Embed Size (px)
Citation preview

Journal of Integrative Plant Biology 2011, 53 (5): 399–408
Research Article
LEUNIG_HOMOLOG and LEUNIG Regulate SeedMucilage Extrusion in ArabidopsisF
Minh Bui, Nathan Lim, Paja Sijacic and Zhongchi Liu∗
Department of Cell Biology and Molecular Genetics, University of Maryland, College Park, Maryland 20742, USA∗Corresponding author
Tel: +1 301 405 1586; Fax: +1 301 314 1284; E-mail: [email protected] Articles can be viewed online without a subscription.Available online on 28 February 2011 at www.jipb.net and www.wileyonlinelibrary.com/journal/jipbdoi: 10.1111/j.1744-7909.2011.01036.x
Abstract
LEUNIG (LUG) and LEUNIG_HOMOLOG (LUH) encode two closely related Arabidopsis proteins, belong-ing to the Gro/TLE family of transcriptional co-repressors. These two genes were previously shownto exhibit partially overlapping functions in embryo and flower development. In this report, the roleof both LUH and LUG on seed mucilage extrusion was examined. Seed mucilage extrusion occursafter the seeds are imbibed, serving as functional aid in seed hydration, germination, and dispersal.While luh-1 mutants exhibited strong defects in seed mucilage extrusion, lug-3 mutants exhibited aminor phenotype in mucilage extrusion. Further characterization indicates that luh-1 does not exhibitany obvious defect in seed epidermal cell differentiation, mucilage synthesis, or mucilage deposition,suggesting a specific role of LUH in mucilage extrusion. This seed mucilage phenotype of luh-1 isidentical to that of mucilage modified 2 (mum2) mutants. MUM2 encodes a β-galactosidase required forthe modification of the mucilage. Quantitative reverse transcription polymerase chain reaction of RNAextracted from siliques detected a slight decrease of MUM2 mRNA in the luh-1 mutant compared to thewild type. Together, LUH and possibly LUG may specifically regulate mucilage extrusion by promotingthe expression of genes required for mucilage maturation.
Bui M, Lim N, Sijacic P, Liu Z (2011) LEUNIG_HOMOLOG and LEUNIG regulate seed mucilage extrusion in Arabidopsis. J. Integr. Plant Biol. 53(5),399–408.
Introduction
Seed mucilage is a functional aid in seed hydration, ger-mination, and dispersal (Fahn 1982; Penfield et al. 2001).The primary component of mucilage is carbohydrate-basedpectin. The three main types of pectins are homogalacturonan(HG), rhamnogalacturonan I (RG I), and rhamnogalacturonanII (Arsovski et al. 2010), which are synthesized, secreted, anddeposited by the seed outer integument, which consists of twocell layers, the outermost epidermal cell layer and a secondsubtending cell layer. In Arabidopsis, the outer integument celldifferentiation is a highly dynamic process and can be dividedinto five stages (Western et al. 2000, 2001; Windsor et al.2000). Fertilization of ovules triggers outer integument cell ex-pansion (stage 1), followed by amyloplast accumulation (stage2), and mucilage synthesis and secretion into the apoplastic
space (stage 3). The mucilage secretion into the apoplasticspace forces cytoplasmic rearrangement, leading to a volcano-shaped cytoplasmic column in the center of the epidermalcells in each seed. At stage 4, starch granule degradation isaccompanied by secondary wall deposition. Epidermal cells aremarked by a central volcano-like secondary cell wall known asthe columella (Western et al. 2000; Windsor et al. 2000). Stage5 is the desiccation of seeds. When seeds are planted, theyabsorb water (imbibition). The mucilage previously trapped inthe apoplastic space expands and erupts through the primarycell wall, generating a gelatin-like ring encapsulating the seeds.
Previous studies have identified a large number of genesrequired for seed coat differentiation and mucilage release.They include TRANSPARENT TESTA GLABRA1 (TTG1),TTG2, TRANSPARENT TESTA2 (TT2), TT8, GLABRA2 (GL2),ENHANCER OF GLABRA3 (EGL3), MYB5, MYB61 and
C© 2011 Institute of Botany, Chinese Academy of Sciences

400 Journal of Integrative Plant Biology Vol. 53 No. 5 2011
APETALA2 (AP2) (Jofuku et al. 1994; Rerie et al. 1994;Penfield et al. 2001; Western et al. 2001; Johnson et al.2002; Zhang et al. 2003; Li et al. 2009; Arsovski et al. 2010).These genes, which encode transcription factors, not onlyregulate seed outer integument cell differentiation but also otherdevelopmental processes such as root hair initiation, trichomedifferentiation, and flower organ identity determination. Mostof these mutants’ seed epidermis exhibit a reduced or absentcolumella.
Another type of mucilage mutant exhibits defects that arespecific to seed mucilage biosynthesis or maturation, and maydefine target genes of transcription factors described above.Mucilage Modified 4 (MUM4) encodes an NDP (nucleosidediphosphate)-rhamnose synthase necessary for rhamnogalac-turonan I (RGI) synthesis (Usadel et al. 2004; Western et al.2004; Oka et al. 2007). mum4 mutants have reduced seedcoat columella size and reduced mucilage extrusion (Westernet al. 2001). MUM4 is upregulated by GL2, that is downstreamof TTG1 and AP2 (Western et al. 2004). On the other hand,MUM2 encodes a member of glycosyl hydrolase Family 35 β-galactosidase (BGAL6) required for mucilage maturation post-synthesis and post-secretion into the apoplastic space (Deanet al. 2007; Macquet et al. 2007). mum2 seed outer integumentsynthesizes normal amounts of mucilage but fails to extrudethe mucilage upon imbibition. MUM2 acts to remove the galac-tose/galactan branches to increase the hydrophilic propertiesof the mucilage, which is needed for normal hydration andexpansion of the mucilage.
