Embed Size (px)
Citation preview

448 SAMUFLR. RUSMFORTH ANDJF.KFRFY R.JOHANSF.N
Fhrb. MTct, Stmphna OMf. Mfdd. Komm. Ilavundfn. Sfr. Plank-
faulwn, O. 1905. On wrnic PeriHinia aiiHMfdd. Komm. HavundfTi. Srr Pliinkinn |(3):3-7.
Prmhkina-Lavrrnko. A. I. 19^3. Planktonic fhritomi oj thf AzSfa. Bot. lmt. Komarova, Mincnw. 190 pp., 9 pi. (In
Tukario, II. 19fi-l. Uiutoni I'liltiirc in aruCuial M-a watcr-ll. Cul-lurr* wiihoul u>iriK M>il fxtracl. BuH. Tokai Hfg. Fith. Rfi.i.ah. 3H;ir)-r>r). i pi.
1968, On ihc<liari)niC'iMr/(Wfro«r«/rirr«m(I*aulwn)emend.anti iis ciu'iirf lorm pumthn forma nov, Bull. Takai Rfg. Fuh.tifi. Lah. 65:1-6. 1 pi.
/ Fhyfol. « . 448-155 (1986)
LIFE HISTORY VARIATIONS IN A PREDOMINANTLY GAMETOPHYTICPOPULATION OV IRIDAEA CORDATA(GIGARriNACEAE. RHODOPHYTA)'
Georgiana May
Drpartmrnt of Butany, Univcrsiiy of California. Berkeley, Calilbmia 94720
ABSTRACT
An Iridaea cordata (Turnfr) Bory population madeup of 83% gametophyles and I77t tetrasporopbytes dem-onstrated little varialion in these proportions over threeyears. These data conlra.it with an expectation that alter-nations of generations, as demonstrated by culture studies,should result in equal numbers of isomorphic generations.Analyses of reproductive and demographic attributes ofthe populatioti were made to better undersland the role ofalternation of generations in this population.
Grouth and mortality rales in the sumtner could notaccount for the success of gamelophyles. hcni'e\'er, site classfrequencies demonstrated that tetrasporophytes were small-er individuals. Sporelings recruited into this intertidalregion and made up 20% of the spring population num-bers. Even wUh ihis In-el of rerruitment, gametophyte-to-tetrasporopbyte ratios did noi change. Perennaling htild-fasts regenerated 80% nj the spring nutnhen atid 90%of the tolal blade area. Consequently, perennation anddifferential iurvtval of gnmetnphyte spores or iporelingsare proposed as mechanisms maintaining gatnetophytedominance in this San Juan Island, \\'a.\hitigton popu-lation.
Key index words: alternation of generations; Iridaeacordata; life history variatmna; population .strurlure
An isomorphic alternation of generations has beendescribed for many red algae using laboratory cul-ture techniques. Much less is known of life historiesin natural populations though relative abundance ofisomorphic phases has often been used as indirectevidence for the life history strategy of the species.For example, some authors have interpreted thefinding of equal numl>ers of isomorphic phases tomean that alternation of generations is an importantmechanism of regeneration (F.dwards 1973, Dawes
' Afffplfd I Juh 1986
el al. 1974), When unequal representation of lifehistory phases is evident, the conclusion may be thatalternation of generations is not as important as oth-er means of regeneration (Dawes etal. 1974,Hansenand Doyle 197(i, Craigie and Pringle 1978). Appar-ently, equal numbers of alternate phases were ex-pected but a life history model was not made explicitin the above papers. The model implied could bestaled as: wilh alternation of isomorphic genera-tions, eventually the numbers of the two phases willeven out. If the non-sporebearing male gameto-phytes are included, the expected ratio will be dif-ferent ihan if they are not.
The purpose of ihis study was to test assumptionsunderlying the question of whether populationstructure is predictive of the process of alternationof generations, assumptions inherent to the life his-tory mtKlel. These assumptions are that (I) the plantsare annual and that the population is regeneratedby spores and thai (2) the two spore prf)ducing phas-es have similar demographic properties of growth,survival and reproduction, 1 studied an intertidalpopulation o^ Iridaea cordata (Turner) Bory on SanJuan Island. Washington because preliminary stud-ies showed that il was predominantly composed ofthe gamelophyte generation and because this specieshas an isomorphic alternation of generations (Kim1976) in culture.
Natural populations do not always have equal pro-portions of isomorphic generations. Numericaldominance of tetrasporophytes in /. cnrdata popu-lations was reported by Ilansenand Doyle (1976) atMonterey, CA, and by Dyck et al. (1985) (or siteson the Oregon coast. Sporophyte dominance ha*been demonstrated for populations of other alga^-Ceramium sp. (F.dwards 1973) in the F.nglish Isles,Gelidinm robu.Uum along the southern California coast(Barilotti and Silverthorne 1972), Hypnea mu.^riformliin India (Rao 1970), and h.ucheitma isnforme on th*Florida coast (Dawes et al, 1974),

LIFE MIS rORV OF IRIDAEA CORDATA 449
Gamelophyte dominance has been reported lessoften ihan has sporophyte dominance. Iridaea cor-datct populations in Washington and Briiish Colum-bia were (M-i% to 959[ gameiophytic (Dyck et al.1985). Far Cboudrus ciispus the raiioof ihe iwo phas-es changes along a vertical gradient with gameto-phyte plants more abundant in upper subiidul /onesand sporophytes dominant in deeper areas (Mathie-son and Burns 1975, Craigie and Pringle 1978).
Both reproductive and demographic mechanismshavf been advanced to accotuii for unequal repre-stMitation of isomorphic phases. Reproduction byasexual spores, rather than by annual alternation ofgeneralions, has been proposed (Dixon 1965, Dawesetal. 1974. Mansenand Doyle 1976, Whittick 1978).Apogamic carpos|)ores and apomeiotic tetrasporescould regenerate stable populations of female ga-metophytesand teirasporophyies, respectively. Kim(1976) showed that, in a few cases. /. cordata's tet-raspores could produce tetrasporophytes. However,data collected in this field study suggest that per-ennation and not asexual spore production is thereproductive mechanism primarily responsible forcontinued gametophyie dominance.
