Embed Size (px)
Citation preview

Kim “Avrama” BlackwellGeorge Mason University
Modeling Signaling Pathways underlying
Synaptic Plasticity

Importance of Signaling Pathways
• Neuromodulators, e.g. Dopamine and Norepinphrine, modulate channel behaviour via intracellular signaling pathways
• Synaptic plasticity, cell excitability, gene regulation and memory are controlled by intracellular signaling pathways
• Intracellular signaling pathways are modelled as biochemical reactions

Mammalian Synaptic Plasticity
• Long Term Synaptic Plasticity– Long lasting, activity dependent change in
synaptic strength– Duration is one hour or more
• Potentiation - increase in synaptic strength
• Depression – decrease in synaptic strength
– Persistence and activity dependence of change makes this an attractive mechanism for memory storage
5 ms
1 mV
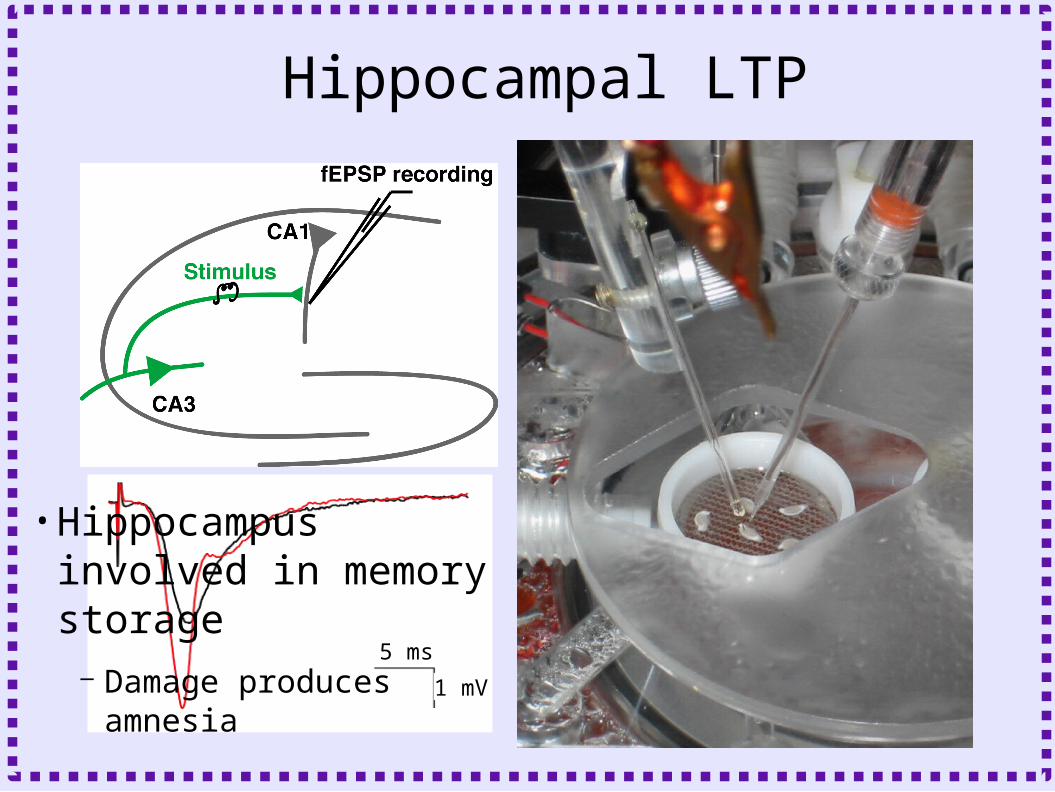
Hippocampal LTP
•Hippocampus involved in memory storage
– Damage produces amnesia

Long-term potentiation of Schaffer collateral-CA1 synapses
• High frequency stimulation of CA1 afferents enhances synaptic transmission – Repetitions of 1 sec of 100 hz– Pathway 1 receives high
frequency stimulation– Stimulus 2 is unstimulated
– High frequency stimulation required for firing of post-synaptic neuron (Hebb’s postulate)
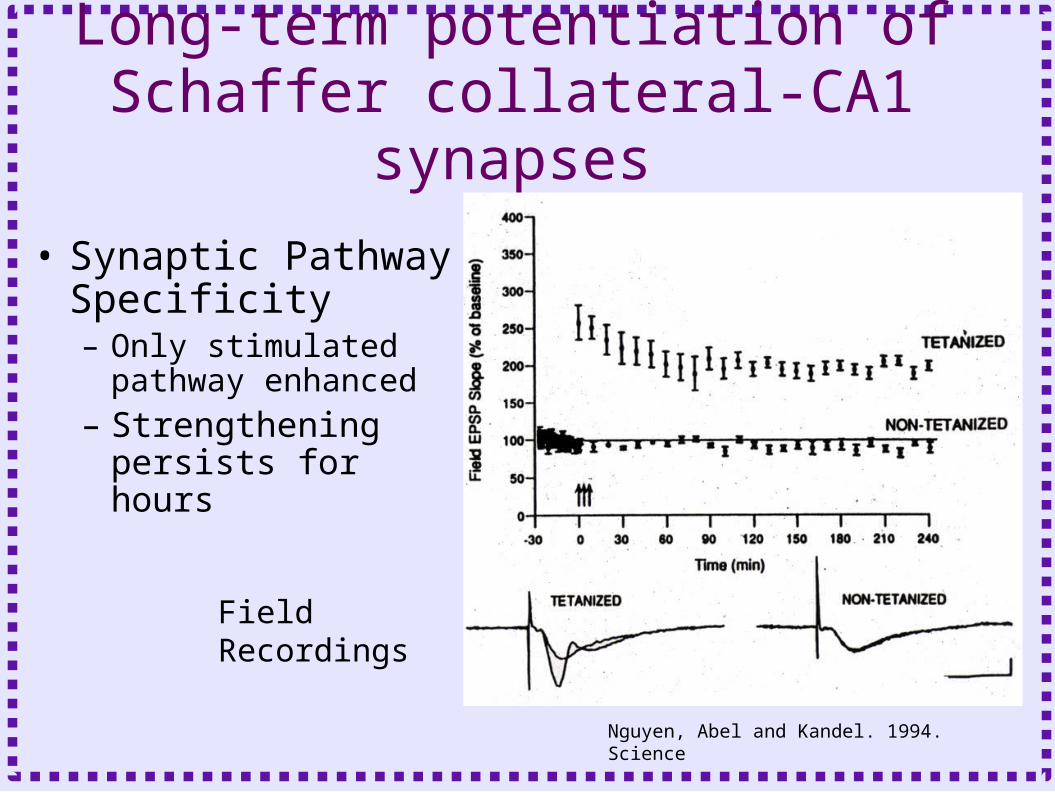
Long-term potentiation of Schaffer collateral-CA1 synapses
• Synaptic Pathway Specificity– Only stimulated pathway
enhanced– Strengthening persists
for hours
Field Recordings
Nguyen, Abel and Kandel. 1994. Science
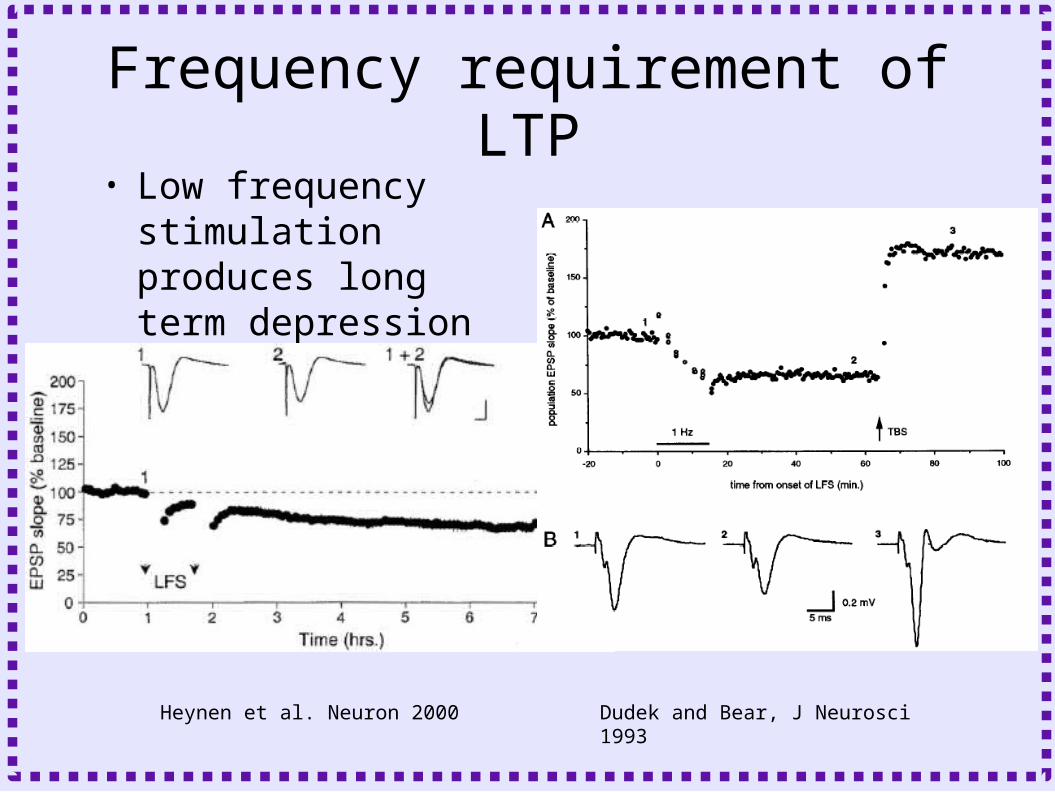
Frequency requirement of LTP• Low frequency
stimulation produces long term depression
Heynen et al. Neuron 2000 Dudek and Bear, J Neurosci 1993
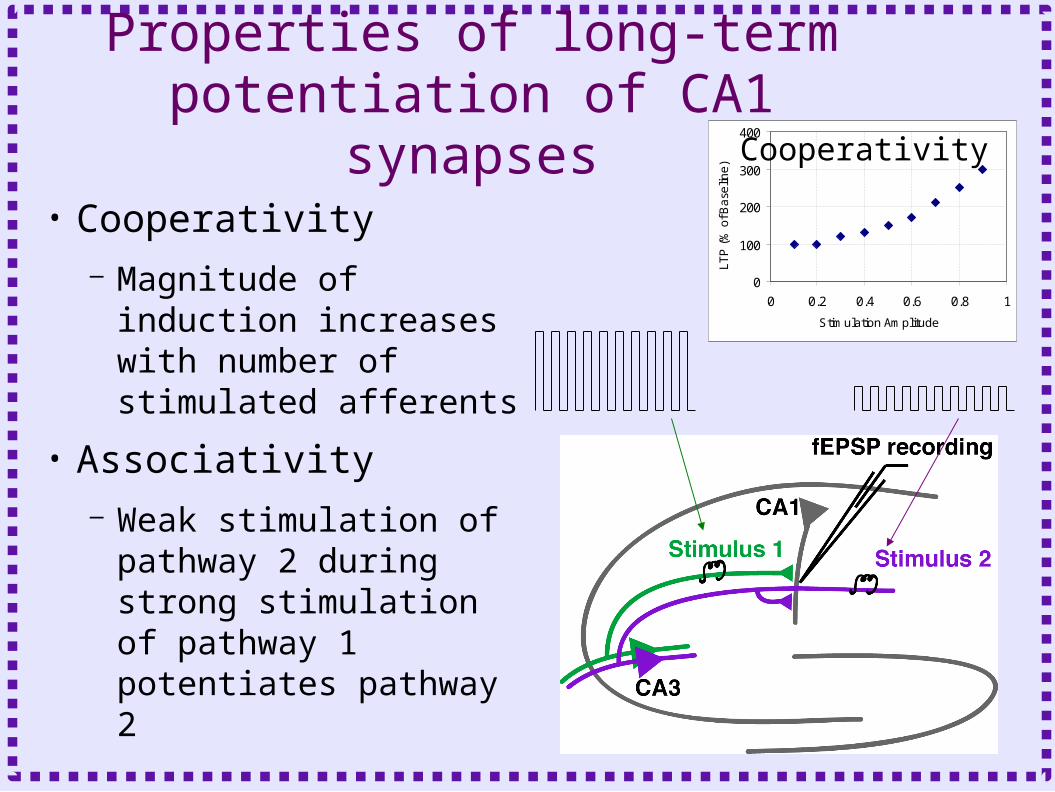
Properties of long-term potentiation of CA1 synapses
• Cooperativity– Magnitude of induction
increases with number of stimulated afferents
• Associativity– Weak stimulation of
pathway 2 during strong stimulation of pathway 1 potentiates pathway 2
0
100
200
300
400
0 0.2 0.4 0.6 0.8 1
Stimulation Amplitude
LT
P (
% o
f Ba
selin
e) Cooperativity

