Embed Size (px)
Citation preview

Validation of Hylodes henselii Peters, 1870, from Southern Brazil andDescription of Acoustic Variation in Eleutherodactylus guentheri
(Anura: Leptodactylidae)
AXEL KWET1,2,3
AND MIRCO SOLE2
1Staatliches Museum fur Naturkunde Stuttgart, Zoology, Rosenstein 1, D-70191 Stuttgart, Germany2Zoological Institute, University of Tubingen, Auf der Morgenstelle 28, D-72076 Tubingen, Germany,
E-mail: [email protected]
ABSTRACT.—Comparison of advertisement calls of Eleutherodactylus guentheri from different localities insouthern and southeastern Brazil revealed considerable acoustic variation among populations. Because of
significant differences in call parameters, we conclude that the southern form is a distinct species, for which
the name Hylodes henselii Peters, 1870, is available. Examination of the holotypes of H. henselii and Hylodesguentheri Steindachner, 1864, along with recently collected material, showed that both species aremorphologically similar, possibly sibling species. Eleutherodactylus henselii is the southernmost member of
the genus Eleutherodactylus occurring in subtropical Atlantic rain forest and Araucaria forest of the Brazilian
states of Rio Grande do Sul and Santa Catarina, and Misiones, Argentina. Bioacoustic data suggest that at leastone additional cryptic species is involved in the Eleutherodactylus guentheri group.
Hylodes henselii Peters, 1870, is currentlyconsidered a synonym of Eleutherodactylus guen-theri (Steindachner, 1864) (Heyer, 1984; Frost,2002), a wide-spread leptodactylid occurring inthe Atlantic forest and Araucaria forest morpho-climatic domains of southern and southeasternBrazil, ranging from northeastern Rio Grando doSul to Espirito Santo (Heyer, 1984), in MisionesProvince, Argentina (Carrizo et al., 1989; Lavillaand Cei, 2001), and presumably in adjacentParaguay (Frost, 2002). Lynch (1976) revised thespecies groups of Eleutherodactylus (Hylodes sensuPeters, 1870) and recognized a binotatus series inwhich he tentatively included Eleutherodactylushenselii as a valid species, among other taxa, likeEleutherodactylus binotatus and E. guentheri. Heyer(1984) doubted the validity of this binotatusgroup and proposed a monophyletic guenthericluster. He regarded H. henselii as a synonym ofE. guentheri and followed Lynch (1976) inconsidering the holotype of henselii to be lost.Heyer (1984) made an arbitrary decision toclarify the nomenclature by designating theholotype of guentheri (NHMW 16515) as theneotype of henselii. In the most recent classifica-tion as proposed by Lynch and Duellman (1997)and Lynch (2001), the guentheri cluster of Heyer(1984) is allocated within a broadly interpretedE. binotatus series.
The holotype of H. henselii (ZMB 6813)collected near Porto Alegre, Rio Grande do Sul,southern Brazil, was recently recovered in thecollection of the Zoological Museum Berlin(Bauer et al., 1995). We compared this specimenwith material identified as E. guentheri from othercollections. Additionally, we analyzed advertise-ment calls assigned to E. guentheri from variouslocalities in Brazil and Argentina. Because ofsignificant differences in many parameters, suchas call length, number of notes per call and noterepetition rate, we conclude that the southernform must be considered a distinct species, forwhich the name H. henselii Peters, 1870, isavailable and should be revalidated. The purposeof this paper is to resurrect E. henselii and todescribe call variation in E. guentheri.
MATERIALS AND METHODS
The following measurements (see Appendix 1)were taken to the nearest 0.1 mm with calipers:snout–vent length (SVL); head length, defined asthe diagonal distance from the tip of the snout tothe right angle of the jaw (HL); greatest headwidth (HW); horizontal eye diameter (ED);horizontal tympanum diameter (TD); interorbitaldistance (IO); internarial distance (IN); eye–nostril distance (EN); thigh length (TL); shanklength (SL); and foot length (FL). Specimensexamined are deposited in the following collec-tions: Biogeographische Sammlung der Univer-sitat des Saarlandes (BGSS, Saarbrucken,Germany), Centro Nacional de InvestigacionesIologicas (CENAI, now in the MACN collection),Museo Argentino de Ciencias Naturales (MACN,
3 Corresponding Author. Present address: StaatichesMuseum fur Naturkunde Stuttgart, Zoology, Rose-nstein 1, D-70191 Stuttgart, Germany; E-mail:[email protected]
Journal of Herpetology, Vol. 39, No. 4, pp. 521–532, 2005Copyright 2005 Society for the Study of Amphibians and Reptiles

Buenos Aires, Argentina), Museu de Ciencias eTecnologia da PUCRS (MCP, Porto Alegre,Brazil), Museo de Ciencias Naturales de La Plata(MLP, La Plata, Argentina), Museu Nacional Riode Janeiro (MNRJ, Rio de Janeiro, Brazil),Naturhistorisches Museum Wien (NMW, Wien,Austria), Staatliches Museum fur NaturkundeStuttgart (SMNS, Stuttgart, Germany), NationalMuseum of Natural History (USNM, Washington,DC), and Museum fur Naturkunde Berlin,formerly Zoologisches Museum Berlin (ZMB,Berlin, Germany).
Most recordings of calls were obtained witha Sony WM-D6C tape recorder and a Sennheisermicrophone (module ME66, K6 power module).The calls were analyzed with a Macintosh-basedsignal analysis software (Canary 1.2, CornellUniversity, Ithaca, NY) at a sampling frequencyof 44.1 kHz. Measurements of the followingacoustic parameters were taken: call duration,note duration, note interval, notes per call, noterepetition rate, dominant frequency, and otherfrequencies. Temporal parameters were mea-sured in seconds (sec) or milliseconds (msec)and spectral parameters in Hertz (Hz). Taperecordings of morphologically identified E. guen-theri (five calls per individual when available)and Eleutherodactylus manezinho, which occurssyntopically, were analyzed from the followinglocations (Brazilian localities arranged fromSouth to North): Brazil: Rio Grande do Sul: SaoFrancisco de Paula: Centro de Pesquisas eConservacao da Natureza Pro-Mata; recordedby AK, (1) voucher specimen SMNS 9005, 9November 1995, 0030 h, 198C air temperature; (2)voucher specimen MCP 3182, 7 December 1997,2330 h, 198C air temperature. Brazil: SantaCatarina: Florianopolis: Ilha de Santa Catarina;recorded by AK, (1) unvouchered, 11 October2002, 2130 h, 20.58C; (2) voucher specimen MCP8281 or 8282, 10 December 2004, 2330 h, 19–208Cair temperature. Brazil: Santa Catarina: Ilhota:Morro do Bau; recorded by AK, unvouchered, 23October 2002, 1000 h, 17.58C. Brazil: SantaCatarina: Guaramirim: Instituto Ra-bugio; re-corded by G. Woehl, three different calls(available at http://www.ra-bugio.org.br/index.html), unvouchered, without data. Brazil: SantaCatarina: Pirabeiraba; recorded by W. R. Heyer,voucher specimen USNM 235727, 10 December1978, 2030 h, without temperature record. Brazil:Sao Paulo: Serra da Bocaina; recorded by W. R.Heyer, voucher specimen USNM 235699, 23December 1976, 2000–2145 h, 19–208C. Brazil:Sao Paulo: Boraceia; recorded by A. S. Rand, twounvouchered specimens, 9 December 1984, 1920h, 19.28C. Argentina: Misiones: DepartamentoSan Pedro: Colonia Victoria; recorded by D.Baldo, two voucher specimens MLP DB 1923–24, 14 October 2002, 2205–2220 h, 19–208C.
