Embed Size (px)
Citation preview

REGULAR PAPER
Leaf developmental stage modulates metabolite accumulationand photosynthesis contributing to acclimation of Arabidopsisthaliana to water deficit
Ilektra Sperdouli • Michael Moustakas
Received: 28 January 2014 / Accepted: 10 March 2014
� The Botanical Society of Japan and Springer Japan 2014
Abstract We examined whether young and mature
leaves of Arabidopsis thaliana in their response to mild
water deficit (MiWD) and moderate water deficit (MoWD),
behave differentially, and whether photosynthetic accli-
mation to water deficit correlates with increased proline
and sugar accumulation. We observed that with increasing
water deficit, leaf relative water content decreased, while
proline and sugar accumulation increased in both leaf-
developmental stages. Under both MiWD and MoWD,
young leaves showed less water loss and accumulated
higher level of metabolites compared to mature leaves.
This, leaf age-related increase in metabolite accumulation
that was significantly higher under MoWD, allowed young
leaves to cope with oxidative damage by maintaining their
base levels of lipid peroxidation. Thus, acclimation of
young leaves to MoWD, involves a better homeostasis of
reactive oxygen species (ROS), that was achieved among
others by (1) increased sugar accumulation and (2) either
increased proline synthesis and/or decreased proline
catabolism, that decrease the NADPH/NADP? ratio,
resulting in a higher level of oxidized state of quinone A
and thus in a reduced excitation pressure, and by (3)
stimulation of the photoprotective mechanism of non-
photochemical quenching, that reflects the dissipation of
excess excitation energy in the form of harmless heat, thus
protecting the plant from the damaging effects of ROS.
Keywords Chlorophyll fluorescence � Excitation
pressure � Non-photochemical quenching � Oxidative
stress � Proline � Soluble sugars
Abbreviations
ETR Electron transport rate
Fv/Fm Potential (maximum) quantum yield of PSII
photochemistry
LWC Leaf relative water content
MDA Malondialdehyde
MiWD Mild water deficit
MoWD Moderate water deficit
NPQ Non-photochemical quenching
PSI, PSII Photosystem I, Photosystem II
PPFD Photosynthetic photon flux density
QA Quinone A
qP Photochemical quenching
ROS Reactive oxygen species
SE Standard error
SWC Soil volumetric water content
UPSII Actual (effective) quantum yield of PSII
photochemistry
Introduction
Soil water deficit is one of the most important factors
limiting crop productivity worldwide, causing several
effects on plant metabolic processes, including water
relations, nutrient uptake and metabolism, and photosyn-
thesis (Shinozaki et al. 2003; Xiong et al. 2012). Under
water deficit, plants close stomata to reduce transpiration
and, due to stomatal closure, CO2 diffusion to the
Electronic supplementary material The online version of thisarticle (doi:10.1007/s10265-014-0635-1) contains supplementarymaterial, which is available to authorized users.
I. Sperdouli � M. Moustakas (&)
Department of Botany, School of Biology, Aristotle University
of Thessaloniki, University Campus, 54124 Thessaloniki, Greece
e-mail: [email protected]
123
J Plant Res
DOI 10.1007/s10265-014-0635-1

chloroplasts decreases significantly, which down-regulates
carbon assimilation. As a result, the consumption of
reduction equivalents (NADPH) decreases, causing an
increase in NADPH/NADP? ratio, generating an oversup-
ply of reduction equivalents (Selmar and Kleinwachter
2013). Over reduction of components within the photo-
synthetic electron transport chain in the chloroplasts cannot
be balanced by availability of the electron acceptor
NADP? and electrons are captured by O2 instead of
NADP?, resulting in the generation of reactive oxygen
species (ROS) (Vankova et al. 2012).
Plants can acclimate to water deficit under some cir-
cumstances by accumulating osmolytes and proteins that
act as osmoprotectants, antioxidants, and/or stress signals
specifically involved in stress tolerance (Kubis et al. 2014;
Pinheiro et al. 2011; Shinozaki and Yamaguchi-Shinozaki
2007). Compatible solutes can counteract negative dehy-
dration effects on the cell structure and enzyme functioning
and detoxify ROS (Kavi Kishor et al. 1995; Kubis et al.
2014; Moustakas et al. 2011; Nanjo et al. 1999; Yoshiba
et al. 1997).
Positive correlations between the capacities for proline
accumulation and drought or salinity tolerance of different
varieties of the same species have been reported (De Ronde
et al. 2004; Hare and Cress 1997; Hayat et al. 2012; Kubis
et al. 2014; Sperdouli and Moustakas 2012b; Yamada et al.
2005). Proline accumulation in plant tissue is the result of a
decrease in proline degradation, increase in proline biosyn-
thesis, decrease in protein synthesis or proline utilization, or
increased hydrolysis of proteins (Hare et al. 1999). Proline
biosynthesis in the chloroplasts under water deficit consumes
NADPH and decreases the NADPH/NADP? ratio, thus
reducing ROS production (Vankova et al. 2012). Under
water deficit conditions, metabolic pathways that decrease
the NADPH/NADP? ratio resulting to reduced ROS pro-
duction are thus stimulated (De Ronde et al. 2004; Hare and
Cress 1997; Selmar and Kleinwachter 2013; Vankova et al.
2012). Proline signaling has been found to stimulate a sol-
uble sugar signaling pathway (Rasheed et al. 2011; Sperdouli
and Moustakas 2012b) and vice versa (Hu et al. 2012). Apart
from proline protection from ROS, sugar protection against
oxidative stress seems to be partly due to activation of spe-
cific ROS scavenging systems, with consequent reduction of
oxidative damage (Pinheiro and Chaves 2011).
Chlorophyll fluorescence measurements have become
widely used to study the functioning of the photosynthetic
apparatus and are a powerful tool to study the plant’s
response to water deficit (Massacci et al. 2008; Sperdouli
and Moustakas 2012a, b; Woo et al. 2008).
