Embed Size (px)
Citation preview

NATURE IMMUNOLOGY VOLUME 7 NUMBER 11 NOVEMBER 2006 1127
Learning from the lost: new insights into TCR specificityNathan J Felix & Paul M Allen
Contacts between T cell receptors and peptide–major histocompatibility complex molecules require specific amino acid interactions. However, some amino acids have no direct involvement in contact but nevertheless are critical for specificity.
Peripheral T cells are the ‘lucky ones’ that have survived the challenges of positive and
negative selection in the thymus, which makes them useful for studying T cell recognition of antigen bound to major histocompatibility complex molecules (MHC), a key factor in both positive and negative selection. However, many unresolved issues remain about how antigen and MHC specificity for a given T cell is achieved. T cell receptors (TCRs) have a ‘germline-encoded preference’ for interaction with MHC molecules, the details of which remain elusive1. No common binding mode between TCR and peptide-MHC has been found despite the solution of over two dozen TCR-peptide-MHC crystal structures2,3. Missing from most of those analyses are the T cells ‘lost’ as a result of thymic selection (TCRs with affinity for peptide-MHC that is too high or too low). In this issue of Nature Immunology, Huseby et al. have taken a new approach by examining T cells normally eliminated by nega-tive selection and thus absent from the periph-eral TCR repertoire4.
The study Huseby et al. presents two new find-ings: first, the number of high-affinity interac-tions between a given TCR and side chains of both peptide and MHC in the central area of the contact surface is much higher for T cells that have undergone negative selection; and second, certain amino acid side chains on the peptide-MHC surface can actually ‘disrupt’ the TCR-peptide-MHC interaction, even though the side chains are not involved in binding to the TCR. Notably, such disrupting side chains neverthe-less directly contribute to the specificity of T cell recognition.
Although previous biophysical analyses of several different TCR-peptide-MHC interac-tions have yielded some important insights5,6, no single parameter (such as ‘on rate’, ‘off rate’ or heat capacity) has fully correlated with all of the observed biological responses1. Complicating analysis of the TCR-peptide-MHC interaction is its relatively low affin-ity. Although surface plasmon resonance is widely used for measuring affinity, the affin-ity of biologically relevant TCR-peptide-MHC interactions (1 mM to 1 μM) is near or below the limit of sensitivity and/or practicality of surface plasmon resonance. In contrast to many receptor-ligand interactions involving monomorphic ligands and receptors, a typi-cal peripheral repertoire of TCRs will have the potential to interact with multiple MHC molecules and a limitless number of peptide antigens.
One approach for studying TCR-peptide-MHC interactions is direct amino acid sub-stitution to determine the effect on binding. However, any systematic analysis of TCR-peptide-MHC interactions deemed adequate would require a large number of amino acid substitutions in the peptide and the MHC. Although amino acid substitution of a given peptide is readily accomplished syntheti-cally7,8, such methodology for a given MHC is much more difficult. Only a limited number of single substitutions of MHC molecules have been produced and studied biochemically, and the difficulty in producing the required amounts of a large number of individual amino acid substitutions for a given MHC molecule makes this approach unfeasible.
The Kappler and Marrack group took a dif-ferent approach. They applied the technique of covalently linking peptides to MHC class II molecules, which fixes and limits the bound peptide9. They also used mice expressing I-Ab
molecules with a covalently linked model peptide, 3K, as the only peptide-MHC class II molecule (‘I-Ab-SP’ mice)10. This single peptide-MHC complex expressed in I-Ab-SP mice generates a large repertoire of T cells that has undergone positive but not negative selec-tion because of the lack of any endogenous self peptide–MHC complexes. The generation of T cells reactive to 3K peptide from normal C57BL/6 and I-Ab-SP mice has shown that the 3K-reactive T cells from C57BL/6 mice are highly specific, whereas those from the I-Ab-SP mice are cross-reactive to both peptide and MHC10. Notably, one of the cross-reactive T cells from an I-Ab-SP mouse could recog-nize both MHC class I and class II molecules, demonstrating the broad reactivity of these non–negatively selected T cells.
