Embed Size (px)
Citation preview

Vol. 147, No. 2JOURNAL OF BACTERIOLOGY, Aug. 1981, p. 563-568002 1-9193/81/080563-06$02.00/0
Origin of Haemophilus influenzae R FactorsRAINER LAUFS,* FRIEDRICH-CHRISTIAN RIESS, GERHARD JAHN, RUDIGER FOCK, AND
PAUL-MICHAEL KAULFERS
Institute of Medical Microbiology and Immunology, University of Hamburg, D 2000 Hamburg 20, FederalRepublic of Germany
Received 3 March 1981/Accepted 8 May 1981
The Haemophilus influenzae R plasmids specifying resistance against one,two, or three antibiotics which have emerged in different parts of the world wereshown to have closely related but not identical plasmid cores. The gene forampicillin resistance in the H. influenzae plasmid pKRE5367 is part of a trans-poson similar to Tn3, which was transposed from pKRE5367 onto RSF1010 inEscherichia coli. An indigenous H. influenzae plasmid (pW266) was isolated. Itsproperties correspond to those of the H. influenzae R plasmids, except for thepresence of a drug resistance transposon. The in vitro-generated H. influenzae Rplasmids carrying an ampicillin resistance transposon, a tetracycline resistancetransposon, and a transposon for combined tetracycline-chloramphenicol resist-ance resembled the natural isolates. The findings support the hypothesis that theR plasmids of H. influenzae are of multiclonal evolutionary origin.
The conjugative Haemophilus influenzae Rplasmids which have been studied are closelyrelated to one another and have most of theirbase sequences in common, independent of theirgeographical origins and their antibiotic resist-ance markers (4, 7). The sudden emergence ofthese R plasmids in different countries raises thequestion of how they originated. This paperexamines whether the H. influenzae R plasmidscould have arisen in independent genetic eventsas a result of the transposition of different re-sistance genes onto closely related indigenous H.influenzae plasmids.
MATERIALS AND METHODSBacterial strains. The bacterial strains and plas-
mids used in this study are listed in Table 1.Media. The liquid medium for H. influenzae con-
tained 3.5% (wt/vol) brain heart infusion (Oxoid Ltd.)supplemented with 10 pg of hemin ml-', 10 pg of L-histidine ml-', and 10 pg of NAD ml-'. Chocolate agarwas used as a solid medium, sometimes supplementedwith 10 to 30 ,ug of ampicillin or chloramphenicol ml-'.Cultures on solid media were incubated at 37°C in aCO2 incubator (CO2 concentration, 3 to 8% [vol/vol]).Cultures in broth were shaken in an air incubator at360C.
Conjugation. Exponential-phase cultures of thedonor strain were mixed 1:1 with the recipient strainand filtered on 0.4-pm membrane filters (MilliporeCorp.). The filters were incubated overnight on agarplates.
Transformation. The transformation methodused has been described (8).DNA-DNA duplex studies. The procedure fol-
lowed for DNA-DNA duplex studies has been reportedby us (7).
Isolation of unlabeled plasmid DNA. The pro-cedure used for isolation of unlabeled plasmid DNAhas been described in detail (7). Bacteriophage lambdaDNA was prepared according to Kleckner et al. (10).
Preparation of labeled plasmid DNA. Plasmidand phage DNAs were labeled to high specific radioac-tivities (>5 x 106 cps/min per pg) in vitro, using[3H]dTTP as the substrate and the nick translationactivity of Escherichia coli DNA polymerase I asdescribed by Rigby et al. (14).Cleavage of plasmid DNA. Restriction endonu-
cleases were obtained from Boehringer MannheimCorp. (West Germany). Digested plasmid DNA waselectrophoresed in 0.7% agarose. EcoRI-generatedfragments of lambda DNA were included in each gelas a molecular weight standard.Agarose gel electrophoresis of DNA. Cleared
lysates of bacterial strains were prepared, and sampleswere subjected to electrophoresis through a 0.7% aga-rose gel as reported by Meyers et al. (13). PlasmidsRldrd-19 (13), RP4 (1), Sa (13), RSF1010 (5), andColEl (13) were used as internal molecular weightstandards.DNA contour length and electron microscope
DNA heteroduplex and homoduplex analysis.The methods used for the studies with the electronmicroscope have been described by us in detail (7).