Previous works have shown that MiWD affected pho-
tosystem II (PSII) functioning, while under MoWD con-
ditions, photosynthetic acclimation was observed,
suggesting that PSII activity does not decrease in a
drought-dependent way (Sperdouli and Moustakas 2012a,
b). However, compared to the increasing knowledge about
the enhanced accumulation of certain metabolites, under
water deficit, revealed by studies using Arabidopsis, little
attention has been paid to the differential behavior of
young and mature leaves to water deficit. Young leaves of
A. thaliana were found to acclimate better to water deficit
by dissipating the excess excitation energy (Sperdouli and
Moustakas 2012c, 2014), and this seems to be an important
mechanism in order to avoid possible photodamage to PSII
under water deficit (Jung 2004; Sperdouli and Moustakas
2014). Thus, in the present study, we evaluated whether
differences in the tolerance of A. thaliana young and
mature leaves, to MiWD and MoWD, correlate with dif-
ferential metabolite accumulation of proline and sugars. In
summary, the overall hypothesis was that young leaves of
A. thaliana would show higher photosynthetic acclimation
to water deficit than mature leaves and thus would accu-
mulate more proline and sugars.
Materials and methods
Plant material, growth conditions, and water deficit
treatment
Arabidopsis thaliana ecotype Columbia (Col-0) plants were
grown in a growth chamber (EF7, Conviron, MN, Canada)
with controlled environmental conditions under a long day
photoperiod 14 h/10 h, with 40 ± 5 %/60 ± 5 % humidity,
temperature 22 ± 1 �C/19 ± 1 �C and light intensity of
120 ± 20 lmol photons m-2 s-1. Water deficit was
imposed on 4 week-old Arabidopsis plants (nine to ten
leaves per plant) by withholding water for a period up to
10 days. All plants, i.e. well watered plants (control plants)
and plants under water deficit, were sampled at the same day
for analysis. Two categories of water deficit, besides well
watered, were characterized: MiWD and MoWD (Sperdouli
and Moustakas 2012b). Watering of MoWD plants was
stopped 10 days before sampling, of MiWD plants 6 days
before sampling while control plants were watered 3 h
before sampling. The two developmental leaf stages that
were examined were fully developed mature leaves and
developing young leaves. As young leaves were assigned
those in the center of the leaf rosette with 1.5–2 cm length,
while the average length of mature leaves in the rosette was
4.1 ± 0.5 cm (Sperdouli and Moustakas 2014).
Soil and leaf water status
Soil volumetric water content (SWC) in m3 m-3 was
measured with a 5TE (Decagon Devices, Pullman, WA,
USA) soil moisture sensor, coupled to a ProCheck
J Plant Res
123

(Decagon Devices, Pullman, WA, USA) read out device.
Plant water status was determined by measuring leaf water
content (LWC) by the electronic moisture balance (MOC-
120H, Shimadzu, Tokyo, Japan) as previously described by
Sperdouli and Moustakas (2012a), using the formula:
(FW - DW)/DW 9 100 % where FW refers to fresh
weight and DW refers to dry weight.
Lipid peroxidation measurements
The level of lipid peroxidation in A. thaliana mature leaves
and developing young leaves from each treatment/control was
measured as malondialdehyde (MDA) content determined by
reaction with 2-thiobarbituric acid reactive substances as
described by Moustakas et al. (2011), according to Heath and
Packer (1968). Tissue was homogenized in 0.3 % TBA in
10 % trichloracetic acid at 4 �C and centrifuged for 10 min at
10,000g. The concentration of MDA was calculated from the
difference of the absorbance at 532 and 600 nm spectropho-
tometrically (PharmaSpec UV-1700; Shimadzu, Tokyo,
Japan), using the extinction coefficient of 155 mmol-1 cm-1,
by the formula, MDA = A532 - A600/155 9 40, and
expressed as nmol (MDA) g dry weight (DW) -1.
Proline and sugar determination
A. thaliana young and mature leaves from each treatment/
control, were cut into small pieces, weighed, placed sepa-
rately in glass vials containing 10 ml of 80 % (v/v) ethanol,
and heated at 60 �C for 30 min. The extract was then fil-
tered and diluted with 80 % (v/v) ethanol up to 20 ml. The
concentration of free proline was determined in this extract
as described by Moustakas et al. (2011), following the
acid-ninhydrin reagent method (Bates et al. 1973). Proline
concentration was determined using calibration curve
with standard dilutions of L-proline and the formula:
proline = [(A518 ? 0.0013)/(7.2665) 9 (15)] g-1 sample-1,
and expressed as lmol g DW-1.
Ethanol extracts from each treatment/control of A. tha-
liana young and mature leaves, as used for proline assay,
were diluted with 80 % (v/v) ethanol for the assay of
sugars, as described previously by Moustakas et al. (2011).
Briefly, the diluted extracts were added, drop-by-drop, in
2 ml anthrone reagent test tubes in an ice bath and left to
mix the content. Fully mixed samples were incubated in a
water bath at 90 �C for 15 min and cooled, and absorbance
was read at 625 nm using a PharmaSpec UV-1700 spec-
trophotometer (Shimadzu, Tokyo, Japan). Soluble sugars
were determined using calibration curve with standard
dilutions of sucrose and the formula: sugars =
[(A625 ? 0.0099)/(7.8109) 9 (15 9 3)] g-1 sample-1, and
expressed as lmol g DW-1.
Chlorophyll fluorescence measurements
Chlorophyll fluorescence was measured at room tempera-
ture in dark-adapted (20 min) A. thaliana young and
mature leaves using an imaging-PAM fluorometer (Walz,
Effeltrich, Germany), as described by Sperdouli and
Moustakas (2012b). Five areas of interest were selected,
one in the center of the leaf, two in the outer zone of the
front and two in the outer zone of the back of the leaf. First
Fo (minimum chlorophyll a fluorescence in the dark) and
Fm (maximum chlorophyll a fluorescence in the dark)
values were measured with dark-adapted samples, from
which the potential (maximum) quantum yield derived (Fv/
Fm). Fm was obtained with a saturating pulse of white light
(2,400 lmol photons m-2 s-1, 800 ms). This was fol-
lowed by exposure at 130 lmol photons m-2 s-1 with
repetitive measurements of F0o (minimum chlorophyll a
fluorescence in the light) and F0
m (maximum chlorophyll
a fluorescence in the light) every 20 s, from which auto-
matically values of other chlorophyll fluorescence param-
eters were calculated by the Imaging Win software. The
actinic illumination of 130 lmol photons m-2 s-1 was
selected to match that of the growth light of A. thaliana
plants, and low enough to avoid photoinhibition. In the
presence of actinic illumination, the fluorescence yield Fs
(steady state chlorophyll a fluorescence) was measured.
