Embed Size (px)
Citation preview

THE JOURNAL OF BIOLODICAL CHEMIBTRY Vol. 249, No. 1, Issue of Jmuary IO, pp. 72-80, 1974
Printed in U.S.A.
Ketone Bodies as Precursors of Sterols and
Fatty Acids in the Developing Rat*
(Received for publication, June 4, 1973)
JOHN EDMOND
From the Department of Biological Chemistry, UCLA School of Medicine. Los Angeles, California 90024
SUMMARY
Four substances, &hydroxy[3-i4C]butyrate, [3-W]aceto- acetate, [2-“C]acetate and [2-W]mevalonate, were compared by subcutaneous injection in 9- to 12-day-old rats as pre- cursors of lipids in the central nervous system and in other organs. Subcutaneously injected mevalonate is a poor substrate for sterol synthesis in the central nervous system as compared to the utilization by the liver and kidneys, the latter taking far the largest share of available mevalonate for sterol biosynthesis. The data obtained demonstrated that 3-hydroxybutyrate was the preferred substrate for sterol and fatty acid biosynthesis in the three organs of ectodermal origin: the brain, spinal cord, and the skin. It is postulated that ketone bodies play a major role in the central nervous system during myelination not only as sources of energy, but also of carbon for lipid biosynthesis.
Although the early experiments of Waelsch et al. (1, 2) indi- cated that the unsaponifiable lipids (mostly sterols) in the brain of the developing rat were probably synthesized in that organ, subsequent work by others demonstrated that not only labeled plasma cholesterol (3-9) but also long chain fatty acids, originat- ing either in the diet or given intravenously, were transferred to the brain intact and were deposited at specific sites and at all ages (10-13).
Comparisons of acetate and mevalonate as precursors of sterols in the central nervous system showed, surprisingly, a poor utilization of mevalonate for sterol synthesis by this tissue (14-23). Further, homogenates of the brain of developing animals-supposedly actively synthesizing sterols during the myelination process-could not carry the sequence of sterol synthesis beyond squalene, whether acetate or mevalonate was the precursor (15, 24-26) ,r although the brain homogenates were made by the same methods as used for the preparation of liver homogenates (27) which are known to synthesize sterols from either acetate or mevalonate.
To resolve some of these paradoxes, in the experiments pre-
* This research was supported in part by United States Pub- lic Health Service Research Grants HL-12745 and HD-05061, U. C. L. A. Mental Retardation Center Grant HD-04612, and the Department of Mental Hygiene, State of California.
1 G. Popjak, J. Edmond, and T. Altshuler, in preparation.
sented here acetate, acetoacetate, P-hydroxybutyrate, and mevalonate were compared as precursors of lipids in the brain, spinal cord, and other organs of rats between the 9th and 12th day after birth. Tests of ketone bodies seemed particularly worthwhile as Sir Hans Krebs and his colleagues have demon- strated recenbly that the level of ketone bodies in the blood of developing rats was 6 to 10 times higher than the level in adult animals (28, 29) and that 48 to 76% of the energy requirement of the brain of the young could be accounted for by the uptake of ketone bodies from the blood (29). Ketone bodies were in fact used preferentially for sterol and fatty acid synthesis by tissues of ectodermal origin: the brain, spinal cord, and the skin.
MATERIALS AKD METHODS
Sodium uL-/3-hydroxy[3-14C]butyrate (specific activity 12.2 Ci per mole), ethyl [3-%]acetoacetate (specific activity 13.1 Ci per mole), sodium [2-14C]acetate (specific activity 59 Ci per mole), and RS-[2-14C]mevalonic acid lactone (specific activity 10.3 Ci per mole) were obtained from the Amersham-Searle Corporation. Hexamethyldisilazane, trimethylchlorosilane, Ad- sorbosil-1-ADN (25 %), cholesterol (about 98 $& pure), desmos- terol (90% pure), a mixture of lanosterol (65%) and dihydro- lanosterol (35y0), and the methyl esters of myristate, palmitate, stearate, and arachidate (all 99% pure) were obtained from Applied Science Laboratories, Inc. Squalene, grade I (98% pure), was obtained from the Sigma Chemical Company. A standard mixture of fatty acid methyl esters (No. 13) was ob- tained from the Hormel Institute, Austin, Minnesota. Digi- tonin, Baker grade, was obtained from the J. T. Baker Chemical Company. Silica Gel H (E. Merck) for thin layer chromatog- raphy was obtained from Brinkmann Instrument Company. All chemicals and solvents were analytical reagents or their equivalents. Gas chromatography liquid phases, 10% Dexsil 300 GC and 15% FFAP on high performance Chromosorb W (AWIDMCS, 80 to 100 mesh), were obtained from Varian Aerograph.
Ethyl [3J4C]acetoacetate was converted to the sodium salt by hydrolysis at 50” for 4 hours with 2 eq of NaOH. The prepara- tion was neutralized by the addition of 1 eq of HCl and diluted with a 0.9% NaCl solution. The specific activity of the sodium [2-14C]acetate was adjusted to 6.25 Ci per mole by dissolving 0.25 mCi of the freeze-dried solid (specific activity 59 Ci per mole) in sterile 0.9% NaCl solution containing 35.76 pmoles of sodium acetate. RS-[2-14C]Mevalonic acid lactone was con- verted to the potassium salt as described (30).
Pregnant albino Sprague-Dawley rats were obtained from the
72
by guest on January 19, 2020http://w
ww
.jbc.org/D
ownloaded from

73
Simonsen Laboratories, Inc., Gilroy, California, 5 days before parturition. On the 1st day after birth the litter size was re- duced to eight and on the 9th day the young were injected sub- cutaneously between the scapulae. In one experiment paired animals from each of three litters were administered 20 ~1 (12.5 PCi) of either sodium nL-/%hydroxy[3J4C]butyrate, sodium [3J4C]acetoacetate, sodium [2-14C]acetate, or potassium RS- [2-14C]mevalonate. Littermates were sacrificed at 4, 24, and 72 hours after injection. In a second experiment the members of a single litter were injected subcutaneously with 5 PCi of sodium nL-@hydroxy[3-14C]butyrate and killed at varying times after the injection. One member of this litter was given a second dose of 5 rc.Ci of n&hydroxy[3-14C]butyrate at 48 hours and killed 96 hours after the first injection. The animals were anesthetized with ether and a sample of blood was taken by cardiac puncture witha heparinized syringe. The liver, kidneys, and lungs, the brain including the cerebellum, the spinal cord from the medulla and including the cauda equina, and a section of skin from the hindquarters were dissected. Each organ or tissue specimen was weighed and transferred to a 50.ml glass centrifuge tube for saponification.
Samples were saponified at 70” with 3.5 N NaOH in 50% aqueous ethanol. Unsaponifiable material was extracted four times with light petroleum (b.p. 30-60”). The combined pe- troleum extracts were concentrated to 10 ml with a stream of N2. The aqueous residue, after evaporation of the ethanol, was acidified to pH 1 and the fatty acids were extracted with 4 equal volumes of diethyl ether. Portions of the extracts were assayed for radioactivity in a Packard Tri-Carb liquid scintilla- tion spectrometer, model 3302, with the standard 2,5-diphenyl- oxazole and 1,4-bis[2-(4-methyl-5-phenyloxazolyl)]benzene mix- ture in toluene. Counting efficiency was determined with [14C]toluene standards. All samples were counted in triplicate.
