Figure 7.10 (no addition of
EDTA to external bulk phase) indicating a reduced pH.
7.2.4 Dependency of proton translocation rates on pH
It was found that the proton pumping rates are sensitive to pH environment as shown in
Figure 7.12. Polymersomes were equilibrated in a phosphate buffer (5 mM) at fixed pH
such that the internal (pHin) and external pH (pHout) were equal. Under an acidified
polymersome environment at pH 6.5, a proton pumping rate of 1.3±0.4 × 104 H+/s was
observed. In contrast, a basified environment at pH 7.9 showed significant inhibition of
proton translocation rate to 8.3±2.5 × 102 H+/s. These results demonstrate that even a
slight change of pH from physiological pH 7.2 to acidic pH 6.5 can lead to a 4-fold
enhancement. This is despite the apparent higher proton “back-pressure” in the more
acidic environment (pH 6.5).26 The rate dependency on pH environment is attributed to
the intrinsic pH-dependent kinetics of cytochrome c oxidase. These results are
consistent with previously reported findings showing that the proton pumping rate is
dependent on pHin which determines the kinetics of internal electron transfer within
cytochrome c oxidase linking proton translocation.27

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-21
Figure 7.12. Dependency of proton translocation rates on pH. Proton pumping rates under equilibrated physiological (pH 7.2), acidic (pH 6.5) and basic (pH 7.9) conditions. Error bars indicate standard deviation. All sample measurements were made in a 5 mM sodium dihydrogen phosphate, 5 mM ethylenediaminetetraacetic acid buffer at the desired pH.
7.2.5 Proton pumping quantum efficiencies ( )
To interpret changes in fluorescence intensity in terms of quantum yield ( ) for proton
translocation, the rate of photon emission of the actinic light source was determined to
be 1.40±0.02 × 1016 photons/s at a distance of 2.5 cm between sample and light source.
In the context of these studies, quantum yield is expressed as the ratio of proton
translocation vs. absorbed photons. The number of photons absorbed could be
determined by correcting for the optical density of the Ru(II) complex antenna (see
Appendix F). A for the synthetic photosynthetic-respiratory hybrid system was
estimated to be 1.1±0.3 × 10-9% at pH 7.2. This poor may be a result of poor

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-22
electronic communication between Ru(II) 8-cyt c and cytochrome c oxidase in the
membrane system as the rate limiting step.
Polymersomes offer the advantage of greater stability for weeks to months
compared to liposomes of hours to days due to lower chain mobilities which ‘freeze’ the
structure.28 However, in this hybrid system, this proves to be disadvantageous by
simultaneously reducing the rates of diffusion of both reconstituted cytochrome c
oxidase and 8-cyt c. This reduces the probability of 8-cyt c:CcOx binding. The rate of
replenishing electrons to cytochrome c oxidase across the electron transport chain is
therefore dramatically reduced and dependent on the rate of reduction of 8-cyt c at the
single enzyme level. These findings were also supported as the of intramolecular
electron transfer for 8-cyt c was 1.1±0.3 × 10-3% as discussed in Chapter 6 in the
polymersome membrane environment which is 1 × 106 fold more efficient than proton
pumping . Consequently, the apparent turnover number of cytochrome c oxidase
proton translocation is also significantly inhibited and determined to be
6.2±1.9 × 10-11 H+/CcOx/s as the rate limiting step is directly correlated to the for
proton pumping. In comparison, the turnover number of cytochrome c oxidase in rat
liver mitochondria is 160 H+/CcOx/s.29
7.3 Conclusion and Future Work
In this Chapter, a synthetic hybrid photosynthetic-respiratory system that generates a
proton potential upon photoexcitation has been constructed via self-assembly.
Reconstitution of the hybrid enzyme cascade consisting of bioconjugate 8-cyt c,
cytochrome c oxidase in the polymersome membrane of polyelectrolyte PS140-b-PAA48
was characterised by transmission electron microscopy and confocal laser scanning
microscopy. Irradiation with actinic light converts photon energy into an

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-23
electrochemical potential via electron transfer. Charge transfer occurs across an
artificial electron transport chain within a polymersome membrane coupled with overall
vectorial proton translocation and resulting pH changes were monitored by steady-state
fluorescence spectroscopy using internally entrapped HPTS. From these investigations,
it was also found that cytochrome c oxidase appears to have the same orientation
(average >50% native orientation) in the polymersome membrane encountered in the
mitochondria. This biased orientation is attributed to the alignment of cytochrome c
oxidase with the curvature of the polymersomes. The photosynthetic-respiratory system
demonstrates the construction of an artificial hybrid organelle capable of storing
chemical energy in the form of a proton potential across a membrane with a proton
pumping rate of 3.3 × 103 H+/s at physiological pH 7.2 ( pH 0.2 units) correlating
to a of 1.1±0.3 × 10-9%. Proton translocation rates were investigated using
pH-dependent studies showing that a four-fold increase in proton pumping rate is
observed under acidic conditions of pH 6.5. This synthetic photoactive electron
transport chain capable of storing chemical energy as a proton gradient brings vesicle
and polymersome chemists towards satisfying one of the three requirements10a for an
artificial model protocell, which include (1) catalyst (storing of chemical energy), (2)
self-reproducing vesicles10b and (3) self-reproducing informational substance (genetic
information).10b
For future work, analogous Ir(III)-bisterpyridine based bioconjugates can be used
in contrast to Ru(II) bioconjugates to improve proton pumping rates and due to their
improved photoreduction abilities. Additionally, it is possible to utilise this system and
harness the potential to drive biomimetic processes such as generation of ATP11b,13 and
ATP-linked processes by potentially reconstituting transmembrane ATP synthase.
Alternatively, reagents can be encapsulated within the interior cavity of the

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-24
proteo-polymersome to induce pH sensitive reactions which can be activated upon
photoinduced pH changes.
7.4 References
(1) Sauvage, J. P.; Collin, J. P.; Chambron, J. C.; Guillerez, S.; Coudret, C.; Balzani, V.; Barigelletti, F.; De Cola, L.; Flamigni, L. Chem. Rev. 1994, 94, 993.
(2) Cordes, M.; Giese, B. Chem. Soc. Rev. 2009, 38, 892.(3) Purves, W. K.; Sadava, D.; Orians, G. H.; Heller, C. H. Life: The Science of
Biology; 7th ed.; Sinauer Associates and W. H. Freeman: U.S.A., 2003.(4) Nugent, J. H. A. Eur. J. Biochem. 1996, 237, 519.(5) Rich, P. R. Biochem. Soc. Trans. 2003, 31, 1095.(6) Wikstrom, M. K. F. Nature 1977, 266, 271.(7) (a) Bloch, D.; Belevich, I.; Jasaitis, A.; Ribacka, C.; Puustinen, A.; Verkhovsky,
M. I.; Wikström, M. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 529. (b) Jasaitis, A.; Verkhovsky, M. I.; Morgan, J. E.; Verkhovskaya, M. L.; Wikström, M. Biochemistry 1999, 38, 2697.
(8) (a) Deatherage, J. F.; Henderson, R.; Capaldi, R. A. J. Mol. Biol. 1982, 158, 487. (b) Love, B.; Chan, S. H. P.; Stotz, E. J. Biol. Chem. 1970, 245, 6664.
(9) Yoshikawa, S.; Shinzawa-Itoh, K.; Nakashima, R.; Yaono, R.; Yamashita, E.; Inoue, N.; Yao, M.; Fei, M. J.; Libeu, C. P.; Mizushima, T.; Yamaguchi, H.; Tomizaki, T.; Tsukihara, T. Science 1998, 280, 1723.
(10) (a) Szostak, J. W.; Bartel, D. P.; Luisi, P. L. Nature 2001, 409, 387. (b) Kurihara, K.; Tamura, M.; Shohda, K.-i.; Toyota, T.; Suzuki, K.; Sugawara, T. Nat. Chem. 2011, 3, 775.
(11) (a) Steinberg-Yfrach, G.; Liddell, P. A.; Hung, S.-C.; Moore, A. L.; Gust, D.; Moore, T. A. Nature 1997, 385, 239. (b) Steinberg-Yfrach, G.; Rigaud, J.-L.; Durantini, E. N.; Moore, A. L.; Gust, D.; Moore, T. A. Nature 1998, 392, 479.
(12) Bhosale, S.; Sisson, A. L.; Talukdar, P.; Fürstenberg, A.; Banerji, N.; Vauthey, E.; Bollot, G.; Mareda, J.; Röger, C.; Würthner, F.; Sakai, N.; Matile, S. Science2006, 313, 84.
(13) Choi, H.-J.; Montemagno, C. D. Nano Lett. 2005, 5, 2538.(14) (a) Nardin, C.; Thoeni, S.; Widmer, J.; Winterhalter, M.; Meier, W. Chem.
Commun. 2000, 1433. (b) Palivan, C. G.; Fischer-Onaca, O.; Delcea, M.; Itel, F.; Meier, W. Chem. Soc. Rev. 2012, 41, 2800.
(15) Vriezema, D. M.; Garcia, P. M. L.; Sancho Oltra, N.; Hatzakis, N. S.; Kuiper, S. M.; Nolte, R. J. M.; Rowan, A. E.; van Hest, J. C. M. Angew. Chem. Int. Ed.2007, 46, 7378.
(16) Peterson, J. R.; Smith, T. A.; Thordarson, P. Chem. Commun. 2007, 1899.(17) Ferguson-Miller, S.; Babcock, G. T. Chem. Rev. 1996, 96, 2889.(18) Hvasanov, D.; Wiedenmann, J.; Braet, F.; Thordarson, P. Chem. Commun. 2011,
47, 6314.(19) Damiano, E.; Bassilana, M.; Rigaud, J. L.; Leblanc, G. FEBS Lett. 1984, 166,
120.(20) Agostiano, A.; Mavelli, F.; Milano, F.; Giotta, L.; Trotta, M.; Nagy, L.; Maroti,
P. Bioelectrochemistry 2004, 63, 125.

Chapter 7 Self-Assembled Light-Driven Proton Pumping Studies
7-25
(21) (a) Valderrama, B.; García-Arellano, H.; Giansanti, S.; Baratto, M. C.; Pogni, R.; Vazquez-Duhalt, R. FASEB J. 2006, 20, 1233. (b) Michael J, D. Biochem. Biophys. Res. Commun. 2003, 305, 761.
(22) (a) Walde, P.; Ichikawa, S. Biomol. Eng. 2001, 18, 143. (b) Mui, B.; Chow, L.; Hope, M. J. Methods Enzymol. 2003, 367, 3.
(23) Meeuwissen, S. A.; Kim, K. T.; Chen, Y.; Pochan, D. J.; van Hest, J. C. M. Angew. Chem. Int. Ed. 2011, 50, 7070.
(24) Choi, J.; Rubner, M. F. Macromolecules 2005, 38, 116.(25) Madden, T. D.; Hope, M. J.; Cullis, P. R. Biochemistry 1984, 23, 1413.(26) Westerhoff, H. V.; Scholte, B. J.; Hellingwerf, K. J. Biochim. Biophys. Acta,
Bioenerg. 1979, 547, 544.(27) Faxén, K.; Brzezinski, P. Biochim. Biophys. Acta, Bioenerg. 2007, 1767, 381.(28) Discher, D. E.; Eisenberg, A. Science 2002, 297, 967.(29) Hochli, L.; Hackenbrock, C. R. Biochemistry 1978, 17, 3712.

Chapter 8
Experimental

Chapter 8 Experimental
8-2
8 Experimental
8.1 Chemicals, Equipment and General Methods
8.1.1 Chemicals, solvents and materials
Chemicals were purchased from Sigma Aldrich with the exceptions of ammonium
acetate, sodium dihydrogen phosphate, sodium hydroxide, sodium bicarbonate,
anhydrous sodium sulphate, citric acid, iodine, pyridine and hydrazine monohydrate
were purchased from Ajax Finechem Pty. Ltd., 4-nitrobenzaldehyde (Hopkins and
Williams Ltd.) and ammonium hexafluorophosphate (Acros Organics), ruthenium(III)
trichloride hydrate (Precious Metals Online), Oregon Green® 488 carboxylic acid,
succinimidyl ester *5-isomer* and Oregon Green® 488 maleimide (Invitrogen).
Bis(4’-(4-hydroxymethylphenyl)-2,2’:6’:2”-terpyridine)iridium(III)tris(hexafluorophosp
hate) was kindly provided by Dr. Daniel C. Goldstein, N-propargyl maleimide by Dr.
Joshua R. Peterson and 4,4’-bipyridinium-N,N-di(maleimidopropyl) hexaflurophosphate
provided by Mr. Dithepon Pornsaksit. Silica was purchased from Davisil (40-63 μM).
Thin layer chromatography plates (Kieselgel 60 F-254 pre-coated sheets 0.25 mm) were
purchased from Merck. Neutral alumina oxide was purchased from Merck (Alumina
Oxide 90 active neutral, 20 – 230 mesh). Dichloromethane (CH2Cl2) and methanol
(CH3OH) were distilled before use. Dry solvents such as acetonitrile (CH3CN),
dichloromethane, diethyl ether (Et2O) and tetrahydrofuran (THF) were obtained from a
Pure Solv dry solvent system (Innovative Technology, Inc. model #PS-MD-7). Dry
methanol was distilled and stored over calcium chloride. Dry N,N-dimethylformamide
(DMF) was purchased from Sigma Aldrich and used directly from the bottle. Deuterated
solvents for NMR were obtained from Cambridge Isotope Laboratories. For
aggregation studies and preparation of all salt buffers, ultra pure water (R > 18 × 106 )
was used. Water and organic solvent was filtered through a 0.45 m cellulose

Chapter 8 Experimental
8-3
membrane filter (Minisart RC 25, Sartonius Stedim Biotech) prior to polymer
aggregation studies. Diblock copolymer, polystyrene140-b-poly(acrylic acid)48 (PDI =
1.10), was purchased from Encapson (The Netherlands, catalogue number 1036).
Chloroform was distilled and passed through neutral alumina oxide prior to lipid
dissolution. Lipids, extrusion kit (Avanti® Mini-Extruder) and accessories were
purchased from Avanti Polar Lipids. All other chemicals were used as received.
Confocal fluorescence laser-scanning microscopy petri dishes (FluoroDish) were
purchased from World Precision Instruments.
8.1.2 Enzymes and phosphate buffers
Yeast cytochrome c from Saccharomyces cerevisiae (catalogue number C2436), bovine
serum albumin (BSA, catalogue number A-0281), equine heart cytochrome c (catalogue
number C2506), equine skeletal muscle myoglobin (catalogue number M0630), bovine
testes calmodulin (catalogue number P1431) and poly-L-lysine hydrobromide (1000-
5000 Da, catalogue number P0879) were purchased from Sigma Aldrich. Cytochrome c
oxidase was purchased from NBS Biologicals (catalogue number 11197-B). Yeast iso-1
cytochrome c was purified following previously published procedures prior to
bioconjugation reactions.1 Acropora millepora green fluorescent protein (amGFP) was
expressed and purified as previously described and was provided by Dr. Jörg
Wiedenmann (University of Southampton, UK) and Dr. Chris Marquis (University of
New South Wales, Australia).2 For the preparation of salt buffers, ultrapure water (R >
18 × 106 , MilliQ Ultrapure Water System, Millipore) was used. Phosphate buffers
were prepared using concentrated stock solutions of sodium dihydrogen phosphate (1 M,
pH ~4), sodium chloride (5 M), ethylenediaminetetraacetic acid (EDTA, 200 mM) and
diluted with ultrapure water as necessary. Buffer pH values were adjusted with aqueous

Chapter 8 Experimental
8-4
sodium hydroxide (1 M) or hydrochloric acid (1 M) using a Scholar 425 pH meter
(Corning) and filtered using a 0.45 m (Millipore, 47 mm regenerated cellulose) prior to
use. Bioconjugation reactions were prepared as above except a stock solution of sodium
dihydrogen phosphate (97 mM), EDTA (97 mM), pH 7.0 was used to eliminate the need
for adjusting pH of small volume solutions.
8.1.3 Spectroscopy and melting points
A Varian Cary 50 Bio UV-Vis or Cary 5 UV-Vis-NIR spectrometer was used for UV-
Vis spectra measurements. Fluorescence spectra were recorded using a Varian Cary
Eclipse spectrometer with excitation and emission slits at 5 nm, excitation filter “auto”,
emission filter “open” and PMT voltage set to “medium”, unless otherwise stated. NMR
spectra (1H and 13C) were recorded in the designated solvents using a Bruker Avance
DPX (300 MHz) spectrophotometer. Multiplicities are assigned as singlet (s), doublet
(d), triplet (t), quartet (q), pentet (p), multiplet (m) and denoted as broad (br) where
appropriate. Chemical shifts are measured in parts per million (ppm), internally
referenced relative to tetramethylsilane (SiMe4, 1H and 13C = 0 ppm) or residual solvent
peaks (CD3CN: 1H = 1.94 and 13C = 1.32; DMSO-d6: 1H = 2.50, 13C = 39.52; CDCl3:
1H = 7.26 ppm, 13C = 77.16 ppm). IR spectra were recorded on a Shimadzu FTIR-
8400S, ThermoNicolet Avatar model 370 FT-IR or a Perkin Elmer Spotlight 400 FT-IR
spectrometer equipped with a microscope and attenuated total reflectance (ATR)
accessories with diamond crystal inset. Intensity abbreviations are as follows: weak (w),
medium (m), strong (s). Low resolution Electrospray Ionisation (ESI) mass spectra were
recorded on a Waters Micromass ZQ electrospray instrument or on a Shimadzu
Prominence UFLC system equipped with a Shimazdu 2010 EV LC-MS detector and a
Shimazdu FRC-10A fraction collector. High resolution ESI mass spectrometry was

Chapter 8 Experimental
8-5
performed on a Thermo Linear Quadropole Ion Trap Fourier Transform Ion Cyclotron
Resonance (LQT FT Ultra) mass spectrometer in electrospray mode with a 7 T
superconducting magnet or a Thermo Scientific LTQ Orbitrap XL hybrid Fourier
Transform Mass Spectrometer (FTMS) in static nanospray mode. MALDI-TOF mass
spectra were recorded on an Applied Biosystems Voyager DE STR MALDI reflectron
TOF MS (protein and bioconjugate measurements were made in linear mode). Melting
points were recorded on a Mel-temp II hot stage apparatus.
8.1.4 Protein and bioconjugate purification equipment
Protein and bioconjugate purification was performed using a GE Healthcare
ÄKTApurifier. Cation exchange chromatography (CEX) was performed using a strong
cation exchange column (TSKgel SP-5PW, Supelco). Immobilised metal affinity
chromatography (IMAC) was performed using either a Ni2+ charged HisTrap HP (GE
Healthcare, 1 mL) or an Acrosep Hypercel (Pall, 1 mL) column. Size exclusion
chromatography (SEC) was performed using either a Superdex 75 10/300 GL (GE
Healthcare) or Sephadex G25 desalting (Superfine, GE Healthcare) column.
Programmed elution gradients are expressed in column volumes (CV) to ease
comparison between columns of different size. Flow rates were based on manufacturer
recommendations for flow and pressure limits and were typically between 0.5 to
1 mL/min. Fractions were collected in 1 mL Eppendorf tubes using either a Frac 901 or
Frac 902 collector (GE Healthcare). Protein solutions were concentrated using 3 000
molecular weight cut-off (MWCO) centrifuge concentrators (Amicon Ultra-15, Amicon
Ultra-4 or Amicon Ultra-0.5, Millipore) and centrifugation at 4 000 rpm in a Sigma 2-6
for large volume samples (<50 mL) or 13 800 rpm in a Sigma 1-14 bench top centrifuge
(John Morris Scientific) for small volume samples (<1.5 mL). Samples were dialysed

Chapter 8 Experimental
8-6
into MilliQ water using Slide-A-Lyzer Mini Dialysis Units (3 500 MWCO, Pierce) or
Tube-O-Dialyzer Dialysis Units (4 000 MWCO, G Biosciences).
8.1.5 Preparation of samples for analysis by MALDI-TOF mass spectrometry
Protein and bioconjugate samples for MALDI were prepared by diluting 5 L of protein
with either 5 L of a saturated solution of sinapic acid and -cyano-4-hydroxycinnamic
acid (10:1 w/w ratio in acetonitrile/water/trifluoroacetic acid (70:30:0.03, v/v/v)) or a
solution of caffeic acid (10 mg/mL in acetonitrile/water/trifluoroacetic acid (80:20:0.1,
v/v/v)). Prepared samples were spotted by adding a 0.5 L droplet (× 2) onto a MALDI
target plate. At times, ZipTips (C4, 0.6 L bed volume, Millipore) were used for
desalting by priming the tip with acetonitrile/water/trifluoroacetic acid (80:20:0.1, v/v/v,
3 × 10 L), washing with acetonitrile/water/trifluoroacetic acid (2:98:0.1, v/v/v,
3 × 10 L), loading the column (5 × 10 L), washing with
acetonitrile/water/trifluoroacetic acid (2:98:0.1, v/v/v, 3 × 10 L) and eluting with
2.5 L of a caffeic acid solution (10 mg/mL in acetonitrile/water/trifluoroacetic acid
(80:20:0.1, v/v/v)) directly onto the MALDI target plate.
8.1.6 Transmission electron microscopy (TEM) studies
TEM micrographs were recorded on a JEOL 1400 (80 kV for polymersomes and
100 kV for liposomes) instrument and cryo-TEM micrographs were recorded on a
JEOL 2100 (200 kV) instrument. Conventional TEM samples were prepared by placing
20 L of sample onto a formvar-coated copper grid and the excess water was blotted
away after 2 min with a filter paper. For statistical analysis, a population of 100 micelles
or polymersomes were measured from TEM micrographs for determination of the
average and standard deviation of diameters. For analysis of polymersome-to-micelle

Chapter 8 Experimental
8-7
(p/m) ratio, a population of 100 aggregates from TEM micrographs were selected and
aggregate morphologies displaying distinct contrast within individual particles
(indicating the presence of bilayer and cavity formation) were classified as vesicles
whereas micelles were defined as particles showing uniform contrast. Ratio
measurements were repeated in triplicate. For cryo-TEM, 3 L of sample was directly
placed onto glow-discharged holey carbon grids (Quantifoil, Germany). Grids were
blotted once at a blotting angle of 2 mm for 2 s under 100% relative humidity at 25 oC
and subsequently plunged into liquid ethane using the automated vitrobot (F.E.I, The
Netherlands). Vitrified samples were stored in liquid nitrogen upon cryo transfer for
cryo-TEM investigation.3
8.1.7 Confocal laser-scanning microscopy studies
Optical and confocal laser microscopy experiments were carried out with an Olympus
Fluoview FV1000, fitted with monochromatic laser light sources for fluorescence
measurements. Images were acquired on a confocal laser scanning microscope with a
40x 0.9NA water-immersion objective. Excitation was at 488 nm using an Ar+ laser.
Detection was in the range 500-600 nm using internal PMTs (gains were set to 468 V).
The confocal pinhole (aperture) was set to auto and images with a 640×640 pixel
resolution were recorded at a scan rate of 40 s/pixel with a total acquisition time of
16 s.
8.1.8 amGFP (amFP497) (Acropora millepora) expression and isolation
The following procedure was performed in the laboratory of Dr. Jörg Wiedenmann at
the University of Southampton. The coding sequence of the green fluorescent protein
(GFP)-like protein from the reef coral Acropora millepora (amFP497)4 was introduced

Chapter 8 Experimental
8-8
in the plasmid pQE32 (Qiagen, Hilden, Germany), resulting in the addition of an N-
terminal 6×histidine tag to the recombinant protein. Bacteria (Escherichia coli, M15
pREP4) were transformed with the plasmid and grown at 37 ºC in 2YT medium to an
optical density of 0.6. Expression of the protein was induced with isopropyl- -D-
thiogalactopyranosid (IPTG) and the culture was incubated at 20 ºC on a shaker at
220 rpm for 12 h. Cultures were slightly agitated further for 7 days at 4 ºC to increase
the yield of soluble protein.5 Subsequently, cells were harvested and the recombinant
protein was purified by immobilised metal ion chromatography using TalonTM matrix
(Clontech; Palo Alto, USA), following the protocol of the manufacturer. Yields were
determined by UV-Vis spectroscopy ( 476 = 31.42 mM-1cm-1).
8.1.9 Gel electrophoresis
Gel electrophoresis was performed using Invitrogen Novex® NuPage® 12% Bis-Tris,
1 mm, 10-well gels, SeeBlue® Plus2 molecular weight marker, NuPage® LDS sample
buffer (4×), NuPage® sample reducing agent (10×), NuPage® MES SDS running
buffer, SimplyBlueTM safestain and the gels run using a Zoom Dual Power supply
(model ZP10002, Invitrogen). Samples for gel electrophoresis were prepared by dilution
in Novex® NuPage® LDS sample buffer (Invitrogen). Samples were reduced (to
eliminate disulfide dimers) by adding NuPage® sample reducing agent (active
ingredient dithiothreitol (DTT)). Samples were heated at 70 oC for 10 min to denature
the protein. Novex® NuPage® gels (12% Bis-Tris, 10-wells) were then loaded with
1-3 g of protein per well, run at a constant 200 V for 40 min and stained according to
the procedure included with SimplyBlueTM safestain.