The calculated parameters included the photochemical
quenching, qP, a measure of the fraction of open PSII
reaction centers, representing the redox state of quinone A
(QA), the primary electron acceptor of PSII, and it was
calculated as (F0m–Fs)/(F
0m–F
0o) (Genty et al. 1989). The
NPQ parameter, which was calculated as (Fm–F0m)/F
0m,
estimates the non-photochemical quenching that reflects
heat dissipation of excitation energy in the antenna system
(Bilger and Bjorkman 1990). The relative PSII electron
transport rate (ETR) was calculated as c 9 UPSII (actual
quantum yield of photochemistry) 9 PPFD (photosyn-
thetic photon flux density); for the coefficient c, the value
of 0.42 was assumed [considering equal distribution of
photons between PSII and PSI (0.5) multiplied by the
homogeneous absorption factor of 0.84, that contains
assumptions on leaf absorption of PPFD] (Schreiber et al.
1994).
Statistical analysis
Each treatment/control was analyzed with six replicates. A
standard error (SE) was calculated and data were expressed
in mean ± SE of six replicates. Chlorophyll fluorescence
measurements represent averaged values (n = 6) from two
independent experiments with three leaf samples (each
with five areas of interest) from three different plants, per
J Plant Res
123
treatment per experiment. One-way ANOVA (StatView
computer package) was carried out and means were sepa-
rated using Duncan’s least significant ranges (LSR) at
95 %. For the analysis of the interaction of leaf age
(mature, young) and water deficit treatment (Control,
MiWD, MoWD) a two-way analysis of variance using the
StatView-ANOVA computer package (Abacus Concepts,
Inc Berkley, CA, USA) was performed. A linear regression
analysis was also performed (Sperdouli and Moustakas
2012b, c).
Results
Soil and leaf water status
In the two categories of water deficit, besides well watered,
that were examined water deficit was induced gradually by
withholding water. In MiWD, SWC was 66–68 % of
control (well watered), while in MoWD 50–52 % (Fig. S1).
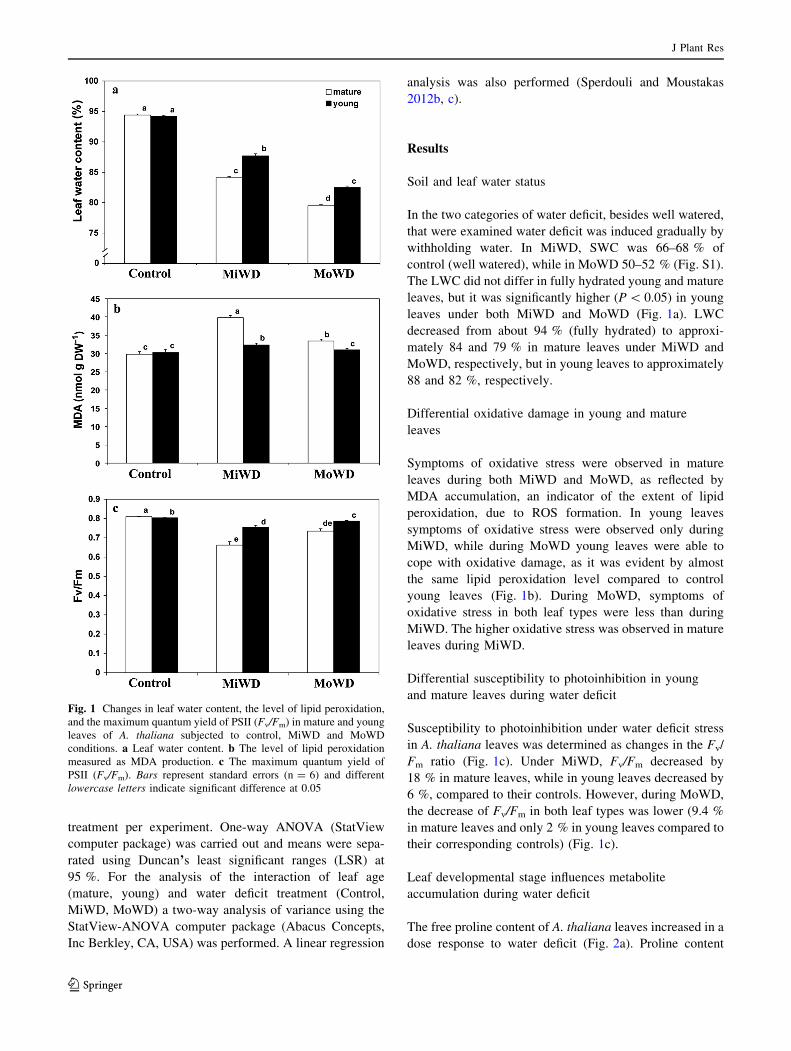
The LWC did not differ in fully hydrated young and mature
leaves, but it was significantly higher (P \ 0.05) in young
leaves under both MiWD and MoWD (Fig. 1a). LWC
decreased from about 94 % (fully hydrated) to approxi-
mately 84 and 79 % in mature leaves under MiWD and
MoWD, respectively, but in young leaves to approximately
88 and 82 %, respectively.
Differential oxidative damage in young and mature
leaves
Symptoms of oxidative stress were observed in mature
leaves during both MiWD and MoWD, as reflected by
MDA accumulation, an indicator of the extent of lipid
peroxidation, due to ROS formation. In young leaves
symptoms of oxidative stress were observed only during
MiWD, while during MoWD young leaves were able to
cope with oxidative damage, as it was evident by almost
the same lipid peroxidation level compared to control
young leaves (Fig. 1b). During MoWD, symptoms of
oxidative stress in both leaf types were less than during
MiWD. The higher oxidative stress was observed in mature
leaves during MiWD.
Differential susceptibility to photoinhibition in young
and mature leaves during water deficit
Susceptibility to photoinhibition under water deficit stress
in A. thaliana leaves was determined as changes in the Fv/
Fm ratio (Fig. 1c). Under MiWD, Fv/Fm decreased by
18 % in mature leaves, while in young leaves decreased by
6 %, compared to their controls. However, during MoWD,
the decrease of Fv/Fm in both leaf types was lower (9.4 %
in mature leaves and only 2 % in young leaves compared to
their corresponding controls) (Fig. 1c).