RESULTSDNA sequence homology between H. in-
fluenzae R plasmids isolated in differentcountries. H. influenzae R plasmids recentlyisolated in different countries were analyzed forcommon polynucleotide base sequences. Therelative DNA sequence homology withpKRE5367 (ampicillin resistance [Apr]) was de-termined by using the single-strand-specific en-
563
on Decem
ber 25, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

TABLE 1. Bacterial strains and their plasmidsMol wt of
Strain Plasmid present plasmid Resistance pattern Source/reference Country" of(lO6) origin
H. influenzaeHK539 pHK539 36 Tc Ap (TnI893) Kilian (9) DKHC234 pRI234 34 Tc Cm (Tn1894) van Klingeren et al. (16) NLKRE5367 pKRE5367 30 Ap (Tn1891) Laufs and Kaulfers (12) FRGFR16017 pFR16017 33 Tc (Tn1892) Kaulfers et al. (8) FRGLU121 pLU121 31.5 Tc Kaulfers et al. (8) FRGHi706 pHi7O6 38 Tc Cm Ap Braveny et al. (2) FRGBS6 pBS6 36 Tc Ap This laboratory PLVe7558 pVe7558 29 Ap This laboratory FRGW266 pW266 27 _b This laboratory FRG
"DK, Denmark; NL, Netherlands; FRG, West Germany; PL, Poland.b-, None.
donuclease Si (3) (Table 2). Each of the twotetracycline resistance (Tcr) -specifying plasmidspFR16017 and pLU121 shared 70% of their basesequences with pKRE5367, but when comparedwith each other they had only 70% of theirpolynucleotide sequences in common.The R plasmids pFR16017 (Tcr), pKRE5367
(Apr), and pVe7558 (Apr) were compared byrestriction enzyme analysis with the enzymes
BamHI, BglI, BglII, EcoRI, and Hpal. Thenumber and size of the fragments differed in allthree R plasmids (Table 3). Plasmids pKRE5367and pVe7558, specifying the same resistance,also showed significant differences.The molecular sizes of the plasmids, which
were measured by agarose gel electrophoresisand by electron microscopy, differed between 27and 38 megadaltons (Mdal) (Table 1). The re-
lationship of the R plasmids pKRE5367 (Apr)and pLU121 (Tcr) was confirmed by the analysisof heteroduplex molecules between these H. in-fluenzae R plasmids in the electron microscope(Fig. 1).Demonstration of drug resistance trans-
posons. The 5.5-Mdal plasmid RSF1030 (6) wasused as a molecular probe for the detection ofthe ampicillin resistance transposon. PlasmidRSF1030 contains the whole Tn3, representing58% of its total DNA. As a molecular probe forthe detection of the tetracycline resistance trans-poson, 3H-labeled bacteriophage lambda 173(10) DNA, containing the whole of TnlO, was
used (Table 1). The TnlO represents 19.3% ofthe X 173, b221 rexl73::TnlO c1857 DNA. Thepresence of the ampicillin and the tetracyclinetransposons was verified by electron micro-graphs of heteroduplex molecules between am-picillin resistance-specifying and tetracycline re-
sistance-specifying plasmids. The heteroduplexmolecules of pKRE5367 DNA (Apr) withpLU121 DNA (Tcr) indicates the presence of an
TABLE 2. Hybridization between 3H-labeledpKRE5367, RSF1 030, and phage X 173 DNA and
whole-cell DNA from H. influenzae isolatescarrying plasmids"
DNA sequence homology with'H-labeled plasmid and phage A
Source of unlabeled 173 DNA (%)DNA
pKRE5367 RSF1030 A 173(Ap') (Apr) (TcO)
pKRE5367 (Apr) 100 51 0pVe7558 (Apr) 78 42 1pFR16017 (Tcr) 70 2 19pLU121 (Tcr) 70 3 20pHK539 (Tcr Apr) 80 53 21pBS (Tcr Apr) 66 46 20pRI234 (Tcr Cmr) 63 0 17pHi7O6 (Tcr Cmr 76 37 23
Apr)pW266b 83 0 0a The degree of DNA-DNA duplex formation was
assayed by the Si endonuclease method as described(3).bNo drug resistance.
