Embed Size (px)
Citation preview

1349
Laser Doppler Flowmetry in CA1 Sector ofHippocampus and Cortex After
Transient Forebrain Ischemia in Gerbils
Toshihiko Kuroiwa, MD; Petra Bonnekoh, MD; and Konstantin-Alexander Hossmann, MD
Background and Purpose: Local differences in the hemodynamic response to transient ischemia could beinvolved in the development of selective vulnerability. These differences were studied in vulnerable andnonvulnerable regions of the brain.
Methods: Five gerbils were subjected to 10 minutes of bilateral forebrain ischemia, and cerebral bloodflow was measured continuously in the frontal cortex and CA1 sector of the hippocampus using laserDoppler flowmetry. Carotid artery pressure was recorded simultaneously with a pressure transducer.
Results: After induction of ischemia, blood flow in the cortex and CA1 sector decreased to 11.8% and18.0% of the baseline value, respectively. After release of the vascular occlusion, blood flow in the cortexreturned to the preischemic level at 7.5 minutes (recovery time), reached the hyperemic peak (123.8%) at12.4 minutes (peak latency), and again decreased to the preischemic level at 27.2 minutes. In the CA1sector, blood flow returned to the preischemic level at 2.1 minutes, reached the hyperemic peak (122.2%)at 5.7 minutes, and decreased again to the preischemic level at 21.3 minutes. In both the cortex and CA1sector, recovery time and peak latency correlated inversely with the amount of residual blood flow duringischemia. Histologically, cortical neurons were not injured but only 14.6% of CA1 neurons survived 1 weekafter ischemia.
Conclusions: CA1 neurons were selectively injured despite the milder percentage decrease of blood flowduring ischemia and the more prompt recovery of flow after ischemia. These findings stress theimportance of intrinsic rather than hemodynamic factors in the pathogenesis of selective vulnerability ofCA1 neurons after transient bilateral forebrain ischemia. (Stroke 1992;23:1349-1354)
KEY WORDS • cerebral blood flow • cerebral ischemia • ultrasonics • gerbils
Neurons of the CA1 sector of the hippocampusare selectively vulnerable to brief periods ofglobal ischemia.1 This phenomenon has re-
ceived considerable attention in the past, and numerousstudies have attempted to elucidate the underlyingmechanisms using various animal models of cerebralischemia.
Two basically different pathophysiological processes,namely, pathoclitic (intrinsic) and hemodynamic (ex-trinsic) factors, have been discussed as playing thedominant role in the development of selective vulnera-bility. Vogt and Vogt2 originally emphasized the impor-tance of inherent morphological or biochemical proper-ties for the "pathoclisis" of selectively vulnerableneurons. Since then many studies have focused onvarious intrinsic factors such as anatomical connections,neurotransmitters, and receptor density, and there isnow substantial evidence that glutamate-mediated exci-tation,3 disturbances of intracellular calcium homeosta-sis,4 and inhibition of protein synthesis5 are of particularimportance.
From the Max-Planck-Institute for Neurological Research, De-partment of Experimental Neurology, Cologne, FRG.
Address for reprints: Prof. Dr. K.-A. Hossmann, Max-Planck-Institute for Neurological Research, Department of ExperimentalNeurology, Gleueler StraBe 50, D-5000 Cologne 41, FRG.
Received January 6,1992; final revision received April 20,1992;accepted June 3, 1992.
The role of hemodynamic (extrinsic) factors has alsobeen stressed since Spielmeyer6 first noted a differencein the vascular supply of selectively vulnerable andresistant brain regions. It is generally held that selectivevulnerability is not caused by a more severe density ofischemia,7 but there are uncertainties about the role ofpostischemic hemodynamic factors, which are known toaffect postischemic recovery.8 Postischemic recruitmentof capillaries is less marked in the CA1 sector than inthe cortex,9 but postischemic hyperemia, measured byquantitative autoradiography, is more pronounced.7
See Editorial Comment, p 1354For these reasons, we became interested in studying
the differences in ischemic and postischemic hemody-namics between selectively vulnerable and tolerantbrain regions. We report the results of continuous,simultaneous laser Doppler flow measurements in thevulnerable CA1 sector of the hippocampus and in theresistant cortex of gerbils subjected to 10 minutes oftransient forebrain ischemia.
Materials and MethodsTwenty adult Mongolian gerbils (Meriones unguicula-
tus) of either sex weighing 50-80 g were used. Beforethe experiment, the animals had free access to waterand food. Atropine sulfate (0.01 mg/g body wt) wasinjected subcutaneously approximately 10 minutes be-
by guest on May 30, 2018
http://stroke.ahajournals.org/D
ownloaded from

1350 Stroke Vol 23, No 9 September 1992
fore anesthesia. Anesthesia was induced with 3%halothane and continued with 1% halothane in a gasmixture of 70% nitrogen and 30% oxygen. Rectaltemperature was monitored and kept close to 37.5°Cwith a feedback-controlled heating system (LM-60, ListMedical Electronic, Darmstadt, FRG). Cranial temper-ature was measured with a needle-type thermistorelectrode (type 513 hypodermic temperature probe,YSI Inc., Yellow Springs, Ohio) inserted under theparietal scalp. Both common carotid arteries were ex-posed, and a 5-0 nylon thread was looped around eachartery and connected to manipulators for production ofarterial occlusion by retraction of the thread. The leftexternal carotid artery was exposed under an operatingmicroscope, and a 27-gauge needle connected to apressure transducer and a microinfusion pump wasinserted with its tip close to the carotid bifurcation.10
Carotid artery pressure was monitored throughout theexperiment, clotting in the needle being prevented bythe infusion of saline at 2-5 /il/min.