Demographic difTerences between isomorphicgenerations are implied for C. crispus by the resultsof Craigie atid Pringle (1978), since the authors ob-served the Iwo phases living in different habitats. 1compared growth, survival and reproduction ratesof gameiophytfs and sporophytes to see if differ-ences in these rates were responsible for gameto-phyte dominance.
rhe following hypotheses were formulated andtested: (1) Alieniatioii of generations is occurringon a longer titne scale than previously considered.The dominant life histt)ry phase might alternate (orreverse) on a year-to-year basis or wheti a cata-strophic event opens siics for coloniz;ttion. (2) Be-cause of perennation from bas;il holdfasts, altertia-tion of getieratiotis is less common than expectedfor annual plants. Perennation acts to stabilize thecurrent gametophyie dominance by dampeningfluctuations in the relative abundances of the twophases caused by sporeling recruitment. The firsttwo hypotheses test the assumption that the plantsare annual and that regeneralion is by spores.
The next two hypotheses test the assumption thatitliernate phases are deniographically equivalent. (3)Alternation of generations may occur, but game-lophytes are better able to survive and grow in thepopulation studied. Having higher growth or sui-vival rates than the sporphyees, gamet<Jphytes be-come more abundant than sporophytes. (4) Alter-nation of generations may occur but sporophyteshave a higher reproductive rate than do gatneto-phyies. 1 etraspores are therefore more numerousthan carpospores and produce the gametophyte-dominani population structure observed.
These hypotheses are not mutually exclusive. Acombination oriilternaiion of generations by spores
and other means of blade regeneration could pro-duce the patterns of gametophyte dominance ob-served in this population.
METHOt>S AND MATERIALS
Field mcaMircmrnts for ihis siutly were made over a three-yrarperitHl. In 1980, prrlimin;iry sampling indicaicd gametophyicdomin,ince; in 1981, summer pritwih. mortaliiy and rrproductionrates were measuretl and holdfaM*' j>eniisiencc noird; in l!>92.whuer-spring reiruiimeiii irom holdfasl* and sjx)rclinKS wasquamificd. The Mudy iiir and ihe l«:hni(|iics fur blade measurc-meni. for Rrowih raie cakiiLitions and for determininn life hivlorv phaM-% were ihe same in alt sampling jx-riods and arc de-scribed beli)w. Sampling simiegies were spetific l<> lheex)ierimentslesiinn ^UlKHhe^es 2, 3 and 4 arul arc deM-ritied for each. Forhyf«nhcsis I. where life hiMorv phase ratios were observed, iam-pllng >.traU-Kies followed ihose iisetl for h>poiheses 2. 3 and 4.
Slud\ <,itf. Ihf siudy sire (Fig, I) was on the south »idr of RockyB:iy, ai Rubin 1 arie Stale Park, l<K.i(cd on thr northeasi side ofSan Juan Ubnd. Washinglon (49'38'45' N, 123*afl"I5' W), Theinterlidal/. cordata [MJpuIaiion investifjaled vs'assiiuatedona nonh-facing kisall suhstraium within the vertical elevations of + I mand -1 m with respect loMLLW. Thesufwtratum included botha gently sloping shelf and n>rk faces. Wave aclion was generallylowai lliissomewhai proiriied site. I h e morphology ofihe plantsfii wiihin Abliott's (1971, 1972) description of /, cordata \-ar.lordnta and /. cnrdata \-ar. yplrndrfu; no blades were attributed lo/, fiiimtU. I'idal elrvaiions were eMimated by standard surveyingtechni(|ues using for refrrenre a U.S. Coast atid Ge<Kiesic Surveymarker located on a near-by bluff,
Bltidf mfasurfmntty Blade sizes were measured as the one-sidedarea of a blade and area was estimated as a product ol length ^width, If a blade consisted of several lotKrs. each lobe was mea-sured se|wrately and the products summed to estimatcihe blade'sKMal area. The pr«xltict of length >< widtli ix Imearly related tothe weight of a thallus for this alga (Hansen 1976), This methodallows rapid field measurement of ihalli without destructive sam-pling. Blades repeate<lly measured for growth rates were taggedar mapi»cd by methods described Ix-low for Hyjwnheses 2 and 3.
Grovlh ralffalrulntwui. Growth rates and relative gTowth rateswere raltulated using tbe following Tormulje (Hunt 1982): (1)Growth r;tte(t, to t,) - Iarra(t,) - area(t,)l/(t, - t,) with growthrate t'\prrssedascm* day ' and (2) relative growth rate(t, toti) -growth nileft, to i,)/iirea(l,) wiih relative growth rate as cin'-da> ' c m •. Growih rate and relative growth rate (t, to i,) arerates calculated for (he ititer\all>etween lime 1 and time 2. Areas(t, . , . i j are blade areas at each measurement time (t, . . . tJ .Sampling limes, in days, represent time elapsed since the begin-ning o( the measurements (t, . . . t^).
Dftfrmination of ptjmfinphyif and irtra^porophytf thalti VtstuI ob-servations were made in ihe field to categtiri^e life history phasesof reproductive ihalli. When nrces«ir\\ pnnicms of blades werereturned to the labf>raton' to confirm the rcproduclive wate bvmaking microscopic observations of rcpnxluciive structures,
\<in-repr«xluciive gamcti>phyte and tetras|x>rophyte individ-uals were diHercntiated by analyzing for the presence cir absenceof K-carrageenan. «-carraReenati is found in gametophytes but isabsent or in very low concentrations in tetrasporophytes whichcontain A-carrageenan (McCandlevs ei al. 1975). Tbe presence of*-carrageenan was deiermined by excising a small (1 cm*) sectionof blade material and using the resortinol-acetal test for 3-6anhydrogalactose characteristic of it-carrageenan (Craigie andLeigh 1978). Controls of ktiown Rametoplutes and sporophyteswerr included in all sets of rests. All blades on a gi\Tn hnidfaMhad the same type of carrageenan.
