Embed Size (px)
Citation preview

www.elsevier.com/locate/chemgeo
Chemical Geology 205 (2004) 367–390
Life at cold seeps: a synthesis of biogeochemical and ecological
data from Kazan mud volcano, eastern Mediterranean Sea
Josef P. Wernea,*, Ralf R. Haeseb,1, Tiphaine Zitterc, Giovanni Aloisid,2,Ioanna Bouloubassie, Sander Heijsf, Aline Fiala-Medionig, Richard D. Pancosta,3,
Jaap S. Sinninghe Damstea, Gert de Langed, Larry J. Forneyf,4,Jan C. Gottschalf, Jean-Paul Foucherh, Jean Masclei, John Woodsideb
the MEDINAUT and MEDINETH Shipboard Scientific Parties5
aDepartment of Marine Biogeochemistry and Toxicology, Royal Netherlands Institute for Sea Research (NIOZ), Den Burg, The NetherlandsbDepartment of Geochemistry, Faculty of Earth Sciences, Utrecht University, Utrecht, The Netherlands
cDepartment of Sedimentology and Marine Geology, Faculty of Earth and Life Sciences, Free University Amsterdam,
Amsterdam, The NetherlandsdLaboratoire d’Oceanographie Dynamique et de Climatologie, Universite Pierre et Marie Curie, Paris, France
eLaboratoire de Chimie et Biogeochimie Marines, Universite Pierre et Marie Curie, Paris, FrancefDepartment of Microbiology, Center for Ecological and Evolutionary Studies, University of Groningen, Haren, The Netherlands
gObservatoire Oceanologique, Universite Pierre et Marie Curie, Banyuls-sur-Mer, FrancehDRO/Environments Sedimentaires, IFREMER, Brest, France
iGeosciences Azur, Observatoire Oceanologique de Villefranche, Villefranche-sur-Mer, France
Received 4 October 2002; received in revised form 30 September 2003; accepted 23 December 2003
Abstract
Recent field observations have identified the widespread occurrence of fluid seepage through the eastern Mediterranean Sea
floor in association with mud volcanism or along deep faults. Gas hydrates and methane seeps are frequently found in cold seep
areas and were anticipated targets of the MEDINAUT/MEDINETH initiatives. The study presented herein has utilized a multi-
disciplinary approach incorporating observations and sampling of visually selected sites by the manned submersible Nautile and
by ship-based sediment coring and geophysical surveys. The study focuses on the biogeochemical and ecological processes and
conditions related to methane seepage, especially the anaerobic oxidation of methane (AOM), associated with ascending fluids
on Kazan mud volcano in the eastern Mediterranean. Sampling of adjacent box cores for studies on the microbiology,
biomarkers, pore water and solid phase geochemistry allowed us to integrate different biogeochemical data within a spatially
0009-2541/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.chemgeo.2003.12.031
* Corresponding author. Current address: Large Lakes Observatory and Department of Chemistry, University of Minnesota Duluth, 10
University Dr. 109 RLB, Duluth, MN 55812, USA. Tel.: +1-218-726-7435; fax: +1-218-726-6979.
E-mail addresses: [email protected] (J.P. Werne), [email protected] (R.R. Haese).1 Address as of October 2003: Geoscience Australia, Canberra.2 Current address: Department of Marine Environmental Geology, GEOMAR, Kiel, Germany.3 Current address: Organic Geochemistry Unit, School of Chemistry, University of Bristol, Bristol, UK.4 Current address: Department of Biological Sciences, University of Idaho, Moscow, ID, USA.5 S. Asjes, K. Bakker, M. Bakker, J.L. Charlou, J.P. Donval, R. Haanstra, P. Henry, C. Huguen, B. Jelsma, S. de Lint, M. van der Maarel, S.
Muzet, G. Nobbe, H. Pelle, C. Pierre, W. Polman, L. de Senerpont, M. Sibuet.

J.P. Werne et al. / Chemical Geology 205 (2004) 367–390368
highly heterogeneous system. Geophysical results clearly indicate the spatial heterogeneity of mud volcano environments.
Results from pore water geochemistry and modeling efforts indicate that the rate of AOM is f6 mol m�2 year�1, which is
lower than at active seep sites associated with conditions of focused flow, but greater than diffusion-dominated sites.
Furthermore, under the non-focused flow conditions at Kazan mud volcano advective flow velocities are of the order of a few
centimeters per year and gas hydrate formation is predicted to occur at a sediment depth of about 2 m and below. The methane
flux through these sediments supports a large and diverse community of micro- and macrobiota, as demonstrated by carbon
isotopic measurements on bulk organic matter, authigenic carbonates, specific biomarker compounds, and macrofaunal tissues.
Because the AOM community appears to be able to completely oxidize methane at the rate it is seeping through the sediments,
ultimate sinks for methane in this environment are authigenic carbonates and biomass. Significant differences in organic
geochemical data between this site and those of other cold seep environments, even within the eastern Mediterranean mud
volcanoes, indicate that the microbiological communities carrying out AOM varies depending on specific conditions such as
methane flux and salinity.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Anaerobic oxidation of methane; Cold seep fauna; Gas hydrate; Mud volcano; Biogeochemistry; Microbiology
1. Introduction
Methane cold seeps have become environments of
intensive study in recent years for several reasons.
Among these are: (1) the potential for methane release
from hydrates and the subsequent impact on climate
(Dickens et al., 1995), (2) slope destabilization associ-
ated with gas hydrate dissociation (Henriet and Mie-
nert, 1998), (3) energy sources for chemosynthetically
based life (Sassen et al., 1993; Sibuet and Olu, 1998),
and (4) as a potential energy resource (Hovland, 2000).
Furthermore, the anaerobic oxidation of methane
(AOM) has been identified in methane seep environ-
ments world-wide, and is believed to scavenge signif-
icant amounts of methane (Reeburgh et al., 1993)
which could otherwise be released to the atmosphere,
potentially contributing to global warming (e.g., Dick-
ens et al., 1995).
Mud volcanoes, which result from the extrusion of
fluid-rich mudflows (mud breccias), are often associ-
ated with methane seepage, and, given the appropriate
pressure and temperature conditions, with gas hydrates
(Brown, 1990; Milkov, 2000). Mud volcanoes have
been identified in the easternMediterranean (Cita et al.,
1981, 1989; Cronin et al., 1997), and active methane
seepage has been documented on these mud volcanoes
(Emeis et al., 1996; Limonov et al., 1996; Woodside et
al., 1998). In 1998 and 1999, the joint French–Dutch
MEDINAUT and the Dutch MEDINETH expeditions
were made to investigate eastern Mediterranean mud
volcanoes and their associated cold seeps. These mul-
tidisciplinary investigations combined visual observa-
tions from dives aboard the manned submersible R/V
Nautile (launched from the support vessel N/O Nadir)
and geophysical/acoustic surveys with shore-based
sedimentological, (bio)geochemical and (micro)bio-
logical investigations on seafloor sediment samples
taken from the Nautile and via box core from the R/V
Professor Logachev (MEDINAUT/MEDINETH Ship-
board Scientific Parties, 2000).
One of the major objectives of the MEDINAUT/
MEDINETH expeditions was an in depth biogeo-
chemical and (micro)biological investigation of
AOM. It is now generally accepted that a microbial
consortium consisting at least of methane-oxidizing
archaea and sulfate-reducing bacteria consumes sulfate
and methane in a 1:1 molar ratio during AOM,
producing sulfide and DIC according to the general-
ized net reaction
SO2�4 þ CH4 ! HS� þ HCO�
3 þ H2O ð1Þ
as proposed by Reeburgh (1976). While the consor-
tium hypothesis for net AOM has been proven through
a number of studies (Boetius et al., 2000; Hinrichs et
al., 1999; Orphan et al., 2001), the specific pathways
of methane-derived carbon incorporation into micro-
bial biomass remain debated (Valentine and Reeburgh,
2000; Sørensen et al., 2001). Furthermore, the specific
organisms remain unknown and uncultured, though
significant advances have been made in narrowing
down the organisms based in large part on phyloge-
J.P. Werne et al. / Chemical Geology 205 (2004) 367–390 369
netic studies (Hinrichs et al., 1999; Boetius et al.,
2000; Orphan et al., 2001; Aloisi et al., 2002). This
paper is a synthesis of the geophysical, (bio)geochem-
ical and (micro)biological studies carried out by mem-
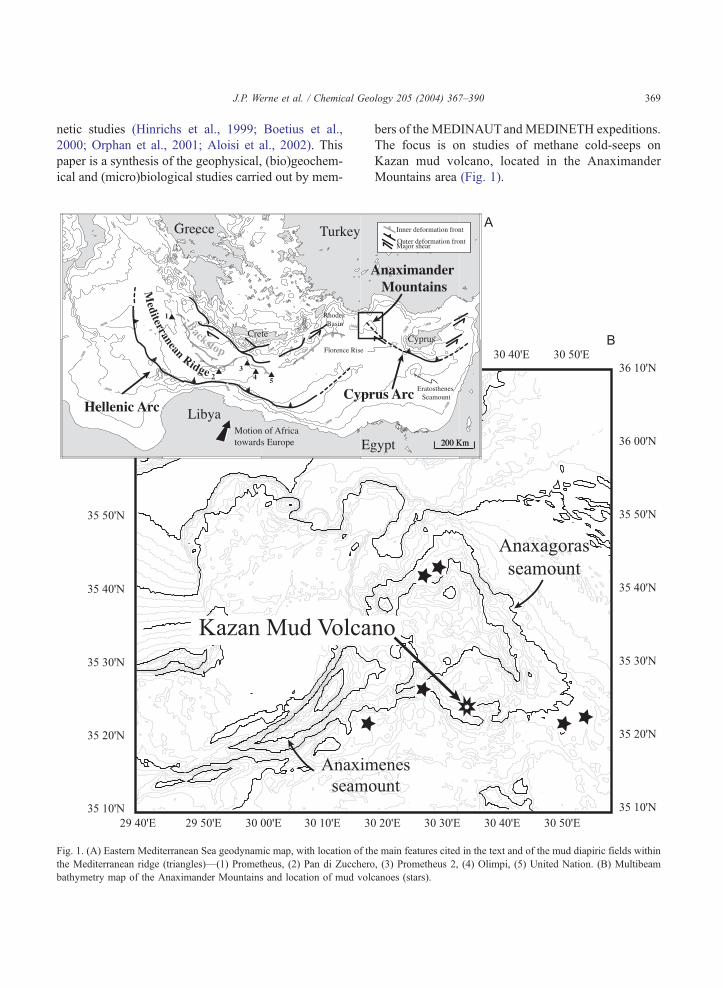
Fig. 1. (A) Eastern Mediterranean Sea geodynamic map, with location of th
the Mediterranean ridge (triangles)—(1) Prometheus, (2) Pan di Zucchero
bathymetry map of the Anaximander Mountains and location of mud volc
bers of the MEDINAUTandMEDINETH expeditions.
The focus is on studies of methane cold-seeps on
Kazan mud volcano, located in the Anaximander
Mountains area (Fig. 1).
e main features cited in the text and of the mud diapiric fields within
, (3) Prometheus 2, (4) Olimpi, (5) United Nation. (B) Multibeam
anoes (stars).