Chapter 8 Experimental
8-9
8.2 X-ray Crystallography
8.2.1 Crystal growth of Ru(II)-bisterpyridine complexes
In general, Ru(II)-bisterpyridine complex (ca. 5 mg) was dissolved in minimal solvent
(DMF) resulting in a concentrated solution. Crystals were grown by slow diffusion of
anhydrous diethyl ether into a solution of complex in N,N-dimethylformamide at room
temperature or 4 oC for complex 7 and 8, respectively. A suitable single crystal was
selected under a polarising microscope (Leica M165Z) for single crystal X-ray
diffraction analysis.
8.2.2 X-ray structure determination
8.2.2.1 Complex 7
The X-ray diffraction measurement for complex 7 was performed on a Bruker kappa
APEX-II CCD diffractometer at 150 K by using graphite-monochromated Mo-K
radiation ( = 0.71075 Å). The crystal was mounted on the goniometer using cryo loops
for intensity measurements, coated with paraffin oil and immediately transferred to the
cold stream using Oxford Cryostream 700 system attachment. Upon obtaining an initial
refinement of unit cell parameters, the data collection strategy was calculated to achieve
a redundancy of at least 4 throughout the resolution range ( - 0.80 Å) at 10 s exposure
time per frame utilising the kappa offsets on the four circle goniometer geometry. Data
integration, reduction with multi-scan absorption correction method was carried out
using Bruker APEX2 Suite software.6 The structure was solved by Direct Methods
program SHELXS-97 and refined by full-matrix least-squares refinement program
SHELXL.7 All non-hydrogen atoms were refined anisotropically and hydrogen atoms
were included by using a riding model. Further crystal and refinement data given in
Appendix A.

Chapter 8 Experimental
8-10
8.2.2.2 Complex 8
The X-ray diffraction measurement for complex 8 was carried out at MX1 beamline at
the Australian Synchrotron Facility, Melbourne. The crystal was mounted on the
goniometer using cryo loop for intensity measurements, coated with paraffin oil and
immediately transferred to the cold stream using a Cryostream attachment. Data was
collected using Si<111> monochromated synchrotron X- = 0.71023 Å)
at 100(2) K and was corrected for Lorentz and polarization effects using the XDS
software.8 The structure was solved by Direct methods and the full-matrix least-squares
refinements was carried out using SHELXL.7 Further crystal and refinement data given
in Appendix A.
8.3 Synthesis of Terpyridine Chromophores
8.3.1 4-nitro-2’-azachalcone (1)9
To a solution of 4-nitrobenzaldehyde (2.08 g, 13.8 mmol) and aqueous sodium
hydroxide (1 M, 13.75 mL) in methanol (60 mL) was added 2-acetylpyridine (1.60 mL,
14.3 mmol). The solution was stirred for 1 h, filtered, and the collected precipitate
washed with cold methanol, dissolved in dichloromethane, washed with water, dried
over anhydrous sodium sulphate and the solvent removed in vacuo. The resulting
yellow solid was recrystallised from ethanol to give azachalcone 1 as yellow crystals
(1.01 g, 29%). 1H NMR (300 MHz, CDCl3) 8.76 (d, J = 4.9 Hz, 1H), 8.43 (d,
J = 16.2 Hz, 1H), 8.28 (d, J = 9.0 Hz, 2H), 8.21 (d, J = 7.9 Hz, 1H), 7.96 – 7.84 (m,
4H), 7.56 – 7.51 (m, 1H). MS (ESI) m/z: ([M + H]+) calcd. for C14H11N2O3, 255.08;
found, 254.96. These results are in agreement with those reported in the literature.9

Chapter 8 Experimental
8-11
8.3.2 1-(2-oxo-2-(2-pyridyl)ethyl)pyridinium iodide (2)10
To a stirred solution of iodine (7.78 g, 30.7 mmol) in dry pyridine (20 mL) under
nitrogen at 60 oC was added 2-acetylpyridine (3.68 g, 30.4 mmol). The resulting
mixture was stirred at 100 oC for 2 h, cooled, then the crystals filtered and washed with
chloroform and diethyl ether to yield pyridinium iodide 2 as black crystals (7.84 g,
79%). 1H NMR (300 MHz, DMSO-d6) 9.00 (d, J = 5.6 Hz, 2H), 8.87 (d, J = 6.0 Hz,
1H), 8.73 (t, J = 7.2 Hz, 1H), 8.27 (t, J = 7.2 Hz, 2H), 8.18 – 8.03 (m, 2H), 7.83 (td,
J = 5.84, 1.5 Hz, 1H), 6.50 (s, 2H). MS (ESI) m/z: ([M - I]+) calcd. for C12H11N2O,
199.08; found, 199.09. These results are in agreement with those reported in the
literature.10
8.3.3 4’-(4-nitrophenyl)-2,2’:6’,2’’-terpyridine (3)9
A solution of azachalcone 1 (0.870 g, 3.42 mmol), pyridinium iodide 2 (1.13 g,
3.47 mmol) and ammonium acetate (3.75 g, 48.6 mmol) in dry methanol (60 mL) was
refluxed for 20 h. The crystals were cooled, filtered and washed with cold methanol
(6 × 50 mL) to yield terpyridine 3 as a purple solid (0.91 g, 75%). 1H NMR (300 MHz,
CDCl3) 8.76 (s, 2H), 8.74 (d, J = 4.9 Hz, 2H), 8.69 (d, J = 7.9 Hz, 2H), 8.38 (d,
J = 8.7 Hz, 2H), 8.06 (d, J = 8.7 Hz, 2H), 7.91 (td, J = 7.7, 1.9 Hz, 2H), 7.42 – 7.35 (m,
2H). MS (ESI) m/z: ([M + H]+) calcd. for C21H15N4O2, 355.12; found, 354.95. These
results are in agreement with those reported in the literature.9
8.3.4 4’-(4-aminophenyl)-2,2’:6’,2’’-terpyridine (4)11
A solution of compound 3 (891.8 mg, 2.52 mmol) in absolute ethanol (60 mL) was
refluxed for 45 min in the presence of 10% Pd/charcoal catalyst (120.6 mg). Hydrazine
monohydrate (4.75 mL, 98.0 mmol) in absolute ethanol (60 mL) was then added drop-

Chapter 8 Experimental
8-12
wise to the reaction mixture and refluxed for 3 h. The solution was filtered over celite
and washed with dichloromethane (150 mL). The organic phase was washed with water
(4 × 100 mL), dried over anhydrous sodium sulphate and solvent removed in vacuo to
yield terpyridine 4 as yellow crystals (658.4 mg, 81%). 1H NMR (300 MHz, CDCl3)
8.73 (d, J = 4.5 Hz, 2H), 8.69 (s, 2H), 8.66 (d, J = 7.9 Hz, 2H), 7.87 (td, J = 7.7, 1.9 Hz,
2H), 7.79 (d, J = 8.7 Hz, 2H), 7.34 (t, J = 6.2 Hz, 2H), 6.80 (d, J = 8.7 Hz, 2H), 5.61 (s,
2H). MS (ESI) m/z: ([M + H]+) calcd. for C21H17N4, 325.14; found, 325.04. These
results are in agreement with those reported in the literature.11
8.3.5 [Ru(tpy)]Cl3 (5)12
A solution of terpyridine (418.9 mg, 1.795 mmol), and ruthenium(III) trichloride hydrate
(722.0 mg, 3.481 mmol) in absolute ethanol (50 mL) was refluxed for 23 h under
nitrogen. The product was collected by filtration and washed with absolute ethanol,
water and diethyl ether yielding [Ru(tpy)]Cl3 5 as a black solid (695 mg, 88%). MS
(ESI) m/z: ([M – 2Cl]+) calcd. for C15H11N3RuCl+, 369.97, found, 369.93. These results
are in agreement with those reported in the literature.12
8.3.6 [Ru(tpy)2](PF6)2 (6)12b
A solution of 2,2’;6’,2”-terpyridine (113.8 mg, 0.488 mmol) and ruthenium(III)
trichloride hydrate (49.6 mg, 0.239 mmol) in ethylene glycol (33 mL) was heated at
110 oC for 21 h. The solution was then diluted with water (150 mL), filtered through
celite and the product was precipitated with ammonium hexafluorophosphate. The solid
was collected by centrifugation, washed with water and recrystallised with
acetonitrile/diethyl ether and collected by filtration and washed with acetonitrile
yielding [Ru(tpy)2](PF6)2 6 as a red solid (103.1 mg, 50%). 1H NMR (300 MHz,

Chapter 8 Experimental
8-13
CD3CN) 8.73 (d, J = 8.1 Hz, 4H), 8.48 (dt, J = 8.1, 1.0 Hz, 4H), 8.40 (t, J = 8.3 Hz,
2H), 7.91 (td, J = 7.8, 1.5 Hz, 4H), 7.32 (dq, J = 5.6, 0.8 Hz, 4H), 7.19 – 7.11 (m, 4H).
MS (ESI) m/z: ([M – 2PF6]2+) calcd. for C30H22N6Ru2+, 284.05; found, 283.80. These
results are in agreement with those reported in the literature.12b
8.3.7 [Ru(tpy)(4’-(4-aminophenyl)-2,2’:6’,2”-tpy)](PF6)2 (7)
Method 1
A solution of terpyridine 4 (273.3 mg, 0.8425 mmol) and [Ru(tpy)]Cl3 5 (373.6 mg,
0.8447 mmol) in ethylene glycol (50 mL) was heated to 110 oC for 20 h under nitrogen.
Reaction mixture was diluted to 150 mL with water and filtered over celite. Filtrate was
precipitated using ammonium hexafluorophosphate, washed with water (3 × 50 mL) and
collected with acetonitrile. Purified over silica using a gradient from 90:9:1
CH3CN:H2O:KNO3 (saturated) to 20:3:1 CH3CN:H2O:KNO3 (saturated). Fractions
precipitated with ammonium hexafluorophosphate and washed with water. Product
further purified over alumina (neutral) using a gradient from acetonitrile to 90:9:1
CH3CN:H2O:KNO3 (saturated). Fractions pooled, precipitated with ammonium
hexafluorophosphate, washed with water (3 × 20 mL) and collected with acetonitrile
yielding complex 7 as a red solid (609.6 mg, 76%). mp >274 oC (decomposed); 1H
NMR (300 MHz, CD3CN) 8.90 (s, 2H), 8.72 (d, J = 7.9 Hz, 2H), 8.60 (d, J = 7.9 Hz,
2H), 8.48 (d, J = 7.9 Hz, 2H), 8.38 (t, J = 8.3 Hz, 1H), 8.00 (dt, J = 8.6, 2.4 Hz, 2H),
7.91 (tt, J = 7.9, 1.3 Hz, 4H), 7.44 (dd, J = 4.9, 0.8 Hz, 2H), 7.30 (dd, J = 4.5, 0.8 Hz,
2H), 7.21 – 7.09 (m, 4H), 6.95 (dt, J = 8.7, 2.4 Hz, 2H); 13C NMR (75 MHz, CD3CN)
159.1, 156.5, 156.0, 153.5, 153.3, 144.9, 138.9, 136.4, 129.8, 128.4, 128.2, 125.3,
124.6, 120.9, 115.8; IR (ATR) max/cm-1 3646 (w), 3593 (w), 3506 (w), 3414 (w), 3121
(w), 1990 (w), 1633 (m), 1597 (m), 1529 (w), 1430 (m); UV-Vis (CH3CN) max/nm

Chapter 8 Experimental
8-14
( /M-1cm-1) 490 (1.71 × 104), 364 (9.80 × 103), 308 (5.00 × 104), 283 (2.55 × 104), 272
(2.86 × 104), 231 (2.67 × 104); HRMS (ESI) m/z: ([M – PF6)]+) calcd. for
C36H27N7P1F6Ru+, 804.1023; found, 804.1006. MS (ESI) m/z: ([M – 2PF6]2+) calcd.
for C36H27N7Ru2+, 329.57; found, 329.49.
Method 2
A solution of terpyridine 4 (179.0 mg, 0.5518 mmol) and [Ru(tpy)]Cl3 5 (248.8 mg,
0.5645 mmol) in ethanol (100 mL) was refluxed for 20 h under nitrogen. Reaction
mixture was filtered over celite, concentrated in vacuo and diluted with water (100 mL).
Filtrate was precipitated using ammonium hexafluorophosphate, washed with water
(3 × 50 mL) and collected with acetonitrile. The product was recrystallised with
acetonitrile/diethyl ether yielding complex 7 as a red solid (138.7 mg, 28%).
Characterisation data was identical to the compound obtained from method 1 above.
8.3.8 [Ru(tpy)(maleimide-hexylcarboxamido-phenyl-tpy)](PF6)2 (8)
A solution of 6-maleimidocaproic acid (53.2 mg, 0.252 mmol), O-(7-azabenzotriazole-
1-yl)-1,1,3,3-tetramethyluronium hexafluorophosphate (HATU, 95.9 mg, 0.252 mmol)
and N,N-diisopropylethylamine (66.8 mg, 0.517 mmol) in dry dimethylformamide
(5 mL) was stirred at room temperature under nitrogen for 1 h. Subsequently, complex 7
(71.7 mg, 0.0756 mmol) in dry dimethylformamide (5 mL) was added to the mixture
and stirred for a further 26 h in the dark at room temperature under nitrogen.
Dichloromethane (50 mL) was added to the solution and the organic phase washed with
aqueous citric acid (10% w/v, 2 × 20 mL), water (3 × 10 mL) and dried over anhydrous
sodium sulphate. Dichloromethane was removed in vacuo and the concentrated red
dimethylformamide phase containing product was precipitated from dry diethyl ether

Chapter 8 Experimental
8-15
and the solid collected by filtration, washed with diethyl ether and collected with
acetonitrile. Product was purified over silica using a gradient from acetonitrile to
70:29:1 CH3CN:H2O:KNO3 (saturated). Fractions pooled, precipitated with ammonium
hexafluorophosphate, washed with water (3 × 20 mL) and collected with acetonitrile
yielding complex 8 as a red solid (27.9 mg, 41%). mp >246 oC (decomposed); 1H NMR
(300 MHz, CD3CN) 8.98 (s, 2H), 8.75 (d, J = 8.3 Hz, 3H), 8.63 (d, J = 7.9 Hz, 2H),
8.50 (d, J = 7.9 Hz, 2H), 8.40 (t, J = 8.3 Hz, 1H), 8.16 (dd, J = 8.7, 1.5 Hz, 2H), 8.00 –
7.85 (m, 6H), 7.43 (dd, J = 5.6, 0.8 Hz, 2H), 7.37 (dd, J = 6.4, 0.8 Hz, 2H), 7.20 – 7.10
(m, 3H), 6.75 (s, 2H), 3.49 (t, J = 7.0 Hz, 2H), 2.41 (t, J = 7.4 Hz, 2H), 1.80 – 1.56 (m,
5H), 1.45 – 1.33 (m, 2H); 13C NMR (75 MHz, CD3CN) 173.60, 172.51, 159.49,
159.41, 159.26, 156.56, 153.61, 153.48, 149.09, 142.43, 139.19, 136.87, 135.37,
132.53, 129.51, 128.61, 128.56, 125.64, 125.56, 124.85, 122.17, 121.17, 38.44, 37.76,
29.17, 27.21, 25.94; IR (ATR) max/cm-1 3647 (w), 3403 (w), 3112 (w), 2933 (w), 2857
(w), 1769 (w), 1699 (s), 1595 (m), 1523 (m), 1449 (m), 1407 (m); UV-Vis (CH3CN)
max/nm ( /M-1cm-1) 485 (2.60 × 104), 410 (4.88 × 103), 308 (7.85 × 104), 282
(4.32 × 104), 272 (4.65 × 104); HRMS (ESI) m/z: ([M – PF6])+ calcd. for
C46H38N8O3P1F6Ru+, 997.1618; found, 997.1741 and ([M – 2PF6])2+ calcd. for
C46H38N8O3Ru2+, 426.0988; found, 426.1050. MS (ESI) m/z: ([M – 2PF6])2+, 426.11.
Prior to bioconjugation, complex 8 was exchanged with chloride salt to increase
solubility and yield.
8.3.9 [Ru(4’-(4-aminophenyl)-2,2’:6’2’’-terpyridine)2](PF6)2 (9)13
A solution of compound 4’-(4-aminophenyl)-2,2’:6’,2’’-terpyridine 4 (141.2 mg,
0.4353 mmol) and ruthenium(III) trichloride hydrate (44.3 mg, 0.214 mmol) in ethylene
glycol (50 mL) was stirred at 110 oC for 21 h. The reaction mixture was diluted with

Chapter 8 Experimental
8-16
water (150 mL) and filtered over celite. The product was precipitated using ammonium
hexafluorophosphate and collected by centrifugation (4 min, 4 000 rpm) and washed
with water (3 × 40 mL). The product was recrystallised with acetonitrile/diethyl ether to
yield terpyridine 9 as red crystals (116.3 mg, 52%). 1H NMR (300 MHz, CD3CN)
8.90 (s, 4H), 8.60 (d, J = 7.9 Hz, 4H), 8.00 (d, J = 8.7 Hz, 4H), 7.91 (td, J = 7.8, 1.5 Hz,
4H), 7.41 (d, J = 4.9 Hz, 4H), 7.20 – 7.09 (m, 4H), 6.95 (d, J = 8.7 Hz, 4H), 4.77 (s,
4H). MS (ESI) m/z: ([M – 2PF6]2+) calcd. for C42H32N8Ru2+, 375.09; found, 374.72.
These results are in agreement with those reported in the literature.13
8.3.10 [Ru(4’-(4-maleimide-hexylcarboxyamido-phenyl)-2,2’:6’2’’-
terpyridine)2](PF6)2 (10)11b
A solution of 6-maleimidocaproic acid (73.8 mg, 0.349 mmol), O-(7-azabenzotriazole-
1-yl)-1,1,3,3-tetramethyluronium hexafluorophosphate (HATU, 136.1 mg,
0.3580 mmol) and N,N-diisopropylethylamine (96.5 mg, 0.746 mmol) in dry
dimethylformamide (5 mL) was stirred at room temperature for 1 h. Complex [Ru(4’-
(4-aminophenyl)-2,2’:6’2’’-terpyridine)2](PF6)2 9 (62.5 mg, 0.0601 mmol) in dry
dimethylformamide (5 mL) was added to the reaction mixture and stirred for an
additional 25 h in the dark. Dichloromethane (50 mL) was added and the organic phase
was washed with 10% w/v citric acid (2 × 10 mL), water (3 × 10 mL), dried over
anhydrous sodium sulfate, filtered and dichloromethane removed in vacuo. The
concentrated dimethylformamide phase was diluted and precipitated from dry diethyl
ether and solid collected by filtration, washed with diethyl ether and collected with
acetonitrile. The dried solid was purified on silica (acetonitrile/saturated aqueous
potassium nitrate/water, 20:1:3 (v/v/v)) followed by concentration of the product
containing fractions, collection of the precipitate, washing with water and collection

Chapter 8 Experimental
8-17
with acetonitrile afforded the complex 10 as a red solid (22.8 mg, 27%). 1H NMR
(300 MHz, CD3CN) 8.98 (s, 4H), 8.63 (d, J = 8.3 Hz, 6H), 8.16 (d, J = 8.7 Hz, 4H),
7.93 (td, J = 7.9, 1.5 Hz, 8H), 7.42 (d, J = 5.7 Hz, 4H), 7.17 (td, J = 6.7, 1.5 Hz, 4H),
6.75 (s, 4H), 3.49 (t, J = 7.0 Hz, 4H), 2.41 (t, J = 7.4 Hz, 4H), 1.78 – 1.68 (m, 4H),
1.68 – 1.57 (m, 4H), 1.46 – 1.31 (m, 4H). MS (ESI) m/z: ([M – 2PF6]2+) calcd. for
C62H54N10O6Ru2+, 568.16; found, 568.14. These results are in agreement with those
previously reported.11b
Prior to bioconjugation, complex 10 was exchanged with chloride salt to increase
solubility and yield.
8.3.11 Anion exchange of Ru(II)-bisterpyridine complexes 8 and 10
Hexafluorophosphate salts of complexes 8 and 10 (10 mg) were dissolved in acetonitrile
to which excess tetrabutylammonium chloride was added until precipitation was
complete. The mixture was centrifuged to a pellet and solution was decanted. The pellet
was resuspended in acetonitrile and recrystallised from diethyl ether followed by
centrifugation (3 ×) to give the chloride adducts of complexes 8 and 10 which were
subsequently used without further purification or characterisation for bioconjugation
synthesis.
8.4 Synthesis of Anthraquinone-Based Acceptors
8.4.1 1-amino-3-azidopropane (11)14
Sodium azide (1.002 g, 15.42 mmol) was added slowly to a solution of
1-amino-3-bromopropane hydrobromide (840.8 mg, 3.840 mmol) in water (50 mL) and
refluxed under nitrogen for 23 h. The solution was allowed to cool to 0 oC, extracted
with diethyl ether (3 × 50 mL), dried with anhydrous sodium sulphate and filtered.