Determination of Specific Activity of So-Hydroxy Sterols-The digitonides were prepared from portions of the extract of the total unsaponifiable material as described (27). The washed precipitates of digitonides were transferred with three l-ml portions of acetone to weighed Whatman glass fiber disks (grade GF/A, 2.4 cm in diameter) that had been washed with methanol and acetone, air-dried, and then stored over silica gel and par- affin wax under vacuum. Pyrex microanalysis filter holders with fritted glass base, supplied by the Millipore Corporation, were used to support the fiber disks during vacuum filtration to collect and partially dry the digitonide precipitate. Each disk was weighed on a microbalance before and after loading with precipitate which had been dried under vacuum overnight. The disks were transferred to glass counting vials containing 6 ml of anhydrous methanol; the digitonides were dissolved by heating and shaking at 50” for 20 min. The solutions were cooled and 4 ml of scintillator solution (3 g of 2,5-bis(2-tert- butylbenzoxazolyl)thiophene per 400 ml of toluene) were added to each vial and the radioactivity assayed in the Packard Tri- Carb liquid scintillation spectrometer. Vials of blanks and [14C]toluene standard samples, each containing a glass fiber disk and the equivalent methanol-scintillator mixture, were included to determine counting efficiency. The specific activity of the digitonin-precipitable sterols was calculated as disintergrations per min per mg of sterol with molecular weight 386. The efficiency of sterol recovery by this method was consistently 90 f 3% on amounts of cholesterol in the range of 0.5 to 6 mg. Eighty to ninety per cent of the radioactivity in the unsaponi- fiable fractions was found in the digitonides. Duplicate specific activity determinations were within 3% of each other.
Gas-Liquid Chromatography-The distribution of the radio- activity in the various sterols and fatty acids was determined by the gas-liquid radiochromatography assembly of PopjLk et al. (31) on IO-foot stainless steel columns (3& inch, internal diam- eter), packed with (a) 10% Dexsil 300 GC and used for the analysis of trimethylsilylated sterols at an oven temperature of 325” and (b) 15% FFAP for the analysis of fatty acid methyl esters at an oven temperature set isothermally at 230” for 16 min after injection and then programmed at 4” per min to 275”. The unsaponifiable fraction was examined for sterols after the alcohols had been converted to the trimethylsilyl ethers (32). Fatty acids were methylated with diazomethane (33).
Thin Layer Chromatography-The silver nitrate-impregnated support Adsorbosil-l-ADN 25% was spread 300 pm thick on glass plates (5 X 20 cm). Coated plates were air-dried for 3 hours, activated at 110” for 1 hour, and stored in a desiccator, all in the dark, then used within 36 hours of preparation. The plates were developed twice with 1 y0 acetone in chloroform to a solvent front of 14 cm from the origin. Plates were dried in air before the second development. Regions of radioactivity were located on the plates with a Packard radiochromatograph scanner, model 7201. Radioactivity was eluted from the scraped off gel with acetone in fritted funnels. After evapora- tion of the acetone with a stream of Ns, 3 ml of a 1% aqueous NaCl solution were added and the mixture was extracted three times with diethyl ether. The ether extracts were combined (12 ml), taken to dryness in a counting vial, and the radioac- tivity was assayed in a scintillation solution of 2,5-bis(2-tert- butylbenzoxazolyl)thiophene (4 g per liter) in 50% methanol in toluene. Quenching of 14C counts was checked by the addition of known amounts of [14C]toluene to each sample after the first counts were taken. The percentage of [14C]squalene in the un- saponifiable fractions was determined, after addition of 200 I.rg of unlabeled squalene, by the separation of squalene from sterols on Silica Gel H plates, 250 pm thick, developed once with ben- zene. The gel from the sterol region (RF 0.08) and the squalene region (RF 0.76), after detection by scanning for radioactivity, was scraped from the plates into fritted funnels and was then eluted with ethyl acetate into counting vials. The ethyl acetate was evaporated with a stream of nitrogen and the radioactivity was measured by scintillation counting.
RESULTS
The four substrates were compared at three time intervals in members of the same litter, two animals being used for each substrate at each time interval after injections. Although the three litters used were born within 8 hours of each other, the rats in the litter chosen for the 72.hour experiment were much smaller than members of the other two litters at 9 days after birth (Table I). These smaller animals attained, nevertheless, on the 12th day near parity in their body and organ weights with the rats from the litters killed at 4 and 24 hours after injection. The tissue extracts of all animals were analyzed for incorporation of 14C from nL-/?-hydroxy[3-14C]butyrate, (3J4C]- acetoacetate, [2-14C]acetate, and RS-[2-14C]mevalonate into the unsaponifiable (neutral) and saponifiable (acidic) lipid com- ponents 4, 24, and 72 hours after the injection of the substrates. There was no truly discernible trend of change in the amount of isotope found in the tissue components with time elapsed after the injections. Nearly all the differences noted could be at- tributed to variations of metabolic activity among individuals of a litter or to variations between litters. For this reason and for the sake of simplifying presentation, the principal data,
by guest on January 19, 2020http://w
ww
.jbc.org/D
ownloaded from

74
Tm~lr I T,\BLI’. III
Body and organ weights of animals used for comparison of four substrates as lipid precursors
Rats from three litters were used 9 days after birth. Two animals in each litter were injected with 12.5 &i of either nL-P-hydroxy[3-14C]butyrate, [3-Wlacetoacetate, [2-Wlacetate, or [2-14C]mevalonate. One litter was sacrificed at 4 hours, the second at 24 hours, and the third at 72 hours after the injections (cf. “Materials and Methods”). The values given are the means, with standard deviation, derived from 24 animals.2 Samples of skin used for analysis weighed 1 to 2.5 g. Samples of blood, 0.4 to 0.9 ml, were obtained for analysis.
Specific activity of digilokrk-precipitable sterols in organs and blood of youtlg rals after itljeclion of 14C-labeled
lipid precursors
OrgaIl Wet weight
The data shown are part of the experiments presented in Tables I and II. The specific activity of the sterols was calculated from that of the digitonides and is expressed as disintegrations X lo+ per mg of sterol of molecular weight 386 (cholesterol). The values given are the means with standard deviation derived from the analysis of data for 4, 24, and 72 hours after injection2 (cf. Tables I and II).
I Substrate
Brain. ...... Spinalcord.. ..... :::::::::.:‘:I Two kidneys .................... Two lungs ...................... Liver ......................... Body weight”. .................
w
877 f 20 102 f 15 218 f 14 318 f 18 547 f 54
17.46 f 1.3 g
a This is the mean body weight of the 24 rats at the termination of the experiment 4, 24, and 72 hours after injections. The mean body weight of the three groups of eight rats used for the 4-, 24-, and 72-hour experiments was at nine days after birth 16.5 f 0.3, 16.7 f 0.7, and 12.22 f 1.16 g, respectively.
T.\HLI.: II
1%’ content of total unsaponi$able components in organs of you?kg rats after subcutaneous injection of substrates
The data were obtained from animals whose body and organ weights are shown in Table I. The experimental conditions are described under “Materials and Methods” and in Table I. The ‘4C content in each organ is expressed as disintegrations X 10-S per min per g of tissue. The values given are the means with standard deviation derived from the analysis of data for 4, 24, and 72 hours after injections2 (6. Table I).
