Embed Size (px)
Citation preview

1
Attenuation of chikungunya vaccine strain 181/clone 25 is determined by 2 amino acid 1 substitutions in the E2 envelope glycoprotein 2 3 Rodion Gorchakova, Eryu Wanga, Grace Leala, Naomi L. Forrestera, Kenneth Plantea, Shannan L. 4 Rossia, Charalambos D. Partidosb, A. Paige Adamsa, Robert Seymoura, James Wegerb, Erin M. 5 Borlandc, Michael B. Shermand, Ann. M. Powersc, Jorge E. Osoriob, e and Scott C. Weavera* 6 7 aInstitute for Human Infections and Immunity, Sealy Center for Vaccine Development, and 8 Department of Pathology, University of Texas Medical Branch, Galveston, TX 77555, United 9 States 10 bInviragen, Inc., Madison, WI 53719, United States 11 cDivision of Vector Borne Infectious Diseases, Centers for Disease Control and Prevention, Fort 12 Collins, CO, United States 13 dSealy Center for Structural Biology and Molecular Biophysics, Department of Biochemistry and 14 Molecular Biology, University of Texas Medical Branch, Galveston, TX 77555, United States 15 eSchool of Veterinary Medicine, University of Wisconsin, Madison, WI 53706 16 17
*Corresponding author at: Galveston National Laboratory, University of Texas Medical 18 Branch, 301 University Boulevard, Galveston, TX 77555-0610, United States. Tel.: +1 409 266 19 6500; fax +1 409 266 6810 20
E-mail address: [email protected] (S. C. Weaver) 21 22
Copyright © 2012, American Society for Microbiology. All Rights Reserved.J. Virol. doi:10.1128/JVI.06449-11 JVI Accepts, published online ahead of print on 28 March 2012
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

2
Abstract 23 24 Chikungunya virus (CHIKV) is a mosquito-borne alphavirus and the etiologic agent of 25
massive outbreaks of arthralgic febrile illness that recently affected millions of people in Africa 26 and Asia. The only CHIKV vaccine that has been tested in humans, strain 181/clone 25, is a live-27 attenuated derivative of Southeast Asian human isolate, strain AF15561. The vaccine was 28 immunogenic in phase I and II clinical trials; however it induced transient arthralgia in 8% of 29 vaccinees. There are 5 amino acid differences between the vaccine and its parent, as well as 5 30 synonymous mutations, none of which involves cis-acting genome regions known to be 31 responsible for replication or packaging. To identify the determinants of attenuation, we 32 therefore tested the 5 nonsynonymous mutations by cloning them individually or in different 33 combinations into infectious clones derived from 2 wild-type (wt) CHIKV strains, La Reunion 34 and AF15561. Levels of virulence were compared with those of the wt strains and the vaccine 35 strain in 2 different murine models: infant CD1 and adult A129 mice. An attenuated phenotype 36 indistinguishable from that of the 181/clone 25 vaccine strain was obtained by the simultaneous 37 expression of 2 E2 glycoprotein substitutions, with intermediate level of attenuation observed for 38 the single E2 mutants. The other 3 amino acid mutations, in nsP1, 6K, and E1, did not have a 39 detectable effect on CHIKV virulence. These results indicate that the attenuation of strain 40 181/clone 25 is mediated by 2 point mutations, explaining the phenotypic instability observed in 41 human vaccinees and also in our studies. 42 43
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

3
Introduction 44 Chikungunya virus (CHIKV) is a mosquito-borne human pathogen in the genus Alphavirus, 45
family Togaviridae. First isolated by Ross during a 1952-53 epidemic of arthralgic disease in 46 Tanzania, CHIKV is now known to occur in many parts of Sub-Saharan Africa as well as in Asia 47 (33, 47). Human disease occurs through two different mechanisms: 1) direct spillover from 48 enzootic transmission cycles in forests of Africa, where arboreal mosquitoes transmit CHIKV 49 among nonhuman primates, and; 2) an urban transmission cycle involving humans as 50 amplification hosts and the peridomestic mosquitoes, Aedes (Stegomyia) aegypti and A. 51 (Stegomyia) albopictus as vectors. Endemic disease caused by enzootic spillover is rarely 52 recognized because chikungunya fever (CHIK) is not easily distinguished from many other 53 tropical diseases such as dengue or malaria without laboratory diagnostics. The urban form of 54 CHIK occurs in both Africa and Asia, and emergences of the urban cycle from African, enzootic 55 strains have probably occurred for centuries, aided initially by transport on sailing ships and 56 more recently by air travel by infected persons (33, 47, 55). Unprecedented epidemic activity has 57 been documented since 2004, when outbreaks first appeared in coastal Kenya (16), followed by 58 transport of the virus to islands in the Indian Ocean (37) and then independently to India (50). 59 These epidemics resulted in thousands of infected travelers returning to Europe, the Americas 60 and Southeast Asia, in some cases initiating urban transmission cycles (13, 34, 40, 42). Since 61 2004, millions of persons have been infected with CHIKV and the epidemics continue to occur 62 in India and Southeast Asia (41). 63
Chikungunya virus infection is typically characterized by an abrupt onset of fever and 64 polyarthralgia, often accompanied by rash (38, 41). Although many patients recover completely 65 within a few weeks of onset, the arthralgia, which is extremely painful and debilitating, can be 66
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

4
persistent, and has been shown to be a major cause of morbidity over one year after the end of 67 epidemics (11). Fatalities following CHIKV infection have also been documented (24), including 68 neurologic disease (5, 9), and excess deaths reported during epidemics suggest that CHIKV 69 infection is at least partially responsible for many fatal outcomes (15, 24, 43). 70
Like other alphaviruses, CHIKV has a single-stranded, positive sense RNA genome of about 71 11.8 kB that encodes 2 open reading frames (ORF): 1) a nonstructural polyprotein ORF encoded 72 in the 5’ 2/3rd of the genome, which is translated directly from genomic RNA to produce 4 73 nonstructural proteins involved in genome replication, capping, and polyprotein processing, and; 74 2) a structural polyprotein ORF that is identical in sequence to the 3’ 1/3 of the genome, is 75 encoded by a subgenomic RNA, and produces the capsid as well as the E2 and E1 envelope 76 glycoproteins (20). Virions are composed of the genomic RNA complexed with 240 copies of 77 the capsid protein, which form nucleocapsids in the cytoplasm of infected cells, and 240 E2/E1 78 heterodimers, which are embedded in the plasma membrane following their transit through the 79 secretory system. Thus, the 70 nm virion is enveloped, with 80 trimeric E2/E1 spikes projecting 80 outward. The E2 protein forms the tips of the spikes and is believed to interact with cellular 81 receptors (52). It is also the target of most alphavirus-neutralizing antibodies (35). 82
Although vaccine candidates have been developed for CHIK, only one has been tested in 83 humans. Levitt et al. (21) used a wild-type (wt) CHIKV strain isolated in Thailand, AF15561, 84 and performed 18 plaque-to-plaque passages in MRC-5 human lung fibroblast cultures and a 85 plaque-clone designated 181/clone 25 (hereafter abbreviated 181/25) was selected for further 86 testing based on its small plaque size, attenuation for infant mice, and reduced viremia in 87 nonhuman primates. Strain 181/25 elicited neutralizing antibodies after a single vaccination of 88 mice or rhesus macaques, and protected against both viremia and disease following wt CHIKV 89
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

5
challenge. This vaccine (called TSI-GSD-218 during human trials) also proved highly 90 immunogenic in humans, with all but one vaccinee developing neutralizing antibodies (8). 91 However, 5/59 vaccinees developed mild, transient arthralgia, suggesting insufficient and/or 92 unstable attenuation, and no further clinical trials have been undertaken. Recent studies of 93 another live-attenuated CHIK vaccine candidate that included direct comparisons with strain 94 181/25 also reported some residual pathogenicity of the latter (32). 95
Subsequent sequence comparisons between CHIKV parental strain AF15561 (GenBank 96 Accession No. EF452493) and the derived, attenuated strain 181/25 (GenBank Acc. No. L37661, 97 listed as TSI-GSD-218) revealed 10 nucleotide differences in the genome, with 5 being 98 nonsynonymous and none involving cis-acting genome regions known to be responsible for 99 replication or packaging (Fig. 1a, Table 1). To further assess the attenuation of strain 181/25 and 100 to determine the mutations responsible for its attenuation, we conducted reverse genetic studies 101 to examine the attenuating effects of the amino acid changes encoded by these 5 nonsynonymous 102 mutations that accompanied its derivation. 103 104
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

6
Methods 105 Cell culture. Vero African green monkey kidney cells (ATCC CCL-81, Bethesda, MD) were 106 maintained in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% fetal 107 bovine serum, non-essential amino acids, sodium pyruvate, and penicillin/streptomycin at 370C 108 under 5% CO2. 109
110 Virus stocks. CHIKV strain 181/25 (21) was passaged once on Vero cells. Stocks of all other wt 111 and mutant CHIKV strains were generated by electroporation of Vero cells with in vitro 112 transcribed RNAs from cDNA infectious clones (see below). Briefly, sub-confluent cell cultures 113 from two T150 flasks were trypsinized, washed 3 times with Dulbecco’s PBS, resuspended in 114 700 µl of PBS, mixed with 10 µl of the in vitro transcribed RNA suspension, and electroporated 115 using a BTX ECM 830 instrument (Harvard Apparatus, Holliston, MA) in 4 mm cuvettes with 3 116 ten-millisecond pulses at 250 V, 1 second intervals. Cells were mixed with 10 ml of DMEM after 117 standing at room temperature for 10 min and seeded into T75 flasks. Cell supernatants 118 containing virus were harvested at 22-26 hours post-electroporation, after the development of 119 visible cytopathic effects (CPE), clarified by centrifugation at 1,000 x G for 5 min, and titered by 120 plaque assay on Vero cells as described previously (1). 121
122 Viral infectious clones plasmids. The CHIKV La Reunion (LR) cDNA infectious clone (45) 123 was provided by Steven Higgs at the University of Texas Medical Branch (UTMB). The CHIKV 124 strain AF15561 (21) sequence was retrieved from GenBank (EF452493, AF15561) and a full-125 length cDNA of this viral sequence was synthesized in 5 fragments by GenScript (Piscataway, 126 NJ). A Sac I restriction enzyme site and a SP6 RNA polymerase promoter were added at the 5’ 127
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

7
terminus, and a polyA tail followed by a Not I restriction site were added at the 3’ terminus. 128 Ambiguous nucleotides present in the GenBank sequence were replaced as described in the 129 Results section. These 5 cDNA fragments were assembled using standard DNA cloning 130 techniques together with the Not I – Sac I DNA fragment from CHIKV LR infectious clone, 131 which contained only the cloning vector, to generate the CHIKV 15561 cDNA infectious clone. 132
The CHIKV vaccine strain 181/25 sequence was obtained from GenBank (L37661, TSI-133 GSD-218). Compared to strain AF15561, its wt parent, there are 5 nonsynonymous mutations in 134 its genome (Table 1 and Fig. 1a). Using PCR-mediated mutagenesis, these mutations were 135 cloned separately or in combinations into the CHIKV LR or 15561 plasmids to generate cDNA 136 infectious clones of the mutants (Figs. 1b, 5). 137
138 Plasmid purification and in vitro transcription. Large-scale preparations of infectious clone 139 plasmids were made using standard methods, and plasmids were purified on CsCl gradients (36). 140 All in vitro synthesized and PCR-derived DNA sequences were confirmed using the BigDye 141 Terminator v3.1 Cycle Sequencing Kit on 3500 Genetic Analyser (Applied Biosystems, Foster 142 City, CA). Plasmids were linearized using the Not I restriction enzyme prior to in vitro 143 transcription using SP6 RNA polymerase (Ambion, Austin, TX) in the presence of RNA cap 144 analog using the manufacturer’s protocol. The quality and integrity of RNA were confirmed by 145 agarose gel electrophoresis in non-denaturing conditions and stored at -800C. 146
147 CD1 mouse infections. All mouse experiments were approved by the Institutional Animal Care 148 and Use Committees of UTMB or the University of Wisconsin (UW). Pregnant CD1 mice were 149 purchased from Charles River Laboratories (Wilmington, MA). Cohorts (n=15-36) of 5-6- or 6-150 7-day-old mice were infected subcutaneously (SC) in the back with 105 PFU of virus. For 151
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

