Embed Size (px)
Citation preview

23
Lect 23-25. Cell Signaling and Regulation of Gene Expression:1. Regulation of gene expression.
a. Review of gene expressionb. Regulation of gene expression in prokaryotesc. Regulation of gene expression in eukaryotes (lecture notes)d. Inducers: Jacob and Monod diploid analysise. Le Chatelier's Principle
2. Growth factors and cell signaling.a. Growth factors and growth factor receptorsb. Tyrosine phosphorylation: a post-translational strategy for
modulating gene expressionc. Phosphates in biologyd. Ras, GTPases, and biological timescalese. MAP kinase cascade
In the last section we talked about transcriptional regulation -- how the temporal expression ofdifferent genes is controlled. We talked in detail about how repressors and activators that bindto DNA affect transcription of the DNA, and I mentioned that the activity of the repressors andactivators is affected by signals that come from outside the cell. I gave an example of a simplesignal -- a diffusible small molecule that enters the cell and travels to a protein on the DNA,where is binds and causes the protein to fall off. In this section, we are going to talk aboutanother signaling strategy in which signals from the outside of the cell are recognized andinformation about the presence of those signals is transmitted from the cell membrane all theway to the nucleus without the signal itself entering the cell. This signaling strategy involves aform of regulation that is post-translational -- that is, it involves chemical changes to proteinsthat occur after the proteins have been synthesized. These chemical changes affect proteinstructure and, therefore, protein activity (e.g., interactions with other proteins or with DNA).
In the course of this section, we are also going to go back over some basic chemical conceptsthat were introduced earlier -- thermodynamics and kinetics -- because these concepts areimportant to understanding how regulation is achieved in biological systems. Some reactionsthat are common biology occur at uncatalyzed rates that are much longer than the existence oflife on earth! For life to exist, there have to be ways to accelerate reactions that would proceedtoo slowly to be compatible with life. There also have to be ways to make reactions that wouldotherwise be unfavorable occur. Many chemical reactions necessary for life to exist would notoccur spontaneously (because the free energy for the reactions is unfavorable) if they were notcoupled to other reactions that are highly favorable. Finally, we are going to connect thesechemical concepts to biological strategies for achieving control over when particular reactionsoccur in a cell. In biology, there must be mechanisms for turning different pathways on and off inresponse to cellular conditions. We are going to talk about how the timescales of chemicalreactions can be manipulated to create”switches”, or mechanisms for turning differentpathways on and off.
24
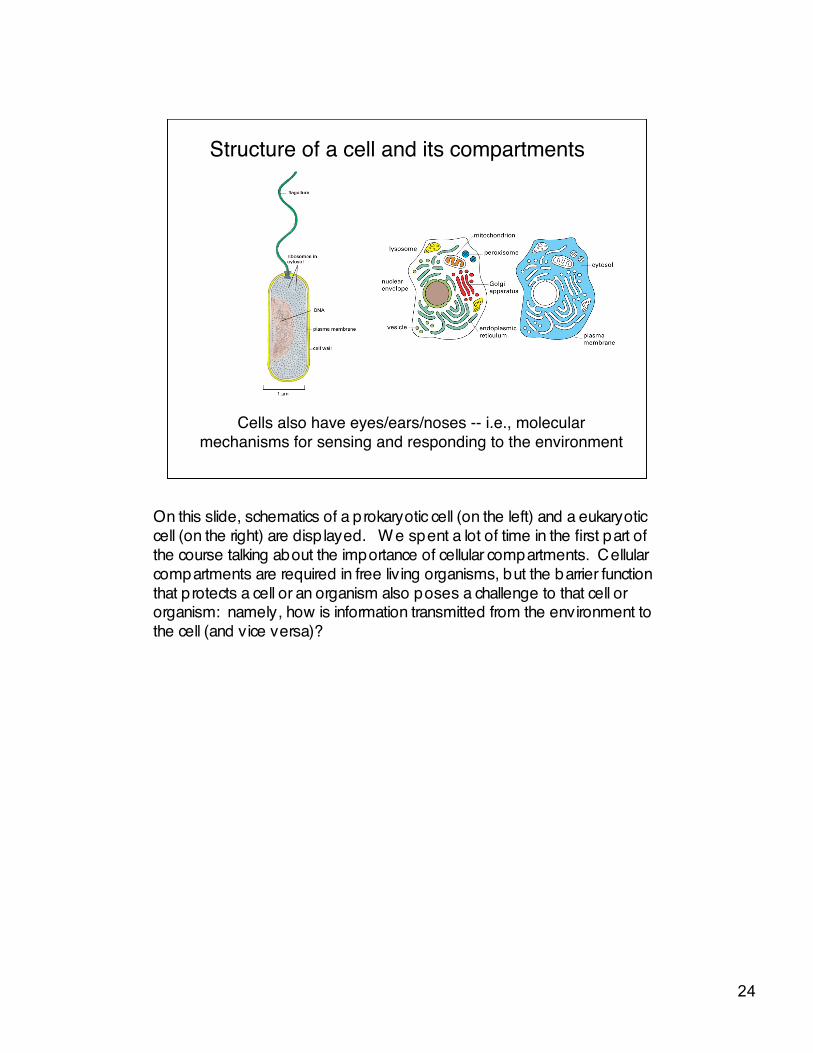
Structure of a cell and its compartments