Eleutherodactylus manezinho, Santa Catarina: Flo-rianopolis: Ilha de Santa Catarina; recorded byAK; (1) voucher specimen MCP 6365, 6 October2002, 2230 h, 20.58C; (2) voucher specimen MCP7647, 17 November 2003, 2230 h, 238C.
RESULTS
Eleutherodactylus henselii (Peters, 1870)Figures 1–2
Hylodes sp. (Cystignathus sp.).—Hensel, 1867:161–162.
Hylodes henselii.—Peters, 1870:648 (holotype:ZMB 6813; type locality: near Porto Alegre,Rio Grande do Sul, Brazil); Bokermann,1966:67 (partim).
Hylodes griseus (non-Hallowell).—Boulenger,1882:206 (partim), 1886:440; Nieden, 1923:460 (partim).
Hylodes gollmeri (non-Peters).—Baumann,1912:143, 150, 161 (partim).
Hylodes ranoides (non-Spix).—Miranda-Ribeiro,1926:53–55 (partim).
Eleutherodactylus griseus (non-Hallowell).—Gorham, 1966:75–76 (partim).
Eleutherodactylus henselii.—Lynch, 1976:3, 7; Har-ding, 1983:162.
Eleutherodactylus guentheri (non-Steindachner).—Cochran, 1955:271–274 (partim); Lynch,1976:7 (partim); Braun and Braun,1980:132; Heyer, 1984:1–42 (partim); Carrizoet al., 1989:10–11; Lavilla, 1992:83; Baueret al., 1995:46; Garcia, 1996:57–68 (partim);Lavilla and Cei, 2001:105–106; Lavillaet al., 2002:106–107.
Eleutherodactylus cf. guentheri.—Kwet, 1999:97–98; 2001:33, 35, 189; Kwet and Di-Bernardo,1999:60–61.
Diagnosis.—Eleutherodactylus henselii is amedium-sized species of the E. guentheri clustersensu Heyer (1984). It is characterized bya prolonged advertisement call with a durationof about 10–25 sec, consisting of a series of morethan 80 notes (up to 170). In other species of theguentheri cluster, the call is much shorter,consisting of 5–10 notes, with a maximum of 35notes with a total duration of less than 3 sec.Eleutherodactylus henselii is a cryptic species thatis not absolutely distinguishable morphological-ly from E. guentheri (for description of E. guentherifrom the type locality see Cochran, 1955).Although E. henselii has a rounded, slightlyprotruding snout profile (Figs. 1A, B), the snoutis more sloping in E. guentheri (Fig. 3B). Thethroat is usually white and unmottled in E.henselii but gray mottled in E. guentheri. OtherEleutherodactylus species occurring syntopicallywith E. henselii are E. binotatus and E. manezinho.
522 A. KWET AND M. SOLE

Eleutherodactylus henselii is distinguished fromthe larger E. binotatus by lacking dorsal longitu-dinal skin crests and the first finger being aboutthe same length as the second (first finger muchlonger than second in E. binotatus). It is distin-guished from E. manezinho by the more slenderbody, smaller finger discs and longer hind legs(shank length about 68–69% SVL in E. henselii;58% SVL in E. manezinho; Garcia, 1996).
Description.—This redescription is based on allavailable specimens from Rio Grande do Sul andsouthern Santa Catarina, which are, because oftheir advertisement calls, referrable to E. henselii(see Appendix 1). Measurements and ratios areonly from specimens which sex could be deter-mined. Eleutherodactylus henselii is characterizedby a distinct sexual dimorphism (SVL 21.0–27.5in males; 28.4–38.4 in females; see Variation).Measurements (in millimeters) for the holotype,ZMB 6813, an adult female faded and in poorcondition (Fig. 1A), are as follows: SVL 32.5; HL12.7; HW 11.4; ED 3.6; TD 2.0; EN 3.8; IO 5.6; IN3.0; TL 20.6; SL 23.4; FL 19.8. Head longer thanwide, its length 42.3% SVL in males, 41.6% infemales, its width 34.6% SVL in males, 35.3% infemales. Snout rounded, protruding in profile
(Figs. 1A, B), subovoid in dorsal view. No cranialcrests, canthus rostralis evident, loreal region flatand nearly vertical. Nostrils not protuberant,much closer to tip of snout than to eye. Eyeslarge, their diameter about 25% head length.Upper eyelids smooth or with small tubercles.Tympanum distinct, its diameter 55–60% eyediameter. Slight supratympanic ridge. Vocal sacsmall, single, subgular. Vocal slits present inmales. Vomerine teeth in two short, narrowlyseparated, transverse rows, located behind andbetween the choanae. Skin on dorsum smooth orindistinctly granulated, usually with two weakdorsolateral glandular folds and a Y- or W-shaped glandular ridge between the shoulders.Skin on throat and venter smooth. Limbs slender,hindlegs elongated in proportion to body. Thighlength on average 61.3% SVL in males, 62.0% infemales; tibia length 67.9% SVL in males, 69.2%in females; and foot length SVL 60.1% SVL inmales, 60.8% in females. Fingers lacking webbingand lateral fringes but possessing fine dermalridges laterally. Finger discs distinct, their widthabout 1.2–1.5 times finger width. Relative lengthsof fingers 1 5 2 , 4 , 3. Subarticular tuberclesdistinct, rounded and moderately raised. Large,
FIG. 1. Eleutherodactylus henselii, lateral views of head of (A) ZMB 6813, (B) SMNS 9125; ventral views of righthand (C) and right foot (D) of SMNS 9125. Scale bar 5 2 mm.
