Embed Size (px)
Citation preview

119
Chapter 4
Kissing Cousins:Genetic Interactions between
Wild and Cultured Salmon
Fred M. Utter
The Fulton spawning channel on the Babine River (Area 4) is one of four constructed spawning channelsdesigned to increase sockeye production. At 4 km long, Channel 2 is one of the largest spawning channelsin the world, producing 67 million sockeye fry in 2001. While the sockeye catch in the Skeena region hasincreased since the spawning channels were constructed in the 1960s, wild runs of sockeye have declinedsignificantly over this period. photo: Garth Traxler

120
The above questions from a public figure reflect wide-spread public concernabout the state of wild and hatchery salmon - concerns that are shared bymany citizens of British Columbia. Mr. Carlson’s inquiries are not frivolous.They echo frustrations about seeming over-regulation and possible defiance ofcommon sense and are based on understandings about genetic relationshipsthat have been widely held for the past 50 years.
When I entered the workforce in the 1950s, there were five species of the genusOncorhynchus native to the US Pacific Northwest (chinook, coho, pink, chum,and sockeye salmon), plus the Asiatic masou salmon. In addition to beingdistinct from one another, the five local species were understood to havevariant “races” within them such as spring-run and fall-run chinook, lake-typeanadromous sockeye and non-migratory kokanee, as well as even- and odd-year pink salmon.
Hatchery fish and ocean ranching were seen as the salvation of fisheriesbecause, then as now, naturally-reproducing runs were declining due tohabitat degradation, harvest pressures, and (in the United States at least)upstream and downstream migratory challenges imposed by dams. Like theabove-noted Germans in Tacoma (presumably contrasted with those fromSeattle), fall-run chinook salmon from, say, the lower Columbia River andPuget Sound were members of a common “race”. One declining segment ofthat race could, it was believed, readily be replenished by surpluses from theother, especially with viable hatchery returns from both groups. Of course,given the massive translocations of these two groups that have now beendocumented, any possible distinction that may have existed historically musthave vanished through hybridization.
This essay probes changes in these understandings that have occurred since Ijoined the work force. First, new insights into the genetic relationships amongand within Oncorhynchus species are outlined. The dynamics of genetic
“...When is a chinook salmon in Puget Sound a ‘species’ ?Are Germans in Tacoma a ‘species’ of human?....Why do ‘wild’ salmonand not hatchery salmon count (under the Endangered Species Act)?”
John Carlson, Republican candidate for
Governor of the State of Washington in 2000.
Introduction
Part Two, Chapter 4: Introduction

121
interactions among translocated and native groups are then examined. Finally,the pertinence of this knowledge to fish culture operations in BritishColumbia is considered.
The present era of molecular genetics began in the 1960s. The resultingabundance of single-gene markers has clarified and quantified relationshipswithin families such as the Salmonidae in a manner that was previouslyimpossible (Utter 1991). Among the consequences of using such molecularmarkers were a better understanding of the relatedness between and amongdifferent species, races and populations of Pacific salmon, and an expansion ofOncorhynchus to include trouts of the Pacific Rim formerly included within thegenus Salmo (Figure 4-1).
These revised perceptions provided a better biological basis for comparisonsamong these salmon and trout species. For example, anadromous rainbowtrout (steelhead) are now more properly grouped as biological cousins of theanadromous Pacific salmon rather than as close relatives of Atlantic salmon.The degree of divergence among the trout species is now recognized as similarto that of the North American salmon, and clear understandings have replacedformerly confusing specific and sub-specific taxonomic relationships.
The finer genetic resolution allowed by molecular markers indicates ancestralgroupings within four Pacific salmon species (Figure 4-2). These imagescontrast with those of Figure 4-1 by being based primarily on frequencies ofshared alleles (i.e., variant forms of single genes) rather than the morediagnostic (invariant or “fixed”) molecular genetic differences that characterizeclosely-related species. Nevertheless, considerable confidence can be placed ingroupings such as these that reflect many variable genes (i.e., loci) as well asrepresentative within-group sampling. The ability to project these groupingson maps is consistent with their primarily geographic character. Variable lifehistory characteristics (e.g., run timings) provide important but secondaryadaptive dimensions to geographic or ancestral group characterizations.
Past and presentunderstandingsof geneticrelationships
Ancestralgroupings withinspecies
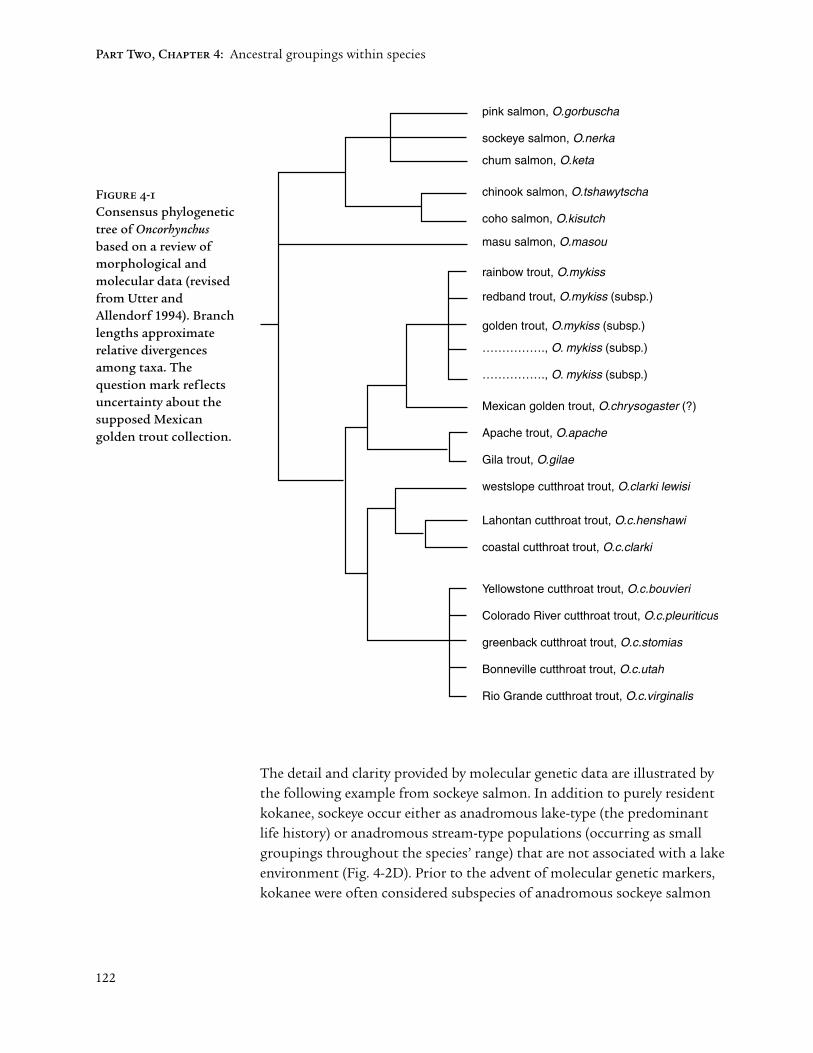
122
The detail and clarity provided by molecular genetic data are illustrated bythe following example from sockeye salmon. In addition to purely residentkokanee, sockeye occur either as anadromous lake-type (the predominantlife history) or anadromous stream-type populations (occurring as smallgroupings throughout the species’ range) that are not associated with a lakeenvironment (Fig. 4-2D). Prior to the advent of molecular genetic markers,kokanee were often considered subspecies of anadromous sockeye salmon
Figure 4-1
Consensus phylogenetictree of Oncorhynchusbased on a review ofmorphological andmolecular data (revisedfrom Utter andAllendorf 1994). Branchlengths approximaterelative divergencesamong taxa. Thequestion mark reflectsuncertainty about thesupposed Mexicangolden trout collection.
