Embed Size (px)
Citation preview

GLUCAGON-LIKE PEPTIDE3 (GLP-2) RECEPTOR EXPRESSION IN THE CENTRAL NERVOUS SYSTEM AND GASTROINTESTINAL TRACT.
Julie A. Lovshin
A thesis submined in conformity with the requirements
for the degree of Doctor of Philosophy.
Graduate Department of the Institute of Medical Science
University of Toronto.
O Copyright by Julie A. Lovshin 200 1.

National Library Bibliolhéque nationaie du Canada
Acquisitions and Acquisitions et Bibliographie Services services bibliographiques
395 Wellington Sbeet 395. rue Weliingtm ORawa ON K1A ON4 ORawa ON KI A ON4 Canada Canada
The author has granted a non- L'auteur a accordé une licence non exclusive licence aiiowing the exclusive permettant à la National Library of Canada to Bibliothèque nationale du Canada de reproduce, loan, distn'bute or seil reproduire, prêter, distribuer ou copies of this thesis in microform, vendre des copies de cette thèse sous paper or electronic formats. la forme de microfiche/fïh, de
reproduction sur papier ou sur format électronique.
The author retains ownership of the L'auteur conserve la propriété du copyright in this thesis. Neither the droit d'auteur qui protège cette thèse. thesis nor substantial extracts fiom it Ni la thèse ni des extraits substantiels may be printed or otherwise de celle-ci ne doivent être imprimés reproduced without the author's ou autrement reproduits sans son permission. autorisation.

ABSTRACT
Glucagon-like Peptide-2 (GLP-2) Receptor Expression in the Central Nervous System and the Gastrointestinal Tract.
Doctor of Philosophy 200 1 Julie A. Lovshin
Graduate Department of the ht i tute of Medical Science University of Toronto
The gut-brain hormones, glucagon-like peptides- 1 (GLP-1) and -3 (GLP-2) are
synthesized from proglucagon in enteroendocrine ce& and neurons, are secreted from the gut
in response to nutrient ingestion and stimulate diverse actions to regulate energy
homeostasis. GLP-1 and GLP-2 transduce their action through cognate membrane bound G-
protein coupled receptors. GLP-2 induces trophic, anti-apoptotic and transit-modulating
events in the put, but GLP-2 action in the brain remains unexplored. To identiQ the
components of the GLP-UGLP-2 receptor axis in the CNS we undertook a combinatorial
approach using 1) RT-PCR, 2) immunocytochemistry, and 3) transgenic models to study the
expression and fùnction of this avis in the bmin in vivo. We studied the developing intestine
and CNS to determine whether this axis was present and functiond during periods of rapid
growth and tissue development.
We have described the spatial distribution of G L P - receptor expression throughout the
adult rodent CNS, and identified 5'-regdatory sequences that confer tissue- and ceil-specific
expression to reporter genes in the Ois. We studied the centrd actions of GLP-2 on food
intake, and demonstrated the satiating effects of a potent GLP-2 analogue, ~ [G~~ ' ] -GLP-~ .
We exarnined the inter-relationship between centrai GLP-1 and GLP-2 responsive systerns,
and discovered functional interdependence of GLP-[Rand GLP-2R signaling in the brain. In
the neonatai rat intestine we detected substantid increases in the relative expression of GLP-

2 and the GLP-2 receptor during gut development and demonstrated that GLP-2 is coupled to
intestinal growth in the neonare. Both GLP-2 and GLP-2 receptor expression were detected
in Létal and neonatal brain, and we demonstrated that cells isolated from the neonatal rat
brainstem respond to GLP-2.
Taken together, these studies demonstrate the Functional expression of the GLP-UGLP-2
receptor avis in the brain. Our finding of integrated GLP-1 and GLP-2 responsive neuronal
systems extends cwrent concepts of centrai GLP- 1 and GLP-2 action and suggests new
strategies for manipuiating the actions of these hvo important regulators of energy
homeostasis in the brain.

CO-AUTHORSHIP
Julie A. Lovshin did the majority of the research presented in this thesis. Experiments
presented in this thesis that were camed out in collaboration with other individuals are noted
in the Merhods section of each chapter. The following thesis contains work published
previously in manuscripts that were CO-authored by Dr. Daniel 1. Drucker, Dr. Patricia
Brubaker, Dr. Theodore Brown, Dr. Bernardo Yusta, Jennifer Estall, Ilias Iliopoulos, Anoush
Migirdicyan, and Liliane Dableh. Versions of original manuscripts that were written by Julie
A. Lovshin and Daniel .i. Drucker appear in Chôpters 1,2,3 of this thesis. Copyright
releases of manuscripts were obtained from publishers and are presented in appendices 1,2,
3, and 4 of this thesis.

ACKNOWLEDGEMENTS
Carrying out a PhD. requires not onIy persona1 dedication and effort, but also substantial
support, sacrifice and understanding h m others. i have been fortunate enough to receive a
wealth of support fiom a number of individuals. Without their combined individual
contributions the h i t ion of this thesis would not have been possible.
i would first like to thank my wondehl husband Dr. Kylen McReelis. In addition to your
continual inspiration and constant suive for exceilence, i would like to thank you for
graciously supporting my endless pursuits in the laboratory. For offsetting disappointment
and setbacks with perpetual optimism, for being my sounding board over the years, and for
always providing me with a smile at the end of the day.
Many thanks to members of my fmily incIuding Brian, Brenden, Stephen, Shona, Helen,
Gerry, Sam and Barb who have al1 supported my efforts; with special thanks to my parents
Albert and Louise.
In addition to academic excellence and scientific exploration, the Drucker laboratory has
provided me with the opportunity to work with a number of talented individuals who 1 am
eratehl to for their heIp, expertise and Friendship. Special thanks to al1 rny colleagues in the - Drucker Laboratory includmg Dr. Louise Scrocchi, Mary Brown, Feng Wang, Dr. Bernardo
Yusta, Dr. Robin Boushey, Dr. Min Nian, and especially to Dr. Laurie Baggio. Best wishes
to Tanya Hansotia, Jemifer Esta11 and Dr. William Harper with their studies in the Drucker
Lab.
1 wouId also like to thank the members of my pro_mrn cornmittee who have taught me
how to criticalIy evaluate my research over the years. Special th& to Dr. David Irwin for
his expertise in sequence analysis and his heIp with analysis of the GLP-2R promoter.

Special thanks also to Dr. Patricia Brubaker for her scientific collaborations and
encouragement over the past years. Finally, a special thanks to Dr. Theodore Brown for his
time, effort, flexibility and help with the very late-night ICV GLP-2 feeding experiments.
Most appropriately, the final person 1 ivould like to thank is Dr. Daniel .J. Drucker.
As my Ph.D. supervisor, Dr. Dmcker has provided me with not only exceptional academic
training, but has instilled the necessary inspiration and confidence within me to pursue a
career in research and medicine. On al1 accounts, Dr. Dmcker has been an exceptional
supervisor, mentor, and teacher, tenaciously set on ensuring al1 of his students achieve
success. Above all else thanks for believing in me. For your kindness, sincerity, patience,
dedication, and most of al1 for providing me with an experience that i will value and treasure
always.

vii
TABLE OF CONTENTS
. . ................................................................................................. Abstract ..II
......................................................................................... Co-Authorship.. iv
.................................................................................... Acknowledgements.. v
........................................................................................... List of Tables.. xi
. . List of Figures ........................................................................................... XII
................................................................................. List of Appendices.. .'uv
............................................................................... List of Abbreviations.. .xv
Dissemination of Work Arising from tbis Thesis ................................................ xix
* - ............................................................................................. Foreword.. m i
.................................................................. CHAPTER 1: INTRODUCTION,. 1
1.1 Proglucagon and Proglucagon-Derived Peptides (PCDPs) in the Intestine and 3 ........................................................................................... Pancreas.. .- 7 ..................................................................... a) Gene expression -
b) Post-translational processing.. ................................................... -4 c) Biological actions of the progiucagon derived peptides. ....................... 6
1.2 Glucagon-like peptide2 (GLP-2) in the Gastrointestinal Tract.. ......................... 7 ......................................................................... a) Introduction.. 7
......................................................... b) Secretion and metabolism.. 8 c) Biological actions in the gastrointestinal tract ................................. 11
(i) stomach.. .................................................................. I 1 ................................................... (iï) srnall and large bowel 1 1 . -
(iii) experimental bowe t rnjury.. ............................................ 12
13 Proglucagon and Proglucagon-Derived Peptides in the Central Nervous System.. ............................................................................................ I 5
............................................................... a) Gene expression.. .S 5 ........................................ b) Regdation of synthesis and secretion.. 16
.............................. c) Biologkal actions in the central nervous system 18 (i) glucagon., ................................................................ 18 (iï) glucagon-like peptide-1.. ............................................... 18


3.3 Methods ..................... ,.,. ........................................................... 54 3.3.1 Characterization of GLP-ZR sequences and transgene construction 3 4 3.3.2 CNS tissue dissections ...................................................... S5 3.3.3 RNA isolation and RT-PCR analysis ....................................... 56 3.3.4 Irnmunocytochemistry ....................................................... 57 3.3.5 Histochemical analysis .... ............. ................................... 3 8 3.3.6 Microscopy .................................................................... 58 3.3.7 Peptides ........................................................................ 58 3.3.8 Analysis of GLP-2R signaling in GLP-2R-transfected Baby Hamster
Kidney (BHK) cells .......................................................... 59 3.3.9 lntracerebroventricular (ICV) peptide injections and food intake ...... 60
3.4 Results .......................................................................................... 60
3.5 Discussion ....................................................................................... 82
CHAPTER 1: ONTOGENY OF THE CLUCACON-LIKE PEPTLDE-2 AND CLUCAGON-LIKE PEPTIDE-2 RECEPTOR AXIS IN THE DEVELOPING RAT CENTRAL NERVOUS SYSTEM ........... 91
Specific Aims of Research ............................................................................ 92
4.1 Research Summary .......................................................................... 92
4.2 introduction .................................................................................... 93
.......................................................................................... 4 3 ~Methods 94 4.3.1 Anirnals ....................................................................... -94
...*..*.*...... .............***...........*....*..*..* 4.3.2 CNS tissue dissection ,., 95 .......................... 4.3.3 Tissue preparation for RIA and HPLC analysis 95
4.3.3 Radioimmunoassay (RIA) analysis ........................................ 95 ....................................... ........... 4.3.5 HPLC analysis ,., ..,.. 96
4.3.6 RNA isolation and semi-quantitative RT-PCR ........................... 97 4.3.7 Fetal and neonatal rat brain tissue experirnents... .....,,....... .......... 98 4.3.8 Peptides ....................................................................... 100 4.3.9 CAMP RiA of neonatal and fetaI rat brain tissues ...................... 100 4.3.10 Statistics ...................................................................... 100
4.4 Results .......................................................................................... 101
4.5 Discussion ..................................................................................... 109

CHAPTER 5: DISCUSSION ............................................................... I l 3
................... 5.1 a) The role(s) of GLP-2 during intestinal development 114 5.2 b) The role(s) of GLP-2 in the brain ......................................... 118 5.3 c) The role(s) of GLP-2 during brain development ........................ 122
......................................................................................... APPENDICES 126 ............................................................................ Appendir L 127 ............................................................................ Appendix 2 129 ............................................................................ Appendix 3 131 ........................................................................... Appendix 4 -133
....................................................................................... REFERENCES 135

LIST OF TABLES
CRAPTER THREE: GLUCAGON-LIKE PEPTIDE3 RECEPTOR EXPRESSION, BIOACTIVITY AND SPECIFICITY IN THE RODENT CENTRAL NERVOUS SYSTEM
paee 3.1 Analysis of endogenous GLP-2R and GLP-2R promoter-lac2 transgene
expression in rodent brain, bowel and peripheral tissues.. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ..77

LIST OF FIGURES
C W T E R ONE: WTRODUCTION Page
1.1 Identified DNA control elements and trans-acting binding-proteins in the 2.3-kb promoter region of the rat proglucagon gene.. .................................................... 3
............................. 1.2 Tissue-specific expression of the marnmalian proglucagon gene.. 5
1.3 The predicted serpentine diagram for the rat GLP-2 receptor ................................ 24
CHAPTER TWO: ONTOGENY OF THE GLUCAGON-LIKE PEPTIDE-2 AND GLUCAGON-LI= PEPTIDE-2 RECEPTOR AXIS IN TIHI DEVELOPINC RAT INTESTINE
2.1 PGDP leveis in intestinal extracts from rats of différent developmental stages. ....... 39
2.2 GLP- in plasma frorn rats throughout development and in fetal intestinal cells ...... 40
3.3 Analysis of rat GLP-ZR, proglrrcagon, and GAPDH mRNA transcripts by RT-PCR.41
2.4 Relative levels of GLP-2 receptor and proglucagon mRNA transcnpts in developing rat gastrointestinal tissues ..................................................................... 43
............................ 2.5 Effect of daily h[~ly']-GLP- administration in neonatal rats 36
CHAPTER THREE: GLUCACON-LIKE PEPTIDE-2 RECEPTOR EXPRESSION, BIOACTIMTY AND SPECIFICITY iN THE RODENT CENTRAL NERVOUS SYSTEM
3.1 Nucleotide sequences at the 5'-end of the GLP-2R mRNA and gene. ................ ..62
3.2 RT-PCR analysis of endogenous GLP-2 receptor and GLP-2R promoter-lac2 transgene expression in adult mouse tissues.. ............................................ ..66
3.3 Analysis of endogenous GLP-2R and GLP-ZR promoter-lac2 transgene expression in the cerebellum ................................................................................... 69
3.4 HistoiogicaI analysis of GLP-2R and GLP-SR promoter-lacztransgene expression in the hippocampus and dentate g p ........................................................ ..72

Page Histological analysis of GLP-2R promoter-lacZ expression in the arnygdaIoid, hypothalarnic and thalamic nuclei of transgenic mice.. ................................. ..35
~ [ G ~ Y ' I G L P - ~ inhibits dark phase food intake in rnice .................................... 79
Exendin (9-39) is a specific GLP-1 receptor antagonist in BKK fibroblasts expressing ................................................................... the cloned GLP-1 receptor.. 8:
Mode1 illustrating the possible interaction behveen GLP-1 and GLP-7 responsive neuronal systems in the regdation of food intake in the murine central nervous
.......................................................................................... system.. 86
CFWPTER FOUR: ONTOGENY OF THE GLUCAGON-LUCE: PEPTIDE-2 AND GLUCAGON-LIKE PEPTIDE-2 RECEPTOR M I S IN THE DEVELOPING RAT CENTRAL NERVOUS SYSTEM
Analysis of IR-GLP-2 throughout rat hypothalamic and brainstem development,,. 102
Molecular foms of IR-GLP-2 in fetal or adult rat hypothalamus by HPLC and .................................................................................. RIA analysis.. 103
Relative levels of rat GLP-2 receptor mRNA transcripts throughout rat brain ................................. developrnent determined by semi-quantitative RT-PCR LOS
Effect of GLP-2 treatment on CAMP accumulation in neonatal . .................................................................. rat bram dispersion cultures 108
EMéct of GLP-2 rreatment on CAMP accumulation in cultured fetal rat hypothalamic (E19), cortical (E14) and braïnstem (E14) cells .......................... 1 10

xiv
LIST OF APPENDICES
Appendix 1. Copyright release agreement From Elsevier Science.. ................... 12 1
Appendix 2, Copyright release agreement From Munksgaard Scientific Journais.,. 129
Appendix 3. Copyright release agreement h m Endocrinology ....................... 13 1
Appendix 4. Copyright release agreement from the American Society for Biochemistry and MoIecular Biology ..................................... ..133

LIST OF ABBREVIATIONS
3v A aa AC ACTH Am' AHi ANOVA BAC Beta 2 bP Sm4 C CAMP CAP CBS cDNA CdxA Cdx2 C d a 3 CNS CP CRE CREB D D3V DEPC DG DM DMEM DMH DM, DNA DP-EV E E 16 E 19 EDTA EGF Ets F FBS FRIC G
third ventride aIanine amino acid arcuate nucleus adrenocorticotropin reIeasing hormone agouti related protein amygdalohippocampal a m analysis of variance bacterial artificial chromosome Beta2MeuroD basic helk-hop-helix Bctor base pairs brain-4 cysteine cyciic 3', 5' adenosine monophosphate CREB-associated proteins CAP-binding sites complementary deoxyribonucleic acid caudal-related chicken homeobox caudal-relatcd homeobox-2 caudal-related homeobox-213 central nervous system basal cerebraI peduncle CAMP response element CAMP response element binding-protein xspartate dorsal third ventncIe diethyl pyrocarbonate dentate gyrus dorsomedial nucleus Dulbecco's Modified Eagle Medium dorsomedial nucleus of the hypothalamus dorsomotor nucleus deoxyribonucleic acid dipeptidyl peptidase four glutamate embryonic day sixteen embryonic day nineteen ethylenediamine tetca-acidic acid epidermal growth factor ubiquitous developmentd transcription factors phenylalanine fetal bovine semm fetal rat intestinai cells glycine

xvi
G-protein GAPDH GATA- 1 GC GDP GH GIP GLL GLP-1 GLP-1 R GLP- 1 R'- GLP-7- GLP-2R GIuc GLUTag G[Y G f CRS GrDG GRP GRPP GUE GTP H Hl0 Hf3 hGH HM= HPA HF'LC HR 1 IBD ICV m-y IGF-1 IGF-II IP IP-1 IP-2 IR ISE Isl k L LiCl LHA
GTP-binding proteins glyceraldehyde-3-p hop hate dehydrogenase a member of the family of zinc finger transcription factors guanine and cytosine guanine dinucleotide phosphate growth hormone glucose-dependent insulinotropic peptide glucagon-like immunoreactivity glucagon-like peptide one glucagon-Iike peptide one receptor glucagon-Iike peptide one receptor knockout (hornozygous) glucagon-Iike peptide two glucagon-like peptide two receptor glucagon glucagon-SV40 T antigen glycine g-protein coupled receptors granular layer of dentate gyms gastrin-releasing peptide glicentin-reIated pancreatic polypeptide glucose upstream element guanine trinucleotide phosphate histidine water, dihydrogen oxide habenular nucleus human growth hormone hepatic nuclear factor hypothalamic-pituitary-adrenal high performance Iiquid chrornatography hilus region isoleucine inflammatory bowel disease intracerebroventricular interferon-y insulin-like growth factor-[ insulin-Iike growth factor41 intraperitoneal intewening peptide one intewening peptide two immunoreactive islet-ce11 specific enhancer islet tim-homeodomain guanine or thymidine leucine lithium chloride lateral hypothalamus

xvii
LHRH LLDM K kb kDa M MC4-R MD MdD MdV MPGF n N N-ProG N-terminal nls NF-Kappa NPY NSAiD NTS P *P PACAP Pau PBS PC PCR PGDPs PKA PMCo PN ProG PS 1 PTH P W PYY Q R RACE RIA RNA RT RT-PCR s SAS SBS
luteinizing hormone releasing hormone laterodorsal thalamic nucleus lysine kilobases kilodalton methionine melanocortin-4 receptor mediodorsal thalamic nucleus dorsal rnedullary nucleus ventral medullary nucleus major proglucagon fragment number asparagine arnino-terminal of proglucagon amino-terminal nuclear localization signal nuclear factor Kappa B neuropeptide Y non-steroidal anti-inflammatory dnig nucleus of the solitary tract proline observed significance level (*P, **P, ***P) pituitary adenylate cyclase-activating polypeptide paired homeobox phosphate buffered saline prohormone convertase polymerase chain reaction proglucagon-derived peptides protein kinase A posterornedial cortical amygdaloid nucleus post-natal proglucagon homologous to a cis-regdatory element in the somatostatin gene parathyroid hormone paraventricular nucIeus peptide tyrosine tryrosine glutamine arginine rapid amplification of cDNA ends radioirnmunoassay ribonucleic acid reverse-transcriptase reverse-transcriptase polymerase chain reaction serine statistical analysis software short bowel syndrome

SCID S.D. S.E.M. SIG sm SON Sel STZ SV40 T TNF-a TPN TSH TRX UC UN-TX UTR v VIE' VLG VM VP \V
W X-G AL
severe-combined immunodeficiency standard deviation standard error of the mean signal peptide sma medullaris suprsoptic nucleus ubiquitous zinc finger transcription factor streptozotocin simian virus 40 large T antigen threonine tumour necrosis factor-alpha total parenteral nutrition thyroid stimulating hormone transcription ulcerative colitis untranslated region untranslated region valine vasoactive intestinal polypeptide ventrolateral geniculate nucleus ventromedial hypothalamic nucleus valine-pyrrolidide adenosine or thymidine tryptophan 5-bromo-3-chloro-3-indoly 1-B-D-galactopyranoside tyrosine
iMETHODOLOGICAL ABBREVIATIONS
metres centimeters, 1.0 .u 10-'rn millimetres, 1.0 x 10-~rn litres micro litres, 1 .O x 10% hour(s) minute(s) roorn temperature gram micro gram 1.0 x lodg nano gram 1 .O x 1 0 - ~ ~ pic0 gram 1.0 x IO-'- g

xix
DISSEMINATION OF WORK ARISiNG FROM TüIS THESlS
CHAPTER ONE:
Publications
Lovshin J, and Drucker DJ 2000 New fiontiers in the biology of GLP-2. Regdatory Peptides 9027-32- Review
Lovshin J, and Drucker DJ 2000 Synthesis, secretion and biologica1 actions of the giucagon-like peptides. Pediatric Diabetes 1:49-57. Review
Drucker DJ, Lovshin J, Baggio L, Nian LM, Adatia F, Boushey RP, Liu Y, Saleh J, Yusta B, Scrocchi L 2000 New developments in the biology of the glucagon-like peptides GLP-1 and GLP-2. Annals of the New York Academy of Science 92 1 :226-32. Review.
CHAPTER TWO:
Publications
Lovshin J, Yusta 8, Iliopoulos 1, Migirdicyan A, Dableh L, Brubaker PL*, and Drucker DJ* 2000 Ontogeny of the glucagon-like peptide-', receptor mis in the developing rat intestine. Endocrinology 14 L:4194-0 1. *Borh are equal senior CO-a~cthor.
Abstracts Lovshin J, Yusta B, Drucker DJ, and Brubaker PL 2000 Ontogeny of the GLP-UGLP-2 receptor a i s d u h g rat intestinal development. Digestive Disease Week 2 000, American Gastroenterological Association, May 2 1-24, San Diego, CA, USA, Abstr. 1692, DDW' 2000.
CHAPTER THREE:
Publications
Lovshin J, Esta11 J, Yusta B, Brown TJ, and Drucker, D J 2001 Elimination of GLP-I receptor signaling enhances GLP-2 action in the rnunne centrai necvous system. journal of Biological Chemistry 276:2 1489-2 1499.

Abstracts Lovshin J, Brown TJ, and Drucker DJ 2000 Tissue-specific control of glucagon-like peptide-2 receptor expression in mice expressing a GLP-ZR- lacZ transgene. Digestive Disease Week 7000, Amencan Gastroenterological Association, May 21-24, San Diego, CA, USA, Abstr. 1693, DD@ 2000.
Oral Presentations
Lovshin J, Estall J, Yusta B, Brown TJ, and Drucker DJ 2001 GLP-1 receptor signaling modulatcs anorectic GLP-2 action in the murine central nervous system. Digestive Disease Week 7001, American Gasteroenterological Association, May S 1-23 Atlanta. GA, USA, p. 139, Abstr. 215, DD@ 2001.
Lovshin J, Estall J, Yusta B, Brown TJ, and Drucker DJ 200 1 Elimination of GLP-IR signaling enhances anorectic GLP-2 action in the murine central nervous system. Laidlaw Manuscnpt Cornpetition. Institute of Medical Science, University of Toronto, May 15, Toronto, Ont.
Lovshin J, and Drucker DJ 2000 Localization of an enterotrophic receptor. Eastern Student Research Forum, February 23-27, University of Miami, Miami, FL.
Awarded Abstracts or Manuscripts
2001 Manuscript Finalist- Laidlaw Manuscript Com~etition. Finalist in manuscript competition, Institute of Medical Science, University of Toronto. May 15,200 1. Toronto, Ont.
2001 Abstract Winner- BBDC/TDA Annual Trainee Awards Comoetition. Second place in trainee abstract competition. Toronto Diabetes Association Annual Meeting, May 200 1. Toronto, Ont.
2000 Abstract Winoer- BBDCITDA Annual Trainee Awards Com~etition. Third place in trainee abstract competition. Toronto Diabetes Association Annual Meeting, May 2000. Toronto, Ont.
2000 BBDC (Banting and Best Diabetes Centre) Traioee Travel Award. Banting & Best Diabetes Centre, University of Toronto. Toronto, Ont.

1999 Abstract Winner- lnstitute of Medical Science for ESRF. Abstract competition for the East Student Research Forum (ESRF). Represented iMS and presented at the ESRF, University of Miami, Febmary 2000. Miami, FL.
Patents Filed
200 l The enhancement of GLP-2 action. (initiaIly filed Febmary 2001). (Inventors include: Dr. Danie1 J. Drucker, Dr. Theodore Brown and Julie Lovshin).
2000 The uses of the GLP-ZR promoter. (initially filed April2000). (Inventors include: Dr. Daniel J. Drucker and Julie Lovshin)
CHAPTER FOUR
f ublications Mantiscript in prepararion.
Abstracts Lovshin J, Eubanks J, Brubaker PL, Drucker DJ 200 1 GLP-2 receptor expression in the developing rat central nervous system. 83rd Annuaf Meeting of the Endocrine Society, June 20-24, Denver, CO, USA, p.529, Abstr. 371, The Endocrine Society Press, Bethesda, MD.

FOREWORD
Glucagon-like peptide-2 (GLP-2) is a 33 amino acid peptide and like glucagon-like
peptide- 1 (GLP-l), is synthesized fiom proglucagon throughout specialized regions of the
brain and enteroendocrine intestinai L-cells of the bowei. GLP-2 is liberated from its
proglucagon precursor in the intestine and brain by the proteolytic actions of prohormone
convertases. The sequence of GLP-2 along with the other peptides encoded in
preproglucagon, was first determined in a hamster preproglucagon cDNA by Graham Bell in
1983 (1). The physiological relevance of GLP- however, remained doubtful for years
despite the cloning of proglucagon cDNAs h m a number of species. Recent investigations
however, have sparked renewed interest and enthusiasm in the biological actions and
therapeutic potential of GLP-2, as studies suggest that GLP-2 is coupled to a nwnber of
actions in the gastrointestinal tract through its high-affinity G-protein coupled receptor, the
GLP-2 receptor.
The following introductory chapter is presented to serve as background to the
proglucagon gene, and the biological actions of the proglucagon-derived peptides. A review
of the biological actions, rnetabolisrn and thenpeutic potential of GLP-2 in the intestine is
provided as a basis for understanding the rationale and biologicai implications of the research
that is presented in individual chapters in this thesis. Sections wiIl also focus on proglucagon
gene expression and the actions of the proglucagon-derived peptides in the central nervous
system. The implications of the research are critically reviewed and presented in a forma1
discussion chapter at the end of this thesis. Finaiiy, future studies and research directions are
also outIined in the discussion and an appendix is provided for specific references.

INTRODUCTION
" Sections of this Chapter have been previously published.
1. Reprinted fiom Regulatory Peptides, volume 90 (1-3), Lovshin, J., Drucker, DJ. New fiontiers in the biology of GLP-2, pages 27-32, Copyright 2000, with permission from EIsevier Science.
2. Reprinted frorn Pediatric Diabetes, volume 1, Lovshin, J., Dmcker, DJ. Synthesis, secretion and bio[ogical actions of gIucagon-like peptides, 49-57, Copyright 2000, with permission h m Munksgaard Scientific Journals.

1.1 Proglucagon and Proglucagon-Derived Peptides (PCDPs) in the Intestine and Pancreas.
(a) Gene expression
Proglucagon is part of a superfarnily of peptide hormones. the glucagon-secretin
superfamily, that are related by considerable sequence similarity. Membcrs of this
superfamily are expressed in the intestine, pancreas and central nervous system ( a s ) (2).
The proglucagon gene is composed of six exons and five introns. It encodes glicentin,
giucagon. intervening peptide-l (IP-1), glucagon-like peptide-1 (GLP-l), intervening-
peptide-2 (W), glucagon-likc peptide-2 (GLP-2) and in mammals is transcribed as a single
messenger RNA (mRNA) transcnpt (l,3-6). -
In the pancreas, proglucagon is produced in the islets of Langerhans, in pancreatic a -cells
(7). in the intestine, proglucagon is synthesized in enterendocrine L-cells (8,9). In some
mammalian species, the proglucagon gene is also expressed in the stomach (IO).
A number of factors that regulate Ievels of proglucagon rnRNA have been identified, and
some of these factors include dibutyryl CAMP (1 1), protein kinase A ( P U ) (12), dietary
fiber (l3), gastrin-releasing peptide (GRP) (14), fasting (15), short chain fatty acids (16),
bowel resection (19) and diabetes ( 17-1 8). Other factors have been identified that regulate
the secretion of the proglucagon-derived peptides including carbohydrates (20-24): fats (25,
26), proteins (21), somatostatin-28 (27), glucose-dependent insutinotropic peptide (GP) (28),
GRP (gastrin-releasing peptide) (29)- and vagal stimulation (30).
There have been a nurnber of cis-acting DNA reguIatory sequences identified that regulate
islet-ce11 specific and enteroendocnne-specifTc control of proglucagon gene expression as
presented in Figure 1.1. Briefly, at least seven regdatory elements have been identified in
the prornoter region of the progiucagon gene designated as G1, G2, G3, G4, G5, CRE (CAMP

Regulatory elements in the promoter region of the rat proglucagon gene.
Pigure 1.1. ldentified DNA control elements and bans-acting binding-proteins in the 2.3-kb promoter region of the rat proglucagon gene, G 1, G2, G3, G4 and G5 (G5 nof shuwn) are elements located in the rat proglucagon promoter region that contain a-celllislet enhancers. CRE and CBS confer responsivity to cAMP and calcium influxes. lSEs direct intestinal specific expression of the rat proglucagon gene in enteroendocrine cells. ISE; intestinal specific enhancer CBS; CAP-binding sites CRE; cAMP response element CREB; cAMP response element binding-protein CAP; CREB-associated protein CES; C/EBP enhancer site IRBP; insulin responsive binding-protein HNF3; hepatic nuclear factor-3 BTS; ubiquitous developmental transcription factors Beta2; Beta2MeuroD basic helix-loop-helix factor Id-1; islet lim-homeodomain protein Brn-4; brain-4 Cdx2; caudal-related homeobox-2 Pax6; paired homeobox-6 TATA; TATA-box Tm transcription [Adapted from Kieffer TJ, and Habener JP 1999 The glucagon-like peptides.
W
Endocrine Reviews 20:876-9 13 with permission].

respouse element) and an ISE (intestinal-specific promoter elernent). Trans-acting DNA
binding-proteins that interact with some of these elements and modulate glucagon gene
transcription have also been identified including Brn4 (3 l), Cdx2 (32,33,34), Pax2 (35),
Pax6 (36), E47 and Betd/Neuro2d (37), Isl-l (38,39), hepatic nuclear factor (HNF) -3p and
HNF-3a (40,41), and members of the Eh family (40). As its name irnplies, the CRE element
confers CAMP responsivity through a CRE binding-protein (CREB) (12,42-44).
Furthemore, the CRE element can also be activated via membrane depolarization or calcium
inflwc (45).
(b) Post-translational processing
The cloning of proglucagon omplementary DNAs (@NA) in the early 1980's facilitated
the analysis of proglucagon post-translationai processing. Studies exarnining the expression
of proglucagon cDNA in non-endocrine ceIl lines resulted in unprocessed proglucagon,
which suggested that there was cell-speci fic machinery responsible for proglucagon
processing (46).
In marnmals, the tissue-specific expression of the proglucagon gene is a result of post-
translational processing by prohomone convenases (47,48). These enzymes are part of the
subtilisin family, and PC2 and PC 113 are the main members of this family of enzymes that
are involved in the proteolytic processing of the proglucagon precursor at both pairs and
single basic amino acid residues (49). In non-mamrna~ian vertebrates, such as birds and fish,
tissue-specific expression of proglucagon occurs as a result of alternative mRNA splicing
(50). In some invertebrates, such as lamprey, multiple proglucagon transcrïpts are produced
(51). As illustrated in Figure 1.2. processing of proglucagon in the a-cells of the pancreatic
istets of Langerhans gives rise predominately to glucagon, GRPP (glicentin-related

Pigure 1.2. Tissue-specilic expression of the mammalian proglucagon gene. (a) The proglucagon gene contains six exons (E 1 -E6) and five introns (IA-IE) and encodes glicentin, GRPP, oxyntomodulin, glucagon, GLP- 1, IP- 1, GLP-2, IP-2, and the MPGF in a single transcript. (b) The proglucagon-derived peptides are liberated from the proglucagon precursor via the proteolyt ic actions of prohormone convertases that are di fferentially expressed in the pancreas, intestine and central nervous syste&; hence the tissue-specific expression of the proglucagon-d&ived peptides. ~ h e prohormone convertases cleave proglucagon at pairs of basic amino acid residues, The major bioactive hormone generated from proglucagon in the pancreas is glucagon, whereas in the intestine and central nervous system the major biologically active peptides generated from proglucagon are GLP-1 and GLP-2. E; exon I; intron UN-TX; untranslated region SIG; signal peptide N-ProG; amino- terminal sequence of proglucagon GZuc; glucagon GLP-1; glucagon-like peptide one GLP-2; glucagon-like peptide two IP-1; intervening-peptide one IP2; intervening peptide two M; methionine Q; glutamine N, histidine K; lysine R; arginine vI MPGF; major proglucagon fragment GRRP; glicentin-related polypeptide. [Adapted from Kieffer TJ, and Habener JP 1999 The glucagon-like peptides. Endocrine Reviews 203876-913 with permission],

pancreatic polypeptide), P-L, and MPGF (major proglucagon hgrnent) (52-55). In the
intestine, proglucagon processing gives rise ro glicentin (hrther processed to oxyntomodulin
and GRPP), GLP- L 157, IP-2, and G L P - ~ I - ~ ~ (48,56-59). GLP- 1 147 is further processed to
yield GLP-1 7-36amide (2 1,60), rendering GLP- 1 bioIogically active.
(c) Biological actions of the proglucagon-derived peptides
The major product of the proglucagon gene in the pancreas is glucagon. Glucagon
controls blood glucose through regulatioti of glycogenolysis and gluconeogenesis in the liver
(6 L), and regulates the release of insulin tiom pancreatic p-cells (62,63).
In the intestine, glicentin is liberated flom proglucagon and is M e r processed to
oxyntomodulin and GRPP. Glicentin is trophic in the intestine when administered to rodents
(64). Oxyntomodulin inhibits pentagastrin-stimulated gastric acid secretion in vitro and in
rats and humans (65-68). Receptors for both of these peptides have yet to be reported and
therefore our understanding of the physioiogical relevance of these peptides remains unclear.
Glucagon-like peptide-1 (GLP-L) is liberated from proglucagon in the intestine, and is an
important incretin that regulates the enteroinsuh axis by potentiating the release of insulin
fiom pancreatic p-ceiis in a glucosc-dependent manner (69). En the pancreas, GLP-1 also
potentiates proinsulin gene transcription and insulin production (70,71). GLP-1 regulates P-
ce11 mass through effects on islet neogenesis and replication (72,73). GLP-I also regulates
nutrient intake in part through effects on gastric transit (74) and gastric acid secretion (75).
Glucagon-like peptide-2 (GLP-2) is also generated fiom proglucagon in the intestine.
Although for quite sorne time the biological actions of GLP-2 rernained unknown, GLP-2 is a
potent stirnuiator of epitheiial mass in the gastrointestinal tract (64). Like GLP-1, GLP-2
also regulates gastric transit (76) and ùihiiits gastnc acid secretion (74).