TAXONOMIC STATUS OF HYLODES HENSELII 523

oval, moderately raised outer and inner meta-carpal tubercles (Fig. 1C). Indistinct, creamnuptial asperities on the inner base of the firstfinger in males. Toes not webbed, laterallyfringed. Relative lengths of toes 1 , 2 , 5 ,3 , 4. Foot texture smooth with rounded, raisedsubarticular tubercles. Inner metatarsal tubercleelliptical, protruding; outer metatarsal tuberclesmall, rounded (Fig. 1D). Tarsal fold absent.
Color in Life.—Color photos of E. henselii (asEleutherodactylus cf. guentheri) are in Kwet(1999:97, 2001:35) and Kwet and Di-Bernardo(1999:61; see also Fig. 3). Dorsal colorationextremely variable, reddish, orange, grayish-, oryellowish-brown, uniform or with symmetricalpattern of indistinct dark markings and lines(Figs. 2). Some specimens with white middorsalpin stripe or broad line. Dorsal region of thesnout usually light brown, contrasting with thedarker brown coloration on loreal region anddorsum. Distinct pattern of dark and light brownvertical bars on upper lip; below eye alwaysa dark broad vertical bar. With or withoutdark interorbital stripe or bar. Black triangularblotch above tympanum, sometimes posteriorlyextended to the groin. Usually two small black
inguinal blotches. Hind legs brownish with 3–5dark cross-bars. Forearms with a few indistinctbrown blotches. Belly and throat dirty yellow orwhite, unmottled. Posterior surface of thighyellowish, uniform or indistinctly marbled withbrown. Outer shank usually with broad darkstripe, sometimes incomplete or interrupted.Coloration in preservative similar.
Variation.—The coloration varies as describedabove. Most specimens are uniformly darkbrown to grey or have a symmetrical dorsalbody pattern of dark markings. Four (MCP 3182,3183, 3683, 6853, SMNS 9125) out of 42 specimensfrom Rio Grande do Sul (about 10%) havea broad, distinct, light middorsal stripe, but mostspecimens are without such a stripe. Skinstructure varying from smooth to slightly tuber-culate. Measurements of adult males (N 5 45) ofE. henselii (in millimeters, mean 6 SD given inparenthesis) are as follows: SVL 21.0–27.5(24.3 6 1.54), HL 9.0–11.8 (10.3 6 0.68), HW 7.0–9.5 (8.460.58), ED 1.8–3.5 (2.660.41), TD 1.1–1.8(1.4 6 0.22), EN 2.2–3.9 (3.2 6 0.31), IO 3.7–5.4(4.6 6 0.41), IN 1.8–2.8 (2.3 6 0.25), TL 12.6–17.7(14.9 6 1.09), SL 14.4–18.5 (16.5 6 0.96), FL 12.2–16.5 (14.6 6 0.86); in adult females (N 5 41): SVL
FIG. 2. Eleutherodactylus henselii in life from Rio Grande do Sul, showing variation in dorsal body pattern;(A, B) adult males, (C, D) adult females.
524 A. KWET AND M. SOLE
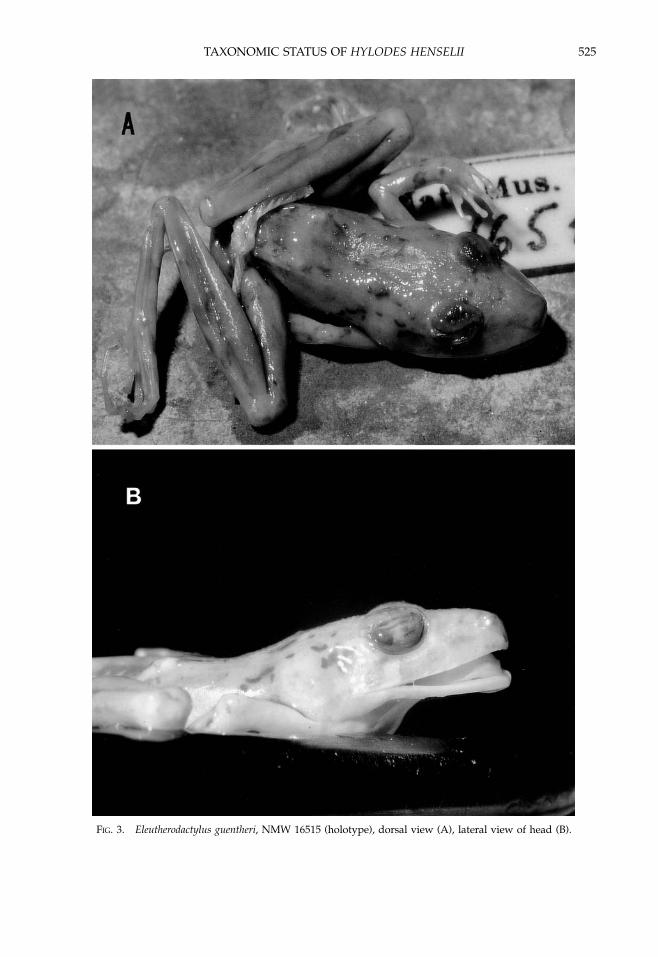
FIG. 3. Eleutherodactylus guentheri, NMW 16515 (holotype), dorsal view (A), lateral view of head (B).
TAXONOMIC STATUS OF HYLODES HENSELII 525

28.4–38.4 (33.4 6 2.39), HL 11.6–16.0 (13.9 6 1.08),HW 9.8–14.1 (11.8 6 0.96), ED 2.3–4.4 (3.3 6 0.45),TD 1.6–2.9 (2.0 6 0.25), EN 3.6–5.3 (4.3 6 0.35), IO4.9–7.7 (5.9 6 0.57), IN 2.5–3.5 (3.0 6 0.26), TL18.1–24.3 (20.7 6 1.53), SL 20.3–26.3 (23.1 61.42),FL 17.2–23.3 (20.3 6 1.43). The measurements incryptic E. guentheri from Rio de Janeiro (seeAppendix 1) are for adult males (N 5 10): SVL20.2–28.3 (24.1 6 2.60), HL 7.9–12.1 (10.1 6 1.33),HW 6.6–9.7 (8.3 6 1.01), ED 1.7–2.7 (2.2 6 0.40),TD 1.3–1.9 (1.5 6 0.18), EN 2.7–3.8 (3.2 6 0.35), IO3.5–5.3 (4.3 6 0.62), IN 1.8–2.7 (2.2 6 0.26), TL12.3–17.5 (14.9 6 1.53), SL 13.5–20.2 (16.4 6 1.90),FL 12.6–17.6 (14.7 6 1.63); for adult females(N 5 8): SVL 31.0–40.6 (34.763.01), HL 12.2–16.3(14.2 6 1.14), HW 11.3–14.5 (12.4 6 1.11), ED 2.8–4.0 (3.460.47), TD 1.7–2.4 (2.160.23), EN 3.8–5.4(4.6 6 0.56), IO 5.4–7.7 (6.3 6 0.71), IN 2.6–4.2 (3.26 0.50), TL 19.7–23.3–24.3 (21.2 6 1.24), SL 21.1–26.1 (23.3 6 1.69), FL 18.4–22.6 (20.9 6 1.54).
