Embed Size (px)
Citation preview

Analytical Biochemistry 447 (2014) 126–132
Contents lists available at ScienceDirect
Analytical Biochemistry
journal homepage: www.elsevier .com/locate /yabio
Laser capture microdissection after c-glutamyl transferasehistochemistry: An optimization for gene expression analysis
0003-2697/$ - see front matter � 2013 Elsevier Inc. All rights reserved.http://dx.doi.org/10.1016/j.ab.2013.11.015
Abbreviations: GGT, c-glutamyl transferase; GMNA, c-glutamyl-4-methoxy-2-naphthylamide; LCM, laser capture microdissection; qRT–PCR, quantitative reversetranscriptase polymerase chain reaction; DEN, diethylnitrosamine; 2AAF, 2-ace-tylaminofluorene; DEPC, diethylpyrocarbonate; RIN, RNA integrity number; rRNA,ribosomal RNA; mRNA, messenger RNA; cDNA, complementary DNA; ANOVA,analysis of variance.⇑ Corresponding author. Fax: +52 (55) 5350 1999.
E-mail address: [email protected] (J.I. Pérez Carreón).
Julia Esperanza Torres Mena a,b, Ricardo Sánchez Rodríguez a, Valeria Quintanar Jurado a,Raúl Mojica Espinosa a, Luis Del Pozo Yauner a, Jorge Meléndez Zajgla a, Saúl Villa Treviño b,Julio Isael Pérez Carreón a,⇑a Instituto Nacional de Medicina Genómica, México, Distrito Federal, Mexicob Departamento de Biología Celular, Centro de Investigación y de Estudios Avanzados del I.P.N, Distrito Federal, Mexico
a r t i c l e i n f o
Article history:Received 27 September 2013Received in revised form 8 November 2013Accepted 11 November 2013Available online 20 November 2013
Keywords:RNACarcinomaAtherosclerosisLiver neoplasmsReverse transcriptase PCRDNA microarrays
a b s t r a c t
c-Glutamyl transferase (GGT) is useful as a marker in pathological conditions, including several types ofcancer. We optimized the histochemical detection of GGT to assay the gene expression profiles of pheno-type-specific cells selected by laser capture microdissection (LCM). For optimization, we used the livers ofrats subjected to hepatocarcinogenesis. This model induced nodules of hepatocytes and tumors with GGTactivity. To obtain sufficient high-quality RNA after histochemistry and LCM, we included an RNase inhib-itor and air-dried the tissue sections. This optimization allowed the visualization of GGT activity in situand a yield of 1.4 to 2.0 lg of total RNA from 15 to 18 mm2 of microdissected tissue (20 mm thickness).The average RNA integrity number in GGT-positive tissue, determined by chip–capillary electrophoresis,was 6.9, and the 28S/18S ribosomal RNA (rRNA) ratio was 1.4. The RNAs were processed for the Rat Gene1.0 ST Array (Affymetrix). Comparable quality control metrics, such as signal intensity and RNA degrada-tion plots, were found between the LCM samples and non-LCM tissue. The increased expression of Ggt1expected in GGT-positive tissue was confirmed by microarrays and quantitative reverse transcriptasepolymerase chain reaction (qRT–PCR). This optimization provided a suitable method for whole-transcriptanalysis of GGT-positive tissue isolated using LCM.
� 2013 Elsevier Inc. All rights reserved.
The enzyme c-glutamyl transferase (GGT) is localized to themembranes of specific cells in diverse organs such as kidney, pan-creas, and liver. Its active site is oriented toward the outer cell sur-face. The strongest expression of GGT is in the brush border of theproximal convoluted tubule of the kidney; pancreatic acinar cellsand bile canaliculi of liver have lower expression than the kidney[1]. GGT is involved in glutathione metabolism; it removes the c-glutamyl group from glutathione and leukotriene C4 and transfersit to acceptor molecules such as amino acids. This distinctive enzy-matic activity is used for the histochemical demonstration of GGTactivity using c-glutamyl-4-methoxy-2-naphthylamide (GMNA) asa synthetic substrate, glycylglycine as the acceptor molecule, andFast Blue BBN as the chromogen in an azo coupling reaction [2].
The final product of the reaction is a red–orange insoluble dye thatcontrasts the GGT-positive cells from the surrounding negativetissue.
Several histological studies report the high expression of GGT inpathologies associated with oxidative stress and inflammation.GGT is used as a tumor marker in experimental liver carcinogenesisin rats [3,4], where it distinguishes altered hepatocyte foci, dys-plastic nodules, and hepatocellular carcinomas. In human cancer,GGT activity is elevated in some tumors of the liver [5], kidney[6], lung [7], colon [8], prostate [9], ovary [10], soft tissue sarcoma[11], and breast [12] as well as in leukemias [13] and melanoma[14]. This enzyme is associated with oxidative stress, drug resis-tance, invasion, and tumor progression [15–17]. In fact, GGT is pro-posed as a common marker for the detection of human epithelialneoplasms [4]. Other non-cancer-related human pathologies withincreased GGT activity include atheromatous diseases [18] and be-nign prostatic hypertrophy [19]. Thus, GGT expression is a usefulmarker for focal lesions in pathological tissue.