J.P. Werne et al. / Chemical Geology 205 (2004) 367–390370
2. Geologic setting and regional tectonics:
formation of the cold seep environment
Eastern Mediterranean tectonics record the initial
stages of collision between Africa and Europe, from
incipient stages south of Crete to more advanced stages
south of Cyprus (and eastwards into the Zagros area
where continental collision between the Arabian prom-
ontory and Europe is now occurring). The complex
deformation of the Eastern Mediterranean results from
the interactions of several plates and micro-plates
(McKenzie, 1972; Reilinger et al., 1997; McClusky
et al., 2000) and develops into two cup-shaped arc
systems, the Hellenic and Cyprus Arcs (Fig. 1). These
two arc systems are characterized by different tectonic
regimes, with the Hellenic arc characterized by more
active subduction (Papazachos and Comninakis, 1978;
Spakman et al., 1988; Chaumillon and Mascle, 1997)
and the Cyprus Arc characterized by a quieter tectonic
and seismic activity (Rotstein and Kafka, 1982; Ben-
Avraham et al., 1988) where subduction may be halted
(Robertson, 1998). The submarine Anaximander
Mountains region (along with the Florence Rise and
the Rhodes Basin) is located at the connection between
these two arc systems, and as a result has been the
subject of several recent studies (Woodside et al., 2000,
2002; Zitter et al., 2003; Ten Veen et al., in press).
A series of mud volcano fields exists within the
Mediterranean Ridge accretionary prism (Cita et al.,
1981, 1989, 1996; Cronin et al., 1997; Mascle et al.,
1999). Two mud volcanoes from the Olimpi field
(Napoli mud volcano and Milano mud volcano) were
investigated in detail during the Ocean Drilling Pro-
gram Leg 160 (Robertson et al., 1998). The Anax-
imander Mountains mud volcano field occurs in a
different context of the eastern Mediterranean colli-
sion zone. This mud volcano field was discovered
from multibeam mapping during the ANAXIPROBE
cruise in 1995 based on high backscattering patches
associated with circular relief (Woodside et al., 1997,
1998), and has received much less vigorous study.
The Anaximander Mountains are a complex of three
seamounts (Fig. 1), rising more than 1000 m above the
seafloor, that rifted southwards from Turkey in late
Miocene–early Pliocene times (Woodside et al., 1997).
The post-Miocene formation of this area is demonstrat-
ed by the absence of evaporites (that were deposited
during the Messinian salinity crisis in most parts of the
Mediterranean Sea Ryan et al., 1973) beneath the
Anaximander Mountains. The gravity signature
(Woodside, 1977; Woodside et al., 1997), lithological
facies (Woodside et al., 1997), and surface deformation
(Zitter et al., 2003) vary between the eastern and
western Anaximander Mountains regions, clearly indi-
cating differences in the geology of the two regions.
The western mountains belong to the Eastern Hellenic
fore-arc deformation zone (Woodside et al., 2000; Ten
Veen et al., in press), whereas the eastern mountains
(the Anaxagoras seamount) form the western limb of
the Cyprus Arc and are affected by a large NW–SE
transpressional wrenching system (Woodside et al.,
2002; Zitter et al., 2003; Ten Veen et al., in press). This
area is intensely crosscut by major N150j and N070jfaulting, inferred to be active shearing based on both
the seismic signature (pop-up flower structures) and the
bathymetric expression.
Most of the mud volcanoes discovered in the Anax-
imander Mountains occur within the Anaxagoras sea-
mount (Fig. 1), associated with both the strike-slip
faults and secondary, probably extensional, faults (Zit-
ter, unpublished results). They are generally identified
as conical topographic features on the order of 50 to
100 m high and 1 km wide (Woodside et al., 1998).
Nonetheless, the mud volcanoes and associated mud-
flows show an important variability in morphology,
size and acoustic characteristics. The shapes vary from
conical for Tuzlukush mud volcano, a quite perfectly
circular dome for Kula mud volcano, an irregular dome
for Kazan mud volcano, to the several km-wide ‘‘mud
pie’’ morphology of the exceptionally large Amster-
dam mud volcano.
3. Kazan mud volcano
Kazan mud volcano is a small dome, about 50 m
high and 1000 m wide, with an elliptical summit at
about 1700 m water depth (Fig. 2). It is located in the
southern part of the Anaxagoras seamount (E30j33.5V,N35j26V) at the interplay between the N150j and
N070j faults. From the sidescan sonar record, the
volcano shows high to very high backscatter (Fig. 2).
The high backscatter signature is believed to be caused
by both the seafloor roughness and the volumetric
heterogeneity of the mudflows produced by clasts
within the mud breccia (Volgin and Woodside,
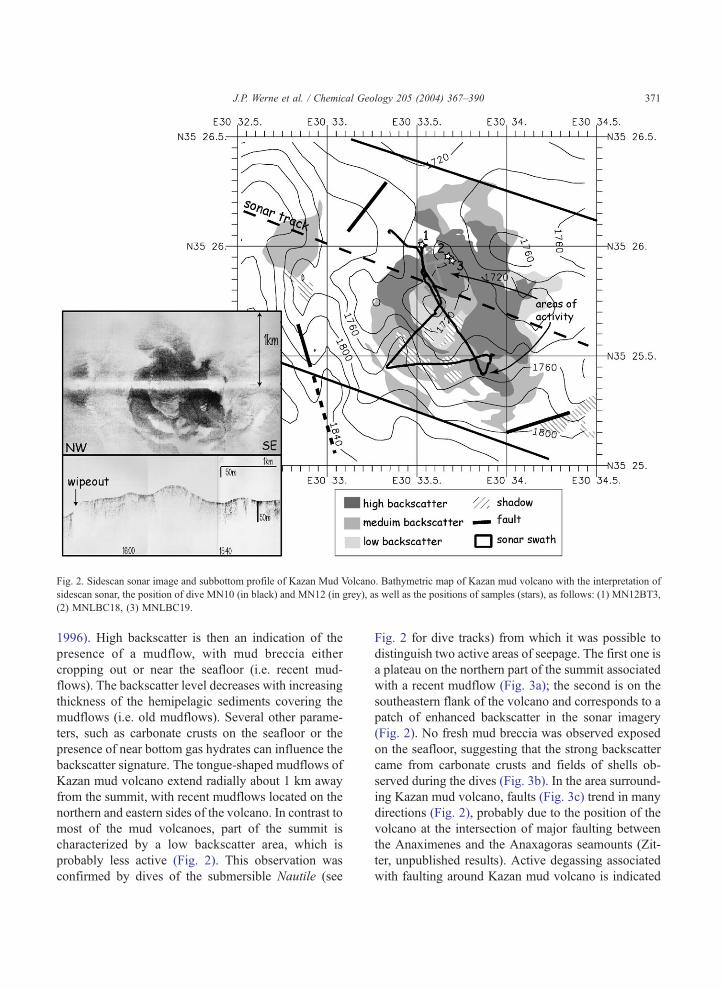
Fig. 2. Sidescan sonar image and subbottom profile of Kazan Mud Volcano. Bathymetric map of Kazan mud volcano with the interpretation of
sidescan sonar, the position of dive MN10 (in black) and MN12 (in grey), as well as the positions of samples (stars), as follows: (1) MN12BT3,
(2) MNLBC18, (3) MNLBC19.
J.P. Werne et al. / Chemical Geology 205 (2004) 367–390 371
1996). High backscatter is then an indication of the
presence of a mudflow, with mud breccia either
cropping out or near the seafloor (i.e. recent mud-
flows). The backscatter level decreases with increasing
thickness of the hemipelagic sediments covering the
mudflows (i.e. old mudflows). Several other parame-
ters, such as carbonate crusts on the seafloor or the
presence of near bottom gas hydrates can influence the
backscatter signature. The tongue-shaped mudflows of
Kazan mud volcano extend radially about 1 km away
from the summit, with recent mudflows located on the
northern and eastern sides of the volcano. In contrast to
most of the mud volcanoes, part of the summit is
characterized by a low backscatter area, which is
probably less active (Fig. 2). This observation was
confirmed by dives of the submersible Nautile (see
Fig. 2 for dive tracks) from which it was possible to
distinguish two active areas of seepage. The first one is
a plateau on the northern part of the summit associated
with a recent mudflow (Fig. 3a); the second is on the
southeastern flank of the volcano and corresponds to a
patch of enhanced backscatter in the sonar imagery
(Fig. 2). No fresh mud breccia was observed exposed
on the seafloor, suggesting that the strong backscatter
came from carbonate crusts and fields of shells ob-
served during the dives (Fig. 3b). In the area surround-
ing Kazan mud volcano, faults (Fig. 3c) trend in many
directions (Fig. 2), probably due to the position of the
volcano at the intersection of major faulting between
the Anaximenes and the Anaxagoras seamounts (Zit-
ter, unpublished results). Active degassing associated
with faulting around Kazan mud volcano is indicated

Fig. 3. Photographs taken during dives of the submersible Nautile to
Kazan mud volcano. (a) Mud breccia showing size variability of
clasts in extrusive flows. (b) Example of abundant authigenic
carbonate crusts and shell fields. (c) Fault scarp demonstrating
recent tectonism on Kazan mud volcano.
J.P. Werne et al. / Chemical Geology 205 (2004) 367–390372
by typical wipeouts in acoustic facies on the subbot-
tom profiler along a scarp about 1 km west of the
volcano (Fig. 2).
4. Methods
4.1. Geophysics
The sidescan sonar and subbottom profiling data
were acquired with a MAK-1 system during the 6th
Training Through Research (TTR6) expedition. The
vehicle (or fish) operating in a Long Range mode at 30-
kHz frequency, is towed at 150 m above the seafloor at
a speed of 1.5 to 2 knots and is able to insonify a swath
2000mwide (1000m on each side). TheMAK-1 fish is
equipped with a sub-bottom profiler (SBP) operating at
a 6-kHz frequency.
4.2. Sampling
Samples were taken from Kazan mud volcano by a
variety of methods. Carbonate crust samples were
taken either by dives of the submersible Nautile during
the MEDINAUT cruise of the R/V Nadir in Novem-
ber–December 1998, or by box cores taken during the
MEDINETH cruise of the R/V Professor Logachev.
Crust sample MNT12BT3 (35j26.007VN, 30j33.53VE,1706 m water depth (Fig. 2) taken from Kazan is the
focus of both organic geochemical and carbonate
geochemical/mineralogical studies, and was sampled
by submersible dives of the Nautile. Two box cores
taken from the summit of Kazan mud volcano during
the 1999 MEDINETH cruise of the R/V Professor
Logachev (MNLBC18 and MNLBC19, 35j25.936VN,30j33.704VE, water depth 1673 m and 35j25.950VN,30j33.679VE, water depth 1673m, respectively, Fig. 2),
were dedicated to geochemical and microbiological
analysis. Pore water geochemistry and molecular iso-
topic geochemistry were studied in core MNLBC19,
and microbiological analyses were performed on both
MNLBC18 and MNLBC19.
It should be noted that the mud breccia sediments
investigated in this study were taken by box core
rather than sampled directly by submersible. There-
fore, the highly precise targeting of active cold seeps
was not possible, and the likelihood of actually
sampling an active seep in this manner is low, given
an estimated radius around the coordinates of f80 m
as a result of a 5% scope (assumed based on known
currents in the Rhodes Gyre area) in the cable lower-
ing the box core, and the extremely variable terrain on
and around Kazan mud volcano (see discussion be-

J.P. Werne et al. / Chemical Geo
low). These samples, therefore, likely represent a mid-
point in the continuum between highly active seep
sites, such as those examined on Hydrate Ridge
(Boetius et al., 2000; Elvert et al., 2001) and those
in areas of ‘‘normal’’ sediment methane levels, such as
the Kattegat (Bian et al., 2001).
4.3. Clay Mineralogy
The matrix clay mineral composition of the mud
breccia from several mud volcanoes, and of a hemi-
pelagic sample as reference, was defined on the basis
of X-ray diffraction (XRD). XRD analyses of the <2
Am-size fraction were carried out at different percen-
tages of relative humidity (0–50–100%) in order to
identify expanding clay minerals in the complex
mixture. Carbonate was removed and an internal
standard was added to prepare samples for analysis.
Semi-quantitative analyses were performed using
the integrated weighted-peak area and the clay
mineral amount was estimated using the samples
measured at 50% humidity. Results from the matrix
of MNLBC19, sampled at 21 cm deep, are presented
here.
4.4. Macrobiology
Preparation for transmission electronic microscopy
(TEM), included fixing small pieces of bivalve gills
(Myrtea sp. andVesicomya sp.) in 3% glutaraldehyde in
0.4 M NaCl, buffering with 0.1 M cacodylate (pH 7.4)
and post-fixing in 1% osmium tetroxide for 1 h in the
same buffer. The specimens were dehydrated through a
graded ethanol series and embedded in Araldite. Semi-
thin sections were stained with toluidine blue. Ultra-
thin sections were contrasted with uranyl acetate and
lead citrate, and observed under a Hitachi H600 trans-
mission electron microscope.
Stable carbon and nitrogen isotope analyses of
the tissues were made on samples frozen and stored
in liquid nitrogen then freeze dried and acidified in
the laboratory. The acidified samples were com-
busted to CO2 and N2 using standard methods
(Bouton, 1991) before being passed through a mass
spectrometer (Finnigan Delta E). Carbon isotopes are
reported relative to PeeDee Belemnite (PDB) and
nitrogen isotopes relative to atmospheric molecular
nitrogen.
4.5. Geochemistry
Pore water analyses are described in detail in Haese
et al. (2003). Carbonate mineralogy and geochemistry
analyses are described in Aloisi et al. (2000). Lipid
concentration and carbon isotopic analyses are de-
scribed in Werne et al. (2002), Pancost et al. (2000,
2001a,b), and Hopmans et al. (2000).
4.6. Microbiology
Three depth intervals spanning the zone of AOM
were investigated (0–6, 6–22, >22 cm) because of the
larger sample size needed relative to geochemical
analyses. Total counts of bacteria as well as aggregates
as described by Boetius et al. (2000) were determined
by epifluorescence microscopy using DAPI (4V,6V-dia-midino-2-phenylindole hydrochloride) and acridin or-
ange dyes. Most probable numbers (MPN) were
determined according to Heijs et al. (1999) using
appropriate culture media for the functional groups of
organisms determined according to the German Col-
lection of Microorganisms and Cell Cultures (www.
dsmz.de). DNA was isolated from sediment samples
following Aloisi et al. (2002), slightly modified with
respect to extraction times and mechanical lysis. DNA
isolates were cleaned by gel-filtration (WIZARD DNA
clean up kit, Promega Benelux, Leiden, The Nether-
lands) and concentrated to a final volume of 120 Al. Thetotal amount of DNAwas measured spectrophotomet-
rically. Archaeal 16S ribosomal RNA genes were
amplified from the total chromosomal DNA pool by
using a group specific archaeal primer set as described
by DeLong (1992). For the analysis of the microbial
communities, terminal-Restriction Fragment Length
Polymorphism (t-RFLP) was applied using FAM and
HEX-labeled primers in Polymerase Chain Reaction
(PCR). After gel purification, 100 ng of PCR product
was cut for 3 h at 37 jC using 2 U of HHA1 digestive
enzyme, and analyzed on a ABI 310 automated
sequencer.
logy 205 (2004) 367–390 373
5. Results and discussion
5.1. Sediment cores
Box cores discussed herein span the upperf30 cm
of sediment. Sediments consist of grey mud breccia