Chapter 8 Experimental
8-18
Solvent was removed in vacuo to yield a clear yellow oil (166.2 mg, 43%). 1H NMR
(300 MHz, CDCl3) 3.39 (t, J = 6.6 Hz, 2H), 2.83 (t, J = 7.5 Hz, 2H), 1.75 (p, J = 6.5
Hz, 2H), 1.42 (br s, 2H). MS (ESI) m/z: ([M + H]+) calcd. for C3H9N4+, 101.08; found,
100.80. These results are in agreement with those previously reported.14
8.4.2 Anthraquinone-2-carboxylic acid N-hydroxysuccinimide ester (12)
Anthraquinone-2-carboxylic acid (604.7 mg, 2.398 mmol) and N-hydroxysuccinimide
(275.8 mg, 2.396 mmol) was dissolved in dry dichloromethane (150 mL). The solution
was cooled to 0 oC and N,N’-dicyclohexylcarbodiimide (687.9 mg, 3.330 mmol) was
added. The reaction mixture was stirred at room temperature under nitrogen in the dark
for 23 h. The resulting cloudy yellow mixture was filtered and solvent removed in
vacuo yielding the NHS-ester of anthraquinone-2-carboxylic acid 12 as a yellow solid
(1.013 g, crude). The crude was used for subsequent reactions without further
purification.
8.4.3 Anthraquinone-2-azidopropylamide (13)
The NHS-ester of anthraquinone-2-carboxylic acid 12 (1.013 g, crude) was suspended
into a cloudy solution in acetone/ethanol (75 mL/50 mL). To this stirring solution, an
aqueous solution of sodium bicarbonate (223.0 mg, 2.655 mmol) and
1-amino-3-azidopropane 11 (166.2 mg, 1.660 mmol) in water (25 mL) was added and
allowed to stir for 23 h at room temperature in the dark under nitrogen. Organic phase
was removed in vacuo and dichloromethane (250 mL) was added, washed with basic
water (3 × 200 mL, pH 14), followed by acidic water (3 × 200 mL, pH 2). Organic
phase was dried with anhydrous sodium sulphate and purified on silica
(dichloromethane) followed by concentration of the product containing fractions

Chapter 8 Experimental
8-19
yielding the anthraquinone 13 as a yellow solid (196.6 mg, 35%). mp >160 oC
(decomposed); 1H NMR (300 MHz, CDCl3) 8.58 (d, J = 2.1 Hz, 1H), 8.44 – 8.25 (m,
4H), 7.89 – 7.81 (m, 2H), 6.61 (br s, 1H), 3.64 (q, J = 6.4 Hz, 2H), 3.50 (t, J = 6.5 Hz,
2H), 1.97 (p, J = 6.5 Hz, 2H); 13C NMR (75 MHz, CDCl3) 182.68, 182.57, 165.87,
139.43, 135.40, 134.67, 134.60, 133.58, 133.26, 128.19, 127.61, 124.93, 49.74, 38.35,
28.87; IR (KBr) max/cm-1 3321 (m), 3288 (s), 3073 (w), 3031 (w), 2928 (m), 2850 (m),
2192 (w), 2113 (s), 1677 (s), 1632 (s), 1590 (s), 1571 (m), 1544 (s), 1472 (m), 1463
(m), 1449 (w), 1436 (w); HRMS (ESI) m/z: ([M + H])+ calcd. for C18H15N4O3+,
335.1144; found, 335.1134 and ([M + Na])+ calcd. for C18H14N4O3Na+, 357.0964;
found, 357.0953. MS (ESI) m/z: ([M - H])- calcd. for C18H13N4O3-, 333.09, found,
332.90.
8.4.4 Anthraquinone-2-bromopropylamide (14)
The NHS-ester of anthraquinone-2-carboxylic acid 12 (691.5 mg, crude) was suspended
into a cloudy solution in acetone/ethanol (75 mL/50 mL). To this stirring solution, an
aqueous solution of sodium bicarbonate (281.6 mg, 3.35 mmol) and
3-bromopropylamine hydrobromide (373.6 mg, 1.707 mmol) in water (25 mL) was
added and allowed to stir for 18 h at room temperature in the dark under nitrogen.
Organic phase was removed in vacuo and dichloromethane (200 mL) was added,
washed with basic water (3 × 100 mL, pH 14), followed by acidic water (3 × 100 mL,
pH 2). Organic phase was dried with anhydrous sodium sulphate and purified on silica
(dichloromethane) followed by concentration of the product containing fractions
yielding the anthraquinone 14 as a yellow solid (344.5 mg, 56%). mp >178 oC
(decomposed); 1H NMR (300 MHz, CDCl3) 8.58 (d, J = 1.5 Hz, 1H), 8.44 – 8.27 (m,
4H), 7.89 – 7.81 (m, 2H), 6.58 (t, J = 7.5 Hz, 1H), 3.70 (q, J = 6.4 Hz, 2H), 3.53 (t,

Chapter 8 Experimental
8-20
J = 6.3 Hz, 2H), 2.27 (p, J = 6.5 Hz, 2H); 13C NMR (75 MHz, CDCl3) 182.65, 182.53,
166.00, 139.38, 135.37, 134.66, 133.55, 133.27, 128.14, 127.59, 124.99, 49.34, 39.20,
34.10, 32.19, 30.92, 25.75, 25.08; IR (KBr) max/cm-1 3318 (m), 3072 (w), 2929 (m),
2851 (w), 1669 (s), 1635 (s), 1591 (s), 1554 (m), 1447 (w), 1413 (w); UV-Vis (CH2Cl2)
max/nm ( /M-1cm-1) 328 (3.38 × 103), 258 (2.96 × 104); HRMS (ESI) m/z: ([M + H])+
calcd. for C18H15NO3Br+, 372.0235; found, 372.0226 and ([M + Na])+ calcd. for
C18H14NO3BrNa+, 394.0055; found, 394.0047. MS (ESI) m/z: ([M + H])+, 373.80.
8.4.5 Attempted synthesis of anthraquinone-2-propylamido-triazole-maleimide
(15)
Anthraquinone-2-azidopropylamide 13 (59.8 mg, 0.179 mmol) and N-propargyl
maleimide (26.8 mg, 0.198 mmol) was dissolved in tetrahydrofuran (10 mL). Copper(II)
sulphate pentahydrate (67.0 mg, 0.268 mmol) and L-ascorbic acid (65.4 mg,
0.371 mmol) in water (2 mL) was slowly added to the reaction mixture. Solution was
stirred in the dark under nitrogen at room temperature for 22.5 h. Organic phase was
removed in vacuo, resuspended in dichloromethane (100 mL), washed with water
(3 × 100 mL), dried with anhydrous sodium sulphate, filtered and solvent removed in
vacuo to yield a yellow oil (34.8 mg). 1H NMR spectroscopy and mass spectrometry
analysis of the crude product did not indicate that the desired compound 15 was present.
8.5 Synthesis and Purification of Bioconjugates
8.5.1 Purification of iso-1 cytochrome c
Crude yeast cytochrome c from Saccharomyces cerevisiae (11.3 mg) was dissolved in
phosphate buffer (5.65 mL, 20 mM, pH 7.0), reduced with dithiothreitol (DTT, 40 L of
1 M stock), loaded onto a Supelco strong cation exchange column (TSKgel SP-5PW,

Chapter 8 Experimental
8-21
10 m resin beads, 7.5 cmh × 0.75 cmd). The protein was eluted using a sodium
chloride gradient from 328 mM to 450 mM in 14.5 mL at pH 7.0 and 1 mL/min. The
main peak (eluting from 15.8 to 18 mL) was collected and concentrated using a
Millipore 3 000 molecular weight cut-off (MWCO) centrifuge concentrator giving pure
iso-1 cytochrome c in 54% yield based on UV-Vis absorbance of the final product
( 410 = 97.6 mM-1cm-1)1,11b. MS (MALDI) m/z: 12 705 ([M]+ requires 12 706).
8.5.2 8-cyt c
A solution of complex (chloride counter ion) 8 (0.900 mg, 0.975 mol) in acetonitrile
(613 L) was added to a solution of 94 mM sodium dihydrogen phosphate, 94 mM
ethylenediaminetetraacetic acid, pH 7.0 (3.19 mL) in water (10.9 mL) at room
temperature. Purified, reduced iso-1 cytochrome c (1.90 mg, 0.150 mol) in water
(351 L) was then added and the resulting solution stirred in darkness at room
temperature for 20 h. The reaction mixture was then concentrated, dialysed into water
and purified by immobilised metal affinity chromatography (IMAC, HisTrapTM HP, GE
Healthcare or Acrosep HypercelTM, Pall) using a gradient from 0 to 125 mM imidazole
in 20 mM sodium dihydrogen phosphate, 0.5 M sodium chloride, pH 7.0 from 7.7 to
19.2 mL at 0.5 mL/min. The product fraction (eluting from 11.1 to 18.9 mL) was
pooled, concentrated and dialysed into water to yield bioconjugate 8-cyt c. (9.33 nmol,
6%). MS (MALDI) m/z: 13 555 ([M – 2Cl]+ requires 13 559).
8.5.3 cyt c-10-BSA dimer
A solution of complex 10 (chloride counter ion) (0.0410 mg, 0.0340 mol) in
acetonitrile (170 L) was added to a solution of 94 mM sodium dihydrogen phosphate,
94 mM ethylenediaminetetraacetic acid, pH 7.0 (723 L) in water (1.87 mL) at room

Chapter 8 Experimental
8-22
temperature. Purified, reduced iso-1 cytochrome c (4.32 mg, 0.340 mol) in water
(636 L) was then added and the resulting solution stirred in darkness at room
temperature for 2 h. Bovine serum albumin (45.9 mg, 0.687 mol) was subsequently
added to the mixture and stirred in darkness at room temperature for an additional 21 h.
The reaction mixture was then concentrated, dialysed into water and purified by
immobilised metal affinity chromatography (IMAC, HisTrapTM HP, GE Healthcare)
using a gradient from 0 to 250 mM imidazole in 20 mM sodium dihydrogen phosphate,
0.5 M sodium chloride, pH 7.0 from in 9 mL at 0.5 mL/min. The product fraction
(eluting from 5.6 to 10.2 mL) was pooled, concentrated and dialysed into water. The
pooled fractions were further purified using a strong cation exchange column (CEX, SP-
5PW, Supelco) using a gradient from 320 to 450 mM sodium chloride in 20 mM sodium
dihydrogen phosphate, pH 7.0 in 14.4 mL at 1 mL/min. The product fraction (eluting
from 12.5 to 14 mL) was pooled, concentrated and dialysed into water to yield
bioconjugate cyt c-10-BSA (0.0340 mol, 30%). MS (MALDI) m/z: 80 609 ([M – 2Cl]+
requires 80 618).
8.5.4 cyt c-10-cyt c dimer
A solution of complex 10 (chloride counter ion) (0.0375 mg, 0.0311 mol) in
acetonitrile (156 L) was added to a solution of 94 mM sodium dihydrogen phosphate,
94 mM ethylenediaminetetraacetic acid, pH 7.0 (665 L) in water (805 L) at room
temperature. Purified, reduced iso-1 cytochrome c (3.97 mg, 0.313 mol) in water
(1.5 mL) was then added and the resulting solution stirred in darkness at room
temperature for 27 h. The reaction mixture was then concentrated, dialysed into water
and purified by immobilised metal affinity chromatography (IMAC, HisTrapTM HP, GE
Healthcare) using a gradient from 0 to 250 mM imidazole in 20 mM sodium dihydrogen

Chapter 8 Experimental
8-23
phosphate, 0.5 M sodium chloride, pH 7.0 in 9 mL at 0.5 mL/min. The product fraction
(eluting from 6 to 10 mL) was pooled, concentrated and dialysed into water to yield
bioconjugate cyt c-10-cyt c (1.90 nmol, 1%). MS (MALDI) m/z: 26 551 ([M – 2Cl]+
requires 26 550).
8.5.5 cyt c-16-GFP dimer
A solution of 4,4’-bipyridinium-N,N-di(maleimidopropyl) hexaflurophosphate 16
(0.78 mg, 1.08 mol) in acetonitrile (344 L) was added to a solution of 94 mM sodium
dihydrogen phosphate, 94 mM ethylenediaminetetraacetic acid, pH 7.0 (1.46 mL) in
water (4.55 mL). Acropora millepora green fluorescent protein (amFP497)4 (1.79 mg,
0.0688 mol) was added at room temperature in darkness and stirred for 3 h. The
reaction mixture was then concentrated to ca. 1 mL and desalted by size exclusion
chromatography (SEC, HiTrapTM desalting, GE Healthcare) using 20 mM sodium
dihydrogen phosphate, pH 7.0 at 0.5 mL/min. The product fraction (eluting from 1.5 to
3.1 mL) was pooled and concentrated (527 L). Purified, reduced iso-1 cytochrome c
(3.68 mg, 0.290 mol) in water (264 L) and a solution of 94 mM sodium dihydrogen
phosphate, 94 mM ethylenediaminetetraacetic acid, pH 7.0 (213 L) was then added to
the desalted product fraction and the resulting solution stirred in darkness at room
temperature for 17 h. The reaction mixture was then concentrated, dialysed into water
and purified by immobilised metal affinity chromatography (IMAC, HisTrapTM HP, GE
Healthcare) using a gradient from 0 to 250 mM imidazole in 20 mM sodium dihydrogen
phosphate, 0.5 M sodium chloride, pH 7.0 in 6 mL at 0.5 mL/min. The product fraction
(eluting from 5.5 to 8.2 mL) was pooled, concentrated and dialysed into water to yield
bioconjugate cyt c-16-GFP (0.49 nmol, 0.1%). MS (MALDI) m/z: 40 245 ([M – 2PF6]+
requires 40 236).

Chapter 8 Experimental
8-24
8.5.6 BSA-10-BSA dimer
A solution of complex 10 (chloride counter ion) (0.0410 mg, 0.0340 mol) in
acetonitrile (170 L) was added to a solution of 94 mM sodium dihydrogen phosphate,
94 mM ethylenediaminetetraacetic acid, pH 7.0 (723 L) in water (2.50 mL) at room
temperature. Bovine serum albumin (90.9 mg, 1.36 mol) was then added and the
resulting solution stirred in darkness at room temperature for 23 h. The reaction mixture
was then concentrated, dialysed into water and purified by immobilised metal affinity
chromatography (IMAC, HisTrapTM HP, GE Healthcare) using a gradient from 0 to
125 mM imidazole in 20 mM sodium dihydrogen phosphate, 0.5 M sodium chloride,
pH 7.0 in 6 mL at 0.5 mL/min. The product fraction (eluting from 5.7 to 8.1 mL) was
pooled, concentrated and dialysed into water to yield bioconjugate BSA-10-BSA
(0.43 nmol, 1%). Conjugate characterised by reduced SDS-PAGE (NuPage®, 12% Bis-
Tris Gel, Invitrogen) gel electrophoresis.
8.5.7 16-GFP
A solution of 4,4’-bipyridinium-N,N-di(maleimidopropyl) hexaflurophosphate 16
(0.590 mg, 0.817 mol) in acetonitrile (344 L) was added to a solution of 94 mM
sodium dihydrogen phosphate, 94 mM ethylenediaminetetraacetic acid, pH 7.0
(1.46 mL) in water (4.26 mL). Acropora millepora green fluorescent protein
(amFP497)4 (1.79 mg, 0.0688 mol) was added at room temperature in darkness and
stirred for 19 h. Excess 2-mercaptoethanol (14 M, 1 L) was added to the reaction
mixture to quench unreacted maleimide and allowed to stir for 1 h in the dark at room
temperature. The reaction mixture was then concentrated to ca. 1 mL and purified by
immobilised metal affinity chromatography (IMAC, HisTrapTM HP, GE Healthcare)
using an imidazole gradient from 0 to 500 mM in 20 mM sodium dihydrogen phosphate,

Chapter 8 Experimental
8-25
0.5 M sodium chloride, pH 7.0 in 16 mL at 0.5 mL/min. The product fraction (eluting
from 10.6 to 14.5 mL) was pooled, concentrated and dialysed into water to yield
bioconjugate 16-GFP (7.74 nmol, 11%). MS (MALDI) m/z: 27 246 ([M – 2PF6]+
requires 27 239).
8.5.8 12-GFP
A solution of anthraquinone 12 (0.567 mg, crude) in N,N-dimethylformamide (353 L)
was added to a solution of 200 mM sodium bicarbonate, pH 8.3 (1.87 mL) in water
(44 L). Acropora millepora green fluorescent protein (amFP497)4 (2.00 mg,
0.0746 mol) was added at room temperature in darkness and stirred for 16.5 h. The
reaction mixture was then concentrated to ca. 1 mL and desalted by size exclusion
chromatography (SEC, HiTrapTM desalting, GE Healthcare) using 20 mM sodium
dihydrogen phosphate, pH 7.0 at 0.5 mL/min. The product fraction (eluting from 1.7 to
3.45 mL) was pooled, concentrated and dialysed into water to yield bioconjugate
12-GFP (20.1 nmol, 27%). MS (MALDI) m/z: 27 037 ([M]+ requires 27 041, singly
labelled) and 27 282 ([M]+ requires 27 275, doubly labelled).
8.5.9 17-GFP
A solution of N-propargyl maleimide 17 (0.068 mg, 0.502 mol) in
N,N-dimethylformamide (251 L) was added to a solution of 94 mM sodium dihydrogen
phosphate, 94 mM ethylenediaminetetraacetic acid, pH 7.0 (1.07 mL) in water (203 L).
Acropora millepora green fluorescent protein (amFP497)4 (1.346 mg, 0.0502 mol) was
added at room temperature in darkness and stirred for 16.5 h. The reaction mixture was
then concentrated to ca. 1 mL and desalted by size exclusion chromatography (SEC,
HiTrapTM desalting, GE Healthcare) using 20 mM sodium dihydrogen phosphate, pH 7.0

Chapter 8 Experimental
8-26
at 0.5 mL/min. The product fraction (eluting from 1.55 to 2.55 mL) was pooled,
concentrated and dialysed into water to yield bioconjugate 17-GFP (24.9 nmol, 50%).
MS (MALDI) m/z: 26 967 ([M + Na]+ requires 26 965).
8.5.10 Attempted ‘click’ synthesis of 13-triazole-GFP
A solution of tetrakis(acetonitrile)copper(I) hexafluorophosphate (0.146 mg,
0.392 mol) and anthraquinone azide 13 (0.100 mg, 0.299 mol) in
N,N-dimethylformamide (98 L) was added to 17-GFP (0.671 mg, 0.0249 mol) in
100 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), pH 7.0 (393 L)
and water (925 L). Reaction mixture allowed to stir for 19 h in the dark with a nitrogen
overlay at 32 oC. Size exclusion chromatography and UV-Vis analysis did not show
evidence of formation of 13-triazole-GFP.
8.5.11 cyt c-Oregon Green 488
A solution of Oregon Green® 488 maleimide (0.46 mg, 0.99 mol) in
dimethylformamide (50 L) was mixed with iso-1 cytochrome c (1.3 mg, 1.0 mol) in
20 mM sodium dihydrogen phosphate, 20 mM ethylenediaminetetraacetic acid, 5%
dimethylformamide, pH 7.0 (1 mL). The mixture was stirred at room temperature for
21.5 h and concentrated. The conjugate was then purified in 20 mM sodium dihydrogen
phosphate buffer, pH 7.0 using size-exclusion chromatography (SEC, HiTrapTM
desalting, GE Healthcare), concentrated and dialysed extensively into water (molecular
weight cut-off 3 000 Da) yielding conjugate cyt c-Oregon Green 488 (13 nmol, 13%).
MS (MALDI) m/z 13 173 ([M + H]+ requires 13 169).

Chapter 8 Experimental
8-27
8.5.12 Cytochrome c Oxidase-Oregon Green 488
A solution of Oregon Green® 488 carboxylic acid, succinimidyl ester *5-isomer*
(0.12 mg, 0.236 mol) in N,N-dimethylformamide (50 L) was mixed with
cytochrome c oxidase (4.04 mg, 0.0202 mol) in 50 mM sodium bicarbonate, pH 8.3
(950 L). The mixture was stirred at room temperature for 160 min and concentrated.
The conjugate was then purified in 20 mM sodium dihydrogen phosphate buffer, pH 7.0
using size-exclusion chromatography (SEC, HiTrapTM desalting, GE Healthcare),
concentrated and dialysed extensively into water (molecular weight cut-off 3 000 Da)
yielding cytochrome c oxidase-Oregon Green 488. The concentration of dye labelled
cytochrome c oxidase was not determined.
8.6 Vesicle Formation and Encapsulation
8.6.1 Phospholipid vesicles (liposomes)
8.6.1.1 Preparation of large unilamellar vesicles
Liposomes were prepared using the film hydration method. In a typical experiment, an
organic solution (7 mg, 350 L) of egg phosphatidylcholine (PC) or 1-palmitoyl-2-
oleoyl-sn-glycero-3-phosphocholine (POPC) from a stock solution in chloroform
(20 mg/mL) was added to a 5 mL round bottom flask. Chloroform was evaporated
under a nitrogen stream and residual solvent was removed under high vacuum for 4 h.
The lipid was then dispersed at room temperature by adding 20 mM sodium dihydrogen
phosphate buffer, pH 7.0 (470 L) to a final lipid concentration of 15 mg/mL and
hydrated for 1 h with regular vortexing. To prepare unilamellar vesicles from the
resulting multilamellar liposome suspension, liposomes were treated according to the
methods below.

Chapter 8 Experimental
8-28
Extrusion method
Liposomal suspension (15 mg/mL) was extruded by passing the sample ten times15
through a 100 nm pore (single or double stacked) polycarbonate filters at room
temperature to size and homogenise the vesicle aggregates.15-16
Freeze-thaw method
Liposomal suspension (15 mg/mL) was subjected to five cycles of freezing (liquid
nitrogen) and thawing (40 oC water bath) followed by passing the sample ten times
through a 100 nm pore (single or double stacked) polycarbonate filter at room
temperature to size and homogenise the vesicle aggregates.17
Dehydration-rehydration method
Liposomal suspension (15 mg/mL) was freeze-dried, rehydrated in water for 1 h with
regular vortexing to a final concentration of 15 mg/mL at room temperature. Sample
was then extruded by passing the sample ten times through a 100 nm pore (single or
double stacked) polycarbonate filter at room temperature to size and homogenise the
vesicle aggregates.18
8.6.1.2 Preparation of enzyme encapsulated lipid vesicles
In a typical experiment, using the method described in Chapter 8.6.1.1, enzymes were
loaded into vesicles by rehydration of the dried lipid by adding cytochrome c (100 M)
or cyt c-Oregon Green 488 (5 M) in 20 mM sodium dihydrogen phosphate buffer,
pH 7.0. Suspension was freeze-dried, rehydrated and non-entrapped enzyme was
removed by size exclusion chromatography (SEC, Superdex 75, GE Healthcare) in
20 mM sodium dihydrogen phosphate buffer, pH 7.0 at 0.5 mL/min. The liposome

Chapter 8 Experimental
8-29
containing fractions eluted from 5.0 to 7.0 mL. To determine encapsulation efficiency
(EE%), liposomes were treated with Triton X-100 (2%, v/v) with a sample to surfactant
ratio of 1:1 (v/v).
8.6.1.3 Dynamic light scattering
The average hydrodynamic diameter (Dh) and size distribution of the prepared
aggregates in an aqueous solution (1 mg/mL) was measured using a Malvern
ZetasizerNano ZS instrument equipped with a 4 mV He-Ne laser operating at
= 632 nm, an avalanche photodiode detector with high quantum efficiency, and an
ALV/LSE-5003 multiple- digital correlator electronics system. Phospholipid based
aggregates were filtered to remove dust by extrusion using a (0.1 m) polycarbonate
membrane prior to measurement. Three measurements were made after stabilisation of
the temperature at 25 oC and the average Dh of the aggregates in these runs were
calculated.
8.6.1.4 Liposome staining for electron microscopy
Vesicles were diluted to a concentration of 1 mg/mL for transmission electron
microscopy (TEM) and measurements were performed within 1 day of preparation. A
droplet of liposomes (20 L) were applied to a formvar-coated copper grid for 1 min
and excess sample was blotted away with filter paper. Liposomes were subsequently
negatively stained according to the methods below.
2% Phosphotungstic acid
Aqueous phosphotungstic acid (2%, 20 L) was applied to the TEM grid for 30 s and
excess stain was blotted away.

Chapter 8 Experimental
8-30
2% Uranyl acetate
Aqueous uranyl acetate (2%, 20 L) was applied to the TEM grid for 45 s and excess
stain was blotted away.
8.6.2 Polymer vesicles (polymersomes)
8.6.2.1 Slow addition
Ultrapure water (0.3 mL) was added dropwise (16 L per 10 min) over a period of 3 h
to a solution of polystyrene140-b-poly(acrylic acid)48 (10 mg) dissolved in either
dioxane or tetrahydrofuran (1 mL), a common solvent for both blocks of the copolymer,
at room temperature. A turbid solution formed and was dialysed extensively (molecular
weight cut-off 4 000 Da) into water to remove dioxane/tetrahydrofuran.
8.6.2.2 Injection method
A solution of 1 mg/mL of polystyrene140-b-poly(acrylic acid)48 dissolved in dioxane or
tetrahydrofuran was injected into water at room temperature or 40 oC with continuous
vortexing to a final solvent to water ratio of either 1:2, 1:6, 1:10 (v/v) and allowed to
equilibrate for 2 days.
8.6.2.3 Enzyme induced polymersomes
In a typical experiment, a 1 mg/mL solution of PS140-b-PAA48 in tetrahydrofuran
(33 l) was injected into an enzyme solution (5 M, 200 L) in sodium dihydrogen
phosphate buffer (20 mM, pH 7.0) or phosphate buffered saline (150 mM, pH 7.2). The
solution was allowed to equilibrate for at least 24 h and extensively dialysed against
water using a 50 kDa molecular weight cut-off membrane over 24 h to remove non-
encapsulated enzymes.

Chapter 8 Experimental
8-31
8.6.2.4 Proton pumping proteo-polymersomes
In a typical experiment, a 1 mg/mL solution of PS140-b-PAA48 in tetrahydrofuran
(33 L) was injected into a 8-cyt c (7.5 M), CcOx (0.75 M),
8-hydroxypyrene-1,3,6-trisulfonic acid (5 M) solution (200 L) in phosphate buffered
saline (PBS, 150 mM, pH 7.5) at 25 oC. The buffered enzyme solution was allowed to
equilibrate for 10 min prior to PS140-b-PAA48 in tetrahydrofuran injection. After
injection, the solution was allowed to equilibrate for at least 24 h and extensively
dialysed into ultrapure water using a 50 kDa molecular weight cut-off membrane over
24 h to remove non-encapsulated enzymes and fluorescent dye at 25 oC. This also
removes both the external and internally encapsulated 150 mM PBS salt buffer from the
proteo-polymersome sample. The concentration of 8-cyt c loaded proteo-polymersomes
was estimated by UV-Vis absorbance ( 410 = 97.6 mM-1cm-1).1b
8.7 Enzyme Activity and Photoreaction Experiments
8.7.1 General photo-induced light reaction equipment
Ruthenium(II)-protein biohybrid photo-reactions were performed using a 16 LED Blue
(465 nm) Flashlight (LDP LLC) and Iridium(III)-protein photoreactions were performed
using a 16 LED White (372 nm) Flashlight (LDP LLC). Power measurements of LED
Flashlights were made using a Newport Power Meter (Model 1918-C). Where indicated,
samples were exposed to light (equipped with a UV filter) from an Oriel Basic Power
Supply (model 68806, 50 – 200 watts) fitted with a xenon arc lamp. amGFP photo-
conversion reactions were made using a custom built LED source consisting of an array
of 3 Cyan Lambertian Luxeon-VTM diodes (Phillips, LXHL-LE5C) emitting in a
490±30 nm band connected to an electrical circuit that was fed a power supply at 2.5 A
(30 W). The array was water-cooled with a tightly attached hard disk drive heat sink.

