Embed Size (px)
Citation preview

LGN Activity Patterns during Ocular Dominance Plasticity in vivo
By
Monica L. Linden
S.B. Mathematics with Computer Science,
S.B. Brain and Cognitive Sciences
Massachusetts Institute of Technology, 2002
SUBMITTED TO THE DEPARTMENT OF BRAIN AND COGNITIVE SCIENCES IN
PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF
DOCTOR OF PHILOSOPHY IN NEUROSCIENCE
AT THE
MASSACHUSETTS INSTITUTE OF TECHNOLOGY
JUNE 2008
© Massachusetts Institute of Technology
All rights reserved
Signature of Author: ____________________________________________________________
Department of Brain and Cognitive Sciences
May 13, 2008
Certified by: ___________________________________________________________________
Mark F. Bear, Ph.D.
Picower Professor of Neuroscience
Thesis Supervisor
Accepted by: __________________________________________________________________
Matthew A. Wilson, Ph.D.
Professor of Neurobiology
Chairman, Committee for Graduate Students


LGN Activity Patterns during Ocular Dominance Plasticity in vivo
By
Monica L. Linden
Submitted to the Department of Brain and Cognitive Sciences
on May 13, 2008 in Partial Fulfillment of the Requirements
for the Degree of Doctor of Philosophy in Neuroscience
ABSTRACT
Perturbations of sensory experience in young animals are known to cause lasting changes in
adult brain function. For example, monocular visual deprivation by lid closure (MC) leads to a
loss of cortical responsiveness of the deprived eye and a concomitant visual impairment. This
ocular dominance (OD) plasticity is a well-studied model of experience-dependent cortical
plasticity. While much is known about the anatomical, physiological and biochemical changes
that occur in primary visual cortex following OD plasticity, the input patterns that lead to these
changes have not been characterized.
Visual input travels from the retina through the dorsal lateral geniculate nucleus (dLGN) of the
thalamus and then into visual cortex. Several models of the thalamic activity patterns which
drive OD plasticity have been proposed, but the assumptions about the pattern and amount of
input activity from thalamus to cortex during deprivation have not been experimentally
validated. Therefore, we performed extracellular recordings from the dLGN of animals during
periods of visual manipulation.
Contrary to previous hypotheses, the present findings demonstrate that MC does not alter the
overall firing rate of neural activity in the dLGN. Instead, MC alters the pattern of neural spike
trains such that there is a decrease in simultaneous firing of neighboring neurons. Moreover, the
elimination of visual input from the retina, a form of deprivation which does not lead to
deprived-eye depression, leads to a dramatic increase in thalamic bursting. Additionally, there
are subtle qualitative differences between dLGN activity in juveniles and adults during MC, and
this may contribute to differences in OD plasticity with age. These findings substantially alter
the interpretation of previous studies and define the activity patterns that govern cortical
plasticity in vivo. Furthermore, this work may have important implications for treatments of
developmental disorders including ambylopia.
Thesis Supervisor: Mark F. Bear, Ph.D.
Title: Picower Professor of Neuroscience


Monica L. Linden [email protected]
Education: Massachusetts Institute of Technology Doctor of Philosophy – June, 2008
Cambridge, MA
Ph.D. in Neuroscience, Department of Brain and Cognitive Sciences.
Major: Systems Neuroscience, Minor: Computational Neuroscience.
Massachusetts Institute of Technology Bachelor of Science – June, 2002
Cambridge, MA
Bachelor of Science degrees in Mathematics with Computer Science and Brain and
Cognitive Sciences, Minor: Ancient and Medieval Studies.
Awards/
Honors: NIH National Research Service Award Predoctoral Fellowship (2007)
Walle Nauta Award for Continuing Dedication to Teaching (2006)
Dean’s Educational and Student Advising Award (2005)
Agnus MacDonald Award for Excellence in Undergraduate Teaching (2005)
National Science Foundation Graduate Research Fellowship (2002)
MIT Brain and Cognitive Sciences Award for Academic Excellence (2002)
Phi Beta Kappa Honor Society
Sigma Xi Research Honor Society
Teaching
Experience: MIT 9.01: Introduction to Neuroscience – Head TA Fall 2004, Fall 2005
Taught one weekly section (~25 students) under Profs. Mark Bear and Sebastian Seung.
Coordinated TAs, multiple guest lecturers, and the writing staff.
MIT 9.00: Introduction to Psychology – TA Fall 2003
Taught two recitation sections (~25 students) under Prof. Jeremy Wolfe.
MIT Orientation for Graduate Teaching Staff – Workshop Facilitator
Fall 2005
Co-facilitated “Problems, Pitfalls, Booby Traps and Surprises (in Teaching)” workshop
during the fall TA orientation, sponsored by the Teaching and Learning Lab.
Research
Experience: MIT Picower Center for Learning and Memory – Bear Lab
Cambridge, MA June, 2003 – 2008
Graduate Student Researcher: Performing in vivo electrophysiology in awake rodents to
understand various aspects of neural plasticity in the visual system.
National Security Agency – Students' Summer Program Government Communications Headquarters Cheltenham, UK
Summer, 2002
Intern: One of two student liaisons selected to represent the US at this British security
agency. Developed mathematical models to analyze systems. Prepared a research report
for internal publication and presented results to internal mathematics community.

MIT Picower Center for Learning and Memory – Tonegawa Lab
Cambridge, MA September, 2001 – March, 2002
Undergraduate Researcher: Data analysis for electrophysiological studies in genetically
manipulated mice.
National Security Agency – Director's Summer Program
Ft. Meade, MD Summer, 2001
Intern: Developed mathematical models to analyze and evaluate data. Prepared a
research report for internal publication and briefed the Director of the Agency. Top
Secret Clearance.
Consulting: Lippincott Williams and Wilkin – Subject Matter Expert Fall 2005
Reviewed manuscripts of ancillary material, specifically “Student Assignments and
Activities” for Neuroscience, Exploring the Brain, 3rd Edition by Bear, Connors and
Paradiso. This is the textbook used in 9.01 – Introduction to Neuroscience, co-authored
by Prof. Bear.
Publications &
Presentations: M.L. Linden, A.J. Heynen, R. Haslinger, M.F. Bear. (2008) Thalamic activity that drives
visual cortical plasticity. Submitted
M.L. Linden, A.J. Heynen, R. Haslinger, E.N. Brown, M.F. Bear. LGN activity patterns
during ocular dominance plasticity. Picower-RIKEN Symposium, November 8 – 9, 2007.
Massachusetts Institute of Technology, Cambridge, MA. Poster Presentation.
M.L. Linden, A.J. Heynen, R. Haslinger, E. N. Brown, M. F. Bear. LGN activity
patterns during ocular dominance plasticity. Society for Neuroscience, Abstracts,
November 3 – 7, 2007. Society for Neuroscience Annual Meeting, San Diego, CA.
Poster Presentation.
M.L. Linden. LGN activity patterns during ocular dominance plasticity. MIT Brain
Lunch Seminar Series, October 15, 2007. Massachusetts Institute of Technology,
Cambridge, MA. Lecture.
M.L. Linden. LGN activity patterns during ocular dominance plasticity. Picower
Institute for Learning and Memory, Annual Retreat, May 30 – June 1, 2007. Cape Cod,
MA. Lecture.
M.G. Shuler, M.L.Linden, M.Bear. (2004) Learning induced, persistent reward-related
activity in the primary visual cortex of adult rodents. Society for Neuroscience,
Abstracts, October 23 – 27, 2004. Society for Neuroscience Annual Meeting, San Diego,
CA.

Acknowledgments
“We were but stones, your light made us stars”
–Low Light, Pearl Jam
There are numerous people whom I’d like to thank, without which my thesis project would not
have been as successful. First of all, I’d like to thank Mark Bear. I thoroughly enjoyed working on the
fundamental problem which he gave to me, and I was constantly driven to work through my frustrations
because I knew there had to be answers. I appreciate all his support throughout the project, even after I
upended his hypotheses. I would also like to thank the rest of my thesis committee for their helpful
insights: Mriganka Sur, Matthew Wilson and especially Harvey Swadlow, who responded patiently to my
emails even when I sent several in the same day. I’d also like to acknowledge my funding sources
including a graduate research fellowship from NSF and an NRSA predoctoral fellowship from
NINDS/NIH.
The Bear Lab provided an incredibly supportive environment throughout my graduate career.
Arnie Heynen was a constant source of scientific expertise and emotional support. I couldn’t have done
this without him. Marshall Shuler patiently taught me how to be an electrophysiologist and was always
there to help me think through my ideas and to challenge my assumptions. Jason Coleman was always
willing to help me and provided invaluable knowledge of the dLGN. Sharing lab space with Jonathan
Whitlock meant my day in lab would be much more fun. Suzanne Meagher always took care of me, and
Erik Sklar fixed everything I managed to break. Our technicians, Caroline Dudley, Cindy Poo, Tina
Udaka, and especially Kathleen Oram who did my histology, were wonderful. Gordon Smith always had
great answers to questions I asked him, even when they had nothing to do with what he was studying, and
even when I asked him quietly during lab meeting. Misha Frenkel, Gül Dölen, Sam Cooke and Emily
Osterweil were great sounding-boards for discussing my problems. Lena Khibnik, Rahmat Muhammad,
and Wendy Chen were all wonderful friends inside and outside the lab, and really helped me to get
through these past few years. I love you, ladies! And I’d like to thank the rest of the Bear lab members
with whom I’ve worked – all of you have had a positive impact on my work and my graduate school
experience.
I’d also like to thank my collaborator, Rob Haslinger, for his assistance with the data analysis,
Brian Blais and Harel Shouval for their insight and assistance interpreting my results, and Mike Long for
helpful conversations regarding my project.
I have to thank Pearl Jam for the 7 concerts I attended in graduate school, and the countless hours
I spent listening to their music during surgeries, experiments, and data analysis.
I couldn’t have accomplished this without the support of my friends. I’d like to thank Gabrielle
Cayton, Wendy Chen, and Sumiko Mekaru for being my fabulous roommates, and their cats Coco, Juliet
and Cobweb, who provided much-needed affection after many stressful days in lab. I’d also like to thank
Melissa Thirer, who has known me my entire life, and has always been there to support me, and Kevin
Matulef for going through all the ups and downs of graduate school with me. In addition to the rest of my
friends in Boston, I’d like to thank my long-distance friends, especially Naomi Eckmann, Shantonu Sen,
Seth Bisen-Hersh, Brian Bucklew, Michael Korn, Dan Winters and Craig Citro, who are never more than
an IM message, email, or phone call away.
Finally, I’d like to thank my family who has always supported whatever I’ve chosen to do. My
grandmothers, Florence and Josette, are a constant source of inspiration and unconditional love. And my
Mom is always there for me, no matter what. I love you all. I’d like to dedicate this thesis to my Dad,
who always knew I could accomplish this, even from the time when my greatest scientific achievement
was comparing fresh squeezed and concentrated orange juice.


Table of Contents
Chapter 1 ..................................................................................................................................... 11
Introduction ............................................................................................................................... 13
dLGN Anatomy ......................................................................................................................... 14
dLGN Physiology ...................................................................................................................... 19
Ocular Dominance Plasticity ..................................................................................................... 29
References ................................................................................................................................. 41
Summary ................................................................................................................................... 50
Chapter 2 ..................................................................................................................................... 51
Abstract ..................................................................................................................................... 53
Introduction ............................................................................................................................... 54
Methods Summary .................................................................................................................... 55
Results ....................................................................................................................................... 56
Discussion ................................................................................................................................. 59
Detailed Methods ...................................................................................................................... 85
References ................................................................................................................................. 89
Chapter 3 ..................................................................................................................................... 91
Abstract ..................................................................................................................................... 93
Introduction ............................................................................................................................... 94
Methods Summary .................................................................................................................... 95
Results ....................................................................................................................................... 96
Discussion ................................................................................................................................. 98
Detailed Methods .................................................................................................................... 113
References ............................................................................................................................... 116
Chapter 4 ................................................................................................................................... 119
Methodological Considerations ............................................................................................... 121
Implications ............................................................................................................................. 125
Future Experiments ................................................................................................................. 131
References ............................................................................................................................... 141
Appendix .................................................................................................................................... 145
Appendix 1 .............................................................................................................................. 147
Appendix 2 .............................................................................................................................. 150
References ............................................................................................................................... 155


11
Chapter 1
INTRODUCTION AND BACKGROUND


13
Introduction
The dorsal lateral geniculate nucleus (dLGN) of the thalamus transmits visual
information from the retina to the visual cortex allowing for visual perception and serving the
“what” and “where” streams of visual information processing. While textbooks often define
thalamic nuclei as mere relay stations between the periphery and the cortex, recent evidence
suggests that important processing takes place within the thalamus (reviewed in Sherman, 2007).
Because the dLGN acts as the gateway into visual cortex, it is important to understand not only
the anatomy and circuitry of this brain area, but also how the dLGN transforms and transmits
retinal information en route to the visual cortex. Therefore, we will begin by reviewing both
dLGN circuitry and dLGN activity. We will draw from studies conducted on several different
species including cats, ferrets, rats and mice. Although the mouse served as the animal model
for the experiments described in the body of this thesis, many of the findings on which the study
is based were discovered in other species (e.g. Rittenhouse et al., 1999). Furthermore, the
literature on the mouse dLGN is not comprehensive, and by including the literature from other
species and assuming that the basic principles are preserved, we can better understand the system
under investigation.
Our interests lie in understanding the role of dLGN input to the visual cortex during
visual manipulations that induce experience-dependent plasticity. Therefore, in addition to
understanding the dLGN, this literature survey will discuss monocular deprivation and the
resulting ocular dominance plasticity, which will serve as our paradigm for exploring
experience-dependent plasticity in the cortex.

14
dLGN Anatomy
The dLGN is contained within the dorsal thalamus, the largest part of the mammalian
diencephalon. Developing from the region between the epithalamus and ventral thalamus, all
dorsal thalamic nuclei have relay cells that project to a region of neocortex in the telencephalon
(Sherman and Guillery, 2006). Specifically, the dLGN is a first-order thalamic relay transmitting
visual information from both retinae to the visual cortex ipsilateral to the nucleus.
The processing that occurs in any region of the brain is contingent on both the inputs to
that brain area and the connections and circuits within the area. In order to understand the
processing that occurs in the dLGN, we must begin by exploring the structural properties of this
brain region. There are two main categories of neurons in the dLGN: relay cells and
interneurons. Rodents lack interneurons in some thalamic nuclei including the ventrobasal
complex and the ventrolateral nucleus (Arcelli et al., 1997). However, using Golgi staining
techniques and electron microscopy, two types of neurons were identified in the mouse dLGN
(Rafols and Valverde, 1973). The larger category of neurons was later identified as the
excitatory relay cells, with the second category, identified as inhibitory interneurons, comprising
15 – 20% of the neuronal population (Fig. 1-1a) (Arcelli et al., 1997).
In several species it is possible to further classify relay cells based on properties including
cell size, axonal or dendritic arbor properties, axon diameter and/or conduction velocity, and
receptive field (RF) properties. Using physiological classifications originally established in the
retina (Enroth-Cugell and Robson, 1966) and later extended to the dLGN (Cleland et al., 1971)
and morphological descriptions established through Golgi techniques (Guillery, 1966), LeVay
and Ferster (1977) provided the first description of morphologically and functionally distinct
relay cell types in the cat. By combining electrophysiological recordings with horseradish

15
peroxidase labeling, Friedlander et al. (1979) directly compared cell function with morphology,
later refining the criteria to segregate cat relay cells into W-, X- and Y-cell categories based on
both morphological and electrophysiological properties (Friedlander et al., 1981).
X-cells have smaller cell bodies than Y-cells, with elongated dendritic trees that remain
in a single dLGN layer. Y-cells have spherical dendritic arbors that can cross between layers.
With thicker dendrites, Y-cells have faster conduction velocities than X-cells. Physiologically,
the larger dendritic arbors of Y-cells lead to larger RFs. Additionally, Y-cells integrate inputs in
a nonlinear fashion, whereas X-cells spatially summate linearly. W-cells are distinguished from
X- and Y-cells based on their location within dLGN (Friedlander et al., 1981).
W-, X- and Y-cell types (and their primate equivalents of konicellular, parvocellular, and
magnocellular cell types, respectively) have been identified in the dLGN of numerous species
including two rodents: the squirrel (Van Hooser et al., 2003) and the rat both morphologically
(Brauer et al., 1979) and physiologically (Lennie and Perry, 1981; Gabriel et al., 1996).
However, there are no studies in the mouse to indicate the presence of distinct relay cell types.
Additionally, electrophysiological recordings from the mouse have shown no indication of
nonlinearly summing Y-cells (Grubb and Thompson, 2003). While the lack of evidence for Y-
cells does not imply that they are not present in the dLGN of mice, they may have a more
homogeneous dLGN than other “higher-order” species.
Another anatomical feature of the dLGN is its layered structure. These layers can be
distinguished by several properties including relay cell type, on- versus off-center RFs and the
eye of origin contributing retinal input. The functional role of the layers is unknown. In fact,
different species show a huge variety in the types of properties which distinguish the layers. For
example, X- and Y-cell types are intermingled in the A and A1 laminae of the cat dLGN, but

16
segregate into distinct layers in the squirrel. This may imply that there is no functional advantage
to the layering (Sherman and Guillery, 2006). Instead, it appears that the properties of the
different layers emerge entirely from the properties of the retinal input (Fukuda and Stone, 1974;
Sur et al., 1987). Additionally, interneurons can modulate input between layers (Sherman and
Spear, 1982) and intracellular recordings of dLGN neurons have shown that the large majority of
cells respond to input presented to either eye (Zhou et al., 2003) , thus layers do not serve as
distinct processing areas within the dLGN. Another indication that the layers may be
unimportant arises from the observation that cortical layers mix the input that remains distinct in
dLGN, for example eye of origin or X- and Y-cells.
Rats and mice do not appear to have layers based on relay cell type or on/off RF
structure, however a “hidden lamination” exists within the rat (Reese, 1988) and mouse dLGN
(Godement et al., 1984; Muir-Robinson et al., 2002; Jaubert-Miazza et al., 2005) with the inputs
from the two eyes remaining separate (Fig. 1-1). The outer “shell” of the dLGN receives
exclusive input from the contralateral eye, surrounding an inner “core” of ipsilateral input. The
segregation of eye-specific input into dLGN has been shown to be an activity dependent,
developmental process in cats and ferrets (Shatz, 1996; Weliky and Katz, 1999; Crowley and
Katz, 2002), as well as in mice (Jaubert-Miazza et al., 2005; Hooks and Chen, 2006).
In order to understand the processing that occurs in the dLGN it is also necessary to
discuss the input to the circuit. Thalamic afferents can be divided into two categories: drivers
and modulators. Drivers determine the qualitative RF properties of the relay cells, thus the
retinal ganglion cell (RGC) input serves as the driving input to dLGN, imbuing dLGN neurons
with RFs responsive to one eye in a center-surround configuration. While the driving input is
most important in determining the response properties of the thalamic relay cells, they comprise

17
the minority of synapses (Van Horn et al., 2000). The majority are modulators which affect the
quantitative aspects of the relay cell response including overall firing rate and size and strength
of the center and surround. Geniculate relay cells receive modulating input from several sources
including corticothalamic feedback, local interneurons, the thalamic reticular nucleus (TRN), and
brainstem.
The driving input to thalamic nuclei defines the information that will pass from the driver
to the cortex. The visual cortex is visual precisely because driving input from the retina passes
through the dLGN to this region of the brain. Rewiring a driver from a different modality into a
thalamic nucleus leads to neurons in cortex with RFs reflecting the driving input and can lead to
behaviors dependent on the new input. For example, rewiring visual input into the auditory
thalamus results in visual RFs in auditory thalamus and cortex and can result in visually driven
behaviors dependent on this rewired pathway (Sur et al., 1988; Sharma et al., 2000; von
Melchner et al., 2000).
The synapses formed by the drivers support the observation that they provide the
strongest input to the dLGN. Originally described by Guillery (1966), these large synapses are
localized to the terminal zones, and exhibit closely packed boutons. These findings were
confirmed by electron microscopy (EM) which shows that the vesicles at the RGC synapses are
round and contained in large synapses (RL synapses) (Wilson et al., 1984).
Each modulatory input to the dLGN has a unique EM profile. The corticothalamic
feedback, arising from layer VI of visual cortex (Gilbert and Kelly, 1975), forms short terminals
on the side branches of dLGN dendrites (Guillery, 1966). The synapses are small and contain
round vesicles (RS) (Erisir et al., 1997a). This feedback is topographic (Murphy and Sillito,
1996). Eliminating cortical input has been shown to have a small effect on dLGN neurons,

18
affecting synchrony between neurons and also the surround antagonism (Sillito et al., 1994;
Cudeiro and Sillito, 1996). However, the effect of corticothalamic feedback may be stronger in
an awake preparation (Sherman and Guillery, 2006).
The TRN sends γ-aminobutyric acid (GABA)-ergic afferents to dLGN (Houser et al.,
1980). In the rat, the terminals form grape-like boutons (Pinault et al., 1995). In the cat, EM
studies show the synapses are symmetric, and contain flattened vesicles (F1 synapses) (Wang et
al., 2001). These inputs are topographic, and may be important during sleep and for modulating
attentional states (Wang et al., 2001).
The local interneurons within the dLGN are also GABA-ergic and also display F1
synapses along their axons (Montero, 1987). However, the distal dendrites of local interneurons
also form synapses with the relay cells, and these synapses are characterized by their F2 profile
(Montero, 1986). F2 synapses have a more random distribution of vesicles than found at F1
terminals.
There are several additional sources of modulatory input to the dLGN including
cholinergic, serotonergic, dopamenergic and noradrenergic inputs from the brainstem and
hypothalamus. Cholinergic input accounts for 30% of the input into the dLGN (Erisir et al.,
1997b; Erisir et al., 1997a). This input forms RS terminals close to the relay cell bodies (Erisir et
al., 1997b).
While relay cells do receive input from all of these sources, some synapses within the
dLGN form a specialized architecture known as a “triad” (reviewed in Sherman, 2004). These
triads are sites where 3 or more synaptic profiles are found very close together, and the synapses
are contained within a glomerulus. A glomerulus is a site free of astrocyctic cytoplasm and
enclosed by an astrocytic sheath (Szentagothai, 1963; Sherman and Guillery, 2006). A triad

19
forms when an RL driving synapse is presynaptic to two sites: (1) a relay cell dendrite and (2) an
interneuron dendrite at an F2 terminal. This F2 terminal on the interneuron is in turn,
presynaptic to the same relay cell dendrite as (1). While corticothalamic feedback and TRN
innervation do not generally occur within glomeruli, cholinergic input is often found there (Erisir
et al., 1997b). This anatomy immediately suggests that the input from the RGCs is modified by
the dLGN, as the relay cell receiving this input is inhibited by an interneuron receiving the same
input, one synapse later. However, the protection of the glial sheath and the absence of
astrocytes at the synapse may allow neurotransmitters to move more freely at these synapses,
rendering them more faithful to their input (Sherman and Guillery, 2006). Furthermore, the
cholinergic input is able to directly modify this retinogeniculate transmission.
Triads exist in the dLGN of several species including cats and rats. While it appears that
only X-cell types are found in cat triads, this may not be the case in rat triads (Lam et al., 2005).
Similar studies have not yet been conducted in the mouse, although early studies using electron
microscopy in the mouse dLGN suggest triads may exist in this species (Rafols and Valverde,
1973).
dLGN Physiology
In addition to understanding the anatomy of dLGN circutiry, it is important to understand
the physiological properties of geniculate neurons and how they transform messages from the
retina to V1. Grubb and Thompson (2003) have characterized the basic response properties of
dLGN neurons in the mouse. Neurons in the mouse dLGN have circular RFs with the classical
center-surround structure found in higher mammals. The RFs have strongly responsive central
regions with diameters of 4 – 20° and weaker surrounds. The median peak spatial frequency for
mouse dLGN neurons is 0.03 cycles/degree, and the mean peak temporal frequency is

20
approximately 4 Hz. While the spatial resolution of the mouse dLGN is much lower than that of
more visual species, the peak temporal frequency is comparable to many species including
humans. Mouse geniculate neurons predominantly sum spatial input in a linear fashion
indicating that mouse dLGN neurons are X-like (Grubb and Thompson, 2003). There is no clear
transformation of the RGC RF properties in the dLGN. This suggests that the role of the dLGN
is to gate and/or modulate retinal input (Sherman and Guillery, 2006).
Retinogeniculate Transmission
Even though the dLGN does not change the basic RF properties transmitted from the
retina, it is possible for the dLGN to transform the message sent from the retina to V1 by
modulating the spike train. Theoretically, there are several ways that the dLGN can change the
retinal input. For example, the spike train originating in the RGCs could have spikes added or
removed, or the temporal patterning of the spikes could be modified before the message is sent to
cortex.
The first location where the visual signal leaving the retina can be altered is at the
retinogeniculate synapse. Simultaneous recordings from RGCs and dLGN neurons have allowed
for studies of the efficacy of the retinogeniculate pathway. The efficacy of retinogeniculate
transmission is defined as the probability that an RGC spike will evoke a spike in a
monosynaptically connected dLGN neuron. dLGN neurons fire approximately 5 ms after RGC
spiking in the anesthetized cat preparation (Mastronarde, 1987). The efficacy of synaptic
transmission from RGC to dLGN is relatively low (approximately 3%) (Usrey et al., 1998).
However, the efficacy is enhanced by short preceding interspike intervals (ISIs) of 3 – 30 ms
(Mastronarde, 1987; Usrey et al., 1998; Rowe and Fischer, 2001). The length of the ISI that will
still result in increased efficacy of the second spike is modulated by the overall firing rate of the

21
RGC (Rowe and Fischer, 2001). Levine and Cleland (2001) further described the efficacy of this
synapse by classifying all recorded dLGN spikes into those evoked from RGC stimuli, and those
that arose from an “anonymous” (non-retinal) source, while also considering if the recorded
RGC spikes were effective at driving the dLGN. They confirmed that short preceding ISIs
enhance the efficacy of RGC transmission, with efficacy dropping off exponentially with ISI
length. They also found that very short ISIs from the RGCs (< 3ms) lead to a refractory period
for dLGN firing. Additionally, if the ISI between a failed RGC input and an anonymous input is
short, the anonymous input will be more likely to lead to a dLGN response. However,
anonymous inputs tend to decrease the likelihood of an RGC input triggering a dLGN response,
possibly due to refractoriness, although they observe no refractory effect when dLGN spikes are
not triggered by RGCs (Levine and Cleland, 2001). Additionally, visual stimulation enhances
the efficacy of the retinogeniculate synapse, perhaps because of the increased synchrony in
RGCs projecting to the same dLGN neuron during vision (Usrey et al., 1999; Rowe and Fischer,
2001)
It is also possible to study retinogeniculate transmission with only a single recording
electrode in the dLGN. Two recent studies have used the method of S-potential recordings to
look at transmission at this synapse in the awake cat (Weyand, 2007) and monkey (Sincich et al.,
2007). S-potentials refer to the retinal excitatory postsynaptic potentials (EPSPs) which can also
be recorded extracellulary from dLGN neurons. Using this technique, they show that the second
EPSP is more likely to evoke a dLGN spike if it was preceded by an EPSP followed by a short
inter-potential interval, confirming the paired retinal and dLGN findings. It was also shown that
during wakefulness, the interval that leads to facilitated transmission is shorter than under
anesthesia, although general efficacy is improved under awake conditions (Weyand, 2007).