Cells also have eyes/ears/noses -- i.e., molecularmechanisms for sensing and responding to the environment
On this slide, schematics of a prokaryotic cell (on the left) and a eukaryoticcell (on the right) are disp layed. W e spent a lot of time in the first part ofthe course talking about the importance of cellular compartments. Cellularcompartments are required in free living organisms, but the barrier functionthat protects a cell or an organism also poses a challenge to that cell ororganism: namely, how is information transmitted from the environment tothe cell (and vice versa)?

25
Problem: Not all signals get in
How do you transduce extracellular signals that cannotenter cells?
Lactose
EpidermalGrowth Factor(53 amino acids)
We just learned about a diffusible small molecule, lactose, and howits presence in a cell affects gene expression. Some molecules thatare present in the environment around a cell have structures that donot allow them to get into the cell. Andrew talked about a class ofsuch molecules -- growth factors -- which are polypeptides thatcirculate in the bloodstream and mediate differentiation andproliferation of cells. The protein shown on the left in the slide aboveis called epidermal growth factor (EGF), and it is essential forembryogenesis. Post-development, it plays a critical role in woundhealing. EGF is also made in large amounts by some tumors whereit stimulates cells to enter cell division, thus promoting tumor growth.You might wonder how a molecule such as EGF, which can’t getthrough the lipid bilayer membrane of cells, can affect intracellularevents. As you will learn, growth factors affect gene expression bybinding receptors on cell surfaces. We will learn how binding eventsthat occur on cell surfaces are transduced to signals that ultimatelyregulate gene expression deep inside cells.

26
A recognition event
The first step in signal transduction involves bindingof the growth factor to the receptor
Recognition 1
Growth factors, which you learned about earlier, are extracellularproteins that are important for communication and coordination ofcells in a multicellular organism. They circulate in the blood. Thesemolecules help orchestrate many processes including cell growth anddivision as well as differentiation of cells into more specialized celltypes. Cells must have a sensitive mechanism for detecting thesemolecules, which are present at very low concentrations in theblood. The first step in the detection mechanism involves binding ofthe growth factor to the extracellular portion of a protein, thereceptor, which is embedded in the cell membrane. This is the“recognition” step. “Recognition” is a term used to denote thespecific interaction of molecules. The interactions between twomolecules (proteins) could involve any of the electrostatic (ionic --charge, dipolar -- partial charge, or Van der Waals -- induced charge)or hydrpphobic forces we talked about as being important instabilizing protein structures. How two molecules can bind to oneanother selectively in the presence of enormous numbers of othermolecules is a fundamental principle in many biological interactions.

