Embed Size (px)
Citation preview

Journal of Applied Ecology
2008 doi 101111j1365-2664200801468x
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society
Blackwell Publishing Ltd
Exploring reservoir dynamics a case study of rabies in
the Serengeti ecosystem
Tiziana Lembo
12
Katie Hampson
234
Daniel T Haydon
5
Meggan Craft
26
Andy Dobson
24
Jonathan Dushoff
7
Eblate Ernest
2
Richard Hoare
8
Magai Kaare
12
Titus Mlengeya
9
Christine Mentzel
2
and Sarah Cleaveland
12
1
Centre for Tropical Veterinary Medicine Royal (Dick) School of Veterinary Studies University of Edinburgh Easter Bush Veterinary Centre Roslin Midlothian EH25 9RG UK
2
Viral Transmission Dynamics Project Tanzania Wildlife Research Institute PO Box 661 Arusha Tanzania
3
Department of Animal and Plant Sciences University of Sheffield Alfred Denny Building Sheffield S10 2TN UK
4
Department of Ecology and Evolutionary Biology Princeton University Princeton NJ 08544-1003 USA
5
Division of Environmental and Evolutionary Biology University of Glasgow Glasgow G12 8QQ UK
6
Department of Ecology Evolution and Behavior University of Minnesota St Paul MN 55108 USA
7
Department of Biology McMaster University Hamilton ON L8S 4K1 Canada
8
Tanzania Wildlife Research InstitutendashMesserli Foundation Wildlife Veterinary Programme PO Box 707 Arusha Tanzania and
9
Tanzania National Parks PO
Box 3134 Arusha Tanzania
Summary
1
Knowledge of infection reservoir dynamics is critical for effective disease control but identifyingreservoirs of multi-host pathogens is challenging Here we synthesize several lines of evidence toinvestigate rabies reservoirs in complex carnivore communities of the Serengeti ecological region innorthwest Tanzania where the disease has been confirmed in 12 carnivore species
2
Long-term monitoring data suggest that rabies persists in high-density domestic dog
Canisfamiliaris
populations (gt 11 dogs km
ndash2
) and occurs less frequently in lower-density (lt 5 dogs km
ndash2
)populations and only sporadically in wild carnivores
3
Genetic data show that a single rabies virus variant belonging to the group of southern Africacanid-associated viruses (Africa 1b) circulates among a range of species with no evidence ofspecies-specific virusndashhost associations
4
Within-species transmission was more frequently inferred from high-resolution epidemiologicaldata than between-species transmission Incidence patterns indicate that spill-over of rabies fromdomestic dog populations sometimes initiates short-lived chains of transmission in other carnivores
5
Synthesis and applications
The balance of evidence suggests that the reservoir of rabies in theSerengeti ecosystem is a complex multi-host community where domestic dogs are the only populationessential for persistence although other carnivores contribute to the reservoir as non-maintenancepopulations Control programmes that target domestic dog populations should therefore have thegreatest impact on reducing the risk of infection in all other species including humans livestockand endangered wildlife populations but transmission in other species may increase the level ofvaccination coverage in domestic dog populations necessary to eliminate rabies
Key-words
carnivore infectious disease multi-host phylogeny rabies reservoir spill-overtransmission
Introduction
Pathogens that infect multiple host species are often (i)economically important (ii) major threats to human health(iii) risk factors for endangered wildlife populations and(iv) causes of emerging human and livestock diseases (DaszakCunningham amp Hyatt 2000 Cleaveland Laurenson amp Taylor
Correspondence author E-mail khampsonshefacukAuthors Tiziana Lembo and Katie Hampson contributed equally tothis studyData deposition The sequences of rabies viruses produced in thisstudy have been deposited in the GenBank data base (Accession nosEU296434ndashEU296446)
2
T Lembo
et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society
Journal of Applied Ecology
2001 Dobson amp Foufopoulos 2001 Taylor Latham ampWoolhouse 2001) The generalist nature of these pathogensposes considerable challenges for understanding infectiondynamics and designing effective control strategies (Haydon
et al
2002) Key questions relate to the identification ofinfection reservoirs the mechanisms by which infections aresustained within reservoirs and the sources and routes oftransmission from reservoirs to species of concern Theseproblems are well-illustrated by bovine tuberculosis in GreatBritain where the role of cattle and badgers
Meles meles
inmaintaining
Mycobacterium bovis
is not fully resolved despitenumerous observational and intervention studies (Woodroffe
et al
2005 Independent Scientific Group on Cattle TB 2007)Many challenges posed by complex reservoir systems are encap-sulated by rabies in the Serengeti where the disease threatenshuman wildlife and domestic animal health (Cleaveland
et al
2007) The complexity of Serengetirsquos abundant and diverse hostcommunities necessitates a multi-faceted approach involvingmultiple data sources and a suite of analytical tools forunderstanding reservoir dynamics
Following the terminology of Haydon
et al
(2002) wedefine a reservoir as a set of epidemiologically connectedpopulations that permanently maintain a pathogen andtransmit infection to particular target populations consideredto require protection The key components of reservoirs ofdirectly transmitted microparasites are therefore the targetpopulations (of concern) and maintenance populations (inwhich sustained transmission occurs) Some reservoir systemsalso involve source populations which provide transmission
links between maintenance and target populations By defini-tion maintenance populations must exceed the minimumpopulation size required for disease persistence [the criticalcommunity size (CCS)
sensu
Bartlett 1960] Populationssmaller than the CCS (non-maintenance populations) cannotmaintain a pathogen independently but together with othermaintenance or non-maintenance populations can constitutepart of a reservoir (Haydon
et al
2002) These definitions areillustrated in Fig 1
Rabies is a classic example of a multi-host pathogen forwhich the identification of reservoirs has proven challenging(Nel 1993 Cleaveland amp Dye 1995 Bingham
et al
1999abJohnson
et al
2003 Bernardi
et al
2005) In Africa and Asiadomestic dog rabies predominates among reported andconfirmed cases and domestic dogs are the reported sourceof infection for over 90 of human cases (World HealthOrganization 1999) however it has been argued that this mayreflect surveillance bias and that the role of wildlife is poorlyunderstood (Swanepoel
et al
1993 Wandeler
et al
1994)Certain mongoose species (
Cynictis penicillata
and
Galerellasanguinea
) maintain distinct rabies virus (RABV) variants inparts of southern Africa (Foggin 1988 King Meredith ampThomson 1993 Swanepoel
et al
1993 Bingham
et al
2001Nel
et al
2005) and wild canids such as bat-eared foxes
Otocyon megalotis
side-striped jackals
Canis adustus
andblack-backed jackals
Canis mesomelas
appear able to sustainrabies cycles in some ecosystems (Thomson amp Meredith1993 Bingham
et al
1999a) but their role as independentmaintenance hosts and infection reservoirs is debated
Fig 1 Potential rabies reservoir systems inthe Serengeti CCS critical community sizeIn this figure we use humans as the targetpopulation but for rabies target populationsinclude humans livestock and endangeredwildlife If the epidemiological situationcorresponds to (a) vaccinating domesticdogs would shift the situation first to (c) andfinally to an overall community insufficientfor rabies maintenance
Rabies reservoir dynamics in the Serengeti
3
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society
Journal of Applied Ecology
(Cleaveland amp Dye 1995 Rhodes
et al
1998 Bingham
et al
1999b)
Wild carnivore communities in the Serengeti comprise26 species (Sinclair amp Arcese 1995) including Canidae andHerpestidae species implicated as independent maintenancehosts of rabies in parts of southern Africa (ie slendermongooses bat-eared foxes side-striped and black-backedjackals) Cleaveland amp Dye (1995) found that rabies appears topersist endemically in high-density domestic dog populations(gt 5 km
ndash2
) to the west of Serengeti National Park (SNP) yetoccurred only sporadically in lower-density domestic dogpopulations (lt 1 km
ndash2
) to the east of SNP and in wild carnivorepopulations They concluded that domestic dog populationsto the west of SNP were likely to be the sole reservoir in theecosystem (Fig 1a) however their study was based on limiteddata and did not consider the possibility of a multi-hostreservoir Furthermore with rapid growth of human andassociated domestic dog populations it seems likely that inthe meantime the lower-density domestic dog populationwhich previously could not support rabies cycles has becomelarge enough to maintain infection Additionally East
et al
(2001) argue that in the Serengeti a distinct rabies virusvariant is maintained in spotted hyaenas
Crocuta crocuta
which undergo atypical infections with no evidence of mor-tality Additional data now allow more detailed examinationof the role of wild carnivores in potential rabies reservoirsystems (i) as maintenance populations independent ofdomestic dogs (Fig 1b) (ii) as part of multi-host maintenancecommunities (Fig 1c) or (iii) as one of several independentmaintenance populations (Fig 1d)
Given the potential complexity of rabies reservoirs incommunities comprising multiple hosts it may not be possibleto reach an exhaustive understanding of their structure How-ever identifying maintenance populations and populationsthat act as sources of infection for target populations hasimportant practical implications Rabies is transmitted toseveral lsquospill-overrsquo hosts that might be considered targetpopulations of concern including humans (Knobel
et al
2005)endangered wildlife (Randall
et al
2006 Vial
et al
2006)and livestock In the Serengeti human rabies deaths occursubsistence farmers suffer livestock losses from rabies andthere is concern about risks to recovering populations ofAfrican wild dog
Lycaon pictus
[rabies was previouslyimplicated in their local extinction (Cleaveland
et al
2007)]There are long-term benefits to controlling rabies infection
directly in maintenance rather than in target populationshuman post-exposure prophylaxis (PEP) although effectiveis costly and often unavailable routine prophylactic vaccinationof livestock is expensive and vaccination of endangeredcanids has been controversial (Dye 1996 Woodroffe 2001Cleaveland
et al
2007) Approaches to control will dependupon how rabies is maintained if domestic dogs are the solemaintenance population or neither domestic dogs nor wildcarnivores maintain rabies independently (Fig 1ac) measurestargeted at domestic dogs should in theory also eliminatehuman and animal rabies However if wild carnivores are thesole maintenance population (Fig 1b) strategies will need totarget wildlife Similarly if domestic dog and wild carnivorepopulations each maintain rabies (Fig 1d) strategies willonly be successful if infection in both populations is controlled
Rabies in the Serengeti is therefore an important thoughchallenging model for addressing questions relating to thestructure and dynamics of complex reservoir systems Herewe present several lines of evidence that advance our under-standing of reservoir dynamics of rabies in multi-host carnivorecommunities of the Serengeti we synthesize data fromlong-term epidemiological records phylogenetic analyses ofvirus isolates from a range of domestic and wild species andstatistical analyses of high-resolution spatiotemporal patternsof rabies incidence
Methods
STUDY
AREA
The study area in the Serengeti ecological region of northwestTanzania was divided into three zones (Fig 2) The first zoneSNP comprises diverse wildlife communities with domestic dogsfound extremely rarely The two other zones are districts adjacent toSNP Serengeti District (SD) to the west and Ngorongoro District(ND) comprising the Loliondo Game Control Area (LGCA) andNgorongoro Conservation Area (NCA) to the east SD is inhabitedby multi-ethnic agro-pastoralist communities and high-densitydomestic dog populations (gt 11 km
ndash2
) ND is a multiple-use controlledwildlife area inhabited by low-density pastoralist communitiesand lower-density domestic dog populations (lt 5 km
ndash2
) We haveobserved a greater variety and abundance of wild carnivores in NDthan SD No physical barriers separate the wildlife-protected areasand human settlements
Estimated densities of human and domestic dog populations arepresented in Table 1 these were extrapolated from human census
Table 1 Estimated domestic dog population densities in Serengeti (SD) and Ngorongoro districts [ND ndash Loliondo Game Control Area(LGCA) only] Confidence intervals about the means are indicated in brackets
AreaArea (km2)
Domestic dogndashhuman ratio
Average human density (kmndash2) at the village level
Average domestic dog density(kmndash2) at the village level
SD urban (Mugumu town) 18middot8 6middot6 619middot8 93middot9SD rural 3128middot0 6middot6 67middot9 (59middot2ndash76middot5) 10middot3 (9middot0ndash11middot6)SD overall 3146middot8 6middot6 75middot2 (58middot4ndash92middot0) 11middot4 (8middot9ndash13middot9)LGCA urban (Loliondo and Sakala) 14middot1 6middot7 144middot9 21middot6LGCA rural 8852middot4 6middot7 17middot0 (11middot1ndash23middot0) 2middot5 (1middot7ndash3middot4)LGCA overall 8866middot5 6middot7 28middot2 (9middot2ndash47middot1) 4middot2 (1middot4ndash7middot0)
4
T Lembo
et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society
Journal of Applied Ecology
data with projected annual growth rates of 2middot6 in SD and 3middot8 inND (National Bureau of Statistics Tanzania 2005) and from humanndashdomestic dog ratios (Wandeler
et al
1988 Brooks 1990) obtainedfrom household questionnaires (from SD in 2003 and LGCA in2004)
Several domestic dog vaccination programmes have beenimplemented in the study area Details of vaccination campaigns arereported in Appendix S2 (Supplementary material) although theirimpact is not analysed here The only vaccination of wildlife (Africanwild dogs) was conducted in 1990 (Gascoyne
et al
1993)
D ISEASE
MONITORING
OPERATIONS
Data presented are the result of disease monitoring operations sincethe late 1980s (Table 2) Passive surveillance was used to detectrabies cases in SNP Sightings of sick and dead carnivores werereported through a network of veterinarians from Tanzania NationalParks (TANAPA) and Tanzania Wildlife Research Institute(TAWIRI) rangers scientists and tour-operators Cases includedanimals that were snared injuredwounded from other or unknowncauses observed with signs of disease or found dead with unknownhistory
Outside SNP passive and active surveillance operations wereemployed Passive surveillance data were available through veterinaryoffice records (government offices TAWIRI TANAPA and NCAAuthority) and district hospitals and medical dispensaries Biteinjuries to humans from suspected rabid dogs were defined based onevaluation by clinicians (from information on clinical signs reportedby bite victims) and when human vaccines were not widely availableon whether PEP had been administered Bite injury incidence wascalculated against human population sizes interpolated from census
data (National Bureau of Statistics Tanzania 2005) Community-based active surveillance measures based on previous studies inrural Kenya (Kitala
et al
2000) and Tanzania (Cleaveland
et al
2003) were implemented in SD using livestock field officersstationed in randomly selected villages to collect information onrabies cases from key informants (village leaders teachers dispensarystaff local healers)
Contact tracing was used to collect high-resolution data on spatialand temporal patterns of disease Visits to incidents involving bitinganimals (to bite victims and owners of biting and bitten animals)were initiated using hospital and medical dispensary records andcase reports from livestock offices and community-based surveillanceIncidents were mapped and witnesses interviewed to make a diagnosis(see below) and obtain case-history information (exposure sourceonset of clinical signs and death resulting exposures) The sameprocedure was followed for all exposures and preceding cases whereidentified so that transmission events between identified individualanimals could be traced and confirmed When multiple incidentsinvolving suspected rabid wildlife were reported on the sameconsecutive days within neighbouring homesteads we assumed asingle animal was involved
Wherever possible brain stem samples were collected from suspectrabies cases and carnivore carcasses whatever the apparent cause ofdeath Sample collection was carried out by research personnelveterinarians and livestock field officers the preferred techniquewas removal via the occipital foramen according to World HealthOrganization recommendations (Barrat 1996) Specimens were frozen(ndash20
deg
C) immediately after collection or placed into phosphate-buffered 50 glycerol solution and preserved either cold (+4
deg
Cndash20
deg
C) or at room temperature (25 plusmn 5
deg
C) when refrigerationfacilities were not promptly available
Fig 2 Map of the Serengeti ecological regionillustrating the distribution of suspectedrabies cases (most were not laboratory-confirmed but diagnosed using epidemio-logical history and clinical criteria)Tables S2 and S3 in Supplementary materialdetail confirmed wildlife cases in SerengetiNational Park (SNP) and Serengeti (SD) andNgorongoro (ND) districts Cases recordedin SNP from January 1995ndashJanuary 2007 aremapped whereas only cases from January2002ndashJanuary 2007 in SD and ND aremapped because GPS data were not availablefor the earlier period Domestic dog cases areshown in black (SD n = 833 ND n = 128)and cases in other carnivores as white circles(SD n = 96 ND n = 27 SNP n = 23)Village boundaries are indicated by greylines LGCA Loliondo Game Control AreaNCA Ngorongoro Conservation Area
Rabies reservoir dynamics in the Serengeti
5
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society
Journal of Applied Ecology
RABIES
D IAGNOSIS
AND
V IRUS
CHARACTERIZATION
Diagnostic tests on brains collected up to 2001 and virus isolationwere carried out at the Agence Franccedilaise de Seacutecuriteacute Sanitaire desAliments (AFSSA) Malzeacuteville France using the fluorescent antibodytest (Dean Abelseth amp Atanasiu 1996) inoculation of murineneuroblastoma cells and mouse inoculation (Barrat
et al
1988)Rabies diagnosis on more recent brain tissues was conducted at theRabies Section of the Centers for Disease Control and Prevention(CDC) Atlanta USA by the fluorescent antibody test (httpwwwcdcgovrabiesdocsstandard_dfa_protocol_rabiespdf) Virussamples were typed at CDC RNA was extracted reverse transcribedamplified by polymerase chain reaction and nucleoprotein genesequences generated as described in Lembo
et al
(2007)When brain tissues were not available for laboratory confirmation
diagnosis of suspected rabies cases was based on history of exposureand clinical evaluation (adapting the six criteria proposed byTepsumethanon Wilde amp Meslin 2005) from information on clinicalsigns reported by villagers livestock field officers researchers andpark veterinarians
1
For owned domestic dogs history of a bite clinical signs(Tepsumethanon
et al
2005) disappearance or death within 10 days
2
For domestic dogs of unknown origin clinical signs (as above) anddisappearance or death
3
For wild carnivores clinical signs (as above) plus tameness lossof fear of humans diurnal activity (for nocturnal species) andunprovoked biting (without eating) of objects and animals
PHYLOGENETIC
ANALYSIS
Nucleotide sequences were edited using
bioedit
7middot0middot0 (Hall 1999)and aligned in
clustal-x
version 1middot83 (Jeanmougin
et al
1998)Phylogenetic relationships among viruses from the study area(Table S1 Supplementary material) and sequence data for RABVsfrom other African countries (Kissi Tordo amp Bourhy 1995 Randall
et al
2004) were estimated using Bayesian Markov chain MonteCarlo (MCMC) methods using
mrbayes
3middot0b4 (Ronquist ampHuelsenbeck 2003) as described in Lembo
et al
(2007) Posteriorprobabilities of 0middot95 or greater were considered significant Graphicoutput was generated using
treeview
version 1middot6middot6 (Page 1996)
STATISTICAL
ANALYSIS
Poisson regression was used to test for a relationship between rabiescases in domestic dogs and in other carnivores Using maximumlikelihood a gamma-distributed spatial infection kernel for domesticdogs k(
d
) was estimated from the distances between source animalsand their known contacts and a probability distribution of thegeneration interval for domestic dogs g(
t
) was estimated fromobserved dates of exposure onset of clinical signs and death (fulldetails in Appendix S2) An algorithm (detailed in Appendix S2)was developed to generate putative epidemic trees by assigningprobabilities to links between possible progenitors and each suspectedcase according to their spatiotemporal proximity and the estimatedspatial infection kernel k(
d
) and generation interval distributiong(
t
) This algorithm assumes all cases are observed and thereforecannot account for unobserved intermediates Cases in all specieswere included in the analysis and all carnivore species were assumedequally likely to transmit rabies Only limited data could be collectedon transmission from wild carnivores therefore progenitors of allspecies were assigned using parameters estimated for domestic dogs
Tab
le 2
Tim
elin
e of
dis
ease
mon
itor
ing
oper
atio
ns in
eac
h zo
ne in
6-m
onth
uni
ts T
AW
IRI
Tan
zani
a W
ildlif
e R
esea
rch
Inst
itut
e T
AN
APA
Tan
zani
a N
atio
nal P
arks
NC
AA
Ngo
rong
oro
Con
serv
atio
nA
rea
Aut
hori
ty
YE
AR
ZO
NE
1987
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
SN
P
TA
NA
PA V
eter
inar
y U
nit
TA
WIR
I V
eter
inar
y P
rogr
amm
e
Vir
al T
rans
mis
sion
D
ynam
ics
Pro
ject
SD
Pas
sive
sur
veill
ance
(do
mes
tic
dog-
bite
inju
ry r
ecor
ds f
rom
hos
pita
ls)
D
omes
tic
dog
vacc
inat
ions
Pas
sive
sur
veill
ance
(V
eter
inar
y re
cord
s
villa
ge r
epor
ts o
ppor
tuni
stic
car
cass
sam
plin
g)
Act
ive
(liv
esto
ck fi
eld-
offic
ers)
Act
ive
(liv
esto
ck fi
eld-
offic
ers)
Con
tact
tra
cing
ND
Pas
sive
(do
mes
tic
dog-
bite
inju
ry r
ecor
ds
from
hos
pita
ls)
Dom
esti
c do
g va
ccin
atio
nsP
assi
ve (
Vet
erin
ary
reco
rds
opp
ortu
nist
ic c
arca
ss
sam
plin
g T
AW
IRI
TA
NA
PA a
nd N
CA
A r
ecor
ds)
Con
tact
tra
cing
A r
esea
rch
proj
ect
inve
stig
atin
g in
fect
ion
dyna
mic
s of
vir
al d
isea
ses
6 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
(k(d) and g(t)) One thousand bootstrapped trees were generatedand the proportional contribution of transmission to each case frompreceding cases was calculated If the date of onset of clinical signswas uncertain in each realization a date was assigned with uniformprobability from the range of uncertainty
From the bootstrap trees the amount of transmission withinand between host types was estimated We pooled all carnivoresexcept domestic dogs because of small sample sizes A null model ofthe expected frequency of within- and between-species transmissionassuming that intra- and interspecific transmission events occurwith the same probability (ie transmission is non-assortative)was estimated as follows the species identity of each case wasrandomly reassigned retaining the original distribution of casesamongst species and the resulting frequency of within- andbetween-species transmission was calculated This procedure wasiterated 1000 times to evaluate the probability that the differencebetween inferred within- and between-species transmissionfrequencies could have been due to chance To investigate whetherthe spatial distribution of hosts influenced the estimated patterns oftransmission we repeated the analysis but randomly reassignedspecies identities within grid cells rather than across the entiredistrict and we repeated the analysis for different sized grids (seeAppendix S2)
Results
RABIES RECOGNIT ION PROBABIL ITY
The sensitivity specificity and positive predictive value ofclinical rabies diagnosis (ie recognition of rabies by villagerslivestock field officers park veterinarians and researchpersonnel) calculated against the gold standard fluorescentantibody test (sensitivity ~99 specificity gt 99 Dean et al1996) are reported in Table 3 As a result of the relatively highrabies recognition probability (gt 74 of animals reported assuspect rabies cases were confirmed positive) and the smallnumber of confirmed cases the analyses presented include allsuspected cases unless otherwise stated
RABIES INCIDENCE IN THE STUDY POPULATIONS
Wildlife monitoring activities in SNP are shown throughoutthe study period in Table 2 and Fig 3a A peak in wild
carnivores reported dead and sick coincided with a caninedistemper outbreak in late 1993 and 1994 (Roelke-Parkeret al 1996) Most rabies cases in wildlife were reported andconfirmed in 1998 and 1999 with only sporadic detection inother years (Fig 3b Table S2 in Supplementary material) Lownumbers of snared animals were reported in 1998 suggestingthat the increase in carnivore carcasses recorded that year(Fig 3a) was not due to increased monitoring No wildlifecases have been confirmed in SNP since 1999 except for aleopard Panthera pardus on the SNPndashNCA boundary inSeptember 2004 when there was an outbreak among adjacentdomestic dog populations (shown in Fig 5b)
Animal-bite injuries in humans from suspected rabiddomestic dogs were reported continuously in high-densitydomestic dog populations to the west of SNP before imple-mentation of dog vaccination programmes whereas in the
Table 3 Sensitivity specificity and positive predictive value of rabiesdiagnosis calculated against the gold standard fluorescent antibodytest Data include only cases for which complete history was available(2002ndash2005)
Clinical diagnosis
Laboratory diagnosis
TotalPositive Negative
Positive 55 19 74Negative 0 33 33Total 55 52 107Sensitivity = 100middot00Specificity = 63middot46 (95 CI 50middot37ndash76middot55)Positive predictive value = 74middot32 (95 CI 64middot37ndash84middot28)
Fig 3 Wildlife disease monitoring in Serengeti National Park(a) carnivore reports and (b) samples retrieved for rabies diagnosisThe peak in 1994 coincided with a canine distemper outbreak inlions (Roelke-Parker et al 1996)
Rabies reservoir dynamics in the Serengeti 7
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
lower-density populations to the east incidence was lowerand there were several periods when no bites were recorded(Fig 4) Gaps in recorded bites by suspected rabid dogs mayalso reflect PEP shortages however when we included alldomestic dog bite injuries there were four periods of at least5 months with no reported bite injuries in ND and no equivalentpattern in SD (Fig S1 Supplementary material)
We traced 1255 suspected rabid animals from January 2002to January 2007 (1044 in SD 211 in ND see Fig 5 for carnivorecases) In both districts cases in wildlife were sporadic andcoincided with outbreaks in domestic dogs (Figs 3b 4 amp 5)The spatial and temporal distribution of suspected cases isshown in Figs 2 and 5
INTER- AND INTRA-SPECIF IC TRANSMISSION
The virus
All of the 57 RABV specimens recovered from a range ofspecies from the study area (including spotted hyaenas) fellwithin the Africa 1b group of canid-associated viruses andrevealed no clustering by host species (Fig 6) The evidentintermingling of viral lineages from domestic dogs andwildlife indicates frequent interspecific transmission Analysesusing statistical parsimony techniques were also consistentwith both within- and between-species transmission events(Lembo et al 2007)
Fig 4 Incidence of bites by suspected rabiddomestic dogs in Serengeti (SD black line)and Ngorongoro (ND grey line) districtsderived from hospital records Incidence isshown only for years before the implementa-tion of domestic dog vaccination programmesin the study populations (July 1987ndashJanuary1997 in SD and January 1994ndashDecember2003 in ND) No data were available for6 months during 1993 in SD
Fig 5 Suspected rabies cases amongcarnivore species in (a) Serengeti (n = 929)and (b) Ngorongoro districts (n = 155)monitored by contact tracing from January2001 to January 2007
8 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Spatiotemporal incidence patterns in hosts
Suspected rabies cases in domestic dogs were significantpredictors of those in other carnivores (pooled cases indomestic cats and wildlife P lt 0middot001 in both districts)Credibility intervals (95) of the expected frequency ofwithin- and between-species transmission events based uponthe null assumption of non-assortative transmission (derivedfrom the bootstrap analysis) are shown in Fig 7 Morewithin-species transmission than expected was estimated inboth districts (domestic dog-to-domestic dog transmissionP = 0middot052 in SD and P = 0middot008 in ND transmission withinother carnivores ie wild carnivores and domestic catsFelis catus P lt 0middot0001 in both districts) Estimated between-species transmission was less than expected (P lt 0middot03 in SD
and P lt 0middot001 in ND) except for transmission from domesticdogs to livestock These patterns were consistent and significantwhen the randomization procedure was carried out on a10 times 10 km grid while on a 5 times 5 km grid the patterns wereevident but no longer significant probably because of reducedstatistical power
The majority of human exposures to suspected rabid animalswere from domestic dogs (Table 4) Similarly domestic dogswere the predominant source of infection for livestock (Fig 7)
Discussion
Demonstrating persistence of infection is a prerequisite foridentifying reservoir(s) Domestic dogs living at high densities(within the SD) are the only species in which rabies has been
Fig 6 Phylogenetic tree of nucleoprotein gene sequences (282 bp 94 deduced amino acids nucleotide positions 1139ndash1420 on the SAD B19genome (Conzelmann et al 1990) for RABV samples from the study area compared with isolates from other areas of Africa The tree isconstructed using Bayesian phylogenetics under the transitional model of nucleotide evolution with a gamma-shaped distribution of rates acrosssites (TIM + Γ Posada amp Crandall 1998 base frequencies = 0middot3253 0middot2134 0middot2360 0middot2253 nucleotide substitution rates = 1middot0000 3middot67230middot4393 0middot4393 8middot1773 1middot0000 Γ = 0middot3390) For samples from the study area the year of collection is indicated within square bracketsPreviously published sequences are designated by strain names (Kissi et al 1995 Randall et al 2004) The tree is rooted with isolate 1500AFSdefined as outgroup representative of the lineage Africa 3 Nodal posterior probabilities gt 0middot95 are shown The scale indicates branch-lengthexpressed as the expected number of substitutions per site Species not definitively identified
Rabies reservoir dynamics in the Serengeti 9
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
recorded continuously throughout the study period whereascases in other species (inside and outside SNP) have beensporadic Although disease monitoring presumably detects alower proportion of cases in wildlife than domestic dogs itseems likely that infection in wildlife populations had it beenpresent continuously would have been detected in more recentyears where the number of samples collected and tested washigher than in previous years when rabies was recorded(Table 2 Fig 3b) These observations provide circumstantialevidence for rabies persistence in domestic dog populationsand lack of persistence in other carnivores Clusters ofwildlife cases that coincided with outbreaks in domestic dogpopulations (Figs 3b 4 amp 5) and explosive but transient(lt 2 months) epidemics previously reported in bat-eared foxes(Maas 1993) are also consistent with chains of transmissionthat are not sustained and contrast with incidence patterns inwild carnivores that are believed to maintain rabies throughindependent cycles elsewhere in Africa For example rabieswas recorded sporadically in bat-eared fox populations inSouth Africa from the 1950s to 1970 then continuouslysuggesting a shift to persistent infection (Thomson amp Meredith1993) and in Zimbabwe outbreaks in jackals although ofteninitiated by domestic dog cases appear to be sustained overseveral years (Bingham et al 1999ab)
