Embed Size (px)
Citation preview

Deep-Sea Research, Vol. 34, Nos 5/6, pp. 741-753, 1987. 0198~)149/87 $3,00 + 0.130 Printed in Great Britain. © 1987 Pergamon Journals Ltd.
Light effects and diel variations of nitrate reductase activity in phytoplankton from the northwest Africa upweiling region
ROSA MARTINEZ,* TltEODORE T. PACKARDt a n d DOLORS BLASCO+
(Received 28 September 1985; in revised form 15 March 1986~ accepted 5 October 1986)
Abstract--Light kinetics and dicl cyclcs of nitrate reductase (NR) activity werc studied in the upwelli.~g ecosystem off northwcst Africa. The activity of the enzyme showed a strong response to light ~t low intensities but became saturated tit light intensities above 15-30% of the incident light intensity. At higher irradianccs, NR activity showed photoinhibition. At sea surface irradiances an average inhibition of 32% in the NR activity was observed.
Diel cycles of NR activity exhibited the following characteristics: a low prc-dawn value (0.(13- 0.009 lamol NO3-N (lag Chl a)-l), a rapid increase with the onset of.daylight, a maximum before noon (tripling the dark value), a secondary maximum in the early afternoon, and an afternoon- evening decreasc that coincided with the decrease in sunlight. To simulate these characteristics a continuous polynomial function of light and time was developed.
INTRODUCTION
NITRATE reduction by phytoplankton and its relationship to nutrients and light are important for ocean mapping (ZIRINO et al. , 1978; Zm~so, 1985), modelling the nitrogen cycle, and understanding phytoplankton physiology. This is expecially true in neritic waters, in the oceanic thermocline, and in upwelling systems--zones where assimilatory nitrate reduction is often the dominant mode of nitrogen assimilation.
Investigations of the enzymatic mechanisms by which nitrate is assimilated have led to the use of nitrate reductase (NR) activity as an index of nitrate reduction (PACKARD et al. , 1971; BLASCO et at . , 1984). Because the enzyme assay is simple, quick and amenable to automation, it provides a tool for mapping nitrate utilization in the same way nutrient salts are mapped (ZmlNO, 1985). However, before this approach can be tested, the diel variation of the nitrate reductase activity in natural phytoplankton assemblages need to be understood (EePLEV et al. , 1970; PACKARD et al. , 1971; PACKARD and BLASCO, 1974).
Diel physiological rhythms, in response to fluctuation in light intensity, have been detected in cultures and natural phytoplankton communities. The list includes photosyn- thesis, P-I curves (HARDING et al . , 1981, 1983); photosynthetic capacity, Pm~,x (TAGUC.I, 1976; MACCAULL and PLA~, 1977; PREZEIAN and LEY, 1980); cellular pigment content (OwENs et al. , 1980; HARDING et al . , 1983); in v ivo chlorophyll a fluorescence (BLAsCo, 1973; OWENS et al . , 1980); nutrient uptake and assimilation (EeeLEV et al. , 1970; PACKARD et al . , 1971; MACISAAC, 1978; TERRY et al . , 1985); carbohydrate and protein content
* E.T.S. Ingenieros C.C.P., Cfitedra de Ecologia, Universidad de Santander, Santander 39071, Spain. t Bigelow Laboratory for Ocean Sciences, West Boothbay Harbor, ME 04575, U.S.A.
741

742 R. MARTINEZ et aL
(HITcHCOCK, 1980); cell division (SMAYI)A, 197S; CttlSItOLM et al., 1978): and enzyme activity (e.g. EPPLEY et al., 1970; PACKARD et al., 1971).
For years it was known that nitrate uptake, and presumably nitrate reduction, arc stimulated by light in a manner similar to the light stimulation of carbon fixation. More recently, with the introduction of the JSN technique, it became apparent that NO~ uptake exhibits diel periodicity (EPPLEY et al., 1970; MACISAAC, 1978). Nitrate uptake and carbon fixation are closely associated, and diet cycles in nitrate uptake, assimilation, nitrate reductase activity, and carbon fixation follow a similar pattern; moreover, nitrate assimilation is inhibited at high light intensities, as is carbon fixation (PRisct~, 1984).
In spite of this general knowledge, the diel cycles of NR activity are not well documented. Past work on the Peru upweiling system (EppLEY et al., 1970; PACKAI~ et al., 1971) suggested a diel cycle. In this work, we (1) confirm earlier findings and present a more detailed description of the diel cycle; (2) show that diel oscillations in shipboard experiments reflect natural cycles in in situ phytoplankton assemblages; (3) characterize the optimum light level for NR activity for different levels of ambient light; and (4) dra~, generalizations about the light and time functions (i.e. NR = f (hv ) + f( t )) that even- tually could be used to model and map nitrate reduction in upwelling ecosystems.
M A T E R I A L S AND METHODS
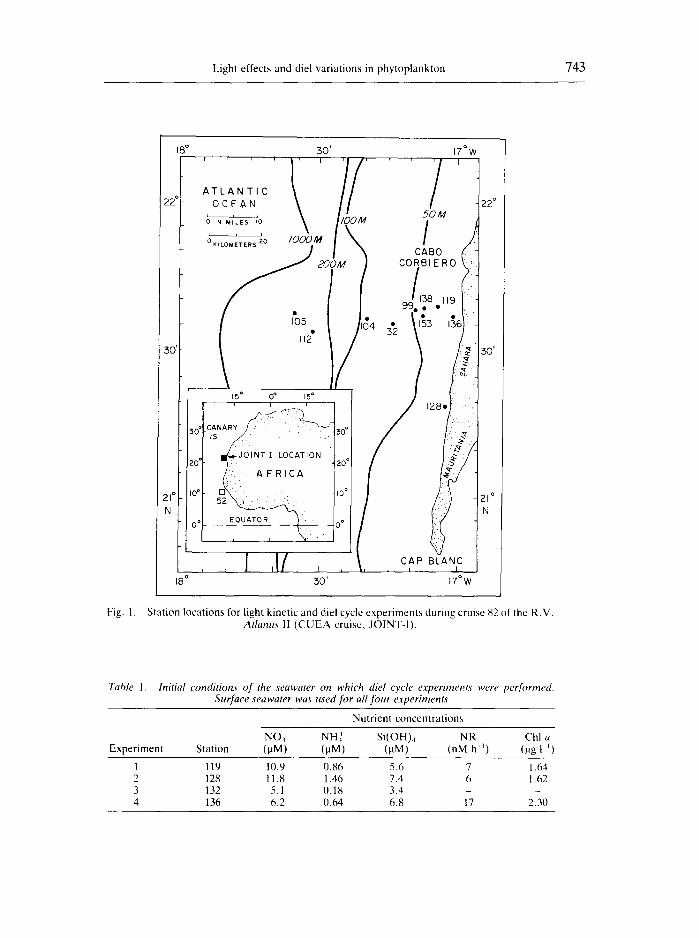
This study was part of the JOINT-I CUEA-12 Experiment off northwest Afric~¢ Figure 1 shows the station locations for the diel cycle and light kinetics experiments. A preliminary report of this research appeared in PACKARt) et al. (1979).