Mechanisms Underlying LTP
• Depolarization of post-synaptic neuron (Hebb’s postulate) explains many properties of LTP– Cooperativity: multiple fibers required for
sufficient depolarization of post-synaptic neuron– Frequency dependence – high frequency required
for depolarization to accumulate– Associativity – high frequency of strong pathway
produces depolarization for weak pathway• Pre-synaptic activation of weak pathway required for
glutamate release
• NMDA receptor channel explains depolarization requirement
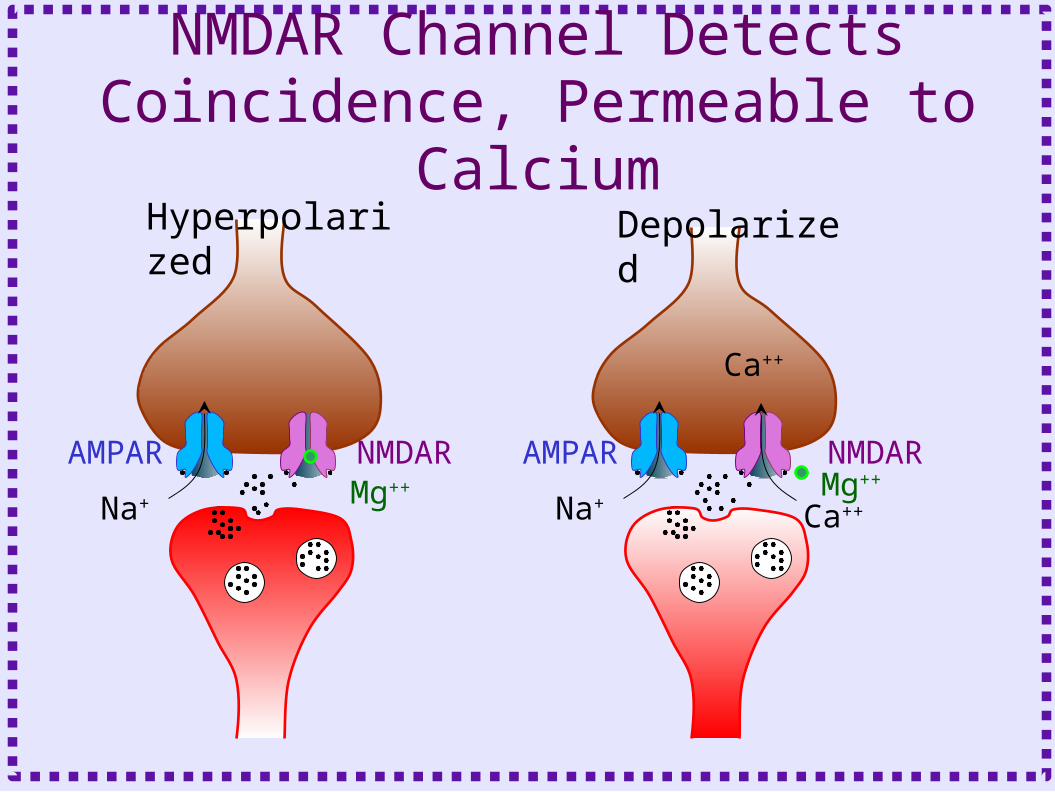
NMDAR Channel Detects Coincidence, Permeable to Calcium
AMPAR NMDARMg++
Na+
AMPAR NMDARMg++
Na+
Hyperpolarized Depolarized
Ca++
Ca++
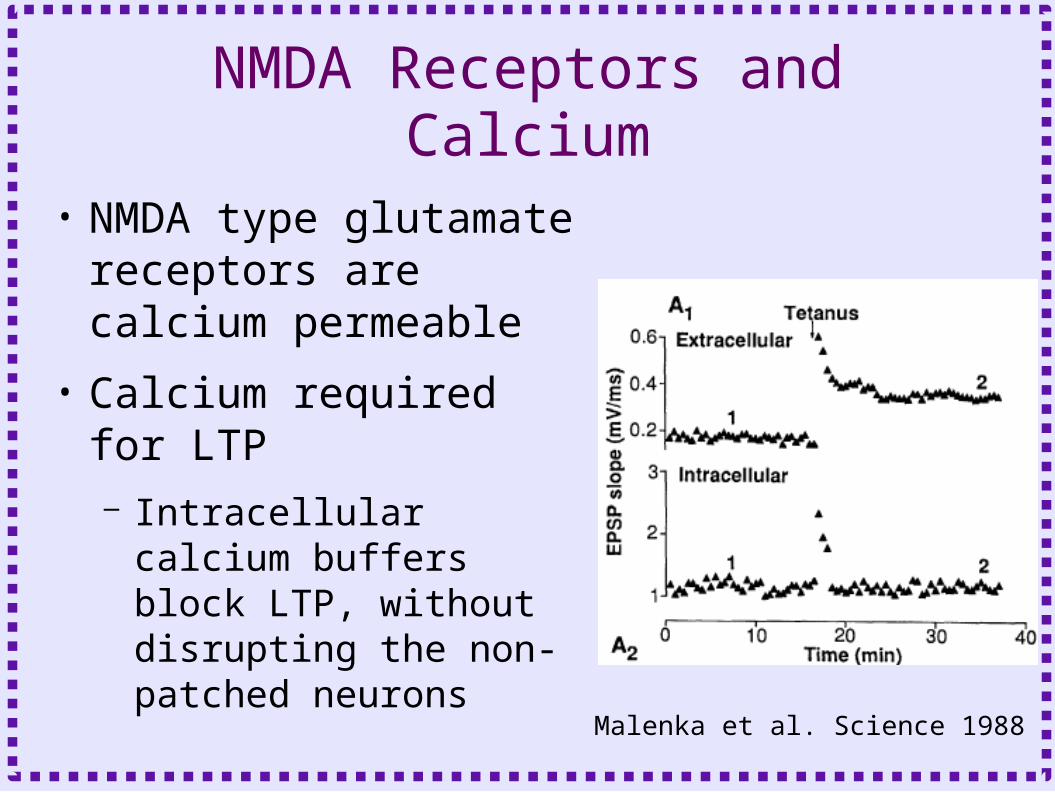
NMDA Receptors and Calcium
• NMDA type glutamate receptors are calcium permeable
• Calcium required for LTP– Intracellular calcium
buffers block LTP, without disrupting the non-patched neurons
Malenka et al. Science 1988
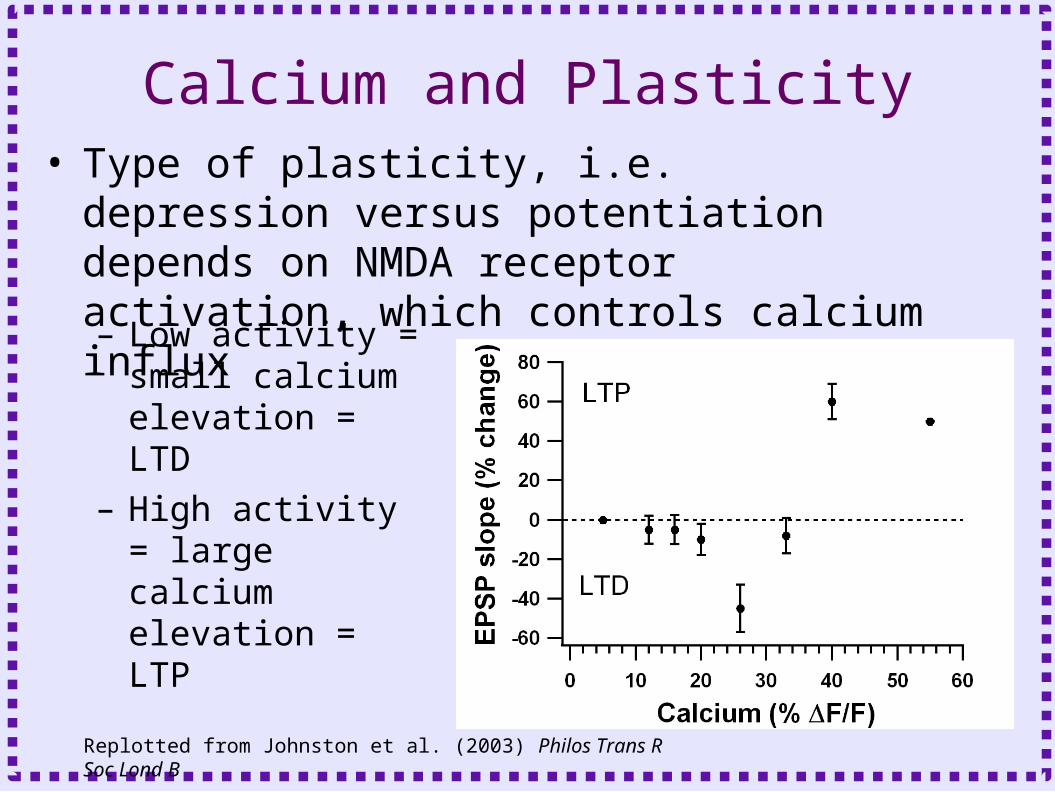