Laser capture microdissection (LCM) allows the precise micro-scopic capture of cell populations from heterogeneous tissue forthe purification of biomolecules such as DNA, RNA, proteins, and

Microdissection of GGT-tissue for gene expression / J.E. Torres Mena et al. / Anal. Biochem. 447 (2014) 126–132 127
metabolites. Several studies denote the importance of couplingLCM and RNA extraction for gene expression analysis [20–22].These studies use staining methods to differentiate the cells ofinterest such as hematoxylin, eosin, methyl green, cresyl violet,and HistoGene. However, these dye-based protocols stain nucleiand cytoplasm to differentiate cellular morphology without phe-notypic specificity between cells of the same type. Specimens canbe stained by immunohistochemistry or immunofluorescence,allowing for the separation of cellular populations by their antigenphenotype [21,23]. Furthermore, the histochemistry of alkalinephosphatase and myofibrillar ATPase were successfully used forLCM and gene expression analysis with reliable cellular specificity[24,25]. An important challenge is to preserve the RNA integrityand quantity from a small amount of starting tissue during tissueprocessing, staining, LCM, and RNA isolation. RNA degradation be-gins within minutes when the tissue is thawed [26]; this process isexacerbated in aqueous solutions at room temperature when ribo-nucleases are active, but RNA degradation may also occur duringLCM. In this study, we optimized the protocol for measuring histo-chemical GGT activity with RNA protection. We obtained GGT-po-sitive cell populations using LCM of the liver of rats subjected to amodel of hepatocarcinogenesis. We collected sufficient quantityand integrity of RNA for gene expression assays such as quantita-tive reverse transcriptase polymerase chain reaction (qRT–PCR)and DNA microarrays.
Materials and methods
Rat tissues
Twenty rats (180–200 g) were subjected to the hepatocarcino-genesis protocol using the resistant hepatocyte model as describedpreviously [27] with some modifications. The rats were initiatedwith a dose of diethylnitrosamine (DEN, 200 mg/kg body weight,i.p.). Then, 2-acetylaminofluorene (2AAF, 25 mg/kg) was orallyadministered for 3 consecutive days starting at day 7, and a 70%partial hepatectomy was performed under ether anesthesia atday 10 as described previously [28]. The rats were sacrificed at4 months, when the livers contained nodules of hepatocytes, andat 9 and 17 months after DEN administration, when tumors haddeveloped as reported [29]. Livers were obtained, washed in salinesolution, frozen in 2-methyl butane with liquid nitrogen, andstored at �70 �C.
GGT histochemistry with RNA protection
Cryosections of liver with a thickness of 20 lm were cut on acryostat (Leica, CM3050S, Germany) at �16 �C and mounted onPEN Membrane Glass Slides (Applied Biosystems, LCM0522, Ger-many). The histochemical protocol was based on the substrates re-ported by Rutenburg and coworkers [2] with modifications
Table 1Modification of GGT histochemical method for LCM
Rutenburg method
Fixation NoneSubstrate concentration 1� GMNA (0.125 mg/ml)
1� Glycylglycine (0.5 mg/ml)1� Fast Blue BBN (0.5 mg/ml)
Buffer 100 mM Tris pH 7.4Reaction time Kidney, 3 min
Liver, 15 minPost-reaction Chelation with CuSO4 and couDrying/mounting Yes/yes
including fixation, increased substrate concentration, the use ofRNase inhibitor diethylpyrocarbonate (DEPC), the use of Mops buf-fer instead of Tris buffer, reduced time of reaction, and drying oftissue samples. All solutions were prepared with RNase-free water,and DEPC was included as described in Table 1.
The staining protocol for each slide was as follows. First, tissueswere fixed with 900 ll of absolute ethanol (molecular biologygrade) and 0.05% (v/v) DEPC for 5 min at �20 �C. After fixation,300 ll of reaction solution was added, which contained 0.25 mg/ml GMNA, 2 mg/ml glycylglycine, and 0.5 mg/ml Fast Blue B. Thesolution was freshly prepared in 50 mM Mops (pH 7.4) with 0.1%(v/v) DEPC. The reaction time was determined by the obvious pres-ence of red-colored precipitate, which unequivocally appearedafter 12 min for liver. Finally, to dry the histological sections, theslides were briefly placed by the flame of a Bunsen burner withoutexcess warming.
LCM
The LCM was performed with microdissection equipment(Arcturus, Veritas 704). This versatile computer-assisted systemenabled ultraviolet (UV) laser cutting of the tissue of interest; thehistological section was supported by a special slide with amembrane that allowed precise dissection. The targeted tissuewas captured by a cap (CapSure Macro LCM Caps, AppliedBiosystems) that becomes adhesive by pulses of infrared laser.The optimal laser settings for LCM of liver tissue were 80 mW pulsepower, 8000 ls duration, and 100 mV intensity. The cutting laserwas set to produce a spot 2 lm in diameter at a power of20 mW. The caps carrying the tissue were placed in a GeneAmptube (Applied Biosystems, N8010611) that contained 320 ll oflysis buffer RLT (Qiagen, RNeasy mini kit) supplemented with 1%(v/v) b-mercaptoethanol. Each tube was inverted to allow totalimmersion of the tissue in the lysis buffer. The tissue was immedi-ately homogenized with the vortex at slow speed for the RNAextraction.