Using another innovative technique, Huseby et al. have undertaken detailed ther-modynamic comparison of two specific and three cross-reactive T cells (from C57BL/6 mice and from I-Ab-SP mice, respectively), all with similar binding affinities for pep-tide-MHC. Their flow cytometry–based approach relies on the generation of mul-tiple covalent peptide-MHC molecules each with a single substitution in either the peptide or the MHC and individual expres-sion of each mutant on the surface of insect cells. They used phycoerythrin-labeled TCR multimers in binding assays with the panel of peptide-MHC mutants (measuring mean fluorescence by flow cytometry) to assess the affinity of a given TCR-peptide-MHC inter-action. Notably, they found a direct relation-ship between the affinity of the interactions as measured by surface plasmon resonance and their flow cytometry–based approach for a defined set of TCR-peptide-MHC interac-tions. They then used that new approach to calculate the change in Gibb’s free energy for
Nathan J. Felix and Paul M. Allen are in the
Department of Pathology and Immunology,
Washington University School of Medicine,
St. Louis, Missouri 63110 USA.
e-mail: [email protected]
N E W S A N D V I E W S©
2006
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eim
mun
olog
y

a large matrix of TCR-peptide-MHC inter-actions (five different TCRs and five distinct amino acid substitutions at each of five pep-tide and ten MHC positions) to identify the amino acid side chains critical for the interac-tions. Classical biochemists may object that this assay is not the correct way to assess the energetic contributions of individual residues in a protein-protein interface, but this assay does seem valid, and the extensive analysis of the more than 375 different combinations evaluated by Huseby et al. would simply not be possible by any other approach.
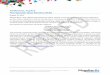
By mapping the ‘hot spots’ of the TCR-pep-tide-MHC interactions11 (alanine substitu-tions resulting in a decrease in binding affinity of more than 1.5 kcal/mol), Huseby et al. have found a substantial difference between TCRs that are specific and those that are cross-reac-tive. Although both TCR ‘types’ seem to have interfaces of similar size with peptide-MHC molecules, specific TCRs have many more ‘hot spots’ than do cross-reactive TCRs: spe-cific TCRs have seven or eight hot spots, yet the cross-reactive TCRs have only two to four (Fig. 1, top). From such data, the authors con-clude that the cross-reactive T cells generate
their binding energy in a very different way than do the specific T cells, a difference that is probably directly involved in the elimination of cross-reactive T cells during negative selec-tion. Such a conclusion provides a molecular basis for the published observation that nega-tive selection, not positive selection, is the main process by which an antigen-specific T cell repertoire is generated10,12.
In their analysis of multiple amino acid substitutions at different MHC or peptide positions, Huseby et al. note that substitution of some side-chain amino acids with alanine has no effect, from which they conclude that the ‘parental’ side chains make no important contacts with the TCR. However, they also note that substitution with amino acids other than alanine are not allowed at those positions, thereby contributing to the overall specificity of the TCR-peptide-MHC interaction. Such side chains (which tolerate replacement with alanine but not with certain other amino acids) are referred to as ‘interface-disrupting’ residues.
The ‘interface-disrupting’ residues do not simply reflect the replacement of small side chains with large or charged ones. Most of
the disrupting residues are located on the MHC surface, although some are on the 3K peptide, and they ‘surround’ the core, high-affinity residues of the TCR-peptide-MHC interface (Fig. 1, bottom). The identifica-tion of ‘interface-disrupting’ residues on the 3K peptide itself is not unexpected, as ‘non-allowable’ peptide substitutions have been reported before. The MHC ‘interface-disrupting’ residues, however, are different in that they have not been identified before, nor has such extensive mutagenesis of an MHC molecule been done before. The demonstra-tion that ‘interface-disrupting’ residues in both the peptide and the MHC could con-tribute to TCR-peptide-MHC specificity is a new finding with important implications for MHC restriction. For example, the inability of a given TCR to recognize different pep-tide-MHC surfaces could be due in part to the presence of residues that interfere with high-affinity binding rather than to lack of specific recognition.