After stabilization of the carotid artery pressure, theleft frontoparietal cortex was exposed and a corticos-tomy approximately 1-1.5 mm in diameter was made 1.5mm posterior to the bregma and 2 mm left of the sagittalsuture to expose the floor of the lateral ventricle at theroof of the hippocampus CA1 sector. To this purposethe arachnoid membrane was incised and the cortex wascarefully suctioned using a surgical vacuum pump. Ex-posure of the hippocampus was confirmed by observa-tion of the lateral ventricular floor and leakage of asmall amount of cerebrospinal fluid.
A laser Doppler probe (PF303, Perimed, Stockholm,Sweden) was then placed stereotaxically over the lateralventricular floor, and another probe of the same type(PF303) was placed on the frontal cortex 1-2 mmanterior to the coronal suture and 2 mm lateral to themidline. Both sites were inspected microscopically toascertain the absence of large vessels. Artificial cerebro-spinal fluid warmed by a heating system to 37.5°C wasused to irrigate the sites of probe placement throughoutthe experiment, and blood flow was measured continu-ously by laser Doppler flowmetry (Periflux Pf2, Per-imed). Flow returned to zero after killing the animalwith an overdose of anesthesia in a separate experimentusing the same measuring unit. Bilateral transient fore-brain ischemia was induced by occluding both commoncarotid arteries for 10 minutes and was discontinued byremoving the threads. Blood flow and carotid arterypressure were monitored before, during, and after re-circulation for 1 hour. In group 1 (n=6), the gerbilswere killed with an overdose of pentobarbital (100mg/kg) after this interval. In group 2 (n=6) the animalswere weaned from anesthesia, returned to their cages,and kept alive for 1 week before being killed with anoverdose of pentobarbital. The brains were then re-moved and fixed by immersion in 4% and 10% bufferedformalin for 1 and 4 days, respectively. Coronal sections5 /xm thick taken at the center of the laser Doppler flowprobe placement sites on the frontal cortex and lateralventricular floor were stained with Luxol fast blue andcresyl violet and with hematoxylin and eosin. In group 2the number of intact neurons in the hippocampus CA1sector was counted and expressed as cells per millimeterof stratum pyramidale.
Nonischemic control animals («=8) were treated in asimilar way except for vascular occlusion. Four controlgerbils were killed at 1 hour and the other four at 1week after sham operation.
All values are expressed as mean±SD. Statisticalevaluation was performed using Bartlett's test for anal-ysis of variance, unpaired Student's / test for compari-son of experimental and control groups, and pairedStudent's t test for comparison of paired data obtainedin each animal.
ResultsControl Groups
Rectal temperature was kept close to 37.5°C andcranial temperature at about 37.0°C during anesthesia.Recordings of laser Doppler blood flow from the frontalcortex and hippocampus were stable except for a slightbut nonsignificant decrease during sham operation (ma-nipulation of the carotid arteries). Mean carotid arterypressure was 71.3±9.6 mm Hg and stable.
No tissue injury was observed in the area of flowmeasurement in the frontal cortex. A defect of thecortex approximately 1.5 mm wide and continuing to thelateral ventricle over the hippocampus CA1 sector wasobserved in the section at the corticostomy site. In the1-hour survival group, the wall of the tissue defect wasmildly edematous and occasionally slightly hemorrhagic.The density of pyramidal neurons in the hippocampusCA1 sector 1 week after the sham operation was244.5 ±18.4/mm.
Experimental GroupsRectal temperature in the experimental groups was
stable and similar to that in the control groups. Cranialtemperature decreased during ischemia by approxi-mately 0.3°C but returned promptly to the preischemiclevel after recirculation and remained at this levelthroughout the experiment.
Typical recordings of laser Doppler blood flow andcarotid artery pressure are shown in Figure 1. Carotidartery pressure was 64.7±8.5 mm Hg before occlusionof the carotid artery (Figure 2). After occlusion, carotidartery pressure decreased promptly to 11.0±5.0 (range,4-16) mm Hg and remained at this level during isch-emia. After release of the vascular occlusion, carotidartery pressure returned promptly to the preischemicbaseline value and remained stable throughout the 1hour of measurement. In only one gerbil, return ofcarotid artery pressure to the baseline level was delayedfor 2 minutes, the pressure after 1 minute of recircula-tion being 60% of the preischemic level.