Chapter 8 Experimental
8-32
8.7.2 Cytochrome c/Cytochrome c oxidase studies
8.7.2.1 Measurement of catalytic activity by ABTS assay
The catalytic activity of yeast iso-1 cytochrome c was measured using 2,2’-azino-bis(3-
ethylbenzothiazoline-6-sulphonic acid) (ABTS). A stock solution of 100 mM hydrogen
peroxide was prepared by dilution of 9.4 L of 30% hydrogen peroxide to 1 mL with
ultra pure water. A second stock solution of 20 mM ABTS was prepared by dissolution
of 6.9 mg ABTS in 629 L of 20 mM sodium dihydrogen phosphate, pH 7.0. Activity of
solution (bulk) cytochrome c and encapsulated cytochrome c (PS140-b-PAA48) were
measured at target concentrations of 200 M ABTS, 0.5 M cytochrome c/encapsulated
cytochrome c and 1 mM hydrogen peroxide by adding 1 L of 20 mM ABTS, 54 L of
0.93 M cytochrome c/encapsulated cytochrome c to 44 L of 20 mM sodium
dihydrogen phosphate, pH 7.0 and initiating the reaction with 1 L of 1 mM hydrogen
peroxide. The reactions were monitored by increasing UV absorbance at 415 nm.19
8.7.2.2 Measurement of biological activity by CcOx assay
Biological activity was measured using horse heart cytochrome c oxidase (CcOx). A
stock solution of cytochrome c oxidase was prepared at 1 mg/mL in 20 mM sodium
dihydrogen phosphate buffer, pH 7.0. A catalytic amount (0.25 M) of cytochrome c
oxidase was added to 2.3 M reduced cytochrome c or bioconjugates (bulk and
encapsulated) in 5 mM sodium dihydrogen phosphate, 5 mM ethylenediaminetetraacetic
acid (EDTA), pH 7.0 by twenty-fold dilution and oxidation of cytochrome c was
monitored by decreasing UV absorbance at 550 nm.

Chapter 8 Experimental
8-33
8.7.2.3 Photo-induced electron transfer studies
Room temperature photo-induced electron transfer measurements of a non-covalent
mixture of complex 6 and cytochrome c and 8-cyt c bioconjugate were conducted in
specialised small volume quartz cuvettes designed for protein samples, allowing
complete exposure to irradiation with a constant area for all experiments
(1.0 cm × 0.3 cm). A solution (80 L) of 5 mM sodium dihydrogen phosphate buffer,
5 mM ethylenediaminetetraacetic acid, pH 7.0 was prepared containing either a 1:1
mixture of cytochrome c (2.3 M) and complex 6 (2.3±0.1 M) or bioconjugate (2.3 M)
in bulk or membrane encapsulated (see Chapter 8.6.2.3) samples. Prior to irradiation,
cuvettes were degassed for 30 min at 0 oC under reduced pressure (120 mbar) and
overlayed with nitrogen in the dark. Samples were irradiated in a nitrogen purged
UV-Vis spectrometer with a 465 nm light (LED) source, placed 2.5 cm from sample
and cytochrome c reduction was monitored by UV absorbance at 550 nm.
8.7.2.4 Photo-induced proton pumping studies
In a typical experiment, a solution of salt-free proteo-polymersomes (see Chapter
8.6.2.4) (1.1±0.1 M 8-cyt c, 113.6 L) was adjusted to a final buffered condition of
5 mM NaH2PO4 and 5 mM EDTA by addition of a stock solution of 94 mM NaH2PO4
and 94 mM EDTA (6.4 L) at a desired pH. The sample was degassed for 20 min at
0 oC under reduced pressure (120 mbar) and overlayed with nitrogen in the dark to
remove oxygen quenching the excited state of Ru(II)-bisterpyridine complex as well as
allowing the equilibration of the buffer in the internal proteo-polymersome
compartment due to porosity of the membrane. A 465 nm light (LED) source was
placed 2.5 cm from the sample over a 1.0 cm × 0.3 cm sample area and irradiated in a
nitrogen purged fluorescence spectrometer. Fluorescence emission measurements were

Chapter 8 Experimental
8-34
monitored over 2 min intervals with an ex = 460 nm and em = 510 nm. Fluorescence
excitation measurements were made with a em = 510 nm. Excitation and emission slit
widths were 5 nm. After irradiation, a drop of aqueous sodium hydroxide (2 L, 1 M)
was added to determine the maximum emission intensity (pH > 9) to calibrate the
8-hydroxypyrene-1,3,6-trisulfonic acid pH titration curve (see Chapter 7).
8.7.2.5 8-hydroxypyrene-1,3,6-trisulfonic acid pH titration
Bulk 8-hydroxypyrene-1,3,6-trisulfonic acid (HPTS) (480 M, 3 L) and polymersome
encapsulated HPTS (200 M in lumen, 3 L) (see Chapter 8.6.2.3) was diluted into
20 mM sodium dihydrogen phosphate buffer (3 mL) at a desired pH value and pH of
phosphate buffers were adjusted using aqueous sodium hydroxide (1 M) or hydrochloric
acid (1 M). Titrations were measured from acidic to basic pH and fluorescence emission
intensities were measured with ex = 460 nm and em = 510 nm.
8.7.2.6 Nitrite reductase activity of cytochrome c
Equine heart cytochrome c (1.06 mM, 189 L) was incubated with 100 nm 1-palmitoyl-
2-oleoyl-sn-glycero-3-phosphocholine (POPC) vesicles (15 mg/mL, 100 L) and
bis(4’-(4-hydroxymethylphenyl)-2,2’:6’:2”-terpyridine)iridium(III)tris(hexafluorophosp
hate) (6 mM, 33 L) in 25 mM sodium dihydrogen phosphate, 5 mM
ethylenediaminetetraacetic acid, pH 5.4 (450 L) under reduced pressure (120 mbar) for
10 min. Sodium nitrite (100 M, 50 L) in 25 mM sodium dihydrogen phosphate, 5 mM
ethylenediaminetetraacetic acid, pH 5.4 was subsequently added and sample irradiated
using a 372 nm light (LED) from a distance of 2.0 cm at room temperature for 3 h
followed by incubation for a further 13 h 45 min under anaerobic conditions
(120 mbar). Solution centrifuged at 13 800 rpm for 30 min using a 3 000 molecular

Chapter 8 Experimental
8-35
weight cut-off centrifuge concentrator and filtrate analysed using the Griess assay
(Griess reagent to sample, 1:1, v/v).
8.7.2.7 Griess Assay
To an aqueous solution containing nitrite (100 L), saturated aqueous solution (50 L)
of sulphanilamide (10 mM) in 20 mM sodium dihydrogen phosphate buffer, pH 7.0 and
orthophosphoric acid (1%, v/v) was added. An aqueous solution (50 L) of
N-(1-Napthyl)ethylenediamine dihydrochloride (400 M) in 20 mM sodium dihydrogen
phosphate buffer, pH 7.0 was added to the mixture and incubated for 10 min. Assay was
monitored by measuring UV absorbance at 540 nm and nitrite concentration was
estimated based on a nitrite calibration curve (see Appendix C).
8.7.3 Green fluorescent protein studies
8.7.3.1 Time-resolved laser spectroscopy
Room temperature fluorescence measurements were performed in the laboratory of Dr.
Timothy W. Schmidt at the University of Sydney with Dr. Raphaël G. C. R. Clady and
Mr. Murad Tayebjee. Time-resolved fluorescence lifetime measurements were recorded
by ultrafast (femtosecond) photoexcitation of sample cuvette (non-degassed) using a
tuneable output TOPAS OPA laser (476 nm) pumped by a Clark-MXR femtosecond
laser operating at 1 kHz. The ~1 mm2 fluorescent spot on the front face of the cuvette
was entirely imaged, with a lens, through the slits of a spectrograph and detected with
an iCCD camera (Acton/Princeton) synchronised to the laser output. The kinetics of
delayed fluorescence were measured in 5 ps slices from 5 to 2180 ps delay for ultrafast
time-resolved photoluminescence and in 50 ps slices for time-correlated single photon

Chapter 8 Experimental
8-36
counting. All spectra are baseline corrected. Fitting of the delayed fluorescence signal
was performed with OriginPro 8.
8.7.3.2 Xenon arc lamp irradiation studies
Room temperature photo-induced electron transfer measurements on a 16-GFP
bioconjugate were conducted in a specialised small volume quartz cuvette. A solution
(80 L) of 20 mM sodium dihydrogen phosphate buffer, 20 mM
ethylenediaminetetraacetic acid, pH 7.0 was prepared containing bioconjugate 16-GFP
(13 M). Prior to irradiation, cuvettes were degassed for 30 min at 0 oC under reduced
pressure (120 mbar) and overlayed with nitrogen in the dark. Samples were irradiated
using a xenon lamp fitted with a 2 mm iris from a distance of 10 cm. Samples were
irradiated for 30 min and monitored by UV-Vis spectroscopy at 600 nm.
8.7.3.3 Cyan LED irradiation studies
A solution (120 L) of a non-covalent mixture of green fluorescent protein (20 M) and
4,4’-bipyridinium-N,N-di(maleimidopropyl) hexafluorophosphate, p-benzoquinone,
anthraquinone-2-carboxylic acid (1mM) or Anthraquinone-GFP (12-GFP, 18 M) in
20 mM sodium dihydrogen phosphate buffer, pH 7.0 were irradiated using a custom
built 3 × 490 nm LED source for 3 h at a distance of 2.0 cm. Photoconversion was
followed by UV-Vis and fluorescence (ca. 1.6 M) spectroscopy post-irradiation.
8.8 References
(1) (a) Peterson, J. R.; Smith, T. A.; Thordarson, P. Org. Biomol. Chem. 2010, 8,151. (b) Peterson, J. R.; Thordarson, P. Chiang Mai J. Sci. 2009, 26, 236.
(2) Hvasanov, D.; Wiedenmann, J.; Braet, F.; Thordarson, P. Chem. Commun. 2011,47, 6314.

Chapter 8 Experimental
8-37
(3) (a) Iancu, C. V.; Tivol, W. F.; Schooler, J. B.; Dias, D. P.; Henderson, G. P.; Murphy, G. E.; Wright, E. R.; Li, Z.; Yu, Z.; Briegel, A.; Gan, L.; He, Y.; Jensen, G. J. Nat. Protocols 2007, 1, 2813. (b) Frederik, P. M.; Hubert, D. H. W. Methods Enzymol. 2005, 391, 431.
(4) Angelo, C.; Denzel, A.; Vogt, A.; Matz, M. V.; Oswald, F.; Salih, A.; Nienhaus, G. U.; Wiedenmann, J. Mar. Ecol. Prog. Ser. 2008, 364, 97.
(5) Wiedenmann, J.; Schenk, A.; Röcker, C.; Girod, A.; Spindler, K.-D.; Nienhaus, G. U. Proc. Natl. Acad. Soc. U. S. A. 2002, 99, 11646.
(6) Bruker APEX2 Suite; Bruker AXS Inc.: Madison, WI, USA, 2007.(7) Sheldrick, G. Acta Crystallogr. Sect. A: Found. Crystallogr. 2008, 64, 112.(8) Kabsch, W. J. Appl. Crystallogr. 1993, 26, 795.(9) Mukkala, V.-M.; Helenius, M.; Hemmilä, I.; Kankare, J.; Takalo, H. Helv.
Chim. Acta 1993, 76, 1361.(10) Mikel, C.; Potvin, P. G. Polyhedron 2002, 21, 49.(11) (a) Lainé, P.; Bedioui, F.; Ochsenbein, P.; Marvaud, V.; Bonin, M.; Amouyal, E.
J. Am. Chem. Soc. 2002, 124, 1364. (b) Peterson, J. R. PhD Thesis, The University of Sydney, 2009.
(12) (a) Sullivan, B. P.; Calvert, J. M.; Meyer, T. J. Inorg. Chem. 1980, 19, 1404. (b) Hofmeier, H.; Andres, P. R.; Hoogenboom, R.; Herdtweck, E.; Schubert, U. S. Aust. J. Chem. 2004, 57, 419.
(13) Ng, W. Y.; Gong, X.; Chan, W. K. Chem. Mater. 1999, 11, 1165.(14) Lewis, W. G.; Magallon, F. G.; Fokin, V. V.; Finn, M. G. J. Am. Chem. Soc.
2004, 126, 9152.(15) Mui, B.; Chow, L.; Hope, M. J. Methods Enzymol. 2003, 367, 3.(16) Das, N.; Gupta, S.; Mazumdar, S. Biochem. Biophys. Res. Commun. 2001, 286,
311.(17) (a) Walde, P.; Marzetta, B. Biotechnol. Bioeng. 1998, 57, 216. (b) Blocher, M.;
Walde, P.; Dunn, I. J. Biotechnol. Bioeng. 1999, 62, 36.(18) Kirby, C.; Gregoriadis, G. Nat. Biotech. 1984, 2, 979.(19) Peterson, J. R.; Smith, T. A.; Thordarson, P. Chem. Commun. 2007, 1899.

Chapter 9
Conclusions and Future Work

Chapter 9 Conclusions and Future Work
9-2
9 Conclusions and Future Work
The development of nanoreactors and artificial cells has gained considerable interest
over the last two decades in order to better understand the origin of life.1 In a landmark
article, Szostak and co-workers have proposed three requirements for a model protocell
as (1) bearing an informational substance (DNA or RNA), (2) a catalyst and (3) a
compartment.2 However, implicitly, the requirement for energy production in a model
protocell system is essential to drive these processes. Nearly all organisms on Earth
source energy directly or indirectly from the Sun. In nature, photosynthesis in the
chloroplasts (thylakoid membrane) of plants achieve this energy production by
converting light energy from the Sun into chemical energy in the form of a
transmembrane electrochemical gradient which produces ATP.3 This Thesis describes
the development of a primitive chloroplast – a system that converts light energy into
chemical energy in the form of an electrochemical gradient ( ). Such a system can in
principle be used as a “nanoreactor battery” where the light induced proton gradient is
used to drive chemical reactions or biochemical processes.
In order to develop a primitive artificial chloroplast, a light-activated
donor-acceptor system based on a Ru(II)-bisterpyridine cytochrome c bioconjugate was
synthesised which has been reported in the Thordarson group.4 Terpyridine ligands
bearing amines with 4’-aryl functionalisation as metal complex precursors were
synthesised based on a modified Kröhnke synthesis with isolation of the
azachalcone/pyridinium iodide precursors. Light-activated chromophores of
Ru(II)-bisterpyridine complexes were prepared in ethylene glycol based on conditions
adapted from Ir(III)-bisterpyridine literature.5 Homo and heteroleptic aniline complexes
7 and 9 were synthesised to allow chemical modification by introducing maleimide
functionality using peptide coupling chemistry. Asymmetric and symmetric maleimide

Chapter 9 Conclusions and Future Work
9-3
complexes 8 and 10 were synthesised such that these electron donating photosensitisers
could be attached to cysteine bearing proteins/enzymes. As a reference compound,
complex 6 was prepared for control room temperature photoreduction studies with yeast
iso-1 cytochrome c (cyt c). Single crystals of complexes 7 and 8 were isolated and
analysed by X-ray crystallography to confirm their structural properties which was
conducted by Dr. Mohan Bhadbhade (UNSW) with analysis of aniline 7 performed
in-house at the UNSW Analytical Centre and maleimide 8 at the Australian Synchotron
facility. At the time of writing, complex 8 is the first reported single crystal X-ray
structure of a Ru(II)-bistepyridine complex with maleimide functionality.
The synthesis of a light-activated donor-acceptor system based on an
asymmetric terpyridine cytochrome c bioconjugate 8-cyt c was prepared for electron
transfer studies in membranes and for use as a component in a polymer membrane
reconstituted electron transfer chain as a photosynthetic-respiratory hybrid. The
synthesis of 8-cyt c was achieved by reacting maleimide functionalised asymmetric
complex 8 with the single cysteine residue (CYS102) of yeast iso-1 cytochrome c under
benign conditions at physiological pH 7. The purification of Ru(II)-bisterpyridine based
bioconjugate could be achieved using immobilised metal affinity chromatography
(IMAC, Ni2+) in high purity with 6% yield following procedures previously developed
in the Thordarson group by Peterson et al.4a, although the exact mechanism remains
unknown. The limitation of low yielding asymmetric bioconjugate 8-cyt c is a result of
the poor water solubility.
As an extension of the this work, the preparation methods of high molecular
weight dimeric bioconjugates were explored in order to improve yields. Several
bioconjugate dimers have been prepared based on combinations of cytochrome c, green
fluorescent protein (Acropora millepora) and bovine serum albumin to probe factors

Chapter 9 Conclusions and Future Work
9-4
affecting dimerisation yield. The role and importance of protein charge to analyse
protein function after amino acid modification has been employed in the literature, such
as the use of protein charge ladders.6 However, the exploitation of global protein charge
to facilitate dimer bioconjugate synthesis via supramolecular interactions has been
neglected. It was found that a high molecular weight heterodimer of complementary
charge cyt c-10-BSA (>80 kDa) was prepared in up to 30% yield. Additionally, it is
noteworthy that like-charged homo and heterodimers were prepared in extremely low
yields of less than 1%. Based on these studies, it is proposed that global protein charge
can be utilised to induce higher dimer yields by electrostatic supramolecular ionic
attraction. However, the absolute purification of dimers proved to be difficult due to the
low yields of the dimers. The bioconjugates were characterised by MALDI-TOF mass
spectrometry, UV-Vis spectroscopy and gel electrophoresis.
Following the work of synthetic donor-biological acceptor systems based on
Ru(II)-cyt c, the alternative biological donor-synthetic acceptor construct was
investigated using green fluorescent protein (GFP) for light-induced electron transfer
studies. Anthraquinone precursors were prepared to function as electron acceptors after
functionalising to the light-activated biological electron donor, GFP. Azide bearing
anthraquinone 13 was prepared for potential Cu(I)-catalysed [3+2] Huisgen
cycloaddition reactions (click reactions) of azides with terminal alkynes to introduce
maleimide functionality and subsequently site-specifically modify GFP at the single
CYS119 residue. However, attempts to functionalise azide 13 with maleimides using
click chemistry proved to be unsuccessful. Instead, the N-hydroxysuccinimide ester of
anthraquinone-2-carboxylic acid (anthraquinone 12) was prepared to non-specifically
modify GFP via amine residues. Electron transfer studies were conducted using
viologen-GFP (16-GFP) and anthraquinone-GFP (12-GFP) bioconjugates, which were

Chapter 9 Conclusions and Future Work
9-5
prepared via cysteine-maleimide coupling (reacting to single CYS119 residue) and
amine modification (non-specific), respectively.
The purification of the resulting bioconjugates was achieved using Ni2+-IMAC
chromatography, resulting in yields of 11% and 27%, respectively. Based on
steady-state UV-Vis and fluorescence spectroscopy studies, evidence of electron
transfer was observed by red photoconversion of the chromophore with p-benzoquinone
and anthraquinone acceptor molecules as a non-covalent mixture. To further confirm
electron/energy transfer processes of non-covalent and bioconjugate GFP mixtures,
time-resolved fluorescence spectroscopy measurements were conducted. Lifetime
measurements showed a GFP fluorescence lifetime of 1.65±0.06 ns. Furthermore, the
fluorescence lifetimes of GFP were quenched to 0.14±0.01 and 0.23±0.04 ns in the
presence of excess (non-covalent) p-benzoquinone and anthraquinone-2-carboxylic
acid, respectively, corresponding to a rate constant of forward electron transfer of
ket = 6.5±0.5 × 109 and 3.7±0.7 × 109 s-1. The donor-acceptor viologen bioconjugate
16-GFP showed no evidence of photoinduced electron transfer as viologen acts as a
poor electron acceptor in 16-GFP, which is consistent with non-covalent steady state
UV-Vis and fluorescence spectroscopy measurements. In contrast, fluorescence lifetime
studies of the anthraquinone bioconjugate 12-GFP demonstrated light-induced
electron/energy transfer processes, with a quenched GFP fluorescence lifetime of
0.28±0.04 ns and an electron transfer rate of ket = 2.9±0.4 × 109 s-1.
Vesicles based on phospholipids and diblock copolymers were investigated to
evaluate their feasibility as compartments for the encapsulation of donor-acceptor
8-cyt c. The first compartment candidate was based on lipid vesicles (liposomes). Lipid
vesicles were formed from either naturally sourced egg L- -phosphatidylcholine (egg
PC) or synthetically equivalent L-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine

Chapter 9 Conclusions and Future Work
9-6
(POPC), using the film hydration method and sized via extrusion to produce
monodisperse unilamellar vesicles of 100 nm in diameter. The validation of
monodispersity and average diameter was determined using dynamic light scattering
techniques. Additionally, unilamellar vesicles were characterised using traditional TEM
microscopy and cryo-TEM techniques and it was found that structural characterisation
of lamellar features using conventional TEM microscopy introduced artificacts as a
result of the necessity of negative stains and drying of specimen. The possibility of
liposomes based on egg PC and POPC as potential compartments were determined by
the encapsulation of model protein cytochrome c, which is used as the redox
metalloprotein for light-induced electron transfer. Encapsulation was confirmed using
confocal laser-scanning microscopy (CLSM) of fluorescently labelled cytochrome c.
However, low encapsulation efficiencies of 1.5±0.1% prevented their further use for
bioconjugate encapsulation studies.
Polymersomes were also formed as potential compartments as an alternative
candidate, using the polyelectrolyte diblock copolymer, polystyrene140-b-poly(acrylic
acid)48 (PS140-b-PAA48). A novel method of polymersome preparation was
demonstrated using the syringe injection method which induces polymersome
aggregates in a facile and biologically friendly manner. Concomitantly, encapsulation of
positively charged biomolecules occurs within the coronal block. It is proposed that the
formation of polymersomes is in a 2:1 polymer:protein ratio which was investigated by
determining the relative ratio of polymersome/micelle formation as a function of
positive charge (enzyme) composition. Additionally, this induced formation is versatile
and can be applied to various positively charged biomolecules with high encapsulation
efficiencies allowing for potential applications as nanoreactors7 or protein therapeutic
vessels including use of polycationic copolymers as non-viral vectors for DNA/RNA

Chapter 9 Conclusions and Future Work
9-7
gene therapy8 and simplified preparation compared to polyelectrolyte microcapsules.9
Polymersomes were characterised by conventional and cryo-TEM microscopy as well
as CLSM. The encapsulated enzymes were demonstrated to remain biologically active
after encapsulation in the presence of organic solvent (tetrahydrofuran), using
spectroscopy studies and the ABTS assay. Due to the high encapsulation efficiencies of
55±1% and 66±7% in 20 mM NaH2PO4 and PBS, respectively, for cytochrome c and
35±1% for GFP, polymersomes were selected as the most suitable compartments for
enzyme encapsulation.
Using the formation of PS140-b-PAA48 polymersomes which allows the
encapsulation of positively charged enzymes within the coronal block of the membrane,
photoinduced electron transfer studies were performed using 8-cyt c. Initially, the
photophysical properties of bulk Ru(II)-cyt c bioconjugate 8-cyt c was investigated. The
photoreduction of 8-cyt c could be followed by monitoring the increase in the 550 nm
absorption band corresponding to heme reduction using steady-state UV-Vis absorption
spectroscopy measurements. It was observed that Ru(II) 8-cyt c could be fully reduced
in 50 min via irradiation with a 465 nm LED light source. The quantum efficiency ( )
of bioconjugate 8-cyt c in bulk solution was estimated to be 5.9±1.5 10-4%.
Subsequently, in order to determine the effect of membrane encapsulation for use as a
component in a semi-synthetic electron transport chain, bioconjugate 8-cyt c was
membrane encapsulated in PS140-b-PAA48. Induced 8-cyt c polymersome formation
was confirmed by TEM microscopy with an average diameter of 290±132 nm. The
encapsulated conjugate 8-cyt c, exhibited a two-fold enhancement of the initial rate of
heme reduction with an estimated of 1.1±0.3 × 10-3%. Molecular modelling of 8-cyt c
showed that electron transfer is occurring over a maximum distance between the
ruthenium and heme centre of 32 Å.