pink salmon, O.gorbuscha
sockeye salmon, O.nerka
chum salmon, O.keta
chinook salmon, O.tshawytscha
coho salmon, O.kisutch
masu salmon, O.masou
rainbow trout, O.mykiss
redband trout, O.mykiss (subsp.)
golden trout, O.mykiss (subsp.)
……………., O. mykiss (subsp.)
……………., O. mykiss (subsp.)
Mexican golden trout, O.chrysogaster (?)
Apache trout, O.apache
Gila trout, O.gilae
westslope cutthroat trout, O.clarki lewisi
Lahontan cutthroat trout, O.c.henshawi
coastal cutthroat trout, O.c.clarki
Yellowstone cutthroat trout, O.c.bouvieri
Colorado River cutthroat trout, O.c.pleuriticus
greenback cutthroat trout, O.c.stomias
Bonneville cutthroat trout, O.c.utah
Rio Grande cutthroat trout, O.c.virginalis
Part Two, Chapter 4: Ancestral groupings within species

123
O. nerka kennerlyi; (e.g., Robertson 1961), and the stream-type anadromousadaptation was more or less ignored. Molecular genetic data now indicatethat
• kokanee and anadromous populations within a lake system have a recentcommon ancestry distinct from ancestries in other lake systems; that is,sockeye and kokanee within most lakes are more closely related than eitheris to sockeye or kokanee, respectively, from other lakes (Foote et al. 1989),and
• stream-type populations appear to be primary colonizers of vacant (e.g.,post-glacial) habitats, and are thus most likely the key to the evolutionaryfuture of the species (Wood 1995; Gustafson and Winans 1999).
This example illustrates the critical role of molecular genetic data in lineageclassification. Life history traits remain crucial attributes within the adaptiveframework of a lineage, but their tendency to evolve independently amonglineages makes them unreliable as primary ancestral markers (Utter et al.1993).
The distinction among subgroups within major lineages lies near or beyondthe threshold of detection by molecular genetic markers. Similar frequenciesof more or less neutral marker alleles reflect recent divergence or intermittentgene flow among subgroups. However, these similarities may mask importantdirectional adaptations at other loci that distinguish subgroups and enrichthe genetic diversity of the lineage. For instance, no subgroup structure wasapparent from a molecular genetic survey of pink salmon populations nearJuneau, Alaska (Fig. 4-3). The low index of genetic diversity (F
ST = .003)
equated to consistently high gene flow among populations of this region,and supported a general understanding of weak regional substructure amongpink salmon populations. However, genetic marking of a late upstream run inAuke Creek revealed a fine substructure (i.e., a distinct subpopulation) thatpersisted over at least five generations (10 years). Similar within-lineagesubstructures have been found in other anadromous salmonids (Hendryet al. 2000). Such revelations imply the existence of pervasive adaptivesubstructures within anadromous salmonid lineages. Optimizing theoccupation of available or emerging habitats, these substructures apparentlydissolve and re-form as habitats shift, like eddies in a stream.
Adaptivedivergencewithin majorgroups
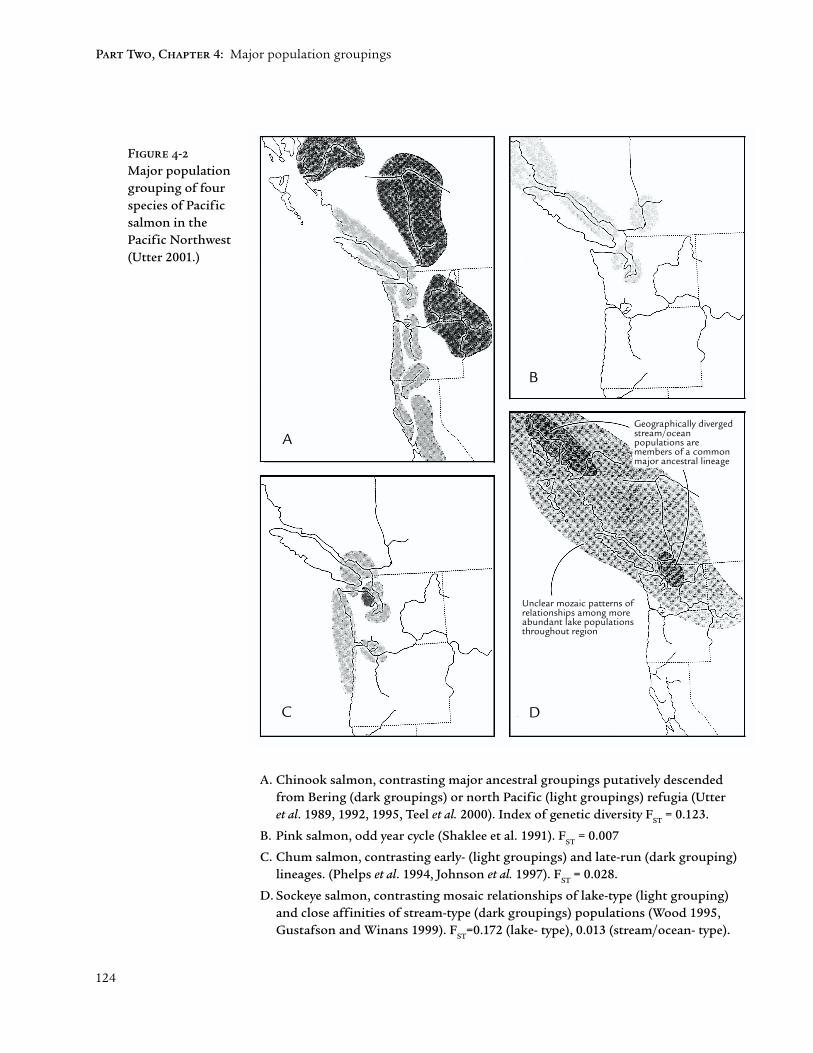
124
A. Chinook salmon, contrasting major ancestral groupings putatively descendedfrom Bering (dark groupings) or north Pacific (light groupings) refugia (Utteret al. 1989, 1992, 1995, Teel et al. 2000). Index of genetic diversity F
ST = 0.123.
