Embed Size (px)
Citation preview

© 2007 by Russia, Protistology
Protistology 4 (4), 339�345 (2006/7) ProtistologyProtistologyProtistologyProtistologyProtistology
Lagenophrys lenticula and L. patina (Peritricha),epibionts of Hyalella azteca (Amphipoda). A studyusing scanning electron microscopy to reveal detailsof the lorica aperture
R. Mayun�Estrada and M. A. Aladro�Lubel
Laboratorio de Protozoolog|a, Depto. Biolog|a Comparada, Fac. Ciencias,
Universidad Nacional Autsnoma de Muxico
Summary
The peritrich ciliates Lagenophrys lenticula and L. patina are epibionts of the amphipod
Hyalella azteca, from Pwtzcuaro Lake, Michoacwn, Mexico. The two species were
studied with scanning electron microscopy, enabling fine characterization of the lips
of the lorica aperture and complete description of each species. Prevalence and
distribution of each lagenophryid species on the basibiont body and their
biogeographical record are updated.
Key words: lorica aperture, lagenophryids, ciliated epibionts, amphipods
Introduction
Members of the genus Lagenophrys are ciliated
protozoa adapted to living as epibionts on crustaceans.
Species of Lagenophrys spend most of their life cycle
attached to their host's exoskeleton by means of the
ventral surface of their distinctive lorica or, in some
cases, by means of a pseudostalk formed from a
narrowed region of the lorica. The free�swimming phase
of the life cycle of Lagenophrys is the migratory larva,
or telotroch, that is characteristic of all sessiline
peritrichs. It is ephemeral, having the function of
seeking out and settling on a new host once it is released.
Extra telotrochs are produced when the host undergoes
ecdysis, as part of the process of abandoning the loricae
on the host's old exoskeleton. Some characters that
define species of Lagenophrys are the structure of the
lorica and the lips of the lorica aperture.
To date, 62 species of Lagenophrys have been
described (Clamp and Kane, 2003), of which 26 use
marine and freshwater amphipods as substrates. Only
10 species of this genus have been studied with scanning
or transmission electron microscopy (Couch, 1973;
Felgenhauer, 1979, 1982; Schadel, 1983, 1985, 1986;
Walker et al., 1986; Clamp, 1988; Claps and Sampsns,
1994; Veltkamp et al., 1994; Roberts and Chubb, 1998;

· R. Mayun�Estrada and M. A. Aladro�Lubel340
Mayun�Estrada and Aladro�Lubel, 2000). Of these
studies, only four have focused on species of Lageno�phrys which attach to amphipods. Two species, L.lenticula (Kellicott, 1885) and L. patina Stokes, 1887,
have been reported as associates of freshwater amphi�
pods. Only optical microscopy has been used to describe
the morphology of L. lenticula (Felgenhauer, 1979).
The structure of the lips of the lorica aperture is a key
diagnostic feature used to identify species of Lageno�phrys. The purpose of this paper is to add new
information that will improve diagnostic features of
these species with emphasis on the details of the lips as
revealed by scanning electron microscopy. In addition,
morphometric data, biogeographical record and
distribution on the host's body are provided.
Material and Methods
The crustacean amphipod Hyalella azteca was
collected from Pwtzcuaro Lake, Michoacwn, Mexico
with the aid of a 5 mm�mesh net over a five month
period through the years 2000�2002 (January and
November, 2000; February and June, 2001, and
February 2002). In order to observe living ciliates
attached to the exoskeleton, amphipods were mainta�
ined in laboratory aquaria at room temperature, using
unfiltered water from the lake with submerged plants
added. To record distribution on the host's body,
preferential habitat, and prevalence, each amphipod
was dissected and separated into 12 units: head (H),
antennae (A), mouth parts (Mp), gnathopods (G),
coxae (C), pereionites (Pe), pleonites (Pn), uronites
(Un), pereiopods (P), pleopods (Pl), uropods (U) and
telson (T). Each part was fixed with 5% formaldehyde
and stained or impregnated with Harris hematoxylin
and protargol in order to reveal the cytological
characters of epibionts. For scanning electron micro�
scopy, material was fixed in 1% glutaraldehyde,
transferred to 2.5% glutaraldehyde in 0.1 M sodium
cacodylate buffer (pH 7.2), critical point dried, and
coated with carbon and gold. Morphometric data of
lagenophryid peritrich cilates were recorded, and the
mean, maximum, minimum and standard deviation
calculated.
