Embed Size (px)
Citation preview

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers,academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research.
Laboratory Selection of Resistance to Spinosad in Culexquinquefasciatus (Diptera: Culicidae)Author(s): Tianyun Su and Min-Lee ChengSource: Journal of Medical Entomology, 51(2):421-427. 2014.Published By: Entomological Society of AmericaURL: http://www.bioone.org/doi/full/10.1603/ME13173
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in thebiological, ecological, and environmental sciences. BioOne provides a sustainable onlineplatform for over 170 journals and books published by nonprofit societies, associations,museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicatesyour acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercialuse. Commercial inquiries or rights and permissions requests should be directed to theindividual publisher as copyright holder.

VECTOR CONTROL, PEST MANAGEMENT, RESISTANCE, REPELLENTS
Laboratory Selection of Resistance to Spinosad inCulex quinquefasciatus (Diptera: Culicidae)
TIANYUN SU1 AND MIN-LEE CHENG
West Valley Mosquito and Vector Control District, 1295 E. Locust Street, Ontario, CA 91761
J. Med. Entomol. 51(2): 421Ð427 (2014); DOI: http://dx.doi.org/10.1603/ME13173
ABSTRACT A southern house mosquito Culex quinquefasciatus Say colony was established fromsurviving late instars and pupae from a semiÞeld evaluation on Natular XRG (a granular formulationcontaining 2.5% spinosad). The initial lethal levels of Natular XRG against this colony were determinedin the laboratory for the Þrst-generation progeny (designated as F1). Selection pressure was appliedat LC70Ð90 levels to 10,000Ð15,000 late third- and early fourth-instar larvae of each generation withNatular XRG. Susceptibility changes in response to selection were determined every other generation,where a gradual and steady decline in susceptibility occurred from generation F1 to F35, followed bysigniÞcant decline from generations F37 to F45. For reference purposes, susceptibility of freshlycollected wild populations as well as a laboratory colony of the same species was also determinedconcurrently, which ßuctuated within a slightly wider range for the wild populations and a tighterrange for the laboratory colony. By comparing with wild populations and laboratory reference colony,tolerance to spinosad was observed from generations up to F9 in the selected population. Resistancelevels increased gradually from generation F11 to F35, and elevated signiÞcantly from generations F37
to F45, when resistance ratios reached 1,415.3- to 2,229.9-fold at LC50 and 9,613.1- to 17,062.6-fold atLC90. Possible mechanisms of resistance development to spinosad were discussed.
KEY WORDS Culex quinquefasciatus, Diptera, Culicidae, spinosad, resistance development
Spinosad is produced by a naturally occurring soil-dwelling bacterium, Saccharopolyspora spinosaMertzand Yao (Mertz and Yao 1990). It consists of spinosynA (C41H65NO10) and spinosyn D (C42H67NO10). As anew class of polyketide-macrolide insecticides, spi-nosad has pesticidal activity after ingestion and cuticleabsorption against a broad spectrum of susceptibleinsect species, acting at the nicotinic acetylcholinereceptors (nAChR) and the �-aminobutyric acid re-ceptors and causing rapid excitation of the insect ner-vous system (Salgado et al. 1998, Thompson et al.2000). Spinosad and spinetoram (spinosyn J and L) arecategorized as group 5 pesticidesÑnAChR allostericmodulators, by the Insecticide Resistance ActingCommittee. The U.S. Environmental ProtectionAgency considers spinosad as a reduced-risk prod-uct because it has minimal or lacks effect on mostnontargets, plants, and the environment at the labelrates. After spinosad formulations are repeatedly usedin the control of agricultural and green-house pests ona variety of crops, development of resistance andcross-resistance has been reported recently (seeSparks et al. 2012, Geng et al. 2013). Low to very highlevels of resistance have been documented in re-sponse to laboratory selections and Þeld applicationsin Diptera: the house ßy Musca domestica L. (Shono
and Scott 2003); the oriental fruit ßy Bactrocera dor-salis (Hendel) (Hsu and Feng 2006), the olive fruit ßyBactrocera oleae (Rossi) (Kakani et al. 2010), and theAmerican serpentine leafminer Liriomyza trifolii(Burgess) (Ferguson 2004); Lepidoptera: the tobaccobudworm Heliothis virescens (F.) (Wyss et al. 2003,Young et al. 2003), the cotton bollworm Helicoverpaarmigeria (Hubner) (Kranthi et al. 2000), the dia-mondback moth Plutella xylostella (L.) (Zhao et al.2002, 2006; Sayyed et al. 2004), the beet armywormSpodoptera exigua (Hubner) (Moulton et al. 2000,Wang et al. 2006), and obliquebanded leafroller Cho-ristoneura rosaceana (Harris) (Sial et al. 2011); Thy-sanoptera: the western ßower thrips, Frankliniella oc-cidentalis (Pergande) (Bielza et al. 2007); andHymenoptera: the braconid wasps Cotesia plutellae(Kurdjumov) (Liu et al. 2007).
Hertlein et al. (2010) recently reviewed the poten-tial use of spinosad-based formulations as naturalproducts to control mosquito larvae. Several spinosadformulations under the trade name Natular, rangingfrom fast release emulsiÞable concentrate and gran-ules to slow release tablets and briquets, have beenregistered for mosquito control in the United Statesand elsewhere (Clarke, Roselle, IL). These productsare expected to play important roles together withother biorational larvicides for the upcoming decades.To maximize the Þeld efÞcacy and product longevity,1 Corresponding author, e-mail: [email protected].