Leaf developmental stage influences metabolite
accumulation during water deficit
The free proline content of A. thaliana leaves increased in a
dose response to water deficit (Fig. 2a). Proline content
Fig. 1 Changes in leaf water content, the level of lipid peroxidation,
and the maximum quantum yield of PSII (Fv/Fm) in mature and young
leaves of A. thaliana subjected to control, MiWD and MoWD
conditions. a Leaf water content. b The level of lipid peroxidation
measured as MDA production. c The maximum quantum yield of
PSII (Fv/Fm). Bars represent standard errors (n = 6) and different
lowercase letters indicate significant difference at 0.05
J Plant Res
123
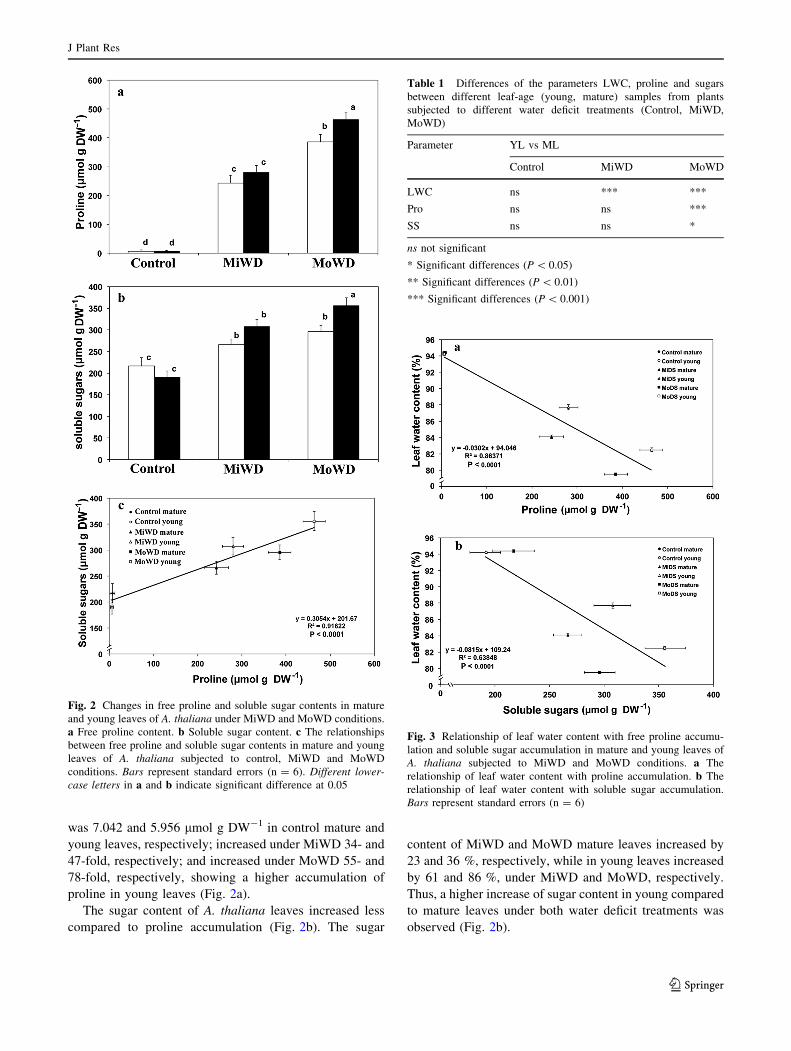
was 7.042 and 5.956 lmol g DW-1 in control mature and
young leaves, respectively; increased under MiWD 34- and
47-fold, respectively; and increased under MoWD 55- and
78-fold, respectively, showing a higher accumulation of
proline in young leaves (Fig. 2a).
The sugar content of A. thaliana leaves increased less
compared to proline accumulation (Fig. 2b). The sugar
content of MiWD and MoWD mature leaves increased by
23 and 36 %, respectively, while in young leaves increased
by 61 and 86 %, under MiWD and MoWD, respectively.
Thus, a higher increase of sugar content in young compared
to mature leaves under both water deficit treatments was
observed (Fig. 2b).
Fig. 2 Changes in free proline and soluble sugar contents in mature
and young leaves of A. thaliana under MiWD and MoWD conditions.
a Free proline content. b Soluble sugar content. c The relationships
between free proline and soluble sugar contents in mature and young
leaves of A. thaliana subjected to control, MiWD and MoWD
conditions. Bars represent standard errors (n = 6). Different lower-
case letters in a and b indicate significant difference at 0.05
Table 1 Differences of the parameters LWC, proline and sugars
between different leaf-age (young, mature) samples from plants
subjected to different water deficit treatments (Control, MiWD,
MoWD)
Parameter YL vs ML
Control MiWD MoWD
LWC ns *** ***
Pro ns ns ***
SS ns ns *
ns not significant
* Significant differences (P \ 0.05)
** Significant differences (P \ 0.01)
*** Significant differences (P \ 0.001)
Fig. 3 Relationship of leaf water content with free proline accumu-
lation and soluble sugar accumulation in mature and young leaves of
A. thaliana subjected to MiWD and MoWD conditions. a The
relationship of leaf water content with proline accumulation. b The
relationship of leaf water content with soluble sugar accumulation.
Bars represent standard errors (n = 6)
J Plant Res
123
Relationship between proline and sugar accumulation
Sugar accumulation in young and mature leaves under both
water deficit treatments was tightly correlated
(R2 = 0.91622, P \ 0.0001) with the magnitude of proline
accumulation (Fig. 2c).
Relationship between leaf’s water content
and metabolite accumulation
While there was a significant difference in LWC between
young and mature leaves during MiWD, this was not
associated with any significant difference between young
and mature leaves in either proline or sugar accumulation
(Table 1). However, under MoWD the significant higher
LWC in young leaves was associated with a significant
difference between leaf age in both proline and sugar
accumulation (Table 1). LWC exhibited significantly neg-
ative linear correlation with both proline (Fig. 3a) and
sugar (Fig. 3b) content. Proline accumulation under MiWD
and MoWD was negatively correlated (R2 = 0.86371,
P \ 0.0001) with LWC in both young and mature leaves
(Fig. 3a). Sugar accumulation was also negatively corre-
lated (R2 = 0.63848, P \ 0.0001) with the magnitude of
LWC in both leaf types (Fig. 3b).