J.P. Werne et al. / Chemical Geology 205 (2004) 367–390374
with millimetric to centimetric mudstone clasts. The
upper few centimeters of sediment are oxidized. In core
MNLBC19, from 22 to 34 cm there is evidence that the
sediment was gas saturated (this interval corresponds to
a temperature of 11.9 jC measured on board).
5.2. Mud matrix clay mineralogy
The clay mineral composition of the mud matrix of
Kazan mud volcano is largely dominated by smectite
(85%), with 10% kaolinite, 3% palygorskite and 2%
illite. In comparison with the composition of Pliocene
sediments in the Mediterranean Sea (Melieres et al.,
1978) and with the hemipelagic core recovered in the
region of the Florence Rise, the matrix of the mud
breccia is considerably enriched in smectite. Such
smectite-rich formation is typically found in the Med-
iterranean Sea in Messinian deposits (Chamley et al.,
1978), which suggest a possible deeper, Messinian
source for these mud breccia matrix sediments. On
the other hand, the high abundance of hydrated clays
seems to indicate the absence of deep burial and
subsequent diagenetic transformation, supporting a
shallow source for the matrix. The smectite in the
Anaximander area could also be derived from the wea-
thering of ophiolitic rocks found on Turkey, Cyprus,
and probably partly beneath the Anaximander Moun-
tains. As discussed below, the predominance of smec-
tite clays has implications for the interpretations of pore
water geochemistry and the occurrence of methane
hydrates.
5.3. Pore water geochemistry
Profiles of measured pore water geochemical con-
stituents are shown in Fig. 4. Since methane is highly
volatile, methane gas bubbles form and may migrate
laterally and upwards during sample retrieval, leading
to highly variable measured methane concentrations,
which may significantly underestimate the in situ con-
centrations (Paull et al., 2000). In order to overcome
this artifact, the in situ methane concentrations were
reconstructed in a recent study on methane dynamics
(Haese et al., 2003) by using the unambiguous sulfate
profile and the advective flow velocity of 4 cm year�1
estimated for site MNLBC19 using best-fit simulations
of measured depth profiles of conservative solutes (Na,
B) (Haese et al., 2003). Additional assumptions re-
quired for reconstruction of a reasonable methane
concentration profile include near-complete methane
oxidation by AOM, an insignificant rate of methano-
genesis below the depth of AOM, and an insignificant
rate of sulfate reduction related to organic matter (OM)
decomposition. These assumptions are justified be-
cause at this site, AOM is restricted to a narrow depth
interval (14–18 cm below sediment surface), as can be
seen from the coinciding discrete depth of the sulfide
maximum, the minimum in the d13C of DIC, and the
depth of sulfate depletion (Fig. 4). Total organic
carbon and biomarker concentrations and carbon iso-
topic composition (Fig. 4 and Table 2) show the
imprint by AOM at approximately the same depth as
the pore water indicators, and the lack of AOM-related
biomarkers in the shallowest sediments justify the
assumption of (near) complete methane oxidation
within a discrete depth interval. Sulfate reduction
related to the mineralization of OM and methano-
genesis should be negligible due to the low amount
(0.2–0.3 wt.%, Fig. 4D) and refractory nature of the
OM present (most of the OM in these mud breccia
sediments is associated with the ascending mud ma-
trix, not with pelagic sedimentation, and is therefore
already largely degraded, Werne et al., 2002). The
most convincing argument for a negligible contribu-
tion of sulfate reduction related to the mineralization of
OM comes from a sulfate/DIC* (=DIC+Ca) molar
ratio of 1 in the depth interval where both variables
show a concentration gradient (Haese et al., 2003).
This clearly reflects the stoichiometery of AOM;
sulfate reduction related to OM mineralization would
result in a ratio of 0.5.
If our assumptions are valid, according to the net-
stoichiometery of AOM proposed by Reeburgh
(1976) (Eq. (1)), the sulfate flux must be exactly
balanced by the methane flux (Borowski et al.,
1996). Once the methane flux is known, the methane
depth distribution can be reconstructed based on our
assumptions with an analytical solution to the well-
established one-dimensional, steady-state transport
equation for solutes accounting for diffusion and
advection (e.g. Berner, 1980). The modeled methane
depth distribution is shown in Fig. 5.
According to the model results by Haese et al.
(2003), a steep increase in the methane concentration
is predicted from the depth of AOM (14–18 cm) down
to a depth of about 1 m before the concentration
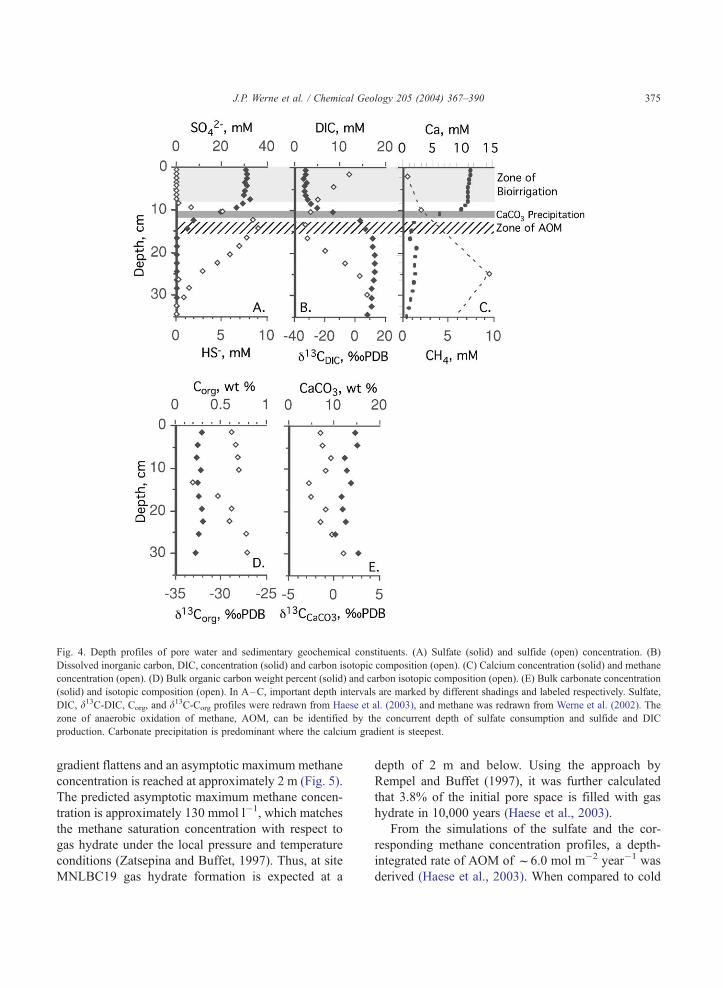
Fig. 4. Depth profiles of pore water and sedimentary geochemical constituents. (A) Sulfate (solid) and sulfide (open) concentration. (B)
Dissolved inorganic carbon, DIC, concentration (solid) and carbon isotopic composition (open). (C) Calcium concentration (solid) and methane
concentration (open). (D) Bulk organic carbon weight percent (solid) and carbon isotopic composition (open). (E) Bulk carbonate concentration
(solid) and isotopic composition (open). In A–C, important depth intervals are marked by different shadings and labeled respectively. Sulfate,
DIC, d13C-DIC, Corg, and d13C-Corg profiles were redrawn from Haese et al. (2003), and methane was redrawn from Werne et al. (2002). The
zone of anaerobic oxidation of methane, AOM, can be identified by the concurrent depth of sulfate consumption and sulfide and DIC
production. Carbonate precipitation is predominant where the calcium gradient is steepest.
J.P. Werne et al. / Chemical Geology 205 (2004) 367–390 375
gradient flattens and an asymptotic maximum methane
concentration is reached at approximately 2 m (Fig. 5).
The predicted asymptotic maximum methane concen-
tration is approximately 130 mmol l�1, which matches
the methane saturation concentration with respect to
gas hydrate under the local pressure and temperature
conditions (Zatsepina and Buffet, 1997). Thus, at site
MNLBC19 gas hydrate formation is expected at a
depth of 2 m and below. Using the approach by
Rempel and Buffet (1997), it was further calculated
that 3.8% of the initial pore space is filled with gas
hydrate in 10,000 years (Haese et al., 2003).
From the simulations of the sulfate and the cor-
responding methane concentration profiles, a depth-
integrated rate of AOM of f6.0 mol m�2 year�1 was
derived (Haese et al., 2003). When compared to cold
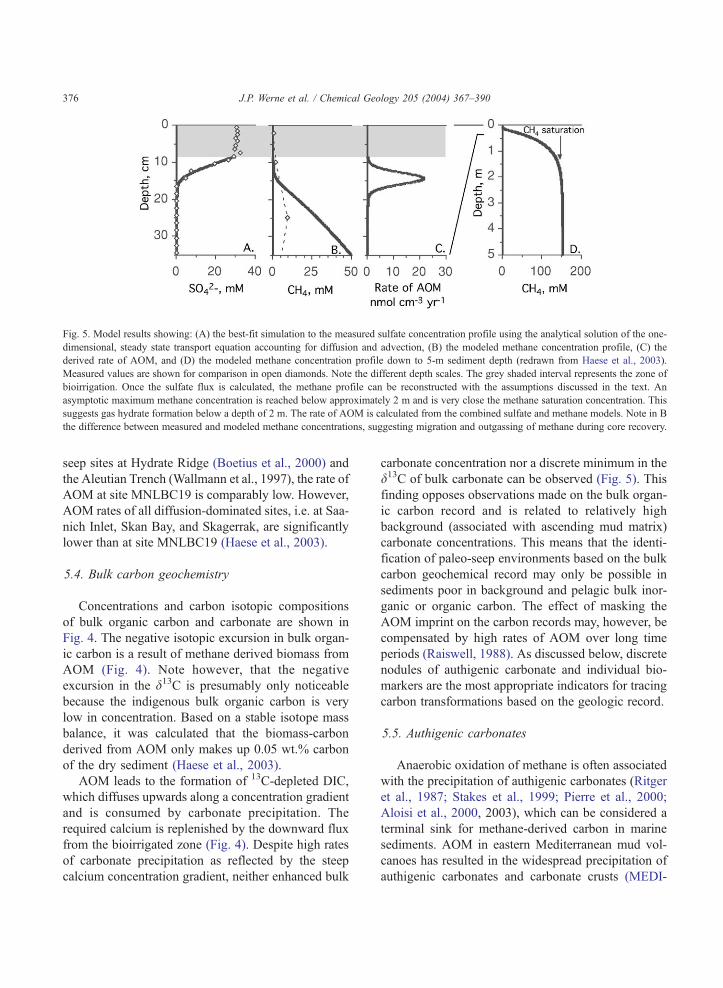
Fig. 5. Model results showing: (A) the best-fit simulation to the measured sulfate concentration profile using the analytical solution of the one-
dimensional, steady state transport equation accounting for diffusion and advection, (B) the modeled methane concentration profile, (C) the
derived rate of AOM, and (D) the modeled methane concentration profile down to 5-m sediment depth (redrawn from Haese et al., 2003).
Measured values are shown for comparison in open diamonds. Note the different depth scales. The grey shaded interval represents the zone of
bioirrigation. Once the sulfate flux is calculated, the methane profile can be reconstructed with the assumptions discussed in the text. An
asymptotic maximum methane concentration is reached below approximately 2 m and is very close the methane saturation concentration. This
suggests gas hydrate formation below a depth of 2 m. The rate of AOM is calculated from the combined sulfate and methane models. Note in B
the difference between measured and modeled methane concentrations, suggesting migration and outgassing of methane during core recovery.
J.P. Werne et al. / Chemical Geology 205 (2004) 367–390376
seep sites at Hydrate Ridge (Boetius et al., 2000) and
the Aleutian Trench (Wallmann et al., 1997), the rate of
AOM at site MNLBC19 is comparably low. However,
AOM rates of all diffusion-dominated sites, i.e. at Saa-
nich Inlet, Skan Bay, and Skagerrak, are significantly
lower than at site MNLBC19 (Haese et al., 2003).
5.4. Bulk carbon geochemistry
Concentrations and carbon isotopic compositions
of bulk organic carbon and carbonate are shown in
Fig. 4. The negative isotopic excursion in bulk organ-
ic carbon is a result of methane derived biomass from
AOM (Fig. 4). Note however, that the negative
excursion in the d13C is presumably only noticeable
because the indigenous bulk organic carbon is very
low in concentration. Based on a stable isotope mass
balance, it was calculated that the biomass-carbon
derived from AOM only makes up 0.05 wt.% carbon
of the dry sediment (Haese et al., 2003).
AOM leads to the formation of 13C-depleted DIC,
which diffuses upwards along a concentration gradient
and is consumed by carbonate precipitation. The
required calcium is replenished by the downward flux
from the bioirrigated zone (Fig. 4). Despite high rates
of carbonate precipitation as reflected by the steep
calcium concentration gradient, neither enhanced bulk
carbonate concentration nor a discrete minimum in the
d13C of bulk carbonate can be observed (Fig. 5). This
finding opposes observations made on the bulk organ-
ic carbon record and is related to relatively high
background (associated with ascending mud matrix)
carbonate concentrations. This means that the identi-
fication of paleo-seep environments based on the bulk
carbon geochemical record may only be possible in
sediments poor in background and pelagic bulk inor-
ganic or organic carbon. The effect of masking the
AOM imprint on the carbon records may, however, be
compensated by high rates of AOM over long time
periods (Raiswell, 1988). As discussed below, discrete
nodules of authigenic carbonate and individual bio-
markers are the most appropriate indicators for tracing
carbon transformations based on the geologic record.
5.5. Authigenic carbonates
Anaerobic oxidation of methane is often associated
with the precipitation of authigenic carbonates (Ritger
et al., 1987; Stakes et al., 1999; Pierre et al., 2000;
Aloisi et al., 2000, 2003), which can be considered a
terminal sink for methane-derived carbon in marine
sediments. AOM in eastern Mediterranean mud vol-
canoes has resulted in the widespread precipitation of
authigenic carbonates and carbonate crusts (MEDI-