22
Slice experiments have also contributed to our understanding of the transformations that
can occur at the retinogeniculate synapse. Using slices prepared from postnal day (p)28 – p33
mice, dLGN neurons were recorded in a dynamic clamp mode during optic tract stimulation
(Blitz and Regehr, 2003). This allowed for a comparison of the α-amino-3-hydroxy-5-methyl-4-
isoxazolepropionic acid (AMPA) and N-methyl-D-asparate (NMDA) components of the dLGN
response. The AMPA component allowed for a precise and faithful following of the RGC input
with short latency responses. However, this component was sensitive to short-term plasticity in
the forms of both presynaptic depression and AMPA receptor (AMPAR) desensitization. The
NMDA component acted at a longer latency and was less precise, but allowed for multiple
dLGN responses and an amplification of the RGC input. Depending on the strength of the
NMDA and AMPA components of the dLGN response, a single dLGN neuron will produce a
reliable response to a repeated pattern of input, with the two components working together to
modulate the response relayed to V1 (Blitz and Regehr, 2003).
Modulation of the Retinogeniculate Synapse
The retinogeniculate synapse is the first location where visual input can be modified on
its path to cortex, and the degree of change that occurs at this synapse can be modulated by input
from other brain areas. Using whole-cell patch-clamp recording methods and calcium imaging,
it has been shown that dopamine (D) (Govindaiah and Cox, 2006) serotonin (5-HT) and GABA
(Chen and Regehr, 2003) can inhibit synaptic strength and decrease synaptic depression. These
neuromodulators can act presynaptically through D2 (Govindaiah and Cox, 2006), 5-HT or
GABAB receptors (Chen and Regehr, 2003) or postsynaptically through GABAA receptors
(McCormick and Bal, 1994).

23
In vivo studies have shown that eye movements can also modulate the dLGN response
(Lal and Friedlander, 1990) as shown in the paralyzed, anesthetized cat preparation using passive
eye movement. Additionally, electrical stimulation of the parabrachial region of the brain stem
leads to an increased release of acetylcholine in the dLGN. This cholinergic activation has been
shown to facilitate the response of both X- and Y-cell types to sinusoidal gratings (Uhlrich et al.,
1995).
Geniculocortical Transmission
In order to understand the plasticity that occurs in visual cortex, it is necessary to
consider not only the input sent from the retina to the dLGN, but also the message transmitted
across the thalamocortical (TC) synapse and received by the cortical neuron. The rules
governing the plasticity at the TC synapse most likely rely on both the input to the cortical
neuron and its response. As discussed above, the message from retina to the dLGN is modified
at the retinogeniculate synapse, and it is not necessarily a simple, faithful relay of visual
information. Below I will describe how visual information is transformed at the TC synapse.
TC transmission is often studied using simultaneous recordings from the dLGN and V1
in a manner similar to studies of retinogeniculate transmission. Alonso et al. (2001) described
the properties shared by connected dLGN and V1 simple cells in the anesthetized cat
preparation. They sampled from numerous pairs of dLGN and V1 simple cells with overlapping
RFs. However, only one-third of their pairs were connected. Neurons are said to be connected if
they have overlapping RFs, with the dLGN spikes preceding the V1 spikes by 2 – 4 ms (Tanaka,
1983). Other RF properties proved important in predicting whether a pair of neurons was
connected. Connected neurons generally had the same RF sign and size and were not just
overlapping in space (Alonso et al., 2001).

24
Efficacy studies similar to those conducted at the retinogeniculate synapse have been
performed at the TC synapse as well. A single spike preceded by at least 10 ms of silence has
about a 1% chance of driving a connected cortical partner (Usrey et al., 2000). If two spikes
arrive within a 10 – 15 ms window, the second spike will show an enhancement in efficacy,
becoming up to 5 times more likely to drive the postsynaptic neuron (Usrey et al., 2000). Usrey
et al. (2000) were also able to record simultaneously from two dLGN neurons and a V1 neuron
to which they were both connected. In this case, if spikes arrived from the two neurons within
7.5 ms, the second spike to arrive showed an enhancement in efficacy.
Seemingly contrary to the increased efficacy of the second spike, other studies have
shown that the TC synapse is depressing in vivo (Jia et al., 2004; Boudreau and Ferster, 2005).
Field potential recordings from rat V1 during electrical stimulation of dLGN show paired-pulse
depression (PPD), with the degree of depression increasing as the frequency of the stimulation
increases (Jia et al., 2004). This short-term synaptic depression is dependent on GABA-
mediated inhibition, and is not affected by NMDA or AMPA receptor blockers. Intracellular
recordings during dLGN stimulation in cat show a decrease in postsynaptic potentials in neurons
monosynaptically receiving dLGN input following repeated stimulation (Boudreau and Ferster,
2005). This depression was not affected by cortical stimulation. However, reducing dLGN
activity (via increased intraocular pressure) greatly enhanced the first evoked PSP, although
subsequent PSPs were depressed. These observations imply that the TC synapse may be in a
state of tonic depression due to spontaneous input, and decreasing that input reduces the
depression. However, the authors who conducted the study note that the degree of PSP
depression is small (never more than 40%). From this they conclude that amount of depression
based on spontaneous dLGN activity alone is near saturation, and has little effect on the efficacy

25
of transmission at this synapse. Further evidence for the tonic depression of the TC response
comes from work in the somatosensory system of the awake rabbit. Using current source-
density analysis, Swadlow et al. (2002) found an increased cortical response when the TC input
was preceded by a long ISI. The long ISI would allow time for the release from tonic
depression, thus enhancing the response.
Even though the depression is small and can be overcome, it is difficult to reconcile this
PPD and PSP depression with the short-latency facilitation observed by Usrey et al. (2000). One
possibility may be choice of anesthesia. As opposed to Swadlow et al. (2002) who recorded
from awake animals and Jia et al. (2004) who used urethane anesthesia, Usrey et al. (2000) used
a barbiturate anesthesia during their recordings. This may have resulted in a decrease in dLGN
spontaneous activity, thus relieving the tonic depression and allowing for facilitation of
transmission at the TC synapse. However, Boudreau and Ferster (2005) also used barbiturates
and observed PSP depression. The difference in this case may be that their results are based on
electrical stimulation, not intrinsic dLGN activity, thus changes to dLGN firing based on
barbiturate anesthesia may not have a large effect on their results. It is important to note that
many of these studies of spike efficacy in the retinogeniculocortical pathway were performed in
the anesthetized, paralyzed preparation and factors such as attentional state, level of arousal, eye
movements, etc. may all influence transmission in the visual pathway. This underscores the
importance of recording from the awake animal.
One important difference between the retinogeniculate and geniculocortical pathways is
that while the retinal response is always precisely and tightly coupled to the dLGN response,
there is much more variability in the cortical response time relative to the dLGN input (Veredas
et al., 2005). Using a theoretical model, Veredas et al. proposed that the difference is due to

26
changes in the EPSP through the visual pathway. Increased EPSP rise time, duration and jitter in
the TC synapse relative to the retinogeniculate synapse can explain the differences in
correlations. While it may seem counterintuitive that the visual system would want to lose the
precise timing as information ascends the visual pathway, the increase in EPSP duration allows
for increased temporal summation at the V1 neuron, which may be important for visual
computation.
Disynaptic Connections
Kara and Reid examined the direct influence of RGCs on cortical firing by
simultaneously recording from RGCs and V1 neurons with overlapping RFs that were
disynaptically connected (2003). They found that a single RGC can account for about 3% of a
V1 neuron’s response, and that the V1 response is enhanced when the RGC spike is preceded by
a short (<10 ms) ISI. However, they found that this facilitation is greater than what would be
expected from a combination of the monosynaptic enhancements in the retinogeniculate and
geniculocortical synapses (Kara and Reid, 2003). This may imply that factors other than the
preceding ISI are important in determining spike efficacy. However, this study may also be
flawed by the use of barbiturate anesthesia (see above) and also the difficulty of assuring
connectivity between RGCs and the cortical neurons to which they project.
Bursts
The phenomenon of thalamic bursting is one important property of thalamic nuclei that
must be considered when evaluating TC transmission. While bursts are commonly observed
throughout the thalamus during sleep states, they have been shown to occur in the dLGN of the
awake cat (Guido and Weyand, 1995), rabbit (Bezdudnaya et al., 2006), and monkey
(Ramcharan et al., 2000). This section will focus on thalamic bursting during sensory

27
stimulation in order to understand how this type of activity could affect information transfer to
cortex.
Bursting results from the activation of T-type Ca2+
channels, which require a long period
of hyperpolarization for deinactivation (Sherman, 2001). Extracellularly, bursts are defined by a
period of quiescence lasting > 100 ms followed by two or more spikes with ISIs less than 4 ms
(Lu et al., 1992). The long silent period is the time in which the cell is hyperpolarized wherein
T-type channels become deinactivated. Following deinactivation, activating the channels
requires only a subthreshold increase in membrane potential. Once the channels are open, Ca2+
can enter the cell, depolarizing it even further to the Na+ channel activation potential, resulting in
a train of two or more spikes riding on the Ca2+
current.
Epochs of thalamic activity dominated by bursting are said to occur when the thalamus is
in a “burst mode”. Burst mode is highly correlated with slow-wave sleep, but also occurs in
awake, “inattentive” animals (Bezdudnaya et al., 2006). In recording from an awake animal,
periods of high bursting activity are accompanied by high voltage irregular hippocampal EEG
activity which is indicative of an inattentive state. Temporal tuning curves recorded from dLGN
neurons during alert versus inattentive states indicate that inattention (i.e. burst mode) flattens
the tuning curve and decreases response to visual stimuli. In this study, RF size remained the
same in both burst and tonic modes (Bezdudnaya et al., 2006). However, work performed in the
anesthetized cat dLGN has shown that burst spikes were restricted to smaller regions in the RF
center (Rivadulla et al., 2003). The discrepancy between these findings is likely to be accounted
for by anesthetic effects or differences in RF mapping methodology. This highlights the
importance of using an awake preparation when possible.

28
The purpose of bursts in signal transmission is unknown. Bursts are often triggered by
retinal spikes, with the first spike and secondary burst spikes being highly correlated with retinal
input (Usrey et al., 1999; Rowe and Fischer, 2001). Bursting may serve as a method of signal
amplification, as additional spikes in the burst are added to the RGC spike train to enhance the
message sent to V1. Larger visual stimuli are more likely to evoke bursts, which implies a role
for the inhibitory surround in burst production (Weyand et al., 2001). In fact, bursting, as
opposed to tonic firing, occurs following stimuli that strongly drive both the excitatory center
and inhibitory surround of the RF (Alitto et al., 2005).
dLGN bursts have also been shown to be related to visual processing. Bursts have been
shown to carry as nearly as much visual information as tonic firing (Reinagel et al., 1999).
Recordings from dLGN in awake cats show that the majority of bursts occur during visual
stimulation and are sometimes related to eye movements (Guido and Weyand, 1995). In the
anesthetized cat preparation, bursts are more prominent during natural scene viewing than while
the animal views a white noise stimuli (Lesica and Stanley, 2004). Bursts are also a more
reliable indication of visual stimuli than tonic spikes in the sense that they will more often occur
at a temporarily precise time following multiple presentations of the same stimuli (Alitto et al.,
2005). Recent work using intracellular in vivo recordings from cat dLGN have confirmed that
bursts can result from natural scene stimuli (Wang et al., 2007).
What effect do bursts have on TC transmission? This question has been investigated
using the somatosensory system of awake rabbits (Swadlow and Gusev, 2001). The first spike in
a burst recorded from the ventrobasal nucleus of the thalamus was found to be more effective
than isolated spikes or “pseudobursts” at driving presumptive cortical interneurons in barrel
cortex (Swadlow and Gusev, 2001). Pseudobursts consist of two or more spikes with ISIs less

29
than 4 ms, but are not preceded by at least 100 ms of silence. Subsequent burst spikes also show
an increased efficacy of transmission, and can lead to cortical spiking (Swadlow and Gusev,
2001). These results confirm what is known about TC transmission as described above. The
long period of quiescence preceding the bursts removes the tonic depression at the TC synapse,
increasing the efficacy of the initial burst spike. Additionally, subsequent burst spikes arrive
after short ISIs, which would also suggest an increased ability to drive cortex.
The existence of bursts has been confirmed in the mouse (Grubb and Thompson, 2005).
The properties of bursts recorded from the anesthetized mouse dLGN are very similar to those
reported in other species. In addition to having similar response properties, the percentage of
spikes in bursts in the mouse (~20%) was approximately the same as reported in other species.
Burst spiking increased in the presence of visual stimuli, and the bursts contained reliable
information about the input (Grubb and Thompson, 2005).
Ocular Dominance Plasticity
The studies described in this thesis examine the patterns of dLGN input during periods of
visual deprivation. In addition to understanding the activity entering and exiting the dLGN, it is
important to understand what is known about the experience-dependent plasticity resulting from
changes in visual experience. Beginning over 40 years ago with the pioneering studies of Hubel
and Wiesel (1963a; 1963b) ocular dominance (OD) plasticity, or the brain’s ability to change the
relative responsiveness to input from the two eyes, has been a well studied model of cortical
plasticity. While the literature surrounding this topic is vast, I will focus on the aspects specific
to the aims of this study: how various inputs affect OD plasticity; mouse OD plasticity;
differences between juvenile and adult plasticity in the mouse; and the effects of deprivation on
retinal and dLGN activity.

30
Wiesel and Hubel (1963a) first studied the effects of visual deprivation in V1 by
recording extracellularly from numerous neurons in kittens with different histories of visual
experience. They classified each neuron’s response based on the relative responsiveness to
contralateral- and ipsilateral-eye inputs (OD scores). In the normally reared kitten, the
distribution of OD scores showed a bias for neurons responding equally to inputs from both eyes,
with a slight overall bias for contralateral input. Monocular lid closure (MC) or monocular
occlusion (MO) with a translucent lens prior to eye-opening led to a loss of responsiveness to the
deprived eye, where the distribution of responses shifted entirely to the non-deprived eye.
Kittens that were allowed normal visual experience until 9 weeks of age followed by MC for 4
months showed a similar OD shift.
Subsequent studies of OD plasticity by other groups found that the OD shift can occur in
less than one day (Mioche and Singer, 1989). Mioche and Singer used chronically implanted
electrodes to follow the response properties of cortical neurons after brief periods of MO with an
eye patch in kittens. A decrease in response to the deprived eye was observed as early as 6 hr
following MO, with a complete loss of the deprived-eye response occurring as early as 12 hours
after deprivation.
Wiesel and Hubel (1965) also compared the effects of binocular lid closure (BC) with
MC. Somewhat surprisingly, they found that BC had a minimal effect on the distribution of OD
scores. From this work they concluded that the amount of plasticity observed in cortex following
suture of one eye depends on whether or not the other eye is sutured. Subsequently, several
studies have used different visual inputs to induce OD plasticity, in an attempt to understand
which input patterns result in cortical plasticity (e.g. Blakemore, 1976; Kratz and Spear, 1976;
Singer et al., 1977; Wilson et al., 1977; Chapman et al., 1986; Greuel et al., 1987).

31
Two studies that were specifically designed to elucidate the role of retinal activity in OD
plasticity compared MC with monocular retinal inactivation (MI) via intraocular injection of
tetrodotoxin (TTX), creating an imbalance in amount of input entering the visual systems
(Chapman et al., 1986; Greuel et al., 1987). Both studies showed that MI shifted responses
towards the non-deprived eye. Additionally, both research groups also recorded from animals
that had one retina inactivated while the other lid was sutured. These studies produced
contrasting results. Chapman et al. (1986) observed an OD shift towards the closed eye (which
has residual retinal activity), and Greuel et al. (1987) observed no OD shift. Therefore these
conflicting results, made it difficult to make conclusions regarding the role of retinal activity in
OD plasticity.
To resolve this conflict, Rittenhouse et al. (1999) directly compared the effects of MC
and MI in the same study. They found that brief periods (2 days) of MI caused only a minimal
OD shift (contrary to the prior studies which used extended periods of MI > 6 days), with MC
leading to a much more extreme shift towards the non-deprived eye. From these results they
concluded that the residual retinal activity that remains during MC, and is relayed from the retina
to V1 via the dLGN, is required to induce a significant OD shift. This residual presynaptic
activity, when paired with weak postsynaptic activation, leads to homosynaptic long term
depression (LTD) and a weakening of the synapses carrying deprived-eye input (Blais et al.,
1999; Rittenhouse et al., 1999). On the other hand, MI, the complete absence of retinal input,
leads to no depression. This was assumed to be due to a lack of input from dLGN to V1. With
no residual input entering cortex, there is no activity to drive the homosynaptic depression.
Subsequent studies have confirmed the connection between deprived-eye depression
following MC and the in vitro model of plasticity, LTD. The molecular and functional changes

32
that result from brief MC, including altered AMPAR phosphorylation and decreased surface
expression, are the same changes as seen following induction of LTD. Furthermore, response
depression from MC occludes the subsequent induction of LTD ex vivo (Heynen et al., 2003).
Additionally, blocking AMPAR endocytosis prevents OD plasticity (Yoon et al., 2008).
Moreover, these AMPAR changes are not found following MI. This implies that MC induces
LTD in the visual cortex, while MI does not.
Ocular Dominance Plasticity in the Mouse
Mice serve as the animal model for the research described in this thesis. While the
preliminary research was performed in cats, the important findings including the effects of MC
and MI have been replicated in the mouse. In recent years it has become clear that plasticity of
binocular connections in visual cortex is highly conserved across extant mammalian species
(reviewed in Horton and Adams, 2005). Furthermore, rodents are phylogenetically closer to
primates than carnivores are, thus they may serve as a better model for understanding the human
brain (Arnason et al., 2002). Even though rodents do lack ocular dominance columns observed in
carnivores and old-world primates, they exhibit robust OD plasticity following MC (described
below).
Among the advantages of using mice as a model system are: that they have been in-bred
to minimize inter-individual variability, they are plentiful and relatively inexpensive, and they
are amenable to genetic manipulations. Furthermore, we can easily perform both acute and
chronic electrophysiological studies from the awake mouse preparation. All these features allow
more rapid progress than in many other species. Plasticity in rodent visual cortex can also be
compared to the somatosensory “barrel” cortex to discern features of cortical plasticity that may
generalize across areas. Additionally, because the dLGN of the mouse is relatively homogenous,

33
lacking multiple relay cell types (Sherman and Guillery, 2006), the findings may be more easily
extrapolated to the other primary thalamic nuclei.
Originally studied by Drager (1978), OD plasticity occurs in the mouse and is similar to
the plasticity observed in other species. In the normally reared mouse, the distribution of OD
scores shows a bias towards the contralateral eye. Converting the distribution of OD scores to a
relative response ratio, she found mice have a contralateral-to-ipsilateral (C/I) ratio of about 2 to
1. In animals that have undergone MC from eye-opening, there is an OD shift towards the non-
deprived eye. While the shift in the hemisphere contralateral to the closed eye is not as extreme
as the shifts observed in cat, the majority of the neurons have responses dominated by the non-
deprived eye. In the hemisphere ipsilateral to the deprived eye, nearly all neurons became
monocular, responding only to the non-deprived eye.
Frenkel and Bear (2004) used chronic field potential recording electrodes to follow
effects of MC and BC in awake, juvenile mice. Similar to Drager’s (1978) finding with single
unit recordings, the normal mouse shows a 2 to 1 ratio of visual evoked potentials (VEPs)
recorded in response to input from the contralateral and ipsilateral eye. MC, but not BC, leads
to a statistically significant change in the C/I VEP ratio, corresponding to an OD shift away from
the deprived eye. One advantage to the chronic recording method is the ability to separate the
changes in the responses of the two eyes. Two distinct responses in V1 lead to the change in the
C/I ratio. First there is a rapid depression of the deprived-eye response, followed by a delayed
potentiation of the non-deprived eye response. The deprived-eye depression peaks after 3 days
of MC, while the open-eye potentiation becomes significant after 7 days of MC (Frenkel and
Bear, 2004).