Apr transposon and of a Tcr transposon (Fig. 1).Transposition ofthe Apr transposon from
pKRE5367 onto RSF1010. To further charac-terize the DNA sequence specifying ampicillinresistance in pKRE5367, the transposable na-ture of that DNA sequence was examined. As arecipient for the ampicillin resistance-specifyinggenes, the small plasmid RSF1010 (Smr, Sur)was chosen and used for transformation of E.coli 1485-lF-Na1r. The H. influenzae plasmidpKRE5367 was transferred by conjugation intothe E. coli containing RSF1010. A 3.2 ± 0.2-Mdal DNA sequence of the 30-Mdal pKRE5367was transposed onto the 5.5-Mdal plasmidRSF1010, selecting for ampicillin-resistantclones. The resulting 8.7 ± 0.2-Mdal hybrid plas-mid (pUWH) was shown to be RSF1O0O::TnA
564 LAUFS ET AL. J. BACTERIOL.
on Decem
ber 25, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

ORIGIN OF H. INFLUENZAE R FACTORS 565
TABLE 3. Cleavage of H. influenzae R plasmid DNAsNo. of fragments in the gel
Restriction enzyme No. of common fragmentspFR16017 pKRE5367 pVe7558
BamHI 1 1 1 1 pFR16017pKRE5367pVe7558
Bgll 4 3 2 2 pKRE5367pVe7558
BglII 5 2 3 0EcoRI 11 6 5 3 pKRE5367
pVe7558HpaI 6 2 2 1 pKRE5367
pVe7558
FIG. 1. (A) Electron micrograph ofheteroduplex betweenpKRE5367 (Apr) andpLU121 (Tcr)plasmid DNA.(B) Tracing of the same heteroduplex: ss, single-stranded DNA; ds, double-stranded DNA. Ap, Ampicillinresistance transposon ofpKRE5367. Tc, Tetracycline resistance transposon ofpL U121. IR, Inverted repeatsequence.
by analysis with restriction enzyme EcoRI (Fig.2) and electron microscope heteroduplex studies(Fig. 3).Isolation of an indigenous H. influenzae
plasmid. The demonstration of a transposableelement as the cause of the ampicillin resistanceof the H. influenzae strain KRE5367 stimulatedthe idea that pKRE5367 might have originatedby the transposition of the Apr transposon froman unknown donor plasmid onto an indigenousH. influenzae plasmid. Therefore, fresh isolates
of H. influenzae without drug resistance wereexamined by agarose gel electrophoresis for thepresence ofextrachromosomal DNA. Among 699H. influenzae strains isolated from healthy per-sons, one H. influenzae type B strain was foundwith the expected indigenous plasmid (pW266).The molecular size of pW266 was found in theelectron microscope to be 27 Mdal (mean of 10molecules measured), and it has 82% base ho-mology with pKRE5367 (Table 2). The molec-ular weight of pW266 (27 Mdal) corresponds to