Sampling \tralff[if\. Si ecific sampling prot(Ki>ls were desigtiedto perform the experiments necessary to test each of the fourhy|M>theses listed previously. They are as follows;

450 C;KOKC;IANA MAY
V \M I I . fifliitr- r tiliu'fliiuif i''i Ifihii\r<, lit tiiihin Tniff Stiitr Park. Jl u
IILKM t iinl.ii.i
Ktr.l l i r
A <lriail<-<) Mi<- in;i|* v h o u i n n i h rir.tli%r<l% ju r l |>i)iiits .lli't .1 K('[i(-I.iti/<'<1 VICM I ] ^ pI h r \xut\\ Mir IS >nt I lir Muiilirri i \ l i i i r r ol HIM kv 11^ \ r.,iiniv*rii
m i i iwt i liiM-t niitp s h i m v l l i r IIK ;tti(in •>) KIK l \ h.iv on San J u u nll ID r h c .SttliitfKis ol ( • r ( l r^ l4
piithrM^ t u t forftni'i plant* h^ivr tiiKh r;iir% «>! lui iiovrr miflc r r i ru i fmrn i r a t h yrar , th ru rr%cjsj | ol ilu-diiniinaiH (ili.isr
or 1.1 tKr HIM tti,ill«itt% iii irl . i l tvr i)uiii)H-r^ ut t h r I MO ph.I vrs sliniili)hr i»hsrrvr<l nti ;i vr.ii-lo-scir b.isis. I tnv<-MiK.ilril th r j>ro|ioi tioMtit ((•'">•*'ophvIrs and trtias(M>iiijilivtr's thai (f>Ni|»<t%r(l ihc loLiI)xipiil.iliiiii(liiriliK t h r r r (hllriciil )>rrio<l%, siiTiiniri I DHO.MtmnicrI ' J S I . jnci \ p t i f i K i " r . s r l \ su r imi r r \\tH'2. In i h r la t ter (w<>iirri(Kls,t pUimrd l(i vjiiiplr all [Jl^K<'(l " r niap)M-il ihalli
111 |»rrliminar% satnptnin. whrii t(.iniihiph>tr <l<iniiiiariir wa%first n o i n l . r rp t rs r i i ta i ion ut r a t h phasr Wiis r \ t in iJ i r t l .il oi i r<l.tir. jtils 11), r.'HO 1 v*o tr.iiisri is. piirallrl lo ih r shor r ur * 0 5m And 0 '> m litl.il hrij;hi w r r r M'I up. r.n h *\.is .if)piii\itiialrl>iQ m IniiK .\( )M>inls I rn apart . JII hl.idrs ol pl.iiits IWIK hiii(( i h rir.ins<'it wr t r i[.iihrrr(i l.ilr liisiors ph;iM* M.IS d r i r r m i n r d lorA\\ Madr* col lcdrr l . fi>r A it it a I of \'U> ttLirlrs
RrprrsriK jiioii ol litr histnrs ph;iM-\ w;is .i^.nii .issrsscrl diinii)(ihr Mimnirr 1'>HI vthrn I lan^rd 'iU'.t bladrs Inr i rpca t rd ohsrr-\.iiiiin (drvrit^ird iinrlri l i i iKiihrsis .'i\, Siii.dl pirc rst i t Mi'.' t.i^ni'dihalli UpproximairK '2 ' 'J < ni 'j w r i r icrtum-d ami usc-d Inriarrai ; r r i i . in ar).d\\is hor 10 plants, lilr hislnrv ph.isr n;is dc i r r -mined h\ itis(M-< it'Hi ut rrptiHhii t i s r strtM tirrrs a lnnr
In spnn(( l''H',?. ph;iM' r a t u ^ w r r r mrasnrcd .1^^111. ['hnil> weremaiiprd r J th r r th^ii tanurt! (tlrw rtl)rd uridri ttv|Mithrsi% '2) ^nrlin Junr I'.'M'J. planc\ s*tlh suttn it rii t>l,idr rn.iirr i.d w r r r \.implr<ltn d r i r r n i i n r Iitr hiMorv phasr I nlur tunalr lv , alM>ul TiO sampir*v . r i r li>st tn j i i a««iilrnt nt iru onnrin (idr In tin* y.A\. 'I'i pl.nit^w r i r viniplril .All additional 11 plains ti.id drv(ln|jr<i rrpii>dur-( u r s tnu till r s ami lite hisrr»r\ ph,iw<oul<l tir d r t r i nnnrd visually.
thti"ttir\i', 2 It (K-rrntiJtioii is .in iin)H)ri.int rnrfhatiistn lorhiarlr r r n r n r r jt inn. ihrii s[Mirrlirn{ r n t nitltirnt stimtkl IM* ur ata lot^rt Irsr l ihaii l'U jiiniial planis 1 rs i imatr i l r n niiriiH-nt Itnni
(n - Till
C.atlirlriphvtrvIfi.-l
ML* S-
17 2 17.7
ditlri (roni 'i.\ -ind l-l rnlms nt
^pnrclit)((s a n d p r r r n i i i i l m n iri>ni rx iMinj ; hnUil.ists in n r d r r t "t o n i | ) a r c t h r s r t w o itirt li.iinsrtis nf i r ^ r n r i a r i o i i I n t ;ni l i l i i tci r in iMin i^ p<-rrnn.itinK l io ld las t s a n d lo r ( - ioK ' i i '< ' ^p^r i liiiK ir*-4 l u i l s . a s in ip l r w s i r r n nt m a p p i n g / inTiiatii U u i d f a s l s w a s d r v i s n l .I llf r r I fans r i ts hrr<- r s t . i l i l i shrd : vAi h M<IS | i r i prii i l l i lilai lo i h r
Nhorr a n d rx t r i id i i iK Hi 111 ftoiii .1 iid.il ( I r v a l m n ol a b o u t t 11.5ni lo - O.H tn. T h r s r IIK l i id rd ;ind r x p a n i l c d ti)«>ii l l i r a t r a viitl-p l r d in s i i tnn i r r I'.IHO Alon j ; r t ir i h r r r t t . i n s r i ts . a loia l ot r i n h tpo in l s . 2 5 - T) ni a p a r t . w e r e n i . i i k r d t>\ itiMlliiiK •• n i r t a l ))iii in ' t )a l ioir f i n l i r d iti t h r nw k (Ki^. I). I l i r p ins w e r e r i l h r r st;ntilrMv>trr l thnilM-t 's Iinlts or tiravv ^ a h a n i / r d ii.nls wiih plasiu v\-p a n d a t i l r s | r i-vrs, I hroii){h i j i h p i i in i . a t o tnpass t r f r T i ' t u c l inewas ) hns r i i ^.t»^^ holdl. ist »*ilhm .1 TiO < ni t^idiiis of t h e piM wa*ttt.ip|Md iiMti(* pnla i ( i i o i d i n . i t r s . tll.it is, rstalil ishiiiK an atl^l(*tri ini l l i r i r l r i r i u r tiiir aii<l .1 ths i j t i i i r t rnn i t h r pin tn l l i r plat i l .l ' \ < r p t III A \vv. I iisrs, iiiflividti.iK tn i i ld In- r^isily d i U n r i i t i a i r d;ind t r i o t a i r d MSIHK 'li '" ' s\Atriti.