34
Frenkel and Bear (2004) also examined the effects of MI on VEPs in V1. Unlike the
deprived-eye depression observed following MC, there was no change in the deprived-eye
response following MI. However, there was a significant potentiation of ipsilateral eye response
in the same hemisphere. This implies that any OD shift observed in cats following MI resulted
not from a loss of response to the silenced retina, but rather an increase in the response of the
other eye. MI appears to protect against the initial stage of the OD shift (the reduction in the
deprived-eye response), while still allowing the open-eye potentiation.
The Concept of a “Critical Period” for Ocular Dominance Plasticity
In their original study of OD plasticity, Wiesel and Hubel (1963a) recorded from one cat
that underwent 3 months of MC in adulthood. There was no significant shift in the distribution
of OD scores for this animal. A careful investigation of the ages in which the cat is most
susceptible to the effects of MC showed that kittens begin to a show an OD shift after 4 weeks of
age, with a peak shift occurring following MC when the animal is 6 – 8 weeks of age (Hubel and
Wiesel, 1970). After this peak, the sensitivity to MC declines, with no shift observed after 3
months of age. Furthermore, after experiencing MC during this period of susceptibility, normal
visual experience in adulthood will not correct the OD shift, rendering the animal amblyopic
(Hubel and Wiesel, 1970).
Rodents have also been shown to have a “critical period” for OD plasticity via MC (e.g.
(Fagiolini et al., 1994; Gordon and Stryker, 1996). Early work in the rat showed that the critical
period begins at around 3 weeks of age and tapers off after 5 weeks (Fagiolini et al., 1994). Four
days of MC in the mouse caused the most robust loss of deprived-eye response at postnatal day
(P)28, with no shift occurring after P32 (Gordon and Stryker, 1996). However, more recent

35
work in rodents has shown that OD plasticity persists in the mature brain (Hofer et al., 2006a;
Iny et al., 2006; Hooks and Chen, 2007).
Several studies in the rat have shown that the critical period in the rat can be
“reactivated.” Cortical infusion of drugs that break down the perineuronal nets in the
extracellular matrix which may prevent anatomical changes from occurring provide one way to
reopen the critical period in adults (Pizzorusso et al., 2002; Pizzorusso et al., 2006). Non-
invasive options including environmental enrichment (Sale et al., 2007) and periods of dark
exposure (He et al., 2007) have also been shown to allow for juvenile-like OD plasticity in
adults.
Recent studies suggest that the classically defined critical period for OD plasticity may
not even exist in the mouse (reviewed in Hofer et al., 2006a). Changes in the expression of the
immediate early gene, Arc, show an expansion of the non-deprived eye input to V1 following
MC well past the traditional end of the critical periods (13 weeks) (Tagawa et al., 2005).
Intrinsic signal optical imaging and extracellular single-unit recording techniques also verified
OD plasticity in the adult mouse, although this OD shift was attributed to a potentiation of the
non-deprived eye input, with no depression of the deprived-eye response (Hofer et al., 2006b).
Furthermore, VEP recordings in the adult mouse show that extended periods of MC (7 days) lead
to both deprived-eye depression and open-eye potentiation, while shorter periods of MC (3 days)
are not accompanied by a loss of the deprived-eye response (Frenkel et al., 2006). While these
findings suggest that the concept of a “critical period” may not exist in the mouse, there is a
qualitative difference between juvenile and adult OD plasticity in this species.

36
Spontaneous Retinal Ganglion Cell Activity
While much is known about the effects of monocular deprivation in V1, fewer studies
have examined the physiological consequences of visual manipulation in the retina and dLGN.
Closing the eyelid leaves spontaneous RGC activity as the driving input to the dLGN. While one
may assume that this activity is purely random, RGC activity does show correlations, even in the
absence of visual input (reviewed in Mastronarde, 1989). Single-unit recordings from the retina
of cats have shown that neighboring RGCs can be highly correlated (Rodieck, 1967). This
correlation depends on the sign of the RGCs (ON- or OFF-center), with neurons of the same sign
showing positive correlation, and neurons with opposite sign showing negative correlations.
Furthermore, the degree of overlap of the RF centers determines the magnitude of the correlation
(Mastronarde, 1989). Shared input from rods or from spiking amacrine neurons, or gap junction
connections between RGCs may account for the correlated firing (Mastronarde, 1989).
Effects of Monocular Deprivation on dLGN Activity
Monocular lid suture only has small effects on the response properties of neurons in the
dLGN. The original study by Wiesel and Hubel (1963b) showed the effects of lid suture from
birth until 3 months of age in kittens. The physiological effects of the lid suture were minimal,
with neurons in the deprived dLGN layers retaining their RF size and shape. Some neurons
showed an increase in response latency, and the authors report a slight decrease of overall
activity in the deprived layer. Anatomically, some atrophy was observed in the layer receiving
deprived-eye input (Wiesel and Hubel, 1963b).
Subsequent studies in the cat show similar findings to those reported by Wiesel and
Hubel (Sherman and Sanderson, 1972; LeVay and Ferster, 1977; Lehmkuhle et al., 1980;
Friedlander et al., 1982). In a study where X- and Y-cells were evaluated separately, there was

37
no physiological effect on the Y cells and a small decrease in contrast sensitivity in the X-cells
(Lehmkuhle et al., 1980). However, there does appear to be a loss of Y-, but not X-, cells in the
deprived laminae (LeVay and Ferster, 1977), corroborating the atrophy observed by Wiesel and
Hubel. Further studies by Friedlander et al. (1982) confirm the morphological shrinkage of the
Y-cells, and find physiological deficits in the remaining Y-cells, probably overlooked in the prior
studies due to difficulties in recording from the smaller cell bodies.
Extended periods of MC in primates also result in small physiological changes. Four
years of monocular lid suture in the galago (Sesma et al., 1984) and five to six years of
monocular lid suture in macaque monkeys (Levitt et al., 2001) led to only subtle changes in
dLGN response properties, however atrophy was observed in both parvo- and magnocellular
cells. One explanation for atrophy being observed in both cell types in the primates but not the
cats is that the parvo- and magnocellular cells are separated into different layers in the primates,
while X- and Y-cell types are intermingled within layers in the cat. This may lead to competition
for retinal input between X- and Y-cells, which seems to result in selective Y-cell atrophy
(Sesma et al., 1984).
Previous Recordings from dLGN Neurons During Deprivation
There are very few studies of the effect of deprivation on dLGN activity during periods
of deprivation. In fact, there are only two reports describing dLGN activity during MI in the
anesthetized cat (Stryker and Harris, 1986; Rittenhouse et al., 1999). In the Stryker and Harris
(1986) study, recordings were made 88 - 100 hours after injection of TTX. At this time point,
the pupil of the injected eye was no longer dilated and the consensual pupilary response was
observed when light was applied to the non-injected eye. Recordings were made ipsilateral to
the injected eye, in the A1 lamina of the dLGN. While recordings from the A and C layers were

38
normal, the deprived layer showed units that were silent or had very low levels of spontaneous
firing, with no evidence of evoked responses. Visual responses through the previously deprived
eye were observed in cortex 10 – 22 hours later (Stryker and Harris, 1986). These results
suggest that the dLGN neurons showed a large reduction in activity during MI. However,
recordings performed at an earlier time-point are necessary to confirm that assumption.
In the study by Rittenhouse et al. (1999) dLGN recordings were made within 48 hours of
TTX injection. They also observed a huge decrease in spontaneous dLGN activity (~80%),
confirming the results of Stryker and Harris (1986). Taken together, these two studies suggest
that the amount of dLGN activity during retinal inactivation is greatly reduced in the
anesthetized cat preparation.
Rittenhouse et al. (1999) also recorded from the dLGN during MC. The spontaneous
firing rate in the dLGN was higher than during MI, although they did not compare MC to an
open eye response rate. They conclude that the important difference between MC and MI is the
amount of spontaneous firing in the dLGN (Rittenhouse et al., 1999). Work from Kaplan et al.
(1987) shows that the dLGN firing rate is lower than the retinal firing rate during both
spontaneous and stimulus-driven conditions. While they do not specifically examine MC and
MI, this would suggest that the dLGN firing rate is highest during open-eye viewing, lower
during MC when only spontaneous retinal activity remains, and near zero during MI. However,
one caveat to all of these studies is that they were performed in the anesthetized cat. To fully
determine the dLGN firing responses during open-eye viewing, MC, and MI, careful studies
need to be performed in the awake animal. The goal of this project is to explore dLGN activity
under these 3 viewing conditions in the awake animal, in order to understand the contribution of
presynaptic input to OD plasticity.

39
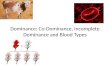
Figure 1-1
Figure 1-1 “Hidden lamination” in the mouse dLGN
The mouse dLGN consists of an outer shell receiving contralateral eye input and an inner core
receiving ipsilateral eye input. a, Confocal image of a coronal section from a mouse brain
following intraocular injections of cholera toxin B (CTB). CTB is an anterograde tracer that fills
neuronal axons. CTB conjugated to Alexa 555 was injected in the ipsilateral eye (indicated in
red), and CTB conjugated to Alexa 647 was injected into the contralateral eye (indicated in
blue). The mouse was a transgenic animal expressing GFP under the GAD67 promoter
(Chattopadhyaya et al., 2004), thus inhibitory interneurons are labeled in green. Scale bar: 200
μm. b, Schematic depicting the contralateral shell and ipsilateral core of the dLGN. Both of
these areas project to the binocular zone of V1.


41
References
Alitto HJ, Weyand TG, Usrey WM (2005) Distinct properties of stimulus-evoked bursts in the
lateral geniculate nucleus. J Neurosci 25:514-523.
Alonso JM, Usrey WM, Reid RC (2001) Rules of connectivity between geniculate cells and
simple cells in cat primary visual cortex. J Neurosci 21:4002-4015.
Arcelli P, Frassoni C, Regondi MC, De Biasi S, Spreafico R (1997) GABAergic neurons in
mammalian thalamus: a marker of thalamic complexity? Brain Res Bull 42:27-37.
Arnason U, Adegoke JA, Bodin K, Born EW, Esa YB, Gullberg A, Nilsson M, Short RV, Xu X,
Janke A (2002) Mammalian mitogenomic relationships and the root of the eutherian tree.
Proc Natl Acad Sci U S A 99:8151-8156.
Bezdudnaya T, Cano M, Bereshpolova Y, Stoelzel CR, Alonso JM, Swadlow HA (2006)
Thalamic burst mode and inattention in the awake LGNd. Neuron 49:421-432.
Blais BS, Shouval HZ, Cooper LN (1999) The role of presynaptic activity in monocular
deprivation: comparison of homosynaptic and heterosynaptic mechanisms. In: Proc Natl
Acad Sci U S A, pp 1083-1087.
Blakemore C (1976) The conditions required for the maintenance of binocularity in the kitten's
visual cortex. J Physiol 261:423-444.
Blitz DM, Regehr WG (2003) Retinogeniculate synaptic properties controlling spike number and
timing in relay neurons. J Neurophysiol 90:2438-2450.
Boudreau CE, Ferster D (2005) Short-term depression in thalamocortical synapses of cat primary
visual cortex. J Neurosci 25:7179-7190.
Brauer K, Schober W, Winkelmann E (1979) Two morphologically different types of retinal
axon terminals in the rat's dorsal lateral geniculate nucleus and their relationships to the
X- and Y-channel. Exp Brain Res 36:523-532.
Chapman B, Jacobson MD, Reiter HO, Stryker MP (1986) Ocular dominance shift in kitten
visual cortex caused by imbalance in retinal electrical activity. Nature 324:154-156.
Chattopadhyaya B, Di Cristo G, Higashiyama H, Knott GW, Kuhlman SJ, Welker E, Huang ZJ
(2004) Experience and activity-dependent maturation of perisomatic GABAergic
innervation in primary visual cortex during a postnatal critical period. J Neurosci
24:9598-9611.
Chen C, Regehr WG (2003) Presynaptic modulation of the retinogeniculate synapse. J Neurosci
23:3130-3135.
Cleland BG, Dubin MW, Levick WR (1971) Sustained and transient neurones in the cat's retina
and lateral geniculate nucleus. J Physiol 217:473-496.

42
Crowley JC, Katz LC (2002) Ocular dominance development revisited. Curr Opin Neurobiol
12:104-109.
Cudeiro J, Sillito AM (1996) Spatial frequency tuning of orientation-discontinuity-sensitive
corticofugal feedback to the cat lateral geniculate nucleus. J Physiol 490 ( Pt 2):481-492.
Drager UC (1978) Observations on monocular deprivation in mice. J Neurophysiol 41:28-42.
Enroth-Cugell C, Robson JG (1966) The contrast sensitivity of retinal ganglion cells of the cat. J
Physiol 187:517-552.
Erisir A, Van Horn SC, Sherman SM (1997a) Relative numbers of cortical and brainstem inputs
to the lateral geniculate nucleus. Proc Natl Acad Sci U S A 94:1517-1520.
Erisir A, Van Horn SC, Bickford ME, Sherman SM (1997b) Immunocytochemistry and
distribution of parabrachial terminals in the lateral geniculate nucleus of the cat: a
comparison with corticogeniculate terminals. J Comp Neurol 377:535-549.
Fagiolini M, Pizzorusso T, Berardi N, Domenici L, Maffei L (1994) Functional postnatal
development of the rat primary visual cortex and the role of visual experience: dark
rearing and monocular deprivation. Vision Res 34:709-720.
Frenkel MY, Bear MF (2004) How monocular deprivation shifts ocular domiance in visual
cortex of young mice. Neuron 44:917-923.
Frenkel MY, Sawtell NB, Diogo AC, Yoon B, Neve RL, Bear MF (2006) Instructive effect of
visual experience in mouse visual cortex. Neuron 51:339-349.
Friedlander MJ, Lin CS, Sherman SM (1979) Structure of physiologically identified X and Y
cells in the cat's lateral geniculate nucleus. Science 204:1114-1117.
Friedlander MJ, Stanford LR, Sherman SM (1982) Effects of monocular deprivation on the
structure-function relationship of individual neurons in the cat's lateral geniculate
nucleus. J Neurosci 2:321-330.
Friedlander MJ, Lin CS, Stanford LR, Sherman SM (1981) Morphology of functionally
identified neurons in lateral geniculate nucleus of the cat. J Neurophysiol 46:80-129.
Fukuda Y, Stone J (1974) Retinal distribution and central projections of Y-, X-, and W-cells of
the cat's retina. J Neurophysiol 37:749-772.
Gabriel S, Gabriel HJ, Grutzmann R, Berlin K, Davidowa H (1996) Effects of cholecystokinin
on Y, X, and W cells in the dorsal lateral geniculate nucleus of rats. Exp Brain Res
109:43-55.
Gilbert CD, Kelly JP (1975) The projections of cells in different layers of the cat's visual cortex.
J Comp Neurol 163:81-105.

43
Godement P, Salaun J, Imbert M (1984) Prenatal and postnatal development of retinogeniculate
and retinocollicular projections in the mouse. J Comp Neurol 230:552-575.
Gordon JA, Stryker MP (1996) Experience-dependent plasticity of binocular responses in the
primary visual cortex of the mouse. J Neurosci 16:3274-3286.
Govindaiah G, Cox CL (2006) Depression of retinogeniculate synaptic transmission by
presynaptic D(2)-like dopamine receptors in rat lateral geniculate nucleus. Eur J Neurosci
23:423-434.
Greuel JM, Luhmann HJ, Singer W (1987) Evidence for a threshold in experience-dependent
long-term changes of kitten visual cortex. Brain Res 431:141-149.
Grubb MS, Thompson ID (2003) Quantitative characterization of visual response properties in
the mouse dorsal lateral geniculate nucleus. J Neurophysiol 90:3594-3607.
Grubb MS, Thompson ID (2005) Visual response properties of burst and tonic firing in the
mouse dorsal lateral geniculate nucleus. J Neurophysiol 93:3224-3247.
Guido W, Weyand T (1995) Burst responses in thalamic relay cells of the awake behaving cat. J
Neurophysiol 74:1782-1786.
Guillery RW (1966) A study of Golgi preparations from the dorsal lateral geniculate nucleus of
the adult cat. J Comp Neurol 128:21-50.
He HY, Ray B, Dennis K, Quinlan EM (2007) Experience-dependent recovery of vision
following chronic deprivation amblyopia. Nat Neurosci 10:1134-1136.
Heynen AJ, Yoon BJ, Liu CH, Chung HJ, Huganir RL, Bear MF (2003) Molecular mechanism
for loss of visual cortical responsiveness following brief monocular deprivation. Nat
Neurosci 6:854-862.
Hofer SB, Mrsic-Flogel TD, Bonhoeffer T, Hubener M (2006a) Lifelong learning: ocular
dominance plasticity in mouse visual cortex. Curr Opin Neurobiol 16:451-459.
Hofer SB, Mrsic-Flogel TD, Bonhoeffer T, Hubener M (2006b) Prior experience enhances
plasticity in adult visual cortex. Nat Neurosci 9:127-132.
Hooks BM, Chen C (2006) Distinct roles for spontaneous and visual activity in remodeling of
the retinogeniculate synapse. Neuron 52:281-291.
Hooks BM, Chen C (2007) Critical periods in the visual system: changing views for a model of
experience-dependent plasticity. Neuron 56:312-326.
Horton JC, Adams DL (2005) The cortical column: a structure without a function. Philos Trans
R Soc Lond B Biol Sci 360:837-862.

44
Houser CR, Vaughn JE, Barber RP, Roberts E (1980) GABA neurons are the major cell type of
the nucleus reticularis thalami. Brain Res 200:341-354.
Hubel DH, Wiesel TN (1970) The period of susceptibility to the physiological effects of
unilateral eye closure in kittens. J Physiol 206:419-436.
Iny K, Heynen AJ, Sklar E, Bear MF (2006) Bidirectional modifications of visual acuity induced
by monocular deprivation in juvenile and adult rats. J Neurosci 26:7368-7374.
Jaubert-Miazza L, Green E, Lo FS, Bui K, Mills J, Guido W (2005) Structural and functional
composition of the developing retinogeniculate pathway in the mouse. Vis Neurosci
22:661-676.
Jia F, Xie X, Zhou Y (2004) Short-term depression of synaptic transmission from rat lateral
geniculate nucleus to primary visual cortex in vivo. Brain Res 1002:158-161.
Kaplan E, Purpura K, Shapley RM (1987) Contrast affects the transmission of visual information
through the mammalian lateral geniculate nucleus. J Physiol 391:267-288.
Kara P, Reid RC (2003) Efficacy of retinal spikes in driving cortical responses. J Neurosci
23:8547-8557.
Kratz KE, Spear PD (1976) Effects of visual deprivation and alterations in binocular competition
on responses of striate cortex neurons in the cat. J Comp Neurol 170:141-151.
Lal R, Friedlander MJ (1990) Effect of passive eye movement on retinogeniculate transmission
in the cat. J Neurophysiol 63:523-538.
Lam YW, Cox CL, Varela C, Sherman SM (2005) Morphological correlates of triadic circuitry
in the lateral geniculate nucleus of cats and rats. J Neurophysiol 93:748-757.
Lehmkuhle S, Kratz KE, Mangel SC, Sherman SM (1980) Effects of early monocular lid suture
on spatial and temporal sensitivity of neurons in dorsal lateral geniculate nucleus of the
cat. J Neurophysiol 43:542-556.
Lennie P, Perry VH (1981) Spatial contrast sensitivity of cells in the lateral geniculate nucleus of
the rat. J Physiol 315:69-79.
Lesica NA, Stanley GB (2004) Encoding of natural scene movies by tonic and burst spikes in the
lateral geniculate nucleus. J Neurosci 24:10731-10740.
LeVay S, Ferster D (1977) Relay cell classes in the lateral geniculate nucleus of the cat and the
effects of visual deprivation. J Comp Neurol 172:563-584.
Levine MW, Cleland BG (2001) An analysis of the effect of retinal ganglion cell impulses upon
the firing probability of neurons in the dorsal lateral geniculate nucleus of the cat. Brain
Res 902:244-254.

45
Levitt JB, Schumer RA, Sherman SM, Spear PD, Movshon JA (2001) Visual response properties
of neurons in the LGN of normally reared and visually deprived macaque monkeys. J
Neurophysiol 85:2111-2129.
Lu SM, Guido W, Sherman SM (1992) Effects of membrane voltage on receptive field properties
of lateral geniculate neurons in the cat: contributions of the low-threshold Ca2+
conductance. J Neurophysiol 68:2185-2198.
Mastronarde DN (1987) Two classes of single-input X-cells in cat lateral geniculate nucleus. II.
Retinal inputs and the generation of receptive-field properties. J Neurophysiol 57:381-
413.
Mastronarde DN (1989) Correlated firing of retinal ganglion cells. Trends Neurosci 12:75-80.
McCormick DA, Bal T (1994) Sensory gating mechanisms of the thalamus. Curr Opin Neurobiol
4:550-556.
Mioche L, Singer W (1989) Chronic recordings from single sites of kitten striate cortex during
experience-dependent modifications of receptive-field properties. J Neurophysiol 62:185-
197.
Montero VM (1986) Localization of gamma-aminobutyric acid (GABA) in type 3 cells and
demonstration of their source to F2 terminals in the cat lateral geniculate nucleus: a
Golgi-electron-microscopic GABA-immunocytochemical study. J Comp Neurol
254:228-245.
Montero VM (1987) Ultrastructural identification of synaptic terminals from the axon of type 3
interneurons in the cat lateral geniculate nucleus. J Comp Neurol 264:268-283.
Muir-Robinson G, Hwang BJ, Feller MB (2002) Retinogeniculate axons undergo eye-specific
segregation in the absence of eye-specific layers. J Neurosci 22:5259-5264.
Murphy PC, Sillito AM (1996) Functional morphology of the feedback pathway from area 17 of
the cat visual cortex to the lateral geniculate nucleus. J Neurosci 16:1180-1192.
Pinault D, Bourassa J, Deschenes M (1995) Thalamic reticular input to the rat visual thalamus: a
single fiber study using biocytin as an anterograde tracer. Brain Res 670:147-152.
Pizzorusso T, Medini P, Berardi N, Chierzi S, Fawcett JW, Maffei L (2002) Reactivation of
ocular dominance plasticity in the adult visual cortex. Science 298:1248-1251.
Pizzorusso T, Medini P, Landi S, Baldini S, Berardi N, Maffei L (2006) Structural and functional
recovery from early monocular deprivation in adult rats. Proc Natl Acad Sci U S A
103:8517-8522.
Rafols JA, Valverde F (1973) The structure of the dorsal lateral geniculate nucleus in the mouse.
A Golgi and electron microscopic study. J Comp Neurol 150:303-332.

46
Ramcharan EJ, Gnadt JW, Sherman SM (2000) Burst and tonic firing in thalamic cells of
unanesthetized, behaving monkeys. Vis Neurosci 17:55-62.
Reese BE (1988) 'Hidden lamination' in the dorsal lateral geniculate nucleus: the functional
organization of this thalamic region in the rat. Brain Res 472:119-137.
Reinagel P, Godwin D, Sherman SM, Koch C (1999) Encoding of visual information by LGN
bursts. J Neurophysiol 81:2558-2569.
Rittenhouse CD, Shouval HZ, Paradiso MA, Bear MF (1999) Monocular deprivation induces
homosynaptic long-term depression in visual cortex. Nature 397:347-350.
Rivadulla C, Martinez L, Grieve KL, Cudeiro J (2003) Receptive field structure of burst and
tonic firing in feline lateral geniculate nucleus. J Physiol 553:601-610.
Rodieck RW (1967) Maintained activity of cat retinal ganglion cells. J Neurophysiol 30:1043-
1071.
Rowe MH, Fischer Q (2001) Dynamic properties of retino-geniculate synapses in the cat. Vis
Neurosci 18:219-231.
Sale A, Maya Vetencourt JF, Medini P, Cenni MC, Baroncelli L, De Pasquale R, Maffei L
(2007) Environmental enrichment in adulthood promotes amblyopia recovery through a
reduction of intracortical inhibition. Nat Neurosci 10:679-681.
Sesma MA, Irvin GE, Kuyk TK, Norton TT, Casagrande VA (1984) Effects of monocular
deprivation on the lateral geniculate nucleus in a primate. Proc Natl Acad Sci U S A
81:2255-2259.
Sharma J, Angelucci A, Sur M (2000) Induction of visual orientation modules in auditory cortex.
Nature 404:841-847.
Shatz CJ (1996) Emergence of order in visual system development. Proc Natl Acad Sci U S A
93:602-608.
Sherman SM (2001) Tonic and burst firing: dual modes of thalamocortical relay. Trends
Neurosci 24:122-126.
Sherman SM (2004) Interneurons and triadic circuitry of the thalamus. Trends Neurosci 27:670-
675.
Sherman SM (2007) The thalamus is more than just a relay. Curr Opin Neurobiol 17:417-422.
Sherman SM, Sanderson KJ (1972) Binocular interaction on cells of the dorsal lateral geniculate
nucleus of visually deprived cats. Brain Res 37:126-131.
Sherman SM, Spear PD (1982) Organization of visual pathways in normal and visually deprived
cats. Physiol Rev 62:738-855.