0022-2585/14/0421Ð0427$04.00/0 � 2014 Entomological Society of America

resistance management strategies must be developedand implemented. Considering the history of resis-tance development in agricultural and green-housepests to spinosad, studies were initiated to exploreresistance development inCulex quiquefasciatusSay inresponse to intensive selections in the laboratory.
Materials and Methods
Spinosad. The pesticide used in this study was sandgranules with 2.5% spinosad (mixture of spinosyn Aand D; Lot# 10062307, Clarke, Roselle, IL) under thetrade name Natular XRG.Mosquitoes.A southern house mosquitoCulex quin-quefasciatusSay colony was established from survivingfourth-instar larvae and pupae collected from a semi-Þeld microcosm test with Natular XRG applied at 14.2kg/ha, the mid dose within the label range of 5.7Ð22.5kg/ha. The initial LC50 and LC90 values against Natu-lar XRG were determined in the laboratory for theÞrst-generation progeny (designated as F1) resultedfrom surviving larvae and pupae. This population wasthereafter subjected to continuous selection (see be-low) for resistance development (Su and Cheng2012).Localwildpopulationsof the samespeciesweresampled by collection of egg rafts using organic infu-sion (2.5% rabbit chow pellets fermented for 2 wk)(West Valley Mosquito and Vector Control District,unpublished data) as an oviposition attractant fromthe same location where the early semiÞeld micro-cosm evaluation was conducted. No Þeld applicationsof any spinosad formulations were made on-site or inthe adjacent areas before and during the study period.The egg collection and larval culture were synchro-nized with the spinosad-selected population, so thatbioassays were able to proceed concurrently. At eachbioassay time (i.e., every other generation of the spi-nosad-selected population), in total 10Ð15 egg raftswere collected and resultingCx. quiquefasciatus larvaewere used in the bioassays. A spinosad-naõve labora-tory colony of Cx. quinquefasciatus, which originatedfrom the multisite collections in 2007 from the south-west portion of San Bernardino County, CA, was usedas a reference population. This colony was also bio-assayed concurrently with spinosad-selected andfreshly collected wild populations to further validateresistance development.Selection. Selection was applied after initial lethal
levels were determined against Natular XRG for F1
generation resulted from surviving larvae and pupaefrom the semiÞeld microcosm test. The selection pres-sure was at LC70Ð90 levels. At each generation, 10Ð15plastic pans (30 cm in length by 16 cm in width by 10cm in depth) each containing �2,000 ml of rearingwater (with cumulated debris, uneaten larval food,and sediments) and 1,000 late third- and early fourth-instar larvae were used for selection treatment. Natu-lar XRG was suspended in tap water at 1 or 10%depending on the estimated resistance level in thepopulation to be treated. Appropriate amounts ofNatular XRG aqueous suspension were added to thepans to cause 70Ð90% larval mortality. Larvae were
continuously exposed to spinosad until they pupated.Surviving pupae were collected and used to propagatethe next generation. Doses for selection were elevatedgradually from generation to generation according tosusceptibility decline in response to successive selec-tions.Bioassay.Bioassays were conducted according to Su
and Mulla (2004). Brießy, four concentrations of testformulation within the dosage range that generated�5Ð95% larval mortality were used in a bioassay withthree replicates at each concentration. For each rep-licate, 25 late third-instar larvae were placed in 100 mltap water in a 120-ml Styrofoam cup. One untreatedcontrol cup was set up for each replicate. Larvae weretransferred from rearing pans to water in test cupsusing a small piece of Nylon window screen. Threedrops of 10% rabbit chow pellet suspension wereadded to each cup as larval food. Bioassay was con-ducted at 25 � 1�C. Larval mortality was read at 24 hafter treatment. Moribund larvae that dove sluggishly,and were unable to resurface themselves in responseto disturbance, were also considered dead. Concen-trationÐmortality data were analyzed using POLO-PCprobit analysis to calculate lethal concentrations(LC), their 95% conÞdential limits (CL), and slopevalue of the concentrationÐresponse curve. Data Þt tothe probit analysis model was determined by hetero-geneity value (H � �2/df; LeOra Software 1987).Resistance Ratio Calculation. Resistance ratios
(RR) at LC50 and LC90 were calculated by dividinglethal levels in spinosad-selected population by thoseof freshly collected wild populations and of laboratoryreference colony. Resistance development was cate-gorized as tolerance (RR � 5), low level (RR � 5Ð20),moderate level (RR � 21Ð100), and high level (RR�100).
Results
Spinosad-Selected Colony. In response to the suc-cessive selections for 45 generations, the susceptibilityto Natular XRG (spinosad) declined signiÞcantly.During generations F1Ð9, the LC levels increasedslightly, from 0.671 to 1.343 ppm for LC50 and from2.254 to 3.628 ppm for LC90. Moderate increases in LClevels occurred during generations F11Ð19 when LC50
ranged from 2.597 to 5.704 ppm while LC90 ßuctuatedbetween 7.206 and 14.24 ppm. During generationsF21Ð35, the LC levels further elevated to 6.167Ð11.51ppm for LC50 and 14.97Ð42.90 for LC90. More pro-found increases in LC levels occurred during gener-ations F37Ð45, when LC50 signiÞcantly increased from42.6 to 693.5 ppm and LC90 from 199.1 to 10,459.1 ppm.The LC levels with 95% CL are presented in Table 1and Fig. 1. Throughout the selection process, the slopevalue, a parameter to reßect the population hetero-geneity in terms of response to spinosad treatment,ranged from 1.09 to 4.64 (Avg. � SE � 2.76 � 0.19).The slope value declined noticeably during genera-tions F37Ð45 (0.97Ð1.92), coincidental to signiÞcant de-creases in susceptibility to spinosad. Most of the data
422 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 51, no. 2

Tab
le1
.Su
scep
tibi
lity
(ppm
prod
uct)
tosp
inos
adin
resp
onse
tose
lect
ion
usin
gN
atul
arX
RG
(2.5
%A
I)in
Cx.