Fig. 6. (a) Photo taken by submersible Nautile of authigenic
carbonate crust MN12BT3 in situ, prior to sampling. (b) Photo of
sample MN12BT3 in the lab after sampling.
J.P. Werne et al. / Chemical Geology 205 (2004) 367–390 377
NAUT/MEDINETH Shipboard Scientific Parties,
2000). The mineralogy and stable carbon and oxygen
isotope compositions of these carbonates have been
discussed in detail by Aloisi et al. (2000), so only data
from carbonates occurring on Kazan mud volcano will
be summarized herein.
Mud volcano-associated authigenic carbonates in
the eastern Mediterranean can be divided into several
categories based on mineralogy and morphology,
including crusts, nodules, and carbonate-cemented
sediments (both lithified pelagites and lithified mud
breccia sediments). Crusts range in thickness from
millimeter scale up to decimeter scale, with variable
morphology ranging from mounds to slabs (Figs. 3b
and 6) (Aloisi et al., 2000). The authigenic carbonate
crusts are composed of dolomite, aragonite, and
calcite in varying proportions, ranging from 70%
aragonite (Type ‘‘A’’ crusts of Aloisi et al., 2000) to
90% calcite (Type ‘‘D’’), and carbonate nodules are
up to 90% dolomite (Aloisi et al., 2000). Both low-
Mg and high-Mg (up to 12 mol% Mg) calcites have
been identified (Aloisi et al., 2000). Many of the
crusts are associated to some extent with macrofauna
such as pogonoforan tube worms and bivalves (Figs.
3b and 6; see discussion of macrofauna below).
The carbon isotopic composition of authigenic
carbonates (d13Ccarb) on Kazan mud volcano ranges
fromf0xto nearly �50x(PDB, Fig. 7; data from
Aloisi et al., 2000). Typically, carbonate crusts are
much more 13C depleted than poorly lithified mud
breccia, which is in turn generally more 13C depleted
than unlithified pelagites, due to the formation of fine-
grained authigenic carbonate minerals (Aloisi et al.,
2000). Controls on cold-seep carbonate d13C are
complex mainly due to uncertainties regarding mixing
ratios of different carbon sources having distinct iso-
topic compositions (bottom water DIC, OM, methane-
derived DIC, DIC in deep fluids) and the extent to
which carbon isotopes fractionate during AOM (Aloisi
et al., 2000). The d13Ccarb at Kazan mud volcano
suggests that methane is the major source of carbonate
carbon for crust formation and that this source is
somewhat diluted, probably by mixing with relatively13C-rich bottom water or rising fluids. OM decompo-
sition could play a minor role in contributing to the
DIC pool (Luff and Wallmann, 2003), however, given
the modeling results of Haese et al. (2003), such an
effect is expected to be minimal.
The oxygen isotopic composition of authigenic
carbonates (d18Ocarb) ranges from values of �1xto
+1x(PDB) in the case of nodules and unlithified
sediments, and from +2xto approximately +4.5xin lithified sediments and crusts (such as sample
MN12BT3) (Fig. 7). The enriched d18Ocarb signal is
interpreted by Aloisi et al. (2000) to indicate carbonate
precipitation from 18O-enriched diagenetic fluids, rath-
er than as a temperature effect. Possible processes
producing 18O-rich fluids include smectite dehydration
and gas hydrate destabilization. Pore water chemistry
and clay mineralogy suggest that the former process
might dominate here. In other areas of brine seepage in
the eastern Mediterranean (such as Nadir Brine Lake
or Napoli mud volcano in the Olimpi field), extreme18O-enrichment of the carbonate crusts is probably due
to precipitation from 18O-enriched fluids sourced from
relic Messinian brines (Aloisi et al., 2000). However,
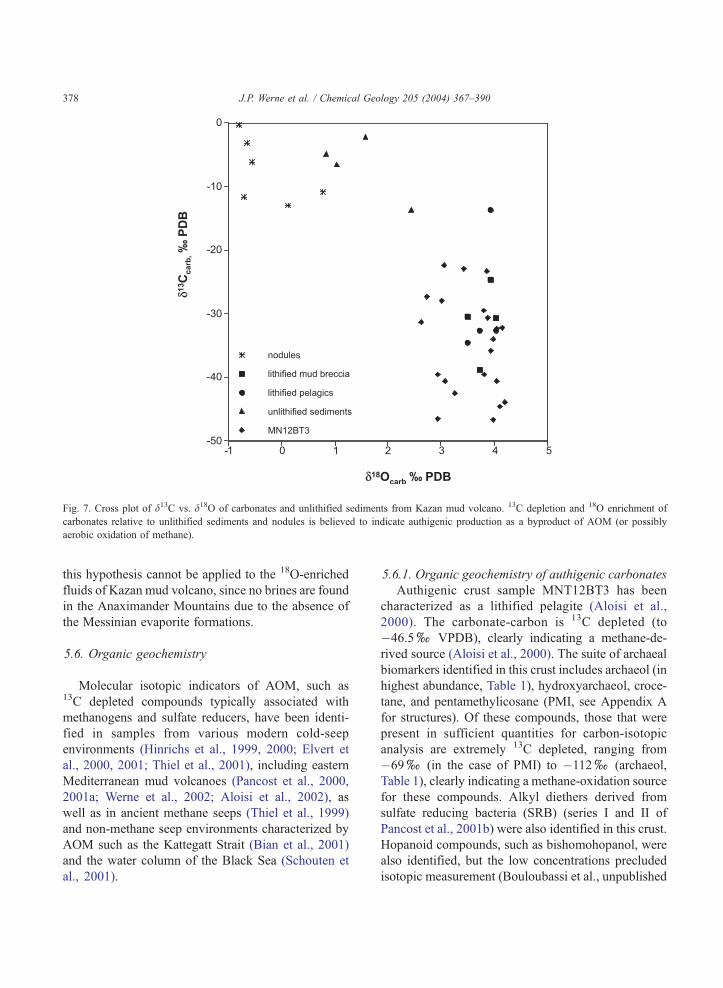
Fig. 7. Cross plot of d13C vs. d18O of carbonates and unlithified sediments from Kazan mud volcano. 13C depletion and 18O enrichment of
carbonates relative to unlithified sediments and nodules is believed to indicate authigenic production as a byproduct of AOM (or possibly
aerobic oxidation of methane).
J.P. Werne et al. / Chemical Geology 205 (2004) 367–390378
this hypothesis cannot be applied to the 18O-enriched
fluids of Kazan mud volcano, since no brines are found
in the Anaximander Mountains due to the absence of
the Messinian evaporite formations.
5.6. Organic geochemistry
Molecular isotopic indicators of AOM, such as13C depleted compounds typically associated with
methanogens and sulfate reducers, have been identi-
fied in samples from various modern cold-seep
environments (Hinrichs et al., 1999, 2000; Elvert et
al., 2000, 2001; Thiel et al., 2001), including eastern
Mediterranean mud volcanoes (Pancost et al., 2000,
2001a; Werne et al., 2002; Aloisi et al., 2002), as
well as in ancient methane seeps (Thiel et al., 1999)
and non-methane seep environments characterized by
AOM such as the Kattegatt Strait (Bian et al., 2001)
and the water column of the Black Sea (Schouten et
al., 2001).
5.6.1. Organic geochemistry of authigenic carbonates
Authigenic crust sample MNT12BT3 has been
characterized as a lithified pelagite (Aloisi et al.,
2000). The carbonate-carbon is 13C depleted (to
�46.5xVPDB), clearly indicating a methane-de-
rived source (Aloisi et al., 2000). The suite of archaeal
biomarkers identified in this crust includes archaeol (in
highest abundance, Table 1), hydroxyarchaeol, croce-
tane, and pentamethylicosane (PMI, see Appendix A
for structures). Of these compounds, those that were
present in sufficient quantities for carbon-isotopic
analysis are extremely 13C depleted, ranging from
�69x(in the case of PMI) to �112x(archaeol,
Table 1), clearly indicating a methane-oxidation source
for these compounds. Alkyl diethers derived from
sulfate reducing bacteria (SRB) (series I and II of
Pancost et al., 2001b) were also identified in this crust.
Hopanoid compounds, such as bishomohopanol, were
also identified, but the low concentrations precluded
isotopic measurement (Bouloubassi et al., unpublished