Results
MORPHOLOGICAL ASPECTS
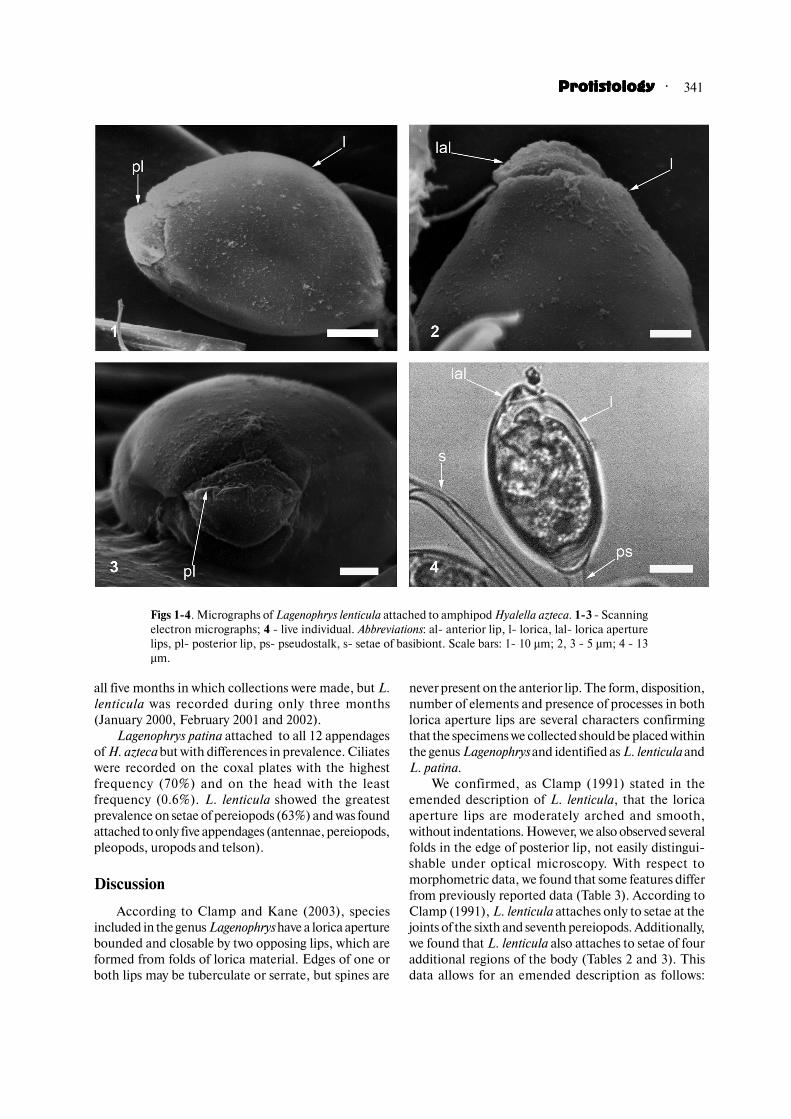
Morphometric data of both species of Lagenophrysare shown in Table 1. Details and disposition of both
lips bordering the lorica aperture in L. lenticula are
shown in figures 1�4. The lips were observed only in the
closed position. Viewed laterally, the posterior lip
showed only fine projections (Fig. 2), but seen from
above (Fig. 3), projections were irregularly disposed and
up to eight in number, with a folded appearance. These
folds were not visible by optical microscopy (Fig. 4).
Both lips of L. patina are shown in figures 5�10,
with the loricastome open to different degrees. The
anterior lip carried from 12 to 17 indentations (Fig. 7),
but specimens with few (Fig. 6) or no teeth (Figs. 5 and
10) were also observed. Teeth were observed across the
entire edge. The posterior lip always had 7 to 18 teeth
(Figs 5�10) distributed along its edge. In some
specimens, these teeth were different in size, but in
others, all were of the same size. Teeth of both lips were
separated from one another by grooves or folds that
penetrated the lips to different depths. In the case of
the posterior lip, this can be seen easily with optical
microscopy when the loricastome is open (Fig. 10).
Observation of teeth on the anterior lip is difficult with
optical microscopy, either because of the shallow depth
of the grooves that delimits each one or because the teeth
are disposed on the inner face of the loricastome.