Specific demographic attributes (high densities andorturnover) are thought to be required for sustained rabiestransmission A threshold density of gt 5 dogs kmndash2 has beensuggested for persistence of rabies in domestic dog populationsin Africa (Foggin 1988 Brooks 1990 Cleaveland amp Dye 1995Kitala et al 2001) Domestic dog populations in the studyregion have grown considerably and even in the lower-density
area (within the ND) the estimated average density hasapproached this threshold (Table 1) and in 2003 populationsizes were sufficient for a sizable epidemic to occur (Fig 5b)Rabies-control measures make it hard to determine whethercurrent densities in ND are high enough for infection to persistin the absence of vaccination but they may now be close
We detected only one canid-associated variant (Africa 1b)from a range of species in the Serengeti and were unable toreplicate East et alrsquos (2001) finding of a distinct virus variantcirculating atypically in spotted hyaenas (Lembo et al 2007)The lack of species-specific virusndashhost associations in thecanid-associated variant that we detected and the high degreeof genetic relatedness between virus samples from differentspecies indicate frequent cross-species transmission This isconsistent with results from southern Africa where phylogeneticbranches from a single geographic area include several hostspecies (Sabeta Bingham amp Nel 2003)
The statistical association between rabies cases in domesticdogs and other species also indicates cross-species transmissionUnfortunately the length of our time series and paucity ofcases detected in species other than domestic dogs precludeanalyses of lsquospill-overrsquo risk and reduce the interpretativepower of time-lagged regressions like those conducted forrabies in North America that could otherwise provide insight(Guerra et al 2003 Gordon et al 2004) In general as theamount of between-species transmission increases the moredifficult it becomes to tease apart the role of differentpotentially independent maintenance hosts We find greaterwithin-species and less between-species transmission thanwould be expected if species interacted randomly This patterncould occur as a consequence of the spatial distribution ofhost species andor preferential contact between conspecificsOur randomization indicates that large-scale patterns in thedistribution of hosts (gt 10 times 10 km) do not explain thislsquoassortativersquo transmission but we cannot make inference ona smaller scale These analyses suggest that occasionallsquospill-overrsquo of infection from domestic dog populations into
Fig 7 The proportion of estimated transmission within andbetween host types (associated with 1255 suspected rabies cases 1044in Serengeti and 211 in Ngorongoro) in Serengeti (light grey) andNgorongoro (dark grey) districts with lsquodogrsquo corresponding to domesticdogs and lsquocarnivorersquo to domestic cats and wild carnivores Credibilityintervals (95) are shown for expected transmission within andbetween host types assuming inter- and intraspecific transmissionwere equally likely and that livestock are dead-end hosts
Table 4 Human exposures to rabies as a result of bites by suspectedrabid animals in Serengeti and Ngorongoro districts (January 2002ndashDecember 2006) Data were obtained by contact tracing
SpeciesSerengeti district ()
Ngorongorodistrict ()
Domestic dogs 466 (85middot5) 84 (70middot0)Domestic cats 25 (4middot6) 6 (5middot0)Livestock 7 (1middot3) 3 (2middot5)Human 2 (0middot4) 1 (0middot8)Spotted hyaena Crocuta crocuta 4 (0middot7) 16 (13middot3)Honey badger Mellivora capensis 11 (2middot0) 6 (5middot0)African wildcat Felis lybica 0 (0) 2 (1middot7)White-tailed mongoose
Ichneumia albicauda2 (0middot4) 1 (0middot8)
Bat-eared fox Otocyon megalotis 2 (0middot4) 0 (0)Small-spotted genet Genetta genetta 1 (0middot2) 1 (0middot8)Jackal spp Canis spp 23 (4middot2) 0 (0)Leopard Panthera pardus 2 (0middot4) 0 (0)Total 545 (100) 120 (100)
10 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
alternative hosts sometimes results in short-lived chains oftransmission as contact is more common among conspecifics(Dobson 2004)
Definitive identification of infection reservoirs is extremelychallenging and rarely possible Despite intensive effortunderstanding of reservoir dynamics in many complexmulti-host systems remains incomplete (Woodroffe et al2005 Biek et al 2006 Independent Scientific Group on CattleTB 2007) Here the balance of evidence implicates domesticdogs as the only maintenance population of rabies in theSerengeti whereas other carnivores are nonessential tomaintenance However because they transmit disease totarget populations (and frequent transmission within andbetween such species may occur) they do constitute part ofthe reservoir (Table 4 Fig 1a) A single maintenance-hostspecies is consistent with broad patterns of rabies infectionobserved elsewhere (Smith 1989 Rupprecht et al 1991Wandeler et al 1994 Smith Orciari amp Yager 1995)
A lesson from this study is that even with considerable datafrom various sources we cannot exhaustively quantify thecontribution of transmission from all non-essential reservoircomponents However unravelling every thread of reservoirdynamics may not be essential Our key finding that wildlifeare not able to maintain rabies cycles or distinct geneticvariants provides new and critical information allowing rabiescontrol measures in the Serengeti to be targeted moreconfidently at domestic dog populations alone The long-term implication is that control of domestic dog rabies in theSerengeti will eliminate rabies in all other species includinghumans livestock and wildlife The absence of wildlifemaintenance hosts even in an area with such abundantwildlife raises the expectation that canine rabies eliminationis a feasible objective throughout much of Africa
Interventions to prevent transmission from domestic dogmaintenance populations would also provide convincingevidence with which to identify reservoirs Such approacheshave demonstrated that cattle were the reservoir for rinderpestafter cattle vaccinations rinderpest disappeared from wildlifeand the wildebeest Connochaetes taurinus population increasednearly ten-fold (Sinclair 1979 Plowright 1982 Dobson 1995)We are currently ring-vaccinating domestic dogs around SNPwhich should provide critical data to evaluate the impact ofalternative hosts on long-term control efforts in particular theireffects on the level of vaccination required for disease control
In conclusion resolving reservoir dynamics in any complexsystem is likely to prove challenging and our experiences inthe Serengeti suggest that consideration needs to be given toidentifying (i) the potential role of different componentsof the system (ii) the key information needed for differentdisease control strategies (iii) the epidemiological approacheslikely to yield most useful information and (iv) the range ofanalytical tools available
Acknowledgements
We are grateful to Tanzania Government ministries TANAPA TAWIRINCA Authority Tanzania Commission for Science and Technology andNational Institute for Medical Research for permissions and TANAPA and
TAWIRI Veterinary Units Viral Transmission Dynamics Project SerengetiLion and Cheetah Projects Frankfurt Zoological Society livestock field-officersof the Ministry of Water and Livestock Development in Mara and ArushaRegions M Magoto P Tiringa M Roelke-Parker R Fyumagwa H Wiickand Mwanza and Arusha Veterinary Investigation Centres for assistance withsample collection We are indebted to J Barrat AFSSA for performingdiagnostic testing and virus isolation and the staff of the Rabies Section of theCDC especially CE Rupprecht A Velasco-Villa LA Orciari IV Kuzminand M Niezgoda and PE Brandatildeo (Universidade de Satildeo Paulo) for technicalassistance with diagnostic tests and virus characterization We thankKLaurenson D Bennett J Pulliam and H Horn for valuable discussions
This work was supported by the joint National Institutes of Health (NIH)National Science Foundation (NSF) Ecology of Infectious Diseases Programunder grant no NSFDEB0225453 Opinions findings conclusions andrecommendations are those of the authors and do not necessarily reflectthe views of NIH or NSF KH was supported by NSF grant DEB-0513994the Pew Charitable Trusts award 2000-002558 to Princeton University theDepartment of Ecology and Evolution Princeton University and the HeinzFoundation TL was supported by the Royal (Dick) School of VeterinaryStudies University of Edinburgh and the University of Edinburgh DevelopmentTrust (visits to CDC) and SC by the Wellcome Trust and the Department forInternational Development Animal Health Programme
References
Barrat J (1996) Simple technique for the collection and shipment of brainspecimens for rabies diagnosis Laboratory Techniques in Rabies (edsF-X Meslin MM Kaplan amp H Koprowski) pp 425ndash432 World HealthOrganization Geneva Switzerland
Barrat J Barrat MJ Picard M amp Aubert MFA (1988) Diagnosis of rabiesby cell-culture Comparative Immunology Microbiology and InfectiousDiseases 11 207ndash214
Bartlett MS (1960) Stochastic Population Models in Ecology and EpidemiologyMethuen London
Bernardi F Nadin-Davis SA Wandeler AI Armstrong J Gomes AABLima FS Nogueira FRB amp Ito FH (2005) Antigenic and geneticcharacterization of rabies viruses isolated from domestic and wild animalsof Brazil identifies the hoary fox as a rabies reservoir Journal of GeneralVirology 86 3153ndash3162
Biek R Walsh PD Leroy EM amp Real LA (2006) Recent common ancestryof Ebola Zaire virus found in a bat reservoir Public Library of SciencePathogens 2 885ndash886
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999a) Theepidemiology of rabies in Zimbabwe 2 Rabies in jackals (Canis adustus andCanis mesomelas) Onderstepoort Journal of Veterinary Research 66 11ndash23
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999b) Theepidemiology of rabies in Zimbabwe 1 Rabies in dogs (Canis familiaris)Onderstepoort Journal of Veterinary Research 66 1ndash10
Bingham J Javangwe S Sabeta CT Wandeler AI amp Nel LH (2001)Report of isolations of unusual lyssaviruses (rabies and Mokola virus)identified retrospectively from Zimbabwe Journal of the South AfricanVeterinary Association 72 92ndash94
Brooks R (1990) Survey of the dog-population of Zimbabwe and its level ofrabies vaccination Veterinary Record 127 592ndash596
Cleaveland S amp Dye C (1995) Maintenance of a microparasite infectingseveral host species rabies in the Serengeti Parasitology 111 S33ndashS47
Cleaveland S Kaare M Tiringa P Mlengeya T amp Barrat J (2003) A dograbies vaccination campaign in rural Africa impact on the incidence of dograbies and human dog-bite injuries Vaccine 21 1965ndash1973
Cleaveland S Laurenson MK amp Taylor LH (2001) Diseases of humansand their domestic mammals pathogen characteristics host range andthe risk of emergence Philosophical Transactions of the Royal Society BBiological Sciences 356 991ndash999
Cleaveland S Mlengeya T Kaare M Haydon DT Lembo T LaurensonMK amp Packer C (2007) The conservation relevance of epidemiologicalresearch into carnivore viral diseases in the Serengeti Conservation Biology21 612ndash622
Conzelmann KK Cox JH Schneider LG amp Thiel HJ (1990) Molecularcloning and complete nucleotide sequence of the attenuated rabies virusSAD B19 Virology 175 485ndash499
Daszak P Cunningham AA amp Hyatt AD (2000) Wildlife ecology ndash Emerginginfectious diseases of wildlife ndash Threats to Biodiversity and human healthScience 287 443ndash449
Dean DJ Abelseth MK amp Atanasiu P (1996) The fluorescent antibodytest Laboratory Techniques in Rabies (eds F-X Meslin MM Kaplan
Rabies reservoir dynamics in the Serengeti 11
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
amp H Koprowski) pp 88ndash95 World Health Organization GenevaSwitzerland
Dobson A (1995) The ecology and epidemiology of rinderpest virus inSerengeti and Ngorongoro Conservation Area Serengeti II DynamicsManagement and Conservation of an Ecosystem (eds ARE Sinclair ampP Arcese) pp 485ndash505 University of Chicago Press Chicago
Dobson A (2004) Population dynamics of pathogens with multiple host speciesAmerican Naturalist 164 S64ndashS78
Dobson A amp Foufopoulos J (2001) Emerging infectious pathogens of wildlifePhilosophical Transactions of the Royal Society B Biological Sciences 3561001ndash1012
Dye C (1996) Serengeti wild dogs what really happened Trends in Ecology ampEvolution 11 188ndash189
East ML Hofer H Cox JH Wulle U Wiik H amp Pitra C (2001) Regularexposure to rabies virus and lack of symptomatic disease in Serengetispotted hyenas Proceedings of the National Academy of Sciences USA 9815026ndash15031
Foggin CM (1988) Rabies and Rabies-related Viruses in Zimbabwe HistoricalVirological and Ecological Aspects PhD thesis University of ZimbabweHarare Zimbabwe
Gascoyne SC King AA Laurenson MK Borner M Schildger B ampBarrat J (1993) Aspects of rabies infection and control in the conservationof the African wild dog (Lycaon pictus) in the Serengeti region TanzaniaOnderstepoort Journal of Veterinary Research 60 415ndash420
Gordon ER Curns AT Krebs JW Rupprecht CE Real LA ampChilds JE (2004) Temporal dynamics of rabies in a wildlife host and the riskof cross-species transmission Epidemiology and Infection 132 515ndash524
Guerra MA Curns AT Rupprecht CE Hanlon CA Krebs JW ampChilds JE (2003) Skunk and raccoon rabies in the eastern United Statestemporal and spatial analysis Emerging Infectious Diseases 9 1143ndash1150
Hall TA (1999) bioedit a user-friendly biological sequence alignment editorand analysis program for Windows 9598NT Nucleic Acids SymposiumSeries 41 95ndash98
Haydon DT Cleaveland S Taylor LH amp Laurenson MK (2002) Identi-fying reservoirs of infection a conceptual and practical challenge EmergingInfectious Diseases 8 1468ndash1473
Independent Scientific Group on Cattle TB (2007) Bovine TB The ScientificEvidence Defra Publications London PB12634 httpwwwdefragovukanimalhtbisgindexhtm
Jeanmougin F Thompson JD Gouy M Higgins DG amp Gibson TJ(1998) Multiple sequence alignment with clustal-x Trends in BiochemicalSciences 23 403ndash405
Johnson N Black C Smith J Un H McElhinney LM Aylan O ampFooks AR (2003) Rabies emergence among foxes in Turkey Journal ofWildlife Diseases 39 262ndash270
King AA Meredith CD amp Thomson GR (1993) Canid and viverrid rabiesviruses in South Africa Onderstepoort Journal of Veterinary Research 60295ndash299
Kissi B Tordo N amp Bourhy H (1995) Genetic polymorphism in the rabiesvirus nucleoprotein gene Virology 209 526ndash537
Kitala PM McDermott JJ Kyule MN amp Gathuma JM (2000) Community-based active surveillance for rabies in Machakos District Kenya PreventiveVeterinary Medicine 44 73ndash85
Kitala PM McDermott J Kyule M Gathuma JM Perry B amp Wandeler A(2001) Dog ecology and demography information to support the planning ofrabies control in Machakos District Kenya Acta Tropica 78 217ndash230
Knobel DL Cleaveland S Coleman PG Fevre EM Meltzer MIMiranda MEG Shaw A Zinsstag J amp Meslin F-X (2005) Re-evaluatingthe burden of rabies in Africa and Asia Bulletin of the World HealthOrganization 83 360ndash368
Lembo T Haydon DT Velasco-Villa A Rupprecht CE Packer CBrandatildeo PE Kuzmin IV Fooks AR Barrat J amp Cleaveland S (2007)Molecular epidemiology identifies only a single rabies virus variantcirculating in complex carnivore communities of the Serengeti Proceedingsof the Royal Society B Biological Sciences 274 2123ndash2130
Maas B (1993) Behavioural Ecology and Social Organisation of the Bat-earedFox in the Serengeti National Park Tanzania University of CambridgeCambridge UK
National Bureau of Statistics Tanzania (2005) 2002 Population and HousingCensus Presidentrsquos Office Planning and Privatization Dar es SalaamTanzania
Nel JAJ (1993) The bat-eared fox-a prime candidate for rabies vectorOnderstepoort Journal of Veterinary Research 60 395ndash397
Nel LH Sabeta CT von Teichman B Jaftha JB Rupprecht CE amp
Bingham J (2005) Mongoose rabies in southern Africa a re-evaluationbased on molecular epidemiology Virus Research 109 165ndash173
Page RDM (1996) treeview an application to display phylogenetic trees onpersonal computers Computer Applications in the Biosciences 12 357ndash358
Plowright W (1982) The effects of rinderpest and rinderpest control on wildlifein Africa Symposia of the Zoological Society of London 50 1ndash28
Posada D amp Crandall KA (1998) modeltest testing the model of DNAsubstitution Bioinformatics 14 817ndash818
Randall DA Marino J Haydon DT Sillero-Zubiri C Knobel DLTallents LA Macdonald DW amp Laurenson MK (2006) An integrateddisease management strategy for the control of rabies in Ethiopian wolvesBiological Conservation 131 151ndash162
Randall DA Williams SD Kuzmin IV Rupprecht CE Tallents LATefera Z Argaw K Shiferaw F Knobel DL Sillero-Zubiri C ampLaurenson MK (2004) Rabies in endangered Ethiopian wolves EmergingInfectious Diseases 10 2214ndash2217
Rhodes CJ Atkinson RPD Anderson RM amp Macdonald DW (1998)Rabies in Zimbabwe reservoir dogs and the implications for disease controlPhilosophical Transactions of the Royal Society B Biological Sciences 353999ndash1010
Roelke-Parker ME Munson L Packer C Kock R Cleaveland SCarpenter M Obrien SJ Pospischil A HofmannLehmann R Lutz HMwamengele GLM Mgasa MN Machange GA Summers BA ampAppel MJG (1996) A canine distemper virus epidemic in Serengeti lions(Panthera leo) Nature 379 441ndash445
Ronquist F amp Huelsenbeck JP (2003) mrbayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 19 1572ndash1574
Rupprecht CE Dietzschold B Wunner WH amp Koprowsky H (1991)Antigenic relationships of lyssaviruses The Natural History of Rabies (edsGM Baer) pp 69ndash100 CRC Press Boca Raton
Sabeta CT Bingham J amp Nel LH (2003) Molecular epidemiology of canidrabies in Zimbabwe and South Africa Virus Research 91 203ndash211
Sinclair ARE (1979) The eruption of the ruminants Serengeti Dynamics ofan Ecosystem (eds ARE Sinclair amp M Norton-Griffiths) pp 82ndash103University of Chicago Press Chicago
Sinclair ARE amp Arcese P (1995) Serengeti II Dynamics Management andConservation of an Ecosystem University of Chicago Press Chicago
Smith JS (1989) Rabies virus epitopic variation use in ecological studiesAdvances in Virus Research 36 215ndash253
Smith JS Orciari LA amp Yager PA (1995) Molecular epidemiology ofrabies in the United States Seminars in Virology 6 387ndash400
Swanepoel R Barnard BJH Meredith CD Bishop GC BrucknerGK Foggin CM amp Hubschle OJB (1993) Rabies in Southern AfricaOnderstepoort Journal of Veterinary Research 60 325ndash346
Taylor LH Latham SM amp Woolhouse MEJ (2001) Risk factors forhuman disease emergence Philosophical Transactions of the Royal SocietyB Biological Sciences 356 983ndash989
Tepsumethanon V Wilde H amp Meslin FX (2005) Six criteria for rabiesdiagnosis in living dogs Journal of the Medical Association of Thailand 88419ndash422
Thomson GR amp Meredith CD (1993) Rabies in bat-eared foxes in SouthAfrica Onderstepoort Journal of Veterinary Research 60 399ndash403
Vial F Cleaveland S Rasmussen G amp Haydon DT (2006) Development ofvaccination strategies for the management of rabies in African wild dogsBiological Conservation 131 180ndash192
Wandeler AI Budde A Capt S Kappeler A amp Matter H (1988) Dogecology and dog rabies control Reviews of Infectious Diseases 10 S684ndashS688
Wandeler AI Nadin-Davis SA Tinline RR amp Rupprecht CE (1994)Rabies epidemiology some ecological and evolutionary perspectivesLyssaviruses (eds CE Rupprecht B Dietzschold amp H Koprowski)pp 297ndash324 Springer-Verlag Berlin
Woodroffe R (2001) Assessing the risks of intervention immobilizationradio-collaring and vaccination of African wild dogs Oryx 35 234ndash244
Woodroffe R Donnelly CA Johnston WT Bourne FJ Cheeseman CLClifton-Hadley RS Cox DR Gettinby G Hewinson RG Le FevreAM McInerney JP amp Morrison WI (2005) Spatial association ofMycobacterium bovis infection in cattle and badgers Meles meles Journal ofApplied Ecology 42 852ndash862
World Health Organization (1999) World Survey of Rabies No 34 for the Year1998 Geneva Switzerland
Received 14 June 2007 accepted 23 January 2008Handling Editor Simon Thirgood
12 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Supplementary material
The following supplementary material is available for thisarticle
Appendix S1 Latin names of wildlife species
Appendix S2 Supplementary details of methods
Table S1 Virus samples from the study area compared inphylogenetic analysis
Table S2 Details of all wildlife confirmed and suspect casesin SNP
Table S3 Details of confirmed wildlife cases in Serengeti andNgorongoro districts
Fig S1 Incidence of bites by domestic dogs in Serengeti andNgorongoro districts
This material is available as part of the online article fromhttpwwwblackwell-synergycomdoifull101111j1365-2664200801468x(This link will take you to the article abstract)
Please note Blackwell Publishing is not responsible for the con-tent or functionality of any supplementary materials suppliedby the authors Any queries (other than missing material)should be directed to the corresponding author for the article

2
T Lembo
et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society
Journal of Applied Ecology
2001 Dobson amp Foufopoulos 2001 Taylor Latham ampWoolhouse 2001) The generalist nature of these pathogensposes considerable challenges for understanding infectiondynamics and designing effective control strategies (Haydon
et al
2002) Key questions relate to the identification ofinfection reservoirs the mechanisms by which infections aresustained within reservoirs and the sources and routes oftransmission from reservoirs to species of concern Theseproblems are well-illustrated by bovine tuberculosis in GreatBritain where the role of cattle and badgers
Meles meles
inmaintaining
Mycobacterium bovis
is not fully resolved despitenumerous observational and intervention studies (Woodroffe
et al
2005 Independent Scientific Group on Cattle TB 2007)Many challenges posed by complex reservoir systems are encap-sulated by rabies in the Serengeti where the disease threatenshuman wildlife and domestic animal health (Cleaveland
et al
2007) The complexity of Serengetirsquos abundant and diverse hostcommunities necessitates a multi-faceted approach involvingmultiple data sources and a suite of analytical tools forunderstanding reservoir dynamics
Following the terminology of Haydon
et al
(2002) wedefine a reservoir as a set of epidemiologically connectedpopulations that permanently maintain a pathogen andtransmit infection to particular target populations consideredto require protection The key components of reservoirs ofdirectly transmitted microparasites are therefore the targetpopulations (of concern) and maintenance populations (inwhich sustained transmission occurs) Some reservoir systemsalso involve source populations which provide transmission
links between maintenance and target populations By defini-tion maintenance populations must exceed the minimumpopulation size required for disease persistence [the criticalcommunity size (CCS)
sensu
Bartlett 1960] Populationssmaller than the CCS (non-maintenance populations) cannotmaintain a pathogen independently but together with othermaintenance or non-maintenance populations can constitutepart of a reservoir (Haydon
et al
2002) These definitions areillustrated in Fig 1
Rabies is a classic example of a multi-host pathogen forwhich the identification of reservoirs has proven challenging(Nel 1993 Cleaveland amp Dye 1995 Bingham
et al
1999abJohnson
et al
2003 Bernardi
et al
2005) In Africa and Asiadomestic dog rabies predominates among reported andconfirmed cases and domestic dogs are the reported sourceof infection for over 90 of human cases (World HealthOrganization 1999) however it has been argued that this mayreflect surveillance bias and that the role of wildlife is poorlyunderstood (Swanepoel
et al
1993 Wandeler
et al
1994)Certain mongoose species (
Cynictis penicillata
and
Galerellasanguinea
) maintain distinct rabies virus (RABV) variants inparts of southern Africa (Foggin 1988 King Meredith ampThomson 1993 Swanepoel
et al
1993 Bingham
et al
2001Nel
et al
2005) and wild canids such as bat-eared foxes
Otocyon megalotis
side-striped jackals
Canis adustus
andblack-backed jackals
Canis mesomelas
appear able to sustainrabies cycles in some ecosystems (Thomson amp Meredith1993 Bingham
et al
1999a) but their role as independentmaintenance hosts and infection reservoirs is debated
Fig 1 Potential rabies reservoir systems inthe Serengeti CCS critical community sizeIn this figure we use humans as the targetpopulation but for rabies target populationsinclude humans livestock and endangeredwildlife If the epidemiological situationcorresponds to (a) vaccinating domesticdogs would shift the situation first to (c) andfinally to an overall community insufficientfor rabies maintenance
Rabies reservoir dynamics in the Serengeti
3
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society
Journal of Applied Ecology
(Cleaveland amp Dye 1995 Rhodes
et al
1998 Bingham
et al
1999b)
Wild carnivore communities in the Serengeti comprise26 species (Sinclair amp Arcese 1995) including Canidae andHerpestidae species implicated as independent maintenancehosts of rabies in parts of southern Africa (ie slendermongooses bat-eared foxes side-striped and black-backedjackals) Cleaveland amp Dye (1995) found that rabies appears topersist endemically in high-density domestic dog populations(gt 5 km
ndash2
) to the west of Serengeti National Park (SNP) yetoccurred only sporadically in lower-density domestic dogpopulations (lt 1 km
ndash2
) to the east of SNP and in wild carnivorepopulations They concluded that domestic dog populationsto the west of SNP were likely to be the sole reservoir in theecosystem (Fig 1a) however their study was based on limiteddata and did not consider the possibility of a multi-hostreservoir Furthermore with rapid growth of human andassociated domestic dog populations it seems likely that inthe meantime the lower-density domestic dog populationwhich previously could not support rabies cycles has becomelarge enough to maintain infection Additionally East
et al
(2001) argue that in the Serengeti a distinct rabies virusvariant is maintained in spotted hyaenas
Crocuta crocuta
which undergo atypical infections with no evidence of mor-tality Additional data now allow more detailed examinationof the role of wild carnivores in potential rabies reservoirsystems (i) as maintenance populations independent ofdomestic dogs (Fig 1b) (ii) as part of multi-host maintenancecommunities (Fig 1c) or (iii) as one of several independentmaintenance populations (Fig 1d)
Given the potential complexity of rabies reservoirs incommunities comprising multiple hosts it may not be possibleto reach an exhaustive understanding of their structure How-ever identifying maintenance populations and populationsthat act as sources of infection for target populations hasimportant practical implications Rabies is transmitted toseveral lsquospill-overrsquo hosts that might be considered targetpopulations of concern including humans (Knobel
et al
2005)endangered wildlife (Randall
et al
2006 Vial
et al
2006)and livestock In the Serengeti human rabies deaths occursubsistence farmers suffer livestock losses from rabies andthere is concern about risks to recovering populations ofAfrican wild dog
Lycaon pictus
[rabies was previouslyimplicated in their local extinction (Cleaveland
et al
2007)]There are long-term benefits to controlling rabies infection
directly in maintenance rather than in target populationshuman post-exposure prophylaxis (PEP) although effectiveis costly and often unavailable routine prophylactic vaccinationof livestock is expensive and vaccination of endangeredcanids has been controversial (Dye 1996 Woodroffe 2001Cleaveland
et al
2007) Approaches to control will dependupon how rabies is maintained if domestic dogs are the solemaintenance population or neither domestic dogs nor wildcarnivores maintain rabies independently (Fig 1ac) measurestargeted at domestic dogs should in theory also eliminatehuman and animal rabies However if wild carnivores are thesole maintenance population (Fig 1b) strategies will need totarget wildlife Similarly if domestic dog and wild carnivorepopulations each maintain rabies (Fig 1d) strategies willonly be successful if infection in both populations is controlled
Rabies in the Serengeti is therefore an important thoughchallenging model for addressing questions relating to thestructure and dynamics of complex reservoir systems Herewe present several lines of evidence that advance our under-standing of reservoir dynamics of rabies in multi-host carnivorecommunities of the Serengeti we synthesize data fromlong-term epidemiological records phylogenetic analyses ofvirus isolates from a range of domestic and wild species andstatistical analyses of high-resolution spatiotemporal patternsof rabies incidence
Methods
STUDY
AREA
The study area in the Serengeti ecological region of northwestTanzania was divided into three zones (Fig 2) The first zoneSNP comprises diverse wildlife communities with domestic dogsfound extremely rarely The two other zones are districts adjacent toSNP Serengeti District (SD) to the west and Ngorongoro District(ND) comprising the Loliondo Game Control Area (LGCA) andNgorongoro Conservation Area (NCA) to the east SD is inhabitedby multi-ethnic agro-pastoralist communities and high-densitydomestic dog populations (gt 11 km
ndash2
) ND is a multiple-use controlledwildlife area inhabited by low-density pastoralist communitiesand lower-density domestic dog populations (lt 5 km
ndash2
) We haveobserved a greater variety and abundance of wild carnivores in NDthan SD No physical barriers separate the wildlife-protected areasand human settlements
Estimated densities of human and domestic dog populations arepresented in Table 1 these were extrapolated from human census
Table 1 Estimated domestic dog population densities in Serengeti (SD) and Ngorongoro districts [ND ndash Loliondo Game Control Area(LGCA) only] Confidence intervals about the means are indicated in brackets
AreaArea (km2)
Domestic dogndashhuman ratio
Average human density (kmndash2) at the village level
Average domestic dog density(kmndash2) at the village level
SD urban (Mugumu town) 18middot8 6middot6 619middot8 93middot9SD rural 3128middot0 6middot6 67middot9 (59middot2ndash76middot5) 10middot3 (9middot0ndash11middot6)SD overall 3146middot8 6middot6 75middot2 (58middot4ndash92middot0) 11middot4 (8middot9ndash13middot9)LGCA urban (Loliondo and Sakala) 14middot1 6middot7 144middot9 21middot6LGCA rural 8852middot4 6middot7 17middot0 (11middot1ndash23middot0) 2middot5 (1middot7ndash3middot4)LGCA overall 8866middot5 6middot7 28middot2 (9middot2ndash47middot1) 4middot2 (1middot4ndash7middot0)
4
T Lembo
et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society
Journal of Applied Ecology
data with projected annual growth rates of 2middot6 in SD and 3middot8 inND (National Bureau of Statistics Tanzania 2005) and from humanndashdomestic dog ratios (Wandeler
et al
1988 Brooks 1990) obtainedfrom household questionnaires (from SD in 2003 and LGCA in2004)
Several domestic dog vaccination programmes have beenimplemented in the study area Details of vaccination campaigns arereported in Appendix S2 (Supplementary material) although theirimpact is not analysed here The only vaccination of wildlife (Africanwild dogs) was conducted in 1990 (Gascoyne
et al
1993)
D ISEASE
MONITORING
OPERATIONS
Data presented are the result of disease monitoring operations sincethe late 1980s (Table 2) Passive surveillance was used to detectrabies cases in SNP Sightings of sick and dead carnivores werereported through a network of veterinarians from Tanzania NationalParks (TANAPA) and Tanzania Wildlife Research Institute(TAWIRI) rangers scientists and tour-operators Cases includedanimals that were snared injuredwounded from other or unknowncauses observed with signs of disease or found dead with unknownhistory