47
Sherman SM, Guillery RW (2006) Exploring the thalamus and its role in cortical function, 2nd
Edition. Cambridge, MA: The MIT Press.
Sillito AM, Jones HE, Gerstein GL, West DC (1994) Feature-linked synchronization of thalamic
relay cell firing induced by feedback from the visual cortex. Nature 369:479-482.
Sincich LC, Adams DL, Economides JR, Horton JC (2007) Transmission of spike trains at the
retinogeniculate synapse. J Neurosci 27:2683-2692.
Singer W, Rauschecker J, Werth R (1977) The effect of monocular exposure to temporal
contrasts on ocular dominance in kittens. Brain Res 134:568-572.
Stryker MP, Harris WA (1986) Binocular impulse blockade prevents the formation of ocular
dominance columns in cat visual cortex. J Neurosci 6:2117-2133.
Sur M, Garraghty PE, Roe AW (1988) Experimentally induced visual projections into auditory
thalamus and cortex. Science 242:1437-1441.
Sur M, Esguerra M, Garraghty PE, Kritzer MF, Sherman SM (1987) Morphology of
physiologically identified retinogeniculate X- and Y-axons in the cat. J Neurophysiol
58:1-32.
Swadlow HA, Gusev AG (2001) The impact of 'bursting' thalamic impulses at a neocortical
synapse. Nat Neurosci 4:402-408.
Swadlow HA, Gusev AG, Bezdudnaya T (2002) Activation of a cortical column by a
thalamocortical impulse. J Neurosci 22:7766-7773.
Szentagothai J (1963) The Structure of the Synapse in the Lateral Geniculate Body. Acta Anat
(Basel) 55:166-185.
Tagawa Y, Kanold PO, Majdan M, Shatz CJ (2005) Multiple periods of functional ocular
dominance plasticity in mouse visual cortex. Nat Neurosci 8:380-388.
Tanaka K (1983) Cross-correlation analysis of geniculostriate neuronal relationships in cats. J
Neurophysiol 49:1303-1318.
Uhlrich DJ, Tamamaki N, Murphy PC, Sherman SM (1995) Effects of brain stem parabrachial
activation on receptive field properties of cells in the cat's lateral geniculate nucleus. J
Neurophysiol 73:2428-2447.
Usrey WM, Reppas JB, Reid RC (1998) Paired-spike interactions and synaptic efficacy of retinal
inputs to the thalamus. Nature 395:384-387.
Usrey WM, Reppas JB, Reid RC (1999) Specificity and strength of retinogeniculate connections.
J Neurophysiol 82:3527-3540.

48
Usrey WM, Alonso JM, Reid RC (2000) Synaptic interactions between thalamic inputs to simple
cells in cat visual cortex. J Neurosci 20:5461-5467.
Van Hooser SD, Heimel JA, Nelson SB (2003) Receptive field properties and laminar
organization of lateral geniculate nucleus in the gray squirrel (Sciurus carolinensis). J
Neurophysiol 90:3398-3418.
Van Horn SC, Erisir A, Sherman SM (2000) Relative distribution of synapses in the A-laminae
of the lateral geniculate nucleus of the cat. J Comp Neurol 416:509-520.
Veredas FJ, Vico FJ, Alonso JM (2005) Factors determining the precision of the correlated firing
generated by a monosynaptic connection in the cat visual pathway. J Physiol 567:1057-
1078.
von Melchner L, Pallas SL, Sur M (2000) Visual behaviour mediated by retinal projections
directed to the auditory pathway. Nature 404:871-876.
Wang S, Bickford ME, Van Horn SC, Erisir A, Godwin DW, Sherman SM (2001) Synaptic
targets of thalamic reticular nucleus terminals in the visual thalamus of the cat. J Comp
Neurol 440:321-341.
Wang X, Wei Y, Vaingankar V, Wang Q, Koepsell K, Sommer FT, Hirsch JA (2007)
Feedforward excitation and inhibition evoke dual modes of firing in the cat's visual
thalamus during naturalistic viewing. Neuron 55:465-478.
Weliky M, Katz LC (1999) Correlational structure of spontaneous neuronal activity in the
developing lateral geniculate nucleus in vivo. Science 285:599-604.
Weyand TG (2007) Retinogeniculate transmission in wakefulness. J Neurophysiol 98:769-785.
Weyand TG, Boudreaux M, Guido W (2001) Burst and tonic response modes in thalamic
neurons during sleep and wakefulness. J Neurophysiol 85:1107-1118.
Wiesel TN, Hubel DH (1963a) Single-Cell Responses in Striate Cortex of Kittens Deprived of
Vision in One Eye. J Neurophysiol 26:1003-1017.
Wiesel TN, Hubel DH (1963b) Effects of Visual Deprivation on Morphology and Physiology of
Cells in the Cats Lateral Geniculate Body. J Neurophysiol 26:978-993.
Wiesel TN, Hubel DH (1965) Comparison of the effects of unilateral and bilateral eye closure on
cortical unit responses in kittens. J Neurophysiol 28:1029-1040.
Wilson JR, Webb SV, Sherman SM (1977) Conditions for dominance of one eye during
competitive development of central connections in visually deprived cats. Brain Res
136:277-287.
Wilson JR, Friedlander MJ, Sherman SM (1984) Fine structural morphology of identified X- and
Y-cells in the cat's lateral geniculate nucleus. Proc R Soc Lond B Biol Sci 221:411-436.

49
Yoon BJ, Heynen AJ, Smith GB, Neve RL, Bear MF (2008) Role of AMPA receptor
endocytosis in visual cortical plasticity.
Zhou Y, Yu H, Yang Y, Shou T (2003) Non-dominant eye responses in the dorsal lateral
geniculate nucleus of the cat: an intracellular study. Brain Res 987:76-85.

50
Summary
The dLGN was classically thought to be a mere relay of visual information from the
retina to the primary visual cortex. However, the anatomical and physiological features of this
nucleus suggest that the signal is transformed from the eye to V1. This transformed signal serves
as the presynaptic input to V1 that drives OD plasticity under MC and MI conditions. While
assumptions regarding the nature of dLGN activity are made based on the nature of the visual
manipulation, no careful study of dLGN activity during MC and MI has been conducted. The
purpose of this thesis research is to examine dLGN activity during MC, MI and normal viewing
conditions to capture the activity that leads to deprived-eye depression. Additionally, we
compare the findings from juvenile and adult mice in an attempt to explain the differences in OD
plasticity as the animal matures.

Chapter 2
LGN ACTIVITY DURING OCULAR DOMINANCE
PLASTICITY IN THE JUVENILE MOUSE
Adapted from
Linden, M.L., Heynen, A.J., Haslinger, R.H., Bear, M.F (2008) Thalamic activity patterns during
ocular dominance plasticity. Submitted.


53
Abstract
Ocular dominance (OD) plasticity is a classic example of experience-dependent cortical
plasticity that results from a brief period of monocular lid closure (MC) during early postnatal
development (Wiesel and Hubel, 1963). The initial phase of the OD shift is characterized by a
decreased cortical response to input from the deprived eye (Mioche and Singer, 1989; Frenkel
and Bear, 2004). While this deprived-eye response depression was originally thought to be
caused by the general reduction of cortical input during MC, monocular retinal inactivation (MI),
which eliminates all retinal activity, does not yield this depression (Rittenhouse et al., 1999;
Frenkel and Bear, 2004). It has therefore been suggested that residual activity in the deprived
retina drives MC-induced cortical depression. This implies that dorsal lateral geniculate nucleus
(dLGN) activity during MC leads to cortical plasticity that subsequently produces changes in
visual function, while dLGN activity during MI does not. However, the actual patterns of dLGN
neural activity resulting in OD plasticity have yet to be characterized. Here we describe neural
activity patterns recorded extracellularly from dLGN of awake mice before and after MC and
MI. We had hypothesized that MC would produce uncorrelated dLGN activity (“noise”)
resulting from spontaneous retinal activity in the absence of visual input, whereas MI would be
accompanied by a marked reduction in dLGN activity. We have verified that dLGN activity
following MC is distinguished by a lack of patterned unit activity. During MI however, instead
of a reduction of neural activity, we unexpectedly found highly patterned activity which is
characterized by thalamic bursting and an increase in simultaneous firing. These findings
provide the first description of dLGN unit activity in the awake mouse that is known to lead to a
loss of visual function. Furthermore, we provide the first evidence that MI does not reduce

54
thalamic input to visual cortex. These findings have important implications for our
understanding of the mechanisms supporting experience-dependent cortical plasticity.
Introduction
Experience-dependent cortical plasticity is a well studied phenomenon of the mammalian
brain. While much is known about the biochemical, electrophysiological and anatomical
changes that occur in the cortex, far less is known about the input activity to cortex that drives
cortical plasticity in vivo. OD plasticity is a particularly well characterized example of
experience-dependent cortical plasticity. Brief periods of MC in young kittens shift OD in
primary visual cortex (V1) (Wiesel and Hubel, 1963), primarily due to a loss of visual
responsiveness to the deprived eye (Mioche and Singer, 1989). Although eyelid closure causes
deprived-eye response depression, completely blocking all input from the eye in kittens by MI
(via intraocular injection of tetrodotoxin, TTX) does not lead to a robust OD shift (Rittenhouse et
al., 1999). Therefore, it has been concluded that OD plasticity requires a basal level of retinal
activity. These findings have recently been replicated using local field potential recordings in
V1 of awake juvenile mice (Frenkel and Bear, 2004). These studies confirm that the OD shift
following MC initially results from depression of the deprived-eye response and that this
depression does not occur following MI.
Since visual input travels from the retina to the dLGN and then into V1, we can conclude
that the patterns or amount of dLGN activity during MC trigger deprived-eye response
depression in vivo, while the patterns and amount of dLGN activity during normal vision (NV)
and MI do not. Although these activity patterns are crucial for understanding how OD plasticity
is initiated in visual cortex, they are essentially unknown. It has been inferred from a limited
number of studies that dLGN activity during deprivation approximates that in the retina (Stryker

55
and Harris, 1986; Kaplan et al., 1987; Rittenhouse et al., 1999). Thus, it has been assumed that
(1) during NV, images formed on the retina translate into spatially and temporally patterned
dLGN activity, (2) patterned activity is lost during MC, leaving only a low level of residual
spontaneous activity, and (3) the absence of retinal activity during MI substantially reduces or
eliminates activity in the dLGN. However, the experiments supporting these assumptions have
all been performed in the paralyzed and anesthetized preparation that does not support OD
plasticity (Freeman and Bonds, 1979). Therefore, the goal of the current study was to understand
how dLGN activity varies under the conditions that trigger rapid shifts in OD in vivo. We chose
to examine this question in the mouse because this species is amenable to awake recordings and
is now preferred for mechanistic studies.
Methods Summary
On postnatal day (P) 25, male mice were anesthetized and surgically prepared for head
restraint and electrophysiological recordings (see Detailed Methods). The animals were
behaviorally habituated to restraint over the next several days, and bundles of electrodes were
inserted in the dLGN at the age of maximal sensitivity to MC (~P28) (Gordon and Stryker,
1996). The location of the recording bundle in the dLGN was initially determined by
observation of visually-driven unit activity in response to stimulation of the contralateral eye,
and was always subsequently confirmed histologically (Fig. 2-1). Baseline recordings were
made with the eye contralateral to the electrode viewing and the ipsilateral eye occluded.
Activity approximating that during NVE was recorded in response to phase-reversing sinusoidal
gratings and natural scene stimuli (see Methods). Unless otherwise indicated, results using
grating stimuli are only illustrated since results from natural scenes did not differ qualitatively.
Following the baseline recording session, animals were briefly anesthetized and either eyelid

56
closure (MC), TTX injection (MI), or no manipulation (control), was performed. After 30
minutes of recovery from anesthesia, stimuli were presented for a second recording session. For
the MI group, effectiveness of TTX was assessed by observing pupilary dilation (Fig. 2-1b) both
before and after the second recording session. At the conclusion of recordings, an electrolytic
lesion was made at the recording site to allow for histological confirmation of electrode
placement (Fig. 2-1d).
Results
Figure 2-2 shows the responses of one representative neuron from each experimental
group. In the baseline condition, each neuron shows a visual response to the grating stimulus as
indicated by an increase in the number spikes immediately following the phase reversal. As
expected, the neuron from the control group shows a qualitatively similar visually-driven
response during both recording sessions (Fig. 2-2a). The neuron from the MC group (Fig. 2-2b)
is no longer visually driven after eyelid closure, but continues to show a high level of
spontaneous activity. Most surprising however, is the activity of the neuron from the MI group
(Fig. 2-2c) which does not show an absence or even a reduction of neural activity following the
TTX injection. Rather, this neuron shows a high level of spontaneous activity after retinal
inactivation, similar to what is seen from the MC neuron.
To quantify these observations, we measured the overall firing rate during the entire
recording session before and after eye manipulation (Fig. 2-3). We found no significant change
in firing rate across any of the experimental groups (p > 0.5, Kruskal-Wallis). Since
measurements of overall firing rate can be considered crude measures of neural activity, as they
do not take into account changes in the temporal pattern of the neural spike train, we next
examined the distribution of inter-spike intervals (ISIs) before and after eye manipulation.

57
Figure 2-4a,b show that there is no change in the ISI distribution in the control or the MC groups
(control: p > 0.2, MC: p > 0.05, Kolmogorov-Smirnov Bootstrap (KS)). In contrast, the MI
group shows a leftward shift in the ISI distribution (Fig. 2-4c), indicating an increased
probability of observing very short ISIs (p < 10-9
, KS). Indeed this corresponds to an increased
probability of observing ISIs from 2 – 4 ms (Fig. 2-4d).
Activity exhibiting very short ISIs is reminiscent of the phenomenon of bursting, which is
common in thalamic relay cells (Sherman, 2001). The criteria for bursts consist of an initial
period of quiescence (> 100 ms) followed by two or more spikes with ISIs < 4 ms (Lu et al.,
1992) (Fig. 2-5a). While originally thought to occur only during sleep states, bursts have now
been shown to occur in awake animals including cats (Guido and Weyand, 1995) and primates
(Ramcharan et al., 2000). Bursts in the dLGN have been related to visual processing, as the
percentage of spikes in bursts increases during natural scene viewing as compared to white-noise
viewing (Lesica and Stanley, 2004). However, bursting has not previously been documented in
the awake mouse preparation.
Since we observed an increase in very short ISIs, we next quantified bursting activity.
Figure 2-5b shows the occurrence of bursts on raster plots from one representative neuron in
each of the three experimental conditions. We observed a small, comparable percentage of burst
spikes in the control and MC conditions, with some of the control bursts apparently related to
visual stimulus presentation. However, as suggested by the shift in ISIs, we found a large
increase in the percentage of spikes in bursts after retinal inactivation. Group analysis (Figure 2-
5c) demonstrated no change in overall bursting in the control and MC groups (p > 0.7 and p >
0.2 respectively, Wilcoxon Sign-Rank (WSR)) but a large increase in the percentage of spikes in
bursts following MI (p < 10-3
, WSR).

58
Because we utilized an acute recording paradigm, it is possible that the observed bursting
was only a transient feature of dLGN firing activity. To address this concern, we chronically
implanted the recording bundle and sampled unit activity throughout the period of retinal
inactivation. Bursting activity persisted throughout the period of MI, up to at least 48 hours (Fig.
2-6). Additionally, the burst percentage returned to baseline values after the TTX washed out, as
confirmed by pupil constriction and a return of visual response (Fig. 2-6d,e).
Our observation of an increase in burst spikes during MI was entirely unexpected.
Although bursting following retinal inactivation or eye enucleation has been described in the
immature ferret dLGN prior to natural eye-opening (Weliky and Katz, 1999), it has long been
assumed that by adolescence, monocular TTX treatment simply reduces activity in the central
visual system. Moreover, it has been suggested that bursts carry specific information about
patterned visual input in mature animals (Lesica and Stanley, 2004). Thus, it was equally
surprising to find that there was no significant decrease in bursts following MC compared to the
NV condition in the adolescent mice.
Occasionally we recorded from the ipsilateral core of the dLGN instead of our intended
target of the contralateral shell. Remarkably, even though the retinal input to this brain area had
not been changed, we found robust changes in the firing properties of these neurons including an
increase in firing rate (p < 0.02, WSR) and an increase in burst percentage (p < 0.01, WSR) (Fig.
2-7).
None of the properties of individual spike trains that we examined differentiated NVE
and MC. Therefore, we next looked to the correlative firing patterns between simultaneously
recorded neurons for an explanation of what triggers OD plasticity. Figure 2-8a shows example
cross-correlograms representing pairs of simultaneously recorded neurons from each of the three

59
experimental groups. Scatter plots showing the amount of correlative firing before and after
visual manipulation are shown in figure 2-8b, with the data summarized in figure 2-8c. Unlike
the control and MI groups, there was a decrease in the simultaneous firing between dLGN
neurons for nearly every pair of neurons following MC (p < 10-3
, WSR).
In the MI group, however, we observed an increase in simultaneous firing (p < 0.01,
WSR). To determine if this increase was a direct result of the increase in burst spikes, we
analyzed the correlative firing of just the initial spikes in the bursts, as well as the correlations
with all burst spikes removed (Fig. 2-8d). The initial bursts spikes were significantly correlated
(p < 0.03) while the non-burst spikes were not (p > 0.9). This implies that the increased in
bursting accounts for the increase in simultaneous firing following MI.
Discussion
MC and MI have very different effects of dLGN firing activity. While neither
manipulation causes a decrease in the amount of firing, MC results in a decrease of correlative
firing between pairs of simultaneously recorded neurons, and MI causes an increase in thalamic
bursting. These results provide insights into the mechanisms which drive OD plasticity.
It has been known for decades that the change in dLGN activity during MC causes rapid
depression of deprived-eye responses in the visual cortex. Theoretically, this could occur as a
consequence of either a decrease or a de-correlation of deprived-eye input to cortex relative to
the seeing eye (Blais et al., 1999). This distinction is important for understanding cortical
plasticity. The former explanation requires a mechanism whereby active inputs “punish”
inactive inputs (heterosynaptic depression); the latter explanation requires a mechanism by
which the activity of poorly correlated inputs triggers their own demise (homosynaptic
depression). Our data reveal no difference in the amount or temporal structure of dLGN activity

60
following eyelid closure. Instead, our findings support the “homosynaptic” hypothesis that
poorly correlated dLGN activity is the trigger for deprived-eye depression in visual cortex. This
supports the recent finding that monocular blur is as effective as MC in driving OD plasticity
(Rittenhouse et al., 2006).
The synaptic depression observed following MC has now been shown to employ
mechanisms of homosynaptic long-term depression (LTD). Surface AMPA receptor (AMPAR)
expression decreases following both brief periods of MC and the induction of LTD (Heynen et
al., 2003). Blocking AMPAR endocytosis in vivo prevents OD plasticity (Yoon et al., 2008).
Also, MC occludes subsequent expression of LTD ex vivo (Heynen et al., 2003). In contrast, MI,
which does not lead to deprived-eye depression, does not engage LTD-like mechanisms (Heynen
et al., 2003). Therefore, we can conclude that patterns of dLGN activity during MC lead to OD
plasticity and LTD in vivo, while patterns of dLGN activity during MI do not. Having
characterized the activity patterns which drive synaptic depression may lead to the development
of novel LTD induction protocols which will allow us to better model cortical plasticity in vitro.
One methodological concern related to the observation of a decrease in correlative firing
stems from the choice of visual stimuli. Because the animals viewed a full-screen sinusoidal
grating, there may have been an artificial increase in simultaneous firing during baseline
viewing, as neurons with non-overlapping receptive fields may be activated together. To address
this concern, the animals were also shown “natural scene” images, in the hopes of better
approximating the types of visual input the animal would be exposed to during a period of MC
that leads to an OD shift. The maintenance of overall dLGN firing activity under all viewing
conditions, the increase in burst spikes during MI, and the decrease in simultaneous firing during
MC all hold when the animal is viewing the natural scene stimuli (Fig. 2-9). This suggests that

61
deprived-eye depression due to homosynaptic mechanisms is in fact what occurs in V1 during
standard OD plasticity protocols.
Our data challenge the interpretation of previous experiments that used MI as a means of
distinguishing between homosynaptic and heterosynaptic hypotheses. The fact that MI induces
substantially less deprived-eye depression than MC (Rittenhouse et al., 1999; Heynen et al.,
2003; Frenkel and Bear, 2004) was taken as evidence that dLGN activity is necessary to trigger
synaptic modifications in visual cortex. Instead of reduced dLGN activity after MI, however, we
find that dLGN neurons are as active and correlated as during normal visual experience, and fire
substantially more spikes in bursts. This finding contrasts with previous reports that MI
decreases dLGN firing rate in the anesthetized kitten (Stryker and Harris, 1986; Rittenhouse et
al., 1999). To determine if this discrepancy is due to the use of anesthesia, we performed MI in
anesthetized mice. Indeed, compared to baseline, a significant decrease in firing rate is observed
following MI in this preparation (Fig. 2-10a,b). Furthermore, we see a substantial increase in the
amount of burst spikes during the baseline condition under anesthesia (Fig. 2-10c). This may
explain the discrepancy between the results from anesthetize cat recordings, where bursts are
more prevalent during natural scene viewing as opposed to white noise (Lesica and Stanley,
2004), and our results where lid closure (presumably resulting in a more “noisy” retinal
response) had no effect on burst percentage. These analyses underscore the importance of using
an awake preparation when determining the patterns of activity which drive cortical plasticity.
Certain behavioral states are correlated with an increase in burst percentage (e.g.
Bezdudnaya et al., 2006). Because we saw an increase in burst percentage following MI, it is
necessary to confirm that this increase was not due to a change in behavioral state. To address
this concern, we analyzed the cortical EEG recorded from a subset of mice in the control and MI

62
groups. Power spectral analysis of EEG recorded from occipital cortex was performed in 10 s
bins using the Welch method of spectral estimation. We defined the delta band as frequencies
below 4 Hz and the theta band as frequencies between 4.5 and 9 Hz. We used the theta /delta
ratio (TDR) as our measure of behavioral state (Maloney et al., 1997). We found no change in
behavioral state following MI, nor did we find a correlation between TDR and the number of
observed bursts (See Appendix 2).
How can we account for the remarkable activity in the dLGN postsynaptic to the inactive
retina? The bursting activity in the dLGN after MI resembles that observed during sleep in this
brain region. Such bursts result from the activation of T-type Ca2+
channels, which require a
long period of hyperpolarization for deinactivation (Sherman, 2001). In the absence of retinal
input, these channels may have more opportunity for deinactivation. Since retinal inputs
comprise such a small percentage of the total input into the dLGN (Sherman and Guillery, 1998),
the input from the other sources, including feedback input from V1, could depolarize dLGN
neurons above the activation potential for Ca2+
channels, which would in turn lead to burst
spikes.
Our results provide a surprising alternate explanation for the lack of deprived-eye
depression following MI. Instead of an absence of input preventing depression, the active,
patterned firing of dLGN neurons may protect against deprived-eye depression by substituting
for normal visual experience. Furthermore, prolonged MI leads to potentiation of the ipsilateral
response (Frenkel and Bear, 2004). The bursting of the ipsilateral core may be the activity that
underlies the potentiation of that connection. While this change in firing properties is surprising
given the lack of change in the retinal input, local dLGN circuitry, intrathalamic circuitry via the
thalamic reticular nucleus, or corticothalamic feedback may be responsible for the change.

63
The dramatically different consequences on visual cortex of deprivation by eyelid closure
or image blur (Rittenhouse et al., 2006) and retinal inactivation (Freeman and Bonds, 1979;
Rittenhouse et al., 1999; Heynen et al., 2003; Frenkel and Bear, 2004) are accounted for by
equally dramatic differences in dLGN activity. Characterization of the activity patterns which
drive homosynaptic depression will allow us to better model and more precisely determine the
mechanism of cortical plasticity. This new knowledge may aid in identifying new treatment
options for amblyopia.