27
Growth factor receptors
Extracellular growthfactor binding domain
Transmembrane domain
Intracellular proteinkinase domain
In the absence of the growth factor, the growth factorreceptor kinase domain is inactive
We said that the signaling molecule, the growth factor, binds to areceptor embedded in the cell membrane. The receptors that bindto growth factors consist of three parts: 1) an extracellular domainthat “recognizes” (i.e., binds to) the growth factor; 2) atransmembrane domain that spans the lipid bilayer of the plasmamembrane; and 3) an intracellular domain that has enzymaticactivity, typically protein kinase activity. The extracellular domaincontains a pocket/cleft/surface that is complementary in shape andcharge characteristics to the growth factor. The transmembranedomain typically consists of hydrophobic residues arrayed in a helicalconformation so that all the hydrogen bonds are internally satisfiedin the nonpolar interior of the bilayer. The intracellular domaincontains an enzymatic activity that is “off” unless the growth factoris bound.

28
Activation of the receptor
inactive, monomeric active, dimeric
ligand
Ligand binding causes the receptor to dimerize, whichactivates the kinase
Many receptors, such as growth factor receptors, exist in twostates, inactive and active. When no ligand (growth factor) isbound, the receptor is monomeric and the intracellular enzymeactivity is “off”. Growth factor binding causes the association ordimerization of two molecules of growth factor receptor. (This isone major mechanism for how extracellular signaling molecules affecttransmembrane receptors; there are other mechanisms.) Thisdimerization turns on the enzymatic activity in the intracellulardomain. In the case of growth factor receptor, the intracellulardomain has tyrosine kinase activity. Protein kinases are enzymesthat transfer phosphate groups to reactive amino acid side chains.In the growth factor receptor, the amino acid side chain that getsphosphorylated is a tyrosine side chain. The tyrosine side chainphosphorylated by growth factor receptor is located in theintracellular domain of the receptor itself. One monomer of thedimeric receptor phosphorylates the other monomer of the dimericreceptor and vice versa. (This is why dimerization activates!) Thisprocess is called autophosphorylation.
Eventually, enzymes called phosphatases dephosphorylate thereceptor turning it off again. Sometimes receptors have both kinaseand phosphatase activity and it is the presence or absence of aligand that determines the activity.

29
Protein kinases and phosphorylation
N
NN
N
NH2
O
OHOH
OPO
O-
O
HN
R O
OH
R PO
O-
O
P-O
O-
O
HN
R O
O
R
PO-O
O-
N
NN
N
NH2
O
OHOH
OPO
O-
O
P-O
OH
O+ +
ATP ADPTyrosine Residue Phosphorylated Tyrosine
Other Side Chains That Can Be Phosphorylated:
HN
R O
OHR
Serine Residue
HN
R O
OHR
Threonine Residue
Kinase
!"#
We said that a protein kinase is an enzyme that catalyzes thephosphorylation of amino acids containing reactive side chains suchas serine, threonine or tyrosine. ATP is the phosphodonorsubstrate. In a kinase-catalyzed reaction, the hydroxyl group of theamino acid side chain attacks the terminal, or gamma, phosphate ofATP, resulting in transfer of the phosphate to the hydroxyl group ofthe side chain. Most receptors for growth factors have tyrosinekinase activity, but other protein kinases phosphorylate serine andthreonine residues. In general, protein kinases phosphorylate oneor a small number of amino acids in a given protein. Kinases are alsospecific. That is, even though there are thousands of proteinsinside cells, each kinase only phosphorylates certain proteins. As wewill see in a later lecture, the substrate specificity of kinases comesfrom their interactions with substrates - they frequently “recognize”(bind to) specific sequences found in their substrates.

30
Why do we need protein kinases?
ΔGoverall <<< 0
N
NN
N
NH2
O
OHOH
OPO
O-
O
PO
O-
O
P-O
O-
O
ATP
! " #
+
N
NN
N
NH2
O
OHOH
OPO
O-
O
P-O
O-
O
OH
OP
O-O
O-+H2O+
Phenol
ADP
Phosphate Ester
H3O++
We have talked a lot about free energy and how to tell whether reactionsare thermodynamically favorable -- i.e., whether they will proceedspontaneously from reactants to products under a standard set ofconditions. We said that the magnitude of the free energy change cantell you how far the equilibrium lies to one side or the other.Phosphorylation reactions are very favorable: the free energy change forhydrolysis of ATP is large and negative. You might wonder, therefore,why we need protein kinases. One reason is that the kinase providesspecificity so that only the correct proteins are phosphorylated. Anotherreason is that the kinase accelerates the rate of the reaction. Eventhough phosphorylation is thermodynamically favorable, it is very slow --too slow to be compatible with a biological process. We need kinases toaccelerate the rate of phosphorylation.