Outside SNP passive and active surveillance operations wereemployed Passive surveillance data were available through veterinaryoffice records (government offices TAWIRI TANAPA and NCAAuthority) and district hospitals and medical dispensaries Biteinjuries to humans from suspected rabid dogs were defined based onevaluation by clinicians (from information on clinical signs reportedby bite victims) and when human vaccines were not widely availableon whether PEP had been administered Bite injury incidence wascalculated against human population sizes interpolated from census
data (National Bureau of Statistics Tanzania 2005) Community-based active surveillance measures based on previous studies inrural Kenya (Kitala
et al
2000) and Tanzania (Cleaveland
et al
2003) were implemented in SD using livestock field officersstationed in randomly selected villages to collect information onrabies cases from key informants (village leaders teachers dispensarystaff local healers)
Contact tracing was used to collect high-resolution data on spatialand temporal patterns of disease Visits to incidents involving bitinganimals (to bite victims and owners of biting and bitten animals)were initiated using hospital and medical dispensary records andcase reports from livestock offices and community-based surveillanceIncidents were mapped and witnesses interviewed to make a diagnosis(see below) and obtain case-history information (exposure sourceonset of clinical signs and death resulting exposures) The sameprocedure was followed for all exposures and preceding cases whereidentified so that transmission events between identified individualanimals could be traced and confirmed When multiple incidentsinvolving suspected rabid wildlife were reported on the sameconsecutive days within neighbouring homesteads we assumed asingle animal was involved
Wherever possible brain stem samples were collected from suspectrabies cases and carnivore carcasses whatever the apparent cause ofdeath Sample collection was carried out by research personnelveterinarians and livestock field officers the preferred techniquewas removal via the occipital foramen according to World HealthOrganization recommendations (Barrat 1996) Specimens were frozen(ndash20
deg
C) immediately after collection or placed into phosphate-buffered 50 glycerol solution and preserved either cold (+4
deg
Cndash20
deg
C) or at room temperature (25 plusmn 5
deg
C) when refrigerationfacilities were not promptly available
Fig 2 Map of the Serengeti ecological regionillustrating the distribution of suspectedrabies cases (most were not laboratory-confirmed but diagnosed using epidemio-logical history and clinical criteria)Tables S2 and S3 in Supplementary materialdetail confirmed wildlife cases in SerengetiNational Park (SNP) and Serengeti (SD) andNgorongoro (ND) districts Cases recordedin SNP from January 1995ndashJanuary 2007 aremapped whereas only cases from January2002ndashJanuary 2007 in SD and ND aremapped because GPS data were not availablefor the earlier period Domestic dog cases areshown in black (SD n = 833 ND n = 128)and cases in other carnivores as white circles(SD n = 96 ND n = 27 SNP n = 23)Village boundaries are indicated by greylines LGCA Loliondo Game Control AreaNCA Ngorongoro Conservation Area
Rabies reservoir dynamics in the Serengeti
5
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society
Journal of Applied Ecology
RABIES
D IAGNOSIS
AND
V IRUS
CHARACTERIZATION
Diagnostic tests on brains collected up to 2001 and virus isolationwere carried out at the Agence Franccedilaise de Seacutecuriteacute Sanitaire desAliments (AFSSA) Malzeacuteville France using the fluorescent antibodytest (Dean Abelseth amp Atanasiu 1996) inoculation of murineneuroblastoma cells and mouse inoculation (Barrat
et al
1988)Rabies diagnosis on more recent brain tissues was conducted at theRabies Section of the Centers for Disease Control and Prevention(CDC) Atlanta USA by the fluorescent antibody test (httpwwwcdcgovrabiesdocsstandard_dfa_protocol_rabiespdf) Virussamples were typed at CDC RNA was extracted reverse transcribedamplified by polymerase chain reaction and nucleoprotein genesequences generated as described in Lembo
et al
(2007)When brain tissues were not available for laboratory confirmation
diagnosis of suspected rabies cases was based on history of exposureand clinical evaluation (adapting the six criteria proposed byTepsumethanon Wilde amp Meslin 2005) from information on clinicalsigns reported by villagers livestock field officers researchers andpark veterinarians
1
For owned domestic dogs history of a bite clinical signs(Tepsumethanon
et al
2005) disappearance or death within 10 days
2
For domestic dogs of unknown origin clinical signs (as above) anddisappearance or death
3
For wild carnivores clinical signs (as above) plus tameness lossof fear of humans diurnal activity (for nocturnal species) andunprovoked biting (without eating) of objects and animals
PHYLOGENETIC
ANALYSIS
Nucleotide sequences were edited using
bioedit
7middot0middot0 (Hall 1999)and aligned in
clustal-x
version 1middot83 (Jeanmougin
et al
1998)Phylogenetic relationships among viruses from the study area(Table S1 Supplementary material) and sequence data for RABVsfrom other African countries (Kissi Tordo amp Bourhy 1995 Randall
et al
2004) were estimated using Bayesian Markov chain MonteCarlo (MCMC) methods using
mrbayes
3middot0b4 (Ronquist ampHuelsenbeck 2003) as described in Lembo
et al
(2007) Posteriorprobabilities of 0middot95 or greater were considered significant Graphicoutput was generated using
treeview
version 1middot6middot6 (Page 1996)
STATISTICAL
ANALYSIS
Poisson regression was used to test for a relationship between rabiescases in domestic dogs and in other carnivores Using maximumlikelihood a gamma-distributed spatial infection kernel for domesticdogs k(
d
) was estimated from the distances between source animalsand their known contacts and a probability distribution of thegeneration interval for domestic dogs g(
t
) was estimated fromobserved dates of exposure onset of clinical signs and death (fulldetails in Appendix S2) An algorithm (detailed in Appendix S2)was developed to generate putative epidemic trees by assigningprobabilities to links between possible progenitors and each suspectedcase according to their spatiotemporal proximity and the estimatedspatial infection kernel k(
d
) and generation interval distributiong(
t
) This algorithm assumes all cases are observed and thereforecannot account for unobserved intermediates Cases in all specieswere included in the analysis and all carnivore species were assumedequally likely to transmit rabies Only limited data could be collectedon transmission from wild carnivores therefore progenitors of allspecies were assigned using parameters estimated for domestic dogs
Tab
le 2
Tim
elin
e of
dis
ease
mon
itor
ing
oper
atio
ns in
eac
h zo
ne in
6-m
onth
uni
ts T
AW
IRI
Tan
zani
a W
ildlif
e R
esea
rch
Inst
itut
e T
AN
APA
Tan
zani
a N
atio
nal P
arks
NC
AA
Ngo
rong
oro
Con
serv
atio
nA
rea
Aut
hori
ty
YE
AR
ZO
NE
1987
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
SN
P
TA
NA
PA V
eter
inar
y U
nit
TA
WIR
I V
eter
inar
y P
rogr
amm
e
Vir
al T
rans
mis
sion
D
ynam
ics
Pro
ject
SD
Pas
sive
sur
veill
ance
(do
mes
tic
dog-
bite
inju
ry r
ecor
ds f
rom
hos
pita
ls)
D
omes
tic
dog
vacc
inat
ions
Pas
sive
sur
veill
ance
(V
eter
inar
y re
cord
s
villa
ge r
epor
ts o
ppor
tuni
stic
car
cass
sam
plin
g)
Act
ive
(liv
esto
ck fi
eld-
offic
ers)
Act
ive
(liv
esto
ck fi
eld-
offic
ers)
Con
tact
tra
cing
ND
Pas
sive
(do
mes
tic
dog-
bite
inju
ry r
ecor
ds
from
hos
pita
ls)
Dom
esti
c do
g va
ccin
atio
nsP
assi
ve (
Vet
erin
ary
reco
rds
opp
ortu
nist
ic c
arca
ss
sam
plin
g T
AW
IRI
TA
NA
PA a
nd N
CA
A r
ecor
ds)
Con
tact
tra
cing
A r
esea
rch
proj
ect
inve
stig
atin
g in
fect
ion
dyna
mic
s of
vir
al d
isea
ses
6 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
(k(d) and g(t)) One thousand bootstrapped trees were generatedand the proportional contribution of transmission to each case frompreceding cases was calculated If the date of onset of clinical signswas uncertain in each realization a date was assigned with uniformprobability from the range of uncertainty
From the bootstrap trees the amount of transmission withinand between host types was estimated We pooled all carnivoresexcept domestic dogs because of small sample sizes A null model ofthe expected frequency of within- and between-species transmissionassuming that intra- and interspecific transmission events occurwith the same probability (ie transmission is non-assortative)was estimated as follows the species identity of each case wasrandomly reassigned retaining the original distribution of casesamongst species and the resulting frequency of within- andbetween-species transmission was calculated This procedure wasiterated 1000 times to evaluate the probability that the differencebetween inferred within- and between-species transmissionfrequencies could have been due to chance To investigate whetherthe spatial distribution of hosts influenced the estimated patterns oftransmission we repeated the analysis but randomly reassignedspecies identities within grid cells rather than across the entiredistrict and we repeated the analysis for different sized grids (seeAppendix S2)
Results
RABIES RECOGNIT ION PROBABIL ITY
The sensitivity specificity and positive predictive value ofclinical rabies diagnosis (ie recognition of rabies by villagerslivestock field officers park veterinarians and researchpersonnel) calculated against the gold standard fluorescentantibody test (sensitivity ~99 specificity gt 99 Dean et al1996) are reported in Table 3 As a result of the relatively highrabies recognition probability (gt 74 of animals reported assuspect rabies cases were confirmed positive) and the smallnumber of confirmed cases the analyses presented include allsuspected cases unless otherwise stated
RABIES INCIDENCE IN THE STUDY POPULATIONS
Wildlife monitoring activities in SNP are shown throughoutthe study period in Table 2 and Fig 3a A peak in wild
carnivores reported dead and sick coincided with a caninedistemper outbreak in late 1993 and 1994 (Roelke-Parkeret al 1996) Most rabies cases in wildlife were reported andconfirmed in 1998 and 1999 with only sporadic detection inother years (Fig 3b Table S2 in Supplementary material) Lownumbers of snared animals were reported in 1998 suggestingthat the increase in carnivore carcasses recorded that year(Fig 3a) was not due to increased monitoring No wildlifecases have been confirmed in SNP since 1999 except for aleopard Panthera pardus on the SNPndashNCA boundary inSeptember 2004 when there was an outbreak among adjacentdomestic dog populations (shown in Fig 5b)
Animal-bite injuries in humans from suspected rabiddomestic dogs were reported continuously in high-densitydomestic dog populations to the west of SNP before imple-mentation of dog vaccination programmes whereas in the
Table 3 Sensitivity specificity and positive predictive value of rabiesdiagnosis calculated against the gold standard fluorescent antibodytest Data include only cases for which complete history was available(2002ndash2005)
Clinical diagnosis
Laboratory diagnosis
TotalPositive Negative
Positive 55 19 74Negative 0 33 33Total 55 52 107Sensitivity = 100middot00Specificity = 63middot46 (95 CI 50middot37ndash76middot55)Positive predictive value = 74middot32 (95 CI 64middot37ndash84middot28)
Fig 3 Wildlife disease monitoring in Serengeti National Park(a) carnivore reports and (b) samples retrieved for rabies diagnosisThe peak in 1994 coincided with a canine distemper outbreak inlions (Roelke-Parker et al 1996)
Rabies reservoir dynamics in the Serengeti 7
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
lower-density populations to the east incidence was lowerand there were several periods when no bites were recorded(Fig 4) Gaps in recorded bites by suspected rabid dogs mayalso reflect PEP shortages however when we included alldomestic dog bite injuries there were four periods of at least5 months with no reported bite injuries in ND and no equivalentpattern in SD (Fig S1 Supplementary material)
We traced 1255 suspected rabid animals from January 2002to January 2007 (1044 in SD 211 in ND see Fig 5 for carnivorecases) In both districts cases in wildlife were sporadic andcoincided with outbreaks in domestic dogs (Figs 3b 4 amp 5)The spatial and temporal distribution of suspected cases isshown in Figs 2 and 5
INTER- AND INTRA-SPECIF IC TRANSMISSION
The virus
All of the 57 RABV specimens recovered from a range ofspecies from the study area (including spotted hyaenas) fellwithin the Africa 1b group of canid-associated viruses andrevealed no clustering by host species (Fig 6) The evidentintermingling of viral lineages from domestic dogs andwildlife indicates frequent interspecific transmission Analysesusing statistical parsimony techniques were also consistentwith both within- and between-species transmission events(Lembo et al 2007)
Fig 4 Incidence of bites by suspected rabiddomestic dogs in Serengeti (SD black line)and Ngorongoro (ND grey line) districtsderived from hospital records Incidence isshown only for years before the implementa-tion of domestic dog vaccination programmesin the study populations (July 1987ndashJanuary1997 in SD and January 1994ndashDecember2003 in ND) No data were available for6 months during 1993 in SD
Fig 5 Suspected rabies cases amongcarnivore species in (a) Serengeti (n = 929)and (b) Ngorongoro districts (n = 155)monitored by contact tracing from January2001 to January 2007
8 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Spatiotemporal incidence patterns in hosts
Suspected rabies cases in domestic dogs were significantpredictors of those in other carnivores (pooled cases indomestic cats and wildlife P lt 0middot001 in both districts)Credibility intervals (95) of the expected frequency ofwithin- and between-species transmission events based uponthe null assumption of non-assortative transmission (derivedfrom the bootstrap analysis) are shown in Fig 7 Morewithin-species transmission than expected was estimated inboth districts (domestic dog-to-domestic dog transmissionP = 0middot052 in SD and P = 0middot008 in ND transmission withinother carnivores ie wild carnivores and domestic catsFelis catus P lt 0middot0001 in both districts) Estimated between-species transmission was less than expected (P lt 0middot03 in SD
and P lt 0middot001 in ND) except for transmission from domesticdogs to livestock These patterns were consistent and significantwhen the randomization procedure was carried out on a10 times 10 km grid while on a 5 times 5 km grid the patterns wereevident but no longer significant probably because of reducedstatistical power
The majority of human exposures to suspected rabid animalswere from domestic dogs (Table 4) Similarly domestic dogswere the predominant source of infection for livestock (Fig 7)
Discussion
Demonstrating persistence of infection is a prerequisite foridentifying reservoir(s) Domestic dogs living at high densities(within the SD) are the only species in which rabies has been
Fig 6 Phylogenetic tree of nucleoprotein gene sequences (282 bp 94 deduced amino acids nucleotide positions 1139ndash1420 on the SAD B19genome (Conzelmann et al 1990) for RABV samples from the study area compared with isolates from other areas of Africa The tree isconstructed using Bayesian phylogenetics under the transitional model of nucleotide evolution with a gamma-shaped distribution of rates acrosssites (TIM + Γ Posada amp Crandall 1998 base frequencies = 0middot3253 0middot2134 0middot2360 0middot2253 nucleotide substitution rates = 1middot0000 3middot67230middot4393 0middot4393 8middot1773 1middot0000 Γ = 0middot3390) For samples from the study area the year of collection is indicated within square bracketsPreviously published sequences are designated by strain names (Kissi et al 1995 Randall et al 2004) The tree is rooted with isolate 1500AFSdefined as outgroup representative of the lineage Africa 3 Nodal posterior probabilities gt 0middot95 are shown The scale indicates branch-lengthexpressed as the expected number of substitutions per site Species not definitively identified
Rabies reservoir dynamics in the Serengeti 9
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
recorded continuously throughout the study period whereascases in other species (inside and outside SNP) have beensporadic Although disease monitoring presumably detects alower proportion of cases in wildlife than domestic dogs itseems likely that infection in wildlife populations had it beenpresent continuously would have been detected in more recentyears where the number of samples collected and tested washigher than in previous years when rabies was recorded(Table 2 Fig 3b) These observations provide circumstantialevidence for rabies persistence in domestic dog populationsand lack of persistence in other carnivores Clusters ofwildlife cases that coincided with outbreaks in domestic dogpopulations (Figs 3b 4 amp 5) and explosive but transient(lt 2 months) epidemics previously reported in bat-eared foxes(Maas 1993) are also consistent with chains of transmissionthat are not sustained and contrast with incidence patterns inwild carnivores that are believed to maintain rabies throughindependent cycles elsewhere in Africa For example rabieswas recorded sporadically in bat-eared fox populations inSouth Africa from the 1950s to 1970 then continuouslysuggesting a shift to persistent infection (Thomson amp Meredith1993) and in Zimbabwe outbreaks in jackals although ofteninitiated by domestic dog cases appear to be sustained overseveral years (Bingham et al 1999ab)
Specific demographic attributes (high densities andorturnover) are thought to be required for sustained rabiestransmission A threshold density of gt 5 dogs kmndash2 has beensuggested for persistence of rabies in domestic dog populationsin Africa (Foggin 1988 Brooks 1990 Cleaveland amp Dye 1995Kitala et al 2001) Domestic dog populations in the studyregion have grown considerably and even in the lower-density
area (within the ND) the estimated average density hasapproached this threshold (Table 1) and in 2003 populationsizes were sufficient for a sizable epidemic to occur (Fig 5b)Rabies-control measures make it hard to determine whethercurrent densities in ND are high enough for infection to persistin the absence of vaccination but they may now be close
We detected only one canid-associated variant (Africa 1b)from a range of species in the Serengeti and were unable toreplicate East et alrsquos (2001) finding of a distinct virus variantcirculating atypically in spotted hyaenas (Lembo et al 2007)The lack of species-specific virusndashhost associations in thecanid-associated variant that we detected and the high degreeof genetic relatedness between virus samples from differentspecies indicate frequent cross-species transmission This isconsistent with results from southern Africa where phylogeneticbranches from a single geographic area include several hostspecies (Sabeta Bingham amp Nel 2003)
The statistical association between rabies cases in domesticdogs and other species also indicates cross-species transmissionUnfortunately the length of our time series and paucity ofcases detected in species other than domestic dogs precludeanalyses of lsquospill-overrsquo risk and reduce the interpretativepower of time-lagged regressions like those conducted forrabies in North America that could otherwise provide insight(Guerra et al 2003 Gordon et al 2004) In general as theamount of between-species transmission increases the moredifficult it becomes to tease apart the role of differentpotentially independent maintenance hosts We find greaterwithin-species and less between-species transmission thanwould be expected if species interacted randomly This patterncould occur as a consequence of the spatial distribution ofhost species andor preferential contact between conspecificsOur randomization indicates that large-scale patterns in thedistribution of hosts (gt 10 times 10 km) do not explain thislsquoassortativersquo transmission but we cannot make inference ona smaller scale These analyses suggest that occasionallsquospill-overrsquo of infection from domestic dog populations into
Fig 7 The proportion of estimated transmission within andbetween host types (associated with 1255 suspected rabies cases 1044in Serengeti and 211 in Ngorongoro) in Serengeti (light grey) andNgorongoro (dark grey) districts with lsquodogrsquo corresponding to domesticdogs and lsquocarnivorersquo to domestic cats and wild carnivores Credibilityintervals (95) are shown for expected transmission within andbetween host types assuming inter- and intraspecific transmissionwere equally likely and that livestock are dead-end hosts
Table 4 Human exposures to rabies as a result of bites by suspectedrabid animals in Serengeti and Ngorongoro districts (January 2002ndashDecember 2006) Data were obtained by contact tracing
SpeciesSerengeti district ()
Ngorongorodistrict ()
Domestic dogs 466 (85middot5) 84 (70middot0)Domestic cats 25 (4middot6) 6 (5middot0)Livestock 7 (1middot3) 3 (2middot5)Human 2 (0middot4) 1 (0middot8)Spotted hyaena Crocuta crocuta 4 (0middot7) 16 (13middot3)Honey badger Mellivora capensis 11 (2middot0) 6 (5middot0)African wildcat Felis lybica 0 (0) 2 (1middot7)White-tailed mongoose
Ichneumia albicauda2 (0middot4) 1 (0middot8)
Bat-eared fox Otocyon megalotis 2 (0middot4) 0 (0)Small-spotted genet Genetta genetta 1 (0middot2) 1 (0middot8)Jackal spp Canis spp 23 (4middot2) 0 (0)Leopard Panthera pardus 2 (0middot4) 0 (0)Total 545 (100) 120 (100)
10 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
alternative hosts sometimes results in short-lived chains oftransmission as contact is more common among conspecifics(Dobson 2004)
Definitive identification of infection reservoirs is extremelychallenging and rarely possible Despite intensive effortunderstanding of reservoir dynamics in many complexmulti-host systems remains incomplete (Woodroffe et al2005 Biek et al 2006 Independent Scientific Group on CattleTB 2007) Here the balance of evidence implicates domesticdogs as the only maintenance population of rabies in theSerengeti whereas other carnivores are nonessential tomaintenance However because they transmit disease totarget populations (and frequent transmission within andbetween such species may occur) they do constitute part ofthe reservoir (Table 4 Fig 1a) A single maintenance-hostspecies is consistent with broad patterns of rabies infectionobserved elsewhere (Smith 1989 Rupprecht et al 1991Wandeler et al 1994 Smith Orciari amp Yager 1995)
A lesson from this study is that even with considerable datafrom various sources we cannot exhaustively quantify thecontribution of transmission from all non-essential reservoircomponents However unravelling every thread of reservoirdynamics may not be essential Our key finding that wildlifeare not able to maintain rabies cycles or distinct geneticvariants provides new and critical information allowing rabiescontrol measures in the Serengeti to be targeted moreconfidently at domestic dog populations alone The long-term implication is that control of domestic dog rabies in theSerengeti will eliminate rabies in all other species includinghumans livestock and wildlife The absence of wildlifemaintenance hosts even in an area with such abundantwildlife raises the expectation that canine rabies eliminationis a feasible objective throughout much of Africa
Interventions to prevent transmission from domestic dogmaintenance populations would also provide convincingevidence with which to identify reservoirs Such approacheshave demonstrated that cattle were the reservoir for rinderpestafter cattle vaccinations rinderpest disappeared from wildlifeand the wildebeest Connochaetes taurinus population increasednearly ten-fold (Sinclair 1979 Plowright 1982 Dobson 1995)We are currently ring-vaccinating domestic dogs around SNPwhich should provide critical data to evaluate the impact ofalternative hosts on long-term control efforts in particular theireffects on the level of vaccination required for disease control
In conclusion resolving reservoir dynamics in any complexsystem is likely to prove challenging and our experiences inthe Serengeti suggest that consideration needs to be given toidentifying (i) the potential role of different componentsof the system (ii) the key information needed for differentdisease control strategies (iii) the epidemiological approacheslikely to yield most useful information and (iv) the range ofanalytical tools available
Acknowledgements
We are grateful to Tanzania Government ministries TANAPA TAWIRINCA Authority Tanzania Commission for Science and Technology andNational Institute for Medical Research for permissions and TANAPA and
TAWIRI Veterinary Units Viral Transmission Dynamics Project SerengetiLion and Cheetah Projects Frankfurt Zoological Society livestock field-officersof the Ministry of Water and Livestock Development in Mara and ArushaRegions M Magoto P Tiringa M Roelke-Parker R Fyumagwa H Wiickand Mwanza and Arusha Veterinary Investigation Centres for assistance withsample collection We are indebted to J Barrat AFSSA for performingdiagnostic testing and virus isolation and the staff of the Rabies Section of theCDC especially CE Rupprecht A Velasco-Villa LA Orciari IV Kuzminand M Niezgoda and PE Brandatildeo (Universidade de Satildeo Paulo) for technicalassistance with diagnostic tests and virus characterization We thankKLaurenson D Bennett J Pulliam and H Horn for valuable discussions
This work was supported by the joint National Institutes of Health (NIH)National Science Foundation (NSF) Ecology of Infectious Diseases Programunder grant no NSFDEB0225453 Opinions findings conclusions andrecommendations are those of the authors and do not necessarily reflectthe views of NIH or NSF KH was supported by NSF grant DEB-0513994the Pew Charitable Trusts award 2000-002558 to Princeton University theDepartment of Ecology and Evolution Princeton University and the HeinzFoundation TL was supported by the Royal (Dick) School of VeterinaryStudies University of Edinburgh and the University of Edinburgh DevelopmentTrust (visits to CDC) and SC by the Wellcome Trust and the Department forInternational Development Animal Health Programme
References
Barrat J (1996) Simple technique for the collection and shipment of brainspecimens for rabies diagnosis Laboratory Techniques in Rabies (edsF-X Meslin MM Kaplan amp H Koprowski) pp 425ndash432 World HealthOrganization Geneva Switzerland
Barrat J Barrat MJ Picard M amp Aubert MFA (1988) Diagnosis of rabiesby cell-culture Comparative Immunology Microbiology and InfectiousDiseases 11 207ndash214
Bartlett MS (1960) Stochastic Population Models in Ecology and EpidemiologyMethuen London
Bernardi F Nadin-Davis SA Wandeler AI Armstrong J Gomes AABLima FS Nogueira FRB amp Ito FH (2005) Antigenic and geneticcharacterization of rabies viruses isolated from domestic and wild animalsof Brazil identifies the hoary fox as a rabies reservoir Journal of GeneralVirology 86 3153ndash3162
Biek R Walsh PD Leroy EM amp Real LA (2006) Recent common ancestryof Ebola Zaire virus found in a bat reservoir Public Library of SciencePathogens 2 885ndash886
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999a) Theepidemiology of rabies in Zimbabwe 2 Rabies in jackals (Canis adustus andCanis mesomelas) Onderstepoort Journal of Veterinary Research 66 11ndash23
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999b) Theepidemiology of rabies in Zimbabwe 1 Rabies in dogs (Canis familiaris)Onderstepoort Journal of Veterinary Research 66 1ndash10
Bingham J Javangwe S Sabeta CT Wandeler AI amp Nel LH (2001)Report of isolations of unusual lyssaviruses (rabies and Mokola virus)identified retrospectively from Zimbabwe Journal of the South AfricanVeterinary Association 72 92ndash94
Brooks R (1990) Survey of the dog-population of Zimbabwe and its level ofrabies vaccination Veterinary Record 127 592ndash596
Cleaveland S amp Dye C (1995) Maintenance of a microparasite infectingseveral host species rabies in the Serengeti Parasitology 111 S33ndashS47
Cleaveland S Kaare M Tiringa P Mlengeya T amp Barrat J (2003) A dograbies vaccination campaign in rural Africa impact on the incidence of dograbies and human dog-bite injuries Vaccine 21 1965ndash1973
Cleaveland S Laurenson MK amp Taylor LH (2001) Diseases of humansand their domestic mammals pathogen characteristics host range andthe risk of emergence Philosophical Transactions of the Royal Society BBiological Sciences 356 991ndash999
Cleaveland S Mlengeya T Kaare M Haydon DT Lembo T LaurensonMK amp Packer C (2007) The conservation relevance of epidemiologicalresearch into carnivore viral diseases in the Serengeti Conservation Biology21 612ndash622
Conzelmann KK Cox JH Schneider LG amp Thiel HJ (1990) Molecularcloning and complete nucleotide sequence of the attenuated rabies virusSAD B19 Virology 175 485ndash499
Daszak P Cunningham AA amp Hyatt AD (2000) Wildlife ecology ndash Emerginginfectious diseases of wildlife ndash Threats to Biodiversity and human healthScience 287 443ndash449
Dean DJ Abelseth MK amp Atanasiu P (1996) The fluorescent antibodytest Laboratory Techniques in Rabies (eds F-X Meslin MM Kaplan
Rabies reservoir dynamics in the Serengeti 11
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
amp H Koprowski) pp 88ndash95 World Health Organization GenevaSwitzerland
Dobson A (1995) The ecology and epidemiology of rinderpest virus inSerengeti and Ngorongoro Conservation Area Serengeti II DynamicsManagement and Conservation of an Ecosystem (eds ARE Sinclair ampP Arcese) pp 485ndash505 University of Chicago Press Chicago
Dobson A (2004) Population dynamics of pathogens with multiple host speciesAmerican Naturalist 164 S64ndashS78
Dobson A amp Foufopoulos J (2001) Emerging infectious pathogens of wildlifePhilosophical Transactions of the Royal Society B Biological Sciences 3561001ndash1012
Dye C (1996) Serengeti wild dogs what really happened Trends in Ecology ampEvolution 11 188ndash189
East ML Hofer H Cox JH Wulle U Wiik H amp Pitra C (2001) Regularexposure to rabies virus and lack of symptomatic disease in Serengetispotted hyenas Proceedings of the National Academy of Sciences USA 9815026ndash15031
Foggin CM (1988) Rabies and Rabies-related Viruses in Zimbabwe HistoricalVirological and Ecological Aspects PhD thesis University of ZimbabweHarare Zimbabwe
Gascoyne SC King AA Laurenson MK Borner M Schildger B ampBarrat J (1993) Aspects of rabies infection and control in the conservationof the African wild dog (Lycaon pictus) in the Serengeti region TanzaniaOnderstepoort Journal of Veterinary Research 60 415ndash420
Gordon ER Curns AT Krebs JW Rupprecht CE Real LA ampChilds JE (2004) Temporal dynamics of rabies in a wildlife host and the riskof cross-species transmission Epidemiology and Infection 132 515ndash524
Guerra MA Curns AT Rupprecht CE Hanlon CA Krebs JW ampChilds JE (2003) Skunk and raccoon rabies in the eastern United Statestemporal and spatial analysis Emerging Infectious Diseases 9 1143ndash1150
Hall TA (1999) bioedit a user-friendly biological sequence alignment editorand analysis program for Windows 9598NT Nucleic Acids SymposiumSeries 41 95ndash98
Haydon DT Cleaveland S Taylor LH amp Laurenson MK (2002) Identi-fying reservoirs of infection a conceptual and practical challenge EmergingInfectious Diseases 8 1468ndash1473
Independent Scientific Group on Cattle TB (2007) Bovine TB The ScientificEvidence Defra Publications London PB12634 httpwwwdefragovukanimalhtbisgindexhtm
Jeanmougin F Thompson JD Gouy M Higgins DG amp Gibson TJ(1998) Multiple sequence alignment with clustal-x Trends in BiochemicalSciences 23 403ndash405
Johnson N Black C Smith J Un H McElhinney LM Aylan O ampFooks AR (2003) Rabies emergence among foxes in Turkey Journal ofWildlife Diseases 39 262ndash270
King AA Meredith CD amp Thomson GR (1993) Canid and viverrid rabiesviruses in South Africa Onderstepoort Journal of Veterinary Research 60295ndash299