Chapter 9 Conclusions and Future Work
9-8
Expanding on photoreduction of cytochrome c, electron transfer studies between
a non-covalent mixture of Ir(III)-bisterpyridine complex 18 and horse heart
cytochrome c using a 372 nm UV light source was investigated to induce nitrite
reductase mimicry behaviour. It was shown that in the presence of zwitterionic
phospholipid (POPC) liposomes (100 nm), a pentacoordinate heme state could be
induced allowing reactions with nitrite anions to form nitric oxide. The photo-activated
nitrite reductase activity of cytochrome c exhibited an 82% conversion of nitrite anions
into nitric oxide after irradiation under anaerobic conditions in an acidic phosphate
buffered environment (pH 5.4). Due to experimental limitations, the exact irradiation
power could not be determined due scattering and absorption by the vacuum flask and
nitrite levels could only measured at a single time point.
After investigation of the photophysical properties of bulk and membrane
encapsulated PS140-b-PAA48 polymersomes, a synthetic hybrid
photosynthetic-respiratory system that generates a proton potential upon photoexcitation
was constructed via self-assembly. Reconstitution of the hybrid enzyme cascade
consisting of bioconjugate 8-cyt c, cytochrome c oxidase in the polymersome
membrane of polyelectrolyte PS140-b-PAA48 was characterised by TEM and CLSM.
Irradiation with visible light (465 nm) converts photon energy into an electrochemical
potential via electron transfer. Photoinduced electron transfer occurs across an
artificial electron transport chain within the polymersome membrane coupled with
overall vectorial proton translocation and resulting pH changes were monitored by
steady-state fluorescence spectroscopy using internally encapsulated HPTS. Following
fluorescence spectroscopy studies, it was observed that cytochrome c oxidase appears to
have the same orientation (average >50% native orientation) in the polymersome
membrane as encountered in the mitochondria. The resulting biased orientation is

Chapter 9 Conclusions and Future Work
9-9
attributed to the alignment of cytochrome c oxidase with the curvature of the
polymersomes. The photosynthetic-respiratory system is capable of storing chemical
energy in the form of a transmembrane proton potential with a proton pumping rate of
3.3 × 103 H+/s at physiological pH 7.2 ( pH 0.2 units) correlating to a of
1.1±0.3 × 10-9%. Proton translocation rates were investigated using pH-dependent
studies showing that a four-fold increase in proton pumping rate is observed under
acidic conditions of pH 6.5 despite the increased proton "back-pressure".
In conclusion, this research has shown that the construction of the synthetic
photoactive electron transport chain capable of storing chemical energy as a proton
gradient brings vesicle and polymersome chemists towards satisfying one of the three
requirements2 for an artificial model protocell, which include (1) catalyst (storing of
chemical energy), (2) self-reproducing vesicles1a and (3) self-reproducing informational
substance (genetic information).1a The semi-synthetic nature of the electron transport
chain, which is reconstituted in an artificial polyelectrolyte membrane demonstrates the
construction of a robust and reproducible primitive chloroplast. This system opens
opportunities for further study including possibly utilising this system to drive
biomimetic processes such as generation of ATP10 and ATP-linked processes by
potentially reconstituting transmembrane ATP synthase or alternatively inducing
pH-sensitive reactions within the lumen.
9.1 References
(1) (a) Kurihara, K.; Tamura, M.; Shohda, K.-i.; Toyota, T.; Suzuki, K.; Sugawara, T. Nat. Chem. 2011, 3, 775. (b) Vriezema, D. M.; Garcia, P. M. L.; SanchoOltra, N.; Hatzakis, N. S.; Kuiper, S. M.; Nolte, R. J. M.; Rowan, A. E.; vanHest, J. C. M. Angew. Chem. Int. Ed. 2007, 46, 7378.
(2) Szostak, J. W.; Bartel, D. P.; Luisi, P. L. Nature 2001, 409, 387.(3) Cordes, M.; Giese, B. Chem. Soc. Rev. 2009, 38, 892.(4) (a) Peterson, J. R.; Smith, T. A.; Thordarson, P. Chem. Commun. 2007, 1899. (b)
Peterson, J. R.; Smith, T. A.; Thordarson, P. Org. Biomol. Chem. 2010, 8, 151.

Chapter 9 Conclusions and Future Work
9-10
(5) Hofmeier, H.; Schubert, U. S. Chem. Soc. Rev. 2004, 33, 373.(6) (a) Gitlin, I.; Carbeck, J. D.; Whitesides, G. M. Angew. Chem. Int. Ed. 2006, 45,
3022. (b) Gitlin, I.; Mayer, M.; Whitesides, G. M. J. Phys. Chem. B 2003, 107,1466.
(7) Minten, I. J.; Claessen, V. I.; Blank, K.; Rowan, A. E.; Nolte, R. J. M.; Cornelissen, J. J. L. M. Chem. Sci. 2011, 2, 358.
(8) Ropert, C. Braz. J. Med. Biol. Res. 1999, 32, 163.(9) Zelikin, A. N.; Becker, A. L.; Johnston, A. P. R.; Wark, K. L.; Turatti, F.;
Caruso, F. ACS Nano 2007, 1, 63.(10) (a) Choi, H.-J.; Montemagno, C. D. Nano Lett. 2005, 5, 2538. (b) Steinberg-
Yfrach, G.; Rigaud, J.-L.; Durantini, E. N.; Moore, A. L.; Gust, D.; Moore, T. A. Nature 1998, 392, 479.

Appendices

Appendices
Appendix A
A.1 Supplementary data for X-ray diffraction analysis of Ru(II) complex 7
Table A.1. Data collection for Ru(II) complex 7
Radiation source: fine-focus sealed tube 7849 independent reflections32252 measured reflections 5932 reflections with I > 2 (I)Graphite monochromator Rint = 0.056
scans, and scans with offsets max = 25.0o, max = 2.7o
Absorption correction: Multi-scan h = -10 10SADABS (Bruker, 2001) k = -10 10Tmin = 0.924, Tmax = 0.974 l = -33 33
Table A.2. Refinement for Ru(II) complex 7
Refinement on F2 Primary atom site location: Structure-invariant direct methods
Least-squares matrix: Full Secondary atom site location: Difference Fourier map
R[F2 > 2 (F2)] = 0.041 Hydrogen site location: Inferred from neighbouring sites
wR(F2) = 0.125 H-atom parameters constrainedS = 0.83 w = 1/[ 2(Fo
2) + (0.1P)2 + 0.7828P] where P =(Fo
2 + 2Fc2)/3
7849 reflections ( / )max = 0.002672 parameters max = 0.62 e Å-3
12 restraints min = -0.52 e Å-3
Table A.3. Fractional atomic coordinates and isotropic or equivalent isotropic displacement parameters (Å2)
x y z U iso*/Ueq Occ. (<1)P1A 0.60250 (16) 0.34843 (13) 0.85755 (4) 0.0454 (3)F1A 0.713 (3) 0.487 (2) 0.8553 (8) 0.108 (9) 0.33 (2)F2A 0.745 (3) 0.257 (3) 0.8420 (10) 0.140 (13) 0.33 (2)F3A 0.499 (3) 0.205 (2) 0.8582 (8) 0.120 (12) 0.33 (2)F4A 0.479 (4) 0.460 (3) 0.8727 (8) 0.110 (10) 0.33 (2)F5A 0.545 (3) 0.349 (3) 0.8037 (4) 0.074 (7) 0.33 (2)F6A 0.664 (4) 0.323 (5) 0.9102 (5) 0.076 (8) 0.33 (2)F1A' 0.642 (3) 0.5117 (7) 0.8666 (5) 0.162 (7) 0.67 (2)F2A' 0.7719 (7) 0.3036 (14) 0.8525 (4) 0.089 (3) 0.67 (2)F3A' 0.5591 (16) 0.1818 (6) 0.8471 (6) 0.108 (5) 0.67 (2)F4A' 0.4243 (10) 0.375 (2) 0.8606 (5) 0.127 (5) 0.67 (2)F5A' 0.5874 (14) 0.3886 (15) 0.8031 (2) 0.064 (3) 0.67 (2)F6A' 0.615 (2) 0.317 (3) 0.9119 (3) 0.091 (5) 0.67 (2)Ru1 0.78387 (3) 0.83275 (3) 0.721565 (10) 0.01946 (11)N1A 0.7035 (3) 1.0282 (3) 0.69429 (11) 0.0243 (7)N2A 0.8119 (3) 0.7998 (3) 0.65361 (11) 0.0255 (7)

Appendices
N3A 0.8671 (3) 0.6207 (3) 0.72202 (12) 0.0256 (7)C1A 0.6451 (4) 1.1439 (4) 0.71774 (15) 0.0269 (9)H1A 0.6418 1.1394 0.7510 0.032*C2A 0.5903 (5) 1.2678 (5) 0.69557 (16) 0.0360 (10)H2A 0.5479 1.3461 0.7132 0.043*C3A 0.5978 (5) 1.2766 (5) 0.64765 (17) 0.0400 (11)H3A 0.5618 1.3616 0.6316 0.048*C4A 0.6584 (5) 1.1603 (5) 0.62311 (16) 0.0384 (11)H4A 0.6645 1.1652 0.5899 0.046*C5A 0.7104 (4) 1.0367 (4) 0.64652 (14) 0.0275 (9)C6A 0.7768 (5) 0.9085 (4) 0.62331 (13) 0.0295 (9)C7A 0.8093 (6) 0.8921 (5) 0.57639 (15) 0.0447 (12)H7A 0.7853 0.9680 0.5547 0.054*C8A 0.8770 (6) 0.7638 (6) 0.56164 (17) 0.0506 (13)H8A 0.8990 0.7509 0.5295 0.061*C9A 0.9133 (5) 0.6529 (5) 0.59337 (16) 0.0450 (12)H9A 0.9609 0.5650 0.5833 0.054*C10A 0.8788 (4) 0.6729 (4) 0.63990 (14) 0.0307 (9)C11A 0.9062 (4) 0.5707 (4) 0.67858 (15) 0.0292 (9)C12A 0.9646 (5) 0.4299 (5) 0.67295 (18) 0.0404 (11)H12A 0.9911 0.3951 0.6428 0.048*C13A 0.9835 (5) 0.3413 (5) 0.71157 (19) 0.0458 (13)H13A 1.0277 0.2465 0.7083 0.055*C14A 0.9388 (5) 0.3891 (5) 0.75472 (18) 0.0403 (11)H14A 0.9468 0.3268 0.7812 0.048*C15A 0.8818 (4) 0.5306 (4) 0.75859 (16) 0.0310 (9)H15A 0.8519 0.5649 0.7884 0.037*N1B 0.9959 (3) 0.9272 (3) 0.74028 (10) 0.0192 (6)N2B 0.7655 (3) 0.8670 (3) 0.78959 (10) 0.0206 (7)N3B 0.5658 (3) 0.7491 (3) 0.73025 (11) 0.0238 (7)N4B 0.6978 (5) 0.9776 (4) 1.08262 (12) 0.0456 (10)H4B1 0.6412 0.9168 1.0985 0.055*H4B2 0.7489 1.0509 1.0973 0.055*C1B 1.1115 (4) 0.9561 (4) 0.71184 (13) 0.0224 (8)H1B 1.0993 0.9270 0.6796 0.027*C2B 1.2456 (4) 1.0258 (4) 0.72752 (14) 0.0279 (9)H2B 1.3249 1.0434 0.7066 0.034*C3B 1.2640 (5) 1.0702 (4) 0.77424 (14) 0.0314 (9)H3B 1.3544 1.1219 0.7857 0.038*C4B 1.1479 (4) 1.0379 (4) 0.80391 (13) 0.0271 (9)H4B 1.1595 1.0648 0.8363 0.033*C5B 1.0154 (4) 0.9666 (4) 0.78651 (12) 0.0195 (8)C6B 0.8845 (4) 0.9287 (4) 0.81554 (13) 0.0229 (8)C7B 0.8775 (4) 0.9458 (4) 0.86366 (13) 0.0249 (8)H7B 0.9625 0.9877 0.8817 0.030*C8B 0.7447 (5) 0.9013 (4) 0.88597 (14) 0.0271 (9)C9B 0.6238 (4) 0.8385 (4) 0.85746 (14) 0.0288 (9)H9B 0.5323 0.8070 0.8713 0.035*

Appendices
C10B 0.6358 (4) 0.8219 (4) 0.80993 (13) 0.0216 (8)C11B 0.5229 (4) 0.7531 (4) 0.77566 (14) 0.0253 (9)C12B 0.3835 (4) 0.6913 (5) 0.78767 (16) 0.0335 (10)H12B 0.3540 0.6950 0.8193 0.040*C13B 0.2894 (5) 0.6250 (5) 0.75311 (17) 0.0409 (11)H13B 0.1938 0.5824 0.7607 0.049*C14B 0.3338 (5) 0.6201 (5) 0.70739 (17) 0.0397 (11)H14B 0.2697 0.5736 0.6833 0.048*C15B 0.4704 (4) 0.6825 (4) 0.69718 (15) 0.0311 (9)H15B 0.5001 0.6792 0.6655 0.037*C16B 0.7339 (5) 0.9204 (4) 0.93706 (14) 0.0309 (9)C17B 0.8121 (5) 1.0334 (5) 0.96206 (14) 0.0347 (10)H17B 0.8753 1.0995 0.9458 0.042*C18B 0.8010 (5) 1.0529 (5) 1.00985 (15) 0.0383 (11)H18B 0.8573 1.1306 1.0261 0.046*C19B 0.7063 (5) 0.9579 (5) 1.03462 (14) 0.0364 (11)C20B 0.6280 (5) 0.8444 (5) 1.01012 (15) 0.0393 (11)H20B 0.5643 0.7787 1.0264 0.047*C21B 0.6411 (5) 0.8250 (5) 0.96246 (14) 0.0365 (10)H21B 0.5866 0.7459 0.9464 0.044*C1D 0.7073 (7) 0.4443 (7) 0.0391 (3) 0.0815 (19)H1D1 0.6725 0.3416 0.0423 0.122*H1D2 0.7287 0.4623 0.0062 0.122*H1D3 0.6271 0.5112 0.0489 0.122*C2D 0.9311 (9) 0.6126 (8) 0.0632 (3) 0.114 (3)H2D1 0.8642 0.6937 0.0718 0.171*H2D2 0.9550 0.6205 0.0301 0.171*H2D3 1.0265 0.6183 0.0830 0.171*N1D 0.8538 (6) 0.4723 (5) 0.07032 (19) 0.0703 (14)C3D 0.8830 (9) 0.3857 (8) 0.1053 (2) 0.086 (2)H3D 0.9706 0.4112 0.1252 0.103*O1D 0.8107 (6) 0.2744 (4) 0.11575 (15) 0.0827 (14)C2E 0.7544 (8) 0.3853 (9) 0.5056 (2) 0.100 (3)H2E1 0.7862 0.4875 0.4999 0.151*H2E2 0.8435 0.3209 0.5046 0.151*H2E3 0.7113 0.3799 0.5367 0.151*C1E 0.5886 (7) 0.1866 (6) 0.4704 (2) 0.0687 (17)H1E1 0.5338 0.1609 0.4403 0.103*H1E2 0.5197 0.1728 0.4961 0.103*H1E3 0.6776 0.1226 0.4750 0.103*N1E 0.6394 (5) 0.3376 (5) 0.46995 (14) 0.0471 (10)C3E 0.5763 (7) 0.4307 (6) 0.4394 (2) 0.0630 (15)H3E 0.6131 0.5301 0.4413 0.076*O1E 0.4765 (4) 0.4022 (4) 0.40915 (12) 0.0604 (10)P1B 0.18071 (13) 0.17081 (13) 0.58486 (4) 0.0365 (3)F1B 0.3310 (3) 0.1981 (3) 0.55689 (10) 0.0581 (8)F2B 0.1036 (3) 0.3115 (3) 0.56081 (10) 0.0577 (8)F3B 0.0298 (3) 0.1440 (3) 0.61339 (10) 0.0574 (8)

Appendices
F4B 0.2581 (4) 0.0315 (3) 0.60954 (10) 0.0708 (9)F5B 0.1129 (3) 0.0685 (3) 0.54266 (10) 0.0582 (8)F6B 0.2477 (3) 0.2766 (3) 0.62676 (9) 0.0537 (7)
Table A.4. Atomic displacement parameters (Å2)U11 U22 U33 U12 U13 U23
P1A 0.0654 (9) 0.0326 (6) 0.0395 (7) 0.0080 (6) 0.0109 (6) 0.0064 (5)F1A 0.171 (17) 0.091 (17) 0.064 (11) 0.052 (10)F2A 0.23 (3) 0.098 (14) 0.106 (16) 0.131 (18) 0.062 (16) 0.050 (12)F3A 0.098 (17) 0.19 (3) 0.066 (9) (17) 0.037 (12)F4A 0.16 (2) 0.102 (17) 0.078 (13) 0.072 (14) 0.071 (14) 0.030 (11)F5A 0.119 (19) 0.068 (11) 0.033 (7) 0.002 (7) 0.012 (5)F6A 0.078 (18) 0.112 (14) 0.037 (8) 0.014 (9)F1A' 0.40 (2) 0.017 (3) 0.068 (8)F2A' 0.043 (4) 0.144 (9) 0.079 (5) 0.005 (3)F3A' 0.092 (7) 0.039 (4) 0.184 (12) 0.023 (4)F4A' 0.085 (6) 0.177 (12) 0.124 (8) 0.071 (7) 0.032 (5) 0.036 (8)F5A' 0.065 (5) 0.078 (7) 0.051 (4) 0.004 (3) 0.020 (3)F6A' 0.097 (11) 0.120 (9) 0.059 (5) 0.020 (8) 0.024 (4) 0.048 (5)Ru1 0.01729 (17) 0.01850 (17) 0.02258 (18) 0.00113 (12) 0.00061 (11)N1A 0.0179 (16) 0.0248 (17) 0.0301 (19) 0.0004 (13) 0.0019 (14)N2A 0.0212 (17) 0.0288 (17) 0.0260 (18)N3A 0.0177 (16) 0.0220 (17) 0.037 (2)C1A 0.020 (2) 0.0219 (19) 0.039 (2) 0.0018 (16) 0.0025 (17) 0.0018 (17)C2A 0.025 (2) 0.030 (2) 0.053 (3) 0.0033 (18) 0.000 (2) 0.005 (2)C3A 0.028 (2) 0.036 (2) 0.055 (3) 0.017 (2)C4A 0.034 (2) 0.046 (3) 0.034 (3) 0.013 (2)C5A 0.021 (2) 0.033 (2) 0.027 (2) 0.0049 (17)C6A 0.030 (2) 0.037 (2) 0.020 (2) 0.0008 (17)C7A 0.060 (3) 0.050 (3) 0.024 (2) 0.000 (2)C8A 0.063 (3) 0.061 (3) 0.028 (3) 0.006 (2)C9A 0.044 (3) 0.048 (3) 0.043 (3) 0.005 (2)C10A 0.027 (2) 0.029 (2) 0.035 (2) 0.0035 (18)C11A 0.0145 (19) 0.028 (2) 0.045 (3) 0.0035 (17)C12A 0.027 (2) 0.028 (2) 0.066 (3) 0.006 (2)C13A 0.027 (2) 0.023 (2) 0.088 (4) 0.0007 (18) 0.001 (2)C14A 0.030 (2) 0.027 (2) 0.064 (3) 0.000 (2) 0.007 (2)C15A 0.022 (2) 0.024 (2) 0.047 (3) 0.0059 (19)N1B 0.0194 (16) 0.0175 (15) 0.0205 (17) 0.0011 (12) 0.0009 (12)N2B 0.0183 (16) 0.0180 (15) 0.0255 (17) 0.0001 (12) 0.0002 (13) 0.0031 (13)N3B 0.0218 (17) 0.0199 (16) 0.0297 (19) 0.0016 (13) 0.0005 (13)N4B 0.057 (3) 0.058 (3) 0.023 (2) 0.004 (2) 0.0054 (18) 0.0053 (17)C1B 0.025 (2) 0.0214 (18) 0.021 (2) 0.0026 (16) 0.0034 (16) 0.0039 (15)C2B 0.021 (2) 0.032 (2) 0.031 (2) 0.0070 (17) 0.0038 (17)C3B 0.023 (2) 0.036 (2) 0.035 (2) 0.0026 (18)C4B 0.027 (2) 0.034 (2) 0.020 (2) 0.0015 (16)C5B 0.0191 (19) 0.0173 (18) 0.022 (2) 0.0009 (15) 0.0040 (15)

Appendices
C6B 0.024 (2) 0.0195 (18) 0.026 (2) 0.0020 (16) 0.0022 (16) 0.0037 (15)C7B 0.025 (2) 0.024 (2) 0.026 (2) 0.0044 (16) 0.0019 (16)C8B 0.033 (2) 0.0212 (19) 0.028 (2) 0.0038 (17) 0.0061 (18) 0.0035 (16)C9B 0.025 (2) 0.028 (2) 0.033 (2) 0.0080 (18) 0.0074 (17)C10B 0.0177 (19) 0.0162 (17) 0.032 (2) 0.0015 (15) 0.0067 (16) 0.0066 (15)C11B 0.021 (2) 0.0183 (19) 0.037 (2) 0.0038 (15) 0.0030 (17) 0.0066 (16)C12B 0.022 (2) 0.035 (2) 0.044 (3) 0.0001 (18) 0.0064 (19) 0.0106 (19)C13B 0.020 (2) 0.043 (3) 0.060 (3) 0.002 (2) 0.003 (2)C14B 0.024 (2) 0.042 (3) 0.052 (3)C15B 0.025 (2) 0.029 (2) 0.038 (3) 0.0005 (18)C16B 0.029 (2) 0.034 (2) 0.030 (2) 0.0041 (18) 0.0038 (18) 0.0041 (18)C17B 0.039 (3) 0.035 (2) 0.031 (2) 0.0088 (19) 0.0025 (18)C18B 0.046 (3) 0.038 (2) 0.031 (2) 0.004 (2) 0.006 (2)C19B 0.038 (3) 0.046 (3) 0.026 (2) 0.019 (2) 0.0053 (19) 0.011 (2)C20B 0.043 (3) 0.049 (3) 0.027 (2) 0.000 (2) 0.009 (2) 0.012 (2)C21B 0.040 (3) 0.040 (2) 0.030 (2) 0.0021 (19) 0.0070 (19)C1D 0.063 (4) 0.066 (4) 0.114 (6)C2D 0.090 (5) 0.068 (4) 0.180 (9) 0.051 (5)N1D 0.077 (4) 0.047 (3) 0.088 (4) 0.013 (3) 0.007 (3)C3D 0.129 (6) 0.072 (4) 0.056 (4) 0.004 (4) 0.004 (3)O1D 0.131 (4) 0.043 (2) 0.078 (3) 0.051 (3) 0.008 (2)C2E 0.097 (6) 0.134 (7) 0.066 (4) 0.035 (4)C1E 0.093 (5) 0.056 (3) 0.057 (4) 0.022 (3) 0.001 (3)N1E 0.045 (2) 0.054 (3) 0.041 (2) 0.011 (2)C3E 0.080 (4) 0.056 (3) 0.051 (3) 0.010 (3)O1E 0.072 (3) 0.064 (2) 0.044 (2) 0.0093 (18)P1B 0.0381 (7) 0.0418 (7) 0.0300 (6) 0.0029 (5) 0.0057 (5)F1B 0.0526 (18) 0.072 (2) 0.0517 (18) 0.0038 (15) 0.0221 (14)F2B 0.070 (2) 0.0518 (17) 0.0508 (18) 0.0158 (15) 0.0033 (14)F3B 0.0541 (18) 0.0559 (17) 0.0631 (19) 0.0229 (14)F4B 0.103 (3) 0.067 (2) 0.0461 (18) 0.0372 (18) 0.0189 (17) 0.0144 (15)F5B 0.069 (2) 0.0598 (18) 0.0450 (17) 0.0037 (14)F6B 0.0385 (15) 0.079 (2) 0.0426 (16) 0.0006 (14) 0.0002 (12)
Table A.5. Geometric parameters (Å, o)P1A—F1A' 1.525 (6) C2B—H2B 0.9500P1A—F2A' 1.551 (6) C3B—C4B 1.383 (5)P1A—F4A 1.555 (9) C3B—H3B 0.9500P1A—F3A' 1.567 (6) C4B—C5B 1.380 (5)P1A—F3A 1.569 (10) C4B—H4B 0.9500P1A—F6A' 1.575 (7) C5B—C6B 1.483 (5)P1A—F1A 1.576 (9) C6B—C7B 1.380 (5)P1A—F4A' 1.583 (7) C7B—C8B 1.407 (5)P1A—F2A 1.584 (9) C7B—H7B 0.9500P1A—F6A 1.586 (10) C8B—C9B 1.403 (6)P1A—F5A 1.587 (9) C8B—C16B 1.469 (5)P1A—F5A' 1.597 (6) C9B—C10B 1.368 (5)