1.2 Glucagon-Like Peptide2 (GLP-2) in the Gastrointestinal Tract.
(a) Introduction.
A potential link between increased secretion of the glucagon-related peptides and the
development of intestinal villus hyperplasia was first established following clinical reports of
patients with glucagon-secreting tumors who presented with small bowel villus hyperplasia
in 1972 (77) and in 1984 (78). Normalization of bowel growth and enteroglucagon levels
followed turnor resection (77). Moreover, enteroglucagon was identified in gel-purifred
tumor extracts by iUA analysis (79). Furthemore, laboratory xenotransplantation of tumor
extracts into nude mice induced bowel hypertrophy (80). These clinical observations
together with laboratory investigation, first suggested the potential enterotrophic properties
of the proglucagon-derived peptides.
The identification of a candidate proglucagon-associated factor responsible for the
induction of bowel hypertrophy remained elusive until the report by Drucker (64) and
colleagues in 1996, who first demonstrated the trophic properties of GLP-2 for the
gastrointestinal epithelium in vivo. This discovery originated in observations of bowel
hypertrophy in gIucagon SV40 T antigen (GLUTag) transgenic mice, overexpressing SV40 T
antigen under the transcriptional regdation of 2.3-kb of the proglucagon gene promoter (8 1).
The prominence of gastrointestinal growth in GLUTag transgenic mice suggested that
eievated piasma PGDP (progiucagon-derived peptide) levels contribute to increased bowel
growth. Moreover, transplantation of proglucagon-producing turnour ce11 lines
subcutaneously into nude mice markedly increased small bowel weight and length,
substantiating a link between PGDPs and induction of gastrointestinal epithelial ce11
proliferation (64).

To identi@ which one of the PGDPs was responsible for bowel hypertrophy, glicentin,
GLP-1, GLP-2, and iP-2 were injected into nude mïce every 12 hours for 10 days (64).
Although glicentin demonstrated modest enterotrophic properties (64), GLP-2 treatment
consistently increased small bowel weight and viltus height in both young and old, age-
rnatched male and female mice (82). G L P - treatrnent of rodents induces crypt-cell
proliferation and inhibits apoptosis in both the crypt and villus cornpartments (82,83).
Trophic effects of GLP-2 have aIso been observed in the large bowel(84) and stornach (249).
In contrat, examination of other tissues foilowing short- and Long-term GLP-2
administration did not reveal any proIiferative effects of GLP- outside the gastrointestinal
tract.
In comparative studies with other intestinal growth Factors (including IGF-1 (insulin-Iike
growth factor 1), IGF-II (insulin-like growth factor 21, hGH (human growth hormone), EGF
(epidermal growth factor), GLP-2 and ~ [ G ~ ~ ' ] - G L P - ~ (a protease resistant GLP-2 analogue)
treatment consistently promoted the greatest increase in small bowel weight in rodents
compared with the intestinotrophic effects of the other growth factors tested (84).
Furthermore, CO-administration of GLP-2 with other growth factors promotes greater
increases in srnall bowel weight and length in rodents compared with GLP-2 treatrnent alone,
The focus of this section will be to review our current understanding of GLP-2 synthesis,
secretion, rnetabolism and the biological actions of GLP-2 in the intestine and in
experimental models of intestinal injury.
b) Secretion and metaboüsm
GLP-2 is synthesized fiom proglucagon in the intestinal L-ceII and is secreted from the L-
cell's basolateral membrane (2). It was recentiy demonstrated that the secretion of GLP- in

rats and in humans is regulated by nutrient ingestion (85,86). Regulation by feeding is
consistent with the well-established incretin action of GLP-L and PYY (peptide tyrosine
tyrosine), which are CO-secreted factors along with GLP-2 from the intestinal L-cell.
Moreover, the secretion of GLP-2 by the intestinal L-ce11 in the distal gut appears to be
differentially responsive to nutrients. Increases in GLP-2 secretion are observed upon
ingestion of fat and carbohydrate but not protein, suggesting that the intestinal L-ce11
differentially secretes GLP-2 and other CO-secreted factors, in a highly selective nument-
dependent marner (85). In addition to feeding, GLP-2 levels are increased following
massive small bowel resection (87,88).
There have been several reports over the past years reporting an association between
uncontrolled diabetes and intestinal hyperplasia, which prompted investigators to examine
GLP-2 synthesis and secretion in the setting of experimental diabetes (89). Interestingly,
increases in GLP-2 levels parallel bowel hypertrophy in untreated diabetic rats (89).
Fwthermore, diabetic rats rnaintained on a fibre-diet experience greater intestinal growth and
have higher GLP-2 plasma levels compared to diabetic rats maintained on a fibre-free diet,
suggesting that GLP-2 might be mediating the effects of fibre on intestinal growth in
experimental diabetes (90).
Consistent with the inhibitory effects of somatostatin-28 on GLP-1 secretion, it was
recently suggested in studies of isolated perfused porciiie ileum (9 1) that somatostatin-28
might also tonically repress GLP-2 secretion.
At least three circulating molecular foims containing GLP-2 imrnunoreactivity have been
detected using GLP-2 specific radioimmunoassays (92). Through the use of high
performance Iiquid chromatography (HPLC), intact ~ ~ p - 2 ' ~ ' ~ and N-terminally degraded

G L P - ~ ~ ~ ~ ~ , as well as the biologically inactive major proglucagon fragment (MPGF), have
been detected in rodent and human plasma (85,92). Consistent with the presence of a
penultimate alanine residue at position 2, GLP-2, like GLP-1, is cleaved at the N-terminus by
dipeptidyl peptidase-IV (DP-IV); hence circulating GLP- consists of the Full-length, -
bioactive ~ L p - 2 ' ' ~ ~ species and the inactive DP-IV truncated G L P - ~ ~ - ~ ' (93,94).
Consistent with the importance of DP-IV for the regulation of GLP-2 activity, GLP-2"33
is substantialIy more bioactive in a strain of Fischer rats harboring a mutation in the gene
encoding DP-IV (93)- The correlation behveen DP-IV and in vivo GLP-2 degradation
prompted the design of protease resistant foms of GLP-2. Replacement of the penultimate
alanine residue with a glycine residue in GLP-2, [Cily']-GLP-2, demonstrated cesistance to
DP- N in vivo in wild-type rats and gmups treated with [GI~']-GLP-~ had heightened ovenll
small bowel weight compared to wild-type groups treated with native GLP-2 (84,93).
Moreover, treatrnent of mice and rats with a specific DP-IV inhibitor, valine-pyrrolidide
(VP), alone or with exoçenous GLP-2 treatrnent significantly increases smaIl bowel weight
(95). These findings indicate that DP-IV is a key metabolic regulator of GLP-2 in vivo.
Factors other than DP-IV act to regulate the metabolism of GLP-2. An observation that the
plasma of patients with chronic renal failure exhibit a marked increase in Ieveis of plasma
GLP-2 led to the hypothesis that the kidneys may play an integral roIe in GLP-2 clearance
(96,97). Subsequent investigation into GLP-2 clearance revealed that nephrectomized rats
extiibit siower clearance rates of GLP-2 and GLP-2 analogues, suggesting that GLP-2
activity is [ikely balanced by the coordinate action of DP-IV activity and renal clearance
(98)-

c) Biolagical Actious in the Castrointestinat Tract.
(i) stomach
Like GLP-1, GLP-2 potently inhibits upper gastrointestinal motility (75) and gastric acid
secretion (74,75). The enterogastrone properties of GLP-2 were Est demonstrated in
studies of centrally induced antsar motitity in pigs (76). Furthennon, infusion of GLP-2 into
human volunteers potently inhibits gastric acid secretion (74). Taken together, these studies
suggest that like GLP-1, GLP-2 rnight also act as an ileal brake (30,99), in that it is reIeased
from the lower small bowel in response to nutrients and may act on the stomach to inhibit
further gastric transit (75, 100). Considering the gastro-inhibitory effects of GLP-1 are
mediated in part through vagal afferents (75, LOO), it is also reasonable to predict that likely
the transit modulating and the inhibitory actions of GLP-2 on acid secretion in the stomach
are also mediated through similar vaga1 connections. The finding that some of the biological
properties of the proglucagon-derived peptides are likeiy mediated through the vagus nerve,
is consistent with the close positioning of proglucagon-containing neurons in the NTS
proximal to vagal afferent connections in the central nervous system (2).
(ii) small and large bowel
In the small and large bowef, multiple studies have demonstrated that GLP-2
stimulates intestinai growth in rodents (64,89,90,95, 10 t -105).
To detemine whether GLP-2 produces physioIogicaUy functional bowel, studies of gene
expression and enzyme activity Ievek, in control and GLP-2-treated rodents have been
undertaken (106). GLP-2-treated rodents exhibit increases in small bowel &VA, protein and
brush border enzymes (106). Furthemore, GLP-2-treated maII bowel exhibits normal to
enhanced nutrient absorption, as assessed by oral and duodenai glucose, maitose and fat-

absorption studies (106). Glycemic profiles are normal in GLP-2-treated rodents, indicating
that GLP- treatment does not disturb the enteroinsular axis (106). The available evidence
suggests that the increased small bowel mass generated fotlowing GLP-2-treatment in
rodents, is both morphologically and physiologically normal.
In addition to its enterotrophic properties, GLP-2 has also been s h o w to up regulate
glucose uptake by the intestine, within 30 minutes of GLP-2 inhsion, via the induction of
sodium glucose transporter-1 (SGLT-1) and glucose transporter-? (GLUT-2) activity (i07-
109).
(iii) experimental bowel injury
The observation that GLP-2 promotes expansion of the intestinal epitheiium has
stimulated considerable interest in the potential therapeutic roie of GLP-2 in the setting of
intestinal injury. Given the demonstrated importance of enteral nutrition in both the
maintenance of the intestinal epithelial mucosa and the stimulation of GLP-2 secretion,
Chance (IO 1) and colleagues examined the trophic effects of GLP-2 in parenterally fed rats.
In contrat to controls, CO-intlsion of GLP-2 completely reversed villus atrophy and mucosal
hypoplasia in the small bowel (IO 1).
As major small bowei resection is associated with increased secretion of intestinal
proglucagon-derived peptides and increased proglucagon gene expression in the intestinal
remnant (87, 1 IO), the effect of GLP-2 on intestinal remnant growth and absorption
following intestinal resection has been investigated. In modeIs of intestinal resection, rodents
receiving GLP-Ztreatment experienced an augmented adaptive response, enhanced
absorptive h c t i o n and improved jejunal weight gain (1 1 1-1 13).

The therapeutic potential of GLP-2 has also been tested in rodents in experimental models
of both small(114, 1 15) and large (1 16, 1 17) bowel inflammation. Small bowel injury that
rapidly ensues following the induction of enteritis with indomethacin treatrnent is
significantly improved by the administration of ~ [ G ~ ~ ' ] - G L P - ~ (114). Furthermore, h [ ~ l ~ ' ] -
GLP-2 improved histological indices of disease activity and markedly reduced the prevalence
of bacterial infection (1 14). In a rat moder ~Fspontaneous chronic gastrointestinal
inflammation, GLP-2 treatment significantly reduced histological lesion scores and mucosal
darnage ( 1 1 5).
In rnodels of chemotherapy-induced enteritis (1 18) and mucositis (1 19), GLP-2 treatment
attenuates epithelial injury, stimulates intestinal groivth, and prevents ce11 death in the crypt
cornpartment, GLP-2 administration to tumor-bearing rodents (1 19, 120) with or without
chemotherapy does not affect tumor growth or progression but does stimulate gut growth.
Similar therapeutic effects of GLP-2 treatment have also been observed in the setting of
large bowel inflammation (colitis) induced by dextran sulphate treatment (1 16). Mice with
dextran sulphate-induced colitis exhibit severe intestinal injury and weight loss that is
markedly attenuated following twice daily CO-administration of ~ [ G L ~ ' ] - G L P - ~ (1 16).
Furthermore, induction of colonic in8 ammation by CD4+ T ce11 transplantation into severe-
cornbined immunodeficiency (SCID) mice para1IeIs a substantial decrease in colonic GLP-2
content (1 17). Positive effects of GLP-2 treatment have aIso been observed following
ischemic bowel injury in rats. intravenous intiision of GLP-2 following superîor mesenteric
artery occlusion enhanced mucosal regair and significantly decreased rnortality (103).
Some clues as to how GLP-2 might be acting in the setting of experimental intestinal
inflammation may be gained from in vitro analysis investigating the putative role of GLP-2

in intestinal permeability (121). In vitro experiments demonstrate that GLP-2 decreases
paracellular transport of ions and srnaIl molecules, and transepithelial movement of
macromolecules, which likely Ieads to improved intestinal bamer function following GLP-2
treatment. Enhanced or maintained ban-ier fùnction may explain in part the ability of GLP-2
treatment to markedly decrease bactenal translocation and sepsis in models of experimental
bowel injury (121). GLP-2 treatment has also been associrtted with reducing intestinal
myeloperoxidase activity and decreasing cytokine production in experimental models of
intestinal injury (1 14, 1 15).
In studies of circulating GLP-2 in the plasma of hurnan IBD (inflammatory bowel disease)
(122) and SBS (short bowel syndrome) patients with a preserved colon (1 23), total plasma
G L P - ~ ' - ~ ~ content is increased cornpared to healthy subjects. This finding suggests that in
some humans with IBD or SBS, increases in bioactive GLP-2 may represent an adaptive
response to intestinal inflammation and disease, tn contrast, some human subjects with a
resected colon exhibit impaired mea1-stimuiated GLP-2 release (124).
The first clinical trial evaluating the efficacy of GLP-2 administration in human patients
was targeted at a population of patients with short bowel syndrome (SBS) with resected
terminal ileum and colonic tissue, lacking an intact postprandial GLP-2 response (125).
Increases in overall intestinal energy absorption, nutritional status, body composition, and
decreased gastric emptying folIowed a 35-day GLP-2 treatment regime (124).
Taken together, these initial investigations examining the effect(s) of GLP-2 treatment in
diverse settings of experimental intestinal injury, and more recently in human chica l trial,
demonstrate that GLP-2 exhibits utility in pceventing bowel injury, in enhancing ihe
reparative response to intestinal injury in both the smdi and large bowel and in enhancing

energy absorption. Human diseases associated with disrupted GLP-2 action or animal
models with dysfunctional GLP-2 receptor signahg have yet to be reported and may provide
additional insight into therapeutic directions for potential GLP-2 therapies.
1.3 Proglucagon and Proglucagon-Derived Peptides (PGDPs) in the Central Nervous System
(a) Gene expression.
Despite growing interest in the actions of the glucagon-like peptides in the CNS,
comparatively little is known about either the synthesis or transcriptional regulation of the
proglucagon gene in the CNS. The primary site of proglucagon gene expression in the CNS
is in the caudal brainstem (126-128).
In marnmals, the brainstem gives rise to a proglucagon mRNA transcript that is uniform in
size and identical in sequence to those genented in the pancreas and intestine (129). In-siru
hybridization studies using GLP- 1 directed o!igonucleotide tiboprobes ( 126- 128) localize
proglucagon mRNA transcripts to neuronal ceII bodies in the nucleus of the solitary tract
(NTS), In-situ hybridization studies (1 27) together with results of immunocytochemistry and
gel chromatographic analyses (128) in the central and caudal brainstem, demonstrate that the
distribution of neurons containing proglucagon RNA and GLP-1 immunoreactive peptide
overlap. These observations suggest that neurons in the nucleus of the solitary tract that
express proglucagon are capabIe of processing proglucagon to yield GLP-1 (128).
Although GLP-1 processing has been exarnined in the brain, currently there have been no
studies examining whether simiiar proglucagon-expressing neurons are also able to process
proglucagon to generate GLP-2. The distribution of the neurons expressing proglucagon in
the NTS, overIaps with the distribution pattern of a group of non-catechoIaminergic, inhibin-

p and somatostatin-IR (immunoreactive) neurons projecting to neurosecretory cells in the
paraventricular (PVN) nucleus in the hypothalamus (128).
In-situ hybridization studies did not detect proglucagon gene expression in the
hypothalamus (128), however proglucagon mRNA transcnpts have been detected in the
hypothalamus by Poly A+ northern blot analysis (129) although notably at levels 100-foId
less than in the brainstem.
ProgIucagon gene expression has also been detected in the dorsal and ventral medullary
reticular nucleus and olfactory bulb (128)- A single report using sensitive RT-PCR analysis
suggests that proglucagon gene expression may be more widespread in the CNS than initially
appreciated (130). In addition to the hypothalamus and brainstem, progiucagon mRNA
transcripts in developing mice brain are present by RT-PCR detection in the cerebelhm, and
cortex in Cetal (El9) mice, as well as in 2,5, and 12-week old mice at trace levels (130).
(b) Regulation of synthesis and secretion
Although a number of studies have focused on the sites of proglucagon biosynthesis and
the distribution of the glucagon-like peptides in the CNS, few studies have examined the
transcriptional regulation of proglucagon gene expression within the brain.
Studies of transgenic mice have demonstrated that -2.3-kb of 5'-flanking sequence of the
rat proglucagon gene, directed transgene expression in the cerebellum, cortex, hypothalamus
and brainstem (13 1). As endogenous proglucagon mRNA transcripts are also detected by
RT-PCR in the same C N S tissues, it appears that the first -2.3-kb of 5'-flanking sequence in
the rat proglucagon gene correctly directs proglucagon gene expression in the brain. Studies
of transgenic mice demonstrate that -1252 bp of the rat proglucagon gene promoter is
sufficient for regulation of proglucagon gene expression in the brain (132). Transgenic

analysis of human proglucagon gene expression demonstrates that -1600 bp of promoter
sequence correctly directs transgene expression to the brain (133).
Similarly, there are only a few studies examining factors regulating the biosynthesis and
secretion of the proglucagon-derived peptides in the brain. Stimulation of fetal (E19) rat
hypothalamic cells by either dibutyryl CAMP or forskolin increases PGDP synthesis and
secretion (134-136). Unlike CAMP stimulation of fetal hypothalamic cells, phorbot myristate
acetate (PMA) treatment only increases PGDP secretion but does not affect PGDP
biosynthesis (134). To determine possible physiological regulators of hypothalamic PGDP
biosynthesis and secretion, fetal hypothalamic cells were treated with other hypothalamic
neuropeptides. The studies demonstrated that somatostatin-14 treatment of fetaI
hypothalamic cells blocked PGDP production and secretion, whereas GRP had no effect
(134-136).
As the excitatory amino acid, glutamate, has previously been demonsated to be a key
mediator of hypothalamic neuroendocrîne hormone secretion, the effect of glutamate on
PGDP secretion was studied (137). Incubation of glutamate with fetal rat hypothalamic ceils
was associated with increased PGDP secretion but not PGDP synthesis, suggesting that
glutamate rnay be a factor involved in the central regulation of PGDP secretion (137). b
Increases in central proglucagon gene expression have also been detected in the
hypothalamus and brainstem isolated fkom streptozotocin-induced diabetic rats (58).
Changes in glucagon-like irnrnunoreactivity (GLI) in the hypothalamus and brainstem occu
at different points throughout the development of STZ-diabetes, suggesting that STZ-
diabetes rnay induce tissue-specific differences in PGDP production within the CNS.

Consistent with the suggested role of GLP-1 in the central regulation of food intake (138),
hypothalamic GLP-1 content is decreased in fasted rats (139).
(c) Biological actions in the central nervous system.
(i) glucagon
Pancreatic-type glucagon has been detected in mammalian brain (l4O), including hurnan
brain (141-143) and cerebrospinal fluid (144), and in the retina of fish, birds and reptiles
(145). Centrally adrninistered glucagon exerts a nurnber of actions in the CNS to modulate
energy balance, including effects on gIucose regulation, thermogenesis, body weight,
feeding, rnacronutrient absorption and metabolisrn, energy expenditure, and respiratory
quotient ( 146- 148).
The paraventricular nucleus of the hypothalamus (PVN) is one of the central targets of
glucagon, where glucagon acts to stimulate autonomie anorectic pathways (148). Glucagon
suppresses glucose-sensitive but not cortical neurons in the Iateral hypothalamus (LHA),
dorsornedial (DM) and ventral media1 (VM) nuclei of the hypothalamus (149), and alters
hypothalamic catecholamine turnover (150). Reversal of central glucagon-induced
hyperglycemia with cholorisondamine suggests that some central glucagon pathways are
mediated through catecholamines in sympathetic ganglion (146).
(ii) glucagon-likc peptide4
Althouph proglucagon-expressing neurons are primarily restricted to the caudal brainstem
the regional distribution of GLP-1 immunoreactive terminal fields are extensive throughout
the CNS. In addition to the NTS, area postrerna, parabrachial nucleus, raphe nuclei, locus
ceruleus, dorsal motor nucleus of the vagus and spinai cord, GLP-1 is expressed in several
forebrain regions including the olfactory bulb, tempomi cortex, caudal hippocampus,

amygdala, lateral spetum, and preoptic area (1 27, 143). The densest locations of GLP- 1 -
immunoreactivity are detected in areas of the CNS that regdate feeding, including the
paraventricular ( P W ) and dorsornedial (DM) nuclei of the hypothalamus, and to a lesser
extent in the supraoptic (SON) and the arcuate (AC) nuclei of the hypothalamus (137, 143)
and cortical amygdala ( 138).
A role for GLP- 1 in the central regulation of food intake was suggested following the
observation that intracerebroventricular (ICV) administration of GLP-1 inhibits short-tenn
food and water intake (138) in fed and Fasted rodents, and subsequsnt meal size (15 1). In
support of this finding, ICV administration of a specific GLP-IR antagonist, exendin (9-39).
increases food intake in satiated rats (138, 152). In addition to rodents. intravenous infusion
of GLP-1 inhibits food intake in humans (153), including diabetic (154) and obese males
(155, 156). Intriguingly, mice with a nul1 mutation in the GLP-IR are not obese and do not
have dismpted feeding behavior (157). Whether GLP-1 mediated anorexia is mediated
through taste aversion is ambiguous. ICV infusions of GLP-1 at doses of 1.0 to 3.0 pg
inducc a conditioned taste aversion (158, 159), however aversive effects tvere not observed at
Iower doses in a different study (160).
Central GLP-1 administration also activates corticotropin-releasing hormone (CRH) -
containing neurons in the hypophysiotrophic part of the PVN, as well as oxytocinergic
magnocellular neurons in the PVN and SON nuclei of the hypothalamus (1 6 1). Following
central GLP-I injection plasma vasopressin levels are increased but oxytocin levels rernain
unaffected, Centrally applied GLP-1 also induces the release of excitatory amino acids,
aspartic acid and glutamine, in the ventral media1 hypothalamic nucieus (162).

In addition to modulating food and water intake, GLP-1 also mediates the response to
visceral illness and interoceptive stress (15 1). The aversive effects of LiCl are blocked by
ICV administration of a GLP-1R antagonist (163), supporting a role for GLP-1 in mediating
the central response to noxious agents (164). The role of GLP-1 in mediating interoceptive
stress is M e r supported by the observation that GLP-IR'- mice have an abnormal response
to amiety and enhanced corticosterone release in response to restraint stress (165).
Furthermore, central administration of GLP-1 to rats under restraint stress increases fecal
output, and is reversed by central administration of exendin (9-39) (166). Central GLP-1
regdation of colonic output is also blocked by administration of thé corticotropin releasing
factor (CRF) receptor antagonist, atressin. These findings suggest that GLP-1 may act
through central CRF-pathways in mediating its effect on colonic motility during restnint
stress.
In addition to satiety and stress, there are a number of reports describing diverse actions
for GLP-1 in the brain, including a role for GLP-1 in the response to brain injury (167). In
the hippocampus, GLP-1 modulates hippocampal neuronal activity in wildtype (168) and P-
amyloid treated rats (169); CO-infhion of a GLP-IR antagonist with P-amyloid protein
inhibits memory impairment and hippocampal neuronal death (170). GLP-1 also modulates
blood pressure in rats, as central administration of GLP-1 increases systolic, diastolic and
mean arterial blood pressure and heart rate, and these increases are blocked by administration
of exendin (9-39) (171-174). GLP-1 might also regulate thermogenesis, as central (ICV) and
pen'pheral p) administration of GLP-1 to rats decreases body temperature (175).
Furthermore, GLP- 1 may activate the hypothalmic-pituitary-adrenal (HPA) axis. GLP- 1
aIso potentiates the secretion of luteinizing hormone-releasing hormone (LHRH) in

hypothalamic GT1-7 cells (139) and GLP-1 administration to cultured mouse pituitary
thyrotrophs increases CAMP accumulation, and thyroid stimulating hormone (TSH) release
in rat anterior pituitary cells (176).
(iii) glucagon-like peptide2
At the outset of this thesis, a role for GLP-2 in the brain was unknown. Although a report
in 1984 (177) suggested that GLP-2 could stimulate adenylate cyclase activity and increase
CAMP levels in rat hypothalamic and pituitary membranes, the biological role(s) if any, of
GLP-2 in the brain remained questionable.
To ascertain the function of GLP-2 in the CNS we first defined and localized GLP-2
targets in the rodent brain. Furthermore we examined the transcriptional properties and
cellular locaIization of a GLP-ZR promater-lac2 cransgene. We also studied the effects of
GLP-2 and a potent GLP-2 analogue, ~ [ G L ~ I - G L P - ~ on feeding behavior in mice and these
studies are presented in Chapter Three of this thesis.
In comparison to our understanding of GLP-1 in the CNS, basic, fundamental questions in
central GLP-2 biology remained unanswered. For instance, whether neurons in the central
nervous system are even able to correctly process proglucagon precursocs to generate
bioactive GLP-~ ' -~ ' had yet to be determined. To address this issue we identified the
molecular species of GLP-2 in the centrai sites of proglucagon synthesis, the brainstem and
hypothalamus, by HPLC and RiA analysis and the results are presented in Chapter Four of
this thesis.
Given the number of diverse actions of the closeiy related GLP-1 hormone in the brain
(138, 15 1, 159, 163, 166, 167, 170, 174, 175, 178-BO), it seems reasonabIe to assume that
Iike GLP-1, GLP-2 may also exert a number of diverse actions in the CNS. It would be a

valuable undertaking then, to identify the tissue- and cell-specific targets of GLP-2
throughout the brain, in order to facilitate the identification oftht biological actions of GLP-
2 intheCNS.
In as much as our understanding of GLP-3 action in the adult brain is limited, currently
there have been no reports exmining the onset of GLP-2 or GLP-2R expression, or the
role(s) if any for this axis in the developing brain. To address this issue we studied GLP-2
expression, as weIl as the relative expression of the GLP-2R in the developing rat brain and
determined whether populations of neonatal neural celIs are responsive to GLP-2, the resdts
of which are the focus of Chapter Four of this thesis.
1.4 The GLucagon-like Peptide-2 Receptor
(a) Introduction
The largest farniIy of ceIl surface receptors that transduce intracellular signaling upon
ligand binding consists of those receptors that are coupled to heterotrimeric guanine
nucleoride binding proteins, G-proteins, and are collective1y known as b-protein oupled
receptors (GPCRs) (18 1). CurrentIy more than 1000 GPCRs have been discovered. All -
rnembers share a simiiar structural architecture of a serpentine, seven transmembrane
spanning region that is joined by intracellular and extraceiIuIar Ioops (181). Receptor
activation by ligand binding causes confornationai changes in core transmembrane domains.
StrucniraI changes in the third intraceMar Loop that interacts with cytoplasmic G-proteins
occur, initiating guanine diphosphate (GDP) release and guanine triphosphate (GTP) binding
by the heterotrirneric G-proteins (18 1).
With a broad specmm of ligands, the physiologicai and biocfiemical implications of

GPCR signaling are diverse (182). GPCEb are involved in a wide range of regdatory
pathways including prolifentive and mitogenic signaling. As GPCR signaling can activate
important cellular pathways, ceil survival is dependent upon stringent control of these
systems. Mechanisms to regdate or terminate GPCR signaling inchde crosstalk,
desensitization, down regulation and sequestration of ligand or mediator molecules (1 82).
The glucagon, GLP-I, and GLP-2 (illustrated in Figure 1.3,) receptors belong to the
secretin-glucagon superfamily of GPCEb and this family of receptors shares considerable
similarity in amino acid sequence, structure, distribution, hnction and membrane topography
(183)- Other members of this receptor superfamily include receptors for vasoactive intestinal
peptide (VIP), gtucose-dependent insulinotropic peptide (GIP), growth-hormone releasing
bctor (GRF), secretin, pituitary adenylate cyclase activating polypeptide (PACAP) and
parathyroid hormone (PTFI) (1 82). Characteristically, this family ofreceptors, B-class, have
a seven transmembrane spanning region and a large extracellular N-terminal domain.
Typically they have six conserved cysteine residues that likely fonn disulphide bonds (184),
usually contain consensus sites for asparagine-linked glycosyIation, and couple to Gsa
heterotrimeric proteins that stimulate increases in CAMP through adenyiate cyclase activation
(183). The focus of this section will be to review the distribution and signaling of the GLP-
2R and transcriptional regulation of the GLP-2R gene.
(b) Clucagon-like peptide-2 receptor expression
The cDNAs encoding the human and the rat GLP-2 receptor were recently cloned using
combined PCR-expression cloning fiom hypothalamic and intestinal cDNA Libraries (185).
The mammalian GLP-2R shares considerabte sequence identity with the glucagon and GLP-

-6-
-.o. =3 Eszi+,eL--
-au.
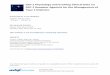
Figure 1.3 The Predicted serpentine diagram for the rat GLP-2 receptor. The architecture of the rai GLP-2 receptor was predicted from the priniary amino acid sequence of the cDNA encoding the rat GLP-2R(185). The rat GLP-2R contains seven putative transmembrane domains mainly composed of aliphatic hydrophobic amino acid residues. The rat GLP-ZR contains a large extracellular N-terminal domain, typical of class B GPCRs. The coloured circles represent single amino acid residues and a colour scheme is provided for reference. [Diagram originally produced by Bernassau JM, Campagne P, and Maigret B in the Viseur program (release 2.35) Copyright 1994, 1995,1996 with permission.]

-1 receptors and with related mernbers of the ghcagon-secretin receptor superfady (185).
The expression o fa single major GLP-2 receptor mRNA transcript {-5.4-kb) is
predominantly restricted to the stomach, small and large bowel, hypothalamus and brainstem
as well as the lung (185, 186). The hurnan GLP-2 receptor gene maps to chromosome
17~13.3, and the hurnan GLP-2 receptor protein is -560 amino acids in length (185).
At the outset of this thesis, a precise definition of GLP-2 responsive ce11 types in the
intestinal mucosa and brain was unknown. Recently a specific antisera raised against the
GLP-2R was developed and was used to locaIize intestinal GLP-2 receptor expression to a
subset of endocrine cells in the human gastrointestinal tract that CO-localize with
chromogranin A, PYY, GIP and GLP-I (186). Focal GLP-ZR* ceIls were also idcntified in
intestinal carcinoid tumors ( l86).
(c) Clucagon-like peptide2 receptor signaling
Consistent with findings in studies of glucagon and GLP-1 receptor signaling (187, GLP-
2 stimulates increased adenylate cyclase activity in fibroblasts transfected with the GtP-2
receptor (188). Activation of AP-I dependent signaling pathways, as exemplified by
induction of transcriptional activity of reporter genes containing AP-1 responsive eiements, is
also observed following GLP-2 stimulation (ISS), although these actions of GLP-2 are IikeIy
indirect and rnediated by the protein kinase A-dependent pathway. Ln contrast to studies
demonstrating activation of calcium influx by either glucagon or GLP-1, there was no
detection of changes of intracellular calcium following activation of the GLP-ZR in baby
hamster kidney (BHK) fibroblasts transfected with the G L P - receptor (188). A modest
stimulation of fibroblast proliferarion and immediate early gene expression was observed
using nanomotar concentrations of GLP-2 in vitro (188). Furthemore, in fïbroblasts

transfected with the GLP-2 receptor, GLP-2 treatment is coupled to the inhibition of cel1ular
apoptosis in a CAMP-dependent protein kinase-independent pathway (1 89).
(d) Regulation of the glucagon-like peptide2 receptor gene
Although the glucagon, GLP-1 and GLP-2 receptors regulate important trophic, and
homeostatic functions, the transcriptional regulation of these genes remains poody studied in
vitro and have yet to be studied in vivo. 5'-flanking sequences of the glucagon receptor gene
have been isolated and studied from rat (190), mouse (191) and human (192). While 5' -
flanking sequences of the human GLP-1 receptor gene have also been isolated and
characterized (193, 194), putative transcriptional regdatory sequences have yet to be
reported for the GLP-2 receptor gene. Hence, currently there is no information regarding the
transcriptional regulation of the GLP-ZR gene in the literature.
Clues to understanding the regulation of the GLP-ZR gene rnay be gained from what is
known about the regulation of the closely related glucagon and GLP-1 receptor genes.
Sequence analysis of the 5'-flanking regions of the glucagon and GLP-1 receptor genes,
reveals that these genes do not contain TATA or CAAT box consensus sequences for basal
transcription initiation. In many TATA-less promoters with high GC (guanine and cytosine)
sequence content, consensus sequences for the ubiquitous Sp 1 transcription factor binding-
protein are found and thought to initiate basal transcription (194, 195). Indeed the genomic
sequences upstream of the putative transcription initiation start sites of the ghcagon and
GLP-1 receptor genes are generally high in GC content (191). For example, the human GLP-
1R gene does not contain TATA or CCAAT box consensus sequences, but contains 74% GC
sequence content proximal to the translation initiation start site (193, 194) of this gene.
Furthemore, three putative Sp1 (-108, -173,-389) recognition sequences are iocated in the

human GLP-1R promoter region (194, 195). Similar to the regdation of the GLP-LR gene, a
GC box is Iocated in the murine glucagon receptor gene in close proximity to the transIation
initiation start codon and four putative Sp 1 recognition sites have been identified in this
promoter (19 1).
In addition to containing consensus sites rnediating basal transcription through putative
Sp 1 transcription factor binding-proteins, elements directing tissue-specific expression of the
gfucagon and GLP-1 receptor genes have been identified. A tissue-specific cis-acting
silencing element that is located in the 5'-flanking sequence of the human GLP-1R gene
(194) conferring pancreatic D-ce11 expression, was recently identified as a PS 1-Iike dernent
(196). Furthemore, a glucose responsive elernent was recently identified in the rat glucagon
receptor gene, which consists of two palindrornic E-boxes, called a G-box (190).
As the glucagon, GLP-1 and GLP-2 receptors exert a diverse nurnber of actions to
regulate nutrient intake, energy disposal, trophic and ami-apoptotic events, the identification
of DNA regdatory sequences that direct the tissue- and cell-specific expression of thesc
genes is a valuable undertaking. Accordingly, we identified regulatory sequences
responsible for directing expression of the rnurine GLP-3R gene in vivo.
At the outset of these thesis studies however, the cDNA encoding the murine GLP-2R was
not cloned and the cDNA encoding the rat GLP-2R gene contained only -25 bp of 5'-
untranslated sequence (5'-UTR). To define the 5'-UTR of the rat gene we used 5'-ppid
amplification of çDNA ends (RACE) reactions and cloned an additionaI250 bp of sequence -
in the 5'-UTR. We then used DNA Fragments fiorn the 5'-UTR of the rat gene as probes to
map and identiQ the 5'-flanking region of the rnurine GLP-2R gene- As currentiy ttiere are
no endogenous ceIl Iines expressing the GLP-2R we directfy exarnined the ability of these

regdatory sequences to drive transcription of a reporter gene in vivo and the results of these
studies are presented in Chapter Three.