quin
quef
asci
atus
Gen
era
tion
s/bio
assa
ydat
esa
Spin
osa
dse
lect
ed
pop
Fre
shly
collect
ed
wild
popula
tion
sR
efe
ren
cela
bora
tory
(spin
osa
d-n
aõve)
colo
ny
LC
50(9
5%C
L)
LC
90(9
5%C
L)
Slo
pe
�2 /d
fL
C50
(95%
CL
)L
C90
(95%
CL
)Slo
pe
�2 /d
fL
C50
(95%
CL
)L
C90
(95%
CL
)Slo
pe
�2 /d
f
F1/
30N
ov.20
100.
671
(0.4
67Ð0
.929
)2.
254
(1.5
48Ð4
.023
)3.
44�
0.23
1.04
0.25
0(0
.142
Ð0.3
72)
0.49
4(0
.336
Ð1.0
95)
4.33
�0.
441.
570.
272
(0.2
28Ð0
.316
)0.
740
(0.6
28Ð0
.917
)2.
95�
0.29
0.96
F3/
12Ja
n.20
111.
120
(0.3
13Ð1
.986
)3.
160
(1.7
96Ð5
.512
)2.
84�
0.34
1.76
0.26
3(0
.223
Ð0.3
02)
0.63
3(0
.545
Ð0.7
63)
3.35
�0.
310.
430.
273
(0.2
24Ð0
.329
)0.
835
(0.6
96Ð1
.063
)2.
64�
0.28
0.69
F5/
17F
eb.20
111.
140
(0.9
83Ð1
.239
)2.
700
(2.3
08Ð3
.309
)3.
41�
0.32
0.72
0.26
4(0
.201
Ð0.3
35)
0.70
8(0
.432
Ð1.3
43)
2.39
�0.
210.
360.
234
(0.1
26Ð0
.366
)0.
798
(0.4
80Ð1
.052
)2.
41�
0.24
1.56
F7/
29M
ar.20
111.
064
(0.9
04Ð1
.225
)2.
867
(2.4
05Ð3
.617
)2.
98�
0.29
0.84
0.19
6(0
.167
Ð0.2
25)
0.50
9(0
.431
Ð0.6
29)
3.08
�0.
280.
910.
324
(0.1
90Ð0
.499
)0.
891
(0.5
59Ð1
.605
)2.
92�
0.30
1.72
F9/
5M
ay20
111.
343
(1.1
57Ð1
.538
)3.
628
(3.0
29Ð4
.618
)2.
97�
0.29
0.60
0.32
0(0
.280
Ð0.3
63)
0.78
6(0
.663
Ð0.9
93)
3.29
�0.
330.
270.
295
(0.2
54Ð0
.340
)0.
856
(0.6
99Ð1
.129
)2.
44�
0.25
0.38
F11
/28
Jun
e20
112.
597
(2.3
61Ð2
.872
)7.
206
(6.0
46Ð9
.079
)2.
48�
0.20
1.22
0.20
6(0
.143
Ð0.2
75)
0.69
1(0
.484
Ð1.3
06)
2.44
�0.
111.
260.
236
(0.1
96Ð0
.275
)0.
808
(0.6
46Ð1
.101
)2.
38�
0.28
0.71
F13
/4
Aug.20
112.
697
(2.3
61Ð3
.117
)7.
326
(5.7
88Ð1
0.41
)2.
95�
0.34
1.82
0.31
7(0
.273
Ð0.3
63)
0.89
1(0
.730
Ð1.1
76)
2.85
�0.
300.
340.
351
(0.3
04Ð0
.402
)0.
979
(0.7
96Ð1
.312
)2.
87�
0.31
0.67
F15
/19
Sept.
2011
3.96
6(3
.298
Ð5.1
09)
14.2
4(9
.61Ð
27.3
9)2.
31�
0.31
0.85
0.54
6(0
.489
Ð0.6
20)
1.20
1(0
.971
Ð1.7
12)
3.75
�0.
520.
130.
301
(0.2
63Ð0
.339
)0.
709
(0.6
04Ð0
.879
)3.
44�
0.34
0.41
F17
/26
Oct
.20
115.
704
(5.0
16Ð6
.454
)14
.18
(11.
89Ð1
8.00
)3.
24�
0.31
0.20
0.27
8(0
.239
Ð0.3
19)
0.78
2(0
.646
Ð1.0
12)
2.86
�0.
280.
750.
340
(0.2
94Ð0
.341
)0.
972
(0.7
88Ð1
.310
)2.
81�
0.30
0.67
F19
/14
Dec.
2011
4.98
7(4
.434
Ð5.6
32)
11.1
5(9
.33Ð
14.2
9)3.
67�
0.37
0.79
0.21
5(0
.186
Ð0.2
44)
0.48
6(0
.420
Ð1.5
85)
3.62
�0.
340.
330.