Substrate
Organ (B-Hydroxy13~~ [3-W- 1 ,I 1 ~ “C]butyrate Acetoacetate [‘- ‘IAcetate M.%%e
disinlegratiors X 10-a per min ger g fissue
Brain. ......... 118 + 39 86 f 28 90 f 7 91 f 26
Spinal cord ....... 444 f 63 193 f 60 233 f 77 141 f 29 Skin .............. 89 f 22 76 f 18 73 f 7 276 f 61
Kidney .......... 24 f 4 24 f 9.3 52 f 6 7253 f 1672 Lung. ............ 16 f 6.4 23 f 10.4 40 f 7.7 873 f 61 Liver. ........... 8.4 f 4 11 f 7.5 21 f 4 1061 f 319
reflecting differences in the extent of utilization of the four sub- strates by the various organs at the three time intervals, are shown as the means =t standard deviation.* Since the experi- mental subjects were rapidly growing animals, not in a state of
2 The detailed data from which the values given in Tables I, II, III, VII, and VIII were calculated are available from the author or as JBC Document Number 73M-708. in the form of one microfiche or nine pages. Orders for supplementary material should specify the title, author, and reference to this paper and the JBC Document number, the form desired (microfiche or photo- copy), and the number of copies desired. Orders should be ad- dressed to The Journal of Biological Chemistry, 9650 Rockville Pike, Bethesda, Maryland 20014, and must be accompanied by remittance to the order of the Journal in the amount of $2.50 per microfiche or photocopy.
Brain. ....... Spinal cord ... Skin. ........ Kidney. ...... Lung. ........ Liver. ....... Blood. .......
disinlegrolions X lo-2 per min per mg slerol no2 wl 386
148 zt 41115 f 40112 & 7 122 A 41 292 f 88126 f 29109 f 1 134 & 38 226 f 42193 zt 60146 f 10 789 * 192
46 f 13 49 f 20 88 SC 2218,640 f 3640 24 f 10 33 f 22 56 f. 21 1,968 f 184 24 f 12 30 f 24 59 f 10 2,796 f 966 26 + 20 36 f 33 55 f 9 2,795 zk 829
equilibrium, synthesis and accumulation of body constituents as opposed to their degradation were to be expected.
Comparison of Acetate, Acetoacetate, /3-Hydroxybutyrate, and Jlevalonate for Biosynthesis of Unsaponifiable Lipids--The data of Table II show that all the organs examined utilized acetate, acetoacetate, and @hydrosybutyrate for the biosynthesis of unsaponifiable substances more efficiently than the liver; the brain, spinal cord, and skin were the most active organs, par- ticularly in respect to the utilization of the two ketone bodies. P-Hydroxybutyrate appears to he the preferred substrate by these three organs of ectodermal origin, the concentration of “C in the unsaponifiable lipids synthgsized from m-P-hydroxy- [3-‘%]butyrate being about 53, 14, and 10 t,imes greater in the spinal cord, brain, and skin, respectively, than in the liver. The isotope content of the unsaponifiable lipids derived from [P4C]acetate in these three organs was similar to that seen after the injection of [3NZ]acetoacetate. The utilization of the ketone bodies by the liver was poorer than that of acetate.
Mevalonate, in contrast to acetate and the ketone bodies, was a very poor substrate for the synthesis of unsaponifiable sub- stances in the central nervous system and skin as compared to utilization in the liver. The surprising finding, apart from the high incorporation of label from the ketone bodies into the un- saponifiable matter of the three organs of ectodermal origin, was the dramatic incorporation of “C from mevalonate into the unsaponifiable matter in the kidneys. At both 4 and 24 hours the total label in the unsaponifiable fraction of the kidneys amounted to 13y0 of the utilizable R-mevalonate. In contrast, liver and lung contained, 4 hours after injection, 5.65 and 2.18%, respectively, of the label derived from mevalonate.
That the data recorded in Table II reflect differences in the relative utilization of the various substrates by the various organs is underlined by the comparison of the specific activities of the digitonin-precipitable sterols (Table III), which accounted for 80 to 90% of the radioactivity in the total unsaponifiable matter in all the samples. The data of Table III show that the specific activities of the sterols in brain, spinal cord, and skin, at all times after the injection of P-hydroxybutyrate and aceto- acetate, were many times higher than the specific activities of
by guest on January 19, 2020http://w
ww
.jbc.org/D
ownloaded from

75
liver and blood sterols. The observations thus offer proof of not only sterol synthesis within these organs but also of the preferential utilization of ketone bodies for such synthesis. The differences between the specific activities of blood sterols and the sterols of the three organs of ectodermal origin after the injection of [2-14C]acetate were not as great as after the injection of the ketone bodies, particularly after the injection of P-hy- droxybutyrate; nevertheless, the data give evidence of sterol synthesis even from this substrate within these three organs.
The poor utilization of mevalonate by brain, spinal cord, and skin as compared to utilization by the liver and kidney suggests a poor transfer of this substrate from the blood to the ectodermal organs. The specific activity of the liver and kidney sterols were 12 and 80 times higher, respectively, than the specific activities of sterols in brain (cf. Table III).
The analysis of the total unsaponifiable matter by gas-liquid radiochromatography, after conversion of the alcohols to tri- methylsilyl ethers, supported fully the data obtained by the analysis of the digitonides and showed that, with the exception of the extracts of liver and lung, 95% or more of the 14C in these extracts was associated with sterols. The analysis indicated that the 14C in sterols in skin, liver, kidney, and lung was asso- ciated mostly with cholesterol. However, t.he brain differs from these organs at this stage in development of the young. The analysis indicated that the radioactivity in sterols was distrib- uted between desmosterol and cholesterol and that desmosterol was present in significant quantity. The unsaponifia.ble matter extracted from the spinal cord was not analyzed, as radioactivity in amounts sufficient only for the preparation of digitonides was available.
The analysis of the total unsaponifiable components in the several organs by gas-liquid radiochromatography showed that. all extracts contained labeled squalene. After the injection of mevalonate 10 y0 of the 14C content in the unsaponifiable fraction in liver was associated with squalene irrespective of the fact that the total radioactivity in this fraction decreased by 72 hours to a value approximately one-half of that found 4 hours after injection. In the experiments with substrates other than mevalonate, and in organs other than the liver, the proportion of the 14C content in the unsaponifiable fraction as squalene changed greatly with time. Thus, only 3, 2, and 0.8% of the 14C in the liver unsaponifiable matter was associated with squa- lene at 4, 24, and 72 hours, respectively, after the injection of P-hydroxybutyrate or acetoacetate. After the injection of acetate, 4.9, 4.5, and 2.4y0 of the 14C content of the unsaponi- fiable fraction in the liver was associated with squalene at 4, 24, and 72 hours, respectively.
The radioactivity in squalene, as a percentage of the 14C in the unsaponifiable components, declined with time irrespective of the precursors used. In kidneys more than one-half of the label in the squalene acquired from [2J4C]mevalonate was lost in 24 hours (5.7 to 2.2’%) while there was no significant change either in the total amount of radioactivity in the unsaponifiable frac- tion or in the specific activity of the digitonin-precipitable sterol. By 72 hours after injection of [2-%]mevalonate 1.1% of the 14C content of the unsaponifiable components was in squalene. The absolute amount of [YJsqualene in the kidneys was re- duced by 72 hours to about one-tenth of that found 4 hours after injection. A very active sterol synthesis in the kidneys was indicated also by the finding that as much as lSyo of the 1% in renal sterols, 4 hours after the injection of mevalonate, co-chromatographed with lanosterol, whereas 20 hours later all the sterol radioactivity was in cholesterol. The 14C content of
the unsaponifiable fraction as squalene in kidney 4, 24, and 72 hours after the injection of acetate was 3.6, 2.7, and 1.4%, respectively. Extensive labeling of squalene in the lung 4 hours after the injection of all theprecursors tested, except mevalonate, is probably good evidence of sterol synthesis within the lung. Thus, as much as 15.6% of the radioactivity found in the non- saponifiable fraction in the lung 4 hours after the injection of [2-14C]acetate was found in squalene. [14C]Squalene amounted to 7.8 and 0.6% of the 14C content in the total unsaponifiable fraction in lung 24 and 72 hours, respectively, after injection of acetate. With either of the ketone bodies as precursor, 11% of the 14C content of the unsaponifiable fraction in lung was in squalene 4 hours after the injections. In contrast the i4C con- tent of the unsaponifiable components in lung as shualene 4, 24, and 72 hours after injection of [2-14C]mevalonate was only 1.06, 0.5, and 0.07 7& respectively.