31
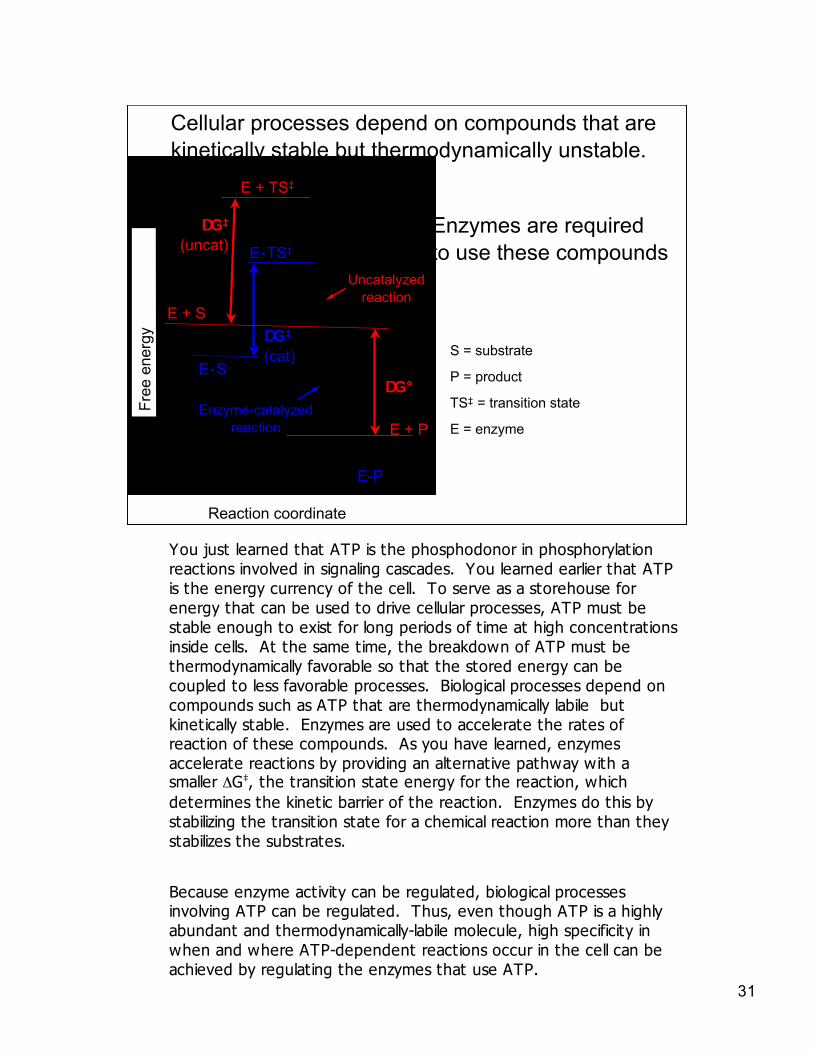
S = substrate
P = product
TS‡ = transition state
E = enzyme
Uncatalyzedreaction
Enzyme-catalyzedreaction
E.S
E.TS‡
E-P
E + S
E + TS‡
E + P
DG‡
(cat)
Reaction coordinate
Free
ene
rgy
DG‡
(uncat)
DG°
Cellular processes depend on compounds that arekinetically stable but thermodynamically unstable.