Blood flow of the cortex and hippocampus CA1 sectordecreased after vascular occlusion and remained constantthroughout the 10 minutes of ischemia (Figure 3). Resid-ual flow in the CA1 sector of the hippocampus wasconsistently higher (18.0±7.3%) than in the cortex(11.8±4.8%), and the difference was significant (/><0.05).After release of the vascular occlusion, return of bloodflow to the baseline level was delayed for some time(recovery time), but thereafter a transient phase of reac-tive hyperemia developed followed by postischemic hypo-perfusion. The time course of these changes was signifi-cantly different between the two structures (Figures 1 and3). In the cortex, blood flow returned to the baseline value
by guest on May 30, 2018
http://stroke.ahajournals.org/D
ownloaded from
Kuroiwa et al Postischemic Recirculation of CA1 and Cortex 1351
LDF cortex
150i
100%
50
0
200
% 100 •
0
LDF hippocampus
CBP
- ? 2x 40
iBCCA occlusion
2min
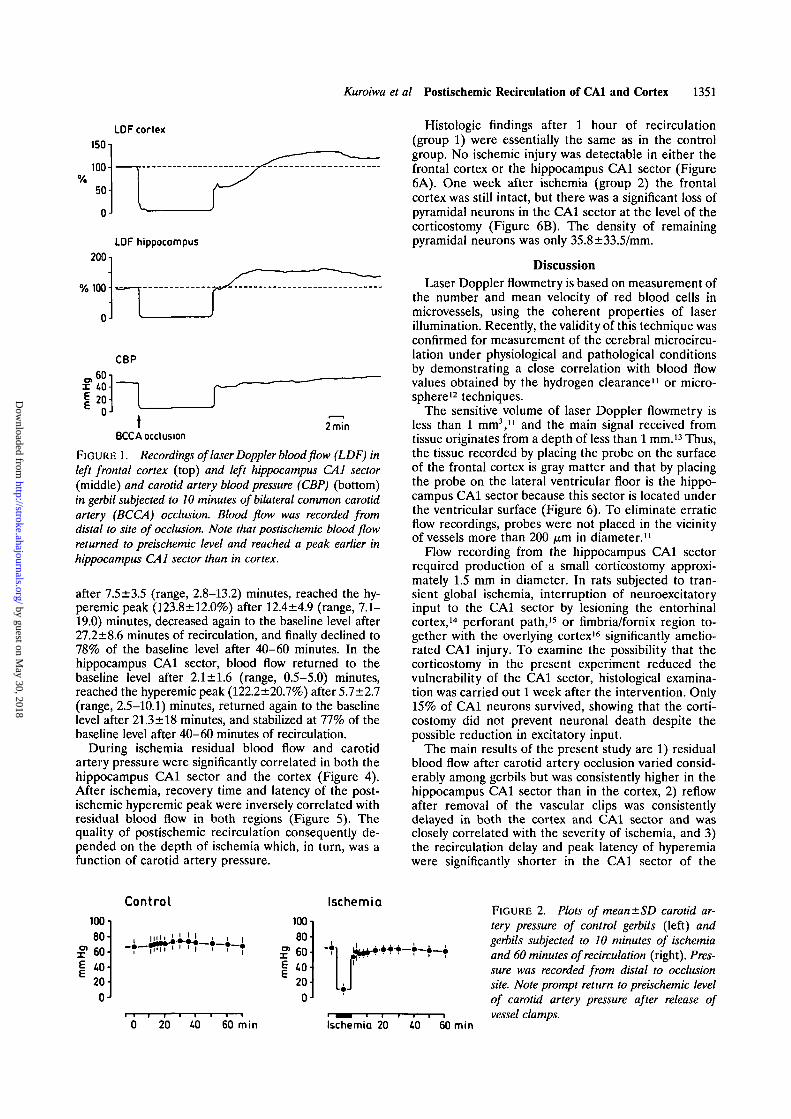
FIGURE 1. Recordings of laser Doppler blood flow (LDF) inleft frontal cortex (top) and left hippocampus CA1 sector(middle) and carotid artery blood pressure (CBP) (bottom)in gerbil subjected to 10 minutes of bilateral common carotidartery (BCCA) occlusion. Blood flow was recorded fromdistal to site of occlusion. Note that postischemic blood flowreturned to preischemic level and reached a peak earlier inhippocampus CA1 sector than in cortex.
after 7.5±3.5 (range, 2.8-13.2) minutes, reached the hy-peremic peak (123.8±12.0%) after 12.4±4.9 (range, 7.1-19.0) minutes, decreased again to the baseline level after27.2±8.6 minutes of recirculation, and finally declined to78% of the baseline level after 40-60 minutes. In thehippocampus CA1 sector, blood flow returned to thebaseline level after 2.1 ±1.6 (range, 0.5-5.0) minutes,reached the hyperemic peak (122.2±20.7%) after 5.1 ±2.1(range, 2.5—10.1) minutes, returned again to the baselinelevel after 21.3 ±18 minutes, and stabilized at 77% of thebaseline level after 40-60 minutes of recirculation.
During ischemia residual blood flow and carotidartery pressure were significantly correlated in both thehippocampus CA1 sector and the cortex (Figure 4).After ischemia, recovery time and latency of the post-ischemic hyperemic peak were inversely correlated withresidual blood flow in both regions (Figure 5). Thequality of postischemic recirculation consequently de-pended on the depth of ischemia which, in turn, was afunction of carotid artery pressure.