Experimental design
Three types of experiments were performed. The first consisted of four time-series experiments in which NR activity, chlorophyll, nitrate and ammonium were measured in deck-incubated samples maintained in 30 I screened Pyrex bottles at sea surface tempera- ture (16.5°C). The screened bottles simulated the 50% /,, light level. To avoid nutrient exhaustion during the experiment, the cultures were enriched with nutrient-salts to 30 p.M NO~ and Si(OH)4, and 4 laM PO]-on the evening before the first measurement. The initial water conditions prior to enrichment are given in Table l.
The second type of experiment consisted of a time-series analysis of samples drawn directly from the sea at one location (time-series Stas Nos 106-118). Five depths were studied, but only two, the surface and 50% 1o levels, are reported here. The third type of experiment consisted of light-kinetics measurements performed on samples held without enrichment in shaded deck incubators at ambient sea surface temperatures (~16°C), Between 0200 and 0400 h the morning after the sample was taken, the seawater was mixed well and divided into eight 2 I screened bottles. The bottles were designed to transmit the following percentages of incident light: 100, 50, 30, 15, 5, 3, 1 and 0%. They were incubated as before in the clear plexiglass deck incubators from before dawn until 1100 h at which time the plankton from each bottle were filtered and assayed for NR activity, The Chl a used to calculate the specific activity was measured at the beginning of the experiment (pre-dawn). Times are given as solar time.
Analytical methods
The NR activity was measured within 30 rain after collecting the samples by a method of EPPLEY et al. (1969) that was modified by PACKARD et al. (1978) to avoid loss of the
Light effects and diel variations in phytoplankton 743
Fig. 1.
18 °
22 °
30
0 ° I
30' ' ' / " // ATLANTIC \ I I
~°2:?,,o \ /,L,
15 ° 0 ° 15"
I ~ ' - 30 I C A N A R Y ~ /
" ~ I L ToN "1NT OCA I 12C / ,o
_ EQUATOR ~ .' O* L I
° 17 W
'/t22o 50M
//1 CABO CORBIERO
CAP BLANC
18 ° 30' 17°W
30'
21 ° N
Station locations for light kinetic and diel cycle experiments during cruise 82 of the R.V. Atlantis 1I (CUEA cruise, JOINT-I).
Table 1. Initial conditions of the seawater on which diel cycle experimet~ts were performed. Surface seawater was used for all four experiments
Nutrient concentrations
NO:~ NH~ Si(OH)4 NR Chl a Experiment Station (pM) (laM) (pM) (nM h i) (lag I R)
1 119 10.9 0.86 5.6 7 1.64 2 128 11.8 1.46 7.4 6 1.62 3 132 5.1 0.18 3.4 - - 4 136 6.2 0.64 6.8 17 2.30
744 R. MARTINEZ et al.
chloroplast-bound NR. Chlorophyll a was determined fluorometrically by the SCOR- UNESCO method (1966) and nutrients were analysed by AutoAnalyze@ (PAcKAm~ et al., 1978)'."
Light measurements
Daily total solar radiation was measured with an Eppley Pyranometer placed on a 3ft pole on the boat-deck of the Atlantis I1. The calibration value was 2.25 mV = 1 Cal cm -2 min -~. Light penetration was determined with a submarine quantum meter (Model Ll185, Lambda Instruments Corp.) as described in BARBER and HUNTSMAN (1975).
R E S t l _ ' l S
Diel cycles
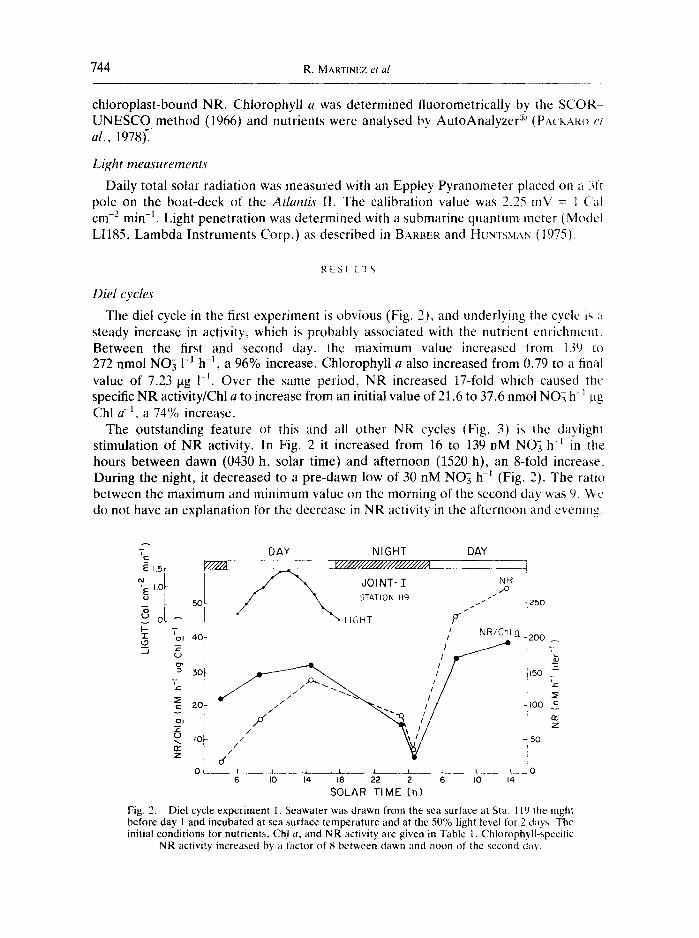
The diel cycle in the first experiment is obvious (Fig. 2), and underlying the cyclc is a steady increase in activity, which is probably associated with the nutrient enrichment Between the first and second day, the maximum value increased from 139 to 272 nmol NO~ 1-1 h -1, a 96% increase. Chlorophyll a also increased from 0.79 to a final value of 7,23 gg 1 -t. Over the same period, NR increased 17-fold which caused the specific NR activity/Chl a to increase from an initial value of 21.6 to 37.6 nmol NO7 h -~ ~tg Chl a -l, a 74% increase.