8
experiments with mutations in the CHIKV strain LR backbone, body weights were measured 152 daily, and blood, hind limb, and brain tissues were harvested (n=2-3) on days 2, 4, 6, 8, and 10 153 post-infection. For experiments with mutations in the AF15561 backbone, blood, hind limb, and 154 brain tissues were harvested (n=6) on days 1-5, and 7 post-infection. Viremia and viral loads 155 were measured by plaque assay on Vero cells. 156
157 A129 mouse infections. Strain A129 mice were bred at UW or UTMB from a breeding pair 158 obtained from B & K ltd. (Grimston, UK). Telemetric devices (Bio Medic Data Systems, Inc., 159 Seaford, DE) were implanted SC into mice one week prior to infections. Cohorts (n=4-9) of 5-8- 160 or 8-10-week-old mice were infected intradermally (ID) in the left rear footpad with 104 PFU of 161 virus. Animals were monitored for 18 days for morbidity and mortality, and body weights and 162 temperatures were measured daily. Mice were bled on days 1-4 (LR mutants) or on days 1 and 3 163 post infection (AF15561 mutants) and viremia was determined by quantitative reverse 164 transcription PCR (qRT-PCR) as described previously (32) or by plaque assay on Vero cells, 165 respectively. Measurements of the perimetatarsal height of the hind feet were taken at 48 hours 166 (LR mutants) or 2, 4 and 7 days (AF15561 mutants) post infection. 167
168 Viral RNA extractions and sequencing. Total RNA was extracted from 200µl of CHIKV strain 169 181/25 viral stocks or 50 µl of blood collected from infected mice using TRIzol LS (Invitrogen, 170 Carlsbad, CA) using the manufacturer’s protocol. cDNA was synthesized using the 171 SuperScriptIII First Strand Kit (Invitrogen) with random hexamer primers. Three µl of reverse 172 transcription reactions were used for PCR reactions with Phusion Hot Start II High-Fidelity 173 DNA Polymerase (New England BioLabs, Ipswich, MA) with CHIKV-specific primers 174
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

9
(sequences available upon request). Amplicons were purified from agarose gels with the 175 QIAquick Gel Extraction Kit (Qiagen, Valencia, CA) and used for sequencing as described 176 above. 177 178
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

10
Results 179 Construction of CHIKV LR clones with strain 181/25 mutations. Alignment of sequences of 180 CHIKV strain AF15561 and its vaccine derivative strain 181/25 revealed 10 nucleotide 181 differences, all within ORFs of the genome. Five of these changes were synonymous and the 182 other 5 led to deduced amino acid differences, 2 in the E2 envelope glycoprotein and one in each 183 of the following proteins: nsP1, 6K, and E1 (Table 1 and Fig. 1a). The 5 synonymous mutations 184 that accompanied derivation of strain 181/25 were assumed to have little or no phenotypic effect, 185 so our efforts focused on the nonsynonymous mutations. Therefore, they were incorporated 186 individually into the genome of the CHIKV strain LR infectious clone (Fig. 1b) derived from a 187 wt human isolate from the La Reunion Island in the Indian Ocean (45). 188
Mutagenized cDNA clones, along with the wt LR clone, were used to produce infectious 189 RNA in vitro, which was electroporated into Vero cells. All rescued mutant viruses were viable, 190 producing CPE in Vero cells within 24 hours of electroporation, and replicating to 5-6 log10 191 plaque forming units (PFU)/ml in cell supernatants, comparable to the wt virus (data not shown). 192 One noticeable phenotype of the LR/E2-82 mutant was a reduced plaque size on Vero cells. This 193 may be attributed to the addition of a positive charge (glycine→arginine) at a putative heparan 194 sulfate binding site of E2, which can lead to increased interaction of the viral particles with the 195 polyanions in agarose media during plaque formation (3, 23). The phenomenon of positive 196 charge acquisition in the E2 glycoprotein is commonly observed when alphaviruses are passaged 197 on cultured cell lines, and often correlates with the acquisition of an attenuated phenotype (7, 14, 198 19). 199
200
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

11
Infection of infant CD1 mice with CHIKV LR mutants. Several mouse models have been 201 developed to study the virulence of CHIKV (6, 10, 26, 56). One utilizes infant outbred CD1 mice, 202 which develop disease similar to that seen in humans (56). We tested the attenuation of the 5 203 single CHIKV LR mutants by infecting SC cohorts (n=15-18) of 6-7-old CD1 mice with 105 204 PFU. Cohorts were also infected with wt CHIKV LR and vaccine strain 181/25. Body weights 205 were measured daily as indicated in Fig. 2, and all cohorts of mice continued to gain weight at 206 similar rates, including those infected with wt LR virus. For this reason, body weights were not 207 monitored further in CD1 mouse experiments. 208
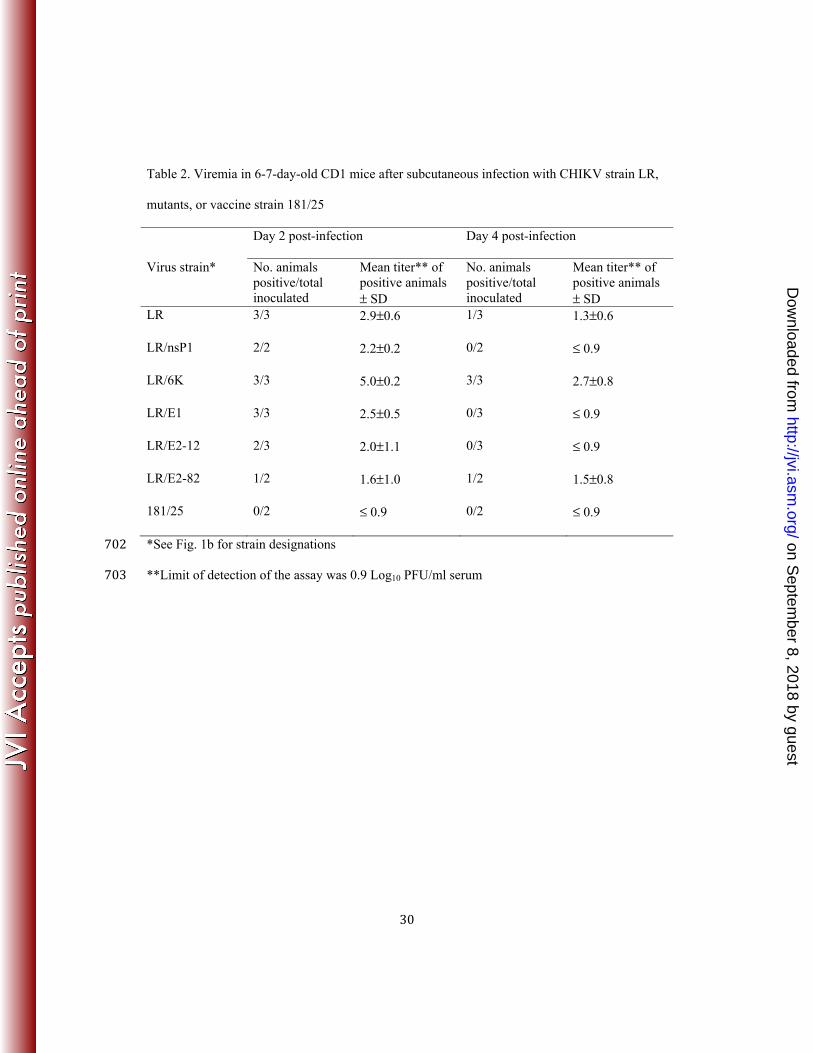
Groups of 2-3 mice from each cohort were sacrificed on days 2, 4, 6, 8, 10 after infection and 209 blood, hind limb, and brain tissues were collected to determine viral loads. No viremia was 210 detected on days 2 and 4 post infection in 181/25-infected mice, whereas 3/3 strain LR-infected 211 mice were positive with a mean viremia titer of 2.9 log10 PFU/ml on day 2 with partial clearance 212 by day 4 with only 1/3 viremic mice (Table 2). The LR/6K mutant was the most virulent, with all 213 mice tested positive on days 2 and 4 after infection and with mean titers higher than those of the 214 wt LR strain. Although it is possible that the 6K mutations slightly increases virulence of 215 CHIKV, a more likely explanation is that this cohort, which was tested independently of the 216 others, was affected by maternal care or health, or a slightly lower age of the animals. The lack 217 of any difference between the triple mutant described below, which includes the 6K mutation, 218 and wild-type CHIKV, supports this explanation. The other 4 single mutants produced 219 intermediate viremia phenotypes (between wt and vaccine strains), in terms of the fraction of 220 positive mice and mean titers, with LR/nsP1- and LR/E1-infected mice exhibiting more virulent 221 phenotypes comparable to that of wt CHIKV at day 2 post infection (Table 2). No viremia was 222 detected in samples tested on day 6 after infection (data not shown). 223
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