Fig. 8. Photo taken by submersible Nautile showing white patches
that were observed on Kazan mud volcano, believed to be
filamentous microbial ‘‘mats’’, perhaps Beggiatoa.
Table 1
Biomarkers and carbon isotopic data from crust MNT12BT3
Compound Relative
abundance
d13C(xVPDB)
Archaeol ++ �112
Hydroxyarchaeol + n.d.
Crocetane + n.d.
PMI:0 + �69
Series I alkyl diethers + to ++ �94 to �99
Series II alkyl diethers + f�70
‘‘Background’’ biomarkers + to +++ �23 to �32
Series I and II alkyl diethers refer to Pancost et al. (2001b).
‘‘Background’’ biomarkers (e.g., hopanoids, fatty acids, steroids,
and n-alkanes derived from bacteria and photoautotrophs) are
probably incorporated into authigenic carbonates from the ascend-
ing mud matrix that was lithified. See Appendix A for compound
structures. ‘‘n.d.’’ indicates no data.
J.P. Werne et al. / Chemical Geology 205 (2004) 367–390 379
results). In other studies, this compound has been
attributed to bacteria oxidizing sulfide (Pancost and
Sinninghe Damste, 2003), which seems reasonable in
this environment as well given the visual observation
of white filamentous Beggiatoa-like bacterial mats by
submersible (Fig. 8; and MEDINAUT/MEDINETH
Shipboard Scientific Parties, 2000) and the identifica-
tion of colorless sulfur bacteria in samples from Kazan
mud volcano sediment (Heijs, unpublished data, see
discussion on microbiology below). Other biomarker
compounds identified in comparable or greater abun-
dance than the AOM consortium biomarkers include
terrestrially derived n-alkanes, sterols, and fatty acids
which are believed to be associated with OM in the
ascending mud breccia based on their typical carbon
isotope compositions (f�24xto �32x, Table 1).
5.7. Organic geochemistry of mud breccia sediments
Themolecular isotopic signature in coreMNLBC19
taken from the mud breccia of Kazan mud volcano
shows similarities to the crust, but also shows some
striking differences. For example, 13C depleted (to
�123.8x) archaeal compounds such as archaeol, hy-
droxyarchaeol, and PMI have been identified in the
sediments (Table 2, some data fromWerne et al., 2002).
We have also identified several non-isoprenoid dialkyl
glycerol diethers attributed to SRB, but only series II
compounds of Pancost et al. (2001b) were identified in
the sediments (compared to dominantly series I in the
crust).
Hopanoid compounds such as diploptene and dip-
lopterol were also identified with ‘‘intermediate’’
degrees of 13C depletion (f�55xto�70x), which
were attributed to aerobic methane-oxidizing bacteria
(Werne et al., 2002) based in part on decreasing down-
core concentration trends. h,h-bishomohopanol was
also identified in surficial sediments (Werne and Sin-
ninghe Damste, unpublished results), as in the crust.
This compound has been attributed to sulfide-oxidizing
bacteria in other studies (Pancost and Sinninghe Dam-
ste, 2003).
A series of glycerol dialkyl glycerol tetraethers
(GDGTs) was also identified in the sediments of Kazan
mud volcano (Werne and Sinninghe Damste, unpub-
lished results). Based on molecular isotopic studies in
other mud volcano cold seep sediments (Pancost et al.,
2001a,b) and in the water column of the Black Sea
(Schouten et al., 2001), it has been proposed that some
of these GDGT compounds are derived at least in part
from anaerobic methane-oxidizing archaea. In Kazan
mud breccia sediments, however, carbon isotope anal-
ysis of C40 isoprenoids released from these GDGTs by
chemical degradation show 13C enriched values, in the
range of �22x, indeed the most 13C enriched com-
pounds in the extractable OM (Werne and Sinninghe
Damste, unpublished results). Such values clearly
indicate a non-methane-related source for these com-
pounds, likely derived primarily from Crenarchaeota
ubiquitously occurring in the pelagic zone of the water
column (Schouten et al., 2000), and are therefore more
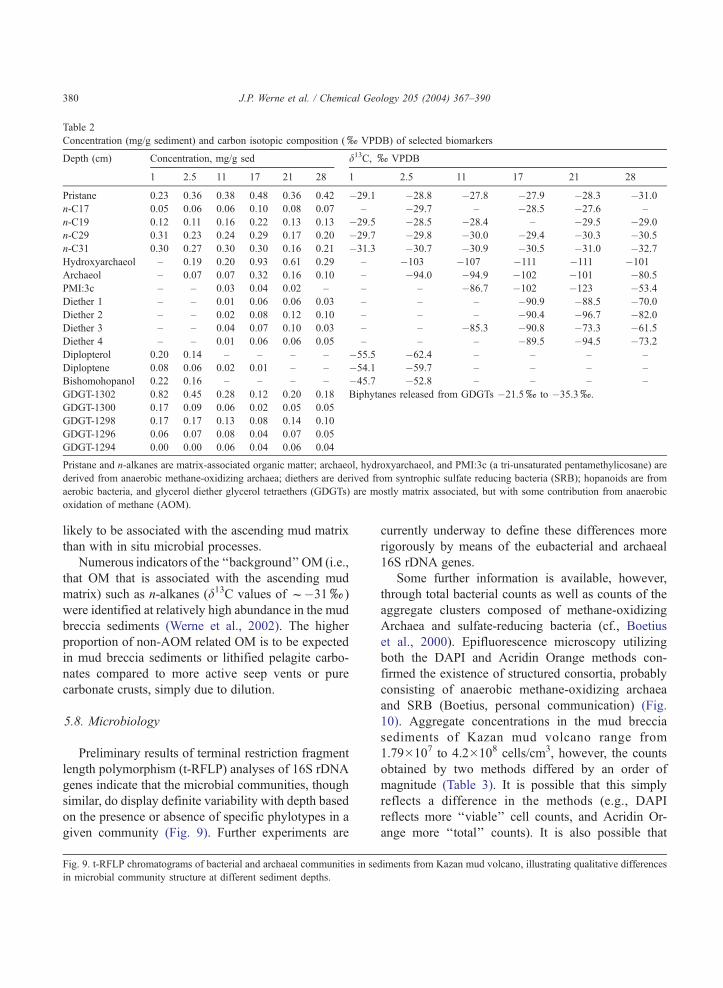
Table 2
Concentration (mg/g sediment) and carbon isotopic composition (xVPDB) of selected biomarkers
Depth (cm) Concentration, mg/g sed d13C, xVPDB
1 2.5 11 17 21 28 1 2.5 11 17 21 28
Pristane 0.23 0.36 0.38 0.48 0.36 0.42 �29.1 �28.8 �27.8 �27.9 �28.3 �31.0
n-C17 0.05 0.06 0.06 0.10 0.08 0.07 – �29.7 – �28.5 �27.6 –
n-C19 0.12 0.11 0.16 0.22 0.13 0.13 �29.5 �28.5 �28.4 – �29.5 �29.0
n-C29 0.31 0.23 0.24 0.29 0.17 0.20 �29.7 �29.8 �30.0 �29.4 �30.3 �30.5
n-C31 0.30 0.27 0.30 0.30 0.16 0.21 �31.3 �30.7 �30.9 �30.5 �31.0 �32.7
Hydroxyarchaeol – 0.19 0.20 0.93 0.61 0.29 – �103 �107 �111 �111 �101
Archaeol – 0.07 0.07 0.32 0.16 0.10 – �94.0 �94.9 �102 �101 �80.5
PMI:3c – – 0.03 0.04 0.02 – – – �86.7 �102 �123 �53.4
Diether 1 – – 0.01 0.06 0.06 0.03 – – – �90.9 �88.5 �70.0
Diether 2 – – 0.02 0.08 0.12 0.10 – – – �90.4 �96.7 �82.0
Diether 3 – – 0.04 0.07 0.10 0.03 – – �85.3 �90.8 �73.3 �61.5
Diether 4 – – 0.01 0.06 0.06 0.05 – – – �89.5 �94.5 �73.2
Diplopterol 0.20 0.14 – – – – �55.5 �62.4 – – – –
Diploptene 0.08 0.06 0.02 0.01 – – �54.1 �59.7 – – – –
Bishomohopanol 0.22 0.16 – – – – �45.7 �52.8 – – – –
GDGT-1302 0.82 0.45 0.28 0.12 0.20 0.18 Biphytanes released from GDGTs �21.5xto �35.3x.
GDGT-1300 0.17 0.09 0.06 0.02 0.05 0.05
GDGT-1298 0.17 0.17 0.13 0.08 0.14 0.10
GDGT-1296 0.06 0.07 0.08 0.04 0.07 0.05
GDGT-1294 0.00 0.00 0.06 0.04 0.06 0.04
Pristane and n-alkanes are matrix-associated organic matter; archaeol, hydroxyarchaeol, and PMI:3c (a tri-unsaturated pentamethylicosane) are
derived from anaerobic methane-oxidizing archaea; diethers are derived from syntrophic sulfate reducing bacteria (SRB); hopanoids are from
aerobic bacteria, and glycerol diether glycerol tetraethers (GDGTs) are mostly matrix associated, but with some contribution from anaerobic
oxidation of methane (AOM).
J.P. Werne et al. / Chemical Geology 205 (2004) 367–390380
likely to be associated with the ascending mud matrix
than with in situ microbial processes.
Numerous indicators of the ‘‘background’’ OM (i.e.,
that OM that is associated with the ascending mud
matrix) such as n-alkanes (d13C values of f�31x)
were identified at relatively high abundance in the mud
breccia sediments (Werne et al., 2002). The higher
proportion of non-AOM related OM is to be expected
in mud breccia sediments or lithified pelagite carbo-
nates compared to more active seep vents or pure
carbonate crusts, simply due to dilution.
5.8. Microbiology
Preliminary results of terminal restriction fragment
length polymorphism (t-RFLP) analyses of 16S rDNA
genes indicate that the microbial communities, though
similar, do display definite variability with depth based
on the presence or absence of specific phylotypes in a
given community (Fig. 9). Further experiments are
Fig. 9. t-RFLP chromatograms of bacterial and archaeal communities in se
in microbial community structure at different sediment depths.
currently underway to define these differences more
rigorously by means of the eubacterial and archaeal
16S rDNA genes.
Some further information is available, however,
through total bacterial counts as well as counts of the
aggregate clusters composed of methane-oxidizing
Archaea and sulfate-reducing bacteria (cf., Boetius
et al., 2000). Epifluorescence microscopy utilizing
both the DAPI and Acridin Orange methods con-
firmed the existence of structured consortia, probably
consisting of anaerobic methane-oxidizing archaea
and SRB (Boetius, personal communication) (Fig.
10). Aggregate concentrations in the mud breccia
sediments of Kazan mud volcano range from
1.79�107 to 4.2�108 cells/cm3, however, the counts
obtained by two methods differed by an order of
magnitude (Table 3). It is possible that this simply
reflects a difference in the methods (e.g., DAPI
reflects more ‘‘viable’’ cell counts, and Acridin Or-
ange more ‘‘total’’ counts). It is also possible that
diments from Kazan mud volcano, illustrating qualitative differences
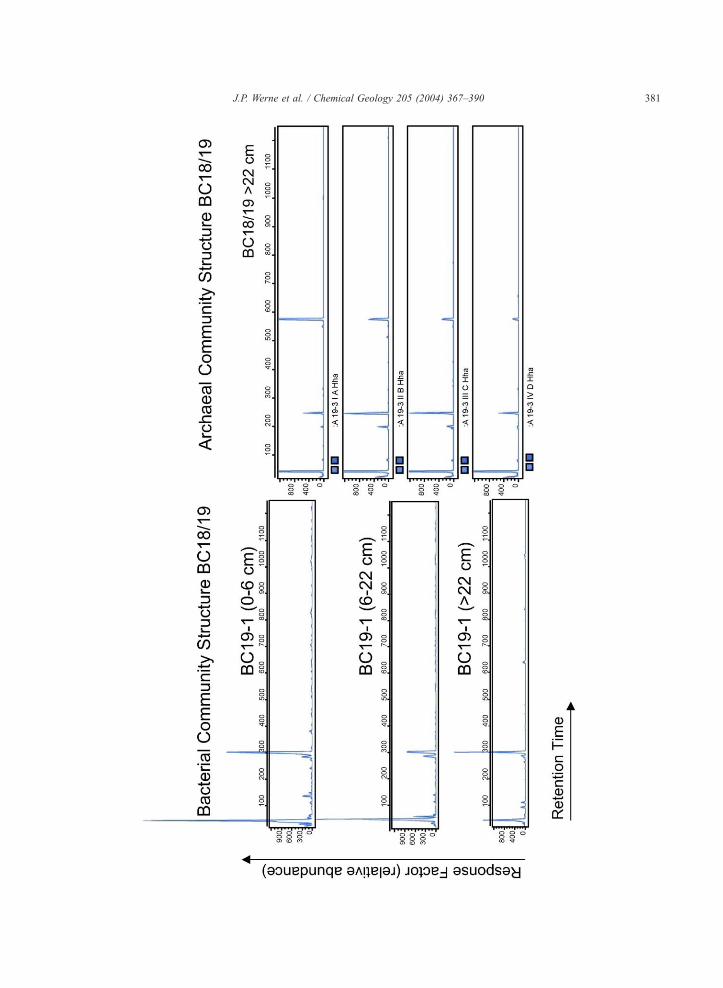
J.P. Werne et al. / Chemical Geology 205 (2004) 367–390 381

Fig. 10. Archaea/SRB aggregates identified using fluorescence in situ hybridization (FISH) and stained with DAPI. Archaea are shown here in
green and SRB are shown in red.
Table 4
Most probable number (MPN) counts
Dilution Water 0–6 cm 6–22 cm >22 cm
WI WII WIII SI SII SIII SI SII SIII SI SII SIII
Sulfate-reducing bacteria
100 � � � / / / / / / / / /
1000 � � � � � � � � � � � �10,000 � � � � � � � � � � � �100,000 � � � � � � � � � � � �
Colorless sulfur bacteria
100 +++ +++ +++ / / / / / / / / /
1000 � � +++ � � � � +++ � � � �10,000 � � +++ � � � � � � � � �100,000 � � � � � � � � � � � �
Methanogens
100 � � � / / / / / / / / /
1000 � � � �x � � � + � � � �10,000 � � � � � + � � �x � � �
J.P. Werne et al. / Chemical Geology 205 (2004) 367–390382
DAPI numbers may be more reliable, due in part to
the very low quantities of DNA that were extractable
from these sediment samples.
Another interesting feature of the microbiological
data is found in the most probable number (MPN)
counts. Data from MPN counts indicate the likely
existence of colorless sulfur bacteria (e.g. sulfide
oxidizers) and methanogens in sediments from Kazan
mud volcano above 22 cm depth, but give no indica-
tion of the existence of SRB or aerobic methane
oxidizers (Table 4). These results suggest a number
of possibilities. First, it is not surprising to see
evidence of colorless sulfur bacteria in the MPN
counts given the visual observation of ‘‘white
patches’’ by submersible dives (Fig. 8) which are
believed to be sulfide oxidizers similar to Beggiatoa.
Second, it is possible that there are no SRB in these
sediments, but this seems very unlikely given the
biomarker and pore water geochemical data discussed.
Alternatively, it is possible that the SRB that are living
syntrophically with the anaerobic methane-oxidizing
archaea are not culturable under the conditions uti-
lized, and therefore do not appear in the MPN counts.
Table 3
Microbial cell and aggregate counts (DAPI and Acridin Orange)
from core MNLBC18/19
Sample
interval
(cm)
DAPI counts
(cells/cm3)
Acridin Orange
counts (cells/cm3)
Aggregate counts
(aggregates/cm3)
0–6 2.14�107 4.2�108 few
6–22 1.79�107 2.6�108 few
>22 2.99�107 1.3�108 F106
aggregates/cm3
As these consortia have proven difficult to culture (cf.
Boetius et al., 2000; Michaelis et al., 2002), the
second hypothesis seems more likely.
100,000 � � � � � � � � � � � �
Methane-oxidizing bacteria
100 � + � / / / / / / / / /
1000 � � � � � � � � � � � �10,000 � � � � � � � � � � � �100,000 � � � � � � � � � � � �Symbols used are: (�) indicates not present; (/) indicates not
analyzed due to pre-dilution of 100 times; (+), (++), and (+++)
indicate the relative level of likely concentration; (x) indicates
sample contaminated by oxygen. WI, WII, WIII and SI, SII, SIII
indicate water and sediment samples used for culture work, run in
triplicate.

l Geology 205 (2004) 367–390 383
5.9. Chemosynthetic macrofauna
Dense communities composed of small bivalves and
vestimentiferan tube worms were discovered on east-
ern Mediterranean mud volcanoes (Fig. 11, cf. Fig. 2b
in MEDINAUT/MEDINETH Shipboard Scientific
Party, 2000). During the 1998 MEDINAUT cruise,
living individuals of several species were sampled by
submersible from the top of Kazan mud volcano
(35j25.983VN, 24j33.594VE, 1707 m water depth;
Fiala-Medioni et al., 2001; Olu-Le Roy et al., in press).
Among the organisms collected were lucinid bivalves:
the most abundant species was identified asMyrtea sp.
(Olu-Le Roy et al., 2001, in press); few samples of
another species were found and described as Lucinoma
kasani (Salas and Woodside, 2002). Other living
bivalves included a small vesicomyid, identified as
J.P. Werne et al. / Chemica
Fig. 11. Photo taken by submersible Nautile showing types of
chemosynthesis-dependent macrofauna observed on Kazan mud
volcano. (a) Vestimentiferan tube worms. (b) Lucinid, vesicomyid,
and mytilid bivalves.
Isorropodon perplexum (Olu-Le Roy et al., 2001, in
press; von Cosel and Salas, 2001) and a small mytilid
attached to carbonate crusts and identified as Idas
modiolaeformis (Olu-Le Roy et al., 2001, in press).
Some tubes of living worms belonging to Lamellibra-
chia sp. (Olu-Le Roy et al., 2001, in press) were also
collected at the site. On top of a strongly reduced
sediment, white-grey bacterial mats (Fig. 8) were
observed as well as many echinidae sea urchins and a
number of unidentified crabs.
TEM studies of the gills of Myrtea sp. and Vesico-
mya sp., as well as trophosomes of vestimentiferan
worms demonstrated that they were composed of
bacteriocytes housing sulfide-oxidizing bacteria of
various morphological types as found in other symbi-
otic organisms from vents or cold seeps (Fiala-Medi-
oni and Felbeck, 1990; Fisher, 1995). In contrast, the
mytilid gill houses both sulfide-oxidizing and meth-
anotrophic symbionts (Fiala-Medioni et al., 2001) as
found in Bathymodiolus puteosrpentis (Cavanaugh et
al., 1992) or B. azoricus (Fiala-Medioni et al., 2002).
Carbon isotopic compositions of the gills of bivalves
also suggested chemosynthesis-based metabolism,
ranging from values �27.7 xto �30.5 xfor M.
sp. and I. perplexum and from �23.6xto �26.6xfor the tube worm Lamellibrachia sp. (Fiala-Medioni
et al., 2001); these values are in the range of those
found for other sulfide-oxidizing symbionts in differ-
ent vent or cold seep sites (Fiala-Medioni and Felbeck,
1990; Kennicutt et al., 1992; Fisher, 1995).
The values measured on the Kazan mud volcano
mytilid I. modiolaeformis are more depleted than those
of the lucinid, ranging from �44.2xto �44.6x(Fiala-Medioni et al., 2001). Such values reflect the
presence of methylotrophic symbionts which may use
methane as an energy source as found for B. childressi,
a cold seep species collected off Louisiana (Nelson and
Fisher, 1995). TEM and isotopic results led to the con-
clusion that these species are mainly relying, through
their symbionts, on reduced sulfur produced in the se-
diment, with the exception of the mytilid which is prob-
ably able to use both sulfide produced in the sediment
and/ormethane expelled in the fluids (Olu-LeRoy et al.,
in press). The chemosynthetic macro- and megafauna
are located at sites where fluids are expelled (Olu et al.,
1997; Sibuet and Olu, 1998), but there is currently no
statistical evidence for any direct relationship between
methane concentrations and living biomass. With the