The outstanding feature of this and all other NR cycles (Fig. 3) is the daylight stimulation of NR activity. In Fig. 2 it increased from 16 to 139 nM NO7 h -~ in the hours between dawn (0430 h, solar time) and afternoon (1520 h), an 8-fold increase. During the night, it decreased to a pre-dawn low of 30 nM NO3 h -~ (Fig. 2). The ratio between the maximum and minimum value on the morning of the second day was 9. We do not have an explanation for the decrease in NR activity in the afternoon and evening.
A
15 f
I--- T lOi t.9 i
,5
"7 a~
g
Z
Fig. 2.
DAY NIGHT DAY - - - - - - - - - ~7//////////////////////A _~
JOINT- I NR i 50 STATION tl9 / ~ t . . . . . 0 ~250
LIGHT fir J
4C / NR/Chl _q 1200 C- lyi 3c ~ 15o -- ! '4:::
200 /111/ l / ~ I J ~c
. / ~ . t / ~ ~ i 0 0 ! m i z
~o -I 50
d I I I I I I I ]0
6 IO 14 ,8 22 2 6 ,0 14.
SOLAR TIME (h)
Diel cycle experiment 1. Seawater was drawn from the sea surface at Sta. 119 the night before day 1 and incubated at sea surface temperature and at the 50% light level for 2 days. The initial conditions for nutrients, Chl a, and NR activity are givcn in Table 1. Chlorophyll-spccilic
NR activity increased by a factor of 8 between dawn and noon of the second day.
~ _'2
!.
t#
'.
LIG
HT
(C
ol
cm -2
rai
n "l
)
o ,
b L,
, r
• N
R/C
hI~_
{ nM
NO
~ h -
~ H
g C
hl- 0
-~)
LIG
HT
(Co
l cm
-2
min
-I)
LIG
HT
(C01
cm
'2m
in
"1)
o ~
o .
,,7
NR
/Chl
o_ (n
M N
O~
h-~M
gChl
g'
~)
NR
/Ch
l g
(nM
N
O~
h-'
pg
Chl
g )
,-a
~'~
(.o
Z
;-&
~ '~
~ ~
~t
~ .--
---~.
....,-
......
.. /1
1 o
=- m
z _
2-
I ~
.%
..... '"
::.,&
'-.
.I~"
11
....
..
--
~-
;;8
C3
O~
Z
~'~
• -I
r~
I I
I I
I I
J I
I
~=
E
¢3"
r.,:
~ o ~
o N
R (
nM
NO
ah
/l
ite
r -I
]
..
..
..
4
---'
L
o o
CH
LOR
OP
HY
LL (
ug
liter
"l)
NR
( n
M N
O 3
li|er
4
h -I
)
t-
~ 4
~-
-
; t
"~--
-4
J--
J
s o
~ H
~
8 O
O
C
HL
OR
OP
HY
LL
(P
9 IH
ef -
~1
CH
LOR
OP
HY
LL0
(pg
hler
~)
NR
(nM
NO
~ I
h ~
hler
J)
746 R. MARTINEZ et al.
It could not be due to nutrient exhaustion, since NO~ in the afternoon (1500 h) of the second day (time of the minimum NR activity) was 4.90 taM NO~, only 20% lower than the initial value.
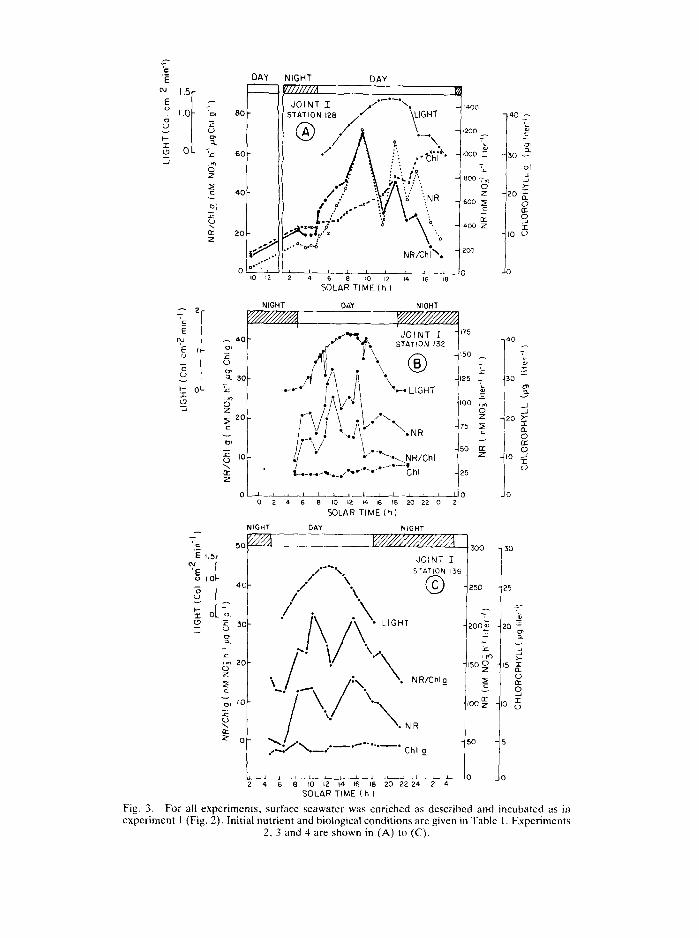
To refine the shape of the diel cycle we increased the sampling frequency in the next experiments. The results are shown in Fig. 3. In experiment 2, the NR activity increased 3-fold from 41 to 121 nM NO~ h -1 during the 24 h between the two maxima. The specific NR activity increased from 8 to 70 nmol NO~ h -~ lag Chl a -] because the Chl a only increased by 2.5 lag 1 -~ during the same period. The low original NR activity suggests some type of repression or inhibition on day 1, possibly caused by the high ammonium concentration (Table 1). In any case, the patterns in experiments 2, 3 and 4 confirmed the results of experiment 1 (Fig. 3). The search for evidence that the induction of NR activity anticipates dawn was accomplished by frequent sampling in the 2 h before dawn. In experiment 2 (Fig. 3A), four samples were taken before 0430 h and no increase in activity was found. In contrast, a sharp increase took place immediately after dawn. Within minutes after the sun appeared, the activity increased by 39% over the initial value; 2 h later it increased 110%, and 2.5 h after that it had increased another 120%. Experiments 3 and 4 (Figs 3B and C) show the same pattern. The ratio between the maximum and the minimum value on the morning of day 2 was 6. We also found in experiments 2, 3 and 4, as in experiment 1, a continual increase in NR, Chl a. and NR/Chl a throughout the 2 days of incubation.