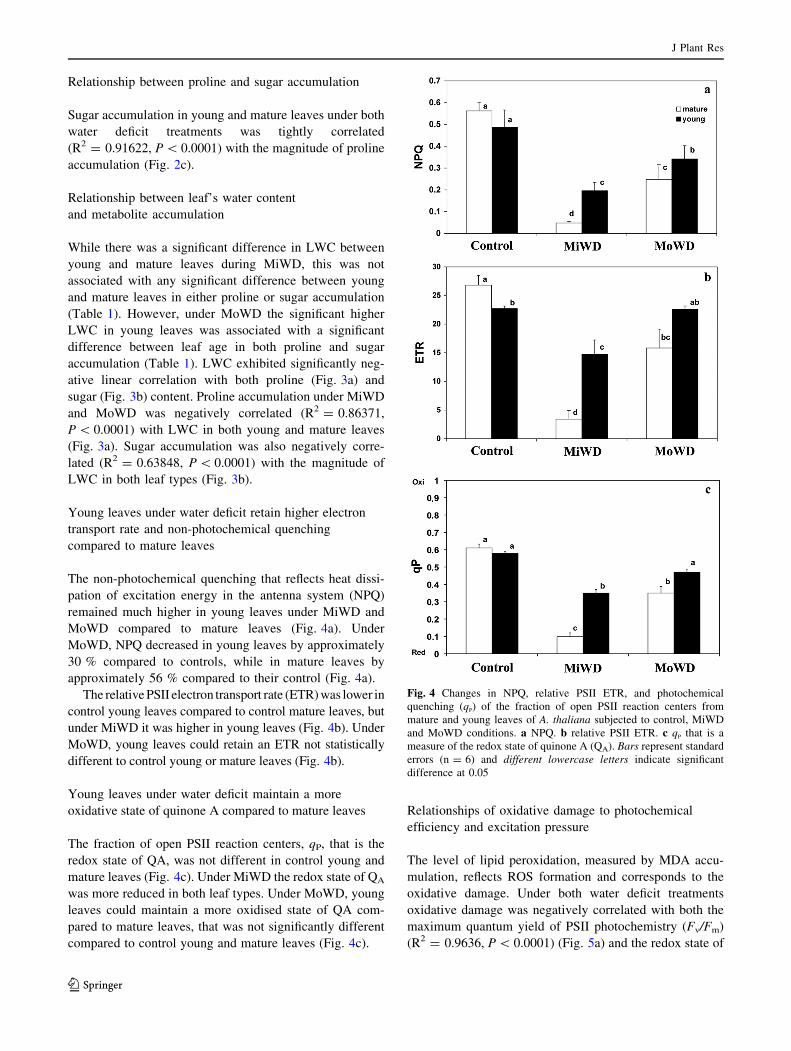
Young leaves under water deficit retain higher electron
transport rate and non-photochemical quenching
compared to mature leaves
The non-photochemical quenching that reflects heat dissi-
pation of excitation energy in the antenna system (NPQ)
remained much higher in young leaves under MiWD and
MoWD compared to mature leaves (Fig. 4a). Under
MoWD, NPQ decreased in young leaves by approximately
30 % compared to controls, while in mature leaves by
approximately 56 % compared to their control (Fig. 4a).
The relative PSII electron transport rate (ETR) was lower in
control young leaves compared to control mature leaves, but
under MiWD it was higher in young leaves (Fig. 4b). Under
MoWD, young leaves could retain an ETR not statistically
different to control young or mature leaves (Fig. 4b).
Young leaves under water deficit maintain a more
oxidative state of quinone A compared to mature leaves
The fraction of open PSII reaction centers, qP, that is the
redox state of QA, was not different in control young and
mature leaves (Fig. 4c). Under MiWD the redox state of QA
was more reduced in both leaf types. Under MoWD, young
leaves could maintain a more oxidised state of QA com-
pared to mature leaves, that was not significantly different
compared to control young and mature leaves (Fig. 4c).
Relationships of oxidative damage to photochemical
efficiency and excitation pressure
The level of lipid peroxidation, measured by MDA accu-
mulation, reflects ROS formation and corresponds to the
oxidative damage. Under both water deficit treatments
oxidative damage was negatively correlated with both the
maximum quantum yield of PSII photochemistry (Fv/Fm)
(R2 = 0.9636, P \ 0.0001) (Fig. 5a) and the redox state of
Fig. 4 Changes in NPQ, relative PSII ETR, and photochemical
quenching (qP) of the fraction of open PSII reaction centers from
mature and young leaves of A. thaliana subjected to control, MiWD
and MoWD conditions. a NPQ. b relative PSII ETR. c qP that is a
measure of the redox state of quinone A (QA). Bars represent standard
errors (n = 6) and different lowercase letters indicate significant
difference at 0.05
J Plant Res
123
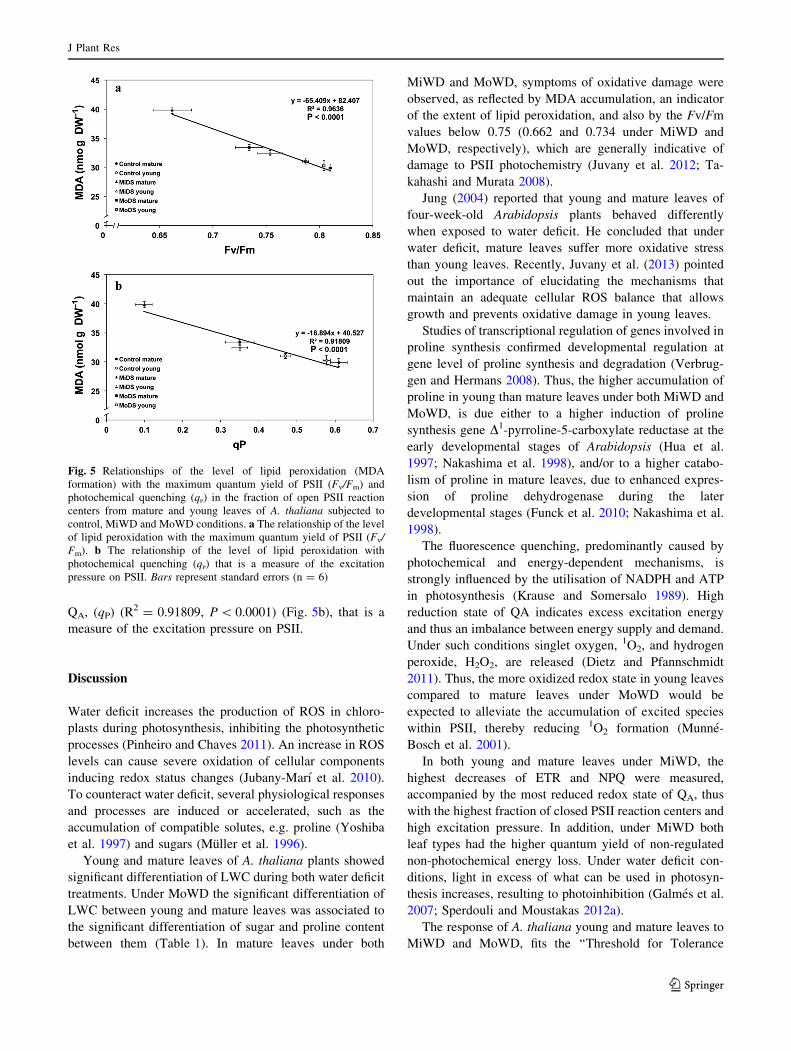
QA, (qP) (R2 = 0.91809, P \ 0.0001) (Fig. 5b), that is a
measure of the excitation pressure on PSII.