Other mutants with similar phenotypes to mum2 includemutants of PATCHY (AtBXL1) which codes for a bifunctional β-D-xylosidase/a-L-arabinofuranosidase. patchy mutants exhib-ited patchy mucilage release and delayed seed germination.PATCHY (AtBXL1) was suggested to trim off rhamnogalactur-onan I arabinan side-chains of the mucilage (Arsovski et al.2009). A subtilisin-like serine protease coded by AtSBT1.7affects mucilage extrusion by either directly degrading a pectinmethylesterase (PME), or proteolytically cleaving and thus acti-vating a PME inhibitor. Thus, AtSBT1.7 prevents excessive de-methylesterification of the mucilage and/or of the primary cellwall (Rautengarten et al. 2008). More recently, mutants of theArabidopsis GALACTURONOSYLTRANSFERASE (GAUT)11was shown to cause reduced mucilage release and lower mu-cilage galacturonic acid levels. gaut11 may affect cell wall mod-ification or biosynthesis of galacturonic acid in mucilage (Caffallet al. 2009). Nevertheless, it is not known what regulators maycontrol the stage- and integument tissue-specific expression ofmucilage modifiers like MUM2, PATCHY (AtBXL1), AtSBT1.7,and GAUT11.
Previously, our lab has been characterizing two homologoustranscriptional co-repressors, LUH and LUG, both belongingto the Groucho/TUP1 family of transcription co-repressors,which included TOPLESS1 in Arabidopsis (Conner and Liu
2000; Long et al. 2006; Liu and Karmarkar 2008; Sitaramanet al. 2008). Both LUH and LUG were shown to regulate floral,leaf, and embryo development, and luh; lug double mutantsare embryo lethal (Sitaraman et al. 2008; Stahle et al. 2009).LUG and LUH directly interact with a co-factor SEUSS (SEU)that bridges the interaction between LUG/LUH co-repressorsand target-specific DNA-binding transcription factors such asAPETALA1 (AP1) (Sridhar et al. 2004, 2006; Sitaraman et al.2008). In this study we showed that mutants of LUH exhibit amucilage extrusion defect similar to that of mum2 mutants. Thissimilarity in phenotype led us to investigate the potential regula-tory relationship between LUH and MUM2 and the involvementof LUG and SEU in this process.
Results
luh-1 mutant seeds exhibit mucilage extrusion defects
Previously we isolated a putative null allele of LUH, luh-1, inthe Col-er background via the Arabidopsis TILLING project(Sitaraman et al. 2008). We reported that luh-1 seeds exhibitedreduced germination rate (Sitaraman et al. 2008). In addition,luh-1 seeds tended to clump to each other when they weresoaked in water. To understand the basis of these defects,we tested mucilage release in luh-1. Dry seeds of luh-1 andthe wild type (both Col-er and L-er accessions) were soaked inliquid containing 0.01% toluidine blue and then observed undera dissecting microscope. A halo of gelatinous mucilage wasobserved that encapsulated each wild-type seed (Figure 1A,
B). In contrast, there was a complete absence of mucilagesurrounding the luh-1 seeds (Figure 1C). This mucilage defect isspecifically caused by a loss of LUH as 35S::LUH fully rescuedthe mucilage defect of luh-1 (Figure 1D).
The released mucilage could be separated into two distinctlayers, the loose and water-soluble outer layer that can beeasily shaken off and the denser inner layer that is more tightlyassociated with the seed (Western et al. 2001). luh-1 lacks bothmucilage layers, while the wild type as well as luh-1; 35S::LUHpossess both mucilage layers (Figure 1A-H). The effect of luh-1 mutation in seed mucilage release is highly penetrant, with95% luh-1 seeds lacking mucilage compared to 1.6% wild-type(Col-er) seeds (Figure 1I–L; Figure 2). luh-1; 35S::LUH is similarto the wild type with only 4.2% seeds lacking mucilage uponimbibition. An elevated LUH expression level caused by thestrong 35S promoter in luh-1; 35S::LUH plants did not increasethe amount of seed mucilage. Thus, LUH expression level isnot proportional to the amount of mucilage produced.
lug but not seu exhibited weak mucilage defects
LUG encodes a protein with a high level of sequence similarityto LUH (Sridhar et al. 2004; Sitaraman et al. 2008). We

Co-repressors Required for Seed Mucilage Release 401
Figure 1. Mucilage extrusion after phenotype of wild type (WT) and luh-1.
Toluidine blue staining of unshaken (A–D) and shaken seeds (E–H), and lower magnification photos of shaken seeds (I–L).
Bars indicate 200 µm in (A–H) and 500 µm in (L), (I–L) have the same magnification.
(A, E, I) Wild type (Col-er).
(B, F, J) Wild type (L-er).
(C, G, K) luh-1.
(D, H, L) luh-1; 35S::LUH.
(M) A wild-type (Col-er) seed pre-treated with ethylene diamine tetra acetate (EDTA).
(N) A luh-1 seed pre-treated with EDTA.
