Embed Size (px)
Citation preview

http://hol.sagepub.com/The Holocene
http://hol.sagepub.com/content/13/1/97The online version of this article can be found at:
DOI: 10.1191/0959683603hl597rp
2003 13: 97The HoloceneIvanka Stefanova and Brigitta Ammann
Lateglacial and Holocene vegetation belts in the Pirin Mountains (southwestern Bulgaria)
Published by:
http://www.sagepublications.com
can be found at:The HoloceneAdditional services and information for
http://hol.sagepub.com/cgi/alertsEmail Alerts:
http://hol.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
http://hol.sagepub.com/content/13/1/97.refs.htmlCitations:
What is This?
- Jan 1, 2003Version of Record >>
at FORDHAM UNIV LIBRARY on September 6, 2014hol.sagepub.comDownloaded from at FORDHAM UNIV LIBRARY on September 6, 2014hol.sagepub.comDownloaded from

The Holocene 13,1 (2003) pp. 97–107
Lateglacial and Holocene vegetation beltsin the Pirin Mountains (southwesternBulgaria)Ivanka Stefanova1* and Brigitta Ammann2
(1Institute of Botany, Bulgarian Academy of Sciences, 23 "Acad. G. Bonchev St.,BG-1113 So� a, Bulgaria; 2Institute of Plant Sciences, 21 Altenbergrain,CH-3013 Bern, Switzerland)
Received 25 June 2001; revised manuscript accepted 16 February 2002
Abstract: Pollen stratigraphy of a core 270 cm long from Lake Dalgoto at 2310 m in the Northern PirinMountains, southern Bulgaria, was treated by optimal partitioning and compared to a broken-stick model toreveal statistically signi� cant pollen zones. The vegetational reconstructions presented here are based on pollenpercentages and pollen in� ux, on comparisons of modern and fossil pollen spectra, and on macrofossil datesfrom other sites in the mountains. During the Younger Dryas (11000–10200 14C yr BP), an open xerophyticherb vegetation with Artemisia and Chenopodiaceae was widely developed around the lake. Deciduous treesgrowing at lower elevations contributed to the pollen rain deposited at the higher-elevation sampling sites.Speci� cally, from 10200 to 8500 yr BP, Quercus, Ulmus, Tilia and Betula expanded rapidly at low and inter-mediate elevations, and between 8500 and 6500 yr BP they extended to higher elevations close to the upperforest limit, which was formed by Betula pendula at about 1900 m. Coniferous species were limited in theregion at this time. After 6500 yr BP, the expansion of conifers (Pinus peuce, P. sylvestris, P. mugo, Abiesalba) at high elevations forced the deciduous trees downward. Between 6500 and 3000 yr BP, the forest limitat 2200 m was formed by P. peuce, and A. alba had its maximum vertical range up to 1900 m. Later theabundance and vertical range of P. peuce and A. alba were reduced. After 3000 yr BP, Picea expanded.
Key words: Vegetation history, altitudinal vegetation belts, climatic change, Pirin Mountains, Bulgaria, lateQuaternary,Younger Dryas, Holocene.
Introduction
The early plant succession in the Pirin Mountains (SW Bulgaria,highest peak 2914 m) is very attractive for research because theBalkans are postulated as Lateglacial refugia for temperate-treetaxa (Bottema, 1974; Beug, 1975; 1982; Huntley and Birks, 1983;Bennett et al., 1991; Tzedakis, 1993; Willis, 1992; 1994a; 1994b;Lang, 1994). Therefore migration distances were short, so thatfactors controlling vegetational changes, such as climatic changeand soil development, may be assessed. So far only the mid- andlate-Holocene vegetational history of the area is known frompollen analysis (Bozilova, 1975; 1977; Stefanova and Bozilova,1992; 1995; Stefanova and Oeggl, 1993; Panovska et al., 1995;Stefanova, 1997; 1998; 1999), and the Lateglacial and early-Holocene vegetational and environmental history has been in-suf� ciently investigated. Numerous cirques produced by local
*Author for correspondence. Present address: Department of Geology andGeophysics, University of Minnesota, Minneapolis, MN 55455, USA (e-mail:[email protected])
Ó Arnold 2003 10.1191/0959683603hl597rp
glaciers contain lakes with natural archives of vegetational andenvironmental change in the Pirin Mountains.
This paper presents the results of a palynological study of LakeDalgoto. The results are discussed with respect to the regionalvegetational and environmental events during the Lateglacial andearly Holocene. Special attention is paid to the dynamics of thealtitudinal belts of vegetation.
Study area
Lake Dalgoto (41°409 N–23°229 E) is located in the northern partof the Pirin Mountains at an altitude of 2310 m a.s.l. (Figure 1).Northern Pirin is the highest and most monolithic part of themountains. It is built up of Precambrian granitic and metamorphicrocks (including marbles; Boyagiev, 1959). The lake is 450 mlong and 150 m wide and has a maximum depth of 10 m (Ivanovand Sotirov, 1964). The surface of 4.55 ha is about 18 timessmaller than the hydrological catchment, which does not containmarble.
at FORDHAM UNIV LIBRARY on September 6, 2014hol.sagepub.comDownloaded from

98 The Holocene 13 (2003)
Figure 1 (A) Bulgaria and location of the Pirin Mountains. (B) Study area in the Pirin Mountains. (C) Position of Lake Dalgoto (1) and spatial relationshipto the sites studied earlier: (2) Lake Popovo-6 (Stefanova and Bozilova, 1995); (3) Mire Praso (Stefanova and Oeggl, 1993); (4) peat bog Goce Delchev(Stefanova, 1997); (5) peat bog Visokata ela (Stefanova, 1997).
The Pirin Mountains belong to the Continental-Mediterraneanarea of the Subtropical climate zone (Nikolova, 1997). In com-parison with the mountain regions in other climatic areas in Bul-garia, the summer and winter temperatures are higher. The meanannual temperature is 3.7°C at the closest weather station, situatedat 1970 m (hut Vihren), and above 2300 m it is negative. Theannual precipitation is 800–1250 mm, with a maximum inNovember–December (Velev, 1997).
The characteristics of the vegetation in the Pirin Mountains aredetermined by their altitude and by their location close to theMediterranean area. According to Veltshev (1997), seven veg-etation belts are represented. Nomenclature for vascular plants fol-lows Flora of the Republic of Bulgaria (Jordanov et al., 1963–95), as follows.
(1) The lowest belt, up to 500 m a.s.l, is represented only in thesouthern part of the mountain by fragments and consists ofcommunities with such Mediterranean elements as Quercuscoccifera, Juniperus excelsa, J. oxicedrus and Phillyrea lati-folia.
(2) The main communities in the belt of xerothermic oak forest(about 500–700 m a.s.l.) are those of Quercus cerris, Q.pubescens, Q. frainetto and Carpinus orientalis.
(3) Communities of Quercus dalechampii and Carpinus betulusare typical in the belt of the mesophilous and xeromesophil-ous oak-hornbeam belt (up to 800–900 or occasionally 1000m a.s.l.), where communities of Pinus nigra, Ostrya carpini-folia and Corylus avellana are also present.
(4) The belt of beech forest (from 900/1000 to 1500/1600 ma.s.l.), where communities of Fagus sylvatica dominate, isrepresented in the Northern Pirin Mountains by fragments.Communities of Abies alba, Pinus nigra, Picea abies andPinus peuce are sporadic.