ONTOGENY OF THE GLUCAGON-LIKE PEPTIDE-2 AND GLUCAGON-LLKE PEPTIDE3 RECEPTOR AXIS IN THE
DEVELOPING RAT INTESTINE
- - -. - -
' 4 version of this chapter has been previously published.
1. Reprinted fiom Endocrinology, volume 141, Lovshin J, Yusta B, IliopouIos 1, Migïrdicyan A, Dabieh L, Brubaker PL, Drucker DJ. Ontogeny of the ghcagon-like peptide- 2 receptor axis in the developing rat intestine, pages 41944201, Copyright 2000, with permission fiom EndocrinoIogy.

SPECiFIC AIMS OF BESEARCE
As GLP-2 stimulates proliferation of the intestinal epitheliwn in the &J bowe1, GLP-2
might also play one or more rotes in controlling the growth, maturation andor the
development of the fetal or neonatal gastrointestinal tract. As both GLP-2 and the GLP-2R
are expressed in the stomach and gastrointestinal epithelium of aduk rats and mice, }ve
hvporhesized that this mis is also ~wres sed and couded to intestinal woivth in the rodent
neonate.
2.1 RESEARCH SLMMARY
GIucagon-like peptide-2 (GLP-2) is secreted by enteroendocrine cells in the small and Iarge
intestine and exerts intestinotrophic effects in the gastrointestinal mucosal epithelium of the
adult rodent. To ascertain whether the GLP-ZGLf -2 receptor axis is expressed and
Functional in the developing intestine, we studied the synthesis of GLP-2 and the expression
of the GLP-2R in the fetal and neonatal rat gut. GLP-2 imrnunoreactivity (GLP-2-IR) was
detected in the fetal rat intestine, and fetal rat intestinal ceII cultures secreted correctly
processed GLP-~'.'~ into die medium. High levels of GLP-z'*~' were also detected in die
circulation of 13 day old neonatal rats (P<O.OOi w. adult). AnaIysis of GLP-2 receptor
expression by RT-PCR demonstrated GLP-2R mRNA transcripts in fetal intestine, and in
neanatal stomach, jejunum, ileum, and colon. The levels o f GLP-ZR mRNA uanscripts were
comparativdy higher in the fetd and neonatal intestine (P<0.05-001 vs. adult) and declined
to adult levels by post-natal day 2 1. Subcutaneous administration o f a degradation-resistant
GLP-2 analogue, ~ [ G ~ ~ ' ] - G L P - ~ once daily for IO days increased somach (0.009 + 0.0003
YS. 0.007 = 0.002 g/g body mass ~ [ G ~ ~ T - G L P - ~ treated vs. conûols; PCO.03 and small
bowel weight (0.043 * 0.0037 us. 0.03 I 10.0030 g/g body mas; PcO.05). h[Gly2]-GLP-2

also increased both small(2.4 * 0.05 vs. 1.8 k 0.17 cmlg body mass; P<0.05) and large
bowel length (0.32 -i. 0.0 1 vs. 0.25 * 0.02 c d g body rnass, ~ [ G ~ ~ ~ I - G L P - treated vs.
controls, respectively; P<O.O5) in neonata1 rats. These findings demonstrate that both
components of the GLP-XGLP-2 receptor axis are expressed in the fetal and neonatal
intestine. The ontogenic regulation and Functional integrity of this axis raises the possibility
that GLP-2 may play a role in the developrnent and/or maturation of the developing rat
intestine.
2.2 INTRODUCTION
GLP-2, located carboxy-terminal to GLP-1 in the proglucagon molecule, appears to
function as a regulator of intestinal mucosal homeostasis (197). Administration of GLP-2 to
adult rodents stimulates crypt cell proIiferation, and inhibits enterocyte apoptosis, resulting in
expansion of the small bowe1 mucosal epithelium (64,82). The observation that intestinal
injury is associated with increased production of the intestinal PGDPs (!IO), taken together
with the finding of increased circulating levels of bioactive GLP-2 in human patients with
inflammatory bowel disease (122), provides indirect evidence supporting a role for GLP-2 as
an endogenous trophic regulator of intestinal mucosal repair in vivo.
Although intestinal proglucagon gene expression is detectable during fetal gut
development (13 1, 198), the bioIogicaI actions of the intestinal PGDPs during fetal and
neonatal life remain unclear. As GLP-2 e.xhibits intestinotrophic activity in adult rodents, it
seems plausible that GLP-2 may play a role in fetal gut development andlor possibiy in the
complex transition From the neonatal to adult gut. The actions of GLP-2 are mediated by the
GLP-2 receptor (GLP-2R), a recently identified novel member of the G-protein coupled
receptor superfamily (185). AIthough GLP-ZR RNA transcripts have been identified in the

adult rodent gastrointestinal tract (185), the developmental onset of GLP-2R expression has
not yet been reported. Furthermore, whether the fetal or neonatal gut is capable of processing
pcogIucagon to bioactive GLP-2Id3 remains unknown. To determine whether the GLP-
UGLP-2R axis is expressed and hnctional during rat development, we have now analyzed
the coordinate expression of GLP-2 and the GLP-2R in fetal and neonatal rats.
2 3 METEODS
2.3.1 A N I m L S
Wistar rats were obtained from Charles River Breeding Laboratory (St. Constant, QC,
Canada). A11 animal experiments described in this thesis were approved by the Animal Care
Committee s f the University Health Network or the University of Toronto.
2.3.2 TlSSUE PREPARATION FOR HIGH PRESSURE LIQUID
CHROMA T O C W H Y (HPL C) AVAL YSIS*
FetaI rat intestinal cells (19-20 d gestation) were dispersed with collagenase,
hyaluronidase and DNAse (40 mg/dl Sigma Blend Type H, 50 rng/dl and 5 mgdi,
respectively; Sigma Chernical Co., St. Louis, MO), and placed into 60 mm culture dishes in
Dulbecco's Minimal Essential Media containing 5% fetal bovine serum and 3.5 g/l glucose
for 24 hr. Cells were then washed and treated with medium containing 0.5% fetal bovine
serurn, i g/l glucose and 20 pulm1 insulin for 2 hr. Cells and ce11 media were collected
separately at the end of the incubation penod (25). Tissues [whoIe intestine fiom fetal
(embryonic day19-20) and post-natal day 1 (PN1) rats, 5 cm of ileum from al1 other rats
(PN 12, PN21, PN42 and adult), and fetal rat intestinal ceIls in culture; n=3-6 each] were
homogenized in 1 N HCl containing 5% HCOOH, 1% mfluoroacetic acid (TFA) and 1%
NaCl, followed by extraction of peptides and small proteins by passage through a cxtridge of

Cl8 silica (SepPak, Waters Associates, Milford, MA). Plasma was collected into a 10%
volume of ~ r a s ~ l o l ' (5000 Knilml):EDTA(12 mg/ml):Diprotin A (34 pgiml). Two volumes
of 1% TFA (pH 2.5 with diethylamine) was added to plasma, and 1 vol of 10% TFA was
added to media pnor to extraction of the peptides on Cl8 silica [t1=3-5 each] (85, (99).
2.3.3 HPLC ANAL YSIS*
GLP-2-related peptides were separated by high performance liquid chromatography using
a C L8 uBonadapak column (Waters Associates) with a 45 min linear gradient of 30 - 60%
soivent B (solvent A = 0.1% TFA in water; solvent B = 0.1% TFA in acetonitnle), followed
by a 10 min purge with 99% solvent B. The flow rate was 1.5 mumin and 0.3 min fractions
were collected (85. 199).
2.3.1 RADIOIMMLWOASSA Y (RIA) *
Radioimmunoassays for proglucagon-derived peptides (PGDPs) were conducted on dried
samples using a panel of site-specific antisera. The N-terminal PGDPs (glicentin,
oxyntomodulin and glucagon) were detected using antiserum K4023 (Glucagon-like
immunoreactivity, GLI; Biospacific, Erneryville, CA), while antiserum 04A was used to
detect ghcagon alone (Immunoreactive glucagon, IRG: Dr. R. Unger, Dallas, TX). GLP-1-
containing peptides were analyzed using an antiserurn for the free C-terminal, Gly-extended
fom of GLP-1 (antisemrn b5; Dr. S. Mojsov, New York, NY), as well as an antiserum that
recognizes the amidated C-terminal end of the molecule (GLP-1 7-36NH2. , hffinity Research
Products Ltd., Mamhead, UK). GLP-2-related peptides were detected using antiserum
UTTH7, which detects the mid-sequence amino acids #25-30, and thus, both ~ ~ p - 2 ' ~ ~ ~ and
the degraded fom, GLP-~'-'~ (85, 199). Tissue protein levels were determined by Lowry
Protein Assay (200).

* Experiments primdly done by Dr. Patricia Brubaker (Professor, Depts. of Physiology and
Medicine, University of Toronto) and her students Ilias Iliopoulos, Anoush Migirdicyan, and
Liliane Dableh.
2.3.5 TISSUE ISOLA TION FOR RE VERTE-TWSCRIPTASE POL YMERASE
CHAIN REACTION (RT-PCR) ANAL YSIS
Rat gastrointestinal tissues were extracted, cleaned of debris, and irnmediately snap-
fiozen in liquid nitrogen and stored at -80°C until RNA was isolated. Fetal (embryonic day
19-20) stomachs and the entire lengths of the fetal small and large intestine were pooled
separately (n=5 each). For neonataI rats, stomachs from PN 1 pups were removed and
pooled, and the intestines were divided into srnail and large bowel, and pooled (n=4 each).
For PN 12 rats, stomachs were removed and RNA prepared from each sample. Intestinal
samples from PN 12 rats consisted of 5 cm of jejunum (10- 15 cm from stomach), 5 cm of
ileum (5 cm proximal to cecum) and the entire length ofthe colon. For al1 other age groups,
separate gastrointestinal RNA preparations were obtained from rats ( PN2 1, PN42 and adults
rats ) consisting of the entire stomach aIone or 5 cm segments ofjejunum (10-15 cm disial ta
the duodenum), ileum (5-cm proximal to the cecum) or colon (5-10 cm distal to cecum).
2.3.6 RNA ISOLA TION AND SEM-QUA NTlTA T I E RT-PCR ANAL YSlS
Total RNA was isolated using a modified guanidiniurn isothiocyanate protocol(20 1) and
RNA integrity was assessed by agarose gel electrophoresis. For first strand complernentary
DNA (cDNA) synthesis, RNA sampIes were treated with DeoxyribonucleaseI (Life
Technologies, Inc., Toronto, Canada) primed with random hexamers (Life Technologies,
Inc.), and reverse-transcribed with sLJPERscRIPT~~ II Reverse Transcriptase (Life
Technologies, Inc.). R T reactions were andyzed to control for genomic DNA and template

contamination. PCR amplification was canied out using Taq DNA polyrnerase (MBI
Fermentas). Oligonucleotide primer pairs for PCR amplification were as follows: rat GLP-
2R 5'-TTGTGAACGGGCGCCAGGAGA-3'ad 5'-GATCTCACTCTCTTCCAGAATCTC-
3', rat proglucagon 5'-GTTTACATCGTGGCTGGATTG-3 ' and 5'-
TGAATTCCTTTGCTGCCTGGC-3' and for rat GAPDH (glyceraldehyde-3-phosphate
dehydrogenase) 5'-TCCACCACCCTGTTGCTGTAG-3' and 5'-
GACCACAGTCCATGACATCACT-3'. Semi-quantitative RT-PCR analysis oFrat GLP-îR
rnRNA transcnpts was achieved by a ~ e a l i n g cDNA at 65°C for 24 cycles using the
indicated primer pairs. The expected PCR product for the rGLP-2R is 1672 bp, which
corresponds to the full length rat GLP-2R cDNA. For rat proglucagon, cDNA was annealed
at 60°C for 22 cycles and, the expected PCR product is 323 bp. For PCR amplification of nt
GAPDH, cDNA was annealed at 60°C for 18 hrs cycles wiîh an expected PCR product of
452 bp. To control for non-specific amplification, PCR reactions were also camed out in the
absence of tint strand cDNA. The conditions for Iinear PCR amplification of rat GLP-2R,
proglucagon and GAPDH PCR products were determined by cWng out multiple PCR
reactions at varying cycle nurnbers (8-28) and different cDNA input concentrations (0.0 1-2
pl) as indicated in Figure 2.3. The linear range for PCR amplification was determined by
plotting the PCR product yield against either the cyde number or cDNA input arnount.
Following PCR amplification, PCR products were separated by gel electrophoresis on a
1% (wt1v)-agarose gel, transferred onto a nyion membrane (GeneScreen, Life Technologies,
inc.) and blots were hybridized overnight with either a 3'~-~abe~ed intemal cDNA probe for
the rat GLP-2R (185) rat proglucagon (1 1) or rat GAPDH (202) in a hybridization buffer
containing formamide. Membranes were washed stringently at 65OC in 0.1 x SSC and 0.1%

SDS. Membranes were then exposed to a Phosphor Screen (Molecular Dynarnics,
Sunnyvale, CA) and PCR products were visualized and quantifred densitometrically using a
STORM 840-phosphorimager (Molecular Dynamics) and ~ r n a ~ e ~ u a ~ ~ ~ ~ software (Version
5.0, Molecular Dynamics).
2.3.7 CL P-2 ADMINISTRA TZON TO NEONA TrlL RA TS
h[Gly2]-GLPI was a generous gifi fiom NPS AlIeliv Corp (Mississauga, ON, Canada).
Pregnant Wistar rats were acclimatized to the animal facilities and delivered pups one week
following arrival. Within 24 hours of birth, all pups in one litter (n=14 pups) were injected
subcutaneously with ~ [ G I ~ ' ] - G L P - ~ (5 pg in a total volume of 100 pl dissolved in phosphate
buffered saline) and al1 pups in a second Iitter (n=Ll) were injected (subcutaneously) with
saline (100 pl) alone. Pups in both litters were injected once a day at approxirnately 5 pm
for a total of 10 days. On day 11, - 70 hours following the last injection, the neonatal rats
were sacrificed by COz anesthesia. Gastrointestinal tissues (stomach, jejunum, ileum and
colon) were removed, cleaned and weighed as previously descnbed (64,82-84). Tissue
samples colIected for histological examination were fixed ovemight in 10% neutral-buffered
Formalin, paraffin embedded and counter-stained in haematoxylin and eosin.
2.3.8 STA TISTICS
Area-under-the-curve for HPLC peaks t a s determined as the sum of the
imrnunoreactivity under the peak. Differences between groups were determined by Student's
unpaired t test or by analysis of variance (AWVA) using n-1 post hoc comparisons, as
appropriate, on a SAS system (Statistical AnaIysis System, Cary, NC).
2.4 RESULTS
The control of proglucagon processing is compIex and the developmenta1

control of proglucagon processing in the intestine has not been extensively studied.
To ascertain whether intestinal processing of proglucagon was comparable in fetal, neonatal
and adult rat intestine, we anaIyzed the intestinal profile of PGDPs using antisera directed
against glicentin/o.uyntomodulin, glucagon, GLP-1, or GLP-2. The fetal rat intestine
contained readily detectable levels of GLl (g~icentin/o~yntomodulin; P<O.OOI vs. adult
levels), with insignificant amounts of glucagon detected in the sarne extracts (Fig. 2.la).
The content of intestinal GLI increased progressively in the neonatal rat, and reached adult
levels by post-natal Day 2 1. Consistent with the known profile of'proglucagon processing in
the intestine (47,48), the two principal bioactive forms of GLP-1, GLP- 1 7*36NH' and GLP-1'-
" were detected in al1 intestinal extracts analyzed (Fig. 2.1 b), with a progressive increase in
the levels of both molecular fonns ofGLP-1 from fetal to adult life. Similarly, the changes
in immunoreactive intestinal GLP-2 panIIeled the pattern obsewed for both GLI and GLP-1,
with relatively lower levels in newbom intestine (P<O.OOZ vs. adult) increasing to adult
levels by post-natal Day 2 1. To determine whether GLP-2 is correctly processed and
secreted by enteroendocrine cells of the developing rat intestine, we analyzed levels of
circulating GLP-2 by high pressure liquid chromatography and ndioimmunoassay. With this
pmtocol, the levels of both bioactive GLP-2'"I and its circulating degradation product, GLP-
25-33 , are determined as a reflection of total GLP-2 secretion (85,92,93, 122). Plasma levels
of GLP-2 in post-natal Day 12 rats were 8-fold higher than in adult rats (P<0.001), and then
declined progressively to Iower IeveIs in oider animals (Fig. 2.2a). The srnall amount of
pIasma avaiiable fkom fetal rats precluded assessrnent of circulating GLP-2 in these animais.
Accordingly, to ascertain whether the fetal rat enteroendocrine ceU exhibits the capacity for
processing and secretion of bioactive G L P - ~ ' - ~ ~ , we assessed whether immunoreactive forms

Fetal Day 1
Fetal Day 1
Fetal Day 1
Day 12 Day 21 Day42 Adult
Day 12 Day 21 Day42 Adult
Day 12 Day 21 Day42 Adult
Figure 2.1. PGDP levels in intestinal extracts from rats ofdifferent developmental stages. (a) GLI ( l) and RG 0). (b) GLP- 17-jm (Fa) and GLP-17-37 a). (c) GLP-2 @). n=3-6. ** PcO.01 and *** P<O.OOI vs. adult levels. GLI; glucagon-like immunoreactivity RG; immunoreactive-gIucagon.

Day 13 Day 22 Day42 Adult
Frrctlon number
Frrctlon number
Figure 2.2. GLP-2 in plasma from rats throughout development and in fetal intestinal cells. (a) Plasma GLP-2 fiom rats of different developmental stages was analyzed by HPLC and RIA, and the area-under-the-curve was quantitated ( ~ 3 ) . *** P<O.OOI vs. adult IeveIs. (b) HPLC analysis of GLP-2- immunoreactive peptides secreted into the media (upperpanei) and contained in the cells (lowerpand) of fetai rat intestine ce11 (FRIC) cultures (n=5). The arrows indicate the elution positions of synthetic rat GLP-21-33 (fraction 72) and G L P - ~ ~ - ~ ~ (fraction 76).

of GLP-2 were detectable in fetal rat intestinal ce11 cultures (56). Bath GLP-2'"' and ~ ~ p - 2 ~ -
33 were detected in the medium fiom fetal rat intestinal cultures (Fig. 2.2b). Furthemore,
~ ~ p - 2 ' ~ ~ ~ and G L P - ~ ~ - ' ~ were also detected in FRIC (fetal rat intestine cell) extracts (Fig.
2.2b). Taken together, these findings clearly demonstrate that the fetal and by inference fiom
plasma studies, the neonatal rat intestine exhibit the capacity to process proglucagon into
bioactive G L P - ~ I - ~ ~ which is then secreted into the circulation.
Glucagon, GLP-1 and GLP-2 exert their actions through unique G-protein coupled
receptors (185,203,204). In contrast to the glucagon and GLP-1 receptors, the GLP-2
receptor is expressed in a highly tissue-specific manner, predominantly in the gastrointestinal
tract (185). To determine whether the GLP-2 receptor is present in the fetal and neonatal rat
intestine, we assessed the relative expression of the GLP-2 receptor and progiucagon genes in
different regions of the rat gastrointestinal tract by semi-quantitative RT-PCR. We have
previously used this methodology to assess the tissue-specific and developmental expression
of both the glucagon and GLP-1 receptors in the mouse (205). To v e r Q that GLP-2R
mRNA banscripts could be detected and assessed semi-quantitatively, we anaiyzed the
reIationship between input cDNA, PCR cycle number, and the relative Ievels of intestinal
GLP-2R mRNA transcripts. The data clearly show a linear relationship behveen the relative
levels of GLPIR mRNA transcnpts and PCR cycle number (Fig. 2.3a). SimiIar results were
obtained for analysis of intestinal rat progiucagon and GAPDH mRNA transcripts in
comparable experiments (Fig. 2.3a). Furthemore, the relative levels of PCR products for al1
three transcripts exhibited a linear relationship between product abundance and input cDNA
(Fig. 2.3b). A representative PCR analysis fiom experiments demonstrating this rdationship
is shown in Figure 2.3~. As the biological actions of GLP-2 may exhibit deveIopmenta1 and

PCR Cycle Number
m
<
Amt. of Input cONA Amt. of Input cDNA
Maure 2.3. Anilyiii of r i t CLP-2R, progluergon, and CAPDBI mRNA iranicripis by RT-PCR. - ~
(a)~crni-quuntitu~ivc rclsrionship b c t & e f i ~ ~ & A e numbtr and omounts o f PCR praduch for rat GLP-ZR. proglucugon, tuid GAIIDH mRNA transcripu;. (b) Lincar retntionships between input cDNA und KR product over a mngc of input cDNA levels for rat GLP-ZR, proglucogon, und CtAPDH mRNA inuiscripis. (c) Rcpracniritivc Southcrn bloi analysis of RT-PCR reactions carricd out ovcr a range o f cDNA conccntmtions for rat GLP-2R, proglucagon, and GAPDH.

region-specific differences in distinct gastrointestinal compartments (197), we studied the
ontogeny of GLP-2R expression in the stomach and both the maIl and large intestine. GLP-
2R transcripts were detected in RNA isolated fiom fetal, neonatal (Day L and Day 12),
weaned (Day 21 and Day 42) and adult rat stomachs and in both the small and large intestine
at al1 ages exarnined (Fig. 2.4.). In the stomach, the relative levels of GLP-2R mRNA
transcripts decreased slightly from fetal to adult levels but levels were not significantly
different in animals of different ages (data not shown). The relative levels ofjejunal GLP-ZR
mRNA were higher in ktal and neonatai compared to adult rats (P<.OOl), reaching adult
levels by post-natal Day 2 1. The reIative abundance of proglucagon mRLVA transcripts
resembled the pattern obtained for GLP-2R RNA in the jejunum, with cornparatively higher
levels observed in fetal and neonatal jejunum (Pc.01 us. adult), followed by a decline to
adult levels in Day 2 1 jejunum (Fig. 7.4.). The developmental expression of the GLP-2R in
the ileum was similar to that observed for jejunum, with higher levels of GLP-2R mRNA
transcripts detected in fetal gut and Day 1 and Day 12 ileum (P<.O5 vs. adult), followed by a
decline to lower levels in older animals. In contrast, progiucagon mRNA transcripts were
Iower in the fetus and Day 1 iieum, increased markedly by Day 12 (Pc.05 LIS. aduit), then
decreased to lower levels in the ileum of oider animals (Fig. 2.1.). In the colon, the levels of
GLP-2R mRNA transcripts were most abundant at Day 12 (P<.OOI vs. adult), followed by a
progressive decrease to adult levels at Day 12. in contrast, proglucagon gene expression in
the colon was not significantly different at various stages of rat development, The
concomitant expression of GLP-2 and the GLP-2R in neonatal rat gastrointestinal tissues
prompted us to detemine whether the GLP-2GLP-2R axis was FunctionaI in the
gastrointestinal tract of the neonatal rat. Accordingly, separate liners of rats were injected

'8 ô 8.0 - P< ** 4 s 6,O '
3 2 fi 'g 4.0
'
2.0 t
# E O - Feu1 ' Day I Day 12 2, ~a~ il2 %Il Male
Figure 2.4. Relative levels of GLP-2 receptor and proglucagon mRNA transcripts in developing rat eastrointestinal tissues. Tissue samples were collected as described in Methoch fkom fetal (embryonic day
2.4b. I 2 # O r
- 19/20), post-natal days 1, 12,21,42, and adult rats. Day I rat intestinal samples were dissected between the small and large bowel. The relative expression of GLP-2R (a,b,d,f) and proglucagon (PwG; proglucagon, c,e,g) mRNA transcripts was expressed relative to the densitometric intensiiy of the GAPDH signal in the same PCR analysis (d-g ure presented on adjacent page). The ratios are expressed as the mean I. S.E.M. Densitometry values were quantified using the Molecular Dynamics Phosphorlmager and ImageQuantTM. **P<.OI, ***P<.OOI vs, levels in adult.
2 . 4 ~ . 4.0 ***
***
3.5
g g 3.0 * 2.5
2.0
q 0.5 .g .E 2 fi 0 ,
1 Day l Day 12 Day 2 l Day 42 Aduli Mulc 2s Fcinl Dny l Day 12 Day 21 Day 42 Adult l

Day l
i Day 12 Day 2 l Day 42 AJult
Mate
U Day l Day 12 Duy 2 1 Doy 42 Adult Male
Feiul Day 1 Day 12 Duy 2 1 Day 42 Adull Mlile
Fctal Day l Day 12 Day 2 1 Day 42 Adult Male
Figure 2.4. Relative levels of GLP-2 receptor and proglucagon mRNA transcripts in developing rat gastrointestinal tissues. (d-g)/'igure legend on previous page.

once daily with either saline or ~[GI~']-GLP-~, a degradation-resistant human GLP-2
analogue (93) for 10 days (Fig. 2.5.). Analysis of intestinal tissues dernonstrated a significant
increase in stomach and smdl bowel weight and small bowel length in the ~ [ G ~ ~ ' ] - G L P - ~ -
treated rats (P<O.OS vs. controls; Fig. 2.5.). The crypt and villus cornpartment appeared
similar in control vs. h[Gly2]-GLP-2-treated nts (Fig 2.5e,f). In contrast, no increase in
colon weight but an increase in colon length was detected following h[GlY2]-GLP-2
administration. These tindings demonstrate that activation of the GLP-ZGLP-2 receptor avis
is coupled to increased intestinal growth in the neonatal rat intestine.
2.5 DISCUSSION
The roles, if any, of glucagon and GLP-I during fetal development are not known. The
proglucagon gene is expressed in the fetal pancreas and intestine (13 1, 198), and both
glucagon and GLP-1 are synthesized in the fetal pancreas and intestine, respectively (206,
207). The results of previous studies have detected GLP-2R expression in adult rats (las),
however the developrnental ontogeny oFGLP-2R expression in the developing gut has not
yet been examined. The data presented here estabiish that the developing rat intestine is
capable of synthesizing, secreting, and responding to the enterotrophic peptide GLP-2.
These findings are in agreement with recent studies demonstrating nutrient-dependent
secretion ofGLP-2 in the neonatal pig (208). As the GLP-2R is also expressed in the fetal
and neonatal rat gastrointestinal tract, these findings mise the possibility that GLP-2 may
play one or more roles dunng intestinal development and differentiation. Although the GLP-
1 receptor is widely expressed in multiple tissues during murine development (205), whether
GLP-1 plays a metabolic role in the fetus remains unknown. As development proceeds
normally in GLP-1 receptorJ- mice (1 57, it is unlikely that GLP-1 plays a major mle in the

Controls
h[Gly2]-ûLP-2 Controls h[Gly2]-GLP-2 Controls Figure 2.5. Effect of daiiy h(GIy2]-CLP-2 administration in neonatal rats. Rat pups were administered either h[Gly2]-GLP-2 (5 pg) or saline subcutaneously once daily for 10 days. On Day 11 rats were euthanized for assessrnent of intestinal lengths and weights, which are expressed as a ratio following normalkation to the body mass of each rat (in grams). The ratios are expressed as mean t SEM. (n=14 for h[GlyZ]- GLP-2 -treated, and n=l1 for saline-treated groups for al1 parameters measured). (a) For stomach weights *P<0.01, h[GLy2]-GLP-2 vs. saline-treated controls. (b) For small bowel weights *Pc0.001, h[GIS]-GLP-2 vs. saline-treated controls. (c) For small bowel Iengths *PcO.OOl, h[GIf]-GLP-2 vs. saline-treated controls. (d) For large bowel Iengths *P<O.OI, h[GlyZ]-GLP-2 vs. saline-treated controls. (e and f ) Histological sections fcom proximal jejunum of neonatal rats treated for 10 days with h[GlyT-GLP-2 (e) or saline (f) (e and f me presented on the next page). ST; stomach SB; srnall bowel LB; large bowel.

Figure 2.5. Effect of daüy h[Gly2J-GLP-2 administration in neonatal rats. (see figure legend on prmbus page).

control of pattern formation or organ developrnent. Similady, although the glucagon receptor
is expressed in the fetal liver (205), glucagon action, as assessed by adenylate cyclase
stimulation, is rnarkedly attenuated in fetal hepatocytes, and develops postnatally (209).
Developmental analysis of enteroendocrine ce11 differentiation has established the
presence of glucagon-immunoreactive fetal L-cells in both the small and large intestine (210-
2 12). Although proglucagon-immunoreactive cells and proglucagon mRNA transcripts are
first detected by fetal day 14 in the n t gut (198), a major upregulation in the levels of L-ce11
density and intestinal proglucagon mR'i'iA transcripts occurs behveen fetal day 17 and 18.
intriguingly, an intestinal profile of glucagon-like immunoreactive peptides is also first
detected in the developing rat intestine between fetal day 17- 19 (207,2 13), suggesting that
the molecuiar machinery required for intestinal processing of proglucagon rnay be highly
regulated at this stage of intestinal development. Consistent with these findings, prohormone
convertase 113, the enzyme required for libention of GLP-1 and GLP-2 From proglucagon
(49, 199,214, 215), is developmentally regulated and expressed in the intestine of the fetal
rat (216). The finding that bioactive GLP-2'-" is synthesized in and secreted from fetal rat
intestinal endocrine cells, taken together with the detection of GLP-2R mRNA transcripts in
fetal intestine, raise the possibility that the GLP-UGLP-2R axis may also be functional
during the period of rapid intestinal development in trtero.
GLP-2 administration to adult rats and rnice prornotes crypt ce11 proliferation and inhibits
apoptosis Ieading to expansion of the mucosal epithelium (82,83,93), however, the putative
action of GLP-7 in the fetal or neonatal gut has remained unclear. Although we have
demonstrated the presence of both bioactive GLP-2 and GLP-ZR mRNA transcripts in the rat
fetât gut, our data do not dlow US to make specific inferences about the bioIogica1 role, if

aay, of GLP-2 during fetal intestinal development. We recently deterrnined that pax6 SEYnN
mice exhibit a marked deficiency of intestinal GLP-2-producing L-cells and a greater than
95% reduction in intestinal proglucagon mRNA (2 17). Nevertheless, tliese mutant
proglucagon-deficient mice eshibit apparentIy noma1 intestinal developrnent (2 17), strongly
suggesting that normal levels of intestinal GLP-2 are not essential for development of the
fetal munne intestine.
The relative 1eveIs of GLP-2R expression were comparatively greater in the fetal and
neonatal rat intestine. and declined to lower levels in older animals. Whether these
differences reflect changes in GLP-2R transcription andior RNA stability or developmental
differences in the numbers of GLP-ZR-positive cells cannot presently be determined, as the
cellular localization of GLP-ZR expression in the rat intestine has not yet been reponed. As
the developing intestine undergoes a complex series of rnolecular changes in the growth,
differentiation and fùnction of the intestinal epithelium in response to enteral nutrition (2 l8),
it seems likely that gut peptides such as GLP-2 that are secreted in a nutrient-dependent
manner may contribute to the development and maturation process of the neonatal intestinal
epithelium in vivo. Although previous experiments demonstrated that GLP-2 is trophic to the
mucosal epithelium of 4-5 week old mice (82), the data presented here extend the window of
intestinotrophic GLP-2 action to the immediate neonatal penod in the developing rat gut.
Whether GLP-2 is essential for one or more aspects of intestinal adaptation in the transition
from the neonatal to adult intestinaI epithelium awaits the development of specific GLP-2
antagonists or a GLP-ZR knockout mouse.
As the fetaI and neonatal intestinal epithelium remains comparatively immature, exhibits
defective barrier h c t i o n and continues to develop postnatally (218), it remains highIy

susceptible to extemal injury. Indeed, premature infants are prone to the development of
necrotizing enteritis, a disease characterïzed by necrotizing infection and destruction of the
intestinal mucosa that kequently requires surgical intervention and prolonged hospitalization
in the neonatal intensive care unit (219). Although GLP-2 reduces bacterial infection (1 14)
and decreases mucosal epithelial pemeability in adult mice (2461, whether GLP-2 exhibits
sirnilar actions in the prernature developing human _eut remains unknown. Nevertheless, Our
demonstration that the rat intestinal GLP-2lGLP-ZR axis is present and functional in the
neonatal penod suggests that a d e for GLP-2 in the prevention or treatment of neonatal
intestinal injury be exarnined in future experiments.

GLUCAGON-LW PEPTIDE-2 RECEPTOR EXPRESSION, BIOACTMTY .AND SPECiFICITY IN THE
RODENT CENTRAL LWRVOUS SYSTEM.
' A version of this chapter bas been previously pubhhed.
1. Reprïnted from The Journal of BiologïcaI Chemistry, volume 276(24), Lovshin J, Esta11 J, Yusta B, Brown TJ, Dmcker DJ. Giucagon-like Peptide (GU)-2 action in the murine central nervous system is enhanced by elhination of GLP-1 receptor signaIing, pages 21489-21499. Copyright 200 1, with permission tiom the Arnerican Society for Biochernistry and Molecdar Biology.