Shown is a thin layer of mucilage that encapsulates the luh-1 seed.
tested if lug-3, a strong loss-of-function mutant, also exhibitedseed mucilage defects. Most lug-3 mutant seeds still releasemucilages (Figure 2). However, lug-3 mutant seeds had asignificant reduction of the outer mucilage layer (Figure 3B). Theinner mucilage layer of lug-3 was thin and often torn (Figure 3F,
J). This weakness became more apparent when the seedswere shaken, as the water was often littered with mucilagefragments. This contrasts with the inner mucilage layer of thewild type seeds that mostly remained intact (Figure 1E). Thus,LUG does have a role, albeit a relatively minor one comparedto LUH, in seed mucilage synthesis or maturation.
Both LUH and LUG directly but independently interact withSEU (Sridhar et al. 2004; Sitaraman et al. 2008). We thustested seu-1 seeds for any seed mucilage defects. seu-1mutants secreted mucilage normally (Figure 3C, G, K) buthad a slightly higher percentage of seeds that lacked mu-cilage (11%) as compared to 6.4% in the wild type (L-er)(Figure 2). This difference however is not statistically significant.Double seu-1; luh-1 mutants exhibit defects identical to luh-1(Figure 3D, H, L), suggesting either that SEU acts in the samepathway as LUH but its function is masked by a redundantgene such as SEUSS_LIKE (SLK) (Bao et al. 2010), or that

402 Journal of Integrative Plant Biology Vol. 53 No. 5 2011
Figure 2. Percentage of seeds that failed to release mucilage
upon imbibition.
Error bar stands for SD based on three biological replicates (except
for ap2–2 with only two biological replicates). “luh-1 (EDTA)” indi-
cates luh-1 seeds treated with ethylene diamine tetra acetate before
they were stained with toluidine blue.
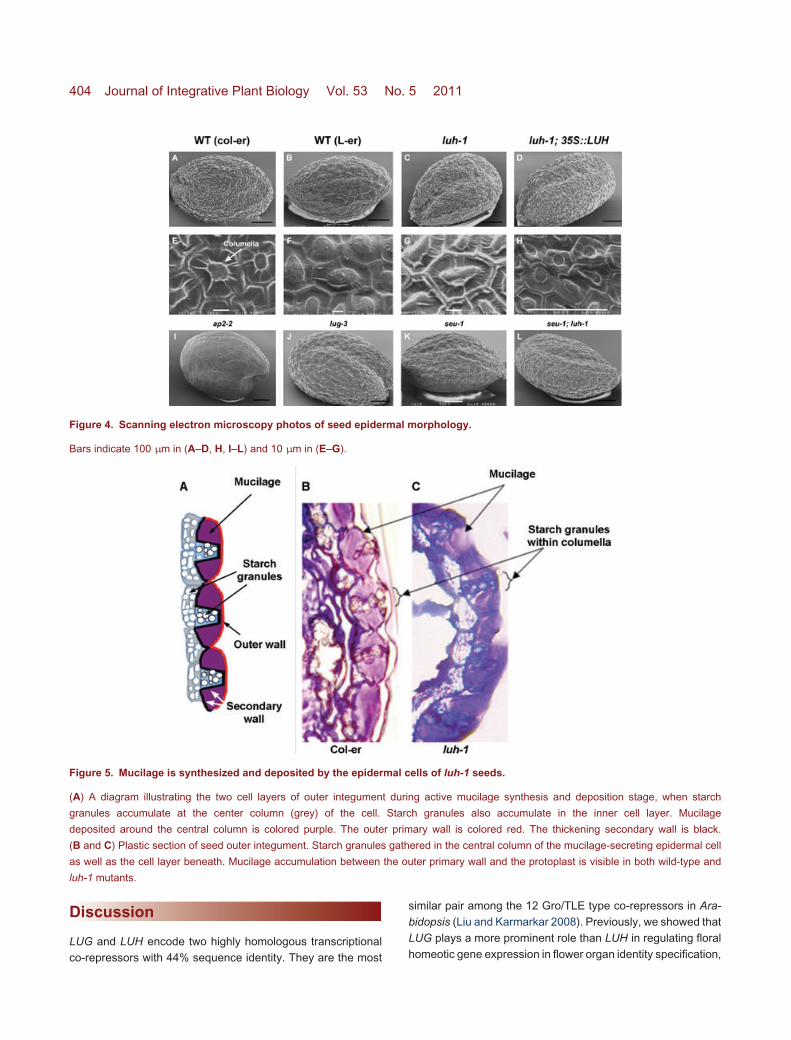
SEU does not play a role in mucilage extrusion. The ap2mutant seeds, which served as a control here, were previouslyreported to exhibit seed mucilage defect due to a failurein outer integument cell differentiation (Jofuku et al. 1994;Western et al. 2001; Ohto et al. 2009). The ap2 mutantseed epidermis lacks columella (Figure 4I) and the underlyingpalisade cells. As a result, ap2 seeds make little to no mucilage(Figure 2; Figure 3A, E, I).
luh-1 seeds do not exhibit significant changes in seedcoat morphology or integument cell differentiation
The absence of seed mucilage release upon imbibition canbe attributed to defects in several dependent or independentprocesses. First, the outer integument cells could develop ordifferentiate improperly, leading to an absence of seed coatlayer and columella, as in ap2 mutants (Figure 4I) (Jofukuet al. 1994; Western et al. 2001). Second, the outer integumentcells could fail to synthesize or secrete mucilage to apoplasticspace, resulting in a reduced or abnormal columella, as inthe case of mum4 (Western et al. 2001, 2004). Third, theouter integument cell layer could synthesize and secrete mu-cilage properly into the apoplastic space but fail to extrudemucilage post-imbibition due to improper modification of mu-cilage, which is similar to mum2 mutants (Western et al. 2001;Dean et al. 2007).