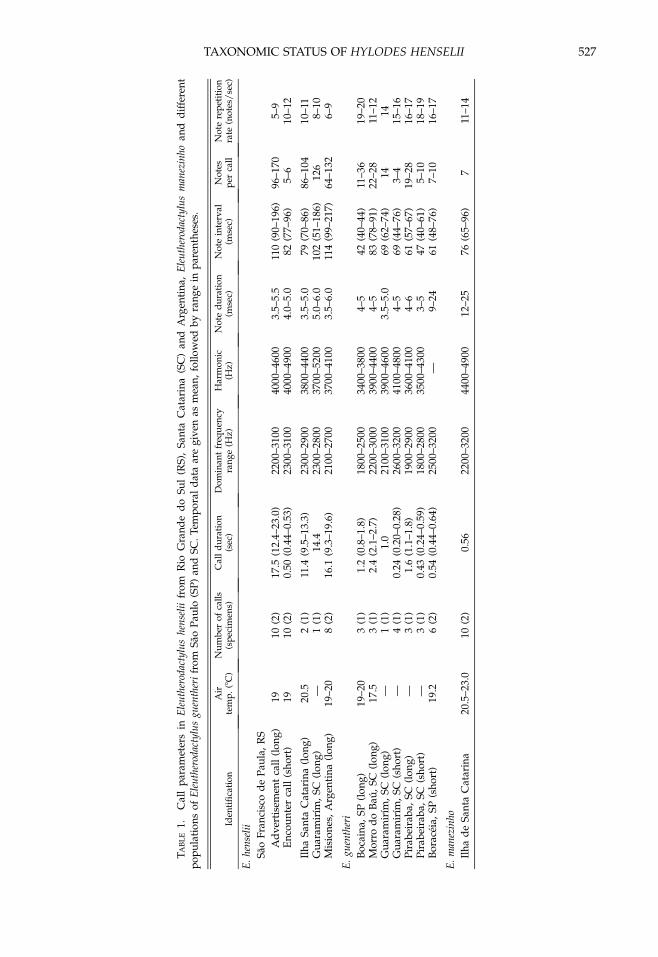
Vocalization.—We recorded two different callsof E. henselii (Table 1; Fig. 4). The prolonged calltype (Figs. 4A, C, 5, 6C; presumably theadvertisement call) was a sporadically given,castanet-like sound beginning quietly, rising inintensity and reaching the maximum energy inthe last third of the call. The whole call lasted10–23 sec and consisted of 86–170 unpulsed notes(each note 3.5-6 msec in duration). Note intervalswere greater at the beginning of the call (about130–230 msec), decreasing towards the end of thecall (70–130 msec). Accordingly, the note repeti-tion rate increased from 5–6 notes per sec to 8–9notes per sec at the end of the call. The second calltype recorded (Figs. 4B, 7C; presumably anaggressive call) lasted 0.5 sec and consisted of5–6 notes with a higher note repetition rate of10–12 per sec. Both call types had a dominantfrequency range from 2100–3100 Hz and pre-sented a weak harmonic structure; anotherfrequency with perceptable energy ranged from4000–4900 Hz and a third harmonic with verylow energy peak was at around 6000 Hz (Fig. 5).There was no frequency modulation.
Call Variation.—Small differences were revealedwhen comparing this advertisement call ofE. henselii from northeastern Rio Grande do Sul(recorded at 198C air temperature) with the callfrom Ilha de Santa Catarina which had (at 20.58Cair temperature) on average a shorter call dura-tion and shorter note intervals (Table 1; notfigured). However, these differences could bedue to temperature effects (temporal parametersdepend on air temperature; Duellman and Trueb,1986), and we refer all citations from southernSanta Catarina (i.e., Ilha de Santa Catarina,Rancho Queimado, Aguas Mornas) to E. henselii.A recording from Guaramirım, northern SantaCatarina (Fig. 6A; available as Eleutherodactylussp1a. at http://www.ra-bugio.org.br/index.html),
and the call from Misiones, Argentina (Fig. 6B), fitthe character states of E. henselii (Fig. 6C)and definitely belong to this species. However,all other calls that are currently assigned to E.guentheri differ significantly from this advertise-ment call and there is also considerable variationbetween different populations (Table 1; Figs. 7, 8).It seems that at least one or two other crypticspecies are involved in the guentheri group.
The advertisement call from Serra da Bocaina(Figs. 6D, 8A), which lies near the type locality ofE. guentheri (about 150 km west of from Rio deJaneiro), should originate from E. guentheri sensustricto. This call is considerably shorter (onmaximum 1.8 sec) than the advertisement call ofE. henselii (Fig. 6C) consisting of only 11–36 noteswith short note intervals of about 40 msec andmuch higher note repetition rates of 19–20 per sec.According to Cochran (1955:273), the call of E.guentheri from Tijuca (Rio de Janeiro) is a ‘‘sort ofquacking, suggesting that of a duck’’ but thisstatement is obviously erroneous. In northernSanta Catarina, there are several recordings thatdiffer from the calls of Serra da Bocaina. The 2–3sec lasting call from Morro do Bau (Fig. 8B) seemsto be identical with the published call fromPirabeiraba (Heyer, 1984) and a call from Guar-amirım (Fig. 8C; available as Eleutherodactylussp25. at http://www.ra-bugio.org.br/index.html).Two different, shorter calls recorded from thesame localities in Pirabeiraba (Fig. 7A) andGuaramirım (Fig. 7B; available as E. guentheri athttp://www.ra-bugio.org.br/index.html) couldbe aggressive calls of the same species becausethey are very similar to the aggressive call of E.henselii (Fig. 7C). However, it seems that all thesevocalizations are not identical with the advertise-ment call of E. guentheri from Serra da Bocaina,which has a much higher note repetition rate.
Another distinct call is that from Boraceia, SaoPaulo (Heyer et al., 1990), which differs sub-stantially in several temporal and spectral char-acteristics (Table 1; Fig. 7D), that is, by havinga longer note duration (each note lasts 10–25msec consisting of 1–2 pulses) and a relativelyhigh dominant frequency, showing notable fre-quency modulation (change in dominant fre-quency from about 2.2 kHz at the beginning ofthe call to 3.2 at the end of the call). Calls ofE. henselii and E. guentheri are never frequencymodulated and have short (3–6 msec lasting)notes without pulse structure (Fig. 5). It isremarkable that the unvouchered recordingsfrom Boraceia are nearly identical to the call ofE. manezinho from Ilha de Santa Catarina (Fig. 7E;see also Castanho and Haddad, 2000). However,further data are needed to make taxonomicalterations and pending further studies allremaining populations should be recognized asE. guentheri.
