Embed Size (px)
Citation preview

Anita. Behav., 1992, 44, 1021-1027
Lifelong patterns of forager behaviour in a tropical swarm-founding wasp: effects of specialization and activity level on longevity
S E A N O ' D O N N E L L & R O B E R T L. J E A N N E Department of Entomology, University of Wisconsin, Madison, W153706, U.S.A.
(Received22 November 1991; initial acceptance 30 January 1992," final acceptance 20 February 1992; MS. number: A6223)
Abstract. The relationships between foraging tenure and specialization on materials, and foraging tenure and foraging activity were measured for marked, known-age workers of the social wasp Polybia oeciden talis. The purpose of the study was to assess differences in cost to foragers, in terms of risk of mortality, associated with material specializations and activity level, and determine whether ontogenetic changes in forager behaviour reflect these differences in cost, such that more risky patterns of behaviour are postponed until later in adult life. Polybia occidentalis workers terminated foraging at an average of 5-9 days after their first trip, independent of their age at first foraging, suggesting that this task entailed a high risk of mortality. Lifetime foraging activity was calculated as foraging rate (mean number of trips per hour), foraging effort (mean number of time-weighted trips per hour), and as the proportion of observation days that an individual was active during her foraging tenure. Foraging tenure was not related to foraging rate or effort, but had a strong negative relationship with the proportion of observation days during which an individual foraged. By the latter measure, risk of mortality increased with foraging activity: The length of foraging tenure was negatively correlated with the proportion of foraging effort devoted to food materials (nectar and insect prey), supporting the hypothesis that gathering food was riskier than gathering nest materials (water and wood pulp). Foragers that switched between food and nest materials exhibited no tendency to specialize on nest materials first and therefore foraging for riskier materials was not always performed later in adult life. These results are discussed in terms of possible factors regulating task performances at the individual level in social insects.
Recently some authors have suggested that behav- ioural specialization by workers increases efficiency and reliability at the colony level in eusocial insects (Jeanne 1986; Franks 1987) and therefore increases colony fitness. However, in some eusocial insects, workers that adopt certain behavioural roles have been shown to suffer high costs or risks. Foraging, in particular, entails a high risk of mortality rela- tive to other roles (Sakagami & Fukuda 1968; Porter & Jorgensen 1981; Schmid-Hempel & Schmid- Hempet 1984; Strassmann 1985). We examined the notion of behaviour-specific costs to workers through studies of foraging in the advanced eusocial wasp Polybia oecidentalis.
Even within behavioural roles, specialization by eusocial hymenopteran workers can be striking. In some species, forager specializations have been shown to persist for at least several days (Heinrich 1976; Rissing 1981; Schmid-Hempel 1984; O'Donnell & Jeanne 1990). In other species,
in which foragers have been found to specialize on materials for longer periods (Forsyth 1978; Strassmann et al. 1984; Post et al. 1988), foraging may be subdivided into distinct roles based on individual material specializations.
Few studies have addressed the possibility of differential costs to foragers that specialize on different materials (Strassmann et al. 1984; Post et al. 1988). The cost we consider is the likelihood of mortality, hereafter called risk, associated with foraging for particular materials. Within colonies, foragers also frequently differ in rate of work or activity level, and several studies have sought a correlation between worker longevity and foraging activity (Schmid-Hempel & Wolf 1988; Calabi & Porter 1989; Wolf & Schmid-Hempel 1989). The goal of this study was to learn whether certain material specializations or high activity levels involved greater risks for P. occidentalis foragers.
0003-3472/92/121021 + 07 $08.00/0 �9 1992 The Association for the Study of Animal Behaviour 1021

t022 Animal Behaviour, 44, 6
M E T H O D S
Study Site and Colonies
The study was conducted between 26 June and 8 August 1989 (rainy season) near the town of Cafias in Guanacaste Province, Costa Rica. The study site is a mixture of gallery forest, seasonally deciduous forest, and open pastures with scattered shrubs. Polybia occidentalis nests are most abundant in the last of these habitats, where many colonies nest within 1.5 m of the ground. A more detailed descrip- tion of the site is found in Daubenmire (1972). We studied four colonies of moderate size (adult popu- lation of several hundred individuals), with all brood stages present, and located within a t-ha area.