In the brain less than 1 7. of the label found in the unsaponi- fiables was associated with squalene irrespective of the pre- cursors administered. However, in contrast to the other organs in which the mass and radioactivity in sterols were found mainly as cholesterol, the brain contained significant quantities of mass and radioactivity as desmosterol. Gas-liquid chromatog- raphy of the trimethylsilyl ethers of brain sterols showed that desmosterol accounted for 22 to 24 y0 of the mass of sterols in all animals. The response of the gas-liquid chromatographic de- tector (thermal conductivity cell) was calibrated with a known mixture of cholesterol and desmosterol. In contrast, analysis of the distribution of 14C in cerebral sterols by gas-liquid radio- chromatography and by thin layer chromatography on silver nitrate-impregnated plates showed that about 46 v/0 of the radio- activity was associated with desmosterol 4 and 24 hours after the injection of any of the precursors. Thus the specific activity of desmosterol at these times was 4 times higher than that of cholesterol. However, by 72 hours the dist)ribution of label in cerebral sterols became identical with the distribution of mass.
Distribution oj Radioactivity in Saponijiable Fractions of Organs-All the organs examined utilized P-hydroxybutyrate, acetoacetate, and acetate for the biosynthesis of fatty acids more efficiently than the liver (Table IV). As in the case of sterol synthesis (cf. Table II) the brain, spinal cord, and skin were the most active organs. The pattern of labeling of fatty acids was the same from acetate as from /3-hydroxybutyrate. The utilization of [2-14C]acetate by the liver in these young ani- mals for sterol and fatty acid synthesis was not much greater than the utilization of ketone bodies.
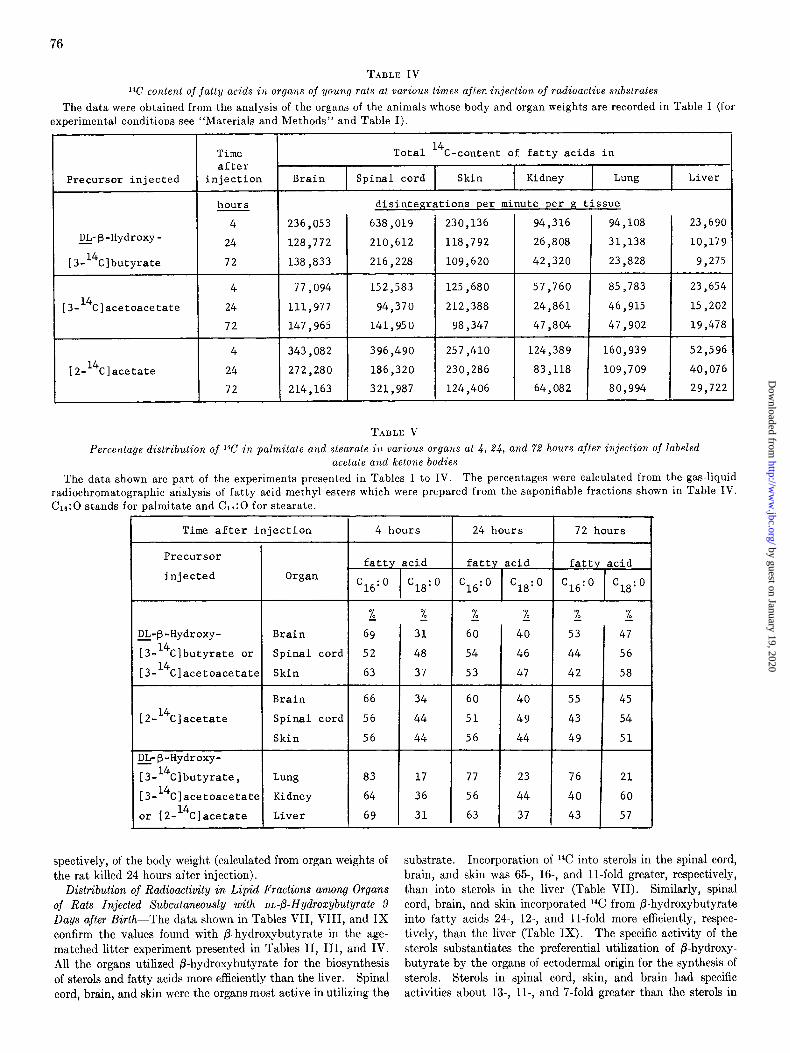
Gas-liquid radiochromatographic analysis showed that, in all the organs, with the possible exception of the skin, all the radio- activity of the fatty acids was associated with palmitate and stearate (Table V). Among the skin fatty acids 6 to 77, of the 14C co-chromatographed with myristate and the remainder with palmitate and stearate. The same trend was discernible in all the organs with time elapsed after injection of the various pre- cursors: palmitate was generally more heavily labeled than stearate, but there was a trend for stearate to acquire in all organs a higher proportion of the label by 72 hours than at 4 hours.
Utilization of DL-P-Hydroxy[S-14C]butyrafe for Synthesis of Sterols and Fatty Acids by Rats of Single Litter-The body and organ weights are shown in Table VI. As much of the skin as possible was removed from each animal for analysis. Skin constituted 22 to 25% of the body weight of each animal. This is considerable compared to spinal cord, brain, kidneys, and liver which account for as little as 0.5, 4.5, 1.3, and 3.8%, re-
by guest on January 19, 2020http://w
ww
.jbc.org/D
ownloaded from
76
TABLE IV
1%’ content of fatty acids in organs of young rats at various times after. injection of radioactive substrates
The data were obtained from the analysis of the organs of the animals whose body and organ weights are recorded in Table I (for experimental conditions see “Materials and Methods” and Table I).
Brain
Total l4 C-content of fatty acids in
Spinal cord Skin Kidney Lung Liver
Time after
injection
hours
4
24
72
4
24
72
4
24
72
Precursor injected
DL- p-Hydroxy - -
[3-14C]butyrate
Jte per g tissue I 236,053
128,772
138,833
77,094
111,977
147,965
343,082
272,280
214,163
disintegrations per m
638,019 230,136
210,612 118,792
216,228 109,620
152,583 125,680
94,370 212,388
141,950 98,347
396,490 257,410
186,320 230,286
321,987 124,406
94,316 94,108 23,690
26,808 31,138 10,179
42,320 23,828 9,275
[3-14C]acetoacetate
57,760
24,861
47,804
[2-14C]acetate
124,389
83,118
64,082
TABLE V
Percentage distribution of 1%’ in palmitate a,ld stearate i/L various orgam at 4, 24, and 72 hours after injection of labeled acetate aud ketone bodies
The data shown are part of the experiments presented in Tables I to IV. The percentages were calculated from the gas-liquid
radiochromatographic analysis of fatty acid methyl esters which were prepared from the saponifiable fractions shown in Table IV. C16:0 stands for palmitate and G8:O for stearate.
c Time after injection I 4 hours 24 hours 72 hours
Precursor
injected fatty acid
c16:o ( c18:o
3 3 2 2 L L
31 31 60 60 40 40
48 48 54 54 46 46
37 37 53 53 47 47
34 34 60 60 40 40
44 44 51 51 49 49
44 44 56 56 44 44
Brain
Spinal cord
Skin
“/ 1
53 47
44 56
42 58
Brain
Spinal cord
Skin
55
43
49
45
54
51
Lung 83 17 17 77 77 23 23 76 21
Kidney 64 36 36 56 56 44 44 40 60
Liver 69 31 31 63 63 37 37 43 57
DL-@-Hydroxy- -
[3-14C]butyrate or
[3-14C]acetoacetats
[ 2-14C]acetate
DL-@-Hydroxy- -
[3-14C]butyrate,
[3 l4 - Clacetoacetats
or [2-lOCIacetate
spectively, of the body weight (calculated from organ weights of
the rat killed 24 hours after injection).