B. Pink salmon, odd year cycle (Shaklee et al. 1991). FST
= 0.007
C. Chum salmon, contrasting early- (light groupings) and late-run (dark grouping)lineages. (Phelps et al. 1994, Johnson et al. 1997). F
ST = 0.028.
D. Sockeye salmon, contrasting mosaic relationships of lake-type (light grouping)and close affinities of stream-type (dark groupings) populations (Wood 1995,Gustafson and Winans 1999). F
ST=0.172 (lake- type), 0.013 (stream/ocean- type).
Figure 4-2
Major populationgrouping of fourspecies of Pacificsalmon in thePacific Northwest(Utter 2001.)
A
B
C D
Unclear mozaic patterns ofrelationships among moreabundant lake populationsthroughout region
Geographically divergedstream/oceanpopulations aremembers of a commonmajor ancestral lineage
Part Two, Chapter 4: Major population groupings

125
The capacity for genetic interaction among and within Oncorhynchus speciesdefies predictions based solely on taxonomic distance. The classicalunderstanding of species as “groups of populations where gene exchangebetween such groups is limited by reproductive isolating mechanisms” (e.g., seediscussions in Avise 1994) often breaks down in this genus. Introgressivehybridization (gene flow between genetically differentiated groups) readilyoccur between native and introduced trout species that have diverged for morethan a million years (Leary et al. 1995). In contrast, translocations amongconspecific anadromous lineages indicate strong resistances to introgressionbetween groups separated by much shorter intervals (Figure 4-2; Utter et al.1995). Complex co-adaptations appear to inhibit introgression among lineagesof anadromous salmon through severe penalties (i.e., outbreeding depression)affecting hybridized offspring (Utter 2001). In contrast, ease of introgressionwithin anadromous lineages suggests much weaker coadaptations andassociated penalties.
In response to the questions raised at the beginning of this chapter, chinooksalmon of Puget Sound indeed behave as a species in the classical sense. Thesituation is more complex with regard to conservation issues involvinghatchery and wild salmon. Critical genetic considerations include whether ornot introgressive hybridization is likely to occur, and, if it does, the adversegenetic interactions between hatchery-adapted (“domesticated”) and wildpopulations. Also, numerous indirect effects (Figure 4-5) almost inevitablyaccompany large-scale culture operations and releases. Those of particularconcern in the discussion below are disease transmission from cultured to wildpopulations and excessive harvest imposed on wild populations in mixed stockfisheries based on more abundant cultured salmon of the same species.
Capacity forgeneticinteraction
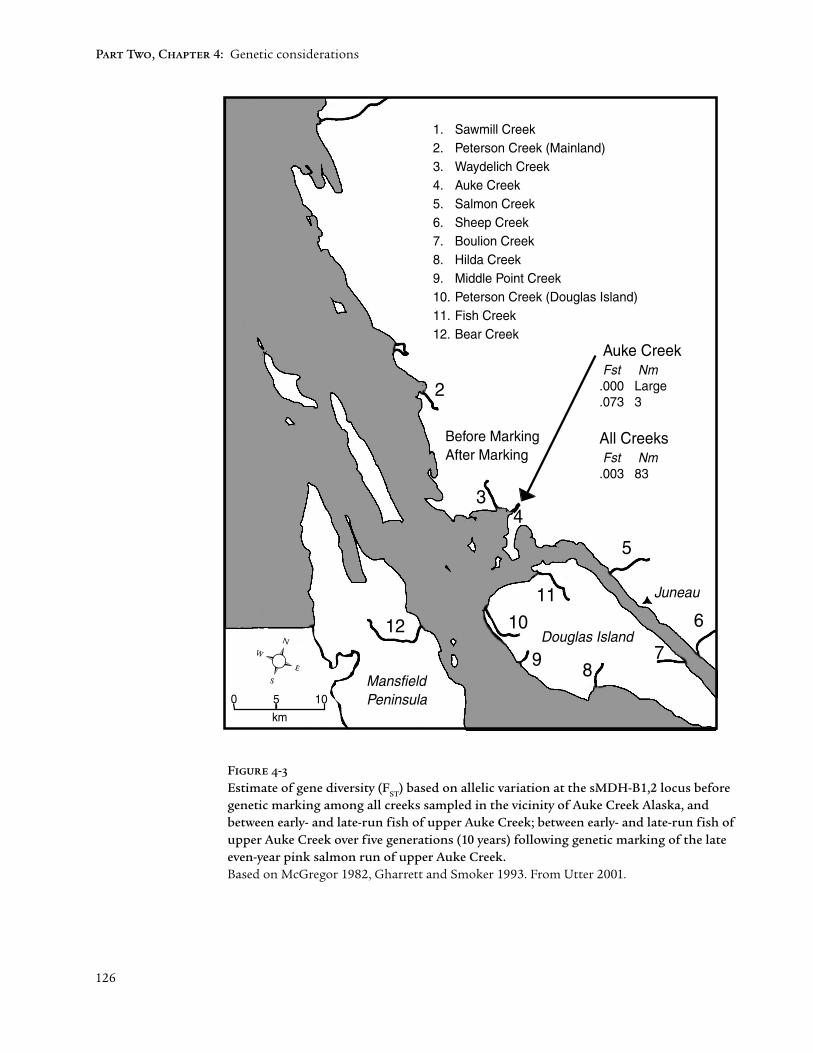
126
1. Sawmill Creek2. Peterson Creek (Mainland)3. Waydelich Creek4. Auke Creek5. Salmon Creek6. Sheep Creek7. Boulion Creek8. Hilda Creek9. Middle Point Creek10. Peterson Creek (Douglas Island)11. Fish Creek12. Bear Creek
2
34
5
6
789
10
11
12Douglas Island
MansfieldPeninsula
Before MarkingAfter Marking
Auke Creek Fst Nm .000 Large .073 3
All Creeks Fst Nm .003 83
�km
0 5 10
Juneau
Figure 4-3
Estimate of gene diversity (FST
) based on allelic variation at the sMDH-B1,2 locus beforegenetic marking among all creeks sampled in the vicinity of Auke Creek Alaska, andbetween early- and late-run fish of upper Auke Creek; between early- and late-run fish ofupper Auke Creek over five generations (10 years) following genetic marking of the lateeven-year pink salmon run of upper Auke Creek.Based on McGregor 1982, Gharrett and Smoker 1993. From Utter 2001.