(5) In the coniferous belt (between 1500/1600 m and 2000/2200m a.s.l.) communities of Pinus sylvestris and Picea abies aremost abundant. The Balkan endemic Pinus peuce and therelict subendemic Pinus heldreichii are present. Communitiesof Abies alba are less common, but Juniperus sibirica and
Vaccinium myrtillus are more so (Velchev and Vassilev,1987; Velchev and Rusakova, 1990).
(6) The vegetation in the subalpine belt (about 2000–2500 ma.s.l.), where Lake Dalgoto is located, is dominated by com-munities of Pinus mugo, Juniperus sibirica and Vacciniummyrtillus.
(7) In the alpine belt (above 2500 m a.s.l.) communities ofSesleria comosa, Festuca airoides, Agrostis rupestris andCarex curvula occur.
The approximate modern altitudinal ranges of the main tree taxaare indicated on the right of Figure 7. The upper forest limit inthe study area, which today is about 2000–2100 m a.s.l., is formedon siliceous rocks by communities of Picea abies and the Balkanendemic species Pinus peuce, and in calcareous areas by com-munities of the Balkan relict subendemic species Pinus heldre-ichii.
The aquatic vegetation is represented by Ranunculus aquatilis,Subularia aquatica and Sparganium angustifolium. In the hygro-phytic vegetation around the lake Carex nigra, Juncus alpinus,Silene pusilla, Parnassia palustris, Geum coccineum, Plantagogentianoides and Epilobium palustre are present.
Methods
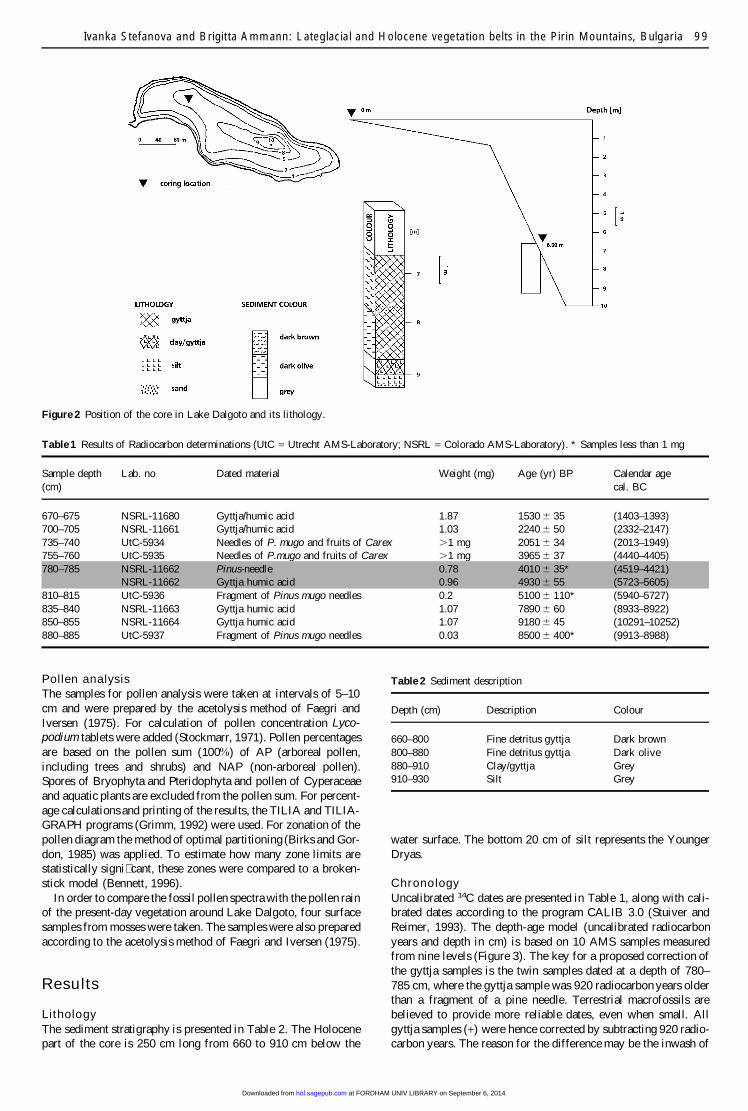
CoringA core 270 cm long was taken with a square-rod piston sampler(Wright, 1991) from the northwestern part of the lake in water6.60 m deep (Figure 2).
DatingAccelerator mass spectrometry 14C dating (AMS) of terrestrialplant macrofossils and of gyttja samples was carried out to estab-lish a local absolute chronology (Table 1). Dates in this paper aregiven as uncalibrated radiocarbon years BP, unless otherwisestated.
at FORDHAM UNIV LIBRARY on September 6, 2014hol.sagepub.comDownloaded from
Ivanka Stefanova and Brigitta Ammann: Lateglacial and Holocene vegetation belts in the Pirin Mountains, Bulgaria 99
Figure 2 Position of the core in Lake Dalgoto and its lithology.
Table 1 Results of Radiocarbon determinations (UtC = Utrecht AMS-Laboratory; NSRL = Colorado AMS-Laboratory). * Samples less than 1 mg
Sample depth Lab. no Dated material Weight (mg) Age (yr) BP Calendar age(cm) cal. BC
670–675 NSRL-11680 Gyttja/humic acid 1.87 1530 6 35 (1403–1393)700–705 NSRL-11661 Gyttja/humic acid 1.03 2240 6 50 (2332–2147)735–740 UtC-5934 Needles of P. mugo and fruits of Carex .1 mg 2051 6 34 (2013–1949)755–760 UtC-5935 Needles of P.mugo and fruits of Carex .1 mg 3965 6 37 (4440–4405)780–785 NSRL-11662 Pinus-needle 0.78 4010 6 35* (4519–4421)
NSRL-11662 Gyttja humic acid 0.96 4930 6 55 (5723–5605)810–815 UtC-5936 Fragment of Pinus mugo needles 0.2 5100 6 110* (5940–5727)835–840 NSRL-11663 Gyttja humic acid 1.07 7890 6 60 (8933–8922)850–855 NSRL-11664 Gyttja humic acid 1.07 9180 6 45 (10291–10252)880–885 UtC-5937 Fragment of Pinus mugo needles 0.03 8500 6 400* (9913–8988)
Pollen analysisThe samples for pollen analysis were taken at intervals of 5–10cm and were prepared by the acetolysis method of Faegri andIversen (1975). For calculation of pollen concentration Lyco-podium tablets were added (Stockmarr, 1971). Pollen percentagesare based on the pollen sum (100%) of AP (arboreal pollen,including trees and shrubs) and NAP (non-arboreal pollen).Spores of Bryophyta and Pteridophyta and pollen of Cyperaceaeand aquatic plants are excluded from the pollen sum. For percent-age calculations and printing of the results, the TILIA and TILIA-GRAPH programs (Grimm, 1992) were used. For zonation of thepollen diagram the method of optimal partitioning (Birks and Gor-don, 1985) was applied. To estimate how many zone limits arestatistically signi� cant, these zones were compared to a broken-stick model (Bennett, 1996).
In order to compare the fossil pollen spectra with the pollen rainof the present-day vegetation around Lake Dalgoto, four surfacesamples from mosses were taken. The samples were also preparedaccording to the acetolysis method of Faegri and Iversen (1975).