Histologic findings after 1 hour of recirculation(group 1) were essentially the same as in the controlgroup. No ischemic injury was detectable in either thefrontal cortex or the hippocampus CA1 sector (Figure6A). One week after ischemia (group 2) the frontalcortex was still intact, but there was a significant loss ofpyramidal neurons in the CA1 sector at the level of thecorticostomy (Figure 6B). The density of remainingpyramidal neurons was only 35.8±33.5/mm.
DiscussionLaser Doppler flowmetry is based on measurement of
the number and mean velocity of red blood cells inmicrovessels, using the coherent properties of laserillumination. Recently, the validity of this technique wasconfirmed for measurement of the cerebral microcircu-lation under physiological and pathological conditionsby demonstrating a close correlation with blood flowvalues obtained by the hydrogen clearance11 or micro-sphere12 techniques.
The sensitive volume of laser Doppler flowmetry isless than 1 mm3,11 and the main signal received fromtissue originates from a depth of less than 1 mm.13 Thus,the tissue recorded by placing the probe on the surfaceof the frontal cortex is gray matter and that by placingthe probe on the lateral ventricular floor is the hippo-campus CA1 sector because this sector is located underthe ventricular surface (Figure 6). To eliminate erraticflow recordings, probes were not placed in the vicinityof vessels more than 200 /u,m in diameter.11
Flow recording from the hippocampus CA1 sectorrequired production of a small corticostomy approxi-mately 1.5 mm in diameter. In rats subjected to tran-sient global ischemia, interruption of neuroexcitatoryinput to the CA1 sector by lesioning the entorhinalcortex,14 perforant path,15 or fimbria/fornix region to-gether with the overlying cortex16 significantly amelio-rated CA1 injury. To examine the possibility that thecorticostomy in the present experiment reduced thevulnerability of the CA1 sector, histological examina-tion was carried out 1 week after the intervention. Only15% of CA1 neurons survived, showing that the corti-costomy did not prevent neuronal death despite thepossible reduction in excitatory input.
The main results of the present study are 1) residualblood flow after carotid artery occlusion varied consid-erably among gerbils but was consistently higher in thehippocampus CA1 sector than in the cortex, 2) reflowafter removal of the vascular clips was consistentlydelayed in both the cortex and CA1 sector and wasclosely correlated with the severity of ischemia, and 3)the recirculation delay and peak latency of hyperemiawere significantly shorter in the CA1 sector of the
100
80
x" 60
| 40-
20
0 J
Control
0 20 40 60 min
100-
80-£ 60-| 40-
20-0-
Ischemia
\rIschemia 20 40 60 min
FIGURE 2. Plots of mean±SD carotid ar-tery pressure of control gerbils (left) andgerbils subjected to 10 minutes of ischemiaand 60 minutes of recirculation (right). Pres-sure was recorded from distal to occlusionsite. Note prompt return to preischemic levelof carotid artery pressure after release ofvessel clamps.
by guest on May 30, 2018
http://stroke.ahajournals.org/D
ownloaded from
1352 Stroke Vol 23, No 9 September 1992
140120100806040200
ControlCortex
UO-i
120-
100-
80-60-40-20-
0-
0 20 40 60 min
IschemiaCortex
11
rJ
140
120
100
80
' 60
40
20
0
140
120
100
* 60
40
20
0
CA1
20 40 60 min
CA1
I I
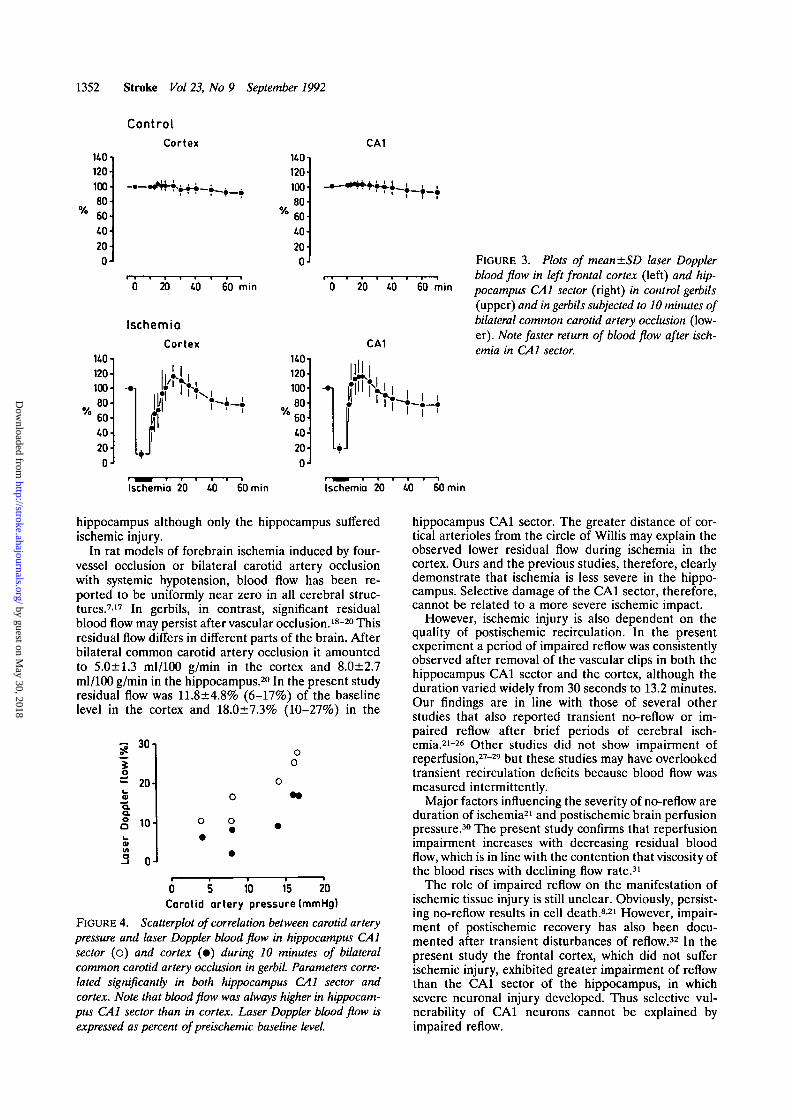