Calcium and Plasticity• Type of plasticity, i.e. depression versus
potentiation depends on NMDA receptor activation, which controls calcium influx
– Low activity = small calcium elevation = LTD
– High activity = large calcium elevation = LTP
Replotted from Johnston et al. (2003) Philos Trans R Soc Lond B
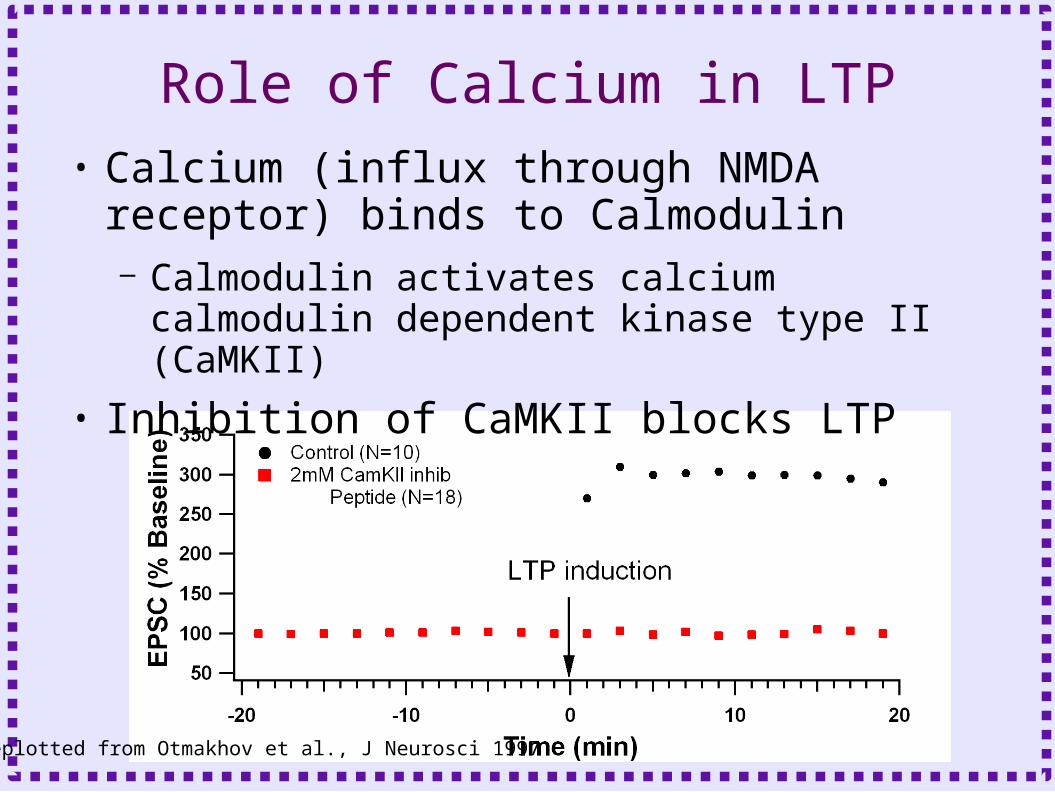
Role of Calcium in LTP• Calcium (influx through NMDA receptor)
binds to Calmodulin– Calmodulin activates calcium calmodulin
dependent kinase type II (CaMKII)
• Inhibition of CaMKII blocks LTP
Replotted from Otmakhov et al., J Neurosci 1997
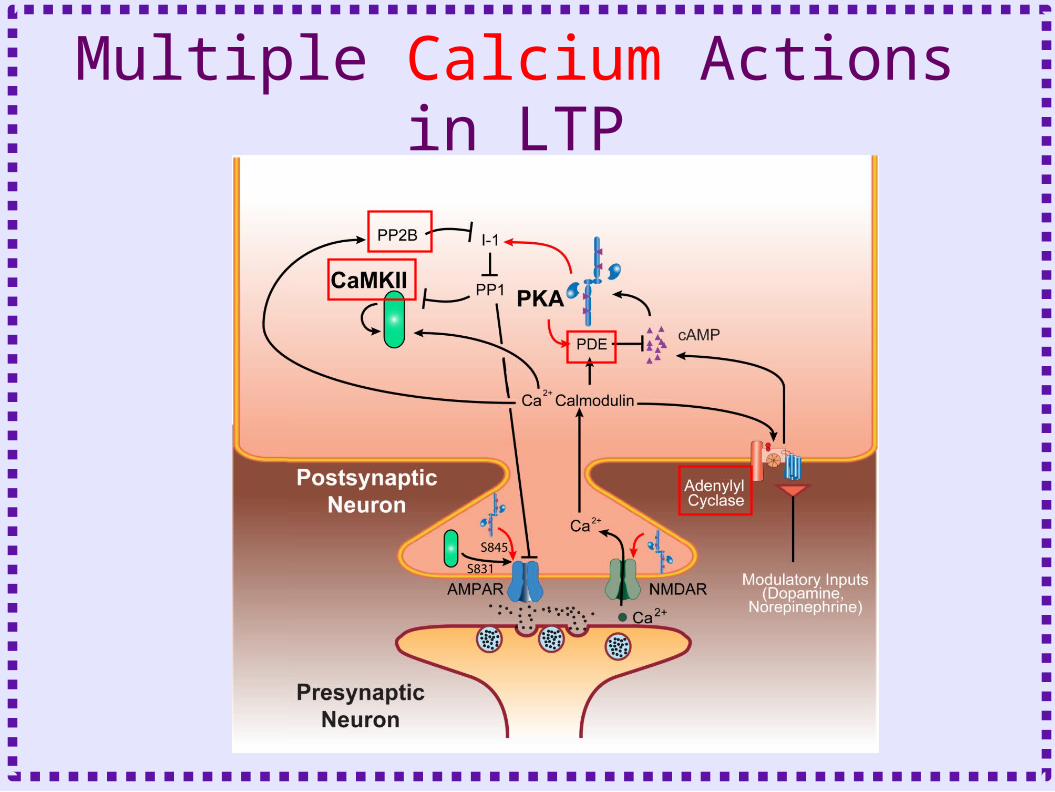
Multiple Calcium Actions in LTP
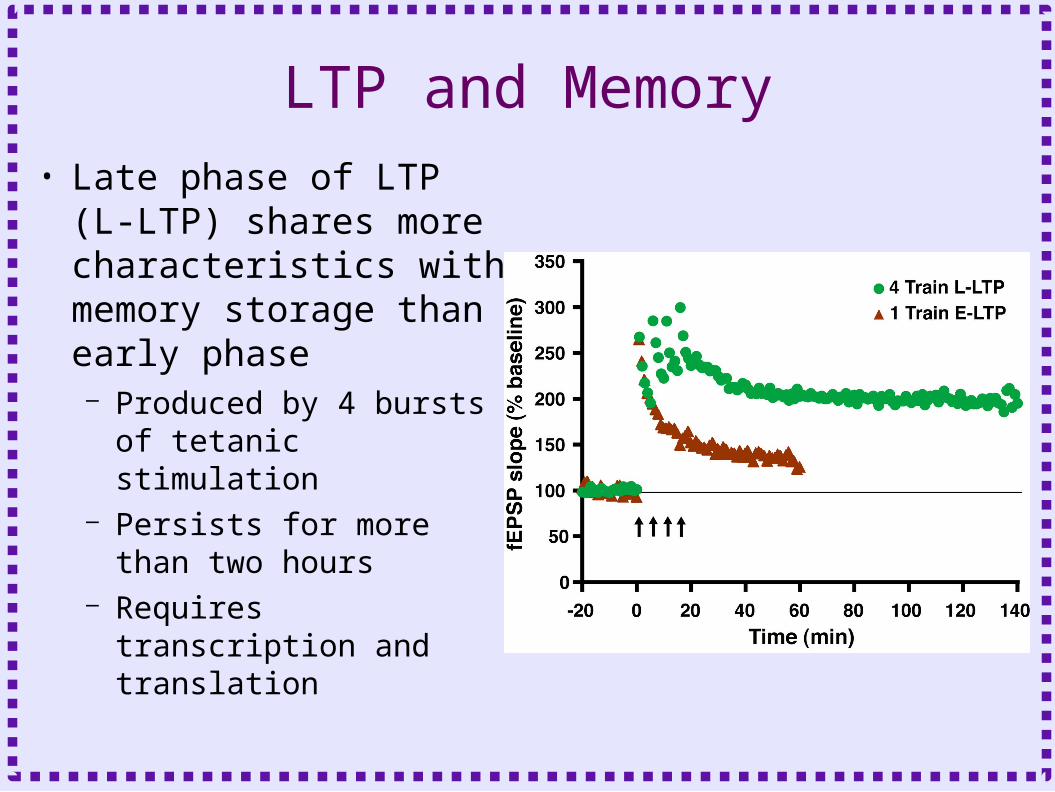
LTP and Memory• Late phase of LTP (L-
LTP) shares more characteristics with memory storage than early phase– Produced by 4 bursts of
tetanic stimulation– Persists for more than
two hours– Requires transcription
and translation
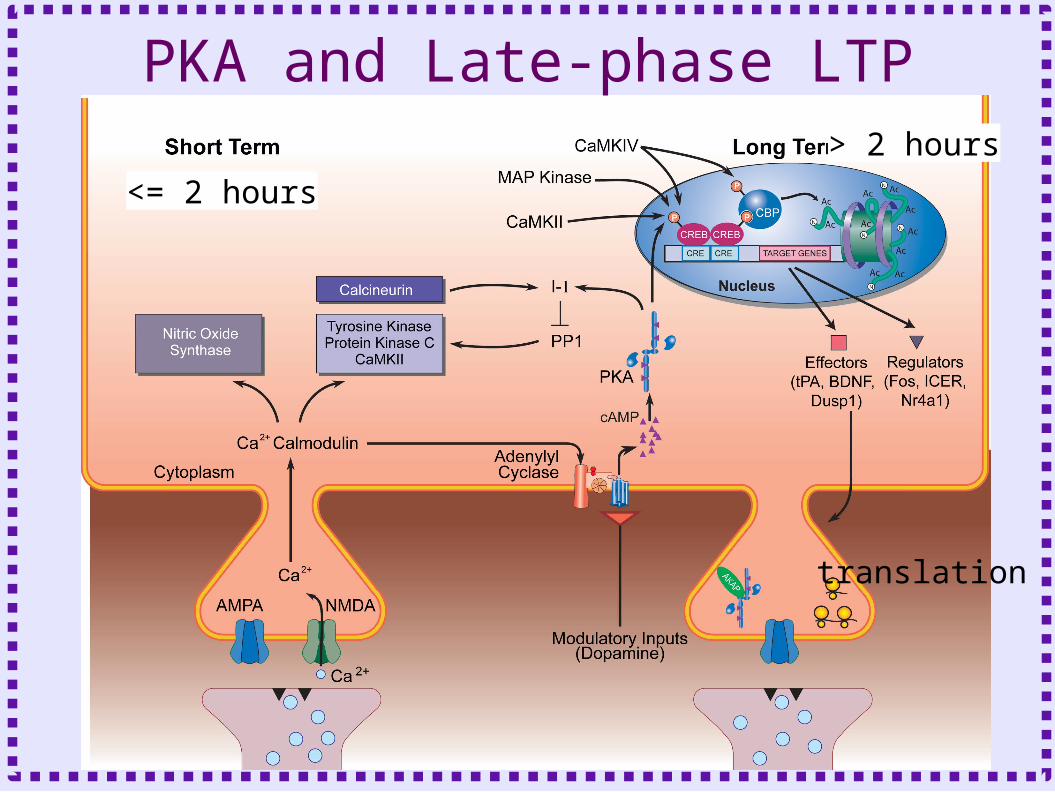
PKA and Late-phase LTP
<= 2 hours
> 2 hours
translation