Heavy metal chelating agents such as ethylene diaminetetra acetate (EDTA) can force the release of trappedmucilage in mucilage extrusion mutants such as mum2
(Dean et al. 2007) and patchy/atbxl1 (Arsovski et al.2009) as absence of Ca++ is believed to reduce thebinding of pectin chains through calcium bridges, causingmore extensive hydration of the mucilage and/or weaken-ing the primary wall. We thus tested if EDTA treatmentof luh-1 seeds could help release the trapped mucilage.Indeed, approximately 51% of EDTA-treated luh-1 seeds couldnow release mucilage, as opposed to only 6% of EDTA-untreated luh-1 seeds (Figure 2). However, the secreted mu-cilage layer is significantly thinner than the wild-type control(Figure 1M, N) but resembles EDTA-treated mum2 mutants(Dean et al. 2007). This suggests that either luh-1 synthesizesmucilage at a reduced level or that luh-1 mucilage has limitedability to expand as mum2.
Scanning electron microscopy (SEM) was used to examineepidermal morphology of dry seeds. Wild-type seeds havehexagonal shaped epidermal cells and a protruding centralcolumella in each epidermal cell (Figure 4A, B, E, F). luh-1seed epidermal cells appeared normal with columella in thecenter of each epidermal cell (Figure 4C, G). However, thecolumella appeared more rectangular and had thicker radialwalls compared to the wild type. luh-1; 35S::LUH seeds werewild type-like (Figure 4D, H). lug-3 and seu-1 single (Figure 4J,
K), and seu-1; luh-1 double (Figure 4L) mutant seed epider-mis also appeared normal. Thus, in contrast to ap2 seeds(Figure 4I), luh-1 does not appear to cause defects in in-tegument cell differentiation or mucilage synthesis/secretion.Normal amounts of mucilage secretion into the apoplasticspace are necessary for cytoplasmic rearrangement leading tovolcano-shaped columella (Western et al. 2001). The normalepidermal morphology and the presence of mucilage releasedby EDTA treatment suggest that the mucilage defect of luh-1likely occurs after mucilage synthesis and secretion andmay reside in mucilage modification/maturation similarly tomum2.
To further confirm that mucilage is properly synthesizedand deposited, plastic sections and histological staining wereperformed on the wild-type and luh-1 seeds. luh-1 mutantsproduced and accumulated mucilage in apoplastic space sim-ilarly to wild-type seeds (Figure 5). Each outer integumentcell has a centrally raised columella. Amyloplasts were vis-ible in the cytoplasm of both wild-type and luh-1 cells to-gether with mucilage accumulation in the apoplastic spaceflanking the columella. Therefore, luh-1 is capable of synthe-sizing and depositing mucilage but fails to extrude it uponimbibition.
Examination of MUM2 and other cell wall modificationenzymes in luh-1
Because luh-1 and mum2-1 mutants exhibited highly similarseed mucilage phenotypes, which include normal epidermal

Co-repressors Required for Seed Mucilage Release 403
Figure 3. Mucilage extrusion phenotypes of ap2, seu, and lug.
Toluidine blue staining of unshaken (A–D) and shaken seeds (E–H), and lower magnification photos of imbibed seeds (I–L).
Bars indicate 200 µm in (A–H) and 500 µm in (I) and (L), (J, K) have similar magnifications as L.
(A, E, I) ap2–2.
(B, F, J) lug-3.
(C, G, K) seu-1.
(D, H, L) seu-1; luh-1.
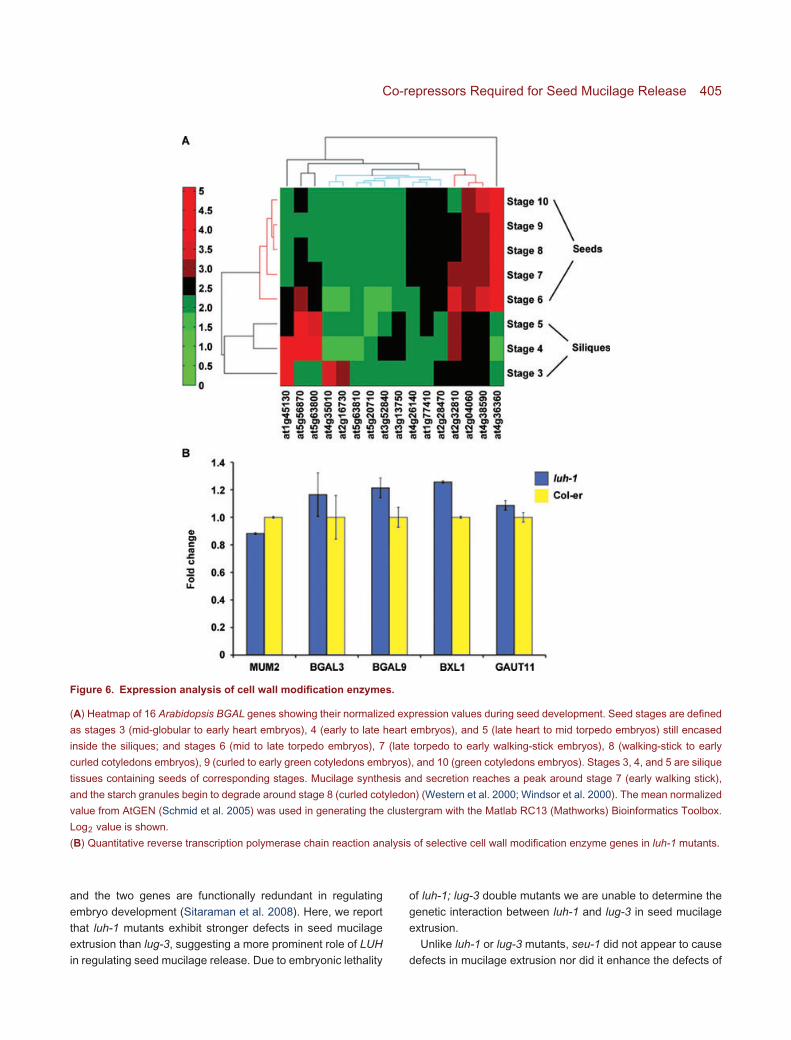
morphology and normal mucilage release upon EDTA treat-ment (Western et al. 2001), LUH and MUM2 may both act inmucilage maturation. While MUM2 encodes a β-galactosidaseinvolved in cleaving the pectin branches, LUH encodes a puta-tive transcriptional co-repressor. LUH may positively regulateMUM2 expression by repressing the expression of a negativeregulator of MUM2. We tested if MUM2 expression is reduced inluh-1 by quantitative reverse transcription polymerase chain re-action (qRT-PCR) using RNA isolated from luh-1 siliques at 7 dpost-anthesis (7 DPA), when mucilage synthesis and modi-fication occurs (Western et al. 2000, 2001). We detected avery slight reduction (12%) of MUM2 in luh-1 (Figure 6B).One possible explanation is that the effect of LUH on MUM2expression is specific in the outer integument tissue and thusdifficult to detect when we extracted RNA from the whole siliquetissue. Alternatively, other BGAL genes in Arabidopsis maybe regulated by LUH, and a reduction of these other BGALgenes in luh-1 is responsible for the mum2-like phenotypein luh-1.
