Embed Size (px)
Citation preview

Konstanz SymposiumChemical Biology
– ks-cb.com
8–9 October 2015Universität Konstanz, A 704
Conference Booklet

We thank our sponsors and cooperation partners for their support:

Content
OrganisationKonstanz Research School Chemical Biology (KoRS-CB)Universität KonstanzFach 63078457 Konstanz – Germany
+49 7531 88-2237/[email protected]: L 901, L 902
Prof. Dr. Andreas Marx/CoordinatorDr. Heike Brandstädter/General Manager
– ks-cb.com
Programme 2
Location Map 4
Speakers & Abstracts 5
Short Talks 33
Notes 46

2
Thursday, 8 October 2015
09.00–09.10 Welcome: Andreas Marx
Session I Chair: Elisa May09.10–09.50 Benjamin G. Davis, University of Oxford: Sugars & Proteins: Towards a Synthetic Biology09.50–10.30 Ernst H.K. Stelzer, Goethe-Universität Frankfurt am Main: The Light Sheet as a Basis for Perfectly Corrected,
Sensorbased and Fully Automated Microscopy
10.30–11.00 Coffee Break
11.00–11.40 Chris Meier, University of Hamburg: Developing Prodrugs of Antivirally Active Nucleoside Triphosphates
– Against all Odds – It Works!11.40–12.20 Stephen Taylor, University of Manchester: How do Cells React When Mitosis Goes Wrong?12.20–13.00 Short Talks (Leila Motiei, Saurav Malhotra, Sabrina Weickert, Franziska Doll)
13.00–14.00 Lunch
Session II Chair: Daniel Summerer14.00–14.40 Georg Pohnert, Friedrich Schiller University Jena: Exploration of Chemical Signals from Marine Plankton that Structure the Marine Environment14.40–15.20 Wenshe Liu, Texas A&M University: Use Structurally Defined Nucleosomes to Profile Epigenetic Targets of Histone
Lysine Deacylases and Demethylases 15.20–15.40 Short Talks (Grzegorz Kubik, Sandra Hess)
15.40–16.10 Coffee Break
Session III Chair: Karin Hauser16.10–16.50 Dirk Trauner, Ludwig-Maximilians-Universität München: Controlling Biological Function with Photopharmacology16.50–17.30 Sheena E. Radford, University of Leeds: Biophysical Studies of Amyloid Formation and its Inhibition17.30–17.50 Short Talks (Andreas Groß, Nassima Meriem Gueddouda)
18.00 Dinner & Postersession
Programme

3
Friday, 9 October 2015
Session IV Chair: Thomas U. Mayer09.10–09.50 Tom W. Muir, Princeton University: ‘Houdini’ Proteins: Discovery and Applications of Ultrafast Inteins09.50–10.30 Erich A. Nigg, Biozentrum, University of Basel: Control of Mitotic Chromosome Segregation by Cooperating Protein Kinases
10.30–11.00 Coffee Break
11.00–11.40 Short Talks (Christoph Paone, Nemanja Milisavljevic, Michele Felletti, Xiaohui Zhao)
11.40–13.00 Lunch
Session V Chair: Andreas Marx13.00–13.40 Elena Dubikovskaya, Ecole polytechnique fédérale de Lausanne: Real-Time Imaging of Enzymatic Activity and Metabolite Absorption13.40–14.20 Petra Schwille, Max Planck Institute of Biochemistry, Martinsried: Synthetic Biology of Cell Division14.20–15.00 Benjamin F. Cravatt, The Scripps Research Institute, La Jolla: Activity-Based Proteomics – Applications for Enzyme and Inhibitor Discovery15.00 Concluding Remarks & Poster Prize Awarding

4
Talks – Lecture Hall A 704Lunch – Mensa, K 6Dinner & Postersession – above the Mensa, K 7Coffee Breaks – A 6
S
ZZS
PZ
T
PM
ML
Y
W
R
L K
H
A B
V
J
G
D
F
KH
E
C
S5
U
N
Parkplatz NordP
Parkplatz OstP
Parkplatz SüdP
P
X5
5
Q1
Q2
Fußweg
Eggerhaldestraße
Fußw
eg
Fußw
eg
Fußweg
Universitätsstraße
Eichbergstraße
Lecture Hall A 704
K/Mensa
Location Map

5
Speakers & Abstracts
Benjamin F. Cravatt The Scripps Research Institute, La Jolla 6
Benjamin G. Davis University of Oxford 8
Elena Dubikovskaya Ecole polytechnique fédérale de Lausanne 10
Wenshe Liu Texas A&M University 12
Chris Meier University of Hamburg 14
Tom W. Muir Princeton University 16
Erich A. Nigg Biozentrum, University of Basel 18
Georg Pohnert Friedrich Schiller University Jena 20
Sheena E. Radford University of Leeds 22
Petra Schwille Max Planck Institute of Biochemistry, Martinsried 24
Ernst K.H. Stelzer Goethe-Universität Frankfurt am Main 26
Dirk Trauner Ludwig-Maximilians-Universität München 28
Stephen Taylor University of Manchester 30

6
Benjamin Cravatt was born 1970 in Houston, Texas. He graduated in 1992 from Stanford University with a Bachelor of Science in Biological Sciences (with Honors) and a Bachelor of Arts in History and finished his Ph.D. in 1996 at The Scripps Research Institute (TSRI). From 1996 to 2001 he functioned as an assis-tant professor at The Scripps Research Institute (The Skaggs Institute for Chemical Biology) in the Depart-ment of Cell Biology. Additionally, he was also an assis-tant professor in the Department of Chemistry from 2000 to 2001 at the same institute. In 2001 he became an associate professor with tenure at the Departments of Cell Biology and Chemistry at TSRI, until 2004, and later a professor in the Departments of Cell Biology and Chemistry from 2004 to 2007. From 2002 to 2010 he was the director of the Helen L. Dor-ris Child and Adolescent Neuro-Psychiatric Disorder Institute.
Benjamin Cravatt is currently professor and chair of the Department of Chemical Physiology and the Norton B. Giulia Chair in Chemical Biology at The Scripps Research Institute. He is a member of the editorial boards of the following journals: Bioorganic Chemistry, Bioorganic Medicinal Chemistry & Bioorganic Medicinal Chemistry Letters, Chemistry and Biology, Molecular Biosystems, ChemBioChem, Curr. Opin. Chem. Biol., ChemMedChem, ACS Chemical Biology, and Cell. Addi-tionally, he serves as a member of the editorial advisory board for Biochemistry and as associate editor for JACS. In 2014, he was elected a member of the Natio-nal Academy of Sciences.
Benjamin F. Cravatt
ContactDepartment of Chemical PhysiologyThe Skaggs Institute for Chemical BiologyThe Scripps Research Institute, SR10710550 North Torrey Pines RoadLa Jolla, CA [email protected] – scripps.edu/cravatt

7
Genome sequencing projects have revealed that euka-ryotic and prokaryotic organisms universally possess a huge number of uncharacterized enzymes. The functio-nal annotation of enzymatic pathways thus represents a grand challenge for researchers in the genome era. To address this problem, we have introduced chemical proteomic and metabolomic technologies that globally profile enzyme activities in complex biological systems. These methods include activity-based protein profiling (ABPP), which utilizes active site-directed chemical probes to determine the functional state of large
numbers of enzymes in native proteomes. In this lec-ture, I will describe the application of ABPP and com-plementary proteomic methods to discover and functio-nally annotate enzyme activities in mammalian physiology and disease, with a special emphasis on enzymes involved in metabolic disorders. I will also pre-sent competitive ABPP platforms for developing selec-tive inhibitors for these enzymes and discuss ongoing challenges that face researchers interested in assig-ning protein function using chemoproteomic methods.
Activity-Based Proteomics – Applications for Enzyme and InhibitorDiscovery
Benjamin F. Cravatt, The Scripps Research Institute, La Jolla

8
Ben Davis got his B.A. (1993) and D.Phil. (1996) from the University of Oxford. During this time he learnt the beauty of carbohydrate chemistry under the supervi-sion of Professor George Fleet. He then spent 2 years as a postdoctoral fellow in the laboratory of Professor Bryan Jones at the University of Toronto, exploring pro-tein chemistry and biocatalysis. In 1998 he returned to the U.K. to take up a lectureship at the University of Durham. In the autumn of 2001 he moved to the Dyson Perrins Laboratory, University of Oxford and received a fellowship at Pembroke College, Oxford. He was pro-moted to Full Professor in 2005. His group’s research centres on the chemical under-standing and exploitation of biomolecular function (Synthetic Biology, Chemical Biology and Chemical Medicine), with an emphasis on carbohydrates and pro-teins. In particular, the group’s interests encompass synthesis and methodology; target biomolecule synthe-sis; inhibitor/probe/substrate design; biocatalysis;
enzyme & biomolecule mechanism; biosynthetic pathway determination; protein engineering; drug deli-very; molecular biology; structural biology; cell biology; glycobiology; molecular imaging and in vivo biology. He sits on the Editorial/Editorial Advisory Boards of Chemical Biology and Drug Design (2006-), the Bio-chemical Journal (Advisory Board 2002–2005, Edito-rial Board 2009–), Chemical Science (2010–2012, 2015–) and ChemBioChem (2011–). He was the Edi-tor-in-Chief of Bioorganic Chemistry (2011–2013) and an Associate Editor of Chemical Science (2012–14). He is the Editor-in-Chief of Current Opinion in Chemical Biology (2011–) and a Senior Editor for ACS Central Science (2014–). In 2005 he was elected the UK representative and Secretary (2005–2013) of the European Carbohydrate Organisation and from 2011–2014 the President of the RSC Chemical Biology Division.
Benjamin G. Davis
ContactUniversity of Oxford · Chemistry Research · LaboratoryMansfield Road · Oxford, OX1 3TAUnited [email protected]– users.ox.ac.uk/~dplb0149