RNA extraction and integrity
Total RNA was extracted using an RNeasy mini kit (Qiagen).This column technology combines a strong protein denaturant,guanidinium thiocyanate, with silica-based membrane purification.The total RNA was eluted with 30 ll of RNase-free water. RNAconcentration and purity were analyzed using a spectrophotome-ter (NanoDrop, ND-1000). All samples had a purity ratio of260/280 nm >2.0. The RNA integrity was assessed by twomeasures. First, an RNA integrity number (RIN) was generated bythe Agilent bioanalyzer 2100 using the RNA 6000 Nano chip.Second, the 28S/18S ribosomal RNA (rRNA) ratio was obtained byanalysis of the gel-like image produced in the bioanalyzer usingthe ImageJ Gel analyzer tool [30].
Optimized method for LCM
Absolute ethanol with 0.05% DEPC2� GMNA (0.25 mg/ml)4� Glycylglycine (2 mg/ml)1� Fast Blue B (0.5 mg/ml)50 mM MOPS pH 7.4 with 0.1% DEPCKidney, 30 sLiver, 12 min
nterstaining NoneYes/no

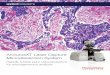
Fig.1. LCM of GGT-stained liver and kidney tissues. GGT histochemical activity wasdetected in frozen tissues using our RNA protection protocol. (A) Rat liver tissuewith GGT-positive nodules. The captured positive (arrow) and negative tissue afterLCM is on the right, and the remaining tissue after microdissection is on the left. (B)Kidney of a normal rat. The remaining tissue after microdissection illustrates thedifferential activity of GGT between the renal medulla and pelvis (left) and thecaptured positive renal medulla (arrow) and the low-level activity in the renalpelvis after LCM (right).
128 Microdissection of GGT-tissue for gene expression / J.E. Torres Mena et al. / Anal. Biochem. 447 (2014) 126–132
qRT–PCR analysis
Complementary DNA (cDNA) was synthesized from 150 ng oftotal RNA in a 5-ll reaction using the High-Capacity RNA to cDNAMaster Mix (Applied Biosystems, USA) according to themanufacturer’s instructors. Real-time quantitative PCR was per-formed using TaqMan Gene Expression Assays according to themanufacturer’s protocol for the gene Ggt1 (Rn01428468_g1). 18SrRNA (Rn03928990) was used as reference of expression. The reac-tion was performed in 15 ml using a 1:10 dilution of cDNA with thenuclease-free water adjusted accordingly.
All samples were run in triplicate. Gene expression was mea-sured by relative quantitation using SDS version 2.4 software (Ap-plied Biosystems, USA). The DDCt method was used to calculate therelative amount of messenger RNA (mRNA) using 18S rRNA as theendogenous control and normal adult liver as a reference control.The same baseline correction and Ct were used for all samples.
Affymetrix microarrays for gene expression
Briefly, first-strand cDNA was obtained from 200 ng of totalRNA using Superscript II reverse transcriptase and poly(T) oligo-mer. Then, the second-strand cDNA was produced. The comple-mentary RNA (cRNA) was obtained by in vitro transcription andused as the template for a second cycle of cDNA synthesis withdUTP incorporation. The obtained cDNA was fragmented using ura-cil–DNA glycosylase. The fragments (40–70mers) were biotin-la-beled by a deoxynucleotide terminal addition reaction. Thelabeled cDNA product was hybridized to the Rat Gene 1.0 ST micro-array (Affymetrix) for 17 h at 45 �C. Samples were washed withlow (SSPE: saline–sodium phosphate–EDTA) and high (100 mMMES and 0.1 M NaCl) stringency buffers and stained with strepta-vidin–phycoerythrin using the Affymetrix Fluidics Station 450FS450_0007 protocol. The arrays were scanned with a GeneChipScanner 3000 7G (Affymetrix), and Expression Console software(Affymetrix) was used to obtain the intensity signal and qualityof the scanned arrays. The software provided summary reportsand metrics, including average background and positive versusnegative area under the curve for the array QC evaluation. The Bio-conductor packages ‘‘affy,’’ ‘‘affyPLM,’’ and ‘‘limma’’ were used togenerate box plots and degradation plots to evaluate the qualityof RNA obtained for the hybridized arrays. The quality controland normalization of arrays were also verified with the open-source package ArrayAnalysis.org [31].
Data analysis
A one-way analysis of variance (ANOVA) was used for compar-ing differences in gene expression and RNA integrity among LCMsamples. Statistical significance was set at P < 0.05.