MUM2/BGAL6 is a member of a gene family with 17members. To identify other BGAL genes with a role inseed mucilage maturation in Arabidopsis, we used AtGENmicroarray data (Schmid et al. 2005) to identify BGALgenes that are highly expressed during seed development(Figure 6A). Out of 16 BGAL genes present in the At-GEN data, approximately seven were expressed at relativelyhigher level during seed development: At4g36360 (BGAL3),At4g38590 (BGAL14), At2g04060, At2g32810 (BGAL9),At1g45130 (BGAL5), At5g56870 (BGAL4), and At5g63800(BGAL6/MUM2) (Figure 6A). Several of these were selectedfor further analysis with semi-qRT-PCR (data not shown) andqRT-PCR (Figure 6B) using gene-specific primers. However,none showed any significant difference in mRNA expressionlevel between wild type and luh-1 (Figure 6B). We also testedBXL1 and GAUT11 due to their specific defects in mucilagemodification. luh-1 mutants showed a slight increase of BXL1expression and no significant change in GAUT11 expressioncompared to the wild type (Figure 6B).
404 Journal of Integrative Plant Biology Vol. 53 No. 5 2011
Figure 4. Scanning electron microscopy photos of seed epidermal morphology.
Bars indicate 100 µm in (A–D, H, I–L) and 10 µm in (E–G).
Figure 5. Mucilage is synthesized and deposited by the epidermal cells of luh-1 seeds.
(A) A diagram illustrating the two cell layers of outer integument during active mucilage synthesis and deposition stage, when starch
granules accumulate at the center column (grey) of the cell. Starch granules also accumulate in the inner cell layer. Mucilage
deposited around the central column is colored purple. The outer primary wall is colored red. The thickening secondary wall is black.
(B and C) Plastic section of seed outer integument. Starch granules gathered in the central column of the mucilage-secreting epidermal cell
as well as the cell layer beneath. Mucilage accumulation between the outer primary wall and the protoplast is visible in both wild-type and
luh-1 mutants.
Discussion
LUG and LUH encode two highly homologous transcriptionalco-repressors with 44% sequence identity. They are the most
similar pair among the 12 Gro/TLE type co-repressors in Ara-bidopsis (Liu and Karmarkar 2008). Previously, we showed thatLUG plays a more prominent role than LUH in regulating floralhomeotic gene expression in flower organ identity specification,
Co-repressors Required for Seed Mucilage Release 405
Figure 6. Expression analysis of cell wall modification enzymes.
(A) Heatmap of 16 Arabidopsis BGAL genes showing their normalized expression values during seed development. Seed stages are defined
as stages 3 (mid-globular to early heart embryos), 4 (early to late heart embryos), and 5 (late heart to mid torpedo embryos) still encased
inside the siliques; and stages 6 (mid to late torpedo embryos), 7 (late torpedo to early walking-stick embryos), 8 (walking-stick to early
curled cotyledons embryos), 9 (curled to early green cotyledons embryos), and 10 (green cotyledons embryos). Stages 3, 4, and 5 are silique
tissues containing seeds of corresponding stages. Mucilage synthesis and secretion reaches a peak around stage 7 (early walking stick),
and the starch granules begin to degrade around stage 8 (curled cotyledon) (Western et al. 2000; Windsor et al. 2000). The mean normalized
value from AtGEN (Schmid et al. 2005) was used in generating the clustergram with the Matlab RC13 (Mathworks) Bioinformatics Toolbox.
Log2 value is shown.