STDP• Spike Timing Dependent Plasticity
– Recent experiments show that relative timing of action potentials plays a critical role in determining sign and amplitude of changes in synaptic efficacy
• These experiments typically involve paired intracellular recordings– AP induced in pre-synaptic neuron
• Release of Glutamate– AP induced in post-synaptic neuron
• Requires AP to propagate backward into the dendritic tree
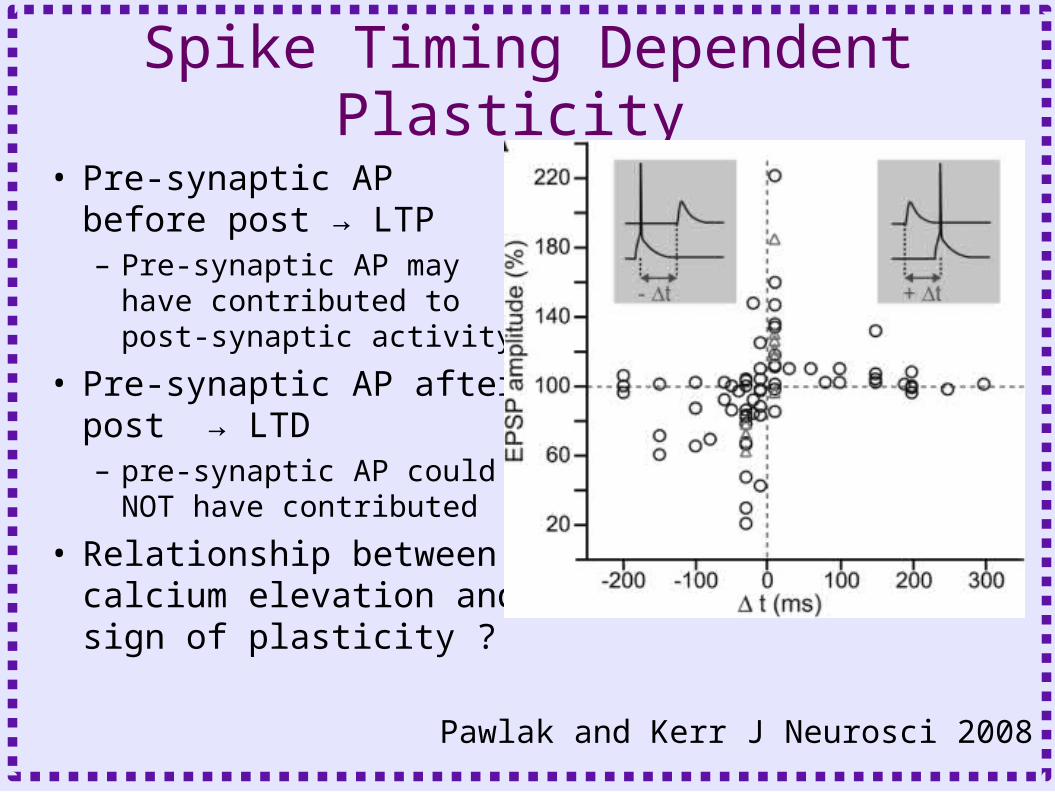
Spike Timing Dependent Plasticity
• Pre-synaptic AP before post → LTP– Pre-synaptic AP may have
contributed to post-synaptic activity
• Pre-synaptic AP after post → LTD– pre-synaptic AP could NOT
have contributed
• Relationship between calcium elevation and sign of plasticity ?
Pawlak and Kerr J Neurosci 2008
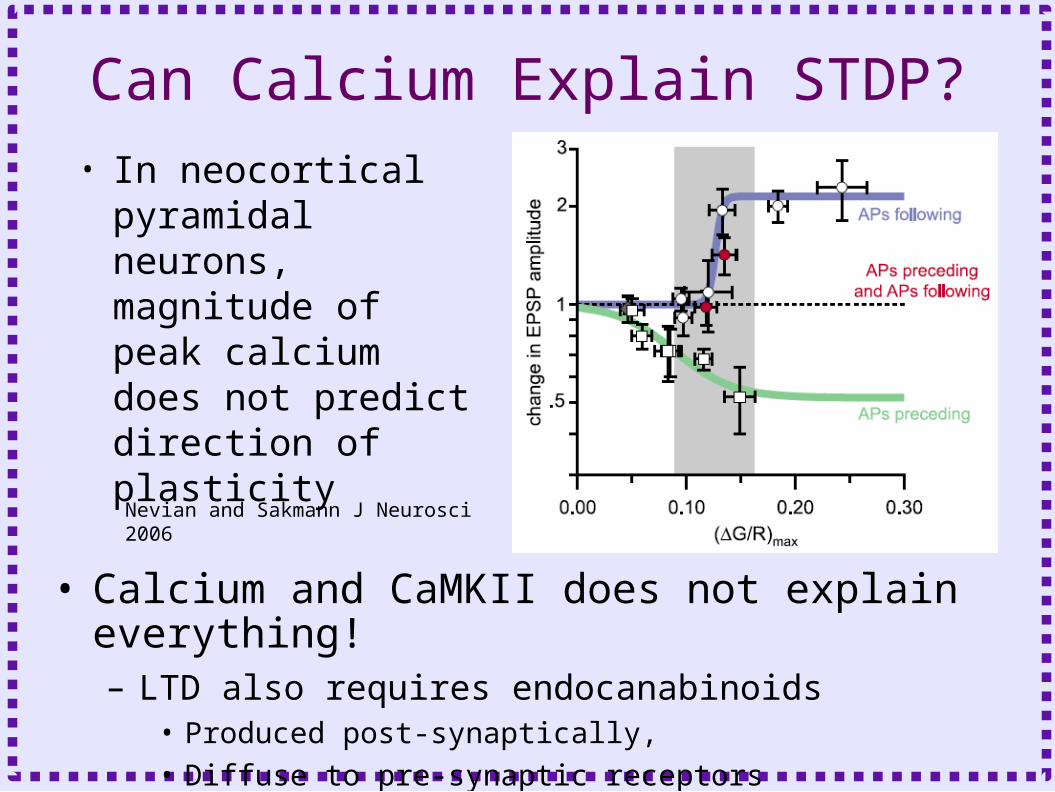
Can Calcium Explain STDP?• In neocortical
pyramidal neurons, magnitude of peak calcium does not predict direction of plasticity
• Calcium and CaMKII does not explain everything!– LTD also requires endocanabinoids
• Produced post-synaptically,
• Diffuse to pre-synaptic receptors
Nevian and Sakmann J Neurosci 2006