12
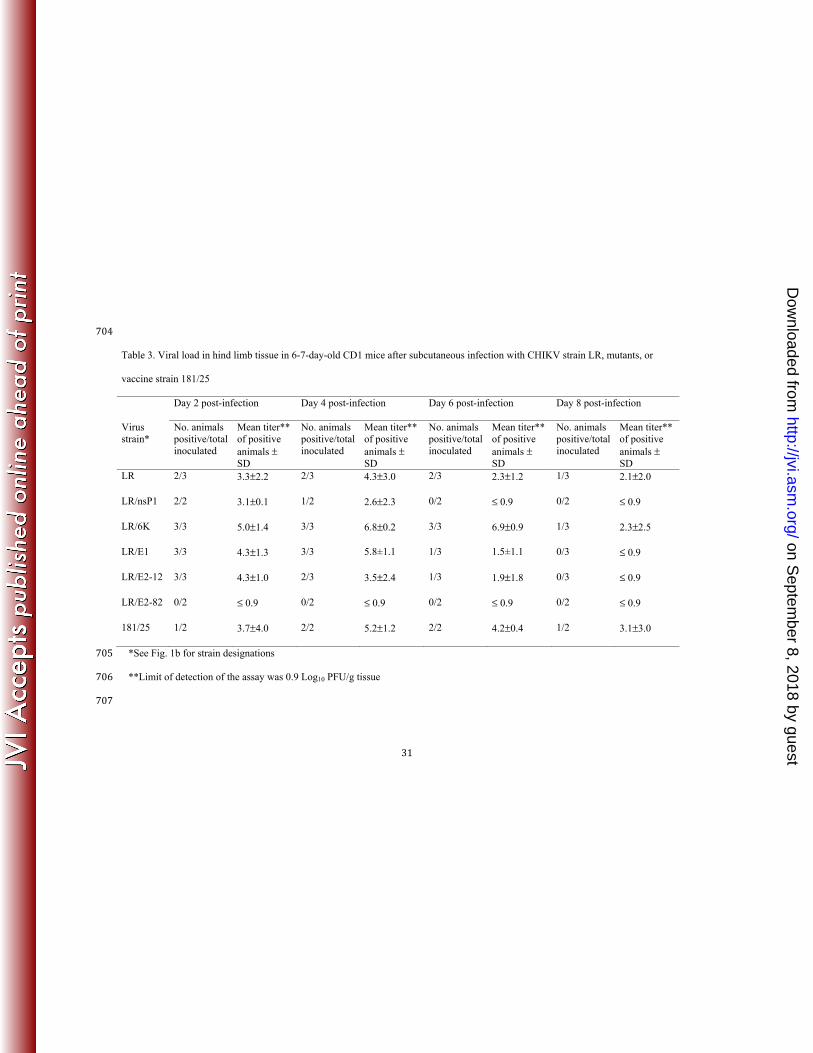
Analysis of viral loads in the hind limb tissues showed high viral titers in both wt- and 224 vaccine-infected mice on days 2, 4, and 6 (Table 3), followed by clearance of the viruses by day 225 10, when only 1 mouse (infected with LR strain) had virus present out of all sampled cohorts 226 (data not shown). High viral loads in the knee tissues of the 181/25-infected mice might correlate 227 with transient arthralgia in some of the vaccinees during human trials (8). The LR/6K mutant 228 produced the highest mean titers, and the LR/E1 and LR/E2-12 mutants also had phenotypes 229 similar to wt CHIKV, with comparable mean hind limb titers on days 2 and 6 after infection. The 230 single mutants, with the exception of LR/6K, exhibited less virulent phenotypes when compared 231 to the 181/25 strain, with lower titers at several time points and clearance by day 8 (Table 3). The 232 LR/E2-82 mutant was the most attenuated, with no detectable virus in the hind limb at any time 233 post infection. The differences in viral titers between the vaccine strain and some of the 234 individual mutants could be due to different genomic backgrounds (AF15561 vs. LR, 235 respectively). 236
CHIKV was detected in the brains of LR-infected CD1 mice up to day 6, while only 1 mouse 237 infected with vaccine strain 181/25 was positive (on day 4) during the time course of the 238 experiment (Table 4). Again, strains LR/6K and LR/E1 were the most virulent mutants, behaving 239 as wt CHIKV. Similar to the results from the limb tissues, there was no virus in the brains of any 240 LR/E2-82 mutant-infected mice at any time point. Also, the LR/E2-12 mutant was detected in 241 the brain of only in 1 mouse on day 4 after infection. All brains sampled were virus-negative at 242 day 10 post infection (data not shown). 243
In summary, infections of infant CD1 mice with the single mutants showed the attenuation 244 phenotype of LR/E2-82 to be close to that of vaccine strain 181/25. To confirm that the 245 remaining 4 nonsynonymous mutations did not strongly influence virulence, we constructed an 246
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

13
additional clone, LR/4x (Fig. 1b), with these 4 amino acid substitutions (in nsP1, E2-12, 6K, and 247 E1) combined in the LR genomic background. The LR/4x mutant virus was successfully rescued 248 in Vero cells and was used to infect 5-6-day old CD1 mice as described above, along with wt LR 249 and 181/25 controls. As expected, the LR/4x mutant behaved like the wt LR strain, showing high 250 viremia titers up to day 4 after infection (Table 5). In contrast to the lack of viremia in the 251 previous experiment, 33% of 181/25-infected mice produced viremia on days 2 and 4 post 252 infection, which may be attributed to the slightly younger age of the mice. 253
The 6K protein mutation was viewed as unlikely to be involved in attenuation; this protein is 254 not known to affect viral replication, it only acts as the signal peptide for E1 glycoprotein, and 255 only small amounts of 6K remain associated with the viral particle (20). Indeed, incorporation of 256 the 6K protein mutation in LR strain backbone, in the form of either LR/6K or LR/4x, did not 257 produce any detectable attenuation. For these reasons the single 6K mutation was not tested 258 further. 259
260 Infection of A129 mice with CHIKV LR mutants. Another mouse model of CHIKV infection 261 is the A129 inbred mouse (6). This strain has impaired type-I interferon signaling due to inactive 262 interferon-alpha/beta receptors. Cohorts (n=4-5) of 8-10-week-old A129 mice were infected ID 263 in the footpad with 104 PFU of vaccine strain 181/25 or the 4 single mutants (Fig. 1b) and 264 monitored for morbidity and mortality. All 181/25-infected mice survived up until day 18 post 265 infection when the experiment was terminated. However, the animals that received wt LR, 266 LR/nsP1, and LR/E1 mutants succumbed to infection by day 4 post infection (Fig. 3). The 267 LR/E2-12 and LR/E2-82 mutant-infected mice had significantly extended survival times (P<0.01, 268 Mantel-Cox test), but all died by day 7 post infection. All 6 groups of mice gradually lost weight 269
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

14
until day 7. However, mice infected with vaccine strain 181/25 began to recover after that time 270 point (Fig. 4a). Morbidity was also reflected in body temperature changes; in moribund mice 271 there was a slight hyperthermia for the first 1-3 days post infection, followed by distinct 272 hypothermia 1-2 days before death (P<0.05 on day 6 and P<0.001 on day 7 for the E2 single 273 mutants, one-way ANOVA with Tukey-Kramer post-test, compared to 181/25) (Fig. 4b). Slight 274 hyperthermia in 181/25-infected mice was sustained until day 5 post infection, and then body 275 temperatures returned to normal (Fig. 4b). 276
Swelling of the footpad, the location of ID injection, is another indication of CHIK disease in 277 murine models. Measurement of the perimetatarsal height of the infected limb is used to assess 278 swelling (10). Such measurements taken at day 2 post infection indicated significant (P<0.05 or 279 less by one-way ANOVA with Tukey-Kramer post-test) swelling induced by mutants LR/nsP1 280 and LR/E1 compared to 181/25- or sham-infected mice (Fig. 4c). Footpad swelling in the wt LR-281 infected mice was also overt. However, because of one animal with an outlier value, it was not 282 significantly different by one-way ANOVA compared to 181/25-infected mice. Animals infected 283 with the LR/E2-82 or LR/E2-12 mutant did not exhibit significant swelling (P>0.05 by one-way 284 ANOVA) compared to controls. Analysis of virus content in the feet should be examined in 285 future CHIK pathogenesis experiments. 286
A129 mice were bled daily for the first 4 days post infection and viremia was determined 287 using qRT-PCR due to the small volume of collected blood. Mice infected with the LR/nsP1 and 288 LR/E1 mutants consistently produced viremia that did not differ significantly (P>0.05, one-way 289 ANOVA, except on day 1 post infection), up to 6.8 log10 PFU/ml for LR/E1 on day 3. The 290 LR/E2-82 mutant- and 181/25-infected mice showed lower and similar viremia levels (P>0.05, 291
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

15
one-way ANOVA), except on day 4 post infection, while the LR/E2-12 mutant-infected mice 292 developed intermediate viremia titers (Table 6). 293
In summary, these A129 mouse experiments revealed partial attenuation of CHIKV caused 294 by expression of the E2-12 and E2-82 mutations in the LR strain background. The LR/nsP1 and 295 LR/E1 mutants showed little or no attenuation and exhibited the virulent, wt phenotype. 296
297 Construction of CHIKV AF15561 infectious clone and its mutants. Different CHIKV strains 298 have been shown to differ in their response to mutations that can enhance mosquito vector 299 infectivity (46, 48). Therefore, to eliminate the possibility that the CHIKV genomic background 300 affected the expression of the attenuation phenotype of the 181/25 strain mutations, we 301 constructed an infectious cDNA clone of strain AF15561, the parental strain of 181/25. This 302 strain shares 92.9% nucleotide and 97.7% amino acid identity with the LR strain. The AF15561 303 strain sequence was retrieved from GenBank (Acc. No. EF452493), furnished with 5’ SP6 RNA 304 polymerase promoter and 3’ polyA tail, and divided in 5 fragments according to availability of 305 convenient restriction sites. These 5 DNA fragments were synthesized in vitro and ligated 306 together in 2 steps using the cloning vector plasmid fragment from the CHIKV LR infectious 307 clone. The GenBank sequence of the AF15561 strain contains 4 ambiguous nucleotides that do 308 not affect the amino acid sequence, and these were assigned in the clone, based on the 181/25 309 sequence, as follows: K9→G, Y412→T, Y1612→C, Y6994→T (nucleotide numbering is from the 5’ 310 of the CHIKV genome). Electroporation of Vero cells with in vitro synthesized RNA of 311 AF15561 strain caused CPE within 24 hours, and infectivity of this RNA was 1.4x105 PFU/4 µg. 312 The viral stock titer was 1.5x107 PFU/ml and produced medium-sized plaques on Vero cells. 313
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

16
These data indicated that the generated AF15561 virus was viable and did not require 314 compensatory mutations to be rescued in Vero cells. 315
The AF15561 clone was mutated to incorporate the 181/25-specific amino acid substitutions 316 (see clone designations in Fig. 5): E2-12 and E2-82 were mutated individually and together in 317 one clone, and the remaining 3 nonsynonymous mutations (nsP1, 6K, E1) were included in one 318 construct. Viral stocks of the mutants were generated in Vero cells as described above, with titers 319 of 6-7 log10 PFU/ml (data not shown). As was observed with the LR/E2-82 virus, the 15561/E2-320 82 mutant produced smaller plaques on Vero cells. 321
322 Infection of infant CD1 mice with CHIKV AF15561 mutants. Cohorts (n=36) of 5-6-day-old 323 CD1 mice were infected SC with 105 PFU of the 15561/3x, 15561/E2-12, 15561/E2-82, or 324 15561/2xE2 mutants, wt CHIKV AF15561, and vaccine strain 181/25 to test the attenuation 325 effects of the vaccine-specific substitutions in the background of the parental AF15561 genome. 326 Six mice from each group were sacrificed on days 1-5 and 7 post infection to harvest hind limb 327 and brain tissues and collect blood. The parent AF15561 virus and 15561/3x mutant produced 328 the highest mean viremias over the course of the experiment, which did not differ significantly 329 (P>0.05, one-way ANOVA), reaching 5-6 log10 PFU/ml on days 1 and 2 post infection, and were 330 not cleared from the bloodstream until day 7 (Supplementary Table 1). From day 3 post infection, 331 number of viremic animals began to decline, and on day 5 no detectable viremia was present in 332 mice infected with 181/25 or with the single and double E2 mutants. The mean titers were 333 similar (P>0.05, one-way ANOVA, except for 15561/E2-12 on day 1) and 100-1000-fold lower 334 than in wt strain 15561-infected mice on days 1-4 after infection. 335
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