357
(0.3
16Ð0
.398
)0.
752
(0.6
49Ð0
.926
)3.
96�
0.44
0.38
F21
/1
Feb.20
127.
206
(6.3
28Ð8
.434
)17
.11
(13.
39Ð2
5.23
)3.
41�
0.44
0.11
0.33
2(0
.225
Ð0.4
50)
0.78
8(0
.556
Ð1.8
17)
3.42
�0.
361.
220.
354
(0.3
12Ð0
.397
)0.
798
(0.6
79Ð0
.999
)3.
63�
0.39
0.58
F23
/13
Mar
.20
129.
579
(8.5
44Ð1
0.80
6)20
.28
(17.
12Ð2
5.53
)3.
94�
0.38
0.22
0.34
2(0
.305
Ð0.3
87)
0.76
0(0
.633
Ð0.9
82)
3.70
�0.
380.
650.
283
(0.2
49Ð0
.325
)0.
703
(0.5
56Ð1
.033
)3.
25�
0.45
0.45
F25
/23
Apri
l20
128.
672
(7.8
15Ð9
.670
)16
.37
(14.
10Ð2
0.11
)4.
64�
0.47
0.59
0.23
6(0
.206
Ð0.2
73)
0.70
0(0
.546
Ð1.0
10)
2.71
�0.
300.
380.
314
(0.2
80Ð0
.358
)0.
707
(0.5
78Ð0
.950
)3.
64�
0.39
0.72
F27
/29
May
2012
7.71
9(6
.764
Ð8.8
51)
20.9
3(1
6.63
Ð26.
96)
3.04
�0.
290.
170.
281
(0.2
51Ð0
.313
)0.
613
(0.5
25Ð0
.760
)3.
78�
0.38
0.64
0.40
8(0
.364
Ð0.4
59)
0.94
8(0
.796
Ð1.2
03)
3.50
�0.
330.
71F
29/
9Ju
ly20
126.
167
(4.5
70Ð7
.659
)14
.97
(11.
77Ð2
2.07
)3.
33�
0.33
1.16
0.33
3(0
.257
Ð0.4
32)
0.71
5(0
.528
Ð1.3
04)
3.87
�0.
361.
030.
394
(0.2
92Ð0
.542
)0.
792
(0.5
69Ð1
.684
)4.
23�
0.38
1.49
F31
/21
Aug.20
127.
151
(6.0
57Ð8
.381
)23
.81
(18.
45Ð3
4.84
)2.
45�
0.29
0.85
0.26
3(0
.233
Ð0.2
94)
0.91
3(0
.763
Ð1.1
62)
2.37
�0.
200.
790.
323
(0.2
93Ð0
.356
)0.
617
(0.5
38Ð0
.742
)4.
55�
0.43
0.98
F33
/13
Oct
.20
1211
.51
(10.
15Ð1
3.00
)31
.10
(25.
35Ð4
1.93
)2.
97�
0.33
0.45
0.29
5(0
.199
Ð0.4
13)
0.64
5(0
.453
Ð1.3
87)
3.77
�0.
371.
310.
333
(0.3
05Ð0
.363
)0.
844
(0.7
36Ð1
.003
)3.
17�
0.22
0.43
F35
/21
Nov.20
1211
.27
(9.4
3Ð13
.25)
42.9
0(3
2.71
Ð64.
77)
2.21
�0.
260.
280.
334
(0.2
93Ð0
.380
)0.
896
(0.7
34Ð1
.185
)2.
99�
0.30
0.42
0.31
4(0
.285
Ð0.3
47)
0.60
4(0
.525
Ð0.7
29)
4.52
�0.
430.
77F
37/
17Ja
n.20
1342
.69
(31.
87Ð6
0.91
)19
9.1
(119
.3Ð5
09.5
)1.
92�
0.15
0.81
0.44
3(0
.318
Ð0.6
65)
1.10
2(0
.717
Ð2.3
13)
3.24
�0.
321.
410.
424
(0.3
77Ð0
.479
)1.
015
(0.8
44Ð1
.310
)3.
38�
0.32
0.57
F39
/5
Mar
.20
1310
0.3
(81.
50Ð1
23.0
)63
5.2
(446
.5Ð1
049.
2)1.
60�
0.16
0.82
0.33
5(0
.293
Ð0.3
82)
0.93
7(0
.761
Ð1.2
55)
2.57
�0.
300.
460.
271
(0.2
35Ð0
.308
)0.
732
(0.6
05Ð0
.57)
2.96
�0.
320.
34F
41/
30A
pri
l20
1324
4.8
(195
.9Ð3
11.8
)1,
631.
6(1
110.
8Ð27
42.7
)1.
56�
0.14
0.75
0.27
2(0
.238
Ð0.3
08)
0.69
9(0
.583
Ð0.9
00)
3.12
�0.
320.
460.
311
(0.2
78Ð0
.348
)0.
704
(0.5
99Ð0
.878
)3.
61�
0.34
0.50
F43
/28
Jun
e20
1341
6.0
(287
.7Ð6
41.1
)8,
765.
8(4
,201
.5Ð3
0,59
3.1)
0.97
�0.
120.
220.
427
(0.3
83Ð0
.479
)0.
935
(0.7
92Ð1
.170
)3.
77�
0.35
0.33
0.38
1(0
.341
Ð0.4
26)
0.84
8(0
.718
Ð1.0
62)
3.69
�0.
340.