(B) Quantitative reverse transcription polymerase chain reaction analysis of selective cell wall modification enzyme genes in luh-1 mutants.
and the two genes are functionally redundant in regulatingembryo development (Sitaraman et al. 2008). Here, we reportthat luh-1 mutants exhibit stronger defects in seed mucilageextrusion than lug-3, suggesting a more prominent role of LUHin regulating seed mucilage release. Due to embryonic lethality
of luh-1; lug-3 double mutants we are unable to determine thegenetic interaction between luh-1 and lug-3 in seed mucilageextrusion.
Unlike luh-1 or lug-3 mutants, seu-1 did not appear to causedefects in mucilage extrusion nor did it enhance the defects of

406 Journal of Integrative Plant Biology Vol. 53 No. 5 2011
luh-1 in seed mucilage extrusion. These results suggest thateither SEU acts in the same pathway as LUH but its function ismasked by a redundant gene such as SEUSS_LIKE (SLK) (Baoet al. 2010), or suggest SEU does not play a role in mucilageextrusion.
Our data strongly support a defect of luh-1 mutants in mu-cilage extrusion rather than mucilage synthesis or deposition.These defects are highly similar to those of mum2, mum1 and,to some extent, patchy/atblx1 and gaut11 (Arsovski et al. 2009;Caffall et al. 2009). Because LUH encodes a transcription co-repressor rather than a cell wall modification enzyme, onepossibility is that LUH regulates the expression of MUM2or other cell wall modification enzymes. In the absenceof LUH, these cell wall modification enzymes are eitherexpressed at a lower level or not expressed at all. However,qRT-PCR failed to reveal a significant reduction of MUM2mRNA in luh-1 silique collected at 7 DPA. One possibilityis that LUH is required to positively regulate other BGALgenes with a similar role as MUM2 or LUH may regulateother enzymes involved in mucilage modification or primarywall modification. Nevertheless qRT-PCR failed to detectany expression differences in BGAL3, BGAL9, AtBXL1,and GAUT11 (Figure 6B). Expression of additional BGALs,At4g38590 (BGAL14), At2g04060, was not detectable usingthe same RNA samples (data not shown). The interpretationof these results could be twofold: either we have not yetexamined the correct target gene, or we failed to discover areduction of MUM2 or other BGAL genes due to the averagingeffect of isolating the entire silique tissue. Perhaps, regulationof MUM2 by LUH only occurs in the outer integument cellsand could only be detected in those cell types. Furtherwork such as in situ hybridization or isolation of outerintegument tissues for qRT-PCR will be necessary to testthese alternative possibilities. Finally, LUH could regulateexpression of genes that regulate MUM2 post-transcriptionallysuch as modification of enzyme activities throughphosphorylation.
The seed mucilage secretory cells of Arabidopsis provide anideal model for the discovery of novel genes as well as novelregulatory networks for directing proper synthesis, secretion,and modification of cell wall components. The dramatic andhighly penetrant phenotype of luh mutants in seed mucilageextrusion identified LUH as a major player in cell wall biol-ogy and a regulator, rather than an enzyme, in this highlyspecific stage of seed coat maturation. Considering that plantcell walls are important for all aspects of plant cell growth,development, its interaction with the environment, and the for-mation of specialized structures such as cotton fibers and seedmucilage, understanding how LUH regulates seed mucilagematuration may open doors for new discoveries in plant cell wallbiology.
Materials and Methods
Plant growth and materials
Arabidopsis thaliana wild-type and mutant plants were grownunder long day conditions (16 h light, 8 h dark) at 20 ◦C and 65%humidity. luh-1, seu-1, lug-3 mutants were previously described(Liu and Meyerowitz 1995; Franks et al. 2002; Sitaramanet al. 2008). luh-1 is isolated in a Columbia erecta (Col-er)background; seu-1 and lug-3 are in a Landsberg erecta (L-er)background.
Morphological characterization
For plastic section, siliques were fixed overnight in 4%paraformaldehyde in 0.1 M sodium phosphate buffer (1×PBS), pH 7.0. Next day, samples were washed twice with1× PBS and dehydrated at 1 h intervals with ethanolseries at 30%, 50%, 70%, 95%, and twice with 100%.Infiltration and embedding steps were done using JB-4Embedding Kit according to the manufacturer’s protocol(Polysciences; (www.polysciences.com/SiteData/docs/123/204a24a728c8ffee6471ef33ede1e5d9/123.pdf). Sections (1–2 µm thick) were made using a manual Sorvall Porter-BlumJB-4 Microtome placed in water on superfrost/plus slides(Fisher Scientific), and allowed to dry on a slide warmer at42 ◦C overnight. Sections were stained with 0.01% toluidineblue O in 0.01% sodium borate for 5 min, de-stained, and thenphotographed under a compound scope.
To view seed mucilage extrusion defects, wild-type andmutant seeds of approximately similar harvest and storagetime were stained with 0.01% toluidine blue O in 0.01% sodiumborate for 20 min. They were gently transferred with a pipette toplates filled with water, visualized, and then photographed un-der a Zeiss Stemi 2000C stereoscope. For EDTA pretreatment,seeds were shaken for 90 min in 0.05 M EDTA, then rinsed, andstained with 0.01% toluidine blue O (in 0.01% sodium borate)before viewing.
For SEM, seeds were dry mounted on aluminum stubs (TedPella), coated with gold-palladium in a Denton DV 503 VacuumEvaporator, and photographed with an Amray 1820D SEM at5.0 kV.