DISTRIBUTION AND PREVALENCE
Altogether, 584 individuals of Hyalella azteca were
collected, of which 67.6% had Lagenophrys patina and
10.9% had L. lenticula attached to the exoskeleton
(Table 2). L. patina was recorded on amphipods during
Table 1. Morphometric data of Lagenophrys lenticula and L. patina epibionts of Hyalella azteca.
Characters Attributes Lorica
length Lorica width Lips of
lorica Pseudostalk
length Pseudostalk
width Zooid length
Zooid width
Macronucleus length
Macronucleus width
L. lenticula Average 47.7 31.2 14.3 14.3 4.2 31.4 24.5 22.9 4.3 Minimum 39.2 22.4 11.2 8.4 2.8 22.4 19.6 16.8 2.8 Maximum 66.6 44.8 16.8 29.6 7.4 51.8 37.0 30.8 5.6 SD 4.6 4.4 1.6 3.7 1.4 5.0 3.6 3.1 1.3 N 90 90 53 43 43 55 55 31 31
L. patina Average 57.9 53.9 14.4 � � 46.6 37.9 29.3 5.5 Minimum 42.0 36.4 14.0 � � 33.6 28.0 19.6 2.8 Maximum 81.4 74.0 16.8 � � 66.6 52.2 36.5 11.2 SD 5.2 5.3 1.0 � � 13.4 5.3 3.3 1.3 N 150 150 105 � � 104 104 101 101
SD = Standard deviation; N= number of individuals. Measurements in µm.
· 341ProtistologyProtistologyProtistologyProtistologyProtistology
all five months in which collections were made, but L.lenticula was recorded during only three months
(January 2000, February 2001 and 2002).
Lagenophrys patina attached to all 12 appendages
of H. azteca but with differences in prevalence. Ciliates
were recorded on the coxal plates with the highest
frequency (70%) and on the head with the least
frequency (0.6%). L. lenticula showed the greatest
prevalence on setae of pereiopods (63%) and was found
attached to only five appendages (antennae, pereiopods,
pleopods, uropods and telson).
Discussion
According to Clamp and Kane (2003), species
included in the genus Lagenophrys have a lorica aperture
bounded and closable by two opposing lips, which are
formed from folds of lorica material. Edges of one or
both lips may be tuberculate or serrate, but spines are
never present on the anterior lip. The form, disposition,
number of elements and presence of processes in both
lorica aperture lips are several characters confirming
that the specimens we collected should be placed within
the genus Lagenophrys and identified as L. lenticula and
L. patina.
We confirmed, as Clamp (1991) stated in the
emended description of L. lenticula, that the lorica
aperture lips are moderately arched and smooth,
without indentations. However, we also observed several
folds in the edge of posterior lip, not easily distingui�
shable under optical microscopy. With respect to
morphometric data, we found that some features differ
from previously reported data (Table 3). According to
Clamp (1991), L. lenticula attaches only to setae at the
joints of the sixth and seventh pereiopods. Additionally,
we found that L. lenticula also attaches to setae of four
additional regions of the body (Tables 2 and 3). This
data allows for an emended description as follows:
Figs 1�4. Micrographs of Lagenophrys lenticula attached to amphipod Hyalella azteca. 1�3 � Scanning
electron micrographs; 4 � live individual. Abbreviations: al� anterior lip, l� lorica, lal� lorica aperture
lips, pl� posterior lip, ps� pseudostalk, s� setae of basibiont. Scale bars: 1� 10 µm; 2, 3 � 5 µm; 4 � 13
µm.

· R. Mayun�Estrada and M. A. Aladro�Lubel342
anterior lip smooth; posterior lip with slight folds on its
edge. The epibiont attaches to setae of several appendages.
For L. patina, Clamp (1990) reported morpholo�
gical variability of the lips. Clamp (1990) observed that
the edge of anterior lip could be both smooth and with
many shallow evenly spaced indentations, and that only
the medial three�fifths in the edge of posterior lip had
several large blunt tooth�like projections, separated by
shallow notches, which were absent in some individuals.
In describing L. patina (misidentified as L. labiata),
Felgenhauer (1979) documented that the borders of
both lips were crenated and irregular. We assume that
these observations correspond to the teeth. We confirm
variation of the two aperture lips of L. patina,
exemplified in this case among individuals that were
attached to hosts from a population of H. azteca in
Michoacwn, Mexico. On both lips we observed the
presence of variable number of teeth. Lips of L. patinavary as follows: the anterior lip was either smooth or
with indentations (number of teeth ranged from 12 to
17), and each one was delimited by grooves on the inner
face of the loricastome. The posterior lip showed 9�18
teeth, whose size increased from the lateral edge to the
medial edge. We also found some morphometric
difference between our samples of L. patina and
published data (Table 4).