9
Sugars and Post-Translational Modifications are critical biological markers that modulate the properties of pro-teins. Our work studies the interplay of proteins, sugars and modifications. Synthetic Biology’s development at the start of this century may be compared with Synthe-tic Organic Chemistry’s expansion at the start of the last; after decades of isolation, identification, analysis and functional confirmation the future logical and free-ranging redesign of biornacromolecules offers tantali-zing opportunities. This lecture will cover emerging areas in our group in chemical protein construction with an emphasis on new bond-forming processes compa-tible with biology:
(i) New methods: Despite 80-years-worth of non-spe-cific, chemical modification of proteins, precise methods in protein chemistry remain rare. The develop-ment of efficient, complete, cherno- and regio-selec-tive methods, applied in benign aqueous systems to redesign the structure and function of proteins both in vitro and in vivo will be presented. (ii) ‘Synthetic Biologics’ and their applications: biomi-micry; functional recapitulation; drug delivery; selective protein degradation; nanomolar inhibitors of bacterial interactions; gene delivery vehicles; radio-dose delivery vehicles; probes of in vivo function and non-invasive presymptopmatic disease diagnosis.
Sugars & Proteins: Towards a Synthetic Biology
Benjamin G. Davis, Department of Chemistry, University of Oxford

10
Elena Dubikovskaya has been appointed Tenure Track Assistant Professor of bio-organic chemistry at the School of Basic Sciences (FSB). She received her MS degree from University of Central Florida under super-vision of D. Howard Miles in the field of medicinal che-mistry of natural products. She then continued her PhD studies in the field of medicinal chemistry and drug delivery in the group of Professor Paul Wender at Stan-ford University. After graduation with a PhD degree in 2008, she moved to University of California at Berkeley where she did her postdoctoral studies in the field of Chemical Biology in the group of Carolyn Bertozzi.
Professor Dubikovskaya is an advocate of an interdisci-plinary approach, combining synthetic chemistry, opti-cal imaging, and an understanding of cellular functions at molecular level to find solutions to fundamental pro-blems in biology and medicine. This scientist has deve-loped, in particular, several new non-invasive imaging techniques that allow studies of molecular signatures of cancer and metabolic diseases. She will perform her research work in the context of the new Chair in biologi-cal chemistry, at EPFL’s Institute of Chemical Sciences and Engineering.
Elena Dubikovskaya
ContactEcole polytechnique fédérale de LausanneInstitut des sciences et ingénierie chimiquesEPFL SB ISIC LCBIMBCH 5307 (Bâtochime)CH-1015 [email protected]– lcbim.epfl.ch/dubikovskaya

11
Detection and quantification of fatty acid fluxes in ani-mal model systems following physiological, pathologi-cal, or pharmacological challenges is key to our under-standing of complex metabolic networks as these macronutrients also activate transcription factors and modulate signaling cascades including insulin sensiti-vity. To enable noninvasive, real-time, spatiotemporal quantitative imaging of fatty acid fluxes in animals, we created a bioactivatable molecular imaging probe based on long-chain fatty acids conjugated to a repor-ter molecule (luciferin). We show that this probe faith-fully recapitulates cellular fatty acid uptake and can be used in animal systems as a valuable tool to localize and quantitate in real time lipid fluxes such as intestinal fatty acid absorption and brown adipose tissue activation. This imaging approach should further our
understanding of basic metabolic processes and pathological alterations in multiple disease models.Proteases regulate most physiological processes by controlling activation, synthesis and turnover of prote-ins. However, quantification of their activity in living organisms has been hampered by lack of suitable research tools. The discovery of bioorthogonal reac-tions has had a tremendous impact on chemical biology, allowing the study of numerous biological processes directly in very complex biological systems such as living animal. Here we describe how novel “split lucife-rin” ligation reaction has important implications for real-time non-invasive imaging of multiple biological pro-cesses and enables sensitive detection of activity of several proteases.
Real-Time Imaging of Enzymatic Activity and Metabolite Absorption
Elena Dubikovskaya, Ecole polytechnique fédérale de Lausanne, Institut des sciences et ingénierie chimiques

12
Wenshe Liu, born 1977 in Anhui province of China, stu-died chemistry at the Peking University (Beijing, China) from 1996 to 2000. He graduated with a PhD degree from Prof. Michael Toney’s group at University of Cali-fornia, Davis in 2005 in the field of biological chemistry. Afterwards, he joined Professor Peter Schultz’s group at Scripps Research Institute at La Jolla as a postdoc-toral fellow in developing novel methods for the genetic incorporation of non-canonical amino acids into prote-ins in both bacteria and eukaryotes. In August 2007, he started his own independent research position as an assistant professor at Texas A&M University.
Currently, Wenshe Liu is a tenured associate professor and is holding the inaugural Emile and Marta Schwei-kert Professorship in Chemistry at Texas A&M Univer-sity. The research of Wenshe Liu centres on two fronts. One is to develop methods for the synthesis of nucle-osomes with defined posttranslational modifications for understanding epigenetic regulation and the other is to develop novel approaches for the synthesis of peptide and small molecule libraries for high-throughput drug discovery.
Wenshe Liu
ContactDepartment of ChemistryTexas A&M UniversityCollege Station, TX [email protected]– chem.tamu.edu/rgroup/liu

13
As the basic building blocks of nucleosomes, four his-tone proteins carry a large number of posttranslational modifications that collaborate to influence a multitude of cellular processes. Aberrant histone modifications are linked to numerous disorders. The most broadly studied histone modifications include acetylation and three levels of methylation on the lysine Nε-amine. Other lysine acylation types such as propionylation, butyrylation, crotonylation, malonylation, and succinyla-tion have also been discovered. Histone deacylases (HDACs) and histone lysine demethylases (HKDMs) are two groups of enzymes that catalyze the removal of acylation and methylation, respectively, from histones. Being crucial epigenetic erasers, they are important drug targets for disease intervention. The FDA has approved two HDAC inhibitors for the treatment of cutaneous T-cell lymphoma. More than ten HDAC inhi-bitors and one HKDM inhibitor are in clinical trials for the treatment of various cancers including cervical, ovarian, lung, prostate, and breast cancers, several
types of lymphoma, refractory leukemia, and multiple myeloma. Although HDACs and HKDMs are important drug targets for disease intervention, critical informa-tion such as their epigenetic targets is not very clear. For HDACs, little is known about their targeted histone lysine sites and their activities toward acylation types other than acetylation. For HKDMs, no systematic infor-mation about their targeted lysine sites and targeted methylation levels is available. Since most HDACs and HKDMs are promiscuous enzymes, a clear knowledge of their targeted modification sites and types (or levels) will allow the explicit prediction of histone modifications when they are inhibited or activated; this information will boost the understanding of cellular and pharmaco-logical effects of the therapeutics that target them, facilitating the discovery of new cures for diseases. In this talk, we will describe our most recent efforts in the synthesis of nucleosomes with most lysine modification types and their applications in probing substrate speci-ficities of HDACs and HKDMs.
Use Structurally Defined Nucleosomes to Profile Epigenetic Targetsof Histone Lysine Deacylases and Demethylases
Wenshe Liu, Texas A&M University

14
Chris Meier was born in 1962 in Berlin (Germany). He studied Chemistry at the University of Marburg/Lahn (Germany), where he received his diploma degree in 1987 and his Ph.D. degree in 1989 in Organic Che-mistry with Professor G. Boche. During that time he became interested in Bioorganic Chemistry. During his postdoctoral studies at the Pasteur-Institute in Paris (France) Chris Meier got involved in the chemistry of nucleosides, oligonucleotides and prodrugs and Medi-cinal Chemistry. In 1991 he returned to the University of Frankfurt/Main (Germany) and finished his habilita-tion in 1996. In 1994 he received the Adolf-Messer-Award for interdisciplinary research. In 1997 he became Associate Professor of Organic Chemistry at the University of Würzburg (Germany) and 1999 he moved to the University of Hamburg (Germany) as a Full Professor of Organic Chemistry. 2007 Chris Meier received the William Prusoff-Award for Young Investi-gators given by the International Society for Antiviral
Research. Since 2010 Chris Meier is Vice Dean for Research of the Faculty of Mathematics, Informatics and Natural Sciences of the University of Hamburg and in January 2014 he was elected as Vice Scientific Director of the Center of Structural Systems Biology (CSSB), Hamburg. Also in 2014 Chris Meier was elec-ted Vice-President of the International Society for Nuc-leoside, Nucleotides and Oligonucleotides (IS3NA). During his career he was invited as a guest professor at the Universities of Montpellier, Toulouse and Shanghai. His research interests are Prodrugs-development of nucleosidemono-, di- and triphosphates of antiviral nucleoside analogues, stereoselective synthesis of car-bocyclic nucleoside analogues, DNA-damage induced by arylamine carcinogens, synthesis of phosphate-bridged bioconjugates, fluorescence-labeling of bio-conjugates, new methods for the synthesis of phospho-rylated nucleosides and oligonucleotides.
Chris Meier
ContactOrganic ChemistryDepartment of ChemistryUniversity of HamburgMartin-Luther-King-Platz 6D-20146 HamburgGermany [email protected] – chemie.uni-hamburg.de/oc/meier