RNA extraction and qRT-PCR analysis
Total RNA was isolated from 7DPA siliques of Col-er and luh-1 plants using an RNeasy Plant Mini Kit (Qiagen). First-strandcDNA was synthesized from 1 µg total RNA using a QuantiSureFirst-strand cDNA kit (Accugen Biosciences). Tenfold dilutedcDNA (1 µL) was used as a template in real-time PCR analysis.qRT-PCR reactions used iQ SYBR Green Supermix (Bio-Rad

Co-repressors Required for Seed Mucilage Release 407
Table 1. Gene-specific primers used in quantitative reverse transcription polymerase chain reaction
Gene Forward primer 5′→3′ Reverse primer 5′→3′ Efficiency (%)
MUM2 TTCTCTTCTCCGGTTCCATCCACT TCCAAGCTTAGGCTCATGGAGGTT 97
BGAL3 ACATACCAGGTGGGACTGAAAGG AGGCTGAGGCTTTTGTACAGTTAAGG 109
BGAL9 GGACGTATCAGGTGGGATTGAAGG GAAGGTGAAGCATCGGTCTCCAAA 87
BXL1 TTCAACGCTAAGGTCACCCAACAAG TCAGCACATGTGGGCTTTCCATT 96
GAUT11 CTCTGTCGCTGGTTTAGTTCTC CTTGTGACCTCTTCCGTGAAGT 97
GAPC1 CCAGTCACTGTTTTCGGCATCA AGCTGCAGCCTTGTCTTTGTCA 98
Laboratories), and were run and analyzed on CFX96 Real-Time System (Bio-Rad Laboratories). Annealed temperaturesfor all primers were at 60 ◦C, except for GAUT11, which wasannealed at 57 ◦C. Gene-specific primers used in qRT-PCR arelisted in Table 1. The housekeeping gene GLYCERALDEHYDE-3-PHOSPHATE DEHYDROGENASE C SUBUNIT 1 (GAPC1,At3g04120) was used as a reference gene. The Ct for eachgene was subtracted from the Ct for GAPC1 to yield the �CtWT
and �Ctluh−1. Formula 2−��Ct, where -��Ct equals �CtWT
minus �Ctluh−1, gave the fold expression difference. The errorbar represents SD based on three technical repeats.
Acknowledgements
We would like to thank Parsa Hosseini for gene expressionanalysis (Figure 6A), and co-advisor Dr William Higgins forhis support and guidance to M. B. The authors wish to thankCourtney Hollender for the cover photo and Tim Maugel forscanning electron microscopy assistance (contribution #97 ofthe Laboratory for Biological Ultrastructure, University of Mary-land, College Park). The work was supported by agrant from theNational Science Foundation (IOB0616096) to Z. L.
Received 6 Feb. 2011 Accepted 14 Feb. 2011
References
Arsovski AA, Haughn GW, Western TL (2010) Seed coat mucilage
cells of Arabidopsis thaliana as a model for plant cell wall research.
Plant Signal Behav. 5, 796–801.
Arsovski AA, Popma TM, Haughn GW, Carpita NC, McCann
MC. Western TL (2009) AtBXL1 encodes a bifunctional β-D-
xylosidase/alpha-L-arabinofuranosidase required for pectic arabi-
nan modification in Arabidopsis mucilage secretory cells. Plant
Physiol.150, 1219–1234.
Bao F, Azhakanandam S, Franks RG (2010) SEUSS and SEUSS-
LIKE transcriptional adaptors regulate floral and embryonic devel-
opment in Arabidopsis. Plant Physiol. 152, 821–836.
Caffall KH, Pattathil S, Phillips SE, Hahn MG, Mohnen D (2009)
Arabidopsis thaliana T-DNA mutants implicate GAUT genes in the
biosynthesis of pectin and xylan in cell walls and seed testa. Mol.
Plant 2, 1000–1014.
Conner J, Liu Z (2000) LEUNIG, a putative transcriptional corepressor
that regulates AGAMOUS expression during flower development.
Proc. Natl. Acad. Sci. USA 97, 12902–12907.
Dean GH, Zheng H, Tewari J, Huang J, Young DS, Hwang YT,
Western TL Carpita NC, McCann MC, Mansfield SD, Haughn GW
(2007) The Arabidopsis MUM2 gene encodes a β-galactosidase
required for the production of seed coat mucilage with correct
hydration properties. Plant Cell 19, 4007–4021.
Fahn A (1982) Plant Anatomy, Ed 3. Pergamon Press.
Franks RG, Wang C, Levin JZ, Liu Z (2002) SEUSS, a member
of a novel family of plant regulatory proteins, represses floral
homeotic gene expression with LEUNIG. Development 129, 253–
263.
Jofuku KD, den Boer BG, Van Montagu M, Okamuro JK (1994) Con-
trol of Arabidopsis flower and seed development by the homeotic
gene APETALA2. Plant Cell 6, 1211–1225.
Johnson CS, Kolevski B, Smyth DR (2002) TRANSPARENT TESTA
GLABRA2, a trichome and seed coat development gene of Ara-
bidopsis, encodes a WRKY transcription factor. Plant Cell 14, 1359–
1375.
Li SF, Milliken ON, Pham H, Seyit R, Napoli R, Preston J, Koltunow,
AM, Parish RW (2009) The Arabidopsis MYB5 transcription factor
regulates mucilage synthesis, seed coat development, and trichome
morphogenesis. Plant Cell 21, 72–89.
Liu Z, Karmarkar V (2008) Groucho/Tup1 family co-repressors in plant
development. Trends Plant Sci. 13, 137–144.
Liu Z, Meyerowitz EM (1995) LEUNIG regulates AGAMOUS expres-
sion in Arabidopsis flowers. Development 121, 975–991.