Enzymes are requiredto use these compounds
You just learned that ATP is the phosphodonor in phosphorylationreactions involved in signaling cascades. You learned earlier that ATPis the energy currency of the cell. To serve as a storehouse forenergy that can be used to drive cellular processes, ATP must bestable enough to exist for long periods of time at high concentrationsinside cells. At the same time, the breakdown of ATP must bethermodynamically favorable so that the stored energy can becoupled to less favorable processes. Biological processes depend oncompounds such as ATP that are thermodynamically labile butkinetically stable. Enzymes are used to accelerate the rates ofreaction of these compounds. As you have learned, enzymesaccelerate reactions by providing an alternative pathway with asmaller ΔG‡, the transition state energy for the reaction, whichdetermines the kinetic barrier of the reaction. Enzymes do this bystabilizing the transition state for a chemical reaction more than theystabilizes the substrates.
Because enzyme activity can be regulated, biological processesinvolving ATP can be regulated. Thus, even though ATP is a highlyabundant and thermodynamically-labile molecule, high specificity inwhen and where ATP-dependent reactions occur in the cell can beachieved by regulating the enzymes that use ATP.
32
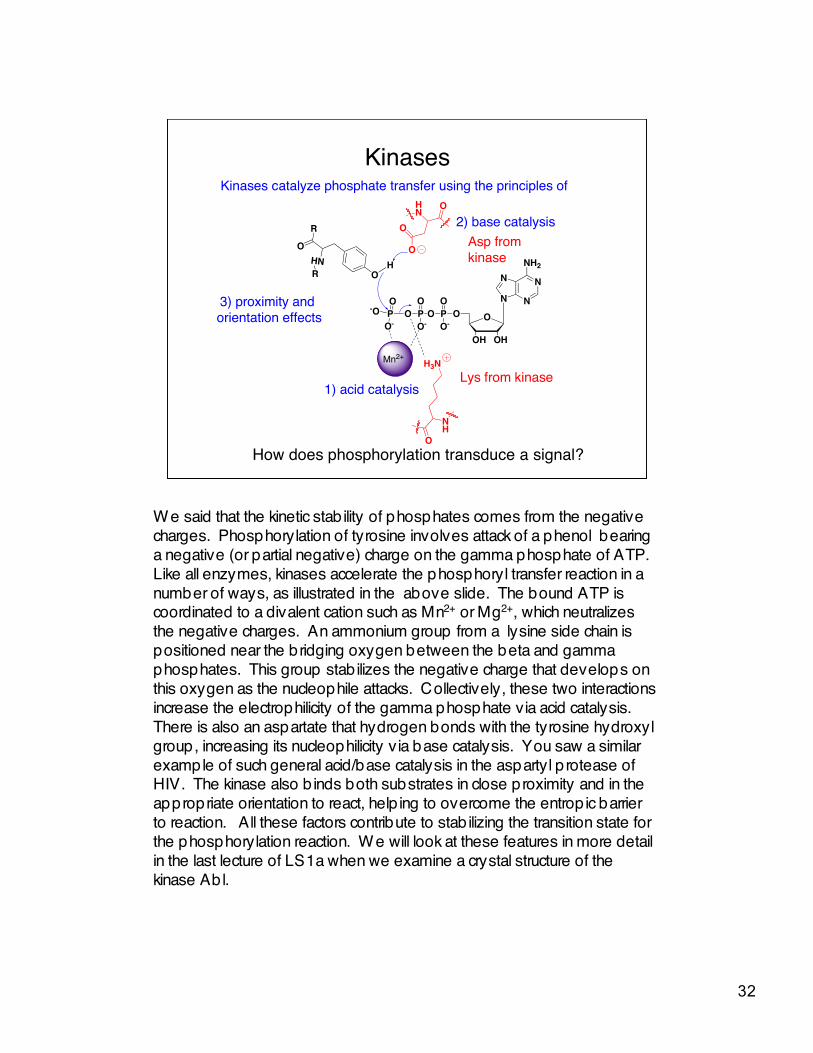
Kinases
Asp fromkinase
Lys from kinase
3) proximity and orientation effects
1) acid catalysis
2) base catalysis
How does phosphorylation transduce a signal?