Appendices
Ru1—N2B 1.971 (3) C9B—H9B 0.9500Ru1—N2A 1.980 (3) C10B—C11B 1.472 (5)Ru1—N3A 2.062 (3) C11B—C12B 1.391 (5)Ru1—N1A 2.062 (3) C12B—C13B 1.372 (6)Ru1—N3B 2.064 (3) C12B—H12B 0.9500Ru1—N1B 2.066 (3) C13B—C14B 1.377 (6)N1A—C1A 1.351 (5) C13B—H13B 0.9500N1A—C5A 1.368 (5) C14B—C15B 1.358 (6)N2A—C6A 1.348 (5) C14B—H14B 0.9500N2A—C10A 1.356 (5) C15B—H15B 0.9500N3A—C15A 1.335 (5) C16B—C17B 1.385 (6)N3A—C11A 1.370 (5) C16B—C21B 1.407 (6)C1A—C2A 1.376 (5) C17B—C18B 1.377 (6)C1A—H1A 0.9500 C17B—H17B 0.9500C2A—C3A 1.373 (6) C18B—C19B 1.406 (6)C2A—H2A 0.9500 C18B—H18B 0.9500C3A—C4A 1.380 (6) C19B—C20B 1.382 (7)C3A—H3A 0.9500 C20B—C21B 1.376 (6)C4A—C5A 1.382 (6) C20B—H20B 0.9500C4A—H4A 0.9500 C21B—H21B 0.9500C5A—C6A 1.467 (6) C1D—N1D 1.533 (8)C6A—C7A 1.386 (6) C1D—H1D1 0.9800C7A—C8A 1.379 (7) C1D—H1D2 0.9800C7A—H7A 0.9500 C1D—H1D3 0.9800C8A—C9A 1.390 (7) C2D—N1D 1.450 (7)C8A—H8A 0.9500 C2D—H2D1 0.9800C9A—C10A 1.383 (6) C2D—H2D2 0.9800C9A—H9A 0.9500 C2D—H2D3 0.9800C10A—C11A 1.462 (6) N1D—C3D 1.295 (8)C11A—C12A 1.389 (6) C3D—O1D 1.229 (8)C12A—C13A 1.376 (7) C3D—H3D 0.9500C12A—H12A 0.9500 C2E—N1E 1.445 (7)C13A—C14A 1.372 (7) C2E—H2E1 0.9800C13A—H13A 0.9500 C2E—H2E2 0.9800C14A—C15A 1.385 (6) C2E—H2E3 0.9800C14A—H14A 0.9500 C1E—N1E 1.429 (7)C15A—H15A 0.9500 C1E—H1E1 0.9800N1B—C1B 1.351 (5) C1E—H1E2 0.9800N1B—C5B 1.359 (5) C1E—H1E3 0.9800N2B—C6B 1.350 (5) N1E—C3E 1.327 (6)N2B—C10B 1.357 (5) C3E—O1E 1.212 (6)N3B—C15B 1.351 (5) C3E—H3E 0.9500N3B—C11B 1.364 (5) P1B—F1B 1.587 (3)N4B—C19B 1.381 (5) P1B—F5B 1.587 (3)N4B—H4B1 0.8800 P1B—F4B 1.595 (3)N4B—H4B2 0.8800 P1B—F6B 1.596 (3)C1B—C2B 1.370 (5) P1B—F2B 1.597 (3)C1B—H1B 0.9500 P1B—F3B 1.600 (3)

Appendices
C2B—C3B 1.383 (6)
F1A'—P1A—F2A' 94.0 (9) C15A—C14A—H14A 120.9F1A'—P1A—F4A 58.7 (10) N3A—C15A—C14A 122.6 (4)F2A'—P1A—F4A 151.6 (15) N3A—C15A—H15A 118.7F1A'—P1A—F3A' 178.5 (7) C14A—C15A—H15A 118.7F2A'—P1A—F3A' 86.7 (5) C1B—N1B—C5B 118.3 (3)F4A—P1A—F3A' 120.8 (13) C1B—N1B—Ru1 127.4 (3)F1A'—P1A—F3A 155.6 (11) C5B—N1B—Ru1 114.2 (2)F2A'—P1A—F3A 109.1 (12) C6B—N2B—C10B 121.1 (3)F4A—P1A—F3A 97.3 (14) C6B—N2B—Ru1 119.5 (2)F3A'—P1A—F3A 24.3 (10) C10B—N2B—Ru1 119.3 (2)F1A'—P1A—F6A' 91.3 (10) C15B—N3B—C11B 118.5 (3)F2A'—P1A—F6A' 91.2 (8) C15B—N3B—Ru1 127.2 (3)F4A—P1A—F6A' 82.5 (11) C11B—N3B—Ru1 114.1 (2)F3A'—P1A—F6A' 90.0 (10) C19B—N4B—H4B1 120.0F3A—P1A—F6A' 80.5 (12) C19B—N4B—H4B2 120.0F1A'—P1A—F1A 27.8 (10) H4B1—N4B—H4B2 120.0F2A'—P1A—F1A 67.9 (12) N1B—C1B—C2B 122.7 (3)F4A—P1A—F1A 86.0 (13) N1B—C1B—H1B 118.6F3A'—P1A—F1A 152.4 (9) C2B—C1B—H1B 118.6F3A—P1A—F1A 176.7 (12) C1B—C2B—C3B 119.2 (4)F6A'—P1A—F1A 100.7 (12) C1B—C2B—H2B 120.4F1A'—P1A—F4A' 92.7 (7) C3B—C2B—H2B 120.4F2A'—P1A—F4A' 173.2 (7) C2B—C3B—C4B 118.5 (4)F4A—P1A—F4A' 35.2 (11) C2B—C3B—H3B 120.7F3A'—P1A—F4A' 86.6 (6) C4B—C3B—H3B 120.7F3A—P1A—F4A' 64.5 (10) C5B—C4B—C3B 120.1 (4)F6A'—P1A—F4A' 89.7 (8) C5B—C4B—H4B 119.9F1A—P1A—F4A' 118.5 (11) C3B—C4B—H4B 119.9F1A'—P1A—F2A 112.4 (13) N1B—C5B—C4B 121.1 (3)F2A'—P1A—F2A 20.3 (15) N1B—C5B—C6B 115.0 (3)F4A—P1A—F2A 171.0 (14) C4B—C5B—C6B 123.9 (3)F3A'—P1A—F2A 68.1 (12) N2B—C6B—C7B 120.5 (3)F3A—P1A—F2A 91.7 (13) N2B—C6B—C5B 112.2 (3)F6A'—P1A—F2A 98.9 (11) C7B—C6B—C5B 127.3 (3)F1A—P1A—F2A 85.0 (14) C6B—C7B—C8B 120.1 (4)F4A'—P1A—F2A 153.1 (13) C6B—C7B—H7B 120.0F1A'—P1A—F6A 86.5 (17) C8B—C7B—H7B 120.0F2A'—P1A—F6A 76.5 (14) C9B—C8B—C7B 117.3 (4)F4A—P1A—F6A 93.0 (17) C9B—C8B—C16B 121.7 (4)F3A'—P1A—F6A 94.9 (17) C7B—C8B—C16B 121.0 (4)F3A—P1A—F6A 91.0 (18) C10B—C9B—C8B 120.9 (4)F6A'—P1A—F6A 15.9 (18) C10B—C9B—H9B 119.6F1A—P1A—F6A 89.5 (18) C8B—C9B—H9B 119.6F4A'—P1A—F6A 105.1 (14) N2B—C10B—C9B 120.2 (4)F2A—P1A—F6A 86.8 (17) N2B—C10B—C11B 112.3 (3)F1A'—P1A—F5A 101.4 (10) C9B—C10B—C11B 127.5 (3)F2A'—P1A—F5A 99.8 (13) N3B—C11B—C12B 120.9 (4)

Appendices
F4A—P1A—F5A 93.7 (16) N3B—C11B—C10B 115.2 (3)F3A'—P1A—F5A 77.2 (10) C12B—C11B—C10B 123.9 (4)F3A—P1A—F5A 83.0 (13) C13B—C12B—C11B 119.0 (4)F6A'—P1A—F5A 162.4 (10) C13B—C12B—H12B 120.5F1A—P1A—F5A 96.1 (13) C11B—C12B—H12B 120.5F4A'—P1A—F5A 77.7 (12) C12B—C13B—C14B 119.9 (4)F2A—P1A—F5A 87.5 (16) C12B—C13B—H13B 120.0F6A—P1A—F5A 172 (2) C14B—C13B—H13B 120.0F1A'—P1A—F5A' 86.2 (6) C15B—C14B—C13B 119.2 (4)F2A'—P1A—F5A' 90.5 (6) C15B—C14B—H14B 120.4F4A—P1A—F5A' 94.8 (9) C13B—C14B—H14B 120.4F3A'—P1A—F5A' 92.5 (6) N3B—C15B—C14B 122.6 (4)F3A—P1A—F5A' 101.2 (10) N3B—C15B—H15B 118.7F6A'—P1A—F5A' 177.0 (10) C14B—C15B—H15B 118.7F1A—P1A—F5A' 77.7 (11) C17B—C16B—C21B 117.3 (4)F4A'—P1A—F5A' 88.9 (6) C17B—C16B—C8B 121.7 (4)F2A—P1A—F5A' 83.4 (10) C21B—C16B—C8B 121.0 (4)F6A—P1A—F5A' 164.5 (13) C18B—C17B—C16B 122.0 (4)F5A—P1A—F5A' 18.6 (9) C18B—C17B—H17B 119.0N2B—Ru1—N2A 177.57 (13) C16B—C17B—H17B 119.0N2B—Ru1—N3A 99.94 (12) C17B—C18B—C19B 120.0 (4)N2A—Ru1—N3A 79.52 (13) C17B—C18B—H18B 120.0N2B—Ru1—N1A 101.75 (12) C19B—C18B—H18B 120.0N2A—Ru1—N1A 78.86 (12) N4B—C19B—C20B 122.0 (4)N3A—Ru1—N1A 158.28 (13) N4B—C19B—C18B 119.4 (4)N2B—Ru1—N3B 79.03 (12) C20B—C19B—C18B 118.6 (4)N2A—Ru1—N3B 103.32 (12) C21B—C20B—C19B 120.9 (4)N3A—Ru1—N3B 89.68 (12) C21B—C20B—H20B 119.5N1A—Ru1—N3B 93.17 (12) C19B—C20B—H20B 119.5N2B—Ru1—N1B 78.97 (12) C20B—C21B—C16B 121.2 (4)N2A—Ru1—N1B 98.68 (12) C20B—C21B—H21B 119.4N3A—Ru1—N1B 93.30 (11) C16B—C21B—H21B 119.4N1A—Ru1—N1B 92.08 (11) N1D—C1D—H1D1 109.5N3B—Ru1—N1B 157.98 (12) N1D—C1D—H1D2 109.5C1A—N1A—C5A 118.1 (3) H1D1—C1D—H1D2 109.5C1A—N1A—Ru1 127.9 (3) N1D—C1D—H1D3 109.5C5A—N1A—Ru1 114.0 (2) H1D1—C1D—H1D3 109.5C6A—N2A—C10A 122.3 (3) H1D2—C1D—H1D3 109.5C6A—N2A—Ru1 119.2 (3) N1D—C2D—H2D1 109.5C10A—N2A—Ru1 118.2 (3) N1D—C2D—H2D2 109.5C15A—N3A—C11A 119.1 (3) H2D1—C2D—H2D2 109.5C15A—N3A—Ru1 127.6 (3) N1D—C2D—H2D3 109.5C11A—N3A—Ru1 113.3 (3) H2D1—C2D—H2D3 109.5N1A—C1A—C2A 122.9 (4) H2D2—C2D—H2D3 109.5N1A—C1A—H1A 118.6 C3D—N1D—C2D 125.0 (6)C2A—C1A—H1A 118.6 C3D—N1D—C1D 118.8 (6)C3A—C2A—C1A 118.9 (4) C2D—N1D—C1D 115.0 (5)C3A—C2A—H2A 120.5 O1D—C3D—N1D 127.5 (7)

Appendices
C1A—C2A—H2A 120.5 O1D—C3D—H3D 116.2C2A—C3A—C4A 119.0 (4) N1D—C3D—H3D 116.2C2A—C3A—H3A 120.5 N1E—C2E—H2E1 109.5C4A—C3A—H3A 120.5 N1E—C2E—H2E2 109.5C3A—C4A—C5A 120.3 (4) H2E1—C2E—H2E2 109.5C3A—C4A—H4A 119.8 N1E—C2E—H2E3 109.5C5A—C4A—H4A 119.8 H2E1—C2E—H2E3 109.5N1A—C5A—C4A 120.6 (4) H2E2—C2E—H2E3 109.5N1A—C5A—C6A 115.4 (3) N1E—C1E—H1E1 109.5C4A—C5A—C6A 124.0 (4) N1E—C1E—H1E2 109.5N2A—C6A—C7A 119.7 (4) H1E1—C1E—H1E2 109.5N2A—C6A—C5A 112.4 (3) N1E—C1E—H1E3 109.5C7A—C6A—C5A 127.8 (4) H1E1—C1E—H1E3 109.5C8A—C7A—C6A 119.0 (4) H1E2—C1E—H1E3 109.5C8A—C7A—H7A 120.5 C3E—N1E—C1E 120.5 (5)C6A—C7A—H7A 120.5 C3E—N1E—C2E 122.0 (5)C7A—C8A—C9A 120.6 (4) C1E—N1E—C2E 117.4 (5)C7A—C8A—H8A 119.7 O1E—C3E—N1E 126.8 (5)C9A—C8A—H8A 119.7 O1E—C3E—H3E 116.6C10A—C9A—C8A 118.9 (4) N1E—C3E—H3E 116.6C10A—C9A—H9A 120.6 F1B—P1B—F5B 89.46 (16)C8A—C9A—H9A 120.6 F1B—P1B—F4B 90.09 (17)N2A—C10A—C9A 119.5 (4) F5B—P1B—F4B 90.71 (17)N2A—C10A—C11A 112.9 (3) F1B—P1B—F6B 90.40 (16)C9A—C10A—C11A 127.6 (4) F5B—P1B—F6B 178.85 (18)N3A—C11A—C12A 120.4 (4) F4B—P1B—F6B 90.43 (17)N3A—C11A—C10A 115.8 (3) F1B—P1B—F2B 90.13 (16)C12A—C11A—C10A 123.8 (4) F5B—P1B—F2B 90.04 (16)C13A—C12A—C11A 119.2 (4) F4B—P1B—F2B 179.22 (18)C13A—C12A—H12A 120.4 F6B—P1B—F2B 88.82 (16)C11A—C12A—H12A 120.4 F1B—P1B—F3B 179.57 (18)C14A—C13A—C12A 120.3 (4) F5B—P1B—F3B 90.97 (16)C14A—C13A—H13A 119.8 F4B—P1B—F3B 89.90 (17)C12A—C13A—H13A 119.8 F6B—P1B—F3B 89.17 (14)C13A—C14A—C15A 118.2 (4) F2B—P1B—F3B 89.88 (17)C13A—C14A—H14A 120.9
A.2 Supplementary data for X-ray diffraction analysis of Ru(II) complex 8
Table A.6. Data collection for Ru(II) complex 8
Tmin = 0.935, Tmax = 0.978 9736 independent reflections65736 measured reflections 8515 reflections with I > 2 (I)Absorption correction: Multi-scan Rint = 0.049

Appendices
Table A.7. Refinement for Ru(II) complex 8
R[F2 > 2 (F2)] = 0.059 180 restraintswR(F2) = 0.144 H atoms treated by a mixture of independent
and constrained refinementS = 1.16 max = 0.97 e Å-3
9736 reflections min = -0.60 e Å-3
814 parameters
Table A.8. Fractional atomic coordinates and isotropic or equivalent isotropic displacement parameters (Å2)
x y z U iso*/Ueq Occ. (<1)Ru1 0.48331 (4) 0.56103 (2) 0.123505 (7) 0.02499 (14)N1A 0.2711 (4) 0.5947 (2) 0.14104 (8) 0.0274 (8)N2A 0.5066 (4) 0.5149 (3) 0.16776 (8) 0.0312 (8)N3A 0.7029 (4) 0.5075 (3) 0.12376 (8) 0.0308 (8)C1A 0.1528 (5) 0.6378 (3) 0.12592 (10) 0.0289 (9)H1A 0.1607 0.6524 0.1044 0.035*C2A 0.0205 (5) 0.6612 (3) 0.14081 (11) 0.0358 (11)H2A 0.6915 0.1296 0.043*C3A 0.0063 (5) 0.6391 (3) 0.17252 (11) 0.0386 (11)H3A 0.6547 0.1830 0.046*C4A 0.1247 (5) 0.5937 (4) 0.18847 (10) 0.0379 (11)H4A 0.1163 0.5776 0.2098 0.045*C5A 0.2565 (5) 0.5718 (3) 0.17255 (10) 0.0319 (10)C6A 0.3894 (5) 0.5267 (3) 0.18784 (10) 0.0348 (10)C7A 0.4081 (6) 0.4999 (4) 0.21942 (11) 0.0478 (13)H7A 0.3274 0.5056 0.2331 0.057*C8A 0.5481 (7) 0.4644 (4) 0.23041 (12) 0.0510 (14)H8A 0.5632 0.4483 0.2519 0.061*C9A 0.6661 (7) 0.4526 (4) 0.20970 (12) 0.0471 (13)H9A 0.7606 0.4291 0.2171 0.057*C10A 0.6410 (5) 0.4764 (3) 0.17783 (11) 0.0360 (11)C11A 0.7503 (5) 0.4682 (3) 0.15227 (11) 0.0342 (10)C12A 0.8889 (5) 0.4234 (3) 0.15600 (13) 0.0405 (12)H12A 0.9175 0.3971 0.1755 0.049*C13A 0.9840 (6) 0.4179 (4) 0.13069 (14) 0.0450 (13)H13A 1.0770 0.3875 0.1328 0.054*C14A 0.9390 (6) 0.4583 (3) 0.10227 (13) 0.0420 (12)H14A 1.0026 0.4560 0.0850 0.050*C15A 0.7999 (5) 0.5024 (3) 0.09930 (11) 0.0334 (10)H15A 0.7717 0.5294 0.0799 0.040*O1B 0.1203 (5) 0.7942 (3) 0.0655 (12)O2B 0.6872 (6) 0.7083 (4) 0.0835 (15)O3B 0.7345 (5) 0.8553 (3) 0.0544 (10)N1B 0.5611 (4) 0.6855 (3) 0.13178 (8) 0.0278 (8)N2B 0.4601 (4) 0.6111 (2) 0.07960 (8) 0.0250 (8)N3B 0.3951 (4) 0.4566 (2) 0.09776 (8) 0.0271 (8)

Appendices
N4B 0.3551 (5) 0.8387 (3) 0.0422 (10)H4B 0.4333 0.8704 0.051*N5B 0.6643 (6) 0.7793 (3) 0.0515 (12)C1B 0.6103 (5) 0.7216 (3) 0.16021 (10) 0.0351 (11)H1B 0.6110 0.6878 0.1787 0.042*C2B 0.6590 (6) 0.8059 (3) 0.16289 (11) 0.0388 (11)H2B 0.6914 0.8284 0.1829 0.047*C3B 0.6594 (6) 0.8570 (4) 0.13562 (11) 0.0426 (12)H3B 0.6933 0.9140 0.1370 0.051*C4B 0.6086 (5) 0.8220 (3) 0.10620 (11) 0.0371 (11)H4B1 0.6079 0.8554 0.0876 0.045*C5B 0.5591 (5) 0.7373 (3) 0.10478 (9) 0.0287 (9)C6B 0.4986 (5) 0.6938 (3) 0.07498 (9) 0.0268 (9)C7B 0.4786 (5) 0.7313 (3) 0.04503 (9) 0.0281 (9)H7B 0.5040 0.7892 0.0422 0.034*C8B 0.4199 (5) 0.6817 (3) 0.01888 (9) 0.0268 (9)C9B 0.3820 (5) 0.5956 (3) 0.02439 (9) 0.0281 (9)H9B 0.3439 0.5613 0.0074 0.034*C10B 0.4006 (5) 0.5605 (3) 0.05516 (9) 0.0264 (9)C11B 0.3622 (5) 0.4730 (3) 0.06542 (9) 0.0267 (9)C12B 0.2963 (5) 0.4108 (3) 0.04535 (10) 0.0333 (10)H12B 0.2729 0.4233 0.0238 0.040*C13B 0.2650 (6) 0.3298 (3) 0.05741 (11) 0.0389 (11)H13B 0.2191 0.2876 0.0441 0.047*C14B 0.3025 (6) 0.3119 (3) 0.08947 (11) 0.0380 (11)H14B 0.2852 0.2573 0.0979 0.046*C15B 0.3662 (5) 0.3772 (3) 0.10872 (10) 0.0316 (10)H15B 0.3901 0.3652 0.1303 0.038*C16B 0.3999 (5) 0.7219 (3) 0.0288 (9)C17B 0.4868 (5) 0.7935 (3) 0.0342 (10)H17B 0.5591 0.8157 0.041*C18B 0.4683 (6) 0.8327 (3) 0.0383 (11)H18B 0.5260 0.8815 0.046*C19B 0.3621 (6) 0.7983 (3) 0.0358 (11)C20B 0.2744 (6) 0.7274 (3) 0.0373 (11)H20B 0.2029 0.7051 0.045*C21B 0.2926 (5) 0.6891 (3) 0.0338 (10)H21B 0.2330 0.6411 0.041*C22B 0.2419 (7) 0.8338 (4) 0.0490 (14)C23B 0.2775 (8) 0.8813 (4) 0.0587 (16)H23A 0.3642 0.9204 0.070*H23B 0.1882 0.9154 0.070*C24B 0.3177 (8) 0.8160 (4) 0.0576 (15)H24A 0.4041 0.7803 0.069*H24B 0.2294 0.7785 0.069*C25B 0.3604 (8) 0.8632 (4) 0.0581 (16)H25A 0.4530 0.8974 0.070*H25B 0.2771 0.9027 0.070*

Appendices
C26B 0.3897 (8) 0.8024 (5) 0.0612 (16)H26A 0.2958 0.7696 0.073*H26B 0.4112 0.8370 0.073*C27B 0.5226 (8) 0.7397 (4) 0.0600 (16)H27A 0.4903 0.6960 0.072*H27B 0.5441 0.7107 0.072*C28B 0.7393 (8) 0.7591 (4) 0.0606 (16)C29B 0.8819 (8) 0.8073 (5) 0.0668 (18)H29B 0.9527 0.8072 0.080*C30B 0.8959 (7) 0.8521 (4) 0.0571 (15)H30B 0.9764 0.8896 0.069*C31B 0.7618 (7) 0.8318 (4) 0.0482 (13)P1A 0.93516 (15) 0.68546 (12) 0.03687 (3) 0.0486 (4)F1A 0.8758 (3) 0.7075 (2) 0.07160 (7) 0.0564 (9)F2A 1.0358 (3) 0.6073 (2) 0.05201 (7) 0.0553 (9)F3A 1.0797 (4) 0.7481 (3) 0.04297 (9) 0.0727 (11)F4A 0.9921 (4) 0.6611 (3) 0.00219 (7) 0.0718 (12)F5A 0.8334 (4) 0.7626 (3) 0.02169 (9) 0.0751 (12)F6A 0.7905 (3) 0.6209 (3) 0.03097 (7) 0.0565 (9)P1B 0.97262 (16) 0.89067 (10) 0.23038 (3) 0.0446 (4)F1B 0.7997 (14) 0.8619 (12) 0.2363 (4) 0.066 (4) 0.56 (3)F2B 0.9861 (16) 0.8163 (10) 0.2048 (4) 0.083 (5) 0.56 (3)F3B 0.908 (3) 0.9526 (9) 0.2022 (4) 0.077 (4) 0.56 (3)F4B 1.143 (2) 0.926 (2) 0.2242 (9) 0.065 (5) 0.44 (3)F5B 0.9643 (16) 0.9714 (10) 0.2542 (4) 0.052 (4) 0.44 (3)F6B 1.041 (2) 0.8326 (8) 0.2589 (4) 0.097 (5) 0.56 (3)F1B' 0.7961 (18) 0.883 (2) 0.2357 (6) 0.094 (7) 0.44 (3)F2B' 0.965 (3) 0.7989 (11) 0.2167 (7) 0.114 (8) 0.44 (3)F3B' 0.925 (3) 0.9322 (18) 0.1977 (3) 0.088 (6) 0.44 (3)F4B' 1.1495 (18) 0.904 (2) 0.2258 (7) 0.082 (7) 0.56 (3)F5B' 0.964 (3) 0.9770 (13) 0.2491 (6) 0.121 (7) 0.56 (3)F6B' 0.999 (3) 0.861 (2) 0.2658 (3) 0.116 (7) 0.44 (3)O1D 0.7385 (5) 0.0191 (3) 0.0741 (13)N1D 0.9267 (7) 0.0640 (4) 0.03604 (15) 0.0696 (15)C1D 0.9186 (11) 0.0556 (2) 0.112 (3)H1D1 0.8688 0.0750 0.168*H1D2 1.0216 0.0607 0.168*H1D3 0.8600 0.0437 0.168*C2D 1.0363 (9) 0.1294 (5) 0.0468 (2) 0.085 (2)H2D1 1.0257 0.1788 0.0329 0.127*H2D2 1.1397 0.1068 0.0463 0.127*H2D3 1.0166 0.1462 0.0684 0.127*C3D 0.8372 (7) 0.0734 (4) 0.01008 (17) 0.0613 (16)H3D 0.8471 0.1240 0.074*O1E 0.3699 (8) 1.0700 (4) 0.11749 (15) 0.104 (2)N1E 0.2293 (9) 0.9481 (5) 0.12085 (19) 0.104 (2)C1E 0.182 (3) 0.9696 (15) 0.1514 (4) 0.313 (11)H1E1 0.2634 1.0010 0.1629 0.470*