By sampling more frequently, we were able to detect in experiments 2, 3 and 4, not only the light activation of NR, but also light inhibition of NR activity around solar noon. In all three experiments, the activity decreased by 50% and then as the light started to decrease, the NR activity increased to about 80% of the maximum. The specific NR/Chl a activity followed a similar pattern.
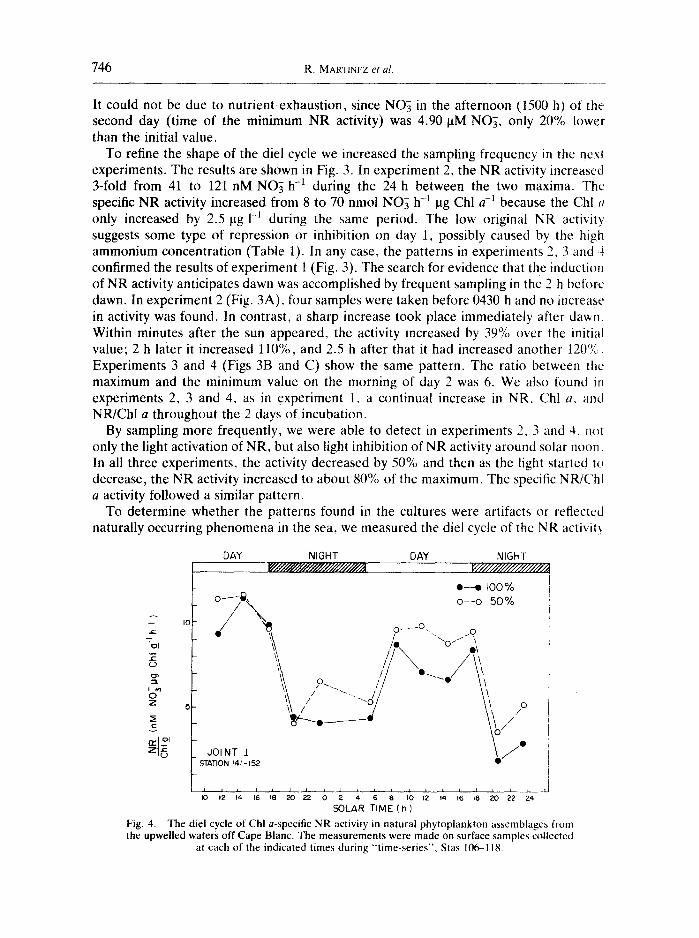
To determine whether the patterns found in the cultures were artifacts or reflected naturally occurring phenomena in the sea, we measured the diel cycle of the NR activity
I 0 T .£:
m JE
Z
E
CE o l
DAY NIGHT DAY NIGHT F///////////////////////////M V// / / / / / / / / / / / / / / / / /~
e - - e 100% - - - i~ , o--o 50%
O- - - 0 \ 0
~ " - o - ~ - - - ~ e ~ o / /
i i ~ i J i * J J i i , i i i i i ~ , ~_L I0 12 I 16 18 2 0 2 2 0 2 4 6 8 I 0 t 2 14 t 6 18 2 0 Z2 2~*
SOLAR TIME ( h ) Fig. 4. The diel cycle of Chl a-specific NR activity in natural phytoplankton assemblages t/tom the upwelled waters off Cape Blanc. The measurements were made on surface samples collected
at each of the indicated times during "'time-series", Stas 106-I 18.
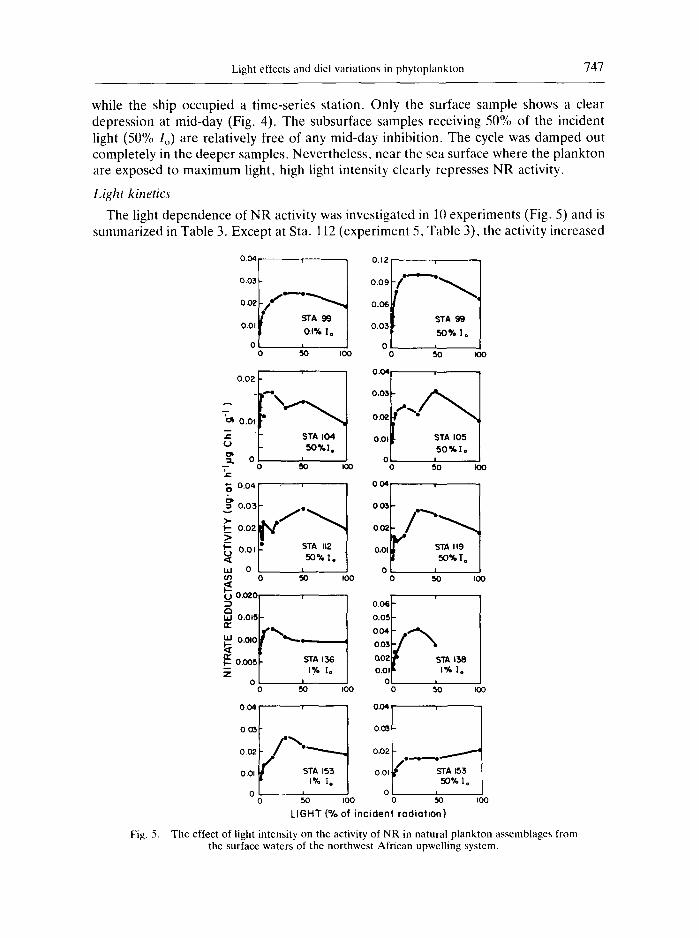
Light effects and diel variations in phytoplankton 7 4 7
while the ship occupied a time-series station. Only the surface sample shows a clear depression at mid-day (Fig. 4). The subsurface samples receiving 50% of the incident light (50% Io) are relatively free of any mid-day inhibition. The cycle was damped out completely in the deeper samples. Nevertheless, near the sea surface where the plankton are exposed to maximum light, high light intensity clearly represses NR activity.
Light kinetics
The light dependence of NR activity was investigated in 10 experiments (Fig. 5) and is summarized in Table 3. Except at Sta. 112 (experiment 5, Table 3), the activity increased
0.04
0.03 ' [
0.02 / ~ " ' ~
~rA ~ I 0,01 I
o ,01% I. I 50 I00
\ 1 ;ooF o / ~ow.,. I
T 0 5O i
-- 0.04 " ' /
0.02
C-- o . o , STA ,2 1
W 0 U~ 5O 100
o=o / j ~ °'°'7,, / ~ o.o,o. "-...______~
~= / '%" I o-- ' I 0 50 I00
005
0.02
0.01
o r ,"" I 0 50 tO0
0.12 F , ,
o[ ,~" 1 O 50 I00
0.04
O.Ol[~ STA ,05 I
o/ r " " I 0 50 I00
0.~, / " ~ ~ o,od~ STA .9 I
O 5O 100
0"06 F T 0.05 r 004 r _ , , ' . ~ o.o~/ \ GO2~P STA 158 O.O/[ = ,1% I.