Part Two, Chapter 4: Genetic considerations

127
The range and longevity of introgression in cultured fish affects its impacton wild populations. Using contrasting hypothetical examples based onFigure 4-3, consider first the case of a localized enhancement activity focusedon increasing and stabilizing late returns of even-year pink salmon to upperAuke Creek. Straying (and thus introgression) to other temporal or geographicsubgroups within or beyond Auke Creek could be minimized or preventedthrough design of the operation. Such an idealized operation would still haveto deal with genetic concerns within the targeted subgroup. One such concernis supportive breeding where the genetically effective size of the overallsubgroup is reduced by magnifying the survival of the small part of thepopulation represented by the cultured parents and their offspring (Rymanand Laikre 1991; Waples and Do 1994). Paradoxically, this effect is increased asthe enhancement becomes more successful. Thus the short-term benefits ofenhancement must be weighed against the longterm loss of genetic variabilityaffecting future adaptability of the subgroup. Harvest of the successfulenhancement would also have to be controlled to prevent over-harvest of co-migrating wild fish from other subgroups. Assuming satisfactory resolution ofboth of these concerns, a localized enhancement program could coexist with ahealthy substructure of natural reproduction (Figure 4-4).
An alternative enhancement program might be a regional facility with broodfish originally obtained throughout the area, and producing a healthy surplusof returning adults free to migrate to nearby streams and interbreed withnative fish. The resulting introgressions then erode largely unrecognizedgeographic and temporal natural substructures with a corresponding declinein natural productivity (Figure 4-4). Although hatchery productioncompensates for this decline in numbers of returning adults, the overallnumber of effective breeders is reduced by the magnified survival of culturedfish contributing to both wild and hatchery reproduction. The fitness of theincreasingly homogeneous wild populations is further eroded by continuedinfusions of hatchery-adapted (i.e. “domesticated”) genotypes into thenaturally reproducing gene pools. Although modification of the cultureoperation toward one without supplementation, as outlined in Figure 4-4,would permit reformation of the original regional and within-streamsubstructures, the regional loss of effective breeders would slow the processconsiderably. Though hypothetical, these alternate scenarios are based onsubstantial empirical observation of Oncorhynchus:
• The validity of the natural substructure within a major lineage is outlinedin Figure 4-3
• The effects of culture operations on pink salmon populations in Prince
Geneticconsiderationsin the design ofenhancementoperations: anexample withinanadromouslineage.

128
William Sound described in Seeb et al. (1999) provide the basis for proposingdifferent effects of local and regional hatchery programs
• An example of regional homogenization induced by extended hatcherymanagement is provided by “tule” fall-run chinook salmon of the lowerColumbia River (Utter et al. 1989)
• Evidence exists for reduced effective population numbers arising from Pacificsalmon hatchery operations (Waples and Teel 1990), and the inescapablereality of culture-induced domestication was reviewed by Waples (1999)
• Considerable evidence has also accumulated for the reduced fitness ofhybridized cultured-wild anadromous salmonids contrasted with native-wildpopulations (e.g., Reisenbichler and McIntyre 1977, Campton et al. 1991,Hindar et al. 1991, Reisenbichler and Rubin 1999)
• The ability of anadromous salmonid substructures to re-form following stressor in vacated habitats has repeatedly been demonstrated (e.g., Withler 1984,Quinn et al. 1996, Kinnison et al. 1998, Hendry et al. 2000).
In addition to introgressive hybridization, other genetic interactions may occurbetween native and cultured salmonids. For example, another direct geneticeffect is altered selection regimes imposed on native fishes as a consequence ofreleases (e.g., Waples 1995). Also, the above-noted indirect genetic effects com-monly accompany large-scale culture operations and releases (Figure 4-5).