Two of the colonies (numbers 4 and 5) were exper- imentally manipulated periodically during the study by removing a portion of the outer nest envelope. The effects of these manipulations have been pub- lished elsewhere (O'Donnell & Jeanne 1992); the manipulations had no significant effect on the behavioural variables addressed here or on longevity and will not be considered further.
2 days. Observation periods at each colony lasted 20-90 min (X= 73 min) in the morning, and 30- 90 min (X= 61 min) in the afternoon. Two colonies were observed each day. No observations were made during periods of heavy rain. All occurrences of marked forager arrivals were noted by recording time of landing and material carried.
Polybia occidentalis foragers gather four materials. Nectar and insect prey are referred to collectively as 'food materials'. Wood pulp and water, used in nest construction and maintenance, are referred to as 'nest materials'. Methods used to identify materials carried by foragers are detailed in Hunt et al. (1987). In some cases, returning foragers entered the nest or left again before their load could be identified; when this occurred we recorded material as unknown. Trips made by foragers carrying unknown material were not included in our analysis.
The four colonies were collected after the termin- ation of behavioural observations, and all adult wasps were placed in Kahle's fixative solution or frozen. Collections were performed after sunset to ensure that all workers were captured; no workers were observed returning to the nest sites the morning following collections.
Introduction of Known-age Workers
Combs containing pupae were removed from two P. occidentalis nests located more than 100 m from the nearest observation colony and incubated at ambient temperature in the laboratory. These combs served as sources of known-age workers for introduction into the observation colonies. Adult wasps eelosing from the source combs were indi- vidually marked with paint on the thorax and were released within 24 h ofeclosion into the observation colonies. Cohorts of known-age wasps were intro- duced at 2- to 5-day intervals from 26 June to 9 July. The total numbers of known-age workers and cohorts added were: Colony 4:88 workers in six cohorts; Colony 5:105 workers in seven cohorts; Colony 10:80 workers in five cohorts; Colony 13: 105 workers in seven cohorts (mean cohort size = 15, range = 8 20). Introduced workers were accepted into the worker populations of the observation colonies.
Behavioural Observations
Behavioural data were collected between 28 June and 8 August 1989. With few exceptions we collected data from each observation colony every
Data Analysis
We defined foraging tenure as the number of days between a worker's first and last observed for- aging trips. This estimate represents a lower bound on true foraging tenure, since workers could have foraged on days preceding or following those on which they were obscrvcd.
In an effort to analyse exposure to risk, we exam- ined two attributes of foraging behaviour measured for the duration of individual foraging tenures: activity level and material specialization. Forager activity level was quantified in three ways: (1) Foraging rate: the mean number of trips made per hour of observation, under the assumption that risk accumulatcs incrementally with each trip, regard- less of trip duration and of what material was collected. (2) Foraging effort: each trip for a given material was weighted by a factor equal to the mean time to collect that material divided by the mean time to collect water, which required the least time. This activity measure was used under the assump- tion that risk of mortality bears some relation to how long the forager spends in the field. Mean time spent in the field on a foraging trip differed widely among the four materials (O'Donncll & Jeanne

O'Donnell & Jeanne. Forager behaviour and longevity in wasps 1023
1990). (3) Proportion of days that an individual was observed foraging during her tenure. The final measure allowed us to assess the effect of long periods of inactivity on forager longevity.