99F
45/
19A
ug.20
1369
3.5
(507
.Ð95
7.7)
10,4
59.1
(6,2
16.3
Ð21,
244.
5)1.
09�
0.10
0.38
0.49
0(0
.438
Ð0.5
53)
1.08
8(0
.912
Ð1.3
83)
3.70
�0.
350.
480.
311
(0.2
35Ð0
.412
)0.
613
(0.4
52Ð1
.207
)4.
34�
0.41
1.32
aG
en
era
tion
sof
spin
osa
d-s
ele
cted
popula
tion
wit
hco
rresp
on
din
gdat
es
of
bio
assa
yfo
ral
lth
ree
popula
tion
s.
March 2014 SU AND CHENG: LABORATORY SELECTION OF RESISTANCE TO SPINOSAD 423
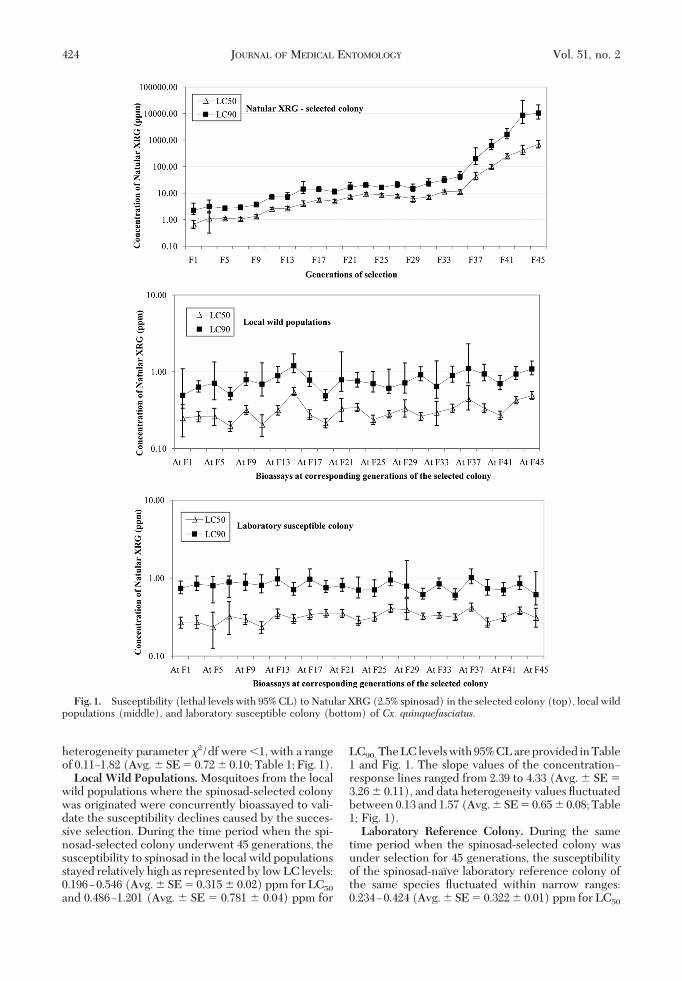
heterogeneity parameter �2/df were �1, with a rangeof 0.11Ð1.82 (Avg. � SE � 0.72 � 0.10; Table 1; Fig. 1).Local Wild Populations.Mosquitoes from the local
wild populations where the spinosad-selected colonywas originated were concurrently bioassayed to vali-date the susceptibility declines caused by the succes-sive selection. During the time period when the spi-nosad-selected colony underwent 45 generations, thesusceptibility to spinosad in the local wild populationsstayed relatively high as represented by low LC levels:0.196Ð0.546 (Avg. � SE � 0.315 � 0.02) ppm for LC50
and 0.486Ð1.201 (Avg. � SE � 0.781 � 0.04) ppm for
LC90. The LC levels with 95% CL are provided in Table1 and Fig. 1. The slope values of the concentrationÐresponse lines ranged from 2.39 to 4.33 (Avg. � SE �3.26 � 0.11), and data heterogeneity values ßuctuatedbetween 0.13 and 1.57 (Avg. � SE � 0.65 � 0.08; Table1; Fig. 1).Laboratory Reference Colony. During the same
time period when the spinosad-selected colony wasunder selection for 45 generations, the susceptibilityof the spinosad-naõve laboratory reference colony ofthe same species ßuctuated within narrow ranges:0.234Ð0.424 (Avg. � SE � 0.322 � 0.01) ppm for LC50
Fig. 1. Susceptibility (lethal levels with 95% CL) to Natular XRG (2.5% spinosad) in the selected colony (top), local wildpopulations (middle), and laboratory susceptible colony (bottom) of Cx. quinquefasciatus.