The above background provides a useful basis for examining the genetic effectsof culture operations on wild salmon populations in British Columbia. Thearguments that follow may be summarized thus:
• Distinct ancestries minimize concerns about introgressive hybridizationsbetween Atlantic salmon and Pacific salmon and trout
• However, taxonomy poorly predicts genetic interactions within Oncorhynchuswhere
• gene flow is strongly inhibited among subspecific anadromous lineages, but
• gene flow occurs freely within subspecific anadromous lineages
• In addition, important within-lineage adaptive substructure lies near orbeyond the threshold of molecular genetic detection where
• enhancement can unwittingly erode this substructure by stimulating over-harvest, reduction of the overall effective population size, and infusion ofnon-nadaptive genotypes
Geneticinteractionsbetween wild andcultured salmon inBritish Columbia:levels of concern
Part Two, Chapter 4: Genetic interactions
The Snootli Hatchery raisescoho and chinook that arereleased into streams in theBella Coola Region (Area 8).The short term gain in fishproduction must be weighedagainst the long term loss ingenetic variability.photo: Terry Brown

129
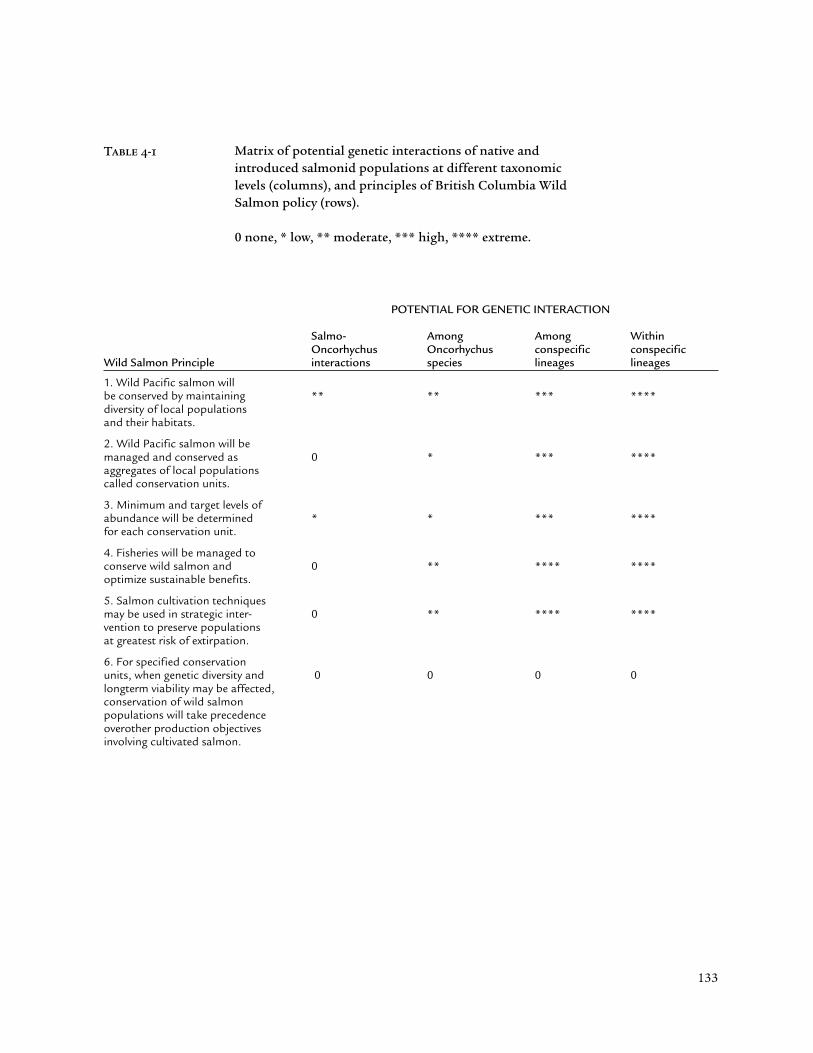
These conclusions, when viewed in the context of the principles for salmonconservation outlined in the Wild Salmon Policy (Fisheries and Oceans Canada2000) point to increasing concern as the potential for gene flow betweencultured and wild fish increases (Table 4-1). For this reason, Atlantic salmonescaping from netpen operations actually represent the lowest level of concern.Despite some experimental evidence for Oncorhynchus spp. x Salmo salarhybridization (e.g., Gross 1999) up to the first generation, the possibility forintrogressive hybridization appears to be virtually non-existent (see discussionsin Chevassus et al. 1983, Galbreath and Thorgaard 1994). Nevertheless, currentevidence for successful colonization by escaped Atlantic salmon in BCindicates a potentially serious problem, namely competition with alreadythreatened native salmon populations (Volpe et al. 2000, Volpe 2001).Although numerous failures at intentional colonization minimized thisconcern in the past (Alverson and Ruggerone 1997), recent evidence warrantsgreater monitoring and, where necessary, remedial action.
Gene flow among Oncorhynchus species is a greater worry. Though not recordedamong salmon species, introgressive hybridization between steelhead andcoastal cutthroat trout has been documented in Washington State with higherincidences related to cultural releases (Campton and Utter 1985, Hawkins
Figure 4-4
Contrastingsubgroupstructure withand withoutsupplementationwithin ananadromoussalmonid lineage.
WITHOUT SUPPLEMENTATION(Overall Productivity = x)
A complex and dynamic set of subgroupsconstantly adjusting to shifting natural habitats,and affected by such variables as times ofmigration and spawning, locations of migrationand spawning, duration of freshwater andmarine residence, and microhabitat distinctions(e.g., temperature, stream flow)
WITH SUPPLEMENTATION(Overall Productivity < x)
A reduced set of subgroups homogenized bygene flow imposed by persistent releases,straying, and outplanting of cultured fish.
A culture operation withisolated reproduction andharvest is not excluded as asubunit
Regeneration of naturalcomplexity followingdiscontinuation ofsupplementation

130
1997). Similar hybridizations between rainbow trout and cutthroat trout havebeen found in British Columbia. Some serious problems with introgressionsbetween released hatchery rainbow and native westlope cutthroat trout havebeen observed in the Kootenay drainage, where hybrids beyond the firstgeneration (identified by nuclear markers) and random mating (identified bymtDNA markers) were observed at each of 11 sites examined (Rubidge et al.2001). A sample of presumably pure coastal cutthroat from Vancouver Islandrevealed high levels of hybridization in an area stocked with rainbow trout(A. Dale and D. Heath, pers. com.).
Concerns about gene flow or displacement among lineages focus firston uncertainties regarding the existence and extent of those lineages(“conservation units”, Principle Two of the Wild Salmon Policy). In other words,in many cases such conservation units are not yet defined. Figure 4-2 providesvaluable but preliminary approximations of such units, many of which arealmost certainly subdivided into further lineages. Salmon culture operationswhich intentionally or potentially release fish must know the populationstructure of wild conspecifics with which cultured fish may interbreed,interact, and who they may even displace.
Cultured fish of distinct ancestry threaten wild populations in several ways.Interbreeding promotes severe outbreeding depression, leading to wastedreproduction and potential displacement (Utter 2001). Continued and large-scale releases further threaten wild populations with excessive harvest in mixedfisheries (e.g., Utter and Ryman 1993). Management to optimize sustainablebenefits and conserve wild salmon (Principle Four of the Wild Salmon Policy)cannot occur under these conditions. These concerns extend to escapes ofnative salmon reared in British Columbia, amounting to 10,000 tons or 20% ofall farmed salmon in 1999 (Egan 2000).