FIGURE 3. Plots of mean±SD laser Dopplerblood flow in left frontal cortex (left) and hip-pocampus CA1 sector (right) in control gerbils(upper) and in gerbils subjected to 10 minutes ofbilateral common carotid artery occlusion (low-er). Note faster return of blood flow after isch-emia in CA1 sector.
Ischemia 20 40 60 min
I '
Ischemia 20 40 60 min
hippocampus although only the hippocampus sufferedischemic injury.
In rat models of forebrain ischemia induced by four-vessel occlusion or bilateral carotid artery occlusionwith systemic hypotension, blood flow has been re-ported to be uniformly near zero in all cerebral struc-tures.717 In gerbils, in contrast, significant residualblood flow may persist after vascular occlusion.18"20 Thisresidual flow differs in different parts of the brain. Afterbilateral common carotid artery occlusion it amountedto 5.0±1.3 ml/100 g/min in the cortex and 8.0±2.7ml/100 g/min in the hippocampus.20 In the present studyresidual flow was 11.8±4.8% (6-17%) of the baselinelevel in the cortex and 18.0±7.3% (10-27%) in the
s.
>ple
r
oa
Lase
r
30-
20-
10-
0-
o
0
o
•
oo
o
• •
0 5 10 15 20Carotid artery pressure (mmHg)
FIGURE 4. Scatterplot of correlation between carotid arterypressure and laser Doppler blood flow in hippocampus CA1sector (o) and cortex (•) during 10 minutes of bilateralcommon carotid artery occlusion in gerbil. Parameters corre-lated significantly in both hippocampus CA1 sector andcortex. Note that blood flow was always higher in hippocam-pus CA1 sector than in cortex. Laser Doppler blood flow isexpressed as percent of preischemic baseline level.
hippocampus CA1 sector. The greater distance of cor-tical arterioles from the circle of Willis may explain theobserved lower residual flow during ischemia in thecortex. Ours and the previous studies, therefore, clearlydemonstrate that ischemia is less severe in the hippo-campus. Selective damage of the CA1 sector, therefore,cannot be related to a more severe ischemic impact.
However, ischemic injury is also dependent on thequality of postischemic recirculation. In the presentexperiment a period of impaired reflow was consistentlyobserved after removal of the vascular clips in both thehippocampus CA1 sector and the cortex, although theduration varied widely from 30 seconds to 13.2 minutes.Our findings are in line with those of several otherstudies that also reported transient no-reflow or im-paired reflow after brief periods of cerebral isch-emia.21-26 Other studies did not show impairment ofreperfusion,27-29 but these studies may have overlookedtransient recirculation deficits because blood flow wasmeasured intermittently.
Major factors influencing the severity of no-reflow areduration of ischemia21 and postischemic brain perfusionpressure.30 The present study confirms that reperfusionimpairment increases with decreasing residual bloodflow, which is in line with the contention that viscosity ofthe blood rises with declining flow rate.31
The role of impaired reflow on the manifestation ofischemic tissue injury is still unclear. Obviously, persist-ing no-reflow results in cell death.8'21 However, impair-ment of postischemic recovery has also been docu-mented after transient disturbances of reflow.32 In thepresent study the frontal cortex, which did not sufferischemic injury, exhibited greater impairment of reflowthan the CA1 sector of the hippocampus, in whichsevere neuronal injury developed. Thus selective vul-nerability of CA1 neurons cannot be explained byimpaired reflow.