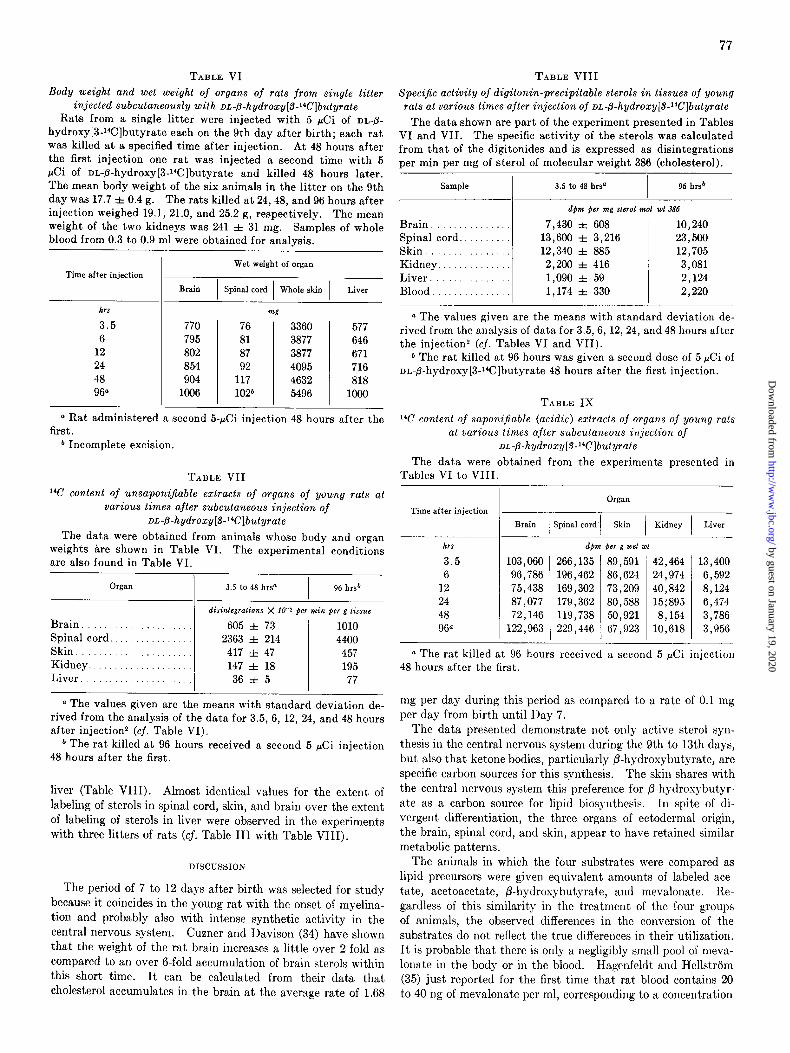
Distribution of Radioactivity in Lipid Fractions among Organs of Rats Injected Subcutaneously with DA-fl-Hydroxybutyrate 9 Days after Birth-The data shown in Tables VII, VIII, and IX confirm the values found with /3-hydroxybutyrate in the age- matched litter experiment presented in Tables II, III, and IV. All the organs utilized P-hydroxybutyrate for the biosynthesis of sterols and fatty acids more efficiently than the liver. Spinal cord, brain, and skin were the organs most active in utilizing the
substrate. Incorporation of 14C into sterols in the spinal cord, brain, and skin was 65-, 16., and ll-fold greater, respectively, than into sterols in the liver (Table VII). Similarly, spinal
cord, brain, and skin incorporated 1% from fi-hydroxybutyrate into fatty acids 24., 12-, and 11-fold more efficiently, respec- tively, than the liver (Table IX). The specific activity of the sterols substantiates the preferential utilization of /3-hydroxy- butyrate by the organs of ectodermal origin for the synthesis of sterols. Sterols in spinal cord, skin, and brain had specific activities about, 13., ll-, and 7-fold greater than the sterols in
by guest on January 19, 2020http://w
ww
.jbc.org/D
ownloaded from
77
TABLE VI TABLE VIII
Body weight and wet weight of organs of rats from single litter injected subcutaneously with DL-fl-hydroxy[3-14C]butyrate
Rats from a single litter were injected with 5 &i of DL-fi-
hydroxy[3-Wlbutyrate each on the 9th day after birth; each rat was killed at a specified time after injection. At 48 hours after the first injection one rat was injected a second time with 5 &i of nn-8-hydroxy[3-Wlbutyrate and killed 48 hours later.
Specific activity of digitonin-precipitable sterols in tissues of young rats at various times ajter injection of DL-&hydroxy[SW?]butyrate
The data shown are part of the experiment presented in Tables VI and VII. The specific activity of the sterols was calculated from that of the digitonides and is expressed as disintegrations per min per mg of sterol of molecular weight 386 (cholesterol).
3.5 to 48 hrs' I
96 hd The mean body weight of the six animals in the litter on the 9th Sample
day was 17.7 f 0.4 g. The rats killed at 24,48, and 96 hours after injection weighed 19.1, 21.0, and 25.2 g, respectively. The mean weight of the two kidneys was 241 f 31 mg. Samples of whole Brain
blood from 0.3 to 0.9 ml were obtained for analysis. Spinal cord.. Skin
Time after injection
hrs
3.5 6
12 24 48 96a
Brain
Wet weight of organ
Spinal cord Whole skin Liver
Kidney. ............ Liver. ............. Blood. .............
770 795 802 854 904
1006
.I
d@n per mg derol m
7,430 f 608 13,600 f 3,216 12,340 f 885
2,200 f 416 1,090 f 59 1,174 f 330
co1
--
W 386
10,240 23,500 12,705
3,081 2,124 2,220
ms 76 3360
Q The values given are the means with standard deviation de- 577
81 3877 646 rived from the analysis of data for 3.5,6, 12, 24, and 48 hours after
87 3877 671 the injection2 (cf. Tables VI and VII).
b 92 4095 716
The rat killed at 96 hours was given a second dose of 5 &i of 48 hours after the first 117 4632 818 nn-&hydroxy[3-“Clbutyrate injection.
102” 5496 1000 TABLE IX
a Rat administered a second 5-&i injection 48 hours after the first.
b Incomplete excision.
TABLE VII
14C content of unsaponijiable extracts of organs of young rats at various times after subcutaneous injection of
or,-&hydroxy[S-“Clbutyrate
The data were obtained from animals whose bodv and orean
14C content of saponijable (acidic) extracts of organs oj young rats at various times after subcutaneous injection of
oL-p-hydroxy[3-14C]butyrate
The data were obtained from the experiments presented in Tables VI to VIII.