The highest levels of genetic concern relate to erosion of substructures withinmajor lineages. An assessment of the Babine Lake Development Project forsockeye salmon (BLDP) provides a useful example of culture-wild interactionswithin a single major lineage (Wood et al. 1998). Babine sockeye represent asingle ancestral grouping characterized by three distinct run timings (early,mid, late; Varnavskaya et al. 1994). Begun in the 1960s, the BLDP consists oftwo major spawning channels in tributaries to Babine Lake, where semi-natural rearing tempers concerns about domestication. Through its initialdecades, the project was an unquestionable numerical success, averaging analmost three-fold increase (to 2.7 million) in adult returns. However, recentdeclines in smolt survival, attributed in part to white spot disease, appear to be
Case study:The Babine LakeDevelopmentProject
Part Two, Chapter 4: Case study

131
moderating this trend, with a forecast of pre-enhancement levels for 1998.Despite attempts to focus harvests on enhanced fish, returns to un-enhancedareas have declined, particularly those whose timing coincided with that of theenhanced fish. At the same time, sizeable surpluses (averaging 46%) haveaccumulated at the two enhanced areas. Therefore, despite initial numericalgrowth and efforts directed toward distinct management of cultured and wildsegments, continuation of present trends by the BLDP will result in reductionsin complexity of subgroup structure, overall productivity, and geneticallyeffective population size. Indirect effects of the BLDP have been particularlyproblematic for non-Babine sockeye populations of the Skeena River, most ofwhich have declined in abundance since BLDP and remain well below minimalsustainable yield despite efforts to reduce their harvest (C. Wood, pers. com.PBS).
A similar trend has been linked to a hatchery program for winter steelhead onthe Stamp/Somass River where, over many years, native summer runs declinedin tandem with intensive hatchery releases. The situation was remedied whenhatchery releases were reduced (S. Pollard, pers. com.).
In closing, Principle Six of the Wild Salmon Policy warrants specialconsideration. At each hierarchical level of genetic interaction (Table 4-1),placing top priority on conserving wild salmon populations reduces geneticconcerns to zero. However, this priority inevitably conflicts with the goals ofsupplementation, as is evident in the BLDP (e.g., see discussion in Rymanet al. 1995). Erosion of substructure appears to be reversible if left alone;presumably, despite losses, enough genetically effective breeders are left(Figure 4-4).
The threatened extinction of non-Babine sockeye through BLDP-inducedmixed-stock fisheries is therefore theoretically reversible. In general, thelongterm viability and productivity of wild salmon populations may best besecured by turning the focus away from supplementation and towards habitatprotection and restoration. Such redirection will permit fuller operation of thenatural processes that have optimized salmon abundance since long before theadvent of humankind.
Conclusions
The Pinkett spawningchannel is one of 4 channelsthat comprise the Babine LakeDevelopment Project on theUpper Skeena River. ThePinkett spawning channelsproduced 25 million fry in2001. photo: Garth Traxler
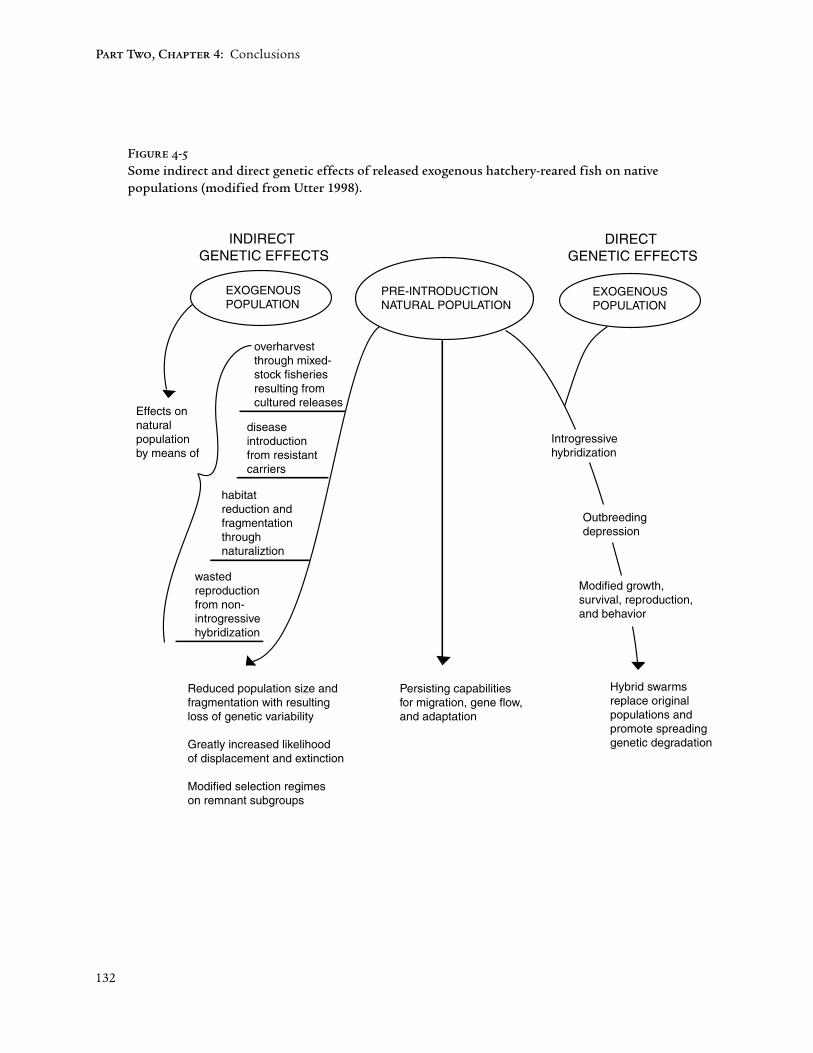
132
Figure 4-5
Some indirect and direct genetic effects of released exogenous hatchery-reared fish on nativepopulations (modified from Utter 1998).
INDIRECT GENETIC EFFECTS
EXOGENOUS POPULATION
Effects on natural population by means of
overharvest through mixed-stock fisheries resulting from cultured releases
disease introduction from resistant carriers
habitat reduction and fragmentation through naturaliztion
wasted reproduction from non-introgressive hybridization
Reduced population size and fragmentation with resulting loss of genetic variability
Greatly increased likelihood of displacement and extinction
Modified selection regimes on remnant subgroups
PRE-INTRODUCTION NATURAL POPULATION
Persisting capabilities for migration, gene flow, and adaptation
DIRECT GENETIC EFFECTS
EXOGENOUS POPULATION
Introgressive hybridization
Outbreeding depression
Modified growth, survival, reproduction, and behavior
Hybrid swarms replace original populations and promote spreading genetic degradation
Part Two, Chapter 4: Conclusions
133
POTENTIAL FOR GENETIC INTERACTION
Salmo- Among Among WithinOncorhychus Oncorhychus conspecific conspecific
Wild Salmon Principle interactions species lineages lineages
1. Wild Pacific salmon willbe conserved by maintaining ** ** *** ****diversity of local populationsand their habitats.
2. Wild Pacific salmon will bemanaged and conserved as 0 * *** ****aggregates of local populationscalled conservation units.