Kinases catalyze phosphate transfer using the principles of
N
NN
N
NH2
O
OHOH
OPO
O-
O
HN
R
O
OR
PO
O-
O
P-O
O-
O
H
O
O
HN
O
NH
O
Mn2+
H3N
W e said that the kinetic stab ility of phosphates comes from the negativecharges. Phosphorylation of tyrosine involves attack of a phenol bearinga negative (or partial negative) charge on the gamma phosphate of ATP.Like all enzymes, kinases accelerate the phosphoryl transfer reaction in anumber of ways, as illustrated in the above slide. The bound ATP iscoordinated to a divalent cation such as Mn2+ or Mg2+, which neutralizesthe negative charges. An ammonium group from a lysine side chain ispositioned near the bridging oxygen between the beta and gammaphosphates. This group stab ilizes the negative charge that develops onthis oxygen as the nucleophile attacks. Collectively, these two interactionsincrease the electrophilicity of the gamma phosphate via acid catalysis.There is also an aspartate that hydrogen bonds with the tyrosine hydroxylgroup, increasing its nucleophilicity via base catalysis. You saw a similarexample of such general acid/base catalysis in the aspartyl p rotease ofHIV. The kinase also b inds both substrates in close proximity and in theappropriate orientation to react, help ing to overcome the entrop ic barrierto reaction. All these factors contribute to stab ilizing the transition state forthe phosphorylation reaction. W e will look at these features in more detailin the last lecture of LS1a when we examine a crystal structure of thekinase Abl.
33
N
O
O
OH
N
O
O
OH
N
O
O
OPO
OO
PO
OO
N
O
O
O
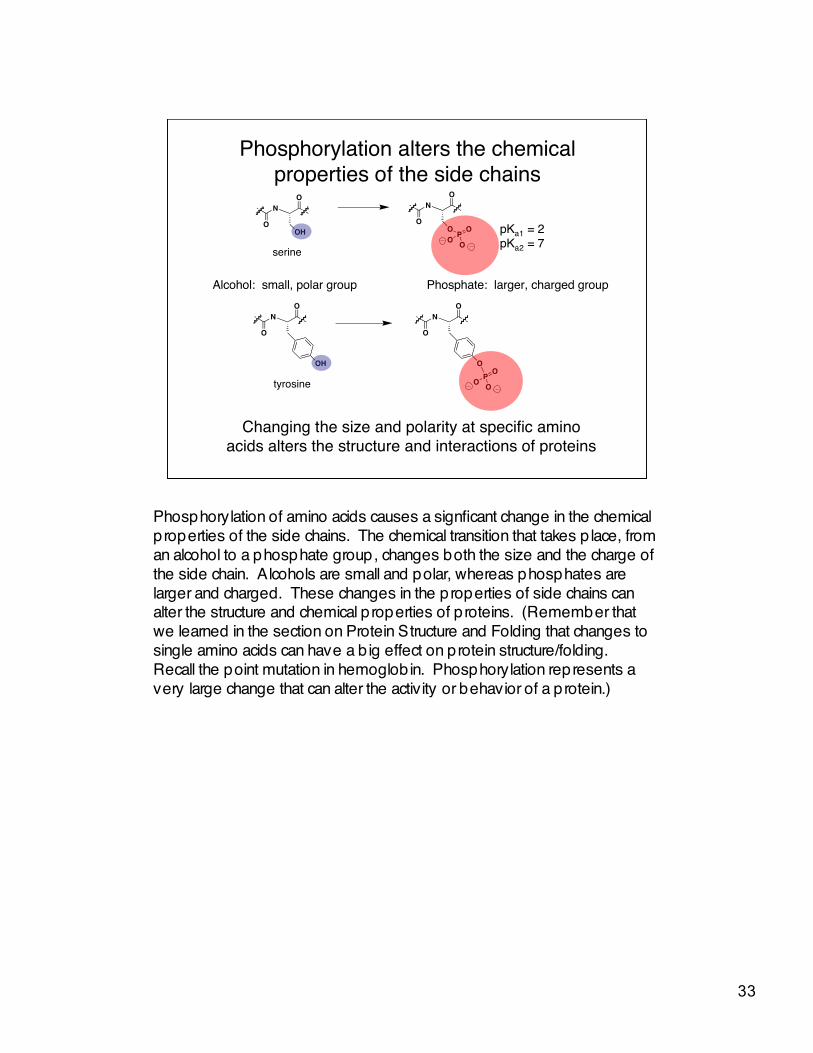
Phosphorylation alters the chemicalproperties of the side chains
Alcohol: small, polar group Phosphate: larger, charged group
serine
tyrosine
pKa1 = 2pKa2 = 7
Changing the size and polarity at specific aminoacids alters the structure and interactions of proteins
Phosphorylation of amino acids causes a signficant change in the chemicalproperties of the side chains. The chemical transition that takes p lace, froman alcohol to a phosphate group, changes both the size and the charge ofthe side chain. Alcohols are small and polar, whereas phosphates arelarger and charged. These changes in the properties of side chains canalter the structure and chemical properties of proteins. (Remember thatwe learned in the section on Protein Structure and Folding that changes tosingle amino acids can have a b ig effect on protein structure/folding.Recall the point mutation in hemoglobin. Phosphorylation represents avery large change that can alter the activity or behavior of a protein.)