J.P. Werne et al. / Chemical Geology 205 (2004) 367–390384
exception of the mytilid, such a relationship appears
indirect for Kazan mud volcano species where no
methylotrophic-like symbionts were observed. Within
Mediterranean mud volcano fields, it has been docu-
mented that concentrations of methane increase with
depth in the uppermost meter of the mud breccias. In
most cases, this appears to be associated with the depth
zone of sulfate reduction (de Lange and Brumsack,
1998) and results in the generation of inorganic carbon
and hydrogen sulfide which can be used as a substrate
by the sulfide oxidizing symbionts of the macrofauna.
6. Summary and biogeochemical synthesis
AOM is a significant biogeochemical process in
many cold-seep sediments, evident in everything from
pore water and sedimentary geochemical profiles to
authigenic carbonates to (chemosynthetic) macrofau-
nal abundance and diversity. In sediments from Kazan
mud volcano, measured and modeled pore water
profiles of the concentrations of sulfate, sulfide, dis-
solved inorganic carbon (DIC) and d13CDIC all indi-
cate that AOM is presently occurring in the sediments
of Kazan mud volcano (Fig. 4, Haese et al., 2003). It
is clear from these measured and modeled pore water
profiles that sulfate and methane are consumed in the
same depth interval, coincident with the production of
sulfide and 13C-depleted DIC. Indeed, d13CDIC values
of �35x(PDB) require the oxidation of methane to
CO2 (Whiticar and Faber, 1986; Alperin et al., 1988;
Whiticar, 1999). 13C depleted authigenic carbonates
and TOC are identified in the zone where AOM is
believed to be occurring based on pore water profiles
(Figs. 4–7). Further evidence of AOM is found in 13C
depleted biomarkers of methane-oxidizing archaea and
SRB (Table 2). Visual observation of microbial con-
sortium aggregates through FISH (Fig. 10) and che-
mosynthetic macrofauna such as bivalves and tube
worms (Fig. 11) confirm the existence of organisms
that are dependent on methane and the byproducts of
AOM (such as the sulfide utilized by bacterial sym-
bionts dwelling in the tissues of the macrofauna).
The porewater (sulfate, sulfide, DIC) and solid
phase (bulkOM, and selected biomarkers) geochemical
profiles suggest that this process is restricted to a
narrow depth interval, where methane is completely
oxidized. This hypothesis is supported by the lack of
MPN response for aerobic methane oxidizers in even
the most surficial sediments of Kazan. The lack of mi-
crobiological evidence for aerobicmethane oxidizers in
the sediments further suggests that there are no aerobic
organisms utilizing methane escaping the AOM zone
(Werne et al., 2002), despite the presence of aerobic
methane-oxidizer biomarkers (diploptene and di-
plopterol) in near surface sediments. An alternative
explanation is that these compounds are derived from
an early, rapid phase of aerobic methane oxidation that
took place immediately after mud breccia emplacement
rather than from presently active bacteria. This hypoth-
esis is more attractive given that microbiological data
indicates the absenceofmethane-oxidizingbacteria cur-
rently living in Kazan mud volcano surficial sediments.
It is also conceivable, however, that the biomarkers
attributed to aerobic methane oxidizing bacteria by
Werne et al. (2002) could be derived from other
bacteria in this environment. Indeed, these biomarkers
are fairly general bacterial biomarkers, and there is
strong evidence for the existence of colorless sulfur
bacteria, both in MPN counts (Table 4) as well as in
visual observations of white patches (which may have
been Beggiatoa-like white filamentous bacteria) made
by Nautile submersible dives (MEDINAUT/MEDI-
NETH Shipboard Scientific Parties, 2000). In addition,
other biomarkers identified in Kazan mud volcano
sediments and other cold seep environments, such as
bishomohopanol, have been attributed to sulfide-oxi-
dizing bacteria (this study; Pancost et al., 2002).
Under such conditions of complete (or near-com-
plete) methane oxidation, only small amounts of meth-
ane are released into the bottom waters. This release is
typically associated with major faults where highest
methane fluxes (due to advective flow) can be expec-
ted. This aspect of fluid fluxes and methane oxidation
processes explains why bottom-water methane concen-
trations are so variable over mud volcanoes in the
eastern Mediterranean and the highest methane con-
centrations in the deep Mediterranean are identified
above Amsterdam mud volcano (Charlou et al., 2003),
which is characterized by intense active tectonism. (Ac-
tive) faults also serve as preferred fluid pathways
(focused flow), which leads to a high spatial heteroge-
neity of solute fluxes, fluid flow, and microbial andma-
crobenthic abundances, as identified through visual
observations made during dives of the submersible
Nautile.

J.P. Werne et al. / Chemical Geology 205 (2004) 367–390 385
The production of authigenic carbonates as a by-
product of AOM represents one of the major terminal
sinks for methane-derived carbon in cold seep envi-
ronments. This process also keeps methane out of the
atmosphere thereby reducing the impact of methane
on global climate change. The high variability in the
carbon isotopic composition of these carbonates can
be explained by the fact that: (1) they form both as a
result of chemical supersaturation and microbial cal-
cification (Aloisi et al., 2000, 2002); and (2) slight
variations in the depth of precipitation of carbonates
within the sediments will substantially alter their
isotopic signature due to the steep gradients in d13Cof the available DIC (Aloisi et al., 2000, 2002). One
implication of these observations is that deriving rates
of carbonate formation from pore water gradients may
underestimate the true calcification rate due to the ad-
ditional contribution from microbes and the very steep
and likely variable gradients in the concentrations and
carbon isotope composition of DIC resulting from
AOM.
Series I diethers are higher in concentration and
more 13C depleted than series II diethers (ca. �97xcompared to �70x ; Table 1) in the authigenic
carbonate investigated here. This pattern differs from
that observed in carbonates from Amsterdam mud
volcano (also in the Anaximander Mountains area)
where the two series are approximately equivalent in
abundance, as well as to authigenic carbonates from
Napoli mud volcano (Olimpi area), where series I
diethers are lower in concentration than series II
diethers (Bouloubassi, unpublished results). Further-
more, in mud breccia sediments from Kazan mud
volcano, only series II diethers were identified. These
observations suggest that different SRB could be par-
ticipating in the microbial consortium with the archaeal
methane-oxidizers in different environments, possibly
as a result of the different salinities of the two environ-
ments. Furthermore, different SRB may be active
syntrophic partners with archaea in seep areas charac-
terized by carbonate precipitation compared to areas
not precipitating carbonate within the same mud vol-
cano environment.
Additional support for the hypothesis that very
different microbial consortia are carrying out AOM in
different environments is found in the differences
between GDGT compounds found on Kazan mud
volcano and those identified elsewhere. While the
isotopic composition of C40 isoprenoids released from
GDGTs of Kazan mud volcano are up to �22x,
significantly 13C-depleted C40 isoprenoids were iden-
tified in authigenic carbonate crusts from Olimpi mud
volcano (Bouloubassi et al., unpublished results), as
well as in seeps from Milano and Amsterdam mud
volcanoes, and crusts, bacterial mats, seeps, and brines
from Napoli mud volcano (Pancost et al., 2001a).
These contrasting results in different cold seep loca-
tions, even within eastern Mediterranean mud volca-
noes, strongly suggest that the specific microbial
communities vary depending on the geochemistry of
the environment, including factors such as methane
concentration and flux, and salinity.
The archaeal/SRB aggregates (as determined by the
Acridin Orange method) show the greatest abundance
in the deepest sediment layer analyzed (>22 cm). This
contrasts with the pore water geochemistry and solid
phase bulk geochemical data, which suggests present-
day maximum AOM rates in the region around 13–15
cm depth. The lipid profiles, however, display a max-
imum in AOM indicators (e.g., 13C depleted specific
biomarkers) at an intermediate sediment depth of 16–
18 cm. The discrepancies in the geochemical and
microbiological data suggest that the historical center
of AOM in this environment may have migrated
vertically in the sediments, likely as a result of varia-
tions in the rate of the methane flux. While such a hy-
pothesis is not unlikely, it should be noted that the
microbiological analyses were taken on a separate adja-
cent core, so slight variations in the depth of AOM are
expected due to the lateral spatial variability in all
aspects of a mud volcano, from fluid venting to surface
topography. Furthermore, it is generally acknowledged
that lipid profiles and bulk OM provide a more ‘‘time-
averaged’’ view of the sedimentary processes, while
pore water geochemistry is thought to give more of a
‘‘snapshot’’ view of present-day conditions. DNA has
been observed to degrade on time-scales from 10min to
100,000 years, depending on specific circumstances.
These differences in the interpretable time frame of the
different geochemical and microbiological indicators
can often make interpretation problematical.
Model calculations estimate a rate of AOM of f6
mol m�2 year�1. The advective flow velocity at Kazan
mud volcano was found to be 4 cm year�1. Compared
to other cold seep sites, the advective flow velocity and
the rate of AOM are rather moderate (Haese et al.,
J.P. Werne et al. / Chemical Geology 205 (2004) 367–390386
2003). Based on the modeled methane concentration
profile and the state of methane saturation with respect
to gas hydrate, hydrate formation is expected at a
sediment depth of 2 m and below at site MNLBC19.
Whereas gas hydrate stability is thermodynamically
favorable on Kazan mud volcano, the rate of hydrate
formation is rather slow (Haese et al., 2003). These
findings may best explain the rare occasions of gas
hydrate recoveries during two expeditions to Kazan
mud volcano.
Acknowledgements
We would like to thank the Captain and crew of the
R/V Nadir, the submersible Nautile, and the R/V
Professor Logachev for their tireless assistance during
sampling. We also thank M. Kienhuis, E. Hopmans,
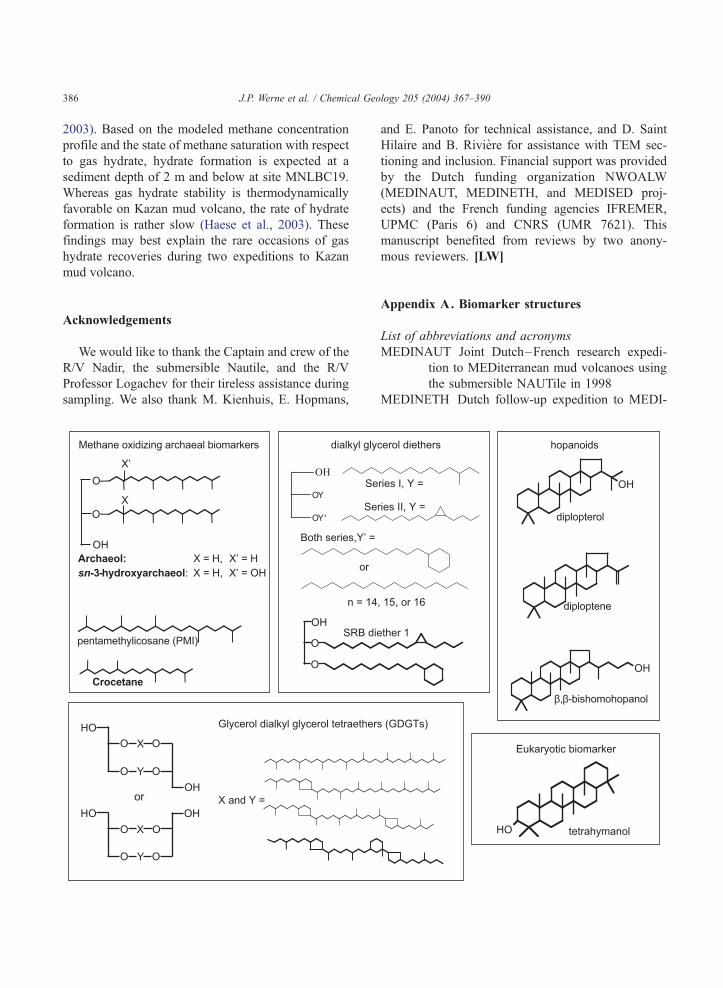
pentamethylicosane (PMI)
dialkyl gly
OH
O
O
Archaeol: X = H, X’ = H
sn-3-hydroxyarchaeol: X = H, X’ = OH
O
O
X
OH
X’
Crocetane
SRB di
OY '
OY
OH
Se
Se
Both series,Y’ =
or
n = 14
O
HO
X O
OH
O
HO
X
Y O
O Y O
O O
OHor
Glycerol dialkyl glycerol tetraether
X and Y =
Methane oxidizing archaeal biomarkers
and E. Panoto for technical assistance, and D. Saint
Hilaire and B. Riviere for assistance with TEM sec-
tioning and inclusion. Financial support was provided
by the Dutch funding organization NWOALW
(MEDINAUT, MEDINETH, and MEDISED proj-
ects) and the French funding agencies IFREMER,
UPMC (Paris 6) and CNRS (UMR 7621). This
manuscript benefited from reviews by two anony-
mous reviewers. [LW]
Appendix A. Biomarker structures
List of abbreviations and acronyms
MEDINAUT Joint Dutch–French research expedi-
tion to MEDiterranean mud volcanoes using
the submersible NAUTile in 1998
MEDINETH Dutch follow-up expedition to MEDI-
cerol diethers
OH
OH
diplopterol
diploptene
β,β-bishomohopanol
HO tetrahymanol
ether 1
ries I, Y =
ries II, Y =
, 15, or 16
s (GDGTs)
hopanoids
Eukaryotic biomarker