The exact number of teeth is not specified in
original descriptions of L. patina, and the degree of
depth in the grooves between teeth is not shown;
therefore, an emended description of L. patina is as
follows:
Anterior lip of loricastome either smooth or with
12�17 teeth, which can be disposed at the internal face
of the lip; posterior lip always with 9�18 teeth. On both
lips, teeth are distributed along the edge. The teeth are
delimited by grooves that extend from the edge to the
basal area of the lips.
Lagenophrys patina and L. lenticula have been
recorded from the continental United States, Canada,
Mexico (Puebla), and Uruguay (Clamp, 1990). L.lenticula was also reported from Mexico (Hidalgo state)
associated with H. azteca (Aladro�Lubel and Swnchez�
Caldersn, 2005; Aladro�Lubel et al., 2006). According
to proposed biogeographical schemes (Morrone et al.,
2002; Morrone, 2005), previous records and the present
study place the two species from the Nearctic Region
to the Neotropical Region and, for Mexico, in the
Mexican Transition Zone at the Transmexican Volcanic
Belt Province.
Scanning electron microscopy study of Lageno�phrys species is very important for precise description
of the species�level characters. Intra� and interspecific
variability with respect to characteristics of the lips of
the lorica aperture exists in some members of the genus
Lagenophrys, and several terms are used to describe any
Ta
ble
2.
Occ
urr
en
ce a
nd
dis
trib
utio
n o
f sp
ec
ies
of L
ag
en
op
hry
s in
the
am
ph
ipo
d H
. a
zte
ca
fro
m P
atzc
uar
o L
ake
, Mic
ho
acan
, M
exi
co
.
Dat
e N
N
H
Sp
eci
es
H
A
Mp
G
C
P
e P
n
Un
P
P
l U
T
Jan
200
0 69
44
47
L.
lent
icul
a L.
pat
ina
0 0 0
2 (
2.8
) 0
0
0
0
0
46
(66.
6)
0 7
(10
.0)
0
3 (
4.3
) 0
7
(10.
0)
44
(63.
0)
7 (1
0.0
) 0
3
(4
.3)
11 (
16.0
) 1
(1
.4)
8 (
11.6
) 0
No
v 2
000
42
0 11
L. le
ntic
ula
L. p
atin
a 0 0
0 1
(2.
3)
0
0
0
0
0 1
0 (2
4.0)
0
1 (
2.3
) 0
1
(2
.3)
0
2 (
4.8
) 0
4
(9
.5)
0
1 (
2.3
) 0
2 (
4.8
) 0 0
Feb
200
1 1
64
13
85
L. le
ntic
ula
L. p
atin
a 0 0
0 0 0
0
0
0
0
76
(46.
0)
0 8
(4.
8)
0
22
(13.
0)
0
16
(10.
0)
13
(8
.0)
15
(9
.0)
0
0
1 (
0.6
) 3
(1
.8)
0 0 Ju
n 2
001
134
0
122
L.
lent
icul
a L.
pat
ina
0 0 0
12
(9
.0)
0
0
0
1 (0
.7)
0 9
2 (6
9.0)
0
28 (
21.0
) 0
5
1 (3
8.0
) 0
6
2 (4
6.0
) 0
4
7 (3
5.0
) 0
0
0
10
(7
.8)
0 6
(4
.4)
Feb
200
2 1
75
7 1
30
L. le
ntic
ula
L. p
atin
a 0
1 (
0.6
) 2
(1.
1)
7 (
4.0
) 0
5
(2.8
) 0
10
(5.
8)
0 122
(70.
0)
0 20
(11
.0)
0
27
(15.
0)
0
32
(18.
0)
6 (
3.4
) 3
7 (2
1.0
) 1
(0
.6)
2 (
1.1
) 0
9 (
5.1
) 0
3 (
1.8
)
Ja
n=
Jan
ua
ry, N
ov=
Nov
em
ber
, Fe
b=
Feb
ruar
y, J
un=
Ju
ne, N
= n
umb
er
tota
l of h
ost
s in
the
sam
ple
, NH
= n
umb
er o
f ho
sts
with
lag
enop
hry
ids.