34
General ways phosphorylation canaffect function:
• Regulation of enzymatic activity• Subcellular localization• Stability• Association with other molecules
Phosphorylation is one of the most common post-translationalmodifications of proteins. It is estimated that 30% of proteins in eukaryoticcells are phosphorylated. Phosphorylation regulates protein function inmany different ways - by controlling enzyme activty, by regulating thelocation of proteins within a cell, by controlling stab ility, or by regulatingassociation with other molecules. All of these effects on protein functioncan be traced back to changes in protein structure and chemical propertiescaused by phosphorylation.
Given the importance of phosphates in our body (DNA, ATP, etc),before we go on to talk about how autophosphorylation of growthfactor receptor leads to a cascade of signaling events, we need tosummarize how the features that make ATP ideal as a biologicalphosphodonor.

35
Westheimer, F. H. Science, 1987, 235, 1173.
As we have seen in this course, nature uses phosphates in manycontexts - in nucleic acids, in protein phosphorylation, in storing energy inATP. W e saw when we talked about DNA that phosphates are stab le -the integrity of our genetic material relies on DNA being resistant tohydrolysis. W e have just said that ATP itself is kinetically stab le. How wethink about the role of ATP in b iology is influenced by an analysisdeveloped by Professor F. W estheimer of Harvard University anddescribed in the reference given on this slide. W e will discuss the centralfeature of the analysis on the next slide.

36
ATP hydrolysis• The activation barrier (DG‡ ) to ATP hydrolysis is
large due to electrostatic repulsion.pKa1 = 2.15
pKa2 = 7.20
pKa3 = 12.15
pKa1 = 3.13
pKa2 = 4.76
pKa3 = 6.40
O
PHO
OH
OH
Phosphoric Acid
HO OH
OOOH
O OH
Citric Acid
Negative charges on the oxygens make ATP kinetically stable.
P
OO
O
+RO O
O
P
O
OO P
O
O
O
RO
!- !-
RO
W estheimer pointed out that the kinetic stab ility of phosphates is a function of thenegative charges that are present at physiological pH. Phosphoric acid has threeionizable groups. One has a pKa around 2 and is fully ionized at physiological pH.The second and third groups have pKas around 7 and 12, respectively.Therefore, a substantial population of the phosphoric acid molecules (~50%)present in solution at physiological pH are doubly ionized; the rest are monoanions.You can infer from the high pKa of the third ionizable goup on phosphoric acid that itmust be highly unfavorable to form a phosphate trianion at physiological pH. Youshould use the Henderson-Hasselbalch equation to convince yourself about this.(In contrast, the trianion of citric acid can be readily formed at physiological pHbecause the anions are further apart and each one can delocalize its electrons into itsadjacent carbonyl.) Hydrolysis of a phosphate dianion involves attack of ahydroxide ion on the phosphorus atom to form the hydrolyzed products via apenta-coordinate high energy transition state containing essentially three negativecharges around the phosphorus atom. Knowing that the thermodynamic stab ility ofphosphate trianion is very low, it is not difficult to infer that the energy of thetransition state would be exceedingly high, resulting in a large ΔG” and hence a slowreaction. Another way of thinking about why phosphate hydrolysis is slow is thatthere is a huge electrostatic barrier to approach of the phosphate dianion by thenegatively charged hydroxide ion. Because like charges repel, you have to do alot of electrochemical work to bring three like charges in close proximity.(Remember the first time you realized how hard you have to work to bring the “like”poles of two magnets together?)