15
Over the last decades a variety of nucleoside analogues were applied in antiviral chemotherapy. However, in some cases the antiviral potency of the nucleoside ana-logues, e.g. 2',3'-dideoxy- or 3'-modified-compounds, is limited due to the lack of efficient intracellular phos-phorylation into their triphosphorylated form by host cell kinases. This problem cannot be solved by using the phosphorylated nucleosides because of their very high polarity. An option to overcome this problem is the use of masked nucleotide analogues, which are able to pass the cell membrane and deliver the corresponding nucleotides intracellularly by e.g. chemical or enzymatic hydrolysis.The cycloSal-prodrug system was developed for nucle-oside monophosphates and was also applied to diffe-rent nucleoside analogs. Recently, we reported on a
first successful approach for the delivery of nucleoside diphosphates called DiPPro-approach. Here, we exten-ded our work to the development of nucleoside triphos-phate prodrugs. So far, no example of such a prodrug system has been reported. A number of d4TTP pro-drugs with different aliphatic masking units have been synthesized via two different routes based on phospho-ramidite (Scheme 1) or H-phosphonate chemistry. In addition, a variety of nucleoside analogues have been investigated. The target prodrug compounds were obtained in yields up to 66 %. Chemical hydrolysis stu-dies, enzymatic cleavage in CEM/0 cell extract, primer extension assays, PCR assays, CEM whole-cell incuba-tions and antiviral HIV tests proved the successful deli-very of NTP.
Developing Prodrugs of Antivirally Active Nucleoside Triphosphates– Against all Odds – It Works !
Chris Meier1, Tristan Gollnest1, Tobias Nack1, Chenglong Zhao1 Jan Balzarini2
1 Organic Chemistry, Department of Chemistry, Faculty of Sciences, University of Hamburg, Martin-Luther-King-Platz 6, D-20146 Hamburg, Germany
2 Rega Institute for Medical Research, Katholieke Universiteit Leuven, Minder broedersstraat 10, 3000 Leuven, Belgium

16
Tom W. Muir received his B.Sc. (Hons, 1st class) in Che-mistry from the University of Edinburgh in 1989 and his Ph.D. in Chemistry from the same institute in 1993 under the direction of Professor Robert Ramage. After postdoctoral studies with Stephen B.H. Kent at the Scripps Research Institute, he joined the faculty at the Rockefeller University in New York City in 1996, where he was, until recently, the Richard E. Salomon Family Professor and Director of the Pels Center of Chemistry, Biochemistry and Structural Biology. In 2011, Tom Muir joined the Princeton Faculty as the Van Zandt Williams Jr. Class of ’65 Professor of Chemistry. He is currently Chairman of the Chemistry Department. Tom Muir has won a number of honors for his research, including; the
Burroughs-Welcome Fund New Investigator Award, the Pew Award in the Biomedical Sciences, the Alfred P. Sloan Research Fellow Award, the Leonidas Zervas Award in Peptide Chemistry, the Irving Sigal Award from the Protein Society, the 2008 Vincent du Vigneaud Award in Peptide Chemistry, the 2008 Blavatnik Award from the New York Academy of Sciences, the 2008 Distinguished Teaching Award from The Rockefeller University, the 2012 Jeremy Knowles Award from the Royal Society of Chemistry and a 2013 Arthur C. Cope Scholar Award from the American Chemical Society. Tom Muir is a Fellow of American Association for the Advancement of Science and the Royal Society of Edin-burgh.
Tom W. Muir
ContactDepartment of ChemistryPrinceton UniversityFrick Chemistry LaboratoryPrinceton, NJ [email protected]– chemists.princeton.edu/muir

17
Inteins are auto-processing domains found in orga-nisms from all domains of life. These proteins are con-summate molecular escape artists that spontaneously excise themselves, in a traceless manner, from proteins in which they are embedded. Chemical biologists have long exploited various facets of intein reactivity to modify proteins in myriad ways for both basic biological research as well as therapeutic applications. While many thousands of inteins have been identified at the sequence level, only a handful of these proteins have been characterized in any biochemical detail. Here I
discuss our recent efforts to mine this genomic data-base, leading to the discovery of ultrafast split inteins that ligate proteins together several orders of magni-tude faster that any inteins previously known. I will also discuss the remarkable folding properties of these split proteins, and how this has led to the development of new methods for the manipulation of protein structure in the test tube and in cells. The latter will be highlighted through the ability to manipulate the chemical structure of cellular chromatin.
‘Houdini’ Proteins: Discovery and Applications of Ultrafast Inteins
Tom W. Muir, Princeton University

18
Erich Nigg was born in Switzerland in 1952 and studied Life Sciences at the ETH in Zurich. He completed his PhD in 1980 focusing on the mobility of membrane proteins. He then pursued research at the University of California in San Diego, U.S.A., where he investigated the influence of oncogenic protein kinases on the cyto-skeleton. Subsequently, he worked at several institu-tions throughout Switzerland, notably the ETH, the Swiss Institute for Experimental Cancer Research (ISREC) and the University of Geneva, before he joined the Max Planck Society in Germany, where he led the Department of Cell Biology in Martinsried/Munich from 1999–2009. Since February 2009 Erich Nigg is the director of the Biozentrum of the University of Basel (http://www.biozentrum.unibas.ch/), one of the most prestigious Life Science research and teaching institu-tions in Switzerland. Erich Nigg was awarded several
prizes and is an elected member of the European Mole-cular Biology Organization (EMBO), the German Aca-demy of Sciences, the Academia Europeae, as well as the European Academy of Cancer Sciences. The early work by Erich Nigg encompassed several fields, prima-rily the structure and dynamics of the nuclear envelope, the mechanisms underlying signal transduction from the plasma membrane to the cell nucleus, and the regu-lation of the cell division cycle. At present, the research conducted in the Nigg laboratory aims at elucidating the molecular mechanisms that control chromosome segregation during cell division and the centrosome duplication cycle. Disruption of these processes are widely thought to contribute to the chromosomal insta-bility of tumour cells. Moreover, centrosome dysfunc-tions are implicated in ciliopathies and primary micro-cephaly.
Erich A. Nigg
ContactBiozentrumUniversity of BaselKlingelbergstrasse 50/704056 [email protected]– biozentrum.unibas.ch/de/forschung/gruppen-
plattformen/overview/unit/nigg

19
The error-free segregation of duplicated chromosomes during cell division is critical for the development and health of all organisms. In humans, extra or missing copies of chromosomes (aneuploidies) are common causes of genetic disorders and birth defects, while chromosomal instability (defined as chromosomal aber-rations that change over time) is typical of most can-cers. Chromosomal instability is generally correlated with increased malignancy and likely to favor not only tumor progression but also the emergence of resis-tance to anti-cancer therapy. Many chromosome aber-rations in tumor cells are thought to result from deregu-lation of the molecular machinery that controls chromosome segregation during M phase. This stage of the cell cycle is governed largely by posttranslational mechanisms, notably protein phosphorylation and ubi-quitin-dependent proteolysis. Central to chromosome segregation is the spindle assembly checkpoint (SAC), a surveillance mechanism that monitors the attachment of all mitotic chromosomes to the microtubule-based
spindle apparatus. High up in the hierarchy of SAC sig-naling is the protein kinase Monopolar spindle 1 (Mps1), which controls the recruitment of several evolu-tionarily conserved SAC components to unattached or misaligned kinetochores. Recently, we and others have discovered that the consensus phosphorylation motif of Mps1 is very similar to that of Polo-like kinase 1 (Plk1). This surprising observation raised the tantalizing possi-bility that human Plk1 cooperates with Mps1 in SAC signaling. Our recent work indicates that this is indeed the case. Specifically, we demonstrate that cooperation between Plk1 and Mps1 involves the phosphorylation of at least two kinetochore-associated common subst-rates. As a result, Plk1 activity enhances Mps1 catalytic activity, as well as the recruitment of SAC components to kinetochores. We conclude that Plk1 strengthens the robustness of SAC establishment at the onset of mitosis and supports SAC maintenance during prolon-ged mitotic arrest.
Control of Mitotic Chromosome Segregationby Cooperating Protein Kinases
Erich A. Nigg, Biozentrum, University of Basel

20
Prof. Georg Pohnert is Professor and Director of the Institute for Inorganic and Analytical Chemistry at the Friedrich Schiller University, Germany. He studied Che-mistry in Karlsruhe and pursued his doctoral studies on algal pheromones in the group of Prof. W. Boland in Bonn. In 1997 he joined the Ganem-group at the Cor-nell University as a postdoc working on the biochemical and biophysical characterisation of E. coli receptors. As group leader at the Max-Planck-Institute for Chemical Ecology he addressed algal defence reactions. In 2005, he was appointed as assistant professor at the EPFL, Lausanne and moved in 2007 to the Friedrich Schiller University in Jena where he holds a chair in Bio-organic Analytics. His research focuses on algal defence reactions, cur-rently on chemically mediated interactions in the
plankton, on biofilm signaling (both with a focus on marine diatoms), on macroalgal defense mechanisms and on the oxylipin chemistry of algae and mosses. His interdisciplinary work gives new insights into the chemi-cally mediated species interactions and the function of natural products. The work aims to understand the role of secondary metabolites as mediators of ecological interactions. Isolation, spectroscopy and organic syn-thesis of natural products are important aspects of his work, and the full picture of the role of the compounds can only be obtained if metabolomics, biochemistry and ecology are brought in as well. He is editor of the Roempp online dictionary, associate editor for Botanica Marina and on the editorial board of Marine Drugs, Natural Products Reports and Bioanalytical Reviews.
Georg Pohnert
ContactFriedrich Schiller UniversityDepartment of Bioorganic AnalyticsLessingstr. 807749 [email protected]– chemgeo.uni-jena.de/Pohnertgroup