I . iH.itions (if 'itYl hofd ias t s wi-ii- i n . i pp rd | a t iua rv 2 - t i . 111^2.f'laiits i r u r n r i . i t i t i K h o r n ho ld las r s ar<- r r l r i r c - d In .is "lioldfiistpl . i t i l s ." OtiK a frw lat j-r . o s r t \ M t i l r t iti^ li l . idrs w r i r t i l i w r v r uInn t d r n t i t i i a l i n n of / omtnln hnldf. is ts was a u i r d 1)V i h r p r r s r t l i fof t n o t r i i u i n r r o u s sitiall l i l . idr \ (• 'J i in ' ) .
Attri .ipptoxitnalrlv I .Ti nm. in April I'.IH'J, rnapprd hohllast*wrr<- rr|n<arrii f he prr'srrii r <ir .ilisrnn- of holdlasts was rf-(iirrlrd iiiirl hla<li-s tni inird and riu'iisurrd as br ior r . Aliwiitc (>•;i tTiapiH-(l liolillast was i r t n r d r d .is niortahtv nt thr plant. No
100
3ao
oo
Holdfast
Sporeimg
8 - n Jon 2 4 - 8 Apr
Kli. 2 Krla1l^r lo i i i f i l i i i l lo l l nt hitltif,|i» i h r spiiriK MtH'J |Kipii1ation ini tnlx-rs .ni.ip|K-il in Api i1 as di si i iltc-it in i lir ii st.
aii<l
21-24 June
plan'

LIFE HISTORY OF miDAK.\ CORDATA 451
*IOOO-lOOO
- 5 0 0
XC
ffl
Eu
ZE
S (
m
-AP
E
•_*tn
51-1000-50
"lOOO-1000
0 3
SPORELING PLANTS
-500
-200- 1000-60
B
0 2
92 6
Fiti. 3A-D.
HOLDFAST PLANTSAPRIL-SIZE CLASS FREQUENCY (%}
Size class frequencies of /. r
•20001501-20001001-1500
-1000
-500
51-1000-50
>2000l5Or-2OOO1001-1500
-1000
-500
51-1000-50
A
0 40 4
092330
92 6SPORELING PLANTS
D
90 3
HOLDFAST PLANTSJUNE-SIZE CLASS FREQUENCY (%)
nm blades for sporcling and holdfast populatioiu. Spring 1982. A April populationplants. B, April jioputaiion, htildfaw rcRcncrated plants. C. June population, sporcling plants. D. June population holdfajt'ed plants.
plani.1 bore reproductive structures. Unmapped plants located insample area* were assumed to be sporclingx and are referred tohere as "sporeliiig plants." TheM* had imallcr holdfasts and fewerblades than those plants arising from established holdfasts.
Six mo (June 21 -24) after initial mapping thalli were relocated.A-s in the previous samplitig period, mortality was recorded forpreviously mapped plants no longer present and new plants ap-pearing in sample area* were counted as s[x>relin(5s.
Si/e class fretjuencies were calculated sefwrately for holdfastsand i^porclings as the fmctitin of the total number of blades ofeach tyix- in each si/e class. These data allowed comparison ofholdfasi and sjjoreling siie class structure independent of theeffpct of greater niimlx-r« of holdfast plants.
HypmhrKn 3. If gametophyies are belter able lo survive andgrow in this (Kipulation. then gametophytic thalli should be largerand have lower mortality rates. The following pnKcdures wereUied to measure these demographic characteristics. In ihe sum-mer 1081. I tagged the largest blade of each plant within 25 cm•)f the transects drscrihed abtwe. Location, tidal height and lifehi*!r)ry phaM* were noted for each lagged blade. The tags weremade of niiml>ered, plastic dynam<vlabrl» threadeti onto smallnylon electrical tics (Secura-tie fasteners, Andfel Corp., Chicago.'L). The ties were fastened around the blade's stipe and no abra-sion of stipes was apparent. In June and early July. 209 bladesWere tagged.
1 agged blades were monitored for growth and mortality.Junethrough late August 1981. at each low tide. Monality of a bladewas recorded when it could no longer be located at its originalposition. Note that blade mortality and not mortality of the entireplant is being measured. Size class frequencies of gametophytrsand sporophytes were calculated in the same way as for holdfastsand sc>oreiings.
IhfNtthesis i. With alternation of generations, if sporophyteshave higher reproductive rates than do gametophytes. tetra-spores released could produce the gametophytc-dominated pop-ulation observed. As an estimate of reproductive rates in spo-mphytes and female gametophytes, 1 examined all tagged bladesfor the presence or absence of reproductive sori through summer1981. Release of carpospores or tetraspores was detected in thefield by the ability of mature blades to release spores and colorwhite paper while blotting blade surfaces. The percentage ofreprfiduciively mature female gametophyles and sfxirophytescould then be calculated, Males and females were differentiatedboth by microscopic examination of portions of blades returnedto ihe lab and by difTereticen in thallus structure observed in ihefield.
Sltttistiml proffdutfs. Tests for differences in percent composi-tion of life history phases were carried out using the G-test forg(M>dness of fit (Stikal and Rohlf 1981). Analysis of variance wascarried out for relative growth rates using SAS GLM progmms

452 CEORGIANA MAY
2 . Hfintr.-f gmu'lh rnu-. (cm^Hn^ 'rm ' ± SD) of filnnl\atiu£ from huliifayti and ipofflnig\. Sfiririg I9H2.