How to Model Signaling Pathways
• Identify and describe biochemical reactions comprising the signaling pathway– Metabotropic Receptors– G proteins– Membrane bound enzyme– Diffusible second messenger– Kinase or phosphatase activation

How to Model Signaling pathways
• Metabotropic Receptors– Protein does not form channel– Protein is linked to GTP binding protein (G
protein)– Effect mediated by
• Activated G protein subunits
• Downstream second messengers
– Receptor bound to neurotransmitter is an enzyme which activates G protein
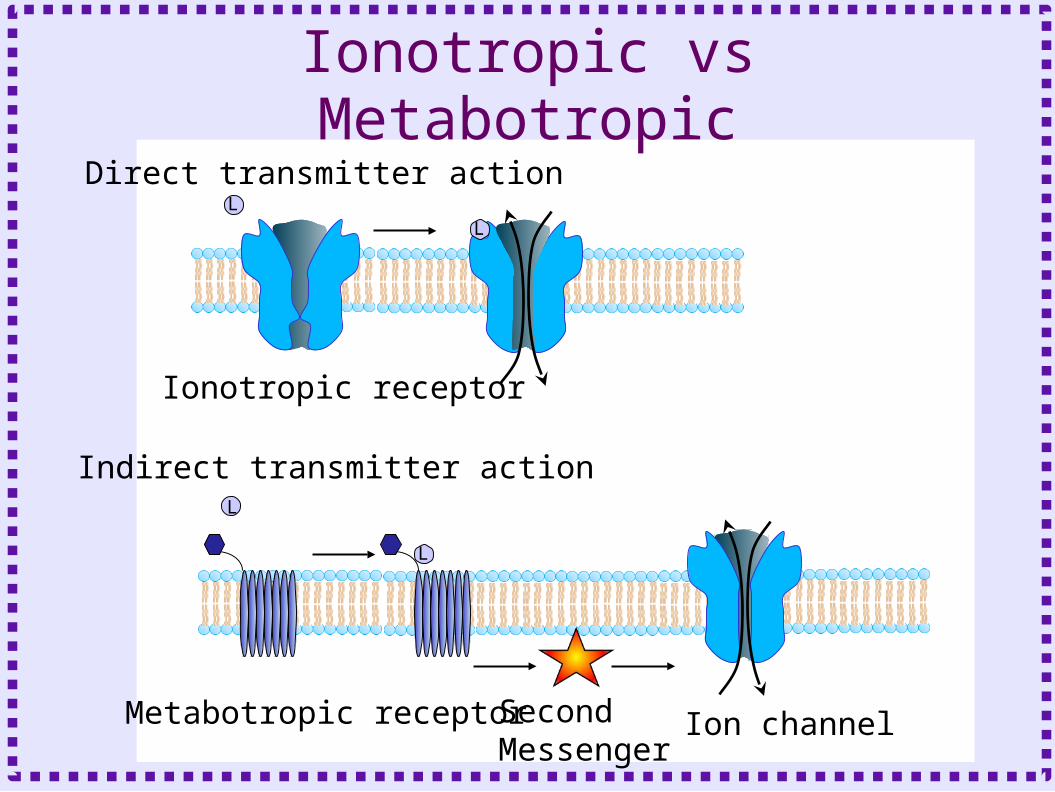
Ionotropic vs Metabotropic
LL
Direct transmitter action
Indirect transmitter action
Ionotropic receptor
Metabotropic receptor Ion channelSecond Messenger
L
L

Heterotrimeric GTP Binding Proteins• Binds to GTP or GDP
– GDP bound form is inactive– GTP bound form is active
• Three subunits– Alpha
• Binds to guanosine nucleotides: GDP or GTP• Many different subtypes
– Beta and Gamma
• Binds to alpha subunit, prevents it from interacting with effector
• Stabilizes G protein in membrane • Can be effectors
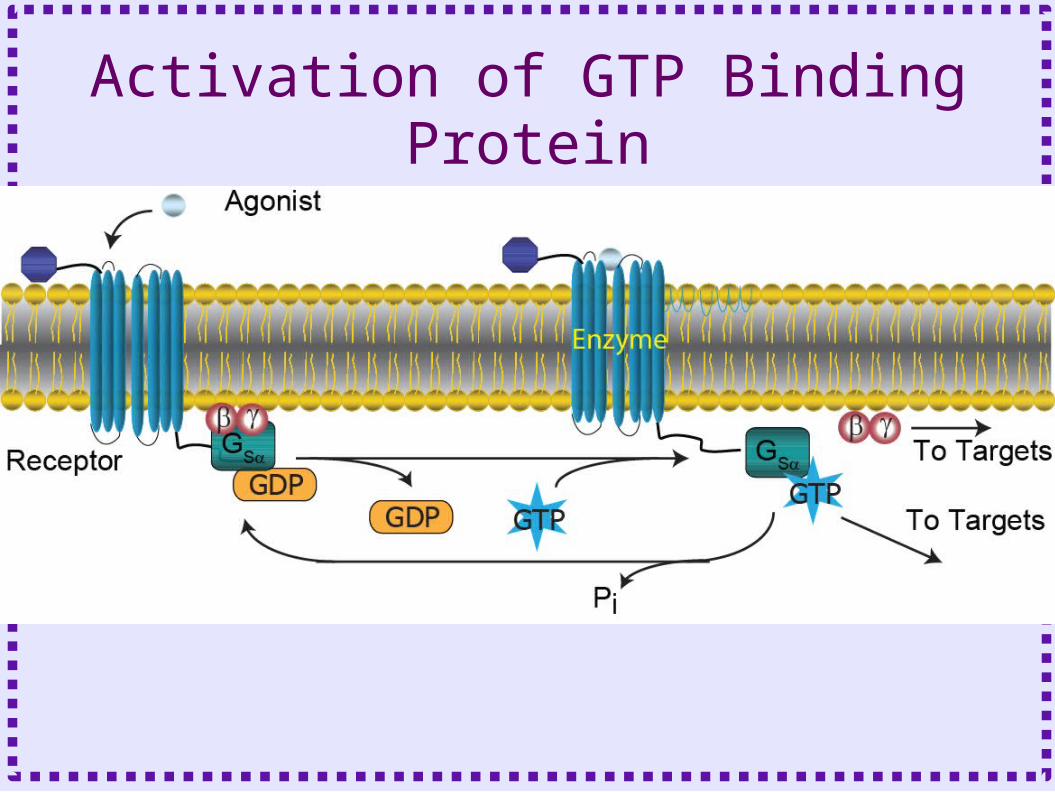
Activation of GTP Binding Protein