21
Unicellular algae from the plankton have established means to interact with other organisms in their environ-ment. Especially interactions mediated by chemical compounds have gained a lot of attention during the last years. Algal exudates and metabolites stored in the cells can mediate feeding activity of herbivores and algal/algal interactions but also communication of an alga with the surrounding microbial community. We int-roduce an approach to address such chemically media-ted interactions based on the metabolomic investiga-tion of the cellular and released metabolites of microalgae. These surveys are based on elaborate
mass spectrometric methods and reveal that algae exhibit a high plasticity of metabolite production during their development. Bioassays that are guided by meta-bolomic evidence reveal that these specific compounds can mediate mate finding, chemical defense or algal/algal interactions in a highly dynamic manner. Examples of diatom defense against herbivores, bacteria and of diatom-diatom interactions will be introduced. Conse-quences for future investigations of plankton ecology, algal physiology and chemical interactions are dis-cussed.
Exploration of Chemical Signals from Marine Plankton that Structurethe Marine Environment
Georg Pohnert, Friedrich Schiller University Jena

22
Sheena Radford was born in Altrincham, a market town in Cheshire, UK. She gained a first class degree in Bio-chemistry at the University of Birmingham in 1984, and completed her PhD in Cambridge under the supervision of Professor Richard Perham, FRS in 1987. She began studies of protein folding whilst a research fellow at the University of Oxford. Sheena became a lecturer at the University of Leeds in 1995, progressing to Reader in 1998 and Professor in 2000. She has been the Direc-tor of the Astbury Centre for Structural Molecular Bio-logy since 2012 and was elected Astbury Chair of Bio-physics the next year. She has been awarded the Biochemical Society Colworth Medal, the AstraZeneca prize of the Royal Society of Chemistry and the Protein Society Carl Brändén Award. She was made a Fellow of the Royal Society of Chemistry in 2003, a Member of EMBO in 2007, a Fellow of the UK Academy of Medical Sciences in 2010 and a Fellow of the Royal Society in 2014. Sheena currently runs a large research labora-tory at the University of Leeds where she supervises or co-supervises twelve PhD students and eleven
postdoctoral research fellows. Her research is focussed on fundamental structural molecular biology, specifi-cally the measurement of the conformational dynamics of proteins and the elucidation of the role that these motions play in protein folding and misfolding in health and disease. Working on both soluble and membrane proteins, she and her group are using a wide range of biophysical methods (including NMR, mass spectrome-try and single molecule methods) to determine how soluble and membrane proteins fold in all-atom detail; the mechanisms by which proteins misfold and cause some of the deadliest human diseases. Her group is now exploiting their fundamental insights to develop new routes towards combatting aggregation disease. Sheena has published more than 220 peer-reviewed papers and has spoken at 130 invited lectu-res at large meetings and seminars in more than 20 countries, in addition to more than 170 seminars and meetings in the UK. She also finds time to serve on many national and international committees, and has significant editorial and reviewing roles.
Sheena E. Radford
ContactAstbury BuildingSchool of Molecular and Cellular BiologyUniversity of LeedsLS2 9JT LeedsUnited [email protected]– astbury.leeds.ac.uk/bmbsgi10

23
References: Ion mobility spectrometry mass spectrometry defines the oligomeric inter-
mediates in amylin amyloid formation and the mode of action of inhibitors.
Young, L.M. et al. (2014) J. Am. Chem. Soc., 136, 660–670.
Visualization of transient protein-protein interactions that promote or inhibit
amyloid assembly. Karamanos, T.K. et al. (2014) Mol. Cell, 55, 214–226
Screening and classifying small molecule inhibitors of amyloid formation
using ion mobility spectrometry-mass spectrometry. Young, L.M. et al.
(2015) Nature Chem., 1, 73–81
Amyloid formation involves the polymerisation of prote-ins and peptides into polymers with a cross-ß fold. How amyloid formation causes disease, and identifying the culprit species involved, remain a significant challenge. This results from the complexity of the aggregation pro-cess and the fact that amyloid assembly is initiated by non-native states of proteins that are partially folded or intrinsically disordered. Structure determination is thus difficult, and identifying the interacting surfaces in these transiently formed and dynamic ensembles is challenging. In this lecture I will describe our recent att-empts to discover new insights into how proteins
aggregate into amyloid, and how to prevent cellular dysfunction caused by amyloid assembly/disassembly mechanisms, using a number of different strategies. Specifically, I will show how we have used different bio-physical and biochemical approaches to map the nature of the earliest protein-protein interactions in amyloid assembly and to re-assess the potential role(s) of fibrils in disease. Finally, using a novel screen developed with E.coli we have been able to discover new highly potent inhibitors of aggregation for some of the most highly aggregating protein sequences known.
Biophysical Studies of Amyloid Formation and Its Inhibition
Sheena E. Radford, Astbury Centre for Structural Molecular Biology, University of Leeds, UK

24
Petra Schwille, born 1968 in Sindelfingen (Germany), studied physics and philosophy at the Universities of Stuttgart and Göttingen (Germany), 1987 to 1993. She graduated from Prof. Manfred Eigen’s group at the Max Planck Institute of Biophysical Chemistry (Göttingen) in 1996 in physics with a thesis on Fluorescence Correla-tion Spectroscopy. Afterwards, she joined Professor Watt Webb’s group at Cornell University (Ithaca, USA) as a Feodor Lynen postdoctoral fellow of the Alexander von Humboldt foundation. In 1999 she started an inde-pendent research group at the MPI in Göttingen, spon-sored by a BMBF “Biofuture” startup grant. In 2002
she followed a call of the Dresden University of Techno-logy (Germany) for the chair of biophysics at the BIO-TEC center. In 2011 she accepted the call by the Max Planck Society to become scientific member and direc-tor at the Max Planck Institute of Biochemistry (Mar-tinsried/Munich). In addition to the development of novel ultrasensitive biophysical techniques to study molecular interactions in biological systems, her research goal is to design biological functionality from the bottom-up, in the framework of synthetic biology of minimal systems.
Petra Schwille
ContactMax Planck Institute of BiochemistryDept. Cellular and Molecular BiophysicsAm Klopferspitz 18D-82152 [email protected]– biochem.mpg.de/en/rd/schwille

25
In recent years, biophysics has accumulated an impres-sive selection of novel techniques to analyze biological systems with ultimate sensitivity and precision. Single molecule imaging, tracking and manipulation have ena-bled us to unravel biological phenomena with unprece-dented analytical power, and to come closer to revealing fundamental features of biological self-orga-nization. The power of physics has always been the reductionist approach, i.e. the possibility to define an appropriate subsystem simple enough to be quantita-tively modeled and described, but complex enough to retain the essential features of its real counterpart. Transferring this approach into biology has so far been extremely challenging, because most “modern” biologi-cal systems usually comprise so many modules and elements, many of them still awaiting to be functionally resolved, that it is a risky task to define truly essential
ones. Nevertheless, the strive for identifying minimal biological systems, particularly of subcellular structures or modules, has in the past years been very successful, and crucial in vitro experiments with reduced comple-xity can nowadays be performed, e.g., on reconstituted cytoskeleton and membrane systems. As a particularly exciting example for the power of minimal systems, self-organization of essential proteins of the bacterial cell division machinery could be shown in a simple assay, consisting of only two protein species, an energy source, and a membrane. In my talk, I will discuss some recent results of our work on membrane-based sys-tems, using single molecule optics and biological reconstitution assays. I will further discuss the perspec-tive of assembling a minimal system to reconstitute bacterial cell division.
Synthetic Biology of Cell Division
Petra Schwille, Max Planck Institute of Biochemistry

26
Ernst H.K. Stelzer is Professor for Physical Biology and Advanced Light Microscopy at the Goethe-Universität (Frankfurt am Main, Germany) since March 2011. He concentrates his efforts on applications and further developments of advanced light microscopies in the modern life sciences, working on various aspects and applications of three-dimensional cell biology, lateral root development in Arabidopsis thaliana and the emb-ryogenesis of Tribolium castaneum. From 1987–2011, he was a Scientific Group Leader at the European Molecular Biology Laboratory (EMBL, Heidelberg, Ger-many), most lately in the Cell Biology and Biophysics unit. Ernst Stelzer has been working in physics, optics, biophysics, cell biology, molecular biology, plant biology and developmental biology for more than 30 years. He has contributed to conventional fluorescence
microscopy, confocal fluorescence microscopy, 4Pi- and theta-microscopy, optical tweezers and optical levitation, laser ablation and light sheet-based fluores-cence microscopy. In more general terms, he is interes-ted in developing three-dimensional microscopies that enable observations under close-to-natural conditions as a function of time. Many of his former Diploma and Ph.D. students as well as Postdocs continue to pursue successful academic careers of their own. He has pub-lished more than 250 papers and was granted several patents that secure the intellectual property of com-mercially available optical instruments. Stelzer has received several prices and honors; this January light sheet-based fluorescence microscopy was honored “Method of the Year 2014” by Nature Methods.
Ernst H.K. Stelzer
ContactPhysical Biology (FB 15, IZN, CEF-MC, BMLS)Goethe-Universität60629 Frankfurt am [email protected] – physikalischebiologie.de