0 50 I00
/
O OZl o-~--~/
O.OI ~ ' STA 153 O/ 150%1°
0 ~ I00 LIGHT (% of incident radiation)
Fig. 5. The effect of light intensity on the activity of NR in natural plankton assemblages from the surface waters of the northwest African upwelling system.
748 R. MARTIN['Z et al.
from a low value in the dark to a maximtim at either the 15 or 30% 1o and then decreased to a lower value at 100% Io. This decrease at high light intensities represented an average loss of 32% from the maximum. The light dependence of the NR activity at the lower light levels fell into two groups (Table 3). Group l (Stas 104, 112,136 and 153, 50% 1,,) had a low ratio of maximum NR activity to dark NR activity that ranged from 1.4 to 19 with a mean of 1.7. In Group 2 (all the other experiments) the ratio ranged from 3.6 to 4.5 with a mean of 3.9. The half-saturation constants (Kj~) for these two groups were also different. Group 1 had a mean Ku of 0.9% 1,, (range, 0.6-1.1) while Group 2 had a mean KI~ of 2.9% Io (range, 1.6-5.4). In units of langleys rain -j, the mean K~ for the tw~ groups were 0.009 + 0.004, and 0.032 + 0.004, respectively.
l~lsctTss~on
Light dependence of NR activi O,
The light kinetics experiments (Fig. 5) show that at low light levels a rectangular hyperbola describes the light dependence of nitrate reductase activity, but at higher light levels inhibition occurs and another function is required. These results are similar to those of MACISAAC and DUGDALE (1972) for NO? uptake and confirm PACKARD'S (1973) experiments with the light dependence of NR activity. To facilitate quantitative compari- son with previous results we calculated values of Kit and Vn .... as in PACKARD (1973) (Table 2).
Table 2. Summary of the conditions" and kinetic parameters for each experiment shown in big. 5
Experiment Station
Source and initial conditions of the seawater used in each experiment
Light Nitrate reductase depth activity Chl a NO~ Nit; (%1,,) (nmol h -1 lag Chl a l) (p,g I ~) (I-tM) ((aM)
1 99 0.1 13 1.4 9.6 0.3 2 99 50 t5 5.(I S.o 0.6 3 104 5o 23 10.5 9.6 o.2 4 105 Nil 15 2.3 3.9 (I.3 5 112 50 - 7.6 1.2 1i7 6 119 50 4 5.0 8.1i II.~ 7 136 I 2 3. I 4.9 0.2 8 138 I o 1.8 9.5 O.3 9 153 1 8.0 5.7 02 10 153 50 -- 9.9 5.4 0.1
Results of the light experiments K~, V,n~× Max NR Dark NR
(ly rain i) Max NR % Repression (%1o) x 103 (nmol h -I lag Chl a -1) Dark NR at I(~)% I,,
2.4 23 26 24 7 3.6 22 1.6 15 93 88 22 4.1 ~2 l.l 10 17 16 g 1.9 46 2.7 22 33 31 8 3.8 42 - - 86 61 1.4 32
4.1 35 28 28 t~ 4.5 31 0.6 5 13 12 7 1.8 20 3.4 35 45 41 11 3.6 5.4 43 29 26 7 4.1 30 1.0 5 17 17 9 1.8 -

Light effects and diel variations in phytoplankton 749
Table 3. Characteristics of diel cycle experiments 2, 3 and 4 (Fig,. 3). Times are given in solar time; nitrate reductase activities are given as percent o f first maximum. All values reflect means and
standard deviations (n = 3)
Dawn First maximum Noon minimum Second maximum Dusk
Time 4:30 9:37 _+ 11:16 11:49 ± 0.032 13:16 ± 0:52 18:3tl NR activity (%) 31t ± 5 1011 49 ± 14 79 ± 16 30.7 +_ tl.05
The K~ values for both NO~ assimilation and NR activity from previous studies range from 0.6 to 14% of surface intensity, or from 0.002 to 0.051 ly rain ~ in absolute light units (MAcISAAC and DUGDALE, 1972; PACKARD, 1979; PRISCU, 1984; BERMAN et al., 1984). Our values for K~ fall within the same range: l).6-5.4% of surface light intensity, or 0.005-0.043 ly min 1 (Table 2). Most of these K~ values were lower than the light percentage from which the phytoplankton were sampled. These low values indicate that light requirements for nitrate reduction in algae are met at low light levels (AzuARA and APARICIO, 1985), and that even at the bottom of the euphotic zone the phytoplankton populations are able to assimilate nitrate (PACKARD, 1973). In the three cases where the Kit exceeded the level of in situ light, the phytoplankton populations may have been inefficient in their use of light or they may have been advected recently to the depth at which they were sampled. The results of NELSON and CONWAY 11979) support the second explanation. In addition SAKSHAUG 11985) suggests that when phytoplankton cells are in a mixed water column, they will adapt to the highest irradiance encountered. BLASCO et al. 119841 also found phytoplankton at the 1% light level that were acclimated to higher irradiances, and concluded that their presence at the 1% light level was transitory.
To understand the light dependence of NR activity, one must consider the concen- tration of NO3 (MALONE et al., 1975; DUGDALE et al., 1981). As WttALEN and ALEXANI)ER (19841 have pointed out, nitrate assimilation is not always light dependent. These authors, working with phytoplankton from an oligotrophic arctic lake, found light dependent NO~ uptake only in NO~ enriched samples (3.07 laM NO3-N) and not under the low in situ concentrations (0.04-0.06 I~M NO3-N). Our samples for the light kinetic experiments had non-limiting nitrate concentrations ranging from 1.2 to 9.6 IJM NO~-N, and under these conditions NR activity showed light dependence.
More controversial is the influence of species composition. In experiments with dinoflagellates, PAASCHE et al. (1984) found light independent NO~ assimilation in some species and light dependence in others. Diatoms are thought to be more light dependent and to have higher uptake requirements than dinoflagellates ([~PPLEY and HARRISON,
1975). The phytoplankton that we worked with were diatom-dominated (BLASCO et al., 1980), and were not limited by nitrate. In this situation we would expect a strong light response, as was the case.