Appendices
H1E2 0.1597 0.9177 0.1629 0.470*H1E3 0.0905 1.0049 0.1493 0.470*C2E 0.1935 (13) 0.8633 (6) 0.1096 (4) 0.168 (5)H2E1 0.0865 0.8608 0.1020 0.253*H2E2 0.2116 0.8226 0.1268 0.253*H2E3 0.2581 0.8491 0.0923 0.253*C3E 0.3286 (9) 0.9985 (5) 0.10787 (16) 0.0691 (18)H3E 0.3727 0.9782 0.0894 0.083*O1W 0.5174 (6) 0.9460 (3) 0.04109 (10) 0.0619 (11)H1W 0.600 (3) 0.974 (3) 0.0291 (10) 0.060*H2W 0.415 (2) 0.957 (4) 0.0296 (10) 0.060*
Table A.9. Atomic displacement parameters (Å2)U11 U22 U33 U12 U13 U23
Ru1 0.02127 (19) 0.0393 (2) 0.01389 (18) 0.00076 (15)N1A 0.0242 (18) 0.041 (2) 0.0164 (16)N2A 0.0287 (19) 0.045 (2) 0.0194 (17) 0.0003 (17)N3A 0.0271 (19) 0.036 (2) 0.0284 (19)C1A 0.025 (2) 0.039 (3) 0.021 (2)C2A 0.027 (2) 0.050 (3) 0.030 (2) 0.003 (2)C3A 0.026 (2) 0.056 (3) 0.034 (2) 0.002 (2) 0.0006 (18)C4A 0.035 (3) 0.058 (3) 0.021 (2) 0.0035 (18)C5A 0.030 (2) 0.048 (3) 0.0171 (19) 0.000 (2)C6A 0.031 (2) 0.050 (3) 0.023 (2) (2)C7A 0.052 (3) 0.072 (4) 0.019 (2) 0.004 (3) 0.000 (2) 0.004 (2)C8A 0.059 (3) 0.070 (4) 0.023 (2) 0.005 (3) 0.009 (2)C9A 0.049 (3) 0.057 (4) 0.033 (3) 0.004 (3) 0.006 (2)C10A 0.032 (2) 0.048 (3) 0.027 (2) 0.000 (2) (18) 0.000 (2)C11A 0.027 (2) 0.040 (3) 0.034 (2)C12A 0.030 (2) 0.041 (3) 0.049 (3) 0.004 (2) 0.004 (2)C13A 0.027 (2) 0.046 (3) 0.061 (3) 0.006 (2)C14A 0.028 (2) 0.041 (3) 0.058 (3) 0.013 (2)C15A 0.028 (2) 0.038 (3) 0.035 (2) 0.0036 (18)O1B 0.060 (3) 0.098 (4) 0.037 (2) 0.020 (2)O2B 0.096 (4) 0.101 (4) 0.054 (3) 0.004 (3) 0.011 (2) 0.039 (3)O3B 0.066 (3) 0.069 (3) 0.0280 (18) 0.003 (2) 0.0022 (16) 0.0094 (17)N1B 0.0205 (17) 0.044 (2) 0.0185 (16) 0.0029 (16)N2B 0.0195 (17) 0.037 (2) 0.0178 (16)N3B 0.0265 (18) 0.036 (2) 0.0190 (16) 0.0027 (16) 0.0019 (13)N4B 0.054 (3) 0.049 (3) 0.0234 (19) 0.0069 (17)N5B 0.063 (3) 0.057 (3) 0.034 (2) 0.003 (2) 0.003 (2) 0.009 (2)C1B 0.026 (2) 0.059 (3) 0.019 (2) 0.002 (2)C2B 0.038 (3) 0.050 (3) 0.027 (2)C3B 0.045 (3) 0.047 (3) 0.035 (3)C4B 0.037 (3) 0.048 (3) 0.026 (2)C5B 0.022 (2) 0.043 (3) 0.020 (2)C6B 0.021 (2) 0.038 (3) 0.021 (2) 0.0001 (18)

Appendices
C7B 0.025 (2) 0.040 (3) 0.019 (2) 0.0009 (19)C8B 0.022 (2) 0.040 (3) 0.0187 (19) 0.0012 (19) 0.0019 (15)C9B 0.025 (2) 0.043 (3) 0.0161 (19) 0.0015 (19)C10B 0.023 (2) 0.039 (2) 0.0178 (19) 0.0007 (19)C11B 0.021 (2) 0.040 (2) 0.0196 (19) 0.0021 (18) 0.0004 (15)C12B 0.033 (2) 0.046 (3) 0.021 (2) 0.0005 (17)C13B 0.042 (3) 0.041 (3) 0.033 (2) 0.000 (2)C14B 0.043 (3) 0.036 (3) 0.035 (2) 0.004 (2)C15B 0.032 (2) 0.038 (3) 0.025 (2) 0.001 (2) 0.0023 (17)C16B 0.029 (2) 0.038 (3) 0.0186 (19) 0.0052 (19)C17B 0.034 (2) 0.044 (3) 0.024 (2)C18B 0.044 (3) 0.045 (3) 0.026 (2) 0.0043 (19)C19B 0.043 (3) 0.043 (3) 0.022 (2) 0.005 (2) 0.0002 (18) 0.0006 (19)C20B 0.038 (3) 0.051 (3) 0.023 (2) 0.001 (2)C21B 0.036 (2) 0.042 (3) 0.022 (2) 0.0001 (19)C22B 0.055 (3) 0.063 (4) 0.029 (3) 0.000 (3) 0.001 (2) 0.007 (2)C23B 0.080 (4) 0.066 (4) 0.029 (3) 0.001 (3) 0.013 (3)C24B 0.068 (4) 0.071 (4) 0.034 (3) 0.004 (3) 0.010 (3)C25B 0.065 (4) 0.072 (4) 0.037 (3) 0.010 (3) 0.006 (3) 0.012 (3)C26B 0.065 (4) 0.081 (5) 0.039 (3) 0.005 (3) 0.004 (3)C27B 0.073 (4) 0.062 (4) 0.047 (3) 0.011 (3)C28B 0.071 (4) 0.071 (4) 0.040 (3) 0.007 (3) 0.007 (3) 0.012 (3)C29B 0.069 (4) 0.093 (5) 0.037 (3) 0.000 (4) 0.008 (3)C30B 0.059 (4) 0.073 (4) 0.039 (3) 0.006 (3)C31B 0.057 (3) 0.060 (4) 0.028 (3) 0.003 (3) 0.002 (2) 0.005 (2)P1A 0.0273 (6) 0.0907 (12) 0.0276 (6) 0.0016 (7) 0.0096 (7)F1A 0.0407 (16) 0.095 (3) 0.0334 (15) 0.0078 (17) 0.0015 (12)F2A 0.0363 (16) 0.094 (3) 0.0347 (15) 0.0112 (17) 0.0000 (16)F3A 0.0401 (18) 0.101 (3) 0.077 (2) 0.006 (2)F4A 0.0397 (17) 0.149 (4) 0.0276 (15) 0.0069 (12) 0.0082 (19)F5A 0.0459 (19) 0.109 (3) 0.070 (2) 0.011 (2) 0.038 (2)F6A 0.0336 (15) 0.105 (3) 0.0303 (15)P1B 0.0431 (7) 0.0605 (9) 0.0292 (6)F1B 0.065 (6) 0.095 (8) 0.037 (6) 0.007 (5)F2B 0.053 (5) 0.088 (7) 0.104 (9) 0.019 (5)F3B 0.073 (7) 0.052 (6) 0.100 (10) 0.016 (5)F4B 0.037 (7) 0.097 (11) 0.059 (10) 0.002 (6)F5B 0.036 (6) 0.075 (7) 0.046 (6) 0.007 (4)F6B 0.108 (10) 0.053 (6) 0.120 (10) 0.035 (5)F1B' 0.059 (8) 0.160 (18) 0.065 (10) 0.027 (7)F2B' 0.133 (14) 0.086 (8) 0.117 (15)F3B' 0.073 (9) 0.168 (16) 0.020 (5) 0.023 (7)F4B' 0.041 (6) 0.149 (19) 0.056 (10)F5B' 0.136 (13) 0.120 (11) 0.107 (12) 0.017 (9)F6B' 0.128 (14) 0.176 (17) 0.039 (6) 0.034 (8)O1D 0.056 (3) 0.073 (3) 0.094 (4) (2) 0.003 (2) 0.001 (3)N1D 0.059 (3) 0.074 (4) 0.076 (4) 0.002 (3) 0.010 (3)C1D 0.103 (7) 0.108 (7) 0.122 (8) 0.053 (6)

Appendices
C2D 0.062 (4) 0.086 (6) 0.105 (6) 0.003 (4) 0.001 (5)C3D 0.050 (3) 0.058 (4) 0.078 (4) 0.010 (3)O1E 0.118 (5) 0.099 (4) 0.097 (4) 0.031 (4)N1E 0.116 (6) 0.088 (5) 0.113 (5) 0.052 (4)C1E 0.42 (2) 0.33 (2) 0.218 (16) 0.233 (17)C2E 0.106 (8) 0.065 (6) 0.332 (17) 0.002 (8)C3E 0.075 (4) 0.077 (5) 0.056 (4) 0.005 (3)O1W 0.078 (3) 0.057 (3) 0.051 (2) 0.003 (2) 0.000 (2)
Table A.10. Geometric parameters (Å, o)Ru1—N2A 1.976 (4) C14B—H14B 0.9300Ru1—N2B 1.985 (3) C15B—H15B 0.9300Ru1—N3B 2.060 (4) C16B—C17B 1.386 (7)Ru1—N1B 2.061 (4) C16B—C21B 1.406 (6)Ru1—N3A 2.068 (4) C17B—C18B 1.387 (6)Ru1—N1A 2.076 (4) C17B—H17B 0.9300N1A—C1A 1.346 (5) C18B—C19B 1.399 (7)N1A—C5A 1.371 (5) C18B—H18B 0.9300N2A—C10A 1.350 (6) C19B—C20B 1.374 (7)N2A—C6A 1.361 (6) C20B—C21B 1.389 (6)N3A—C15A 1.356 (6) C20B—H20B 0.9300N3A—C11A 1.376 (6) C21B—H21B 0.9300C1A—C2A 1.378 (6) C22B—C23B 1.524 (7)C1A—H1A 0.9300 C23B—C24B 1.539 (9)C2A—C3A 1.376 (7) C23B—H23A 0.9700C2A—H2A 0.9300 C23B—H23B 0.9700C3A—C4A 1.379 (7) C24B—C25B 1.531 (7)C3A—H3A 0.9300 C24B—H24A 0.9700C4A—C5A 1.391 (6) C24B—H24B 0.9700C4A—H4A 0.9300 C25B—C26B 1.536 (8)C5A—C6A 1.458 (6) C25B—H25A 0.9700C6A—C7A 1.380 (6) C25B—H25B 0.9700C7A—C8A 1.382 (8) C26B—C27B 1.516 (9)C7A—H7A 0.9300 C26B—H26A 0.9700C8A—C9A 1.383 (8) C26B—H26B 0.9700C8A—H8A 0.9300 C27B—H27A 0.9700C9A—C10A 1.382 (7) C27B—H27B 0.9700C9A—H9A 0.9300 C28B—C29B 1.438 (9)C10A—C11A 1.467 (7) C29B—C30B 1.332 (8)C11A—C12A 1.383 (7) C29B—H29B 0.9300C12A—C13A 1.375 (7) C30B—C31B 1.471 (8)C12A—H12A 0.9300 C30B—H30B 0.9300C13A—C14A 1.375 (8) P1A—F3A 1.588 (4)C13A—H13A 0.9300 P1A—F5A 1.591 (4)C14A—C15A 1.380 (7) P1A—F4A 1.597 (3)C14A—H14A 0.9300 P1A—F2A 1.597 (4)C15A—H15A 0.9300 P1A—F1A 1.598 (3)

Appendices
O1B—C22B 1.225 (7) P1A—F6A 1.606 (4)O2B—C28B 1.229 (7) P1B—F2B' 1.530 (15)O3B—C31B 1.213 (6) P1B—F3B' 1.540 (12)N1B—C1B 1.355 (5) P1B—F5B' 1.550 (14)N1B—C5B 1.380 (6) P1B—F6B' 1.552 (12)N2B—C6B 1.339 (6) P1B—F1B' 1.557 (14)N2B—C10B 1.361 (5) P1B—F4B' 1.564 (13)N3B—C15B 1.338 (6) P1B—F2B 1.578 (8)N3B—C11B 1.384 (5) P1B—F6B 1.579 (10)N4B—C22B 1.341 (7) P1B—F3B 1.589 (11)N4B—C19B 1.416 (6) P1B—F1B 1.590 (10)N4B—H4B 0.8600 P1B—F5B 1.598 (11)N5B—C31B 1.387 (7) P1B—F4B 1.601 (14)N5B—C28B 1.394 (7) O1D—C3D 1.254 (8)N5B—C27B 1.447 (8) N1D—C3D 1.301 (8)C1B—C2B 1.372 (7) N1D—C2D 1.438 (9)C1B—H1B 0.9300 N1D—C1D 1.466 (10)C2B—C3B 1.384 (7) C1D—H1D1 0.9600C2B—H2B 0.9300 C1D—H1D2 0.9600C3B—C4B 1.387 (6) C1D—H1D3 0.9600C3B—H3B 0.9300 C2D—H2D1 0.9600C4B—C5B 1.377 (7) C2D—H2D2 0.9600C4B—H4B1 0.9300 C2D—H2D3 0.9600C5B—C6B 1.480 (5) C3D—H3D 0.9300C6B—C7B 1.377 (6) O1E—C3E 1.221 (9)C7B—C8B 1.403 (6) N1E—C3E 1.299 (9)C7B—H7B 0.9300 N1E—C1E 1.398 (13)C8B—C9B 1.393 (7) N1E—C2E 1.420 (10)C8B—C16B 1.485 (6) C1E—H1E1 0.9600C9B—C10B 1.392 (6) C1E—H1E2 0.9600C9B—H9B 0.9300 C1E—H1E3 0.9600C10B—C11B 1.463 (6) C2E—H2E1 0.9600C11B—C12B 1.375 (6) C2E—H2E2 0.9600C12B—C13B 1.382 (7) C2E—H2E3 0.9600C12B—H12B 0.9300 C3E—H3E 0.9300C13B—C14B 1.384 (7) O1W—H1W 0.993 (2)C13B—H13B 0.9300 O1W—H2W 0.993 (2)C14B—C15B 1.383 (6)
N2A—Ru1—N2B 178.23 (15) H23A—C23B—H23B 108.2N2A—Ru1—N3B 102.57 (15) C25B—C24B—C23B 110.5 (5)N2B—Ru1—N3B 79.10 (14) C25B—C24B—H24A 109.5N2A—Ru1—N1B 99.56 (15) C23B—C24B—H24A 109.5N2B—Ru1—N1B 78.77 (14) C25B—C24B—H24B 109.5N3B—Ru1—N1B 157.87 (13) C23B—C24B—H24B 109.5N2A—Ru1—N3A 78.79 (15) H24A—C24B—H24B 108.1N2B—Ru1—N3A 101.87 (14) C24B—C25B—C26B 113.7 (6)N3B—Ru1—N3A 90.05 (14) C24B—C25B—H25A 108.8N1B—Ru1—N3A 94.73 (14) C26B—C25B—H25A 108.8

Appendices
N2A—Ru1—N1A 78.93 (15) C24B—C25B—H25B 108.8N2B—Ru1—N1A 100.42 (14) C26B—C25B—H25B 108.8N3B—Ru1—N1A 94.12 (14) H25A—C25B—H25B 107.7N1B—Ru1—N1A 89.60 (14) C27B—C26B—C25B 114.1 (5)N3A—Ru1—N1A 157.71 (14) C27B—C26B—H26A 108.7C1A—N1A—C5A 117.9 (4) C25B—C26B—H26A 108.7C1A—N1A—Ru1 128.4 (3) C27B—C26B—H26B 108.7C5A—N1A—Ru1 113.7 (3) C25B—C26B—H26B 108.7C10A—N2A—C6A 121.6 (4) H26A—C26B—H26B 107.6C10A—N2A—Ru1 119.3 (3) N5B—C27B—C26B 114.4 (5)C6A—N2A—Ru1 119.0 (3) N5B—C27B—H27A 108.7C15A—N3A—C11A 117.1 (4) C26B—C27B—H27A 108.7C15A—N3A—Ru1 128.6 (3) N5B—C27B—H27B 108.7C11A—N3A—Ru1 114.2 (3) C26B—C27B—H27B 108.7N1A—C1A—C2A 123.1 (4) H27A—C27B—H27B 107.6N1A—C1A—H1A 118.5 O2B—C28B—N5B 123.6 (6)C2A—C1A—H1A 118.5 O2B—C28B—C29B 128.6 (6)C3A—C2A—C1A 119.2 (4) N5B—C28B—C29B 107.8 (5)C3A—C2A—H2A 120.4 C30B—C29B—C28B 108.7 (5)C1A—C2A—H2A 120.4 C30B—C29B—H29B 125.6C2A—C3A—C4A 119.0 (4) C28B—C29B—H29B 125.6C2A—C3A—H3A 120.5 C29B—C30B—C31B 108.1 (6)C4A—C3A—H3A 120.5 C29B—C30B—H30B 125.9C3A—C4A—C5A 119.8 (4) C31B—C30B—H30B 125.9C3A—C4A—H4A 120.1 O3B—C31B—N5B 124.2 (5)C5A—C4A—H4A 120.1 O3B—C31B—C30B 129.1 (5)N1A—C5A—C4A 121.0 (4) N5B—C31B—C30B 106.7 (4)N1A—C5A—C6A 115.4 (4) F3A—P1A—F5A 91.0 (2)C4A—C5A—C6A 123.5 (4) F3A—P1A—F4A 90.5 (2)N2A—C6A—C7A 119.6 (4) F5A—P1A—F4A 90.5 (2)N2A—C6A—C5A 112.9 (4) F3A—P1A—F2A 89.6 (2)C7A—C6A—C5A 127.4 (4) F5A—P1A—F2A 179.3 (2)C6A—C7A—C8A 119.2 (5) F4A—P1A—F2A 89.5 (2)C6A—C7A—H7A 120.4 F3A—P1A—F1A 90.8 (2)C8A—C7A—H7A 120.4 F5A—P1A—F1A 90.1 (2)C7A—C8A—C9A 120.4 (5) F4A—P1A—F1A 178.5 (2)C7A—C8A—H8A 119.8 F2A—P1A—F1A 89.97 (18)C9A—C8A—H8A 119.8 F3A—P1A—F6A 179.1 (2)C10A—C9A—C8A 118.9 (5) F5A—P1A—F6A 89.8 (2)C10A—C9A—H9A 120.6 F4A—P1A—F6A 89.50 (19)C8A—C9A—H9A 120.6 F2A—P1A—F6A 89.5 (2)N2A—C10A—C9A 120.0 (5) F1A—P1A—F6A 89.15 (17)N2A—C10A—C11A 112.9 (4) F2B'—P1B—F3B' 93.0 (12)C9A—C10A—C11A 127.0 (5) F2B'—P1B—F5B' 170.1 (19)N3A—C11A—C12A 122.1 (4) F3B'—P1B—F5B' 93.9 (17)N3A—C11A—C10A 114.6 (4) F2B'—P1B—F6B' 94.5 (11)C12A—C11A—C10A 123.3 (4) F3B'—P1B—F6B' 169.3 (14)C13A—C12A—C11A 119.7 (5) F5B'—P1B—F6B' 77.7 (19)

Appendices
C13A—C12A—H12A 120.2 F2B'—P1B—F1B' 87.7 (14)C11A—C12A—H12A 120.2 F3B'—P1B—F1B' 86.7 (14)C12A—C13A—C14A 118.7 (5) F5B'—P1B—F1B' 85.7 (14)C12A—C13A—H13A 120.7 F6B'—P1B—F1B' 86.1 (13)C14A—C13A—H13A 120.7 F2B'—P1B—F4B' 95.7 (18)C13A—C14A—C15A 120.2 (5) F3B'—P1B—F4B' 92.9 (15)C13A—C14A—H14A 119.9 F5B'—P1B—F4B' 91.0 (14)C15A—C14A—H14A 119.9 F6B'—P1B—F4B' 93.8 (16)N3A—C15A—C14A 122.2 (5) F1B'—P1B—F4B' 176.6 (18)N3A—C15A—H15A 118.9 F2B'—P1B—F2B 22.2 (13)C14A—C15A—H15A 118.9 F3B'—P1B—F2B 74.5 (11)C1B—N1B—C5B 117.5 (4) F5B'—P1B—F2B 167.3 (15)C1B—N1B—Ru1 127.9 (3) F6B'—P1B—F2B 114.4 (12)C5B—N1B—Ru1 114.6 (3) F1B'—P1B—F2B 98.5 (11)C6B—N2B—C10B 121.9 (4) F4B'—P1B—F2B 84.6 (13)C6B—N2B—Ru1 119.5 (3) F2B'—P1B—F6B 76.1 (11)C10B—N2B—Ru1 118.6 (3) F3B'—P1B—F6B 166.8 (11)C15B—N3B—C11B 117.9 (4) F5B'—P1B—F6B 97.8 (14)C15B—N3B—Ru1 127.7 (3) F6B'—P1B—F6B 23.8 (13)C11B—N3B—Ru1 114.4 (3) F1B'—P1B—F6B 100.2 (12)C22B—N4B—C19B 128.3 (5) F4B'—P1B—F6B 81.0 (16)C22B—N4B—H4B 115.9 F2B—P1B—F6B 93.2 (7)C19B—N4B—H4B 115.9 F2B'—P1B—F3B 106.2 (11)C31B—N5B—C28B 108.2 (5) F3B'—P1B—F3B 14.5 (14)C31B—N5B—C27B 125.5 (5) F5B'—P1B—F3B 80.1 (13)C28B—N5B—C27B 125.4 (5) F6B'—P1B—F3B 155.2 (14)N1B—C1B—C2B 122.8 (4) F1B'—P1B—F3B 81.2 (13)N1B—C1B—H1B 118.6 F4B'—P1B—F3B 97.5 (16)C2B—C1B—H1B 118.6 F2B—P1B—F3B 88.7 (8)C1B—C2B—C3B 119.4 (4) F6B—P1B—F3B 177.4 (9)C1B—C2B—H2B 120.3 F2B'—P1B—F1B 77.3 (14)C3B—C2B—H2B 120.3 F3B'—P1B—F1B 92.6 (11)C2B—C3B—C4B 119.1 (5) F5B'—P1B—F1B 95.4 (11)C2B—C3B—H3B 120.4 F6B'—P1B—F1B 81.7 (11)C4B—C3B—H3B 120.4 F1B'—P1B—F1B 11.8 (17)C5B—C4B—C3B 119.4 (5) F4B'—P1B—F1B 171.3 (16)C5B—C4B—H4B1 120.3 F2B—P1B—F1B 90.4 (8)C3B—C4B—H4B1 120.3 F6B—P1B—F1B 92.2 (9)C4B—C5B—N1B 121.8 (4) F3B—P1B—F1B 89.5 (10)C4B—C5B—C6B 124.2 (4) F2B'—P1B—F5B 162.5 (16)N1B—C5B—C6B 114.0 (4) F3B'—P1B—F5B 101.8 (14)N2B—C6B—C7B 120.8 (4) F5B'—P1B—F5B 8.2 (17)N2B—C6B—C5B 113.1 (4) F6B'—P1B—F5B 69.6 (16)C7B—C6B—C5B 126.1 (4) F1B'—P1B—F5B 84.0 (11)C6B—C7B—C8B 119.7 (4) F4B'—P1B—F5B 92.8 (12)C6B—C7B—H7B 120.2 F2B—P1B—F5B 175.3 (11)C8B—C7B—H7B 120.2 F6B—P1B—F5B 90.3 (10)C9B—C8B—C7B 118.2 (4) F3B—P1B—F5B 87.8 (10)