Discussion
Water deficit increases the production of ROS in chloro-
plasts during photosynthesis, inhibiting the photosynthetic
processes (Pinheiro and Chaves 2011). An increase in ROS
levels can cause severe oxidation of cellular components
inducing redox status changes (Jubany-Marı et al. 2010).
To counteract water deficit, several physiological responses
and processes are induced or accelerated, such as the
accumulation of compatible solutes, e.g. proline (Yoshiba
et al. 1997) and sugars (Muller et al. 1996).
Young and mature leaves of A. thaliana plants showed
significant differentiation of LWC during both water deficit
treatments. Under MoWD the significant differentiation of
LWC between young and mature leaves was associated to
the significant differentiation of sugar and proline content
between them (Table 1). In mature leaves under both
MiWD and MoWD, symptoms of oxidative damage were
observed, as reflected by MDA accumulation, an indicator
of the extent of lipid peroxidation, and also by the Fv/Fm
values below 0.75 (0.662 and 0.734 under MiWD and
MoWD, respectively), which are generally indicative of
damage to PSII photochemistry (Juvany et al. 2012; Ta-
kahashi and Murata 2008).
Jung (2004) reported that young and mature leaves of
four-week-old Arabidopsis plants behaved differently
when exposed to water deficit. He concluded that under
water deficit, mature leaves suffer more oxidative stress
than young leaves. Recently, Juvany et al. (2013) pointed
out the importance of elucidating the mechanisms that
maintain an adequate cellular ROS balance that allows
growth and prevents oxidative damage in young leaves.
Studies of transcriptional regulation of genes involved in
proline synthesis confirmed developmental regulation at
gene level of proline synthesis and degradation (Verbrug-
gen and Hermans 2008). Thus, the higher accumulation of
proline in young than mature leaves under both MiWD and
MoWD, is due either to a higher induction of proline
synthesis gene D1-pyrroline-5-carboxylate reductase at the
early developmental stages of Arabidopsis (Hua et al.
1997; Nakashima et al. 1998), and/or to a higher catabo-
lism of proline in mature leaves, due to enhanced expres-
sion of proline dehydrogenase during the later
developmental stages (Funck et al. 2010; Nakashima et al.
1998).
The fluorescence quenching, predominantly caused by
photochemical and energy-dependent mechanisms, is
strongly influenced by the utilisation of NADPH and ATP
in photosynthesis (Krause and Somersalo 1989). High
reduction state of QA indicates excess excitation energy
and thus an imbalance between energy supply and demand.
Under such conditions singlet oxygen, 1O2, and hydrogen
peroxide, H2O2, are released (Dietz and Pfannschmidt
2011). Thus, the more oxidized redox state in young leaves
compared to mature leaves under MoWD would be
expected to alleviate the accumulation of excited species
within PSII, thereby reducing 1O2 formation (Munne-
Bosch et al. 2001).
In both young and mature leaves under MiWD, the
highest decreases of ETR and NPQ were measured,
accompanied by the most reduced redox state of QA, thus
with the highest fraction of closed PSII reaction centers and
high excitation pressure. In addition, under MiWD both
leaf types had the higher quantum yield of non-regulated
non-photochemical energy loss. Under water deficit con-
ditions, light in excess of what can be used in photosyn-
thesis increases, resulting to photoinhibition (Galmes et al.
2007; Sperdouli and Moustakas 2012a).
The response of A. thaliana young and mature leaves to
MiWD and MoWD, fits the ‘‘Threshold for Tolerance
Fig. 5 Relationships of the level of lipid peroxidation (MDA
formation) with the maximum quantum yield of PSII (Fv/Fm) and
photochemical quenching (qP) in the fraction of open PSII reaction
centers from mature and young leaves of A. thaliana subjected to
control, MiWD and MoWD conditions. a The relationship of the level
of lipid peroxidation with the maximum quantum yield of PSII (Fv/
Fm). b The relationship of the level of lipid peroxidation with
photochemical quenching (qP) that is a measure of the excitation
pressure on PSII. Bars represent standard errors (n = 6)
J Plant Res
123

Model’’, with a lag time required (during MiWD) for the
induction of a tolerance mechanism (Barcelo and Pos-
chenrieder 2002; Sperdouli and Moustakas 2012a). This
lag-time was required for a substantial increase of proline
and sugar accumulation that was achieved only during
MoWD in young leaves.
Acclimation of young leaves to MoWD involves ROS
homeostasis that results to a higher level of oxidized state
of QA, which alleviates the accumulation of excited spe-
cies within PSII, thereby reducing ROS formation (Munne-
Bosch et al. 2001; Vankova et al. 2012). Increased sugar
accumulation in young leaves under MoWD has contrib-
uted to activation of ROS scavenging systems, with con-
sequent reduction of oxidative damage (Pinheiro and
Chaves 2011; Smeekens et al. 2010). Transgenic Arabi-
dopsis plants overexpressing LcMYB1 gene, show
enhanced expression levels of D1-pyrroline-5-carboxylate
synthase and accumulated greater amounts of proline and
sugars and less MDA under salt stress (Cheng et al. 2013).