526 A. KWET AND M. SOLE
TA
BL
E1.
Cal
lp
aram
eter
sin
Ele
uth
erod
acty
lus
hen
seli
ifr
om
Rio
Gra
nd
ed
oS
ul
(RS
),S
anta
Cat
arin
a(S
C)
and
Arg
enti
na,
Ele
uth
erod
acty
lus
man
ezin
hoan
dd
iffe
ren
tp
op
ula
tio
ns
of
Ele
uth
erod
acty
lus
guen
ther
ifr
om
Sao
Pau
lo(S
P)
and
SC
.T
emp
ora
ld
ata
are
giv
enas
mea
n,
foll
ow
edb
yra
ng
ein
par
enth
eses
.
Iden
tifi
cati
on
Air
tem
p.
(8C
)N
um
ber
of
call
s(s
pec
imen
s)C
all
du
rati
on
(sec
)D
om
inan
tfr
equ
ency
ran
ge
(Hz)
Har
mo
nic
(Hz)
No
ted
ura
tio
n(m
sec)
No
tein
terv
al(m
sec)
No
tes
per
call
No
tere
pet
itio
nra
te(n
ote
s/se
c)
E.
hen
seli
i
Sao
Fra
nci
sco
de
Pau
la,
RS
Ad
ver
tise
men
tca
ll(l
on
g)
1910
(2)
17.5
(12.
4–23
.0)
2200
–310
040
00–4
600
3.5–
5.5
110
(90–
196)
96–1
705–
9E
nco
un
ter
call
(sh
ort
)19
10(2
)0.
50(0
.44–
0.53
)23
00–3
100
4000
–490
04.
0–5.
082
(77–
96)
5–6
10–1
2
Ilh
aS
anta
Cat
arin
a(l
on
g)
20.5
2(1
)11
.4(9
.5–1
3.3)
2300
–290
038
00–4
400
3.5–
5.0
79(7
0–86
)86
–104
10–1
1G
uar
amir
ım,
SC
(lo
ng
)—
1(1
)14
.423
00–2
800
3700
–520
05.
0–6.
010
2(5
1–18
6)12
68–
10M
isio
nes
,A
rgen
tin
a(l
on
g)
19–2
08
(2)
16.1
(9.3
–19.
6)21
00–2
700
3700
–410
03.
5–6.
011
4(9
9–21
7)64
–132
6–9
E.
guen
ther
i
Bo
cain
a,S
P(l
on
g)
19–2
03
(1)
1.2
(0.8
–1.8
)18
00–2
500
3400
–380
04–
542
(40–
44)
11–3
619
–20
Mo
rro
do
Bau
,S
C(l
on
g)
17.5
3(1
)2.
4(2
.1–2
.7)
2200
–300
039
00–4
400
4–5
83(7
8–91
)22
–28
11–1
2G
uar
amir
ım,
SC
(lo
ng
)—
1(1
)1.
021
00–3
100
3900
–460
03.
5–5.
069
(62–
74)
1414
Gu
aram
irım
,S
C(s
ho
rt)
—4
(1)
0.24
(0.2
0–0.
28)
2600
–320
041
00–4
800
4–5
69(4
4–76
)3–
415
–16
Pir
abei
rab
a,S
C(l
on
g)
—3
(1)
1.6
(1.1
–1.8
)19
00–2
900
3600
–410
04–
661
(57–
67)
19–2
816
–17
Pir
abei
rab
a,S
C(s
ho
rt)
—3
(1)
0.43
(0.2
4–0.
59)
1800
–280
035
00–4
300
3–5
47(4
0–61
)5–
1018
–19
Bo
race
ia,
SP
(sh
ort
)19
.26
(2)
0.54
(0.4
4–0.
64)
2500
–320
0—
9–24
61(4
8–76
)7–
1016
–17
E.
man
ezin
ho
Ilh
ad
eS
anta
Cat
arin
a20
.5–2
3.0
10(2
)0.
5622
00–3
200
4400
–490
012
–25
76(6
5–96
)7
11–1
4
TAXONOMIC STATUS OF HYLODES HENSELII 527

Geographical Distribution.—Eleutherodactylushenselii occurs in the subtropical Atlantic rainforest and parts of the Araucaria forest domainin southern Brazil and Misiones, Argentina, ataltitudes from about sea level up to 1200 m. Allbibliographical citations for Eleutherodactylusguentheri from northeastern Rio Grande do Sul,that is, municipalities Cambara do Sul, Canela,Caxias do Sul, Gramado, Maquine, Sao Francisco
de Paula, Sao Leopoldo, Terra de Areia, andTorres (Braun and Braun, 1980; Heyer, 1984;Kwet and Di-Bernardo, 1999; Kwet, 2001), referto E. henselii, but the species is not known fromPorto Alegre as stated in Bauer at al. (1995) andFrost (2002). The nearest recordings are fromabout 100 km northeast of Porto Alegre. To ourknowledge, all citations of E. guentheri from thecoastal mountain range of southeastern SantaCatarina (including Ilha de Santa Catarina;Garcia, 1996) also refer to E. henselii. In north-eastern Santa Catarina, there are several speciesinvolved, besides E. henselii and presumably E.manezinho (based on a recording from Guara-mirım named ra-das-pedras, Eleutherodactylussp., which is available at http://www.ra-bugio.org.br/index.html), at least one species whichcould be E. guentheri. All citations of E. guentherifrom the Argentine province Misiones (Carrizoet al., 1989; Lavilla, 1992; Lavilla and Cei, 2001;Lavilla et al., 2002) and from southeastern Para-guay (Frost, 2002) should belong to E. henselii.
FIG. 6 Oscillograms of the advertisement call ofEleutherodactylus henselii from Guaramirım, SantaCatarina (A), Misiones, Argentina (B) and Pro-Mata,Rio Grande do Sul (C) in comparison with theadvertisement call of Eleutherodactylus guentheri fromSerra da Bocaina, Sao Paulo (D).
FIG. 4. Audiospectrograms (above) and oscillograms (below) of two call types of Eleutherodactylus henseliifrom Pro-Mata, Rio Grande do Sul; (A) prolonged advertisement call, (B) short aggressive call. (C) depicts theexpansion of the last 20 notes of the advertisement call, as the audiospectrogram (A) is visually misleading.
FIG. 5. Power spectrum (above), audiospectrogram(middle) and oscillogram (below) of a single note of theadvertisement call of Eleutherodactylus henselii fromPro-Mata.