VOL. 147, 1981
on Decem
ber 25, 2019 by guesthttp://jb.asm
.org/D
ownloaded from
566 LAUFS ET AL.
enzyme analysis of the in vitro-generated Tc-ApR plasmids with BamHI, BglI, Bgl[I, EcoRI,HindIII, Hpal, and Sall revealed a great simi-larity to pFR16017, indicating that the Apr trans-poson of pKRE16017 or of pVe7558 had trans-posed onto pFR16017. The in vitro-generatedAp-Tc R plasmids had almost the same restric-tion endonuclease cleavage pattern as the natu-ral Ap-Tc R plasmid pHK539.The conjugation of the in vitro-generated
plasmids specifying combined Tc-Ap resistancewith the strain HC234 (pRI234) specifying com-bined tetracycline-chloramphenicol resistanceresulted in plasmids which were indistinguish-able in the electron microscope from that found
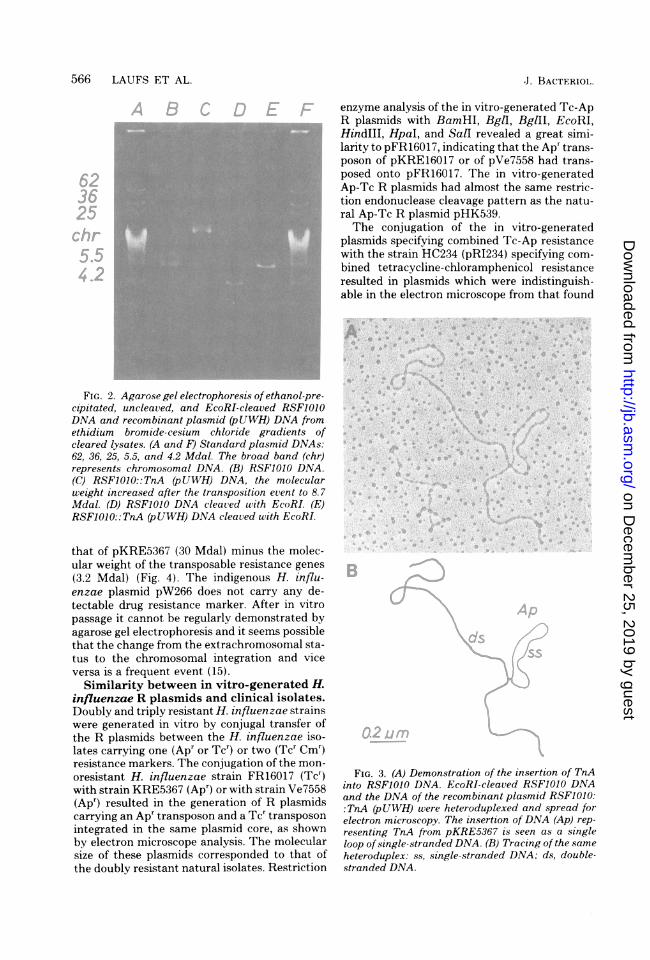
FIG. 2. Agarose gel electrophoresis of ethanol-pre-cipitated, uncleaved, and EcoRI-cleaved RSF1010DNA and recombinant plasmid (pUWH) DNA fromethidium bromide-cesium chloride gradients ofcleared lysates. (A and F) Standard plasmid DNAs:62, 36, 25, 5.5, and 4.2 Mdal. The broad band (chr)represents chromosomal DNA. (B) RSF1010 DNA.(C) RSFO010::TnA (pUWH) DNA, the molecularweight increased after the transposition event to 8.7Mdal. (D) RSF1010 DNA cleaved with EcoRI. (F)RSF1OO::TnA (pUWH) DNA cleaved with EcoRI.
that of pKRE5367 (30 Mdal) minus the molec-ular weight of the transposable resistance genes(3.2 Mdal) (Fig. 4). The indigenous H. influ-enzae plasmid pW266 does not carry any de-tectable drug resistance marker. After in vitropassage it cannot be regularly demonstrated byagarose gel electrophoresis and it seems possiblethat the change from the extrachromosomal sta-tus to the chromosomal integration and viceversa is a frequent event (15).Similarity between in vitro-generated H.
influenzae R plasmids and clinical isolates.Doubly and triply resistant H. influenzae strainswere generated in vitro by conjugal transfer ofthe R plasmids between the H. influenzae iso-lates carrying one (Apr or Tcr) or two (Tcr Cmr)resistance markers. The conjugation of the mon-oresistant H. influenzae strain FR16017 (Tcr)with strain KRE5367 (Apr) or with strain Ve7558(Apr) resulted in the generation of R plasmidscarrying an Apr transposon and a Tcr transposonintegrated in the same plasmid core, as shownby electron microscope analysis. The molecularsize of these plasmids corresponded to that ofthe doubly resistant natural isolates. Restriction
FIG. 3. (A) Demonstration of the insertion of TnAinto RSF1010 DNA. EcoRI-cleaved RSF1010 DNAand the DNA of the recombinant plasmid RSF1010::TnA (pUWH) were heteroduplexed and spread forelectron microscopy. The insertion of DNA (Ap) rep-resenting TnA from pKRE5367 is seen as a singleloop of single-stranded DNA. (B) Tracing of the sameheteroduplex: ss, single-stranded DNA; ds, double-stranded DNA.