27
– researcherid.com/rid/A-7648-2011 or – scholar.google.com/citations?user=
EV5RvqkAAAAJ
In light sheet–based fluorescence microscopy (LSFM), optical sectioning in the excitation process minimizes fluorophore bleaching and phototoxic effects. Since biological specimens survive long-term three-dimensi-onal imaging at high spatiotemporal resolution, light sheet-based microscopes (LSM) have become an indispensable tool in developmental, three-dimensional cell and plant biology. LSFM is based on two main opti-cal paths. The detection path consists of a microscope objective, a spectral filter, a tube lens and a camera. The excitation path is perpendicular to the detection path and directs a light sheet into the side of the speci-men. The thin light sheet and the focal plane of the detection objective overlap. LSFM provides at least three important degrees of freedom, which are usually not available in an epifluorescence microscope: a) the axial and b) lateral locations of the light sheet and c) the axial location of the focal plane. Further, probably less important, degrees of freedom are the tilt and the inc-line of the light sheet. LSFM takes full advantage of modern cameras, massively parallelizing the data acquisition process and recording ten to one hundred images per second with a high dynamic range. LSFM does not rely on traditional features that are required for ergonomic reasons. A powerful multiple-sensors-based image processing pipeline is, therefore, an inher-ent feature. Traditional fluorescence microscopy enforces specimen preparation schemes that rely on hard and flat surfaces. LSFM places the specimen in the center and arranges the optics around it.
Specimens can be prepared in new ways, their three-dimensional integrity is maintained, and they can be used in experiments hitherto regarded as impossible. Fluorescence microscopy has several basic limitations. First, the excitation light is absorbed not only by fluoro-phores but also by many endogenous organic com-pounds, which are degraded much like fluorophores and thus unavailable for vital metabolic processes. Second, the number of fluorophores in any volume ele-ment at any given time is finite, and fluorophores can degrade upon excitation. As a consequence, the num-ber of photons that are retrieved from a fluorophore-labeled specimen is limited. Finally, life on Earth is adapted to the solar flux, which is less than 1.4 kW/m2. This might not be a hard limit, but it indicates that irradi-ance should not exceed 1 nW/µm2 = 100 mW/cm2 when dynamic biological processes are observed. When imaging living biological samples, these challen-ges must be addressed. LSFM is perhaps the best technology we have so far, which makes a sincere and honest effort to address these challenges: 1) it provi-des optical sectioning, 2) a true axial resolution, 3) reduces fluorophore bleaching and 4) phototoxicity at almost any scale, 5) allows one to record millions of pixels in parallel and 6) dramatically improves the viabi-lity of the specimen. Stelzer, E. H.K. (2015). Light-sheet fluorescence microscopy for quantitative biology. Nature Methods, 12(1), 23–27.
The Light Sheet as a Basis for Perfectly Corrected, Sensor-Basedand Fully Automated Microscopy
Ernst H.K. Stelzer, Physical Biology, Goethe-Universität Frankfurt am Main

28
Dirk Trauner was born and grew up in Linz, Austria. After studying biology and biochemistry at the University of Vienna, he joined Professor Johann Mulzer’s group at the Free University of Berlin to pursue natural product synthesis. Subsequently, he became a postdoctoral fel-low with Professor Samuel J. Danishefsky at the Memo-rial Sloan-Kettering Cancer Center in New York City. In 2000, Dirk joined the University of California, Berkeley,
where he rose through the ranks to become an Associ-ate Professor of chemistry (with tenure). In the summer of 2008, he moved to the University of Munich, where he currently resides as a Professor of Chemistry and Chemical Biology. His research interests range from organic synthesis and natural product chemistry to chemical neurobiology, optogenetics and photophar-macology.
Dirk Trauner
ContactFakultät für Chemie und PharmazieLudwig-Maximilians-Universität MünchenButenandtstr. 5–13, Haus F81377 Mü[email protected] – cup.uni-muenchen.de/oc/trauner

29
Light can be used to control biological events with unmatched temporal and spatial precision. A case in point is optogenetics, which is currently revolutionizing neuroscience. Optogenetics relies on natural photore-ceptors that typically employ retinal as the chromo-phore. Recently, the incorporation of synthetic pho-toswitches, such as azobenzenes, into naturally “blind” receptors has been explored as well. These molecules can bind covalently or non-covalently to a wide variety
of proteins, including ion channels, GPCRs, enzymes, molecular motors, and components of the cytoskeleton, effectively turning them into photoreceptors. As such, photoswitchable molecules add another dimension to pharmacology. I will discuss the advantages and disad-vantages of photopharmacology and its potential in biology and medicine, in particular with respect to restoring vision and fighting cancer.
Controlling Biological Function With Photopharmacology
Dirk Trauner, Ludwig-Maximilians-Universität München, Germany
30
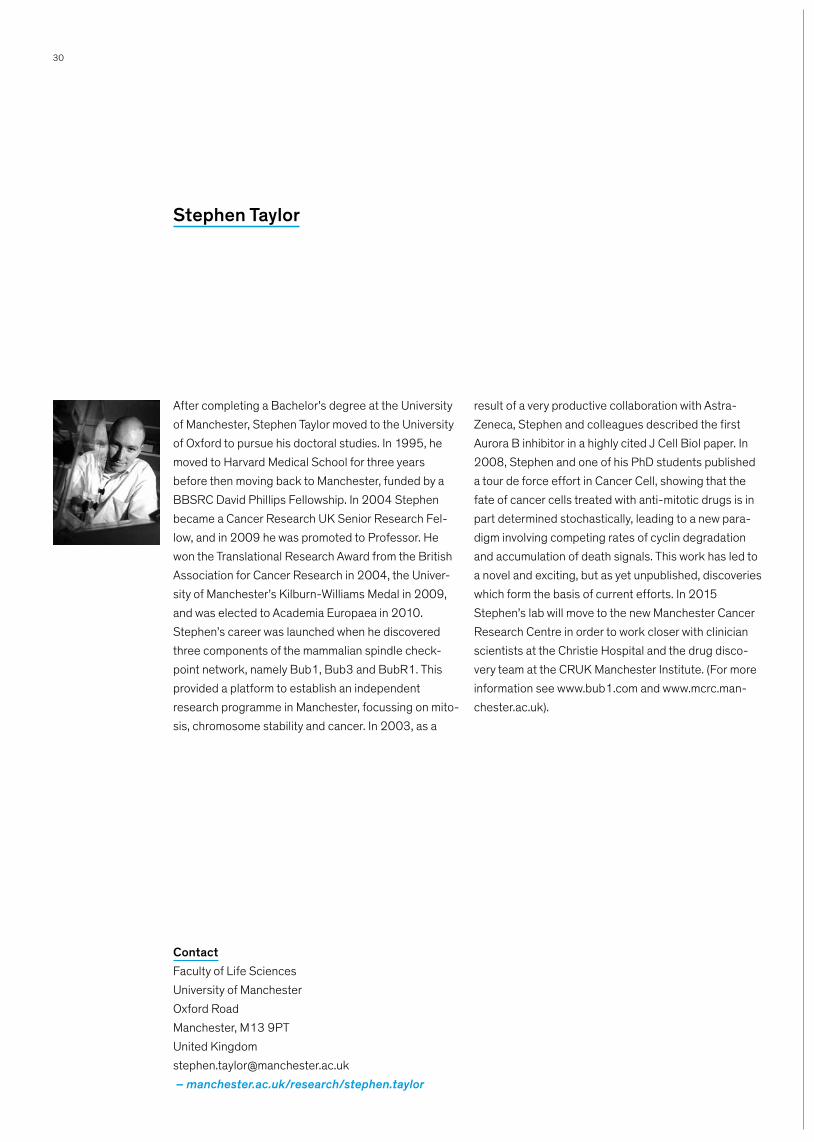
After completing a Bachelor’s degree at the University of Manchester, Stephen Taylor moved to the University of Oxford to pursue his doctoral studies. In 1995, he moved to Harvard Medical School for three years before then moving back to Manchester, funded by a BBSRC David Phillips Fellowship. In 2004 Stephen became a Cancer Research UK Senior Research Fel-low, and in 2009 he was promoted to Professor. He won the Translational Research Award from the British Association for Cancer Research in 2004, the Univer-sity of Manchester’s Kilburn-Williams Medal in 2009, and was elected to Academia Europaea in 2010. Stephen’s career was launched when he discovered three components of the mammalian spindle check-point network, namely Bub1, Bub3 and BubR1. This provided a platform to establish an independent research programme in Manchester, focussing on mito-sis, chromosome stability and cancer. In 2003, as a
result of a very productive collaboration with Astra-Zeneca, Stephen and colleagues described the first Aurora B inhibitor in a highly cited J Cell Biol paper. In 2008, Stephen and one of his PhD students published a tour de force effort in Cancer Cell, showing that the fate of cancer cells treated with anti-mitotic drugs is in part determined stochastically, leading to a new para-digm involving competing rates of cyclin degradation and accumulation of death signals. This work has led to a novel and exciting, but as yet unpublished, discoveries which form the basis of current efforts. In 2015 Stephen’s lab will move to the new Manchester Cancer Research Centre in order to work closer with clinician scientists at the Christie Hospital and the drug disco-very team at the CRUK Manchester Institute. (For more information see www.bub1.com and www.mcrc.man-chester.ac.uk).
Stephen Taylor
ContactFaculty of Life SciencesUniversity of Manchester Oxford RoadManchester, M13 9PTUnited [email protected] – manchester.ac.uk/research/stephen.taylor

31
Chemical biology tools open up excellent opportunities to study various aspects of cell biology, ranging from cell cycle control and mitosis to signaling pathways and gene expression; and from migration and wound healing, to apoptosis and autophagy. Indeed, there is a long and rich history of small, drug-like molecules fuel-ling discovery in the biosciences. For example, studies with colchicine led to the discovery of tubulin, and phal-loidin has long been used to stain the actin cytoskele-ton. The quest for new therapeutics has also provided the research community with excellent chemical biology tools. For example, efforts to identify new anti-inflamm-atory drugs, led to the discovery of p38α/ß, thus provi-ding excellent tools for dissecting MAPK signaling pathways. More recently, mitosis research has been aided enormously by development of inhibitors targe-ting kinesin-related motor proteins (e.g. Eg5 and Cenp-E) and mitotic kinases (e.g. Plk1, Aurora B, Haspin and Mps1). The most famous mitotic inhibitors however are
the taxanes, microtubule stabilizers that are used as frontline chemotherapy agent to treat a variety of can-cers. And indeed, the desire to develop better anti-mitotic chemotherapeutics was the driving force behind developing many of the targeted mitotic inhibitors men-tioned above. Perhaps surprisingly however, how taxol and other microtubule toxins yield patient benefit remains rather unclear. Our current research therefore is aimed at understanding how cells react when treated with these various anti-mitotic agents. Surprisingly, our efforts have led us to the oncogenic transcription factor c-Myc, and we now have compelling evidence that Myc is a major determinant of mitotic cell fate in response to both traditional microtubule toxins and 2nd generation inhibitors. Importantly, this has opened up new opportunities for sensitizing cancer cells to these important drugs. My talk will focus on these issues.
How do Cells React When Mitosis Goes Wrong ?
Stephen Taylor, Faculty of Life Sciences, University of Manchester