Results
LithologyThe sediment stratigraphy is presented in Table 2. The Holocenepart of the core is 250 cm long from 660 to 910 cm below the
Table 2 Sediment description
Depth (cm) Description Colour
660–800 Fine detritus gyttja Dark brown800–880 Fine detritus gyttja Dark olive880–910 Clay/gyttja Grey910–930 Silt Grey
water surface. The bottom 20 cm of silt represents the YoungerDryas.
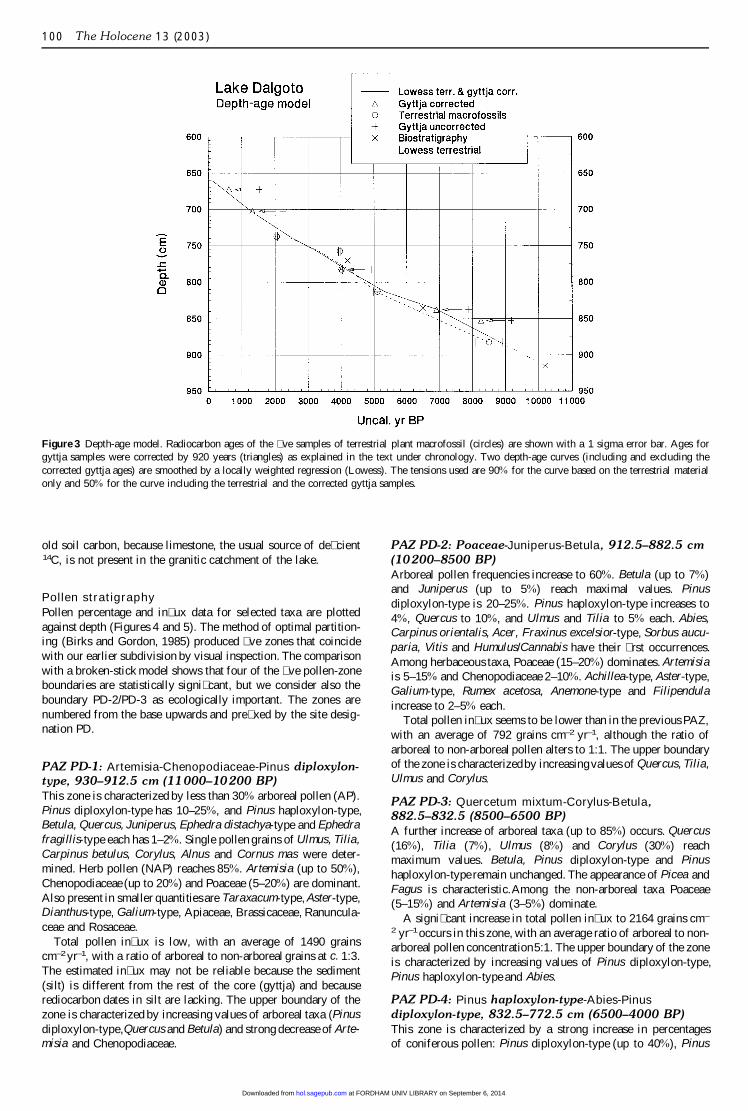
ChronologyUncalibrated 14C dates are presented in Table 1, along with cali-brated dates according to the program CALIB 3.0 (Stuiver andReimer, 1993). The depth-age model (uncalibrated radiocarbonyears and depth in cm) is based on 10 AMS samples measuredfrom nine levels (Figure 3). The key for a proposed correction ofthe gyttja samples is the twin samples dated at a depth of 780–785 cm, where the gyttja sample was 920 radiocarbon years olderthan a fragment of a pine needle. Terrestrial macrofossils arebelieved to provide more reliable dates, even when small. Allgyttja samples (+) were hence corrected by subtracting 920 radio-carbon years. The reason for the difference may be the inwash of
at FORDHAM UNIV LIBRARY on September 6, 2014hol.sagepub.comDownloaded from
100 The Holocene 13 (2003)
Figure 3 Depth-age model. Radiocarbon ages of the � ve samples of terrestrial plant macrofossil (circles) are shown with a 1 sigma error bar. Ages forgyttja samples were corrected by 920 years (triangles) as explained in the text under chronology. Two depth-age curves (including and excluding thecorrected gyttja ages) are smoothed by a locally weighted regression (Lowess). The tensions used are 90% for the curve based on the terrestrial materialonly and 50% for the curve including the terrestrial and the corrected gyttja samples.
old soil carbon, because limestone, the usual source of de� cient14C, is not present in the granitic catchment of the lake.
Pollen stratigraphyPollen percentage and in� ux data for selected taxa are plottedagainst depth (Figures 4 and 5). The method of optimal partition-ing (Birks and Gordon, 1985) produced � ve zones that coincidewith our earlier subdivision by visual inspection. The comparisonwith a broken-stick model shows that four of the � ve pollen-zoneboundaries are statistically signi� cant, but we consider also theboundary PD-2/PD-3 as ecologically important. The zones arenumbered from the base upwards and pre� xed by the site desig-nation PD.
PAZ PD-1: Artemisia-Chenopodiaceae-Pinus diploxylon-type, 930–912.5 cm (11000–10200 BP)This zone is characterized by less than 30% arboreal pollen (AP).Pinus diploxylon-type has 10–25%, and Pinus haploxylon-type,Betula, Quercus, Juniperus, Ephedra distachya-type and Ephedrafragillis-type each has 1–2%. Single pollen grains of Ulmus, Tilia,Carpinus betulus, Corylus, Alnus and Cornus mas were deter-mined. Herb pollen (NAP) reaches 85%. Artemisia (up to 50%),Chenopodiaceae (up to 20%) and Poaceae (5–20%) are dominant.Also present in smaller quantities are Taraxacum-type, Aster-type,Dianthus-type, Galium-type, Apiaceae, Brassicaceae, Ranuncula-ceae and Rosaceae.
Total pollen in� ux is low, with an average of 1490 grainscm–2 yr–1, with a ratio of arboreal to non-arboreal grains at c. 1:3.The estimated in� ux may not be reliable because the sediment(silt) is different from the rest of the core (gyttja) and becauserediocarbon dates in silt are lacking. The upper boundary of thezone is characterized by increasing values of arboreal taxa (Pinusdiploxylon-type,Quercus and Betula) and strong decrease of Arte-misia and Chenopodiaceae.
PAZ PD-2: Poaceae-Juniperus-Betula, 912.5–882.5 cm(10200–8500 BP)Arboreal pollen frequencies increase to 60%. Betula (up to 7%)and Juniperus (up to 5%) reach maximal values. Pinusdiploxylon-type is 20–25%. Pinus haploxylon-type increases to4%, Quercus to 10%, and Ulmus and Tilia to 5% each. Abies,Carpinus orientalis, Acer, Fraxinus excelsior-type, Sorbus aucu-paria, Vitis and Humulus/Cannabis have their � rst occurrences.Among herbaceous taxa, Poaceae (15–20%) dominates. Artemisiais 5–15% and Chenopodiaceae 2–10%. Achillea-type, Aster-type,Galium-type, Rumex acetosa, Anemone-type and Filipendulaincrease to 2–5% each.
Total pollen in� ux seems to be lower than in the previous PAZ,with an average of 792 grains cm–2 yr–1, although the ratio ofarboreal to non-arboreal pollen alters to 1:1. The upper boundaryof the zone is characterizedby increasingvalues of Quercus, Tilia,Ulmus and Corylus.