Organ
Time after injection
Brain 1 Spinal card.1 Skin ( Kidney 1 Liver
weights are shown in Table VI. The experimental conditions are also found in Table VI.
Organ 3.5 to 48 hrsa 96 hrsb
disintegrahns X IO-= per min per g tirme
Brain. 605 f 73
I
1010 Spinal cord.. . 2363 f 214 4400
hrs
3.5 6
12 24 48 96a
103,060 96,786 75,438 87,077 72,146
122,963
266,135 89,591 42,464 13,400 196,462 86,624 24,974 6,592 169,302 73,209 40,842 8,124 179,362 80,588 15:895 6,474 119,738 50,921 8,154 3,786 229,446 67,923 10,618 3,956
- - Skin. 417 f 47 457 Kidney. . 147 f 18 195 Liver. 36 + 5 77
a The values given are the means with standard deviation de- rived from the analysis of the data for 3.5, 6, 12, 24, and 48 hours after injection2 (cf. Table VI).
b The rat killed at 96 hours received a second 5 FCi injection 48 hours after the first.
liver (Table VIII). Almost identical values for the extent of labeling of sterols in spinal cord, skin, and brain over the extent of labeling of sterols in liver were observed in the experiments with three litters of rats (cf. Table III with Table VIII).
DISCUSSION
The period of 7 to 12 days after birth was selected for study because it coincides in the young rat with the onset of myelina- tion and probably also with intense synthetic activity in the central nervous system. Cuzner and Davison (34) have shown that the weight of the rat brain increases a little over 2-fold as compared to an over g-fold accumulation of brain sterols within this short time. It can be calculated from their data that cholesterol accumulates in the brain at the average rate of 1.68
u The rat killed at 96 hours received a second 5 &i injection 48 hours after the first.
mg per day during this period as compared to a rate of 0.1 mg per day from birth until Day 7.
The data presented demonstrate not only active sterol syn- thesis in the central nervous system during the 9th to 13th days, but also that ketone bodies, particularly P-hydroxybutyrate, are specific carbon sources for this synthesis. The skin shares with the central nervous system this preference for P-hydroxybutyr- ate as a carbon source for lipid biosynthesis. In spite of di- vergent differentiation, the three organs of ectodermal origin, the brain, spinal cord, and skin, appear to have retained similar metabolic patterns.
The animals in which the four substrates were compared as lipid precursors were given equivalent amounts of labeled ace- tate, acetoacetate, P-hydroxybutyrate, and meva1onat.e. Re- gardless of this similarity in the treatment of the four groups of animals, the observed differences in the conversion of the substrates do not reflect the true differences in their utilization. It is probable that there is only a negligibly small pool of meva- lonate in the body or in the blood. Hagenfeldt and Hellstrom (35) just reported for the first time that rat blood contains 20 to 40 ng of mevalonate per ml, corresponding to a concentration
by guest on January 19, 2020http://w
ww
.jbc.org/D
ownloaded from

78
of 0.15 to 0.3 pM. In contrast, the concentration of ketone bodies in whole blood of 6- to IO-day-old rats has been reported by Page et al. (28) to be about 1.1 mM (0.hydroxybutyrate, 0.67 f 0.25 mM; acetoacetate, 0.43 f 0.09 mM), which accord- ing to Hawkins et al. (29) rises to 1.55 and 2.2 mM in 16. and 18-day-old rats. The blood volume of a 17-g rat (the approxi- mate weight of the animals used in the esperiments) is about 1 ml (36). Thus, the amount of circulating mevalonate is only 0.15 to 0.3 nmole as compared to 1 to 2 pmoles of ketone bodies. Assuming that the rates of absorption of mevalonate, acetate, acetoacetate, and fl-hydroxybutyrate from subcutaneous de- posits were similar, one is led to the conclusion that the injected R-[14C]mevalonate (0.61 pmole) was barely “diluted” with unlabeled circulating substance whereas the injected [14C]aceto- acetate and P-hydroxybutyrate were “diluted” probably several- fold by the circulating endogenous ketone bodies. It is thus probable that the specific radioactivity of the circulating meva- lonate was much higher than that of the ketone bodies after the injections of the respective labeled substances and this empha- sizes further the specific role of ketone bodies as carbon source for lipid biosynthesis in the central nervous system and skin.
The relatively poor utilization of intraperitoneally injected mevalonate for sterol biosynthesis in the brain has been noted before (22, 23, 25) and was attributed to its absorption into the portal venous system and subsequent avid utilization by the liver, leaving very little substrate for other organs. Indeed, Ramsey et al. (25) demonstrated a superior conversion of meva- lonate into brain sterols after intracerebral injection. In the experiments reported here all the injections were given subcu- taneously and hence the first organ to be exposed to the highest concentrations of the 14C-labeled precursors was expected to be the.lung (excepting the subcutaneous tissues in the vicinity of the injections) with a uniform distribution to all the other organs thereafter through the arterial blood. The very large differences observed among the organs in respect to the utiliza- tion of mevalonate for sterol biosynthesis even after subcutaneous injection could be accounted for either by differences in the transfer of mevalonate from blood to the various organs or to differences in the enzymic complements of the various organs, or to a combination of these two. It is reported in an adjoining communication (37) that mevalonate can provide indirectly carbon atoms for the synthesis of n-fatty acids, in all probability through a shunt of sterol intermediates starting with the hydroly- sis of 3,3-dimethylallyl pyrophosphate and the recycling of 5 mevalonate carbon atoms to 3-hydroxy-3-methylglutaryl-CoA and the cleavage of the latter to free acetoacetate and acetyl-CoA (37). Thus, the possibility exists that the labeling of cerebral sterols from mevalonate did not result entirely from the direct ut’ilization of mevalonate, but at least in part from the utiliza- tion of labeled acetoacetate generated through the postulated “[runs-methylglutaconate shunt” (37). It can be calculated that label from fl-hydroxybutyrate or acetoacetate is incor- porated into fatty acids and sterols in a ratio 1.32 : 1 (based on values for disintergrations per min per g of brain 4 hours after injections). Thus, it can be predicted using this ratio and the data presented in the adjoining communication (37) for the extent of incorporation of 14C from [2-14C]mevalonate into fatty acids by brain (disintergrations per min per g) that about 30% of the 14C found in brain sterols could be attributed to indirect labeling. Studies are presently underway to investigate this prediction.
pccted. The results corroborate recent reports (36, 38) that the uptake of mcvalonate by the kidney exceeded that by the liver in adult mice, rats, and rabbits. The extraordinarily high utilization of mevalonate for sterol synthesis in the kidneys, amounting to 13% of the total utilizable R-mevalonate by the pair of organs, constituting no more than about 2.5% of the total body weight, is the more surprisiilg as the utilization of acetate for sterol synthesis in these organs is unremarkable. It is known, however, that the S-mevalonate, unutilizable by animals on account of the stereospecificity of mevalonate kinase, is escreted in the urine (39, 40). Overloading of an animal with RS-mevalonate (4 pmoles per g) results in the excretion in the urine of not only S- but also of R-mevalonate.3 It seems, therefore, not unreasonable to suppose that the kidneys, on account of a renal clearance of mevalonate, are exposed to exceptionally high concentrations of mevalonate and that the very high utilization of mevalonate found in the kidneys is not so much a reficction of an esceptionnlly active sterol metabolism in these organs as of the specific clearing function of these organs.