32

33
Short Talks
Franziska Doll AG Zumbusch, Universität Konstanz 34
Michele Felletti AG Hartig, Universität Konstanz 35
Andreas Groß AG Drescher, Universität Konstanz 36
Nassima Meriem Gueddouda University of Bordeaux and INSERM, Pessac 37
Sandra Hess AG Mecking, Universität Konstanz 38
Grzegorz Kubik AG Summerer, Universität Konstanz 39
Saurav Malhotra AG Mayer, Universität Konstanz 40
Nemanja Milisavljevic Institute of Organic Chemistry and Biochemistry AS CR, Prague 41
Leila Motiei Weizmann Institute of Science, Rehovot 42
Christoph Paone AG Hauck, Universität Konstanz 43
Sabrina Weickert AG Drescher, Universität Konstanz 44
Xiaohui Zhao AG Marx, Universität Konstanz 45

34
The post-translational modification of intracellular pro-teins with ß-linked N-acetylglucosamine, termed O-GlcNAcylation, is widespread and plays an important role in regulating the proteins’ structure and function. Dysregulation of protein O-GlcNAcylation is related to severe illnesses such as Alzheimer’s disease or type 2 diabetes. Hence, the visualization of the O-GlcNAcyla-tion-state of specific proteins inside living cells is of great interest. To achieve this goal, we combined meta-bolic glycoengineering with Fluorescence Lifetime Ima-ging-Förster Resonance Energy Transfer (FLIM-FRET) microscopy. In our approach, EGFP is genetically fused to a protein of interest. A recently developed N-acetyl-glucosamine derivative bearing a cyclopropene-tag is incorporated into the cellular glycome in order to target protein O-GlcNAcylation. In a second step, it reacts with a dye-tetrazine conjugate in a bioorthogonal Diels-Alder reaction with inverse electron demand. The
glycosylation of the protein of interest then enables FRET between the donor EGFP and the acceptor dye. FLIM allows for the detection of FRET with high cont-rast even in the presence of a large excess of acceptor fluorophores. This strategy was successfully applied to visualize the glycosylation of specific proteins inside living cells. O-GlcNAc-transferase, the key regulator of protein O-GlcNAcylation, was chosen to establish this method by performing Western blot analysis and FLIM-FRET microscopy on fixed and living cells. To verify the broad applicability of our approach, we additionally visualized the intracellular glycosylation of the insulin receptor ß-subunit and the tumor suppressor p53. This is the first time that the glycosylation state of specific intracellular proteins was visualized directly inside living cells. Our new technique provides a general tool and will be used to study influences of glycosylation on the function and dynamics of proteins.
Franziska DollImaging Protein-Specific Glycosylation Inside Living Cells Using FLIM-FRET Microscopy
Franziska Doll, Annette Buntz, Anne-Katrin Späte, Verena F. Schart, Valentin Wittmann and Andreas Zumbusch*University of Konstanz and Konstanz Research School Chemical Biology (KoRS-CB), Contact: [email protected]

35
The recent description of a new class of small endonu-cleolytic ribozymes termed “Twister” opened new ave-nues into the field of RNA biology and synthetic biology. We present for the first time a series of approaches and designs for the employment of the twister ribozyme as an expression platform in the construction of artificial riboswitches that are able to control gene expression in E. coli. In our design the ribozyme motif sequesters the ribosome binding site allowing control of translation ini-tiation. The twister ribozyme reveals to be an outstan-dingly flexible expression platform and enables the construction of many different one- and two-input regulators for controlling gene expression in a ligand-dependent manner. We first generated one-input dependent riboswitches some of which outperform previous artificial genetic regulators, by attaching apta-mer domains to two different sites of the catalytic
domain. The fact that the twister ribozyme scaffold offers at least two independent sites for attaching ligand-sensing aptamer domains, opens the way to the development of Boolean logic operators at the post-transcriptional level by constructing compact two-input riboswitches that sense and respond to two small molecular signals at once. Using both a rational design and a screening strategies we were able to generate a broad range of binary Boolean logic gates such as AND, NAND, OR, NOR, ANDNOT and ORNOT opera-tors. Besides the important implications for synthetic biology, our results represent the first proof for the involvement of twister ribozymes in the control of gene expression in bacteria. The observed versatility in gene-regulatory setups hints at possible roles of naturally occurring ribozyme motifs.
Michele FellettiTwister Ribozymes as Highly Versatile Expression Platforms for Artificial Riboswitches
Michele Felletti1, 2, Julia Stifel1, Sophie Geiger1, Lena A. Wurmthaler1, 2 and Jörg S. Hartig1, 2
1 Department of Chemistry, University of Konstanz, Germany
2 Konstanz Research School Chemical Biology (KoRS-CB), University of Konstanz, Germany

36
The determination of structure and dynamics of biomac-romolecules like proteins in their natural environment is crucial for understanding their function. A powerful tool to gain structural information of proteins is intracellular electron paramagnetic resonance (in-cell EPR). This technique utilizes site-directed labeling of the target biomacromolecule by a stable paramagnetic label. How-ever, intracellular application of EPR is often hampered by the short lifetime of the commonly used radical nit-roxide spin labels because of the reducing milieu inside a cell.[1, 2] To overcome this limitation, we introduce the Gd(III) based spin label Gd(III)[2,6-pyridine-bis(methylamino)-N,N,N’,N’-tetraacetic acid] (Gd-PyMTA) for intracellular
distance measurements with the EPR technique double electron-electron resonance (DEER).[3] Gd-PyMTA tur-ned out to be cell compatible and was proven to be inert in cell extract of Xenopus laevis oocytes. The polyproline (PP)-peptide H-AP10CP10CP10-NH2 was site-direc-tedly spin labeled with Gd-PyMTA at both cysteine moie-ties and microinjected into Xenopus laevis oocytes. These EPR experiments suggest that the spin labeled PP-peptide is inserted into cell membranes coinciding with a conformational change of the PP-helix type II to PP-helix type I, which cannot be observed in cell extract. Thus, this study shows that in-cell EPR distance measu-rements are capable to monitor conformational changes in cellula.[3]
Andreas GroßConformational Change of a Spin Labeled Peptide in Cellula Monitored by EPR Distance Measurements
Andreas Groß1, Mian Qi2, Gunnar Jeschke3, Adelheid Godt2, Malte Drescher1
1 Department of Chemistry, and KoRS-CB, University of Konstanz, DE
2 Faculty of Chemistry and Center for Molecular Materials; Bielefeld University, DE
3 Department of Chemistry and Applied Biosciences, ETH Zurich, CH
[1] Azarkh, M.; Okle, O.; Eyring, P.; Dietrich, D. R.; Drescher, M. J. Magn. Reson. 2011, 212, 450.
[2] Krstić, I.; Hänsel, R.; Romainczyk, O.; Engels, J. W.; Dötsch, V.; Prisner, T. F. Angew. Chem. Int. Ed. 2011, 50, 5070
[3] Qi, M.; Groß, A.; Jeschke, G.; Godt, A.; Drescher, M. J. Am. Chem. Soc. 2014, 136, 15366.

37
G-quadruplexes (G4) are RNA and DNA secondary structures formed by the stacking of guanine quartets in G-rich sequences. Computational studies argue that more than 370 000 sequences in human genome are able to form G4. Some of these sequences are found in key regions such as 5’-UTR of messenger RNA, onco-genes promoters, telomeres … [1] Their existence was probed using structure-specific antibodies [2] and through their interactions with different proteins. Indeed, helicases like WRN, BLM, FANCJ or PIF1, show abilities to unwind different G-quadruplexes in vitro.
Mutations in these helicases are responsible for genome instability and for increase in risks of cancer [3]. So targeting G4 is of great interest to understand their impact on gene expression and more specifically on oncogenesis. Here, we present a fluorescence-based reliable, inexpensive and fast assay to study G4/heli-case interactions (figure). We use the nuclear isoform of Pif1 from S.cerevisiae, a 5’ to 3’ helicase. This assay allows to monitor the unfolding properties of this enzyme towards DNA quadruplexes and to screen for ligands that target G-quadruplexes. [4]
Nassima Meriem GueddoudaA Fluorescent Based G4 Helicase Assay
Nassima Meriem Gueddouda1, 2, Oscar Mendoza, Anne Bourdoncle1, 2, Jean-Baptiste Boulé3, Jean-Louis Mergny1,
1 University of Bordeaux, ARNA laboratory, Bordeaux, France, [email protected].
2 INSERM, U869, IECB, Pessac, France.
3 CNRS, UMR7196, Museum National d’Histoire Naturelle, 75005 Paris, France.
[1] Huppert, J. L. Balasubramanian, S. Nucleic Acids Research. 2005, 33(9), 2908–2916.
[2] Biffi, J. McCafferty, J. Balasubramanian, S. Nature Chemistry. 2013, 6(1), 75–80.
[3] Bochman, M. L. Paeschke, K, Zakian, V. A. Nature Reviews. Genetics. 2012, 13(11), 770–80.
[4] Mendoza, O. Gueddouda, N. M. Boule, J.-B. Bourdoncle, A. Mergny, J.-L. Nucleic Acids Research. 2015, 1–10.
Figure: Illustration of the helicase reaction relying on a G4/duplex DNA system. The substrate displays a sin-gle-stranded, a G4 and a duplex regions. When the duplex is formed, the fluorescence of the FAM donor (orange symbol) labelling the single strand is quenched. In the first step of the reaction, the Pif1p helicase binds to the 5’ single-strand end. After ATP addition, the
helicase proceeds in a 5’ to 3’ direction, unwinding first the G-quadruplex structure and then the duplex DNA. The unwinding of the duplex releases the labelled sin-gle-stranded probe, which emits a strong fluorescence while it is no longer quenched by the dabcyl acceptor (purple symbol) on the complementary strand. [4]