by guest on May 30, 2018
http://stroke.ahajournals.org/D
ownloaded from
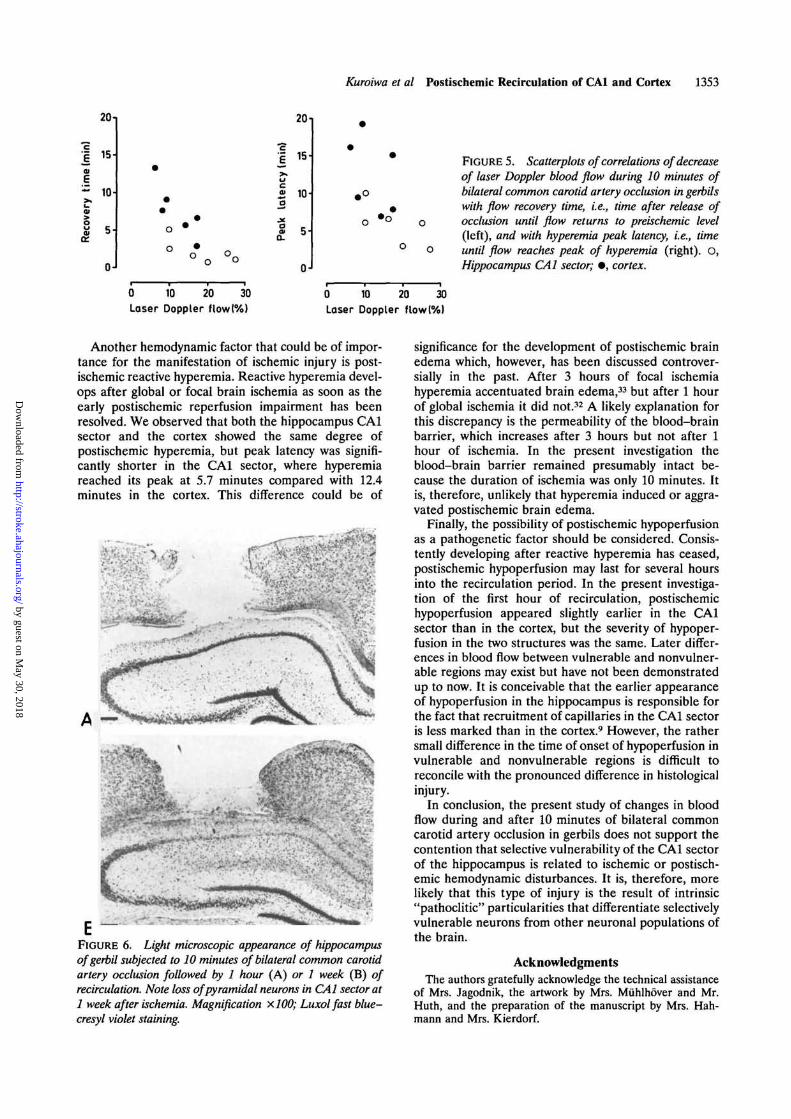
Kuroiwa et al Postischemic Recirculation of CAl and Cortex 1353
20
01
~ 10c>o
S 5
20
#
0
Laser
••
o • *
10 20 30
Doppler flow(%)
c
Ica
J>JC
aa.
15
10
5
0
»o*
0 10 20 30Laser Doppler flowl%)
FIGURE 5. Scatterplots of correlations of decreaseof laser Doppler blood flow during 10 minutes ofbilateral common carotid artery occlusion in gerbilswith flow recovery time, i.e., time after release ofocclusion until flow returns to preischemic level(left), and with hyperemia peak latency, i.e., timeuntil flow reaches peak of hyperemia (right). O,Hippocampus CAl sector; • , cortex.
Another hemodynamic factor that could be of impor-tance for the manifestation of ischemic injury is post-ischemic reactive hyperemia. Reactive hyperemia devel-ops after global or focal brain ischemia as soon as theearly postischemic reperfusion impairment has beenresolved. We observed that both the hippocampus CAlsector and the cortex showed the same degree ofpostischemic hyperemia, but peak latency was signifi-cantly shorter in the CAl sector, where hyperemiareached its peak at 5.7 minutes compared with 12.4minutes in the cortex. This difference could be of
A —
&
-E "FIGURE 6. Light microscopic appearance of hippocampusofgerbil subjected to 10 minutes of bilateral common carotidartery occlusion followed by 1 hour (A) or 1 week (B) ofrecirculation. Note loss of pyramidal neurons in CAl sector at1 week after ischemia. Magnification X100; Luxolfast blue-cresyl violet staining.
significance for the development of postischemic brainedema which, however, has been discussed controver-sially in the past. After 3 hours of focal ischemiahyperemia accentuated brain edema,33 but after 1 hourof global ischemia it did not.32 A likely explanation forthis discrepancy is the permeability of the blood-brainbarrier, which increases after 3 hours but not after 1hour of ischemia. In the present investigation theblood-brain barrier remained presumably intact be-cause the duration of ischemia was only 10 minutes. Itis, therefore, unlikely that hyperemia induced or aggra-vated postischemic brain edema.