Direct and Indirect Action of G Proteins
• Direct action – G subunit directly gates channel– Limited spatial extent– Usually G
• Indirect action– G protein binds to enzyme– Enzyme produces intracellular second
messenger– Wide spatial extent due to diffusible
second messenger
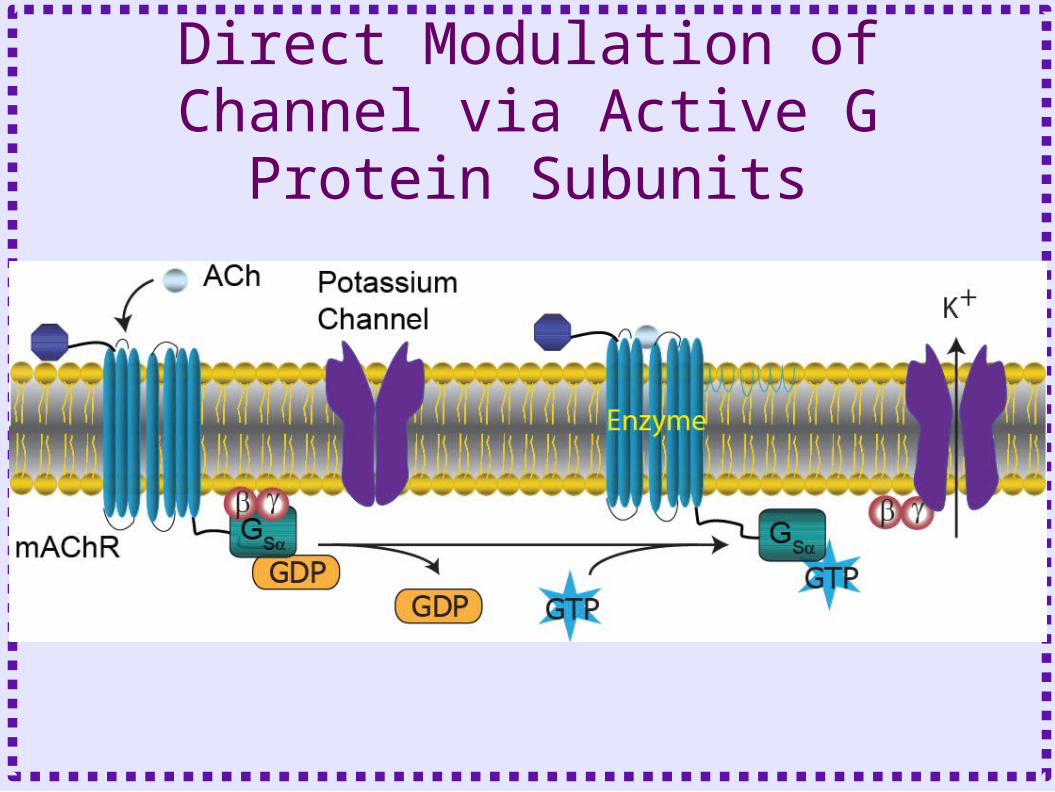
Direct Modulation of Channel via Active G Protein Subunits
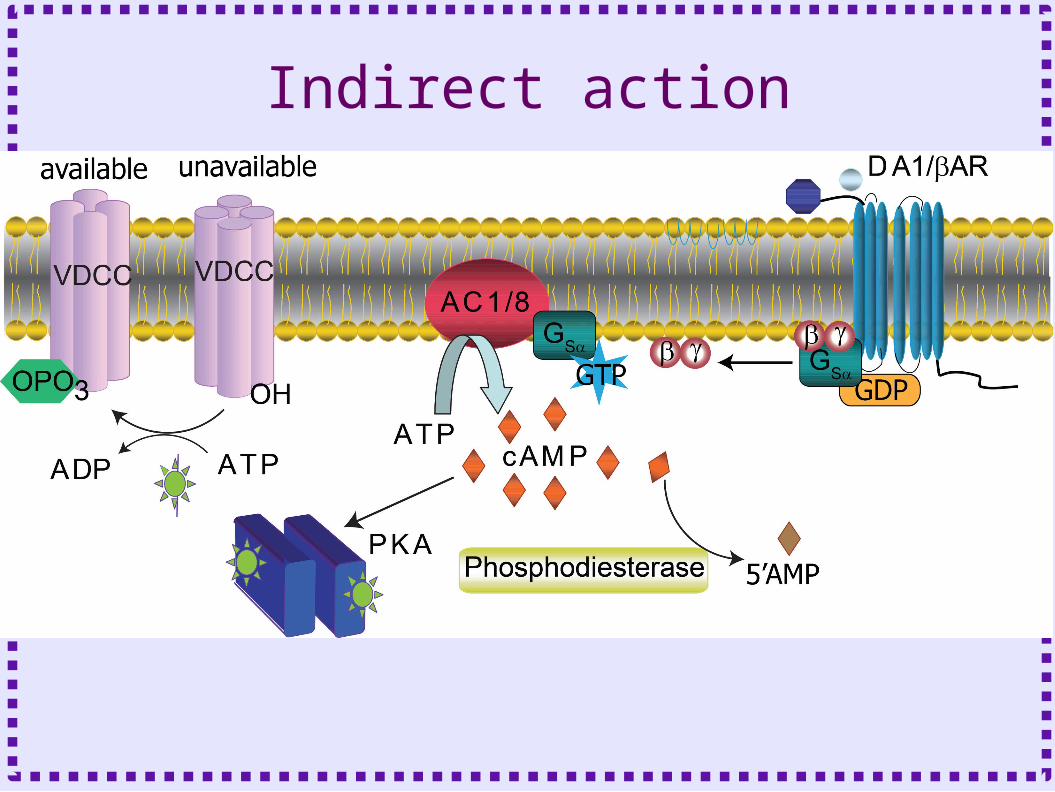
Indirect action

Enzymes Activated by G proteins
• Adenylyl Cyclase– Also activated by calcium-calmodulin– Produces cAMP– Activates protein kinase A– Activates cyclic nucleotide gated channels (IH)
• Phospholipase C– Produces diacylgylcerol and Inositol
triphosphate– DAG activates protein kinase C– IP3 causes calcium release from the ER
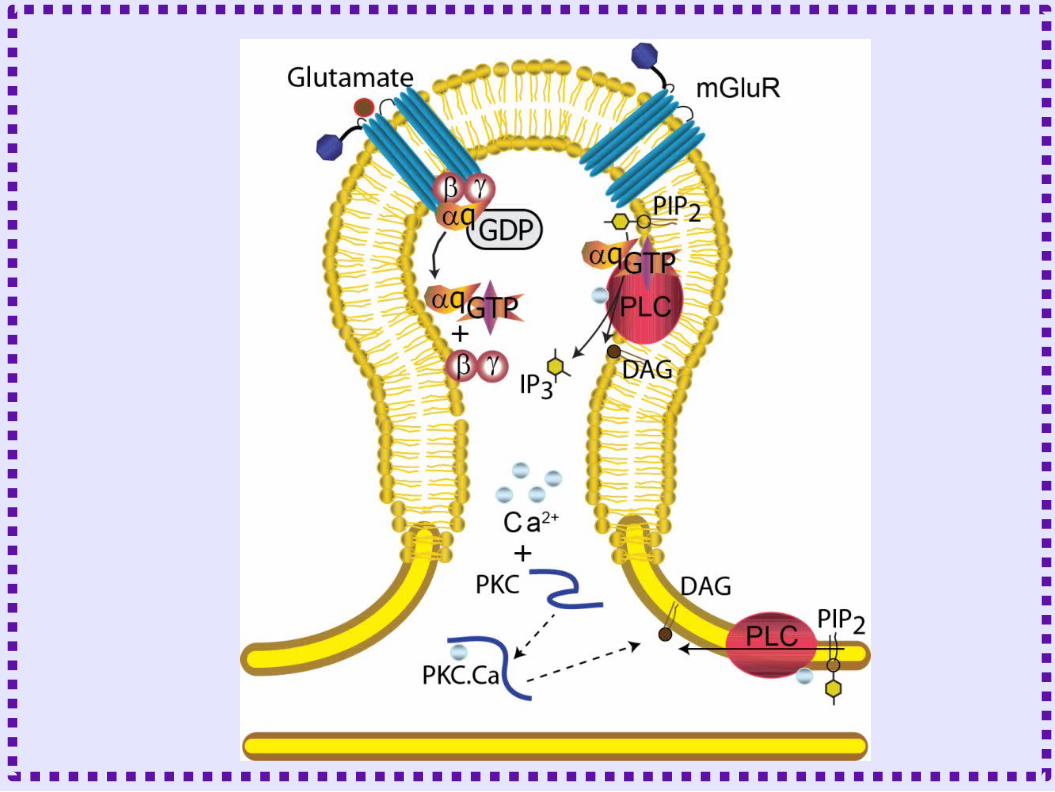

Biochemical Reactions
• Bimolecular Reactions– Stoichiometric interactions between
substrate molecules to form product molecule
• Formation of bond between the substrate molecules
• Stoichiometric implies that the reaction specifies the number of each molecule required for reaction
• Molecules are consumed in order to make product

Biochemical Reactions
• Bimolecular Reactions– Reaction order is the number of
simultaneously interacting molecules• First order reaction: single substrate becomes
product
• Rate constants: rate (units: per sec) at which substrate becomes product
• Ratio of rate constants gives concentration of substrates and products at equilibrium

substra te product
product
substra te
K
K
K
K
b
f
f
b
[ ]
[ ]
• First order reaction:
• At equilibrium:
Bimolecular Reactions

Bimolecular Reactions• Differential equations express rate of
change of molecule quantity with respect to time– Rate constants give frequency of transitions– Equations describe behavior of large
numbers of molecules (mass action kinetics)
– In closed system, mass is conserved, thus:• Substrate = initial value - produce
dproduct
d tK substra te K productf b [ ] [ ]

• Second order reaction:
• Each molecule of product requires 1 molecule of subs1 and 1 molecule of subs2
• Conservation of mass applies to both substrates– Subs1(t) = subs1(t=0) - product(t)– Subs2(t) = subs1(t=0) - product(t)
subs subs product
K
K
f
b
1 2
dproduct
d tK subs subs K productf b [ ][ ] [ ]1 2
Bimolecular Reactions

• Third order reaction:
• Order of reaction is number of molecules needed for product
• Substrate 2 is consumed twice as fast as substrate 1– Subs1(t) = subs1(t=0) - product(t)– Subs2(t) = subs1(t=0) - 2 product(t)
subs subs product
K
K
f
b
1 2 2
dproduct
d tK subs subs K productf b [ ][ ] [ ]1 2 2
Bimolecular Reactions