65
Figure 2-1
Figure 2-1 Methodology.
a, Experimental setup. Data were obtained from awake, head-restrained mice viewing grating
stimuli. b, Intraocular delivery of tetrodotoxin (TTX) was confirmed by pupilary dilation. Left
column: non-injected eye, right column: TTX-injected eye. Lower row: circles outline the pupil.
c, Schematic of recording electrode placement in the dLGN. d, Histological confirmation of
recording electrode track in dorsal LGN. Arrow indicates site of electrolytic lesion (10 µA; 10 s)
made at the end of recording session.


67
Figure 2-2
Figure 2-2 dLGN responses before and after visual manipulation.
a – c, Peristimulus time histograms and raster plots from representative neurons for each
experimental group (80 stimulus trials) are presented. Stimuli were presented at 0° or 90°, 1 Hz
phase reversing (▼). Arrowheads in this and subsequent figures indicate time of stimulus phase
reversal. Spike waveforms are recording session averages. Scale bar: 100 µV, 500 µs. Left
column data were obtained during baseline, right column after eye manipulation. Neuron (a) is
from the control group, (b) MC group, and (c) MI group. Note the loss of visual responsiveness
with MC and MI, and that the total number of spikes recorded is comparable before and after
each manipulation.


69
Figure 2-3
Figure 2-3 Firing rate before and after visual manipulation.
Monocular lid closure and inactivation have no effect on dLGN firing rate (p > 0.5, Kruskal-
Wallis (KW)). Firing rate was calculated for the full duration of visual stimulation across all
stimulus conditions. Connected gray circles represent the same neuron recorded before and after
eye manipulation ((O)pen, (C)losed, (I)nactived). Black lines indicate median values (control: n
= 22 neurons (9 animals), p > 0.2 Wilcoxon Sign-Rank (WSR); MC: n = 24 neurons (12
animals), p > 0.3; MI: n = 19 neurons (8 animals), p > 0.3). See also Fig. 2-9a.


71
Figure 2-4
Figure 2-4 Interspike interval distributions before and after visual manipulation.
ISI distributions are not affected in control (a) and MC groups (b). Plots represent mean
cumulative ISI distribution during baseline (black), and after eye manipulation (gray) (control: p
> 0.1 Kolmogorov-Smirnov Bootstrap (KS); MC: p > 0.08). c – d, MI increases the probability
of observing short ISIs. c, cumulative probability distributions for MI as described in (a,b) (p <
10-5
). d, Probability density function; the curves differ significantly from 2 - 4 ms (p < 0.01
WSR).


73
Figure 2-5
Figure 2-5 Monocular inactivation increases the percentage of spikes in bursts.
a, Schematic of burst firing. A burst is defined by at least 100 ms of quiescence followed by two
or more spikes with ISIs < 4 ms. b, Raster plots of 80 stimulus trials from neurons nearest the
median in each experimental group. Stimuli were presented at 0° or 90°, 1 Hz phase reversing
(▼). Black squares represent spikes in bursts; gray squares non-burst spikes. Note bursts on the
leading edge of the visual response in the open eye condition and the marked increase in bursts
in the MI condition. c, MI increases the percentage of spikes in bursts (p < 10-3
KW; MI n = 19
neurons (8 animals), p < 10-3
WSR). Connected gray circles represent the same neuron recorded
before and after eye manipulation. Black lines indicate the median values (control: n = 22
neurons (9 animals), p > 0.7; MC: n = 24 neurons (12 animals), p > 0.2). See also Fig. 2-9b.


75
Figure 2-6
Figure 2-6 An increase in the percentage of spikes in bursts persists throughout an extended
period of monocular inactivation.
a – d, Raster plots of 80 stimulus trials from representative neurons prior to, during and
following MI. Stimuli were presented at 0° or 90°, 1 Hz phase reversing (▼). Black squares
represent spikes in bursts; gray squares non-burst spikes. a, baseline condition 2 hrs prior to
TTX injection; b – c, 24 and 48 hrs after TTX injection, respectively; d, recovery from MI, 120
hrs after injection. Note that the neurons are visually responsive only during the baseline and
recovery conditions (a, d), and the marked increase in bursts throughout the period of
inactivation (c – d). e, The percentage of spikes in bursts remains elevated at least 48 hrs
following TTX injection and returns to baseline values following recovery from MI (KW effect
of time, p < 0.01). Gray circles represent individual neurons. Black lines indicate the median
values. At 2, 24 and 48 hrs, the percentage of spikes in bursts is significantly different from both
the baseline and recovery (120 hrs) time points (p < 0.05 for all comparisons, Mann-Whitney U-
test (MWU)). The baseline and recovery time points are not significantly different (p > 0.7,
MWU), nor are the time points during MI (p > 0.4 for all comparisons, MWU).


77
Figure 2-7
Figure 2-7 Inactivation of the contralateral eye changes the firing patterns of the dLGN
ipsilateral core.
a, Raster plots of 80 stimulus trials from neurons nearest the median in each experimental group.
Stimuli were presented at 0° or 90°, 1 Hz phase reversing (▼). Black squares represent spikes in
bursts; gray squares non-burst spikes. Note bursts on the leading edge of the visual response in
when the ipsilateral (center panel) but not contralateral (left panel) eye is viewing. Following MI
of the contralateral eye (right panel), a visual response is maintained in addition to the marked
increase in bursts and overall firing. b, Contralateral MI increases dLGN firing in the ipsilateral
core (n = 9 neurons (4 animals), p < 0.02). Connected gray circles represent the same neuron
recorded before and after eye manipulation. Black lines indicate the median values. c,
Contralateral MI increases the percentage of spikes in bursts in the ipsilateral core (n = 9 neurons
(4 animals), p < 0.01 WSR).


79
Figure 2-8
Figure 2-8 Monocular lid closure and monocular inactivation have opposite effects on
correlative dLGN firing.
a, Cross-correlograms from three pairs of simultaneously recorded neurons before (black line)
and after (gray line) visual manipulation. b, Scatter plots of the area under the cross-
crosscorrelogram before and after visual manipulation for each pair of simultaneously recorded
neurons. Gray line represents unity. Note that following MC (center panel) nearly all points fall
below the unity line, indicating a decrease in correlative firing. c, MC and MI have opposite
effects on spike correlation (p < 10-4
KW). Bars represent the median change in area under the
peak of the cross-correlogram (± 10 ms) following visual manipulation. Error bars show the
interquartile range. MC and MI induce significant changes in correlation (control: n = 22 neuron
pairs (6 animals), p > 0.2 WSR; MC: n = 20 neuron pairs (6 animals), p < 10-3
; MI: n = 18
neuron pairs (6 animals), p < 0.01). See also Fig. 2-9c. d, Bursts contribute to increased
correlation following MI. Data are represented as in c. Cross-correlograms using all the data
are plotted in the “All Spikes” bar which recapitulates I from c. “Bursts” are cross-correlograms
of only the first spikes from all bursts (n = 18 neuron pairs (6 animals), p < 0.03). “Non-Bursts”
are cross-correlograms of all spikes not contained in bursts (p > 0.9).


81
Figure 2-9
Figure 2-9 The effects of monocular lid closure and monocular inactivation on dLGN activity
when assessed using natural scene stimuli are comparable to that observed using phase-reversing
sinusoidal grating stimuli.
a, Monocular lid closure (C) and inactivation (I) have no effect on dLGN firing rate (p > 0.7,
KW). Connected gray circles represent the same neuron recorded before and after eye
manipulation. Black lines indicate the median values (control: n = 22 neurons (9 animals), p >
0.3 WSR; MC: n = 24 neurons (12 animals), p > 0.8; MI: n = 19 neurons (8 animals), p > 0.5).
b, MI increases the percentage of spikes in bursts (p < 0.01 KW; control: n = 22 neurons (9
animals), p > 0.8; MC: n = 24 neurons (12 animals), p > 0.8; MI: n = 19 neurons (8 animals), p <
0.01 WSR). c, MC leads to a decrease in spike correlation (p < 10-4
, KW; MC: n = 20 neuron
pairs (6 animals), p < 10-3
WSR). Bars represent the median change in area under the cross-
correlogram following visual manipulation. Error bars show the interquartile range (control: n =
22 neuron pairs (6 animals), p > 0.9; MI: n = 18 neuron pairs (6 animals), p < 0.02).


83
Figure 2-10
Figure 2-10 Monocular inactivation decreases firing rate if the animal is under Nembutal
anesthesia.
a, Peristimulus time histograms and raster plots (80 stimulus trials) for one representative neuron
are presented. Stimuli were presented at 90°, 1 Hz phase reversing (▼). Spike waveforms are
averages over the entire recording session. Scale bar: 100 mV, 500 µs. Left panel: responses
during baseline, right panel: after MI under barbiturate anesthesia. b, Neuronal firing rate
decreases after MI. Connected gray circles represent the same neuron recorded before and after
manipulation ((O)pen, (I)nactivated). Black lines indicate the median values (n = 9 neurons (3
animals), p < 0.05 WSR). c, Percentage of spikes in bursts decreases after MI (p < 0.03). Notice
that during baseline, burst spikes are much more prevalent under barbiturate anesthesia than in
the awake mouse preparation (see Fig. 2-5c).


85
Detailed Methods
Subjects
Juvenile, male C57/BL6 mice (Charles River Laboratories) were group housed, on a 12
hr/12 hr light/dark cycle, with food and water available ad libitum. All animals were treated
according to NIH and MIT guidelines for animal use.
Surgical preparation for acute in vivo recording
P25 animals were anesthetized with 50 mg/kg ketamine and 10 mg/kg xylazine (i.p.).
Using cyanoacrylate, a fixation post was attached to the skull anterior to bregma. The skull
above the dLGN (2.0 mm posterior to bregma, 2.0 mm lateral to the midline) was demarcated for
future acute recording and the skull surrounding the location was encircled by a plastic ring
affixed with cyanoacrylate. An EEG electrode was placed in occipital cortex, and a reference
electrode was placed in frontal cortex. Electrodes were secured with cyanoacrylate and dental
cement was used to cover the entire skull exposure outside of the plastic ring.
Following surgery animals were monitored for signs of infection or discomfort.
Habituation to the restraint apparatus began ≥ 24 hours post-recovery. Animals remained in the
restraint system for ≥ 3 habituation sessions lasting ≥ 30 minutes each prior to the acute
recording session.
Acute recording
Animals were placed in the restraint apparatus and anesthetized with Isoflurane (1.5-
3.0% in 100% oxygen). A craniotomy was performed over the dLGN of one hemisphere.
Isoflurane was discontinued and the animal was allowed to recover from anesthesia while
remaining in the restraint system. A recording bundle consisting of seven 0.009” (22.86 μm)
diameter microwires was lowered into the dLGN. Microwires were 99.95% tungsten with

86
Formvar coating (California Fine Wire Co., Grover Beach, CA). Bundles were hand-made and
surrounded by a fine coat of cyanoacrlate to prevent splaying. Visually-driven unit activity was
used to aid in the placement of the bundle. Single unit activity and simultaneous EEG activity
were recorded using “Recorder” system software (Plexon, Inc., Dallas, TX). Single units were
discriminated offline using “Offline Sorter” (Plexon, Inc.) following methods adapted from
Csicsvari et al. (1998). The digital signal was high-pass filtered at 300 Hz. A recording
threshold was manually placed at a level to minimize noise without losing spikes. Spike
waveforms were sampled at 40 kHz and extended from 500 μs prior to threshold crossing to
1000 μs after crossing. Files recorded before and after visual manipulation were combined prior
to spike sorting to “blind” the user to the condition of each spike. Spike waveforms were aligned
to the peak. Principle component analysis (PCA) was applied to the waveform shapes.
Graphically, units were identified in either two or three dimensions using the first three principle
components, and hand-drawn polygons were used to define the cluster borders. Single units
were required to show refractory periods of at least 1 ms. In order to confirm the same unit was
recorded during the baseline and post-manipulation recording session, the cluster was visualized
in three dimensions using the first two principal components and time. Clusters that were not
stable through time were not included in further analysis. In experiments where the animals
were anesthetized throughout the recording session (see Fig. 2-10) Nembutal was administered
s.c. (100 mg/kg).
Chronic recordings
Once the recording bundle was placed in the contralateral shell of the dLGN (as
described above), it was affixed in place with dental cement to allow for chronic recordings.

87
Stimulus delivery
The visual stimuli were generated using custom MATLAB software (The Mathworks,
Natick, MA) and the Psychophysics Toolbox function set (psychtoolbox.org). The video
monitor, suitably linearized by -correction, was positioned 16 cm from the subjects’ eyes and
centered on the midline, occupying 82.5° x 100° of the visual field. Visual stimuli consisted of
full-screen sinusoidal gratings (0.05 cycles/°, 100% contrast), alternating in phase (phase
reversed) at a temporal frequency of 1, 2 and 4 Hz in both horizontal and vertical orientations.
Animals also viewed natural scene movies (excerpts from Microcosmos: Le peuple de l’herbe,
presented in grayscale, consisting of one 30s segment shown 10 times and one 4 minute segment
shown twice) between presentations of the sinusoidal gratings, with all stimuli presented in a
pseudorandom fashion.
Eyelid closure
Mice were anesthetized by inhalation of Isoflurane (1.5 – 3.0% in 100% oxygen).
Eyelids were held closed with Vetbond tissue adhesive (3M, St. Paul, MN).
Acute TTX injection
Mice were anesthetized by inhalation of Isoflurane (see above). A small puncture was
made in the vitreous chamber using a 30-guage needle. TTX (1 µL, 1 mM, Sigma, St. Louis,
MO) was then injected into the vitreous chamber using a microsyringe (10 µl, Hamilton Co,
Reno, NV). Following syringe withdrawal, the eye was rinsed with sterile eye drops. The
efficacy of retinal blockade by TTX was confirmed by a tonic and fully dilated pupil.
Chronic TTX injection
Under administration of Isoflurane anesthesia (1.5 – 3.0% in 100% oxygen), the superior
part of the conjunctiva was exposed, and the eyeball was stabilized with a 7-0 silk suture.

88
Ophthalmic ointment was applied to keep the eye moist. A small puncture was made in the
vitreous chamber using a 30-guage needle. A glass micropipette attached to a microinjection
apparatus (MMP, World Precision Instruments, Sarasota, FL) was inserted. TTX (300 nL, 2.85
mM, Sigma, St. Louis, MO, in NeuroSeal, NeuroNexus Technologies, Ann Arbor, MI) was
injected into the vitreous chamber. Calcium chloride was then applied at the insertion site to
polymerize the NeuroSeal. Following micropipette withdrawal, the eye was rinsed with sterile
eye drops.
Comparison of pre- and post-manipulation firing rates and burst percentages
Three groups of neurons were recorded under baseline and post-manipulation conditions.
To determine the statistical significance of any manipulation-induced firing rate or burst
percentage changes a two step procedure was used. We first tested if the median firing rate
change differed significantly between groups using a non-parametric Kruskal-Wallis (KW) test.
Second, we tested if the median firing rate change for each of the groups was significantly
different from zero using non-parametric Wilcoxon Sign-Rank (WSR) tests. Non-parametric
tests were used as the data were not normally distributed.
Comparison of pre- and post-manipulation ISI distributions
Mean ISI distributions for each group and condition were constructed using a
bootstrapping method and compared using Kolmogorov-Smirnov (KS) tests. First, for each
group, a neuron was randomly selected with replacement. Next an ISI was randomly selected
with replacement from the set of all ISIs recorded from that neuron. This procedure was
repeated N times where N was the mean number of ISIs recorded from all neurons in the group.
The resulting set of ISIs, a random sample of the group’s mean ISI distribution, was subjected to
a KS test. This procedure was repeated 1000 times, and the mean of the p-value recorded.

89
Comparison of pre- and post-manipulation pairwise correlation
To determine if correlated firing, beyond that predicted by chance, existed between pairs
of neurons, the Bernoulli correlation coefficient between the neurons’
spike trains was calculated (Aertsen et al., 1989). ( ) is the firing probability per bin of
neuron 1 (2) and the probability of joint firing at time lag . Positive (or negative)
departures of from zero indicate synchronous (or anti-synchronous) firing. To check the
statistical significance of manipulation induced correlation changes, the change in across each
lag was calculated and summed over lags from -10 ms to 10 ms for each pair of neurons before
and after manipulation. KW and WSR tests were then applied as above.
References
Aertsen AM, Gerstein GL, Habib MK, Palm G (1989) Dynamics of neuronal firing correlation:
modulation of "effective connectivity". J Neurophysiol 61:900-917.
Bezdudnaya T, Cano M, Bereshpolova Y, Stoelzel CR, Alonso JM, Swadlow HA (2006)
Thalamic burst mode and inattention in the awake LGNd. Neuron 49:421-432.
Blais BS, Shouval HZ, Cooper LN (1999) The role of presynaptic activity in monocular
deprivation: comparison of homosynaptic and heterosynaptic mechanisms. In: Proc Natl
Acad Sci U S A, pp 1083-1087.
Csicsvari J, Hirase H, Czurko A, Buzsaki G (1998) Reliability and state dependence of
pyramidal cell-interneuron synapses in the hippocampus: an ensemble approach in the
behaving rat. Neuron 21:179-189.
Freeman RD, Bonds AB (1979) Cortical plasticity in monocularly deprived immobilized kittens
depends on eye movement. In: Science, pp 1093-1095.
Frenkel MY, Bear MF (2004) How monocular deprivation shifts ocular domiance in visual
cortex of young mice. Neuron 44:917-923.
Gordon JA, Stryker MP (1996) Experience-dependent plasticity of binocular responses in the
primary visual cortex of the mouse. J Neurosci 16:3274-3286.
Guido W, Weyand T (1995) Burst responses in thalamic relay cells of the awake behaving cat. J
Neurophysiol 74:1782-1786.

90
Heynen AJ, Yoon BJ, Liu CH, Chung HJ, Huganir RL, Bear MF (2003) Molecular mechanism
for loss of visual cortical responsiveness following brief monocular deprivation. Nat
Neurosci 6:854-862.
Kaplan E, Purpura K, Shapley RM (1987) Contrast affects the transmission of visual information
through the mammalian lateral geniculate nucleus. J Physiol 391:267-288.
Lesica NA, Stanley GB (2004) Encoding of natural scene movies by tonic and burst spikes in the
lateral geniculate nucleus. J Neurosci 24:10731-10740.
Lu SM, Guido W, Sherman SM (1992) Effects of membrane voltage on receptive field properties
of lateral geniculate neurons in the cat: contributions of the low-threshold Ca2+
conductance. J Neurophysiol 68:2185-2198.
Maloney KJ, Cape EG, Gotman J, Jones BE (1997) High-frequency gamma
electroencephalogram activity in association with sleep-wake states and spontaneous
behaviors in the rat. Neuroscience 76:541-555.
Mioche L, Singer W (1989) Chronic recordings from single sites of kitten striate cortex during
experience-dependent modifications of receptive-field properties. J Neurophysiol 62:185-
197.
Ramcharan EJ, Gnadt JW, Sherman SM (2000) Burst and tonic firing in thalamic cells of
unanesthetized, behaving monkeys. Vis Neurosci 17:55-62.
Rittenhouse CD, Shouval HZ, Paradiso MA, Bear MF (1999) Monocular deprivation induces
homosynaptic long-term depression in visual cortex. Nature 397:347-350.
Rittenhouse CD, Siegler BA, Voelker CC, Shouval HZ, Paradiso MA, Bear MF (2006) Stimulus
for rapid ocular dominance plasticity in visual cortex. J Neurophysiol 95:2947-2950.
Sherman SM (2001) Tonic and burst firing: dual modes of thalamocortical relay. Trends
Neurosci 24:122-126.
Sherman SM, Guillery RW (1998) On the actions that one nerve cell can have on another:
distinguishing "drivers" from "modulators". Proc Natl Acad Sci U S A 95:7121-7126.
Stryker MP, Harris WA (1986) Binocular impulse blockade prevents the formation of ocular
dominance columns in cat visual cortex. J Neurosci 6:2117-2133.
Weliky M, Katz LC (1999) Correlational structure of spontaneous neuronal activity in the
developing lateral geniculate nucleus in vivo. In: Science, pp 599-604.
Wiesel TN, Hubel DH (1963) Single-Cell Responses in Striate Cortex of Kittens Deprived of
Vision in One Eye. J Neurophysiol 26:1003-1017.
Yoon BJ, Heynen AJ, Smith GB, Neve RL, Bear MF (2008) Role of AMPA receptor
endocytosis in visual cortical plasticity.

Chapter 3
LGN ACTIVITY DURING OCULAR DOMINANCE
PLASTICITY IN THE ADULT MOUSE


93
Abstract
The juvenile brain is generally considered to be more plastic than the brain of an adult.
Binocular neurons in the primary visual cortices (V1) of young animals lose their responsiveness
to an eye deprived of vision by monocular lid closure (MC) (Wiesel and Hubel, 1963). This
ocular dominance (OD) plasticity is a classic example of experience-dependent cortical plasticity
that is traditionally thought to occur only in juvenile animals (Hubel and Wiesel, 1970). While
the end of this “critical period” for OD plasticity has been proposed to be related to maturation of
inhibition in the cortex (reviewed in Hensch, 2004), changes in presynaptic input to V1 could
also contribute to differences in plasticity between juvenile and adult plasticity. Current research
suggests that mice do not have a critical period for OD plasticity, as mouse V1 is susceptible to
OD plasticity into adulthood (reviewed in Hofer et al., 2006a). However, qualitative differences
between juvenile and adult OD plasticity have been observed (Frenkel et al., 2006). To
determine if changes in input patterns to V1 contribute to differences in juvenile and adult
plasticity, we recorded extracellulary from neurons in the dorsal lateral geniculate nucleus
(dLGN) of adult animals during MC and compared the results to findings from juveniles
(Chapter 2). Both juveniles and adults show a similar decrease in correlative firing following lid
closure. However, MC in adults leads to a reduction in burst activity, a difference we were
unable to observe in juveniles at a level of statistical significance. These findings suggest that
presynaptic input to V1 from dLGN may contribute to differences in juvenile and adult cortical
plasticity. This may have important implications in studies of “critical period” mechanisms for
experience-dependent cortical plasticity and may also be important in the treatment of disorders
including amblyopia.

94
Introduction
OD plasticity is a well-studied model of experience-dependent cortical plasticity.
Closing one eye (MC) for short periods of time in young animals leads to changes in V1, such
that the majority of binocular neurons show responses now dominated by the non-deprived eye
(Wiesel and Hubel, 1963). This OD shift results primarily from a loss of response to deprived-
eye input (Mioche and Singer, 1989; Frenkel and Bear, 2004). The critical period for the loss of
deprived-eye response is restricted to early postnatal life in kittens and higher mammals (Hubel
and Wiesel, 1970). Cortical changes, including a maturation of inhibitory circuitry have been
put forth as putative mechanisms which lead to the close of the critical period (Kirkwood and
Bear, 1994; Huang et al., 1999; Rozas et al., 2001).
Recent studies suggest the mouse does not have a critical period as classically defined
(reviewed in Hofer et al., 2006a). For example, changes in the expression of the immediate early
gene, Arc, indicating an expansion of non-deprived eye input, are present following MC in mice
as old as 13 weeks, well past the traditional end of the critical periods (Tagawa et al., 2005).
Adult OD plasticity has also been demonstrated with intrinsic signal optical imaging and
extracellular single-unit recording techniques, although the OD shift observed in this study
resulted from potentiation of the non-deprived eye input, not a depression of the deprived eye
response (Hofer et al., 2006b). Furthermore, visually evoked potential recordings in the adult
mouse demonstrate that extended periods of MC (7 days) lead to both deprived-eye depression
and open-eye potentiation, while, unlike the case in juveniles, shorter periods of MC (3 days) are
not accompanied by a loss of the deprived-eye response (Frenkel et al., 2006). While these
findings suggest that the concept of a “critical period,” as classically defined, may not exist in the
mouse, there is a qualitative difference between juvenile and adult OD plasticity in this species.