J. BACTERIOL.
on Decem
ber 25, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

ORIGIN OF H. INFLUENZAE R FACTORS 567
FIG. 4. Agarose gel electrophoresis ofethanol-pre-cipitated plasmid DNA from cleared lysates. (A andE) Standard plasmid DNAs as in Fig. 2. (B) Lysatefrom strain KRE5367 (Apr). (D) Lysate from strainW266 harboring the indigenous plasmid pW266.
in the natural isolates with triple resistanceagainst ampicillin, tetracycline, and chloram-phenicol (manuscript in preparation). In eachcase the self-annealed molecules showed the Tc-Cm transposon (TnI894) (7), as well as a struc-ture similar to Tn3. The in vitro-generated plas-mids specifying triple resistance also containeda DNA sequence which had the characteristicsof the Tcr transposon present in pFR16017 (8).
DISCUSSIONThe cores of the H. influenzae R plasmids
isolated in different parts of the world havesimilar base sequences, but they are not identi-cal. For example, each of the two tetracyclineresistance-specifying H. influenzae R plasmidspFR16017 and pLU121 has 70% base sequence
homology with the reference R plasmidpKRE5367, but they show only 70% base se-
quence homology with each other. The restric-tion endonuclease patterns of the R plasmidsand the analysis of heteroduplex molecules inthe electron microscope reflect these differences.These findings support but do not prove the ideathat the H. influenzae R plasmids are of multi-clonal evolutionary origin.The drug resistance-specifying genes found in
the R plasmids resemble those of the transpos-able elements Tn3 and TnlO, which were origi-nally present in the plasmids RI and R100,respectively, which are both in the FII incom-patibility group (6, 11). This was shown in theelectron microscope and by using RSF1030 andX:TnlO as molecular probes. We have demon-
strated that the ampicillin resistance-specifyinggene of the H. influenzae plasmid pKRE5367 ispart of a transposon. It was transposed frompKRE5367 onto the small E. coli plasmidRSF1010. This observation stimulated thesearch for an indigenous H. influenzae plasmidas the naturally occurring recipient plasmid forthe drug resistance transposons. Indeed, such anindigenous H. influenzae plasmid (pW266) wasfound, and its molecular properties correlatedwith those which could be predicted. It has 82%base sequence homology with pKRE5367, andthe molecular size corresponds to that of thecores of the H. influenzae R plasmids. Thefunctions of the indigenous plasmid for its hostbacteria remained unknown, and drug resistancemarkers could not be detected.
After the isolation of monoresistant H. influ-enzae R plasmids (4, 8), doubly (7) and nowtriply (2) resistant strains have emerged. Theydiffer from each other mainly by the integrationof additional transposons. The in vitro-gener-ated doubly and triply resistant H. influenzae Rplasmids have been similar to the natural iso-lates as shown by the morphology of self-an-nealed molecules and by the cleavage with en-donucleases, indicating that the transpositionevents necessary for their generation in vitromay reflect the events in nature.The findings support the hypothesis that the
H. influenzae R plasmids could have simulta-neously arisen as a result of independent trans-position events of resistance genes onto differentbut closely related indigenous H. influenzaeplasmids in several parts of the world.
ACKNOWLEDGMENTSThis work was supported by the Deutsche Forschungsge-
meinschaft, Bonn-Bad Godesberg, West Germany.We thank I. Richter and A. Koppe for excellent technical
assistance.