528 A. KWET AND M. SOLE

At present, there are no locality records for anyE. guentheri cluster member from the westernregions of Rio Grande do Sul (most parts of theAraucaria plateau) and Santa Catarina, but E.henselii (or E. guentheri) occurs in western Parana(Bernarde and Machado, 2001).
Natural History.—Eleutherodactylus henselii liveson mountain slopes with subtropical Atlanticrain forest and in Araucaria forests on the marginof the Araucaria plateau. It is a ground-living frogwith diurnal and nocturnal activity. During thebreeding season from October to January malescall at night, rarely during the day, on the groundor from fallen trees and stones. Calling activityincreases during warm periods with rainfall.Eggs are deposited terrestrially, below trunks orstones, as described for E. guentheri (Lutz, 1946).A big female of E. henselii (MCP 3183, SVL 37.7mm, weigth 4.9 g), captured in activity on 4December 1997 at 16 h, was dissected andcontained 26 large (3 mm diameter) and 45 small(1 mm diameter) yellowish eggs.
DISCUSSION
Steindachner (1864) described Hylodes guen-theri on the basis of a single specimen from Brazilwithout specific locality data. The holotypecollected by Natterer (NHMW 16515; Fig. 3) isa small specimen, presumably a small male ora subadult as stated by Heyer (1984), withfollowing measurements (in millimeters): SVL22.1, HL 9.2, HW 7.4; ED 2.7; TD 1.5; EN 3.2; IO4.0; IN 1.8; TL 14.4; SL 15.5; FL 13.9. Bokermann(1966) indicated that the holotype probably camefrom Rio de Janeiro, which was accepted byHeyer (1984). Steindachner (1864) provided anaccurate description of the holotype and later
described and figured six additional specimensas varieties of E. guentheri (Steindachner, 1867).One of us (AK) examined six individuals de-posited in Vienna that had been collected in thesurroundings of Rio de Janeiro by the NovaraExpedition from 1857–1859 (NMW 4292:1,4292:2, 4294:1, 4294:2) by Natterer (NMW 4296,13281; locality according to the original label:Brazil). These specimens represent the onlymaterial available from this locality and, there-fore, should be identical with the materialdescribed by Steindachner (1867). However, ourexamination revealed that only four specimens(NMW 4292:1, 4292:2, 4296, and 13281) areE. guentheri, whereas NMW 4294:1 and NMW4294:2 have well developed foot webbing, notbelonging to this species.
Hylodes henselii was described by Wilhelm C.H. Peters (1870), based on a specimen collectedby Reinhold Hensel near Porto Alegre in theBrazilian state of Rio Grande do Sul (Fig. 1A).Hensel (1867) provided a short description of thisfrog but did not make taxonomic decisionsbecause the specimen was dead and in poorcondition when found. Boulenger (1882, 1886),who considered H. guentheri Steindachner, 1864,a synonym of Hylodes gollmeri Peters, 1863,placed H. henselii in the synonymy of Hylodesgriseus Hallowell, 1861 (‘‘1860’’). This arrange-ment was followed by Nieden (1923) andMiranda-Ribeiro (1926). Cochran (1955) revali-dated H. guentheri (as E. guentheri) but did notevaluate the status of H. henselii. Therefore, thisspecies continued to be a synonym of Eleuther-odactylus griseus in the species list of Gorham(1966). Bokermann (1966), who listed H. henselii(Peters, 1870) in his catalogue of the typelocalities of Brazilian anurans, confused it withPhysalaemus henselii (Peters, 1872) and put bothtaxa together. However, Physalaemus henselii(originally described as Paludicola henselii), which
FIG. 8. Audiospectrograms (above) and oscillo-grams (below) of the advertisement call of Eleuther-odactylus guentheri from Serra da Bocaina, Sao Paulo(A), Morro do Bau, Santa Catarina (B), and Guara-mirım, Santa Catarina (C).
FIG. 7. Audiospectrograms (above) and oscillo-grams (below) of short call types of members of theEleutherodactylus guentheri cluster from Pirabeiraba,Santa Catarina (A), Guaramirım, Santa Catarina (B),Pro-Mata, Rio Grande do Sul (C), Boraceia, Sao Paulo(D), and of the advertisement call of Eleutherodactylusmanezinho from Ilha de Santa Catarina (E).
TAXONOMIC STATUS OF HYLODES HENSELII 529

is also from Rio Grande do Sul and dedicated toHensel, is a species distinct from E. henselii.
When Savage (1974) allocated H. griseus to thesynonymy of Eleutherodactylus fitzingeri, a MiddleAmerican species, the south Brazilian H. henseliiremained as a valid but uncharacterized species.Lynch (1976), who revised the genus Eleuther-odactylus, assigned H. henselii tentatively to thebinotatus group but stated that, ‘‘according to G.Peters (unpubl. data),’’ its holotype is lost anddata are inadequate to definitely make a taxo-nomic decision. This erroneous communicationof the then curator at the Berlin herpetologicalcollection, Gunther Peters, might be a result ofthe fact that on the label of H. henselii the originalnames had been crossed out and replaced by thename H. griseus. Harding (1983:162) accepted E.henselii as a full species but erroneously quotedSanta Catarina as the type locality. In a revisionof the E. guentheri species cluster, Heyer (1984)placed H. henselii in the synonymy of H. guentheriand designated NMW 16515 as a neotype,following the statement of Lynch (1976) that theholotype of henselii is lost. This arrangement wasaccepted by all subsequent authors (e.g., Frost,1985, 2001; Lavilla, 1992), but our bioacousticdata lead to the conclusion that the recoveredholotype of H. henselii (ZMB 6813, Bauer et al.,1995) belongs to a cryptic species that must beremoved from the synonymy of E. guentheri.
Carrizo et al. (1989) published the first recordof E. guentheri from Misiones, Argentina, basedon three specimens (MACN 33083, MACN33084, CENAI 4708). One of us (AK) examinedthis material. CENAI 4708 (in millimeters: SVL36.4; HL 13.9; HW 13.1; ED 3.9; TD 2.3; IO 2.5; IN2.7; TL 20.5; SL 23.9; FL 22.2) is an adult femalefitting the description of E. guentheri (respectively,E. henselii). Both MACN 33083 (SVL 21.4; HL 8.9;HW 8.2; ED 3.1; TD 1.5; IO 2.2; IN 1.9; TL 14.4; SL14.8; FL 13.6) and MACN 33084 (SVL 24.3; HL8.8; HW 8.8; ED 3.2; TD 1.6; IO 2.0; IN 2.0; TL15.3; SL 16.6; FL 15.4) are adult males, varying insize, pattern and skin texture but seem to belongto the same species. Besides this material, Lavillaet al. (2002) reported two additional specimensfrom Misiones (Iguazu) assessing this species asinsufficiently known in Argentina. Our analysisof acoustic recordings from Misiones (Fig. 6B)assigned to E. guentheri showed that they actuallybelong to E. henselii.