Levels o f dark N R activity
The "dark" NR activity we found in our kinetic experiments ranged between 7 and 61 nm NO~ h -l p,g Chl a -1, and the Max NR/"Dark" NR ratios had values between 1.4 and 4.5 (Table 2). These values fall in the range encountered for other phytoplankton (BERMAN et al., 1984; KUENZLER et al., 1979; PACKARD and BLASCO, 1974) and macro- a lgae (DAv1DSON and STEWART, 1984). At Stas 99 and 112, we found levels of dark activity considerably higher (2 and 4 times as high) than the average (14 nmol N h -1

750 R. MARTINEZ et al.
lag Chl a-l). Possible explanations for the higher dark NR activity at these stations include the foll6wing: (1) these stations were NO~ limited or had recently experienced NO~ limitation; (2) the phytoplankton assemblages there belonged to taxa which characteristically assimilate NO~ in the dark; (3) NR activity was inhibited by amino nium, more in the light than in the dark (CuLLIMORE, 1981; BALCtt, 1985); (4) the activity which is a capacity, did not reflect the physiological rate (LILLO, 1983).
From our results it is not clear whether the dark activity reflects in situ dark nitrate reduction or simply the activation of the reduction capacity. Even if nitrate uptake in the dark were detected, it would not be evidence of nitrate reduction, because phytoplankton are able to take up and store nitrate without actually reducing it (DoRTCJ~, 1982'). Furthermore, physiological studies such as those by FLO~ES et al. (1983), LARA et aL (1984) and DAVIDSON and STEWART (1984), argue against assimilatory nitrate reduction taking place in the dark.
Light inhibition o f NR activity
Repressed NR activity occurs at high light levels (Fig. 5). This is consistent with other observations of light inhibition of NR or of nitrate uptake (MAcISAAC and DUGDAt.~:, 1972; PACKARD, 1973; Pulser,:, 1984). However, we found the inhibition in eutrophic waters which contrasts with MAclSAAC and DCGDALE'S (1972) observations of light inhibition of nitrate uptake only in oligotrophic waters.
In this work, the depth of the sample did not have any clear relationship with the light inhibition. Theoretically, phytoplankton cells taken from near-surface waters could be expected to have a higher light threshold for inhibition than those coming from deeper zones (0.1 or 1% light depths), as the first should be adapted to higher light intensities, as STEEMAN-NIELSEN (1975) found for photosynthesis and PLATret al. (1982) have found for stratified waters. Our results are likely due to the water column mixing in the northwest African upwelling (FRIEBERTSHAUSE~ et al., 1975), and can be considered as reflecting adaptive strategies similar to those used by polar phytoplankton.
Diel cycles
We observed a diel cycle in NR activity that clearly followed the diel variation in light intensity. Such diel cycles appear to be characteristic for plants and algae growing on NO3 under natural conditions, although there are situations when the cycle is suppressed (DORTCH et al., 1985). The onset of NR activation coincides with dawn. We failed to find the pre-dawn rise that had been reported earlier (EePLEY et al., 1970; PACKARD and BLASCO, 1974). However, since the sampling frequency around dawn in all of these early studies was not great enough to determine conclusively the presence of a pre-dawn rise and since the experiments were done in different waters, the question still requires further investigation. Nevertheless, our results to date support the theory that NR activity at moderate light levels and under nutrient-unlimited conditions is a direcl function of light.
The characteristics of the cycle are summarized in Table 3. The amplitude of the first peak relative to the dark base line averaged 3.3, the amplitude of the second peak relative to the same base line was 2.6, and the relative values in the trough averaged 1.6, a drop of 48% from the first peak. WALLACE and PATE (1965) studied the NR diel cycle in Pisum arvense and reported a maximum NR/dark NR ratio equal to 3, which is the same as we found. In other studies in algal cultures and natural phytoplankton populations, the
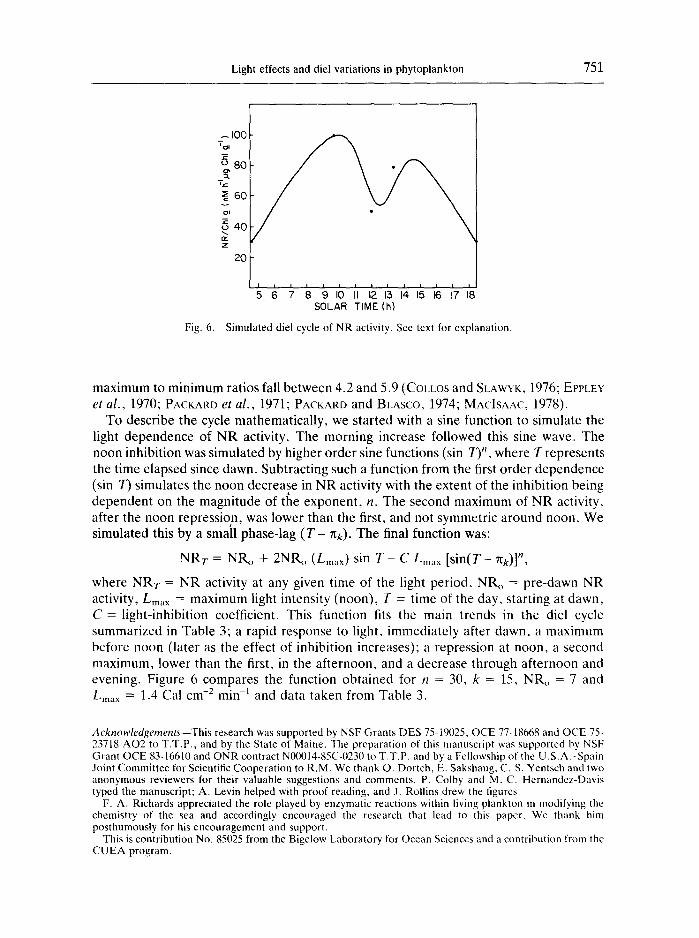
Light effects and diel variations in phytoplankton 751
Fig. 6.
.•_•100 o 80
T,=
6O
6 4o Z
2O
i I ! I i i i I i i i I
5 7 8 9 I0 II 12 15 14 15 16 17 18 SOLAR TIME (h)
Simulated diel cycle of NR activity. See text for explanation.
maximum to minimum ratios fall between 4.2 and 5.9 (COLLOS and SLAWYK, 1976; EPPLEY et al., 1970; PACKARD et al., 1971; PACKARD and BLASCO, 1974; MAClSAAC, 1978).