3. Minimum and target levels ofabundance will be determined * * *** ****for each conservation unit.
4. Fisheries will be managed toconserve wild salmon and 0 ** **** ****optimize sustainable benefits.
5. Salmon cultivation techniquesmay be used in strategic inter- 0 ** **** ****vention to preserve populationsat greatest risk of extirpation.
6. For specified conservationunits, when genetic diversity and 0 0 0 0longterm viability may be affected,conservation of wild salmonpopulations will take precedenceoverother production objectivesinvolving cultivated salmon.
Table 4-1 Matrix of potential genetic interactions of native andintroduced salmonid populations at different taxonomiclevels (columns), and principles of British Columbia WildSalmon policy (rows).
0 none, * low, ** moderate, *** high, **** extreme.

134
Alverson, D.L. and G.T. Ruggerone. 1997. Escaped farmed salmon: environmental and ecologicalconcerns. Pages 1-99 in, Salmon aquaculture review. Volume 3. Part B. British ColumbiaEnvironmental Assessment Office.
Campton, E.E., F.W. Allendorf, B.J. Behnke, F.M. Utter, M.W. Chilcote, S.A Leider and J.J. Loch.1991. Comments: reproductive success of hatchery and wild steelhead. Transactions of theAmerican Fisheries Society 120:816-827.
Carlson, J. 1998. A lot more than species is endangered. The Sun, Bremerton, Washington, U.S.A.March 4th, 1998.
Chevassus, B., R. Guyomard, D. Chourrout, and E. Quillet. 1983. Production of viable hybrids insalmonids by triploidization. Genet. Sel. Evol. 15:519-532.
Egan, D. 2000. Salmon farming overview: 1999. Figures from presentation to the B.C. SalmonFarmers Association Annual General Meetings, Vancouver, B.C. June 15, 2000.
Fisheries and Oceans Canada. 1995. Wild salmon policy: a discussion paper. Available http://www-comm.pac.dfo-mpo.gc.ca/development/wsp-sep-consult/wsp/wsp1.htm.
Foote, C.J., C.C. Wood, and R.E. Withler. 1989. Biochemical genetic comparison of sockeyesalmon and kokanee, the anadromous and non-anadromous forms of Oncorhynchus nerka.Canadian Journal of Fisheries and Aquatic Sciences 46:149-158.
Gharrett, A.J. and W.W. Smoker. 1993. Genetic components in life history traits contribute topopulation structure. Pages 197-202 in, Genetic Conservation of Salmonid Fisheries (J. Cloudand G.H. Thorgaard, eds). Plenum Press, New York.
Gross, M.R. 1998. One species with two biologies: Atlantic salmon (Salmo salar) in the wild andin aquaculture. Canadian Journal of Fisheries and Aquatic Sciences 55 (Suppl. 1):131-144.
Gustafson, R.G. and G.A. Winans. 1999. Distribution and population genetic structure of river-and sea-type sockeye salmon in western North America. Ecology of Freshwater Fish 8:181-193.
Hawkins, D. 1997. Hybridization between coastal cutthroat trout (Oncorhynchus clarki clarki) andsteelhead (O. mykiss). PhD dissertation, University of Washington.
Hendry, A.P., J.K. Wenburg, P. Bentzen, E.C. Volk and T.P. Quinn. 2000. Rapid evolution ofreproductive isolation in the wild: evidence from introduced salmon. Science 290:516-520.
Hindar, K., N. Ryman and F. Utter. 1991. Genetic effects of cultured fish on natural fishpopulations. Canadian Journal of Fisheries and Aquatic Sciences 48:945-957.
Johnson, O.W., W.S. Grant, R.G. Kope, K. Neely, F.W. Waknitz and R.S. Waples. 1997. Statusreview of chum salmon from Washington, Oregon, and California. U.S. Department ofCommerce. NOAA Technical Memorandum NMFS-NWFSC-32.
Kinnison, M.T., M. Unwin, N. Boustead, and T. Quinn. 1998. Population-specific variation inbody dimensions of adult chinook salmon (Oncorhynchus tshawytscha) from New Zealand andtheir source population, 90 years after introductions. Canadian Journal of Fisheries andAquatic Sciences 55:5554-5563.
Leary, R.F., F.W. Allendorf and K.G. Sage, 1995. Hybridization and introgression betweenintroduced and native fish. Pages 91-101 in, Uses and effects of cultured fishes in aquaticecosystems (J.L. Schramm and R.G. Piper, eds.). American Fisheries Society Symposium 15.Bethesda, Maryland.
McGregor, A.J. 1982. A biochemical genetic analysis of pink salmon (Oncorhynchus gorbuscha)from selected streams in northern Southeast Alaska. Master of Science thesis, University ofAlaska, Juneau. 94 pp.
Phelps, S.R., L.L. LeClair, S. Young, and H.L. Blankenship. 1994. Genetic stock structure ofwestern Alaska chum salmon and a comparison with Russian far east stocks. CanadianJournal of Fisheries and Aquatic Sciences. 51 (Suppl. 1):65-83.
Quinn, T., J.L. Nielsen, C. Gan, M.J. Unwin, R. Wilmot, C. Guthrie and F.M. Utter. 1996. Originand genetic structure of chinook salmon (Oncorhynchus tshawytscha) transplanted fromCalifornia to New Zealand: allozyme and mtDNA evidence. Fishery Bulletin 94:506-521.
References
Part Two, Chapter 4: References

135
Reisenbichler, R.R. and J. McIntyre. 1977. Genetic differences in growth and survival of juvenilehatchery and wild steelhead trout, Salmo gairdneri. Journal of the Fisheries Research Board ofCanada 34:123-128.
Reisenbichler, R.R. and S.P. Rubin. 1999. Genetic changes from artificial propagation of Pacificsalmon affect the productivity and viability of supplemented populations. (ICES Journal ofMarine Science 56:459-466.