The recording from Boraceia (Heyer et al.,1990), currently assigned to E. guentheri, isdistinct from that of E. guentheri from near thetype locality, which suggests that it may repre-sent another species. The overall similarity of theBoraceia call with that of E. manezinho couldeither mean that both recordings belong to thisspecies or that they represent two different butnearly related species of the guentheri cluster. The
acoustic data contradict the morphological find-ings (Garcia, 1996; Castanho and Haddad, 2000),which suggests that E. manezinho is a member ofthe lacteus group of Lynch (1976). However,although not being a cryptic species, E. manezinhois morphologically so similar to E. guentheri that,in a large series of Eleutherodactylus from Ilhade Santa Catarina labeled as E. guentheri(BGSS 4381-94), we identified two specimens(BGSS 4382, 4389) belonging to E. manezinho.In his study on E. guentheri and related species,Heyer (1984) described the monophyletic guen-theri cluster, consisting of Eleutherodactylus epi-pedus, Eleutherodactylus erythromerus, E. gualteri,E. guentheri, Eleutherodactylus nasutus, and Eleu-therodactylus oeus. Eleutherodactylus izecksohni waslater included by Caramaschi and Kisteumacher(1988), and E. henselii also belongs to this cluster.However, there are more calls referred to E.guentheri than available names and it seems thatat least two or three other cryptic species areinvolved. These results confirm Heyer et al.(1996), who described a cryptic Leptodactylusspecies based solely on acoustic data and statedthat sibling species pairs might be more commonthan currently acknowledged. Further studiesare needed to clarify the distribution of E. henselii,E. guentheri and related species.
Acknowledgments.—W. Engels, M. Di-Bernardo,and A. Zillikens provided the opportunity forour field studies in Brazil. W. R. Heyer and M.Sanchez made comments on the manuscript. Foraccess to material under their care, we thank P.Muller and M. Monzel (BGSS), G. Carrizo(MACN), M. Di-Bernardo (MCP), J. P. PombalJr. (MNRJ), F. Tiedemann (NMW), W. RonaldHeyer (USNM), and R. Gunther (ZMB). W. R.Heyer, A. S. Rand, and D. Baldo kindly allowedus to use their tape recordings. This study wassupported by German grants of the DAAD,LGFG, DLR-SHIFT, and DLR-FAPERGS.
LITERATURE CITED
BAUER, A. M., R. GUNTHER, AND M. KLIPFEL. 1995. TheHerpetological Contributions of Wilhelm C. H.Peters (1815–1883). Thomson-Shore, Ithaca, NY.
BAUMANN, F. 1912. Brasilianische Batrachier des BernerNaturhistorischen Museums nebst Untersuchungenuber die geographische Verbreitung der Batrachierin Brasilien. Zoologisches Jahrbuch fur Systematik,Abteilung Systematik, Geographie und Biologie derTiere 33:87–172.
BERNARDE, P. S., AND R. A. MACHADO. 2001. Riqueza deespecies, ambientes de reproducao e temporadade vocalizacao da anurofauna em Tres Barrasdo Parana, Brasil (Amphibia: Anura). Cuadernosde Herpetologia 14:93–104.
BOKERMANN, W. C. A. 1966. Lista Anotada das Local-idades Tipo de Anfıbios Brasileiros. Servico deDocumentacao-RUSP, Sao Paulo, Brazil.
530 A. KWET AND M. SOLE

BOULENGER, G. A. 1882. Catalogue of the BatrachiaSalientia s. 2nd ed. Ecaudata in the Collection of theBritish Museum, London.
———. 1886. A synopsis of the reptiles and batra-chians of the province Rio Grande do Sul, Brazil.Annals and Magazine of Natural History, series 5,18:423–445.
BRAUN, P. C., AND C. A. S. BRAUN. 1980. Lista previa dosanfıbios do estado do Rio Grande do Sul, Brasil.Iheringia, Serie Zoologia 56:121–146.
CARAMASCHI, U., AND G. KISTEUMACHER. 1988. A newspecies of Eleutherodactylus (Anura: Leptodactyli-dae) from Minas Gerais, southeastern Brazil.Herpetologica 44:423–426.
CARRIZO, G., E. V. DE OLMEDO, AND J. SOROKA. 1989. Unnuevo leptodactilido (Amphibia: Anura) para laArgentina, Eleutherodactylus guentheri Steindachner,1864. Boletın de la Asociacion HerpetologicaArgentina 5:10–11.
CASTANHO, L. M., AND C. F. B. HADDAD. 2000. Newspecies of Eleutherodactylus (Amphibia: Leptodacty-lidae) from Guaraquecaba, Atlantic Rain Forest ofBrasil. Copeia 2000:777–781.
COCHRAN, D. M. 1955. Frogs of southeastern Brazil.United States National Museum Bulletin 206:1–424.
DUELLMAN, W. E., AND L. TRUEB. 1986. Biology of Amphi-bians. John Hopkins Univ. Press, Baltimore, MD.
FROST, D. R. (ED.) 1985. Amphibian Species of theWorld—A Taxonomic and Geographical Reference.Allen Press, Lawrence, KS.
———. 2002. Amphibian Species of the World: Anonline reference. V2.21 (15 July 2002). http://research.amnh.org/herpetology/amphibia/index.html.
GARCIA, P. C. A. 1996. Nove especie de EleutherodactylusDumeril & Bibron, 1891 do estado de SantaCatarina, Brasil (Amphibia; Anura; Leptodactyli-dae). Biociencias 4:57–68.
GORHAM, S. W. 1966. Liste der rezenten Amphibien undReptilien. Ascaphidae, Leiopelmatidae, Pipidae,Discoglossidae, Pelobatidae, Leptodactylidae, Rhi-nophrynidae. Das Tierreich 85. De Gruyter, Berlin,Germany.
HARDING, K. A. 1983. Catalogue of New WorldAmphibians. Pergamon Press, Oxford.
HENSEL, R. 1867. Beitrage zur Kenntniss der Wirbelth-iere Sudbrasiliens. Wiegmanns Archiv fur Natur-geschichte 33:120–162.
HEYER, W. R. 1984. Variation, systematics, and zooge-ography of Eleutherodactylus guentheri and closelyrelated species (Amphibia: Anura: Leptodactyli-dae). Smithsonian Contributions to Zoology402:1–42.
HEYER, W. R., A. S. RAND, C. A. G. CRUZ, O. L. PEIXOTO,AND C. E. NELSON. 1990. Frogs of Boraceia. Arquivosde Zoologia 31:230–410.