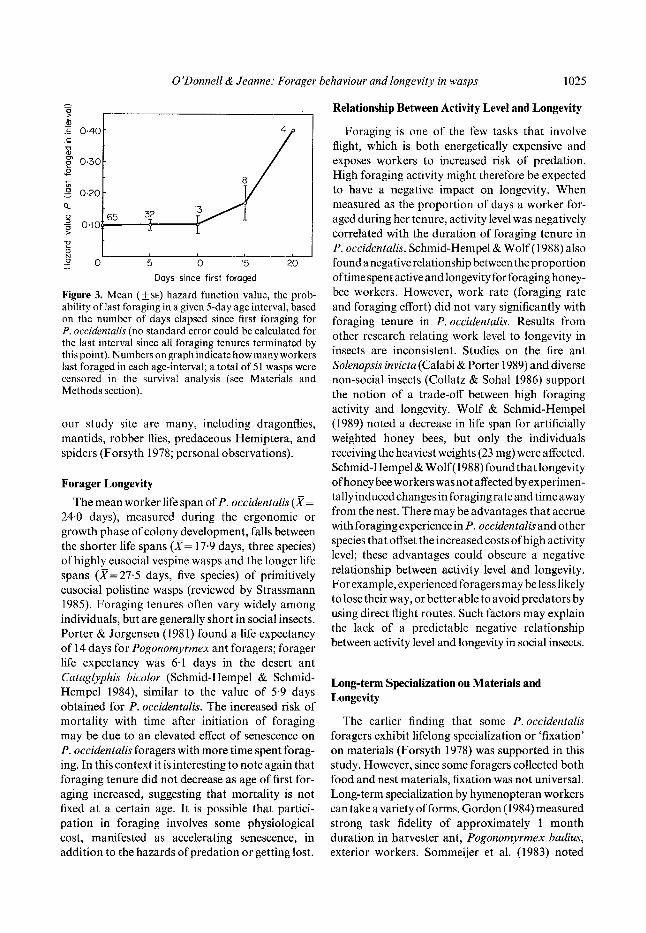
We used life table methods of survival analysis (SAS Institute 1985) to study the effects of material specialization and activity level on the length of foraging tenure. By definition, behavioural attri- butes associated with shorter foraging tenures involved greater risk. We assumed that the observed termination of foraging was due to mor- tality, rather than to changes in worker behaviour such as switching to other tasks. Therefore marked workers present when the colonies were collected were presumed not to have completed their forag- ing careers and were censored in the survival analy- sis. Censoring permits inclusion in the survival analysis of individuals whose true longevity is unknown because observations were terminated before their demise (for further discussion see SAS Institute 1985). The log-rank test was used to assess differences in survivorship distributions among colonies and to test for effects of behavioural vari- ables on forager longevity. This test is appropriate when examining censored data (D. Heisey, per- sonal communication). The hazard function, the probability that a worker last foraged in a given age interval, was used to examine changes in risk of mortality over time.
R E S U L T S
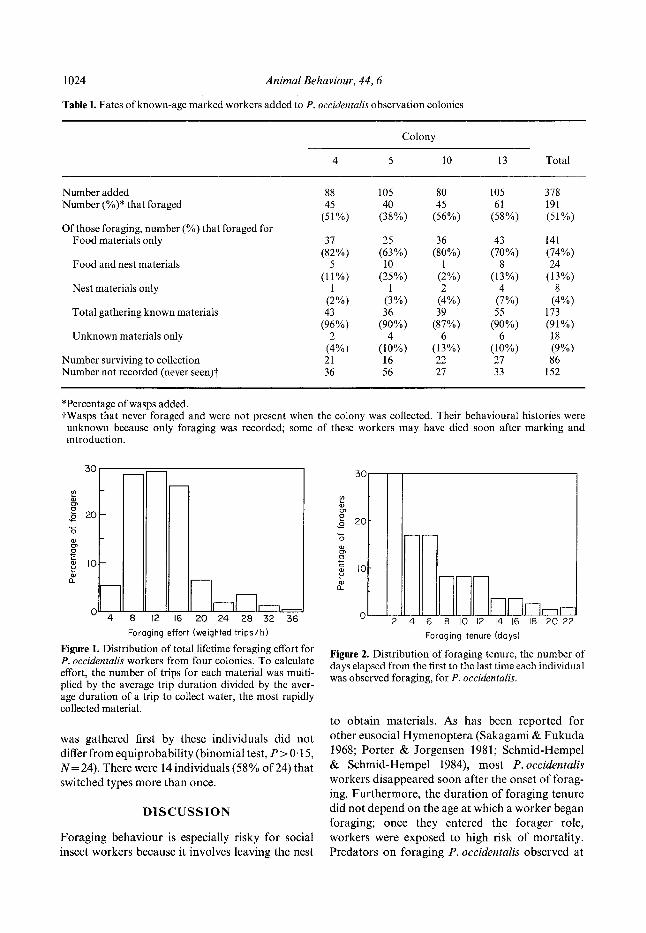
Approximately half of the workers (N = 191) added to the observation colonies foraged during the study (Table I). Of these, 173 workers made trips where the material carried was identified and were included in the analysis of foraging behaviour. The overall mean worker life span was 24 days. Workers began foraging at a minimum of 5-8 days of age, although some individuals did not begin foraging until 25-40 days of age (ranges over colonies). Colonies did not differ in the distributions of adult age at onset of foraging (X_+ SE = 19-1 _+ 0"53 days), of foraging tenure (5-9_+ 0.39 days), or of foraging activity (foraging effort: 11 '2_+ 0.44 weighted trips/ hour; foraging rate: 1.5+0.20 unweighted trips/ hour; Kruskal-Waltis test, df=3, P>0.10 in all cases). Workers within colonies, on the other hand, were highly variable in both foraging effort and the duration of foraging tenure (Figs 1 and 2). Many foragers disappeared from the worker force within a few days of the onset of foraging (Fig. 2) and
tenure did not vary with the age at which workers first foraged (univariate log-rank test, ~2=0-58, P > 0-25).