424 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 51, no. 2
and 0.604Ð1.015 (Avg. � SE � 0.794 � 0.02) ppm forLC90. The 95% CL are provided in Table 1 and Fig. 1.The slope values of the concentrationÐresponse weresimilar to those in local wild populations, ßuctuatingfrom 2.38 to 4.55 (Avg. � SE � 3.36 � 0.14). The dataheterogeneity values ranged from 0.34 to 1.72 (Avg. �SE � 0.78 � 0.08; Table 1; Fig. 1).Resistance Ratios. RR is a function of the spinosad
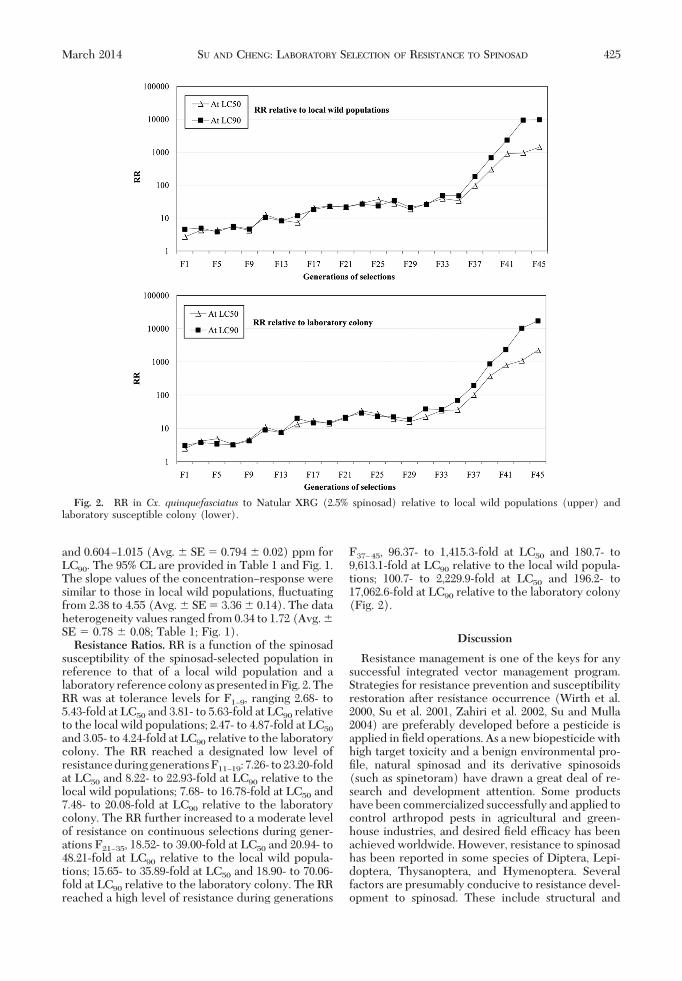
susceptibility of the spinosad-selected population inreference to that of a local wild population and alaboratory reference colony as presented in Fig. 2. TheRR was at tolerance levels for F1Ð9, ranging 2.68- to5.43-fold at LC50 and 3.81- to 5.63-fold at LC90 relativeto the local wild populations; 2.47- to 4.87-fold at LC50
and 3.05- to 4.24-fold at LC90 relative to the laboratorycolony. The RR reached a designated low level ofresistanceduringgenerationsF11Ð19: 7.26- to23.20-foldat LC50 and 8.22- to 22.93-fold at LC90 relative to thelocal wild populations; 7.68- to 16.78-fold at LC50 and7.48- to 20.08-fold at LC90 relative to the laboratorycolony. The RR further increased to a moderate levelof resistance on continuous selections during gener-ations F21Ð35, 18.52- to 39.00-fold at LC50 and 20.94- to48.21-fold at LC90 relative to the local wild popula-tions; 15.65- to 35.89-fold at LC50 and 18.90- to 70.06-fold at LC90 relative to the laboratory colony. The RRreached a high level of resistance during generations
F37Ð45, 96.37- to 1,415.3-fold at LC50 and 180.7- to9,613.1-fold at LC90 relative to the local wild popula-tions; 100.7- to 2,229.9-fold at LC50 and 196.2- to17,062.6-fold at LC90 relative to the laboratory colony(Fig. 2).
Discussion
Resistance management is one of the keys for anysuccessful integrated vector management program.Strategies for resistance prevention and susceptibilityrestoration after resistance occurrence (Wirth et al.2000, Su et al. 2001, Zahiri et al. 2002, Su and Mulla2004) are preferably developed before a pesticide isapplied in Þeld operations. As a new biopesticide withhigh target toxicity and a benign environmental pro-Þle, natural spinosad and its derivative spinosoids(such as spinetoram) have drawn a great deal of re-search and development attention. Some productshave been commercialized successfully and applied tocontrol arthropod pests in agricultural and green-house industries, and desired Þeld efÞcacy has beenachieved worldwide. However, resistance to spinosadhas been reported in some species of Diptera, Lepi-doptera, Thysanoptera, and Hymenoptera. Severalfactors are presumably conducive to resistance devel-opment to spinosad. These include structural and
Fig. 2. RR in Cx. quinquefasciatus to Natular XRG (2.5% spinosad) relative to local wild populations (upper) andlaboratory susceptible colony (lower).
March 2014 SU AND CHENG: LABORATORY SELECTION OF RESISTANCE TO SPINOSAD 425

functional similarities of spinosyn A and D. The targetspecies may easily develop resistance by means ofbehavioral avoidance, reduced absorption, decreasedtarget site, target afÞnity, or both, enhanced metab-olism (detoxication), or increased cell efßux. Anotherreason contributing to resistance development couldbe related to the mode of action. Spinosad activatesnAChR in competition with the normal neurotrans-mitter nACh. Sublethally dosed targets thus couldrecover (T. S. et al., unpublished data) once the nor-mal signal transduction resumes. This sublethal expo-sure issue is more likely to occur in terrestrial arthro-pods, exposure of which is optional, whereas aquatictargets are exposed continuously once the entire hab-itat is treated.