Kissi B Tordo N amp Bourhy H (1995) Genetic polymorphism in the rabiesvirus nucleoprotein gene Virology 209 526ndash537
Kitala PM McDermott JJ Kyule MN amp Gathuma JM (2000) Community-based active surveillance for rabies in Machakos District Kenya PreventiveVeterinary Medicine 44 73ndash85
Kitala PM McDermott J Kyule M Gathuma JM Perry B amp Wandeler A(2001) Dog ecology and demography information to support the planning ofrabies control in Machakos District Kenya Acta Tropica 78 217ndash230
Knobel DL Cleaveland S Coleman PG Fevre EM Meltzer MIMiranda MEG Shaw A Zinsstag J amp Meslin F-X (2005) Re-evaluatingthe burden of rabies in Africa and Asia Bulletin of the World HealthOrganization 83 360ndash368
Lembo T Haydon DT Velasco-Villa A Rupprecht CE Packer CBrandatildeo PE Kuzmin IV Fooks AR Barrat J amp Cleaveland S (2007)Molecular epidemiology identifies only a single rabies virus variantcirculating in complex carnivore communities of the Serengeti Proceedingsof the Royal Society B Biological Sciences 274 2123ndash2130
Maas B (1993) Behavioural Ecology and Social Organisation of the Bat-earedFox in the Serengeti National Park Tanzania University of CambridgeCambridge UK
National Bureau of Statistics Tanzania (2005) 2002 Population and HousingCensus Presidentrsquos Office Planning and Privatization Dar es SalaamTanzania
Nel JAJ (1993) The bat-eared fox-a prime candidate for rabies vectorOnderstepoort Journal of Veterinary Research 60 395ndash397
Nel LH Sabeta CT von Teichman B Jaftha JB Rupprecht CE amp
Bingham J (2005) Mongoose rabies in southern Africa a re-evaluationbased on molecular epidemiology Virus Research 109 165ndash173
Page RDM (1996) treeview an application to display phylogenetic trees onpersonal computers Computer Applications in the Biosciences 12 357ndash358
Plowright W (1982) The effects of rinderpest and rinderpest control on wildlifein Africa Symposia of the Zoological Society of London 50 1ndash28
Posada D amp Crandall KA (1998) modeltest testing the model of DNAsubstitution Bioinformatics 14 817ndash818
Randall DA Marino J Haydon DT Sillero-Zubiri C Knobel DLTallents LA Macdonald DW amp Laurenson MK (2006) An integrateddisease management strategy for the control of rabies in Ethiopian wolvesBiological Conservation 131 151ndash162
Randall DA Williams SD Kuzmin IV Rupprecht CE Tallents LATefera Z Argaw K Shiferaw F Knobel DL Sillero-Zubiri C ampLaurenson MK (2004) Rabies in endangered Ethiopian wolves EmergingInfectious Diseases 10 2214ndash2217
Rhodes CJ Atkinson RPD Anderson RM amp Macdonald DW (1998)Rabies in Zimbabwe reservoir dogs and the implications for disease controlPhilosophical Transactions of the Royal Society B Biological Sciences 353999ndash1010
Roelke-Parker ME Munson L Packer C Kock R Cleaveland SCarpenter M Obrien SJ Pospischil A HofmannLehmann R Lutz HMwamengele GLM Mgasa MN Machange GA Summers BA ampAppel MJG (1996) A canine distemper virus epidemic in Serengeti lions(Panthera leo) Nature 379 441ndash445
Ronquist F amp Huelsenbeck JP (2003) mrbayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 19 1572ndash1574
Rupprecht CE Dietzschold B Wunner WH amp Koprowsky H (1991)Antigenic relationships of lyssaviruses The Natural History of Rabies (edsGM Baer) pp 69ndash100 CRC Press Boca Raton
Sabeta CT Bingham J amp Nel LH (2003) Molecular epidemiology of canidrabies in Zimbabwe and South Africa Virus Research 91 203ndash211
Sinclair ARE (1979) The eruption of the ruminants Serengeti Dynamics ofan Ecosystem (eds ARE Sinclair amp M Norton-Griffiths) pp 82ndash103University of Chicago Press Chicago
Sinclair ARE amp Arcese P (1995) Serengeti II Dynamics Management andConservation of an Ecosystem University of Chicago Press Chicago
Smith JS (1989) Rabies virus epitopic variation use in ecological studiesAdvances in Virus Research 36 215ndash253
Smith JS Orciari LA amp Yager PA (1995) Molecular epidemiology ofrabies in the United States Seminars in Virology 6 387ndash400
Swanepoel R Barnard BJH Meredith CD Bishop GC BrucknerGK Foggin CM amp Hubschle OJB (1993) Rabies in Southern AfricaOnderstepoort Journal of Veterinary Research 60 325ndash346
Taylor LH Latham SM amp Woolhouse MEJ (2001) Risk factors forhuman disease emergence Philosophical Transactions of the Royal SocietyB Biological Sciences 356 983ndash989
Tepsumethanon V Wilde H amp Meslin FX (2005) Six criteria for rabiesdiagnosis in living dogs Journal of the Medical Association of Thailand 88419ndash422
Thomson GR amp Meredith CD (1993) Rabies in bat-eared foxes in SouthAfrica Onderstepoort Journal of Veterinary Research 60 399ndash403
Vial F Cleaveland S Rasmussen G amp Haydon DT (2006) Development ofvaccination strategies for the management of rabies in African wild dogsBiological Conservation 131 180ndash192
Wandeler AI Budde A Capt S Kappeler A amp Matter H (1988) Dogecology and dog rabies control Reviews of Infectious Diseases 10 S684ndashS688
Wandeler AI Nadin-Davis SA Tinline RR amp Rupprecht CE (1994)Rabies epidemiology some ecological and evolutionary perspectivesLyssaviruses (eds CE Rupprecht B Dietzschold amp H Koprowski)pp 297ndash324 Springer-Verlag Berlin
Woodroffe R (2001) Assessing the risks of intervention immobilizationradio-collaring and vaccination of African wild dogs Oryx 35 234ndash244
Woodroffe R Donnelly CA Johnston WT Bourne FJ Cheeseman CLClifton-Hadley RS Cox DR Gettinby G Hewinson RG Le FevreAM McInerney JP amp Morrison WI (2005) Spatial association ofMycobacterium bovis infection in cattle and badgers Meles meles Journal ofApplied Ecology 42 852ndash862
World Health Organization (1999) World Survey of Rabies No 34 for the Year1998 Geneva Switzerland
Received 14 June 2007 accepted 23 January 2008Handling Editor Simon Thirgood
12 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Supplementary material
The following supplementary material is available for thisarticle
Appendix S1 Latin names of wildlife species
Appendix S2 Supplementary details of methods
Table S1 Virus samples from the study area compared inphylogenetic analysis
Table S2 Details of all wildlife confirmed and suspect casesin SNP
Table S3 Details of confirmed wildlife cases in Serengeti andNgorongoro districts
Fig S1 Incidence of bites by domestic dogs in Serengeti andNgorongoro districts
This material is available as part of the online article fromhttpwwwblackwell-synergycomdoifull101111j1365-2664200801468x(This link will take you to the article abstract)
Please note Blackwell Publishing is not responsible for the con-tent or functionality of any supplementary materials suppliedby the authors Any queries (other than missing material)should be directed to the corresponding author for the article

Rabies reservoir dynamics in the Serengeti
3
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society
Journal of Applied Ecology
(Cleaveland amp Dye 1995 Rhodes
et al
1998 Bingham
et al
1999b)
Wild carnivore communities in the Serengeti comprise26 species (Sinclair amp Arcese 1995) including Canidae andHerpestidae species implicated as independent maintenancehosts of rabies in parts of southern Africa (ie slendermongooses bat-eared foxes side-striped and black-backedjackals) Cleaveland amp Dye (1995) found that rabies appears topersist endemically in high-density domestic dog populations(gt 5 km
ndash2
) to the west of Serengeti National Park (SNP) yetoccurred only sporadically in lower-density domestic dogpopulations (lt 1 km
ndash2
) to the east of SNP and in wild carnivorepopulations They concluded that domestic dog populationsto the west of SNP were likely to be the sole reservoir in theecosystem (Fig 1a) however their study was based on limiteddata and did not consider the possibility of a multi-hostreservoir Furthermore with rapid growth of human andassociated domestic dog populations it seems likely that inthe meantime the lower-density domestic dog populationwhich previously could not support rabies cycles has becomelarge enough to maintain infection Additionally East
et al
(2001) argue that in the Serengeti a distinct rabies virusvariant is maintained in spotted hyaenas
Crocuta crocuta
which undergo atypical infections with no evidence of mor-tality Additional data now allow more detailed examinationof the role of wild carnivores in potential rabies reservoirsystems (i) as maintenance populations independent ofdomestic dogs (Fig 1b) (ii) as part of multi-host maintenancecommunities (Fig 1c) or (iii) as one of several independentmaintenance populations (Fig 1d)
Given the potential complexity of rabies reservoirs incommunities comprising multiple hosts it may not be possibleto reach an exhaustive understanding of their structure How-ever identifying maintenance populations and populationsthat act as sources of infection for target populations hasimportant practical implications Rabies is transmitted toseveral lsquospill-overrsquo hosts that might be considered targetpopulations of concern including humans (Knobel
et al
2005)endangered wildlife (Randall
et al
2006 Vial
et al
2006)and livestock In the Serengeti human rabies deaths occursubsistence farmers suffer livestock losses from rabies andthere is concern about risks to recovering populations ofAfrican wild dog
Lycaon pictus
[rabies was previouslyimplicated in their local extinction (Cleaveland
et al
2007)]There are long-term benefits to controlling rabies infection
directly in maintenance rather than in target populationshuman post-exposure prophylaxis (PEP) although effectiveis costly and often unavailable routine prophylactic vaccinationof livestock is expensive and vaccination of endangeredcanids has been controversial (Dye 1996 Woodroffe 2001Cleaveland
et al
2007) Approaches to control will dependupon how rabies is maintained if domestic dogs are the solemaintenance population or neither domestic dogs nor wildcarnivores maintain rabies independently (Fig 1ac) measurestargeted at domestic dogs should in theory also eliminatehuman and animal rabies However if wild carnivores are thesole maintenance population (Fig 1b) strategies will need totarget wildlife Similarly if domestic dog and wild carnivorepopulations each maintain rabies (Fig 1d) strategies willonly be successful if infection in both populations is controlled
Rabies in the Serengeti is therefore an important thoughchallenging model for addressing questions relating to thestructure and dynamics of complex reservoir systems Herewe present several lines of evidence that advance our under-standing of reservoir dynamics of rabies in multi-host carnivorecommunities of the Serengeti we synthesize data fromlong-term epidemiological records phylogenetic analyses ofvirus isolates from a range of domestic and wild species andstatistical analyses of high-resolution spatiotemporal patternsof rabies incidence
Methods
STUDY
AREA
The study area in the Serengeti ecological region of northwestTanzania was divided into three zones (Fig 2) The first zoneSNP comprises diverse wildlife communities with domestic dogsfound extremely rarely The two other zones are districts adjacent toSNP Serengeti District (SD) to the west and Ngorongoro District(ND) comprising the Loliondo Game Control Area (LGCA) andNgorongoro Conservation Area (NCA) to the east SD is inhabitedby multi-ethnic agro-pastoralist communities and high-densitydomestic dog populations (gt 11 km
ndash2
) ND is a multiple-use controlledwildlife area inhabited by low-density pastoralist communitiesand lower-density domestic dog populations (lt 5 km
ndash2
) We haveobserved a greater variety and abundance of wild carnivores in NDthan SD No physical barriers separate the wildlife-protected areasand human settlements
Estimated densities of human and domestic dog populations arepresented in Table 1 these were extrapolated from human census
Table 1 Estimated domestic dog population densities in Serengeti (SD) and Ngorongoro districts [ND ndash Loliondo Game Control Area(LGCA) only] Confidence intervals about the means are indicated in brackets
AreaArea (km2)
Domestic dogndashhuman ratio
Average human density (kmndash2) at the village level
Average domestic dog density(kmndash2) at the village level
SD urban (Mugumu town) 18middot8 6middot6 619middot8 93middot9SD rural 3128middot0 6middot6 67middot9 (59middot2ndash76middot5) 10middot3 (9middot0ndash11middot6)SD overall 3146middot8 6middot6 75middot2 (58middot4ndash92middot0) 11middot4 (8middot9ndash13middot9)LGCA urban (Loliondo and Sakala) 14middot1 6middot7 144middot9 21middot6LGCA rural 8852middot4 6middot7 17middot0 (11middot1ndash23middot0) 2middot5 (1middot7ndash3middot4)LGCA overall 8866middot5 6middot7 28middot2 (9middot2ndash47middot1) 4middot2 (1middot4ndash7middot0)
4
T Lembo
et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society
Journal of Applied Ecology
data with projected annual growth rates of 2middot6 in SD and 3middot8 inND (National Bureau of Statistics Tanzania 2005) and from humanndashdomestic dog ratios (Wandeler
et al
1988 Brooks 1990) obtainedfrom household questionnaires (from SD in 2003 and LGCA in2004)
Several domestic dog vaccination programmes have beenimplemented in the study area Details of vaccination campaigns arereported in Appendix S2 (Supplementary material) although theirimpact is not analysed here The only vaccination of wildlife (Africanwild dogs) was conducted in 1990 (Gascoyne
et al
1993)
D ISEASE
MONITORING
OPERATIONS
Data presented are the result of disease monitoring operations sincethe late 1980s (Table 2) Passive surveillance was used to detectrabies cases in SNP Sightings of sick and dead carnivores werereported through a network of veterinarians from Tanzania NationalParks (TANAPA) and Tanzania Wildlife Research Institute(TAWIRI) rangers scientists and tour-operators Cases includedanimals that were snared injuredwounded from other or unknowncauses observed with signs of disease or found dead with unknownhistory
Outside SNP passive and active surveillance operations wereemployed Passive surveillance data were available through veterinaryoffice records (government offices TAWIRI TANAPA and NCAAuthority) and district hospitals and medical dispensaries Biteinjuries to humans from suspected rabid dogs were defined based onevaluation by clinicians (from information on clinical signs reportedby bite victims) and when human vaccines were not widely availableon whether PEP had been administered Bite injury incidence wascalculated against human population sizes interpolated from census
data (National Bureau of Statistics Tanzania 2005) Community-based active surveillance measures based on previous studies inrural Kenya (Kitala
et al
2000) and Tanzania (Cleaveland
et al
2003) were implemented in SD using livestock field officersstationed in randomly selected villages to collect information onrabies cases from key informants (village leaders teachers dispensarystaff local healers)
Contact tracing was used to collect high-resolution data on spatialand temporal patterns of disease Visits to incidents involving bitinganimals (to bite victims and owners of biting and bitten animals)were initiated using hospital and medical dispensary records andcase reports from livestock offices and community-based surveillanceIncidents were mapped and witnesses interviewed to make a diagnosis(see below) and obtain case-history information (exposure sourceonset of clinical signs and death resulting exposures) The sameprocedure was followed for all exposures and preceding cases whereidentified so that transmission events between identified individualanimals could be traced and confirmed When multiple incidentsinvolving suspected rabid wildlife were reported on the sameconsecutive days within neighbouring homesteads we assumed asingle animal was involved
Wherever possible brain stem samples were collected from suspectrabies cases and carnivore carcasses whatever the apparent cause ofdeath Sample collection was carried out by research personnelveterinarians and livestock field officers the preferred techniquewas removal via the occipital foramen according to World HealthOrganization recommendations (Barrat 1996) Specimens were frozen(ndash20
deg
C) immediately after collection or placed into phosphate-buffered 50 glycerol solution and preserved either cold (+4
deg
Cndash20
deg
C) or at room temperature (25 plusmn 5
deg
C) when refrigerationfacilities were not promptly available
Fig 2 Map of the Serengeti ecological regionillustrating the distribution of suspectedrabies cases (most were not laboratory-confirmed but diagnosed using epidemio-logical history and clinical criteria)Tables S2 and S3 in Supplementary materialdetail confirmed wildlife cases in SerengetiNational Park (SNP) and Serengeti (SD) andNgorongoro (ND) districts Cases recordedin SNP from January 1995ndashJanuary 2007 aremapped whereas only cases from January2002ndashJanuary 2007 in SD and ND aremapped because GPS data were not availablefor the earlier period Domestic dog cases areshown in black (SD n = 833 ND n = 128)and cases in other carnivores as white circles(SD n = 96 ND n = 27 SNP n = 23)Village boundaries are indicated by greylines LGCA Loliondo Game Control AreaNCA Ngorongoro Conservation Area
Rabies reservoir dynamics in the Serengeti
5
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society
Journal of Applied Ecology
RABIES
D IAGNOSIS
AND
V IRUS
CHARACTERIZATION
Diagnostic tests on brains collected up to 2001 and virus isolationwere carried out at the Agence Franccedilaise de Seacutecuriteacute Sanitaire desAliments (AFSSA) Malzeacuteville France using the fluorescent antibodytest (Dean Abelseth amp Atanasiu 1996) inoculation of murineneuroblastoma cells and mouse inoculation (Barrat
et al
1988)Rabies diagnosis on more recent brain tissues was conducted at theRabies Section of the Centers for Disease Control and Prevention(CDC) Atlanta USA by the fluorescent antibody test (httpwwwcdcgovrabiesdocsstandard_dfa_protocol_rabiespdf) Virussamples were typed at CDC RNA was extracted reverse transcribedamplified by polymerase chain reaction and nucleoprotein genesequences generated as described in Lembo
et al
(2007)When brain tissues were not available for laboratory confirmation
diagnosis of suspected rabies cases was based on history of exposureand clinical evaluation (adapting the six criteria proposed byTepsumethanon Wilde amp Meslin 2005) from information on clinicalsigns reported by villagers livestock field officers researchers andpark veterinarians
1
For owned domestic dogs history of a bite clinical signs(Tepsumethanon
et al
2005) disappearance or death within 10 days
2
For domestic dogs of unknown origin clinical signs (as above) anddisappearance or death
3
For wild carnivores clinical signs (as above) plus tameness lossof fear of humans diurnal activity (for nocturnal species) andunprovoked biting (without eating) of objects and animals
PHYLOGENETIC
ANALYSIS
Nucleotide sequences were edited using
bioedit
7middot0middot0 (Hall 1999)and aligned in
clustal-x
version 1middot83 (Jeanmougin
et al
1998)Phylogenetic relationships among viruses from the study area(Table S1 Supplementary material) and sequence data for RABVsfrom other African countries (Kissi Tordo amp Bourhy 1995 Randall
et al
2004) were estimated using Bayesian Markov chain MonteCarlo (MCMC) methods using
mrbayes
3middot0b4 (Ronquist ampHuelsenbeck 2003) as described in Lembo
et al
(2007) Posteriorprobabilities of 0middot95 or greater were considered significant Graphicoutput was generated using
treeview
version 1middot6middot6 (Page 1996)
STATISTICAL
ANALYSIS
Poisson regression was used to test for a relationship between rabiescases in domestic dogs and in other carnivores Using maximumlikelihood a gamma-distributed spatial infection kernel for domesticdogs k(
d
) was estimated from the distances between source animalsand their known contacts and a probability distribution of thegeneration interval for domestic dogs g(
t
) was estimated fromobserved dates of exposure onset of clinical signs and death (fulldetails in Appendix S2) An algorithm (detailed in Appendix S2)was developed to generate putative epidemic trees by assigningprobabilities to links between possible progenitors and each suspectedcase according to their spatiotemporal proximity and the estimatedspatial infection kernel k(
d
) and generation interval distributiong(
t
) This algorithm assumes all cases are observed and thereforecannot account for unobserved intermediates Cases in all specieswere included in the analysis and all carnivore species were assumedequally likely to transmit rabies Only limited data could be collectedon transmission from wild carnivores therefore progenitors of allspecies were assigned using parameters estimated for domestic dogs
Tab
le 2
Tim
elin
e of
dis
ease
mon
itor
ing
oper
atio
ns in
eac
h zo
ne in
6-m
onth
uni
ts T
AW
IRI
Tan
zani
a W
ildlif
e R
esea
rch
Inst
itut
e T
AN
APA
Tan
zani
a N
atio
nal P
arks
NC
AA
Ngo
rong
oro
Con
serv
atio
nA
rea
Aut
hori
ty
YE
AR
ZO
NE
1987
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
SN
P
TA
NA
PA V
eter
inar
y U
nit
TA
WIR
I V
eter
inar
y P
rogr
amm
e
Vir
al T
rans
mis
sion
D
ynam
ics
Pro
ject
SD
Pas
sive
sur
veill
ance
(do
mes
tic
dog-
bite
inju
ry r
ecor
ds f
rom
hos
pita
ls)
D
omes
tic
dog
vacc
inat
ions
Pas
sive
sur
veill
ance
(V
eter
inar
y re
cord
s
villa
ge r
epor
ts o
ppor
tuni
stic
car
cass
sam
plin
g)
Act
ive
(liv
esto
ck fi
eld-
offic
ers)
Act
ive
(liv
esto
ck fi
eld-
offic
ers)
Con
tact
tra
cing
ND
Pas
sive
(do
mes
tic
dog-
bite
inju
ry r
ecor
ds
from
hos
pita
ls)
Dom
esti
c do
g va
ccin
atio
nsP
assi
ve (
Vet
erin
ary
reco
rds
opp
ortu
nist
ic c
arca
ss
sam
plin
g T
AW
IRI
TA
NA
PA a
nd N
CA
A r
ecor
ds)
Con
tact
tra
cing
A r
esea
rch
proj
ect
inve
stig
atin
g in
fect
ion
dyna
mic
s of
vir
al d
isea
ses
6 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
(k(d) and g(t)) One thousand bootstrapped trees were generatedand the proportional contribution of transmission to each case frompreceding cases was calculated If the date of onset of clinical signswas uncertain in each realization a date was assigned with uniformprobability from the range of uncertainty
From the bootstrap trees the amount of transmission withinand between host types was estimated We pooled all carnivoresexcept domestic dogs because of small sample sizes A null model ofthe expected frequency of within- and between-species transmissionassuming that intra- and interspecific transmission events occurwith the same probability (ie transmission is non-assortative)was estimated as follows the species identity of each case wasrandomly reassigned retaining the original distribution of casesamongst species and the resulting frequency of within- andbetween-species transmission was calculated This procedure wasiterated 1000 times to evaluate the probability that the differencebetween inferred within- and between-species transmissionfrequencies could have been due to chance To investigate whetherthe spatial distribution of hosts influenced the estimated patterns oftransmission we repeated the analysis but randomly reassignedspecies identities within grid cells rather than across the entiredistrict and we repeated the analysis for different sized grids (seeAppendix S2)
Results
RABIES RECOGNIT ION PROBABIL ITY
The sensitivity specificity and positive predictive value ofclinical rabies diagnosis (ie recognition of rabies by villagerslivestock field officers park veterinarians and researchpersonnel) calculated against the gold standard fluorescentantibody test (sensitivity ~99 specificity gt 99 Dean et al1996) are reported in Table 3 As a result of the relatively highrabies recognition probability (gt 74 of animals reported assuspect rabies cases were confirmed positive) and the smallnumber of confirmed cases the analyses presented include allsuspected cases unless otherwise stated
RABIES INCIDENCE IN THE STUDY POPULATIONS
Wildlife monitoring activities in SNP are shown throughoutthe study period in Table 2 and Fig 3a A peak in wild
carnivores reported dead and sick coincided with a caninedistemper outbreak in late 1993 and 1994 (Roelke-Parkeret al 1996) Most rabies cases in wildlife were reported andconfirmed in 1998 and 1999 with only sporadic detection inother years (Fig 3b Table S2 in Supplementary material) Lownumbers of snared animals were reported in 1998 suggestingthat the increase in carnivore carcasses recorded that year(Fig 3a) was not due to increased monitoring No wildlifecases have been confirmed in SNP since 1999 except for aleopard Panthera pardus on the SNPndashNCA boundary inSeptember 2004 when there was an outbreak among adjacentdomestic dog populations (shown in Fig 5b)
Animal-bite injuries in humans from suspected rabiddomestic dogs were reported continuously in high-densitydomestic dog populations to the west of SNP before imple-mentation of dog vaccination programmes whereas in the
Table 3 Sensitivity specificity and positive predictive value of rabiesdiagnosis calculated against the gold standard fluorescent antibodytest Data include only cases for which complete history was available(2002ndash2005)
Clinical diagnosis
Laboratory diagnosis
TotalPositive Negative
Positive 55 19 74Negative 0 33 33Total 55 52 107Sensitivity = 100middot00Specificity = 63middot46 (95 CI 50middot37ndash76middot55)Positive predictive value = 74middot32 (95 CI 64middot37ndash84middot28)
Fig 3 Wildlife disease monitoring in Serengeti National Park(a) carnivore reports and (b) samples retrieved for rabies diagnosisThe peak in 1994 coincided with a canine distemper outbreak inlions (Roelke-Parker et al 1996)
Rabies reservoir dynamics in the Serengeti 7
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
lower-density populations to the east incidence was lowerand there were several periods when no bites were recorded(Fig 4) Gaps in recorded bites by suspected rabid dogs mayalso reflect PEP shortages however when we included alldomestic dog bite injuries there were four periods of at least5 months with no reported bite injuries in ND and no equivalentpattern in SD (Fig S1 Supplementary material)
We traced 1255 suspected rabid animals from January 2002to January 2007 (1044 in SD 211 in ND see Fig 5 for carnivorecases) In both districts cases in wildlife were sporadic andcoincided with outbreaks in domestic dogs (Figs 3b 4 amp 5)The spatial and temporal distribution of suspected cases isshown in Figs 2 and 5
INTER- AND INTRA-SPECIF IC TRANSMISSION
The virus
All of the 57 RABV specimens recovered from a range ofspecies from the study area (including spotted hyaenas) fellwithin the Africa 1b group of canid-associated viruses andrevealed no clustering by host species (Fig 6) The evidentintermingling of viral lineages from domestic dogs andwildlife indicates frequent interspecific transmission Analysesusing statistical parsimony techniques were also consistentwith both within- and between-species transmission events(Lembo et al 2007)
Fig 4 Incidence of bites by suspected rabiddomestic dogs in Serengeti (SD black line)and Ngorongoro (ND grey line) districtsderived from hospital records Incidence isshown only for years before the implementa-tion of domestic dog vaccination programmesin the study populations (July 1987ndashJanuary1997 in SD and January 1994ndashDecember2003 in ND) No data were available for6 months during 1993 in SD
Fig 5 Suspected rabies cases amongcarnivore species in (a) Serengeti (n = 929)and (b) Ngorongoro districts (n = 155)monitored by contact tracing from January2001 to January 2007
8 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Spatiotemporal incidence patterns in hosts
Suspected rabies cases in domestic dogs were significantpredictors of those in other carnivores (pooled cases indomestic cats and wildlife P lt 0middot001 in both districts)Credibility intervals (95) of the expected frequency ofwithin- and between-species transmission events based uponthe null assumption of non-assortative transmission (derivedfrom the bootstrap analysis) are shown in Fig 7 Morewithin-species transmission than expected was estimated inboth districts (domestic dog-to-domestic dog transmissionP = 0middot052 in SD and P = 0middot008 in ND transmission withinother carnivores ie wild carnivores and domestic catsFelis catus P lt 0middot0001 in both districts) Estimated between-species transmission was less than expected (P lt 0middot03 in SD
and P lt 0middot001 in ND) except for transmission from domesticdogs to livestock These patterns were consistent and significantwhen the randomization procedure was carried out on a10 times 10 km grid while on a 5 times 5 km grid the patterns wereevident but no longer significant probably because of reducedstatistical power
The majority of human exposures to suspected rabid animalswere from domestic dogs (Table 4) Similarly domestic dogswere the predominant source of infection for livestock (Fig 7)
Discussion
Demonstrating persistence of infection is a prerequisite foridentifying reservoir(s) Domestic dogs living at high densities(within the SD) are the only species in which rabies has been
Fig 6 Phylogenetic tree of nucleoprotein gene sequences (282 bp 94 deduced amino acids nucleotide positions 1139ndash1420 on the SAD B19genome (Conzelmann et al 1990) for RABV samples from the study area compared with isolates from other areas of Africa The tree isconstructed using Bayesian phylogenetics under the transitional model of nucleotide evolution with a gamma-shaped distribution of rates acrosssites (TIM + Γ Posada amp Crandall 1998 base frequencies = 0middot3253 0middot2134 0middot2360 0middot2253 nucleotide substitution rates = 1middot0000 3middot67230middot4393 0middot4393 8middot1773 1middot0000 Γ = 0middot3390) For samples from the study area the year of collection is indicated within square bracketsPreviously published sequences are designated by strain names (Kissi et al 1995 Randall et al 2004) The tree is rooted with isolate 1500AFSdefined as outgroup representative of the lineage Africa 3 Nodal posterior probabilities gt 0middot95 are shown The scale indicates branch-lengthexpressed as the expected number of substitutions per site Species not definitively identified
Rabies reservoir dynamics in the Serengeti 9
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
recorded continuously throughout the study period whereascases in other species (inside and outside SNP) have beensporadic Although disease monitoring presumably detects alower proportion of cases in wildlife than domestic dogs itseems likely that infection in wildlife populations had it beenpresent continuously would have been detected in more recentyears where the number of samples collected and tested washigher than in previous years when rabies was recorded(Table 2 Fig 3b) These observations provide circumstantialevidence for rabies persistence in domestic dog populationsand lack of persistence in other carnivores Clusters ofwildlife cases that coincided with outbreaks in domestic dogpopulations (Figs 3b 4 amp 5) and explosive but transient(lt 2 months) epidemics previously reported in bat-eared foxes(Maas 1993) are also consistent with chains of transmissionthat are not sustained and contrast with incidence patterns inwild carnivores that are believed to maintain rabies throughindependent cycles elsewhere in Africa For example rabieswas recorded sporadically in bat-eared fox populations inSouth Africa from the 1950s to 1970 then continuouslysuggesting a shift to persistent infection (Thomson amp Meredith1993) and in Zimbabwe outbreaks in jackals although ofteninitiated by domestic dog cases appear to be sustained overseveral years (Bingham et al 1999ab)
Specific demographic attributes (high densities andorturnover) are thought to be required for sustained rabiestransmission A threshold density of gt 5 dogs kmndash2 has beensuggested for persistence of rabies in domestic dog populationsin Africa (Foggin 1988 Brooks 1990 Cleaveland amp Dye 1995Kitala et al 2001) Domestic dog populations in the studyregion have grown considerably and even in the lower-density
area (within the ND) the estimated average density hasapproached this threshold (Table 1) and in 2003 populationsizes were sufficient for a sizable epidemic to occur (Fig 5b)Rabies-control measures make it hard to determine whethercurrent densities in ND are high enough for infection to persistin the absence of vaccination but they may now be close