January-April 2.1 ± 5.4 (n = 54) —April-June 0.07 i 0.3S (n - 35) 0.06 ± 0.25 (n - 24)
• Holdfast plants were identified in January.'• BecauK- s[)orcling plants cnuld n<ii IK- identified uniil April,
nu dala are available fnr sporeling growth rates frotn January tr>April.
(SAS 1982): raies were not iransformed pri()r to analysis. Mur-lality curves were cnmpuied and analysed using BMUP PI L pn»-grams(BMDP 1983).
RESULTS
Hypothem I. The proportions of j^amclophytcs andtetrasporophytcindividuaU varied little from ca. 85%gamc'lophyles and ca. \77c ictrasporophytcs during3 years of observation. Thcw proportions differedsignificantly (Z' > 0.05) from both 2:1 and 1:1 ratiosof gametophytes to iporophytfs (Table 1). Of thegametophytes, representation of males and femaleswas approximately ecjual (at P > 0.05, 46.27c fe-males. 53.87c males, n = 173) in summer 1981.
Hypothesis 2. Perennating holdfasts represented8O9E and sporelings» 2091. of the individuals in thespring 1982 population (Fig. 2). Cametophytes andtetrasporophytes were found in both the perennat-ing and sporeling populations: the population as awhole had the same proportions of gametophytesand sptjrophytes in 1982 ai in the previous two years(Table 1). Plants from mapped holdfasts grew rap-idly in the period from winter to early spring whileboth holdfast and sporeling plants grew at the samerate in late spring (Table 2). Blades that regeneratedfrom holdfasts occupied larger size classes than bladesfrom sporeling plants (Fig. 3), and by June, 90^ ofthe blade area for the population was attributed toregeneration from holdfasts.
Hspothesh 3. Using the tagging techniques to assessgrowth rates, I found that tetrasporophyte bladesexpanded faster (P > 0.05) than gametophytes ofeitnrr sex during ihe first growth measurement pe-riod. In the following periods, areas of all taggedblades decreased at similar rates (Table 3).
An analysis of size class frequencies was made toexamine growth in blades other than those tagged.Smaller blades, especially those less than 50 cm^represented an increasing proportion of the popu-lation numbers through most of the summer. Ca-metophytes were better represented in the largersi/e classes than were tetrasporophytes during sum-mer 1981. Fxcept in mid-summer, no sporophyticblades were larger than 1500 cm', and at no timewere any greater than 2500 cm' (Table 4). Thoughthe largest size classes contain only a small fractionof all gametophytic thalli, these blades contributedup to 2Q% of the total population's blade area. Sum-mer 1980 size class data were in general agreementwith 1981 results (data not shown),
TABI h 3. Hflnttvr grmi-lh rnf/i (fiit'nnyph^U\ nntt fetrn.'iftorof/h'stfi in tummrr /9iV/
C..iMlrli,|i
' • - " " " ' • • ' " • • I ' -
6/22-6/29 -0.01 + 0.017/02-7/15 -0.02 ± 0.017/20-7/28 -0.02 i 0.017/30-8/12 -0.03 + 0.01
Int.-.
M..lr
tO.Ol ±-0.05 ±-0.03 ±-0.04 ±
'•fm '
0.030.470.010.01
± SE) of
-0.02 ±-0.02 ±-0.03 ±
0.23'0.010.010.01
* Kelaiivr growth nties ol" [etra ^> »r«lphvU• (lifTiTt-d (/' > 0.05)Irom growth rates lor male and Fc-male gainetophyics onlyjune22-29.
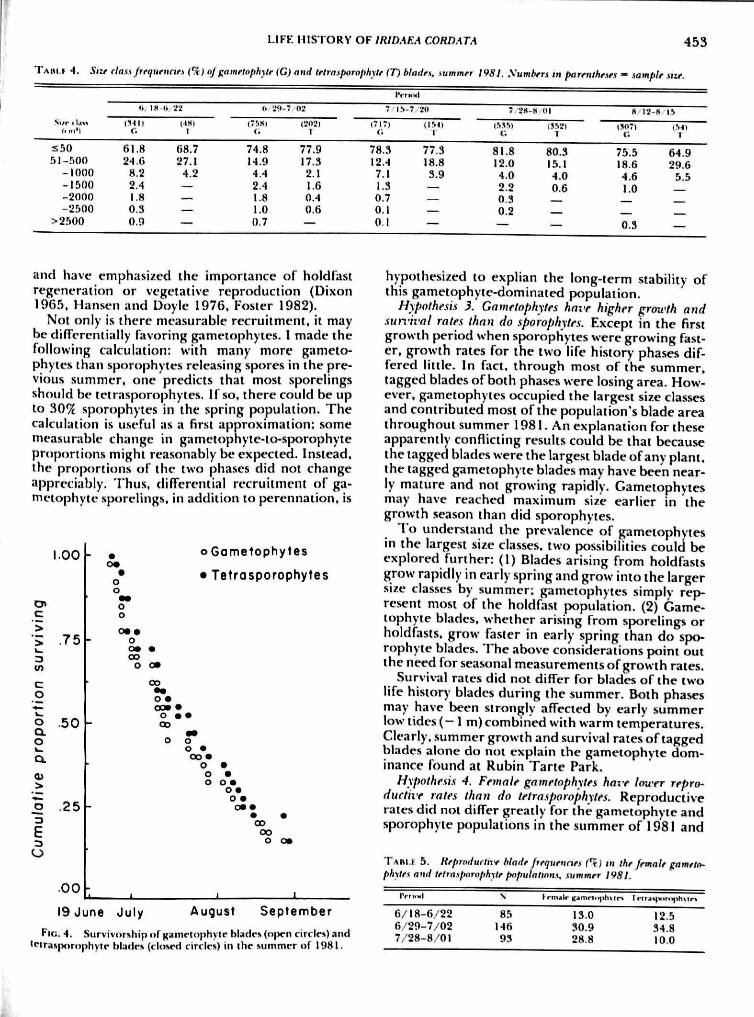
Survivorship in tagged blades did not differ forthe two life history phases over the summer 1981(Fig. A). Moreover, the shape of the curve suggestsa constant probability of blade mortality throughthe summer (Solbrig and Solbrig 1979).