Relationship Between Activity Level and Foraging Tenure
Survivorship statistics for foragers were calcu- lated at 5-day intervals. Survival analysis of forag- ing tenure showed no significant difference among colonies (log-rank test for equality among colonies, df=3, Z2=3.58, P>0-3). The risk of mortality increased over foraging tenure, since the hazard function, the probability that a worker last foraged in a given age interval, rose with the number of days since first foraging (Fig. 3). Foraging tenure was not significantly associated with foraging effort (mean weighted trips per observation hour; univariate log- rank test, df= 1, Z 2 = 1.39, P > 0" 10) or with foraging rate (the unweighted number of foraging trips per observation hour; univariate log-rank test, dr= 1, Z2=4.4, P>0-10). However, the proportion of observation days during a forager's tenure on which she was observed foraging was negatively associated with the length of her tenure (workers with tenures greater than 3 days, univariate log-rank test, elf= 1, Z 2 = 8-14, P < 0.005).
Specialization on Materials Foraging for food materials was riskier than for-
aging for nest materials: considering all foragers, tenure was negatively correlated with the proportion of foraging effort (weighted trips) devoted to food (Spearman correlation, r = -0"39, P<0 '001, N= 173). The majority of foragers in each colony gathered only food materials or only nest materials (Table I). Foragers that specialized exclusively on food materials began foraging at a younger age (_~= 18.6 days, N = 141) than those that gathered nest materials exclusively (X~= 24.1 days, N: -8 ) (t-test, t = -2 .2 , df= 147, P < 0-05).
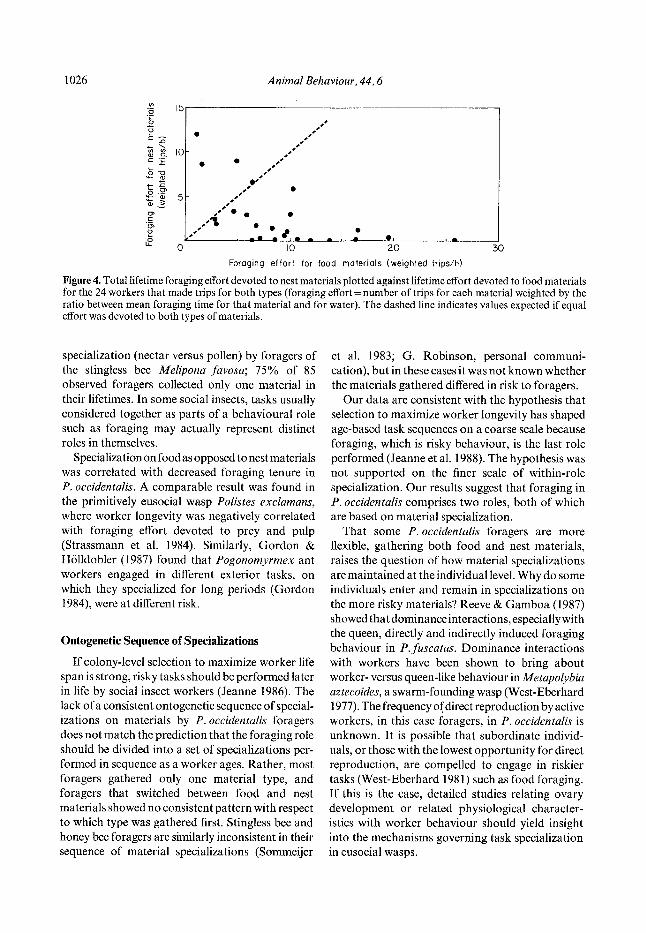
Some workers (N= 24) gathered both food and nest materials. For many of the workers that gathered both material types, there was a tendency to devote the preponderance of foraging effort to one type of material over their entire foraging career (Fig. 4).