SPECLFIC A i M S OF RESEARCH
Although the glucagon, GLP-1 and GLP-2 receptors mediate diverse actions to regulate
key metabolic functions, the molecular mechanisms goveming the transcriptional control of
these genes is poorly detined. We hv~othesized that the eneration o f a trans~enic morise
eq-es sin.^ a sensitive reporter gene under the transcri~tional repzrlation o f the 5 -fIankinq
repion of the GLP-2R Pene ~votcld allow ILS to identifi DNA seariences directine tisstie-
specific ervression of the GLP-ZR eene in the rodent CNS. Cornparison of endogenous
GLP-2R expression in the rodent CNS, characterized using RT-PCR and
immunocytochemistry, with GLP-2R promoter-lac2 transgenic expression will assist us in
identifjing DNA sequences in the 5'-flanking region of the GLP-2R gene governing CNS-
specific GLP-ZR expression in vivo.
3.1 RESEARCH S U W Y
Giucagon-like peptide-2 (GLP-2) regulates energy homeostasis via effects on nutrient
absorption, and maintenance of gut mucosal epithelial integrity. The biological actions of
GLP-2 in the central nervous system (CNS) remain poorly understood. We studied the sites
of endogenous GLP-2 receptor (GLP-2R) expression, the localization of transgenic LacZ
expression under the control of the mouse GLP-ZR promoter and the actions of GLP-2 in the
murine CNS. GLP-ZR expression was detected in multiple extrahypothalamic regions of' the
mouse and rat C N S including ce11 groups in the cerebellum, medulla, amygdala,
hippocampus, dentate gynis, pons, cerebral cortex and pituitary, An -1.5-kb fragment of the
mouse GLP-ZR prornoter directed Lac2 expression to the gastrointestinal tract and CNS
regions in the mouse that exhibited endogenous GLP-2R expression inc1uding cerebellum,
amygdala, hippocampus and dentate gynis. intracerebroventncular injection of GLP-2

significantly inhibited food intake during dark phase feeding in wildtype mice. Disruption of
glucagon-like peptide-1 (GLP-1) receptor signaling with the antagonist exendin (9-39) in
wildtype mice, or genetically in GLP-1 receptor4- mice, significantly potentiated the
anorectic actions of GLP-2. These findings illustrate that CNS GLP-2R expression is not
resûicted to hypothalarnic nuclei, and demonstrate that the anorectic effects of GLP-2 are
transient and modulated by the presence or absence of GLP-1 receptor signaling in vivo.
3.2 INTRODUCTION
In the central nervous system, the glucagon-like peptides are synthesized predominantly
in the caudal bninstem and to a lesser extent, in the hypothalamus ( 126, 127, 129). The
GLP-1 receptor is expressed more widely throughout the CNS (128,205), and GLP-I has
been shown to regulate appetite, hypothalamic pituitary function, and the central responsc to
aversive stimulation (138, 139, 163, 176,220,221). Peripheml administration of GLP-1 or
the lizard GLP-1 analogue exendin-4 also reduces food intake and body weight (222,223)
suggesting that gut-derived GLP-1 provides signals that influence feeding behavior either
directly to the brain, or indirectly, Iikeiy via vagal afferents.
In contrast to the increasing number of studies describing CNS actions of GLP-1, rnuch
less is known about the potential function(s) of GLP-2 in the brain. Recently,
intracerebrovenûicular (ICV) infusion of GLP-2 in rats inhibited food intake (224), similar to
results obtained following [CV infusion of GLP-1 (138,225)- Unexpectedly, the anorectic
effects of GLP-2 in rats were completely inhibited by the GLP-1 receptor antagonist exendin
(9-39) (224). These findings implied that CNS GLP-2 may exert its effects via the GLP-1
receptor to inhibit food intake, or alternatively, exendin (9-39) may also fùnction as a CNS
GLP-SR antagonist. Furthemore, although expression of the rat GLP-2R was reported to be

restricted to the dorsomedial nucleus of the hypothalamus by in-situ hybridization (224),
other studies have reported a more widespread distribution of GLP-2R rnRNA transcripts in
various regions of the rat CNS (226).
To understand the biological function and rnechanisms regulating control of GLP-2R
expression in the brain, we have now studied GLP-2R expression and GLP-2 action in the
rodent CNS using a combination of immunohistochemical, RT-PCR, transgenic and ce11
based analyses.
3.3 METHODS
Al1 animal experiments were approved and carried out smctly in accordance with the
Canadian Council on Animal Care guidelines and the Animal Care Cornmittee at the
Toronto General Hospital, University Health Network, (Toronto, ON), Animals were
allowed to acclimatize to the animal care facilities for at Ieast one week prior to any
expenmental procedure.
3.3.1 CHrlRACTERIZA TION OF GLP-2R SEQUEiVCES AND TUVSCENE
CONSTRUCTION
A genomic clone containing the Y-flanking, 5'-untranslated and coding regions of the
rnurine GLP-7R gene was isotated fiorn a 129 SVJ mouse genomic library. To identiQ
additional GLP-ZR nucleotide sequences 5' to the transIation start site (M), the 5'-end of the
rat GLP-2R cDNA was generated and characterized using adaptor-modified complementary
DNA from rat brain (Clontech Laboratories iNC., Pdo Alto, CA) in 5'-RACE (rapid
amplification of cDNA ends) expenments. SeparateIy, a 1,516 bp (base-pair) fragment of the
mouse GLP-2R gene was subcloned f?om the mouse genomic library (incyte Genomics, St.
Louis, MO), sequenced and ligated immediately 5'- to a cDNA encoding LacZ with a nudear

localization signal (a gift fiom A. Nagy, Toronto, ON). The GLP-2R promoter-CacZ
transgene was gel-purïfied and used for generation of transgenic mice. In total, eight founder
animals were identified by Southern blot and PCR analysis and mated with non-transgenic
mice to detemine germline transmission of the transgene. Three tmsgenic founder mice
(dcsignated #2, #3, #4) exhibited germline transmission, and were used to genemte lines for
further analysis of transgene expression.
3.3.2 CNS TISSUE DISSECTIONS
Male Sprague-Dawley rats (300-500g) or GLP-2R promoter-lac2 transgenic mice were
sacrificed by CO? inhalation and quickly decapitated. The brains were rapidly removed and
placed ventral side up on a chilled glass plate. The pituitary glands were also removed and
frozen in liquid nitrogen. The amygdala, cerebral cortex, cerebellum, pons/midbrain, and
medulla were dissected and fiozen in liquid nitrogen. The amygdala was dissected by first
producing a 3 mm thick coronal section by making a coronal cut at the optic chiasm and at
the posterior edge of the mammillary bodies, A cut connecting the rhinal fissures formed
the dorsal boundary of the amygdaloid block and cuts made continuous with the lateral
ventncles to the lateral hypothalamic sulci formed the medial boundanes of the amygdatoid
blocks respectively. The cerebral cortex was also taken from this coronal section and
consisted primanly of parietal and fiontai cortex. The cerebellum was removed and a
coronal cut was made at the posterior edge of the pons. The neural tissue posterior io this cut
comprised the medulla, which also contained the anterior most portion of the spinal cord.
The midbrain block, which also included the pons, extended fiom the posterior edge of the
marnmillary bodies to the postenor edge of the pons, with the cerebelIar and cerebral
cortices, hippocmpus, and amygdala removed,

3.3.3 RNA ISOW TION AND RT-PCR ANAL YSfS
Total RNA was isotated from CNS tissues using Trizoln' reagent (Gibco BRL, Toronto,
Ontario) and fiom peripheral tissues using a modified guanidinium thiocynate procedure
(20 1) and dissolved in ribonuclease-hee water. RNA integrity was assessed on a 1%-(wt/vl)-
agarose gel containing tormaldehyde and visualized on a UV-transilIurninator (Fischer
Scientific, MontreaI, Quebec) using ethidiurn bromide staining. For RT-PCR experiments
RNA samples were treated with DNase 1 (GibcoBRL), primed with random hexamers (Gibco
BRL), and reverse-transcribed with SUPERSCRWT~' II Reverse Transcriptase (Gibco
BRL). To controi for contamination, reactions were also carricd out in the absence of
SLIPERSCRIPT~. Following firs t-rond cDNA synthesis, samples were heared with
Ribonuclease H (MW Fermentas, Vilnius, Lithuania) to remove RNA. For subsequent PCR
amplification, first stnnd cDNA was used as template. Oligonucleotide primer pairs,
annealing temperature and cycle number for PCR amplification were as foIlows: for rat GLP-
2R: 5'-TTGTGAACGGGCGCCAGGAGA-3'and 5'-
GATCTCACTCTCTTCCAGAATCTC-3' were annealed at 65OC for 40 cycles; for mouse
GLP-ZR: 5'-CTGCTGGTTTCCATCAAGCAA-3' and 5'-
TAGATCTCACTCTCTTCCAGA-3' were ameaied at 65'C for 30 cycles, for rat GAPDH:
(giyceraldehyde-3-phosphate dehydrogenase) 5'-TCCACC ACCCTGTTGCTGTAG-3' and
5'-GACCACAGTCCATGACATCACT-3' were amealed at 60°C for 30 cycles ; for GLP-
2R-Lac2 mrtsgene: 5'-CGCTGA'ITTGTGTAGTCGGR-3' and 5'-
CTTATTCGCCTTGCAGCACAT-3' were annealed at 63°C for 40 cycles. The expected
PCR product for the mouse and rat GLP-2R cDNA is - 1.6-kb, corresponding to full Length
GLP-2R (1 85, L 86). The predicted LacZ PCR product is - 580 bp, and for rat GAPDH the

expected PCR product is - 450 bp. To control for non-specific amplification, PCR reactions
were also camed out in the absence of first strand cDNA. Following amplification, PCR
products were separated by gel electrophoresis, transferred ont0 a nylon membrane
(GeneScreen, Life Technologies) and hybridized with a 32~-labeled 1) intemal cDNA probe
for rat GLP-2R (185, 186), or 2) an intemal LacZ oligonucleotide (5'-
TCAGGAAGATCGCACTCCAGC-3'), or 3) an interna1 cDNA probe for n t GAPDH (202).
Following hybridization, membranes were washed stringently and hybridization signals were
quantified on a STORM 840-phosphorimager (Molecular Dynamics, Sunnyvale, CA) using
~ m a ~ e ~ u a ~ ~ ~ ~ software, Version 5.0, Molecular Dynamics.
3.3.4 IMMUNOCYTOCHEMISTR Y
Rats or mice were deeply anesthetized foilowing intraperitoneal injections of sodium
pentobarbital. Following transcardial perfusion with 0.9% sodium chloride, animals were
perfused with 4% neutral-buffered Formatin for -15 minutes. Brains were removed and
post-fixed at room temperature for 4 hrs or overnight at 4°C. Brains were then cryopreserved
overnight in a 20% sucrose in PBS (phosphate-buffered saline) at CC, frozen in dry ice
vapor and either stored at -80°C or sectioned immediateIy at IO pm on a cryostat. Al1
sections were collected and thaw-mounted onto Superfrost plus slides (Fischer Scientific,
Montreal, Quebec). For detection ofp-gdactosidase inununopositive cells, slides were
incubated for 4 hrs at 37°C in 1:8000 dilution of potyclona1 anti-P-Galactosidase antiserum
(ICN Pharmaceuticals Inc., Costa Mesa, CA). Polyclonal GLP-2R antiserum (1:800) that
recognizes the GLP-2R but not the glucagon, glucose-dependent inhibitory poIypeptide, or
GLP- 1 receptors (1 86) was a gift fiom NPS AIIelix Corp. (Mississauga, Canada). To control
for non-specific immunopositivity, GLP-2R antiserum was also preabsorbed overnight at 4OC

in the presence of recombinant GLP-SR inimunogen (186) and ICC (immunocytochemistry)
studies were carried out with a) GLP-2R a n t i s e m b) preimrnune semm c) antibody diluent
alone, or dl preabsorbed GLP-2R antiserum. Al1 siides were counterstained in haematoxylin.
3.3.5 HISTOCHEMICAL ANGI; YSIS
Brains were isoiated from mice and ptaced in 2% paraformaldehyde 10.2%
glutaraldehyde in PBS fixative for 1 hr at room temperature. Sixty minutes Later, brains were
rinsed in PBS and transferred to 4°C in 15% sucrose in PBS soIution for 4 hours to ovemight,
and subsequently to a 30% sucrose in PBS solution for 4 hours to ovemight for
cryopreservation. Brains were then frozen in dry ice vapor and stored at -80°C. Tissues
were sectioned at 10 prn in a -25 to -30°C cryostat, and subsequently thaw-mounted and
stored at -80°C. Prior to 5-bromo-4-chloro-3-indolyl-~-D-galactop yranosid (X-GAL,
Bioshop Canada, Burlington Ontario) staining, slides were slowly warrned to room
temperature and rinsed in PBS. S tides were treated with X-GAL solution overnight at 37°C.
Following treatrnent with X-GAL solution slides were rinsed in PBS, counterstained witli
eosin and dehydnted in an ethanol series.
3.3.6 MICROSCOPY
Al1 dides were visualized and captured using a video camera (JVC) with a Yi" chip device
adapted (0.63X c-mount) to a light microscope Leica (Leica Ltd, Cambridge). Mapification
is reported as the objective magnification multiplied by the c-mount magnification multiplied
by the electronic magnification (electronic magnification was corrected for by dividing the
diagonal of the image captured by the carnera chip size).
3.3.7 PEPTlDES
Recombinant ~[GL~']GLP-~, a dipeptidyl peptidase IV @P N)-resistant GLP-2 analogue

(93,227), was a gift fiom NPS Allelk Corp. (Mississauga, Ontario), Hurnan GLP-1 (7-
36)NHr, exendin-4 and exendin (9-39) were purchased fiom California Peptide Research Inc.
(Napa, CA). Forskolin and 3-isobutyl-1-methylxanthine (IBMX) were obtained from Sigma
Chernical Co., (St. Louis, MO).
3.3.8 ANALYSIS OF GLP-2R SIGNALING IN GLP-ZR TRANSFECTED BABY
HAMSTER KIDNEY (BHK) CELLS*
Baby hamster kidney fibroblast (BHK) cclls stably transfected with either the rat GLP-1
or GLP-2 receptor were propagated as previously described (188) and levels of intracellular
CAMP were assayed following exposure to individual peptides in Dulbecco's Modified
Eagles Medium containing 100 pM IBMX (Sigma Chemical Co., St. Louis, MO) as reported
(188,227). Cells were incubated for 5 minutes with exendin (9-39) or medium alone before
addition of an agonist (GLP-1, ~ [ G I ~ I G L P - 2 , or exendin-4). The treated cells were then
incubated at 37°C for 10 minutes. Absolute ethanol (-20°C) was added to terminate the
reaction and the plates were stored at -80°C until the ce11 extracts were collected (2-4 hours
Iater). Forskolin was used as a positive control. CycIic 3', 5' adenosine monophosphate
(CAMP) radioimmunoassays (Biomedical Technoiogies, Stoughton, MA) were performed on
dried aliquots of extract and data was normalized to cAMPlwel1. Al1 treatrnents were
performed in triplicate or quadruplicate and the data is expressed as mean + SD. E& values
were calculated using GraphPad Prism 3.00 (GraphPad Software Inc., San Diego, CA).
* Experiments carried out by Jennifer Esta11 and Bemardo Yusta (Banting & Best Diabetes
Centre, The Toronto Hospital, The University of Toronto),

3.3.9 INTRACEREBROVENTRICULAR (ICW PEPTIDE INJECTIONS AND FOOD
INTAKE~
For ICV injections adult male CD1 mice randomized into multiple experimental groups
were anesthetized by inhalation of rnethoxyflurane (Metophane; Janssen, Toronto, Ontario)
(L57). [CV injections were canied out as previously described (159). Anesthetized mice
were injected with equal volumes of saline or peptide dissolved in saline (10 ji) with a 2.5
mm X 30 gauge needle attached to a Hamilton syringe at 0.5 mm-1 .O mm posterior and 1.0
mm lateral to bregma, Following injection, anirnals were allowed to recover for -15 minutes
until the observation of a righting-response. Mice were then weighed, given a premeasured
quantity of rodent chow and food intake was quantified at 1,2,4 and 22 hours. The accuracy
of ICV injection was verified at autopsy analysis by detection of bromophenol dye in the
lateral ventricles ofselected animals. Anirnals were injected with peptide at either 7 pm (for
dark phase feeding studies) or at 10 am foliowing an overnight fast of 15 hours (for fasting
studies).
t~xperiments were carried out by Julie Lovshin and Dr. Theodore Brown (Assoc. Professor,
Depts. of Obstetrics and G ynecology, University of Toronto).
3.4 RESULTS
The observation that GLP-2R mRNA transcripts were restncted to the dorsornedial
nucleus of the rat hypothalamus as demonstrated by in-situ hybridization (224) differed from
men t reports of more widespread expression of the GLP-2R in multiple regions of the CNS
(226). We detected GLP-2R mEWA transcripts not oniy in rat hypothalamus but also in
brainstem by RT-PCR (186). Accordingiy, we re-examined the localization of rodent CNS
GLP-2R expression using a combination of RT-PCR and irnmunohistochemistry anaIyses.

Furthemore, we compared the localization of endogenous GLP-2R mRNA transcripts and
GLP-2R immunopositivity with the regions of LacZ expression in tissues isolated frorn GLP-
2R promoter-lacZ transgenic rnice.
To identify DNA regdatory sequences important for control of CNS GLP-2R expression,
we focused initially on characterization of the 5'-end of the GLP-2R mRNA transcnpt. As
GLP-2R cDNA sequences upstream of the translation start site had not been previously
reported (1 85), we camed out s'-RACE experiments using cDNA ternplate from n t brain to
identih 5'-untranslated sequences of the rat GLP-2R. Multiple RACE reaction products
were consistently obtained that were -500 bp in size. These products were cloned and
sequence analysis demonstrated the presencc of previously identified rat GLP-ZR cDNA
sequences (185) and an additional 104 nucleotides of rat GLP-ZR 5'-untranslated sequences
upstream of the previously reported ATG (Fig. 3.1a).
Using a 213 bp Apa IEma I rat cDNA fragment containing 5'-coding sequences as a
probe, we isolated a -2 kb subclone from a bacteria1 artificial chromosome (BAC) clone
derived From a mouse genomic library. The DNA sequences of the mouse GLP-îR genomic
subclone were aligned with the known rat GLP-2R cDNA sequence (1 85) and with human
GLP-2R genomic sequence we identified in GenBankTMtEBI Data bank, as shown in Figure
3.lb. The rat and mouse sequences exhibit 96% identity over the first 104 bp 5'- to the
initiator ATG codon (rat); the mouse and hurnan GLP-ZR cDNAs exhibit 76% identity over
this same region. Whereas the rat and human GLP-2R sequences contained an upstream
ATG transIation initiation site that would give nse to a GLP-2R protein containing an extra
41 amino acids at the N-terminus, a more distal ATG initiation codon, was identified in the
mouse (Fig. 3. lb). Nevertheiess, transfection studies using a rat GLP-2R cDNA that initiates

Figure 3.1 Nucleotide sequences at the S'-end of the GLP-ZR mRNA and gene.
(a) -230 bp of sequence, including IO4 bp of 5'-untranslated sequences corresponding to the
5'-end of the cDNA encoding the rat GLP-2R obtained from sequencing of RACE products,
is shown. The vertical arrow indicates the 5'-end of the RACE product and the putative start
of transcription, The 5'-untranslated region (5'-UTR) is highlighted in boldfce letters. The
coding sequence is presented in uppercase letters, with the corresponding translated product
presented above in rrppercase bold letters. (b) Shown is the organization of 5'-flanking and
exon 1 sequences in the mouse GLP-2R gene compared with rat exon 1 and hurnan GLP-2R
5'-flanking and 5'-untranslated sequences. Sequence identities are presented in boldface
letters, and dashes indicate gaps introduced to maximize alignment. The DNA sequence is
nurnbered from the putative transcription start site. Potential transcription factor-binding
regions in the putative promoter regions of the human and mouse genes are boxeci. The
predicted translation initiation codons in rat, mouse and human genes are rinderlined. The
predicted translated product of the mouse gene is indicated above the nucleotide sequence.
The vertical arrow indicates the predicted 5'-boundary of intron 1. The Pst [site,
corresponding to the 3'-end of the GLP-2R promoter-lacZ transgene, is indicated by
arrowheads. The letters k and iv are ambiguity codes where k = G or T, and LV = A or T, The
GenBankTMIEBI accession numbers fior the rat GLP-2R sequences derived from Our RACE
experiments is AF338323. The accession number for the nucleotide sequence of our GLP-2R
mouse genomic clone is AF338224. The accession number for the previously published n t
GLP-2R cDNA is AFLO5368. The accession number for GenBankTM/EBI sequences we
identified as corresponding to human GLP-ZR genomic sequence is AC069006. The
accession number for the GenBankTM/EBI submission we identified as corresponding to
mouse GLP-2R genomic sequence is AC0 16464 (c) Construction of the transgene was
achieved by inserting a 1.5-kb Sma W.sl I fiagrnent of the murine GLP-2R gene upstream of
nucIear localization signal-containing lac2 (nls-laca cDNA. The solid black b o . ~ denotes
the presence of GLP-2R 5'-untranslated sequences 5' to the Pst [site shown in Figure 3. L b.
NF-Kappa B; nucIear factor-&
(Figure 3.1 presented on adjacent page)-

3.la. 4 5'-end 5'-UTR rat ~ b p - 2 ~ CDNA aacact tcc t c t c t g g i c i i ggiggagtgc aggiggccac cgcctgcagt acatc t tgga g tg t tgg igg gatgtgcctg c i c t t g t g a a cgggcgccag
M R P Q P S P A V P S W C R E A P V P R V R A Q P V r a t QLP-2R CDNA gag8 A M AGQ CCC CAA CCA AOC CCQ GCA GTG CCC AGT AGA TGC AGA GAG GCA CCC G W CCC CGA GTG AGQ GCA CAO CCA G M
G I P E A Q C P V P L H S Q Q W r a t QLP-ZR cDNA W C ATC CCT GAO GCC CAG 000 CCC GTP CCT CïC CAC TCC CAA CAG ATO
3.1 b. mumm QLP-ZR atgtcttgct ttttcttctg ggcttgct ggcrgcgtag acgtcttggg ggtaggtctg ggrrarrtct h- QLP-ZR ccgccttgtt ctttctcctc agcctgtc gaargcacrg ctgacttagg ga-aggtctg ggrrrrrtct
-203 NF-Kappa B mumm QLP-ZR cccaagattt agga arct tggagattcg gtagatcgct gt---agrgc aactcagrcr h- OLP-2ï1 ccatgctttt gg-9 crgg gccgagaagg aactctgaag actccgtrga ttgctctrga
- l a 3 SPI Spl M T A - 1 + 1 rmt QLP-ZR arcrctt cctctc ga crrggrggrg tgcaggaggc r o u i r OLP-an gtcggcggcc - - - - - - - - - - - - - - - - g gcrracactt cctctctgga crrggrggaa tgcrggrggc huun QLP-PR ccgcctcaga cactctcggc gcagcgtggr gag ttt t gcrrrcrttt cctctgtgga ccragrggar tgcaagagga
-43 gE2a1 C U r a t QLP-ZR crccgcc tgcr gtacrtctt ggrgtgttgg rgggrtgtgc ctgcrcttgt garcgggcgc crggrga ACia CCC mumr OLP-ZR C ~ C C ~ C C ~ ~ ~ C ~ ~ gtacatatt ggagtgttgg rgggrtgtgc ctgcrcttgt graagggcgc crgargg ACQ A W CCC hunn GLP-ZR ggctgca tgcg gtgcrtctt ggicggctag rgagatgtac ccctrcttgt grrggtgcac grggrag AAQ CTG
38 v ~ # t zV r a t QLP-2R CM CCA AOC CCQ QCA QTO CCC AGT AQA MC AQA QAQ QCA CCC GTQ CCC CGA QTG AGG QCA CAO CCA mumm QLP-ZR CAA CCA AQC CCQ OCA GTQ CCC AQT AGA MC AQA QAQ - - - - - - - - - CGT CCC TGC CCC OGQ CGC ACA h- QLP-2R GOA TCG AQC AGQ QCA GGQ CCT GOCi AQA GQA AOC GCO GGA CTC CTQ CCT GQC QTC CAC QAQ CTQ CCC
114 W R R L W Q
r a t QLP-ZR QTQ QQC! ATC CCT QAQ QCC CAQ OCIO CCC OTT CCT CTC CAC TCC CAA CAQ AT0 CQT CTQ CTG TQQ WC mumm QLP-ZR QTW ûûû CTC CCT QCQ QCC CAQ QW C m QAG TCT CTC CAC kCC CAC GQQ CGT CG0 CTC T m QQC h- QLP-ZR ATQ QQC ATC C m OCC CCC TGQ 000 ACC AGT CCT CTC TCC TTC CAC AQQ AAQ TQC TCT CTC Ma W C
i e o p a T P P L s L L L L v s I K Q J ,
rmt QLP-29 O ~ P 000 A- ccc TTC CN? GCC n a c m CTG ma OTT TCC ATC MG CM mumm GLP-ZR QCT Wû ACQ CCC TTC CrC TCC CTG CTT CTQ CTQ QT!F TCC ATC M O CM gtaagracrg- - - - - -rtttttat tcct h u v a GLP-ZR QCT ûûû AûQ CCC TTC Cr(3 ACT CTQ GTC CTQ CTQ OTT TCC ATC M Q CM gtaagagcagttca ttrttattrt tatt
Xho I
3.1~. 5' 1 3' E 5'-UTR

translation fiom the downstreant rat ATG corresponding to the position of the mouse ATG
codon gives rise to a functional GLP-2 receptor (185, 188). Hence the biological
significance of the additional 41 amino acidç predicted to be present in the rat and human but
not the mouse GLP-ZR sequence remains unclear.
Using the mouse genomic GLP-2R sequences, we idcntified genomic sequences in
GenBankTM/EBi Data bank that shared 98% identity over 3,103 nucleotides with our munne
GLP-2R genomic subclone isolated from BAC DNA. Analysis of the murine genomic GLP-
2R sequence in GenBankTM/EBi Data bank also demonstrated the presence of a single
translation initiation codon in the murine gene.
As the glucagon, GLP-1 and GLP-2 receptors are related members of a G-protein coupled
receptor superfamily (183)' we cornpared the sequences of the 5'-untranslated and 5'-
flanking regions of these 3 receptors. We did not find significant sirnilarity using base pair
matching over 5'-untranslated or putative promoter regions. No putative TATA or CAAT
box sequences were identified in the mouse GLP-2R genomic sequences irnmediately 5'- to
the end of the putative 5'-untrdnslated region. Computer analyses identificd several potential
transcription factor recognition sites (TFSEARCH Version 1.3) for CdxA, GATA-1, NF-
Kappa B, and Sp 1 as indicated in Figure 3.1 b. Cornparison of the 5'-flanking regions in the
mouse and hurnan GLP-2R genes reveal70% identity over the first 200-nucleotides 5' to the
start of transcription, however sequence similarity diverged 5'- to this region.
Although the glucagon and GLP- 1 receptor promoters have been characterized in ce11
transfection experiments in vitro (190, 194-196), there is no information availabIe regarding
the transcriptional regdation of the genes encoding the glucagon, GLP-1, or GLP-2 receptors
in vivo. To identiQ GLP-2R regdatory sequences that direct GLP-2R gene transcription to

various regions of the CNS, we ligated a -1 .5-kb Fragment of the rnurine GLP-2R gene
containing 5'-flanking and 5'-untranslated sequences upstream of the lac2 cDNA (Fig. 3. lc)
and generated several lines of GLP-2R prornoter-lac2 transgenic mice. We then assessed
and compared expression of the endogenous murine GLP-2R gene with expression of the
iacZ transgene in different regions of the rnurine CNS. Consistent with the highty tissue-
specific expression of the endogenous rat and human GLP-2R in the GI tract (185, 186), both
lacZ transgene and endogenous GLP-2R mRNA transcripts were detected in stomach (data
not shown), and in both the srna11 and large bowel (Fig, 3.2a). Similarly, we observed GLP-
2R transcripts in several regions of the murine CNS (Fig. 3.2b). Endogenous GLP-ZR
mRNA and transgene-derived lac2 transcripts were identified in the cerebellurn, rnedulla,
pons, amygdala, and cerebral cortex of GLP-2R promoter-lac2 mice (Fig. 3.2b). In contnst,
GLP-2R mRNA transcripts, but not lac2 rnRNA transcripts, were detected in the pituitary
gland. The results of previous studies localized rat and hurnan GLP-2R transcripts and
immunoreactive protein principally to the gastrointestinal tract and CNS (185, 186).
Consistent with the tissue specificity of endogenous GLP-2R expression, the GLP-ZR
promoter-lac2 transgene was not expressed in the liver, kidney, spleen, or heart of transgenic
mice (Fig. 32c). The specificity of transgene expression was further illustrated by
dernonstrating that tissues that did not contain transgene-derived mRNA transcripts also did
not contain ~ a c Z * positive ce11 types (Fig. 3.2d). Furthemore, additional evidence for the
regional specificity of transgene expression in the CNS derives From analysis of numerous
coronal sections of frozen brain tissue from GLP-2R promoter-lacZ transgenic mice, the
rnajority of which did not exhibit any B-galactosidase-Iike immunopositivity when incubated
with a B-galactosidase-specific antisera (Fig. 3.2d and data not shom). Similariy, the

Figure 3.2 RT-PCR anaiysis of endogenous GLP-2 receptor and GLP-2R promoter-
lacZ transgene expression in adult mouse tissues. (a) UT-PCR anaIysis of i) transgene (ds-
l a d ) expression in two different lines of 1.5-kb GLP-îR pronoter-lac2 transgenic mice
(lines 3 and 4) and ii) endogenous GLP-2R mRNA transcripts in gastrointestinal tissues. (b)
RT-PCR analysis of endogenous GLP-2R GLPCR promoter-lacZ (111s-lncZ), and GAPDH
m W A transcnpts in structures isolated fiom the CNS of GLP-2R promoter-lacZ transgenic
mice- (c) Analysis of the tissue-specificity of m s g e n e expression in liver, kidney, Iung,
spleen, and hem From 1 -5-kb GLP-2R prornoter-lac2 transgenic mice. PCR control
designates reaction carried out in the absence of cDNA, whereas RT- control indicates
reactions carried out in the absence of reverse transcriptase. + and - presence or absence of
reverse-transcriptase in fmt strand cDNA synthesis respectively. (d) Analysis of tnnsgene
expression in frozen histological sections (kidney and brain) fiom I .5-kb GLP-2R promoter-
1acZtransgenic mice and littermate control mice incubated with fi-galactosidase antiserum.
(Figure 3.2 presenred orr adjacent page).

3.2a. Duodenum Jejunum Ileum Colon
O 0 i ) - nlslacZ-Line#4
m - - nlstacZ-Line#3 - - + - + +
GLP-ZR - + -
3.2 b. Pituiiary Amygdala Pons Medulla Cortex Cerebellum 1
- 0 8 - CAPDH
- + - + + - + - + -
Jejunum Liver Kidney Lung Spleen Heart RT-control PCR control .-- -- - - - -- -- -
w a I nlsiucZ
Brain
Brain .

majority of coronal sections of fiozen brain tissue incubated with GLP-2R antisenim were
also immunonegative. To localize specific regions and cell types within the rat and mouse
CNS that express the endogenous GLP-2R we used immunopurified antiserurn directed
against the carboxy-terminal region of the rat GLP-2R. The specificity of this antiserum has
been previausIy characterized (186). The antisecum specifically recognizes the GLP-'IR as a
single major producr of - 72 kDa on Western blot analysis and does not exhibit cross-
reactivity against the related glucagon or GLP-1 receptors as demonsmted by the lack of
irnmunopositivity in histological sections of rat liver or pancreas incubated with the GLP-2R
antiserum ( 186). GLP-7R immunoreactive celis were observed in the Purkinje Iayer of the
rat cerebellum with no staining detected in other ce11 types throughout the cerebellum (Fig.
3.3a). GLP-ZR immunoreactive neurons were also detected in the Purkinje ceIl layer of the
murine cerebellum (Fig. 3.3~). In conuast, adjacent sections incubated with preimmune
serum (Fig. 3.3b) or without primary antibody did not exhibit irnmunopositivity in the
Purkinje ce11 Iayer. Similady, preabsorption of GLP-2R antiserum with recombinant GLP-
2R protein completely elirninated GLP-SR immunoreactivity in the cerebellum, Analysis of
transgene expression in GLP-ZR promoter-lac2 tmsgenic mice demonstrated nuclear LacZ
irnmunopositivity in the Purkinje ceIl layer of the cerebellum, consistent with the presence of
a nuclear localization signa1 in the modified IacZ coding sequence (Fig. 3.3e). In contrat,
LacZ* ceIls were not detected in the cerebellum of age-rnatched non-transgenic littermate
controls (Fig. 3.3f). h k i n j e neurons exhibithg positivity for the native GLP-2R in rat and
mouse cerebellum detected with a n t i s e m against the endogenous GLP-2R were
cornparatively more abundant than the number of Lac2 immunopositive Purkinje neurons
identified in transgenic mouse cerebeiIum. As both GLP-2R and GLP-2R prornoter-lacZ

Figure 3.3 Analysis of endogenous GLP-2R and GLP-ZR prornoter-lac2 transgene
expression in the cerebeliurn. Frozen coronal sections of Formalin-perfused rat (a-b), and
mouse (c-d) cerebellum were exposed to anti-GLP-2R antiserum or to preimmune senrm to
detect the specific presence of GLP-2RT cells. Frozen coronal sections of Formalin-perfised
GLP-2R promoter-lacZ le) and lirtemate control (f) mouse cerebellums were incubated with
a polyclonal antibody generated against the P-galactosidase trizyme to detect specific nuclear
LacZ expression in the transgenic cerebellum. Histological regions presented correspond
approximately to the coordinates in Figure 9 1 in the atlas of Franklin and Paxinos (215).
(Figure 3.3 presented on adjacent page).