We detected only one canid-associated variant (Africa 1b)from a range of species in the Serengeti and were unable toreplicate East et alrsquos (2001) finding of a distinct virus variantcirculating atypically in spotted hyaenas (Lembo et al 2007)The lack of species-specific virusndashhost associations in thecanid-associated variant that we detected and the high degreeof genetic relatedness between virus samples from differentspecies indicate frequent cross-species transmission This isconsistent with results from southern Africa where phylogeneticbranches from a single geographic area include several hostspecies (Sabeta Bingham amp Nel 2003)
The statistical association between rabies cases in domesticdogs and other species also indicates cross-species transmissionUnfortunately the length of our time series and paucity ofcases detected in species other than domestic dogs precludeanalyses of lsquospill-overrsquo risk and reduce the interpretativepower of time-lagged regressions like those conducted forrabies in North America that could otherwise provide insight(Guerra et al 2003 Gordon et al 2004) In general as theamount of between-species transmission increases the moredifficult it becomes to tease apart the role of differentpotentially independent maintenance hosts We find greaterwithin-species and less between-species transmission thanwould be expected if species interacted randomly This patterncould occur as a consequence of the spatial distribution ofhost species andor preferential contact between conspecificsOur randomization indicates that large-scale patterns in thedistribution of hosts (gt 10 times 10 km) do not explain thislsquoassortativersquo transmission but we cannot make inference ona smaller scale These analyses suggest that occasionallsquospill-overrsquo of infection from domestic dog populations into
Fig 7 The proportion of estimated transmission within andbetween host types (associated with 1255 suspected rabies cases 1044in Serengeti and 211 in Ngorongoro) in Serengeti (light grey) andNgorongoro (dark grey) districts with lsquodogrsquo corresponding to domesticdogs and lsquocarnivorersquo to domestic cats and wild carnivores Credibilityintervals (95) are shown for expected transmission within andbetween host types assuming inter- and intraspecific transmissionwere equally likely and that livestock are dead-end hosts
Table 4 Human exposures to rabies as a result of bites by suspectedrabid animals in Serengeti and Ngorongoro districts (January 2002ndashDecember 2006) Data were obtained by contact tracing
SpeciesSerengeti district ()
Ngorongorodistrict ()
Domestic dogs 466 (85middot5) 84 (70middot0)Domestic cats 25 (4middot6) 6 (5middot0)Livestock 7 (1middot3) 3 (2middot5)Human 2 (0middot4) 1 (0middot8)Spotted hyaena Crocuta crocuta 4 (0middot7) 16 (13middot3)Honey badger Mellivora capensis 11 (2middot0) 6 (5middot0)African wildcat Felis lybica 0 (0) 2 (1middot7)White-tailed mongoose
Ichneumia albicauda2 (0middot4) 1 (0middot8)
Bat-eared fox Otocyon megalotis 2 (0middot4) 0 (0)Small-spotted genet Genetta genetta 1 (0middot2) 1 (0middot8)Jackal spp Canis spp 23 (4middot2) 0 (0)Leopard Panthera pardus 2 (0middot4) 0 (0)Total 545 (100) 120 (100)
10 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
alternative hosts sometimes results in short-lived chains oftransmission as contact is more common among conspecifics(Dobson 2004)
Definitive identification of infection reservoirs is extremelychallenging and rarely possible Despite intensive effortunderstanding of reservoir dynamics in many complexmulti-host systems remains incomplete (Woodroffe et al2005 Biek et al 2006 Independent Scientific Group on CattleTB 2007) Here the balance of evidence implicates domesticdogs as the only maintenance population of rabies in theSerengeti whereas other carnivores are nonessential tomaintenance However because they transmit disease totarget populations (and frequent transmission within andbetween such species may occur) they do constitute part ofthe reservoir (Table 4 Fig 1a) A single maintenance-hostspecies is consistent with broad patterns of rabies infectionobserved elsewhere (Smith 1989 Rupprecht et al 1991Wandeler et al 1994 Smith Orciari amp Yager 1995)
A lesson from this study is that even with considerable datafrom various sources we cannot exhaustively quantify thecontribution of transmission from all non-essential reservoircomponents However unravelling every thread of reservoirdynamics may not be essential Our key finding that wildlifeare not able to maintain rabies cycles or distinct geneticvariants provides new and critical information allowing rabiescontrol measures in the Serengeti to be targeted moreconfidently at domestic dog populations alone The long-term implication is that control of domestic dog rabies in theSerengeti will eliminate rabies in all other species includinghumans livestock and wildlife The absence of wildlifemaintenance hosts even in an area with such abundantwildlife raises the expectation that canine rabies eliminationis a feasible objective throughout much of Africa
Interventions to prevent transmission from domestic dogmaintenance populations would also provide convincingevidence with which to identify reservoirs Such approacheshave demonstrated that cattle were the reservoir for rinderpestafter cattle vaccinations rinderpest disappeared from wildlifeand the wildebeest Connochaetes taurinus population increasednearly ten-fold (Sinclair 1979 Plowright 1982 Dobson 1995)We are currently ring-vaccinating domestic dogs around SNPwhich should provide critical data to evaluate the impact ofalternative hosts on long-term control efforts in particular theireffects on the level of vaccination required for disease control
In conclusion resolving reservoir dynamics in any complexsystem is likely to prove challenging and our experiences inthe Serengeti suggest that consideration needs to be given toidentifying (i) the potential role of different componentsof the system (ii) the key information needed for differentdisease control strategies (iii) the epidemiological approacheslikely to yield most useful information and (iv) the range ofanalytical tools available
Acknowledgements
We are grateful to Tanzania Government ministries TANAPA TAWIRINCA Authority Tanzania Commission for Science and Technology andNational Institute for Medical Research for permissions and TANAPA and
TAWIRI Veterinary Units Viral Transmission Dynamics Project SerengetiLion and Cheetah Projects Frankfurt Zoological Society livestock field-officersof the Ministry of Water and Livestock Development in Mara and ArushaRegions M Magoto P Tiringa M Roelke-Parker R Fyumagwa H Wiickand Mwanza and Arusha Veterinary Investigation Centres for assistance withsample collection We are indebted to J Barrat AFSSA for performingdiagnostic testing and virus isolation and the staff of the Rabies Section of theCDC especially CE Rupprecht A Velasco-Villa LA Orciari IV Kuzminand M Niezgoda and PE Brandatildeo (Universidade de Satildeo Paulo) for technicalassistance with diagnostic tests and virus characterization We thankKLaurenson D Bennett J Pulliam and H Horn for valuable discussions
This work was supported by the joint National Institutes of Health (NIH)National Science Foundation (NSF) Ecology of Infectious Diseases Programunder grant no NSFDEB0225453 Opinions findings conclusions andrecommendations are those of the authors and do not necessarily reflectthe views of NIH or NSF KH was supported by NSF grant DEB-0513994the Pew Charitable Trusts award 2000-002558 to Princeton University theDepartment of Ecology and Evolution Princeton University and the HeinzFoundation TL was supported by the Royal (Dick) School of VeterinaryStudies University of Edinburgh and the University of Edinburgh DevelopmentTrust (visits to CDC) and SC by the Wellcome Trust and the Department forInternational Development Animal Health Programme
References
Barrat J (1996) Simple technique for the collection and shipment of brainspecimens for rabies diagnosis Laboratory Techniques in Rabies (edsF-X Meslin MM Kaplan amp H Koprowski) pp 425ndash432 World HealthOrganization Geneva Switzerland
Barrat J Barrat MJ Picard M amp Aubert MFA (1988) Diagnosis of rabiesby cell-culture Comparative Immunology Microbiology and InfectiousDiseases 11 207ndash214
Bartlett MS (1960) Stochastic Population Models in Ecology and EpidemiologyMethuen London
Bernardi F Nadin-Davis SA Wandeler AI Armstrong J Gomes AABLima FS Nogueira FRB amp Ito FH (2005) Antigenic and geneticcharacterization of rabies viruses isolated from domestic and wild animalsof Brazil identifies the hoary fox as a rabies reservoir Journal of GeneralVirology 86 3153ndash3162
Biek R Walsh PD Leroy EM amp Real LA (2006) Recent common ancestryof Ebola Zaire virus found in a bat reservoir Public Library of SciencePathogens 2 885ndash886
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999a) Theepidemiology of rabies in Zimbabwe 2 Rabies in jackals (Canis adustus andCanis mesomelas) Onderstepoort Journal of Veterinary Research 66 11ndash23
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999b) Theepidemiology of rabies in Zimbabwe 1 Rabies in dogs (Canis familiaris)Onderstepoort Journal of Veterinary Research 66 1ndash10
Bingham J Javangwe S Sabeta CT Wandeler AI amp Nel LH (2001)Report of isolations of unusual lyssaviruses (rabies and Mokola virus)identified retrospectively from Zimbabwe Journal of the South AfricanVeterinary Association 72 92ndash94
Brooks R (1990) Survey of the dog-population of Zimbabwe and its level ofrabies vaccination Veterinary Record 127 592ndash596
Cleaveland S amp Dye C (1995) Maintenance of a microparasite infectingseveral host species rabies in the Serengeti Parasitology 111 S33ndashS47
Cleaveland S Kaare M Tiringa P Mlengeya T amp Barrat J (2003) A dograbies vaccination campaign in rural Africa impact on the incidence of dograbies and human dog-bite injuries Vaccine 21 1965ndash1973
Cleaveland S Laurenson MK amp Taylor LH (2001) Diseases of humansand their domestic mammals pathogen characteristics host range andthe risk of emergence Philosophical Transactions of the Royal Society BBiological Sciences 356 991ndash999
Cleaveland S Mlengeya T Kaare M Haydon DT Lembo T LaurensonMK amp Packer C (2007) The conservation relevance of epidemiologicalresearch into carnivore viral diseases in the Serengeti Conservation Biology21 612ndash622
Conzelmann KK Cox JH Schneider LG amp Thiel HJ (1990) Molecularcloning and complete nucleotide sequence of the attenuated rabies virusSAD B19 Virology 175 485ndash499
Daszak P Cunningham AA amp Hyatt AD (2000) Wildlife ecology ndash Emerginginfectious diseases of wildlife ndash Threats to Biodiversity and human healthScience 287 443ndash449
Dean DJ Abelseth MK amp Atanasiu P (1996) The fluorescent antibodytest Laboratory Techniques in Rabies (eds F-X Meslin MM Kaplan
Rabies reservoir dynamics in the Serengeti 11
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
amp H Koprowski) pp 88ndash95 World Health Organization GenevaSwitzerland
Dobson A (1995) The ecology and epidemiology of rinderpest virus inSerengeti and Ngorongoro Conservation Area Serengeti II DynamicsManagement and Conservation of an Ecosystem (eds ARE Sinclair ampP Arcese) pp 485ndash505 University of Chicago Press Chicago
Dobson A (2004) Population dynamics of pathogens with multiple host speciesAmerican Naturalist 164 S64ndashS78
Dobson A amp Foufopoulos J (2001) Emerging infectious pathogens of wildlifePhilosophical Transactions of the Royal Society B Biological Sciences 3561001ndash1012
Dye C (1996) Serengeti wild dogs what really happened Trends in Ecology ampEvolution 11 188ndash189
East ML Hofer H Cox JH Wulle U Wiik H amp Pitra C (2001) Regularexposure to rabies virus and lack of symptomatic disease in Serengetispotted hyenas Proceedings of the National Academy of Sciences USA 9815026ndash15031
Foggin CM (1988) Rabies and Rabies-related Viruses in Zimbabwe HistoricalVirological and Ecological Aspects PhD thesis University of ZimbabweHarare Zimbabwe
Gascoyne SC King AA Laurenson MK Borner M Schildger B ampBarrat J (1993) Aspects of rabies infection and control in the conservationof the African wild dog (Lycaon pictus) in the Serengeti region TanzaniaOnderstepoort Journal of Veterinary Research 60 415ndash420
Gordon ER Curns AT Krebs JW Rupprecht CE Real LA ampChilds JE (2004) Temporal dynamics of rabies in a wildlife host and the riskof cross-species transmission Epidemiology and Infection 132 515ndash524
Guerra MA Curns AT Rupprecht CE Hanlon CA Krebs JW ampChilds JE (2003) Skunk and raccoon rabies in the eastern United Statestemporal and spatial analysis Emerging Infectious Diseases 9 1143ndash1150
Hall TA (1999) bioedit a user-friendly biological sequence alignment editorand analysis program for Windows 9598NT Nucleic Acids SymposiumSeries 41 95ndash98
Haydon DT Cleaveland S Taylor LH amp Laurenson MK (2002) Identi-fying reservoirs of infection a conceptual and practical challenge EmergingInfectious Diseases 8 1468ndash1473
Independent Scientific Group on Cattle TB (2007) Bovine TB The ScientificEvidence Defra Publications London PB12634 httpwwwdefragovukanimalhtbisgindexhtm
Jeanmougin F Thompson JD Gouy M Higgins DG amp Gibson TJ(1998) Multiple sequence alignment with clustal-x Trends in BiochemicalSciences 23 403ndash405
Johnson N Black C Smith J Un H McElhinney LM Aylan O ampFooks AR (2003) Rabies emergence among foxes in Turkey Journal ofWildlife Diseases 39 262ndash270
King AA Meredith CD amp Thomson GR (1993) Canid and viverrid rabiesviruses in South Africa Onderstepoort Journal of Veterinary Research 60295ndash299
Kissi B Tordo N amp Bourhy H (1995) Genetic polymorphism in the rabiesvirus nucleoprotein gene Virology 209 526ndash537
Kitala PM McDermott JJ Kyule MN amp Gathuma JM (2000) Community-based active surveillance for rabies in Machakos District Kenya PreventiveVeterinary Medicine 44 73ndash85
Kitala PM McDermott J Kyule M Gathuma JM Perry B amp Wandeler A(2001) Dog ecology and demography information to support the planning ofrabies control in Machakos District Kenya Acta Tropica 78 217ndash230
Knobel DL Cleaveland S Coleman PG Fevre EM Meltzer MIMiranda MEG Shaw A Zinsstag J amp Meslin F-X (2005) Re-evaluatingthe burden of rabies in Africa and Asia Bulletin of the World HealthOrganization 83 360ndash368
Lembo T Haydon DT Velasco-Villa A Rupprecht CE Packer CBrandatildeo PE Kuzmin IV Fooks AR Barrat J amp Cleaveland S (2007)Molecular epidemiology identifies only a single rabies virus variantcirculating in complex carnivore communities of the Serengeti Proceedingsof the Royal Society B Biological Sciences 274 2123ndash2130
Maas B (1993) Behavioural Ecology and Social Organisation of the Bat-earedFox in the Serengeti National Park Tanzania University of CambridgeCambridge UK
National Bureau of Statistics Tanzania (2005) 2002 Population and HousingCensus Presidentrsquos Office Planning and Privatization Dar es SalaamTanzania
Nel JAJ (1993) The bat-eared fox-a prime candidate for rabies vectorOnderstepoort Journal of Veterinary Research 60 395ndash397
Nel LH Sabeta CT von Teichman B Jaftha JB Rupprecht CE amp
Bingham J (2005) Mongoose rabies in southern Africa a re-evaluationbased on molecular epidemiology Virus Research 109 165ndash173
Page RDM (1996) treeview an application to display phylogenetic trees onpersonal computers Computer Applications in the Biosciences 12 357ndash358
Plowright W (1982) The effects of rinderpest and rinderpest control on wildlifein Africa Symposia of the Zoological Society of London 50 1ndash28
Posada D amp Crandall KA (1998) modeltest testing the model of DNAsubstitution Bioinformatics 14 817ndash818
Randall DA Marino J Haydon DT Sillero-Zubiri C Knobel DLTallents LA Macdonald DW amp Laurenson MK (2006) An integrateddisease management strategy for the control of rabies in Ethiopian wolvesBiological Conservation 131 151ndash162
Randall DA Williams SD Kuzmin IV Rupprecht CE Tallents LATefera Z Argaw K Shiferaw F Knobel DL Sillero-Zubiri C ampLaurenson MK (2004) Rabies in endangered Ethiopian wolves EmergingInfectious Diseases 10 2214ndash2217
Rhodes CJ Atkinson RPD Anderson RM amp Macdonald DW (1998)Rabies in Zimbabwe reservoir dogs and the implications for disease controlPhilosophical Transactions of the Royal Society B Biological Sciences 353999ndash1010
Roelke-Parker ME Munson L Packer C Kock R Cleaveland SCarpenter M Obrien SJ Pospischil A HofmannLehmann R Lutz HMwamengele GLM Mgasa MN Machange GA Summers BA ampAppel MJG (1996) A canine distemper virus epidemic in Serengeti lions(Panthera leo) Nature 379 441ndash445
Ronquist F amp Huelsenbeck JP (2003) mrbayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 19 1572ndash1574
Rupprecht CE Dietzschold B Wunner WH amp Koprowsky H (1991)Antigenic relationships of lyssaviruses The Natural History of Rabies (edsGM Baer) pp 69ndash100 CRC Press Boca Raton
Sabeta CT Bingham J amp Nel LH (2003) Molecular epidemiology of canidrabies in Zimbabwe and South Africa Virus Research 91 203ndash211
Sinclair ARE (1979) The eruption of the ruminants Serengeti Dynamics ofan Ecosystem (eds ARE Sinclair amp M Norton-Griffiths) pp 82ndash103University of Chicago Press Chicago
Sinclair ARE amp Arcese P (1995) Serengeti II Dynamics Management andConservation of an Ecosystem University of Chicago Press Chicago
Smith JS (1989) Rabies virus epitopic variation use in ecological studiesAdvances in Virus Research 36 215ndash253
Smith JS Orciari LA amp Yager PA (1995) Molecular epidemiology ofrabies in the United States Seminars in Virology 6 387ndash400
Swanepoel R Barnard BJH Meredith CD Bishop GC BrucknerGK Foggin CM amp Hubschle OJB (1993) Rabies in Southern AfricaOnderstepoort Journal of Veterinary Research 60 325ndash346
Taylor LH Latham SM amp Woolhouse MEJ (2001) Risk factors forhuman disease emergence Philosophical Transactions of the Royal SocietyB Biological Sciences 356 983ndash989
Tepsumethanon V Wilde H amp Meslin FX (2005) Six criteria for rabiesdiagnosis in living dogs Journal of the Medical Association of Thailand 88419ndash422
Thomson GR amp Meredith CD (1993) Rabies in bat-eared foxes in SouthAfrica Onderstepoort Journal of Veterinary Research 60 399ndash403
Vial F Cleaveland S Rasmussen G amp Haydon DT (2006) Development ofvaccination strategies for the management of rabies in African wild dogsBiological Conservation 131 180ndash192
Wandeler AI Budde A Capt S Kappeler A amp Matter H (1988) Dogecology and dog rabies control Reviews of Infectious Diseases 10 S684ndashS688
Wandeler AI Nadin-Davis SA Tinline RR amp Rupprecht CE (1994)Rabies epidemiology some ecological and evolutionary perspectivesLyssaviruses (eds CE Rupprecht B Dietzschold amp H Koprowski)pp 297ndash324 Springer-Verlag Berlin
Woodroffe R (2001) Assessing the risks of intervention immobilizationradio-collaring and vaccination of African wild dogs Oryx 35 234ndash244
Woodroffe R Donnelly CA Johnston WT Bourne FJ Cheeseman CLClifton-Hadley RS Cox DR Gettinby G Hewinson RG Le FevreAM McInerney JP amp Morrison WI (2005) Spatial association ofMycobacterium bovis infection in cattle and badgers Meles meles Journal ofApplied Ecology 42 852ndash862
World Health Organization (1999) World Survey of Rabies No 34 for the Year1998 Geneva Switzerland
Received 14 June 2007 accepted 23 January 2008Handling Editor Simon Thirgood
12 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Supplementary material
The following supplementary material is available for thisarticle
Appendix S1 Latin names of wildlife species
Appendix S2 Supplementary details of methods
Table S1 Virus samples from the study area compared inphylogenetic analysis
Table S2 Details of all wildlife confirmed and suspect casesin SNP
Table S3 Details of confirmed wildlife cases in Serengeti andNgorongoro districts
Fig S1 Incidence of bites by domestic dogs in Serengeti andNgorongoro districts
This material is available as part of the online article fromhttpwwwblackwell-synergycomdoifull101111j1365-2664200801468x(This link will take you to the article abstract)
Please note Blackwell Publishing is not responsible for the con-tent or functionality of any supplementary materials suppliedby the authors Any queries (other than missing material)should be directed to the corresponding author for the article

4
T Lembo
et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society
Journal of Applied Ecology
data with projected annual growth rates of 2middot6 in SD and 3middot8 inND (National Bureau of Statistics Tanzania 2005) and from humanndashdomestic dog ratios (Wandeler
et al
1988 Brooks 1990) obtainedfrom household questionnaires (from SD in 2003 and LGCA in2004)
Several domestic dog vaccination programmes have beenimplemented in the study area Details of vaccination campaigns arereported in Appendix S2 (Supplementary material) although theirimpact is not analysed here The only vaccination of wildlife (Africanwild dogs) was conducted in 1990 (Gascoyne
et al
1993)
D ISEASE
MONITORING
OPERATIONS
Data presented are the result of disease monitoring operations sincethe late 1980s (Table 2) Passive surveillance was used to detectrabies cases in SNP Sightings of sick and dead carnivores werereported through a network of veterinarians from Tanzania NationalParks (TANAPA) and Tanzania Wildlife Research Institute(TAWIRI) rangers scientists and tour-operators Cases includedanimals that were snared injuredwounded from other or unknowncauses observed with signs of disease or found dead with unknownhistory
Outside SNP passive and active surveillance operations wereemployed Passive surveillance data were available through veterinaryoffice records (government offices TAWIRI TANAPA and NCAAuthority) and district hospitals and medical dispensaries Biteinjuries to humans from suspected rabid dogs were defined based onevaluation by clinicians (from information on clinical signs reportedby bite victims) and when human vaccines were not widely availableon whether PEP had been administered Bite injury incidence wascalculated against human population sizes interpolated from census
data (National Bureau of Statistics Tanzania 2005) Community-based active surveillance measures based on previous studies inrural Kenya (Kitala
et al
2000) and Tanzania (Cleaveland
et al
2003) were implemented in SD using livestock field officersstationed in randomly selected villages to collect information onrabies cases from key informants (village leaders teachers dispensarystaff local healers)
Contact tracing was used to collect high-resolution data on spatialand temporal patterns of disease Visits to incidents involving bitinganimals (to bite victims and owners of biting and bitten animals)were initiated using hospital and medical dispensary records andcase reports from livestock offices and community-based surveillanceIncidents were mapped and witnesses interviewed to make a diagnosis(see below) and obtain case-history information (exposure sourceonset of clinical signs and death resulting exposures) The sameprocedure was followed for all exposures and preceding cases whereidentified so that transmission events between identified individualanimals could be traced and confirmed When multiple incidentsinvolving suspected rabid wildlife were reported on the sameconsecutive days within neighbouring homesteads we assumed asingle animal was involved
Wherever possible brain stem samples were collected from suspectrabies cases and carnivore carcasses whatever the apparent cause ofdeath Sample collection was carried out by research personnelveterinarians and livestock field officers the preferred techniquewas removal via the occipital foramen according to World HealthOrganization recommendations (Barrat 1996) Specimens were frozen(ndash20
deg
C) immediately after collection or placed into phosphate-buffered 50 glycerol solution and preserved either cold (+4
deg
Cndash20
deg
C) or at room temperature (25 plusmn 5
deg
C) when refrigerationfacilities were not promptly available
Fig 2 Map of the Serengeti ecological regionillustrating the distribution of suspectedrabies cases (most were not laboratory-confirmed but diagnosed using epidemio-logical history and clinical criteria)Tables S2 and S3 in Supplementary materialdetail confirmed wildlife cases in SerengetiNational Park (SNP) and Serengeti (SD) andNgorongoro (ND) districts Cases recordedin SNP from January 1995ndashJanuary 2007 aremapped whereas only cases from January2002ndashJanuary 2007 in SD and ND aremapped because GPS data were not availablefor the earlier period Domestic dog cases areshown in black (SD n = 833 ND n = 128)and cases in other carnivores as white circles(SD n = 96 ND n = 27 SNP n = 23)Village boundaries are indicated by greylines LGCA Loliondo Game Control AreaNCA Ngorongoro Conservation Area
Rabies reservoir dynamics in the Serengeti
5
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society
Journal of Applied Ecology
RABIES
D IAGNOSIS
AND
V IRUS
CHARACTERIZATION
Diagnostic tests on brains collected up to 2001 and virus isolationwere carried out at the Agence Franccedilaise de Seacutecuriteacute Sanitaire desAliments (AFSSA) Malzeacuteville France using the fluorescent antibodytest (Dean Abelseth amp Atanasiu 1996) inoculation of murineneuroblastoma cells and mouse inoculation (Barrat
et al
1988)Rabies diagnosis on more recent brain tissues was conducted at theRabies Section of the Centers for Disease Control and Prevention(CDC) Atlanta USA by the fluorescent antibody test (httpwwwcdcgovrabiesdocsstandard_dfa_protocol_rabiespdf) Virussamples were typed at CDC RNA was extracted reverse transcribedamplified by polymerase chain reaction and nucleoprotein genesequences generated as described in Lembo
et al
(2007)When brain tissues were not available for laboratory confirmation
diagnosis of suspected rabies cases was based on history of exposureand clinical evaluation (adapting the six criteria proposed byTepsumethanon Wilde amp Meslin 2005) from information on clinicalsigns reported by villagers livestock field officers researchers andpark veterinarians
1
For owned domestic dogs history of a bite clinical signs(Tepsumethanon
et al
2005) disappearance or death within 10 days
2
For domestic dogs of unknown origin clinical signs (as above) anddisappearance or death
3
For wild carnivores clinical signs (as above) plus tameness lossof fear of humans diurnal activity (for nocturnal species) andunprovoked biting (without eating) of objects and animals
PHYLOGENETIC
ANALYSIS
Nucleotide sequences were edited using
bioedit
7middot0middot0 (Hall 1999)and aligned in
clustal-x
version 1middot83 (Jeanmougin
et al
1998)Phylogenetic relationships among viruses from the study area(Table S1 Supplementary material) and sequence data for RABVsfrom other African countries (Kissi Tordo amp Bourhy 1995 Randall
et al
2004) were estimated using Bayesian Markov chain MonteCarlo (MCMC) methods using
mrbayes
3middot0b4 (Ronquist ampHuelsenbeck 2003) as described in Lembo
et al
(2007) Posteriorprobabilities of 0middot95 or greater were considered significant Graphicoutput was generated using
treeview
version 1middot6middot6 (Page 1996)
STATISTICAL
ANALYSIS
Poisson regression was used to test for a relationship between rabiescases in domestic dogs and in other carnivores Using maximumlikelihood a gamma-distributed spatial infection kernel for domesticdogs k(
d
) was estimated from the distances between source animalsand their known contacts and a probability distribution of thegeneration interval for domestic dogs g(
t
) was estimated fromobserved dates of exposure onset of clinical signs and death (fulldetails in Appendix S2) An algorithm (detailed in Appendix S2)was developed to generate putative epidemic trees by assigningprobabilities to links between possible progenitors and each suspectedcase according to their spatiotemporal proximity and the estimatedspatial infection kernel k(
d
) and generation interval distributiong(
t
) This algorithm assumes all cases are observed and thereforecannot account for unobserved intermediates Cases in all specieswere included in the analysis and all carnivore species were assumedequally likely to transmit rabies Only limited data could be collectedon transmission from wild carnivores therefore progenitors of allspecies were assigned using parameters estimated for domestic dogs
Tab
le 2
Tim
elin
e of
dis
ease
mon
itor
ing
oper
atio
ns in
eac
h zo
ne in
6-m
onth
uni
ts T
AW
IRI
Tan
zani
a W
ildlif
e R
esea
rch
Inst
itut
e T
AN
APA
Tan
zani
a N
atio
nal P
arks
NC
AA
Ngo
rong
oro
Con
serv
atio
nA
rea
Aut
hori
ty
YE
AR
ZO
NE
1987
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
SN
P
TA
NA
PA V
eter
inar
y U
nit
TA
WIR
I V
eter
inar
y P
rogr
amm
e
Vir
al T
rans
mis
sion
D
ynam
ics
Pro
ject
SD
Pas
sive
sur
veill
ance
(do
mes
tic
dog-
bite
inju
ry r
ecor
ds f
rom
hos
pita
ls)
D
omes
tic
dog
vacc
inat
ions
Pas
sive
sur
veill
ance
(V
eter
inar
y re
cord
s
villa
ge r
epor
ts o
ppor
tuni
stic
car
cass
sam
plin
g)
Act
ive
(liv
esto
ck fi
eld-
offic
ers)
Act
ive
(liv
esto
ck fi
eld-
offic
ers)
Con
tact
tra
cing
ND
Pas
sive
(do
mes
tic
dog-
bite
inju
ry r
ecor
ds
from
hos
pita
ls)
Dom
esti
c do
g va
ccin
atio
nsP
assi
ve (
Vet
erin
ary
reco
rds
opp
ortu
nist
ic c
arca
ss
sam
plin
g T
AW
IRI
TA
NA
PA a
nd N
CA
A r
ecor
ds)
Con
tact
tra
cing
A r
esea
rch
proj
ect
inve
stig
atin
g in
fect
ion
dyna
mic
s of
vir
al d
isea
ses
6 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
(k(d) and g(t)) One thousand bootstrapped trees were generatedand the proportional contribution of transmission to each case frompreceding cases was calculated If the date of onset of clinical signswas uncertain in each realization a date was assigned with uniformprobability from the range of uncertainty
From the bootstrap trees the amount of transmission withinand between host types was estimated We pooled all carnivoresexcept domestic dogs because of small sample sizes A null model ofthe expected frequency of within- and between-species transmissionassuming that intra- and interspecific transmission events occurwith the same probability (ie transmission is non-assortative)was estimated as follows the species identity of each case wasrandomly reassigned retaining the original distribution of casesamongst species and the resulting frequency of within- andbetween-species transmission was calculated This procedure wasiterated 1000 times to evaluate the probability that the differencebetween inferred within- and between-species transmissionfrequencies could have been due to chance To investigate whetherthe spatial distribution of hosts influenced the estimated patterns oftransmission we repeated the analysis but randomly reassignedspecies identities within grid cells rather than across the entiredistrict and we repeated the analysis for different sized grids (seeAppendix S2)
Results
RABIES RECOGNIT ION PROBABIL ITY
The sensitivity specificity and positive predictive value ofclinical rabies diagnosis (ie recognition of rabies by villagerslivestock field officers park veterinarians and researchpersonnel) calculated against the gold standard fluorescentantibody test (sensitivity ~99 specificity gt 99 Dean et al1996) are reported in Table 3 As a result of the relatively highrabies recognition probability (gt 74 of animals reported assuspect rabies cases were confirmed positive) and the smallnumber of confirmed cases the analyses presented include allsuspected cases unless otherwise stated
RABIES INCIDENCE IN THE STUDY POPULATIONS
Wildlife monitoring activities in SNP are shown throughoutthe study period in Table 2 and Fig 3a A peak in wild
carnivores reported dead and sick coincided with a caninedistemper outbreak in late 1993 and 1994 (Roelke-Parkeret al 1996) Most rabies cases in wildlife were reported andconfirmed in 1998 and 1999 with only sporadic detection inother years (Fig 3b Table S2 in Supplementary material) Lownumbers of snared animals were reported in 1998 suggestingthat the increase in carnivore carcasses recorded that year(Fig 3a) was not due to increased monitoring No wildlifecases have been confirmed in SNP since 1999 except for aleopard Panthera pardus on the SNPndashNCA boundary inSeptember 2004 when there was an outbreak among adjacentdomestic dog populations (shown in Fig 5b)