This study by Huseby et al. has provided both a new technique for studying the ener-getics of the TCR-peptide-MHC interaction and new insights into antigen and MHC specificity. The cross-reactive T cells exam-ined had undergone positive selection but not negative selection; future studies should evaluate other ‘lost’ T cells, such as thymo-cytes that have not undergone either positive or negative selection, with the goal of gaining new perspective on the ‘germline-encoded preference’ of TCR for peptide-MHC. It is still amazing that more ways by which TCRs can interact with peptide-MHC surfaces con-tinue to be identified. Many more normal and ‘lost’ T cells must be studied functionally, bio-physically and structurally before complete understanding of the critical TCR-peptide-MHC interaction is finally achieved.
1. Krogsgaard, M. & Davis, M.M. Nat. Immunol. 6, 239–245 (2005).
2. Rudolph, M.G., Stanfield, R.L. & Wilson, I.A. Annu. Rev. Immunol. 24, 419–466 (2006).
3. Garcia, K.C. & Adams, E.J. Cell 122, 333–336 (2005).
4. Huseby, E.S., Crawford, F., White, J., Marrack, P. & Kappler, J.W. Nat. Immunol. 7, 1191–1199 (2006).
5. Kersh, G.J., Kersh, E.N., Fremont, D.H. & Allen, P.M. Immunity 9, 817–826 (1998).
6. Krogsgaard, M. et al. Mol. Cell 12, 1367–1378 (2003).
7. Donermeyer, D.L., Weber, K.S., Kranz, D.M. & Allen, P.M. J. Immunol. (in the press).
8. Reay, P.A., Kantor, R.M. & Davis, M.M. J. Immunol. 152, 3946–3957 (1994).
9. Fremont, D.H., Hendrickson, W.A., Marrack, P. & Kappler, J. Science 272, 1001–1004 (1996).
10. Huseby, E.S. et al. Cell 122, 247–260 (2005).11. Clackson, T. & Wells, J.A. Science 267, 383–386
(1995).12. Huseby, E.S., Crawford, F., White, J., Kappler, J. &
Marrack, P. Proc. Natl. Acad. Sci. USA 100, 11565–11570 (2003).
1128 VOLUME 7 NUMBER 11 NOVEMBER 2006 NATURE IMMUNOLOGY
Specific TCRKd = 7 µm
Residues involved inhigh-affinity binding
Specificity
WT → Alanine
Effect on ∆∆G
> 1.5 kcal/mol
Cross-reactive TCRKd = 8 µm
Positions that contributeto specificity
Specificity
WT → Alanine
→ Other
amino acids
Effect on ∆∆G
None
> 0.8 kcal/mol
Figure 1 Specific and cross-reactive TCRs have distinct interactions with peptide-MHC surfaces. The positions of critical amino acid interactions for a specific TCR (left) and a cross-reactive TCR (right) are presented on the peptide-MHC surface (peptide is yellow). Red boxes (top) indicate residues critically involved in high-affinity binding, as determined by alanine substitution. The specific TCR has many more ‘hot spots’ than does the cross-reactive TCR, even though they have approximately the same overall affinity. These findings show that the cross-reactive TCRs achieve their binding energy in a different way than do specific T cells (such as through more main-chain interactions) and that this difference most likely is involved in their normal elimination from the repertoire by negative selection. Blue cylinders (bottom) indicate positions of ‘interface-disrupting’ residues that contribute to the specificity. The side chains at these positions are not directly involved in high-affinity binding, as alanine substitution has no effect. They are located on the periphery of the peptide-MHC surface surrounding the high-affinity residues. The ‘interface-disrupting’ residues still contribute to the specificity of TCR-peptide-MHC interactions, as substitution with amino acids other than alanine is disruptive. Kd, dissociation constant; WT, wild-type; ΔΔG, change in Gibb’s free energy.
NEWS AND V IEWS©
2006
Nat
ure
Pub
lishi
ng G
roup
ht
tp://
ww
w.n
atur
e.co
m/n
atur
eim
mun
olog
y









![IMMUNOGLOBULINE E T CELL RECEPTOR T. Strachan e A.P. … · B cell antigen receptor tetramero [ IgH 2 + IgL 2 (Ig oppure Ig )] T cell receptor (TCR) eterodimero TCR /TCR TCR /TCR](https://img.pdfslide.us/doc/110x75/5c017b5c09d3f26f1e8cc6a0/immunoglobuline-e-t-cell-receptor-t-strachan-e-ap-b-cell-antigen-receptor.jpg)