95
The goal of this study was to compare the effects of MC in juveniles and adults to
determine if ages differences in OD plasticity can be accounted for by presynaptic mechanisms,
in addition to possible changes in cortex. We have already shown that MC in juveniles results in
decreased correlative firing of simultaneously recorded dLGN neurons (Chapter 2). This lack of
correlated input to V1 presumptively leads to deprived-eye depression via long-term depression
(LTD)-like mechanisms (Heynen et al., 2003; Yoon et al., 2008). Therefore, we recorded dLGN
activity in the awake, adult mouse preparation, to see if similar changes in firing pattern occur
during MC.
Methods Summary
Adult male mice were anesthetized and surgically prepared for head restraint and
electrophysiological recordings (see Detailed Methods). The animals were behaviorally
habituated to restraint over the next several days, and bundles of electrodes were inserted in the
dLGN when the mice were between the ages of postnatal day (P) 90 and P130. The location of
the recording bundle in the dLGN was initially determined by observation of visually-driven unit
activity in response to stimulation of the contralateral eye, and was always subsequently
confirmed histologically (Fig. 3-1). Baseline recordings were made with the eye contralateral to
the electrode viewing and the ipsilateral eye occluded. Activity approximating that during
normal visual experience was recorded in response to phase-reversing sinusoidal gratings and
natural scene stimuli (see Methods). Unless otherwise indicated, results using grating stimuli are
illustrated. Following the baseline recording session, animals were briefly anesthetized and
eyelid closure (MC) was performed. After 30 minutes of recovery from anesthesia, stimuli were
presented for a second recording session. At the conclusion of recordings, an electrolytic lesion
was made at the recording site to allow for histological confirmation of electrode placement.

96
Results
Figure 3-2 shows the responses of two simultaneously recorded dLGN neurons, with
different baseline firing rates, before and after MC. Prior to lid closure, both neurons are
visually responsive to the grating stimulus, as indicated by an increased likelihood of observing a
spike following phase reversal of the stimulus (indicated by arrowhead). Closing the lid causes
both neurons to lose their visually driven activity. However, the overall amount of spikes does
not change dramatically during MC.
To quantify the effect of MC on firing rate, we measured the overall amount of spiking
activity throughout the recording session before and after MC (Fig. 3-3). As observed in the
juveniles (Chapter 2, reproduced in Fig. 3-3, inset), MC has no significant effect on overall firing
rate (p > 0.1, Wilcoxon Sign-Rank (WSR)). Additionally, there are no significant differences
between the firing rates of juveniles and adults in the baseline condition (p > 0.1, Mann-Whitney
U test (MWU)), or in the degree of change in juveniles and adults following MC (p > 0.7,
MWU).
While firing rate provides information about the overall spiking activity of the neuron, it
does not provide information about temporal patterns within the spike train. One type of spike
pattern common among thalamic nuclei is the thalamic burst (reviewed in Sherman, 2001).
Bursts are detected in extracellular recordings when spike patterns consist of at least 100 ms of
silence followed by at least two spikes with inter-spike intervals less than 4 ms (Lu et al., 1992).
In the awake, juvenile mouse dLGN we found no change is the percentage of spikes in bursts
following MC (Chapter 2, reproduced in Fig. 3-4, inset). In the adult dLGN however, MC did
significantly reduce the burst percentage (Fig. 3-4; p < 0.04, WSR). We did not however, find a
difference between juveniles and adults either before MC (p > 0.2, MWU) or in the degree of

97
change following MC (p > 0.8, MWU). This suggests subtle differences between juveniles and
adults in both the baseline burst percentage, and the percentage of spikes in bursts during MC.
While we were able to detect differences between normal viewing and MC when looking
at single spike train properties in the adult, this was not present in young animals. In juveniles,
the most robust change in neural activity during MC was a change in the amount of correlative
firing between pairs of simultaneously recorded neurons (Chapter 2). Therefore, we performed
similar analyses in the adult. Figure 3-5a shows the change in cross-correlations for a
representative pair of simultaneously recorded neurons before and after MC. A scatter plot
depicting the amount of correlative firing before and after MC for the entire data set, including
juvenile animals, is shown in figure 3-5b. (Juvenile data from Chapter 2 reproduced for
comparison.) MC causes a significant decrease in correlative firing in adults (p < 0.05, WSR)
and juveniles, with no significant difference between the two groups in the baseline condition (p
> 0.3, MWU) or in the amount of change in correlative firing between juveniles and adults
following MC (p > 0.4, MWU).
Because full-field sinusoidal gratings may be unnatural visual stimuli, as in the juvenile
study (Chapter 2), we also showed the animal natural scene movies before and after MC. Figure
3-6 shows the results for activity collected while the adult animals viewed the movies. While the
firing rate (Fig. 3-6a) and burst percentage results (Fig. 3-6b) were the same as with the grating
stimuli, the decrease in correlation following MC is not reach statistically significance (p > 0.06,
WSR). The difference between the grating and natural scene conditions appears to be due to a
difference in the amount of correlation during the baseline condition (p < 0.02, WSR) as there is
less correlative activity during natural scene viewing, with no difference during MC (p > 0.1,

98
WSR) (data not shown). This may be a result of less receptive field overlap in the adult dLGN,
possibly as a result of receptive field refinement as the animal continues to develop.
Discussion
The effects of MC on dLGN firing activity are subtlety different between juvenile and
adult mice. While MC results in no change in firing rate in both juveniles and adults, it does
cause a decrease in the percentage of spikes in bursts in adults that was not observed in juveniles.
Furthermore, the decrease in correlative firing between pairs of simultaneously recorded neurons
following MC is observed in both juveniles and adults when assessed using sinusoidal grating
stimuli, but is not present in adults when assessed using natural scene movies, as it was in
juveniles.
While the decrease in burst percentage following MC was not seen in the juvenile, this is
not a surprising finding. Literature from both the cat (Lesica and Stanley, 2004) and mouse
(Grubb and Thompson, 2005) suggests that bursts convey visual information. Both of these
studies observed an increase in burst percentage when the animal was presented with visual
stimuli, as opposed to viewing white noise stimuli (Lesica and Stanley, 2004) or a blank screen
(Grubb and Thompson, 2005).
There are several potential reasons for the difference in dLGN bursting between juvenile
and adult animals. Bursts result from the activation of T-Type Ca2+
channels, which require a
long period of hyperpolarization to deinactivate. Following deinactivation, a small membrane
depolarization will activate the channel, eliciting the burst (Sherman, 2001). The difference
between adults and juveniles may result from changes in the conditions that allow for the
hyperpolarization and/or changes in the potential sources of subsequent depolarization.

99
Completely eliminating retinal input via intraocular injection of tetrodotoxin (TTX)
increases the percentage of spikes in bursts (Chapter 2). Therefore, if retinal activity were higher
in adults than juveniles during MC, bursts may occur less often because the conditions for
extended periods of hyperpolarization are less prevalent. Retinal activity may be higher in adults
if there is a developmental change in the amount of spontaneous retinal activity or if adults can
“see” better through the eyelid. Conversely, if juveniles can “see” better through the eyelid,
visually driven bursts may be more prevalent during MC. Additionally, the other sources of
input to the dLGN may be less active, thus less likely to activate the Ca2+
channel. The dLGN
receives the majority of its input from cortical feedback. Thus, the well-documented maturation
of cortical inhibition may decrease cortical feedback in the adult, which may in turn lead to less
bursting (reviewed in Hensch et al., 1998).
In addition to the bursting results, both juveniles and adults showed similar decreases in
correlative firing when assessed with sinusoidal gratings. Therefore, this may be an important
contribution to OD plasticity observed throughout the animal’s life. However, adults did not
show a statistically significant decrease in correlative firing when assessed with natural scene
movies. This may explain why deprived-eye depression is reported as either absent (Hofer et al.,
2006b) or slower (Frenkel et al., 2006) in adults. The difference between the sinusoidal grating
and natural scene stimuli in the baseline condition could result from a development of more
refined receptive fields in adults, thus decreasing the amount of correlation observed during the
movie. Cats show a maturation of spatio-temporal receptive field properties that persists past the
end of the critical period for OD plasticity in kittens (Cai et al., 1997). This type of study has not
been conducted in the mouse, but may explain this difference between juveniles and adults.

100
Alternatively, the sample size for the data set may just be too low to detect a statistically
significant decrease.
Regardless of the cause for the differences in dLGN activity between adults and
juveniles, the existence of these subtle differences suggest that changes in presynaptic activity
between juveniles and adults may be an important factor contributing to the qualitative and
quantitative differences in cortical plasticity with age. It is possible that the mechanisms for
juvenile plasticity depend heavily on the de-correlation between inputs from the deprived eye,
while in adulthood, the decrease in bursting activity, alone or in conjunction with a decrease in
correlative activity, is the main driver of deprived-eye depression. This study, in addition to
previous cortical studies, suggests that the changes in OD plasticity as an animal matures involve
an interplay between differences in input patterns to cortex and changes within cortex itself.
Moreover, this may be a principle governing maturation of plasticity throughout the cortex, and
may be an important consideration when designing treatments for disorders such as amblyopia.

101
Figure 3-1
Figure 3-1 Methodology.
a, Experimental setup. Data were obtained from awake, head-restrained mice viewing grating
stimuli. b, Schematic of recording electrode placement in the dLGN


103
Figure 3-2
Figure 3-2 dLGN responses before and after visual manipulation.
a - b, Peristimulus time histograms and raster plots from two simultaneously recorded neurons
(80 stimulus trials) are presented. Stimuli were presented at 0° or 90°, 1 Hz phase reversing
(▼). Arrowheads in this and subsequent figures indicate time of stimulus phase reversal. Spike
waveforms are recording session averages. Scale bar: 100 µV, 500 µs. Left column data were
obtained during baseline, right column after lid closure. Note the loss of visual responsiveness
with MC, and that the total number of spikes recorded is comparable before and after each
manipulation.


105
Figure 3-3
Figure 3-3 dLGN firing rate before and after MC in adult mice.
Monocular lid closure has no effect on dLGN firing rate in adult animals (n = 18 neurons (6
animals), p > 0.1 Wilcoxon Sign-Rank (WSR)). Firing rate was calculated for the full duration
of visual stimulation across all stimulus conditions. Connected gray circles represent the same
neuron recorded before and after eye manipulation ((O)pen, (C)losed). Black lines indicate
median values. Inset: Results from juvenile MC experiment (Fig. 2-3: n = 24 neurons (12
animals), p > 0.3). There are no significant differences between the firing rates of juveniles and
adults in the baseline condition (p > 0.1, Mann-Whitney U test (MWU)), or in the degree of
change in juveniles and adults following MC (p > 0.7, MWU). See also Fig. 3-6a.


107
Figure 3-4
Figure 3-4 Percentage of spikes in burst before and after MC in adult dLGN.
Monocular lid closure decreases the percentage of spikes in bursts in adults (n = 18 neurons (6
animals), p < 0.04 Wilcoxon Sign-Rank (WSR)). Connected gray circles represent the same
neuron recorded before and after eye manipulation ((O)pen, (C)losed). Black lines indicate
median values. Inset: Results from juvenile MC experiment (Fig. 2-5c: n = 24 neurons (12
animals), p > 0.2). There is no significant difference between juveniles and adults either before
MC (p > 0.2, MWU) or in the degree of change following MC (p > 0.8, MWU). See also Fig. 3-
6b.


109
Figure 3-5
Figure 3-5 Monocular lid closure decreases correlative dLGN firing in adult animals.
a, Cross-correlograms from a pair of simultaneously recorded neurons before (black line) and
after (gray line) MC. b, Scatter plots of the area under the cross-crosscorrelogram before and
after MC for each pair of simultaneously recorded neurons. Filled circles are pairs recorded in
adult mice, open circles from juveniles (reproduced from Fig. 2-8b). Gray line represents unity.
Note that following MC nearly all points fall below the unity line, indicating a decrease in
correlative firing. See also Fig. 3-6c. c, MC decreases spike correlation (adults: n = 14 neuron
pairs (6 animals), p < 0.05 WSR; juveniles (Fig. 2-8c): n = 20 neuron pairs (6 animals), p < 10-3
).
Bars represent the median change in area under the peak of the cross-correlogram (± 10 ms)
following visual manipulation. Error bars show the interquartile range. There is no significant
difference between the two groups in the baseline condition (p > 0.3, MWU) or in the amount of
change in correlative firing between juveniles and adults following MC (p > 0.4, MWU). See
also Fig. 3-6d.


111
Figure 3-6
Figure 3-6 The effects of monocular lid closure on dLGN activity when assessed using natural
scene stimuli are comparable to that observed using phase-reversing sinusoidal grating stimuli.
a, Monocular lid closure (C) has no effect on dLGN firing rate (n = 18 neurons (8 animals), p >
0.3 WSR). Connected gray circles represent the same neuron recorded before and after eye
manipulation. Black lines indicate the median values. b, MC decreases the percentage of spikes
in bursts (n = 18 neurons (8 animals), p < 0.05) c, Scatter plots of the area under the cross-
crosscorrelogram before and after MC for each pair of simultaneously recorded neurons. Note
that following MC many of the points fall below the unity line, indicating a decrease in
correlative firing. d, A decrease in spike correlation is observed following MC, although this did
not reach statistical significance (n = 14 neuron pairs (6 animals), p > 0.06). Bars represent the
median change in area under the cross-correlogram following visual manipulation. Error bars
show the interquartile range.


113
Detailed Methods
Subjects
Adult, male C57/BL6 mice (Charles River Laboratories) were group housed, on a 12
hr/12 hr light/dark cycle, with food and water available ad libitum. All animals were treated
according to NIH and MIT guidelines for animal use. Methods for obtaining data from juvenile
mice are described in Chapter 2 – Detailed Methods.
Surgical preparation for acute in vivo recording
P90 – P125 animals were anesthetized with 50 mg/kg ketamine and 10 mg/kg xylazine
(i.p.). Using cyanoacrylate, a fixation post was attached to the skull anterior to bregma. The
skull above the dLGN (2.0 mm posterior to bregma, 2.0 mm lateral to the midline) was
demarcated for future acute recording and the skull surrounding the location was encircled by a
plastic ring affixed with cyanoacrylate. An EEG electrode was placed in occipital cortex, and a
reference electrode was placed in frontal cortex. Electrodes were secured with cyanoacrylate and
dental cement was used to cover the entire skull exposure outside of the plastic ring.
Following surgery animals were monitored for signs of infection or discomfort.
Habituation to the restraint apparatus began ≥ 24 hours post-recovery. Animals remained in the
restraint system for ≥ 3 habituation sessions lasting ≥ 30 minutes each prior to the acute
recording session.
Acute recording
Animals were placed in the restraint apparatus and anesthetized with Isoflurane (1.5-
3.0% in 100% oxygen). A craniotomy was performed over the dLGN of one hemisphere.
Isoflurane was discontinued and the animal was allowed to recover from anesthesia while
remaining in the restraint system. A recording bundle consisting of seven 0.009” (22.86 μm)

114
diameter microwires was lowered into the dLGN. Microwires were 99.95% tungsten with
Formvar coating (California Fine Wire Co., Grover Beach, CA). Bundles were hand-made and
surrounded by a fine coat of cyanoacrlate to prevent splaying. Visually-driven unit activity was
used to aid in the placement of the bundle. Single unit activity and simultaneous EEG activity
were recorded using “Recorder” system software (Plexon, Inc., Dallas, TX). Single units were
discriminated offline using “Offline Sorter” (Plexon, Inc.) following methods adapted from
Csicsvari et al. (1998). The digital signal was high-pass filtered at 300 Hz. A recording
threshold was manually placed at a level to minimize noise without losing spikes. Spike
waveforms were sampled at 40 kHz and extended from 500 μs prior to threshold crossing to
1000 μs after crossing. Files recorded before and after visual manipulation were combined prior
to spike sorting to “blind” the user to the condition of each spike. Spike waveforms were aligned
to the peak. Principle component analysis (PCA) was applied to the waveform shapes.
Graphically, units were identified in either two or three dimensions using the first three principle
components, and hand-drawn polygons were used to define the cluster borders. Single units
were required to show refractory periods of at least 1 ms. In order to confirm the same unit was
recorded during the baseline and post-manipulation recording session, the cluster was visualized
in three dimensions using the first two principal components and time. Clusters that were not
stable through time were not included in further analysis. All animals were between P90 and
P130 on the day of recording.
Stimulus delivery
The visual stimuli were generated using custom MATLAB software (The Mathworks,
Natick, MA) and the Psychophysics Toolbox function set (psychtoolbox.org). The video
monitor, suitably linearized by -correction, was positioned 16 cm from the subjects’ eyes and

115
centered on the midline, occupying 82.5° x 100° of the visual field. Visual stimuli consisted of
full-screen sinusoidal gratings (0.05 cycles/°, 100% contrast), alternating in phase (phase
reversed) at a temporal frequency of 1, 2 and 4 Hz in both horizontal and vertical orientations.
Animals also viewed natural scene movies (excerpts from Microcosmos: Le peuple de l’herbe,
presented in grayscale, consisting of one 30s segment shown 10 times and one 4 minute segment
shown twice) between presentations of the sinusoidal gratings, with all stimuli presented in a
pseudorandom fashion.
Eyelid closure
Mice were anesthetized by inhalation of Isoflurane (1.5 – 3.0% in 100% oxygen).
Eyelids were held closed with Vetbond tissue adhesive (3M, St. Paul, MN).
Comparison of pre- and post-manipulation firing rates and burst percentages
Three groups of neurons were recorded under baseline and post-manipulation conditions.
To determine the statistical significance of any manipulation-induced firing rate or burst
percentage changes a two step procedure was used. We first tested if the median firing rate
change differed significantly between groups using a non-parametric Kruskal-Wallis (KW) test.
Second, we tested if the median firing rate change for each of the groups was significantly
different from zero using non-parametric Wilcoxon Sign-Rank (WSR) tests. Non-parametric
tests were used as the data were not normally distributed.
Comparison of pre- and post-manipulation pairwise correlation
To determine if correlated firing, beyond that predicted by chance, existed between pairs
of neurons, the Bernoulli correlation coefficient between the neurons’
spike trains was calculated (Aertsen et al., 1989). ( ) is the firing probability per bin of
neuron 1 (2) and the probability of joint firing at time lag . Positive (or negative)

116
departures of from zero indicate synchronous (or anti-synchronous) firing. To check the
statistical significance of manipulation induced correlation changes, the change in across each
lag was calculated and summed over lags from -10 ms to 10 ms for each pair of neurons before
and after manipulation. KW and WSR tests were then applied as above.
References
Aertsen AM, Gerstein GL, Habib MK, Palm G (1989) Dynamics of neuronal firing correlation:
modulation of "effective connectivity". J Neurophysiol 61:900-917.
Cai D, DeAngelis GC, Freeman RD (1997) Spatiotemporal receptive field organization in the
lateral geniculate nucleus of cats and kittens. J Neurophysiol 78:1045-1061.
Csicsvari J, Hirase H, Czurko A, Buzsaki G (1998) Reliability and state dependence of
pyramidal cell-interneuron synapses in the hippocampus: an ensemble approach in the
behaving rat. Neuron 21:179-189.
Frenkel MY, Bear MF (2004) How monocular deprivation shifts ocular domiance in visual
cortex of young mice. Neuron 44:917-923.
Frenkel MY, Sawtell NB, Diogo AC, Yoon B, Neve RL, Bear MF (2006) Instructive effect of
visual experience in mouse visual cortex. Neuron 51:339-349.
Grubb MS, Thompson ID (2005) Visual response properties of burst and tonic firing in the
mouse dorsal lateral geniculate nucleus. J Neurophysiol 93:3224-3247.
Hensch TK (2004) Critical period regulation. Annu Rev Neurosci 27:549-579.
Hensch TK, Fagiolini M, Mataga N, Stryker MP, Baekkeskov S, Kash SF (1998) Local GABA
circuit control of experience-dependent plasticity in developing visual cortex. Science
282:1504-1508.
Heynen AJ, Yoon BJ, Liu CH, Chung HJ, Huganir RL, Bear MF (2003) Molecular mechanism
for loss of visual cortical responsiveness following brief monocular deprivation. Nat
Neurosci 6:854-862.
Hofer SB, Mrsic-Flogel TD, Bonhoeffer T, Hubener M (2006a) Lifelong learning: ocular
dominance plasticity in mouse visual cortex. Curr Opin Neurobiol 16:451-459.
Hofer SB, Mrsic-Flogel TD, Bonhoeffer T, Hubener M (2006b) Prior experience enhances
plasticity in adult visual cortex. Nat Neurosci 9:127-132.
Huang JZ, Kirkwood A, Pizzorusso T, Porciatti V, Bear MF, Maffei L, Tonegawa S (1999)
Brain-derived neurotrophic factor is a key regulator of visual cortical development and
plasticity. Cell:submitted.

117
Hubel DH, Wiesel TN (1970) The period of susceptibility to the physiological effects of
unilateral eye closure in kittens. J Physiol 206:419-436.
Kirkwood A, Bear MF (1994) Hebbian synapses in visual cortex. J Neurosci 14:1634-1645.
Lesica NA, Stanley GB (2004) Encoding of natural scene movies by tonic and burst spikes in the
lateral geniculate nucleus. J Neurosci 24:10731-10740.
Lu SM, Guido W, Sherman SM (1992) Effects of membrane voltage on receptive field properties
of lateral geniculate neurons in the cat: contributions of the low-threshold Ca2+
conductance. J Neurophysiol 68:2185-2198.
Mioche L, Singer W (1989) Chronic recordings from single sites of kitten striate cortex during
experience-dependent modifications of receptive-field properties. J Neurophysiol 62:185-
197.
Rozas C, Frank H, Heynen AJ, Morales B, Bear MF, Kirkwood A (2001) Developmental
inhibitory gate controls the relay of activity to the superficial layers of the visual cortex. J
Neurosci 21:6791-6801.
Sherman SM (2001) Tonic and burst firing: dual modes of thalamocortical relay. Trends
Neurosci 24:122-126.
Tagawa Y, Kanold PO, Majdan M, Shatz CJ (2005) Multiple periods of functional ocular
dominance plasticity in mouse visual cortex. Nat Neurosci 8:380-388.
Wiesel TN, Hubel DH (1963) Single-Cell Responses in Striate Cortex of Kittens Deprived of
Vision in One Eye. J Neurophysiol 26:1003-1017.
Yoon BJ, Heynen AJ, Smith GB, Neve RL, Bear MF (2008) Role of AMPA receptor
endocytosis in visual cortical plasticity.