J.P. Werne et al. / Chemical Geology 205 (2004) 367–390 387
NAUT, focused on higher resolution geo-
physical surveys and sediment sampling in
1999 (MEDiterranean NETHerlands)
SBP Sub-Bottom Profiler
DAPI 4V,6V-DiAmidino-2-PhenylIndole
hydrochloride
MPN Most Probable Numbers
PCR Polymerase Chain Reaction
HPLC High Performance Liquid Chromatography
t-RFLP terminal-Restriction Fragment Length
Polymorphism
ANAXIPROBE Two phase research program carried
out in 1995 and 1996 to investigate the
origin and evolution of the Anaximander
mountains
AOM Anaerobic Oxidation of Methane
DIC Dissolved Inorganic Carbon
FISH Fluorescence in situ Hybridization
d13Cx stable carbon isotope composition of sub-
stance x
(V)PDB (Vienna) Pee Dee Belemnite, international
standard against which carbon and oxygen
isotope ratios are calibrated
PMI Pentamethylicosane
SRB Sulfate-Reducing Bacteria
GDGT Glycerol Dialkyl Glycerol Tetraether
C40 Carbon chain with 40 carbons
TOC Total Organic Carbon
TEM Transmission Electron Microscopy
OM Organic Matter
References
Aloisi, G., Pierre, C., Rouchy, J.-M., Foucher, J.-P., Wood-
side, J., and the MEDINAUT Shipboard Scientific Party,
2000. Methane-related authigenic carbonates of eastern Med-
iterranean Sea mud volcanoes and their possible relation to
gas hydrate destabilisation. Earth Planet. Sci. Lett. 184,
321–338.
Aloisi, G., Bouloubassi, I., Heijs, S.K., Pancost, R.D., Pierre, C.,
Sinninghe Damste, J.S., Gottschal, J.C., Forney, L.J., Rouchy,
J.-M., 2002. CH4-consuming microorganisms and the formation
of carbonate crusts at cold-seeps. Earth Planet. Sci. Lett. 203,
195–203.
Alperin, M.J., Reeburgh, W.S., Whiticar, M.J., 1988. Carbon
and hydrogen isotope fractionation resulting from anaero-
bic methane oxidation. Global Biogeochem. Cycles 2,
279–288.
Ben-Avraham, Z., Kempler, D., Ginzburg, A., 1988. Plate tectonics
in the Cyprus Arc. Tectonophysics 146, 231–240.
Berner, R.B., 1980. Early Diagenesis: A Theoretical Approach.
Princeton Univ. Press, Princeton, NY, p. 241.
Bian, L., Hinrichs, K., Xie, T., Brassell, S.C., Iversen, N., Fossing,
H., Jorgensen, B.B., Hayes, J.M., 2001. Algal and archaeal
polyisoprenoids in a recent marine sediment: molecular isotopic
evidence for anaerobic oxidation of methane. Geochem. Geo-
phys. Geosyst. 2 (1), doi:10.1029/2000GC000112.
Boetius, A., Ravenschlag, K., Schubert, C.J., Rickert, D., Widdel,
F., Gieseke, A., Amann, R., Jørgensen, B.B., Witte, U., Pfann-
kuche, O., 2000. A marine microbial consortium apparently me-
diating anaerobic oxidation of methane. Nature 407, 623–626.
Borowski, W.S., Paull, C.K., Ussler III, W. 1996. Marine pore water
sulfate profiles indicate in situ methane flux from underlying gas
hydrate. Geology 24, 655–658.
Bouton, T.W., 1991. Stable carbon isotope ratios of natural materi-
als: I. Sample preparation and mass spectrometric analysis. In:
Coleman, D., Fry, B. (Eds.), Carbon Isotope Techniques. Aca-
demic Press, San Diego, pp. 155–172.
Brown, K.M., 1990. The nature and hydrogeologic significance of
mud diapers and diatremes for accretionary systems. J. Geo-
phys. Res. 95 (B6), 8969–8982.
Cavanaugh, C.M., Wirsen, C., Jannasch, H.J., 1992. Evidence for
methylotrophic symbionts in a hydrothermal vent mussel (Bival-
via: Mytilidae) from the Mid-Atlantic Ridge. Appl. Environ.
Microbiol. 58, 3799–3803.
Chamley, H., Dunoyer-de-Segonzac, G., Melieres, F., 1978. Clay
minerals in Messinian sediments of the Mediterranean Sea.
DSDP 42, 389–395.
Charlou, J.L., Donval, J.P., Zitter, T., Roy, N., Jean-Baptiste, P.,
Foucher, J.P., Woodside, J., MEDINAUT Scientific Party,
2003. Evidence of methane venting and geochemistry of brines
on mud volcanoes of the eastern Mediterranean Sea. Deep-Sea
Res., Part 1, Oceanogr. Res. Pap. 50, 941–958.
Chaumillon, E., Mascle, J., 1997. From foreland to fore-arc
domains: new multichannel seismic reflection survey of the
Mediterranean Ridge accretionary complex (Eastern Mediterra-
nean). Mar. Geol. 138, 237–259.
Cita, M.B., Ryan, W.B.F., Paggi, L., 1981. Prometheus mud brec-
cia. An example of shale diapirism in the Western Mediterra-
nean Ridge. Ann. Geol. Pays Hell. 30, 543–569.
Cita, M.B., Camerlenghi, A., Erba, E., McCoy, F.W., Castadrori, D.,
Cazzani, A., Guasti, G., Gambastiani, M., Lucchi, R., Nolli, V.,
1989. Discovery of Mud diapirism on Mediterranean Ridge. A
preliminary report. Boll. Soc. Geol. Ital. 108, 537–543.
Cita, M.B., Ivanov, M.K., Woodside, J.M., 1996. The Mediterra-
nean ridge diapiric belt. Mar. Geol. 132, 1–6.
Cronin, B.T., Ivanov, M.K., Limonov, A.F., Egorov, A., Akhmanov,
G.G., Akhmetjanov, A.M., Kozlova, E., TTR-5, S.S.P., 1997.
New discoveries of mud volcanoes on the Eastern Mediterra-
nean Ridge. J. Geol. Soc. (Lond.) 154, 173–182.
de Lange, G.J., Brumsack, H.J., 1998. The occurrence of gas
hydrates in Eastern Mediterranean mud dome structures as in-
dicated by pore-water composition. Gas Hydrates: Relevance to
World Margin Stability and Climate Change. Spec. Publ.-Geol.
Soc., vol. 137, pp. 167–175.
DeLong, E.F., 1992. Archaea in coastal marine environments. Proc.
Natl. Acad. Sci. U. S. A. 89, 5685–5689.

J.P. Werne et al. / Chemical Geology 205 (2004) 367–390388
Dickens, G.R., O’Neil, J.R., Rea, D.K., Owen, R.M., 1995. Disso-
ciation of oceanic methane hydrate as a cause of the carbon
isotope excursion at the end of the Paleocene. Paleoceanography
10, 965–971.
Elvert, M., Suess, E., Greinert, J., Whiticar, M.J., 2000. Archaea
mediating anaerobic methane oxidation in deep-sea sediments at
cold seeps of the eastern Aleutian subduction zone. Org. Geo-
chem. 31, 1175–1187.
Elvert, M., Greinert, J., Suess, E., Whiticar, M., 2001. Carbon iso-
topes of biomarkers derived from methane-oxidizing microbes at
Hydrate Ridge, Cascadia convergent margin. In: Paull, C., Dil-
lon, W. (Eds.), Natural Gas Hydrates: Occurrence, Distribution,
and DetectionGeophys. Monogr., vol. 124. Am. Geophys. Un.,
pp. 115–129.
Emeis, K.C., Robertson, A.H.F., Richter, C., 1996. Proc. Ocean
Drill. Program, Initial Rep., vol. 160. Ocean Drilling Program,
College Station, TX. 972 pp.
Fiala-Medioni, A., Felbeck, H., 1990. Autotrophic process in inver-
tebrate nutrition: bacterial symbiosis in bivalve molluscs. In:
Kinne, R.K.H., Kinne-Saffran, E., Beyenbach, K.W. (Eds.), An-
imal Nutrition and Transport Processes: 1. Nutrition in Wild
and Domestics Animals, Comparative Physiology, vol. 5.
Karger, Basel, pp. 49–59.
Fiala-Medioni, A., Sibuet, M., Gottschal, J.C., Mariotti, A., 2001.
Chemosynthesis-based communities associated with fluids in
deep mediterranean mud-volcanoes. Proceedings, CIESM Con-
ference. Monte Carlo, Sept. 24–28.
Fiala-Medioni, A., McKiness, Z.P., Dando, P., Boulegue, J., Mari-
otti, A., Alayse-Danet, A.M., Robinson, J.J., Cavanaugh, C.M.,
2002. Ultrastructural, biochemical, and immunological charac-
terization of two populations of the mytilid mussel, Bathymo-
diolus azoricus, from the Mid-Atlantic Ridge. Evidence for a
dual symbiosis. Mar. Biol. 141, 1035–1043.
Fisher, C.R., 1995. Toward an appreciation of hydrothermal vent
animals: their environment, physiological ecology, and tissue
stable isotope values. Seafloor Hydrothermal Systems: Physical,
Chemical, Biological, and Geological Interactions. American
Geophysical Union, 297–316.
Haese, R.R., Meile, C., Van Cappellen, P., De Lange, G.J., 2003.
Carbon geochemistry of cold seeps: methane fluxes and trans-
formation in sediments from Kazan mud volcano, eastern Med-
iterranean Sea. Earth Planet. Sci. Lett. 212, 361–375.
Heijs, S.K., Jonkers, H.M., van Gemerden, H., Schaub, B.M., Stal,
L.J., 1999. The buffering capacity towards sulfide in sediments
of a coastal lagoon (Bassin D’Arcachon France): relative impor-
tance of chemical and biological processes. Estuar., Coast. Shelf
Sci. 49, 21–35.
Henriet, J.P., Mienert, J. (Eds.), 1998. Gas Hydrates: Relevance to
World Margin Stability and Climate Change. Special Publica-
tion-Geological Society, vol. 137. 338 pp.
Hinrichs, K.-U., Hayes, J.M., Sylva, S.P., Brewer, P.G., Delong,
E.F., 1999. Methane-consuming archaebacteria in marine sedi-
ments. Nature 398, 802–805.
Hinrichs, K.-U., Summons, R.E., Orphan, V., Sylva, S.P., Hayes,
J.M., 2000. Molecular and isotopic analysis of anaerobic meth-
ane-oxidizing communities in marine sediments. Org. Geochem.
31, 1685–1701.
Hopmans, E.C., Schouten, S., Pancost, R.D., van der Meer, M.T.J.,
Sinninghe Damste, J.S., 2000. Analysis of intact tetraether lipids
in archaeal cell material and sediments by high performance
liquid chromatography/atmospheric pressure chemical ioniza-
tion mass spectrometry. Rapid Commun. Mass Spectrom. 14,
585–589.
Hovland, M., 2000. Are there commercial deposits of marine
hydrates and ocean sediments?. Energy Explor. Exploit. 18,
339–347.
Kennicutt, M.C., Burke Jr., R.A., MacDonald, I.R., Brooks, J.M.,
Denoux, G.J., Macko, S.A., 1992. Stable isotope partitioning in
seep and vent organisms: chemical and ecological significance.
Chem. Geol. 101, 293–310.
Limonov, A.F., Woodside, J.M., Cita, M.B., Ivanov, M.K., 1996.
The Mediterranean Ridge and related mud diapirism: a back-
ground. Mar. Geol. 132, 7–20.
Luff, R., Wallmann, K., 2003. Fluid flow, methane fluxes, carbon-
ate precipitation and biogeochemical turnover in gas hydrate-
bearing sediments at Hydrate Ridge, Cascadia Margin: numer-
ical modeling and mass balances. Geochim. Cosmochim. Acta
67, 3403–3421.
Mascle, J., Huguen, C., Benkhelil, J., Chamot-Rooke, N., Chaumil-
lon, E., Foucher, J.-P., Griboulard, R., Kopf, A., Lamarche, G.,
Volkonskaia, A., et al., 1999. Images may show start of Euro-
pean–African plate collision. Eos, Transactions, American Geo-
physical Union, 80, 37, 421, 425, 428.
McClusky, S., Balassanian, S., Barka, A., Demir, C., Ergintav, S.,
Georgiev, I., Gurkan, O., Hamburger, M., Hurst, K., Kahle, H.,
Kastens, K., Kekelidze, G., King, R., Kotzev, V., Lenk, O.,
Mahmoud, S., Mishin, A., Nadariya, M., Ouzounis, A., Para-
dissis, D., Peter, Y., Prilepin, M., Reilinger, R., Sanli, I., Seeger,
H., Tealeb, A., Toksoz, M.N., Veis, G., 2000. Global Positioning
System constraints on plate kinematics and dynamics in the
eastern Mediterranean and Caucasus. J. Geophys. Res., B: Solid
Earth 105/3, 5695–5719.
McKenzie, D.P., 1972. Active tectonics of the Mediterranean re-
gion. Geophys. J. R. Astron. Soc. 30, 109–185.
MEDINAUT/MEDINETH Shipboard Scientific Parties, D.P., 2000.
Linking Mediterranean brine pools and mud volcanism. EOS,
Trans. Am. Geophys. Un. 81, 625–632.
Melieres, F., Chamley, H., Coumes, F., 1978. X-ray mineralogy
studies, Leg 42A, deep sea drilling project, Mediterranean
Sea. DSDP 42, 361–383.
Michaelis, W., Seifert, R., Nauhaus, K., Treude, T., Thiel, V., Blu-
menberg, M., Knittel, K., Gieseke, A., Peterknecht, K., Pape, T.,
Boetius, A., Amann, R., Jorgensen, B.B., Widdel, F., Peckmann,
J., Pimenov, N., Gulin, M., 2002. Microbial reefs in the Black
Sea fueled by anaerobic oxidation of methane. Science 297,
1013–1015.
Milkov, A.V., 2000. Worldwide distribution of submarine mud vol-
canoes and associated gas hydrates. Mar. Geol. 167, 29–42.
Nelson, D.C., Fisher, C.R., 1995. Chemoautotrophic and Methano-
trophic Endosymbiotic Bacteria at Deep-Sea Vents and Seeps.
In: Karl, D. (Ed.), Microbiology of Deep-Sea Hydrothermal
Vents. CRC Press, Boca, pp. 125–166.
Olu, K., Lance, S., Sibuet, M., Fiala-Medioni, A., Dinet, A., 1997.
Spatial distribution of cold seep communities as indicators of