Long JA, Ohno C, Smith ZR, Meyerowitz EM (2006) TOPLESS
regulates apical embryonic fate in Arabidopsis. Science 312, 1520–
1523.
Macquet A, Ralet MC, Loudet O, Kronenberger J, Mouille G,
Marion-Poll A, North HM (2007) A naturally occurring mutation
in an Arabidopsis accession affects a β-D-galactosidase that in-
creases the hydrophilic potential of rhamnogalacturonan I in seed
mucilage. Plant Cell 19, 3990–4006.
Ohto MA, Floyd SK, Fischer RL, Goldberg RB, Harada JJ (2009)
Effects of APETALA2 on embryo, endosperm, and seed coat
development determine seed size in Arabidopsis. Sex Plant Reprod.
22, 277–289.

408 Journal of Integrative Plant Biology Vol. 53 No. 5 2011
Oka T, Nemoto T, Jigami Y (2007). Functional analysis of Ara-
bidopsis thaliana RHM2/MUM4, a multidomain protein involved in
UDP-D-glucose to UDP-L-rhamnose conversion. J. Biol. Chem.
282, 5389–5403.
Penfield S, Meissner RC, Shoue DA, Carpita NC, Bevan MW (2001)
MYB61 is required for mucilage deposition and extrusion in the
Arabidopsis seed coat. Plant Cell 13, 2777–2791.
Rautengarten C, Usadel B, Neumetzler L, Hartmann J, Bussis D,
Altmann T (2008) A subtilisin-like serine protease essential for
mucilage release from Arabidopsis seed coats. Plant J. 54, 466–
480.
Rerie WG, Feldmann KA, Marks MD (1994) The GLABRA2 gene
encodes a homeo domain protein required for normal trichome
development in Arabidopsis. Genes Dev. 8, 1388–1399.
Schmid M, Davison TS, Henz SR, Pape UJ, Demar M, Vingron M,
Scholkopf B, Weigel D, Lohmann JU (2005) A gene expression
map of Arabidopsis thaliana development. Nat. Genet. 37, 501–
506.
Sitaraman J, Bui M, Liu Z (2008) LEUNIG_HOMOLOG and LEUNIG
perform partially redundant functions during Arabidopsis embryo
and floral development. Plant Physiol. 147, 672–681.
Sridhar VV, Surendrarao A, Gonzalez D, Conlan RS, Liu Z (2004)
Transcriptional repression of target genes by LEUNIG and SEUSS,
two interacting regulatory proteins for Arabidopsis flower develop-
ment. Proc. Natl. Acad. Sci. USA 101, 11494–11499.
Sridhar VV, Surendrarao A, Liu Z (2006) APETALA1 and SEPA-
LLATA3 interact with SEUSS to mediate transcription repression
during flower development. Development 133, 3159–3166.
Stahle MI, Kuehlich J, Staron L, von Arnim AG, Golz JF (2009)
YABBYs and the transcriptional corepressors LEUNIG and LE-
UNIG_HOMOLOG maintain leaf polarity and meristem activity in
Arabidopsis. Plant Cell 21, 3105–3118.
Usadel B, Kuschinsky AM, Rosso MG, Eckermann N, Pauly M
(2004) RHM2 is involved in mucilage pectin synthesis and is
required for the development of the seed coat in Arabidopsis. Plant
Physiol. 134, 286–295.
Western TL, Burn J, Tan WL, Skinner DJ, Martin-McCaffrey L,
Moffatt BA, Haughn GW (2001) Isolation and characterization of
mutants defective in seed coat mucilage secretory cell development
in Arabidopsis. Plant Physiol. 127, 998–1011.
Western TL, Skinner DJ, Haughn GW (2000) Differentiation of mu-
cilage secretory cells of the Arabidopsis seed coat. Plant Physiol.
122, 345–356.
Western TL, Young DS, Dean GH, Tan WL, Samuels AL, Haughn
GW (2004) MUCILAGE-MODIFIED4 encodes a putative pectin
biosynthetic enzyme developmentally regulated by APETALA2,
TRANSPARENT TESTA GLABRA1, and GLABRA2 in the Ara-
bidopsis seed coat. Plant Physiol. 134, 296–306.
Windsor JB, Symonds VV, Mendenhall J, Lloyd AM (2000) Ara-
bidopsis seed coat development: Morphological differentiation of
the outer integument. Plant J. 22, 483–493.
Zhang F, Gonzalez A, Zhao M, Payne CT, Lloyd A (2003) A net-
work of redundant bHLH proteins functions in all TTG1-dependent
pathways of Arabidopsis. Development 130, 4859–4869.
(Co-Editor: Chun-Ming Liu)





![Identification and Characterization of Arabidopsis …...Identification and Characterization of Arabidopsis Seed Coat Mucilage Proteins1[OPEN] Allen Yi-Lun Tsai2, Tadashi Kunieda3,](https://img.pdfslide.us/doc/110x75/5e93c006afc9c34a843ac831/identiication-and-characterization-of-arabidopsis-identiication-and-characterization.jpg)




![SALT-OVERLY SENSITIVE5 Mediates Arabidopsis Seed Coat … · SALT-OVERLY SENSITIVE5 Mediates Arabidopsis Seed Coat Mucilage Adherence and Organization through Pectins1[W][OPEN] Jonathan](https://img.pdfslide.us/doc/110x75/5e3585e8c9781e4b2f69c445/salt-overly-sensitive5-mediates-arabidopsis-seed-coat-salt-overly-sensitive5-mediates.jpg)


