PAZ PD-3: Quercetum mixtum-Corylus-Betula,882.5–832.5 (8500–6500 BP)A further increase of arboreal taxa (up to 85%) occurs. Quercus(16%), Tilia (7%), Ulmus (8%) and Corylus (30%) reachmaximum values. Betula, Pinus diploxylon-type and Pinushaploxylon-type remain unchanged. The appearance of Picea andFagus is characteristic.Among the non-arboreal taxa Poaceae(5–15%) and Artemisia (3–5%) dominate.
A signi� cant increase in total pollen in� ux to 2164 grains cm–
2 yr–1occurs in this zone, with an average ratio of arboreal to non-arboreal pollen concentration5:1. The upper boundary of the zoneis characterized by increasing values of Pinus diploxylon-type,Pinus haploxylon-type and Abies.
PAZ PD-4: Pinus haploxylon-type-Abies-Pinusdiploxylon-type, 832.5–772.5 cm (6500–4000 BP)This zone is characterized by a strong increase in percentagesof coniferous pollen: Pinus diploxylon-type (up to 40%), Pinus
at FORDHAM UNIV LIBRARY on September 6, 2014hol.sagepub.comDownloaded from

Ivanka Stefanova and Brigitta Ammann: Lateglacial and Holocene vegetation belts in the Pirin Mountains, Bulgaria 101
haploxylon-type (up to 15%) and Abies (up to 25%). Quercus,Ulmus, Tilia and Corylus decrease to 2–3% each. Herb species(NAP) are low (up to 10%). Poaceae is 2%, and Artemisia,Achillea-type, Aster-type, Galium-type, Silene-type and Ranuncu-laceae are up to 1–2% each.
Total pollen in� ux increases to an average of 2682 granscm–2 yr–1, with arboreal pollen accounting for 84% of this � gure.The upper boundary of the zone is marked by decreasing valuesof Pinus haploxylon-type and Abies.
PAZ PD-5: Pinus diploxylon-type-Picea-Fagus, 772.5–660 cm (4200–0 BP)A decrease of Pinus haploxylon-type to 6–11% and Abies to2–5% characterize this uppermost zone. Pinus diploxylon-type(up to 60–83%) is dominant. The participation of Picea (13%)and Fagus (6%) is signi� cant. Single pollen grains of Juglans andCastanea are present. Herb species increase up to 20%. Poaceaeis 2–3%. Permanent presence of anthropophytes is characteristic–Cerealia, Triticum, Secale, Plantago lanceolata, Polygonumaviculare and Scleranthus. In� ux values increase to an average of3421 grains cm–2 yr–1.
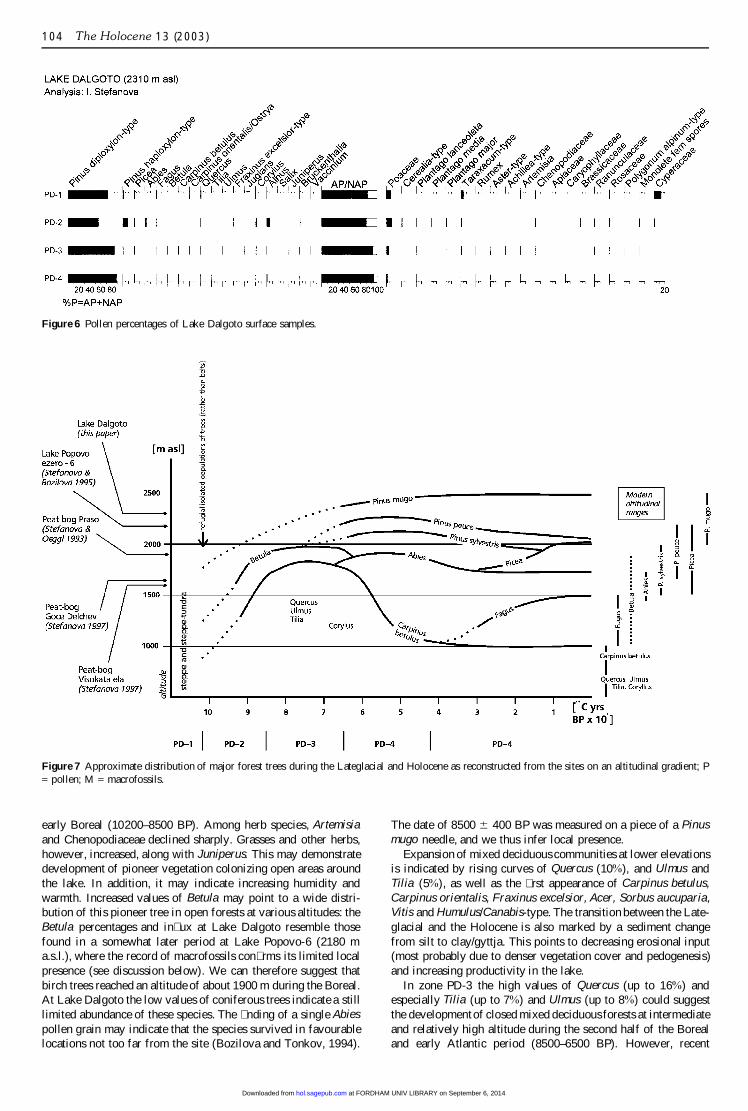
Recent pollen depositionRecent pollen rain at Lake Dalgoto, estimated from the pollencontained within moss samples, is dominated by 82–94% arborealpollen (Figure 6). The main pollen type is Pinus diploxylon-typewith 73–89%, followed by 2–9% Pinus peuce (Pinus haploxylon-type), 2% Picea, 1–3% Fagus, Abies, Betula, Quercus, Carpinusbetulus, Carpinus orientalis, Alnus and Corylus. Single pollengrains of Ulmus, Tilia and Fraxinus excelsior-type occur, andJuniperus, Vaccinium, Bruckenthalia and Salix have values under1%. Among the herb species, Poaceae dominates with 2–9%.
Pollen deposition in a sedimentary basin consists of local,extralocal, regional and extraregionalcomponents (Tauber, 1965),depending essentially upon the area of the sedimentation basin(Janssen, 1973; Jacobson and Bradshaw, 1981). Lakes and mireswith a diameter under 100 m are characterized by local andextralocal components. In contrast, in highland zones above theecotone of the forest limit regional and extraregional componentsin the pollen deposition depend in part on wind direction (Lang,1993; Oeggl, 1994).
Results from the vegetation survey around Lake Dalgoto makeit possible to separate the components of recent pollen deposition.In the surroundingsof the lake are species such as Juniperus sibir-ica, Vaccinium myrtillus, Salix herbaceae and Bruckenthalia spic-ulifolia. They are conceivably representative of the local andextralocal components (up to several hundred metres). Pinusmugo relates to those components too, and it is highly probablethat a signi� cant part of Pinus diploxylon-type pollen originatesfrom this species.
All other tree taxa represented in recent pollen deposition growat lower altitudes and have regional and extraregional origin (upto several km). Trees present in the lower woodlands but notrepresented in the pollen rain include Acer and Fraxinus ornus-type. Presence of these taxa in the fossil pollen spectra must there-fore indicate that their past distributions in the Northern PirinMountains were wider and probably at higher altitudes.