J.P. Werne et al. / Chemical Geology 205 (2004) 367–390 389
fluid expulsion patterns through mud volcanoes on the Barbados
accretionary prism. Deep-Sea Res., Part 1, Oceanogr. Res. Pap.
44, 811–841.
Olu-Le Roy, K., Sibuet, M., Levitre, G., Gofas, S., Fiala-Medioni,
A., Vacelet, J., and the Medinaut scientific shipboard party (J.P.
Foucher, J.W., J. Mascle, G. De Lange, G. Aloisi, J.L. Charlou,
J.P. Donval, R. Haese, P. Henry, S. De Lint, M. van der Maarel,
G. Nobbe, C. Pierre, R. Pancost), 2001. Cold seep communities
in the deep Mediterranean Sea (South of Crete and Turkey):
faunal composition and spatial distribution. CIESM Congress
Proceedings. Monte Carlo, Sept. 24–28, pp. 36, 40.
Olu-Le Roy, K., Sibuet, M., Levitre, G., Gofas, S., Salas, C.,
Fiala-Medioni, A., Fouchet J.P., Woodside J., in press. Cold
seep communities in the deep eastern Mediterranean Sea:
composition and spatial distribution on mud volcanoes. Deep
Sea Research.
Orphan, V.J., House, C.H., Hinrichs, K.-U., McKeegan, K.D.,
DeLong, E.F., 2001. Methane-consuming archaea revealed by
directly coupled isotopic and phylogenetic analysis. Science
293, 484–487.
Pancost, R.D., Sinninghe Damste, J.S., 2003. Carbon isotopic com-
positions of prokaryotic lipids as tracers of carbon cycling in
diverse settings. Chem. Geol. 195, 29–58.
Pancost, R.D., Sinninghe Damste, J.S., de Lint, S., van der Maarel,
M.J.E.C., Gottschal, J.C., and the MEDINAUT Shipboard Sci-
entific Party, 2000. Biomarker evidence for widespread anaerobic
methane oxidation in Mediterranean sediments by a consortium
of methanogenic archaea and bacteria. Appl. Environ. Microbiol.
66, 1126–1132.
Pancost, R.D., Hopmans, E.C., Sinninghe Damste, J.S., and the
MEDINAUT Shipboard Scientific Party, 2001. Archaeal lip-
ids in Mediterranean cold seeps: molecular proxies for an-
aerobic methane oxidation. Geochim. Cosmochim. Acta 65,
1611–1627.
Pancost, R.D., Bouloubassi, I., Aloisi, G., Sinninghe Damste, J.S.,
and the MEDINAUT Shipboard Scientific Party, 2001. Three
series of non-isoprenoidal dialkyl glycerol diethers in cold-seep
carbonate crusts. Org. Geochem. 32, 695–707.
Papazachos, B.C., Comninakis P.E., 1978. Deep structures and
tectonics of the Eastern Mediterranean. Tectonophysics 45,
285–296.
Paull, C.K., Lorenson, T.D., Dickens, G., Borowski, W.S., Ussler
III, W., Kvenvolden, K.A., 2000. Comparison of in situ and
core gas measurements in ODP Leg 164 bore holes. In: Hol-
der, G.D., Bishnoi, P.R. (Eds.), Gas Hydrates: Challenges for
the Future. Annals of the New York Academy of Sciences,
vol. 912, pp. 23–31.
Pierre, C., Rouchy, J.M., Gaudichet, A., 2000. Diagenesis in the gas
hydrate sediments of the Blake Ridge: mineralogy and stable
isotope compositions of the carbonate and sulphide minerals. In:
Paull, C.K., Matsumoto, R., Wallace, P.J. (Eds.), Proc. Ocean
Drill. Program Sci. Results, vol. 164, pp. 139–146.
Raiswell, R., 1988. Chemical model for the origin of minor lime-
stone-shale cycles by anaerobic methane oxidation. Geology 16,
41–644.
Reeburgh, W.S., 1976. Methane consumption in Cariaco Trench
waters and sediments. Earth Planet. Sci. Lett. 28, 337–344.
Reeburgh, W.S., Whalen, S.C., Alperin, M.J., 1993. The role of
methylotrophy in the global methane budget. In: Murrell, J.C.,
Kelley, D.P. (Eds.), Microbial Growth on C-1 Compounds. In-
tercept, Andover, UK, pp. 1–14.
Reilinger, R.E., McClusky, S.C., Oral, M.B., King, R.W., Toksoz,
M.N., 1997. Global positioning system measurements of pres-
ent-day crustal movements in the Arabia –Africa –Eurasia
plate collision zone. J. Geophys. Res., B: Solid Earth 102,
9983–9999.
Rempel, A.W., Buffett, B.A., 1997. Formation and accumulation
of gas hydrate in porous media. J. Geophys. Res. 102,
10151–10164.
Ritger, S., Carson, B., Suess, E., 1987. Methane-derived authi-
genic carbonates formed by subduction-induced pore-water ex-
pulsion along the Oregon/Washington margin. GSA Bull. 98,
147–156.
Robertson, A.H.F., 1998. Tectonic significance of the Eratosthenes
Seamount: a continental fragment in the process of collision
with a subduction zone in the eastern Mediterranean (Ocean
Drilling Program Leg 160). Tectonophysics 298, 63–82.
Robertson, A.H.F., Emeis, K.C., Carmenlenghi, A. (Eds.), 1998.
Proceedings ODP, Scientific Results TX (Ocean Drilling Pro-
gram), College station. 817 pp.
Rotstein, Y., Kafka, A.L., 1982. Seismotectonics of the southern
boundary of Anatolia, eastern Mediterranean region: subduction,
collision and arc jumping. J. Geophys. Res., B 87, 7694–7706.
Ryan, W.B.F., Hsu, K.J., et al., 1973. Initial Report of the Deep-Sea
Drilling Project, Leg XIII. U.S. Government Printing Office,
Washington.
Salas, C., Woodside, J., 2002. Lucinoma kazani n.sp. (Mollusca:
bivalvia): evidence of a living benthic community associated
with a cold seep in the Eastern Mediterranean Sea. Deep-Sea
Res., Part 1, Oceanogr. Res. Pap. 49, 991–1005.
Sassen, R., Roberts, H.H., Aharon, A., Larkin, J., Chinn, E.W.,
Carney, R., 1993. Chemosynthetic bacterial mats at cold hydro-
carbon seeps, Gulf of Mexico continental slope. Org. Geochem.
20, 77–89.
Schouten, S., Hopmans, E.C., Pancost, R.D., Sinninghe Damste,
J.S., 2000. Widespread occurrence of structurally diverse tet-
raether membrane lipids: evidence for the ubiquitous presence
of low-temperature relatives of hyperthermophiles. Proc. Natl.
Acad. Sci. 97, 14421–14426.
Schouten, S., Wakeham, S.G., Sinninghe Damste, J.S., 2001. An-
aerobic methane oxidation by archaea in the euxinic water col-
umn of the Black Sea. Org. Geochem. 32, 1277–1281.
Sibuet, M., Olu, K., 1998. Biogeography, biodiversity and fluid
dependence of deeps cold seep communities at active and pas-
sive margins. Deep-Sea Res., Part 2, Top. Stud. Oceanogr. 45,
517–567.
Sørensen, K.B., Finster, K., Ramsing, N.B., 2001. Thermodynamic
and kinetic requirements in anaerobic methane oxidizing con-
sortia exclude hydrogen, acetate, and methanol as possible elec-
tron shuttles. Microb. Ecol. 42, 1–10.
Spakman, W., Wortel, M.J.R., Vlaar, N.J., 1988. The Hellenic sub-
duction zone: a tomographic image and its geodynamic impli-
cations. Geophys. Res. Lett. 15, 60–63.
Stakes, D.S., Orange, D., Paduan, J.B., Salamy, K.A., Maher, N.,

J.P. Werne et al. / Chemical Geology 205 (2004) 367–390390
1999. Cold-seeps and authigenic carbonate formation in Mon-
terey Bay, California. Mar. Geol. 159, 93–109.
ten Veen, J.H., Woodside, J.M., Zitter, T.A.C., Dumont, J.F.,
Mascle, J., Volkonskaya, A., in press, Neotectonic evolution
of the Anaximander Mountains at the junction of the Hellenic
and Cyprus arcs. Tectonophysics.
Thiel, V., Peckmann, J., Seifert, R., Wehrung, P., Reitner, J.,
Michaelis, W., 1999. Highly isotopically depleted isoprenoids:
molecular markers for ancient methane venting. Geochim. Cos-
mochim. Acta 63, 3959–3966.
Thiel, V., Peckmann, J., Richnow, H.H., Luth, U., Rietner, J.,
Michaelis, W., 2001. Molecular signals for anaerobic methane
oxidation in Black Sea seep carbonates and a microbial mat.
Mar. Chem. 73, 97–112.
Valentine, D.L., Reeburgh, W.S., 2000. New perspectives on anaer-
obic methane oxidation. Environ. Microbiol. 2, 477–484.
Volgin, L., Woodside, J., 1996. Sidescan sonar images of mud
volcanoes from the Mediterranean Ridge: possible causes of
variations in backscatter intensity. Mar. Geol. 132, 39–53.
von Cosel, R., Salas, C., 2001. Vesicomyidae (Mollusca: Bivalvia)
of the genera Vesicomya, Waisiuconcha, Isorrpodon and Callo-
gonia in the eastern Atlantic and the Mediterranean. Sarsia 86,
333–366.
Wallmann, K., Lionke, P., Suess, E., Bohrmann, G., Sahling, H.,
Schluter, M., Dahlmann, A., Lammers, S., Greinert, J., von
Mirbach, N., 1997. Quantifying fluid flow, solute mixing,
and biogeochemical turnover at cold vents of the eastern
Aleutian subduction zone. Geochim. Cosmochim. Acta 61,
5209–5219.
Werne, J.P., Baas, M., Sinninghe Damste, J.S., 2002. Molecular
isotopic tracing of carbon flow and trophic relationships in a
methane-supported benthic microbial community. Limnol. Oce-
anogr. 47, 1694–1701.
Whiticar, M.J., Faber, E., 1986. Methane oxidation in sediment and
water column environments—Isotope evidence. Org. Geochem.
10, 759–768.
Whiticar, M.J., 1999. Carbon and hydrogen isotope systematics of
bacterial formation and oxidation of methane. Chem. Geol. 161,
291–314.
Werne, J.P., Baas, M., Sinninghe Damste, J.S., 2002. Molecular
isotopic tracing of carbon flow and trophic relationships in a
methane-supported benthic microbial community. Limnol. Oce-
anogr. 47, 1694–1701.
Whiticar, M.J., 1999. Carbon and hydrogen isotope systematics of
bacterial formation and oxidation of methane. Chem. Geol. 161,
291–314.
Woodside, J.M., 1977. Tectonic element and crust of the eastern
Mediterranean. Mar. Geophys. Res. 3, 317–354.
Woodside, J.M., Ivanov, M.K., Limonov, A.F., 1997. Neotectonics
and fluid flow through seafloor sediments in the Eastern Med-
iterranean and Black Seas: Part I. Eastern Mediterranean
SeaIOC Technical Series (Intergovernmental Oceanographic
Commission, Unesco) UNESCO 1997, Paris, International,
vol. 48. 128 pp.
Woodside, J.M., Ivanov, M.K., Limonov, A.F.Anaxiprobe Ship-
board Scientifis Parties, 1998. Shallow gas and gas hydrates
in the Anaximander Mountains region, eastern Mediterranean
Sea. In: Henriet, J.-P., Mienert, J. (Eds.), Gas Hydrates: Rele-
vance to World Margin Stability and Climate ChangeGeological
Society Special, vol. 137, pp. 177–193.
Woodside, J., Mascle, J., Huguen, C., Volkonskaia, A., 2000. The
Rhodes Basin, a post-Miocene tectonic trough. Mar. Geol. 165,
1–12.
Woodside, J.M., Mascle, J., Zitter, T.A.C., Limonov, A.F., Ergun,
M., Volkonskaia, A., 2002. The Florence Rise, the Western Bend
of the Cyprus Arc. Mar. Geol. 185, 177–194.
Zatsepina, O.Y., Buffett, B.A., 1997. Phase equilibrium of gas hy-
drate: implications for the formation of hydrate in the deep sea
floor. Geophys. Res. Lett. 24, 1567–1570.
Zitter, T.A.C., Woodside, J.M., Mascle, J., 2003. The Anaximander
Mountains: a clue to the tectonics of Southwest Anatolia. Geol.
J. 38, 375–394.





