37
Reaction Kinetics
Asp fromkinase
Lys fromkinase
Rate of reaction = Collision frequency
X Probability thatmolecules collide inthe right orientation
Probability thatmolecules collidewith enough energyto react
X
= [ SM ] (depends on P)
XProbability thatmolecules collide inthe right orientation
Probability thatmolecules collidewith enough energyto react
XMolecularvelocity
(depends onT)
X Reactioncross-section(depends on
size and shape)
X
= [ SM ] Rateconstant (k)
XN
NN
N
NH2
O
OHOH
OPO
O-
O
HN
R
O
OR
PO
O-
O
P-O
O-
O
O
O
HN
O
H
NH
O
Mn2+
H3N
Enzymes change the rate of a reaction by affecting the rate constant (k). As you knowfrom David’s lectures, k is proportional to exp(-ΔG‡/RT). If you rep lace ΔG‡ with ΔH‡ -TΔS‡, you can easily derive that k is proportional to exp(- ΔH‡/RT).exp(ΔS ‡/R). As you willsee in this slide, the four effects that David introduced as being key to how enzymesaccelerate reactions can be thought of as affecting either the ΔH‡ or the ΔS ‡ term in the kexpression.
The rate of reaction can also be broken down into the set of terms shown on this slide.The factors that involve how fast a reaction will occur are: 1) the collision frequency (sincein order to react, the molecules need to come together); 2) the probability that themolecules collide in the right orientation (since if the right ends of molecules don’t collide,nothing will happen); and 3) the probability that the molecules collide with enough energyto react (since even if the right ends of the molecules collide, nothing will happen if thereisn’t enough energy for bonds to be broken and formed). The collision frequencydepends on the concentration of the starting materials, how fast the molecules are movingin solution (which depends on temperature), and the shape and size of the molecules(i.e., a property that we call the “reaction cross-section”, which simply means that thebigger the reacting parts of the molecule are, the more likely they are to bump into eachother at a given concentration and velocity).
(continues next page.)

38
Reaction Kinetics
Asp fromkinase
Lys fromkinase
Rate of reaction = Collision frequency
X Probability thatmolecules collide inthe right orientation
Probability thatmolecules collidewith enough energyto react
X
= [ SM ] (depends on P)
XProbability thatmolecules collide inthe right orientation
Probability thatmolecules collidewith enough energyto react
XMolecularvelocity
(depends onT)
X Reactioncross-section(depends on
size and shape)
X
= [ SM ] Rateconstant (k)
XN
NN
N
NH2
O
OHOH
OPO
O-
O
HN
R
O
OR
PO
O-
O
P-O
O-
O
O
O
HN
O
H
NH
O
Mn2+
H3N
As you know, enzymes increase reaction rates through proximity and orientation effectsas well as other factors such as acid and base catalysis. Enzymes effectively influencethe collision frequency by trapping the substrates when they collide and holding themclose together (within bonding distance). Enzymes also influence the probability thatmolecules will collide in the right orientation to react by fixing the substrates in the rightorientation. Incidentally, both of these effects alter the ΔS ‡ term in the rate constant (kproportional to exp(- ΔH‡/RT).exp(ΔS ‡/R). Acid and base catalysis effects can beconsidered as enzymes influencing the probability that the molecules collide with enoughenergy to react. As you might expect, acid and base catalysis thus modify the ΔH‡ term inthe rate constant.
W hat you want to learn from this slide is that the four effects about how an enzymecatalyzes a reaction all directly perturb the rate constant of that reaction. Proximity andorientation effects affect the ΔS ‡ term while acid and base catalysis affect the ΔH‡ term.The overall effect is then to stab ilize the transition state(s) of the reaction, provide apathway with a lower ΔG‡, and therefore accelerate the rate at which a reaction proceeds.





