Analysis of the fossil pollen spectra from Lake Dalgoto mustalso take into account the different pollen productivity of theplants and their ability to disperse pollen. Pinus (with exception ofPinus peuce, according to Stefanova, 1996), Quercus, Carpinus,Betula, Alnus and Corylus have high pollen productivity and gooddispersal capacity (Tauber, 1965; Andersen, 1970; 1974;Bradshaw, 1981). In contrast, Tilia, Acer and Fraxinus have poorpollen-dispersal capacities (Andersen, 1970). Between those twogroups are Abies, Fagus and Picea (Andersen, 1970; 1974).Results of the present studies of recent pollen rain at Lake Dal-
goto, along with previous investigations in other subalpine anddifferent forests (beech and coniferous), con� rm these conclusions(Stefanova, 1996).
Discussion
Linking chrono- and biostratigraphyThe oldest minerogenic sediments at 930–910 cm are not dated.Pollen analysis allows biostratigraphic comparison with other 14Cdated diagrams. The percentage of tree taxa in this zone PD-1 isbelow 30%, while herb taxa like Artemisia, Chenopodiaceae andPoaceae may take 80%. The upper boundary with zone PD-2 ischaracterized by decreasing Artemisia and Chenopodiaceae andincreasing Betula and Quercus. Zone R-3 in the pollen diagramof Lake Suhoto in the Rila Mountains (Bozilova et al., 1990) hasidentical characteristics. The radiocarbon date of 10575 6 220BP there indicates that this zone corresponds to the YoungerDryas (11000–10200 BP). With this comparison we can considerthe boundary between zones PD-1 and PD-2 as 10200 BP.
The upper boundary of zone PD-3 is characterized by anincrease of Pinus diploxylon-type, Pinus haploxylon-type andAbies, dated in other pollen diagrams in the Northern Pirin Moun-tains at 6700/6500 BP (Stefanova and Oeggl, 1993; Stefanova andBozilova, 1995).
The following conclusion can be made about the boundaries ofthe pollen zones (Figure 4):
PD-1/PD-2 at 10 200 BPPD-2/PD-3 at 8500 BPPD-3/PD-4 at 6500 (6700) BPPD-4/PD-5 at 4000 BP
Compared with the division of Holocene into chronozones(Mangerud et al., 1974) accepted for Bulgarian mountain regions(Bozilova, 1982; Bozilova et al., 1996), zone PD-1 correlates withthe Younger Dryas, PD-2 with the Preboreal and early Boreal,PD-3 with the late Boreal and early Atlantic, PD-4 with the lateAtlantic and Subboreal, and PD-5 with the late Subboreal andSubatlanthic.
Vegetational historyDuring the Younger Dryas (11000–10200 BP), as represented atthe base of the diagram (PD-1) an open xerophyticherb vegetationwas widely developed probably both around Lake Dalgoto and atlower altitudes, It was dominated by Artemisia and Cheno-podiaceae, together with species of the Asteraceae (Achillea,Aster, Centaurea), Caryophyllaceae (Dianthus, Silene), Apiaceae,Ranunculaceae and Brassicaceae. The trees were of low densityeven at lower elevations, as is re� ected in their low percentages(under 30%; Figure 4) and low in� ux values (Figure 5). Compari-son with the recent pollen spectra, where the pollen percentageof trees reaches 82–94%, con� rms this suggestion (Figure 6).Probably with the exception of Juniperus and Salix (S. herbacea),the arboreal pollen was a product of long-distance transport.Pollen of Pinus, Betula, Quercus, Ulmus, Tilia, Corylus, Carpinusbetulus and Alnus suggest that a variety of tree species found suit-able environmental conditions and existed in small stands at loweraltitudes and provided pollen to the site by distant transport. It isextremely dif� cult to pinpoint the location of such sheltered refug-ial sites so, in Figure 7 we symbolize them just as dots duringthe end of the Lateglacial.
The open vegetation around Lake Dalgoto during the YoungerDryas led to active soil erosion in the Dalgoto catchment area,producing more terrigenous materials in the lake sediments.
A major shift in NAP from 70% to 40% (and a correspondingincrease of tree pollen) in zone PD-2 suggests the gradual refores-tation at low and intermediate elevations during the Preboreal and
at FORDHAM UNIV LIBRARY on September 6, 2014hol.sagepub.comDownloaded from

102 The Holocene 13 (2003)
Figure 4 Pollen percentages of Lake Dalgoto. (a) Trees and shrubs (AP). (b) Herbs and aquatics (NAP). Statistically signi� cant pollen boundaries areindicated by solid lines and the ecologically important limit by a dashed line; the core segments are indicated by . beside the lithology column.
at FORDHAM UNIV LIBRARY on September 6, 2014hol.sagepub.comDownloaded from

Ivanka Stefanova and Brigitta Ammann: Lateglacial and Holocene vegetation belts in the Pirin Mountains, Bulgaria 103
Figure 5 Pollen in� ux of Lake Dalgoto. (a) Trees and shrubs (AP). (b) Herbs and aquatics (NAP).
at FORDHAM UNIV LIBRARY on September 6, 2014hol.sagepub.comDownloaded from
104 The Holocene 13 (2003)
Figure 6 Pollen percentages of Lake Dalgoto surface samples.
Figure 7 Approximate distribution of major forest trees during the Lateglacial and Holocene as reconstructed from the sites on an altitudinal gradient; P= pollen; M = macrofossils.
early Boreal (10200–8500 BP). Among herb species, Artemisiaand Chenopodiaceae declined sharply. Grasses and other herbs,however, increased, along with Juniperus. This may demonstratedevelopment of pioneer vegetation colonizing open areas aroundthe lake. In addition, it may indicate increasing humidity andwarmth. Increased values of Betula may point to a wide distri-bution of this pioneer tree in open forests at various altitudes: theBetula percentages and in� ux at Lake Dalgoto resemble thosefound in a somewhat later period at Lake Popovo-6 (2180 ma.s.l.), where the record of macrofossils con� rms its limited localpresence (see discussion below). We can therefore suggest thatbirch trees reached an altitude of about 1900 m during the Boreal.At Lake Dalgoto the low values of coniferous trees indicate a stilllimited abundance of these species. The � nding of a single Abiespollen grain may indicate that the species survived in favourablelocations not too far from the site (Bozilova and Tonkov, 1994).
The date of 8500 6 400 BP was measured on a piece of a Pinusmugo needle, and we thus infer local presence.
Expansion of mixed deciduous communities at lower elevationsis indicated by rising curves of Quercus (10%), and Ulmus andTilia (5%), as well as the � rst appearance of Carpinus betulus,Carpinus orientalis, Fraxinus excelsior, Acer, Sorbus aucuparia,Vitis and Humulus/Canabis-type. The transition between the Late-glacial and the Holocene is also marked by a sediment changefrom silt to clay/gyttja. This points to decreasing erosional input(most probably due to denser vegetation cover and pedogenesis)and increasing productivity in the lake.