Ontogenetic Sequence of Material Specializations For the 24 workers that collected both food and
nest materials, we found no evidence for a con- sistent sequence of material specializations. The likelihood with which either food or nest materials
1024 Animal Behaviour, 44, 6
Table I. Fates of known-age marked workers added to P. oeeidentalis observation colonies
Colony
4 5 10 13 Total
Number added 88 105 80 105 378 Number (%)* that foraged 45 40 45 61 191
(51%) (38%) (56%) (58%) (51%) Of those foraging, number (%) that foraged for
Food materials only 37 25 36 43 141 (82%) (63%) (80%) (70%) (74%)
Food and nest materials 5 10 1 8 24 (11%) (25%) (2%) (13%) (13%)
Nest materials only 1 1 2 4 8 (2%) (3%) (4%) (7%) (4%)
Total gathering known materials 43 36 39 55 173 (96%) (90%) (87%) (90%) (91%)
Unknown materials only 2 4 6 6 18 (4%) (10%) (13%) (10%) (9%)
Number surviving to collection 21 16 22 27 86 Number not recorded (never seen)? 36 56 27 33 152
*Percentage of wasps added. ?Wasps that never foraged and were not present when the colony was collected. Their behavioural histories were
unknown because only foraging was recorded; some of these workers may have died soon after marking and introduction.
50
2 2 o ~6
g
~o 13-
I .
8 12 16 20 24 28 32 36
Foroging effort (weighled tr ips/h)
Figure 1. Distribution of total lifetime foraging effort for P. occidentalis workers from four colonies. To calculate effort, the number of trips for each material was multi- plied by the average trip duration divided by the aver- age duration of a trip to collect water, the most rapidly collected material.
was gathered first by these individuals did not differ f rom equiprobabi l i ty (binomial test, P > 0-15, N = 24). There were 14 individuals (58% of 24) tha t switched types more than once.
D I S C U S S I O N
Foraging behav iour is especially risky for social insect workers because it involves leaving the nest
30
u~
c~ o
~6
g ~3
y
20
4 6 8 I0 12 14 16 18 20 22 Foraging tenure (days)
Figure 2. Distribution of foraging tenure, the number of days elapsed from the first to the last time each individual was observed foraging, for P. occidentalis.
to obta in materials. As has been reported for other eusocial Hymenop te r a (Sakagami & F u k u d a 1968; Por te r & Jorgensen 1981; Schmid-Hempel & Schmid-Hempel 1984), most P. occidentalis workers d isappeared soon after the onset of forag- ing. Fur the rmore , the dura t ion of foraging tenure did not depend on the age at which a worker began foraging; once they entered the forager role, workers were exposed to h igh risk of mortal i ty. Predators on foraging P. occidentalis observed at
O'Donnell & Jeanne." Forager behaviour and longevity in wasps 1025
.=- 0.40
. -
0.30 ,2 "6
0,20 cL
~ o.loi -o
Q O I
65 32 T I
i i
s ,'o r5 ab Days since first foraged
Figure 3. Mean (+SE) hazard function value, the prob- ability of last foraging in a given 5-day age interval, based on the number of days elapsed since first foraging for P. occidentalis (no standard error could be calculated for the last interval since all foraging tenures terminated by this point). Numbers on graph indicate how many workers last foraged in each age-interval; a total of 51 wasps were censored in the survival analysis (see Materials and Methods section).
our study site are many, including dragonflies, mantids, robber flies, predaceous Hemiptera, and spiders (Forsyth 1978; personal observations).