Appendices
C9B—C8B—C16B 122.2 (4) F1B—P1B—F5B 92.7 (9)C7B—C8B—C16B 119.7 (4) F2B'—P1B—F4B 106 (2)C10B—C9B—C8B 120.5 (4) F3B'—P1B—F4B 85.5 (17)C10B—C9B—H9B 119.7 F5B'—P1B—F4B 81.9 (17)C8B—C9B—H9B 119.7 F6B'—P1B—F4B 99.7 (17)N2B—C10B—C9B 118.9 (4) F1B'—P1B—F4B 165 (2)N2B—C10B—C11B 113.3 (3) F4B'—P1B—F4B 12 (3)C9B—C10B—C11B 127.8 (4) F2B—P1B—F4B 91.9 (15)C12B—C11B—N3B 121.2 (4) F6B—P1B—F4B 90.3 (16)C12B—C11B—C10B 124.2 (4) F3B—P1B—F4B 87.9 (16)N3B—C11B—C10B 114.6 (4) F1B—P1B—F4B 176.5 (16)C11B—C12B—C13B 119.7 (4) F5B—P1B—F4B 84.9 (15)C11B—C12B—H12B 120.2 C3D—N1D—C2D 122.0 (6)C13B—C12B—H12B 120.2 C3D—N1D—C1D 120.6 (7)C12B—C13B—C14B 119.4 (4) C2D—N1D—C1D 117.5 (7)C12B—C13B—H13B 120.3 N1D—C1D—H1D1 109.5C14B—C13B—H13B 120.3 N1D—C1D—H1D2 109.5C15B—C14B—C13B 118.5 (5) H1D1—C1D—H1D2 109.5C15B—C14B—H14B 120.7 N1D—C1D—H1D3 109.5C13B—C14B—H14B 120.7 H1D1—C1D—H1D3 109.5N3B—C15B—C14B 123.2 (4) H1D2—C1D—H1D3 109.5N3B—C15B—H15B 118.4 N1D—C2D—H2D1 109.5C14B—C15B—H15B 118.4 N1D—C2D—H2D2 109.5C17B—C16B—C21B 118.0 (4) H2D1—C2D—H2D2 109.5C17B—C16B—C8B 120.6 (4) N1D—C2D—H2D3 109.5C21B—C16B—C8B 121.5 (4) H2D1—C2D—H2D3 109.5C16B—C17B—C18B 121.5 (4) H2D2—C2D—H2D3 109.5C16B—C17B—H17B 119.3 O1D—C3D—N1D 124.8 (7)C18B—C17B—H17B 119.3 O1D—C3D—H3D 117.6C17B—C18B—C19B 119.5 (5) N1D—C3D—H3D 117.6C17B—C18B—H18B 120.3 C3E—N1E—C1E 118.0 (10)C19B—C18B—H18B 120.3 C3E—N1E—C2E 123.6 (9)C20B—C19B—C18B 120.1 (4) C1E—N1E—C2E 116.8 (12)C20B—C19B—N4B 123.3 (4) N1E—C1E—H1E1 109.5C18B—C19B—N4B 116.5 (4) N1E—C1E—H1E2 109.5C19B—C20B—C21B 120.0 (4) H1E1—C1E—H1E2 109.5C19B—C20B—H20B 120.0 N1E—C1E—H1E3 109.5C21B—C20B—H20B 120.0 H1E1—C1E—H1E3 109.5C20B—C21B—C16B 121.0 (4) H1E2—C1E—H1E3 109.5C20B—C21B—H21B 119.5 N1E—C2E—H2E1 109.5C16B—C21B—H21B 119.5 N1E—C2E—H2E2 109.5O1B—C22B—N4B 124.0 (5) H2E1—C2E—H2E2 109.5O1B—C22B—C23B 121.8 (5) N1E—C2E—H2E3 109.5N4B—C22B—C23B 114.1 (5) H2E1—C2E—H2E3 109.5C22B—C23B—C24B 110.0 (5) H2E2—C2E—H2E3 109.5C22B—C23B—H23A 109.7 O1E—C3E—N1E 126.4 (7)C24B—C23B—H23A 109.7 O1E—C3E—H3E 116.8C22B—C23B—H23B 109.7 N1E—C3E—H3E 116.8

Appendices
C24B—C23B—H23B 109.7 H1W—O1W—H2W 109.4 (3)
N2A—Ru1—N1A—C1A 178.9 (4) N3A—Ru1—N3B—C15B 76.8 (4)N2B—Ru1—N1A—C1A 0.6 (4) N1A—Ru1—N3B—C15BN3B—Ru1—N1A—C1A N2A—Ru1—N3B—C11B 177.8 (3)N1B—Ru1—N1A—C1A 79.1 (4) N2B—Ru1—N3B—C11BN3A—Ru1—N1A—C1A N1B—Ru1—N3B—C11BN2A—Ru1—N1A—C5A 1.7 (3) N3A—Ru1—N3B—C11BN2B—Ru1—N1A—C5A N1A—Ru1—N3B—C11B 98.2 (3)N3B—Ru1—N1A—C5A 103.7 (3) C5B—N1B—C1B—C2BN1B—Ru1—N1A—C5A Ru1—N1B—C1B—C2BN3A—Ru1—N1A—C5A 3.4 (6) N1B—C1B—C2B—C3BN2B—Ru1—N2A—C10A C1B—C2B—C3B—C4B 0.8 (8)N3B—Ru1—N2A—C10A 89.7 (4) C2B—C3B—C4B—C5BN1B—Ru1—N2A—C10A C3B—C4B—C5B—N1BN3A—Ru1—N2A—C10A 2.2 (4) C3B—C4B—C5B—C6B 178.3 (4)N1A—Ru1—N2A—C10A C1B—N1B—C5B—C4B 1.6 (6)N2B—Ru1—N2A—C6A 67 (5) Ru1—N1B—C5B—C4BN3B—Ru1—N2A—C6A C1B—N1B—C5B—C6BN1B—Ru1—N2A—C6A 85.9 (4) Ru1—N1B—C5B—C6B 1.0 (4)N3A—Ru1—N2A—C6A 178.8 (4) C10B—N2B—C6B—C7B 0.3 (6)N1A—Ru1—N2A—C6A Ru1—N2B—C6B—C7BN2A—Ru1—N3A—C15A 177.1 (4) C10B—N2B—C6B—C5B 179.2 (4)N2B—Ru1—N3A—C15A Ru1—N2B—C6B—C5B 1.3 (5)N3B—Ru1—N3A—C15A 74.3 (4) C4B—C5B—C6B—N2B 179.0 (4)N1B—Ru1—N3A—C15A N1B—C5B—C6B—N2BN1A—Ru1—N3A—C15A 175.3 (4) C4B—C5B—C6B—C7BN2A—Ru1—N3A—C11A 1.1 (3) N1B—C5B—C6B—C7B 177.4 (4)N2B—Ru1—N3A—C11A 179.4 (3) N2B—C6B—C7B—C8BN3B—Ru1—N3A—C11A C5B—C6B—C7B—C8B 180.0 (4)N1B—Ru1—N3A—C11A 99.9 (3) C6B—C7B—C8B—C9B 0.7 (6)N1A—Ru1—N3A—C11A C6B—C7B—C8B—C16BC5A—N1A—C1A—C2A 1.4 (7) C7B—C8B—C9B—C10B 0.7 (6)Ru1—N1A—C1A—C2A C16B—C8B—C9B—C10BN1A—C1A—C2A—C3A C6B—N2B—C10B—C9B 1.1 (6)C1A—C2A—C3A—C4A Ru1—N2B—C10B—C9B 179.1 (3)C2A—C3A—C4A—C5A 0.8 (8) C6B—N2B—C10B—C11BC1A—N1A—C5A—C4A Ru1—N2B—C10B—C11BRu1—N1A—C5A—C4A 176.6 (4) C8B—C9B—C10B—N2BC1A—N1A—C5A—C6A C8B—C9B—C10B—C11B 177.8 (4)Ru1—N1A—C5A—C6A C15B—N3B—C11B—C12B 2.5 (6)C3A—C4A—C5A—N1A Ru1—N3B—C11B—C12BC3A—C4A—C5A—C6A 177.7 (5) C15B—N3B—C11B—C10BC10A—N2A—C6A—C7A 0.8 (7) Ru1—N3B—C11B—C10B 1.9 (4)Ru1—N2A—C6A—C7A N2B—C10B—C11B—C12B 177.9 (4)C10A—N2A—C6A—C5A 178.2 (4) C9B—C10B—C11B—C12BRu1—N2A—C6A—C5A 1.6 (6) N2B—C10B—C11B—N3BN1A—C5A—C6A—N2A C9B—C10B—C11B—N3B 179.5 (4)C4A—C5A—C6A—N2A N3B—C11B—C12B—C13B

Appendices
N1A—C5A—C6A—C7A 177.0 (5) C10B—C11B—C12B—C13B 179.8 (4)C4A—C5A—C6A—C7A C11B—C12B—C13B—C14BN2A—C6A—C7A—C8A 2.5 (8) C12B—C13B—C14B—C15B 1.9 (7)C5A—C6A—C7A—C8A C11B—N3B—C15B—C14BC6A—C7A—C8A—C9A Ru1—N3B—C15B—C14B 178.1 (3)C7A—C8A—C9A—C10A C13B—C14B—C15B—N3BC6A—N2A—C10A—C9A C9B—C8B—C16B—C17BRu1—N2A—C10A—C9A 172.6 (4) C7B—C8B—C16B—C17B 23.9 (6)C6A—N2A—C10A—C11A 178.7 (4) C9B—C8B—C16B—C21B 24.2 (6)Ru1—N2A—C10A—C11A C7B—C8B—C16B—C21BC8A—C9A—C10A—N2A 3.7 (8) C21B—C16B—C17B—C18B 0.7 (7)C8A—C9A—C10A—C11A C8B—C16B—C17B—C18BC15A—N3A—C11A—C12A C16B—C17B—C18B—C19BRu1—N3A—C11A—C12A 175.1 (4) C17B—C18B—C19B—C20B 1.5 (7)C15A—N3A—C11A—C10A 179.7 (4) C17B—C18B—C19B—N4BRu1—N3A—C11A—C10A C22B—N4B—C19B—C20B 19.8 (8)N2A—C10A—C11A—N3A 5.5 (6) C22B—N4B—C19B—C18BC9A—C10A—C11A—N3A (5) C18B—C19B—C20B—C21BN2A—C10A—C11A—C12A N4B—C19B—C20B—C21B 177.3 (5)C9A—C10A—C11A—C12A 9.4 (8) C19B—C20B—C21B—C16B 0.1 (7)N3A—C11A—C12A—C13A 0.5 (8) C17B—C16B—C21B—C20B 0.0 (7)C10A—C11A—C12A—C13A 179.2 (5) C8B—C16B—C21B—C20B 179.7 (4)
C11A—C12A—C13A—C14A 0.7 (8) C19B—N4B—C22B—O1B 3.0 (10)
C12A—C13A—C14A—C15A C19B—N4B—C22B—C23B
C11A—N3A—C15A—C14A 1.3 (7) O1B—C22B—C23B—C24BRu1—N3A—C15A—C14A N4B—C22B—C23B—C24B 105.3 (6)C13A—C14A—C15A—N3A C22B—C23B—C24B—C25BN2A—Ru1—N1B—C1B C23B—C24B—C25B—C26BN2B—Ru1—N1B—C1B 178.5 (4) C24B—C25B—C26B—C27BN3B—Ru1—N1B—C1B 177.8 (3) C31B—N5B—C27B—C26BN3A—Ru1—N1B—C1B C28B—N5B—C27B—C26B 120.3 (6)N1A—Ru1—N1B—C1B 77.8 (4) C25B—C26B—C27B—N5BN2A—Ru1—N1B—C5B C31B—N5B—C28B—O2BN2B—Ru1—N1B—C5B C27B—N5B—C28B—O2BN3B—Ru1—N1B—C5B C31B—N5B—C28B—C29B 5.4 (7)N3A—Ru1—N1B—C5B 100.9 (3) C27B—N5B—C28B—C29B 175.1 (6)N1A—Ru1—N1B—C5B O2B—C28B—C29B—C30B 177.6 (7)N2A—Ru1—N2B—C6B 19 (5) N5B—C28B—C29B—C30BN3B—Ru1—N2B—C6B 179.1 (3) C28B—C29B—C30B—C31BN1B—Ru1—N2B—C6B C28B—N5B—C31B—O3B 174.5 (6)N3A—Ru1—N2B—C6B C27B—N5B—C31B—O3B 4.8 (10)N1A—Ru1—N2B—C6B 86.9 (3) C28B—N5B—C31B—C30BN2A—Ru1—N2B—C10B C27B—N5B—C31B—C30BN3B—Ru1—N2B—C10B 1.1 (3) C29B—C30B—C31B—O3BN1B—Ru1—N2B—C10B C29B—C30B—C31B—N5B 5.1 (7)

Appendices
N3A—Ru1—N2B—C10B 88.9 (3) C2D—N1D—C3D—O1DN1A—Ru1—N2B—C10B C1D—N1D—C3D—O1D 0.7 (11)N2A—Ru1—N3B—C15B C1E—N1E—C3E—O1EN2B—Ru1—N3B—C15B 178.9 (4) C2E—N1E—C3E—O1EN1B—Ru1—N3B—C15B 179.6 (3)

Appendices
Appendix B
Estimation of dimer bioconjugate yield by gel electrophoresis
A typical calculation for the dimer bioconjugate example cyt c-10-cyt c. After collection
of product band containing a mixture of unreacted protein, dimer and monomer
bioconjugate (determined by MALDI-TOF mass spectrometry), the concentration of
mixture was estimated by Beer-Lambert law,
[Proteincyt c] = 0.841 a.u./(97.6 mM-1cm-1 1 cm) (cyt c: 410 = 97.6 mM-1cm-1)
= 8.62 10-3 mM
Amount of mixture = 8.62 10-3 mM 220 L
= 1.89 10-9 mol
To estimate the fraction of dimer in the mixture, gel electrophoresis was performed to
separate the components. The estimation of relative concentration of bands within the
lane was determined using the ‘gel analysis’ function in ImageJ v1.42q as shown in
Figure B.1.
Figure B.1. Estimated relative composition of bands in purified bioconjugate mixture cyt c-10-cyt c.
Therefore, amount of dimer = composition amount of mixture
= 0.119 1.89 10-9 mol
= 2.25 10-10 mol
Yield = (amount of dimer/starting ligand concentration) 100%

Appendices
= (2.25 10-10 mol/3.1325 10-10 mol) 100%
= 0.7%
1%

Appendices
Appendix C
Standard nitrite curve using the Griess assay
Table C.1. Nitrite anion standard curved based on absorbance using Griess assay.Nitrite concentration / M Average absorbance /a.u.a
0 0.0032±0.00261 0.0386±0.00182 0.0765±0.00023 0.1118±0.00074 0.1488±0.00155 0.1950±0.00566 0.2111±0.0020
a Monitored absorbance at 540 nm. Error bars are standard deviation.
Figure C.1. Standard nitrite anion concentration standard curve.

Appendices
Appendix D
Theoretical enzyme encapsulation efficiency of egg PC liposomes
In a representative calculation for egg L- -phosphatidylcholine liposomes,
Areahead = 69 Å2 (=0.69 nm2)1 and vesicle d = 105 nm
Surface areavesicle = 4 r2 = 4 (105/2)2 = 34636.0 nm2
Therefore, number of phospholipid molecules = surface areavesicle/Areahead 2 because
of a bilayer.
= (34636.0/0.69) 2 molecules/vesicle
= 100394.2 molecules/vesicle (MW of egg PC = 770.12 g/mol)
Hence, one vesicle = number of phospholipid molecules/NA
= 100394.2/6.023 1023 mol
= 1.67 10-19 mol
Stock (100 L): 100 M iso-1 cyt c = 1 10-8 mol; 15 mg/mL egg PC = 1.5 mg
Moles of phospholipid = mass of lipid/MWegg PC
= 1.5 10-3/770.12 mol
= 1.95 10-6 mol
Number of vesicles = moles of total phospholipid/moles of phospholipid for one vesicle
= 1.95 10-6 mol/1.67 10-19 mol
= 1.16 1013 vesicles
Volumevesicle = (4/3) r3 = 606131 nm3
= 6.06 10-16 cm3
= 6.06 10-16 mL
Therefore, total lumen volume = volumevesicle number of vesicles
= 6.06 10-16 mL 1.16 1013 vesicles
= 7.03 10-6 L

Appendices
[iso-1 cyt c]lumen = 100 M
Therefore, total proteins in lumen = [iso-1 cyt c]lumen total lumen volume
= 100 10-6 M 7.03 10-6 L
= 7.03 10-10 mol
Hence, encapsulation efficiency (%) = proteins in lumen/proteins total
= 7.03 10-10 mol/1 10-8 mol 100%
= 7.0%
Determination of encapsulation efficiency via fluorescence
With the following representative calculation, the encapsulation efficiency (EE%) was
determined for dye labelled cytochrome c and amGFP.
EE% = total number of enzymes encapsulated/number of enzymes added
For 5 M of cyt c added for a 200 L buffer solution:
ncyt c added = (200 × 10-6 L) × (5 × 10-6 M) = 1 × 10-9 mol
Now, enzymes encapsulated determined by estimating total fluorescence of sample after
dialysis; for 1 a.u. fluorescence emission intensity ( ex = 488 nm, em= 513 nm),
equivalent to 4.2 × 10-3 M cyt c-dye (determined experimentally). Therefore, since
sample has fluorescence intensity of 849 a.u. in PBS (166 L) after dialysis,
nencapsulated = 849 × (4.2 × 10-9 M) × (166 × 10-6 L) = 5.9 × 10-10 mol
EE% = (nencapsulated/ncyt c added) × 100 = (5.9 × 10-10/1 × 10-9) × 100 = 59%

Appendices
Appendix E
pH titration of 8-hydroxypyrene-1,3,6-trisulfonic acid (HPTS)
Figure E.1. pH titration showing the changes in the excitation spectra ( em = 510 nm) of pyranine encapsulated in a diluted (0.003 mg/mL) PS140-b-PAA48 polymer micelles in 20 mM sodium dihydrogen phosphate with 20 mM ethylenediaminetetraacetic acid. Legend corresponds to pH values.

Appendices
Photoreduction scattering by EDTA by-products of HPTS
Figure E.2. Photoreduction scattering from EDTA by-products under various conditions. (a) Excitation spectra of HPTS encapsulated in a poly-L-lysine: PS140-b-PAA48 polymersome in 5 mM sodium dihydrogen phosphate, 5 mM EDTA at pH 7.2.This experiment shows the characteristic (ratiometric) changes in the HPTS excitation spectra with the 405 nm maxima moving up and the 460 nm maxima moving down following slight acidification after irradiation of this system in the absence of Ru(II)-bisterpyridine chromophore and hence no EDTA by-product formation after irradiation. (b) Excitation spectra of 8-cyt c:CcOx:polymersome in 50 mM unbuffered potassium chloride, 5 mM EDTA, pH 7.2. In this experiment, there is an overall increase in excitation spectra baseline due to scattering caused by EDTA by-products from photoreduction resulting in increase in both the 405 nm and 460 nm excitation maxima for HPTS. (c) UV-Vis spectra of bulk 8-cyt c after photoreduction in 5 mM sodium dihydrogen phosphate, 5 mM EDTA, pH 7.2 showing increased scattering due to EDTA by-products. (d) UV-Vis spectra of 8-cyt c:polymersome after photoreduction in 5 mMsodium dihydrogen phosphate, 5 mM EDTA, pH 7.2 showing increased scattering due to EDTA by-products.

Appendices
Appendix F
F.1 Optical density correction
The number of photons absorbed by a chromophore can be estimated based on
transmittance using the formula TA log where A is absorbance and T is
transmittance.2
Figure F.1. Transmittance of light based on absorbance of chromophore absorption maxima.
In a typical calculation, for a Ru(II)-bisterpyridine chromophore with an absorbance of
0.0115 a.u, an equivalent 97.9% transmittance of photons is observed. Therefore, 2.1%
of incident photons are absorbed by the complex participating in photoexcitation, which
is the quantity used for subsequent quantum efficiency calculations.
F.2 Quantum efficiency of photoreduced 8-cyt c bioconjugates
In a typical calculation, assume that the initial rate of heme reduction is within the linear
region.
Absorbancephotoreduction = 0.007373 a.u. ( 550 = 24.3 mM-1cm-1)

Appendices
Abs = b[8-cyt c]
[8-cyt c] = 3.03 10-7 M (in an 80 L solution)
moles of reduced 8-cyt c = 2.427 10-11 mol
= 2.427 10-11 NA electrons in 1860 s
= 1.46 1013 electrons in 1860 s
Therefore, rate of heme reduction is 7.85 109 electrons/s
%efficiency = (rate of heme reduction/incident photons) 100% ( incident =
1.4 1016 photons/s)
= (7.85 109/1.4 1016) 100%
= 5.59 10-5 %
= %100)correctiondensity opticalphotons(incident
reductionhemeofrate
= (7.85 109/[1.4 1016 0.095]) 100% (For 0.046 a.u., 9.5% photons
contributing to excitation)
= 5.9 10-4%
F.3 Determinatin of proton pumping rate (H+/s)
With the following representative calculation, proton pumping rate was determined.
Vout
Vin

Appendices
Estimation of number of polymer molecules within membrane of one polymersome:
Vmembrane = Vout – Vin
Vout = 3
34
outd where dout = diameter of vesicles (dout = 367±185 nm) and
membrane thickness = 98±35 nm.
Vout = 2.06 × 10-19 m3
Vin = 3
34
ind
Vout = 8.09 × 10-20 m3
Hence, Vmembrane = 1.25 × 10-19 m3
Therefore, one polymersome occupies total volume of
Vmembrane = 1.25 × 10-19 m3.
As a result, number of polymer molecules in one polymersome nPS-b-PAA;
nPS-b-PAA =PAAbPS
membrane
VV
= 4.76 × 107 molecules
= 7.90 × 10-17 mol
mpolymersome = 1.42 × 10-12 g (for one polymersome).
Estimation of number of polymersomes in reaction sample:
Now, the mass of PS-b-PAA in 120 mL for a 1:6 THF:Buffer solution with
1 mg/mL polymer dissolved in THF and MWPS-b-PAA = 18054 g/mol is 0.02 mg
(6.67 × 1014 molecules).
Total number of polymersomes (Npolymersomes) in reaction sample is then;
Npolymersomes = 6.67 × 1014 molecules/4.76 × 107 molecules
= 1.40 × 107 polymersomes
Estimation of total encapsulated volume in reaction sample:
As a result, total encapsulated volume in a reaction solution (Vtotal inside);
Vtotal inside = Npolymersomes × Vin
= 1.13 × 10-12 m3
=1.13 × 10-9 L
dPS-b-PAA 0.27 nm
lPS-b-PAA 46 nm
VPS-b-PAA = ld 2)2
(
VPS-b-PAA = 2.63 × 10-27 m3

Appendices
Estimation of proton pumping rate:
Initial average internal pH for pH 7.2 buffered experiment was pH 7.28 and
increased to pH 7.44 over 3360 s of irradiation.
Hence, [H+] = 1.63 × 10-8 M protons pumped.
Therefore, nH+ = [H+] × Vtotal inside
= 1.85 × 10-17 mol
= 1.12 × 107 H+ pumped across membrane over 3360 s.
Rate of proton pumping = 3.33 × 103 H+/s.
F.4 Quantum efficiency of proton pumping
= %100)correctiondensity opticalphotons(incident
ratepumpingproton ( incident =
1.4 1016 photons/s)
= (3333/[1.4 1016 0.021]) 100% (For 0.0115 a.u., 2.1% photons contributing to
excitation)
= 1.1 10-9%

Appendices
Appendix G
Abbreviations
ABTS 2,2'-azino-bis(3-ethylbenzothiazoline-6-sulphonic acid)
alpha
beta
br broad
ca. circa
NA Avogadro constant
J coupling constantoC degrees Celsius
d deuterium
s singlet (NMR)
s strong (IR)
d doublet
t triplet
q quartet
p pentet
m medium (IR)
m multiplet (NMR)
w weak (IR)
g grams
mg milligrams
h hours
Hz hertz
MHz megahertz
nuclear magnetic resonance chemical shift (ppm)
ppm parts per million
mp melting point
m/z mass to charge ratio
MS mass spectrometry
IR infrared
UV-Vis ultraviolet visible spectroscopy
L litre

Appendices
mL millilitre
L microlitre
M molar
mM millimolar
M micromolar
nM nanomolar
mol mole
mmol millimole
mol micromole
nmol nanomole
min minutes
molar absorptivity
r.t. room temperature
cmd centimeters diameter (chromatography column diameter)
cmh centimeters height (chromatography column height)
CLSM confocal laser-scanning microscopy
TEM transmission electron microscopy
FPLC fast protein liquid chromatography
HPLC high-performance liquid chromatography
ESI electrospray ionisation
PBS phosphate buffered saline
HEPES 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid
DIPEA N,N-diisopropylethylamine
DMF N,N-dimethylformamide
CH2Cl2 dichloromethane
CDCl3 chloroform
CH3CN acetonitrile
DMSO dimethyl sulfoxide
MeOH methanol
EtOH ethanol
THF tetrahydrofuran
TLC thin layer chromatography

Appendices
MALDI-TOF matrix assisted laser desorption ionisation time of flight mass
spectrometry
UFTRPL ultrafast time-resolved photoluminescence
TCSPC time-correlated single photon counting
ESI electrospray ionisation
SEC size exclusion chromatography
CEX strong cation exchange chromatography
IMAC immobilised metal affinity chromatography
PC L- -phosphatidylcholine
POPC L-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine
PS-b-PAA polystyrene-b-poly(acrylic acid)
EDTA ethylenediaminetetraacetic acid
BSA bovine serum albumin
cyt c cytochrome c
CcOx cytochrome c oxidase
GFP green fluorescent protein
bpy 2,2'-bipyridine
tpy 2,2':6',2"-terpyridine
HPTS 8-hydroxy-1,3,6-pyrenetrisulfonate
HATU O-(7-azabenzotriazole-1-yl)-1,1,3,3-tetramethyluronium
hexafluorophosphate
HOAc glacial acetic acid
DTT dithiothreitol
References
(1) Huang, C.; Mason, J. T. Proc. Natl. Acad. Soc. U. S. A. 1978, 75, 308.(2) Skoog, D. A.; West, D. M.; Holler, J. F.; Crouch, S. R. Fundamentals of
Analytical Chemistry 8th ed.; Thomson Brooks/Cole: Belmont, California, 2004.