Hypathesis 4. Reproductive sori were present andspores were released Irom 10 to 35^ of both taggedfemale gametophytes and sporophytes, July throughlate August 1981 (Table 5). Although the propor-tion of blades releasing spores is approximately thesame for the two phases, the total number of repro-ductively mature gametophytes is much greater thanthe number of mature sporophytes.
DISCUSSION
Hypothesis 1. Alternnlhn of gen frntUuu occurs, hut ona linger time scale than pmwusly npprennteil. Re-markably little variation was observed in the rep-resentation of life history phases over 3 years andthe phase dominating the Rubin Tarte populationdid not reverse from year-to-year. Catastrophicevents which might dislodge the dominant and pe-rennating phase were not observed. Blades of bothphases are shed and regenerated each year withoutchange in population structure, largely because ofperennation from existing (and gametophytic) hold-fast.s.
Hypothesis 2. Iridaea cordata plants art annual andpopulations are rpf^etierated each year hy spores. Regen-eration from perennating holdfasts is ihe primarymechanism of renewing the spring blade population,most of which are gametophytes. llansen (1977)came to a similar conclusion for tetrasporophyte-dominated populations in California. Holdfasts couldlimit spore settlement (Hansen 1977, Foster 1982)by occupying "safe sites" (Harper 1977) or, havingbeen initiated the previous summer, holdfast bladesmay get an earlier start in spring growth. Becauseof perennation from holdfasts, alternation of gen-erations must be occurring at a much lower levelthan expected for a population of annual plants es-tablished each year by spores.
Sporeling recruitment cannot, however, be ig'nored. The level of sporeling recruitment reportedhere is comparable to that of Cunnill (1980) forPrhetia fmti^ata where recruitment replaces plantslost to mortality. Previous comparisons t»f red algalrecruitment and perennation have been qualitative
LIFE HISTORY OF IRIDAEA COHDATA 453
4. Sizf cla.^\ frfi)Ufncif.y l*^i nJ pninrtophylr {Gj tiitd Iftrasporophylf iT) bladfs. iutntnfr t9St. Sumbfrs in parfnthfifs " samplf
l-ni'l
5 5 0
51 -:.oo-1000-1500-2000-2500
>2r>oo
II 1(4 1
c
f)l.824.68.'i2.41.80.30.9
1
6K27,
4,
n
.1
,12
74.814.94.42.41.81.00.7
• . " .» -7 n : ;
77.917.32.11.60.40.6—
7 l > -
I T I T )
78.312.47.11.30,70,10,1
7 -JO
V
•J7.S18.83.9—
—
7 2«-(t
r.
81.812.04.02.20.30.2
ul
l "
80.315.14.00.6
—
« 12-B
.V.7,
75.518.64.61.0
0.3
I
64.929.6
5.5
—
and have emphasized the importance of holdfastregeneration or vegetative reproduction (Dixon1965. Hansen and Doyle 1976. Foster 1982).
Not only is there measurable recruitment, it maybe differentially favoring gametophytes. 1 made thefollowing calculation: with many more gameto-phytes than sporophytes releasing spores in the pre-vious summer, one predicts that most sporelingsshould be tetrasporophytes. If so, there could be upto 30^ sporophytes in the spring population. Thecalculation is useful as a first approximation: somemeasurable change in gametopliyte-to-sporophyteproportions might reasonably be expected. Instead,the proportions of the two phases did not changeappreciably. Thus, differential recruitment of ga-metophyte sporelings. in addition to perennation, is
1.00 -
•> .751-
W,
co
o .50oa
.25 -3
O
.00 -
- •
ooooo a *
o
oo0
-
I 1
oaooo aosa *
O 1CDo
o
*
oo •« j m
0cc
Gametophyies
Tetrosporophytes
*) *> o a
o ao ao a *
0000o oa
1 1
19 June July
FIG. 4. Survivnrshijxif
August September
ptophyic l>laclc% (npcn t irclcs) and(cl(w.cd circles) in the summer of 1981.
hypothesized to explian the long-term stability ofthis gametophyte-dominated population.
Hypothesis 3. Gametophytes have higher growth andsurvival rates than do sporophytes. Except in the firstgrowth period when sporophytes were growing fast-er, growth rates for the two life history phases dif-fered little. In fact, through most of the summer.tagged blades of both phases were losing area. How-ever, gametophytes occupied the largest size classesand contributed most ofthe population's blade areathroughout summer 1981. An explanation for theseapparently conflicting results could be that becausethe tagged blades were the largest blade of any plant,the tagged gametophyte blades may have been near-ly mature and not growing rapidly. Cametophytesmay have reached maximum size earlier in thegrowth season than did sporophytes.
1 o understand the prevalence of gametophytesin the largest size classes, two possibilities could beexplored further: (1) Blades arising from holdfastsgrow rapidly in early spring and grow into the largersize classes by summer; gametophytes simply rep-resent most of the holdfast population. (2) Game-tophyte blades, whether arising from sporelings orholdfasts, grow faster in early spring than do spo-rophyte blades. Ihe above considerations point outthe need for seasonal measurements of growth rates.
Survival rates did not differ for blades of the twolife history blades during the summer. Both phasesmay have been strongly affected by early summerlow tides (- 1 m)combined with warm temperatures.Clearly, summer growth and survival rates of taggedblades alone do noi explain the gametophyie dom-inance found at Rubin Tarte Park.
Hypothesis 4. Female gametophytes have lower repro-ductive rates than do tetrasporophytes. Reproductiverates did not differ greatly for the gametophyte andsporophyte populations in the summer of 1981 and
T,\Ri,f 5. tifprvduftnf hladfftfqufncifs f'tj in thf fftnalf gamfto-ph\tf\ and tftrn\pttTapHytf popuiatioHi. .lummfr 1981.
trnulr uj
fi/18-6/22fi 29-7/027 28-8/01
8514693
13,030.928.8
12.534.8l O . Q

454 ORORCIANA MAY
rates measured are comparable to those of otherreports (Dawes et al. I974» Mathieson and Burns1975, Norall et al. 1981). Keeping in mind the al-ternation of generations demonstrated in culture(Kim 1976), an explanation of gametophyte domi-nance would have to profwse that, in the field, fe-male gametophyles' reproduction was inhibited (Ed-wards, I973)orthat they were apogamic. Inhibitionof carpospore formation relative to tetraspore for-mation is not supported here as both phases repro-duced at comparable rates.