Forager Longevity
The mean worker life span ofP. occidentalis (.Y= 24.0 days), measured during the ergonomic or growth phase of colony development, falls between the shorter life spans (X= 17.9 days, three species) of highly eusocial vespine wasps and the longer life spans (X=27.5 days, five species) of primitively eusocial polistine wasps (reviewed by Strassmann 1985). Foraging tenures often vary widely among individuals, but are generally short in social insects. Porter & Jorgensen (1981) found a life expectancy of 14 days for Pogonomyrmex ant foragers; forager life expectancy was 6.1 days in the desert ant Cataglyphis bicolor (Schmid-Hempel & Schmid- Hempel 1984), similar to the value of 5-9 days obtained for P. occidentalis. The increased risk of mortality with time after initiation of foraging may be due to an elevated effect of senescence on P. occidentalis foragers with more time spent forag- ing. In this context it is interesting to note again that foraging tenure did not decrease as age of first for- aging increased, suggesting that mortality is not fixed at a certain age. It is possible that partici- pation in foraging involves some physiological cost, manifested as accelerating senescence, in addition to the hazards of predation or getting lost.
Relationship Between Activity Level and Longevity
Foraging is one of the few tasks that involve flight, which is both energetically expensive and exposes workers to increased risk of predation. High foraging activity might therefore be expected to have a negative impact on longevity. When measured as the proportion of days a worker for- aged during her tenure, activity level was negatively correlated with the duration of foraging tenure in P. occidentalis. Schmid-Hempel & Wolf (1988) also found a negative relationship between the proportion of time spent active and longevity for foraging honey- bee workers. However, work rate (foraging rate and foraging effort) did not vary significantly with foraging tenure in P. oecidentalis. Results from other research relating work level to longevity in insects are inconsistent. Studies on the fire ant Solenopsis invieta (Calabi & Porter 1989) and diverse non-social insects (Collatz & Sohal 1986) support the notion of a trade-off between high foraging activity and longevity. Wolf & Schmid-Hempel (1989) noted a decrease in life span for artificially weighted honey bees, but only the individuals receiving the heaviest weights (23 mg) were affected. Schmid-Hempel & Wolf(1988) found that longevity of honey bee workers was not affected by experimen- tally induced changes in foraging rate and time away from the nest. There may be advantages that accrue with foraging experience in P. occidentalis and other species that offset the increased costs of high activity level; these advantages could obscure a negative relationship between activity level and longevity. For example, experienced foragers may be less likely to lose their way, or better able to avoid predators by using direct flight routes. Such factors may explain the lack of a predictable negative relationship between activity level and longevity in social insects.
Long-term Specialization on Materials and Longevity
The earlier finding that some P. occidentalis foragers exhibit lifelong specialization or 'fixation' on materials (Forsyth 1978) was supported in this study. However, since some foragers collected both food and nest materials, fixation was not universal. Long-term specialization by hymenopteran workers can take a variety of forms. Gordon (1984) measured strong task fidelity of approximately 1 month duration in harvester ant, Pogonomyrmex badius, exterior workers. Sommeijer et al. (1983) noted
1026 Animal Behaviour, 44, 6
15
~ z
~ ,o
o
se Sr �9 r
s S �9 �9 s,~s"
,O" os S
s~ r �9 �9 �9
, - . . . . s . . . . �9 " o , . .
I0 20
Foraging effort for food materials (weighted trips/h)
30
Figure 4. Total lifetime foraging effort devoted to nest materials plotted against lifetime effort devoted to food materials for the 24 workers that made trips for both types (foraging effort = number of trips for each material weighted by the ratio between mean foraging time for that material and for water). The dashed line indicates values expected if equal effort was devoted to both types of materials.
specialization (nectar versus pollen) by foragers of the stingless bee Melipona favosa; 75% of 85 observed foragers collected only one material in their lifetimes. In some social insects, tasks usually considered together as parts of a behavioural role such as foraging may actually represent distinct roles in themselves.
Specialization on food as opposed to nest materials was correlated with decreased foraging tenure in P. occidentalis. A comparable result was found in the primitively eusocial wasp Polistes exclamans, where worker longevity was negatively correlated with foraging effort devoted to prey and pulp (Strassmann et al. 1984). Similarly, Gordon & H611dobler (1987) found that Pogonomyrmex ant workers engaged in different exterior tasks, on which they specialized for long periods (Gordon 1984), were at different risk.