Animal-bite injuries in humans from suspected rabiddomestic dogs were reported continuously in high-densitydomestic dog populations to the west of SNP before imple-mentation of dog vaccination programmes whereas in the
Table 3 Sensitivity specificity and positive predictive value of rabiesdiagnosis calculated against the gold standard fluorescent antibodytest Data include only cases for which complete history was available(2002ndash2005)
Clinical diagnosis
Laboratory diagnosis
TotalPositive Negative
Positive 55 19 74Negative 0 33 33Total 55 52 107Sensitivity = 100middot00Specificity = 63middot46 (95 CI 50middot37ndash76middot55)Positive predictive value = 74middot32 (95 CI 64middot37ndash84middot28)
Fig 3 Wildlife disease monitoring in Serengeti National Park(a) carnivore reports and (b) samples retrieved for rabies diagnosisThe peak in 1994 coincided with a canine distemper outbreak inlions (Roelke-Parker et al 1996)
Rabies reservoir dynamics in the Serengeti 7
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
lower-density populations to the east incidence was lowerand there were several periods when no bites were recorded(Fig 4) Gaps in recorded bites by suspected rabid dogs mayalso reflect PEP shortages however when we included alldomestic dog bite injuries there were four periods of at least5 months with no reported bite injuries in ND and no equivalentpattern in SD (Fig S1 Supplementary material)
We traced 1255 suspected rabid animals from January 2002to January 2007 (1044 in SD 211 in ND see Fig 5 for carnivorecases) In both districts cases in wildlife were sporadic andcoincided with outbreaks in domestic dogs (Figs 3b 4 amp 5)The spatial and temporal distribution of suspected cases isshown in Figs 2 and 5
INTER- AND INTRA-SPECIF IC TRANSMISSION
The virus
All of the 57 RABV specimens recovered from a range ofspecies from the study area (including spotted hyaenas) fellwithin the Africa 1b group of canid-associated viruses andrevealed no clustering by host species (Fig 6) The evidentintermingling of viral lineages from domestic dogs andwildlife indicates frequent interspecific transmission Analysesusing statistical parsimony techniques were also consistentwith both within- and between-species transmission events(Lembo et al 2007)
Fig 4 Incidence of bites by suspected rabiddomestic dogs in Serengeti (SD black line)and Ngorongoro (ND grey line) districtsderived from hospital records Incidence isshown only for years before the implementa-tion of domestic dog vaccination programmesin the study populations (July 1987ndashJanuary1997 in SD and January 1994ndashDecember2003 in ND) No data were available for6 months during 1993 in SD
Fig 5 Suspected rabies cases amongcarnivore species in (a) Serengeti (n = 929)and (b) Ngorongoro districts (n = 155)monitored by contact tracing from January2001 to January 2007
8 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Spatiotemporal incidence patterns in hosts
Suspected rabies cases in domestic dogs were significantpredictors of those in other carnivores (pooled cases indomestic cats and wildlife P lt 0middot001 in both districts)Credibility intervals (95) of the expected frequency ofwithin- and between-species transmission events based uponthe null assumption of non-assortative transmission (derivedfrom the bootstrap analysis) are shown in Fig 7 Morewithin-species transmission than expected was estimated inboth districts (domestic dog-to-domestic dog transmissionP = 0middot052 in SD and P = 0middot008 in ND transmission withinother carnivores ie wild carnivores and domestic catsFelis catus P lt 0middot0001 in both districts) Estimated between-species transmission was less than expected (P lt 0middot03 in SD
and P lt 0middot001 in ND) except for transmission from domesticdogs to livestock These patterns were consistent and significantwhen the randomization procedure was carried out on a10 times 10 km grid while on a 5 times 5 km grid the patterns wereevident but no longer significant probably because of reducedstatistical power
The majority of human exposures to suspected rabid animalswere from domestic dogs (Table 4) Similarly domestic dogswere the predominant source of infection for livestock (Fig 7)
Discussion
Demonstrating persistence of infection is a prerequisite foridentifying reservoir(s) Domestic dogs living at high densities(within the SD) are the only species in which rabies has been
Fig 6 Phylogenetic tree of nucleoprotein gene sequences (282 bp 94 deduced amino acids nucleotide positions 1139ndash1420 on the SAD B19genome (Conzelmann et al 1990) for RABV samples from the study area compared with isolates from other areas of Africa The tree isconstructed using Bayesian phylogenetics under the transitional model of nucleotide evolution with a gamma-shaped distribution of rates acrosssites (TIM + Γ Posada amp Crandall 1998 base frequencies = 0middot3253 0middot2134 0middot2360 0middot2253 nucleotide substitution rates = 1middot0000 3middot67230middot4393 0middot4393 8middot1773 1middot0000 Γ = 0middot3390) For samples from the study area the year of collection is indicated within square bracketsPreviously published sequences are designated by strain names (Kissi et al 1995 Randall et al 2004) The tree is rooted with isolate 1500AFSdefined as outgroup representative of the lineage Africa 3 Nodal posterior probabilities gt 0middot95 are shown The scale indicates branch-lengthexpressed as the expected number of substitutions per site Species not definitively identified
Rabies reservoir dynamics in the Serengeti 9
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
recorded continuously throughout the study period whereascases in other species (inside and outside SNP) have beensporadic Although disease monitoring presumably detects alower proportion of cases in wildlife than domestic dogs itseems likely that infection in wildlife populations had it beenpresent continuously would have been detected in more recentyears where the number of samples collected and tested washigher than in previous years when rabies was recorded(Table 2 Fig 3b) These observations provide circumstantialevidence for rabies persistence in domestic dog populationsand lack of persistence in other carnivores Clusters ofwildlife cases that coincided with outbreaks in domestic dogpopulations (Figs 3b 4 amp 5) and explosive but transient(lt 2 months) epidemics previously reported in bat-eared foxes(Maas 1993) are also consistent with chains of transmissionthat are not sustained and contrast with incidence patterns inwild carnivores that are believed to maintain rabies throughindependent cycles elsewhere in Africa For example rabieswas recorded sporadically in bat-eared fox populations inSouth Africa from the 1950s to 1970 then continuouslysuggesting a shift to persistent infection (Thomson amp Meredith1993) and in Zimbabwe outbreaks in jackals although ofteninitiated by domestic dog cases appear to be sustained overseveral years (Bingham et al 1999ab)
Specific demographic attributes (high densities andorturnover) are thought to be required for sustained rabiestransmission A threshold density of gt 5 dogs kmndash2 has beensuggested for persistence of rabies in domestic dog populationsin Africa (Foggin 1988 Brooks 1990 Cleaveland amp Dye 1995Kitala et al 2001) Domestic dog populations in the studyregion have grown considerably and even in the lower-density
area (within the ND) the estimated average density hasapproached this threshold (Table 1) and in 2003 populationsizes were sufficient for a sizable epidemic to occur (Fig 5b)Rabies-control measures make it hard to determine whethercurrent densities in ND are high enough for infection to persistin the absence of vaccination but they may now be close
We detected only one canid-associated variant (Africa 1b)from a range of species in the Serengeti and were unable toreplicate East et alrsquos (2001) finding of a distinct virus variantcirculating atypically in spotted hyaenas (Lembo et al 2007)The lack of species-specific virusndashhost associations in thecanid-associated variant that we detected and the high degreeof genetic relatedness between virus samples from differentspecies indicate frequent cross-species transmission This isconsistent with results from southern Africa where phylogeneticbranches from a single geographic area include several hostspecies (Sabeta Bingham amp Nel 2003)
The statistical association between rabies cases in domesticdogs and other species also indicates cross-species transmissionUnfortunately the length of our time series and paucity ofcases detected in species other than domestic dogs precludeanalyses of lsquospill-overrsquo risk and reduce the interpretativepower of time-lagged regressions like those conducted forrabies in North America that could otherwise provide insight(Guerra et al 2003 Gordon et al 2004) In general as theamount of between-species transmission increases the moredifficult it becomes to tease apart the role of differentpotentially independent maintenance hosts We find greaterwithin-species and less between-species transmission thanwould be expected if species interacted randomly This patterncould occur as a consequence of the spatial distribution ofhost species andor preferential contact between conspecificsOur randomization indicates that large-scale patterns in thedistribution of hosts (gt 10 times 10 km) do not explain thislsquoassortativersquo transmission but we cannot make inference ona smaller scale These analyses suggest that occasionallsquospill-overrsquo of infection from domestic dog populations into
Fig 7 The proportion of estimated transmission within andbetween host types (associated with 1255 suspected rabies cases 1044in Serengeti and 211 in Ngorongoro) in Serengeti (light grey) andNgorongoro (dark grey) districts with lsquodogrsquo corresponding to domesticdogs and lsquocarnivorersquo to domestic cats and wild carnivores Credibilityintervals (95) are shown for expected transmission within andbetween host types assuming inter- and intraspecific transmissionwere equally likely and that livestock are dead-end hosts
Table 4 Human exposures to rabies as a result of bites by suspectedrabid animals in Serengeti and Ngorongoro districts (January 2002ndashDecember 2006) Data were obtained by contact tracing
SpeciesSerengeti district ()
Ngorongorodistrict ()
Domestic dogs 466 (85middot5) 84 (70middot0)Domestic cats 25 (4middot6) 6 (5middot0)Livestock 7 (1middot3) 3 (2middot5)Human 2 (0middot4) 1 (0middot8)Spotted hyaena Crocuta crocuta 4 (0middot7) 16 (13middot3)Honey badger Mellivora capensis 11 (2middot0) 6 (5middot0)African wildcat Felis lybica 0 (0) 2 (1middot7)White-tailed mongoose
Ichneumia albicauda2 (0middot4) 1 (0middot8)
Bat-eared fox Otocyon megalotis 2 (0middot4) 0 (0)Small-spotted genet Genetta genetta 1 (0middot2) 1 (0middot8)Jackal spp Canis spp 23 (4middot2) 0 (0)Leopard Panthera pardus 2 (0middot4) 0 (0)Total 545 (100) 120 (100)
10 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
alternative hosts sometimes results in short-lived chains oftransmission as contact is more common among conspecifics(Dobson 2004)
Definitive identification of infection reservoirs is extremelychallenging and rarely possible Despite intensive effortunderstanding of reservoir dynamics in many complexmulti-host systems remains incomplete (Woodroffe et al2005 Biek et al 2006 Independent Scientific Group on CattleTB 2007) Here the balance of evidence implicates domesticdogs as the only maintenance population of rabies in theSerengeti whereas other carnivores are nonessential tomaintenance However because they transmit disease totarget populations (and frequent transmission within andbetween such species may occur) they do constitute part ofthe reservoir (Table 4 Fig 1a) A single maintenance-hostspecies is consistent with broad patterns of rabies infectionobserved elsewhere (Smith 1989 Rupprecht et al 1991Wandeler et al 1994 Smith Orciari amp Yager 1995)
A lesson from this study is that even with considerable datafrom various sources we cannot exhaustively quantify thecontribution of transmission from all non-essential reservoircomponents However unravelling every thread of reservoirdynamics may not be essential Our key finding that wildlifeare not able to maintain rabies cycles or distinct geneticvariants provides new and critical information allowing rabiescontrol measures in the Serengeti to be targeted moreconfidently at domestic dog populations alone The long-term implication is that control of domestic dog rabies in theSerengeti will eliminate rabies in all other species includinghumans livestock and wildlife The absence of wildlifemaintenance hosts even in an area with such abundantwildlife raises the expectation that canine rabies eliminationis a feasible objective throughout much of Africa
Interventions to prevent transmission from domestic dogmaintenance populations would also provide convincingevidence with which to identify reservoirs Such approacheshave demonstrated that cattle were the reservoir for rinderpestafter cattle vaccinations rinderpest disappeared from wildlifeand the wildebeest Connochaetes taurinus population increasednearly ten-fold (Sinclair 1979 Plowright 1982 Dobson 1995)We are currently ring-vaccinating domestic dogs around SNPwhich should provide critical data to evaluate the impact ofalternative hosts on long-term control efforts in particular theireffects on the level of vaccination required for disease control
In conclusion resolving reservoir dynamics in any complexsystem is likely to prove challenging and our experiences inthe Serengeti suggest that consideration needs to be given toidentifying (i) the potential role of different componentsof the system (ii) the key information needed for differentdisease control strategies (iii) the epidemiological approacheslikely to yield most useful information and (iv) the range ofanalytical tools available
Acknowledgements
We are grateful to Tanzania Government ministries TANAPA TAWIRINCA Authority Tanzania Commission for Science and Technology andNational Institute for Medical Research for permissions and TANAPA and
TAWIRI Veterinary Units Viral Transmission Dynamics Project SerengetiLion and Cheetah Projects Frankfurt Zoological Society livestock field-officersof the Ministry of Water and Livestock Development in Mara and ArushaRegions M Magoto P Tiringa M Roelke-Parker R Fyumagwa H Wiickand Mwanza and Arusha Veterinary Investigation Centres for assistance withsample collection We are indebted to J Barrat AFSSA for performingdiagnostic testing and virus isolation and the staff of the Rabies Section of theCDC especially CE Rupprecht A Velasco-Villa LA Orciari IV Kuzminand M Niezgoda and PE Brandatildeo (Universidade de Satildeo Paulo) for technicalassistance with diagnostic tests and virus characterization We thankKLaurenson D Bennett J Pulliam and H Horn for valuable discussions
This work was supported by the joint National Institutes of Health (NIH)National Science Foundation (NSF) Ecology of Infectious Diseases Programunder grant no NSFDEB0225453 Opinions findings conclusions andrecommendations are those of the authors and do not necessarily reflectthe views of NIH or NSF KH was supported by NSF grant DEB-0513994the Pew Charitable Trusts award 2000-002558 to Princeton University theDepartment of Ecology and Evolution Princeton University and the HeinzFoundation TL was supported by the Royal (Dick) School of VeterinaryStudies University of Edinburgh and the University of Edinburgh DevelopmentTrust (visits to CDC) and SC by the Wellcome Trust and the Department forInternational Development Animal Health Programme
References
Barrat J (1996) Simple technique for the collection and shipment of brainspecimens for rabies diagnosis Laboratory Techniques in Rabies (edsF-X Meslin MM Kaplan amp H Koprowski) pp 425ndash432 World HealthOrganization Geneva Switzerland
Barrat J Barrat MJ Picard M amp Aubert MFA (1988) Diagnosis of rabiesby cell-culture Comparative Immunology Microbiology and InfectiousDiseases 11 207ndash214
Bartlett MS (1960) Stochastic Population Models in Ecology and EpidemiologyMethuen London
Bernardi F Nadin-Davis SA Wandeler AI Armstrong J Gomes AABLima FS Nogueira FRB amp Ito FH (2005) Antigenic and geneticcharacterization of rabies viruses isolated from domestic and wild animalsof Brazil identifies the hoary fox as a rabies reservoir Journal of GeneralVirology 86 3153ndash3162
Biek R Walsh PD Leroy EM amp Real LA (2006) Recent common ancestryof Ebola Zaire virus found in a bat reservoir Public Library of SciencePathogens 2 885ndash886
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999a) Theepidemiology of rabies in Zimbabwe 2 Rabies in jackals (Canis adustus andCanis mesomelas) Onderstepoort Journal of Veterinary Research 66 11ndash23
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999b) Theepidemiology of rabies in Zimbabwe 1 Rabies in dogs (Canis familiaris)Onderstepoort Journal of Veterinary Research 66 1ndash10
Bingham J Javangwe S Sabeta CT Wandeler AI amp Nel LH (2001)Report of isolations of unusual lyssaviruses (rabies and Mokola virus)identified retrospectively from Zimbabwe Journal of the South AfricanVeterinary Association 72 92ndash94
Brooks R (1990) Survey of the dog-population of Zimbabwe and its level ofrabies vaccination Veterinary Record 127 592ndash596
Cleaveland S amp Dye C (1995) Maintenance of a microparasite infectingseveral host species rabies in the Serengeti Parasitology 111 S33ndashS47
Cleaveland S Kaare M Tiringa P Mlengeya T amp Barrat J (2003) A dograbies vaccination campaign in rural Africa impact on the incidence of dograbies and human dog-bite injuries Vaccine 21 1965ndash1973
Cleaveland S Laurenson MK amp Taylor LH (2001) Diseases of humansand their domestic mammals pathogen characteristics host range andthe risk of emergence Philosophical Transactions of the Royal Society BBiological Sciences 356 991ndash999
Cleaveland S Mlengeya T Kaare M Haydon DT Lembo T LaurensonMK amp Packer C (2007) The conservation relevance of epidemiologicalresearch into carnivore viral diseases in the Serengeti Conservation Biology21 612ndash622
Conzelmann KK Cox JH Schneider LG amp Thiel HJ (1990) Molecularcloning and complete nucleotide sequence of the attenuated rabies virusSAD B19 Virology 175 485ndash499
Daszak P Cunningham AA amp Hyatt AD (2000) Wildlife ecology ndash Emerginginfectious diseases of wildlife ndash Threats to Biodiversity and human healthScience 287 443ndash449
Dean DJ Abelseth MK amp Atanasiu P (1996) The fluorescent antibodytest Laboratory Techniques in Rabies (eds F-X Meslin MM Kaplan
Rabies reservoir dynamics in the Serengeti 11
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
amp H Koprowski) pp 88ndash95 World Health Organization GenevaSwitzerland
Dobson A (1995) The ecology and epidemiology of rinderpest virus inSerengeti and Ngorongoro Conservation Area Serengeti II DynamicsManagement and Conservation of an Ecosystem (eds ARE Sinclair ampP Arcese) pp 485ndash505 University of Chicago Press Chicago
Dobson A (2004) Population dynamics of pathogens with multiple host speciesAmerican Naturalist 164 S64ndashS78
Dobson A amp Foufopoulos J (2001) Emerging infectious pathogens of wildlifePhilosophical Transactions of the Royal Society B Biological Sciences 3561001ndash1012
Dye C (1996) Serengeti wild dogs what really happened Trends in Ecology ampEvolution 11 188ndash189
East ML Hofer H Cox JH Wulle U Wiik H amp Pitra C (2001) Regularexposure to rabies virus and lack of symptomatic disease in Serengetispotted hyenas Proceedings of the National Academy of Sciences USA 9815026ndash15031
Foggin CM (1988) Rabies and Rabies-related Viruses in Zimbabwe HistoricalVirological and Ecological Aspects PhD thesis University of ZimbabweHarare Zimbabwe
Gascoyne SC King AA Laurenson MK Borner M Schildger B ampBarrat J (1993) Aspects of rabies infection and control in the conservationof the African wild dog (Lycaon pictus) in the Serengeti region TanzaniaOnderstepoort Journal of Veterinary Research 60 415ndash420
Gordon ER Curns AT Krebs JW Rupprecht CE Real LA ampChilds JE (2004) Temporal dynamics of rabies in a wildlife host and the riskof cross-species transmission Epidemiology and Infection 132 515ndash524
Guerra MA Curns AT Rupprecht CE Hanlon CA Krebs JW ampChilds JE (2003) Skunk and raccoon rabies in the eastern United Statestemporal and spatial analysis Emerging Infectious Diseases 9 1143ndash1150
Hall TA (1999) bioedit a user-friendly biological sequence alignment editorand analysis program for Windows 9598NT Nucleic Acids SymposiumSeries 41 95ndash98
Haydon DT Cleaveland S Taylor LH amp Laurenson MK (2002) Identi-fying reservoirs of infection a conceptual and practical challenge EmergingInfectious Diseases 8 1468ndash1473
Independent Scientific Group on Cattle TB (2007) Bovine TB The ScientificEvidence Defra Publications London PB12634 httpwwwdefragovukanimalhtbisgindexhtm
Jeanmougin F Thompson JD Gouy M Higgins DG amp Gibson TJ(1998) Multiple sequence alignment with clustal-x Trends in BiochemicalSciences 23 403ndash405
Johnson N Black C Smith J Un H McElhinney LM Aylan O ampFooks AR (2003) Rabies emergence among foxes in Turkey Journal ofWildlife Diseases 39 262ndash270
King AA Meredith CD amp Thomson GR (1993) Canid and viverrid rabiesviruses in South Africa Onderstepoort Journal of Veterinary Research 60295ndash299
Kissi B Tordo N amp Bourhy H (1995) Genetic polymorphism in the rabiesvirus nucleoprotein gene Virology 209 526ndash537
Kitala PM McDermott JJ Kyule MN amp Gathuma JM (2000) Community-based active surveillance for rabies in Machakos District Kenya PreventiveVeterinary Medicine 44 73ndash85
Kitala PM McDermott J Kyule M Gathuma JM Perry B amp Wandeler A(2001) Dog ecology and demography information to support the planning ofrabies control in Machakos District Kenya Acta Tropica 78 217ndash230
Knobel DL Cleaveland S Coleman PG Fevre EM Meltzer MIMiranda MEG Shaw A Zinsstag J amp Meslin F-X (2005) Re-evaluatingthe burden of rabies in Africa and Asia Bulletin of the World HealthOrganization 83 360ndash368
Lembo T Haydon DT Velasco-Villa A Rupprecht CE Packer CBrandatildeo PE Kuzmin IV Fooks AR Barrat J amp Cleaveland S (2007)Molecular epidemiology identifies only a single rabies virus variantcirculating in complex carnivore communities of the Serengeti Proceedingsof the Royal Society B Biological Sciences 274 2123ndash2130
Maas B (1993) Behavioural Ecology and Social Organisation of the Bat-earedFox in the Serengeti National Park Tanzania University of CambridgeCambridge UK
National Bureau of Statistics Tanzania (2005) 2002 Population and HousingCensus Presidentrsquos Office Planning and Privatization Dar es SalaamTanzania
Nel JAJ (1993) The bat-eared fox-a prime candidate for rabies vectorOnderstepoort Journal of Veterinary Research 60 395ndash397
Nel LH Sabeta CT von Teichman B Jaftha JB Rupprecht CE amp
Bingham J (2005) Mongoose rabies in southern Africa a re-evaluationbased on molecular epidemiology Virus Research 109 165ndash173
Page RDM (1996) treeview an application to display phylogenetic trees onpersonal computers Computer Applications in the Biosciences 12 357ndash358
Plowright W (1982) The effects of rinderpest and rinderpest control on wildlifein Africa Symposia of the Zoological Society of London 50 1ndash28
Posada D amp Crandall KA (1998) modeltest testing the model of DNAsubstitution Bioinformatics 14 817ndash818
Randall DA Marino J Haydon DT Sillero-Zubiri C Knobel DLTallents LA Macdonald DW amp Laurenson MK (2006) An integrateddisease management strategy for the control of rabies in Ethiopian wolvesBiological Conservation 131 151ndash162
Randall DA Williams SD Kuzmin IV Rupprecht CE Tallents LATefera Z Argaw K Shiferaw F Knobel DL Sillero-Zubiri C ampLaurenson MK (2004) Rabies in endangered Ethiopian wolves EmergingInfectious Diseases 10 2214ndash2217
Rhodes CJ Atkinson RPD Anderson RM amp Macdonald DW (1998)Rabies in Zimbabwe reservoir dogs and the implications for disease controlPhilosophical Transactions of the Royal Society B Biological Sciences 353999ndash1010
Roelke-Parker ME Munson L Packer C Kock R Cleaveland SCarpenter M Obrien SJ Pospischil A HofmannLehmann R Lutz HMwamengele GLM Mgasa MN Machange GA Summers BA ampAppel MJG (1996) A canine distemper virus epidemic in Serengeti lions(Panthera leo) Nature 379 441ndash445
Ronquist F amp Huelsenbeck JP (2003) mrbayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 19 1572ndash1574
Rupprecht CE Dietzschold B Wunner WH amp Koprowsky H (1991)Antigenic relationships of lyssaviruses The Natural History of Rabies (edsGM Baer) pp 69ndash100 CRC Press Boca Raton
Sabeta CT Bingham J amp Nel LH (2003) Molecular epidemiology of canidrabies in Zimbabwe and South Africa Virus Research 91 203ndash211
Sinclair ARE (1979) The eruption of the ruminants Serengeti Dynamics ofan Ecosystem (eds ARE Sinclair amp M Norton-Griffiths) pp 82ndash103University of Chicago Press Chicago
Sinclair ARE amp Arcese P (1995) Serengeti II Dynamics Management andConservation of an Ecosystem University of Chicago Press Chicago
Smith JS (1989) Rabies virus epitopic variation use in ecological studiesAdvances in Virus Research 36 215ndash253
Smith JS Orciari LA amp Yager PA (1995) Molecular epidemiology ofrabies in the United States Seminars in Virology 6 387ndash400
Swanepoel R Barnard BJH Meredith CD Bishop GC BrucknerGK Foggin CM amp Hubschle OJB (1993) Rabies in Southern AfricaOnderstepoort Journal of Veterinary Research 60 325ndash346
Taylor LH Latham SM amp Woolhouse MEJ (2001) Risk factors forhuman disease emergence Philosophical Transactions of the Royal SocietyB Biological Sciences 356 983ndash989
Tepsumethanon V Wilde H amp Meslin FX (2005) Six criteria for rabiesdiagnosis in living dogs Journal of the Medical Association of Thailand 88419ndash422
Thomson GR amp Meredith CD (1993) Rabies in bat-eared foxes in SouthAfrica Onderstepoort Journal of Veterinary Research 60 399ndash403
Vial F Cleaveland S Rasmussen G amp Haydon DT (2006) Development ofvaccination strategies for the management of rabies in African wild dogsBiological Conservation 131 180ndash192
Wandeler AI Budde A Capt S Kappeler A amp Matter H (1988) Dogecology and dog rabies control Reviews of Infectious Diseases 10 S684ndashS688
Wandeler AI Nadin-Davis SA Tinline RR amp Rupprecht CE (1994)Rabies epidemiology some ecological and evolutionary perspectivesLyssaviruses (eds CE Rupprecht B Dietzschold amp H Koprowski)pp 297ndash324 Springer-Verlag Berlin
Woodroffe R (2001) Assessing the risks of intervention immobilizationradio-collaring and vaccination of African wild dogs Oryx 35 234ndash244
Woodroffe R Donnelly CA Johnston WT Bourne FJ Cheeseman CLClifton-Hadley RS Cox DR Gettinby G Hewinson RG Le FevreAM McInerney JP amp Morrison WI (2005) Spatial association ofMycobacterium bovis infection in cattle and badgers Meles meles Journal ofApplied Ecology 42 852ndash862
World Health Organization (1999) World Survey of Rabies No 34 for the Year1998 Geneva Switzerland
Received 14 June 2007 accepted 23 January 2008Handling Editor Simon Thirgood
12 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Supplementary material
The following supplementary material is available for thisarticle
Appendix S1 Latin names of wildlife species
Appendix S2 Supplementary details of methods
Table S1 Virus samples from the study area compared inphylogenetic analysis
Table S2 Details of all wildlife confirmed and suspect casesin SNP
Table S3 Details of confirmed wildlife cases in Serengeti andNgorongoro districts
Fig S1 Incidence of bites by domestic dogs in Serengeti andNgorongoro districts
This material is available as part of the online article fromhttpwwwblackwell-synergycomdoifull101111j1365-2664200801468x(This link will take you to the article abstract)
Please note Blackwell Publishing is not responsible for the con-tent or functionality of any supplementary materials suppliedby the authors Any queries (other than missing material)should be directed to the corresponding author for the article

Rabies reservoir dynamics in the Serengeti
5
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society
Journal of Applied Ecology
RABIES
D IAGNOSIS
AND
V IRUS
CHARACTERIZATION
Diagnostic tests on brains collected up to 2001 and virus isolationwere carried out at the Agence Franccedilaise de Seacutecuriteacute Sanitaire desAliments (AFSSA) Malzeacuteville France using the fluorescent antibodytest (Dean Abelseth amp Atanasiu 1996) inoculation of murineneuroblastoma cells and mouse inoculation (Barrat
et al
1988)Rabies diagnosis on more recent brain tissues was conducted at theRabies Section of the Centers for Disease Control and Prevention(CDC) Atlanta USA by the fluorescent antibody test (httpwwwcdcgovrabiesdocsstandard_dfa_protocol_rabiespdf) Virussamples were typed at CDC RNA was extracted reverse transcribedamplified by polymerase chain reaction and nucleoprotein genesequences generated as described in Lembo
et al
(2007)When brain tissues were not available for laboratory confirmation
diagnosis of suspected rabies cases was based on history of exposureand clinical evaluation (adapting the six criteria proposed byTepsumethanon Wilde amp Meslin 2005) from information on clinicalsigns reported by villagers livestock field officers researchers andpark veterinarians
1
For owned domestic dogs history of a bite clinical signs(Tepsumethanon
et al
2005) disappearance or death within 10 days
2
For domestic dogs of unknown origin clinical signs (as above) anddisappearance or death
3
For wild carnivores clinical signs (as above) plus tameness lossof fear of humans diurnal activity (for nocturnal species) andunprovoked biting (without eating) of objects and animals
PHYLOGENETIC
ANALYSIS
Nucleotide sequences were edited using
bioedit
7middot0middot0 (Hall 1999)and aligned in
clustal-x
version 1middot83 (Jeanmougin
et al
1998)Phylogenetic relationships among viruses from the study area(Table S1 Supplementary material) and sequence data for RABVsfrom other African countries (Kissi Tordo amp Bourhy 1995 Randall
et al
2004) were estimated using Bayesian Markov chain MonteCarlo (MCMC) methods using
mrbayes
3middot0b4 (Ronquist ampHuelsenbeck 2003) as described in Lembo
et al
(2007) Posteriorprobabilities of 0middot95 or greater were considered significant Graphicoutput was generated using
treeview
version 1middot6middot6 (Page 1996)
STATISTICAL
ANALYSIS
Poisson regression was used to test for a relationship between rabiescases in domestic dogs and in other carnivores Using maximumlikelihood a gamma-distributed spatial infection kernel for domesticdogs k(
d
) was estimated from the distances between source animalsand their known contacts and a probability distribution of thegeneration interval for domestic dogs g(
t
) was estimated fromobserved dates of exposure onset of clinical signs and death (fulldetails in Appendix S2) An algorithm (detailed in Appendix S2)was developed to generate putative epidemic trees by assigningprobabilities to links between possible progenitors and each suspectedcase according to their spatiotemporal proximity and the estimatedspatial infection kernel k(
d
) and generation interval distributiong(
t
) This algorithm assumes all cases are observed and thereforecannot account for unobserved intermediates Cases in all specieswere included in the analysis and all carnivore species were assumedequally likely to transmit rabies Only limited data could be collectedon transmission from wild carnivores therefore progenitors of allspecies were assigned using parameters estimated for domestic dogs
Tab
le 2
Tim
elin
e of
dis
ease
mon
itor
ing
oper
atio
ns in
eac
h zo
ne in
6-m
onth
uni
ts T
AW
IRI
Tan
zani
a W
ildlif
e R
esea
rch
Inst
itut
e T
AN
APA
Tan
zani
a N
atio
nal P
arks
NC
AA
Ngo
rong
oro
Con
serv
atio
nA
rea
Aut
hori
ty
YE
AR
ZO
NE
1987
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
SN
P
TA
NA
PA V
eter
inar
y U
nit
TA
WIR
I V
eter