The
dat
a co
rres
pon
ds
resp
ect
ivel
y to
num
be
r an
d
fre
que
ncy
% (
in p
are
nth
esis
) of
hos
ts th
at h
arb
ore
d e
pib
ion
ts. H
= h
ead
, A=
ant
enn
ae,
Mp
= m
outh
pa
rts,
G=
gna
thop
od
s, C
= c
oxae
, Pe=
pe
reio
nite
s, P
n=
ple
oni
tes,
Un
= u
roni
tes,
P=
per
eio
pod
s,
Pl=
ple
opo
ds,
U=
uro
pod
s, T
= t
els
on.

· 343ProtistologyProtistologyProtistologyProtistologyProtistology
kind of protuberance in the lorica aperture lips (rounded
tubercles, folds or grooves that extend from the edge to
the basal area of the lip, indentations and/or notches).
Therefore, we propose that species be categorized into
the following general groups: a) species with both lips
smooth, b) species with one of the two lips with some
kind of processes, and c) species with both lips carrying
some type of processes. The new arrangement proposed
here would be useful for all kinds of studies of Lage�nophrys species.
Figs 5�10. Micrographs of Lagenophrys patina attached to Hyalella azteca. 5�9 � Scanning electron
micrographs; 10 � micrograph of empty lorica of L. patina seen by light microscopy. Abbreviations: c�
collar, lr� lorica rim, t� teeth; for explanation of other symbols see figures 1�4. Scale bars: 5 � 10 µm; 6�
8 � 5 µm; 9 � 2 µm; 10 � 13 µm.

· R. Mayun�Estrada and M. A. Aladro�Lubel344
ACKNOWLEDGMENTS
We are very grateful to Bisl. Armando Zepeda R.
(Lab. Microscop|a Electrsnica, Facultad de Medicina,
UNAM), who kindly processed all SEM material and
to Bisl. Francisco Pasos (Depto. Biolog|a Celular y
Tisular, Facultad de Medicina, UNAM) for technical
assistance with photographs. We would like to thank Dr.
T. Nyborg (Department of Earth and Biological
Sciences, Loma Linda University) and Dr. F. Vega
(Instituto de Geolog|a, UNAM) for help with English
grammar and expression.
References
Aladro�Lubel M.A. and Swnchez�Caldersn G.
2005. Ciliados epibiontes de la vegetacisn sumergida y
de los invertebrados de la Laguna de Tecocomulco. In:
La Laguna de Tecocomulco. Geo�Ecolog|a de un
Desastre. (Eds. Huizar A.R., Jimunez F. E. and Juwrez
C.). Publ. Esp. 3. I. Geolog|a, UNAM. pp. 169�178.
Aladro�Lubel M. A., Mayun�Estrada R. and
Reyes�Santos M. 2006. Registro actualizado de ciliados
(Agosto, 2004). Listados faunisticos de Muxico. XI.
Inst. Biol. UNAM.
Clamp J.C. 1988. A new species of Lagenophrys(Ciliophora: Peritrichia: Lagenophryidae) ectocom�
mensal on the wood�boring isopod Limnoria (Flabel�
lifera: Limnoridae). Trans. Am. Micros. Soc. 107, 12�
16.
Clamp J.C. 1990. Redescription of three species
of Lagenophrys (Ciliophora: Peritricha: Lagenophry�
idae) and a new North American species of Lageno�phrys from hypogean amphipods. Trans. Am. Microsc.
Soc. 109, 1�31.
Clamp J.C. 1991. Revision of the family Lageno�
phryidae Bztschli, 1889 and description of the family
Usconophryidae n. fam. (Ciliophora, Peritricha). J.
Protozool. 38, 4, 355�377.
Clamp J.C. and Kane J.R. 2003. Redescription of
four species of lagenophryid peritrichs (Ciliophora)
from Australia and New Guinea, with descriptions of
two new species. Rec. Aust. Mus. 55, 153�168.
Claps M.C. and Sampons M.R. 1994. First record
of Lagenophrys discoidea Kellicott (Ciliophora,
Peritrichia, Lagenophryidae), ectocommensal of
Argentinian ostracods. Iheringia, Ser. Zool. Porto
Alegre. 76, 167�170.
Couch J.A. 1973. Ultrastructural and protargol
studies of Lagenophrys callinectes (Ciliophora: Peritri�
chida). J. Protozool. 20, 638�647.
Felgenhauer B.E. 1979. A note on the scanning
electron microscopy and hosts of the widespread
peritrich ciliate Lagenophrys labiata. Trans. Amer.