Ontogenetic Sequence of Specializations
If colony-level selection to maximize worker life span is strong, risky tasks should be performed later in life by social insect workers (Jeanne 1986). The lack of a consistent ontogenetic sequence of special- izations on materials by P. oecidentalis foragers does not match the prediction that the foraging role should be divided into a set of specializations per- formed in sequence as a worker ages. Rather, most foragers gathered only one material type, and foragers that switched between food and nest materials showed no consistent pattern with respect to which type was gathered first. Stingless bee and honey bee foragers are similarly inconsistent in their sequence of material specializations (Sommeijer
et al. 1983; G. Robinson, personal communi- cation), but in these cases it was not known whether the materials gathered differed in risk to foragers.
Our data are consistent with the hypothesis that selection to maximize worker longevity has shaped age-based task sequences on a coarse scale because foraging, which is risky behaviour, is the last role performed (Jeanne et al. 1988). The hypothesis was not supported on the finer scale of within-role specialization. Our results suggest that foraging in P. oecidentalis comprises two roles, both of which are based on material specialization.
That some P. occidentalis foragers are more flexible, gathering both food and nest materials, raises the question of how material specializations are maintained at the individual level. Why do some individuals enter and remain in specializations on the more risky materials? Reeve & Gamboa (1987) showed that dominance interactions, especially with the queen, directly and indirectly induced foraging behaviour in P.fuscatus. Dominance interactions with workers have been shown to bring about worker- versus queen-like behaviour in Metapolybia aztecoides, a swarm-founding wasp (West-Eberhard 1977). The frequency of.direct reproduction by active workers, in this case foragers, in P. oecidentalis is unknown. It is possible that subordinate individ- uals, or those with the lowest opportunity for direct reproduction, are compelled to engage in riskier tasks (West-Eberhard 1981) such as food foraging. If this is the case, detailed studies relating ovary development or related physiological character- istics with worker behaviour should yield insight into the mechanisms governing task specialization in eusocial wasps.

O'Donnell & Jeanne: Forager behaviour and longevity in wasps 1027
A C K N O W L E D G M E N T S
We t h a n k Werner and Lilly H a g n a u e r for their ongoing suppor t of our research, especially for permiss ion to work on their land. La r ry Phelps pro- vided valuable assistance in the field. Jos6 Ar tu ro Le6n and the Costa Rican office of the Organiz- a t ion for Tropical Studies were helpful in obta in ing field supplies. Dennis Heisey's advice on statistical analysis of survivorship da ta was greatly appreci- ated. T h a n k s to Susan Bulova, Emil ia Mart ins , Joan S t rassmann and an a n o n y m o u s referee for reading and comment ing on earlier versions of the manuscr ipt . Our research was funded by N S F grant BNS-8517519, R.L.J. principal invest igator; and the Univers i ty of Wisconsin College of Agriculture and Life Sciences.
R E F E R E N C E S
Calabi, P. & Porter, S. D. 1989. Worker longevity in the fire ant Solenopsis invicta: ergonomic considerations of correlations between temperature, size and metabolic rates. J. Insect Physiol., 35, 643-649.
Collatz, K. G. & Sohal R. S. (Eds) 1986. Insect Aging. New York: Springer Verlag.
Daubenmire, R. 1972. Phenology and other characteristics of tropical semi-deciduous forest in North-Western Costa Rica. J. Ecol., 60, 147-170.
Forsyth, A. B. 1978. Studies on the behavioral ecology of polygnous social wasps. Ph.D. thesis, Harvard University, Cambridge, Massachusetts.
Franks, N. R. 1987. The organization of working teams in social insects. Trends Ecol. Evol., 2, 72-75.
Gordon, D. M. 1984. The persistence of role in workers of the harvester ant, Pogonomyrmex badius. Psyche, 91, 251-265.
Gordon, D. M. & H611dobler, B. 1987. Worker longevity in harvester ants (Pogonomyrmex). Psyche, 94, 341-346.
Hcinrich, B. 1976. The foraging specializations of indi- vidual bumblebees. Ecol. Monogr., 46, 105 128.