38
Microalgae are attractive as a feedstock as they contain unique compositions of, for example, fatty acids. Bene-ficially, they gain energy from photosynthesis like higher plants, but they have the advantage to grow in brackish or salt water independent from season and they do not compete with food production by consuming arable land. Additionally, microalgae can double within 24 hours and their lipid content can reach up to 70 wt-% of their dry weight.[1] Current efforts to technically use microalgae oil focus on the generation of fuels with a molecular structure ideally identical to crude oil based products.[2] In this contribution we discuss a different approach for the utilization of algae by translating the unique molecular structures of algae oil fatty acids into
higher value chemical intermediates and materials. The issues of algae mass workup and the compatibility of catalytic conversions with the specific multicompo-nent mixtures obtained are discussed for the example of carbonylation (Scheme 1). These direct practical considerations are aided by a mechanistic understan-ding. The behavior of multiple unsaturated components is of particular interest here. To this end, multifunctional intermediates could be enriched in Phaeodactylum tri-cornutum without compromising yields via genetic modification. Moreover, it was possible to tune the fatty acid composition only via the algaes’ growth conditions.[3, 4] These intermediates are further investigated in dif-ferent selective reactions.
Sandra K. HessHigher Value Chemicals from Crude Algae Oil LipidsSandra K. Hess, Philipp Roesle, Florian Stempfle, Julia Zimmerer, Stefan Mecking*Department of Chemistry, University of Konstanz, GermanyDaniela Ewe, Carolina Río-Bártulos, Bernard Lepetit, Angelika Eckert, Peter Kroth*Department of Biology, University of Konstanz, Germany
Literature [1] A. J. A. van Maris, A. A. Winkler, D. Porro, J. P. van Dijken, J. T. Pronk, Appl. Environ. Microbiol. 2004, 70, 2898–2905.
[2] U.S. DOE 2010. National Algal Biofuels Technology Roadmap. U.S. Department of Energy, Office of Energy Efficiency and Renewable Energy, Biomass
Program.
[3] P. Roesle, F. Stempfle, S. K. Hess, J. Zimmerer, C. Río Bártulos, B. Lepetit, A. Eckert, P. G. Kroth, S. Mecking, Angew. Chem. Int. Ed. 2014, 53, 6800-
6804.
[4] Sandra K. Hess, Masterthesis, University of Konstanz, 2014.
Scheme 1: Utilization of
algae as exemplified by the
generation of an aliphatic
long-chain polyester.[3]

39
Epigenetic modification at the 5-position of cytosine is a key regulatory element of mammalian gene expression with important roles in genome stability, development, and disease.[1] The repertoire of cytosine modifications, long confined to only 5 methylcytosine (mC), has recently been expanded by the discovery of 5-hydroxymethyl-, 5-formyl-, and 5-carboxylcytosine. These are key inter-mediates of active mC demethylation but may additionally represent new epigenetic marks with distinct biological roles.[2] This leap in chemical complexity of epigenetic cytosine modifications has not only created a pressing need for analytical approaches that enable unraveling of their functions, it has also created new challenges for such analyses with respect to sensitivity and selectivity. The crucial step of any such approach that defines its analytic potential is the strategy used for the actual diffe-rentiation of the cytosine 5-modifications from one ano-ther. Transcription-activator-like effectors (TALEs) have recently emerged as a new platform for the design of programmable DNA binding domains,[3] consisting of concatenated repeats, each of which recognizes one nucleobase through a repeat variable diresidue (RVD). RVDs with the amino acids NI, NN (NH), NG, and HD the-reby recognize A, G, T, and C, respectively.[4] TALEs are strand-specific DNA major groove binders that recognize pyrimidines via the face of the 5-position. RVD HD reco-gnizes C through a hydrogen bond between the aspartate and the cytosine 4-amino group.[5] We report direct diffe-rentiation between single genomic C and mC in vitro by TALEs[6] based on the mC sensitivity of RVD HD[7]. Fur-thermore TALEs can quantitatively detect mC levels at single positions and little depend on the sequence con-text or on the position of the mC.[8] Though RVD HD diffe-rentiates only between C and mC and cannot differenti-ate between mC and oxidized mCs, the engineering of RVDs holds the potential to provide new, individual selec-tivities for each of the five cytosine nucleobases, that is,
an expanded programmability of nucleobase recognition. Indeed first RVDs with individual selectivities were identi-fied in a pilot study involving C, mC, and hmC.[9] It should be pointed out that, programmable sequence selectivity is the prerequisite for targeting epigenetic nucleobases at user-defined loci in vivo. The combination of TALEs with functional protein domains such as fluorophores[10] or TET dioxygenases[11] thus offers potential for the in vivo detection of cytosine modifications, or for coupling their presence to the control of specific chromatin pro-perties.[12]
References: [1] Jones, Nat. Rev. Genet. 2012, 13, 484–492; Law et al., Nat. Rev.
Genet. 2010, 11, 204–220; Rodriguez-Paredes et al., Nat. Medicine
2011, 17, 330–339.
[2] Ito et al., Science 2011, 333, 1300–1303; He et al., Science 2011,
333, 1303–1307; Maiti et al., J. Biol. Chem. 2011, 286, 35334–
35338.
[3] Boch et al., Annu. Rev. Phytopath.2010, 48, 419–436; Bogdanove
et al., Science 2011, 333, 1843–1846.
[4] Moscou et al., Science 2009, 326, 1501; Boch et al., Science 2009,
326, 1509–1512; Yang et al., Cell Res. 2014, 24, 628–631.
[5] Deng et al., Science 2012, 335, 720–723; Mak et al., Science 2012,
335, 716–719.
[6] Kubik et al., Angew. Chem., Int. Ed. Engl. 2014, 53, 6002–6006.
[7] Bultmann et al., Nucleic Acids Res. 2012, 40, 5368–5377;
Valton et al., J. Biol. Chem. 2012, 287, 38427–38432.
[8] Kubik et al., Chembiochem 2015, 16, 228–231.
[9] Kubik et al., J. Am. Chem. Soc. 2015, 137, 2–5.
[10] Thanisch et al., Nucleic Acids Res. 2014, 42, e38; Ma et al., Proc.
Natl. Acad. Sci. U. S. A. 2013, 110, 21048–21053; Miyanari et al.,
Nat. Struct. Mol. Biol. 2013, 20, 1321–1324.
[11] Maeder et al., Nat. Biotechnol. 2013, 31, 1137–1142.
[12] Kubik et al., ACS Chem. Biol. 2015, Article ASAP.
Grzegorz KubikDeciphering Epigenetic Cytosine Modifications by Direct Molecular Recognition with TALEs
Grzegorz Kubik and Daniel Summerer, University of Konstanz
Reprinted from ACS Chem.
Biol., 2015, 10 (7), pp
1580–1589. Copyright
© 2015 American Chemical
Society

40
The vertebrate oocyte pool is sustained through a long-lasting arrest at the prophase of meiosis I (MI). The robustness of this arrest and responsiveness to appro-priate stimuli for entry into the meiotic divisions are cri-tical for reproductive health. Here we demonstrate that the ubiquitin E3-ligase anaphase-promoting-complex/cyclosome (APC/C) along with its co-activator Cdh1 is essential for the entry of Xenopus oocytes into M-phase of MI. Further, we identify the catalytic subunit of protein-phosphatase-6 (PP6c) as the critical subst-rate whose APC/CCdh1-mediated destruction is prere-quisite for MI entry. Preventing PP6c destruction
blocks meiotic entry and impairs activating autophos-phorylation of Aurora-A, a cell-cycle kinase critical for the activation of meiotic translation events essential for M-phase entry. Restoring meiotic translation rescues the meiotic resumption defect of Cdh1-depleted oocy-tes. Thus, our studies identify a crucial APC/CCdh1-PP6c-Aurora-A signaling axis in the resumption of ver-tebrate female meiosis and discover that the essential function of the APC/C in triggering cell-cycle transi-tions is not limited to M-phase exit but also applies to M-phase entry in vertebrate oocytes.
Saurav MalhotraThe Anaphase Promoting Complex/Cyclosome is Essential for Entry into Meiotic M-Phase
Saurav Malhotra1, P.K. Vinod2, 3, Olaf Stemmann4 & Thomas U. Mayer1
1 Department of Molecular Genetics and Konstanz Research School Chemical Biology
(KoRS-CB), University of Konstanz,Universitätsstr. 10, 78457 Konstanz, Germany.
2 OCISB, Department of Biochemistry, University of Oxford, South Parks Road, Oxford OX13QU, UK.
3 Current address: CCNSB, International Institute of Information Technology, Hyderabad, India, 500032.
4 Department of Genetics, University of Bayreuth, Universitätsstr. 30, 95440 Bayreuth, Germany.