RNA transcripts were detected in extrahypothalamic regions of the rat CNS (Fig. 3,2b), we
next exarnined these regions for GLP-2R imrnunoreactivity. GLP-2R immunoreactive cells
were detected in the hippocampus and dentate gyms of the rat CNS (Fig. 3.4a-c). Numerous
GLP-2R immunoreactive cells were observed in the pyramidal ce11 layer of the rostral rat
hippocarnpus, including the CA1, C M , CA3 fields; in the caudal hippocampus GLP-2R-
cells were also present in the CA3 fields. GLP-ZR immunoreactive staining was also
observed in scattered ceIl types located in the ganular layer and polyrnorphic hilus region of
the rat dentate gyrus. In contrast, preabsorption of antiserum with recombinant GLP-2R or
the use of preimmune antiserum did not result in detectable GLP-2R immunopositivity in
adjacent senal sections (data not shown). Following detection of GLP-2R immunopositivity
in the rat hippocampus and dentate gyrus, we examined these same structures in the CNS of
GLP-2R promoter-lacZ transgenic mice. Nuclear LacZ immunopositivity was clearly visible
within simiIar ce11 types (that also exhibited endogenous GLP-2R irnrnunoreactivity)
positioned in the hippocampus and dentate gyrus in GLP-2R promoter-lac2 transgenic mice
(Fig. 3.4d-h). Corresponding age-matched littermate non-transgenic control sections did not
exhibit Lac2 immunopositivity under identical staining conditions. In contrast to the extent
of endogenous GLP-ZR immunoreactivity observed in the pyramidal layers of the rat
hippocampus, LacZ-immunopositive cells in the CA1, CA2 and CA3 fields of the transgenic
hippocampus were Iess abundant, with the highest density of positive cells observed in the
CA3 field (Fig. 3.4h). Nevertheless, P-galactosidase cells were dense in the granular tayer
of the transgenic dentate gynis (Fig. 3.4g)- Cells with nuclear B-galactosidase staining were
also observed in the polyrnorphic hilus region of the transgenic dentate gyrus, consistent with
the distribution of GLP-2R-ceIIs observed in the rat dentate gynis using GLP-2R antiserum.

Figure 3.4 Histological analysis of GLP-2R and GLP-ZR promoter-lac2 transgene
expression in the hippocampus and dentate gyrus. (a-c) Frozen coronal sections of
perfused rat brain were incubated with anti-GLP-2R antiserurn to detect the presence of
GLP-2RT ceIl types in the hippocampus and dentate g p s . Endogenous GLP-2R
immunostaining is shown in the rostral hippocampus (a), granular layer of dentate "gynrs
(GrDG, b), and CA3 field of hippocampus (c). (d-h) Frozen coronal sections of perfused
GLP-2R promoter-lac2 rnouse brain were incubated with anti-P-galactosidase antibody to
identib transgene-positive nuclei in the transgenic hippocampus and dentate gyrus. (d). The
bo-red regions in (d) correspond to areas containing LacZ- cells, including the transgenic
dentate gyrus (DG? e and g) and the CA1 field (f) of the transgenic hippocampus using P- galactosidase immunocytochernistry. The caudal CA3 field OP the transgenic hippocampus
using X-GAL histochemical analysis is shown in (h). The relative magnifications (X24, L 20,
and 240) are shown in each panel. Histological regions represented in (a-c) correspond
approximately to the coordinates in Figs.40 and 11 in the atlas of Franklin and Paxinos (245);
those in (d, e, g) correspond approximately to the coordinates in Fig. 43; that in ( f )
correspond approximately to the coordinates in Fig. 53; and that in (h) correspond
approximately to the coordinates in Fig. 59. HR; hilus region cp; basal cerebral peduncle.
(Figure 3.4 presented on adjacent page).


Examination of the CNS in GLP-2R promoter-lacZ transgenic mice, revealed additional
structures that exhibited intense B-galactosidase activity. The amygdala contained a number
of LacZt cells detected bihemispherically by both histochemical and immunocytochemica1
staining (Fig. 3.5a-c). Positive cell staining was principally restricted to the posterolatera1
and posteromedial cortical amygdaloid nucleus and to the amygdalohippocampal area. The
lateral and media1 amygdaloid nuclei did not contain positive staining.
The results of previous GLP-2R in situ hybridization studies demonstrated restricted
hypothalamic GLP-ZR expression exclusiveIy in the caudal part of the rat dorsornedial
nucleus of the hypothalamus (224). Immunocytochemical analysis of the transgenic
hypothalamus revealed occasional rare LacZc nuclei of the dorsomedial nucleus (Fig. 3.5d).
A few rare nuclcar LacZt nuclei were also detected throughout the ventromedial
hypothalamic nucleus in transgenic mice (Fig. 3.5e,f).
Neural populations exhibiting nuclear B-galactosidase activity were also localized to the
thalamic nuclei of GLP-2R promoter-lacZ transgenic mice (Fig. 3Sg,h). Nuclear Lac2
irnmunopositivity was observed in seiect cells in the mediodorsal thalarnic nucleus, and in
the ventrolateral and dorsomedial part of the laterodorsal thalarnic nucleus.
Immunopositivity was also observed in the ventrolateraI geniculate nucleus with the majority
of positive cells in the parvoceMar regions (Fig. 3.50. A few irnrnunopositive cells were
identified in the caudate putamen, dorsal fornix and septofimbrial nucleus of GLP-2R
promoter-lad mice. NucIear P-galactosidase positivity was also detected in celIs at the base
of the corpus callosum in the ventral endopiriform nucleus and in the piriform cortex (data
not shown). A surnmary of the sites of endogenous GLP-2 receptor and GLP-2R promoter-
IacZ transgene expression fs shown in TabIe 3.1,

Figure 3.5. Histological analysis of GLP-2R promoter-lac2 expression in the
amygdaloid, hypothalamic and thalamic nuclei of transgenic mice. (a) Histochemical
analysis of Lac2 positivity in the amygdaloid nucleus of GLP-2R promoter-lac2 transgenic
mice using X-GAL staining. The bo-red area corresponds to LacZ' nuclei in the
arnygdalohippocampal area and the nrrows point to LacZt nuclei in the caudal CA3 region of
the hippocampus and the posteromedial cortical amygdaloid nucleus- (b,c)
immunocytochemical anaIysis of the amygdalohippocampal m a (b) and the posteromedial
(c) cortical amygdaloid nucleus using an antibody directed against the B-galactosidase
enzyme. (d-t) Imrnunocytochemical analysis of the hypothalamus of GLP-2R promoter-lac2
transgenic mice using P-galactosidase antiserum, specifically in the dorsomedial nucleus (d)
and in the ventromedia1 nucleus (mapification x 60 (e) and 240 (0). (g-i)
[mmunocytochemical analysis of thalamic nuclei in GLP-2R prornoter-lacZ transgenic mice
using P-galactosidase antisemm, specifically in the mediodorsal nuclei (g) the dorsomedial
part of the Iaterodorsal thalamic nucIeus (h) and the ventrolateral geniculate nucleus (i). The
awuws point to LacZ' nuclei. The relative magnifications (;Y24,60, 120 and 240) are
indicated. Histological regions presented in (a-c) correspond approxirnately to the
coordinates in Figs. 5 1 and 52 in the atlas of Franklin and Paxinos (245); those in (d-fj
correspond approxirnately to the coordinates in Figs. 46 and 47; that in (g) corresponds
approximately to the coordinates in Fig. 38; that in (h) corresponds approximately to Fig. 41
and that in (i) correspond approximately to Figs. 46 and 47. AHi; amygdalohippocampal arca
PMCo; posterornedial cortical amygdaloid nucleus cp; basal cerebrai pedunde DM;
dorsomedial hypothalamic nucleus Vhc ventromedial hypothalamic nucleus 3 y; third
ventricle D3Y; dorsal third ventricle HL?; habenular nucleus sm; stria rneduIIaris MD;
mediodorsat thalarnic nucleus LLD1Ci; laterodorsal thalmic nucleus VLG; ventrolated
genicdate nucleus.
(FEgire 3.5 presented on adjucenf page).


Table 3.1.
Analysis of endogenous GLP-2R and GLP-2R promoter-lacZ transgene expression in brain, bowel and penpheral rodent tissues,
I I Brain Bowel Peripheral tissues
PT AM HC DG HY P MD CX CB ST DU JE IL CL LV KD LG SP HT
+ + + + + + c + + + + - i f + - - Endogenous GLP-R
Endogenous GLP-2R and GLP-2R promoter-kucZ transgene expression was examined in
rodent bnin, gastrointestinal tract and peripherd tissues using a combination of RT-PCR and
immunocytochemistry analysis. AnaIysis of 1.5-kb GLP-ZR promoter-lacZ transgene
expression in brain tissues was performed in three independent transgenic lines using X-GAL
histochemical analysis. Transgene expression in gastrointestinal tissues was also perfonned
in al1 three independent transgenic Lines using RT-PCR analysis. Transgene expression was
obsewed in the amygdala, hippocampus, and dentate gyrus by histochemistry in al1 three
lines. Immunocytochemistry for P-Gdactosidase was used to ana1yze in more detail the
cellular localization of CNS transgene expression in one line of GLP-2R promoter-lac2
transgenic mice. PT; pituitary il1l.l; amygdala HC; hippocampus DG; dentate gyrus HY;
hypothalamus P; pons MD; medulla C,Y; cortex Cl?; cerebellum ST; stomach DI/; duodenum
JE; jejunum IL; ileum CL; colon L V; liver KD; kidney LG; lung SP; spleen HT; hem.
+ expression; - lack of expression.

As intracerebrovenhicular GLP-2 administration inhibited dark phase feeding in rats
(224), we compared the effects of a DP W-resistant GLP-2 anaiope, ~ [ G ~ ~ ' ] - G L P - ~ (93)
with the GLP-1 analogue, exendin-4 on feeding afier a prolonged fast, or during dark phase
feeding in mice. The GLP-1 analogue exendin-4, but not ~ [ G ~ ~ ' ] G L P - ~ , inhibited food
intake in fasted mice (Fig. 3.6a). In contrast. both exendin-4 and ~ [ G ~ ~ ' ] - G L P - ~
significantly inhibited dark phase food intake (Fig. 3.6b), although the inhibitory effects of
exendin-4 were significantly more potent than those of the GLP-2 analogue. Similarly,
whereas the inhibitocy effects of exendin-4 were sustained over 24 hours, the inhibitory
effects of h[~l$]-GLP-2 on food intake were transient and not detectabIe after more than 4
hours (data not shown).
The effects of GLP-2 on food intake in rats were completely blocked by the GLP-1
receptor antagonist exendin (9-39), suggesting that exendin (9-39) might Function as a GLP-7
aniagonist in vivo (224). Remarkably, the inhibitocy effects of ICV ~ [ G ~ ~ ' ] G L P - ~ on food
intake in wildtype mice were significantly more pronounced in the presence of co-
administered exendin (9-39) at multiple time intervals (Fig. 3.6c), including the first hour. In
contrast, in the absence of exendin (9-39), TCV ~ [ G ~ ~ ' ] - G L P - ~ did not inhibit food intake in
the first hour after peptide injection (Fig. 3.6b,c). Furthemore, as little as 0.05 ug of exendin
(9-39) was sufficient for significant potentiation of the anorectic effects of ~ [ G L ~ ~ I - G L P - on
the inhibition of dark phase food intake at the 1-2 hour time point (Fig. 3.6d).
The demonstration that the GLP-1 receptor antagonist exendin (9-39) significantly
enhances the anorectic effect of GLP-2 in wildtype rnice implied a role for GLP- t receptor
signaling in the regulation of CNS G L P - action. Accordingly, we next examined the effects
of ICV L ~ [ G ~ ~ ~ ] G L P - ~ in mice with complete genetic disruption of GLP-I receptor signding

Wildtype Mice O vehicle
50 pg h[Gly21GLP-2
a 1pg Exendin 4
Figure 3.6 h[Gly21GLP-2 inhibits dark phase food intake in mice. (a-b) Mice were randomized into three groups and received an intracerebroventricular injection of vehicle (PBS), h[Gly2]GLP-2 (50 pg dissolved in PBS) or exendin-4 (1 pg dissolved in PBS) following a 15 hour fast (a) or pnor to the start of dark phase (b). Following recovery, food intake was measured al 1,2, and 4 hours (Hr) d e r injection Values are expressed as mean f S.E.M., (n=6 per group for (a) and n=8-20 for (b)). (3.66.-c are presented on page~fol lo~ngl . (c) Gmups of wildtype mice were administered either vehicle (PBS), h[GIyqGLP-2 (50 pg dissolved in PBS), h[Glf]GLP-2 and exendin (9-39) (Ex (9-39)) (50 pg of each peptide dissolved in PBS) or exendin (9-39) alone (50 pg dissolved in PBS) via intracereborventncular injection. (f) Groups of wildtype rnice were administered intracerebroventriculxty vehicle (PBS), h[GIy2]GLP-2 alone (50 pg dissolved in PBS), or h[Gly2]GLP-2 (50 pg dissolved in PBS) and exendin (9-39) (0.05 or 0.5 or 5 pg dissolved in PBS). (e) Groups of wildtype mice (+/+) and GLP-IR4- mice (4) were injected intracerebroventriculariy with either vehicle (PBS) or h[Gly2]GLP-2 (50 pg dissolved in PBS) just prior to the start of dark phase and food intake was measured at 1,2, and 4 hours. Data are expressed as mean + SE.M., (n=5-8). Treatments at each tirne point for aii panels were compared using a one-way analysis of variance followed by a least sigaificant ciifference muItiple range test using SPSS for Whdows version 5.0.1 (SPSS hc., Chicago IL). *P<O.OJ compared to controls (PBS), or to cornparisons between groups indicated by brackets.

Dark Phase 1
Wildtype Mice vebicle 50 pg h[GlyZ]GLP-2 1 pg Exendin 4
0.7 1 1 Dark Phase /
0-1 Hr 1-2 Hr 2 4 Hr 1 Wiidtype Mice 1

Wildtype Mice 0 vebicle
50 pg h[CIyZjCLP-2 O 50 pg h[GlyZIGLP-2 + 0.05 pg EX (9-39)
50 pg b[ClyzlCLP-2 + 0.5 pg Ex (9-39) 50 pg h(ClyzlGLP-2 + 5 pg Ex (9-39)
Dark Phase 1
CLP-IR4- Mice (4-) O. vebicle RI SO pg h[ClytJCLP-2

(157). Remarkably, GLP-IR'- mice exhibited a significantly greater sensirivity to the
inhibitory actions of the GLP-2 analogue on food intake, as can clearly be seen at the 0-1
hour and 2-4 hour time points, compared to the anorectic effects of an identical arnount of
ICV ~ [ G ~ ~ ' ] G L P - ~ adrninistered in wildtype control mice (Fig. 3.6e).
As the GLP-2-mediated inhibition of food intdce was blocked by the GLP-1 receptor
antagonist exendin (9-39) in rats (224), we examined whether exendin (9-39) hnctioned as a
rat GLP-2 receptor antagonist using cells expressing the cloned rat GLP-2 receptor in vitro.
Although ~ [ G I ~ ' ] G L P - ~ increased CAMP accumulation in a dose-dependent manner in BHK-
GLP-2R cells, increasing amounts of exendin (9-39), from 50-1000 nM had no effect on
h[~ly ' ]~~~-2-s t imula ted CAMP formation (Fig. 3.7). Exendin (9-39) alone did not
stimulate CAMP accumulation in BHK-GLP-2R cells (data not shown). Furthermore, the
GLP-2R responded specifically to ~ [ G I ~ ' ] G L P - ~ as no CAMP accumulation was detected
following incubation of BK-GLP-ZR cells with GLP-1 or exendin-4 (Fig. 3.7~~). In
contrast, exendin (9-39) decreased GLP-1-stimulated CAMP accumulation in a concentration-
dependent manner in BHK-GLP-IR cells (Fig. 3.7b), consistent with its known actions as a
GLP-1 receptor antagonist. Furthermore. the actions of h[~lJ]-GLP-2 were specific for
cells expressing the GLP-2 receptor, as ~[GI~']GLP-S had no effect on CAMP accumulation
in BHK-GLP-IR cells (Fig. 3.7).
3.5 DISCUSSION
Several lines of evidence support a role for glucagon-like peptides in the control of food
intake. Intracerebroventricular administration of GLP-1 agonists inhibits food intake in mice
and rats (138, 157,225), whereas peripheral administration of GLP-1 reduces appetite and
size of meal ingestion in human subjects (154,222). Furthermore, intracerebroventricuIar

BHK-GLP-SR
O 0.5
O 0.01 [peptide] 0.1 1 10 (nM) 100 1000
O. l.7 O 0.01 0.1 1 IO i00100o [peptide] (nM)
Figure 3.7 Exendin (9-39) is a specific CLP-1 receptor antagonist in BHK iibroblasts expressing the cloned CLP-1 receptor. Stably transfected BHK cells expressing either the rat GLP-2 (a) or GLP-I (b) receptor werc pre-treated for 5 minutes with exendin (9-39) (Ex (9-39)) or medium alone before a 10 minute incubation with a rcceptor agonist: human GLP-I (hGLP-1)-(7-36)-NH,, exendin-4 , or h[GIy2]GLP-2. Peptides were diluted in Dulbecco's Modofoed Eagle's Medium containing 100 uM IBMX. The CAMP concentration in aliquots of ethanol extrack was determined using n RIA and norrnalized to show cAMP/well, EC,, values for h[Gly2]GLP-2 in the presence of 0,50,500, or 1000 nM Exendin (9-39) were 0.044,0.059,0.076, and 0.081 nM, respectively. A one-way analysis of variance showed no significant difference between pretreatment conditions with exendin (9-39) al al1 h[Gly2]GLP-2 concentrations on the GLP-2R. A two- factor analysis of variance analysis of the exendin (9-39) antagonism of the GLP- I R showcd a significant différence between pre-treatment conditions, hGLP- 1(7-36)-NH2 concentration, and the interaction between thcm (P< 0,001 for al1 groups). The EC,, value of human GLP-l(7-36)-NH, alone was 0.041 nM, whercas in the presence of 500 nM or 1000 nM exendin (9-39) it was 0.57 and 4.28 nM, respcctively. The EC,, of exendin-4 in BHK-GLP-IR cells was 0.041 nM, Represenlative data from two separaie experinlents perfonned in triplicate are shown. The concentration of peptide is plotted on a logarithmic scale. Error bars represent niean f S.D.

(KV) administration of the GLP-1 receptor antagonist exendin (9-39) increases food intake
in short t e m studies (1381, and promotes weight gain in rats afier 6 days of ICV
administration in vivo (152). Nevertheless, GLP-1 receptor4- mice are not obese, do not eat
more than wildtype mice, and fail to deveIop obesity even following several months of high
fat feeding (157,228). Hence it remains unclear whether the anorectic effects of ICV GLP-1
represent a highly specific effect on C N S feeding centers, or a non-specific effect on C N S
nuclei that mediate the response to aversive stimulation (158, 164,221,239,230). The
avaiIable evidence suggests that although ICV GLP-1 msient ly iahibits food intake (229),
GLP-1 does not appear to be an essential regulator of long-term body weight homeostasis in
vivo.
The report that intmcerebroventricular injection of GLP-2 inhibits food intake in rats (224)
provides new information about a possible roIe for GLP-2 in the CNS. Although our data
demonstrate ha1 GLP-2 transientiy inhibits dark phase teeding in mice, in contrast to the
inhibition of food intake observed with exendin-4. we did not detect significant effects of
GLP-2 on inhibition of food intake in mice after a 15 hour fast. Furtherrnore, our data clearly
show that the effect of GLP-2 on dark phase food intake is not bIocked but is significantly
enhanced in wildtype mice in the presence of exendin (9-39), a GU- i receptor antagonist
(23 1). Consistent with the results obtained in wildtype mice following GLP-I receptor
blockade with exendin (9-39)? GLP-2 more potently inhibited food intake in C t ~ p - 1 ~ ~ -
compared to control wildtype mice. Hence our data clearly demonsuate that in contrast to
results obtained in rats (224), the effects of GLP-2 on food intake in mice are not attenuated
by disruption of GLP-1 receptor signaling. Moreover, transient blockade of the GLP-1
receptor with exendin (9-39) or compleie disruption of GLP-1R signaiing in GLP-1~'- mice

was associated with enhanced sensitivity to the inhibitory response to GLP-2. These findings
provide new evidence for potentia1 functionaI crosstalk between GLP- 1 and GLP-2 receptor
signaliig networks regulating food intake in vivo, as itlustrated in Figure 3.8. Whether GLP-
, IR signaling modulates GLP-2R action directly through interceliular connections or more
indirectly through intervening interneuronal connections or by other mechanisrns and
pathways yet to be described, remains to be determined. Nevertheiess, our data strongiy
implicate the GLP-I R in tonic repression of GLP-3R signaling in the murine central nervous
system.
The finding that the GLP-1 receptor antagonist exendin (9-39) eliminated the GLP-2-
mediated feeding response in rats implied that exendin (9-39) might also be a functional
antagonist of GLP-2. action in the CNS (224). These observations suggested that either
exendin (9-39) blocks the CNS action of GLP-3 at the Ievel of rhe GLP-2 receptor, or
altematively, GLP-2 may mediate its effects indirectly, through downstream activation of the
GLP-1 receptor. Our data in wiIdtype mice CO-injected with ~[GL~ ' ]GLP-~ and exendin (9-
39), taken together with studies using ctoned GLP-2 and GLP-1 receptors, clearly
demonsuates that exendin (9-39) is not a functiona1 antagonist of the GLP-2 receptor in vivo
or in vitro. Furthemore, the demonstration that h i ~ 1 ~ q ~ ~ p - 2 inhibits food intake in GLP-
1 R-' mice, taken together with the enhanced sensiuvity to GLP-2 action following inhibition
of GLP-1 signaling, provides new evidence demonstrating that the effects of GLP-2 on
feeding do not require the GLP-1 receptor but may be modulated in part through the
functional activity of GLP-1 receptor s ignahg In the rat C N S proghcagon mRNA
transcripts have b e n Iocaiized primarily in the caudal par^ of the nucIeus of the s o l i t q tract,
dorsal and ventral rnedulla and the olfactory bulb (126, 128), and to a Iesser extent, in the


hypothalamus (129). in contrast, GLP-1 receptor mRNA transcripts and GLP-1 binding sites
are more widely distributed throughout the CNS in the olfactory bulb, temporal cortex,
hypothalamus, amygdala, hippocarnpus, preoptic area, thalamus, substantia nigra,
parabrachial nuclei, locus ceruleus, nucleus of the solitary tract and the area postrcma (127,
128,232). Moreover, GLP-1 immunoreactive tracts originating from the brainstem project to
several forebrain nuclei including the dorsomedial and paraventricular nuclei of the
hypothalamus, thalamic and cortical areas (127). Hence, the available evidence demonstrates
both GLP-IR expression and GLP-1-immunoreactive tracts in multiple regions of the CNS,
inchding the hypothalamus, amygdala, hippocampus, thalamus and multiple hindbrain
regions. As GLP-1 and GLP-2 are CO-synthesized from a common proglucagon precursor, it
seems likely that many of the GLP-1-immunopositive tracts originating from the brainstern
also contain GLP-2,
The GLP-2R was originally cloned from intestinal and hypothalamic cDNA libraries
(185). GLP-2R mRNA transcripts were detected in rat hypothalamus and bninstern by RT-
PCR analysis ( 1 86), and GLP-îR expression was exclusively localized to the compact part of
the dorsomedial nucleus of the hypothalamus by in-situ hybridization (224). We have now
extended these studies in the rat CNS to demonstrate more widespread distribution GLP-2R
expression in thalamic, hippocampal, cortical and hindbrain regions in addition to the
hypothalamic sites of GLP-2R expression. Our studies are consistent with previous reports
dernonstrating both GLP-IR and GLP-2R expression in hypothalamus, midbrain,
hippocarnpus, striatum and cortex (226), raising the possibility that in the rat CNS, the GLP-
IR and GLP-2R are expressed in either identical or proximal neural cells in these same brain
regions. Furthemore, the demonstrated specificity of the GLP-2R antiserum (M), taken

together with the colocalization of endogenous GLP-2R immunoreactivity and nuclear
localization of Lac2 transgene expression in multiple CNS regions of the mouse, provides
additional evidence supporting a more widespread GLP-2R expression pattern extending
beyond the hypothalamus.
The mechanisms regulating expression of the receptors for glucagon, GLP-1, and GLP-2
have not been extensively examined, AIthough promoter sequences directing expression of
the glucagon and GLP-1 receptor sequences have been analyzed in cell-based transfection
studies (190, 194), the DNA regulatory sequences mediaung tissue-specific control of these
receptor genes in vivo have not yec been identified. Furthetmore, our analysis of the 5'-ends
of the receptor coding regions, 5'-unmnsIated, and 5 ' 4 anking region DNA sequences does
not reveal significant shared nucleotide identity across the glucagon, GLP-1 or GLP-2
receptors, providing indirect evidence for the evolution of distinct control mechanisms
regulating the transcription of each receptor gene. This observation is supported by a recent
examination of the evolution of the receptor DNA sequences for the proglucagon-derived
peptides (233), which suggests that these receptors Iikely evolved independently of each
other.
Our results extend the previously reported GLP-2 receptor sequence at the 5'-end and
identib both the 5'-untransIated region and the location of intron 1. Furthemore, we provide
hnctional evidence for the transcriptional activity of DNA regdatory sequences in the
mouse GLP-ZR 5'-flanking region. Our hdings demonstrate that a -1.5-kb fragment of the
mouse GLP-2 receptor gene containing 5'-flanking and Y-untranslated sequences directs
transgene expression specifica1Iy to the GI tract and brain, in agreement with tlie restricted
pattern of tissue-specific expression demonstrated for the endogenous GLP-2 receptor (185,

186). The identification of potential Sp 1 binding sites in the proximal GLP-2R prornoter is
intriguing in light of studies suggesting the hctional importance of Sp 1 binding sites for
basal GLP-1 receptor transcription in transfection studies in vitro (195). Furthermore,
several studies have demonstrated an important role for both GATA factors and caudal
proteins (Fig. 3. lb) in the regulation of both intestine- and enteroendocrine-specific gene
transcription (33, 234-236). Hence, future studies exarnining the potential hnctional
importance of these sites for regulation of GLP-2R gene transcription appear warranted.
The regional and tissue-specific localization of GLP-2R promoter-lacZ expression was
highly correlated with the expression of the endogenous murine GLP-2R, with the exception
of the pituitary gland and lung. The abundance olcelIs expressing the endogenous GLP-2R
appeared comparatively greater than the number of celIs expressing the GLP-2R promoter-
lacZ transgene in regions such as the hippocampus and cerebellum. These findings imply
that additional DNA regulatoty sequences not present in the 1.5-kb GLP-2R 5'-flanking
region are required to correctly specib transgene expression in al1 cells and tissues
expressing the endogenous GLP3R receptor. Furthermore, the interpretation of the
localization data may be further complicated in that unlike the endogenous GLP-ZR, the
nuclear Lac2 reporter protein would not be transported ta sites distal from the neural nuclei
that transcnbed the GLP-2R promoter in vivo. Nevertheless, the excellent correlation
between gastrointestina1 tissues and CNS regions expressing both the endogenous GLP-2R
and the lac2 transgene suggest that putacive regulato~sequences encoded within the first
1.5-kb of the mouse GLP-2R prornoter may be use fur for hture studies directing transgenes
to specific GLP-2R' ce11 populations in the murine CNS and p t ,

in the GI tract, GLP-1 and GLP-2 exert both overlapping and distinct actions in the
regulation of nutrient absorption and glucose homeostasis (187). Although both GLP-1 and
GLP-2 inhibit gastric emptying and acid secretion, the mechanisms underlying the comrnon
actions of these peptides have not been delineated. Despite these overlapping actions
however, no previous studies have implicated a role for GLP-1 receptor signaling in the
regulation of GLP-2 action in vivo. Our data generated independently using either the GLP- 1
receptor antagonist exendin (9-39) or GLP-I R'- mice, clearly show that blockade or
disruption of GLP-1 signaling enhances the sensitivity to GLP-2 action in the muine CNS.
Given the overlapping actions and expression patterns of GLP-1 and GLP-2 in peripheral
tissues and brain, it seems reasonable to search for additionaI interactions of GLP-1 and
GLP-2 receptor signaling systems in the control of shared physiologicaI actions such as
gastric emptying or acid secretion (74,76).
Following the initial description of GLP-1 action in the rodent CNS as a satiety factor,
multiple additional actions for GLP-1 in the CNS have emerged, including regulation of
hypothalamic pituitary tünction (139, 161, 165, I76), modulation of the extent of brain injury
(170), and transduction of the CNS response to aversive stimulation (15 1,220). Our data
demonstrating expression of the GLP-2R in multiple regions of the rodent CNS are
consistent with previous findings dernonsmting extensive projections of GLP-1 and GLP-2-
immunopositive nerve fibers to comparable regions of the rat brain (127). Taken together,
the avaiIable data clearly implies additional potential roles for CNS GLP-2, beyond
hypothalamic regulation of food intake, that ment careful anaIysis in hture studies.

CHAPTER FOUR:
ONTOCENY OF THE GLUCAGON-LIKE PEPTIDE-2
AND CLUCACON-LUCE PEPTIDE-2 RECEPTOR AXIS
IN THE DEVELOPING RAT CENTRAL NERVOUS SYSTEM.

SPECLFIC A i M S OF RESEARCH
The observation of widespread expression of the GLP-ZR throughout the adult rodent
CNS as detaiied in Chapter Three, coupled with the demonstrated functional integrity of the
GLP-2/GLP-2R axis in the neonatal rat intestine as described in Chapter Two, suggested to
us that the GLP-UGLP-2R axis might be present in the developing rodent CNS. Taken
together, Ive hwothesized thar rhe GLP-2/GLP-2R mis is mesent and fi~ncrionallv intact in
the rodent CIVS during develo~menf.
4.1 RESEARCH SUMMARY
in the central nervous system, the proglucagon-derived peptides, GLP-1 and GLP-2, are
synthesized in the brainstem and hypothalamus. Recent studies in the adult rat suggest that
both GLP- 1 and GLP-2 act as satiety factors when applied centrally. We have dernonstrated
that the GLP-2 receptor is expressed not only in the hypothalamus, but also throughout the
adult rodent CNS including hypothalamic, thalamic, cortical, arnygdaloid, hippocampai and
cerebellar nuclei. Although the GLP-2fGLP-2 receptor axis is present and functionaIIy intact
in the developing rat gastrointestinal tract, whether a central GLP-ZGLP-2R avis is present
during development remains unclear.
We assessed the expression of GLP-2 using radioimrnunoassay (RIA) and high pressure
liquid chromatography (HPLC) in the developing rat brainstem and hypothalamus. We aIso
studied GLP-2 receptor expression using semi-quantitative RT-PCR in the cortex,
hippocampus and cerebellum of fetal, early post natal and adult rats. Immunoreactive GLP-2
was detected in the brainstem and hypothalamus throughout development, including prenatal
(E 19) and early post-natal (PN O,2,4,8,IO, 16) time points. The highest Ievels of GLP- in
the brainstem were detected in fetal tissues (P<0.001 vs. al1 other time points), whereas in the
hypothalamus, immunoreactive GLP-2 IeveIs increased steadily during earIy neonatal t h e

points with peak levels occurring in older animais (Day 42 and adults) (PcO.01 vs. E19/20 or
Day O). To determine the molecular identity of GLP-2 in the brain, we performed HPLC and
RIA analysis and found that both GLP-2IJ3 and GLP-2'" wcre present in fetal and adult rat
brain extracts, GLP-2 receptor mRNA transcripts were detected in both fetal and e d y
neonatal cortex, hippocampus and cerebellum and were most abundant at post natal days 10-
16. Furthemore, GLP-2 stimulateri increases of CAMP in neonatal rat brainstem dispersion
cultures, suggesting that at least in some regions of the CNS the GLP-YGLP-2R axis may be
biologically active during brain development.
4.2 INTRODUCTlON
The cDNA encoding the proglucagon gene in the brain was originalIy isolated From a
human neonatal brainstem cDNA library (129). [n the fetai rat CNS, proglucagon is also
expressed in the hypothalamus (134, 135). Proglucagon processing in the adult rat brain is
similar to that of the intestine, generating glicentin, oxyntomodulin and the çlucagon-like
peptides (135). Studies examining proglucagon processing in the fetal brain however, have
detected both glicentin (oxynotomodu~in) and small arnounts oCgIucagon, suggesting that in
the fetal rat brain, proglucagon undergoes an intermediate profile of pancreatic and intestinal
processing ( 135).
In the adult rat bnin, immunoreactive GLP-L and the GU-1R receptor are present (127,
128) throughout the CNS. Similarly, GLP-2 receptor expression has been detected by RT-
PCR analysis at multiple sites in the CNS (237). UnIike the recent studies examining the
closely related VIP (vasoactive intestinal peptide) (238), and PACAP (pituitary adenyIate
cyclase activating polypeptide) (239-24 1) receptors during brain deveIopment, the central

actions of GLP-I and GLP-2 and their cognate receptors have not yet been examined during
neonatal brain development.
Given the recent interest in the role of glucagon-like peptides-1 and -2 in the central
control of food intake, and in Iighe of the expression of the GLP-2/GLP-2R axis in the
developing rat intestine, we determined whether a similar GLP-2IGLP-2 receptor axis is
present and biological1y functional during brain development.
Accordingly, we studied GLP-2 expression in the hypothalamus and brainstem by
radioimmunoassay. in order to detennine the molecular identity of the iR-GLP-2 species in
rat brain and to determine whether GLP-2 post-translationai processing differed throughout
development, we studied the molecular forrns of GLP-2 contt-ibuting to total IR-GLP-2 in
fetal and adult rat brainstem and hypothalamus via HPLC and MA analyses. To assess
relative GLP-2 receptor expression throughout early rat brain deveiopment and to deterrnine
whether there were spatial and age-specific differences in CNS GLP-2R expression, we
measured the retative Ievel of GLP-IR mRNA transcripts in developing rat conex,
hippocampus and brainstem. Furthemore, as an initial fiinctional assessment of the GLP-
2GLP-2R axis in the deveIoping rat brain, we tested whettier GLP-2 couId activate adenylate
cyclase in neural cells isolated from neonatal and fetal rat brain.
4 3 METHODS
4.3.2 ANIMALS
Pregnant fernales, lactating fernates with pups and aduit male Wistar rats were obtained
fiom Charles River Breeding Laboratories (St. Constant, QC, Canada). Animals were
housed in a climate-controlled roorn with a 12 hr Iightf 12 hr dark photoperiod. Food (Purina
rat chow) and tap water were fieely avaiIabIe. AnimaIs were allowed at least 3 days to

acclimatize before use. The first 24 hrs afier birth was designated as post-natal (PN) day 0.
4.3.2 CNS TISSUE DISSECTION
AduIt Wistar rats were euthanized by COz inhalation and quickly decapitated. For earlier
developmental time points, fetal and neonatal pups were decapitated. Brains were removed
and placed ventral side down on an ice-chilled g l a s plate. A coronal cut was made at the
posterior edge of the cerebral hemispheres and the cerebellum was removed from the
resulting hindbrain block by rnaking cuts along the cerebellar peduncles (cerebeilurn not
present in fetal sarnples). The hippocampus was removed fiom the rest of the forebrain by
cutting along the midsaggital fissure, to expose the underIying hippocampal formation. The
hippocampus was separated frorn the cortex. The cortex was blocked at the anterior and
posterior edge to avoid inclusion of olfactory bulb and amygdaloid tissue, respectively.
Samples were either imrnediately snap-frozen in liquid nitrogen and stored at -80°C until
processing for RNA isolation or processed imrnediately for RIA and HPLC analysis (see
beIow).
4.3.3 TISSUE PREPARA TION FOR RL4 AND HPLC ANAL YSES*
For RIA and HPLC analyses, tissues were homogenized in 1 N HCI containing 5%
HCOOH, 1% trifluoroacetic acid ( V A ) and 1% NaCl, followed by extraction of peptides
and small proteins by passage through a cartridge of Cl8 silica (SepPak, Waters Associates,
MiIford, MA). Plasma was collected into a 10% volume of TED (Trasylol, EDTA, Diprotin
A) and 2 volumes of 1% V A (pH 2.5 with diethylamine) was added ptior to extraction of
the peptides on C 18 silica (85, 199).
43.4 RW ANALYSIS*
For RIA andysis of IR-GLP-2 in developing rat brain, brainstem and hypothalamus

were isolated from rats at various developmentai time points. For fetal (embryonic day
19/20) rats, hypothalamus and brainstem were each pooled from 5 fetuses to make n=l. For
newborn rats (PN O), hypothalamus and brainstem were each pooled fiom 6-7 pups to make
n=L, while for PN 12 rats, hypothalamus and bninstem were each pooled fiom 3 pups to
make n=l. For PN 21 rats, hypothaIamus and brainstem were each pooled fiom 2 pups to
make n=l, and for PN 42 rats and aduIt rats, each hypothalamus and brainstem consisted of
an experimental sample, with 3 sampIes studied for each age group. GLP-2-related peptides
were detected using antisemm UTTH7, which detects the mid-sequence amino acids #25-30,
and thus, both GLP-p3 and the degraded form, G L P - ~ ~ " ~ , as well as prohormone precursors
of GLP-2 including proglucagon and the major proglucagon Fragment (85, 199). Tissue
protein levels were determined by Bradford assay and IR-GLP-2 (pg) was normalized to
protein (pg),
1.3.5 HPLC ANAL YSIS*
For HPLC separation of fetal rat brain extracts, hypothalami and brainstem were pooled
from 14 rat fetuses to rnake n= 1. For aduit samples, hypothalamus and brainstem were
isolated from a single brain to constitute one sample each. GLP-2-related peptides were
separated by high performance liquid chomatography using a C 18 donadapak column
(Waters Associates) with a 45 min linear gradient of 30 - 60% solvent B (solvent A = 0.1%
TFA in water; solvent B = 0.1% TFA in acetonitrile), followed by a 10 min purge wïth 99%
solvent B. The flow rate was 1.5 mumin and 0.5 ml fractions were collected (85, 199). Data
are expressed per hypothalamus or brainstem.
* Experiments primarily carried out by Dr. Patricia Brubaker (Professor, Depts. of
PhysioIogy and Medicine, University of Toronto).