Hunt, J. H., Jeanne, R. L., Baker, I. & Grogan, D. E. 1987. Nutrient dynamics of a swarm-founding social wasp species, Polybia occidentalis (Hymenoptera: Vespidae). Ethology, 75, 291-305.
Jeanne, R. L. 1986. The evolution of the organization of work in social insects. Monitore Zoologico Italiano N.S.,20, 119-133.
Jeanne, R. L., Downing, H. A. & Post, D. C. 1988. Age polyethism and individual variation in Polybia occidentalis, an advanced eusocial wasp. In: Inter- individual behavioral variability in social insects (Ed. by R. L. Jeanne), pp. 323-357. Boulder, Colorado: Westview Press.
O'Donnell, S. & Jeanne, R. L. 1990. Forager specialization and the control of nest repair in Polybia oecidentalis Olivier (Hymenoptera: Vespidae), a tropical swarm- founding wasp. Behav. Ecol. Sociobiol., 27, 359-364.
O'Donnell, S. & Jeanne, R. L. 1992. The effects ofcotony characteristics on longevity and foraging behavior of individual wasps (Polybia oeeidentalis, Hymenoptera: Vespidae). Insects Soc., 39, 73-80.
Porter, S. D. & Jorgensen, C. D. 198l. Foragers of the harvester ant Pogonomyrmex owyheei: a disposable caste? Behav. Ecol. Sociobiol., 9, 247-256.
Post, D. C., Jeanne, R. L. & Erickson, E. H. Jr. 1988. Variation in behavior among workers of the primitively social wasp Polistes fuscatus variatus. In: Interindividual Behavioral Variability in Social Insects" (Ed. by R. L. Jeanne), pp. 283-319. Boulder, Colorado: Westview Press.
Reeve, H. K. & Gamboa, G. J. 1987. Queen regulation of worker foraging in paper wasps: a social feedback control system. Behavior, 102, 147 167.
Rissing, S. W. 1981. Foraging specializations of indi- vidual seed-harvester ants. Behav. Ecol. Sociobiol., 9, 149-152.
Sakagami, S. F. & Fukuda, H. 1968. Life tables for worker honeybees. Res. Pop. Ecol., 10, 127-139.
SAS Institute. 1985. SAS Users Guide." Statistics. Version 5 edn. Cary, North Carolina: SAS Institute.
Schmid-Hempel, P. 1984. Individually different foraging methods in the desert ant Cataglyphis bicolor (Hymenoptera, Formicidae). Behav. Ecol. Sociobiol., 14, 263-271.
Schmid-Hempel, P. & Schmid-Hempel, R. 1984. Life duration and turnover of foragers in the ant Cata- glyphis bicolor (Hymenoptera, Formicidae). lnsectes Soc., 31,345-360.
Schmid-Hempel, P. & Wolf, T. 1988. Foraging effort and life span of workers in a social insect. J. Anim. Ecol., 57, 509-522.
Sommeijer, M. J., De Rooy, G. A., Punt, W. & De Bruijn, L. L. M. 1983. A comparative study of foraging behavior and pollen resources of various stingless bees (Hym., Meliponinae) and honeybees (Hym., Apinae) in Trinidad, West Indies. Apidol., 14, 205-224.
Strassmann, J. E. 1985. Worker mortality and the evolution of castes in the social wasp Polistes exclamans. Insectes soc., 32, 275-285.
Strassmann, J. E., Meyer, D. C. & Matlock, R. L. 1984. Behavioral castes in the social wasp, Polistes exelamans (Hymenoptera: Vespidae). Sociobiology, 8, 211-224.
West-Eberhard, M. J. 1977. The establishment of repro- ductive dominance in social wasp colonies. Proceedings of the VIII Congress on International Union Study of Social Insects, pp. 223 227.
West-Eberhard, M. J. 1981. Intragroup selection and the evolution of insect societies. In: Natural Selection and Social Behavior (Ed. by R. D. Alexander & D. W. Tinkle), pp. 3 17. New York: Chiron Press.
Wolf, T. J. & Schmid-Hempel, P. 1989. Extra loads and foraging life span in honeybee workers. J. Anim. Ecol., 58, 943-954.





