Results
GGT activity and LCM in histological sections
The optimized method for GGT histochemistry was tested in nor-mal kidneys and livers of carcinogen-treated rats. We found similarenzymatic activity as the original Rutenburg method, which in-volved a red enzymatic precipitate in positive cells (Fig. 1). The kid-neys displayed a spectrum of GGT activity; activity was high in therenal medulla, medium in the renal cortex, and low/diffuse in the re-gion of renal pelvis. In the livers of the hepatocarcinogenic rats sac-rificed at 4 months, GGT activity was observed in two types ofnodules, remodeling and persistent, as described previously[32,33]. In this model, the rats sacrificed after 9 months showed
hepatocellular tumors that were GGT positive. In normal liver fromcontrol rats, activity of GGT was microscopically visualized in biliaryepithelial cells as reported elsewhere [2,34]. The livers of rats sacri-ficed at 4 and 9 months were used for LCM, and the negative and po-sitive GGT tissue regions of nodules at 4 months were collected.
RNA integrity and Affymetrix microarrays
To compare RNA integrity of liver sections after GGT histochem-istry using the two methods described in Table 1, the presence ofrRNA was verified by gel electrophoresis. The RNA degraded duringthe Rutenburg histochemistry procedure. However, the optimizedmethod protected the RNA during the first 2 h but failed to protectit at 4 h (Fig. 2A). The GGT-positive and GGT-negative tissue areasof nodules were microdissected within the first hour post-GGTstaining. The dissected area of tissue ranged from 10 to 20 mm2
(20 mm thickness), with a yield of 1 to 2 mg of RNA (Table 2). TheRNA integrity was verified by capillary electrophoresis, and twomeasurements were used: the RIN and the 28S/18S rRNA ratio(Fig. 2B and C). The average RIN of frozen liver was 7.9 (28S/18S ra-tio of 1.6), and it decreased after LCM in GGT-positive and GGT-negative areas to 6.9 and 6.6, respectively (28S/18S ratios of 1.4and 1.3) (Table 2). RNA samples with a RIN above 6.0 were used

Fig.2. RNA integrity of liver processed by GGT histochemistry and LCM. (A) Agarose gel electrophoresis of RNA obtained from 0 to 4 h after GGT histochemistry using theRutenburg method or the optimized protocol (Table 1). (B, C) Capillary electrophoresis of RNA obtained after GGT histochemistry and LCM. (B) Representativeelectropherograms from the GGT-negative (lower activity) and GGT-positive microdissected samples. The RIN for each sample is shown. (C) Image of RNA bands denoting theabundance of 28S rRNA and the calculated 28S/18S ratios. For each RNA electrophoresis, a sample of RNA from frozen tissue was included. L, ladder.
Microdissection of GGT-tissue for gene expression / J.E. Torres Mena et al. / Anal. Biochem. 447 (2014) 126–132 129
for Affymetrix microarrays, and the quality control of the arrayswas analyzed using Affymetrix Expression Console software andby the website ArrayAnalysis.org [31]. The 15 LCM samplesshowed a satisfactory raw signal intensity that was comparableto the non-LCM samples (Fig. 3A). We displayed the RNA degrada-tion in a plot where the intensities of probe sets were ordered from
the 50 end to the 30 end (Fig. 3B). Because RNA degradation startedfrom either the 50 or 30 end, RNA that was too degraded produced acurve with a pronounced slope [35]. The obtained graph showedagreement between rat arrays with a similar profile from 50 to 30
in both LCM samples (average RIN = 6.8) and non-LCM samples (ar-rays 16 to 20, average RIN = 7.9).

Table 2RNA integrity and quantity of LCM liver samples.
Liver LCM–GGT
Frozen (n = 12) Pos (n = 15) Neg (n = 7)
LCM area (mm2) 18.4 ± 12.2 15.7 ± 5.3Concentration (ng/ll) 1032.2 ± 536.2 67.4 ± 44.1 47.4 ± 12.9Total RNA lg/extraction 30.9 ± 16.1 2.0 ± 1.37 1.4 ± 0.4RIN 7.9 ± 0.9 6.9 ± 0.4* 6.6 ± 0.6*
28S/18S ratio 1.6 ± 0.2 1.4 ± 0.2* 1.3 ± 0.2*
Note: Means ± standard deviations of the indicated numbers of animals and inde-pendent RNA extractions (n) are shown.* P < 0.01 (statistically significant difference compared with the frozen sample byone-way ANOVA).
130 Microdissection of GGT-tissue for gene expression / J.E. Torres Mena et al. / Anal. Biochem. 447 (2014) 126–132
Ggt1 mRNA expression
The expression level of Ggt1 was obtained from normalizedAffymetrix arrays of microdissected nodules and tumor samplesin rat livers (Fig. 4). Expression was also measured by TaqManqRT–PCR. Both sources of data showed a similar gene expressionprofile. As observed for GGT activity in histological sections, therewas increased gene expression in the nodules at 4 months and inthe tumors at 9 and 17 months. The GGT-positive nodules showedhigher Ggt1 expression than did the GGT-negative regions of thenodules. Based on the qRT–PCR (Fig. 4B), this difference corre-sponded to a 52-fold increase in Ggt1 expression in positive nod-ules compared with negative nodules. These data were inagreement with the LCM criteria for histological GGT activity.