4.3.6 RNA ISOLA TION AND SEMI-QUANTITA T N E R T-PCR
For embryonic day 16 or 19 samples, fetuses were rapidly removed fiom surrounding
tissues, quickly decapitated and hippocampi and cortex were removed. Each fetal
hippocampal or cortical sample consisted of tissue pooled From 2 fetuses to make n=l. For
a11 neonatal time points (PNO, PN2, PN4), a cerebellum was pooled from 2 pups to make
n= 1, hippocampi (both hemispheres) were pooled from 2 pups to make n=l, cortex (both
hemispheres) was collected frorn a single pup to make n=l . For older tirne points. (PN8,
PN10, PN16) each brain consisted of a single sample.
RNA was isolated h m rat brain tissues using TrizoITM reagent (Gibco BRL, Life
Technologies Inc., Toronto, Canada). RNA was quantified using a spectrophotometer and
RNA integrity was assessed by gel electrophoresis and ethidiuni bromide fluoresence.
Following quantification of intact RNA, 10 pg of RNA was used for first strand
complementary DNA (cDNA) synthesis. Superscnpt iiTM Reverse Transcriptase (GibcoBRL)
was used to generate first strand cDNA following DNasel (GibcoBRL) treaûnent of sample
RNA to remove genomic DNA contamination. RNA samples were then divided into hvo,
and one half (5 pg) was treated with reverse transcriptase (designated R n while it was
omitted from the other sample (5 pg) (designated RT) to control for non-specific
ampIification. To remove residuaI RNA following cDNA synthesis, al1 samples were treated
with ribonuclease H (MBI Fermentas, Vilnius, Lithuania) and stored at -80°C untiI
amptification. For PCR amplification 1-2 pl of first strand cDNA (RT" and RT) was
annealed with gene-specific primers, dNTPS, PCR b d e r and amplified using Taq
poIyrnerase (MBI Fermentas). For iinear amplification of rat GLP-ZR, cDNA was annealed
with the following primer pairs 1) 5'- TTGTGAACGGGCGCCAGGAGA-3' and 2) 5'-

GATCTCACTCTCTTCCAGAATCTC 3' for 22 cycles at 65OC. The expected PCR product
for rat GLP-2R is 1672 base pairs. For linear amplification of rat GAPDH (glyceraldehyde-
3-phosphate dehydrogenase) cDNA was annealed with the following primer pairs 1) 5'-
TCCACCACCCTGTTGCTGTAG-3' and with
2) 5'- GACCACAGTCCATGACATCACT 3' for 18 cycles at 60°C. The expected P CR
product for rat GAPDH is 452 bp. The linear range for PCR amplification of rat GLP-2R and
rat GAPDH was previousIy determined by plotting the PCR product yield against either the
cycle number or cDNA input amount (186) (242). Following PCR amplification, PCR
products were separated on a 1% (wtfvol) agarose gel by electrophoresis and transfened
overnight onto a nylon membrane (Genescreen, Life Technologies, Inc.). DNA was
permanently cross-linked to membranes by exposure to UV-light. Membranes were
hybridized with 1) a 3'~-labeled intemal cDNA probe for rat GLP-2R or 2) a 3?P-labeled
internai probe for rat GAPDH in an aqueous solution containing Formalin, SDS (sodium
deodecyl sulphate), sodium phosphate and EDTA overnight at SOC. 16-18 hours later,
membranes were washed at 60-65°C and exposed to a phosphoscreen (Molecular Dynamics,
[nc., Sunnyvale, CA) for visualization and densitometric analysis. Quantification of
hybridized PCR product density was determined using a Phosphoimager (Storm 840
Phosphoirnager, Molecular Dynamics,) and ImageQuant software (version 5.0, Molecular
Dynamics). Relative levels of the rat GLP-R were determined by dividing rat GLP-2R
signals by the corresponding rat GAPDH signals, which were amplified fiom the same
cDNA samples.
4.3.7 FE TAL AND NEONA TAL RAT BRAlN TISSUE EXPERlMENTS
For dispersion of neonatat rat brain tissues, a post-natal day 8 Wistar rat pup was

decapitated, and the brain was quickly removed ont0 an ice-chilled glass plate. Brainstem,
cerebellum, cortex, hippocampus and hypothalamus were dissected with a scalpel blade as
described above. Brain tissues were transferred to Eppendorf tubes on ice containing DMEM
(Dulbecco's Modified Eagle Medium)(Life Technologies, Inc.) supplemented with 100 pM
iBMX (Sigma Aldrich, Oakville, Canada) and a proteinase inhibitor, Aprotinin (~ ra s~ lo l@,
Bayer Inc., Etobicoke, Canada). Tissues were gentIy triturated with a 16 or 18 gauge needle,
and further diluted with medium. Prior to peptide treatment (n=l for each treatment), equal
0.3 ml samples of dispersed tissue were aliquoted to n= 16.
For fetal rat brain cultures, timed-pregnant Wistar rats (embryonic day 19 or 14) were
sacrificed by asphyxiation and fetuses were quickly dissected and washed in sterile ice-
chilled PBS (phosphate buffered saline). Fetal rat hypothalami (E19) were dissected by
scalpel blade in stenle conditions, and brainstem (E14) and neocortex (E14) were isolated by
microdissection under sterile conditions using a dissecting microscope.
Fetal rat hypothalarnic cells (€19) were isolated from -14-16 rat pups, pooled together
and cultured as previously described (134, 135,243) in 6-well plates. Prior to peptide
treatment 24 hours later (n=5-6 for each treatment), cells were washed in PBS and msiently
s e m starved 1-2 hours in DMEM.
For fetal rat brainstern and cortical cultures, brainstem and neocortex were microdissected
fiom -14-16 rat pups, finely triturated by surgical scissors in media and were fiirther purified
from cellular debns by enzymatic dissociation (Papain Dissociation System, Worthington
Biochemical Corp., Lakewood, NJ). FolIowing purification, El4 brainstem and cortical cells
were pIated at a density of 1 . 0 ~ 1 0 ~ cells per well in 50% Minimai Essential Medium (Giko
BRL) and 50% DMEM contaïning 10% FBS (fetal bovine s e m ) 2 m M @utamine,

penicilih (100 Ufml), and streptomycin ( IO pdrnl) in polyornithine coated 24-well culture
piates placed in a humidified s tede culture hood at 37°C and 5% CO?. Photomicrographs of
cultures were captured 44 and 66 hours after initial pIating. Pnor to peptide treatrnent (n=6
for each treatment) 72 hours Iater, cells were washed in PBS and transientiy serum starved
for 1-2 hours in DMEM alone.
4.3.8 PEPTIDES
Recombinant n t GLP-2'"3, and exendin (9-39) were purchased from Califomia Peptide
Research Inc. (Napa, CA). Forskolin and LBMX (3-isobutyl- I methyixanthine) were
purchased from Sigma Aldrich (St. Louis, MO, USA).
4.3.9 CAMP RIA of FETAL AND NEONA TAL RA T BRAIN TISSUES
To rneasure intracellular CAMP accumulation folIowing addition of various peptide
treatrnents to neonatal rat brain tissues and fetal rat bnin cultures, either rat GLP-2 (1 nM, 10
nM, 100 nM), n t exendin (9-39) (100 nM) alone or in combination with n t GLP-2 ( t O nM),
or forskolin (LOO PM) or medium alone was added. The dispersed neonatal brain tissues or
fetal brain cultures celis were then stimulated at 37°C for 20 minutes. Forlowing peptide
stimulation, reactions were terminated by the addition of 1 ml ofice-cold ethanol; medium
was colIected and stored at -80°C. To rneasure CAMP levels in media extracts, sarnples were
sheared with a 25-gauge needle and cellular debris was separated by centrifugation. Media
extracts ( I O pl of total) were vacuum-dried and CAMP concentrations were measured by
radioirnmunoassay (BTI, Stoughion, MA).
4.3.10 STATISTICS
For HPLC anaiysis, area-under-the-curve was determined as the sum of the
irnmunoreactivity under the peak. Statistical analysis was carried out using SAS (statistical

analysis software) or GraphPad Prism 3.00 (GraphPad Software Inc., San Diego, CA).
Differences between groups were determined by analysis of variance (ANOVA), and hrther
analysis was camed out using post-hoc comparisons.
4.4 RESULTS
RIA analysis of developing rat brain tissues indicated that small arnounts of IR-GLP-2
were present in both fetal (E19) and neonatal rat brainstem (1.68 k 0.09 pg/pg protein at
E 19/20) and hypothalamus (1 .O7 * 0.25 pgpg protein at E 19/20) as s h o w in Figure 4.1.
Within the hypothalamus, iR-GLP-2 levers were similar (1.32 k 0.32 pg/pg protein at PNO
vs. 2.06 k 0.72 pg/pg protein at PN21) throughout early fetal (E 19) and neonatal penods
(PNO-PN21) and peaked at post-natal day 42 (5.59k 0.60 pg/pg protein; P<O.O1 vs. E19120,
PNO).
Analysis of rat brainstem extracts indicated a markedly different trend of total
immunoreactive GLP-2, with the highest arnounts detected in fetal extracts (1.68 * 0.09
pg/pg protein; P~O.001 vs. al1 other time points), and relatively lower but similar amounts
present throughout neonatal and adulthood time points. Although previous studies (127)
have demonstrated that proglucagon neurons in the brainstem are capable of correctly
processing proglucagon to bioactive GLP-1 7-3i=16~mid~ , currently it is unknown whether these
same proglucagon-synthesizing neurons are capable of correctly processing proglucagon to
yield bioactive GLP-2'-j3. To determine the identity of the rnolecular species connibuthg to
total immunoreactive GLP-2, we studied developing and adult rat brainstem and
hypothalamus by HPLC and RIA analysis as presented in Figure 4.2. In fetal and adult rat
hypothalamic extracts, we detected biologicaIIy active, G L P - ~ ' - ~ ~ and its degradation
product, GLP'"). These findings suggest that proglucagon-expressing neumns in fetal and

*** Brainstem 1 1
E 19-20 PN O PN 12 PN 21 PN 42 I Adult 1
E19-20 PN O PN 12 PN 21 PN 42 Adult
Figure 4.1. Analysis of IR-GLP-2 throughout rat hypothalamic and brainstem developmenî. (a) Brainstem were removed fiom fetai, neonatai, weaned and adult male rats at diffèrent developmental time points, and extracts were analyzed for mid-sequence GLP-2 irnmunoreactivity and nomdized to protein (r-4)). (b) Hypothalami were also isolated fiom fetai (19-20), eady neonatai (PN 0, 12,21), weaned (PN42) and addt rats (A). AU smpIes were processed and IR-GLP-2 was measured by RIA and normaked to protein content. *P<O.OS, **P<O.OI, ***P<0,001 vs. PN 42 leveIs.

Petal Hypothalamus 3-33
5 4 3 2 -
1 O - ,- - -- 7 - - . - - v- - --- y - - - - - ., - - - - -- - --
68 69 70 71 72 73 74 75 76 Fractions
Adult Hypothalamus 1-33
Fractions Figure 4.2. MolecuJar forms of IR-GLP-2 in fetal or ndult rat hypothalamus by HPLC and RIA analysis. To determine the molecular idcntity of GLP-2 in the fetitl and adulr rat hypothalamus contributing IO total IR-GLP-2, hypothalami from fetal and adult rat brains were removed, processed and separated by HPLC. HPLC fractions wcrc thcn analyzed for IR-GLP-2 by RIA using C
an antibody recognizing mid-sequencc GLP-2. T h arroua indicatc the elution positions of synthetic rat GLP-2 (fraction 72) O W
and the prcdicted elution position of GLP-2 3-33 (fractian 74). Values are rcpresenlative of IR-GLP-2 per fractian pcr tissue.

adult hypothalamus have the necessary celIuIar machinery to correctly process proglucagon
to biologically active ~ ~ ~ - 2 l - j ~ .
Bath adult and fetal hypothalamus had relatively simiIar ratios of GLP-21J3: GLP~"). In
the rat brainstern, we also detected G L P - ~ ' - ~ ~ and GLP-L~-~ ' in fetal and adult brainstem
extracts, suggesting that proglucagon-expressing neurons in the rat brainstem are also
capable olliberating biologically active GLP-2Id3 from proglucagon throughout n t brain
developrnent (data not shown).
GLP-1 and GLP-2 actions are mediated through activation of their cognate heptahelical
transrnembrane G-protein coupIed recep tors ( 1 85,204). The spatial- and age-speci fic
expression of the glucagon and GLP-1 receptors in the murine brain has been previously
studied (205). These studies dernonstrated divergent regional and developmental expression
patterns for the expression of the gIucagon and GLP-1 receptor in the developing rodent
brain (205). In order to gain an understanding of the developrnental expression of the GLP-2
receptor within region-specific sites of the neonatal rat brain, we studied the expression of
GLP-2R mRNA transcripts relative to an interna1 control (GAPDH) using semi-quantitative
RT-PCR analysis of RNA from rat cerebelhm, cortex and hippocampus. In al1 CNS regions
exarnined, we detected GLP-îR mRNA uanscripts in fetal samples. The highest level of
GLP-2R expression was detected in the cortex, followed by the hippocampus and cerebellum
(Figure 4.3.). GLP-ZR mRNA transcripts were more abundant throughout early
cortical deveIopment in cornparison to &e relative leveis of GLP-2R expression present in
adult rat cortex. GLP-2R expression was aiso detected at al1 time points examined in the
developing rat hippocampus, including fetat, early neonatal and adult periods. Consistent
with the developing rat cortex and hippocampus, the GLP-2R was detected by RT-PCR


ONd +

analysis throughout rat cerebellar development. Amounts of relative GLP-2R expression in
the developing cerebellum appeared to be comparatively similar throughout al1 points
examined with peak expression occurring at post-natal day 16.
To ascertain whether the GLP-XLP-ZR axis is bioactive in the developing rat brain, we
tested the ability of GLP-2 to activate adenylate cyclase and induce an increase in CAMP
accumuIation in dispersed neonatal or cultured fetal neural cells. Forskolin treatment of
dispersed neonatal cerebellar (n=5), cortical (n=3), hypothalamic ( ~ 2 ) and brainstem (n=5)
neural cells resulted in increases in CAMP. Furthemore, we demonstrated (n=5) that GLP-2
(10 nM) consistently increased CAMP in cells isolated from PN 8 rat brainstem as presented
in Figure 4.4. In contrast, we did not detect a GLP-2-induced increase in CAMP in dispersed
neonatal cerebellar, cortical or hypothalamic neural cells. We therefore tested the specificity
of the GLP-2-induced increase in CAMP in the neonatal rat brainstem by CO-administering an
antagonist for the closely related GLP-1 receptor, exendin (9-39).
As indicated in Figure 44b, exendin (9-39) did not block the GLP-2-induced increase in
CAMP in neural cells isolated from PN 8 n t bziinstem, suggesting that the GLP-2
stimulation of CAMP was likely mediated through GLP-2R activation and not through the
closely related GLP-1R. As both GLP-2 and the GLP-2 receptor are expressed in the fetal rat
brain, we wanted to address whether the GLP-UGLP-2 receptor axis might be biologically
Functional in the fetal rat brain. We tested the effect of GLP-2 in a fetal (E19) rat
hypothalarnic culture system that was previously used to study proglucagon gene expression
in the fetal rodent brain (134, 135). Fetal (EI9) rat hypothalamic cultures responded to
forskolin stimuIation as indicated in Figure 4.5. however, there was no effect of GLP-2 (10
nM) on 1eveIs of CAMP.

PN8 Rat Brain 1 PN8 Rat Brain CI Control 1 10 nM GLP-2 1 100 pM Forskolin
Cerebellurn Brainstem Hypothalamus Cortex
4.4b. PN8 Rat Brainstem
U Control 10 nM GLP-2
m 10 nM GLP-2+ 100 nM Exendin (9-39)
a 100 FM Forskolin
U Control GLP-2 GW-2 + Forskolin
Exendin (9-39) Figure 4.4. E f k t of CLP-2 on c A i i accumuhtion in neonatai rat braia tissues, (a) Cerebellurn and brainstern were harvested and transiently dispersed in culture as described in Methou3 fiom neonatai rat brains on pst-natal &y 8. Following isolation. either GLP-2 (10 nM) or Forskolin (100 IiM) or media alone (control) was added, and cells were stimulated for 20 minutes at 3TC, reacrions were teminated by the addition of ice-cold ethanol. Media was extracted and CAMP was measured by RIA, CAMP values were expressed as mean k S.EM per 10 pl of tissue exmct Peptide treatments were repeated in quadniplicate and the values presented are representative of one of n=S urperiments. Pc0.05 vs. conml levels. (b) To determioc the pharmacologicai specificity of the GLP-2 induced incr~sse in cAMEJ in neonatal rat brainstem (PN8) we c~administered GLP-2 (IO nM) with the GLP-1R antagonist, exendin (9-39) (100 nM), and stimulated neonatal rat brainstem neural ceiis for 20 minutes at 37°C and subsequentiy rneasured the resuItant hcrease in CAMP per 10 pl of media extract via RIA analysis. The CAMP levels were expressed as the mean i SEM., peptide freatments were perfonned r 4 5 , and the values presented represent n=3 experiments. PC0.05 vs. conml levels-

We next assessed whether GLP-2 could increase CAMP in the fetal rat brainstem. We
therefore isolated neural cells from fetal rat brainstem (E14) as demonstrated in Figure 4.52.
Administration of GLP-2 did not increase CAMP in cultured fetal rat brainstem cells, whereas
administration of the potent CAMP secretagogue, forskolin, potently increased CAMP in
these cells.
Taken together, these studies suggest that the GLP-ZGLP-2R is biologically active within
at least one region of the neonatal rat brain and that GLP-2R signaling in the neonatal
brainstem is coupled to activation of adenylate cyclase.
4.5 DISCUSSION
Despite increasing awareness of the biological propenies of the glucagon-like peptides in
the central nervous system, the role(s) of these homones throughout brain development
rernains unexpIored. Although previous studies have primarily focused on glicentin,
oxyntomodulin (13 1, 135), and more recently the biosynthesis of GLP-1 in the brain (127,
128), currently there have been no studies examining the production and processing ofGLP-
2 in either the adult or fetal brain. Our finding that bioactive GLP-2"33 is processed correctly
and secreted by the fetal rat brainstem and hypothalamus, prornpted us to question whether
GLP-2 receptors might be coordinately expressed at similar tirne points duting early brain
development.
The region- and age-specific expression of the GLP-2 receptor lias not yet been examined
throughout brain development. In our present study, we detected GLP-2R mRNA
transctipts throughout the developing brain in both fetal and neonatal periods, nising the
possibiiity that GLP-2 may be bioIogicaIIy reIevant during brain development. As the
relative levels of GLP-2R expression in the neonatal cortex, hippocampus or cerebellum

4Sa. 8 1 El9 Rat Hypothalamus
~ S C . Conîrol 1 O n m GLP-2 Fonkolin
El4 Rat Brainstem - *
Bninabm (E14) Cmu (El4)
Figure 4.5 Efiect of CLP-2 treatmeat on CAMP accumulitior in cultured fetal rat hypotbalamic (E19), cortical (E14) and brainstem @14) ceUr (a) Hypothaiami were isolateci and pooled h m -14- 16 feial (E19) rats and ûansiently cuitured ovenùght in suppiemented DMEM at 37°C. 18 hom later, c e k were senim starved in DMEM alone for 1-2 hrs and ueated with either GLP-2 (IO nM) or Foskolin ( 100 PM) or media alone (contrai). FoUowing stimulation at 3PC for 20 minutes, reactions were stopped by ethanol addition, media was extracted and CAMP kvek were determined by radioirnmu~loassay. Ceik were treated with the different peptides to ir=5, and data are expresseci as mean * S-EM, n=l experiment. *P<O.OS vs control Ieveis. (b-c) Brainstern and neocortex were microdissected and p i e d h m h m 44-16 fetal rats (E14) under sterile conditions, neural cells were enzymaticaily dissociated. The appearance of in vitro neural ceus were show 40 k and 66 hr foiiowing isoiation (b). FoIIowing hansieut s e n i m starvation, corticai and bcanistem cuituces were maicd with either GLP-2 (1 nM. 10 KIM, or 100 nu), Forskoün (100 pM) or media alone (contd). Afkrpeptide stjmuiation of cells at 3PC for 20 minutes, ethanoi was added to terminare reactions and CAMP levek m cuiaire media was determined by RIA. AU values preseoted are mean * S-EM, all peptide treatmentr are n=4 for n=l exp&enL *PcO.OS vs, connoC Cevek.

appeared to change throughout brain development, GLP-2R signaling may be relevant during
cortical, hippocampal and cerebellar development. It is not possible to predict however,
whether these changes in gene expression are reflective ofspecific changes in the
transcriptional activation of the GLP-2R gendor stability of the GLP-2R mRNA transcript.
Tt is possible that the changes in GLP-2R expression in the neonatal rat brain might reflect
differences in the number of cells expressing the GLP-2R in the developing brain.
Although we detected rnRNA transcripts for both proglucagon and the GLP-2R
throughout the fetal and neonatal rat brain, the existing data precludes us from speculating on
the specific biological roles, if any, of GLP-2 throughout brain development. Interestingly,
we were unable to demonstrate an effect of GLP-2 treatment on CAMP accumulation in
isolated fetal hypothalamus, cortex or bninstem tissues. This preliminary analysis suggests
that the GLP-2GLP-2R axis may not be hct ional in regions of the fetal rat brain tissues,
Aiternatively, the Iack of response to GLP-2 stimulation in these fetal brain tissues could be
attributable to unsuitable tissue culture conditions used that are not optimal for GLP-2R
expression. It is ais0 possible that the GLP-UGLP-2 receptor axis is present in these feta1
tissues but is coupled to intracellular signaling systems other then adenylate cyciase.
Cunently we do not have a mode1 of disrupted GLP-2 receptor signaling or specific GLP-2
receptor antagonists, which might assist us in gaining an understanding of the physiological
relevance of this hormone during brain development.
Although we recently demonstrated the widespread distribution of the GLP-2 receptor
throughout the aduIt brain, whether these same or different celIs express the GLP-2R during
brain development rernains unknown. Our finding that GLP-2 increases CAMP in neural

cells transiently isolated h m neonatal rat brainstem, suggests that some regions of the
neonatal CNS may be responsive to GLP-2.
The development, differeotiation and ceilular remodeling of the central nervous system is
complex and not Fully understood. The ability to identiQ novel factors involved in this
process is therefore highly relevant. As we previously demonstrated that the neonatal
intestine responds to GLP-2 (2421, we questioned whether a GLP-2lGLP-2R wis might aIso
be present in the developing CNS. It is unclear as to whether GLP-2 inhibits food intake in
the neonate as demonstrated recently in the adult rat and rnouse. Furthemore. additional
studies are required to delineate whether the observed patterns ofGLP-2R expression in the
developing cortex, hippocarnpus and cerebellum have specific implications for bnin
development, or whether cenml GLP-2 networks are regulating actions in the developing
intestine. Nevertheless, the responsiveness to GLP-2 in ce1ls transiently isolated from the
neonatal rat brainstem, suggests that at least one region of the developing CNS likely has an
intact GLP-2lGLP-2 receptor mis that may be of physiological relevance dunng the
development of the central nervous system.

CHAPTER FIVE:
DISCUSSION

5.1 TEE ROLE(S) OF GLP-2 D W G LNTFSI'INAL DEVELOPMENT
As GLP-2 exerts trophic and cytoprotective actions in the adult intestine, and inhibitory
actions on gastric acid secretion and rnoti1ity in the aduh stornach, we wanted to examine the
GLP-UGLP-2 receptor axis throughout gastrointestinat deveIoprnent to determine if this ~ ~ i s
might be hnctionally coupled in the neonate. The results of our studies suggest that both
proglucagon and the GLP-2R are coordinately expressed in the fetal and neonatal intestine.
Furthermore, Our assessment using semi-quantitative RT-PCR suggests that in the neonate.
the relative expression of proglucagon and the GLP-2R may be increascd.
lVhat are the limitations of this studv?
To quanti@ the relative expression of the GLP-ZR (and proglucagon) at different
developmental tirne points in the rat bowel, we used semi-quantitative RT-PCR to compare
the amount of GLP-2R expression relative to the expression of a housekeeping gene,
GAPDH. Our results then, are dependent on the constant expression of GAPDH throughout
intestinal development. Whether GAPDH represents the optimal interna1 control gene for
studies of intestinal development remains unclear. To confirm whether our assessment of
relative GLP-2R expression during intestinal developrnent is accurate, it might be usehl to
compare the expression of the GLP-2R relative to the expression of other housekeeping
genes in addition to GAPDH. Furthermore, to eliminate the dependency on housekeeping
genes in our study, an interna1 competitor (cornpetitive RT-PCR) that is amplified using the
same oligonucleotide primer pairs as the GLP-2R could be used to quanti@ GLP-2R
expression.
In addition, quantification of specific mRiVA transcnpts by semi-quantitative RT-PCR
relies on the h e a r amplification of input cDNA- AIthough we examined the relationship

between PCR cycle number and input cDNA over a broad range for the amplification of rat
GLP-2R, proglucagon, and GAPDH mRNA transcripts in adult ïatestinal tissues, we did not
examine these same relationships in fetal or neonatal tissues. As a result, our assessment of
the relative abundance of GLP-2R and proglucagon rnRNA transcripts in fetal and neonatal
intestinal tissues may not have been exarnined under Iinear conditions.
Furthemore, as we sampled only the mid portion of the adult jejunum (5 cm) versus the
entire length of fetal and neonatal small bowel, the perceived differences in the relative
abundance of GLP-ZR mRNA transcripts throughout development might be a result of the
differential expression of this receptor throughout the length of bowel. Quantitative in-sitir
hybridization or in-sirit PCR could be used to mess regional differences in GLP-2R
expression throughout the length of the bowel. Moreover, our data only measures changes in
the relative amount of proghcagon and GLP-îR mRNA transcripts. As a consequence, the
data do not allow us to detennine whether the changes in the relative abundance of these
transcripts throughout intestinal d~velopment are a result of 1) specific transcriptional
activation of these genes, 2) increased mRNA stability of these transcnpts, or 3) an increase
in the number of celis expressing these genes. Furthemore, the data do not provide any
information regarding the expression of the protein products of these genes. Western blot
anaIysis and semi-quantitative immunocytochemistry could be used to ascertain the reIative
expression and distribution of GLP-2 and GLP-2R protein throughout gut developrnent.
Interestingly, in our study we found that the circulating levels of ~ ~ p - 2 ' ~ ~ ~ taken fkorn
post-natal (PN) day 12 rats were significantly increased relative to levels detected in the
plasma of older rats. The observed increases in plasma GLP-2 levels, however, may be
unrelated to GLP-2 action directly and could be a consequence of other factors. For

example, the relative increase in circulating GLP-2 rnight be a result of decreased clearance
rates in the neonate relative to the adult rodent resulting in increased levels of GLP-2 plasma
content.
Taken together, these data provide an approximation of the relative expression OP
proglucagon, and the GLP-2R throughout maturation of the rodent bowel. To ascertain the
precise levels of expression of these genes in a region-specific manner throughout intestinai
development, the suggestions provided above rnay facilitate a more thorough and quantitative
analysis.
The demonstration that GLP-2 and the GLP-2 receptor are expressed in fetaI and neonatal
bowel, suggests that this mis might be functional during development.
Wlrat mipht be the roles of CLP-2 dur in^ intestinal develo~rnent?
As GLP-2 is trophic in the adult rodent bowel, to determine whether this peptide has
simiiar effects in neonatal rats, we adrninistered exogenous GLP-2 to newborn rats for ten
days. We found that GLP-2 administration was coupied to growth in the neonatal rat
stomach, small and large bowel. Following our study, other groups have studied GLP-:!
action during intestinal development in neonatai piglets (244). In these studies, CO-infusion
of GLP-2 to preterm and tenn piglets maintained on total parenteral nutrition (TPN),
suppressed intestinal atrophy associated with TPN feeding through decreased proteolysis and
apoptosis (244).
Taken together, these studies demonstrate that the neonatal intestine is capable of
responding to exogenous GLP-2 administration and that GLP-2 treatrnent in the neonate is
coupled to gastrointestinal growth and decreased apoptosis.