Chapter 4
DISCUSSION: Methodological Considerations,
Implications and Future Studies


121
The purpose of this research was to characterize dLGN activity patterns during normal
viewing, monocular lid closure (MC), and monocular retinal inactivation (MI) in the awake
animal. We were essentially testing the hypothesis that the dLGN was a faithful relay of retinal
input, such that when we closed the eye the dLGN would be “noisy” and less active, reflecting
spontaneous retinal input, and after MI the dLGN would, like the retina, be silent. However, we
discovered that the activity patterns of dLGN neurons are not the direct reflection of retinal
input. Neither MC nor MI decreased the overall firing rate in dLGN. MC led to a decrease in
the correlative firing of simultaneously recorded neurons, while MI led to an increase in bursting
activity and concomitant increase in correlative firing. In adults, MC reduced the bursting
activity in addition to de-correlating the activity of simultaneously recorded neurons. These
results have several implications for understanding cortical plasticity. After discussing the
methodological considerations of these experiments, I will summarize some implications and
describe avenues for future experiments.
Methodological Considerations
As with any scientific endeavor, compromises are often made in experimental design.
This research was no exception, and there are several caveats to the methodology worthy of
consideration.
The majority of the experiments were conducted using acute recording methods. The
major advantage to this strategy is that it is easier to obtain dLGN units for recording in an acute
preparation. The main drawback however, is that we were only able to look at the effects of
deprivation within hours of manipulation. While this was less than ideal, noticeable changes in
OD scores have been observed with as little as 6 hrs of deprivation (Mioche and Singer, 1989),
so the activity we were recording should correspond to the neural activity that does in fact drive

122
OD plasticity. A chronic recording method would have allowed us to explore the effects over
many hours to days of deprived eye viewing conditions. It is possible that dLGN activity
changes with increased periods of deprivation, and we were unable to track those changes with
the acute preparation. Because we suspected that dLGN activity may have changed over time
during MI, we did perform chronic recordings during that manipulation. However, the activity
did not change in any qualitative or quantitative way. Therefore, we assume a similar case
would hold for animals undergoing MC.
Another advantage to the acute recording preparation is that we believe we were able to
hold the same neurons before and after visual manipulation. To ensure that we recorded from
the same neuron, spike sorting was conducted blind to the source of the spike (pre- or post-
manipulation). Clusters were then checked for consistency between recording sessions. Because
we recorded nearly continuously throughout the experiment, stable clusters indicate that we were
holding the same unit throughout the recording sessions. This allowed us to use paired statistics
when analyzing the data. Had we used a chronic preparation and sampled activity over several
days, we could not be certain that we were recording from the same unit across days, and
statistically comparisons would have been unpaired, between groups.
Another methodological consideration related to the use of acute recordings is the choice
of head-fixing the animals. The head-fixed preparation provides a marked advantage over
freely-moving recordings because we can control the visual stimuli presented to the animal.
However, chronically implanted electrodes would have allowed us to record from animals
exploring the environment in the same manner to that which occurs during traditional OD
experiments. Additionally, we could have recorded from the animal during sleep states to see if
dLGN activity during sleep was affected by the visual manipulations. One final drawback to the

123
acute, head-fixed recording technique is that we were unable to suture the eyelids without
compromising the electrode position. For this reason, we used surgical glue to close the eyelid,
approximating lid suturing.
Another important consideration was that we were unable to measure receptive fields
(RFs) from the neurons we were recording. Mapping the RFs would have provided us with
valuable information, particularly for the analysis of simultaneously recorded neurons.
However, because the animals were awake and could move their eyes, it was not feasible to map
RF location. One option would have been to anesthetize the animal during RF mapping.
However, without also paralyzing the animals there may still have been eye movements, and
paralyzing and respirating the animal may have affected the subsequent awake recording
sessions. Another option would have been to track the eye movements of the mouse to attempt
to map RFs in the awake animal, but this was not attempted.
We also unable to confirm that the dLGN neurons we were recording from were in fact
relay cells, and not inhibitory interneurons. To address this issue, we did attempt to validate that
the neurons were projecting to primary visual cortex by stimulating either V1 or the optic
radiations and looking for an antidromic response in dLGN. However, we never observed single
units responsive to the antidromic stimulation. Therefore, it is possible that interneurons were
included in our data set, but because they account for only approximately 20% of the neuronal
population (Arcelli et al., 1997) and they have smaller cell bodies (Sherman and Guillery, 2006)
implying we would record from them less often, it is unlikely they made a significant
contribution to our findings.
Another concern relates to our spike sorting method. Because our spike sorting method
is somewhat subjective, our analysis will be subject to both type I and type II errors. That is, we

124
will sometimes include false spikes in our data in addition to sometimes omitting spikes. These
errors can contribute to changes in firing rate and ISI distributions due to simple changes in the
amount of spikes. They will also contribute to changes in burst percentage because spurious
spikes during periods of quiescence may prevent the observation of a burst, false spikes during
bursts would falsely inflate the burst percentage, missed spikes during quiescence could lead to
false classification of bursts, and missed spikes during bursts would decrease the burst
percentage. However, because we are blind to the condition of the spikes (pre- or post-
manipulation), both conditions will be equally affected by these errors. Furthermore, we looked
at relative differences before and after manipulation, and since both conditions should be equally
affected, the relative differences should not change. While our absolute values may vary slightly
from the true values, we do not make claims about absolute values.
There are at least two additional controls that we could have performed during our
experiments. One useful control would have been intraocular saline injections. The saline
injection would have controlled for any effects of the pain from intraocular TTX injection.
Following saline injection, we would not expect the bursts that occurred after MI. However, we
do not feel that the bursts observed following intraocular TTX resulted from changes in
behavioral state because our chronic recordings indicate that the bursts persist throughout the
period of MI, up to 48 hours, a time at which the animal should have fully recovered from any
discomfort due to injection.
A second useful control would have been a complete “lights out” condition to record
purely spontaneous activity. Our experimental setup did not allow for this type of recording.
Recording purely spontaneous activity would have allowed us to compare the effect of MC to the
complete absence of visual input.

125
Implications
In spite of the drawbacks to our experimental design, there are several implications of our
findings. We will discuss the implications of our results on cortical plasticity in juvenile and
adult mice, the implications of the increase in bursting following MI, and the potential hazards of
using anesthetics when studying dLGN function.
Cortical Plasticity
OD plasticity serves as a model for plasticity throughout the cortex. Deprived-eye
depression is known to mimic and occlude LTD (Heynen et al., 2003). Our results suggest that
the activity driving deprived-eye depression in vivo is a decrease in correlative firing of neurons
in dLGN, not a decrease in overall dLGN firing activity. This implies that cortical LTD in vivo
results from a de-correlation of presynaptic input.
The depression of presynaptic input can theoretically occur as a result of either
homosynaptic or heterosynaptic mechanisms (reviewed in Kerr and Abraham, 1996).
Heterosynaptic LTD occurs when postsynaptic activation is paired with very low levels of
presynaptic activity, while homosynaptic LTD results from active presynaptic synapses causing
their own depression by not correlating with postsynaptic responses. In order to distinguish
between homosynaptic and heterosynaptic mechanisms for deprived-eye depression, Rittenhouse
et al. (1999) compared the effects of MC and MI, but assumed that because MI silenced the
retina, the dLGN was also inactive. Because MC and not MI result in deprived-eye depression,
the mechanism was assumed to be homosynaptic, as weak levels of presynaptic activity did not
depress (MI), but the “noise” during MC did (Blais et al., 1999; Rittenhouse et al., 1999).
Blais et al. (1999) fit the assumptions of the Rittenhouse study to the Bienenstock,
Cooper, and Munro (BCM) (Bienenstock et al., 1982) model for homosynaptic plasticity. One

126
feature of this model is that the more residual “noise” in the presynaptic input, the faster that
input will depress. Since the dLGN was assumed to be more “noisy” during MC, and silent
during MI, the greater depression following MC fit well with the model.
However, we now know that dLGN neurons are not silent during MI, and we assume that
there is no deprived-eye depression following MI because the dLGN activity is highly patterned.
While this may invalidate the comparison assumed in the Rittenhouse study (1999), we can still
conclude that deprived-eye depression results from homosynaptic mechanisms. Homosynaptic
LTD is still the likely mechanisms for deprived-eye depression because the overall amount of
input during MC did not change. Heterosynaptic mechanisms function by “punishing” inputs
with decreased activity. MC resulted in a de-correlation of presynaptic input, and this de-
correlated input likely depressed itself, resulting in homosynaptic LTD. The conclusive study
however, described below (Future Experiments), would require the silencing of dLGN input.
The standard protocols for inducing LTD in vivo and in vitro are not physiologically
relevant (reviewed in Holscher, 1999; and Albensi et al., 2007). Because we have recorded the
neural activity that leads to homosynaptic LTD in vivo, this activity could be used to design more
physiologically relevant LTD induction protocols (see Future Experiments). Inducing LTD with
physiologically-derived induction protocols will allow researchers to study the mechanisms of
experience-dependent plasticity that actually occur in vivo. Additionally, these mechanisms may
be conserved throughout cortex. Moreover, because there are subtle differences between
juvenile and adult dLGN activity during MC, which may contribute to the differences in juvenile
and adult OD plasticity, understanding the differences between these input patterns may
elucidate mechanisms of experience-dependent plasticity in the adult. This may also help in the

127
development of treatments for disorders such as ambylopia and strabismus, conditions which are
very difficult to treat in adulthood.
MI-Induced Thalamic Bursting
In addition to the studies described above, several other studies assumed that MI
decreased dLGN activity (e.g. Chapman et al., 1986; Greuel et al., 1987; Catalano et al., 1997;
Caleo et al., 1999; Desai et al., 2002; Heynen et al., 2003; Frenkel and Bear, 2004; Young et al.,
2007). The findings of these studies warrant reinterpretation given our results.
Chapman et al. (1986) and Greuel et al. (1987) performed the first studies looking at the
effects of retinal inactivation on OD plasticity (for details, see Chapter 1). These studies found
OD shifts towards the non-deprived eye after extended periods of MI. While they attributed the
shift to a disconnection of deprived-eye input, their experimental design did not allow them to
distinguish deprived-eye depression from potentiation of the non-deprived eye. Based on the
findings of Frenkel and Bear (2004), who show open-eye potentiation following MI, and our
results regarding bursting in both the contralateral and ipsilateral segments of dLGN, we can be
relatively certain that their OD shift was in fact the result of potentiation of the non-deprived eye
input. These studies also looked at the OD shift following MI with lid closure of the fellow
eye. The results of these studies can be reinterpreted if we assume that when one eye undergoes
MI, the fellow eye is driven towards an increase in synaptic strength. Thus in Greuel et al.
(1987), no shift occurs because the increased synaptic strength occludes deprived-eye
depression. In Chapman et al. (1986), the increased synaptic strength surpassed the depressive
effect of MC.
Catalano et al. (1997) showed that NMDA-receptor subunit 1 (NR1) expression was
decreased in layer IV columns of cat and ferret V1 receiving input from the deprived eye during

128
MI. They did not observe this decrease in layer II/III, nor was it observed in layer IV of dark-
reared animals. They concluded that NR1 activity was reduced in columns receiving deprived-
eye input because those neurons were silenced. They attribute the silencing of the layer IV
neurons to the lack of dLGN input following MI and local inhibition from non-deprived eye
columns.
However, we know that the dLGN input is not silent, but we do not know the effect of
this input on cortical activity. On the one hand, the increased dLGN bursting observed during
MI may lead to an increase in activity in V1, as bursts are known to enhance efficacy of
thalamocortical (TC) transmission (Swadlow and Gusev, 2001). In fact, even though the
expression of T-Type Ca++
channels is lower in cortex than in the thalamus, hippocampus, and
other regions that highly express this channel required for bursting (Klugbauer et al., 1999), it is
possible that V1 would burst in response to dLGN bursting. However, dLGN bursting may
actually decrease activity in V1. Swadlow and Gusev (2001) looked specifically at TC
connections onto presumptive interneurons and showed that bursting enhanced the probability of
driving inhibitory neurons. Other studies in the barrel cortex have demonstrated that TC neurons
provide a stronger drive to inhibitory neurons than onto excitatory cells (Cruikshank et al.,
2007). Thus, increased inhibition may result in an overall decrease of cortical activity following
MI, and may explain the decreased NR1 expression. Experiments to characterize cortical
activity patterns during MI are described below (Future Experiments).
Desai et al. (2002) looked at the effects of synaptic scaling following MI in the
monocular segment of V1 in rats. They found an increase in mini excitatory postsynaptic
potential (mEPSP) amplitude in layer IV neurons following MI from postnatal day (P)14 – P16.
From P21 – P23, MI resulted in no change in mEPSPs in layer IV, while an increase in mEPSPs

129
was observed in layer II/III. They conclude that this scaling results from homeostatic changes
due to lack of dLGN input during MI, and that this scaling is lost at P21 due to undetermined
“critical period” mechanisms. There are some experimental differences between these studies
and ours that may account for the differences. They were examining activity at a much younger
age and were examining plasticity in the monocular segment of V1. While it is possible that MI
does not result in dLGN bursting at this age, these results could be interpreted with the presence
of a bursting input as well. Again, if bursting input leads to the silencing of cortical activity due
to increased inhibition, synaptic scaling may result. Moreover, the observed synaptic scaling
may actually be a potentiation of the response due to the increase in bursting. Furthermore, if
there is a developmental increase in bursting, perhaps bursting is not robust prior to P21, but
does drive layer IV activity after that time. This would explain the lack of scaling in layer IV,
while activity from IV to II/III may be decreased during bursting. Again, recordings of cortical
activity during MI are necessary to confirm these hypotheses.
Frenkel and Bear (2004), as described above, saw a lack of deprived-eye depression
accompanied by an increase in the non-deprived eye response following MI in the mouse. While
they interpreted this finding as a lack of depression due to the absence of retinal input, we now
know the lack of depression is likely due to patterned dLGN input following MI, with the
ipsilateral potentiation likely resulting from bursting in the ipsilateral core of the dLGN. Heynen
et al. (2003) similarly misinterpreted the lack of AMPAR endocytosis following MI as resulting
from a lack of dLGN input, when in fact the bursting presynaptic input would be highly unlikely
to result in LTD-like changes in V1.
Young et al. (2007) have modeled the effects on cortical receptive field properties
following retinal lesions. Although induction of small retinal lesions is different than MI, it is

130
unknown if retinal lesions decrease dLGN activity, or like MI, increase dLGN bursting. Because
their model assumed that the lesion resulted in the absence of presynaptic input to V1, it is
unknown if their conclusions regarding the nature of intracortical plasticity following lesioning
would hold. Without access to the specifics of their model, it is hard to predict the effect of
bursting input on their conclusions, but presumably changing the nature of the cortical input in
their model may alter their conclusions.
As is apparent from these examples, our work elucidating the activity in dLGN neurons
during MI provides critical insight to the interpretations of studies that have used this type of
manipulation. Hopefully this will lead to an increased understanding of the mechanisms of
cortical plasticity.
Effects of Anesthesia
Another critical finding of our work was that dLGN activity under barbiturate anesthesia
does not accurately represent the activity observed in the awake animal. We replicated the
results from anesthetized studies by recording from Nembutal-anesthetized mice (Chapter 2).
Under anesthesia, almost all dLGN activity was visually driven, with very little residual
spontaneous activity. Furthermore, visual stimulation often resulted in bursting activity, with
approximately 20% of spikes in bursts, similar to what was observed by Grubb and Thompson
(2005). Moreover, MI in the anesthetized preparation nearly eliminated all spiking activity,
confirming previous studies in the cat (Stryker and Harris, 1986; Rittenhouse et al., 1999). Most
importantly though, none of these results accurately reflect our recordings from awake mice,
where only approximately 5% of spikes were in bursts under normal viewing conditions, and MI
had no effect on overall firing rate, but significantly increased dLGN bursting.

131
In Chapter 1, I describe a conflict between studies showing that the TC synapse is
tonically depressed (Swadlow et al., 2002; Jia et al., 2004) and studies showing paired-pulse
facilitation at this synapse (Usrey et al., 2000). One distinguishing factor between the two
findings is the type of anesthesia used in the studies, with facilitation only present in animals that
were anesthetized with barbiturates. Taken together with our findings, it seems that barbiturate
anesthetics have a profound impact on dLGN firing.
The mechanisms of action for barbiturates is through activation of the GABAA receptor.
By increasing inhibitory tone throughout the brain, the spontaneous activity in dLGN appears to
be blocked, either through the activation of local inhibition within or projecting to the dLGN, or
by blocking excitatory inputs into dLGN. In the absence of spontaneous activity, visual input is
more likely to drive bursts, presumably because T-Type Ca++
are more easily deinactivated.
Additionally, without spontaneous dLGN activity the tonic depression at the TC synapse is
lifted.
Our findings suggest that studies of dLGN activity, TC transmission, and experience-
dependent cortical plasticity should be avoided in animals under barbiturate anesthesia, and
when they are the results should be evaluated with caution. Our results should encourage other
researchers to record activity from the awake animal whenever possible.
Future Experiments
Even though there are several important implications to the work we’ve already
completed, there are several potential future experiments that would enhance and refine the data
collected in this thesis. Some avenues of future research would require new experimental
designs, and other could potentially be investigated using the data already recorded. Several
potential ideas are described below.

132
Cortical Activity during Visual Deprivation
Our studies describe the activity in dLGN during MC and MI. However, the plasticity
that occurs in cortex depends on cortical activity patterns in addition to the presynaptic input.
Therefore, to fully understand the mechanisms that contribute to OD plasticity, it is necessary to
record cortical activity during deprivation.
The “ideal” experimental protocol would consist of chronically implanted recording
electrodes in both dLGN and layer IV of the binocular segment of V1 with electrodes recording
from neurons that are monosynaptically connected. This would require mapping the receptive
fields of the dLGN and V1 neurons during electrode placement and performing cross-correlation
analysis to verify connectivity (Tanaka, 1983). Once the electrodes were positioned to record the
connected neuron pairs, they would be affixed to the animal with dental acrylic. Following a
baseline recording session and either MC or MI, subsequent recording sessions could be
conducted throughout the period of deprivation.
This experimental design would accomplish several goals. First, we would increase the
data set for chronic recordings from the dLGN during MI, as well as establish a data set of
chronic recordings during MC. Additionally, we would record data sets of chronic recordings
from V1 during both MC and MI. Finally, the most important advantage to this design would be
to follow the correlations between monosynaptically connected pairs of dLGN and V1 neurons
before and during extended periods of MC and MI. Furthermore, the data obtained from pairs
during baseline may elucidate properties of TC transmission in the awake mouse.
This experiment is particularly difficult in practice, due to the difficulty of finding
monosynaptically connected pairs of neurons (Alonso et al., 2001) and holding them for chronic
recording. Several modifications of the experiment may be easier and will still yield useful data

133
sets. One option would be to attempt the paired recordings in the acute preparation. While this
eliminates the ability to record from the neurons during extended periods of deprivation,
connected pairs are less likely to be lost during the cementing process of the chronic preparation.
However, using microdrives might overcome this difficulty. Another option would be to record
chronically (or acutely) without specifically searching for connected pairs, while a final option
would be to record only from V1.
We would hypothesize that during MC, activity between pairs of dLGN and V1 neurons
would show a decrease in correlative firing, which may be accompanied by an overall decrease
in cortical activity. These conditions would support LTD under a Hebbian or BCM-like model
of synaptic plasticity. It is much more difficult to speculate on the cortical activity following MI.
The cortex may be quiet due to increased drive on inhibitory neurons (described above).
Furthermore, studies of immediate early gene (IEG) expression in V1 following MI show a
reduction in zif268 but not cJun expression in the hemisphere of V1 contralateral to the injected
eye in rodents (Worley et al., 1991). While it is difficult to interpret these results, zif268
expression is related to NMDAR activation (Worley et al., 1991) and may be related to synaptic
plasticity, while cJun activity is more indicative of overall activity. This implies that MI may not
have a strong effect on overall cortical activity, but may decrease the amount of plasticity in V1.
This fits with the finding that there is no deprived-eye depression following MI (Frenkel et al.,
2006). Even if simultaneous dLGN and V1 recordings are not feasible, looking at the V1
activity alone may be sufficient to resolve these questions and understand the effects of MI and
dLGN bursting. While recordings from paired dLGN and V1 neurons would provide the most
informative way to evaluate the effects of dLGN bursting on V1 activity, even recordings from

134
dLGN and V1 neurons that are not connected would be useful since bursting activity within the
dLGN is correlated (Chapter 3).
Recordings from the Monocular Zone of the dLGN
The dLGN is divided into a binocular zone which contains neurons that project to the
binocular region of V1, and a monocular zone, containing neurons that project to the monocular
region of V1. Each region of the dLGN receives cortical feedback from the region of V1 it
projects to. Therefore, if binocular and monocular V1 change differently following MC and/or
MI, they may have distinct effects of dLGN firing.
There is no change in VEP amplitude in the monocular zone following MC, presumably
due to the lack of interaction between deprived and non-deprived eye inputs in the monocular
zone (unpublished data, Frenkel and Bear). However, recent studies have shown increased
spontaneous firing rate and changes in mEPSC amplitude following MC and MI in slices of
monocular V1 studied ex vivo (Maffei and Turrigiano, 2008) and increased evoked response as
measured with calcium imaging in vivo (Mrsic-Flogel et al., 2007), suggesting plastic changes
occur in this region. Even if plastic changes are occurring in the monocular zone, they may be
different from those in the binocular zone, thus the feedback these two brain regions send to the
two segments of dLGN may differ.
Our work is focused on changes that lead to ocular dominance plasticity in the binocular
region of V1, therefore we are primarily interested in neurons recorded from the binocular zone
of the dLGN. Because of the curved shape of the dLGN, one is more likely to penetrate the
binocular zone of the dLGN when inserting an electrode from the dorsal surface. However,
inserting electrodes at the appropriate angle of penetration would facilitate recordings from the
monocular zone.

135
Monocular zone recordings may be particularly interesting because the different changes
between binocular and monocular V1 will lead to different feedback into the two zones of
dLGN. Studies of the differences between the binocular and monocular zones of the dLGN may
elucidate how much of the differences in spike patterns following MC and MI were due to feed-
forward, as opposed to feedback inputs.
Details of dLGN Bursting in the Absence of Visual Input
It would be interesting to determine which brain areas are contributing to the increased
bursting during MI. Potential input sources include cortical feedback, the thalamic reticular
nucleus (TRN), and brain stem input. Inactivating one or more of these regions may help
elucidate the source of the bursts.
In addition to understanding the source of the bursting input, we would like to understand
the purpose of the bursts. One way to address the effects of MI in the absence of bursting is to
study OD plasticity in mice with the T-Type Ca++
channel knocked-out (KO) in the thalamus
(Anderson et al., 2005). One hypothesis would be that the KO mice would not show potentiation
of the non-deprived eye in the absence of bursting in the ipsilateral core. However, because
sleep is disrupted in these animals (Anderson et al., 2005), plasticity may be affected by that as
well.
dLGN Activity in the Freely Behaving Animal, Particularly during Sleep
In typical experiments studying OD plasticity, the animals are returned to their home
cage during the period of visual deprivation. To best capture the activity patterns that result in
OD plasticity, we would want to observe dLGN activity while the animal is freely behaving in
his home cage. The behavioral state of the animal will affect the dLGN firing activity, and it is
possible that by restricting ourselves to recording from head-fixed animals, we are missing

136
behavioral states where activity during MC and MI may differ from normal viewing in different
ways than those we observed in the present study. To monitor behavioral state in freely
behaving animals we would need to implant hippocampal or cortical EEG electrodes and nuchal
muscle EMG electrodes in addition to our electrode bundle in dLGN.
Studying dLGN activity during sleep may also be informative in terms of understanding
activity patterns that contribute to OD plasticity. Sleep has been shown to enhance the effects of
OD plasticity in cats (Frank et al., 2001). Furthermore, non-rapid eye movement (NREM) sleep
has been shown to enhance LTD (reviewed in Benington and Frank, 2003). Because deprived-
eye depression utilizes LTD-like mechanisms (Heynen et al., 2003), taken together these
findings suggest that activity during NREM sleep may significantly contribute to deprived-eye
depression in vivo. Additionally, NREM activity is dominated by bursting activity, and we
observed a decrease in bursting following MC in adult animals (but not juveniles) and an
increase in bursting following MI. Perhaps bursting activity in the awake animal affects bursting
during sleep and contributes to the overall effects of visual deprivation.
Plasticity in the dLGN
As described in Chapter 1, there is very little evidence for plasticity at the
retinogeniculate synapse following MC (Wiesel and Hubel, 1963; Sesma et al., 1984; Levitt et
al., 2001). However, using a chronic recording preparation, we could test for plasticity in the
dLGN of the mouse. Recordings following eyelid opening after MC or recover from MI would
allow us to test for changes in visual responsiveness.
Verification of Homosynaptic LTD in V1
In homosynaptic LTD, active synapses depress due to their own activity patterns while in
heterosynaptic LTD, inactive synapse depress due to higher activity in other synapses (Bear and