Discussion
We optimized the classical method of Rutenburg and coworkersfor histochemical demonstration of GGT expression [2]. This meth-od has been widely used in several histological studies in rats andhumans that describe GGT activity in normal adult tissues, fetal tis-
Fig.3. Intensities and RNA degradation plots obtained in microarrays using LCM sampleTumors at 9 months (T 9 mo) and nodules at 4 months were processed for LCM accordinwere directly processed for RNA isolation. (A) Box plot of the raw intensities of 20 microathe microarrays.
sues, and neoplasias [36–39]. The optimized staining protocol con-sists of an enzymatic reaction that uses the same substratesdescribed by Rutenburg and coworkers. Its efficacy was tested tosatisfy several criteria. First, a positive GGT reaction in the tissuesections was required. Second, the protocol needed to preservethe tissue morphology at the macroscopic and microscopic levels.Third, the sections needed to be compatible with the LCM proce-dure. Finally, the RNA was preserved in the tissue of interest andwas suitable for gene expression analyses. There were three mainmodifications from the original Rutenburg protocol: (i) increasedsubstrate concentration for reduced reaction time; (ii) inclusionof RNase inhibitor DEPC, which was compatible with enzymaticactivity; and (iii) selection of Mops as a buffer that is compatiblewith GGT activity and DEPC. With these modifications, we col-lected GGT-positive hepatocytes from carcinogen-treated rat liv-ers. The tissue sections were suitable for LCM of specific lesions,and sufficient high-quality RNA was obtained for gene expressionexperiments such as DNA microarrays (Affymetrix technology)and qRT–PCR (TaqMan technology). The differential expression ofGgt1 between GGT-positive tissue (nodules and tumors) andGGT-negative tissue (normal liver and the negative regions ofGGT nodules) was confirmed by DNA microarrays and qRT–PCR.
The selection of cell-specific populations from tissue is impor-tant for precise gene expression profiling. The majority of tissuesin normal and pathological conditions, such as neoplasia and in-flamed organs, present a heterogeneous microenvironment inwhich different cell types might be involved. LCM is a useful histo-logical tool for obtaining specific cell types for genetic research.Several studies have successfully performed LCM for gene expres-sion analysis and obtained precise data in normal cells such asbreast epithelium [40] and neuronal subtypes [41] and in patholog-ical conditions such as pre-neoplastic foci in liver [42] and prostatecancers [22]. These studies use routine histological dyes such asmethyl green, hematoxylin and eosin (H&E), HistoGene, and cresylviolet acetate that can differentiate cell types and subcellular struc-tures; however, they cannot distinguish different enzymatic pheno-
s. Total RNA of liver samples was processed for Affymetrix microarrays as follows.g to positive (Pos) or negative (Neg) GGT activity. Tumors at 17 months (T 17 mo)
rrays. (B) RNA degradation plot denoting similar intensity of 50 and 30 probes among

Fig.4. Ggt1 expression in LCM samples. Tumors at 9 months (T 9 mo) and nodules at 4 months were processed for LCM according to positive (Pos) or negative (Neg) GGTactivity. Tumors at 17 months (T 17 mo) and normal livers (NL) were directly processed for RNA isolation. (A) Signal intensity for Ggt1 probes from normalized microarraydata. (B) qRT–PCR. Relative expression of Ggt1 with respect to 18S rRNA was calculated using the DDCt method. Data were normalized to the expression in control livers andare expressed as the means ± standard deviations. Statistically significant differences obtained by one-way ANOVA are indicated as ⁄⁄⁄⁄P < 0.0001 compared with NL and Negand as ⁄⁄P < 0.01 and ⁄P < 0.05 compared with NL.
Microdissection of GGT-tissue for gene expression / J.E. Torres Mena et al. / Anal. Biochem. 447 (2014) 126–132 131
types within the same cell type. This differentiation is critical incancer research, particularly in the study of pre-tumor lesions,which are distinguished from surrounding cells by tumor-specificmarkers. An important way to select phenotype-specific cells isby immunohistochemical staining of frozen sections; however,the standard protocols usually require several hours of immersionin aqueous media, resulting in activation of endogenous RNasesand RNA degradation [23,43]. RNases in liver are rapidly activatedonce tissue is hydrated; for example, RNA degradation occurs whentissue sections are incubated in the aqueous solutions used in theRutenburg protocol for GGT activity (Fig. 2A).