Once newly developed spinosad products wereavailable for mosquito control, studies to explore thepotential for resistance development and character-istics of spinosad resistance were prompted by seren-dipitous observations of survivingCx. quinquefasciatuslarvae and pupae in a microcosm evaluation of a nat-ural population to Natular XRG applied at a midrangedosage (14.2 kg/ha). The subsequent generations ofdescendants of the Natular XRG survivors were sub-jected to successive selections at LC70Ð90 at late thirdor early fourth instars for 45 generations. SigniÞcantlevels of resistance were conÞrmed relative to bothlocal wild populations and a laboratory reference col-ony. Tolerance (RR � 5.0) occurred at F1Ð9, low levelsof resistance were observed at F11Ð19, moderate levelsat F21Ð35, and high levels at F37Ð45. In response tosuccessive selection for 45 generations, resistance lev-els increased in an exponential pattern, indicating thata recessive gene might be involved. This was true forspinosad-resistant house ßyM. domestica (Shono andScott 2003) and most other species studied, which isin agreement with a target site-related resistancemechanism (see Sparks et al. 2012, Geng et al. 2013).This aspect warrants further investigations in the fu-ture.
The susceptibility to spinosad of the local wild pop-ulation of Cx. quinquefasciatus varied slightly within(95% CL) and between (SE of LC levels) samples inthe laboratory bioassay, but nevertheless the variabil-ity was mostly greater than that in the laboratoryreference colony. This scenario is attributable to thefact that local populations randomly sampled are ex-pected to have higher population heterogeneity ascompared with an inbred population in the laboratorycolony. The bioassay of the spinosad-selected line,local wild populations, and laboratory reference col-ony all showed low data heterogeneity values (�2/df),mostly being �1 (LeOra Software 1987). As noticed inearly studies, the variability (range of 95% CL) at theLC90 was greater than at the LC50 (Wirth et al. 2000,Su et al. 2001, Zahiri et al. 2002, Su and Mulla 2004).
High levels of resistance occurred in response tosuccessive selection at sublethal doses in a conÞnedpopulation in the laboratory. These conditions maynot happen in the Þeld, provided spinosad product isapplied at the full dose and in rotation with otherproducts with different modes of actions. Further-
more, gene ßow between the treated populations andadjacent untreated populations should dilute the fre-quency of resistance gene(s); hence, resistance de-velopment may not occur rapidly in the Þeld. Avoid-ance of sublethal exposures is a key for prevention ofresistance development. In separate laboratory stud-ies, we have observed that �50% of mosquito larvaerecovered when larvae were treated with NatularXRG at LC50 (24 h at 25 � 1�C), if the exposure timewas shorter than 7.2 h (RT50 � 7.2 h; T. S. et al.,unpublished data). In contrast to terrestrial insectpests that can avoid contacting or feeding on treatedplants, mosquito larvae and pupae are constantly ex-posed to spinosad in treated water through feedingand contact. However, sublethal dosing may occur inmosquito control, if the spinosad product is appliedbelow the maximum lethal rates, or the volume ofbreeding source water is increased by ßooding orprecipitation posttreatment.
In summary, the risk of resistance development tospinosad in mosquitoes does exist, even though spi-nosad along with spinetoram is the only pesticide withunique mode of action in group 5 by Insecticide Re-sistance Acting Committee. Moderate to high levels ofresistance to spinosad in mosquitoes seem fairly easyto induce. Future studies are warranted to examinecross-resistance, genetic and biochemical mecha-nisms, stability of resistance, biological Þtness in re-sistant population, as well as resistance managementtactics.
Acknowledgments
The assistance in colony maintenance and bioassay by thelaboratory staff, particularly Assistant Vector Ecologist Jen-nifer Thieme with the West Valley Mosquito and VectorControl District, Ontario, CA, is duly acknowledged.
References Cited
Bielza, P., V. Quinto, E. Fernandez, C. Gravalos, and J.Contreras. 2007. Genetics of spinosad resistance inFrankliniella occidentalis (Thysanoptera: Thripidae). J.Econ. Entomol. 100: 916Ð920.
Ferguson, J. S. 2004. Development and stability of insecti-cide resistance in the leaf-miner Liriomyza trifolii (Dip-tera: Agromyzidae) to cyromazine, abamectin, and spi-nosad. J. Econ. Entomol. 97: 112Ð119.
Geng, C. X., G. B.Watson, and T. C. Sparks. 2013. Nicotinicacetylcholine receptors as spinosyn targets for insect pestmanagement. Adv. Insect Physiol. 44: 101Ð210.
Hertlein, M. B., C. Mayrotas, C. Joussesume, M. Lysandrou,G. D. Thompson, W. Jany, and S. A. Ritchie. 2010. Areview of spinosad as a natural product for larval mos-quito control. J. Am. Mosq. Control Assoc. 26: 67Ð87.
Hsu, J. C., andH. T. Feng. 2006. Development of resistancein Oriental fruit ßy (Diptera: Tephritidae) in laboratoryselection and cross-resistance. J. Econ. Entomol. 99: 931Ð936.
Kakani, E. G., N. E. Zygouridis, K. T. Tsoumani, N. Seraphi-des, F. G. Zalom, and K. D. Mathiopoulos. 2010. Spi-nosad resistance development in wild olive fruit ßyBactrocera oleae (Diptera: Tephritidae) populations inCalifornia. Pest Manage. Sci. 66: 447Ð453.
426 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 51, no. 2

Kranthi, K. R., S. Shakir, and S. K. Banerjee. 2000. Baselinetoxicity of spinosad on the cotton bollworm, Helicoverpaarmigera (Hub.) in India. Resistant Pest Manage. Newsl.11: 9Ð12.