HEYER, W. R., J. M. GARCIA-LOPEZ, AND A. J. CARDOSO.1996. Advertisement call variation in the Leptodac-tylus mystaceus species complex (Amphibia: Lep-todactylidae) with a description of a new siblingspecies. Amphibia-Reptilia 17:7–31.
KWET, A. 1999. Die Pfeiffrosche im Araukarienwald-schutzgebiet Pro-Mata. Elaphe 7:92–100.
———. 2001. Frosche im brasilianischen Araukarien-wald. Anurengemeinschaft des Araukarienwaldesvon Rio Grande do Sul: Diversitat, Reproduktion
und Ressourcenaufteilung. Natur und Tier-Verlag,Munster, Germany.
KWET, A., AND M. DI-BERNARDO. 1999. Pro-Mata:Anfıbios-Amphibien-Amphibians. EDIPUCRS,Porto Alegre, Brazil.
LAVILLA, E. O. 1992. Tipos portadores de nombre ylocalidades tipo de Anfibios de Argentina. Actazoologica Lilloana 42:61–101.
LAVILLA, E. O., AND J. M. CEI. 2001. Amphibians ofArgentina. A Second Update, 1987–2000. MuseoRegionale Scienze Naturali, Torino, Italy.
LAVILLA, E. O., J. S. BARRIONUEVO, AND J. D. BALDO. 2002.Los anfıbios insuficientemente conocidos en Argen-tina. Una reevaluacion. Cuadernos de Herpetologıa16:99–118.
LUTZ, B. 1946. The development of Eleutherodactylusguentheri Stdnr. 1864 (Salientia). I. Notes on theexternal development of Eleutherodactylus guentheriStdnr. 1864. Boletim do Museu Nacional Rio deJaneiro, Zoologia 71:1–12.
LYNCH, J. D. 1976. The species group of the SouthAmerican frogs of the genus Eleutherodactylus(Leptodactylidae). Occasional Papers of the Muse-um of Natural History, Univ. of Kansas 61:1–24.
———. 2001. Four osteological synapomorphies with-in Eleutherodactylus (Amphibia: Leptodactylidae)and their bearing on subgeneric classifications.Revista de la Academia Colombiana de Ciencias25:127–136.
LYNCH, J. D., AND W. E. DUELLMAN. 1997. Frogs of thegenus Eleutherodactylus (Leptodactylidae) in west-ern Ecuador: systematics, ecology, and biogeogra-phy. Natural History Museum, Univ. of Kansas,Special Publication 23:1–236.
MIRANDA-RIBEIRO, A. 1926. Notas para servirem aoestudo dos Gymnobatrachios (Anura) Brasileiros.Arquivos do Museu Nacional do Rio de Janeiro27:1–227.
NIEDEN, F. 1923. Anura I. Subordo Aglossa undPhaneroglossa. Sectio 1. Arcifera. Das Tierreich, 46.Lieferung. De Gruyter and Co., Berlin, Germany.
PETERS, W. 1870. Uber neue Amphibien des Koniglichzoologischen Museums. Monatsberichte der BerlinerAkademie der Wissenschaften 1870:641–652.
SAVAGE, J. M. 1974. On the leptodactylid frog calledEleutherodactylus palmatus (Boulenger) and thestatus of Hylodes fitzingeri O. Schmidt. Herpetolog-ica 30:289–299.
STEINDACHNER, F. 1864. Batrachologische Mittheilungen.Verhandlungen der kaiserlich-koniglichen zoolog-ischen botanischen Gesellschaft 14:239–288.
———. 1867. Reise der osterreichischen FregatteNovara um die Erde in den Jahren 1857, 18581859, unter den Befehlen des Commodore B. vonWullerstorf-Urbair. Zoologischer Theil Bd. 1. Am-phibien. Kaiserlich-Konigliche Staatsdruckerei,Wien, Germany.
Accepted: 6 April 2005.
APPENDIX 1
Specimens Examined
Eleutherodactylus henselii: Brazil: Rio Grande do Sul:near Porto Alegre (ZMB 6813, holotype); Canela (SMNS
TAXONOMIC STATUS OF HYLODES HENSELII 531

9125); Maquine (MCP 6192); Sao Francisco de Paula:Pro-Mata (MCP 1752, 2452–53, 2769, 3182–83, 3416,3454, 3486, 3674, 3678–80, 3682–83, 3691–92, 3843, 3875,4938, 6639–40, 6649–53, 6655, 6680, 6686, 7198, 7201;SMNS 9004–06, 9507); Sao Leopoldo (BGSS 5665–66);Torres (MCP 324–25, 327). Santa Catarina: Florianop-olis: Ilha de Santa Catarina (BGSS 4381, 4383–88, 4390–94; MCP 1704–06, 8281-82); Rancho Queimado (MCP1707); Teresopolis (5 Quecaba, Aguas Mornas) (NMW4293:1–8, 15826:1–61, 19418; USNM 137696–99). Ar-gentina: Misiones: General Belgrano: Arroyo Monyolito(CENAI 4708); Dos de Mayo (MACN 33083–84).
Eleutherodactylus guentheri: Brazil: Santa Catarina:Blumenau (NMW 15826:62); Ilhota: Morro do Bau(MCP 1356, 1708–11); Pirabeiraba (USNM 235723–26).Parana: Guaratuba (MCP 7683–87); Pinhao (MCP 2017,2023, 2043). Minas Gerais: Belmiro Braga (MNRJ 27508);Monte Verde (MNRJ 30365–70). Sao Paulo: Jureia
(USNM 303306–09); Ribeirao Branco (MNRJ 18706–07); Salesopolis: Boraceia (USNM 235630–33); Sao Josedo Barreiro (USNM 235699–702); Ilha de Sao Sebastiao(BGSS 4348–80, MNRJ 23673–76). Rio de Janeiro (MNRJ1835, 18706–07, 23673–76, 37508, 30365–67, 31665–66;NMW 4292:1–2, 4296, 13281); Angra dos Reis (USNM096505–08); Rio dos Macacos (NMW 16515, holotype);Teresopolis (MNRJ 1835); Tijuca (USNM 097404–07).
Eleutherodactylus binotatus: Brazil: Rio Grande do Sul:Torres (MCP 329). Santa Catarina: Florianopolis: Ilha deSanta Catarina (MCP 6368–69, 6376, 6414, 6441).Parana: Guaratuba (MCP 7649). Sao Paulo: Ilha deSao Sebastiao (BGSS 4330–44).
Eleutherodactylus manezinho: Brazil: Santa Catarina:Florianopolis: Ilha de Santa Catarina (BGSS 4382,4389; MCP 6364–67, 6413, 6425–29, 6442, 6445, 7646–48, 7695).
532 A. KWET AND M. SOLE





