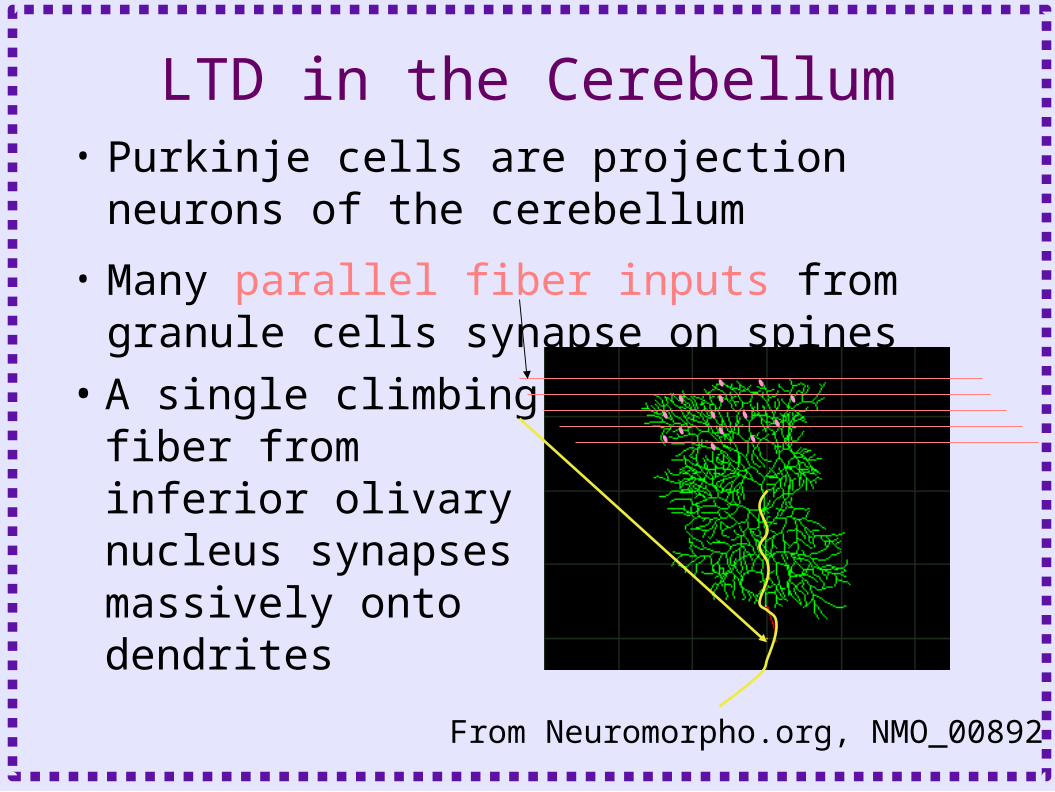
LTD in the Cerebellum• Purkinje cells are projection neurons of
the cerebellum
• Many parallel fiber inputs from granule cells synapse on spines
• A single climbing fiber from inferior olivary nucleus synapses massively onto dendrites
From Neuromorpho.org, NMO_00892
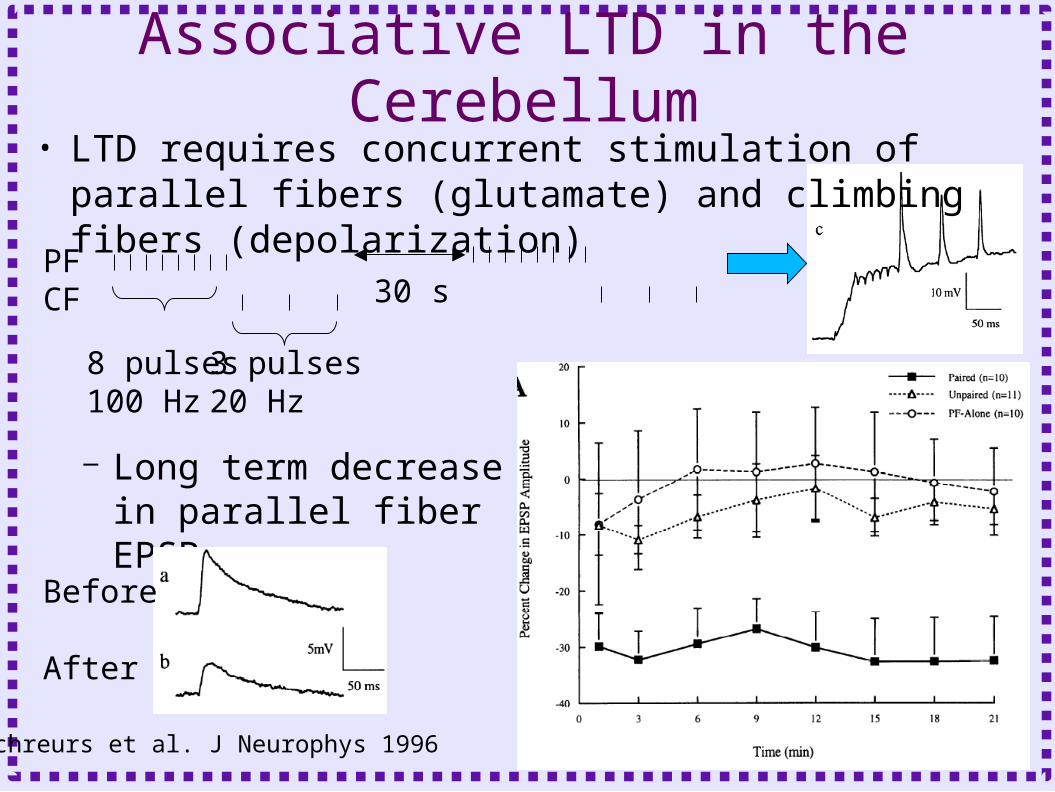
Associative LTD in the Cerebellum• LTD requires concurrent stimulation of parallel
fibers (glutamate) and climbing fibers (depolarization)
PFCF
8 pulses100 Hz
3 pulses20 Hz
30 s
– Long term decrease in parallel fiber EPSP
Schreurs et al. J Neurophys 1996
Before
After
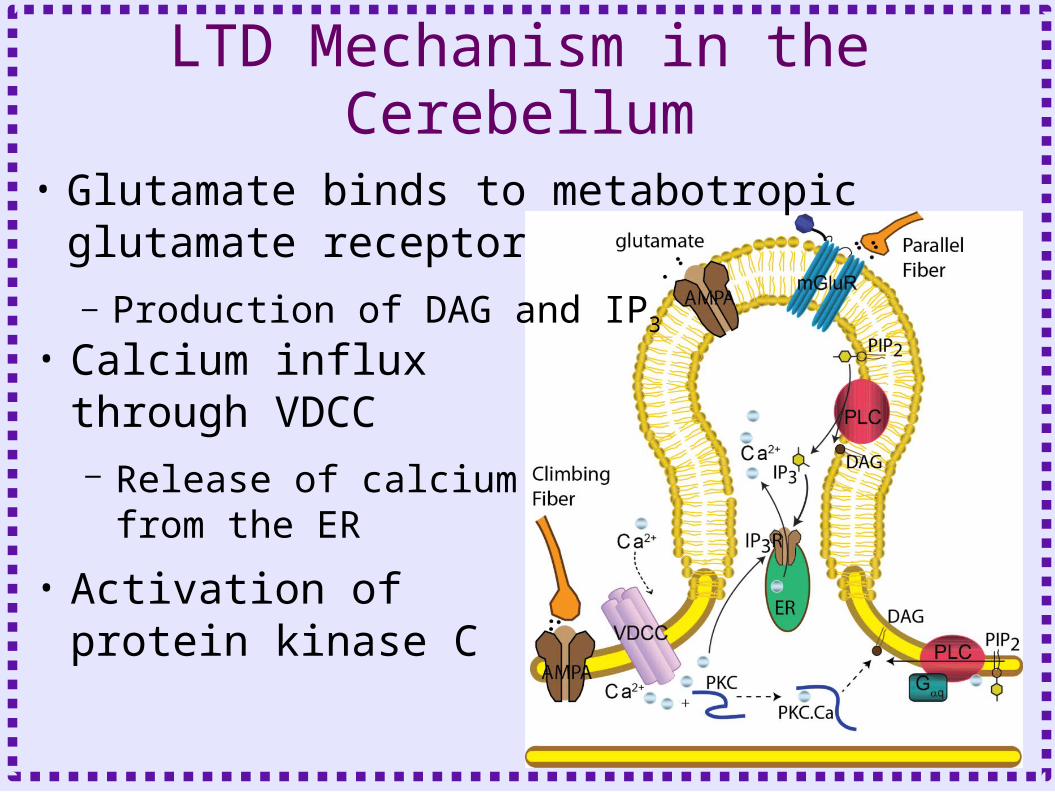
LTD Mechanism in the Cerebellum
• Calcium influx through VDCC– Release of calcium from
the ER
• Activation of protein kinase C
• Glutamate binds to metabotropic glutamate receptor– Production of DAG and IP3

General rules• One differential equation for each molecule
in the system of biochemical reactions• Two terms on the right hand side of a
differential equation for each set of arrows– Both terms must be in two different differential
equations
• Conservation equations can replace some differential equations
• Michaelis Menten approximation reduces number of equations



XPPAUT example
• General purpose ODE solver commonly used in neuroscience
• http://www.math.pitt.edu/~bard/xpp/xpp.html
• Xppaut mglu-ip3.ode– Evaluate role of aG decay– Evaluate role of IP3 decay

Three Types of Objects in Chemesis/Kinetikit
• Pools of molecules– Keep track of concentration
• Uni- and Bi-molecular Reactions– Transformation of one or more molecules
into equal number of another molecule
• Enzyme reactions– One enzyme molecule can transform
multiple copies of substrate into equal number of product