In zone PD-3 the high values of Quercus (up to 16%) andespecially Tilia (up to 7%) and Ulmus (up to 8%) could suggestthe developmentof closed mixed deciduous forests at intermediateand relatively high altitude during the second half of the Borealand early Atlantic period (8500–6500 BP). However, recent
at FORDHAM UNIV LIBRARY on September 6, 2014hol.sagepub.comDownloaded from

Ivanka Stefanova and Brigitta Ammann: Lateglacial and Holocene vegetation belts in the Pirin Mountains, Bulgaria 105
surface samples collected around the lake show only low pollenpercentages of Quercus between 1–2% and single pollen grainsof Tilia and Ulmus transported from the presumable correspond-ing belt of the modern deciduous oak forests from c. 600 m toc. 1100 m a.s.l.
The only macrofossil � nds so far in the Pirin Mountains thatthrow light on the upper limit of the mixed oak deciduous forestsduring the � rst half of the Holocene are from the small mire Prasoat 1900 m and Lake Popovo-6 at 2180 m. The two records unfor-tunately only go back to the 7200 BP (Stefanova and Oeggl, 1993;Stefanova and Bozilova, 1995). Communities of Betula pendulagrew at Praso at 1900 m a.s.l, where they formed the upper forestlimit between 7200 and 6500 BP (Stefanova, 1999). Single macro-fossils of Acer and Quercus were found there together with highpollen percentages (Quercus up to 36%, Tilia up to 25% and Acer1%) for the same period. At Lake Popovo-6 at 2180 m per-centages of the same taxa were lower than at Lake Dalgoto butstill remarkably high: Quercus (10%), Tilia (6%) and Acer (1%).The pollen-source area (related to the basin size; Jacobson andBradshaw, 1981) of the Praso mire at 1900 m (diameter 25 m)should be more local and extralocal, whereas than for LakePopovo-6 should be more regional and extraregional. The occur-rence of Quercus, Acer (with macroremains) and Tilia was thusa widespread phenomenon. We therefore concluded that in theearly Atlantic (and most probably in the second half of the Boreal)the mixed deciduous forest grew close to the upper forest limit at1900 m, which was formed by communities of Betula pendula.Thus the deciduous forest up to 800 m higher than today’s limitat 1100 m. At Lake Dalgoto, high pollen percentages of Tilia andUlmus, which are known to have poor pollen dispersal capacities,may indicate a higher abundance of these taxa in the mixeddeciduous forests during 8500–6500 BP, in comparison with therecent mixed deciduous forest, where Quercus species aredominant.
The participation of Corylus up to 30% in the zone PD-3 pre-sumes the development of this species not only as undergrowthin the deciduous forest but also in the open, where it could formmonodominant communities. The Corylus maximum occurred justwhen the oak values decreased and just before the expansion ofconifers in zone PD-4. If this change marked the beginning of thecooler conditions that brought the conifers after degrading the oakforest, than Corylus could occupy the openings prior to coniferexpansion.
Coniferous species remained restricted. The � rst appearance ofPicea and Fagus is characteristic for zone PD-3. After 6500 BPthe expansion of conifers (Pinus and Abies) began, leading to thelimitation of Betula communities and forcing mixed deciduousforests to a lower altitude. The increased values of Pinusdiploxylon-type can be considered partly as a result of Pinus mugocommunities in the subalpine belt. Its needles were used for radi-ocarbon dating (Table 1). Maximum pollen percentages of Pinuspeuce and Abies suggest that these trees were more importantcomponents of the coniferous belt than today. Macrofossil datafrom other sites in the Northern Pirin show that about 6500 BPPinus peuce was established as a dominant tree in the upper partof the coniferous belt and rose above 2200 m a.s.l., where theupper forest limit was formed. Abies also had its maximum verti-cal range of distribution between 6500 and 4800 BP (Stefanovaand Oeggl, 1993). After 5100 BP (at Lake Dalgoto) or 4800 BP(at Praso mire and Popovo Lake) the abundance of Pinus peucedecreased.
The main features in the vegetation dynamics after 4000 BP(PAZ PD-5) were changes in the coniferous belt. The participationof Pinus peuce (Pinus haploxylon-type) and Abies in the compo-sition of this belt became reduced (Figure 4). The upper limit ofAbies moved down, as is indicated at Lake Dalgoto by pollen(around 3965 BP) and at Praso mire by macrofossils (around 3950
BP). For Pinus peuce the altitudinal range changed after 3000 BP,as is indicated by macrofossils at Lake Popovo-6 and Goce Del-chev peat bog (Figure 7).
The expansion of Picea began after 3000 BP, overcoming thecompetition of Abies, Pinus sylvestris and P. peuce, which formedforest earlier because of the closer proximity of their glacial refu-gia. Picea then became a dominant tree in the upper part of theconiferous belt together with Pinus peuce. The late developmentof the spruce forest in the Pirin Mountains might be a result ofremoteness of Lateglacial refugia of the species in the south-eastern Alps and adjacent Dinaric mountains (Lang, 1994) or fromthe southwestern part of the Romanian Carpathians (Farcaset al., 1999).
High values of Pinus diploxylon-typeand single � nds of Pinusstomata re� ect a further development of Pinus mugo communitiesin the subalpine belt. A slight increase in pollen percentages ofFagus marks an enlargement of its communities and formation ofa fragmented belt in the mountain (today between 900/1000 and1500/1600 m a.s.l., but no macroremains were found).
The picture of the reconstructed changes in the vegetation after3000 BP is rather complicated because of human activity in themountains. The increase in the anthropophytes (Cerealia,Triticum, Secale, Plantago lanceolata, Scleranthus, Polygonumaviculare, etc.) in zone PD-5 as well as the decrease in the totalparticipationof trees and especially of the coniferous communitiescon� rm this view.
Climatic reconstructionThe wide participation of Artemisia and Chenopodiaceae in thecomposition of herb vegetation during the Younger Dryas is anindication of a cold and probably dry climate, which limited thedevelopment of trees. The upward expansion of trees during thePreboreal and the establishment of new taxa is a response towarmer and more humid climatic and edaphic condition. Theearly-Holocene forest vegetation in the Pirin Mountains wasdominated by the thermophilous deciduous communities, and par-ticipation of boreal communities was limited mainly to birch.Recent modelling of early-Holoceneclimate in the Near East indi-cates that the summers were warmer and drier because of highsummer insolation (Kutzbach et al., 1993). Most probably theseclimatic conditions limited the expansion of conifers. The lateexpansion of conifers in the Pirin Mountains after 6500 BP canbe explained by a change in the seasonality of the climate by6000 BP, when summers became signi� cantly cooler and winterswarmer (Huntley and Prentice, 1993).
Conclusions
In summary, we can say that, mainly, two phenomena of the Holo-cene vegetation history in the Pirin Mountains are unexpected andneed an explanation. (1) The belt of mesophilous forests (withQuercus, Ulmus, Tilia) expanded upward from 8500 to 6500 BP.(2) The conifers (Abies and especially Picea) expanded only inthe late Holocene.
Both observations may have a climatic as well as anecological/competitional cause. First, the insolation maximum forsummers during the early Holocene may have favoured the riseof the upper limit of the deciduous tree taxa to about 1900 m a.s.l.(compared to 1100 m a.s.l. today). Before 6500 BP, the absenceof conifers (Abies, Pinus peuce, P. mugo, Picea) eliminated oralleviated potential competition with the deciduous trees. Second,long distance to glacial refuges cannot be the cause for the latedevelopment of the coniferous belt, because the mountains of theBalkan peninsula offered favourable sites for tree survival (Beug,1975; Huntley and Birks, 1983; Birks and Line, 1993; Bennettet al., 1991). The pines (Pinus sylvestris at low and intermediate
at FORDHAM UNIV LIBRARY on September 6, 2014hol.sagepub.comDownloaded from

106 The Holocene 13 (2003)
altitudes, P. peuce and P. mugo at high altitudes) and Abies seemto have had scattered stands over thousands of years before theyexpanded around 6500 BP. This means they had very long estab-lishment times sensu Bennett (1983a; 1983b; 1988a; 1988b;1996).