TO describe the cycle mathematically, we started with a sine function to simulate the light dependence of NR activity. The morning increase followed this sine wave. The noon inhibition was simulated by higher order sine functions (sin 7)", where T represents the time elapsed since dawn. Subtracting such a function from the first order dependence (sin T) simulates the noon decrease in NR activity with the extent of the inhibition being dependent on the magnitude of the exponent, n. The second maximum of NR activity, after the noon repression, was lower than the first, and not symmetric around noon. We simulated this by a small phase-lag ( T - nk). The final function was:
NRT = NRo + 2NRo (Lm~lx) sin T - C L ..... [ s in(T- ~k)] n,
where NRT = NR activity at any given time of the light period, NRo = pre-dawn NR a c t i v i t y , L m a x = maximum light intensity (noon), T = time of the day, starting at dawn, C = light-inhibition coefficient. This function fits the main trends in the diel cycle summarized in Table 3; a rapid response to light, immediately after dawn, a maximum before noon (later as the effect of inhibition increases); a repression at noon, a second maximum, lower than the first, in the afternoon, and a decrease through afternoon and evening. Figure 6 compares the function obtained for n = 30, k = 15, NR,, = 7 and Lmax = 1.4 Cal c m -2 min -1 and data taken from Table 3.
Acknowledgements--This research was supported by NSF Grants DES 75-19025, OCE 77-18668 and OCE 75- 23718 AO2 to T.T.P., and by the State of Maine. The preparation of this manuscript was supported by NSF Grant OCE 83-16610 and ONR contract N00014-85C-0230 to T.T.P. and by a Fellowship of the U.S.A.-Spain Joint Committee for Scientific Cooperation to R.M. We thank Q. Dortch, E. Sakshaug, C. S. Yentsch and two anonymous reviewers for their valuable suggestions and comments. P. Colby and M. C. Hernandcz-Davis typed the manuscript; A, Levin helped with proof reading, and J. Rollins drew the figurcs.
F. A. Richards appreciated the role played by enzymatic reactions within living plankton in modifying the chemistry of the sea and accordingly encouraged the research that lead to this paper. We thank him posthumously for his encouragement and support.
This is contribution No. 85025 from the Bigelow Laboratory for Ocean Sciences and a contribution from thc CUEA program.

752 R. MART~NEZ et al.
R E F E R E N C E S
AZUARA M. D. and P. APARICIO (1985) Spectral dependence ot photoregulation of inorganic nitrogeJ~ metabolism in Chlamydomonas reinhardii. Plant Physiology, 77, 95-98,
BAI,CH W. M. (1985) Lack of an effect of light on methylamine uptake by phytoplankton, l,imnology am/ Oceanography, 30, 665-674.
BARBER R. T. and S. A. HUNTSMAN 11975) Joint-I Carbon, chlorophyll and light extinction---R/V Atlami,~ I I Cruise 82. C.U.E.A. Data Report 14, t65 pp.
BI~,RMAN T., B. F. SHERR, D. LUYNNti and J. J. M('CArrtlv (1984) The characteristics o1 ammonium a~d nitrate uptake by phytoplankton in Lake Kinneret. Limnology and Oceanography, 29,287-291.
BLAS('O D. (1973) Estudio de las variaciones de la relation fluorescencia in vivo/clorofila y su aplicacion cn oceanografia. Influencia de ht limitacion de diferentes nutrientes, efecto del dia y Roche y dependencia de la especie estudiada, lnvestigaciones Pesqueras, 37,533-556,
BLASCO D., M. ESTRADA and B. H. JONES (1980) Relationship between thc phytoplankton distribution u~td composition and the hydrography in the Northwest African upwelling near Cabo Corbeiro. De~7~-Sc, Research, 27, 79%821.
BLASCO D., J. J. MACISAAC, T. T. PA¢KARD and R. C. DUGDALE 119841 Relationship between nitralt: reductase and nitrate uptake in phytoplankton in the Peru upwelling region. Limnoh~.qv and Ocea,,',- graphy, 29, 275-286.
CHISHOLM S. W., F. AZAM and R. EPPI,EY 11978) Silicic acid incorporation in marinc diatoms on light :da~k cycles: Use as an assay for phased cell division. Limnology and Oceanography, 23, 518-52t,L
COLLOS Y. and G. SLAWYK (1976) Significance of cellular nitrate content in natural populations ol mannc phytoplankton growing in shipboard cultures. Marine Biology, 34, 27-32.
CULLIMORE J. V. (1981) Glutamine synthetase of Chlamydomonas. Rapid reversible deactivation. Planta, 152, 587-591.
DAVIDSON I. R. and W. D. D. STEWART ( 19841 Studies on nitrate reductase activity in Laminaria digitata. The role of nitrate availability in the regulation of enzyme activity. Journal of Experimental Marine BiOlog) and Ecology, 79, 6,%-78.
DORTCH Q. (1982) Effect of growth conditions on accumulation of internal nitrate, amino acids, and protein m three marine diatoms. Journal of Experimental Marine Biology and Ecology, 61,243-264.
DORTCH Q., J. R. CLAYTON, S. S. TttORESEN, J. S. CIJ-:VH,AND, S. L. BRESSLER and S. I. AHMf~D (1985) Nitrogen storage and use of biochemical indices to assess nitrogen deficiency and growth rate in natured plankton populations. Journal of Marine Research, 43,437-464.
DUGDALE R. C., B. H. JONES, J. J. MA('ISAAC and J. J. GOF, RIN(; (19811 Adaptation of nutrient assimilation Canadian Bulletin of Fisheries and Aquatic Science, 210, 234-2511.
EPPLEY R. W. and W. G. HARRISON 11975) Physiological ecology of (7onyaula:c polyedra, a red w~ltcr dinoflagellate off Southern California. In: Toxic dino]tagellate blooms, V. R. Locicf-r¢/, editor, Mass~ chusetts Science and Technology Foundation, Wakefield, Massachusetts, U.S.A., pp. l l - 2 1
EPPLI-:Y R. W., J. L. COATSWORTH and L. SOLORZANO (1969) Studies on nitrate reductasc in matin~_ phytoplankton. Limnology and Oceanography, 14, 194-2tl5.
EPPLEY R. W., T. T. PACKARD and J. J. MA¢'IsAAC 1197111 Nitrate reductase in Peru current phytoplankton. Marine Biology, 6, 195-199.
FLORES E., M. G. GUERRERO, J. M. RGMERO and M. LOSAI~A (1983) Regulatory interaction o1 photosynthc tic nitrate utilization and carbon dioxide fixation in the cyanobacteria Anacystis nidulans. Biochimica ,,~ Biophysica Acta, 725, 529-532.
FRIEBERTSHAUSER M. A., L. A. CODISPOIh D. D. BISHOP. G. E. FRIteDRICIt and A. A. W~-sli~A~;~ ',, (1~175i Joint I Hydrographic Station Data, C.U.E.A. Data Report 18, 243 pp.