Recycling females by unfertilized carpospores, pu-tative evidence for apogamy, has been demonstratedfor some heteromorphic members of the Cigarti-nales (DeCew and West 1981, Polanshek and West1977) and might lead to a population that was dom-inated by female gametophytes. However, equalnumbers of males and females were found. Thoughthese data provide some insight into the relativereproductive rates of gametophytes and tetraspo-rophytes, they point out a need for accompanyingdata of spjre viability, settlement patterns and es-tablishment rates.
In summary, I monitored an Iridaea cordata pop-ulation for possible changes in population structureand investigated mechanisms by which the observedgametophyte dominance might be maintained. Twomodels, mechanistic and stochastic, can be proposedand tested to explain observations of phase-domi-nated populations. The models are presented tostimulate further thought and research. For theRubin Tarte population, a mechanistic model bestfits the data; a stochastic mrxiel may be more appli-cable to other /. cordata populations.
The mechanistic model propfwes the following:(I) Differential spore recruitment or sporeling sur-vival could explain recruitment without change inpopulation structure. Though others have exam-tned spore settlement (Barilotti and Silverthorne1972. Prince and Kingsbury 1973), the possibilityof differential spore recruitment is unexplored. Astudy "f gra/er preference might profitably focuson this developmental stage, (2j Turn-over rates ofgametophvte holdfasts may be lowt-r than for spo-rophytes, because they are longer-lived or more re-sistant to grazers. Hansen and Doyle (1976) made asimilar suggestion for sporophyte-dominated pop-ulations in California. (3) As discussed under hy-pothesis 3, gamet<jphyies may grow faster than spo-rophytes in the early spring. With the exception ofphotosynthetic rates of Ptilota serrata measured byMathieson and Norall (1975), physiological attri-butes have rarely been compared for isomorphic redalgal phases.
A stochastic model predicts that chance eventsaffect ratios of life history phases in a study popu-lation. For example, a disaster (Har(x*r 1977) thairemoved holdfasts would allow a change in the pop-ulation siruciure with the nearest spore source de-termining which phase was re-established. Such an
event may have affected Hansen'.s (197G) Ano Nue-vo population recently; Dyck('tal.(1985) report thatthe /. cordata population has changed from sporo-phyte- to gamctophyte-dominated. F.ffects of disas-ters or catastrophes have been noted for brown algalpopulations (Kirkman 1981, Dayton and Tegner1984) but not for red algae. A stochastic model isnot supported for the Rubin Iarte population be-cause with sporeling recruitment and turn-over ofperennating plants, fluctuations in population struc-ture should be observed from year to year.
The stochastic and mechanistic models are pop-ulation-level modelsand their applicability could varyfrom place to place. The stochastic model predictsthat both phases arc equally able to survive and re-produce. The mechanistic mode! predicts that Hfehistory phase.s differ in demographic or physiolog-ical responses to their environment. Within a species,population structure may depend not only on thelife history phases' ability to grow and reproducebut on life history variations governed by largerscale environmental change.s such as latitudinal gra-dients (Guines and Lubchenco 1982).
Sexual reproduction of some species may be in-hibited at the higher latitudes, producing popula-tions of tetrasporophytes and slcrilo plants (Dixon1965, Knaggs 1969, F.dwards 1973. Whittick 1978).Iridaea rorriata phase ratios vary along the Pacificcoast but a latitudinal gradient of phase ratios i.s notstrongly supported (Dyck et al. 1985)- However,pr)pulations have not been studied at the extremesof the /. cordata range, from Alaska south to Mexico(Abbott 1971, 1972).
In conclusion, the following general hypothesisfor the life history of Iridara cordata is propo.scd forfurther testing: Alternation of generations—ol ne-cessity by spores—depends on two factors; one. theturn-<»vcr of older, perennating plants and two, theability of alternate phases to recruit and reproduce.With high levels fjf perennation, alternation of gen-erations will occur at a lower level.
Alternation of generations may still be importantwhere phases are spatially or temporally separated.Phases may be spatially separated because of de-mographic or physiological differences. Phases maybe temporally separated because of stochasticchanges. Populations dominaled by one phase canbe maintained by a combination of perennation anddifferential spore survival.
Deviations from a life hi.story of alternating gen-eralions, such as apogamy, may IK" important in somepopulations of red algae but oihrr mechanisms canexplain over-representation ofone life history phase.Population structure at any given time is not, there-fore, predictive of population dynamics and life his-tory variations.
I thank Friday ll»rb<ir Marine Lain for %up|Kirl andihank the l>epnrtmeni nf RoMnv. L'nivrrsily of CHlilley. California for travel und t ompu i r r fuiHU. Ihi* work

LIFE HISTORY OF IRtDAEA CORDATA 455
made possible and enjoyable by ihr generous support and cn-roiiraKemrnt from ihr following individuals: Anne Sylvester.Donna Kn/lowski. Bob Wual»nd, Kaihy Anil Milter, Bob HaiRht.and ihemcmbfrsol the Fhycologi,' f^roupai Berkeley. Commentsirom IWO anonymous reviewers were helpful.
Abbori.l.A. 1971. Onrhc sjx-cirsof/ni/dm (Rluxiophyta)rromthe Pacific toasi of Nnrih America. Syf.us 4:51-72.'
1972. Field studies which evaluate criteria used in sep-arating s|K*cirN of tndiifa (Rhodophyta). t» Abbott. I. A. ScKuroRi, M. [Eds.] CO»I/M7>I(/IO»H ti> Ihf Syitfmalia of Brnthic Ma-riitf Algftf of thf Sorth Pacific. Japanese Society of Phycology,Kobr, pp. 25S-6-).
Barilotti. C. D. Sc Silveribonic. W. 1972. A resource manage-ment srudy of Gflidiurn rnbii.'iiutn. Pror. ttit. Sfawffd Symp. 7:255-r.l.
BMDP Stattuical Softu'arf. Dixon. W. S. (F.d.) University of Cali-fornia Press. Berkeley, Los Angeles. London. 1983. 744 pp.