Acclimation of young leaves to MoWD is also achieved by
stimulation of photoprotective mechanisms such as NPQ
(Havaux et al. 2000; Neubauer and Yamamoto 1992). NPQ
reflects the dissipation of excess excitation energy in the
form of harmless heat, thus protecting the plant from the
damaging effects of ROS (Hideg et al. 2008). It seems that
photoprotection was achieved in young leaves under
MoWD, since NPQ was regulated in such a way that the
remained open fraction of PSII reaction centers was non
significantly different to controls (Lambrev et al. 2012),
thus young leaves under MoWD had the same excitation
pressure to controls. It is concluded that under MoWD, a
significant higher proline and sugar accumulation in young
leaves, contributed in maintaining their base levels of lipid
peroxidation. ROS homeostasis of young leaves during
MoWD resulted to acclimation, that was achieved among
others by (1) increased sugar accumulation and (2) either
increased proline synthesis that converts excess NADPH
into NADP? and/or decreased proline catabolism, resulting
to a higher level of oxidized state of QA, and by (3)
stimulation of the photoprotective NPQ mechanism that
protects the plant from the damaging effects of ROS.
Young leaves of MoWD A. thaliana plants possessed not
only an ETR almost the same with controls, but also the
same excitation pressure to controls. Thus, we can con-
clude from our results that photosynthetic acclimation of
young leaves to water deficit is correlated to increased
proline and sugar accumulation.
Acknowledgments We are grateful to Prof. Dr. Stefanos Sgardelis
(Department of Ecology, Aristotle University of Thessaloniki) for the
help in statistical analysis. This work was supported by the European
Fund of Regional Growth and the Hellenic General Secretariat for
Research and Technology under the project No. 09FR47 to Michael
Moustakas.
References
Barcelo J, Poschenrieder C (2002) Fast root growth responses, root
exudates, and internal detoxification as clues to the mechanisms
of aluminium toxicity and resistance: a review. Environ Exp Bot
48:75–92
Bates IS, Waldren RP, Teare ID (1973) Rapid determination of free
proline for water stress studies. Plant Soil 39:205–207
Bilger W, Bjorkman O (1990) Role of the xanthophyll cycle in
photoprotection elucidated by measurements of light-induced
absorbance changes, fluorescence and photosynthesis in leaves
of Hedera canariensis. Photosynth Res 25:173–186
Cheng L, Li X, Huang X, Ma T, Liang Y, Ma X, Peng X, Jia J, Chen
S, Chen Y, Deng B, Liu G (2013) Overexpression of sheepgrass
R1-MYB transcription factor LcMYB1 confers salt tolerance in
transgenic Arabidopsis. Plant Physiol Biochem 70:252–260
De Ronde JA, Cress WA, Kruger GHJ, Strasser RJ, Van Staden J
(2004) Photosynthetic response of transgenic soybean plants,
containing an Arabidopsis P5CR gene, during heat and drought
stress. J Plant Physiol 161:1211–1224
Dietz KJ, Pfannschmidt T (2011) Novel regulators in photosynthetic
redox control of plant metabolism and gene expression. Plant
Physiol 155:1477–1485
Funck D, Eckard S, Muller G (2010) Non-redundant functions of two
proline dehydrogenase isoforms in Arabidopsis. BMC Plant Biol
10:70
Galmes J, Abadia A, Medrano H, Flexas J (2007) Photosynthesis and
photoprotection responses to water stress in the wild-extinct
plant Lysimachia minoricensis. Environ Exp Bot 60:308–317
Genty B, Briantais JM, Baker NR (1989) The relationship between
the quantum yield of photosynthetic electron transport and
quenching of chlorophyll fluorescence. Biochim Biophys Acta
990:87–92
Hare PD, Cress WA (1997) Metabolic implications of stress-induced
proline accumulation in plants. Plant Growth Regul 21:79–102
Hare PD, Cress WA, Van Staden J (1999) Proline synthesis and
degradation: a model system for elucidating stress-related signal
transduction. J Exp Bot 50:413–434
Havaux M, Bonfils JP, Lutz C, Niyogi KK (2000) Photodamage of the
photosynthetic apparatus and its dependence on the leaf devel-
opmental stage in the npq1 Arabidopsis mutant deficient in the
xanthophyll cycle enzyme violaxanthin de-epoxidase. Plant
Physiol 124:273–284
Hayat S, Hayat Q, Alyemeni MN, Wani AS, Pichtel J, Ahmad A
(2012) Role of proline under changing environments. A review.
Plant Signal Behav 7:1456–1466
Heath RL, Packer L (1968) Photoperoxidation in isolated chloroplasts
I. Kinetics and stoichiometry of fatty acid peroxidation. Arch
Biochem Biophys 125:189–198
Hideg E, Kos P, Schreiber U (2008) Imaging of NPQ and ROS
formation in tobacco leaves: heat inactivation of the water–water
cycle prevents down-regulation of PSII. Plant Cell Physiol
49:1879–1886
Hu M, Shi Z, Zhang Z, Zhang Y, Li H (2012) Effects of exogenous
glucose on seed germination and antioxidant capacity in wheat
seedlings under salt stress. Plant Growth Regul 68:177–188
Hua XJ, Van de Cotte B, Van Montagu M, Verbruggen N (1997)
Developmental regulation of pyrroline-5-carboxylate reductase
gene expression in Arabidopsis. Plant Physiol 114:1215–1224
Jubany-Marı T, Munne-Bosch S, Alegre L (2010) Redox regulation of
water stress responses in field-grown plants. Role of hydrogen
peroxide and ascorbate. Plant Physiol Biochem 48:351–358
Jung S (2004) Variation in antioxidant metabolism of young and
mature leaves of Arabidopsis thaliana subjected to drought.