The main challenge when combining LCM and gene expressionanalyses using DNA microarrays was to obtain sufficient RNA ofadequate integrity [44]. Some studies reduced the time of stainingprotocols [23], and the inclusion of RNase inhibitors seems to bemandatory during the immersion of tissue in aqueous solutions[45]. We tested two RNase inhibitors during the optimization ofGGT histochemistry. Ambion RNAsecure, an inhibitor compatiblewith RT–PCR and in vitro transcription in Tris-buffered reactions,could protect RNA after tissue recovery. However, its inclusion af-fected the staining procedure through the formation of precipitatewith components of the reaction. DEPC was not compatible withTris or Hepes buffers [46]. However, when DEPC was combinedwith ethanol for fixation and with Mops buffer during reaction (Ta-ble 1), the GGT reaction and preservation of RNA was obtained. Thetime of reaction for visible GGT activity in tissue with the opti-mized method is comparable to or even shorter than the one re-ported by Rutenburg and coworkers (Table 1). To avoid possibleRNA degradation, the post-reaction procedures used in the Ruten-burg method, such as chelation with CuSO4 and counterstaining,were not implemented. Another important aspect in this optimiza-tion was the drying step of GGT-stained tissue. This step wasimportant for capturing the tissue with the laser-activated adhe-sive caps and for avoiding hydration of the tissue. However, we ob-served that the RNA integrity was affected after staining; a reduced28S/18S rRNA ratio was found 2 h after staining, and clear degrada-tion was observed after 4 h (Fig. 2A).
In addition, the majority of LCM systems prohibit the use ofmounting medium and coverslips for the laser-assisted dissection;thus, the tissue is exposed to rehydration, compromising the RNAquality. Indeed, laboratory humidity is suggested as a detrimentto RNA integrity during LCM [47]. RNA stabilization by supplying
an argon atmosphere seems to be a promising alternative to avoidhumidity [20]; however, we suggest that the drying step beforeLCM offers RNA stability up to 1 h, which is sufficient time forthe LCM procedure.
We propose that this optimization allows precise gene expres-sion analyses in tissues that contain GGT-positive cells, such aspancreas, kidney, liver, and fetal tissues, and pathologies, such ashuman neoplasias and atheromatous lesions. Because GGT is abroad marker of liver carcinogenesis [34], we obtained precisegene expression signatures in the nodules and tumors from the liv-ers of carcinogen-treated rats.
In conclusion, we optimized the method for histological demon-stration of GGT for gene expression analysis of LCM samples. Dueto the importance of GGT as a pathological marker, this procedurewill facilitate more accurate genome-associated studies related tothis enzyme.
Acknowledgments
We greatly appreciate the technical assistance of Karol CarrilloSánchez, Nayeli Belem Gabiño López, and Dan Jerzain GutiérrezFuentes from the high-technology units at INMEGEN. Thanks alsogo to Evelia Arce Popoca, Sergio Hernandez García, and Ruth Pach-eco for their technical assistance at CINVESTAV. This work wassupported by Fondo Sectorial de Investigación en Salud y Seguri-dad Social SSA/IMSS/ISSSTE-CONACYT SALUD-2009-01-115431and Institutional Project 77-10/2011/I.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.ab.2013.11.015.
References
[1] J.P. Ward, c-Glutamyl transpeptidase: a sensitive indicator of renal ischaemicinjury in experimental animals and renal homograft rejection in man, Ann. R.Coll. Surg. Engl. 57 (1975) 248–261.
[2] A.M. Rutenburg, H. Kim, J.W. Fischbein, J.S. Hanker, H.L. Wasserkrug,A.M. Seligman, Histochemical and ultrastructural demonstration ofc-glutamyl transpeptidase activity, J. Histochem. Cytochem. 17 (1969)517–526.

132 Microdissection of GGT-tissue for gene expression / J.E. Torres Mena et al. / Anal. Biochem. 447 (2014) 126–132
[3] R. Cameron, J. Kellen, A. Kolin, A. Malkin, E. Farber, c-Glutamyltransferase inputative premalignant liver cell populations during hepatocarcinogenesis,Cancer Res. 38 (1978) 823–829.
[4] S. Fiala, E.C. Trout Jr., C.A. Teague, E.S. Fiala, c-Glutamyltransferase, a commonmarker of human epithelial tumors?, Cancer Detect Prev. 3 (1980)471–485.
[5] M. Tsutsumi, D. Sakamuro, A. Takada, S.C. Zang, T. Furukawa, N. Taniguchi,Detection of a unique c-glutamyl transpeptidase messenger RNA speciesclosely related to the development of hepatocellular carcinoma in humans: anew candidate for early diagnosis of hepatocellular carcinoma, Hepatology 23(1996) 1093–1097.
[6] K. Arai, S.H. Sumi, K. Yoshida, T. Komoda, A precursor form of human kidney c-glutamyl transferase in normal and cancerous tissues, and its possible post-translational modification, Biochim. Biophys. Acta 1253 (1995) 33–38.
[7] S.L. Blair, P. Heerdt, S. Sachar, A. Abolhoda, S. Hochwald, H. Cheng, M. Burt,Glutathione metabolism in patients with non-small cell lung cancers, CancerRes. 57 (1997) 152–155.
[8] I. Pettersen, J.H. Andersen, K. Bjornland, O. Mathisen, R. Bremnes, M. Wellman,A. Visvikis, N.E. Huseby, Heterogeneity in c-glutamyltransferase mRNAexpression and glycan structures: search for tumor-specific variants inhuman liver metastases and colon carcinoma cells, Biochim. Biophys. Acta1648 (2003) 210–218.
[9] H.F. Frierson Jr., D. Theodorescu, S.E. Mills, M.H. Hanigan, c-Glutamyltranspeptidase in normal and neoplastic prostate glands, Mod. Pathol. 10(1997) 1–6.