inar
y P
rogr
amm
e
Vir
al T
rans
mis
sion
D
ynam
ics
Pro
ject
SD
Pas
sive
sur
veill
ance
(do
mes
tic
dog-
bite
inju
ry r
ecor
ds f
rom
hos
pita
ls)
D
omes
tic
dog
vacc
inat
ions
Pas
sive
sur
veill
ance
(V
eter
inar
y re
cord
s
villa
ge r
epor
ts o
ppor
tuni
stic
car
cass
sam
plin
g)
Act
ive
(liv
esto
ck fi
eld-
offic
ers)
Act
ive
(liv
esto
ck fi
eld-
offic
ers)
Con
tact
tra
cing
ND
Pas
sive
(do
mes
tic
dog-
bite
inju
ry r
ecor
ds
from
hos
pita
ls)
Dom
esti
c do
g va
ccin
atio
nsP
assi
ve (
Vet
erin
ary
reco
rds
opp
ortu
nist
ic c
arca
ss
sam
plin
g T
AW
IRI
TA
NA
PA a
nd N
CA
A r
ecor
ds)
Con
tact
tra
cing
A r
esea
rch
proj
ect
inve
stig
atin
g in
fect
ion
dyna
mic
s of
vir
al d
isea
ses
6 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
(k(d) and g(t)) One thousand bootstrapped trees were generatedand the proportional contribution of transmission to each case frompreceding cases was calculated If the date of onset of clinical signswas uncertain in each realization a date was assigned with uniformprobability from the range of uncertainty
From the bootstrap trees the amount of transmission withinand between host types was estimated We pooled all carnivoresexcept domestic dogs because of small sample sizes A null model ofthe expected frequency of within- and between-species transmissionassuming that intra- and interspecific transmission events occurwith the same probability (ie transmission is non-assortative)was estimated as follows the species identity of each case wasrandomly reassigned retaining the original distribution of casesamongst species and the resulting frequency of within- andbetween-species transmission was calculated This procedure wasiterated 1000 times to evaluate the probability that the differencebetween inferred within- and between-species transmissionfrequencies could have been due to chance To investigate whetherthe spatial distribution of hosts influenced the estimated patterns oftransmission we repeated the analysis but randomly reassignedspecies identities within grid cells rather than across the entiredistrict and we repeated the analysis for different sized grids (seeAppendix S2)
Results
RABIES RECOGNIT ION PROBABIL ITY
The sensitivity specificity and positive predictive value ofclinical rabies diagnosis (ie recognition of rabies by villagerslivestock field officers park veterinarians and researchpersonnel) calculated against the gold standard fluorescentantibody test (sensitivity ~99 specificity gt 99 Dean et al1996) are reported in Table 3 As a result of the relatively highrabies recognition probability (gt 74 of animals reported assuspect rabies cases were confirmed positive) and the smallnumber of confirmed cases the analyses presented include allsuspected cases unless otherwise stated
RABIES INCIDENCE IN THE STUDY POPULATIONS
Wildlife monitoring activities in SNP are shown throughoutthe study period in Table 2 and Fig 3a A peak in wild
carnivores reported dead and sick coincided with a caninedistemper outbreak in late 1993 and 1994 (Roelke-Parkeret al 1996) Most rabies cases in wildlife were reported andconfirmed in 1998 and 1999 with only sporadic detection inother years (Fig 3b Table S2 in Supplementary material) Lownumbers of snared animals were reported in 1998 suggestingthat the increase in carnivore carcasses recorded that year(Fig 3a) was not due to increased monitoring No wildlifecases have been confirmed in SNP since 1999 except for aleopard Panthera pardus on the SNPndashNCA boundary inSeptember 2004 when there was an outbreak among adjacentdomestic dog populations (shown in Fig 5b)
Animal-bite injuries in humans from suspected rabiddomestic dogs were reported continuously in high-densitydomestic dog populations to the west of SNP before imple-mentation of dog vaccination programmes whereas in the
Table 3 Sensitivity specificity and positive predictive value of rabiesdiagnosis calculated against the gold standard fluorescent antibodytest Data include only cases for which complete history was available(2002ndash2005)
Clinical diagnosis
Laboratory diagnosis
TotalPositive Negative
Positive 55 19 74Negative 0 33 33Total 55 52 107Sensitivity = 100middot00Specificity = 63middot46 (95 CI 50middot37ndash76middot55)Positive predictive value = 74middot32 (95 CI 64middot37ndash84middot28)
Fig 3 Wildlife disease monitoring in Serengeti National Park(a) carnivore reports and (b) samples retrieved for rabies diagnosisThe peak in 1994 coincided with a canine distemper outbreak inlions (Roelke-Parker et al 1996)
Rabies reservoir dynamics in the Serengeti 7
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
lower-density populations to the east incidence was lowerand there were several periods when no bites were recorded(Fig 4) Gaps in recorded bites by suspected rabid dogs mayalso reflect PEP shortages however when we included alldomestic dog bite injuries there were four periods of at least5 months with no reported bite injuries in ND and no equivalentpattern in SD (Fig S1 Supplementary material)
We traced 1255 suspected rabid animals from January 2002to January 2007 (1044 in SD 211 in ND see Fig 5 for carnivorecases) In both districts cases in wildlife were sporadic andcoincided with outbreaks in domestic dogs (Figs 3b 4 amp 5)The spatial and temporal distribution of suspected cases isshown in Figs 2 and 5
INTER- AND INTRA-SPECIF IC TRANSMISSION
The virus
All of the 57 RABV specimens recovered from a range ofspecies from the study area (including spotted hyaenas) fellwithin the Africa 1b group of canid-associated viruses andrevealed no clustering by host species (Fig 6) The evidentintermingling of viral lineages from domestic dogs andwildlife indicates frequent interspecific transmission Analysesusing statistical parsimony techniques were also consistentwith both within- and between-species transmission events(Lembo et al 2007)
Fig 4 Incidence of bites by suspected rabiddomestic dogs in Serengeti (SD black line)and Ngorongoro (ND grey line) districtsderived from hospital records Incidence isshown only for years before the implementa-tion of domestic dog vaccination programmesin the study populations (July 1987ndashJanuary1997 in SD and January 1994ndashDecember2003 in ND) No data were available for6 months during 1993 in SD
Fig 5 Suspected rabies cases amongcarnivore species in (a) Serengeti (n = 929)and (b) Ngorongoro districts (n = 155)monitored by contact tracing from January2001 to January 2007
8 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Spatiotemporal incidence patterns in hosts
Suspected rabies cases in domestic dogs were significantpredictors of those in other carnivores (pooled cases indomestic cats and wildlife P lt 0middot001 in both districts)Credibility intervals (95) of the expected frequency ofwithin- and between-species transmission events based uponthe null assumption of non-assortative transmission (derivedfrom the bootstrap analysis) are shown in Fig 7 Morewithin-species transmission than expected was estimated inboth districts (domestic dog-to-domestic dog transmissionP = 0middot052 in SD and P = 0middot008 in ND transmission withinother carnivores ie wild carnivores and domestic catsFelis catus P lt 0middot0001 in both districts) Estimated between-species transmission was less than expected (P lt 0middot03 in SD
and P lt 0middot001 in ND) except for transmission from domesticdogs to livestock These patterns were consistent and significantwhen the randomization procedure was carried out on a10 times 10 km grid while on a 5 times 5 km grid the patterns wereevident but no longer significant probably because of reducedstatistical power
The majority of human exposures to suspected rabid animalswere from domestic dogs (Table 4) Similarly domestic dogswere the predominant source of infection for livestock (Fig 7)
Discussion
Demonstrating persistence of infection is a prerequisite foridentifying reservoir(s) Domestic dogs living at high densities(within the SD) are the only species in which rabies has been
Fig 6 Phylogenetic tree of nucleoprotein gene sequences (282 bp 94 deduced amino acids nucleotide positions 1139ndash1420 on the SAD B19genome (Conzelmann et al 1990) for RABV samples from the study area compared with isolates from other areas of Africa The tree isconstructed using Bayesian phylogenetics under the transitional model of nucleotide evolution with a gamma-shaped distribution of rates acrosssites (TIM + Γ Posada amp Crandall 1998 base frequencies = 0middot3253 0middot2134 0middot2360 0middot2253 nucleotide substitution rates = 1middot0000 3middot67230middot4393 0middot4393 8middot1773 1middot0000 Γ = 0middot3390) For samples from the study area the year of collection is indicated within square bracketsPreviously published sequences are designated by strain names (Kissi et al 1995 Randall et al 2004) The tree is rooted with isolate 1500AFSdefined as outgroup representative of the lineage Africa 3 Nodal posterior probabilities gt 0middot95 are shown The scale indicates branch-lengthexpressed as the expected number of substitutions per site Species not definitively identified
Rabies reservoir dynamics in the Serengeti 9
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
recorded continuously throughout the study period whereascases in other species (inside and outside SNP) have beensporadic Although disease monitoring presumably detects alower proportion of cases in wildlife than domestic dogs itseems likely that infection in wildlife populations had it beenpresent continuously would have been detected in more recentyears where the number of samples collected and tested washigher than in previous years when rabies was recorded(Table 2 Fig 3b) These observations provide circumstantialevidence for rabies persistence in domestic dog populationsand lack of persistence in other carnivores Clusters ofwildlife cases that coincided with outbreaks in domestic dogpopulations (Figs 3b 4 amp 5) and explosive but transient(lt 2 months) epidemics previously reported in bat-eared foxes(Maas 1993) are also consistent with chains of transmissionthat are not sustained and contrast with incidence patterns inwild carnivores that are believed to maintain rabies throughindependent cycles elsewhere in Africa For example rabieswas recorded sporadically in bat-eared fox populations inSouth Africa from the 1950s to 1970 then continuouslysuggesting a shift to persistent infection (Thomson amp Meredith1993) and in Zimbabwe outbreaks in jackals although ofteninitiated by domestic dog cases appear to be sustained overseveral years (Bingham et al 1999ab)
Specific demographic attributes (high densities andorturnover) are thought to be required for sustained rabiestransmission A threshold density of gt 5 dogs kmndash2 has beensuggested for persistence of rabies in domestic dog populationsin Africa (Foggin 1988 Brooks 1990 Cleaveland amp Dye 1995Kitala et al 2001) Domestic dog populations in the studyregion have grown considerably and even in the lower-density
area (within the ND) the estimated average density hasapproached this threshold (Table 1) and in 2003 populationsizes were sufficient for a sizable epidemic to occur (Fig 5b)Rabies-control measures make it hard to determine whethercurrent densities in ND are high enough for infection to persistin the absence of vaccination but they may now be close
We detected only one canid-associated variant (Africa 1b)from a range of species in the Serengeti and were unable toreplicate East et alrsquos (2001) finding of a distinct virus variantcirculating atypically in spotted hyaenas (Lembo et al 2007)The lack of species-specific virusndashhost associations in thecanid-associated variant that we detected and the high degreeof genetic relatedness between virus samples from differentspecies indicate frequent cross-species transmission This isconsistent with results from southern Africa where phylogeneticbranches from a single geographic area include several hostspecies (Sabeta Bingham amp Nel 2003)
The statistical association between rabies cases in domesticdogs and other species also indicates cross-species transmissionUnfortunately the length of our time series and paucity ofcases detected in species other than domestic dogs precludeanalyses of lsquospill-overrsquo risk and reduce the interpretativepower of time-lagged regressions like those conducted forrabies in North America that could otherwise provide insight(Guerra et al 2003 Gordon et al 2004) In general as theamount of between-species transmission increases the moredifficult it becomes to tease apart the role of differentpotentially independent maintenance hosts We find greaterwithin-species and less between-species transmission thanwould be expected if species interacted randomly This patterncould occur as a consequence of the spatial distribution ofhost species andor preferential contact between conspecificsOur randomization indicates that large-scale patterns in thedistribution of hosts (gt 10 times 10 km) do not explain thislsquoassortativersquo transmission but we cannot make inference ona smaller scale These analyses suggest that occasionallsquospill-overrsquo of infection from domestic dog populations into
Fig 7 The proportion of estimated transmission within andbetween host types (associated with 1255 suspected rabies cases 1044in Serengeti and 211 in Ngorongoro) in Serengeti (light grey) andNgorongoro (dark grey) districts with lsquodogrsquo corresponding to domesticdogs and lsquocarnivorersquo to domestic cats and wild carnivores Credibilityintervals (95) are shown for expected transmission within andbetween host types assuming inter- and intraspecific transmissionwere equally likely and that livestock are dead-end hosts
Table 4 Human exposures to rabies as a result of bites by suspectedrabid animals in Serengeti and Ngorongoro districts (January 2002ndashDecember 2006) Data were obtained by contact tracing
SpeciesSerengeti district ()
Ngorongorodistrict ()
Domestic dogs 466 (85middot5) 84 (70middot0)Domestic cats 25 (4middot6) 6 (5middot0)Livestock 7 (1middot3) 3 (2middot5)Human 2 (0middot4) 1 (0middot8)Spotted hyaena Crocuta crocuta 4 (0middot7) 16 (13middot3)Honey badger Mellivora capensis 11 (2middot0) 6 (5middot0)African wildcat Felis lybica 0 (0) 2 (1middot7)White-tailed mongoose
Ichneumia albicauda2 (0middot4) 1 (0middot8)
Bat-eared fox Otocyon megalotis 2 (0middot4) 0 (0)Small-spotted genet Genetta genetta 1 (0middot2) 1 (0middot8)Jackal spp Canis spp 23 (4middot2) 0 (0)Leopard Panthera pardus 2 (0middot4) 0 (0)Total 545 (100) 120 (100)
10 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
alternative hosts sometimes results in short-lived chains oftransmission as contact is more common among conspecifics(Dobson 2004)
Definitive identification of infection reservoirs is extremelychallenging and rarely possible Despite intensive effortunderstanding of reservoir dynamics in many complexmulti-host systems remains incomplete (Woodroffe et al2005 Biek et al 2006 Independent Scientific Group on CattleTB 2007) Here the balance of evidence implicates domesticdogs as the only maintenance population of rabies in theSerengeti whereas other carnivores are nonessential tomaintenance However because they transmit disease totarget populations (and frequent transmission within andbetween such species may occur) they do constitute part ofthe reservoir (Table 4 Fig 1a) A single maintenance-hostspecies is consistent with broad patterns of rabies infectionobserved elsewhere (Smith 1989 Rupprecht et al 1991Wandeler et al 1994 Smith Orciari amp Yager 1995)
A lesson from this study is that even with considerable datafrom various sources we cannot exhaustively quantify thecontribution of transmission from all non-essential reservoircomponents However unravelling every thread of reservoirdynamics may not be essential Our key finding that wildlifeare not able to maintain rabies cycles or distinct geneticvariants provides new and critical information allowing rabiescontrol measures in the Serengeti to be targeted moreconfidently at domestic dog populations alone The long-term implication is that control of domestic dog rabies in theSerengeti will eliminate rabies in all other species includinghumans livestock and wildlife The absence of wildlifemaintenance hosts even in an area with such abundantwildlife raises the expectation that canine rabies eliminationis a feasible objective throughout much of Africa
Interventions to prevent transmission from domestic dogmaintenance populations would also provide convincingevidence with which to identify reservoirs Such approacheshave demonstrated that cattle were the reservoir for rinderpestafter cattle vaccinations rinderpest disappeared from wildlifeand the wildebeest Connochaetes taurinus population increasednearly ten-fold (Sinclair 1979 Plowright 1982 Dobson 1995)We are currently ring-vaccinating domestic dogs around SNPwhich should provide critical data to evaluate the impact ofalternative hosts on long-term control efforts in particular theireffects on the level of vaccination required for disease control
In conclusion resolving reservoir dynamics in any complexsystem is likely to prove challenging and our experiences inthe Serengeti suggest that consideration needs to be given toidentifying (i) the potential role of different componentsof the system (ii) the key information needed for differentdisease control strategies (iii) the epidemiological approacheslikely to yield most useful information and (iv) the range ofanalytical tools available
Acknowledgements
We are grateful to Tanzania Government ministries TANAPA TAWIRINCA Authority Tanzania Commission for Science and Technology andNational Institute for Medical Research for permissions and TANAPA and
TAWIRI Veterinary Units Viral Transmission Dynamics Project SerengetiLion and Cheetah Projects Frankfurt Zoological Society livestock field-officersof the Ministry of Water and Livestock Development in Mara and ArushaRegions M Magoto P Tiringa M Roelke-Parker R Fyumagwa H Wiickand Mwanza and Arusha Veterinary Investigation Centres for assistance withsample collection We are indebted to J Barrat AFSSA for performingdiagnostic testing and virus isolation and the staff of the Rabies Section of theCDC especially CE Rupprecht A Velasco-Villa LA Orciari IV Kuzminand M Niezgoda and PE Brandatildeo (Universidade de Satildeo Paulo) for technicalassistance with diagnostic tests and virus characterization We thankKLaurenson D Bennett J Pulliam and H Horn for valuable discussions
This work was supported by the joint National Institutes of Health (NIH)National Science Foundation (NSF) Ecology of Infectious Diseases Programunder grant no NSFDEB0225453 Opinions findings conclusions andrecommendations are those of the authors and do not necessarily reflectthe views of NIH or NSF KH was supported by NSF grant DEB-0513994the Pew Charitable Trusts award 2000-002558 to Princeton University theDepartment of Ecology and Evolution Princeton University and the HeinzFoundation TL was supported by the Royal (Dick) School of VeterinaryStudies University of Edinburgh and the University of Edinburgh DevelopmentTrust (visits to CDC) and SC by the Wellcome Trust and the Department forInternational Development Animal Health Programme
References
Barrat J (1996) Simple technique for the collection and shipment of brainspecimens for rabies diagnosis Laboratory Techniques in Rabies (edsF-X Meslin MM Kaplan amp H Koprowski) pp 425ndash432 World HealthOrganization Geneva Switzerland
Barrat J Barrat MJ Picard M amp Aubert MFA (1988) Diagnosis of rabiesby cell-culture Comparative Immunology Microbiology and InfectiousDiseases 11 207ndash214
Bartlett MS (1960) Stochastic Population Models in Ecology and EpidemiologyMethuen London
Bernardi F Nadin-Davis SA Wandeler AI Armstrong J Gomes AABLima FS Nogueira FRB amp Ito FH (2005) Antigenic and geneticcharacterization of rabies viruses isolated from domestic and wild animalsof Brazil identifies the hoary fox as a rabies reservoir Journal of GeneralVirology 86 3153ndash3162
Biek R Walsh PD Leroy EM amp Real LA (2006) Recent common ancestryof Ebola Zaire virus found in a bat reservoir Public Library of SciencePathogens 2 885ndash886
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999a) Theepidemiology of rabies in Zimbabwe 2 Rabies in jackals (Canis adustus andCanis mesomelas) Onderstepoort Journal of Veterinary Research 66 11ndash23
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999b) Theepidemiology of rabies in Zimbabwe 1 Rabies in dogs (Canis familiaris)Onderstepoort Journal of Veterinary Research 66 1ndash10
Bingham J Javangwe S Sabeta CT Wandeler AI amp Nel LH (2001)Report of isolations of unusual lyssaviruses (rabies and Mokola virus)identified retrospectively from Zimbabwe Journal of the South AfricanVeterinary Association 72 92ndash94
Brooks R (1990) Survey of the dog-population of Zimbabwe and its level ofrabies vaccination Veterinary Record 127 592ndash596
Cleaveland S amp Dye C (1995) Maintenance of a microparasite infectingseveral host species rabies in the Serengeti Parasitology 111 S33ndashS47
Cleaveland S Kaare M Tiringa P Mlengeya T amp Barrat J (2003) A dograbies vaccination campaign in rural Africa impact on the incidence of dograbies and human dog-bite injuries Vaccine 21 1965ndash1973
Cleaveland S Laurenson MK amp Taylor LH (2001) Diseases of humansand their domestic mammals pathogen characteristics host range andthe risk of emergence Philosophical Transactions of the Royal Society BBiological Sciences 356 991ndash999
Cleaveland S Mlengeya T Kaare M Haydon DT Lembo T LaurensonMK amp Packer C (2007) The conservation relevance of epidemiologicalresearch into carnivore viral diseases in the Serengeti Conservation Biology21 612ndash622
Conzelmann KK Cox JH Schneider LG amp Thiel HJ (1990) Molecularcloning and complete nucleotide sequence of the attenuated rabies virusSAD B19 Virology 175 485ndash499
Daszak P Cunningham AA amp Hyatt AD (2000) Wildlife ecology ndash Emerginginfectious diseases of wildlife ndash Threats to Biodiversity and human healthScience 287 443ndash449
Dean DJ Abelseth MK amp Atanasiu P (1996) The fluorescent antibodytest Laboratory Techniques in Rabies (eds F-X Meslin MM Kaplan
Rabies reservoir dynamics in the Serengeti 11
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
amp H Koprowski) pp 88ndash95 World Health Organization GenevaSwitzerland
Dobson A (1995) The ecology and epidemiology of rinderpest virus inSerengeti and Ngorongoro Conservation Area Serengeti II DynamicsManagement and Conservation of an Ecosystem (eds ARE Sinclair ampP Arcese) pp 485ndash505 University of Chicago Press Chicago
Dobson A (2004) Population dynamics of pathogens with multiple host speciesAmerican Naturalist 164 S64ndashS78
Dobson A amp Foufopoulos J (2001) Emerging infectious pathogens of wildlifePhilosophical Transactions of the Royal Society B Biological Sciences 3561001ndash1012
Dye C (1996) Serengeti wild dogs what really happened Trends in Ecology ampEvolution 11 188ndash189
East ML Hofer H Cox JH Wulle U Wiik H amp Pitra C (2001) Regularexposure to rabies virus and lack of symptomatic disease in Serengetispotted hyenas Proceedings of the National Academy of Sciences USA 9815026ndash15031
Foggin CM (1988) Rabies and Rabies-related Viruses in Zimbabwe HistoricalVirological and Ecological Aspects PhD thesis University of ZimbabweHarare Zimbabwe
Gascoyne SC King AA Laurenson MK Borner M Schildger B ampBarrat J (1993) Aspects of rabies infection and control in the conservationof the African wild dog (Lycaon pictus) in the Serengeti region TanzaniaOnderstepoort Journal of Veterinary Research 60 415ndash420
Gordon ER Curns AT Krebs JW Rupprecht CE Real LA ampChilds JE (2004) Temporal dynamics of rabies in a wildlife host and the riskof cross-species transmission Epidemiology and Infection 132 515ndash524
Guerra MA Curns AT Rupprecht CE Hanlon CA Krebs JW ampChilds JE (2003) Skunk and raccoon rabies in the eastern United Statestemporal and spatial analysis Emerging Infectious Diseases 9 1143ndash1150
Hall TA (1999) bioedit a user-friendly biological sequence alignment editorand analysis program for Windows 9598NT Nucleic Acids SymposiumSeries 41 95ndash98
Haydon DT Cleaveland S Taylor LH amp Laurenson MK (2002) Identi-fying reservoirs of infection a conceptual and practical challenge EmergingInfectious Diseases 8 1468ndash1473
Independent Scientific Group on Cattle TB (2007) Bovine TB The ScientificEvidence Defra Publications London PB12634 httpwwwdefragovukanimalhtbisgindexhtm
Jeanmougin F Thompson JD Gouy M Higgins DG amp Gibson TJ(1998) Multiple sequence alignment with clustal-x Trends in BiochemicalSciences 23 403ndash405
Johnson N Black C Smith J Un H McElhinney LM Aylan O ampFooks AR (2003) Rabies emergence among foxes in Turkey Journal ofWildlife Diseases 39 262ndash270
King AA Meredith CD amp Thomson GR (1993) Canid and viverrid rabiesviruses in South Africa Onderstepoort Journal of Veterinary Research 60295ndash299
Kissi B Tordo N amp Bourhy H (1995) Genetic polymorphism in the rabiesvirus nucleoprotein gene Virology 209 526ndash537
Kitala PM McDermott JJ Kyule MN amp Gathuma JM (2000) Community-based active surveillance for rabies in Machakos District Kenya PreventiveVeterinary Medicine 44 73ndash85
Kitala PM McDermott J Kyule M Gathuma JM Perry B amp Wandeler A(2001) Dog ecology and demography information to support the planning ofrabies control in Machakos District Kenya Acta Tropica 78 217ndash230
Knobel DL Cleaveland S Coleman PG Fevre EM Meltzer MIMiranda MEG Shaw A Zinsstag J amp Meslin F-X (2005) Re-evaluatingthe burden of rabies in Africa and Asia Bulletin of the World HealthOrganization 83 360ndash368
Lembo T Haydon DT Velasco-Villa A Rupprecht CE Packer CBrandatildeo PE Kuzmin IV Fooks AR Barrat J amp Cleaveland S (2007)Molecular epidemiology identifies only a single rabies virus variantcirculating in complex carnivore communities of the Serengeti Proceedingsof the Royal Society B Biological Sciences 274 2123ndash2130
Maas B (1993) Behavioural Ecology and Social Organisation of the Bat-earedFox in the Serengeti National Park Tanzania University of CambridgeCambridge UK
National Bureau of Statistics Tanzania (2005) 2002 Population and HousingCensus Presidentrsquos Office Planning and Privatization Dar es SalaamTanzania
Nel JAJ (1993) The bat-eared fox-a prime candidate for rabies vectorOnderstepoort Journal of Veterinary Research 60 395ndash397
Nel LH Sabeta CT von Teichman B Jaftha JB Rupprecht CE amp
Bingham J (2005) Mongoose rabies in southern Africa a re-evaluationbased on molecular epidemiology Virus Research 109 165ndash173
Page RDM (1996) treeview an application to display phylogenetic trees onpersonal computers Computer Applications in the Biosciences 12 357ndash358
Plowright W (1982) The effects of rinderpest and rinderpest control on wildlifein Africa Symposia of the Zoological Society of London 50 1ndash28
Posada D amp Crandall KA (1998) modeltest testing the model of DNAsubstitution Bioinformatics 14 817ndash818
Randall DA Marino J Haydon DT Sillero-Zubiri C Knobel DLTallents LA Macdonald DW amp Laurenson MK (2006) An integrateddisease management strategy for the control of rabies in Ethiopian wolvesBiological Conservation 131 151ndash162
Randall DA Williams SD Kuzmin IV Rupprecht CE Tallents LATefera Z Argaw K Shiferaw F Knobel DL Sillero-Zubiri C ampLaurenson MK (2004) Rabies in endangered Ethiopian wolves EmergingInfectious Diseases 10 2214ndash2217
Rhodes CJ Atkinson RPD Anderson RM amp Macdonald DW (1998)Rabies in Zimbabwe reservoir dogs and the implications for disease controlPhilosophical Transactions of the Royal Society B Biological Sciences 353999ndash1010
Roelke-Parker ME Munson L Packer C Kock R Cleaveland SCarpenter M Obrien SJ Pospischil A HofmannLehmann R Lutz HMwamengele GLM Mgasa MN Machange GA Summers BA ampAppel MJG (1996) A canine distemper virus epidemic in Serengeti lions(Panthera leo) Nature 379 441ndash445
Ronquist F amp Huelsenbeck JP (2003) mrbayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 19 1572ndash1574
Rupprecht CE Dietzschold B Wunner WH amp Koprowsky H (1991)Antigenic relationships of lyssaviruses The Natural History of Rabies (edsGM Baer) pp 69ndash100 CRC Press Boca Raton
Sabeta CT Bingham J amp Nel LH (2003) Molecular epidemiology of canidrabies in Zimbabwe and South Africa Virus Research 91 203ndash211
Sinclair ARE (1979) The eruption of the ruminants Serengeti Dynamics ofan Ecosystem (eds ARE Sinclair amp M Norton-Griffiths) pp 82ndash103University of Chicago Press Chicago
Sinclair ARE amp Arcese P (1995) Serengeti II Dynamics Management andConservation of an Ecosystem University of Chicago Press Chicago
Smith JS (1989) Rabies virus epitopic variation use in ecological studiesAdvances in Virus Research 36 215ndash253
Smith JS Orciari LA amp Yager PA (1995) Molecular epidemiology ofrabies in the United States Seminars in Virology 6 387ndash400
Swanepoel R Barnard BJH Meredith CD Bishop GC BrucknerGK Foggin CM amp Hubschle OJB (1993) Rabies in Southern AfricaOnderstepoort Journal of Veterinary Research 60 325ndash346
Taylor LH Latham SM amp Woolhouse MEJ (2001) Risk factors forhuman disease emergence Philosophical Transactions of the Royal SocietyB Biological Sciences 356 983ndash989
Tepsumethanon V Wilde H amp Meslin FX (2005) Six criteria for rabiesdiagnosis in living dogs Journal of the Medical Association of Thailand 88419ndash422
Thomson GR amp Meredith CD (1993) Rabies in bat-eared foxes in SouthAfrica Onderstepoort Journal of Veterinary Research 60 399ndash403
Vial F Cleaveland S Rasmussen G amp Haydon DT (2006) Development ofvaccination strategies for the management of rabies in African wild dogsBiological Conservation 131 180ndash192
Wandeler AI Budde A Capt S Kappeler A amp Matter H (1988) Dogecology and dog rabies control Reviews of Infectious Diseases 10 S684ndashS688
Wandeler AI Nadin-Davis SA Tinline RR amp Rupprecht CE (1994)Rabies epidemiology some ecological and evolutionary perspectivesLyssaviruses (eds CE Rupprecht B Dietzschold amp H Koprowski)pp 297ndash324 Springer-Verlag Berlin
Woodroffe R (2001) Assessing the risks of intervention immobilizationradio-collaring and vaccination of African wild dogs Oryx 35 234ndash244
Woodroffe R Donnelly CA Johnston WT Bourne FJ Cheeseman CLClifton-Hadley RS Cox DR Gettinby G Hewinson RG Le FevreAM McInerney JP amp Morrison WI (2005) Spatial association ofMycobacterium bovis infection in cattle and badgers Meles meles Journal ofApplied Ecology 42 852ndash862
World Health Organization (1999) World Survey of Rabies No 34 for the Year1998 Geneva Switzerland
Received 14 June 2007 accepted 23 January 2008Handling Editor Simon Thirgood
12 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Supplementary material
The following supplementary material is available for thisarticle
Appendix S1 Latin names of wildlife species
Appendix S2 Supplementary details of methods
Table S1 Virus samples from the study area compared inphylogenetic analysis
Table S2 Details of all wildlife confirmed and suspect casesin SNP
Table S3 Details of confirmed wildlife cases in Serengeti andNgorongoro districts
Fig S1 Incidence of bites by domestic dogs in Serengeti andNgorongoro districts
This material is available as part of the online article fromhttpwwwblackwell-synergycomdoifull101111j1365-2664200801468x(This link will take you to the article abstract)
Please note Blackwell Publishing is not responsible for the con-tent or functionality of any supplementary materials suppliedby the authors Any queries (other than missing material)should be directed to the corresponding author for the article

6 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
(k(d) and g(t)) One thousand bootstrapped trees were generatedand the proportional contribution of transmission to each case frompreceding cases was calculated If the date of onset of clinical signswas uncertain in each realization a date was assigned with uniformprobability from the range of uncertainty