Plant Sci 166:459–466
J Plant Res
123

Juvany M, Muller M, Munne-Bosch S (2012) Leaves of field-grown
mastic trees suffer oxidative stress at the two extremes of their
lifespan. J Integr Plant Biol 54:584–594
Juvany M, Muller M, Munne-Bosch S (2013) Photo-oxidative stress
in emerging and senescing leaves: a mirror image. J Exp Bot
64:3087–3098
Kavi Kishor PB, Hong Z, Miao GH, Hu CAA, Verma DPS (1995)
Overexpression of D1-pyrroline-5-carboxylate synthetase
increases proline production and confers osmotolerance in
transgenic plants. Plant Physiol 108:1387–1394
Krause GH, Somersalo S (1989) Fluorescence as a tool in photosyn-
thesis research: application in studies of photoinhibition, cold
acclimation and freezing stress. Phil Trans R Soc Lond
323:281–293
Kubis J, Floryszak-Wieczorek J, Arasimowicz-Jelonek M (2014)
Polyamines induce adaptive responses in water deficit stressed
cucumber roots. J Plant Res 127:151–158
Lambrev PH, Miloslavina Y, Jahns P, Holzwarth AR (2012) On the
relationship between non-photochemical quenching and photo-
protection of photosystem II. Biochim Biophys Acta
1817:760–769
Massacci A, Nabiev SM, Pietrosanti L, Nematov SK, Chernikova TN,
Thor K, Leipner J (2008) Response of the photosynthetic
apparatus of cotton (Gossypium hirsutum) to the onset of drought
stress under field conditions studied by gas-exchange analysis
and chlorophyll fluorescence imaging. Plant Physiol Biochem
46:189–195
Moustakas M, Sperdouli I, Kouna T, Antonopoulou CI, Therios I
(2011) Exogenous proline induces soluble sugar accumulation
and alleviates drought stress effects on photosystem II function-
ing of Arabidopsis thaliana leaves. Plant Growth Regul
65:315–325
Muller J, Boller T, Wiemken A (1996) Pools of non-structural
carbohydrates in soybean root nodules during water stress.
Physiol Plantarum 98:723–730
Munne-Bosch S, Jubany-Marı T, Alegre L (2001) Drought-induced
senescence is characterized by a loss of antioxidant defences in
chloroplasts. Plant Cell Environ 24:1319–1327
Nakashima K, Satoh R, Kiyosue T, Yamaguchi-Shinozaki K,
Shinozaki K (1998) A gene encoding proline dehydrogenase is
not only induced by proline and hypoosmolarity, but is also
developmentally regulated in the reproductive organs of Arabi-
dopsis. Plant Physiol 118:1233–1241
Nanjo T, Kobayashi M, Yoshiba Y, Sanada Y, Wada K, Tsukaya H,
Kakubari Y, Yamaguchi-Shinozaki K, Shinozaki K (1999)
Biological functions of proline in morphogenesis and osmotol-
erance revealed in antisense transgenic Arabidopsis thaliana.
Plant J 18:185–193
Neubauer C, Yamamoto H (1992) Mehler-peroxidase reaction
mediates zeaxanthin formation and zeaxanthin-related fluores-
cence quenching in intact chloroplasts. Plant Physiol
99:1354–1361
Pinheiro C, Chaves MM (2011) Photosynthesis and drought: can we
make metabolic connections from available data? J Exp Bot
62:869–882
Pinheiro C, Antonio C, Ortuno MF, Dobrev PI, Hartung W, Thomas-
Oates J, Ricardo CP, Vankova R, Chaves MM, Wilson JC (2011)
Initial water deficit effects on Lupinus albus photosynthetic
performance, carbon metabolism, and hormonal balance: meta-
bolic reorganization prior to early stress responses. J Exp Bot
62:4965–4974
Rasheed R, Wahid A, Farooq M, Hussain I, Basra SMA (2011) Role
of proline and glycinebetaine pretreatments in improving heat
tolerance of sprouting sugarcane (Saccharum sp.) buds. Plant
Growth Regul 65:35–45
Schreiber U, Bilger W, Neubauer C (1994) Chlorophyll fluorescence
as a non-intrusive indicator for rapid assessment of in vivo
photosynthesis. In: Schulze ED, Caldwell MM (eds) Ecophys-
iology of photosynthesis. Series ecological studies, vol 100.
Springer, Berlin, pp 49–70
Selmar D, Kleinwachter M (2013) Stress enhances the synthesis of
secondary plant products: the impact of stress-related over-
reduction on the accumulation of natural products. Plant Cell
Physiol 54:817–826
Shinozaki K, Yamaguchi-Shinozaki K (2007) Gene networks
involved in drought stress response and tolerance. J Exp Bot
58:221–227
Shinozaki K, Yamaguchi-Shinozaki K, Seki M (2003) Regulatory
network of gene expression in the drought and cold stress
responses. Curr Opin Plant Biol 6:410–417
Smeekens S, Ma J, Hanson J, Rolland F (2010) Sugar signals and
molecular networks controlling plant growth. Curr Opin Plant
Biol 13:274–279
Sperdouli I, Moustakas M (2012a) Spatio-temporal heterogeneity in
Arabidopsis thaliana leaves under drought stress. Plant Biol
14:118–128
Sperdouli I, Moustakas M (2012b) Interaction of proline, sugars, and
anthocyanins during photosynthetic acclimation of Arabidopsis
thaliana to drought stress. J Plant Physiol 169:577–585
Sperdouli I, Moustakas M (2012c) Differential response of photo-
system II photochemistry in young and mature leaves ofArabidopsis thaliana to the onset of drought stress. Acta Physiol
Plant 34:1267–1276
Sperdouli I, Moustakas M (2014) A better energy allocation of
absorbed light in photosystem II and less photooxidative damage
contribute to acclimation of Arabidopsis thaliana young leaves
to water deficit. J Plant Physiol 171:587–593
Takahashi S, Murata N (2008) How do environmental stresses
accelerate photoinhibition? Trends Plant Sci 13:178–182
Vankova R, Dobra J, Storchova H (2012) Recovery from drought
stress in tobacco. An active process associated with the reversal
of senescence in some plant parts and the sacrifice of others.
Plant Signal Behav 7:19–21
Verbruggen N, Hermans C (2008) Proline accumulation in plants: a
review. Amino Acids 35:753–759
Woo NS, Badger MR, Pogson BJ (2008) A rapid, non-invasive
procedure for quantitative assessment of drought survival using
chlorophyll fluorescence. Plant Methods 4:27
Xiong J, Zhang L, Fu G, Yang Y, Zhu C, Tao L (2012) Drought-
induced proline accumulation is uninvolved with increased nitric
oxide, which alleviates drought stress by decreasing transpiration
in rice. J Plant Res 125:155–164
Yamada M, Morishita H, Urano K, Shiozaki N, Yamaguhi-Shinozaki
K, Shinozaki K, Yoshiba Y (2005) Effects of free proline
accumulation in petunias under drought stress. J Exp Bot
56:1975–1981
Yoshiba Y, Kiyosue T, Nakashima K, Yamaguchi-Shinozaki K,
Shinozaki K (1997) Regulation of levels of proline as an
osmolytes in plants under water stress. Plant Cell Physiol
18:1095–1102
J Plant Res
123





