As GLP-2 exerts actions unreIated to gastrointestinal growth in the adult bowel, GLP-2
might exhibit additional actions independent of growth in the neonatal bowel. Indeed, GLP-
2 might regulate gastrointestinal motility, gastnc acid secretion or epithelial integrity in the
developing intestine. As the regulation of these functions may be of even greater biological
significance during times of rapid growth and during the introduction of enteral nutrients, it
is not unreasonable to assume that GLP-2 couId be coordinating one or more of these actions
in the neonatal intestine.
Although we have found that GLP-2 is coupled to intestinal growth in the neonatal rat
bowel, whether GLP-2 exerts novel actions in the maturing rat intestine remains to be
determined. Inferences into novel GLP-2 action in the developing intestine might be gained
fiom DNA microarray or differential display studies. While our findings in the neonatal rat
intestine are interesting, the ontogeny of the GLP-YGLP-IR mis in the rat intestine can not
be directly correlated to human intestinaI development, as the timing of rat and hurnan
intestinal development differ significantly. As RiAs (85, 122) have been developed that
recognize hurnan GLP-2, and the cDNA sequence encoding the human GLP-2R is h o w n ,
analysis of the relative expression of the GLP-UGLP-2R axis in human neonates could be
initiated. As it was previousiy demonstrated that enteroglucagon levels are higher in both the
human fetal and neonatal circulation, we predict that the circulating levels of plasma GLP-2
may also be increased in the hurnan neonate (247,248).
Fially, an appreciation for the reIevance of GLP-2 action in gut development awaits the
generation of mice with genetic disntption of the GLP-2R gene or the development of GLP-
2R antagonists. As intestinaI development pmceeds normaIIy in mice deficient in

enteroendocrine production of proglucagon (217), it appears that the role(s) of GLP-2 in gut
development if any, rnay be compensated for by other factors.
5.2 TtiE ROLEIS) OF GLP-2 IN THE BRMN
Although the bioIogica1 actions of GLP-2 in the adult intestine have been studied, the
role(s), if any, of GLP-2 in the adult CNS has not yet been explored.
What are the action(s) of CLP-2 in the ceritral nervous svstem?
Coinciding with the completion of our studies, a recent report suggested that GLP-2, like
GLP-1, is a central regulator of feeding behavior (224). Central administration of GLP-2 (10
pg) to rats following an overnight fast or at the onset of feeding inhibits short-term food
intake (224). We demonstrated that central intracerebroventricular (ICV) administration of a
potent GLP-2 analogue, ~ [GI~ ' ] -GLP-~ , to mice inhibits food intake at the onset of dark
phase feeding but not following an overnight fast (237). As GLP-2 did not decrease
saccharin preference in a conditioned taste aversion assay (224), it seems unlikely that the
anorectic effects of GLP-2 on food intake are mediated through malaise or behavioral
suppression. As the distribution of GLP-2 immunoreactivity in the hypothalamic PVN and
AC nuclei corresponds identically with expression patterns described for the closely related
GLP-1 neuropeptide (224), it is possible that these two peptidergic systems are both
anatomically and fwictionally integrated. The anorectic effect(s) of GLP-2 in the CNS
however, occurs presumably through GLP-2 receptors expressed in the dorsomediai nucleus
of the hypothaIamus (D-MH) (224). NotabIy, centrai GLP-2 administration was found to
induce c-fos (an indicator of neuronal activity) expression in the caudaI part of the DMH, but
not the supraoptic (SON), paraventricular (PV) or the ventromedial (VM) nuclei of the

hypothalamus (224). Furthermore, unlike GLP- 1, central GLP-2 administration does not
inhibit water intake (224), suggesting that the anorectic action(s) of GLP-2 in the CNS is
independent of GLP- 1.
Taken together, these studies suggest a role for GLP-2 in the central regdation of short-
term food intake and appetite suppression. These studies however, are based upon the
injection of large amounts of peptide directly into the rodent CNS. Thus, whether these
observations will hold tme in other species, including humans, or with physiologicat doses of
peptide, rernain to be determined. Furthermore, whether the site(s) of GLP-2 action in the
hypothaIamus represents a potential pharmaceutical target for the treatment of feeding
disorders, such as obesity and anorexia, in humans is unknown, As o u initial studies
exarnined the exogenous administration of GLP-2 to rodents at supraphysiological levels, the
physiological relevance of centrally produced GLP-2 remains to be determined. The
generation of a GLP-2 receptor knockout mouse and or the development of GLP-2 receptor
antagonists wiII provide hrther insight into the importance of GLP-2 action on feeding
behavior and may facilitate the identification of novel actions of GLP-2 in the brain.
CVhm biolo~ical actioris. in addition to the centrai reaiilation o f food intake, could CLP-2
be merring iri the central riervous svstem?
At the outset of these thesis studies, it was ambiguous as to whether GLP-2R expression
was exciusively localized to the dorsornedial nucleus of the hypothalamus (224) or whether
the GLP-2R was more widely distributed in the C N S cocsistent with the distribution of the
closely related GLP-1R (226). Our current studies have demonstrated that similar to the
distribution of the GLP-IR, the GLP-2R is expressed throughout the rodent central nervous
system. We found widespread distribution of endogenous GLP-2R expression in

anatomically and functionally distinct areas including cortical, thalarnic, hypothalarnic,
amygdaloid, hippocampal and cerebellar nuciei, suggesting that GLP-2 might have broad
functions in the CNS, possibly as a neuromodulator or neurotransmitter, as h a previously
been suggested for GLP-1 (128).
We correlated our findings of endogenous GLP-ZR expression in the rodent C N S with our
GLP-2R promoter-IacZ transgenic model. There was excellent cellular correlation between
sites of endogenous GLP-2R expression with the sites of expression of a reporter gene in the
CNS of our GLP-2R promoter-lacZ transgenic model. Histological analysis suggested that
P-galactosidase-immunopositive cells in our GLP-ZR promoter-lacZ transgenic mice, were
less abundant than GLP-2R-immunopositive ceIls observed in the rat CNS. The observed
differences between reporter gene expression in Our transgenic model, and endogenous GLP-
2R expression are unlikely to be a consequence of decreased transcriptional activity as a
result of transgene integration, as multiple transgenic lines displayed similar patterns of
transgene expression in the CNS. Our results suggest that important regulatory elements
present in the murine GLP-ZR gene are absent fiom the transgene and as a result, endogenous
GLP-ZR expression patterns are not fuily recapitulated in our transgenic model.
Alternatively, the observed discrepancies between endogenous GLP-ZR expression and
reporter gene expression in our transgenic mice, might reflect differences in the sites of GLP-
2R mRNA expression versus sites of GLP-2R protein expression in the CNS.
The closely related hormone GLP-L has a number of actions in the CNS including
appetite suppression, mediation of interoceptive stress, blood pressure regulation, the
response to brain injury, and hippocampal activation. As the GLP-1 receptor is wideIy
expressed throughout the centrai nervous system in a pattern simiIar to the GLP-ZR, it not

unreasonable to assume that GLP-2 rnight share overlapping hc t ions with GLP- 1 in the
CNS, as it does in the periphery (e.g. regulation of gastric motility and gastric acid secretion).
To ascenain whechcr central GLP-2 administration promotes actions unrelated to appetite
suppression, there are a number of physiological and behavioral parameters that could be
examined following central GLP-2 treatrnent, including blood pressure levels, response to
brain injury, and the response to stress.
Is there a relationshi~ between GLP-I and GLP-2 receptor svstems in the brain?
The anorectic effect(s) of centrally applied GLP-2 on food intake was reported to be
resistant to 1 hr post-treatrnent with a potent feeding stimulator, neuropeptide Y (NPY) (224).
Unexpectedly however, in the same study, the effect(s) of GLP-2 on food intake was biocked
by CO-administration of the GLP-IR antagonist, exendin (9-39) (221), raising the possibility
that exendin (9-39) also acts as a GLP-2 receptor antagonist. This is the first report
suggesting that the specific GLP- 1 receptor antagonist exendin (9-39) can aIso block GLP-2
action. Considering that specific antagonists have not yet been defined for the GLP-2
receptor, we wanted to assess the specificity of exendin (9-39) for the GLP-2 receptor.
We demonstrated that CO-administration of exendin (9-39) with the GLP-2 analogue,
~ [ G I ~ ' ] - G L P - ~ did not block, but rather enhanced central anorectic GLP- action(s),
suggesting that exendin (9-39) is not a hnctional GLP-2R antagonist AIthough the
discrepancies between our studies and those of Tang-Christensen et al. (224) may be
explained in part by species-specific differences, differences in methodology or perhaps
differences between native GLP-2 and the ~ [ G ~ ~ ' ] - G L P - ~ analogue, our studies using celk
that stably express the GLP-2R clearly demonstrate that exendin (9-39) is not a
pharmacologicai antagonist of the GLP-2R

As either dismption of GLP-1R signaling via a receptor antagonist (exendin (9-39)) or
genetic disruption ( G L P - ~ R ~ - mice) enhanced the anorectic actions of GLP-2, Our studies
suggest the finctional integration of GLP-IR and GLP-2R systems in the CNS. As
GLP-IR-'- rnice exhibit normal feeding behavior and do not become obese; it is Our
hypothesis that functionally integrated GLP-2 responsive anorectic systems may be up
regulated and compensating in GLP-IR" mice. Since GLP-1 and GLP-2 share overlapping
actions in the stomach, it will be interesting to determine whether similar signaling
integration occurs between these two systems in the periphery.
Cleariy we are at an early stage in Our understanding of GLP-2 biology in the central
nervous system. The recent finding that central GLP-2 administration (224,237) inhibits
food intake, taken together with the finding of widespread distribution of the GLP-ZR
throughout the rodent brain (237), suggests that GLP-2 may have several hnctions in the
Ois, and provides incentive for further studies examining the biological roIe(s) ofGLP-2 in
the brain.
5.3 THE ROLE(S) OF GLP-2 DURING BRAIN DEVELOPMENT.
Although the spatial- and age-specific expression of the glucagon and GLP-1 receptors in
the murine brain has been previously studied (205), there have been no studies examining the
expression or the biological significance of the GLP-ZR in the developing neonatal rodent
brain. To determine the ontogeny of the GLP-2R a i s in the developing rodent brain we
examined GLP-ZR expression within fetal and neonatal rat cerebellum, cortex and
hippocampus using serni-quantitative RT-PCR We have previously demonstrated that this
method can be used to measure relative GLP-2R expression in the deveIoping rat intestine

(242) as well as in the adult hypothalamus and brainstem (186). The demonstration that there
is coordinate expression O~GLP-2"33 and the GLP-2R in the developing rat brain, raises the
possibility that the GLP-2/GLP-2R axis is hnctional during early brain development.
IMiat are the [inlitutions of this studv?
Considering the widespread distribution of the GLP-2R throughout the rat brain and the
region- and age-specific cellular remodeling in the neonatal rat CNS, we studied the relative
expression of the GLP-2R in tissues from various neural origins including the forebrain
(cortex and hippocampus) and hindbrain (cerebellum). As the detection of the relative
abundance of GLP-2R uanscripts in the developing rat brain was carried out using semi-
quantitative RT-PCR analysis, the same concems and limitations that were raised previously
in this discussion can be applied to this study.
This initial assessment of GLP-2R expression in the developing rat brain should not be
considered quantitative as each time point represents only a singIe sample. As RT-PCR is a
sensitive technique and single sarnples can be variable, an accumte assessment of the relative
expression of the GLP-2R in the developing rat brain may be gained by analyzing additional
samples at each time point.
As GLP-2 is produced in the rat brainstem and hypothalamus, it may be interesting to
evaluate the relative expression of the GLP-2R in these tissues. Considering the only
ascribed biological fiinction for GLP-2 in the brain reIates to the hypothalamus and the
regdation of food intake, exarnining the ontogeny oCGLP-2R expression in the neonatd
hypothalamus may provide ches as to whether GLP-2 could potentiaIIy regulate food intake
in the neonate.

Although our studies dernonstrate that celb isolated from neonatal rat brainstem, but not
tiom neonatal rat hypothalamus, cortex or cerebehrn respond to GLP-2, this does not
preclude a role for GLP-2 in developing cortical, cerebelIar or hypothalamic tissues. Our
inability to detect increases in CAMP upon GLP-2 treatment in these tissues could be due to
depietion of the limited number of GLP-2R expressing celis in these tissues during
mechanica1 dispersion of the tissues into culture. Alternatively, our cell culture conditions
may not have provided a suitable environment to support GLP-2R expression in these cell
types. These concerns rnay be further addressed by RT-PCR or Western blot analysis.
Furthemore, it is aIso possible that in the neonatal cerebellum, cortex and hippocampus,
GLP-2R signaling is not coupled to CAMP.
This study is a descriptive, prelirninary analysis of GLP-2R expression during brain
devetopment. Nevenheless, the finding of GLP-2R transcripts in fetaI and neonatal
cerebellum, cortex and hippocampus coupkd with the detection of GLP'-" in the fetal and
neonatai rat bninstem and hypothalamus, raises the possibiiity that a GLP-2/GLP-2R axis
may be present in the developing rat brain.
CVliat are the rulehl of GLP-2 in the d e v e l o ~ i n ~ brain?
AIthough our studies establish that cells isolated from the neonata1 rat brainstem respond
to GLP-~'-'' via increases in CAMP, we cm not make specific inferences as ta the biological
actions of GLP-2 in the deveIoping rat brain. Our initiai studies demonstrate the bct ional
integrity of the GLP-YGLP-2Raxis in celLs isolated from rat brainstem at post-natal day
eight. Whether more rigorous analyses of GLP-2R expression in fetal and neonatd CNS
tissues \vil1 reved the funcrional integrity of the GLP-UGLP-2R axis in tissues other than the
brainstem, rernains to be determined.

As GLP-2 stimulates growth in the adult intestine through increased crypt ce11
proliferation it is tempting to speculate that G L P - rnight exert similar trophic properties in
the neonatai CNS. in the adult intestine, GLP-2 also acts to increase intestinal mass through
the inhibition of cellular apoptosis, As it was recently suggested that endogenous GLP-L
plays a hnctional role in B-amyloid protein-induced apoptosis in the adult rat hippocampus
(170), it is not unreasonable to suggest that GLP-2, like GLP-1, might aIso be involved in
amyloid protein-induced ce11 death in the hippocampus. A role for GLP-2 in neurond
apoptosis during the remodeling of the neonatal CNS is also conceivable in Iight of the fact
that we readiiy detected GLP-2R transcripts in the fetal and neonatal rat brain.
Although our studies demonstrate GLP-2R expression in the neonatal CNS, we are unable
to predict whether this correlates with a biological role for GLP-2 during brain development.
Furthemore, whether the expression of the GLP-2/GLP-2R avis is critical for brain
development is unknown and awaits the development of GLP-2R antagonists and the
generation of mice with genetically disrupted GLP-ZR signaling. As mice with genetic
disruption of GLP-IR (GLP-IR'? signaling develop normally, it is unlikely that mice with a
nu11 mutation for the GLP-2R ( ~ ~ p - 2 ~ ~ 3 will exhibit abnormal brain development ifGLP-2
receptor systems are functionally integrated with GLP-IR systems. In order to fürther
investigate the biological actions of GLP-2 in the bnin, it will likely be necessary to perform
behavioral, physiological and pharmacological chaIIenges in GLP-;IR''- mice to unmask
specific phenotypes in the brains of these mice.

APPENDICES

APPENDIX ONE:
Copyright release agreement from Elsevier Science.


APPENDIX TWO:
Copyright release agreement from Munksgaard Scientific Journalr

. . ?erpitmlan gtantwl .by th ce rlaht MUN . ewnir. providu mmp1r9, JI* Ir glvrn to th. otlglnol iOwrc and -W..
d m Muniugwnl Im«nrtisnri ~ - ~ - E b a b O E 2 l 4 8 r u b l l ~ r L t d bpmm knmrrfI~ D=6QP=bml&
I Damark

APPENDIX THREE:
Copyright release agreement from Endocrinology.

Aug 31 O 1 04:41p Endocr inr
The mmumipt in questh is:

Copyright release agreement from The American Society for Biochernistry and Molecular Biology.

- .---. rtm cri#in aa r nui iyvpsl~ -:2l tfl-Pt-~nr

REFERENCES
BeU GI, Santerre RF, Mullenbach GT 1983 Hamster preproglucagon contains the
sequence of glucagon and two related peptides. Nature 3O2:7 16-7 18
Kieffer TJ, Habener JF 1999 The glucagon-like peptides. Endocr Rev 209376-9 13
Lund PK, Goodman RH, Dee PC, Habener JI? 1982 Pancreatic preproglucagon
cDNA contains two glucagon-related coding sequences arranged in tandem. Proc Natl
Acad Sci USA 79:345-349
Heinrich G, Cros P, Lund PK, Bentley RC, Habener JF 1984 Pre-proglucagon
messenger ribonucleic acid: Nucleotide and encoded amino acid sequences of the rat
pancreatic complementary deoxyribonucleic acid. Endocnnology 1 152 176-2 18 1
Lopez LC, Frazier ML, Su CJ, Kumar A, Saunders CF 1983 Mammalian
pancreatic preproglucagon contains three gIucagon-related peptides. Proc Natl Acad
Sci USA 805485-5489
Novak U, Wilks A, BueU G, McEwen S 1987 Identical mRNA for preproglucagon
in pmcreas and gut. Eur J Biochem I64:553-558
Baum J, Simmons Jr BE, Unger RH, Madison LL 1962 Localization of glucagon
in the alpha cells in the pancreatic islets by immunofluorescent techniques. Diabetes
11:371-374
Orci L, Bordi C, Unger RH, Perrelet A 1988 Glucagon- and glicentin-producing
cells. Glucagon- and glicentin-producing celis. Ln: Ghcagon I. edited by PJ Lefébre
Berlin. Springer, 1983, Chapter 457-79
Eissele R, Goke R, Willemer S, Harthus H-P, Vermeer H, Arnold R, Goke B
1992 Glucagon-like peptide 1 ceHs in the gastrointestinal tract and pancreas of rat,
pig, and man. Eur J Clin Invest 22:283-29 t
Irwin DM, Sivarajah P, 1998 Dog stomach and pancreatic proglucagons are
encoded by identical mRNA transcripts. 80th dnnual Meeting of the Endocrine
Society, June 21-24, New Orteans, LA, USA, p. 278, Abstr. 114, The Endocrine
Society Press, Bethesda MD
Drucker DJ, Brubaker PL 1989 Progiucagon gene expression is reguIated by a
cyclic AMP-dependent pathway in rat intestine. Proc Natl Acad Sci USA 86:3953-
3957

Drucker DJ, Jin T, Asa SL, Young TA, Brubaker PL 1994 Activation of
proglucagon gene transcription by protein kinase A in a novel mouse enteroendocrine
cell line. Mol Endocrinol8: 1646-1655
Reimer RA, McBurney MI 1996 Dietary fiber modulates intestinal proglucagon
messenger ribonucleic acid and postprandial secretion of glucagon-like peptide-1 and
insuIin in rats. Endocrinology 137:3948-3956
Lu F, Jin T, Drucker DJ 1996 Activation of proglucagon gene expression by
gastrin-releasing peptide in a mouse enteroendocrine cell line. Endocrinology
137:3710-3716
Hoyt EC, Lund PK, Winesett DE, Fuller CR, Ghatei MA, Bloom SR, Ulshen MR
1996 Effects of fasting, refeeding and intraluminal trïglyceride on proglucagon
expression in jejunum and iIeum. Diabetes 45:434-439
Tappenden KA, Thomson AB, Wild GE, iMcBurney MI 1996 Short-chain fatty
acids increase proglucagon and ornithine decarboxylase messenger RNAs after
intestinal resection in rats. J Parenter Enteral Nutr 20:357-362
Reimer RA, Field CJ, McBurney MI 1997 Ontogenic changes in proglucagon
mRNA in BB diabetes prone and normal rats weaned ont0 a chow diet. Diabetologia
401871-878
Reimer FU, Glen S, Field CJ, McBurney Mi 1998 Proglucagon and glucose
transporter mRNA is altered by diet and disease susceptibility in 30-day-old
biobreeding (BB) diabetes-prone and normal rats, Pediatr Res 44:68-73
Ulshen MH, Hoyt EC, Fuller CR, Ghatei MA, Bloom SR, Lund PK 1996
increased ileal proglucagon expression aAer jejunectomy is not suppressed by
inhibition ofbowel growth. Dig Dis Sci 41:677-683
Kreymann B, Chatei MA, Williams G, Bloom SR 1987 Glucagon-like peptide-1 7-
36: A physiological incretin in man. Lancet 2:1300-1304
Orskov C, Rabenhoj L, Wettergren A, Kofod H, Eiolst JJ 1994 Tissue and plasma
concentrations of amidated and glycine-extended glucagon-like peptide 1 in humans.
Diabetes 43:535-539
Ghatei MA, Uttenthal LO, Christofides ND, Bryant MG, Bloom SR 1983
MoIecuIar f o m of human enteroglucagon in tissue and phsrna: Plasma responses to

nutrient stimuli in health and in disorders of the upper gastrointestinal tract. I Clin
Endocrinol Metab 57:488-495
Herrmann C, Goke R, Richter Ç, Fehmann HC, Arnold R, Goke B 1995
GIucagon-like peptide1 and glucose-dependent insulin-releasing potypeptide plasma
levels in response to nutrients. Digestion 56:L 17-126
Elliott RM, Morgan LM, Tredger JA, Deacon S, Wright J, Marks V 1993
GIucagon-like peptide- l(7-36)amide and glucose-dependent insulinotropic
polypeptide secretion in response to nutrient ingestion in man: Acute post-prandial
and 24-h secretion patterns. .! Endocrinol 138: 159- 1 66
Rocca AS, Brubaker PL 1995 Stereospecific effects of fatty acids on proglucagon-
derived peptide secretion in fetal rat intestinal cultures. Endocrinology 1365593-
5599
R~berge JN, Brubaker PL 1991 Secretion ofproglucagon-derïved peptides in
response to intestinal luminal nutrients. Endocrinology 128:3 169-3 174
Sakurai H, Dobbs RE, Unger RH 1975 The effect of somatostatin on the response
of GLI to the intraduodenal administration of glucose, protein, and fat. Diabetologia
1 1 :427430
Brubaker PL 199 1 Regulation of intestinal proglucagon-derived peptide secretion by
intestinal regulatory peptides. Endocrinology 128:3 175-3 182
Roberge JN, Gronau K4, Brubaker PL 1996 Gasbin-releasing peptide is a novel
rnediator of proxima1 nutrient-induced proglucagon-derived peptide secretion kom
the distal gut. Endocrinotogy 1372383-2388
Rocca AS, Brubaker PL 1999 Role of the vagus nerve in mediating proximal
nutrient-induced glucagon- Like peptide- l secretion. Endocrinology 140: 1687-1694
Hussain MA, Lee J, Miller CP, Habener JF 1997 POU dornain transcription factor
brain 4 confers pancreatic a-cell-specific expression of the proglucagon gene through
interaction with a novel proximal promoter G1 eIement. Mo1 Ce11 Bi01 I7:7186-7194
Laser B, Meda P, Constant 1, Philippe J 1996 The caudal-related homeodomain
protein Cdx-13 regulates glucagon gene expression in islet cells. I Biol Chem
27 1 :28984-28994

Jin T, Drucker D J 1996 Activation of proglucagon gene transcription through a
nover promoter element by the caudal-related homeodomain protein cdx-U3. Mo1
Ce11 Bi01 16:19-28
Ritz-Laser B, Estreicher A, Klages N, Saule S, Philippe J 1999 Pax-6 and Cdu-2/3
interact to activate glucagon gene expression un the G 1 control eiernent. J Biol Chcm
274:41244132
Ritz-Laser B, Estreicher A, Gauthier B, Philippe J 2000 The paired homeodomain
transcription factor Pax- is expressed in the endocrine pancreas and transactivates
the glucagon gene promoter. J Biol Chem 275:32708-32715
Hussain MA, Habener JF 1999 GIucagon gene transcription activation mediated by
synergistic interactions of pax-6 and cd.2 with the p300 coactivator. J Bi01 Chem
273: 19945-19954
Dumonteil E, Laser B, Constant 1, Philippe J 1998 Differential regulation of the
glucagon and insulin I gene promoters by the basic helix-loop-helix transcription
factors E47 and BETA2. I Biol Chem 273: L9945-19954
Wang M, Drucker DJ 1995 The LIM domain horneobox gene isI-1 is a positive
regulator of idet cell-speci fiç proglucagon gene transcription. J Biol C hem
270: 12646- 12652
Ahlgren U, Pfaff SL, Jesseii TM, Edlund T, Edlund H 1997 Independent
requirement for ISL 1 in formation of pancreatic mesenchyme and islet cells. Nature
385257-260
Philippe J 1995 Hepatocyte-nuclear factor 3B gene transcripts generate protein
isofoms wvith different transactivation properties on the glucagon gene. Mo1
Endocrinology 91368-374
Diedrich T, Furstenau U, Knepet W 1997 Ghcagon gene G3 enhancer: Evidence
that activity depends on combination of an islet-specific factor and a winged helix
protein. Bi01 Chem 378:89-98
Knepet W, ChaGtz J, Fhbener .IF 1990 Transcriptional activation of the rat
gIucagon gene by the cyclic AMP-responsive element in pancreatic isIet cells- Mol
Ce11 Biol 1016799-6804

Miller CP, Lin JC, Habener JF 1993 Transcription of the rat glucagon gene by the
cyclic lZMP response element-binding protein CREB is modulated by adjacent
CREB-associated proteins. Mol Ce11 Bi01 13:7080-7090
Gajic D, Drucker DJ 1993 Multiple cis-acting dornains mediate basal and adenosine
3', 5'-monophosphate-dependent glucagon gene transcription in a rnouse
neuroendocnne ce11 line. Endocrinology 132: 1055- 1062
Schwaninger kI, Lux G, Blume R, Oetjen E, Hidaka H, Knepel W 1993
Membrane depolanzation and calcium infll~v induce glucagon gene transcription in
pancreatic islet cells through the cyclic AMP-responsive element. J Bi01 Chem
268:5 168-5 177
Drucker DJ, Philippe J, Mojsov S 1988 Proglucagon gene expression and
posttranslational processing in a hamster islet ce11 line. Endocnnology 123: 186 1-1867
Mojsov S, Heinrich G, Wilson IB, Ravauola M, Orci L, Habener JF 1986
Preproglucagon gene expression in pancreas and intestine diversifies at the Levet of
post-translational processing, J Bi01 Chem 261:11880-11889
Orskov C, Holst JJ, Knuhtsen S, Baldissera FGA, Poulsen SS, Nielsen OV 1986
Glucagon-like peptides GLP-1 and GLP-2, predicted products of the glucagon gene.
are secreted separately fiom pig small intestine but not pancreas. EndocrinoIogy
119:1167-1475
Rouille Y, Martin S, Steiner DF 1995 Differential processing of proglucagon by the
subtilisin-like prohormone convertases PC2 and PC3 to generate either glucagon or
glucagon-like peptide. J Bi01 Chern 270:26488-26496
Imin DM, Wong J 1995 Trout and chicken proglucagon: Alternative splicing
generates mRNA transcripts encoding glucagon-iike peptide 2. Mol Endocrinol
9967-277
Irwin DM, Huner O, Youson JH 1999 Lamprey proglucagon and the origin of
glucagon-like peptides. Mol Bi01 EvoI 16: 1548-1557
Orskov C, Holst JJ, Poulsen SS, Kirkegaard P 1987 Pancreatic and intestinal
processing of proglucagon in man. Diabetologia 30:874-88 1

Drucker DJ, Lee YC, Asa SL, Brubaker PL 1992 Inhibition of pancreatic glucagon
gene expression in mice bearing a subcutaneous glucagon-producing GLUTag
transplantable tumor. Mol Endocrinol6:2 175-2 183
Ehrlich P, Tucker D, Asa SL, Brubaker PL, Drucker DJ 1994 Inhibition of
pancreatic proglucagon gene expression in mice bearing subcutaneous endocrine
tumors. Am J Physiol Endocrinol Metab 267:E662-E671
Tucker JD, Dhanvantari S, Brubaker PL 1996 Proglucagon processing in islet and
intestinal ce11 lines. Regul Pept 6229-35
Brubaker PL, Vranic M 1987 Fetal rat intestinal cells in monolayer culture: A new
in vitro system to study the glucagon-like immunoreactive peptides, Endocrinology
120:1976-1985
Brubaker P 1988 Control of glucagon-like immunoreactive peptide secretion Gom
fetrii rat intestinal cultures. Endocrinology 123:220-226
Brubaker PL, So DCY, Drucker DJ 1989 Tissue-specific differences in the levels
of proglucagon-derived peptides in streptozotocin-induced diabetes. Endocrinology
124:3003-3009
Brubaker PL, Lee YC, Drucker DJ 1992 Alterations in proglucagon processing and
inhibition of proglucagon gene expression in glucagon-SV40 T antigen transgenic
mice. J Biol Chem 267:20728-20733
Mojsov S, Kopczynski MG, Habener JF 1990 Both amidated and nonamidated
fonns of glucagon-like peptide I are synthesized in the rat intestine and the pancreas.
J Biol Chem 2653001-8008
Moore MC, Connolly CC, Cherrington AD 1998 Autoregulation of hepatic glucose
production. Eur J Endocrinol 138:240-248
Kawai K, Yokota C, Ohashi S, Watanabe Y, Yamashita K 1995 Evidence that
glucagon stimulates insulin secretion through its own receptor in rats. Diabetologia
3 8 1274-276
Samols E, Marri G, Marks V 1966 interrelationship of glucagon, insulin, and
glucose: The insuhogenic effect of glucagon. Diabetes 15:855-865

Drucker DJ, Ehrlich P, Asa SL, Brubaker PL 1996 induction of intestinal
epithelial proliferation by glucagon-like peptide 2. Proc Natl Acad Sci USA 93:7911-
7916
Veyrac M, Ribard D, Daures JP, Mion H, Lequellec A, Martinez J, BataiUe D,
Michel H 1989 Inhibitory effect of the C-terminal octapeptide of oxyntomodulin on
pentagastrin-stimulated gastric acid secretion in man. Scand J Gas troenterol24: 1238-
1242
Bado A, Cloarec D, Moizo L, Laigneau J-P, BataiUe D, Lewin MJM 1993
Neurotensin and oxyntomodulin-(30-37) potentiate PYY regulation of gasmc acid
and sornatostatin secretions, Am J Physiol Gastrointest Liver Physiol265:GI 13-G117
Jarrousse Cl Niel H, Audousset-Puech M-P, ~Martinez J, Bataille D 1986
Oxyntomodulin and its C-terminal octapeptide inhibit liquid meal-stimulated acid
secretion. Peptides 7:253-256
SchjoIdager BTG, Baldissera FGA, Mortensen PE, Holst JJ, Christiansen J 1988
Oxyntomodulin: A potential hormone from the distal gut. Pharmacokinetics and
effects on gasmc acid and insulin secretion in man. Eur J Clin tnvest 18:499-503
Thorens 5, Waeber G 1993 Glucagon-like peptide-1 and the control of insulin
secretion in the normal state and in NIDDM. Diabetes 42: 12 19- 1225
Drucker DJ, Philippe J, Mojsov S, Chick WL, Habener SF 1987 Glucagon-like
peptide 1 stimulates insulin gene expression and increases cyclic AMP levels in a rat
islet ce11 line. Proc Natl Acad Sci USA 84:3434-3438
Fehmann H-C, Habener JF 1992 Insulinotropic hormone glucagon-like peptide-I(7-
37) stimulation of proinsulin gene expression and proinsulin biosynthesis in
insulinoma BK-1 cells. Endocrinology 130: 159-L66
Xu G, Stoffers DA, Habener JF, Bonner-Weir S 1999 Exendin-4 stimulates both
beta-cell replication and neogenesis, resulting in increased beta-cell mass and
improved glucose tolerance in diabetic rats. Diabetes 48:2270-2276
Stoffers DA, Kieffer TJ, Hussain MA, Drucker DJ, Egan JM, Bonner-Weir S,
Habener JF 2000 Insulinotropic glucagon-Iike peptide-1 agonists stimulate
expression of homeodomain protein IDX-1 and increase B-ce11 mass in mouse
pancreas. Diabetes 49:741-748

Wojdemann M, Wettergren A, Hartmann B, Hilsted L, Holst JJ 1999 Inhibition
of sham feeding-stimulated human gastric acid secretion by glucagon-like peptide-2.
J Clin Endocrinol Metab 84% 13-25 17
Wettergren A, Wojdemann M, Meisner S, Stadil F, Holst JJ 1997 The inhibitory
effect of glucagon-Iike peptide-1 (GLP-1) 7-36 amide on gastric acid secretion in
humans depends on an intact vagal innervation. Gut 40597-60 1
Wojdemann M, Wttergren A, Hartmann B, Holst JJ 1998 Glucagon-like
peptide-2 inhibits centrally induced anûal motility in pigs. Scand J Gastroenterol
33:828-532
Gleeson MH, Bloom SR, Polak trm, Henry K, Dowling RH 1971 Endocrine
tumour in kidney affecting small bowel structure, motility, and absorptive fiinction.
Gut 12773-782
Stevens FM, Flanagaa RW, O'Gormm D, Buchanan KD 1984 Glucagonorna
syndrome demonstrating giant duodenal villi. Gut 25:784-79 1
Bloom SR 1972 An enteroglucagon tumour. Gut l3:52O-j23
Bloom SR, Polak JM 1983 The hormonal pattern of intestinal adaptation [a major
role for enteroglucagon]. Scand J Gastroenterol 17:93-103
Dmcker DJ 1994 Molecular pathophysiology of glucagon-SV40 T antigen
üansgenic mice. Am I Physiol Endocrinol Metab 267:E629-E635
Tsai C-H, WU M, Asa SL, Brubaker PL, Drucker DJ 1997 Intestinal g r o h -
promoting properties of glucagon-like peptide 2 in mice. Am I Physiol Endocrinol
Metab 273:E77-E84
Tsai C-A, Hi1 M, Drucker DJ 1997 Biological deteminants of intestinotrophic
properties of GLP-2 in vivo. Am J PhysioI Gastrointest Liver PhysioI 272:G662-
G668
Drucker DJ, Deforest L, Brubaker PL 1997 Intestinal response to growth factors
administered aIone or in combination with h[Gly2]-Glucagon-like peptide 2. Am 5
Physiol Gastrointest Liver Physiol273:G1252-GI262
Xiao Q, Boushey RP, Drucker DJ, Brubaker PL 1999 Secretion of the
intestinotropic hormone glucagon-Iike peptide 2 is differentiaIIy regulated by
nutrients in humans. Gastroenterolcgy L 17:99- 1 OS

Hartmann B, Johnsen AH, Orskov C, Adelhorst K, Thim L, Holst JJ 2000
Structure, measurement, and secretion of human glucagon-like peptide-2. Peptides
2 L:73-8O
Taylor RG, Verity K, Fuller PJ 1990 Ileal glucagon gene expression: ontogeny and
response to massive small bowel resection. Gastroenterology 99:724-729
Taylor RG, FuUer PJ 1994 Humoral regdation of intestinal adaptation. Baillieres
Clinical Endocrinology and Metabolism 8: 165- 183
Fischer KD, Dhanvantari S, Drucker DJ, Brubaker PL 1997 Intestinal growth is
associated with elevated levels of glucagon-like peptide-2 in diabetic rats. Am J
Physiol Endocrinol Metab 273:E8 15-E820
Thulesen J, Hartmann B, Nielsen C, Holst JJ, Poulsen SS 1999 Diabetic intestinal
growth adaptation and glucagon-like peptide 2 in the rat: effects of dietary fibre. Gut
45:672-678
Hansen L, Hartmann B, Bisgaard T, Mineo H, Jorgensen PN, Holst JJ 2000
Sornatostatin restrains the secretion of glucagon-like peptide-1 and -2 from isolated
perfused porcine ileum, Am J PhysioI Endocrinol Metab 2ïS:ElO 10-El0 18
Brubaker PL, Crivici A, h o A, Ehrlich P, Tsai C-H, Drucker DJ 1997
Circulating and tissue foms of the intestinal growth factor, glucagon-like peptide 2.
Endocrinology 138:48374843
Drucker DJ, Shi Q, Crivici A, Sumner-Smith M, Tavares W, Hill M, Deforest L,
Cooper S, Brubaker PL 1997 Regdation of the biological activity of glucagon-like
peptide 2 by dipeptidyl peptidase IV. Nature Biotechnology 15:673-677
Hartmann B, Harr MB, Jeppesen PB, Wojdemann LM, Deacon CF, Mortensen
PB, Holst JJ 2000 In vivo and in vitro degradation of glucagon-like peptide-2 in
hurnans. J Clin Endocrinol Metab 85:2884-2888
Hartmann B, Thulesen J, Kissow H, Thulesen S, Orskov C, Ropke C, Poulsen
SS, Eolst JJ 2000 Dipeptidyi peptidase IV inhibition enhances the intestinotrophic
effect of glucagon-Iike peptide-2 in rats and mice. Endocrinology 141:40 13-4020
Orskov C, Andreasen J, Holst JJ 1992 Al1 products of proglucagon are elevated in
plasma Eom uremic pahents. J C h Endocrino1 Metab 74:379-384