41
Base-modified RNAs are crucial tools in RNA chemical biology.[1] Most common method for the synthesis of nucleic acids is the solid phase synthesis which suffers from drawbacks such as low functional group tolerance and low yields.[2] Alternative approach is by enzymatic incorporation of modified nucleoside triphosphates.[3]
Routinely used enzyme for RNA synthesis is T7 RNA polymerase since it does not require special conditions nor the presence of complex transcription factors for its function.[4] Our objective is to synthesize novel nucleo-side triphosphates and use them for construction of functionalized RNAs (Fig. 1).
Nemanja MilisavljevicSynthesis of Base Modified Nucleoside Triphosphates for the Construction of Modified RNAs
Nemanja Milisavljevic, Pavla Perlíková and Michal Hocek*Institute of Organic Chemistry and Biochemistry AS CR, Flemingovo nam. 2, 166 10, Prague 6, Czech Republic, [email protected]
This work was supported by Gilead Sciences Inc.
References: [1] M. Phelps, A. Morris, P. Beal, ACS Chem. Bio. 2012, 7, 100–109.
[2] H. El Sagheer, T. Brown, Proc. Natl. Acad.Sci. U.S.A. 2010, 107, 15329–15334.
[3] Y. Jao, A. Salic, Proc. Natl. Acad.Sci. U.S.A. 2008, 105, 15779–15784.
[4] J. Milligan, D. Groebe, G. Witherell, O. Uhlenbeck, Nucleic Acid Res. 1987, 15, 8783–8798.
Fig. 1: Novel nucleoside triphosphates and in vitro transcription reaction.

42
Fluorescent molecular sensors have become valuable tools in the analytical biosciences owing to their high sensitivity and their ability to track proteins in their native environment. A major limitation in using these probes, however, is the lack of a general and easily applicable method for providing them with high selecti-vity and high signal-to-noise ratios. In addition, many of these sensors are designed according to the “lock and key” paradigm; therefore, they cannot be used to ana-lyze biomolecule combinations.
In our group, we aim to address these problems by developing novel classes of fluorescent molecular sen-sors. [1, 2, 3] I present our recent development of fluores-cent switches that light up in the presence of specific protein biomarkers, as well as probes that can shed light on analyte combinations in biofluids. In addition, the design of a sensory system that utilizes both speci-fic and non-specific interactions for distinguishing bet-ween protein isoforms will be discussed.
Leila MotieiCommunicating with Fluorescent Molecular Sensors in a Biological Environment
Leila Motiei, David Margulies Department of Organic Chemistry, Weizmann Institute of Science, 76100, Rehovot (Israel), E-Mail: [email protected]
References:
[1] L. Motiei, Z. Pode, A. Koganitsky, D. Margulies, Angew. Chem. Int. Ed. 2014, 53, 9289–9293.
[2] L. Unger-Angel, B. Rout, T. Ilani, M. Eisenstein, L. Motiei, D. Margulies, Chem. Sci. 2015, in press, DOI: 10.1039/C5SC01038A.
[3] B. Rout, L. Motiei, D. Margulies, Synlett, 2014, 25, 1050–1054.

43
Cell adhesion and migration depend on integrin-initia-ted cell adhesion sites, so-called focal adhesions. Focal adhesions are multi-protein complexes, where a cha-racteristic core set of proteins is assembled in a hierar-chical fashion guided by protein phosphorylation. A key signaling molecule at focal adhesions is the focal adhe-sion kinase (FAK), which is critical for the dynamic regu-lation of focal adhesion complexes and, therefore, important for cell migration processes during embryo-genesis, tissue homeostasis, and cancer metastasis. FAK is anchored to these subcellular sites via interac-tions with the focal adhesion proteins paxillin and talin. We postulated that a chemical compound, which could displace FAK from focal adhesions, would interfere with FA dynamics and would block cell migration. Using high-throughput screening and automated image ana-lysis, we screened ~17,000 compounds of a commer-cial library for effects on FAK localization at focal adhe-sion sites. We identified two compounds that displaced FAK from focal adhesion sites, but that did not affect
the localization of vinculin, another FA marker protein. We named these compounds FoolLocker1 and FootLo-cker2 for their effects on FA dynamics. Microscopic analysis demonstrated that besides FAK, several known FAK-binding partners (paxillin, p130CAS, and Grb2) were also displaced from FAs by FootLocker, whereas the localization of most FA core proteins (including talin, vinculin, zyxin, or α–actinin) was not affected. In addi-tion, FootLockers were able to inhibit cell migration within minutes, but the effect was reversible upon wash-out of the compound. Structure-activity investi-gations revealed the critical molecular determinants for cellular activity of FootLocker and allowed the inclusion of additional functionality by chemical synthesis. The combined characteristics of FootLocker derivatives make these compounds attractive chemical tools to decipher the sequence of events during focal adhesion complex formation, but also form the basis for novel applications in modulating cell migration.
Christoph PaoneFootLocker – Small Molecule Disruptors of Integrin-Initiated Focal Adhesion Complexes
Christoph Paone, Alexander Buntru, Christof R. Hauck, Lehrstuhl Zellbiologie, Universität Konstanz

44
Multivalency is a key concept in biological systems lea-ding to targeted, specific and strong interactions which play a role in many biological processes, among them signaling, adhesion, etc. The interplay of multiple ligand-receptor moieties has the potential to enormously incre-ase binding affinities compared to a monovalent binding situation. Knowledge of the topology of binding sites is thus crucial for the design of high-affinity multivalent ligands, e.g., as potent inhibitors. An electron paramag-netic resonance (EPR) spectroscopy based method for distance determination between ligand binding sites with a divalent spin labeled GlcNAc ligand and the model protein wheat germ agglutinin (WGA) was pre-sented by Braun and coworkers.[1] However, in order to apply this approach to any protein receptor also without
a structural model at hand, it is desirable to determine reliable distances with monovalent ligand molecules and a spin label which allows straight forward assignment of precise distance information to the protein structure. Therefore, we present a newly synthesized C-glycoside of GlcNAc as monovalent ligand for WGA carrying a conformationally unambiguous spin label side chain. The ligand and its binding behavior are characterized using EPR spectroscopy. Distance measurements using double electron-electron resonance (DEER) spectro-scopy are in accordance with the X-ray crystallography structure of WGA.[2] Based on this proof of concept study with the model system WGA it is possible to apply our approach to many multivalent receptor-ligand sys-tems.
Sabrina WeickertTopology of Multivalent Binding SitesSabrina Weickert1, Torben Seitz1, Valentin Wittmann1 and Malte Drescher1
1 Department of Chemistry, University of Konstanz, 78457 Konstanz, Germany, [email protected]
[1] P. Braun et al., Angew. Chem. Int. Ed. 2011, 50(36), 8428–8431.
[2] D. Schwefel et al., J. Am. Chem. Soc. 2010, 132(25), 8704–8719.

45
Branched ubiquitin chains in which a single ubiquitin moiety is modified at various lysine sites by two or more Ubs, attract researchers’ attention since their existence was initially confirmed in yeast. Together with other types of chains containing distinct linkages and diffe-rent lengths, they serve numerous signals in protein ubi-quitylation. However, so far, only a few of defined chains and their functions in cellular processes were deciphe-red. To elucidate their structure-function relationships, generation of linkage-specific chains in a large amount is required. Due to the limitations of enzymatical methods, such as lack of linkage-specific Ub-conjugating enzymes and assembling the mixture of undefined chains, we provide here a facile and efficient bioothogonal chemistry method to generate defined branched Ub chains and all
seven homogeneous chains. Through site-directed mutagenesis, different lysine residues were replaced by cysteins. The resulting ubiquitin bearing thiol groups could be modified with propargyl acrylate so that termi-nal alkynes were easily introduced at specific sites. Fur-thermore, the unnatural amino acid (azidohomoalanine) was introduced at C-terminus of Ub through selective pressure incorporation. Then click reaction was perfor-med to generate required chains which were further analyzed by western blot using linkage-specific anti-body, purified through size exclusion chromatography and analyzed by mass spectrometry. Moreover, we found our synthesized chains are still biologically active, as K11-linkage homogeneous chain has the ability to prevent degradation of XErp1 and cyclin B in Xenopus egg extracts.
Xiaohui ZhaoGeneration of Protease Resistant Branched Ubiquitin ChainsXiaohui Zhao, Daniel Schneider, Saurav Malhotra, Tatjana Schneider, Thomas U. Mayer, Martin Scheffner and Andreas Marx, (Departments of Chemistry and Biology, Konstanz Research School Chemical Biology, University of Konstanz, 78457 Konstanz, Germany)
References:
[1] Nat. Biotech. 2003, 21, 921;
[2] Angew. Chem. Int. Ed. 2013, 52, 192;
[3] Angew. Chem. Int. Ed. 2014, 53, 12925
Konstanz Research School Chemical Biology and DAAD are gratefully acknowledged.

46
Notes
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
....................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
– ks-cb.com

47
Notes
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
....................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
– ks-cb.com

48
Notes
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
....................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
– ks-cb.com

49
Notes
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
....................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
...................................................................................................................................................................................................................
– ks-cb.com

ContactKonstanz Research School Chemical Biology (KoRS-CB)[email protected]
Dr. Heike BrandstädterGeneral Manager+49 7531 [email protected]
Katharina MagerkurthProject Assistant+49 7531 [email protected]
– chembiol.uni-konstanz.de uni.k
n · w
wa-
grafi
k, w
wa-
druc
k · F
oto:
© M
ende
· 0
9/2
015


![arXiv:1802.03810v1 [cond-mat.mes-hall] 11 Feb 2018Budapest University of Technology and Economics, H-1111 Budapest, Hungary 4Department of Physics, University of Konstanz, D-78457](https://img.pdfslide.us/doc/110x75/5e72edead0f78059304c712c/arxiv180203810v1-cond-matmes-hall-11-feb-2018-budapest-university-of-technology.jpg)
![2008 05 white presentation at 39th konstanz seminar konstanz 7 may 2008 [mode de compatibilité]](https://img.pdfslide.us/doc/110x75/559877471a28ab39058b461e/2008-05-white-presentation-at-39th-konstanz-seminar-konstanz-7-may-2008-mode-de-compatibilite.jpg)

