Robertson, O.H. 1961. Prolongation of the life span of kokanee salmon (Oncorhynchus nerkakennerlyi) by castration before beginning of gonal development. Proceedings of the NationalAcademy of Sciences, U.S.A. 47:609-621.
Rutridge, E.B., P. Corbett and E.B. Taylor. 2001. A molecular analysis of hybridization betweennative west slope cutthroat trout and introduced rainbow trout in Southeastern BritishColumbia, Canada. Journal of Fish Biology 59 (supplement A): 42-54.
Ryman, N. and L. Laikre. 1991. Effects of supportive breeding on the genetically effectivepopulation size. Conservation Biology 5:325-329.
Ryman, N., F.M. Utter and L. Laikre. 1995. Protection of intraspecific biodiversity of exploitedfishes. Reviews in Fish Biology and Fisheries 5:417-446.
Seeb, J.E., C. Habicht, W.D. Templin, L.W. Seeb, J. Shaklee and F.M. Utter. 1999. Allozyme andmitochondrial DNA variation describe ecologically important genetic structure of even-yearpink salmon inhabiting Prince William Sound, Alaska. Ecology of Freshwater Fish 8:122-140.
Shaklee, J.B., D.C. Klaybor, S. Young and B.A. White. 1991. Genetic stock structure of odd-yearpink salmon Oncorhynchus gorbuscha (Walbaum), from Washington and British Columbia andpotential mixed-stock fisheries applications. Journal of Fish Biology 39 (suppl. A):21-34.
Teel, D.J., G.B. Milner, G.A. Winans, and W.S. Grant. 2000. Genetic population structure andorigin of life history types in chinook salmon in British Columbia, Canada. Transactions ofthe American Fisheries Society 129:194-209.
Utter, F.M. In press. Genetic impacts of fish introductions. In, Genetic Principles and Practicesfor Fisheries Scientists (E. Hallerman, Ed).
Utter, F.M. 1998. Genetic problems of hatchery-reared progeny released into the wild, and howto deal with them. Bulletin of Marine Science 62:623-640.
Utter, F.M. 2001. Patterns of subspecific anthropogenic introgression in two salmonid genera.Reviews in Fish Biology and Fisheries 10:265-279.
Utter, F.M. 1991. Biochemical genetics and fishery management: an historical perspective.Journal of Fish Biology 39 (Suppl A):1-20.
Utter, F.M. and F.W. Allendorf. 1994. Phylogenetic relationships among species of Oncorhynchus:a concensus view. Conservation Biology 8:864-867.
Utter, F.M. and N. Ryman. 1993. Genetic markers and mixed-stock fisheries. Fisheries 18(8):11-21.
Utter, F.M., G.B. Milner, G. Stahl, and D. Teel. 1989. Genetic population structure of chinooksalmon, Oncorhynchus tshawytscha, in the Pacific Northwest. Fishery Bulletin 87:239-264.
Utter, F.M., R.S. Waples and D. Teel. 1992. Genetic isolation of previously indistinguishablechinook salmon populations of the Snake and Klamath rivers: limitations of negative data.Fishery Bulletin 90:770-777.
Utter, F.M., J.E. Seeb and L.W. Seeb. 1993. Complementary uses of ecological and biochemicalgenetic data in identifying and conserving salmon populations. Fisheries Research 18: 59-76.
Utter, F.M., D. Chapman and A. Marshall. 1995. Genetic population structure and history ofchinook salmon of the upper Columbia River. Pages 149-165 in, Evolution and the AquaticEcosystem: Defining Unique Units in Population Conservation (J. Nielsen, ed). AmericanFisheries Society Symposium 17, Bethesda, Maryland.
Varnavskaya, N.V., C.C. Wood, R.J. Everett, R.L. Wilmot, V.S. Varnavsky, V.V. Midanaya, andT.P. Quinn. 1994. Genetic differentiation of subpopulations of sockeye salmon (Oncorhynchusnerka) within lakes of Alaska, British Columbia, and Kamchatka, Russia. Canadian Journal ofFisheries and Aquatic Sciences (Suppl. 1)51:147-157.

136
Volpe, J.P. 2000. Do we know what we don’t know: Atlantic salmon in British Columbia: areview, pages 28-33 in, Aquaculture and the protection of wild salmon (P. Gallaugher and C.Orr, eds) Simon Fraser University.
Volpe, J.P. 2001. Super-unnatural. Report published by the David Suzuki Foundation.Vancouver, BC.
Volpe, J.P., E.B. Taylor, D.W. Rimmer and B.W. Glickman. 2000. Evidence of naturalreproduction of aquaculture escaped Atlantic salmon (Salmo salar) in a coastal BritishColumbia river. Conservation Biology 14:899-904.
Waples, R.S. 1995. Genetic effects of stock transfers of fish. Pages 51-69 in, Proceedings of theWorld Fisheries Congress, Theme 3. Oxford and IBH, New Delhi.
Waples, R.S. 1999. Dispelling some myths about hatcheries. Fisheries 24(2):12-21.
Waples, R.S. and C. Do. 1994. Genetic risks associated with supplementation of Pacificsalmonids: captive broodstock programs. Canadian Journal of Fisheries and Aquatic Sciences51 (Suppl. 1):310-329.
Waples, R.S. and D. Teel. 1990. Conservation genetics of Pacific salmon. I. Temporal changes inallele frequency. Conservation Biology 4:144-156.
Withler, F.C. 1982. Transplanting Pacific salmon. Canadian Technical Report of Fisheries andAquatic Sciences 1079.
Wood, C.C. 1995. Life history variation and population structure in sockeye salmon. Pages 195-216 in, Evolution and the Aquatic Ecosystem: defining unique units in populationconservation (J. Nielsen, ed). American Fisheries Society Symposium 17, Bethesda, Maryland.
Wood, C.C., D.T. Rutherford, D. Bailey and M. Jakubowski. 1998. Assessment of sockeye salmonproduction in Babine Lake, British Columbia with forecast for 1998. Canadian TechncialReport of Fisheries and Aquatic Sciences 2241.
Wood, C.C. Personal communication. Pacific Biological Station. Department of Fisheries &Oceans. February, 2001.
Part Two, Chapter 4: References





