Although observations were made at intervals varying from 3.5 to 96 hours after the injection of labeled substrates, no co11- sistent trend in the total amount of 14C in the unsaponifiable lipids was discernible with time elapsed after the injections; an exception to this was observed only in the 1*C content of the unsaponifiablc components in liver and blood.2 The observa- tions suggest that accumulation, as opposed to degradation, of sterols dominates the picture in these young animals as was noted many years ago in fetuses (41). A definite time-dependent change was observed in the distribution of 14C among the com- ponents of the unsaponifiable lipids: the longer t,he time after the injection of the labeled substrate, the greater the proportion of isotope in cholesterol. In all the organs examined, escepting the brain, all the radioactivity found ill sterol digitonides 24 and 72 hours after the administ,ration of a labeled precursor was in cholesterol. In the brain of the animals used in this study 22 to 24oj, of the mass of the sterols was attributable to desmosterol in confirmation of the data of Fumagalli and I’aoletti (42). However, 4 hours after the injection of any of the four labeled substrates as much as 46% of the isotope was found in cerebral desmosterol, but by 72 hours the distribution of isotope corre- sponded precisely to the proportions of cholesterol and desmos- terol by mass.
A comparison of the extent of incorporation of ‘“C into spinal cord and brain shows that between 9 and 12 days after birth the spinal cord has a greater capacity than the brain to utilize ketone bodies for the synthesis of sterols and fatty acids. This was expected as the spinal cord, unlike the brain at this time, is in the process of laying down myelin lamellae from as early as 4 days after birth (43). Thus, the spinal cord during active myelination exhibit,s a greater utilization of these precursors for lipid synthesis than the brain which is at the stage immediately before the onset of myelination.
The novel observation of these experiments was the preferen- tial utilization of ketone bodies for synthetic purposes in the central nervous system and skin as compared to other organs. The significantly higher conversion of /3-hydroxybutyrate, than of acetoacetate, to sterols and fatty acids in the three organs of ectodermal origin was nevertheless surprising on three counts. First, it has been shown by Klee and Sokoloff (44) that only the
The dramatic incorporation of 1% from [2-14C]mevalonate into 3 J. Edmond, H. L. Ngan, and G. PopjBk, unpublished observa- renal sterols, exceeding that found in liver sterols, was unex- tions.
by guest on January 19, 2020http://w
ww
.jbc.org/D
ownloaded from

n-isomer of P-hydroxybutyrate can function as substrate for the mitochondrial fl-hydroxybutyrate dehydrogenase. Since in the experiments equal amounts of on-fi-hydroxybutyrate and acetoacetate, containing identical amounts of 14C in correspond- ing positions at C-3, were administered, it was expected that if n-P-hydroxybutyrate and acetoacetate were used with equal efficiency for lipid biosynthesis, then on-P-hydroxy[3J4C]butyr- ate would give only half the amount of isotope found in lipids as compared to the amount of i4C incorporated from [3-14C]- acetoacetate. Without exception, nL-P-hydroxy[3-14C]butyrate was a more effective lipid precursor in the central nervous sys- tem and skin than [3-r4C]acetoacetate. Second, Hawkins et al. (29) have found that the concentration of P-hydroxybutyrate in the blood of 16-day-old rats was 5 times greater than that of acetoacetate. Thus, one would expect that after the adminis- tration of equal amounts of fi-hydroxybutyrate and acetoacetate of equal radioactivity, the specific activity of the circulating fl-hydrosybutyrate would be substantially lower than that of acet.oacetate. Third, Hawkins et al. (29) found also that the rate of uptake of acetoacetate by the brain in suckling rats was twice as great as that of P-hydroxybutyrate. These three fac- tors taken together should have favored acetoacetate over @-hydrosybutyrate as a substrate for lipid biosynthesis in the central nervous system, yet the opposite was found. Current experiments seek an explanation for the differentiation between t,hese two substrates.
The incorporation of label from [2-i4C]acetate into sterols and fatty acids was remarkably similar to that seen for [3J4C]aceto- acetate in all the organs excepting the liver which incorporated more isotope into sterol and fatty acids from acetate than from acetoacetate. The more efficient utilization of acetate than acetoacetate was, however, not apparent in the central nervous system and skin in which acetate gave a labeling pattern equiva- lent to that seen after the administration of acetoacetate. The results suggest that while acetate is probably activated to acetyl- CoA in the liver and used in that form for fatty acid and sterol biosynthesis there, the acetyl-CoA is rapidly converted in the liver into ketone bodies, which are then released into the blood and preferentially utilized by the central nervous system and skin. Supporting evidence for this contention may be found in the work of McGarry and Foster (20) who have demonstrated that (a) the liver is the main ketogenic organ; (b) the liver is capable of synthesizing in 24 hours an amount of acetoacetate equivalent to one-half of its weight; and (c) the rate of conver- sion of [i4C]acetate into acetoacetate in the liver is 20 times faster than its conversion into sterols.
The difference between the utilization of ketone bodies in the liver and the central nervous system can be readily csplained by a difference in the enzymic complement,s of these organs. Mali- ler (45) and others (46, 47) have concluded that free acetoacet’ate was metabolically inert in the liver because of the absence of succingl-Cod :3-osoacid Co-A transferase (EC 2.8.3.5. ) and the weakness, or absence, of acetoacetate: CoA lygase (aceto- acetyl-CoA synthetase) there.4 In contrast, the bran-devoid of 3-hgdroxy-3-methylglutaryl-Cob acetoacetate lyase (EC 4.1.3.4. )-is well endowed with all the enzymes needed for the utilization of P-hydroxybutyrate and acetoacetate, particularly during the neonatal period. Thus, the levels of both n( -)-fi-
4 A recent report by Stern (48) indicates, however, that the liver of adult fed rats contains acetoacetyl-CoA synthetasc. Unpub- lished experiments in this laboratory suggest that the acetoacetyl- CoA synt,hetase is very weak in the liver of newborn rats, but that it develops after weaning.
79
hydroxybutyrate dehydrogenase (EC 1.1.1.30) and 3-oxoacid CoA-transferase rise rapidly after birth in rat brain to values 3 to 4 times higher than the levels found in adults (28). The high levels of acetoacetyl-Coil thiolase activities in the brain of the rat during development (49) are also consistent with a high utilization of ketone bodies. Hawkins et al. (29) have demon- strated not only a high concentration of ketone bodies in the blood of suckling rats but also a high transfer of acetoacetate and P-hydroxybutyrate from the blood to the brain in these animals. They found that the concentration of ketone bodies in the venous blood returning from the brain of 16- to 18- day-old rats was 0.32 to 0.44 mM less than in the arterial blood. Furthermore, at the physiological concentrations of ketone bodies the arteriovenous differences were 12 to 17 times greater in suckling animals than in adults. Hawkins et al. (29) calcu- lated from their data that as much as 48 to 76% of the energy requirements of the brain of the, 16. to 18.day-old rat can be accounted for by the metabolism of ketone bodies. These data taken together with the results presented here indicate that t,he ketone bodies are not only major metabolic fuels for energy pro- duction, but are also sources of carbon for the synthesis of sterols and fatty acids in the central nervous system of the suckling rat. The skin, which accounts for 22% of the body weight in young animals, shares with the central nervous system a great avidity for ketone bodies and high synthetic activity.
The experiments or1 whole animals brought to light a beautiful interdependence of the organs of the body. The liver is com- mitted to the production of ketone bodies, which it can hardly utilize, whereas the central nervous system, which does not make free ketone bodies, depends totally 011 the ketogenic organs for their supply. In fact, the uptake of ketone bodies by the brain is proportional to their concentration (29) maintained in the blood by the ketogenic organs. The dependence of the central nervous system 011 ketone bodies during the myelination process, rather than on carbohydrate, is underscored by the finding that there is a net efflux of lactate and pyruvate from the brain of suckling rats in quaruities equivalent to as much as 60% of the amount of glucose imported (29). It is postulated, consequently, that ketone bodies play a major role in the central nervous sys- tem by providing the principal lipid precursors during the crucial period of myelination.