17
All 6 groups of mice produced similarly high viral loads in hind limb tissue up to day 4 post 336 infection, with peak mean titers of 6-7 log10 PFU/g on day 2 (Supplementary Table 2). On days 5 337 and 7 post infection, there was a decline in titers and number of positive animals. The absence of 338 a difference in viral loads in hind limb tissue of mice infected with the vaccine strain versus the 339 wt CHIKV suggest that this phenotype is not a good indicator of attenuation. 340
Viral loads in the brains of mice infected with wt AF15561, 15561/3x, and 15561/E2-12 341 were the highest and similar to each other (P>0.05, one-way ANOVA) during the time course of 342 the experiment (Supplementary Table 3). In contrast, 181/25- and 15561/2xE2-infected mice 343 produced the lowest titers that did not differ significantly (P>0.05, one-way ANOVA), mostly 344 having none to 2 positive animals out of 6. The 15561/E2-82 mutant generated intermediate viral 345 loads in the brain. 346
In summary, infection of infant CD1 mice with AF15561-derived viruses demonstrated a 347 general trend of 15561/3x mutant exhibiting a phenotype that was similar to wt AF15561 virus, 348 and the 15561/2xE2 mutant resembling that of the 181/25 vaccine strain, with single E2 mutants 349 showing intermediate levels of virulence. 350
351 Infection of A129 mice with CHIKV AF15561 mutants. To study the combined effect of the 352 E2-12 and E2-82 mutations using adult mice, we infected cohorts (n=9) of 5-8-week-old A129 353 mice ID with 104 PFU of the 15561/2xE2, 15561/3x, 15561/E2-12, and 15561/E2-82 mutants, as 354 well as wt CHIKV strain AF15561 and the derived vaccine strain 181/25. All of 15561/2xE2- 355 and vaccine-infected mice survived, but animals infected with wt and 15561/3x began to die on 356 day 4 after infection, and succumbed to infection by day 8 with no difference compared to the wt 357
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

18
strain (P>0.05, Mantel-Cox test) (Fig. 6). The 15561/E2-82-infected mice died by day 9, and the 358 15561/E2-12-infected mice had 56% survival, with the last death on day 9 post infection. 359
Body weights declined gradually in mice infected with wt, 15561/3x, and 15561/E2-82 360 viruses until death, and there was no significant difference between 15561/3x and wt (P>0.05, 361 one-way ANOVA). The 15561/E2-12-infected mice also lost weight until day 9 post infection, 362 then surviving mice began to gain weight (Fig. 7a). The 15561/2xE2- and vaccine-infected mice 363 exhibited a slight weight loss until days 7 and 9, respectively, and then recovered, with no 364 difference compared to the 181/25 strain (P>0.05, one-way ANOVA), except on day 9 post 365 infection, when there was an increase in the mean weight of the 15561/2xE2 cohort. 366
Patterns of body temperature changes were nearly identical to those produced by the LR-367 derived CHIKV strains. In mice that suffered fatal infections, there was slight hyperthermia 368 during the first few days followed by hypothermia preceding death (Fig. 7b). The 15561/2xE2- 369 and 181/25-infected mice had the same pattern (P>0.05, one-way ANOVA) of slight 370 hyperthermia until day 5 post infection as the 181/25 cohort, followed by a return to normal 371 values. 372
The height of the injected footpad was measured on days 2, 4, and 7 post infection. Footpad 373 swelling was more severe on days 2 or 4 for mice infected with wt, 15561/3x, and 15561/E2-82 374 viruses, compared to 181/25-infected mice by one-way ANOVA with Tukey-Kramer post-test 375 (Fig. 7c). Foodpad swelling peaked on day 2 after infection and then gradually decreased in wt- 376 and 15561/3x-infected mice. Generation of virulent revertants (see below) by day 4 post 377 infection might explain the sharp peak in footpad swelling in 15561/E2-82-infected mice. The 378 mean footpad heights of 15561/E2-12- and 15561/2xE2-infected mice were not significantly 379 different (P>0.05) from those of vaccine strain-infected mice on all days tested. 380
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

19
Viremia was evaluated by plaque assay on days 1 and 3 post infection. Most of the infected 381 mice produced viremia, with AF15561 and 15561/3x having the highest titers of about 5 log10 382 PFU/ml on day 1, and about 8 log10 PFU/ml on day 3 (Table 7) that did not differ significantly 383 (P>0.05, one-way ANOVA). Mice infected with the 15561/2xE2 and 181/25 strains had 100-384 1,000-fold lower viremia on days 1 and 3, respectively, with no significant differences (P>0.05, 385 one-way ANOVA). Intermediate levels were observed in mice infected with single E2 mutants, 386 especially on day 3 post infection. 387
In summary, infections of A129 mice with AF15561-derived viruses demonstrated that 388 simultaneous expression of E2 substitutions at amino acid positions 12 and 82 is necessary and 389 sufficient to recapitulate the attenuation phenotype of the 181/25 vaccine strain, as shown by 390 survival of infected mice, body weight and temperature changes, footpad swelling, and viremia. 391
392 Sequencing of 181/25 viral stock and E2 mutants isolates. To confirm that our stock of 181/25 393 vaccine strain corresponded to the sequence published in GenBank, we extracted RNA from 0.2 394 ml of the stock and performed RT-PCR to produce 6 overlapping amplicons covering the full 395 genome of the virus. Direct sequencing of the amplicons revealed no amino acid differences. 396 However, nucleotide position 978 contained a mixture of 181/25-specific T and AF15561-397 specific C, indicating a mixed threonine/isoleucine nsP1-301 population (Table 1). The lack of 398 predominance of the isoleucine residue in the 181/25 population we tested, further suggests that 399 it is not an important attenuation determinant. However, we cannot rule out the possibility that 400 this mutation exerts an effect on the quasispecies population even though it is not present in the 401 majority of CHIKV genomes. Two other nucleotide changes were also found in our 181/25 stock, 402
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

20
both synonymous: R3706→A and T6043→Y. These mutations may have occurred during additional 403 cell culture passages of the 181/25 stock available in our laboratory. 404
Since our investigation indicated the importance of the E2-12 and E2-82 substitutions in the 405 attenuation of strain 181/25, we looked for evidence of reversion in infected mice. Blood 406 samples collected on day 3 post infection from A129 mice injected with 15561/E2-12, 15561/E2-407 82, and 15561/2xE2 viruses were used to extract total RNA and amplify via RT-PCR genome 408 fragments covering both of these mutations. The resultant amplicons were sequenced directly 409 and the presence of revertants was investigated by examining sequence electropherogram peaks. 410 All 8 samples of the 15561/2xE2 mutant showed the presence of both E2 mutations as indicated 411 by mixed nucleotide peaks. Two of the 8 strain 15561/E2-12 blood samples and all of the 9 strain 412 15561/E2-82 samples had partial reversions to the wt C at position 8576, or wt G at position 413 8785, respectively (presence of mixed nucleotide peaks). These data indicate the wt sequences 414 were selected in vivo, partially explaining the highly virulent phenotype of these 2 mutants in the 415 A129 mice model as well as the inadequate attenuation stability suggested by previous human 416 trials of strain 181/25 (8). 417
418 Discussion 419
420 For decades, the generation of live attenuated viral vaccine strains by passaging virulent wt 421
viruses in cell culture has been the mainstay of vaccine development (2, 44). These passages are 422 usually accompanied by the acquisition of several adaptive mutations in the viral genome, with 423 one or a few of these mutations responsible for the attenuated phenotype that is desired for a 424 vaccine strain (12, 18, 49). This cell culture virus passaging procedure was utilized by Levitt et 425
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

21
al. to produce the 181/25 CHIKV vaccine strain to protect against CHIK (21), a debilitating 426 illness that has affected millions of people during recent outbreaks (41). The vaccine exhibited 427 reduced virulence and proved to be highly immunogenic. However, it caused transient arthralgia 428 in 8% of the vaccinees (8). Sequence comparisons of CHIKV strains 181/25 and AF15561, the 429 vaccine’s parent, identified 5 synonymous and 5 nonsynonymous mutations, all in ORFs of the 430 genome (Table 1). 431
We sought to identify the attenuation determinants of strain 181/25 to understand its 432 reactogenicity in humans and to better understand the limitations of attenuation based on cell 433 culture adaptation. Using a reverse genetics approach, we generated an array of CHIKV mutants 434 containing different combinations of 181/25-specific amino acid substitutions in the genetic 435 backbone of two wt CHIKV strains, and tested their level of attenuation in 2 mouse models. 436
The first set of mutants was based on the CHIKV LR genome (Fig. 1b) derived from a 437 French patient returning from La Reunion Island during the 2005 epidemic (45). For a more 438 accurate assessment of the significance of the 181/25 mutations – in their wt genomic 439 background – we also assembled an infectious clone of the vaccine’s parent strain AF15561 (21), 440 and used it to generate the second set of comparable mutants (Fig. 5). The viruses were tested in 441 infant outbred CD1 mice, a model for transient CHIKV infection that resembles human disease 442 histopathologically (56). The mutants, wt and vaccine strains were compared for their generation 443 of viremia and viral loads in hind limb and brain tissues. Body weight change proved to be 444 uninformative for this model. Inbred A129 mice, which lack type I interferon receptors and are 445 sensitive to differences in CHIKV virulence (6, 29, 32), were also used and monitored for 446 survival, body weight and temperature, footpad swelling, and viremia. Strain 181/25 infection 447
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

22
was well tolerated by both mouse strains and ages, and the animals survived with only transient 448 signs of illness (29, 32). 449
We found no evidence that vaccine-specific amino acid substitutions in the nsP1, 6K, or E1 450 genes, when expressed individually or together, mediated attenuation. By all measures, these 451 constructs exhibit virulence comparable to wt viruses, whether expressed in the LR or AF15561 452 genomic background. We also tested another construct with 4 combined 181/25-specific 453 mutations, all except E2-82, and when infected with this mutant, CD1 mice developed viremia 454 similar to wt. Sequencing of our 181/25 vaccine stock, which revealed a ca. 1:1 mixture of the 455 vaccine- and wt-specific nucleotides at the nsP1 position, also suggested that it did not affect 456 virulence. 457
The attenuated phenotype of the 181/25 vaccine strain was recapitulated by the 15561/2xE2 458 virus, which incorporated both E2 glycoprotein substitutions at amino acid positions 12 and 82. 459 CD1 and A129 mice infected with this double mutant exhibited indistinguishable survival, 460 viremia, and all phenotypes tested when compared to the vaccine strain. When these E2 461 mutations were expressed individually, each exhibited an intermediate level of attenuation in 462 most experiments, with the E2-82 mutation demonstrating a higher degree of attenuation than 463 E2-12 overall. Sequencing of virus isolates of 15561/E2-12 and 15561/E2-82 derived from the 464 serum of A129 mice 3 days post infection revealed a high rate of reversion for these single 465 mutants: 9/9 mice had mixed mutant-wt populations in the case of E2-82, and 2/8 in the case of 466 E2-12 virus. The ability of these point mutations to rapidly revert to wt may explain, at least in 467 part, the intermediate/virulent phenotypes observed for these single E2 mutants. In contrast, the 468 mouse isolates of 15561/2xE2 double mutant showed no evidence of any reversion, underscoring 469 the inherent stability advantage of multiple mutations. The stabilizing effect of multiple 470
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