Craigie. J. S. & Lei^h. C. 1978. Carragecnan* and agar^. tnHellrbust,J. A. Sc Craigie.J. S. \¥As.\ttfniithookofPhyrologicalMfthod.s, PhyxiologiralandBhifhfmicaf Atfthods. Cambridge Univ,Prew. Cambridge. London, New Vork. Melbourne, pp. 109-31.
Craigie. J. S. &: Pringle. J. D. 1978. Spatial dtsiribution of tet-rasfxtrophytcs and gametophytes in four maritime popula-tions of Chondrui cmpui. Can.J. Bot. 56:2910-14.
Dauci. C. J.. Mathieson. A. C. & Cheney. D. P. 1974. Ecologicalstudies of Floridian Euchfuma (Rhodophyta, GigarlinaU-s) LSeaM)nal growth and reproduction. Bull Mar. Sci. 24:235-73.
Dayton, P. K. & legner, M. J. 1984. Oitasiropbic storms. ElNino, and patch stability in a Southern California kelp com-munity. Scifncf (^Vash.. DC.) 224:283-4.
ileCew, r. C. & Wesi, J. A. 1981. Life histories in the Phyllo-phoraceae (Rh<Klophyia. Gigartinalrs) from the Pacific coaMol North America I. C\mnof;ongruslinfari.s9ttdC, tfptoph\llus.J. Phyrol. 17:240-50. '
Dixon. P. S. 1965. Pcreniiation, vegetative propagation and algallife histories, with special reference to.ii/>rtrflg(j/»i(.v and Ol herRhodophyia. J!»(. Gctihoburg. 3:67-74.
liyck. L.. DeWrcede, R. E. & Garbary. D. 1985. Life historyphases in Indafa cnriinta (Gigartinacear): relative abundanceand distribution from British Columbia to California.ya/»./.Phycol. (Sorm) 33:225-32.
Edward*. P. 1973. Life history studieiofsck-ctcd British Cccaw-tum sj>ccie».y, Phycot. 9;181-4.
Power, M. S. 1982. Factor* controlling ihc intertidal/onationof/rirfrtr(i/?(ifrr//rt (Rhodophyla).y. Phyral. 18:285-94.
Caincs. S. D. & LulKhcnco, J. 1982. A unified approach tomarine plant-herbivore interactions II. BiogeographyMnim.Wn- E,i>l. 5vw, 13:111-38.
Gunnill. F. C. 1980. Demography of the intertidal brown algat*fhvtta fa\tifpatn in Soulheni C«ilifornia. USA. .\Mr, Bml. {Bfrl.)59:169-79.
Hansen. J .E. 1976. Population biology of/nrfoM forrfato Rho-dophyia: Gigartinaccae. Ph.D. ihcsis, Univ. of California.Santa Cruz, xiii + 341 pp.
1977. Ecology and natural history of/ni/n/'(iforrfii/fl(Gig-ariinales, Rhodophyta) growih./ Ph\T(>l. 13:395-402.
Hansen.J.E.& Doyle. W.T. 1976. Ecology and natural historyof tndafa cordala (Rhodophyta: Giganinaceae): populationstructure.//»/ivrrt/. 12;273-8.
Harper.J. L. 1977. Population Biotof^ oJPtattts. Academic Press.London. New York. San Francisco. 892 pp.
Hunt, R. 1982. Plant Grm-thCunva. a Fumtional .\ppToach to PlantGrauih Analysu. University Park Press. Baltimore. 248 pp.
Kim. D. H. 1976. A study of the development of cwocarps andtctrasporangial sori in Giganinaccae (Rhodophyta, Ginrti-nales). Soi<a Hfdwigia 27:1 -146.
Kirkman. H. 1981. Thr first year in the life history and tbesurvival of the juvenile marine macrophyte. Eckloma radiata( rum.)J . Agardh./ Exp. Mar. Bid Ecol. 55:243-54.
Knaggs. K. W. 1969. A review of floridcophycran life historiesand the culture techniques cmploved in their investigation.Sm^a Hfd-ii-ifTia 18:293-330. '
McCandless. E. L.. Craigie. J. S. & Hansen, J. E. 1975. Carra-geenans of gametangial and tetnisporangial stages of I rida facordata (Gigartinaceae). CanJ. Bol. 53:2315-18.
Mathiestm. A. C. Sc Bums. R. L. 1975. Ecological studies ofeconomic red algae V. Growth and reproduction of naturaland harvested populations of Chondrus critpuji Stackhousc inNew Hampshire./ Exp. Mar. Biol. Ecol. 17:137-56.
Mathieson. A. C. & Norall. T. L. 1975. Physiological studies ofsubtidal red algae./ Exp. Mar. Biol. Ecol. 20:237-47.
NoraII,T. L.. Mathieson, A. C.&Kilar.J. A, 1981. Rcproduaiwecology of four subtidal red algae.7 f-xp- ^tar. BioL Ecol. 54-119-36.
Polanshek. A. R. & West, J. A. 1977. Culture and hybridizationstudies on Gigartim papillata (Rhodophyta). / PAvro/. IS:141-49.
Prince.J. S. & Kingsbur>. J. M. 1973. The ecolog) of Chortdruscrt.fpus at Plymouth. Massachusetts II. Field studies. Am IBol. 60:964-75. "^
Rao.K. R. 1970. Studies on gniwth cycle and phyrocoltoid con-tent in ttypufa muinformu (Wulf) Lamour. Bot. Mar. (Bfrl J13:163-5.
SAS Institute Inc. 1982. 5 . « f V r ( Guidr StatiMm, t9S2 Edition,Cary. NC: SAS Institute Inc.. 1982. 584 pp.
Sokal. R. R.&Rohlf. F.J. 1981. Biomftry. (2nd ed.) W. H. F r« -man and Co., San Francisco, pp. 691-731.
Solbrig. O. T. Sc Solbrig, D. J. 1979. Introduction to PopulationBiolofntandEi'olution. Addison-U'cslcy Publishing Co.. Read-ing. Massachusetts. 467 pp.
Whittick. A. 1978. The life history of Ca//ifAflifimfln«)rTmAo«/»(Rhtxlophyta: Ceramiacrae) in Newfoundland. Can. /. BOL56:2497-9. "






