There are two more general conclusions from this study. First,mountain ecosystems and their development since the last glacialmaximum (LGM) are interesting because of their many ecotonesacross short distances. These may be recorded over thousands ofyears because the formerly glaciated mountains offer lakes asnatural archives at various altitudes. Yet conclusions from pollenstudies alone are more dif� cult than in the lowlands because ofthe importance of pollen transported over long distances. Theimportance of the plant-macrofossil (or stomata) analysis for theestimation of presence or absence of taxa is emphasized (Wick,1996; Birks and Birks, 2000). For the Pirin Mountains, the recordof macroremains of Quercus and Acer at 1900 m about 7200–6500 BP illustrates this point by showing that temperate treeswere indeed present 800 m above their present range, althoughnot necessarily as high as the present conifer tree-limit at 2200m, as the high pollen percentages alone might suggest.
Second, the modern altitudinal ranges of taxa do not fullyre� ect a species’ ecological tolerance, as indicated by the examplein the Pirin Mountains before 6500 BP: Tilia, Quercus and Ulmusreached altitudes about 800 m higher than today. Modern altitudi-nal ranges can be restricted by competition and especially by thehistory of competition (in our example: the absence of the conifersin the early Holocene, and their expansion around 6500 BP). Ifspecies’ ecological tolerance cannot be fully estimated from itsmodern range, its value as a climatic indicator is hampered.
Acknowledgements
We appreciate the cooperation of the staff of Regional Of� ce ofNational Park Pirin in arranging for the � eldwork. The core wasobtained with the aid of H.E. Wright, E. Bozilova, A. Velchev,D. Dimitrov, E. Kozuharova, D. Uzunov, S. Tonkov and R. Kol-chakov. W. Tinner made the depth-age model. W.O. van derKnaap applied a broken-stick model to the pollen data. We wouldlike to thank Jacqueline F.N. van Leeuwen, J. Atanassova, L.Wick and W. Tinner, and the two reviewers of the manuscript forhelpful comments. Our most cordial thanks go to Herbert E.Wright for fruitful discussions and improvement of the manu-script. The study was supported by the Swiss National ScienceFoundation (Grant No. 7BUPJ041258) and the National ScienceFund in So� a, Bulgaria (B-625/96).
References
Andersen, S.T. 1970: The relative pollen productivity and representationof North European trees and correction of tree pollen spectra. GeologicalSurvey of Denmark II, Series 96, 1–99.—— 1974: The differential pollen productivity of trees and its signi� cancefor the interpretation of a pollen diagram from a forested region. In Birks,J.H. and West, R., editors, Quaternary plant ecology, Oxford: Blackwell,109–15.Bennett, K.D. 1983a: Devensian late-glacial and Flandrian vegetationalhistory at Hockham Mere, Norfolk, England. II. Pollen accumulation rates.New Phytologist 95, 489–504.—— 1983b: Postglacial population expansion of forest trees in Norfolk,UK. Nature 303, 164–67.—— 1988a: Holocene geographic spread and population expansion ofFagus grandifolia in Ontario, Canada. Journal of Ecology 76, 547–57.—— 1988b: Post-glacial vegetation history: ecological considerations. InHuntley, B. and Webb, T., editors, Vegetation history, Dordrecht: Kluwer,699–724.
—— 1996: Determination of the number of zones in a biostratigraphicalsequence. New Phytologist 132, 155–70.Bennett, K.D., Tzedakis, P.C. and Willis, K.J. 1991: Quaternary refugiaof north European trees. Journal of Biogeography 18, 103–15.Beug, H.-J. 1975: Changes of climate and vegetation belts in the moun-tains of Mediterranean Europe since the Holocene. Bulletin of Geology19, 101–10.—— 1982: Vegetation history and climatic changes in central and south-ern Europe. In Harding, A.F., editor, Climatic changes in later prehistory,Edinburgh: Edinburgh University Press, 85–102.Birks, H.H. and Birks, H.J.B. 2000: Future uses of pollen analysis mustinclude plant macrofossils. Journal of Biogeography 27, 31–35.Birks, H.J.B. and Gordon, A.D. 1985: Numerical methods in Quaternarypollen analysis. London: Academic Press.Birks, H.J.B. and Line, J.M. 1993: Glacial refugia of European trees –a matter of chance? Dissertationes Botanicae 196, 283–91.Bottema, S. 1974: Late Quaternary vegetation history of northern Greece.Thesis, University of Groningen.Boyagiev, S. 1959: On the geology of the Pirin Mountain (in Bulgarianwith English summary). Annuaire de la Direction Generale desRecherches Geologiques So�a 8, 89–125.Bozilova, E. 1975: Pollen analytical investigation in Northeastern PirinMountain. Annuaire del’ Universite de So�a, Faculte de Biologie 69(2),20–26.—— 1977: The Late Holocene history of vegetation in Northwestern PirinMountain. I. Fitologija 7, 18–24.—— 1982: Holocene chronostratigraphy in Bulgaria. Striae 16, 88–90.Bozilova, E. and Tonkov, S. 1994: The postglacial distribution patternsof Abies in Bulgaria. Dissertationes Botanicae 234, 215–23.Bozilova, E., Filipova, M., Filipovich, L. and Tonkov, S. 1996: Bulgaria.In Berglund, B.E., Birks, H.J.B., Ralska-Jasiewiczowa, M. and Wright,H.E., editors, Palaeoecological events during the last 15000 years, Chich-ester: John Wiley, 701–28.Bozilova, E., Tonkov, S. and Pavlova, D. 1990: Pollen and plant macro-fossil analysis of the Lake Suho Ezero in the South Rila Mountains.Annuaire del’ Universite So�a, Faculte de Biologie 80(2), 48–57.Bradshaw, R.H.W. 1981: Quantitative reconstruction of local woodlandvegetation using pollen analysis from a small basin in Norfolk, England.Journal of Ecology 67, 941–55.Faegri, K. and Iversen, J. 1975: Textbook of pollen analysis. Oxford:Blackwell.Farcas, S., de Beaulieu, J-L., Reille, M., Coldea, G., Diaconeasa, B.,Goeury, C. and Jull, T. 1999. First 14 C dating of Late Glacial andHolocene pollen sequences from Romanian Carpathes. Comtes rendusAcademie Sciences Paris, Sciences de la vie 332, 799–807.Grimm, E.C. 1992: Tilia 1.11 and Tilia-graph 1.17. Spring� eld: IllinoisState Museum, Research and Collection Center.Huntley, B. and Birks, H.J.B. 1983: An atlas of past and present pollenmaps of Europe 0–13000 years ago. Cambridge: Cambridge UniversityPress.Huntley, B. and Prentice, I.C. 1993: Holocene vegetation and climatesof Europe. In Wright, H.E. Jr, Webb, T. III, Ruddimann., W.F., Street-Perrott., F.A. and Bartlein, P.J., editors, Global climates since the lastglacial maximum, Minneapolis: University of Minnesota Press, 136–69.Ivanov, K. and Sotirov, A. 1964: Proceedings of the Institute ofHydrology and Meteorology XVI (in Bulgarian). So� a: Nauca i izkustvo.Jacobson, G.L. and Bradshaw, R.H.W. 1981: The selection of sites forpalaeovegetational studies. Quaternary Research 16, 80–96.Janssen, C.R. 1973: Local and regional pollen deposition. In Birks, H.J.B.and West, R.G., editors, Quaternary plant ecology, Oxford: Blackwell,31–42.Jordanov, D., Velchev, V. and Kozhuharov, S., editors 1963–95: Floraof the Republic of Bulgaria I–X (in Bulgarian). So� a: Publ. House of Bulg.Acad. Sci.Kutzbach, J.E., Guetter, P.J., Behling, P.J. and Selin, R. 1993: Simu-lated climatic changes: results of the COHMAP climate-model experi-ments: In Wright, H.E. Jr, Webb, T. III, Ruddimann, W.F., Street-Perrott,F.A. and Bartlein, P.J., editors, Global climates since the last glacialmaximum, Minneapolis: University of Minnesota Press, 136–69.Lang, G. 1993: Holozane Veranderungen der Waldgrenze in SchweizerAlpen – Metodische Ansatze und gegenwartiger Kenntnisstand. Dissert-ationes Botanicae 196, 317–27.