23
mutations in attenuation has been described for several other viruses, including Venezuelan 471 equine encephalitis virus (VEEV) (7, 18), Ross River virus (25), Sindbis virus (SINV) (22), and 472 poliovirus (27). 473
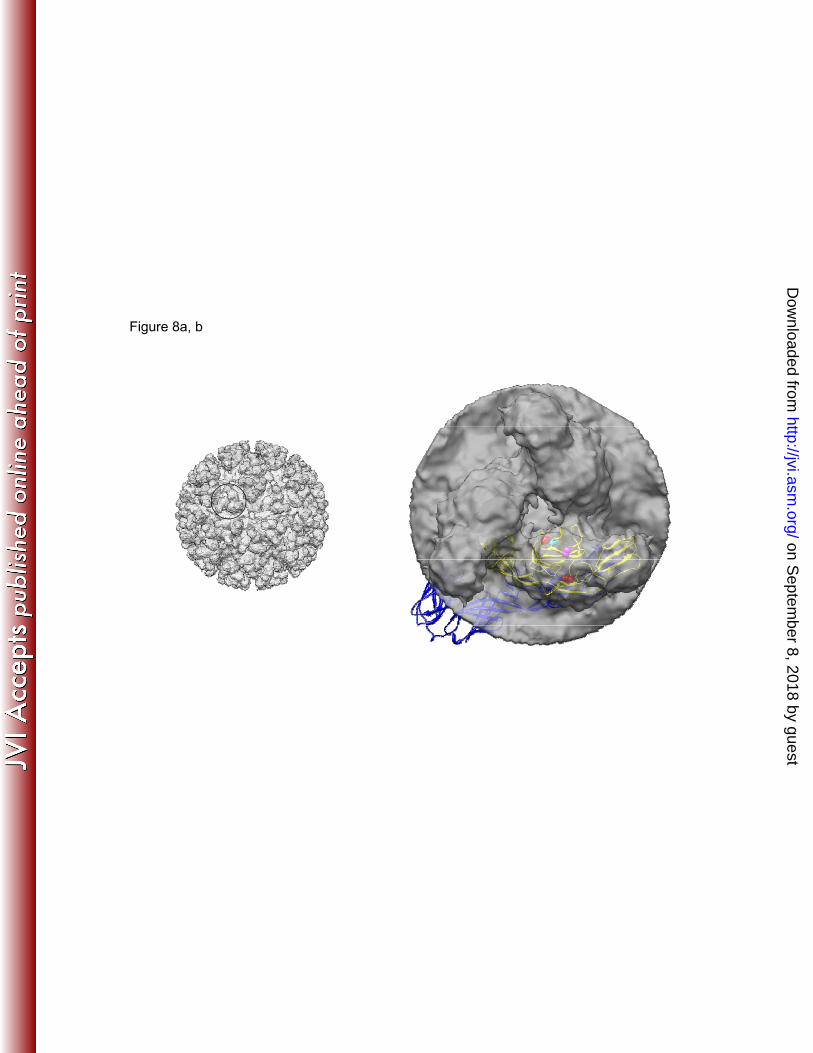
The importance of the Gly→Arg E2-82 mutation in attenuation of CHIKV was not surprising 474 considering findings in other alphaviruses. Cell culture passage typically selects for increased 475 positive charge on the surface of alphavirus spikes comprised principally of the E2 envelope 476 glycoprotein (3, 7, 14, 19, 23). These changes typically increase the binding of alphaviruses to 477 heparan sulfate and other glycosaminoglycans present at high concentrations on the surface of 478 cultured cells. However, they are believed to be attenuating in vivo because they prevent the 479 spread of virus through tissues rich in negatively charged extracellular components. Based on the 480 crystal structure of the mature CHIKV E3-E2-E1 glycoprotein complex (52), amino acid position 481 82 lies on the outer surface of E2 domain A (Fig. 8). Substitutions in the E2 protein of SINV 482 responsible heparan sulfate binding (4, 19) map to the same area, further suggesting that this 483 substitution was selected during cell culture passage for glycosaminoglycan binding. The E2-12 484 position maps to the E2 beta-ribbon and is buried between the E2 domain A and the beta-ribbon 485 (Fig. 8). However, this polar-to-hydrophobic (Thr→Ile) mutation may affect the secondary 486 structure of the beta-ribbon, and thereby affect the nearby E2 domain A, possibly stabilizing the 487 effect of the E2 arginine-82 substitution. These putative glycosaminoglycan binding properties of 488 these mutations should be confirmed experimentally. 489
Our work emphasizes the limitations of the traditional cell culture passaging method for 490 vaccine development, which leads to virus attenuation based on either single or multiple point 491 mutations alone. Due to the high rate of mutagenesis observed in RNA viruses, there is a high 492 rate of reversion to wt sequences, restoring the virulent phenotype. The sequence of a virus 493
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

24
isolate from a viremic human vaccinee who received the 181/25 vaccine, TSI-GSD-218-VR1 494 (GenBank Acc. No. EF452494), showed reversion to wt glycine at position 82 of E2 495 glycoprotein, in addition to other novel mutations. Such instability might explain occasional 496 reactogenicity that was observed in 181/25 vaccine trials (8). A similar situation is observed with 497 the TC-83 live-attenuated VEEV vaccine strain, which also depends only on only 2 point 498 mutations for its attenuation and also causes a high rate of adverse reactions in vaccinees (18, 31). 499
Recent progress in alphaviral genetics has led to the development of more complex and 500 stable approaches for vaccine strain attenuation. Several alternative strategies based on directed 501 manipulation of the alphaviral genome, such as chimeras between two distinct alphaviruses or 502 internal ribosome entry site-dependent translation of structural proteins, have proven to be stably 503 attenuated, safe and effective in recent studies (17, 28, 32, 51, 53, 54). 504
In conclusion, our results indicate that attenuation of the CHIKV 181/25 vaccine strain is 505 mediated by 2 mutations in E2 glycoprotein at amino acid positions 12 and 82. The presence of 506 both mutations is required and sufficient for attaining a well attenuated phenotype. The 507 identification of these 2 attenuating E2 mutations may be useful for the generation of other live-508 attenuated CHIK vaccines. 509
510 Acknowledgements 511
512 We thank Joan Kenney for assistance with statistical analyses. This research was supported 513
by National Institutes of Health grant AI082202. 514 515
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

25
References 516 1. Beaty, B. J., C. H. Calisher, and R. E. Shope. 1989. Arboviruses, p. 797-855. In N. J. 517 Schmidt and R. W. Emmons (ed.), Diagnostic procedures for viral, rickettsial and 518 chlamydial infections, 6th edition. American Public Health Association, Washington, 519 D. C. 520 2. Berge, T. O., I. S. Banks, and W. D. Tigertt. 1961. Attenuation of Venezuelan equine 521 encephalomyelitis virus by in vitro cultivation in guinea pig heart cells. Am. J. Hyg. 522 73:209-218. 523 3. Brault, A. C., A. M. Powers, E. C. Holmes, C. H. Woelk, and S. C. Weaver. 2002. 524 Positively charged amino acid substitutions in the E2 envelope glycoprotein are 525 associated with the emergence of Venezuelan equine encephalitis virus. J. Virol. 526 76:1718-1730. 527 4. Byrnes, A. P., and D. E. Griffin. 2000. Large-plaque mutants of Sindbis virus show 528 reduced binding to heparan sulfate, heightened viremia, and slower clearance from 529 the circulation. Journal of virology 74:644-651. 530 5. Casolari, S., E. Briganti, M. Zanotti, T. Zauli, L. Nicoletti, F. Magurano, C. Fortuna, 531 C. Fiorentini, M. Grazia Ciufolini, and G. Rezza. 2008. A fatal case of encephalitis 532 associated with Chikungunya virus infection. Scand J Infect Dis 40:995-996. 533 6. Couderc, T., F. Chretien, C. Schilte, O. Disson, M. Brigitte, F. Guivel-Benhassine, 534 Y. Touret, G. Barau, N. Cayet, I. Schuffenecker, P. Despres, F. Arenzana-535 Seisdedos, A. Michault, M. L. Albert, and M. Lecuit. 2008. A mouse model for 536 Chikungunya: young age and inefficient type-I interferon signaling are risk factors 537 for severe disease. PLoS Pathog 4:e29. 538 7. Davis, N. L., N. Powell, G. F. Greenwald, L. V. Willis, B. J. Johnson, J. F. Smith, and 539 R. E. Johnston. 1991. Attenuating mutations in the E2 glycoprotein gene of 540 Venezuelan equine encephalitis virus: construction of single and multiple mutants in 541 a full-length cDNA clone. Virology 183:20-31. 542 8. Edelman, R., C. O. Tacket, S. S. Wasserman, S. A. Bodison, J. G. Perry, and J. A. 543 Mangiafico. 2000. Phase II safety and immunogenicity study of live chikungunya 544 virus vaccine TSI-GSD-218. The American journal of tropical medicine and hygiene 545 62:681-685. 546 9. Ganesan, K., A. Diwan, S. K. Shankar, S. B. Desai, G. S. Sainani, and S. M. Katrak. 547 2008. Chikungunya encephalomyeloradiculitis: report of 2 cases with neuroimaging 548 and 1 case with autopsy findings. AJNR Am J Neuroradiol 29:1636-1637. 549 10. Gardner, J., I. Anraku, T. T. Le, T. Larcher, L. Major, P. Roques, W. A. Schroder, S. 550 Higgs, and A. Suhrbier. 2010. Chikungunya virus arthritis in adult wild-type mice. 551 Journal of virology 84:8021-8032. 552 11. Gerardin, P., A. Fianu, D. Malvy, C. Mussard, K. Boussaid, O. Rollot, A. Michault, 553 B. A. Gauzere, G. Breart, and F. Favier. Perceived morbidity and community 554 burden after a Chikungunya outbreak: the TELECHIK survey, a population-based 555 cohort study. BMC Med 9:5. 556 12. Goni, S. E., J. A. Iserte, A. M. Ambrosio, V. Romanowski, P. D. Ghiringhelli, and M. 557 E. Lozano. 2006. Genomic features of attenuated Junin virus vaccine strain 558 candidate. Virus Genes 32:37-41. 559
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