LeOra Software. 1987. POLO-PC: a userÕs guide to probit orlogit analysis. LeOra Software, Berkeley, CA.
Liu, S. S., Z. M. Li, Y. Q. Liu, M. G. Feng, and Z. H. Tang.2007. Promoting selection of resistance to spinosad in theparasitoid Catesia plutellae by integrating resistance ofhosts to the insecticide into the selection process. Biol.Control 41: 246Ð255.
Mertz, F. P., and R. Yao. 1990. Saccharopolyspora spinosasp. nov. isolated from soil collected in a sugar mill rumstill. Int. J. Syst. Evol. Microbiol. 40: 34Ð39.
Moulton, J. K., D. A. Pepper, and T. J. Dennehy. 2000. Beetarmyworm (Spodoptera exigua) resistance to spinosad.Pest Manage. Sci. 56: 842Ð848.
Salgado, V. L., J. J. Sheets, G. B. Watson, and A. L. Schmidt.1998. Studies on the mode of action of spinosad: theinternal effective concentration and the concentrationdependence of neural excitation. Pestic. Biochem.Physiol. 60: 103Ð110.
Sayyed, A. H., D. Omar, andD. J.Wright. 2004. Genetics ofspinosad resistance in a multi-resistant Þeld-selected pop-ulation of Plutella xylostella. Pest Manage. Sci. 60: 827Ð32.
Shono, T., and J. G. Scott. 2003. Spinosad resistance in thehouseßy,Musca domestica, is due to a recessive factor onautosome 1. Pestic. Biochem. Physiol. 75: 1Ð7.
Sial, A. A., J. F. Brunner, and S. F. Garczynski. 2011. Bio-chemical characterization of chlorantraniliprole andspinetoram resistance in laboratory-selected oblique-banded leafroller, Choristoneura rosaceana (Harris)(Lepidoptera: Tortricidae). Pestic. Biochem. Physiol.99: 274Ð279.
Sparks, T. C., J. E.Dripps, G. B.Watson, andD. Paroonagian.2012. Resistance and cross-resistance to the spinosyn Ð Areview and analysis. Pestic. Biochem. Physiol. 102: 1Ð10.
Su, T., and M. L. Cheng. 2012. Resistance development inCulex quinquefasciatus to spinosad: a preliminary report.J. Am. Mosq. Control Assoc. 28: 263Ð267.
Su, T., and M. S. Mulla. 2004. Documentation of high levelBacillus sphaericus-resistance in tropical Culex quinque-
fasciatuspopulations from Thailand. J. Am. Mosq. ControlAssoc. 20: 405Ð411.
Su, T., B. A. Soliman, J. D. Chaney, M. S. Mulla, and J. W.Beehler. 2001. SusceptibilityofCulexmosquitoesbreed-ing in dairy ponds before and after treatment with Ba-cillus sphaericus formulation. Proc. Papers 69th Ann. Conf.Mosq. Vector Control Assoc. Calif. 69: 110Ð116.
Thompson, G. D., R. Dutton, and T. C. Sparks. 2000. Spi-nosad-a case study: an example from a natural productsdiscovery programme. Pest Manage. Sci. 56: 696Ð702.
Wang,W., J. C.Mo, J. A. Cheng, P. J. Zhuang, andZ.H. Tang.2006. Selection and characterization of spinosad resis-tance in Spodoptera exigua (Hubner) (Lepidoptera: Noc-tuidae). Pestic. Biochem. Physiol. 84: 180Ð187.
Wirth, M. C., G. P. Georghiou, J. I. Malik, and G. H. Abro.2000. Laboratory selection for resistance toBacillus spha-ericus inCulexquinquefasciatus(Diptera: Culicidae) fromCalifornia, USA. J. Med. Entomol. 37: 534Ð40.
Wyss, C. F., H. P. Young, J. Shukla, and R. M. Roe. 2003.Biology and genetics of a laboratory strain of the tobaccobudworm, Heliothis virescens (Lepidoptera: Noctuidae),highly resistant to spinosad. Crop Prot. 22: 307Ð314.
Young, H. P., W. D. Bailey, and R. M. Roe. 2003. Spinosadselection of a laboratory strain of the tobacco budworm,Heliothis virescens (Lepidoptera: Noctuidae), and char-acterization of resistance. Crop Prot. 22: 265Ð273.
Zahiri, N. S., T. Su, and M. S. Mulla. 2002. Strategies formanagement of resistance in mosquitoes to the microbialcontrol agent Bacillus sphaericus. J. Med. Entomol. 39:513Ð520.
Zhao, J. Z., Y. X. Li, H. L. Collins, L. Gusukuma-Minuto,R.F.L. Mau, G. D. Thompson, and A. M. Shelton. 2002.Monitoring and characterization of diamondback mothresistance to spinosad. J. Econ. Entomol. 95: 430Ð436.
Zhao, J. Z., H. L. Collins, Y. X. Li, R.F.L. Mau, G. D. Thomp-son, M. Hertlein, J. T. Andaloro, R. Boykin, and A. M.Shelton. 2006. Monitoring of diamondback moth (Lep-idoptera: Plutellidae) resistance to spinosad, indoxacarb,and emamectin benzoate. J. Econ. Entomol. 99: 176Ð181.
Received 3 September 2013; accepted 6 December 2013.
March 2014 SU AND CHENG: LABORATORY SELECTION OF RESISTANCE TO SPINOSAD 427





