Ruiz-Grande C, Pintado J, Alarcon C, Castilia C, Valverde 1, Lopez-Novoa SM
1990 Renal catabolism of human glucagon-like peptides 1 and 2. Can J Physiol
Phannacol 68: 1568-1573
Tavares W, Drucker DJ, Brubaker PL 1999 Enzymatic and renal-dependent
catabolism of the intestinotropic hormone ghcagon-Iike peptide-2 in the rat. Am J
Physiol Endocrinol Metab 278:E 134-E 139
Dumoulin V, Moro F, Barcelo A, Dakka T, Cuber JC 1998 Peptide Y Y , glucagon-
like peptide-1, and neurotensin responses to Iuminal factors in the isolated vascularly
perfused rat ileum. Endocrinology 139:3780-3786
Wettergren A, Schjoldager B, Mortensen PE, Myhre J, Christiansen J, Holst JJ
1993 Truncated GLP- 1 (proglucagon 78- 107-amide) inhibits gastric and pancreatic
functions in man. Dig Dis Sci 38:665-673
Chance WT, Foley-Nelson T, Thomas 1, Balasubramaniam A 1997 Prevention of
parenteral nutrition-induced gut hypolasia by coinfusion of glucagon-like peptide2
Am 1 Physiol Gastrointest tiver Physiol273:G559-G563
Kato Y, Yu D, Schwartz MZ 1999 Glucagon-Iike peptide-2 enhances small
intestinal absorptive Function and mucosai mass in vivo. 1 Pediatr Surg 34: 18-20
Prasad R, Alavi K, Schwartz MZ 2000 Glucagon-like peptide-2 analogue enhances
intestinal mucosal mass after ischemia and reperfusion. J Pediatr Surg 32357-359
Chance WT, Sheriff S, McCarter F, Ogle C 200 1 Glucagon-like peptide-2
stimulates gut mucosal growth and immune response in burned rats. J Burn Care
Rehabil22: 136-143
Litvak DA, Hellmich MR, Evers BM, Banker NA, Townsend Jr CM 1998
Glucagon-like peptide 2 is a potent growth factor for small intestine and coIon. 1
Gastrointest Surg 2: 146-150
Brubaker PL, Izzo A, Hill M, Drucker DJ 1997 Intestinal Fiinction in mice with
small bowel growth induced by giucagon-like peptide-2. Am J Physiol Endocrinol
Metab 272:E 1050-E 1058
Cheeseman CI, Tsang R 1996 The effect of gastric inhiiitory polypeptide and
glucagon Iike peptides on intestinal hexose transport. Am 1 Physiol Gastrointest Liver
Physio127 133477-G482

Cheeseman C I 1997 Upregulation of SGLT-1 transport activity in rat jejunum
induced by GLP-2 infusion in vivo, Am J Physiol Regulatory Integrative Comp
Physio1273:R1965-RL971
Cheeseman CI, O'Neil D 1998 Basolateral D-glucose transport activity along the
crypt-villus axis in rat jejunum and upregulation induced by gastric inhibitory peptide
and glucagon-like peptide-2. Experimental Physiology 83:605-6 16
Fuller PJ, Beveridge DJ, Taylor RG 1993 Ileal proglucagon gene expression in the
rat: characterization in intestinal adaptation using in situ hybridization.
Gastroenterology 104:459-466
Scott RB, Kirk D, Maclriaughton WK, Meddings JB 1998 GLP-2 augments the
adaptive response to massive intestinal resection in rat. Am J Physiol Gastrointest
Liver Physiol275:G9 1 1 -G9Z 1
Sigalet DL, Martin GR 2000 Hormonal thempy for short bowel syndrome. J Pediatr
Surg 35360-363; discussion 364
Thulesen J, Hartmann B, Kissow H, Jeppensen PB, Orskov C, Holst JJ, Poulsen
SS 200 1 intestinal growth adaptation and glucagon-like peptide 2 in rats with ileal-
jejunal transposition or small bowe1 resection. Dig Dis Sci 46:379-388
Boushey RP, Yusta B, Drucker DJ 1999 GIucagon-like peptide 2 decreases
rnortality and reduces the severity of indomethacin-induced rnurine enteritis. Am J
Physiol Endocrinol Metab 277:E937-E947
Alavi K, Schwartz MZ, Palazzo SP, Prasad R 2000 Treatment of inflamrnatory
bowel disease in a rodent model with the intestinal growth factor glucagon-like
peptide-2. J Pediatr Surg 35347-85 L
Drucker DJ, Yusta B, Boushey RP, Deforest L, Brubaker PL 1999 Human
[Gly2]-GLP-2 reduces the severity of colonic injury in a murine model of
expenmental colitis. Am J Physiol Gastrointest Liver Physiol276:G79-G91
Schmidt PT, Hartmann B, Brengenholt S, Hoist JJ, Clasesson MH 2000
Deficiency of the intestinal growth factor, glucagon-like peptide 2, in the colon of
SCiD mice with inflamrnatory bowel disease induced by transplantation of CD4+
cells. Scand J Gastroenterol35522-527

Tavakkolizadeh A, Shen R, Abraham P, Kormi N, Seifert P, Edelman ER,
Jacobs DO, Zinner MJ, Ashley SW, Whang EE 2000 Glucagon-like peptide 2: a
new treatrnent for chemotherapy-induced enteritis. J Surg Res 9 1 :77-82
Boushey RP, Yusta B, Drucker DJ 200 1 Glucagon-Iike peptide-2 (GLP-2) reduces
chemotherapy-associated mortality and enhances ce11 swvivaf in cells expressing a
transfected GLP-2 receptor. Cancer Res 6 1:687-693
Chance WT, Sheriff S, Fotey-Nelson T, Thomas 1, Balasubramaniam A 2000
Maintaining gut integrity during parenteral nutrition of tumor-bearïng rats: effects of
glucagon-like peptide 2. Nutr Cancer 37:215-222
Benjamin MA, McKay DM, Yang P-C, Perdue MH 2000 Glucagon-like peptide-2
enhances epithelial banfer function of both transcellular and paracellular pathways in
the mouse. Gut 17: 1 12- 1 19
Xiao Q, Boushey RP, Cino M, Drucker DJ, Brubaker PL 2000 Circulating levels
of glucagon-like peptide-2 in hurnan subjects with infl ammatory boule1 disease. Am J
Physiol Regulatory Integrative Comp Physiol278:R1057-R 1063
Jeppesen PB, Hartmann B, Thulesen J, Hansen BS, Holst JJ, Poulsen SS,
Mortensen PB 2000 Eievated plasma giucagon-like peptide 1 and 2 concentrations in
ileum resected short bowel patients with a preserved colon. Gut 47:370-276
Jeppesen PB, Hartmann 8, Hansen BS, Thulesen J, Holst JJ, Mortensen PB
1999 Impaired meal-stimulated glucagon-like peptide-2 response in ileal resected
short bowel patients with intestinal failure. Gut 45559-563
Jeppesen PB, Hartmann B, Thutesen J, Graff J, Hansen BS, Tofteng F,
Lohmann J, Poulsen SS, Madsen JL, Holst JJ, Mortensen PB 2001 Treaûnent of
short bowel patients with glucagon-like peptide-2 (GLP-2), a newly discovered
intestinotrophic, anti-secretory, and transit-modulating peptide. Gastroenterology
120:806-8 15
Han VKM, Hynes MA, Jin C, Towle AC, Lauder JM, Lund PK 1986 Cellular
localization of proglucagon/glucagon-like peptide I messenger RNAs in rat brain. J
Neurosci Res 16:97-107

Larsen PJ, Tang-Christensen M, Holst JJ, Orskov C 1997 Distribution of
glucagon-like peptide- 1 and other preproglucagon-derived peptides in the rat
hypothalamus and brainstem. Neuroscience 77:257-270
Merchenthaler 1, Lane M, Shugbrue P 1999 Distribution of pre-pro-glucagon and
glucagon-like peptide1 receptor messenger RNAs in the rat central nervous system. J
Comp Neurol403:X 1-280
Drucker DJ, Asa S 1988 GIucagon gene expression in vertebrate brain. J Biol Chem
263: 13475- 13478
Lee YC, Campos RV, Drucker DJ 1993 Region- and age-specific differences in
proglucagon gene expression in the central nervous system of wild-type and
glucagon-simian virus-40 T-antigen transgenic mice. Endocrinology 133: 17 1 - 177
Lee YC, Brubaker PL, Drucker DJ 1990 Developmental and tissue-specific
regulation of proglucagon gene expression. Endocrinology 13722 17-2222
Efrat S, Teitelman G, Anwar M, Ruggiero D, Hanahan D 1988 Glucagon gene
regdatory region directs oncoprotein expression to neurons and pancreatic alpha
ceIls. Neuron i:605-6 13
Nian Ml Drucker DJ, lmin D 1999 Divergent regulation of human and rat
proglucagon gene promoters in vivo. Am J Physiol Gastrointest Liver Physiol
277:G829-(3837
Stobie-Hayes KM, Brubaker PL 1992 ControI of proglucagon-derived peptide
synthesis and secretion in fetal rat hypothalamus. NeuroendocrinoIogy 56:340-347
Lui EY, Asa SL, Drucker DJ, Lee YC, Brubaker PL 1990 GLucagon and related
peptides in fetal rat hypothalmus in vivo and in vitro. Endocrinology 126:llO-117
Brubaker PL, Stobie KM, Roberge JN, Lui EYT, Drucker DJ 1991 Progiucagon-
derived peptides in the neuroendocrine system. Adv Exp Med Bi01 29 1 : 143- 159
Stobie-Hayes K, Brubaker PL 1995 Role of glutamate in regulating hypothaiarnic
proghcagon-derïved peptide secretion in vitro. Life Sci 56: 1325- 133 I
Turton MD, O'Shea D, Gunn 1, Beak SA, Edwards CMB, Meeran K, Choi SJ,
Taylor CM, Heath MM, Lambert PD, Wiiding RH, Smith DM, Ghritei MA,
Herbert J, Btoom SR 1996 A roIe for glucagon-like peptide- t in the central
regdation of feeding. Nature 37959-72

Beak SA, Heath MM, Small CJ, Morgan DGA, Ghatei MA, Taylor AD,
Buckingham JC, Bloom SR, Smith DiM 1998 Glucagon-like peptide-1 stimulates
luteinizing hormone releasing hormone secretion in a rodent hypothalamic neuronal
cell line (GTl-7). J Clin Invest LOL:1334-1341
Sasaki H, Ebitani 1, Tominaga M, Yamatani K, Yawata Y, Hara LM 1980
Glucagon-like substance in the canine brain. Endocrinol Japon 1 : 135- 140
Dorn A, Rinne A, Bernstein H-G, Hahn H-J, Ziegler M, Dammert K 198 1
immunoreactive glucagon in neurons of various parts of the human brain.
Demonstration by imrnunofluorescence technique. Acta Histochem 69243-247
Watanabe A, Fujiwara M, Nagashima H 1986 Glucagon-like polypeptide and
insulin contents in the brain from acuee hepatic failure dogs. Res Exp Med 186203-
208
Jin S-LC, Han VKM, Simmons JG, Towle AC, Lauder JM, Lund PK 1988
Distribution of glucagon-like peptide 1 (GLP-l), glucagon, and glicentin in the rat
brain: An immunocytochemica~ study. J Cornp Neurol27 L:5 19-532
Graner JL, Abraira C 1985 Glucagon in the cerebrospinal fluid. N Engl J Med
3 I2:994-995
Tornqvist K, Loren 1, Hakanson R, Sundler F 198 1 Peptide-containing neurons in
the chicken retina. Exp Eye Res 3355-64
Amir S 1986 Central glucagon-induced hyperglycemia is mediated by combined
activation of the adrenal rnedulla and sympathetic nerve endings. Physiol Behav
37563-566
Billington CJ, Bartness TJ, Briggs J, Levine AS, Morley JE 1987 Glucagon
stimulation of brown adipose tissue growth and thermogenesis. Am J Physiol
Integrative Cornp Physiol252:RI60-Ri65
Atrens DM, Menedez JA 1993 Glucagon and the paraventncular hypothalamus:
modulation of energy balance. Brain Res 630:245-25 1
Inokuchi A, Oomura Y, Shimizu N, Yamamoto T 1986 Centra1 action of giucagon
in rat hypothalamus. Am I Physiol Regdatory Integrative Comp Physioi XO:Rl2O-
R126

Fuxe K, Anderssson K, Hokfelt, T., Mutt V, Ferland L, Agnati W, Ganten D,
Said S, Eneroth P, Gustafsson JA 1979 Localization and possible function of
peptidergic neurons and their interactions with central catechcolamine neurons, and
the central actions of gut hormones. Federation Proc 38:2333-2340
Van Dijk G, Thiele TE 1999 GIucagon-like peptide-1 (7-36) amide: a central
regulator of satiety and interoceptive stress. Neuropeptides 33:406-414
Meeran K, O'Shea D, Edwards CM, Turton MD, Heath LMM, Gunn 1, Abusnana
S, Rossi M, Small CJ, Goldstone AP, Taylor GM, Sunter D, Steere J, Cboi SJ,
Ghatei MA, Bloom SR 1999 Repeated intracerebroventricular administration of
glucagon-like peptide-1-(7-36) amide or exendin-(9-39) alters body weight in the rat.
Endocrinology 140:244-250
Flint A, Raben A, Astrup A, Holst JJ 1998 Glucagon-like peptide 1 promotes
satiety and suppresses energy intake in humans. J Clin Invest 10 1 5 15-520
Gutnviller JP, Drewe J, Coke B, Schmidt H, Rohrer 0, Lareida J, Beglinger C
1999 Glucagon-Like peptide-1 promotes satiety and reduces food intake in patients
with diabetes mellitus type 2. Am J Physiol Regulatory Integrative Comp Physiol
276:R1541-15144
Naslund E, Bogefors J, Skogar S, Gryback P, Jacobsson H, Holst JJ, Hellstrom
PM 1999 GLP-1 slows solid gastric ernptying and inhibits insulin, glucagon, and
PYY release in humans. Am J Physiol Regulatory Integrative Comp Physiol
277:Rg 10-9 16
Naslund E, Barkeling B, King N, Gutinak BI, Blundell JE, Holst JJ, Rosner S,
Hellstrom PM 1999 Energy intake and appetite are suppressed by glucagon-like
peptide-1 (GLP-1) in obese men. Int 1 Obes ReIat Metab Disord 23:304-3 11
Scrocchi LA, Brown TJ, MacLusliy N, Brubaker PL, Auerbach AB, Joyner AL,
Drucker DJ 1996 Glucose intolerance but normal satiety in mice with a nuIl
mutation in the glucagon-Iike peptide receptor gene. Nature Med 2: 1254-1258
Thiele TE, Van Dijk G, Campfield LA, Smith FJ, Burn P, Woods SC, Bernstein
H, Seeley RI 1997 Centra1 infusion of GLP-t, but not Ieptin, produces conditioned
taste aversion in rats. Am J Physiol Regulatory integrative Comp Physiol272:R726-
R730

Van Dijk G, Thiele TE, Seeley RJ, Woods SC, Berstein IL 1997 Glucagon-like
peptide- 1 and satiety. Nature 3852 14
McMahon LR, WeUman PJ 1998 PVN infusion of GLP-1-(7-36) amide suppresses
feeding but does not induce aversion or alter locomotion in rats. Am J PhysioI
Regulatory lntegrative Comp Physiol274:R23-R29
Larsen PJ, Tang-Christensen M, Jessop DS 1997 Central administration of
glucagon-like peptide-1 activates hypothalamic neuroendocrine neurons in the rat.
Endocrinology 138:4445-4455
Calvo JC, Gisolfi CV, Blazquez E, Mora F 1995 Glucagon-like peptide-l(7-
36)amide induces the release of aspartic acid and glutamine by the ventrornedial
hypothalamus of the conscious rat. Brain Res Bull 38:435-439
Seeley RJ, Blake K, Rushing PA, Benoit S, Eng J, Woods SC, D'Alessi0 D 2000
The role of CNS glucagon-like peptide-1 (7-36) amide receptors in mediating the
visceral illness effects of lithium chloride. J Neurosci 2O:l6 16-2 1
Rinaman L 1999 A functional role for central glucagon-like peptide-1 receptors in
lithium chloride-induced anorexia. Am J Physiol Regulatory lntegrative Comp
Physiol277:R1537-R1540
MacLusky N, Cook S, Scrocchi L, Shin J, Kim J, Vaccarino F, Asa SL, Drucker
DJ 2000 Neuroendocrine hnction in rnice with complete disniption of GLP-1
receptor signaling. Endocrinology 141:752-762
Gulpinar MA, Bozkurt A, Coskun T, Ulusoy NB, Yegen BC 2000 Glucagon-like
peptide (GLP-1) is involved in the central modulation of fecal output in rats. Am J
Physiof Gastrointest Liver Physiol278:G924-G929
Chowen JA, Roderiquez de Fonseca F, Alvarez E, Navarro M, Garcia-Sequra
LM, Blazquez E 1999 Increased glucagon-like peptide-1 receptor expression in glia
after mechanical lesion of the rat brain. Neuropeptides 33:212-215
Oka J-C, Coto T, Kameyama T 1999 Glucagon-like peptide-1 modulates neuronat
activity in the rat's hippocampus. Neuroreport 10:1643-1646
Oka CI, Suzuki N, Goto T, Kameyama T 1999 Endogenous GLP-1 modulates
hippocarnpal neuronal activity in Beta-amyloid protein-treated rats, Neuroreport
1 O:296 1-2964

Oka J, Suzuki E, Kondo Y 2000 Endogenous GLP-1 is involved in beta-amyloid
protein-induced mernory impairment and hippocampal neuronal death in rats. Brain
Res 878: 194-198
Barragan JM, Rodriguez RE, Blazquez E 1994 Changes in arterial blood pressure
and heart rate induced by glucagon-like peptide-1-(7-36 amide) in rats. Am 1 Physiol
Endocrinol Metab 266:E459-E466
Barragan JM, Rodriguez RE, Eng J, Blazquez E, Defiie LHK 1996 [nteractions
of exendin-(9-39) with the effects of glucagon-like peptide-1-(7-36) amide and of
exendin-4 on arterial blood pressure and heart rate in rats. Regul Peptides 67:63-68
Barragan JM, Eng J, Rodriguez R, Blazquez E 1999 Neural contribution to the
effect of glucagon-like peptide- 1-(7-36) amide on arterial blood pressure in rats. Am J
Physiol Endocrinol Metab 277:E784-E79 1
Bojanowska E, Stempniak B 2000 Effects of centrally or systemically injected
glucagon-like peptide-1 (7-36) amide on release of neurohypophysial hormones and
blood pressue in the rat. Regul Pept 91:75-8 1
O'Shea D, Gunn 1, Chen X, Bloom SR, Herbert J 1996 A role for central
glucagon-like peptide-l in temperature regdation. Neuroreport 7930-832
Beak SA, Small CJ, Ilovaiskaia 1, Hurley JD, Ghatei IMA, Bloom SR, Smith DM
1996 Glucagon-like peptide-1 (GLP-1) releases thyrotropin (TSH): characterization
of binding sites for GLP-1 on alpha-TSH cells. Endocrinology 137:4130-4138
Hoosein NM, Gurd RS 1984 Human glucagon-Iike peptides 1 and 2 activate rat
brain adenylate cyclase. FEBS Lea 178:83-86
Zueco JA, Esquifino AI, Chowen JA, Alvarez E, CastriIlon PO, Blazquez E 1999
Coexpression of glucagon-like peptide-1 (GLP-1) receptor, vasopressin, and oxytocin
mRNAs in neurons of the rat hypothalarnic supraoptic and paraventricular nuclei:
effect of GLP-l(7-36)mide on vasopressin and oxytocin reiease. J Neurochem
72: 10-16
Blazquez E, Aivarez E, Navarro M, Roncero 1, Rodriguez-Fonseca F, Chowen
JA, Zueco JA 1998 Glucagon-like peptide-1 (-36) amide as a novel neuropeptide.
Mol Neurobiol [8:157-173

Imeryuz N, Yegen BC, Bozkurt A, Coskun T, Villanueva-Pennacarrillo ML,
üiusoy NB 1997 Glucagon-like peptide-1 inhibits gastric emptying via vagal
afferent-mediated central mechanism. Am 1 Physiol Gastrointest Liver Physiol
273:G920-G927
Hamm HE 2001 How activated receptors couple to G proteins. Proc Natl Acad Sci
USA 98148 19-43 12
Bockaert J, Pin J P 1999 MolecuIar tinkering of G-protein coupled receptors: an
evolutionary success. EMBO J 18: 1723- 1729
Shenvood NM, Krueckl SL, McRory JE 2000 The origin and function of the
pituitary adenylate cyclase-activating polypeptide (PACAP)/glucagon superfamily.
Endocr Rev 2 1 :6 19-670
Xiao Q, Jeng W, Wheeler MB 2000 Characterization of glucaon-like peptide-1
receptor-binding determinants. J Mol Endocrino1 25:32 1-335
Munroe DG, Gupta AK, Kooshesh P, Rizkalla G, Wang H, Demchyshyn L,
Yang Z-J, Kamboj RK, Chen H, McCallurn K, Sumner-Smith hl, Drucker DJ,
Crivici A 1999 Prototypic G-protein coupled receptor for the intestinotrophic factor
glucagon-like peptide 2. Proc Natl Acad Sci USA 96: 1569-1573
Yusta B, Huang L, Munroe D, WolffG, Fantaske Ft, Sharma S, Demchyshyn L,
Asa SL, Drucker DJ 2000 Enteroendocrine localization of GLP-2 receptor
expression. Gastroenterology 1 19:744-755
Drucker DJ 1998 The glucagon-like peptides. Diabetes 47: 159- 169
Yusta B, Somwar Et, Wang F, Munroe D, Grinstein S, U p A, Drucker DJ 1999
Identification of glucagon-Iike peptide-2 (GLP-2)-activated signaling pathways in
baby hamster kidney frbrobfasts expressing the rat GLP-2 receptor. J Bi01 Chem
274:30459-30467
Yusta B, Boushey RP, Drucker DJ 2000 The glucagon-like peptide-2 receptor
mediates direct inhibition of cellular apoptosis via a CAMP-dependent protein kinase-
independent pathway. J Biol Chem 27535345-35352
Portois L, Maget B, Tastenoy M, Perret J, Svoboda M 1999 Identification of a
gIucose response element in the promoter of the rat glucagon receptor gene. J Bi01
Chem 2748 18 1-8 190

Geiger A, Decaux JF, Burcelin R, Le Cam A, Salazar G, Charron MJ, Girard J,
Kervran A 2000 Structural and functional characterizations of the 5'-flanking region
of the mouse glucagon receptor gene: compa&m with the rat gene. Biochem
Biaphys Res Commun 16:9 13-92 1
Buggy J, Hull J, Yoo-Warren H 1995 Isolation and suucturd analysis of the 5'
flanking region of the gene encoding the hman glucagon receptor. Biochem Biophys
Res Commun 208:339-344
Witmen A, Walkenbach P, Fuller PJ, Lankat-Buttgereit B, Goke R, Coke B 1998
The genomic organization of the human GLP-1 receptor gene. Exp Clin Endocrinol
Diabetes 106:299-302
Lankat-Buttgereit B, Coke B 1997 CIoning and characterization of the 5'-flanking
sequences (promoter region) of the human GLP-1 receptor gene. Peptides 18:6 17-624
Wildhage 1, Trusbeim H, Goke 6, Lankat-Buttgereit B 1999 Gene expression of
the human glucagon-like peptide-1 receptor is regdated by Sp 1 and Sp3.
Endocnnology 140:624-63 1
Galehshahi FS, Gake B, Lankat-Buttgereit B 1998 Contribution of a PSI-like
elernent to the tissue- and ceIl-specific expression of the human GLP-1 receptor gene,
FEBS Lett 336: 163-168
Drucker DJ 1999 Glucagon-like Peptide 2. Trends Endocrinoi Metab 10:153-156
Jin S-LC, Hynes MA, Lund PK 1990 Ontogeny of glucagon messenger RNA and
encoded precursor in the rat intestine. Regul Pept 29:117-13 1
Dhanvantari S, Seidah NG, Brubaker PL 1996 RoIe of prohormone convertases in
the tissue-specific processing of proglucap. Moi Endacrinol 10:342-355
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ 195 I Protein rneasurement with
the folin phenol reagent. J Biol Ctiem 193265-275
Chomczynski P, Sacchi N 1987 Single-step method of RNA isolation by acid
guanidinium thiocyanate-phenol-chlotoform extraction. Anal Biochem 162: 156- 159
Piechaczyk kt, Blanchard JM, Marty L, Dani C, Panabieres F, El Sabouty S,
Fort P, Jeanteur P 1984 Post-transcriptionai regdation of glyceraldehyde-3-
phosphate- dehydrogenase gene expression in rat tissues. Nucleic Acids Res t 2:69SI-
6963

Jelinek W, Lok S, Rosenberg GB, Smith RA, Grant FJ, Biggs S, Bensch PA,
Kuijper JL, Sheppard PO, Sprecher CA, O'Hara PJ, Foster D, Walker KM,
Chen LHJ, McKernan PA, Kindsvogel W 1993 Expression cloning and signaling
properties of the rat glucagon receptor. Science 259: 1614- 1616
Thorens B 1992 Expression cloning of the pancreatic B cell receptor for the gluco-
incretin hormone gIucagon-like peptide 1. Proc Nat1 Acad Sci USA 89:863l-8645
Campos RV, Lee YC, Drucker DJ 1994 Divergent tissue-specific and
developmental expression of receptors for giucagon and glucagon-like peptide-1 in
the mouse. Endocrinology lM:2 156-2 164
Ktona A, Girard JR, Kinebanyan MF, Picon L 198 1 Hyperglycaernia induced by
glucose infusion in the unrestrained pregnant rat during the last three days of
gestation: metabolic and hormonal changes in the rnother and the fetuses.
Diabetologia 2 1569-574
Kreymann B, Ghatei MA, Domin J, Kanse S, Bloorn SR 1991 Developmentai
patterns of glucagon-like peptide-1-(7-36) amide and peptide-YY in rat pancreas and
gut. Endocrinology 129: LOO 1-1005
Burrin DG, Stoll B, Jiang R, Chang X, Hartmann B, Holst JJ, Greeley CH, Jr.,
Reeds PJ 2000 Minimal enteral nutrient requirements for intestinal growth in
neonatal piglets: how much is enough? Am J Clin Nutr 71:1603-16 10
Vinicor F, Higdon G, Clark JF, Clark CM, Jr. 1976 Development of glucagon
sensitivity in neonatal rat liver. J Ctin invest 58571-578
Aiumets J, Hakanson R, Sundler F 1983 Ontogeny of endocrine cells in porcine gut
and pancreas. Gastroenterology 85: 1359-1372
Upchurch BH, Fung BP, Rindi C, Ronco Cl Leiter AB 1996 Peptide YY
expression is an early event in colonic endocrine ceII differentiation: evidence frorn
normal and transgenic mice. Development 122: 1 157- 1 163
Roth KA, Kim S, Gordon SI 1992 Immunocytochemical studies suggest two
pathways for enteroendocnne ce11 differentiation in the colon. Am J Physiol
Gastrointest Liver Physiol263:G174-G180
Brubaker P 1987 Ontogeny ofglucagon-like immunoreactive peptides in rat
intestine. Regul Peptides 17:3 19-326

Rothenberg ME, Eilertson CD, Klein K, Zhou Y, Lindberg 1, McDonald JK,
Mackin RB, Noe BD 1995 Processing of mouse proglucagon by recombinant
prohormone convertase 1 and immunopurified prohormone convertase 2 in vitro. J
Bi01 Chem 270: 10136-10146
Rothenberg ME, Eilertson CD, Klein K, Mackin RB, Noe BD 1996 Evidence for
redundancy in propeptidelprohormone convertase activities in processing
proglucagon: An antisense study. Mol Endocrinol 10:33 1-331
Lee YC, Damholt AB, Billestrup N, Kisbye T, Galante P, mi ch el sen B, Kofod H,
Nielsen JH 1999 Developmental expression of proprotein convertase 113 in the rat.
Mol Cell Endocrinol 15527-35
Hill MF, Asa SL, Drucker DJ 1999 Essential requirement for Pax6 in control of
enteroendocrine proglucagon gene transcription, Mol Endocrinol 13: 1474-1486
Montgomery RK, Mulberg AE, Grand RJ 1999 Development of the human
gastrointestinal tract: hventy years of progress. Gastroenterology 1 16:702-73 1
Grosfeld JL, Cheu H, Schlatter M, West KW, Rescorla FJ 1991 Changing trends
in necrotizing enterocolitis. Experience with 302 cases in two decades. Ann Surg
3 13:300-6: discussion 306-307
Seeley RJ, Woods SC, Dt.-essio D 2000 Targeted gene disruption in endocrine
research: The case of GLP-1 and neuroendocrine fiction. Endocnnology 14 1:473-
475
Rinaman L 1999 Interoceptive stress activates glucagon-like peptide-1 neurons that
project to the hypothalamus. Am J Physiol Regdatory Integrative Cornp Physiol
777:R582-R590
Toft-Nielsen MB, Madsbad S, Fioist JJ 1999 Continuous subcutaneous infusion of
glucagon-like peptide 1 lowers plasma glucose and reduces appetite in type 2 diabetic
patients. Diabetes Care 22: 1137-1 143
Szayna M, Doyle ME, Betkey JA, HoUoway HW, Spencer RG, Gteig NH, Egan
JM 2000 Exendin-4 decelerates food intake, weight gain, and fat deposition in
Zucker rats. Endocnnology 14 1: 1936-194 1

Tang-Christensen M, Larsen PJ, Thulesen J, Romer J, Vrang N 2000 The
proglucagon-derived peptide, glucagon-like peptide-2, is a neurotransmitter involved
in the regulation of food intake. Nat Med 6302-807
Tang-Christensen M, Larsen PJ, Coke R, Fink-Jensen A, Jessop DS, MoUer M,
Sheikh SP 1996 Central administration of GLP-1(7-36) amide inhibits food and
water intake in rats. Am J Physiol Regdatory Integrative Comp Physiol271:R848-
R856
White RB, Broqua Pl Meyer J, Junien J-L, Aubert ML 2000 Regional expression
of GLP-1 and GLP-2 receptors in the rat brain. 8 h d Annttal Meeting of the
Endocrine Sociefy, June 2 1-24, Toronto, Ontario, Canada, p.271, Abstr. 1 1 15, The
Endocrine Society Press, Bethesda, MD
DaCambra MP, Yusta B, Sumner-Smith BI, Crivici A, Drucker DJ, Brubaker
PL 2000 Structural determinants for activity of glucagon-like peptide-?.
Biochemistry 39:8888-8894
Scrocchi LA, Drucker DJ 1998 Effects of aging and a high fat diet on body weight
and glucose control in GLP- 1 R-1- mice. Endocrinology 139:3 127-3 132
Donahey JCK, Van Dijk G, Woods SC, Seeley RJ 1998 Intraventricular GLP-1
reduces short- but not long-term food intake or body weight in Lean and obese rats.
Brain Research 779:75-83
Thiele TE, Seeley RJ, D'Alessi0 D, Eng J, Bernstein IL, Woods SC, Van Dijk G
19% Central infusion of glucagon-like peptide- 1-(7-36) amide (GLP- 1) receptor
antagonist attenuates lithium chloride-induced c-Fos induction in rat brainstem. Brain
Res 801:164-170
Coke R, Fehmann H-C, Linn T, Schmidt H, Krause M, Eng J, Coke B 1993
Exendin-4 is a high potency agonist and truncated exendin-(9-39)-amide an
antagonist at the glucagon-Iike peptide 1-(7-36)-amide receptor of insulin-secreting
p-cells. J Bi01 Chem 268: 1965O-I9655
Alvarez E, Roncero 1, Chowen JA, Thorens B, Blazquez E 1996 Expression of the
gtucagon-like peptide-1 receptor gene in rat brain. J Newochem 66:920-927

Sivarajah P, Wheeler MB, Invin DM 2001 Evolution of receptors for proghcagon-
derived peptides: isolation of frog glucagon receptors. Comp Biochem Physiol
128:517-527
Traber PG, SiIberg DG 1996 Intestine-specific gene transcription. Annu Rev
Physiol58:275-297
Boylan MO, Jepeal LI, Jarboe LA, Wolfe MM 1997 Cell-specific expression o l the
glucose-dependent insulinotropic polypeptide gene in a mouse neuroendocrine turnor
ceII line. J Bi01 Chem 272: 17438-17443
Dusing MR, Brickner AG, Lowe SY, Cohen MB, Wiginton DA 2000 A
duodenum-specific enhancer regulates expression dong three axes in the small
intestine. Am J Physiol Gastrointest Liver Physiol279:G 1080-10893
Lovshin J, Estall J, Yusta E, Brown TJ, Drucker DJ 200 1 Glucagon-like peptide-2
action in the murine central nervous system is enhanced by elimination of GLP-1
receptor signaling. J Biot Chem 276:2 1489-2 1499
Karacay B, O'Dorisio MS, Summers hl, Bruce J 2000 Regulation of vasoactive
intestinal peptide receptor expression in developing nervous systems. Ann N Y Acad
Sci 931:165-174
D'Agata V, Cavallaro S, Stivala F, Canonico P 1996 Tissue-specific and
developmental expression of pitutitary adenylate cyclase-activating polypeptide
(PAPCAP) receptors in rat brain. Eur J Neurosci 8:3 10-3 18
Zhou CJ, Kikuyama S, Nakajo S, Arirnurn A, Shioda S 2000 Ontogeny of PACAP
receptor and its splicc variants in the nt brain. Ann N Y Acad Sci 92 i:375-376
Basile M, Vaudry D, Coulouarn Y, Jegou S, Lihrmann 1, Fournier A, Vaudry H,
Gonzalez BJ 2000 Distribution of PACAP receptor mRNAs and PACAP
binding sites in the rat brain during development. Ann N Y Acad Sci 921:304-307
Lovshin J, Yusta B, Iliopoulos 1, Migirdicyan A, Dableh L, Brubaker PL,
Drucker DJ 2000 Ontogeny of the glucagon-like peptide-2 receptor axis in the
deveIoping rat intestine. EndocrinoIogy 14 1 :4194-420 1
Zingg 8, Patel Y 1982 Biosynthesis of immunoreactive sornatostatin by
hypothalarnic neurons in culture. J Clin invest 70: 1 101-1 109

244. Burrrin DG, StoU B, Jiang R, Petersen Y, ElnifJ, Buddington RK, Schmidt M,
Holst JJ, Hartmann B, Sangild PT 2000 GLP-2 stimulates intestinal growth in
premature TPN-fed pigs by suppressing proteolysis and apoptosis. Am J Physiol
Gastro and Liver Physiol279:G 1249-1256
245 Frankün K and Paxinos G 1997 The Mouse Brain in Sterotaric Coordinates,
Academic Press, Inc., San Diego, CA
246 Benjamin MA, McKay DM, Yang PC, Cameron H, Purdue MJ3 2000 Glucagon-
like peptide2 enhances intestinal epithelial bamer Function of both transcellular and
paracellular pathways in the rnouse. Gut 47: 1 12-1 19.
247 Adrian TE, Soltesz G, MacKenzie 12, Bloom SR, Aynsley-Green A 1995
Gastrointestinal and pancreatic hormones in the human €etus and mother at 18-2 1
weeks of gestation. Bi01 Neonate 67:47-53.
248 Lucas A, Bloom SR, Green AA 1985 Gastrointestinal peptides and the adaptation to
extrauterine nutrition, C m 1 Physiol PharmacoI 63527-537.
249 Chatei IMA, Coodland RA, Taheri S, Mandir N, Brynes AE, Jordinson M,
Bloom SR 2001 Proglucagon-derived peptides in intestinal epithelial proliferation:
glucagon-like peptide-2 is a major mediator of intestinal epithelial proliferation in
rats. Dig Dis Sci 46: 175.5-1263.