Finally, the possibility of postischemic hypoperfusionas a pathogenetic factor should be considered. Consis-tently developing after reactive hyperemia has ceased,postischemic hypoperfusion may last for several hoursinto the recirculation period. In the present investiga-tion of the first hour of recirculation, postischemichypoperfusion appeared slightly earlier in the CAlsector than in the cortex, but the severity of hypoper-fusion in the two structures was the same. Later differ-ences in blood flow between vulnerable and nonvulner-able regions may exist but have not been demonstratedup to now. It is conceivable that the earlier appearanceof hypoperfusion in the hippocampus is responsible forthe fact that recruitment of capillaries in the CAl sectoris less marked than in the cortex.9 However, the rathersmall difference in the time of onset of hypoperfusion invulnerable and nonvulnerable regions is difficult toreconcile with the pronounced difference in histologicalinjury.
In conclusion, the present study of changes in bloodflow during and after 10 minutes of bilateral commoncarotid artery occlusion in gerbils does not support thecontention that selective vulnerability of the CAl sectorof the hippocampus is related to ischemic or postisch-emic hemodynamic disturbances. It is, therefore, morelikely that this type of injury is the result of intrinsic"pathoclitic" particularities that differentiate selectivelyvulnerable neurons from other neuronal populations ofthe brain.
AcknowledgmentsThe authors gratefully acknowledge the technical assistance
of Mrs. Jagodnik, the artwork by Mrs. Miihlhover and Mr.Huth, and the preparation of the manuscript by Mrs. Hah-mann and Mrs. Kierdorf.
by guest on May 30, 2018
http://stroke.ahajournals.org/D
ownloaded from

1354 Stroke Vol 23, No 9 September 1992
References1. Ito U, Spatz M, Walker JT, Klatzo I: Experimental ischemia on
Mongolian gerbils: I. Light microscopical observations. Acta Neu-ropathol (Berl) 1975;32:209-223
2. Vogt C, Vogt O: Der Begriff der Pathoklise. J Psychol Neurol1925;31:245-255
3. Jorgensen MB, Diemer NH: Selective neuron loss after cerebralischemia in the rat: Possible role of transmitter glutamate. ActaNeurol Scand 1982;66:536-546
4. Meldrum B, Evanos M, Griffiths T, Simon R: Ischaemic braindamage: The role of excitatory activity and of calcium entry. Br JAnaesth 1985;57:44-46
5. Bodsch W, Takahashi K, Barbier A, Grosse Ophoff B, HossmannK-A: Cerebral protein synthesis and ischemia. Prog Brain Res1985;63:197-210
6. Spielmeyer W: Zur Pathogenese ortlich selektiver Gehirnveran-derungen. Z Gesamle Neurol Psychiatr 1925;99:756-776
7. Pulsinelli WA, Levy DE, Duffy TE: Regional cerebral blood flowand glucose metabolism following transient forebrain ischemia.Ann Neurol 1982;ll:499-509
8. Crowell RM, Olsson W: Impaired microvascular filling after focalcerebral ischemia in the monkey: Modification by treatment. Neu-rology 1972;22:500-504
9. Imdahl A, Hossmann K-A: Morphometric evaluation of postisch-emic capillary perfusion in selectively vulnerable area of gerbilbrain. Acta Neuropathol (Berl) 1986;69:267-271
10. Kuroiwa T, Bonnekoh P, Hossmann K-A: Threshold of carotidartery back pressure for delayed neuronal injury in the hippocam-pus after bilateral common carotid artery occlusion in gerbils.J Neurol Sci 1990;97:251-259
11. Haberl R, Heizer ML, Marmarou A, Ellis EF: Laser-Dopplerassessment of brain microcirculation: Effect of systemic alter-ations. Am J Physiol 1989;256:H1247-H1254
12. Eyre JA, Essex TJ, Flecknell PA, Bartholomew PH, Sinclair JI: Acomparison of measurements of cerebral blood flow in the rabbitusing laser Doppler spectroscopy and radionuclide labeled micro-spheres. Clin Phys Physiol Meas 1988;9:65-74
13. Bonner RT, Clem TR, Berliner RW: Laser-Doppler continuousreal-time monitor of pulsatile and mean blood flow in tissue micro-circulation, in Chen SH, Chu B, Nossal R (eds): Scattering Tech-niques Applied to Supramolecular and Non-equilibrium System. NewYork, Plenum Publishing Corp, 1981, pp 685-702
14. Jorgensen MB, Johansen FF, Diemer NH: Removal of the entorhi-nal cortex protects hippocampal CA-1 neurons from ischemic dam-age. Acta Neuropathol (Berl) 1987;73:189-194
15. Wieloch T, Lindvall O, Blomqvist P, Gate FH: Evidence for ame-lioration of ischaemic neuronal damage in the hippocampal for-mation by lesion of the perforant path. Neurol Res 1985;7:24-26
16. Buchan AM, Pulsinelli WA: Septo-hippocampal deafferentationprotects CAl neurons against ischemic injury. Brain Res 1990;512:7-14
17. Kagstrom E, Smith ML, Siesjo BK: Recirculation in the rat brainfollowing incomplete ischemia. J Cereb Blood Flow Metab 1983;3:183-192
18. Mies G, Kloiber O, Drewes LR, Hossmann K-A: Cerebral bloodflow and regional potassium distribution during focal ischemia ofgerbil brain. Ann Neurol 1984;16:232-237