26
13. Grandadam, M., V. Caro, S. Plumet, J. M. Thiberge, Y. Souares, A. B. Failloux, H. J. 560 Tolou, M. Budelot, D. Cosserat, I. Leparc-Goffart, and P. Despres. 2011. 561 Chikungunya virus, southeastern France. Emerging infectious diseases 17:910-913. 562 14. Heil, M. L., A. Albee, J. H. Strauss, and R. J. Kuhn. 2001. An amino acid substitution 563 in the coding region of the E2 glycoprotein adapts Ross River virus to utilize 564 heparan sulfate as an attachment moiety. Journal of virology 75:6303-6309. 565 15. Josseran, L., C. Paquet, A. Zehgnoun, N. Caillere, A. Le Tertre, J. L. Solet, and M. 566 Ledrans. 2006. Chikungunya disease outbreak, Reunion Island. Emerging infectious 567 diseases 12:1994-1995. 568 16. Kariuki Njenga, M., L. Nderitu, J. P. Ledermann, A. Ndirangu, C. H. Logue, C. H. 569 Kelly, R. Sang, K. Sergon, R. Breiman, and A. M. Powers. 2008. Tracking epidemic 570 Chikungunya virus into the Indian Ocean from East Africa. The Journal of general 571 virology 89:2754-2760. 572 17. Kenney, J. L., S. M. Volk, J. Pandya, E. Wang, X. Liang, and S. C. Weaver. 2011. 573 Stability of RNA virus attenuation approaches. Vaccine 29:2230-2234. 574 18. Kinney, R. M., G. J. Chang, K. R. Tsuchiya, J. M. Sneider, J. T. Roehrig, T. M. 575 Woodward, and D. W. Trent. 1993. Attenuation of Venezuelan equine encephalitis 576 virus strain TC-83 is encoded by the 5'-noncoding region and the E2 envelope 577 glycoprotein. Journal of virology 67:1269-1277. 578 19. Klimstra, W. B., K. D. Ryman, and R. E. Johnston. 1998. Adaptation of Sindbis 579 virus to BHK cells selects for use of heparan sulfate as an attachment receptor. J. 580 Virol. 72:7357-7366. 581 20. Kuhn, R. J. 2007. Togaviridae: The viruses and their replication, p. 1001-1022. In D. 582 M. Knipe and P. M. Howley (ed.), Fields' Virology, Fifth Edition. Lippincott, Williams 583 and Wilkins, New York. 584 21. Levitt, N. H., H. H. Ramsburg, S. E. Hasty, P. M. Repik, F. E. Cole, and H. W. 585 Lupton. 1986. Development of an attenuated strain of chikungunya virus for use in 586 vaccine production. Vaccine 4:157-162. 587 22. Lustig, S., A. C. Jackson, C. S. Hahn, D. E. Griffin, E. G. Strauss, and J. H. Strauss. 588 1988. Molecular basis of Sindbis virus neurovirulence in mice. J Virol 62:2329-2336. 589 23. Martin, D. H., W. H. Dietz, O. Alvaerez, Jr., and K. M. Johnson. 1982. 590 Epidemiological significance of Venezuelan equine encephalomyelitis virus in vitro 591 markers. Am. J. Trop. Med. Hyg. 31:561-568. 592 24. Mavalankar, D., P. Shastri, T. Bandyopadhyay, J. Parmar, and K. V. Ramani. 593 2008. Increased mortality rate associated with chikungunya epidemic, ahmedabad, 594 India. Emerging infectious diseases 14:412-415. 595 25. Meek, A. D., S. G. Faragher, R. C. Weir, and L. Dalgarno. 1989. Genetic and 596 phenotypic studies on Ross River virus variants of enhanced virulence selected 597 during mouse passage. Virology 172:399-407. 598 26. Morrison, T. E., L. Oko, S. A. Montgomery, A. C. Whitmore, A. R. Lotstein, B. M. 599 Gunn, S. A. Elmore, and M. T. Heise. 2011. A mouse model of chikungunya virus-600 induced musculoskeletal inflammatory disease: evidence of arthritis, tenosynovitis, 601 myositis, and persistence. Am J Pathol 178:32-40. 602 27. Omata, T., M. Kohara, S. Kuge, T. Komatsu, S. Abe, B. L. Semler, A. Kameda, H. 603 Itch, M. Arita, E. Wimmer, and A. Nomoto. 1986. Genetic analysis of the 604 attenuation phenotype of poliovirus type 1. J. Virol. 58:348-358. 605
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

27
28. Paessler, S., R. Z. Fayzulin, M. Anishchenko, I. P. Greene, S. C. Weaver, and I. 606 Frolov. 2003. Recombinant Sindbis/Venezuelan equine encephalitis virus is highly 607 attenuated and immunogenic. Journal of virology 77:9278-9286. 608 29. Partidos, C. D., J. Weger, J. Brewoo, R. Seymour, E. M. Borland, J. P. Ledermann, 609 A. M. Powers, S. C. Weaver, D. T. Stinchcomb, and J. E. Osorio. 2011. Probing the 610 attenuation and protective efficacy of a candidate chikungunya virus vaccine in mice 611 with compromised interferon (IFN) signaling. Vaccine 29:3067-3073. 612 30. Pettersen, E. F., T. D. Goddard, C. C. Huang, G. S. Couch, D. M. Greenblatt, E. C. 613 Meng, and T. E. Ferrin. 2004. UCSF Chimera--a visualization system for exploratory 614 research and analysis. J Comput Chem 25:1605-1612. 615 31. Pittman, P. R., R. S. Makuch, J. A. Mangiafico, T. L. Cannon, P. H. Gibbs, and C. J. 616 Peters. 1996. Long-term duration of detectable neutralizing antibodies after 617 administration of live-attenuated VEE vaccine and following booster vaccination 618 with inactivated VEE vaccine. Vaccine 14:337-343. 619 32. Plante, K., E. Wang, C. D. Partidos, J. Weger, R. Gorchakov, K. Tsetsarkin, E. M. 620 Borland, A. M. Powers, R. Seymour, D. T. Stinchcomb, J. E. Osorio, I. Frolov, and 621 S. C. Weaver. 2011. Novel Chikungunya Vaccine Candidate with an IRES-Based 622 Attenuation and Host Range Alteration Mechanism. PLoS Pathog 7:e1002142. 623 33. Powers, A. M., and C. H. Logue. 2007. Changing patterns of chikungunya virus: re-624 emergence of a zoonotic arbovirus. The Journal of general virology 88:2363-2377. 625 34. Rezza, G., L. Nicoletti, R. Angelini, R. Romi, A. C. Finarelli, M. Panning, P. 626 Cordioli, C. Fortuna, S. Boros, F. Magurano, G. Silvi, P. Angelini, M. Dottori, M. G. 627 Ciufolini, G. C. Majori, and A. Cassone. 2007. Infection with chikungunya virus in 628 Italy: an outbreak in a temperate region. Lancet 370:1840-1846. 629 35. Roehrig, J. T. 1993. Immunogens of encephalitis viruses. Vet Microbiol 37:273-284. 630 36. Sambrook, J., and D. Russell. 2001. Molecular Cloning: A Laboratory Manual, 3rd 631 Edition. Cold Spring Harbor Press, Cold Spring Harbor, N.Y. 632 37. Schuffenecker, I., I. Iteman, A. Michault, S. Murri, L. Frangeul, M. C. Vaney, R. 633 Lavenir, N. Pardigon, J. M. Reynes, F. Pettinelli, L. Biscornet, L. Diancourt, S. 634 Michel, S. Duquerroy, G. Guigon, M. P. Frenkiel, A. C. Brehin, N. Cubito, P. 635 Despres, F. Kunst, F. A. Rey, H. Zeller, and S. Brisse. 2006. Genome 636 microevolution of chikungunya viruses causing the Indian Ocean outbreak. PLoS 637 Med 3:e263. 638 38. Schwartz, O., and M. L. Albert. 2010. Biology and pathogenesis of chikungunya 639 virus. Nat Rev Microbiol 8:491-500. 640 39. Sherman, M. B., and S. C. Weaver. 2010. Structure of the recombinant alphavirus 641 Western equine encephalitis virus revealed by cryoelectron microscopy. Journal of 642 virology 84:9775-9782. 643 40. Shu, P. Y., C. F. Yang, C. L. Su, C. Y. Chen, S. F. Chang, K. H. Tsai, C. H. Cheng, and J. 644 H. Huang. 2008. Two imported chikungunya cases, Taiwan. Emerging infectious 645 diseases 14:1326-1327. 646 41. Simon, F., E. Javelle, M. Oliver, I. Leparc-Goffart, and C. Marimoutou. 2011. 647 Chikungunya virus infection. Curr Infect Dis Rep 13:218-228. 648 42. Soon, Y. Y., I. Junaidi, V. Kumarasamy, Y. K. Chem, R. Juliana, and K. B. Chua. 649 2007. Chikungunya virus of Central/East African genotype detected in Malaysia. 650 Med J Malaysia 62:214-217. 651
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

28
43. Tandale, B. V., P. S. Sathe, V. A. Arankalle, R. S. Wadia, R. Kulkarni, S. V. Shah, S. 652 K. Shah, J. K. Sheth, A. B. Sudeep, A. S. Tripathy, and A. C. Mishra. 2009. Systemic 653 involvements and fatalities during Chikungunya epidemic in India, 2006. J Clin Virol. 654 44. Theiler, M., and H. H. Smith. 1937. Use of yellow fever virus modified by in vitro 655 cultivation for human immunization. J. Exp. Med. 65:787-800. 656 45. Tsetsarkin, K., S. Higgs, C. E. McGee, X. De Lamballerie, R. N. Charrel, and D. L. 657 Vanlandingham. 2006. Infectious clones of Chikungunya virus (La Reunion isolate) 658 for vector competence studies. Vector Borne Zoonotic Dis 6:325-337. 659 46. Tsetsarkin, K. A., R. Chen, G. Leal, N. Forrester, S. Higgs, J. Huang, and S. C. 660 Weaver. 2011. Chikungunya virus emergence is constrained in Asia by lineage-661 specific adaptive landscapes. Proceedings of the National Academy of Sciences of the 662 United States of America 108:7872-7877. 663 47. Tsetsarkin, K. A., R. Chen, M. Sherman, and S. C. Weaver. 2011. Chikungunya 664 Virus: Evolution and Genetic Determinants of Emergence. Curr Opin Virol:in press. 665 48. Tsetsarkin, K. A., C. E. McGee, S. M. Volk, D. L. Vanlandingham, S. C. Weaver, and 666 S. Higgs. 2009. Epistatic roles of E2 glycoprotein mutations in adaption of 667 chikungunya virus to aedes albopictus and ae. Aegypti mosquitoes. PLoS One 668 4:e6835. 669 49. Vialat, P., R. Muller, T. H. Vu, C. Prehaud, and M. Bouloy. 1997. Mapping of the 670 mutations present in the genome of the Rift Valley fever virus attenuated MP12 671 strain and their putative role in attenuation. Virus research 52:43-50. 672 50. Volk, S. M., R. Chen, K. A. Tsetsarkin, A. P. Adams, T. I. Garcia, A. A. Sall, F. Nasar, 673 A. J. Schuh, E. C. Holmes, S. Higgs, P. D. Maharaj, A. C. Brault, and S. C. Weaver. 674 2010. Genome-scale phylogenetic analyses of chikungunya virus reveal independent 675 emergences of recent epidemics and various evolutionary rates. Journal of virology 676 84:6497-6504. 677 51. Volkova, E., E. Frolova, J. R. Darwin, N. L. Forrester, S. C. Weaver, and I. Frolov. 678 2008. IRES-dependent replication of Venezuelan equine encephalitis virus makes it 679 highly attenuated and incapable of replicating in mosquito cells. Virology 377:160-680 169. 681 52. Voss, J. E., M. C. Vaney, S. Duquerroy, C. Vonrhein, C. Girard-Blanc, E. Crublet, A. 682 Thompson, G. Bricogne, and F. A. Rey. 2010. Glycoprotein organization of 683 Chikungunya virus particles revealed by X-ray crystallography. Nature 468:709-712. 684 53. Wang, E., O. Petrakova, A. P. Adams, P. V. Aguilar, W. Kang, S. Paessler, S. M. 685 Volk, I. Frolov, and S. C. Weaver. 2007. Chimeric Sindbis/eastern equine 686 encephalitis vaccine candidates are highly attenuated and immunogenic in mice. 687 Vaccine 25:7573-7581. 688 54. Wang, E., E. Volkova, A. P. Adams, N. Forrester, S. Y. Xiao, I. Frolov, and S. C. 689 Weaver. 2008. Chimeric alphavirus vaccine candidates for chikungunya. Vaccine 690 26:5030-5039. 691 55. Weaver, S. C., and W. K. Reisen. 2009. Present and future arboviral threats. 692 Antiviral Res 85:328-345. 693 56. Ziegler, S. A., L. Lu, A. P. da Rosa, S. Y. Xiao, and R. B. Tesh. 2008. An animal 694 model for studying the pathogenesis of chikungunya virus infection. The American 695 journal of tropical medicine and hygiene 79:133-139. 696 697
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