[10] M.H. Hanigan, H.F. Frierson Jr., J.E. Brown, M.A. Lovell, P.T. Taylor, Humanovarian tumors express c-glutamyl transpeptidase, Cancer Res. 54 (1994)286–290.
[11] S.N. Hochwald, D.M. Rose, M.F. Brennan, M.E. Burt, Elevation of glutathioneand related enzyme activities in high-grade and metastatic extremity softtissue sarcoma, Ann. Surg. Oncol. 4 (1997) 303–309.
[12] J.R. Durham, H.F. Frierson Jr., M.H. Hanigan, c-Glutamyl transpeptidaseimmunoreactivity in benign and malignant breast tissue, Breast Cancer Res.Treat. 45 (1997) 55–62.
[13] M. Tager, A. Ittenson, A. Franke, A. Frey, H.G. Gassen, S. Ansorge, c-Glutamyltranspeptidase–cellular expression in populations of normal humanmononuclear cells and patients suffering from leukemias, Ann. Hematol. 70(1995) 237–242.
[14] E. Maellaro, S. Dominici, B. Del Bello, M.A. Valentini, L. Pieri, P. Perego, R.Supino, F. Zunino, E. Lorenzini, A. Paolicchi, M. Comporti, A. Pompella,Membrane c-glutamyl transpeptidase activity of melanoma cells: Effects oncellular H2O2 production, cell surface protein thiol oxidation, and NF-kBactivation status, J. Cell Sci. 113 (2000) 2671–2678.
[15] A. Corti, M. Franzini, A. Paolicchi, A. Pompella, c-Glutamyltransferase of cancercells at the crossroads of tumor progression, drug resistance, and drugtargeting, Anticancer Res. 30 (2010) 1169–1181.
[16] A. Pompella, A. Corti, A. Paolicchi, C. Giommarelli, F. Zunino, c-Glutamyltransferase, redox regulation, and cancer drug resistance, Curr.Opin. Pharmacol. 7 (2007) 360–366.
[17] A. Pompella, V. De Tata, A. Paolicchi, F. Zunino, Expression of c-glutamyltransferase in cancer cells and its significance in drug resistance,Biochem. Pharmacol. 71 (2006) 231–238.
[18] J.E. Mason, R.D. Starke, J.E. Van Kirk, c-Glutamyl transferase: a novelcardiovascular risk biomarker, Prev. Cardiol. 13 (2010) 36–41.
[19] K. Yoshida, K. Arai, N. Kobayashi, Y. Uchijima, H. Saitoh, Purification andproperties of c-glutamyl transpeptidase from the tissue of human benignprostatic hypertrophy, J. Urol. 146 (1991) 895–899.
[20] M. Clement-Ziza, A. Munnich, S. Lyonnet, F. Jaubert, C. Besmond, Stabilizationof RNA during laser capture microdissection by performing experiments underargon atmosphere or using ethanol as a solvent in staining solutions, RNA 14(2008) 2698–2704.
[21] L. Fink, T. Kinfe, M.M. Stein, L. Ermert, J. Hanze, W. Kummer, W. Seeger, R.M.Bohle, Immunostaining and laser-assisted cell picking for mRNA analysis, Lab.Invest. 80 (2000) 327–333.
[22] D.M. Kube, C.D. Savci-Heijink, A.F. Lamblin, F. Kosari, G. Vasmatzis, J.C.Cheville, D.P. Connelly, G.G. Klee, Optimization of laser capturemicrodissection and RNA amplification for gene expression profiling ofprostate cancer, BMC Mol. Biol. 8 (2007) 25.
[23] F. Fend, M.R. Emmert-Buck, R. Chuaqui, K. Cole, J. Lee, L.A. Liotta, M. Raffeld,Immuno-LCM: laser capture microdissection of immunostained frozensections for mRNA analysis, Am. J. Pathol. 154 (1999) 61–66.
[24] S.B. Sonne, M.D. Dalgaard, J.E. Nielsen, C.E. Hoei-Hansen, E. Rajpert-De Meyts,L.M. Gjerdrum, H. Leffers, Optimizing staining protocols for laser
microdissection of specific cell types from the testis including carcinomain situ, PLoS One 4 (2009) e5536.
[25] C.R. Vanderburg, M.S. Clarke, Laser capture microdissection ofmetachromatically stained skeletal muscle allows quantification of fibertype specific gene expression, Mol. Cell. Biochem. 375 (2013) 159–170.
[26] D. Ibberson, V. Benes, M.U. Muckenthaler, M. Castoldi, RNA degradationcompromises the reliability of microRNA expression profiling, BMC Biotechnol.9 (2009) 102.
[27] D.B. Solt, A. Medline, E. Farber, Rapid emergence of carcinogen-inducedhyperplastic lesions in a new model for the sequential analysis of livercarcinogenesis, Am. J. Pathol. 88 (1977) 595–618.
[28] A. Marche-Cova, S. Fattel-Fazenda, A. Rojas-Ochoa, E. Arce-Popoca, S. Villa-Trevino, Follow-up of GST-P during hepatocarcinogenesis with DEN–2AAF inF344 rats, Arch. Med. Res. 26 (1995) S169–S173.