19. Paschen W, Djuricic BM, Bosma HJ, Hossmann K-A: Biochemicalchanges during graded brain ischemia in gerbils: II. Regional eval-uation of cerebral blood flow and brain metabolites. J Neurol Sci1983;58:37-44
20. Suzuki R, Yamaguchi T, Kirino T, Orzi F, Klatzo I: The effects of5-min ischemia in Mongolian gerbils: I. Blood-brain barrier, cere-bral blood flow, and local glucose utilization changes. Acta Neuro-pathol (Berl) 1983;60:207-216
21. Ames A, Wright RL, Kowada M, Thurston JM, Majno G: Cerebralischemia: II. The no-reflow phenomenon. Am J Pathol 1968;52:437-453
22. Chiang J, Kowada M, Ames A III, Wright RL, Majno G: Cerebralischemia: III. Vascular changes. Am J Pathol 1968;52:455-476
23. Fischer EG, Ames A, Lorenzo A: Cerebral blood flow immediatelyfollowing brief circulatory stasis. Stroke 1979;10:423-427
24. Ginsberg MD, Myers RE: The topography of impaired micro-vascular perfusion in the primate brain following total circulatoryarrest. Neurology 1972;22:998-1011
25. Harrison MJG, Sedal L, Arnold J, Russel RWR: No-reflow phe-nomenon in the cerebral circulation of the gerbil. J Neurol Neuro-surg Psychiatry 1975;38:1190-1193
26. Ito U, Ohno K, Yamaguchi T, Tomita H, Inaba Y, Kashima M:Transient appearance of "no-reflow" phenomenon in Mongoliangerbils. Stroke 1980;ll:517-521
27. Levy DE, Brierley JB, Plum F: Absence of no-reflow after unilat-eral carotid artery occlusion in the gerbil, in Harper AM, JennettWB, Miler JD, Rowen JO (eds): Blood Flow and Metabolism in theBrain. Proceedings of the 7th International Symposium on CerebralBlood Flow and Metabolism. Edinburgh, Churchill Livingstone Inc,1975, pp 12.28-12.31
28. Little JR, Kerr FWL, Sundt TM Jr: Microcirculatory obstruction infocal cerebral ischemia: Relationship to neuronal alterations. MayoClin Proc 1975;50:264-270
29. Miller CL, Lampard DG, Alexander K, Brown WA: Local cerebralblood flow following transient cerebral ischemia. I. Onset ofimpaired perfusion within the first hour following global ischemia.Stroke 1980;l 1:534-541
30. Fischer EG: Impaired perfusion following cerebrovascular stasis:A review. Arch Neurol 1973;29:361-366
31. Gaehtgens P, Marx P: Hemorrheological aspects of the pathophys-iology of cerebral ischemia. J Cereb Blood Flow Metab 1987;8:259-265
32. Hossmann K-A, Kleihues P: Reversibility of ischemic brain dam-age. Arch Neurol 1973;29:375-384
33. Kuroiwa T, Shibutani M, Okeda R: Blood-brain barrier disruptionand exacerbation of ischemic brain edema after restoration ofblood flow in experimental focal cerebral ischemia. Acta Neuro-pathol (Berl) 1988;76:62-70
Editorial Comment
The phenomenon of selective vulnerability of theCAl sector of the hippocampus to ischemia has beenknown for several years. While it is not necessarilywidely believed that differences in ischemic or post-ischemic blood flow account for the selective vulnera-bility, the discontinuous nature of previously usedblood flow measurement techniques has not totallyeliminated this possibility. The study by Kuroiwa et alhas used a continuous indicator of blood flow, laserDoppler flowmetry, to further enlighten this question.Using two laser Doppler probes, they have providedcontinuous, simultaneous measurements of changes inblood flow in the hippocampus and cortex of ischemic
and postischemic gerbil brains. The results show quiteconvincingly that flow differences in the 1-hour post-ischemic period are not the cause of the selectivevulnerability of the hippocampus. The authors are tobe commended on their creative use of a new tech-nique to clarify an old problem. Their results furthersupport the importance of other, intrinsic "patho-clitic" differences in the pathogenesis of selectivehippocampal cell death.
Earl F. Ellis, Guest EditorDepartment of Pharmacology and Toxicology
Medical College of VirginiaRichmond, Va.
by guest on May 30, 2018
http://stroke.ahajournals.org/D
ownloaded from

T Kuroiwa, P Bonnekoh and K A Hossmannischemia in gerbils.
Laser doppler flowmetry in CA1 sector of hippocampus and cortex after transient forebrain
Print ISSN: 0039-2499. Online ISSN: 1524-4628 Copyright © 1992 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Stroke doi: 10.1161/01.STR.23.9.1349
1992;23:1349-1354Stroke.
http://stroke.ahajournals.org/content/23/9/1349World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://stroke.ahajournals.org//subscriptions/
is online at: Stroke Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer available in the
Permissions in the middle column of the Web page under Services. Further information about this process isOnce the online version of the published article for which permission is being requested is located, click Request
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office.Stroke Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on May 30, 2018
http://stroke.ahajournals.org/D
ownloaded from