29
698 699
Table 1. Mutations accompanying the generation of CHIKV vaccine strain 181/clone 25 (AKA
TSI-GSD-218) from its parent strain AF15561
Gene Nucleotide position Amino acid position Amino acid substitution
nsP1 978 301 Thr→Ile
nsP2 3706*
nsP4 7259*
E3 8463*
E2 8576 12 Thr→Ile
8760*
8785 82 Gly→Arg
6K 9935 42 Cys→Phe
E1 10446
11204 404 Ala→Val
*synonymous mutations 700 701
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
30
Table 2. Viremia in 6-7-day-old CD1 mice after subcutaneous infection with CHIKV strain LR,
mutants, or vaccine strain 181/25
Day 2 post-infection Day 4 post-infection
Virus strain* No. animals positive/total inoculated
Mean titer** of positive animals ± SD
No. animals positive/total inoculated
Mean titer** of positive animals ± SD
LR 3/3 2.9±0.6 1/3 1.3±0.6
LR/nsP1 2/2 2.2±0.2 0/2 ≤ 0.9
LR/6K 3/3 5.0±0.2 3/3 2.7±0.8
LR/E1 3/3 2.5±0.5 0/3 ≤ 0.9
LR/E2-12 2/3 2.0±1.1 0/3 ≤ 0.9
LR/E2-82 1/2 1.6±1.0 1/2 1.5±0.8
181/25 0/2 ≤ 0.9 0/2 ≤ 0.9
*See Fig. 1b for strain designations 702 **Limit of detection of the assay was 0.9 Log10 PFU/ml serum 703
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
31
704 Table 3. Viral load in hind limb tissue in 6-7-day-old CD1 mice after subcutaneous infection with CHIKV strain LR, mutants, or
vaccine strain 181/25
Day 2 post-infection Day 4 post-infection Day 6 post-infection Day 8 post-infection
Virus strain*
No. animals positive/total inoculated
Mean titer** of positive animals ± SD
No. animals positive/total inoculated
Mean titer** of positive animals ± SD
No. animals positive/total inoculated
Mean titer** of positive animals ± SD
No. animals positive/total inoculated
Mean titer** of positive animals ± SD
LR 2/3 3.3±2.2 2/3 4.3±3.0 2/3 2.3±1.2 1/3 2.1±2.0
LR/nsP1 2/2 3.1±0.1 1/2 2.6±2.3 0/2 ≤ 0.9 0/2 ≤ 0.9
LR/6K 3/3 5.0±1.4 3/3 6.8±0.2 3/3 6.9±0.9 1/3 2.3±2.5
LR/E1 3/3 4.3±1.3 3/3 5.8±1.1 1/3 1.5±1.1 0/3 ≤ 0.9
LR/E2-12 3/3 4.3±1.0 2/3 3.5±2.4 1/3 1.9±1.8 0/3 ≤ 0.9
LR/E2-82 0/2 ≤ 0.9 0/2 ≤ 0.9 0/2 ≤ 0.9 0/2 ≤ 0.9
181/25 1/2 3.7±4.0 2/2 5.2±1.2 2/2 4.2±0.4 1/2 3.1±3.0
*See Fig. 1b for strain designations 705 **Limit of detection of the assay was 0.9 Log10 PFU/g tissue 706 707
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
32
Table 4. Viral load in brain tissue in 6-7-day-old CD1 mice after subcutaneous infection with CHIKV strain LR, mutants, or vaccine
strain 181/25
Day 2 post-infection Day 4 post-infection Day 6 post-infection Day 8 post-infection
Virus strain*
No. animals positive/total inoculated
Mean titer** of positive animals ± SD
No. animals positive/total inoculated
Mean titer** of positive animals ± SD
No. animals positive/total inoculated
Mean titer** of positive animals ± SD
No. animals positive/total inoculated
Mean titer** of positive animals ± SD
LR 3/3 3.9±1.3 2/3 2.3±1.3 2/3 3.1±2.1 0/3 ≤ 0.9
LR/nsP1 1/2 2.3±2.0 0/2 ≤ 0.9 1/2 2.8±2.6 1/2 2.2±1.8
LR/6K 3/3 4.2±0.4 2/3 3.9±2.8 3/3 4.3±1.2 2/3 3.7±2.6
LR/E1 2/3 3.0±1.8 2/3 4.6±3.3 2/3 2.4±1.3 1/3 1.7±1.3
LR/E2-12 0/3 ≤ 0.9 1/3 1.8±1.6 0/3 ≤ 0.9 0/3 ≤ 0.9
LR/E2-82 0/2 ≤ 0.9 0/2 ≤ 0.9 0/2 ≤ 0.9 0/2 ≤ 0.9
181/25 0/2 ≤ 0.9 1/2 3.7±3.9 0/2 ≤ 0.9 0/2 ≤ 0.9
*See Fig. 1b for strain designations 708 **Limit of detection of the assay was 0.9 Log10 PFU/g tissue 709 710
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
33
Table 5. Viremia in 5-6-day-old CD1 mice after subcutaneous infection with CHIKV strain LR, mutant, or vaccine strain 181/25
Day 1 post-infection Day 2 post-infection Day 3 post-infection Day 4 post-infection
Virus strain*
No. animals positive/total inoculated
Mean titer** of positive animals ± SD
No. animals positive/total inoculated
Mean titer** of positive animals ± SD
No. animals positive/total inoculated
Mean titer** of positive animals ± SD
No. animals positive/total inoculated
Mean titer** of positive animals ± SD
LR 3/4 3.3±2.0 4/4 3.7±0.9 4/4 2.6±1.0 2/4 2.2±1.6
LR/4x 4/4 4.1±1.0 4/4 3.2±0.4 4/4 3.7±1.5 2/4 1.9±1.2
181/25 3/3 4.3±0.2 1/3 1.5±1.0 0/3 ≤ 0.9 1/3 1.5±1.0
*See Fig. 1b for strain designations 711 **Limit of detection of the assay was 0.9 Log10 PFU/ml serum 712 713 on S
eptember 8, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
34
Table 6. Viremia in 10-week-old A129 mice after intradermal infection with CHIKV strain LR mutants or vaccine strain 181/25
Day 1 post-infection Day 2 post-infection Day 3 post-infection Day 4 post-infection
Virus strain*
No. animals positive/total inoculated
Mean titer** of positive animals ± SD
No. animals positive/total inoculated
Mean titer** of positive animals ± SD
No. animals positive/total inoculated
Mean titer** of positive animals ± SD
No. animals positive/total inoculated
Mean titer** of positive animals ± SD
LR/nsP1 5/5 2.9±0.8 5/5 5.2±0.3 4/4 6.7±0.4 NS NS
LR/E1 5/5 4.1±0.2 5/5 6.3±1.0 2/2 6.8±0.2 NS NS
LR/E2-12 5/5 2.5±0.7 5/5 4.2±0.6 5/5 4.7±0.8 5/5 4.6±0.4
LR/E2-82 2/5 0.2±0.1 5/5 2.4±0.4 5/5 3.1±0.4 5/5 3.8±0.2
181/25 4/4 0.5±0.4 4/4 3.3±0.4 4/4 2.9±0.4 4/4 2.6±0.2
*See Fig. 1b for strain designations 714 **Limit of detection of the assay was 0.1 Log10 PFU/ml serum 715 NS – no survivors 716 717
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
35
Table 7. Viremia in 5-8-week-old A129 mice after intradermal infection with CHIKV strain
AF15561, mutants, or vaccine strain 181/25
Day 1 post-infection Day 3 post-infection
Virus strain* No. animals positive/total inoculated
Mean titer** of positive animals ± SD
No. animals positive
Mean titer** of positive animals ± SD
15561 8/9 5.0±0.8 9/9 8.2±1.0
15561/3x 7/9 4.6±2.2 9/9 7.7±0.9
15561/E2-82 8/9 3.3±1.1 9/9 7.0±0.8
15561/E2-12 8/9 2.9±0.9 9/9 6.5±1.3
15561/2xE2 7/9 2.8±1.2 8/9 4.9±1.5
181/25 7/9 2.5±1.0 9/9 5.0±0.5
*See Fig. 5 for strain designations 718 Limit of detection of the assay was 0.9 Log10 PFU/ml serum 719 720 721 722
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

36
Figure legends 723 724 Fig. 1. Schematic representation of the CHIKV genome. (a) Positions of nonsynonymous 725
mutations in the vaccine strain 181/25. (b) Mutations cloned in CHIKV La Reunion genomic 726 backbone. Numbers correspond to amino acid positions of the mutations in the respective viral 727 protein. SG – subgenomic. 728
729 Fig. 2. Mean body weight in 6-7-day-old CD1 mice (n=10) after SC infection with 105 PFU 730
of CHIKV strain LR, mutants, or vaccine strain 181/25. 731 732 Fig. 3. Survival of 8-10-week-old A129 mice (n=4-5) after ID infection with 104 PFU of 733
CHIKV strain LR mutants or vaccine strain 181/25. Animals were monitored for 18 days. 734 735 Fig. 4. Morbidity in 8-10-week-old A129 mice (n=4-5) after ID infection with 104 PFU of 736
CHIKV strain LR mutants or vaccine strain 181/25. (a) Body weight change. (b) Body 737 temperature change. (c) Footpad swelling at day 2 post infection. Asterisks indicate P values by 738 one-way ANOVA with Tukey-Kramer post-test, compared to 181/25 or PBS control groups, 739 *P<0.05, **P<0.01. 740
741 Fig. 5. Schematic representation of CHIKV AF15561 genomes with cloned mutations. 742
Numbers correspond to amino acid positions of the mutations in the respective viral protein. 743 744
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

37
Fig. 6. Survival of 5-8-week-old A129 mice (n=9) after ID infection with 104 PFU of CHIKV 745 strain AF15561, mutants, or vaccine strain 181/25. Animals were monitored for 14 days. 746
747 Fig. 7. Morbidity in 5-8-week-old A129 mice (n=9) after ID infection with 104 PFU of 748
CHIKV strain AF15561, mutants, or vaccine strain 181/25. (a) Body weight change. (b) Body 749 temperature change. (c) Footpad swelling. *** P<0.001 by one-way ANOVA with Tukey-750 Kramer post-test, compared to 181/25 group. 751
752 Fig. 8. Crystal structure of the mature envelope glycoprotein complex of CHIKV fitted into a 753
cryo-electron microscopy (cryoEM) reconstruction map of Western equine encephalitis virus 754 (WEEV) glycoprotein spikes on the surface of the viral particle (39), prepared using UCSF 755 Chimera software (30). (a) CryoEM map of WEEV. (b) Top view of the three E2-E1 756 heterodimers forming the spike, magnified from a circle in panel a, with CHIKV E2-E1 crystal 757 structure fitted into one of the heterodimers. (c) Same spike with CHIKV crystal structure as in 758 panel b, rotated 90 degrees. (d) Ribbon diagram of the CHIKV heterodimer (MMDB ID: 86612, 759 chains B and F (52)) in the same orientation as in panel b. E2 is shown in yellow, E1 is in blue. 760 E2 amino acid positions T12 and G82 are highlighted as red spheres. E2 positions 76 and 114 are 761 residues described in the literature as responsible for heparan sulfate binding for SINV (4, 19) 762 are shown as magenta and cyan spheres, respectively. 763 764
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
Figure 1a
Th Il Thr > Ile Al V lThr > Ile Thr > Ile
Cys > Phe
Ala > ValGly > Arg
y
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