HARDING L. W., B. W. MEESON, B. B. PREZELIN and B. M. SWI-:ENEY (1981) Diel periodicity of photosymhe~ sis in marine phytoplankton. Marine Biology, 61, 95-1115.
HARDING L. W., B. W. MEESON and M. A. TYLER (1983) Photoadaption and diel periodicity of photosynthe- sis in the dinoflagellate Prorocentrum mariaelebouriae. Marine Ecology Progre~s Serie,s, 13, 73-85.
HITCHCOCK G. L. (1980) Diel variation in chlorophyll a, carbohydrate and protein content of the marine diatom Skeletonema costatum. Marine Biology, 57, 271-278.
KUENZLER E. J., D. W. STANLEY and J. P. KOENINGS (1979) Nutrient kinetics of phytoplankton in the Pamlico River, North Carolina. Water Research Institute University of North Carolina Report No. 139, 163 pp.
LARA C.. J. M. ROMERO, E. FLORES, M. G. GUERRERO and M. LOSAI)A (1984) Regulation on nitrate utilization by CO2 fixation products in the cyanobacterium Anacystis nidulans. Advances in Photosynthetic Research, 2, 715-718.
LILLO C. (19831 Diurnal variations of nitrate reductase activity and stability in barley leaves. Phmt Physiology, 58, 184-188.
MACCAULL W. A. and T. PI,A'Fr (1977) Die[ variations in the photosynthetic parameters of coastal marine phytoplankton. Limnology and Oceanography, 22,723-731.

Light effects and diel variations in phytoplankton 753
MACISAAC J. J. (1978) Diel cycles of inorganic nitrogen uptake in a natural phytoplankton population dominated by Gonyaulax polyedra. Limnology and Oceanography, 27, 1141-1153.
MACISAAC J. J. and R. C. DUGDALE (19721 Interactions of light and inorganic nitrogen in controlling nitrogen uptake in the sea. Deep-Sea Research, 19, 2(1%232.
MAI,ONE T. C., C. GARSIDE, K. C. HAINES and O. A. ROELS (1975) Nitrate uptake and growth of Chaetoceros sp. in large outdoor continuous cultures. Limnology and Oceanography, 20, 9-19.
NLLSON D. M. and H. L. CONWAY (19791 Effects of the light regime on nutrient assimilation by phytoplankton in the Baja California and Northwest African upwelling systems. Journal of Marine Research, 37, 3O 1-318.
OW~NS T. G., P. G. FALKOWSKI and T. E. WUm,VD(iE (1980) Diel periodicity in cellular chlorophyll content in marine diatoms. Marine Biology, 59, 71-77.
PAASCHE E., 1. BRYCFSON and K. TANGEN (1984) lntcrspecilic variation in dark nitrogen uptake by dinoflagellates. Journal of Phyeology, 20, 394-4t11.
PACKARD T. T. (19731 The light dependence of nitrate reductasc in marine phytoplankton. Limnology and Oceanography, 18,466-469.
PACKARD T. T. (19791 Half-saturation constants for nitrate reductase and nitrate translocation in marine phytoplankton. Deep-Sea Research, 26,321-326.
PACKARD T. T, and D. BLASCO (1974) Nitrate rcductase in upwclling regions. 2. Ammonia and light dependence. Tethys, 6,269-2811.
PACKARD T. T., D. BI.ASCO, J. J. MA('ISAA(" and R. C. DU(;DALE 11971) Variations of nitrate reductasc activity in marine phytoplankton. Investigaeiones Pesqueras, 35,209-219.
PACKARD T. T., R. C. DUGDALE, J. J. GOERIN(i and R. T. BARBER 11978) Nitrate rednctase activity in the subsurface waters of thc Peru Current. Journal of Marine Research, 36, 59-76.
PACKARD T. T., V. JONES and D. BLAS('O (1979) Nitrate reductasc activity in the N.W. African upwelling area. C.U.E.A. Technical Report No. 511, 10(1 pp.
PIAVF T., W. O. HARRISON, B. IRWIN, E. P. HORNE and C. L. GAIJ+(;OS (1982) Phytosynthesis and photoadaptation of marine phytoplankton in the Arctic. Deep-Sea Research, 29, 1159-1170.
PREZF, I.IN B. B. and A. C. LrY (198(I) Photosynthesis and chlorophyll a fluorescence rhythms of marine phytoplankton. Marine Biology, 55,295-307.
PRISCU J .C. (1984) A comparison of nitrogen and carbon metabolism in the shallow and deep-water phytoplankton populations of a subalpine lake: response to photosynthetic photon flux density. ,lournal Of Plankton Research, 6,733-749.
SAKASHAUG (1985) The physiological ecology of polar phytoplankton. 6th Conf. Comit6 Arctique Inter- national, Fairbanks, 13-15 May 1985.
SCOR-UNESCO (19661 Dctermination of photosynthetic pigments in sea water. Monographs on Ocean- ographic Methodology, 1, 69 pp.
SMAYDA T. J. (1975) Phased cell division in natural populations of the marine diatom Dityhnn hrightwelli and the potential signilicance of diel phytoplankton behavior in the sea. Deep-Sea Research, 22. 151-165.
SrEEMAN-NH~LSt~N E. (19751 Marine photosynthesis. Elsevier, Amsterdam, 141 pp. TA¢;UCHI S. (1976) Short-term variability of photosynthesis in natural marine phytoplankton populations.
Marine Biology, 37, 197-2117. TL'RR¥ K. L., J. HIRATA and E, A. LAWS (19851 Light-, nitrogen-, and phosphorus-limited growth of
Phaeodaco'lum tricornutum Bohlin strain TFX-I: chemical composition, carbon partitioning and the diel periodicity of physiological processes. Journal (~f Experimental Marine Biology and Ecology, 86, 85-100.
WALLACF, W. and J. S. PATE (1965) Nitrate reductase in the field pea (Pisum arvense L.). Analytical Botany,- 29,655-671.
WUALt~N S. C. and V. ALEXANDER (1984) Diel variations in inorganic carbon and nitrogen uptake bv phytoplankton in an arctic lake. Journal of Plankton Research, 6, 571-590.
ZIRINO A. (1985) Mapping strategies in chemical oceanography. Advances in Chemistry Series 209, ACS, 467 PP.
ZIRINO A., C. CLAVELL and P. F. St(I.IGMAN (1978) Real-time analysis in chemical oceanography. Naval Research Reviews, 31, 26-38.





