137
Abraham, 1996; Blais et al., 1999) (described above). In our study, we found that depression
occurs during MC, not due to a decrease in activity, but due to a decrease in correlation of the
deprived-eye input. While our findings suggest that this de-correlation itself contributes to the
depression of those synapses, the definitive study to distinguish homo- and hetero-synaptic LTD
requires silencing one input pattern. If the silenced input does not depress, you can conclusively
determine that the depression uses homosynaptic mechanisms.
This was the intent of the Rittenhouse et al. (1999) study. However, this study assumed
that by silencing the retina, dLGN would be silent as well, and this assumption was also used in
the theoretical modeling work which led to the conclusion that a homosynaptic learning rule
drove deprived-eye depression (Blais et al., 1999). Because we now know that MI does not
silence dLGN activity, it would be informative to conduct an experiment where the contralateral
shell of the dLGN is silenced. Silencing only the contralateral segment of the dLGN is
nontrivial. One potential approach may be through expression of the ivermectin (IVM)-gated
Cl− channel (GluCl) (reviewed inLuo et al., 2008). GluCl channels require the expression of two
subunits to be functional. One could imagine a system where one subunit is expressed
throughout the brain, but the other subunit is introduced via intraocular delivery of a virus which
travels in an anterograde fashion, restricted to shell of the contralateral dLGN. This region can
then be silenced by intracranial delivery of IVM. VEP recordings from these animals could
confirm that the silenced input does not lead to depression, confirming that the plasticity is
homosynaptic.
Physiologically Relevant LTD Induction Protocols
LTP and LTD are popluar for synaptic plasticity in the brain. However, the induction
protocols for these models are rarely physiologically relevant (reviewed in Holscher, 1999; and

138
Albensi et al., 2007). Because the OD plasticity resulting from MC is known to mimic and
occlude LTD (Heynen et al., 2003), our recordings of dLGN activity during MC represent the
actual presynaptic activity patterns that drive plasticity in vivo. Therefore, a “playback” of this
activity in a slice preparation should induce LTD in vitro.
Initially this experiment could be attempted in the IV to II/III synapse in visual cortex
because of the ease of making visual cortical slices. However, the Yuste laboratory has recently
developed a visual thalamocortical slice preparation (MacLean et al., 2006). Using this
preparation one could “playback” our data into the contralateral shell of the dLGN. The activity
recorded during MC should result in LTD in cortex, while the activity recorded during normal
viewing and MI should not. This new LTD induction protocol should better approximate the
activity patterns that drive plasticity in vivo and using this protocol in vitro may provide new
insight into the mechanisms of cortical plasticity.
Other Visual Manipulations
It may be informative to record from the dLGN (and ideally simultaneously from V1)
during other forms of visual deprivation including binocular lid closure (BC), binocular retinal
inactivation (BI), and complete darkness (DE). While the activity patterns during BC and BI
may not be different in dLGN, cortical activity is most likely different under these conditions,
thus cortical feedback to dLGN neurons may be affected. Also, the DE condition would serve as
a comparison to MC, as discussed above.
Secondary Verification
A final series of future experiments would be experiments to confirm our findings. Using
extracellular recordings, we have shown that MC and MI do not reduce the overall firing rates in
dLGN. Because the results were surprising and somewhat unexpected, we would like to verify

139
this through an independent method. In addition to electrophysiological recordings, there are
molecular markers that one can use to monitor neural activity at the cellular level. Under
specific conditions, the expression of IEGs can be used as such a marker. The IEG cFos, a
transcription factor, one marker for neural activity that is expressed in dLGN and V1 (Herdegen
et al., 1998). Thus, we may be able to confirm the electrophysiological recordings by monitoring
levels of cFos expression during visual deprivation.
Basal cFos levels are low in both dLGN and V1 (reviewed in Herdegen et al., 1998).
However, cFos expression can be induced by introducing visual stimuli after a period of dark
exposure (reviewed in Kaczmarek and Chaudhuri, 1997). Therefore, in our proposed
experiments, after an extended period of dark exposure, mice would be exposed to a period of (1)
light, (2) light with MC, (3) light with MI, or (4) MI in the dark. Immunofluorescent staining
and confocal imaging would allow us to visualize and quantify cFos expression at the cellular
level. Next, we would make comparisons both within the same animal between its two
hemispheres (which receive different visual inputs) and between animals with different visual
histories. If dLGN activity levels are the same during MC, MI and light conditions, we would
expect all of these conditions to show higher levels of cFos expression than the dark conditions.
We could also use this technique to examine activity levels in V1. Prior to these experiments, it
may be necessary to confirm that dLGN firing patterns are the same as in the present study when
assessed following a period of dark exposure.
We have performed an extensive pilot study, but, unfortunately the quality of cFos
staining has been variable and unreliable. An alternative approach for visualizing cFos activity
would be to use transgenic mice that harbor a fos-GFP fusion gene under the control of the
native fos promoter (Barth, 2007). The mice would be exposed to the same conditions as

140
described above, and visualization of GFP levels could be used to determine the level of cFos
activation.
More Advanced Spike-Train Analysis
In addition to the experiments proposed above, there are some studies that can possibly
be done by applying new analyses to the data set. Examinations of the raster plots and peri-
stimulus time histograms for activity recorded during MC do not seem to indicate that there is an
effect of visual input. However, no careful analysis has been applied to the spike trains. One
possibility would be to apply a generalized linear model (GLM) to the data (Truccolo et al.,
2005). This technique models the spike train as a point-process and models the relative effects
of spiking history, simultaneously recorded spike trains, and extrinsic factors like visual input. If
the visual input was a significant factor in the model, we could conclude that the neuron was
responding to visual stimuli through the eyelid.
Recent advances in GLM methodology now allow for the analysis of between-trial
variability of spike trains (Czanner et al., 2008). We did not examine the response to the
sinusoidal gratings on a trial-by-trial basis. It may be possible that one difference between
viewing and MC is the amount of trial-to-trial variability. Additionally, we may find short term-
plasticity effects in the dLGN, such as an increased response to repeated stimuli, if we evaluated
the data in this fashion.
Conclusions
In conclusion, we have provided the first recordings of dLGN activity that leads to OD
plasticity in the awake animal. Characterizing this activity can allow for new ways to study
experience-dependent cortical plasticity. This will lead to a more complete understanding of the
mechanisms that allow for these changes. We have also discovered that dLGN activity bursts

141
following an elimination of retinal input. Bursting does not occur following MI in the
anesthetized animal, underscoring the importance of recording from the awake preparation.
These findings allow for new interpretations of old studies, and should guide future research in
cortical plasticity. Furthermore, the results may prove invaluable in the treatments of
developmental disorders including ambylopia.
References
Albensi BC, Oliver DR, Toupin J, Odero G (2007) Electrical stimulation protocols for
hippocampal synaptic plasticity and neuronal hyper-excitability: are they effective or
relevant? Exp Neurol 204:1-13.
Alonso JM, Usrey WM, Reid RC (2001) Rules of connectivity between geniculate cells and
simple cells in cat primary visual cortex. J Neurosci 21:4002-4015.
Anderson MP, Mochizuki T, Xie J, Fischler W, Manger JP, Talley EM, Scammell TE,
Tonegawa S (2005) Thalamic Cav3.1 T-type Ca2+ channel plays a crucial role in
stabilizing sleep. Proc Natl Acad Sci U S A 102:1743-1748.
Arcelli P, Frassoni C, Regondi MC, De Biasi S, Spreafico R (1997) GABAergic neurons in
mammalian thalamus: a marker of thalamic complexity? Brain Res Bull 42:27-37.
Barth AL (2007) Visualizing circuits and systems using transgenic reporters of neural activity.
Curr Opin Neurobiol 17:567-571.
Bear MF, Abraham WC (1996) Long-term depression in hippocampus. Annu Rev Neurosci
19:437-462.
Benington JH, Frank MG (2003) Cellular and molecular connections between sleep and synaptic
plasticity. Prog Neurobiol 69:71-101.
Bienenstock EL, Cooper LN, Munro PW (1982) Theory for the development of neuron
selectivity: orientation specificity and binocular interaction in visual cortex. J Neurosci
2:32-48.
Blais BS, Shouval HZ, Cooper LN (1999) The role of presynaptic activity in monocular
deprivation: comparison of homosynaptic and heterosynaptic mechanisms. In: Proc Natl
Acad Sci U S A, pp 1083-1087.
Caleo M, Lodovichi C, Maffei L (1999) Effects of nerve growth factor on visual cortical
plasticity require afferent electrical activity. Eur J Neurosci 11:2979-2984.
Catalano SM, Chang CK, Shatz CJ (1997) Activity-dependent regulation of NMDAR1
immunoreactivity in the developing visual cortex. J Neurosci 17:8376-8390.

142
Chapman B, Jacobson MD, Reiter HO, Stryker MP (1986) Ocular dominance shift in kitten
visual cortex caused by imbalance in retinal electrical activity. Nature 324:154-156.
Cruikshank SJ, Lewis TJ, Connors BW (2007) Synaptic basis for intense thalamocortical
activation of feedforward inhibitory cells in neocortex. Nat Neurosci 10:462-468.
Czanner G, Eden UT, Wirth S, Yanike M, Suzuki WA, Brown EN (2008) Analysis of between-
Trial and within-Trial Neural Spiking Dynamics. J Neurophysiol.
Desai NS, Cudmore RH, Nelson SB, Turrigiano GG (2002) Critical periods for experience-
dependent synaptic scaling in visual cortex. Nat Neurosci 5:783-789.
Frank MG, Issa NP, Stryker MP (2001) Sleep enhances plasticity in the developing visual cortex.
Neuron 30:275-287.
Frenkel MY, Bear MF (2004) How monocular deprivation shifts ocular domiance in visual
cortex of young mice. Neuron 44:917-923.
Frenkel MY, Sawtell NB, Diogo AC, Yoon B, Neve RL, Bear MF (2006) Instructive effect of
visual experience in mouse visual cortex. Neuron 51:339-349.
Greuel JM, Luhmann HJ, Singer W (1987) Evidence for a threshold in experience-dependent
long-term changes of kitten visual cortex. Brain Res 431:141-149.
Grubb MS, Thompson ID (2005) Visual response properties of burst and tonic firing in the
mouse dorsal lateral geniculate nucleus. J Neurophysiol 93:3224-3247.
Herdegen T, Claret FX, Kallunki T, Martin-Villalba A, Winter C, Hunter T, Karin M (1998)
Lasting N-terminal phosphorylation of c-Jun and activation of c-Jun N-terminal kinases
after neuronal injury. J Neurosci 18:5124-5135.
Heynen AJ, Yoon BJ, Liu CH, Chung HJ, Huganir RL, Bear MF (2003) Molecular mechanism
for loss of visual cortical responsiveness following brief monocular deprivation. Nat
Neurosci 6:854-862.
Holscher C (1999) Synaptic plasticity and learning and memory: LTP and beyond. J Neurosci
Res 58:62-75.
Jia F, Xie X, Zhou Y (2004) Short-term depression of synaptic transmission from rat lateral
geniculate nucleus to primary visual cortex in vivo. Brain Res 1002:158-161.
Kaczmarek L, Chaudhuri A (1997) Sensory regulation of immediate-early gene expression in
mammalian visual cortex: implications for functional mapping and neural plasticity.
Brain Res Brain Res Rev 23:237-256.
Kerr DS, Abraham WC (1996) LTD: many means to how many ends? Hippocampus 6:30-34.

143
Klugbauer N, Marais E, Lacinova L, Hofmann F (1999) A T-type calcium channel from mouse
brain. Pflugers Arch 437:710-715.
Levitt JB, Schumer RA, Sherman SM, Spear PD, Movshon JA (2001) Visual response properties
of neurons in the LGN of normally reared and visually deprived macaque monkeys. J
Neurophysiol 85:2111-2129.
Luo L, Callaway EM, Svoboda K (2008) Genetic dissection of neural circuits. Neuron 57:634-
660.
MacLean JN, Fenstermaker V, Watson BO, Yuste R (2006) A visual thalamocortical slice. Nat
Methods 3:129-134.
Maffei A, Turrigiano GG (2008) Multiple modes of network homeostasis in visual cortical layer
2/3. J Neurosci 28:4377-4384.
Mioche L, Singer W (1989) Chronic recordings from single sites of kitten striate cortex during
experience-dependent modifications of receptive-field properties. J Neurophysiol 62:185-
197.
Mrsic-Flogel TD, Hofer SB, Ohki K, Reid RC, Bonhoeffer T, Hubener M (2007) Homeostatic
regulation of eye-specific responses in visual cortex during ocular dominance plasticity.
Neuron 54:961-972.
Rittenhouse CD, Shouval HZ, Paradiso MA, Bear MF (1999) Monocular deprivation induces
homosynaptic long-term depression in visual cortex. Nature 397:347-350.
Sesma MA, Irvin GE, Kuyk TK, Norton TT, Casagrande VA (1984) Effects of monocular
deprivation on the lateral geniculate nucleus in a primate. Proc Natl Acad Sci U S A
81:2255-2259.
Sherman SM, Guillery RW (2006) Exploring the thalamus and its role in cortical function, 2nd
Edition. Cambridge, MA: The MIT Press.
Stryker MP, Harris WA (1986) Binocular impulse blockade prevents the formation of ocular
dominance columns in cat visual cortex. J Neurosci 6:2117-2133.
Swadlow HA, Gusev AG (2001) The impact of 'bursting' thalamic impulses at a neocortical
synapse. Nat Neurosci 4:402-408.
Swadlow HA, Gusev AG, Bezdudnaya T (2002) Activation of a cortical column by a
thalamocortical impulse. J Neurosci 22:7766-7773.
Tanaka K (1983) Cross-correlation analysis of geniculostriate neuronal relationships in cats. J
Neurophysiol 49:1303-1318.

144
Truccolo W, Eden UT, Fellows MR, Donoghue JP, Brown EN (2005) A point process
framework for relating neural spiking activity to spiking history, neural ensemble, and
extrinsic covariate effects. J Neurophysiol 93:1074-1089.
Usrey WM, Alonso JM, Reid RC (2000) Synaptic interactions between thalamic inputs to simple
cells in cat visual cortex. J Neurosci 20:5461-5467.
Wiesel TN, Hubel DH (1963) Effects of Visual Deprivation on Morphology and Physiology of
Cells in the Cats Lateral Geniculate Body. J Neurophysiol 26:978-993.
Worley PF, Christy BA, Nakabeppu Y, Bhat RV, Cole AJ, Baraban JM (1991) Constitutive
expression of zif268 in neocortex is regulated by synaptic activity. Proc Natl Acad Sci U
S A 88:5106-5110.
Young JM, Waleszczyk WJ, Wang C, Calford MB, Dreher B, Obermayer K (2007) Cortical
reorganization consistent with spike timing-but not correlation-dependent plasticity. Nat
Neurosci 10:887-895.

Appendix
1. DETAILED METHODS FOR dLGN RECORDINGS
2. RESPONSE TO CONCERNS


147
Appendix 1 – Detailed Methods for dLGN Recordings
Surgical Preparation
Prior to surgery:
For each surgery, you will need 1 head post, 1 plastic ring, 2 “gold pin” electrodes, and 1 “plumb
line.”
To make the head post, cut a ¾” x 18 wire nail to 1 cm and smooth the cut edge with the dremel.
To make the plastic ring, cut a ring from a 100 – 1000 μL pipette tip using a razor blade.
To make the electrodes, solder a small length of 0.01” diameter bare silver wire into a male gold
wire contact.
To make the plumb line, remove the sharp point and plastic portion of a 30g needle, leaving just
the needle shaft.
Materials
Needles (26g and 30g) and 1 mL syringes: 1 syringe plus 1 26g needle for anesthesia. 1 syringe
plus 1 26g needle for lidocaine. 1 syringe plus 1 30g needle for Zip Kicker. Additional 30g
needles for glue application.
Ketamine HCl, Ketaject
Xylazine, Xylaject
Lidocaine HCl 2%
Petrolatum Ophthalmic Ointment, Puralube Vet Ointment
70% EtOH
Alcohol Swabs
Iodine Swabs
“Zip Kicker”
Loctite Glue #495
Loctite Glue #454
Ortho Jet Acrylic Jet powder
Ortho Jet Acrylic Liquid
Electric razor
Scissors
Razor blade
Q-tips
Air duster
Forceps
Hex drivers for stereotaxic apparatus
Skin marker

148
Rechargeable High Speed Microdrill
Drill bit (0.7 mm)
Wire cutters
Spatula
Aluminum Dish (for Ortho Jet Arcylic)
Methods
Anesthetize the mouse with a cocktail of 50 mg/kg ketamine and 10 mg/kg xylazine ip.
Shave the fur on the mouse head removing as much as possible from the snout back past the ears,
getting as close to the eyes as possible. Avoid whiskers.
Clean the surgical area with iodine and alcohol (x3). Apply ophthalmic ointment to the eyes.
Inject 0.1 mL of lidocaine subcutaneously above the skull.
Using scissors and forceps, cut open the skin over the skull.
Clean the skull surface with q-tips.
Use a razor blade to roughen the surface of the skull. Clean the surface with EtOh. Dry
thoroughly with air duster.
Using the 495 glue, affix the head post anterior to bregma (on the frontal suture). Solidify glue
with zip kicker.
Place the mouse into the stereotaxic apparatus. Tighten head post with hex driver. Level the
skull, and ensure the midline is straight.
Reinforce head post with 454 glue and zip kicker.
Using the skin marker, mark the location of 2.1 posterior of bregma, 2.2 lateral of midline.
Using 454 glue and zip kicker, affix the plastic ring around this point.
Trim gold pin electrodes with wire cutters. Place the two gold pin electrodes in the hemisphere
opposite the plastic ring by thinning the skull with the microdrill at the insertion location,
pushing the pin through the skull, and affixing in place with 454 glue and zip kicker. The
reference electrode should be over frontal cortex, and the EEG electrode should be over occipital
cortex (on lambda).
Using the stereotaxic electrode holder to hold the plumb line, glue the plumb line slightly
posterior to the plastic ring using 454 glue and zip kicker.
Mix the Ortho Jet acrylic powder and liquid in an aluminum dish. Cover all surfaces outside of
the plastic ring with Ortho Jet acrylic using the spatula.

149
Habituation
Animals are placed in the recording apparatus and head fixed for at least 3 30+ minute sessions,
while the animals views a gray screen on the monitor. During the first session no occluder is
used. During the second and third session, one eye, then the other is occluded.
Recording
Materials
Electrode bundle
Isoflurane
Saline
Microdrill
Drill bit (0.7 mm)
30g needle
Air duster
Surgical Eye Spears
Absorbent paper points
Methods
Animals are placed in the recording apparatus and anesthetized with 1.5-3% Isoflurane in 100%
oxygen.
Using the microdrill, perform a small craniotomy at the site of the marker. A 30g needle can be
used to remove sharp edges from the craniotomy. Remove bone particles with the air duster.
Clean the area and stop any bleeding with saline, surgical eye spears and paper points.
Remove animal from Isoflurane.
Align the electrode bundle to plumb line. Slowly lower electrode bundle into the brain. Be
aware of dimpling. Record during electrode penetration to hear cortex and hippocampus. Once
past the hippocampus, dimpling may occur again at thalamic surface. Listen for response to
sinusoidal grating.

150
Appendix 2 – Response to Concerns
Here I will provide detailed responses to two concerns of my thesis committee: the
possible correlation between behavioral state and increased correlation following MI, and the
sharp peaks observed when cross-correlation analysis was performed.
Behavioral State and Bursting
Certain behavioral states are correlated with an increase in burst percentage (e.g.
Bezdudnaya et al., 2006). Because we saw an increase in burst percentage following MI, it is
necessary to confirm that this increase was not due to a change in behavioral state. To address
this concern, we analyzed the cortical EEG recorded from 3 mice in the control and MI groups.
Power spectral analysis of EEG recorded from occipital cortex was performed in 10 s bins using
the Welch method of spectral estimation. We defined the delta band as frequencies below 4 Hz
and the theta band as frequencies between 4.5 and 9 Hz. We used the theta /delta ratio (TDR) as
our measure of behavioral state (Maloney et al., 1997).
Our results are summarized in Figure A-1. Fig. A-1a shows the mean TDRs before and
after manipulation in the control and MI groups. First, the average TDR was calculated across
all 10 s bins for each animal in each condition. Bars represent the mean over the 3 averages from
each group. There are no differences between groups (p > 0.5, Kruskal-Wallis). This suggests
there is no change in behavioral state following MI. Fig. A-1b,c show the relationship between
the TDR in each 10 s bin to the number of bursts recorded during that bin from each neuron
recorded from the animals. While we do see a peak in bursts at TDRs around 1, possibly
representing an inattentive state, there is no clear correlation between TDR and the number of
observed bursts. For example, high levels of bursting occur when the TDR is high, indicating

151
that bursts can be observed when the animal is aroused. Furthermore, there is no evidence that
MI affects the TDRs observed, nor does it affect the relationship between TDR and bursting.
Sharp Correlogram Peaks
Alonso et al. (1996) have reported dLGN neurons with highly synchronous firing.
Neurons correlated at the 1 ms timescale were recorded from electrodes 100 – 400 μm apart and
had very similar, overlapping receptive fields. When neurons had overlapping receptive fields,
they showed correlations on average of about 28% (Alonso et al., 1996).
We analyzed the sharp peaks in our correlograms to make the case that we were not
recording from the same neurons on multiple electrodes. Correlograms were originally
constructed using 1 ms bins. For those that were highly peaked, we reconstructed the
correlogram using .1 ms bins. Similar to the Alonso et al. study, we found an average correlation
of 22%. Because our electrodes are closely bundled together, and because rodent receptive fields
are larger than those in the cat, we would suspect our neurons to have overlapping fields, so
these results should not be surprising.
Furthermore, when we considered the correlations after MC and MI, we found that the
large majority of neurons that were tightly correlated (in .1 ms bins) prior to manipulation, no
longer showed a tight correlation at the .1 ms timescale following either manipulation (10 out of
12 neuron pairs). If we were in fact recording from the same neuron on two electrodes, the
amount of correlative firing observed should not be affected by visual manipulation. This
suggests that the observed peaks were related to driven retinal ganglion cell input, which is
disrupted by MC and MI. In the control group however, 8 of the 11 neuron pairs remained
tightly correlated following anesthesia, again suggesting convergent retinal ganglion cell input.
2 pairs in the MI group that were not tightly correlated at the .1 ms level became tightly

152
correlated following MI. There are no examples of this in the MC or the control group,
suggesting that this occurred as a result of the TTX. (The example in Fig. 2-8a, right panel, was
one of these pairs. Prior to MI, the tight correlation observed at the 1 ms bin size was not
observed at the .1 ms bin size, however we did observe this following MI.)
However, we acknowledge that it still possible that we were recording spikes from the
same neuron on more than one electrode (particularly in the 2 of 12 pairs that did not change
after manipulation). While this would have a small impact on our data sets, we do not believe it
would change our overall conclusions.

153
Figure A-1
Figure A-1 Increased bursting is not associated with a change in behavioral state.
a, Mean theta/delta ratios from 3 animals in the control group (left) and the MI group (right).
White bars are the baseline or (O)pen eye condition. Black bars are the post-manipulation
conditions of (O)pen eye for the control group and (I)nactivated eye for the MI group. Power-
spectral analysis was performed in 10 s bins across the entire recording session and averaged for
each animal. Bars represent the means of the three animals; error bars show standard error.
There are no differences in theta/delta ratio (p > 0.5, Kruskal-Wallis). b – c, Scatter plots of
theta/delta ratio vs the number of bursts. Each circle represents a single 10 s bin over which
power-spectral analysis was performed for the animal and the number of bursts were counted for
the unit. Open circles represent data from the baseline condition and filled gray circles are the
post-manipulation conditions of (b) open eye viewing and (c) MI. Data represent (b) 9 neurons
from 3 animals and (c) 10 neurons from 3 animals. While bursting peaks at a theta/delta ratio
around 1, there is no clear correlation between theta/delta ratio and the amount of bursts, nor is
there an apparent effect of MI.


155
References
Alonso JM, Usrey WM, Reid RC (1996) Precisely correlated firing in cells of the lateral
geniculate nucleus. Nature 383:815-819.
Bezdudnaya T, Cano M, Bereshpolova Y, Stoelzel CR, Alonso JM, Swadlow HA (2006)
Thalamic burst mode and inattention in the awake LGNd. Neuron 49:421-432.
Maloney KJ, Cape EG, Gotman J, Jones BE (1997) High-frequency gamma
electroencephalogram activity in association with sleep-wake states and spontaneous
behaviors in the rat. Neuroscience 76:541-555.