Compartment-Like Objects• Keep track of molecule quantities and
concentrations•Similar to compartment calculating voltage
• Requires geometry/morphology values•length
•Radius
• Takes messages from reaction objects, enzyme objects, calcium release objects and current influx
•Integrates all increases and decreases
•Divides quantity by volume to calculate concentration

Compartment-Like Objects• Rxnpool (chemesis)
•Morphology fields:
• Len (length)
• Radius
• Surface area and volume (vol) calculated from shape (cylinder or shell)
• area of outer surface
• area of inner surface (can be zero)
• area of side surface
•UNITS fields!!!
• units: 1 for SI, 1e-3 for mmole, etc
• Dunits (length): 1 for SI, 1e-3 for mmeter, etc
• Iunits: 1e-12 to convert from nA, msec to A, sec

Compartment-Like Objects– rxnpool (Chemesis)
• dC/dt = A - B C
• A = change in quantity independent of present quantity
• B = rate of change
• Receives messages with quantities A and/or B from other objects (enzymes, reactions, also calcium influx)
• RXN0 (A), RXN1 (B), RXN2 (A and B)
•For concentration inputs
• RXN0MOLES (A), RXN2MOLES (A and B)
•For quantity inputs
• CURRENT (valence current)

Compartment-Like Objects• Keep track of molecule quantities and
concentrations– conservepool (Chemesis)
• C = Ctot - Ci
• Quantity is remainder after all other forms of molecule accounted for
• Also has volume and units fields
– pool (Kinetikit)
• dC/dt = A - B C
• Or C = Ctot - Ci(if flag is set to conserve)
• Can also implement stochastic reactions

Concentration Pools• chemesis
• genesis #1 > showobject rxnpool
• genesis #2 > showobject conservepool
• genesis #3 > showobject pool

Enzyme and Reaction objects
• Calculate changes due to reactions– mmenz (Chemesis)
• Use if MM assumptions are met
• Fields: Km and Vmax
• Inputs: enzyme, substrate concentration
• Calculates Vmax times [Enzyme] times
[substrate] divided by ([substrate] + Km)
• Send messages RXN0 or RXN0moles to rxnpool
• Empirical feedback modification of enzyme activity can be added

Enzyme and Reaction objects• Calculate changes due to enzyme
reactions• Stores ES substrate concentration
• Has fields for volume
• Fields: Kcat, Kf, Kb
– Enzyme (Chemesis)• Fields: units, surface areas (as rxnpool)
• Inputs: enzyme, substrate quantity
• Calculates change in product, enzyme, substrate
– Enz (kinetikit)• Inputs: enzyme, substrate quantity
• Can implement stochastic reactions

Enzyme and Reaction objects
• Calculate changes due to reactions– reaction (Chemesis) or reac (kinetikit)
• Fields: kf, kb
• Inputs (messages): substrates and products
• Calculates:– forward rate constant times substrate molecules– backward rate constant times product molecules
• send messages RXN0 - RXN2 to rxnpool

Enzyme and Reaction objects
• Genesis #4> showobject mmenz
• Genesis #5> showobject enzyme– Compartment dimensions allows
membrane bound enzyme to have different volume than substrate and products
• Genesis #5> showobject enz
• Genesis #6> showobject reaction
• Genesis #7> showobject reac

Creating Chemesis Simulation
• Create rxnpool pool1
• Create conservepool pool2
• Setfield pool1 Cinit initvalue ...
• Addmsg pool1 pool2 CONC Conc

Creating Chemesis Simulation
• Create reaction rxn1• Setfield rxn1 kf kfvalue kb kbvalue• Addmsg pool1 rxn1 SUBSTRATE Conc• Addmsg pool2 rxn1 SUBSTRATE Conc• Addmsg pool3 rxn1 PRODUCT Conc• Addmsg rxn1 pool1 RXN2 kbprod
kfsubs– To substrate – kbprod is first
• Addmsg rxn1 pool3 RXN2 kfsubs kbprod– To product – kfsubs is first

Chemesis Example
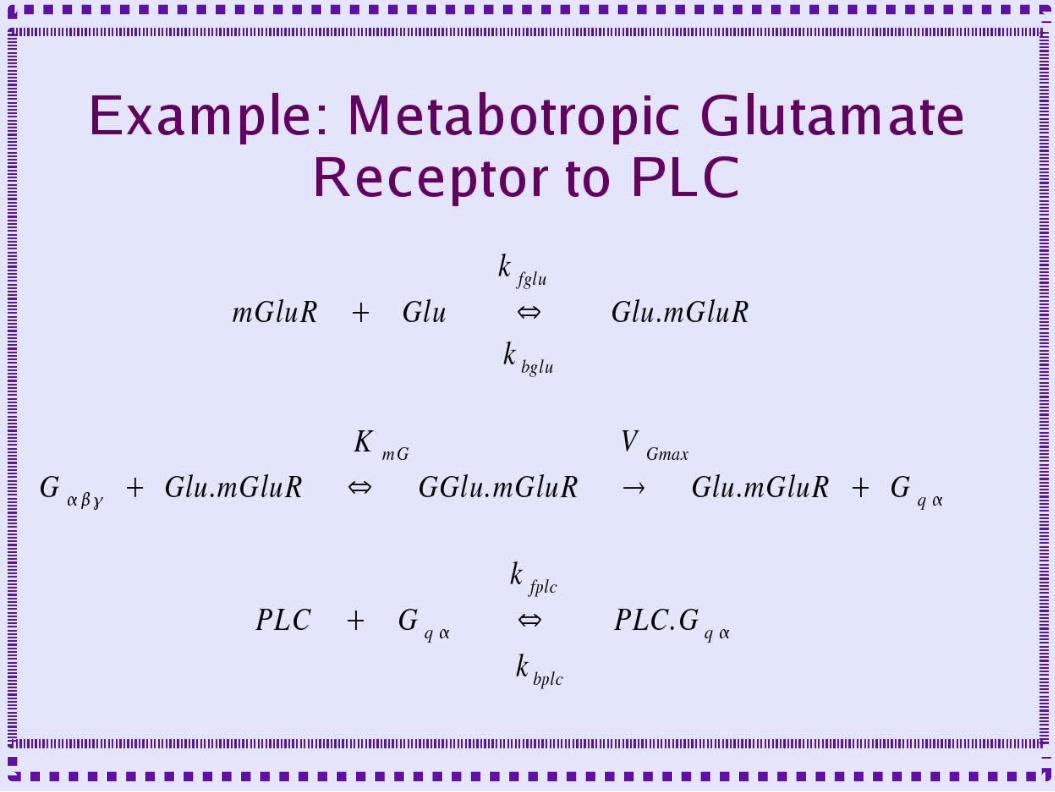
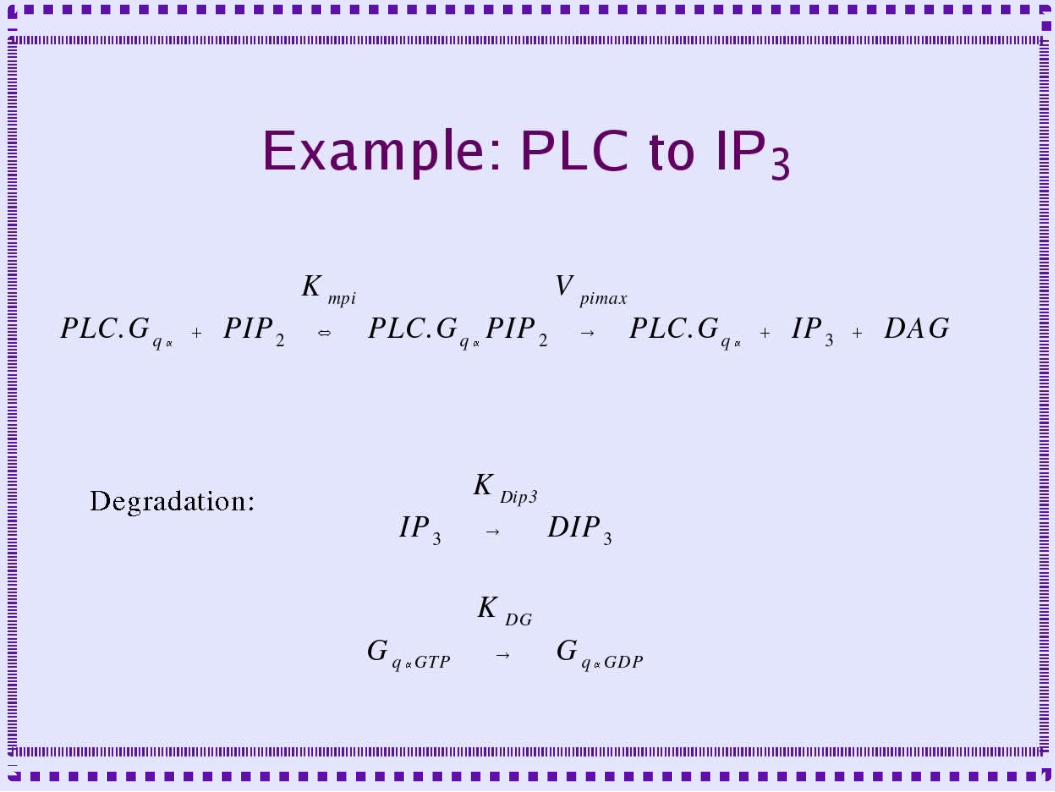
• Metabotropic receptor to PLC to IP3
– mglu-ip3-chemesis.g for complete example– Setclock – to determine the time step– Include param.g – set of parameters used– Create several instances of rxnpool,
conservepool, reaction, enzyme and mmenz
– Include graphs.g to plot some output– Step - to run simulation





