LITERATURE CITED
1. Barth, P. T., N. Datta, R. W. Hedges, and N. J.Grinter. 1976. Transposition of a deoxyribonucleic acidsequence encoding trimethoprim and streptomcycin re-sistance from R483 to other replicons. J. Bacteriol. 125:800-810.
2. Braveny, I. K. Machka, K. Bartmann, K. Fabricius,F. Daschner, K. F. Petersen, H. Grimm, U. Ull-mann, and H. Freiesleben. 1980. Antibiotikaresistenzvon Haemophilus influenzae in der BundesrepublikDeutschland. Dtsch. Med. Wochenschr. 105:1341-1344.
3. Crosa, J. H., J. Brenner, and S. Falkow. 1973. Use ofa single-strand specific nuclease for analysis of bacterialand plasmid deoxyribonucleic acid homo- and hetero-duplexes. J. Bacteriol. 115:904-911.
4. Elwell, L. P., J. R. Saunders, M. H. Richmond, and S.Falkow. 1977. Relationships between some R-plasmidsfound in Haemophilus influenzae. J. Bacteriol. 131:356-362.
5. Heffron, F., C. Rubens, and S. Falkow. 1977. Trans-
VOL. 147, 1981
on Decem
ber 25, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

568 LAUFS ET AL.
position of a plasmid deoxyribonucleic acid sequencethat mediates ampicillin resistance: identity of labora-tory-constructed plasmids and clinical isolates. J. Bac-teriol. 129:530-533.
6. Heffron, F., R. Sublett, R. W. Hedges, A. Jacob, andS. Falkow. 1975. Origin of the TEM beta-lactamasegene found on plasmids. J. Bacteriol. 122:250-256.
7. Jahn, G., R. Laufs, P.-M. Kaulfers, and H. Kolenda.1979. Molecular nature of two Haemophilus influenzaeR factors containing resistances and the multiple inte-gration of drug resistance transposons. J. Bacteriol.138:584-597.
8. Kaulfers, P.-M., R. Laufs, and G. Jahn. 1978. Molecularproperties of transmissible R factors of Haemophilusinfluenzae determining tetracycline resistance. J. Gen.Microbiol. 105:243-252.
9. Kilian, M. 1976. A taxonomic study of the genus Hae-mophilus with the proposal of a new species. J. Gen.Microbiol. 93:9-62.
10. Kleckner, N., D. F. Barker, D. G. Ross, and D. Bot-stein. 1978. Properties of the translocatable tetracy-cline-resistance element TnlO in Escherichia coli andbacteriophage lambda. Genetics 90:427-450.
J. BACTERIOL.
11. Kleckner, N., R. K. Chan, B. K. Tye, and D. Botstein.1975. Mutagenesis by insertion of a drug-resistanceelement carrying an inverted repetition. J. Mol. Biol.97:561-575.
12. Laufs, R., and P.-M. Kaulfers. 1977. Molecular charac-terization of a plasmid specifying ampicillin resistanceand its relationship to other R factors from Haemophi-lus influenzae. J. Gen. Microbiol. 103:277-286.
13. Meyers, J. A., D. Sanchez, L. P. Elwell, and S. Fal-kow. 1976. Simple agarose gel electrophoretic methodfor the identification and characterization of plasmiddeoxyribonucleic acid. J. Bacteriol. 127:1529-1537.
14. Rigby, P. W. J., M. Dieckmann, C. Rhodes, and P.Berg. 1977. Labeling deoxyribonucleic acid to highspecific activity in vitro by nick translation with DNApolymerase I. J. Mol. Biol. 113:237-251.
15. Stuy, J. H. 1980. Chromosomahly integrated conjugativeplasmids are common in antibiotic-resistant Haemo-philus influenzae. J. Bacteriol. 142:925-930.
16. van Klingeren, B. J., D. A. van Embden, and M.Dessens-Kroon. 1977. Plasmid-mediated chloram-phenicol resistance in Haemophilus influenzae. Anti-microb. Agents Chemother. 11:383-387.
on Decem
ber 25, 2019 by guesthttp://jb.asm
.org/D
ownloaded from





