&knowledgment-I am indebted to Professor G. l’opjak for his support during the course of this study and for his help with the preparation of this manuscript.
1.
2.
3.
4.
5. 6.
7.
8.
9. 10.
SI~ULY, W. M., ~~~~~~~~~~ H., .\ND STOYINOFF, V. A. (1940) J. Viol. Chem. 136, 281-296 297-302
WIELSCH, H., SPERRY, W. M., .\ND STOY.INOFF, V. A. (1941) J. Viol. Chem. 140, 885-897
CHKVILLIER, F., .\ND PETIT, L. (1966) Exp. Neural. 16, 250- 254
CLIRICNIKJILG, II., CHJJKOFF, I. L., .ZND MORRIS, M. I>. (19G3) J. Neurochem. 10, 135-143
DOHHING, J. (1903) J. i\‘eurochem. 10, 739-742 PI:TY:ILSOIV, N. A., .\ND CH.UKOFF, I. L. (19B3) J. Neurochem.
10, 17-23 DIVISON, A. N., DOHHING, J., MORGIN, It. S., :~ND PIYLING-
WRIGHT, G. (1958) J. Neurochem. 3, 89-9-1 D.\VISOX, A. N., I~OIJIUNG, J., h/IowxN, R. S., .\ND PIYLING-
WIIIGIIT, G. (1959) Laricet 1, G58-GGO D~~VISON, A. N., .\ND W.\.tua, M. (1959) XValure 183, lGOG-lGO7 I~Ho~I.;sH\~.~RI~~R, G. A., AND MEID, J. F. (1969) Biochim.
Biophys. Acla 167, 461-467
by guest on January 19, 2020http://w
ww
.jbc.org/D
ownloaded from

80
11.
12.
13.
14.
15.
16.
17. 18.
DHOPESHH.ARKAR, G. A., AND MEAD, J. F. (1970) Biochim. 30. GOODMaN, D. S., AND POPJ~K, G. (1960) J. Lipid Res. 1, 286- Biophys. Acta 210, 250-256 300
DHOPESHwARKBR,G. A., SUIlRAM.4NIAN, C., AND MEND, J. F. 31. POPJ~~K, G.,-LOGE, A. E., AND MOORE, D. (1962) J. Lipid Res. (1971) Biochim. Biophys. Acta 239, 162-167 3, 364-371
DHOPESHWARKAR, G. A., SUBR.~MANIAN, C., AND MEAD, J. F. 32. GIVER, It. C., AND SWEELEY, C. C. (1966) J. Amer. Chem. (1971) Biochim..Biophys. Acta 231, 8-14
NAQVI, S. H. M., JONES, J. R., AND NICHOLAS, H. J. (1970) J. Amer. Oil Chem. Sot. 4’7, 92A
RAMSEY, R.B., JONES, J. P.,N~QvI,S.H.M., ANDNICHOLAS, H. J. (1971) Lipids 6, 154-161
GSRBTTINI, S., PAOLETTI, P., AND PAOLETTI, II. (1959) Arch. Biochem. Biophys. 80, 210-211
NICHOLAS, H. J. (1961) J. Kansas Med. Sot. 62, 358-361 KOREY, S. It., .~ND STEIN, A. (1961) in Regional Neurochem-
istry, (KETY, S. S., AND ELICES, J., eds) pp. 175-189, Perga- mon Press, Oxford
33.
34.
35.
36.
37.
19.
20.
21. 22.
23.
24.
25.
26. 27.
K.~NDUTSCH, A. A., AND SXJCIER, S. E. (1969) Arch. Biochem. Biophys. 136, 201-208
38.
MCG~RRY, J. D., .~ND FOSTER, D. W. (1969) J. Biol. Chem. 244, 4251-4256
KaBaRA, J. J. (1965) J. Amer. Oil Chem. Sot. 42, 1003-1008 GARATTINI, S., P~OLETTI, P., AND PAOLETTI, It. (1959) Arch.
Biochem. Biophys. 64, 253-255 KABARA, J. J., AND OKIT~, G. T. (1961) J. Neurochem. 7, 298-
304
39. 40.
41.
KELLEY,M. T., AEXEL, R.T., HERNDON, B.L., ANDNICHOLAS, H. J. (1969) J. Lipid Res. 10, 166-i-174
RAMSEY, R.B., JONES, J.P., NAQVI,~. H.M., AND NICHOLAS, H. J. (1971) Lipids 6, 225-232
PSOLETTI, E. G. (1971) Advan. Exp. Med. Biol. 13, 41-51 POPJ~K, G. (1969) Methods Enzymol. 16, 393-454
42. 43. 44.
45. 46.
47.
28. PAGE, M. A., KREBS, II. A., AND WILLIAMSON, D. H. (1971) Biochem. J. 121, 49-53 48.
29. HAWKINS, It. A., WILLIAMSON, D. H., AND KREBS, H. A. (1971) Biochem. J. 122, 13-18 49.
Sot. 88, 3643-3647 VOGEL, A. I. (1967) Practical Organic Chemistry, 3rd Ed, p.
971, Longmans, Green and Co., Ltd., London CUZNER, M. L., AND DAVISON, A. N. (1968) Biochem. J. 106,
29-34 HAGENFELDT, L., AND HELLSTR~M, K. (1972) Life Sci. 11,
669476 EDGREN, B., AND HELLSTR~M, K. (1972) Acta Physiol. Stand.
86, 250-256 EDMOND, J., AND POPJ.&K, G. (1974) J. Biol. Chem. 249,
66-71 CUZZOPOLI, M., HELLSTRBM, K., AND SVENSSON, B. (1972)
Metabolism 21, 1161-1170 GOULD, It. G., AND POPJ~K, G. (1957) Biochem. J. 66, 51P CORNFORTH,~~. H., FLETCHER, K., HELLIG, H., AND POPJ~~K,
G. (1960) Nature 186, 923 POPJ~K, G., AND BEECKMANS, M.-L. (1950) Biochem. J. 46,
547-561 FUM~GALLI, It., AND P.IOLETTI, R. (1963) Life Sci. 6, 291-295 JACOBSON, S. (1968) J. Comp. Neural. 121, 5-29 KLEE, C. B., AND SOKOLOFF, L. (1967) J. Biol. Chem. 242,
3880-3883 MAHLER, H. R. (1953) Fed. Proc. 12, 694-702 STERN, J. It., COON, M. J., AND DEL CAMPILLO, A. (1953)
Nature 1’71, 28-30 WILLIAMSON, D. H., AND HEMS, It. (1970) in Essays in Cell
Metabolism (BARTLEY, W., KORNBERG, H. L., AND &UBYLE, J. R., eds) pp. 251-281, Wiley-Interscience, London
STERN, J. R. (1971) Biochem. Biophys. Res. Commun. 44, lOOl- 1007
MIDDLETON, B. (1971) Biochem. J. 126, 70P
by guest on January 19, 2020http://w
ww
.jbc.org/D
ownloaded from

John EdmondKetone Bodies as Precursors of Sterols and Fatty Acids in the Developing Rat
1974, 249:72-80.J. Biol. Chem.
http://www.jbc.org/content/249/1/72Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/249/1/72.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on January 19, 2020http://w
ww
.jbc.org/D
ownloaded from





