[29] J.I. Perez-Carreon, C. Lopez-Garcia, S. Fattel-Fazenda, E. Arce-Popoca, L.Aleman-Lazarini, S. Hernandez-Garcia, V. Le Berre, S. Sokol, J.M. Francois, S.Villa-Trevino, Gene expression profile related to the progression ofpreneoplastic nodules toward hepatocellular carcinoma in rats, Neoplasia 8(2006) 373–383.
[30] T.J. Collins, ImageJ for microscopy, BioTechniques 43 (2007) 25–30.[31] L.M. Eijssen, M. Jaillard, M.E. Adriaens, S. Gaj, P.J. de Groot, M. Muller, C.T.
Evelo, User-friendly solutions for microarray quality control and pre-processing on ArrayAnalysis.org, Nucleic Acids Res. 41 (2013) W71–W76.
[32] K. Enomoto, E. Farber, Kinetics of phenotypic maturation of remodeling ofhyperplastic nodules during liver carcinogenesis, Cancer Res. 42 (1982) 2330–2335.
[33] R.P. Mazzantini, A. de Conti, F.S. Moreno, Persistent and remodeling hepaticpreneoplastic lesions present differences in cell proliferation and apoptosis, aswell as in p53, Bcl-2, and NF-kB pathways, J. Cell Biochem. 103 (2008) 538–546.
[34] M.H. Hanigan, H.C. Pitot, c-Glutamyl transpeptidase—its role inhepatocarcinogenesis, Carcinogenesis 6 (1985) 165–172.
[35] K.J. Archer, C.I. Dumur, S.E. Joel, V. Ramakrishnan, Assessing quality ofhybridized RNA in Affymetrix GeneChip experiments using mixed-effectsmodels, Biostatistics 7 (2006) 198–212.
[36] Z. Albert, Z. Rzucidlo, H. Starzyk, Biochemical and histochemical investigationsof the c-glutamyl transpeptidase in embryonal and adult organs of man, ActaHistochem. 37 (1970) 74–79.
[37] N. Fleming, P. Groscurth, G.S. Kistler, The activity and distribution of c-glutamyl transpeptidase (c-GT) in human foetal organs, Histochemistry 51(1977) 209–218.
[38] M.H. Hanigan, H.F. Frierson Jr., P.E. Swanson, B.R. De Young, Altered expressionof c-glutamyl transpeptidase in human tumors, Hum. Pathol. 30 (1999) 300–305.
[39] M. Vanderlaan, W. Phares, c-Glutamyltranspeptidase: a tumour cell markerwith a pharmacological function, Histochem. J. 13 (1981) 865–877.
[40] M. Morrogh, N. Olvera, F. Bogomolniy, P.I. Borgen, T.A. King, N. Olvera, Tissuepreparation for laser capture microdissection and RNA extraction from freshfrozen breast tissue, BioTechniques 43 (2007). 41–42, 44, 46, passim.
[41] L. Luo, R.C. Salunga, H. Guo, A. Bittner, K.C. Joy, J.E. Galindo, H. Xiao, K.E. Rogers,J.S. Wan, M.R. Jackson, M.G. Erlander, Gene expression profiles of laser-captured adjacent neuronal subtypes, Nat. Med. 5 (1999) 117–122.
[42] C. Michel, C. Desdouets, B. Sacre-Salem, J.C. Gautier, R. Roberts, E. Boitier, Livergene expression profiles of rats treated with clofibric acid: comparison ofwhole liver and laser capture microdissected liver, Am. J. Pathol. 163 (2003)2191–2199.
[43] S. Suzuki, M. Asamoto, K. Tsujimura, T. Shirai, Specific differences in geneexpression profile revealed by cDNA microarray analysis of glutathione S-transferase placental form (GST-P) immunohistochemically positive rat liverfoci and surrounding tissue, Carcinogenesis 25 (2004) 439–443.
[44] W.Z. Wang, F.M. Oeschger, S. Lee, Z. Molnar, High quality RNA from multiplebrain regions simultaneously acquired by laser capture microdissection, BMCMol. Biol. 10 (2009) 69.
[45] R.J. Buckanovich, D. Sasaroli, A. O’Brien-Jenkins, J. Botbyl, J.R. Conejo-Garcia, F.Benencia, L.A. Liotta, P.A. Gimotty, G. Coukos, Use of immuno-LCM to identifythe in situ expression profile of cellular constituents of the tumormicroenvironment, Cancer Biol. Ther. 5 (2006) 635–642.
[46] S.L. Berger, Diethyl pyrocarbonate: an examination of its properties in bufferedsolutions with a new assay technique, Anal. Biochem. 67 (1975) 428–437.
[47] G.A. Ordway, A. Szebeni, M.M. Duffourc, S. Dessus-Babus, K. Szebeni, Geneexpression analyses of neurons, astrocytes, and oligodendrocytes isolated bylaser capture microdissection from human brain: detrimental effects oflaboratory humidity, J. Neurosci. Res. 87 (2009) 2430–2438.