Micros. Soc. 98, 4, 591�595.
Felgenhauer B.E. 1982. A new species of Lageno�phrys (Ciliophora: Peritrichida) from the freshwater
shrimp Palaemonetes kadiakensis. Trans. Am. Microsc.
Soc. 101, 2, 142�150.
Mayun�Estrada R. and Aladro�Lubel M.A. 2000.
First record of Lagenophrys dennisi (Ciliophora:
Peritrichia) on the exoskeleton of crayfish Cambarelluspatzcuarensis. J. Eukaryot. Microbiol. 47, 57�61.
Morrone J.J. 2005. Biogeographic areas and
transition zones of Latin America and the Caribbean
islands based on pangeobiographic and cladistic
analyses of the entomophauna. Ann. Rev. Entomol. 51,
467�494.
Morrone J.J., Espinosa D. and Llorente J. 2002.
Mexican biogeographic provinces: preliminary scheme,
general characterizations and synonymies. Acta Zool.
Mex. (n. s.). 85, 83�108.
Table 3. Morphometric comparison of Lagenophrys lenticula (Kellicott, 1885) specimens from the present study with
previously described ones. Characters Clamp (1991) Present study Lorica length 44.7�56.6 39.2�66.6 Lorica width 34.9�47.9 22.4�44.8 Lips of lorica 16.0�18.9 11.2�16.8 Pseudostalk length 11.1�16.6 8.4�29.6 Pseudostalk width �� 2.8�7.4 Zooid length �� 22.4�51.8 Zooid width �� 19.6�37.0 Macronucleus length �� 16.8�30.8 Macronucleus width �� 2.8�5.6 Edge of posterior lip �� Up to eight slight folds
irregularly disposed Host appendages Setae at the joints
of the host's sixth and seventh pereiopods
Setae of antennae, pereiopods, pleopods,
uropods and telson
Morphometric data in µm; the values corresponds to minimum and maximum.
Table 4. Morphometric comparison of Lagenophrys patina Stokes, 1887 specimens obtained in the present study with
previously described ones. Characters Clamp 1990 Present study Lorica length 45.6�74.9 42.0�81.4 Lorica width 43.7�77.6 36.4�74.0 Lips of lorica 15.4�23.6 14.0�16.8 Number of teeth on anterior lip
Smooth, occasionally with several to many
shallow evenly spaced indentations
in edge
Smooth or with 12 to 17 teeth across the
entire edge, delimited by grooves
Number of teeth on posterior lip
Medial three�fifths of edge with several
large, blunt tooth like projections separated
by shallow notches
7�18 teeth distributed along its edge,
delimited by grooves
Zooid length �� 33.6�66.6 Zooid width �� 28.0�52.2 Macronucleus length
9.2�48.0 19.6�36.5
Macronucleus width
2.6�9.7 2.8�11.2
Host appendages All surfaces of body 12 appendages Morphometric data in µm; the values corresponds to minimum and maximum.

· 345ProtistologyProtistologyProtistologyProtistologyProtistology
Address for correspondence: R. Mayun�Estrada. Ap. Postal 70�374, C.P. 04510, Ciudad Universitaria, Muxico,
D.F. Mexico. E�mail: [email protected]
Editorial responsibility: Mark Farmer
Roberts G.N. and Chubb J.C. 1998. The distribution
and location of the symbiont Lagenophrys aselli on the
freshwater isopod Asellus aquaticus. Freshwater Biol.
40, 671�677.
Scha del H. 1983. Three new peritricha on
Gammaridae. Arch. Protistenk. 127, 115�126.
Scha del H. 1985. Epizoische Einzeller auf
Flohkrebsen. 2. Besiedler der Gammaridenbeine.
Mikrokosmos. 74, 269�273.
Schadel H. 1986. Sesshapte Wimpertiere auf
Wasserasseln. Mikrokosmos. 75, 293�301.
Veltkamp C.J., Chubb J.C., Birch S.P. and Eaton
J.W. 1994. A simple freeze dehydration method for
studying epiphytic and epizoic communities using the
scanning electron microscope. Hydrobiologia. 288, 33�
38.
Walker M.H., Roberts E.M. and Usher M.L. 1986.
The fine structure of the trophont and stages in telotroch
formation in Circolagenophrys ampulla (Ciliophora,
Peritrichida). J. Protozool. 33, 2, 246�255.





