Figure 1b T301 → I
LR/nsP1
LR/E2-12
T12 → I
LR/E2-82
G82 → R
LR/6K
A404 → V
C42 → F
LR/6K
T301 > I A404 → V
LR/E1
301 404
T12 → I C42 → F
LR/4x
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
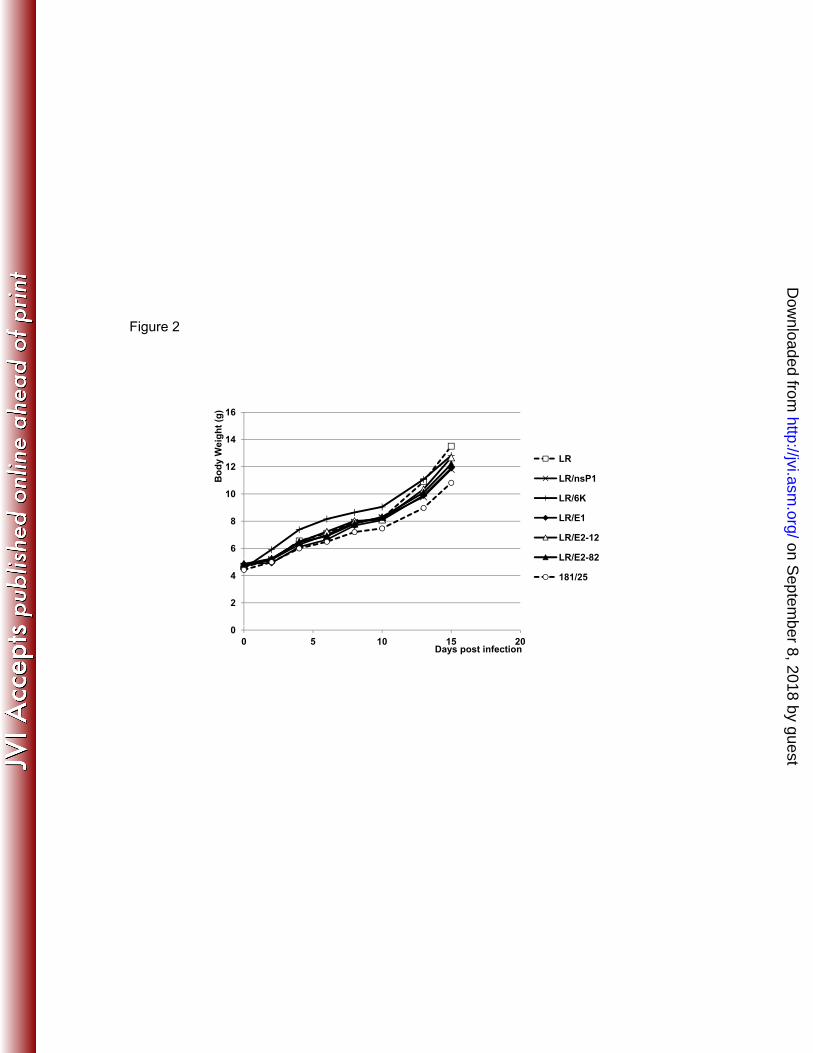
Figure 2
16
t (g)
10
12
14
Bod
y W
eigh
t
LR
LR/nsP1
6
8
10 LR/6K
LR/E1
LR/E2-12
LR/E2-82
2
4
LR/E2 82
181/25
00 5 10 15 20
Days post infection
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
Figure 3
80
100
% S
urvi
val
LR
LR/nsP1
20
40
60 LR/E1
LR/E2-12
LR/E2-82
181/25
00 1 2 3 4 5 6 7 8 9 10
Days post infection
181/25
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
Figure 4a
15
0
5
10
15
Wei
ght C
hang
e
LR
15
-10
-5
0
% B
ody
W LRLR/nsP1LR/E1LR/E2-12LR/E2-82
-25
-20
-15
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18Days post infection
181/25
Days post infection
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
Figure 4b
40
45
mpe
ratu
re (O
C)
LR
30
35Tem LR/nsP1
LR/E1
LR/E2-12
LR/E2-82
20
25
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
LR/E2 82
181/25
Days post infection
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
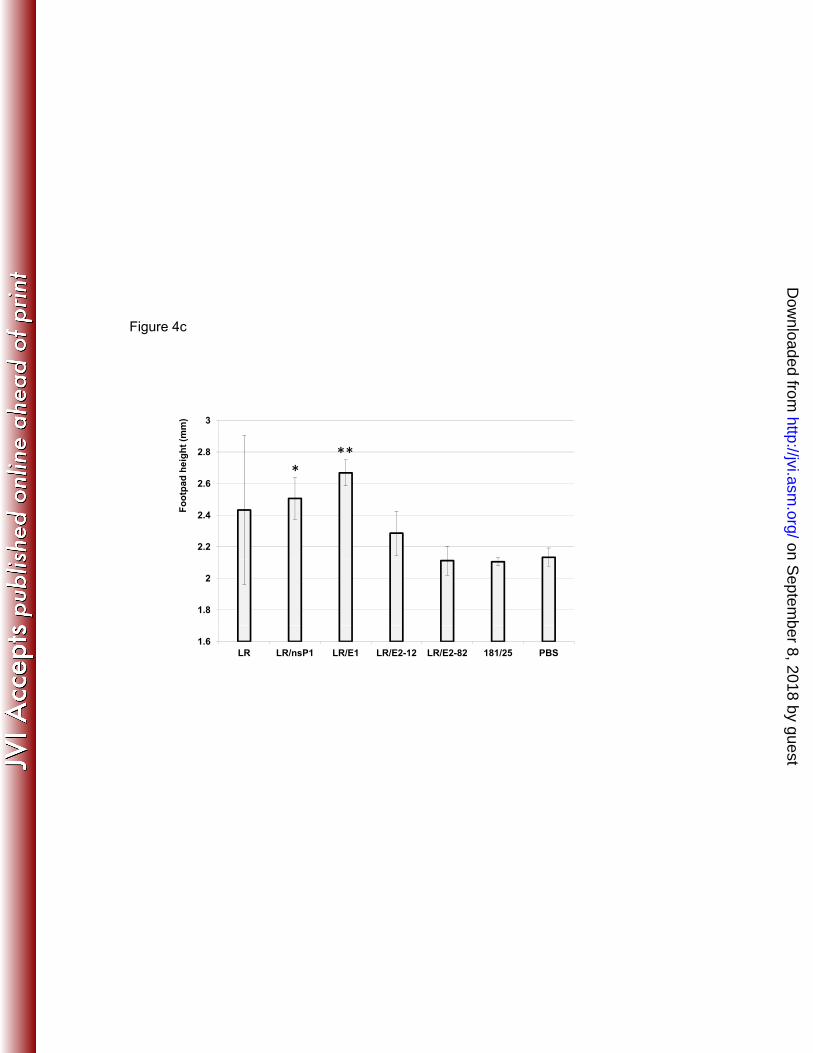
Figure 4c
3
m)
2.6
2.8
tpad
hei
ght (
mm
***
2.2
2.4Foot
1.8
2
1.6LR LR/nsP1 LR/E1 LR/E2-12 LR/E2-82 181/25 PBS
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

Figure 5
T12 → I
15561/E2-12
12
15561/E2-82
G82 → R
15561/2xE2
T12 → I G82 → R
T301 → I A404 → V
15561/3x
C42 → F
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
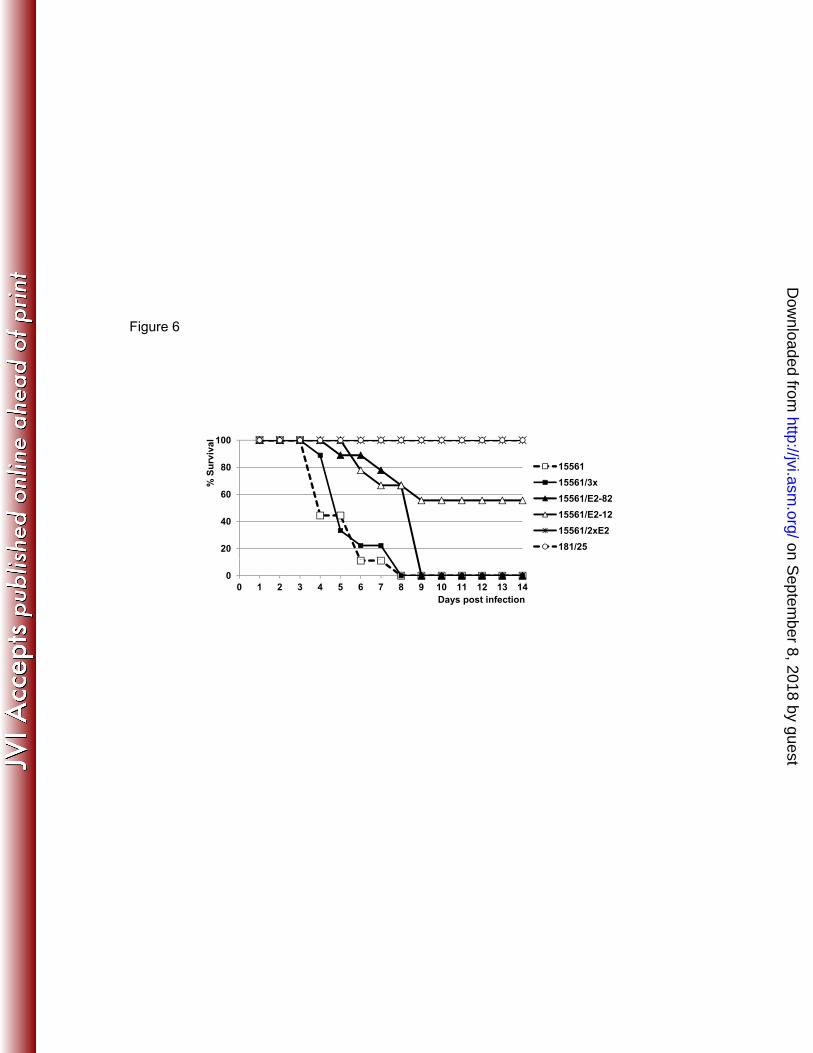
Figure 6
60
80
100
% S
urvi
val
1556115561/3x
20
40
60 15561/E2-8215561/E2-1215561/2xE2181/25
00 1 2 3 4 5 6 7 8 9 10 11 12 13 14
Days post infection
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
Figure 7a
5
10
ange
10
-5
0
5
ody
Wei
ght C
ha
15561
15561/3x
15561/E2-82
25
-20
-15
-10
% B
o
15561/E2-12
15561/2xE2
181/25
-30
-25
0 2 4 6 8 10 12 14Days post infection
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

Figure 7b
42 0)
33 0
36.0
39.0
42.0
mpe
ratu
re (O
C)
15561
15561/3x
24 0
27.0
30.0
33.0
Te 15561/E2-82
15561/E2-12
15561/2xE2
18.0
21.0
24.0
0 2 4 6 8Days post infection
181/25
ays post ect o
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
Figure 7c
3.5
4.0
heig
ht (m
m)
2 days p.i.4 days p.i.7 days p.i.
***
3.0Foot
pad
****** ***
2.0
2.5
1.515561 15561/3x 15561/E2-82 15561/E2-12 15561/2xE2 181/25
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
Figure 8c, d
E2-G82 E2-G114
E2-T12
E2-A76
on Septem
ber 8, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from