at FORDHAM UNIV LIBRARY on September 6, 2014hol.sagepub.comDownloaded from

Ivanka Stefanova and Brigitta Ammann: Lateglacial and Holocene vegetation belts in the Pirin Mountains, Bulgaria 107
—— 1994: Quartare Vegetationsgeschichte Europas. Jena: GustavFisher Verlag.Mangerud, J., Andersen, S., Berglund, B. and Donner, J. 1974. Quat-ernary stratigraphy of Norden, a proposal for terminology andclassi� cation. Boreas 3, 109–26.Nikolova, M. 1997: Geographical situation. In Yordanova, M. andDonchev, D., editors, Geography of Bulgaria. Physical geography. Socio-economic geography (in Bulgarian with English summary), So� a:Publishing House of Bulgarian Academy of Sciences, 108–109.Oeggl, K. 1994: The palynological record of human impact on highlandzone ecosystems. In Biagi, P. and Nandris, J., editors, Highland zoneexploitations in southern Europe, Monogra� e di ‘Natura Bresciana’ 20,107–22.Panovska, H., Bozilova, E. and Tonkov, S. 1995: A palaeoecologicalinvestigation on the vegetation history in the Southern Pirin Mts. In Bozi-lova, E. and Tonkov, S., editors, Advances in Holocene palaeoecology inBulgaria, So� a-Moscow: Pensoft, 32–46.Stefanova, I. 1996: Relationship between recent pollen deposition andvegetation in Northern Pirin Mountains. Phytologia Balcanica 2(2), 61–65.—— 1997: New data on the Late Holocene vegetative succession in theNorthern Pirin Mountains: pollen and macrofossil analysis of depositionsfrom peat bogs Goce Delchev and Visokata Ela. Phytologia Balcanica3(2–3), 53–63.—— 1998: Postglacial formation and development of coniferous belt inthe Northern Pirin Mountains I (in Bulgarian with English summary). InProceedings scienti� c papers, Jubilee Scienti�c Conference ‘70th Anniver-sary of the Forest Research Institute’, 6–7 October 1998, So� a, 326–30.—— 1999: The Holocene forest limit in the Northern Pirin Mountains(south-western Bulgaria) – palaeoecological evidence from pollen analy-sis, macrofossil plant remains and 14C dating. Acta Palaeobotanica 2,615–18.Stefanova, I. and Bozilova, E. 1992: Model of palynological investigationwith the application of correction factors from the area of north-easternPirin Mountains (in Bulgarian with English summary). Annuaire del’Universite So�a, Faculte de Biologie 81(2), 31–43.—— 1995: Studies on the Holocene history of vegetation in the NorthernPirin Mts. (southwestern Bulgaria). In Bozilova, E. and Tonkov, S.,
editors, Advances in Holocene palaeoecology in Bulgaria, So� a-Moscow:Pensoft, 9–31.Stefanova, I. and Oeggl, K. 1993: Zur holozanen VegetationsgeschichteSW-Bulgariens: Das Moor Praso im Pirin-Gebirge. Berichte des naturwis-senschaftlich-medizinischen Verein Innsbruck 80, 69–80.Stockmarr, J. 1971: Tablets with spores used in pollen analysis. Pollenet Spores 13, 615–21.Stuiver, M. and Reimer, P.J. 1993: Extended 14C database and revisedCALIB radiocarbon calibration program. Radiocarbon 35, 215–30.Tauber, H. 1965: Differential pollen dispersion and the interpretation ofpollen diagrams. Danmarks Geologiske Undersogelse II R, 89, 1–69.Tzedakis, P.C. 1993: Long-term tree populations in northwest Greece inresponse to Quaternary climatic cycles. Nature 364, 437–40.Velchev, V. and Rusakova, V. 1990: Ecological peculiarities and phytoc-oenological characteristics of Pinus peuce Griseb. in the Pirin and RilaMountains (in Bulgarian with English summary). Annuaires del’ Univers-ite So�a, Faculte de Biologie 80(2), 58–80.Velchev, V. and Vassilev, P. 1987: Ecological and phytocoenologicalinvestigation of the greybark pine (Pinus heldreichii Christ.) in northernpart of Pirin (in Bulgarian with English summary). Annuaires del’ Univ-ersite So�a, Faculte de Biologie 78(2), 57–96.Velev, St. 1997: Climate division (in Bulgarian with English summary). InYordanova, M. and Donchev, D. editors, Geography of Bulgaria. Physicalgeography. Socio-economic geography, So� a: Publishing House of theBulgarian Academy of Sciences, 269–83.Veltshev, V. 1997: Types of vegetation (in Bulgarian with Englishsummary). In Yordanova, M. and Donchev, D., editors, Geography ofBulgaria. Physical geography. Socio-economic geography, So� a: Pub-lishing House of the Bulgarian Academy of Sciences, 269–83.Wick, L. 1996: Late-Glacial and Early-Holocene palaeoenviroment inBrianza, N Italy. II. Quaternario 9(2), 653–60.Willis, K.J. 1992: The late Quaternary vegetation history of northwestGreece. II. Rezina marsh. New Phytologist 121, 101–17.—— 1994a: The vegetational history of the Balkans. Quaternary ScienceReviews 13, 769–88.—— 1994b: Altitudinal variation in the Late Quaternary vegetationalhistory of Northwest Greece. Historical Biology 9, 103–16.Wright, H.E. Jr 1991: Coring tips. Journal of Paleolimnology 6, 37–49.
at FORDHAM UNIV LIBRARY on September 6, 2014hol.sagepub.comDownloaded from
![The influence of refugial population on Lateglacial and ...people.geo.su.se/barbara/pdf/Feurdean_et_al_2007_RPP_f[1].pdf · The influence of refugial population on Lateglacial and](https://img.pdfslide.us/doc/110x75/5e0a4f17dfca9e635f10a958/the-influence-of-refugial-population-on-lateglacial-and-1pdf-the-influence.jpg)




