From the bootstrap trees the amount of transmission withinand between host types was estimated We pooled all carnivoresexcept domestic dogs because of small sample sizes A null model ofthe expected frequency of within- and between-species transmissionassuming that intra- and interspecific transmission events occurwith the same probability (ie transmission is non-assortative)was estimated as follows the species identity of each case wasrandomly reassigned retaining the original distribution of casesamongst species and the resulting frequency of within- andbetween-species transmission was calculated This procedure wasiterated 1000 times to evaluate the probability that the differencebetween inferred within- and between-species transmissionfrequencies could have been due to chance To investigate whetherthe spatial distribution of hosts influenced the estimated patterns oftransmission we repeated the analysis but randomly reassignedspecies identities within grid cells rather than across the entiredistrict and we repeated the analysis for different sized grids (seeAppendix S2)
Results
RABIES RECOGNIT ION PROBABIL ITY
The sensitivity specificity and positive predictive value ofclinical rabies diagnosis (ie recognition of rabies by villagerslivestock field officers park veterinarians and researchpersonnel) calculated against the gold standard fluorescentantibody test (sensitivity ~99 specificity gt 99 Dean et al1996) are reported in Table 3 As a result of the relatively highrabies recognition probability (gt 74 of animals reported assuspect rabies cases were confirmed positive) and the smallnumber of confirmed cases the analyses presented include allsuspected cases unless otherwise stated
RABIES INCIDENCE IN THE STUDY POPULATIONS
Wildlife monitoring activities in SNP are shown throughoutthe study period in Table 2 and Fig 3a A peak in wild
carnivores reported dead and sick coincided with a caninedistemper outbreak in late 1993 and 1994 (Roelke-Parkeret al 1996) Most rabies cases in wildlife were reported andconfirmed in 1998 and 1999 with only sporadic detection inother years (Fig 3b Table S2 in Supplementary material) Lownumbers of snared animals were reported in 1998 suggestingthat the increase in carnivore carcasses recorded that year(Fig 3a) was not due to increased monitoring No wildlifecases have been confirmed in SNP since 1999 except for aleopard Panthera pardus on the SNPndashNCA boundary inSeptember 2004 when there was an outbreak among adjacentdomestic dog populations (shown in Fig 5b)
Animal-bite injuries in humans from suspected rabiddomestic dogs were reported continuously in high-densitydomestic dog populations to the west of SNP before imple-mentation of dog vaccination programmes whereas in the
Table 3 Sensitivity specificity and positive predictive value of rabiesdiagnosis calculated against the gold standard fluorescent antibodytest Data include only cases for which complete history was available(2002ndash2005)
Clinical diagnosis
Laboratory diagnosis
TotalPositive Negative
Positive 55 19 74Negative 0 33 33Total 55 52 107Sensitivity = 100middot00Specificity = 63middot46 (95 CI 50middot37ndash76middot55)Positive predictive value = 74middot32 (95 CI 64middot37ndash84middot28)
Fig 3 Wildlife disease monitoring in Serengeti National Park(a) carnivore reports and (b) samples retrieved for rabies diagnosisThe peak in 1994 coincided with a canine distemper outbreak inlions (Roelke-Parker et al 1996)
Rabies reservoir dynamics in the Serengeti 7
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
lower-density populations to the east incidence was lowerand there were several periods when no bites were recorded(Fig 4) Gaps in recorded bites by suspected rabid dogs mayalso reflect PEP shortages however when we included alldomestic dog bite injuries there were four periods of at least5 months with no reported bite injuries in ND and no equivalentpattern in SD (Fig S1 Supplementary material)
We traced 1255 suspected rabid animals from January 2002to January 2007 (1044 in SD 211 in ND see Fig 5 for carnivorecases) In both districts cases in wildlife were sporadic andcoincided with outbreaks in domestic dogs (Figs 3b 4 amp 5)The spatial and temporal distribution of suspected cases isshown in Figs 2 and 5
INTER- AND INTRA-SPECIF IC TRANSMISSION
The virus
All of the 57 RABV specimens recovered from a range ofspecies from the study area (including spotted hyaenas) fellwithin the Africa 1b group of canid-associated viruses andrevealed no clustering by host species (Fig 6) The evidentintermingling of viral lineages from domestic dogs andwildlife indicates frequent interspecific transmission Analysesusing statistical parsimony techniques were also consistentwith both within- and between-species transmission events(Lembo et al 2007)
Fig 4 Incidence of bites by suspected rabiddomestic dogs in Serengeti (SD black line)and Ngorongoro (ND grey line) districtsderived from hospital records Incidence isshown only for years before the implementa-tion of domestic dog vaccination programmesin the study populations (July 1987ndashJanuary1997 in SD and January 1994ndashDecember2003 in ND) No data were available for6 months during 1993 in SD
Fig 5 Suspected rabies cases amongcarnivore species in (a) Serengeti (n = 929)and (b) Ngorongoro districts (n = 155)monitored by contact tracing from January2001 to January 2007
8 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Spatiotemporal incidence patterns in hosts
Suspected rabies cases in domestic dogs were significantpredictors of those in other carnivores (pooled cases indomestic cats and wildlife P lt 0middot001 in both districts)Credibility intervals (95) of the expected frequency ofwithin- and between-species transmission events based uponthe null assumption of non-assortative transmission (derivedfrom the bootstrap analysis) are shown in Fig 7 Morewithin-species transmission than expected was estimated inboth districts (domestic dog-to-domestic dog transmissionP = 0middot052 in SD and P = 0middot008 in ND transmission withinother carnivores ie wild carnivores and domestic catsFelis catus P lt 0middot0001 in both districts) Estimated between-species transmission was less than expected (P lt 0middot03 in SD
and P lt 0middot001 in ND) except for transmission from domesticdogs to livestock These patterns were consistent and significantwhen the randomization procedure was carried out on a10 times 10 km grid while on a 5 times 5 km grid the patterns wereevident but no longer significant probably because of reducedstatistical power
The majority of human exposures to suspected rabid animalswere from domestic dogs (Table 4) Similarly domestic dogswere the predominant source of infection for livestock (Fig 7)
Discussion
Demonstrating persistence of infection is a prerequisite foridentifying reservoir(s) Domestic dogs living at high densities(within the SD) are the only species in which rabies has been
Fig 6 Phylogenetic tree of nucleoprotein gene sequences (282 bp 94 deduced amino acids nucleotide positions 1139ndash1420 on the SAD B19genome (Conzelmann et al 1990) for RABV samples from the study area compared with isolates from other areas of Africa The tree isconstructed using Bayesian phylogenetics under the transitional model of nucleotide evolution with a gamma-shaped distribution of rates acrosssites (TIM + Γ Posada amp Crandall 1998 base frequencies = 0middot3253 0middot2134 0middot2360 0middot2253 nucleotide substitution rates = 1middot0000 3middot67230middot4393 0middot4393 8middot1773 1middot0000 Γ = 0middot3390) For samples from the study area the year of collection is indicated within square bracketsPreviously published sequences are designated by strain names (Kissi et al 1995 Randall et al 2004) The tree is rooted with isolate 1500AFSdefined as outgroup representative of the lineage Africa 3 Nodal posterior probabilities gt 0middot95 are shown The scale indicates branch-lengthexpressed as the expected number of substitutions per site Species not definitively identified
Rabies reservoir dynamics in the Serengeti 9
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
recorded continuously throughout the study period whereascases in other species (inside and outside SNP) have beensporadic Although disease monitoring presumably detects alower proportion of cases in wildlife than domestic dogs itseems likely that infection in wildlife populations had it beenpresent continuously would have been detected in more recentyears where the number of samples collected and tested washigher than in previous years when rabies was recorded(Table 2 Fig 3b) These observations provide circumstantialevidence for rabies persistence in domestic dog populationsand lack of persistence in other carnivores Clusters ofwildlife cases that coincided with outbreaks in domestic dogpopulations (Figs 3b 4 amp 5) and explosive but transient(lt 2 months) epidemics previously reported in bat-eared foxes(Maas 1993) are also consistent with chains of transmissionthat are not sustained and contrast with incidence patterns inwild carnivores that are believed to maintain rabies throughindependent cycles elsewhere in Africa For example rabieswas recorded sporadically in bat-eared fox populations inSouth Africa from the 1950s to 1970 then continuouslysuggesting a shift to persistent infection (Thomson amp Meredith1993) and in Zimbabwe outbreaks in jackals although ofteninitiated by domestic dog cases appear to be sustained overseveral years (Bingham et al 1999ab)
Specific demographic attributes (high densities andorturnover) are thought to be required for sustained rabiestransmission A threshold density of gt 5 dogs kmndash2 has beensuggested for persistence of rabies in domestic dog populationsin Africa (Foggin 1988 Brooks 1990 Cleaveland amp Dye 1995Kitala et al 2001) Domestic dog populations in the studyregion have grown considerably and even in the lower-density
area (within the ND) the estimated average density hasapproached this threshold (Table 1) and in 2003 populationsizes were sufficient for a sizable epidemic to occur (Fig 5b)Rabies-control measures make it hard to determine whethercurrent densities in ND are high enough for infection to persistin the absence of vaccination but they may now be close
We detected only one canid-associated variant (Africa 1b)from a range of species in the Serengeti and were unable toreplicate East et alrsquos (2001) finding of a distinct virus variantcirculating atypically in spotted hyaenas (Lembo et al 2007)The lack of species-specific virusndashhost associations in thecanid-associated variant that we detected and the high degreeof genetic relatedness between virus samples from differentspecies indicate frequent cross-species transmission This isconsistent with results from southern Africa where phylogeneticbranches from a single geographic area include several hostspecies (Sabeta Bingham amp Nel 2003)
The statistical association between rabies cases in domesticdogs and other species also indicates cross-species transmissionUnfortunately the length of our time series and paucity ofcases detected in species other than domestic dogs precludeanalyses of lsquospill-overrsquo risk and reduce the interpretativepower of time-lagged regressions like those conducted forrabies in North America that could otherwise provide insight(Guerra et al 2003 Gordon et al 2004) In general as theamount of between-species transmission increases the moredifficult it becomes to tease apart the role of differentpotentially independent maintenance hosts We find greaterwithin-species and less between-species transmission thanwould be expected if species interacted randomly This patterncould occur as a consequence of the spatial distribution ofhost species andor preferential contact between conspecificsOur randomization indicates that large-scale patterns in thedistribution of hosts (gt 10 times 10 km) do not explain thislsquoassortativersquo transmission but we cannot make inference ona smaller scale These analyses suggest that occasionallsquospill-overrsquo of infection from domestic dog populations into
Fig 7 The proportion of estimated transmission within andbetween host types (associated with 1255 suspected rabies cases 1044in Serengeti and 211 in Ngorongoro) in Serengeti (light grey) andNgorongoro (dark grey) districts with lsquodogrsquo corresponding to domesticdogs and lsquocarnivorersquo to domestic cats and wild carnivores Credibilityintervals (95) are shown for expected transmission within andbetween host types assuming inter- and intraspecific transmissionwere equally likely and that livestock are dead-end hosts
Table 4 Human exposures to rabies as a result of bites by suspectedrabid animals in Serengeti and Ngorongoro districts (January 2002ndashDecember 2006) Data were obtained by contact tracing
SpeciesSerengeti district ()
Ngorongorodistrict ()
Domestic dogs 466 (85middot5) 84 (70middot0)Domestic cats 25 (4middot6) 6 (5middot0)Livestock 7 (1middot3) 3 (2middot5)Human 2 (0middot4) 1 (0middot8)Spotted hyaena Crocuta crocuta 4 (0middot7) 16 (13middot3)Honey badger Mellivora capensis 11 (2middot0) 6 (5middot0)African wildcat Felis lybica 0 (0) 2 (1middot7)White-tailed mongoose
Ichneumia albicauda2 (0middot4) 1 (0middot8)
Bat-eared fox Otocyon megalotis 2 (0middot4) 0 (0)Small-spotted genet Genetta genetta 1 (0middot2) 1 (0middot8)Jackal spp Canis spp 23 (4middot2) 0 (0)Leopard Panthera pardus 2 (0middot4) 0 (0)Total 545 (100) 120 (100)
10 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
alternative hosts sometimes results in short-lived chains oftransmission as contact is more common among conspecifics(Dobson 2004)
Definitive identification of infection reservoirs is extremelychallenging and rarely possible Despite intensive effortunderstanding of reservoir dynamics in many complexmulti-host systems remains incomplete (Woodroffe et al2005 Biek et al 2006 Independent Scientific Group on CattleTB 2007) Here the balance of evidence implicates domesticdogs as the only maintenance population of rabies in theSerengeti whereas other carnivores are nonessential tomaintenance However because they transmit disease totarget populations (and frequent transmission within andbetween such species may occur) they do constitute part ofthe reservoir (Table 4 Fig 1a) A single maintenance-hostspecies is consistent with broad patterns of rabies infectionobserved elsewhere (Smith 1989 Rupprecht et al 1991Wandeler et al 1994 Smith Orciari amp Yager 1995)
A lesson from this study is that even with considerable datafrom various sources we cannot exhaustively quantify thecontribution of transmission from all non-essential reservoircomponents However unravelling every thread of reservoirdynamics may not be essential Our key finding that wildlifeare not able to maintain rabies cycles or distinct geneticvariants provides new and critical information allowing rabiescontrol measures in the Serengeti to be targeted moreconfidently at domestic dog populations alone The long-term implication is that control of domestic dog rabies in theSerengeti will eliminate rabies in all other species includinghumans livestock and wildlife The absence of wildlifemaintenance hosts even in an area with such abundantwildlife raises the expectation that canine rabies eliminationis a feasible objective throughout much of Africa
Interventions to prevent transmission from domestic dogmaintenance populations would also provide convincingevidence with which to identify reservoirs Such approacheshave demonstrated that cattle were the reservoir for rinderpestafter cattle vaccinations rinderpest disappeared from wildlifeand the wildebeest Connochaetes taurinus population increasednearly ten-fold (Sinclair 1979 Plowright 1982 Dobson 1995)We are currently ring-vaccinating domestic dogs around SNPwhich should provide critical data to evaluate the impact ofalternative hosts on long-term control efforts in particular theireffects on the level of vaccination required for disease control
In conclusion resolving reservoir dynamics in any complexsystem is likely to prove challenging and our experiences inthe Serengeti suggest that consideration needs to be given toidentifying (i) the potential role of different componentsof the system (ii) the key information needed for differentdisease control strategies (iii) the epidemiological approacheslikely to yield most useful information and (iv) the range ofanalytical tools available
Acknowledgements
We are grateful to Tanzania Government ministries TANAPA TAWIRINCA Authority Tanzania Commission for Science and Technology andNational Institute for Medical Research for permissions and TANAPA and
TAWIRI Veterinary Units Viral Transmission Dynamics Project SerengetiLion and Cheetah Projects Frankfurt Zoological Society livestock field-officersof the Ministry of Water and Livestock Development in Mara and ArushaRegions M Magoto P Tiringa M Roelke-Parker R Fyumagwa H Wiickand Mwanza and Arusha Veterinary Investigation Centres for assistance withsample collection We are indebted to J Barrat AFSSA for performingdiagnostic testing and virus isolation and the staff of the Rabies Section of theCDC especially CE Rupprecht A Velasco-Villa LA Orciari IV Kuzminand M Niezgoda and PE Brandatildeo (Universidade de Satildeo Paulo) for technicalassistance with diagnostic tests and virus characterization We thankKLaurenson D Bennett J Pulliam and H Horn for valuable discussions
This work was supported by the joint National Institutes of Health (NIH)National Science Foundation (NSF) Ecology of Infectious Diseases Programunder grant no NSFDEB0225453 Opinions findings conclusions andrecommendations are those of the authors and do not necessarily reflectthe views of NIH or NSF KH was supported by NSF grant DEB-0513994the Pew Charitable Trusts award 2000-002558 to Princeton University theDepartment of Ecology and Evolution Princeton University and the HeinzFoundation TL was supported by the Royal (Dick) School of VeterinaryStudies University of Edinburgh and the University of Edinburgh DevelopmentTrust (visits to CDC) and SC by the Wellcome Trust and the Department forInternational Development Animal Health Programme
References
Barrat J (1996) Simple technique for the collection and shipment of brainspecimens for rabies diagnosis Laboratory Techniques in Rabies (edsF-X Meslin MM Kaplan amp H Koprowski) pp 425ndash432 World HealthOrganization Geneva Switzerland
Barrat J Barrat MJ Picard M amp Aubert MFA (1988) Diagnosis of rabiesby cell-culture Comparative Immunology Microbiology and InfectiousDiseases 11 207ndash214
Bartlett MS (1960) Stochastic Population Models in Ecology and EpidemiologyMethuen London
Bernardi F Nadin-Davis SA Wandeler AI Armstrong J Gomes AABLima FS Nogueira FRB amp Ito FH (2005) Antigenic and geneticcharacterization of rabies viruses isolated from domestic and wild animalsof Brazil identifies the hoary fox as a rabies reservoir Journal of GeneralVirology 86 3153ndash3162
Biek R Walsh PD Leroy EM amp Real LA (2006) Recent common ancestryof Ebola Zaire virus found in a bat reservoir Public Library of SciencePathogens 2 885ndash886
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999a) Theepidemiology of rabies in Zimbabwe 2 Rabies in jackals (Canis adustus andCanis mesomelas) Onderstepoort Journal of Veterinary Research 66 11ndash23
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999b) Theepidemiology of rabies in Zimbabwe 1 Rabies in dogs (Canis familiaris)Onderstepoort Journal of Veterinary Research 66 1ndash10
Bingham J Javangwe S Sabeta CT Wandeler AI amp Nel LH (2001)Report of isolations of unusual lyssaviruses (rabies and Mokola virus)identified retrospectively from Zimbabwe Journal of the South AfricanVeterinary Association 72 92ndash94
Brooks R (1990) Survey of the dog-population of Zimbabwe and its level ofrabies vaccination Veterinary Record 127 592ndash596
Cleaveland S amp Dye C (1995) Maintenance of a microparasite infectingseveral host species rabies in the Serengeti Parasitology 111 S33ndashS47
Cleaveland S Kaare M Tiringa P Mlengeya T amp Barrat J (2003) A dograbies vaccination campaign in rural Africa impact on the incidence of dograbies and human dog-bite injuries Vaccine 21 1965ndash1973
Cleaveland S Laurenson MK amp Taylor LH (2001) Diseases of humansand their domestic mammals pathogen characteristics host range andthe risk of emergence Philosophical Transactions of the Royal Society BBiological Sciences 356 991ndash999
Cleaveland S Mlengeya T Kaare M Haydon DT Lembo T LaurensonMK amp Packer C (2007) The conservation relevance of epidemiologicalresearch into carnivore viral diseases in the Serengeti Conservation Biology21 612ndash622
Conzelmann KK Cox JH Schneider LG amp Thiel HJ (1990) Molecularcloning and complete nucleotide sequence of the attenuated rabies virusSAD B19 Virology 175 485ndash499
Daszak P Cunningham AA amp Hyatt AD (2000) Wildlife ecology ndash Emerginginfectious diseases of wildlife ndash Threats to Biodiversity and human healthScience 287 443ndash449
Dean DJ Abelseth MK amp Atanasiu P (1996) The fluorescent antibodytest Laboratory Techniques in Rabies (eds F-X Meslin MM Kaplan
Rabies reservoir dynamics in the Serengeti 11
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
amp H Koprowski) pp 88ndash95 World Health Organization GenevaSwitzerland
Dobson A (1995) The ecology and epidemiology of rinderpest virus inSerengeti and Ngorongoro Conservation Area Serengeti II DynamicsManagement and Conservation of an Ecosystem (eds ARE Sinclair ampP Arcese) pp 485ndash505 University of Chicago Press Chicago
Dobson A (2004) Population dynamics of pathogens with multiple host speciesAmerican Naturalist 164 S64ndashS78
Dobson A amp Foufopoulos J (2001) Emerging infectious pathogens of wildlifePhilosophical Transactions of the Royal Society B Biological Sciences 3561001ndash1012
Dye C (1996) Serengeti wild dogs what really happened Trends in Ecology ampEvolution 11 188ndash189
East ML Hofer H Cox JH Wulle U Wiik H amp Pitra C (2001) Regularexposure to rabies virus and lack of symptomatic disease in Serengetispotted hyenas Proceedings of the National Academy of Sciences USA 9815026ndash15031
Foggin CM (1988) Rabies and Rabies-related Viruses in Zimbabwe HistoricalVirological and Ecological Aspects PhD thesis University of ZimbabweHarare Zimbabwe
Gascoyne SC King AA Laurenson MK Borner M Schildger B ampBarrat J (1993) Aspects of rabies infection and control in the conservationof the African wild dog (Lycaon pictus) in the Serengeti region TanzaniaOnderstepoort Journal of Veterinary Research 60 415ndash420
Gordon ER Curns AT Krebs JW Rupprecht CE Real LA ampChilds JE (2004) Temporal dynamics of rabies in a wildlife host and the riskof cross-species transmission Epidemiology and Infection 132 515ndash524
Guerra MA Curns AT Rupprecht CE Hanlon CA Krebs JW ampChilds JE (2003) Skunk and raccoon rabies in the eastern United Statestemporal and spatial analysis Emerging Infectious Diseases 9 1143ndash1150
Hall TA (1999) bioedit a user-friendly biological sequence alignment editorand analysis program for Windows 9598NT Nucleic Acids SymposiumSeries 41 95ndash98
Haydon DT Cleaveland S Taylor LH amp Laurenson MK (2002) Identi-fying reservoirs of infection a conceptual and practical challenge EmergingInfectious Diseases 8 1468ndash1473
Independent Scientific Group on Cattle TB (2007) Bovine TB The ScientificEvidence Defra Publications London PB12634 httpwwwdefragovukanimalhtbisgindexhtm
Jeanmougin F Thompson JD Gouy M Higgins DG amp Gibson TJ(1998) Multiple sequence alignment with clustal-x Trends in BiochemicalSciences 23 403ndash405
Johnson N Black C Smith J Un H McElhinney LM Aylan O ampFooks AR (2003) Rabies emergence among foxes in Turkey Journal ofWildlife Diseases 39 262ndash270
King AA Meredith CD amp Thomson GR (1993) Canid and viverrid rabiesviruses in South Africa Onderstepoort Journal of Veterinary Research 60295ndash299
Kissi B Tordo N amp Bourhy H (1995) Genetic polymorphism in the rabiesvirus nucleoprotein gene Virology 209 526ndash537
Kitala PM McDermott JJ Kyule MN amp Gathuma JM (2000) Community-based active surveillance for rabies in Machakos District Kenya PreventiveVeterinary Medicine 44 73ndash85
Kitala PM McDermott J Kyule M Gathuma JM Perry B amp Wandeler A(2001) Dog ecology and demography information to support the planning ofrabies control in Machakos District Kenya Acta Tropica 78 217ndash230
Knobel DL Cleaveland S Coleman PG Fevre EM Meltzer MIMiranda MEG Shaw A Zinsstag J amp Meslin F-X (2005) Re-evaluatingthe burden of rabies in Africa and Asia Bulletin of the World HealthOrganization 83 360ndash368
Lembo T Haydon DT Velasco-Villa A Rupprecht CE Packer CBrandatildeo PE Kuzmin IV Fooks AR Barrat J amp Cleaveland S (2007)Molecular epidemiology identifies only a single rabies virus variantcirculating in complex carnivore communities of the Serengeti Proceedingsof the Royal Society B Biological Sciences 274 2123ndash2130
Maas B (1993) Behavioural Ecology and Social Organisation of the Bat-earedFox in the Serengeti National Park Tanzania University of CambridgeCambridge UK
National Bureau of Statistics Tanzania (2005) 2002 Population and HousingCensus Presidentrsquos Office Planning and Privatization Dar es SalaamTanzania
Nel JAJ (1993) The bat-eared fox-a prime candidate for rabies vectorOnderstepoort Journal of Veterinary Research 60 395ndash397
Nel LH Sabeta CT von Teichman B Jaftha JB Rupprecht CE amp
Bingham J (2005) Mongoose rabies in southern Africa a re-evaluationbased on molecular epidemiology Virus Research 109 165ndash173
Page RDM (1996) treeview an application to display phylogenetic trees onpersonal computers Computer Applications in the Biosciences 12 357ndash358
Plowright W (1982) The effects of rinderpest and rinderpest control on wildlifein Africa Symposia of the Zoological Society of London 50 1ndash28
Posada D amp Crandall KA (1998) modeltest testing the model of DNAsubstitution Bioinformatics 14 817ndash818
Randall DA Marino J Haydon DT Sillero-Zubiri C Knobel DLTallents LA Macdonald DW amp Laurenson MK (2006) An integrateddisease management strategy for the control of rabies in Ethiopian wolvesBiological Conservation 131 151ndash162
Randall DA Williams SD Kuzmin IV Rupprecht CE Tallents LATefera Z Argaw K Shiferaw F Knobel DL Sillero-Zubiri C ampLaurenson MK (2004) Rabies in endangered Ethiopian wolves EmergingInfectious Diseases 10 2214ndash2217
Rhodes CJ Atkinson RPD Anderson RM amp Macdonald DW (1998)Rabies in Zimbabwe reservoir dogs and the implications for disease controlPhilosophical Transactions of the Royal Society B Biological Sciences 353999ndash1010
Roelke-Parker ME Munson L Packer C Kock R Cleaveland SCarpenter M Obrien SJ Pospischil A HofmannLehmann R Lutz HMwamengele GLM Mgasa MN Machange GA Summers BA ampAppel MJG (1996) A canine distemper virus epidemic in Serengeti lions(Panthera leo) Nature 379 441ndash445
Ronquist F amp Huelsenbeck JP (2003) mrbayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 19 1572ndash1574
Rupprecht CE Dietzschold B Wunner WH amp Koprowsky H (1991)Antigenic relationships of lyssaviruses The Natural History of Rabies (edsGM Baer) pp 69ndash100 CRC Press Boca Raton
Sabeta CT Bingham J amp Nel LH (2003) Molecular epidemiology of canidrabies in Zimbabwe and South Africa Virus Research 91 203ndash211
Sinclair ARE (1979) The eruption of the ruminants Serengeti Dynamics ofan Ecosystem (eds ARE Sinclair amp M Norton-Griffiths) pp 82ndash103University of Chicago Press Chicago
Sinclair ARE amp Arcese P (1995) Serengeti II Dynamics Management andConservation of an Ecosystem University of Chicago Press Chicago
Smith JS (1989) Rabies virus epitopic variation use in ecological studiesAdvances in Virus Research 36 215ndash253
Smith JS Orciari LA amp Yager PA (1995) Molecular epidemiology ofrabies in the United States Seminars in Virology 6 387ndash400
Swanepoel R Barnard BJH Meredith CD Bishop GC BrucknerGK Foggin CM amp Hubschle OJB (1993) Rabies in Southern AfricaOnderstepoort Journal of Veterinary Research 60 325ndash346
Taylor LH Latham SM amp Woolhouse MEJ (2001) Risk factors forhuman disease emergence Philosophical Transactions of the Royal SocietyB Biological Sciences 356 983ndash989
Tepsumethanon V Wilde H amp Meslin FX (2005) Six criteria for rabiesdiagnosis in living dogs Journal of the Medical Association of Thailand 88419ndash422
Thomson GR amp Meredith CD (1993) Rabies in bat-eared foxes in SouthAfrica Onderstepoort Journal of Veterinary Research 60 399ndash403
Vial F Cleaveland S Rasmussen G amp Haydon DT (2006) Development ofvaccination strategies for the management of rabies in African wild dogsBiological Conservation 131 180ndash192
Wandeler AI Budde A Capt S Kappeler A amp Matter H (1988) Dogecology and dog rabies control Reviews of Infectious Diseases 10 S684ndashS688
Wandeler AI Nadin-Davis SA Tinline RR amp Rupprecht CE (1994)Rabies epidemiology some ecological and evolutionary perspectivesLyssaviruses (eds CE Rupprecht B Dietzschold amp H Koprowski)pp 297ndash324 Springer-Verlag Berlin
Woodroffe R (2001) Assessing the risks of intervention immobilizationradio-collaring and vaccination of African wild dogs Oryx 35 234ndash244
Woodroffe R Donnelly CA Johnston WT Bourne FJ Cheeseman CLClifton-Hadley RS Cox DR Gettinby G Hewinson RG Le FevreAM McInerney JP amp Morrison WI (2005) Spatial association ofMycobacterium bovis infection in cattle and badgers Meles meles Journal ofApplied Ecology 42 852ndash862
World Health Organization (1999) World Survey of Rabies No 34 for the Year1998 Geneva Switzerland
Received 14 June 2007 accepted 23 January 2008Handling Editor Simon Thirgood
12 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Supplementary material
The following supplementary material is available for thisarticle
Appendix S1 Latin names of wildlife species
Appendix S2 Supplementary details of methods
Table S1 Virus samples from the study area compared inphylogenetic analysis
Table S2 Details of all wildlife confirmed and suspect casesin SNP
Table S3 Details of confirmed wildlife cases in Serengeti andNgorongoro districts
Fig S1 Incidence of bites by domestic dogs in Serengeti andNgorongoro districts
This material is available as part of the online article fromhttpwwwblackwell-synergycomdoifull101111j1365-2664200801468x(This link will take you to the article abstract)
Please note Blackwell Publishing is not responsible for the con-tent or functionality of any supplementary materials suppliedby the authors Any queries (other than missing material)should be directed to the corresponding author for the article
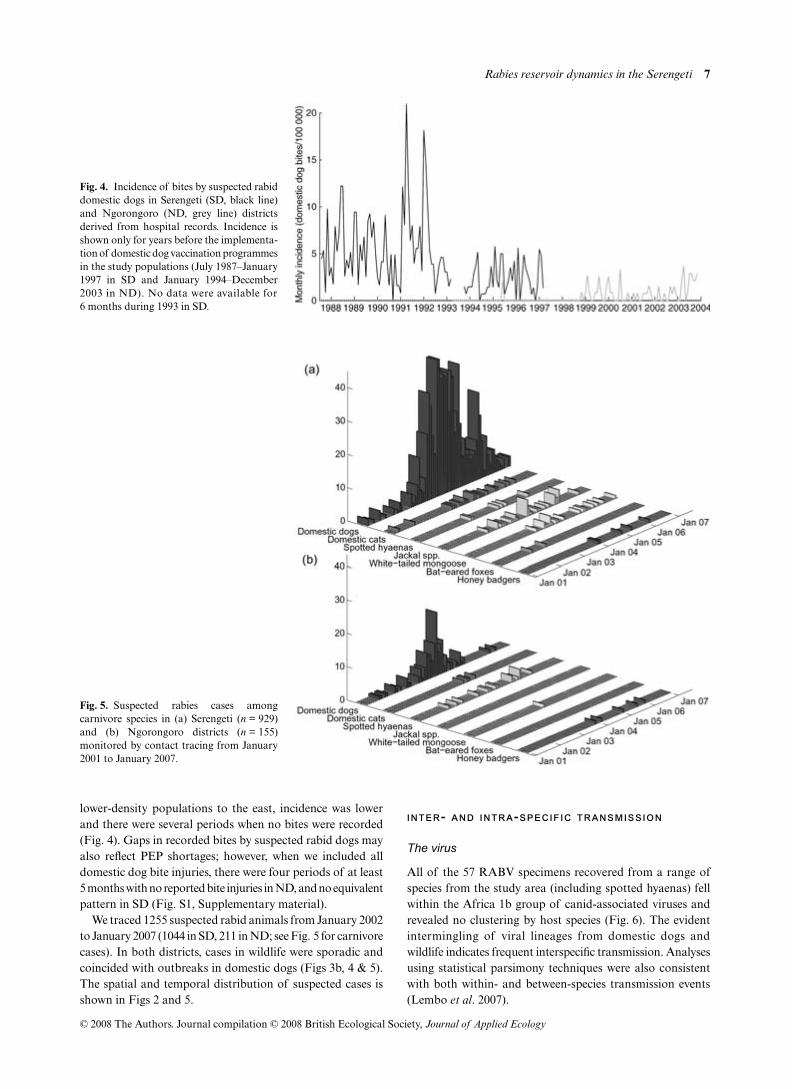
Rabies reservoir dynamics in the Serengeti 7
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
lower-density populations to the east incidence was lowerand there were several periods when no bites were recorded(Fig 4) Gaps in recorded bites by suspected rabid dogs mayalso reflect PEP shortages however when we included alldomestic dog bite injuries there were four periods of at least5 months with no reported bite injuries in ND and no equivalentpattern in SD (Fig S1 Supplementary material)
We traced 1255 suspected rabid animals from January 2002to January 2007 (1044 in SD 211 in ND see Fig 5 for carnivorecases) In both districts cases in wildlife were sporadic andcoincided with outbreaks in domestic dogs (Figs 3b 4 amp 5)The spatial and temporal distribution of suspected cases isshown in Figs 2 and 5
INTER- AND INTRA-SPECIF IC TRANSMISSION
The virus
All of the 57 RABV specimens recovered from a range ofspecies from the study area (including spotted hyaenas) fellwithin the Africa 1b group of canid-associated viruses andrevealed no clustering by host species (Fig 6) The evidentintermingling of viral lineages from domestic dogs andwildlife indicates frequent interspecific transmission Analysesusing statistical parsimony techniques were also consistentwith both within- and between-species transmission events(Lembo et al 2007)
Fig 4 Incidence of bites by suspected rabiddomestic dogs in Serengeti (SD black line)and Ngorongoro (ND grey line) districtsderived from hospital records Incidence isshown only for years before the implementa-tion of domestic dog vaccination programmesin the study populations (July 1987ndashJanuary1997 in SD and January 1994ndashDecember2003 in ND) No data were available for6 months during 1993 in SD
Fig 5 Suspected rabies cases amongcarnivore species in (a) Serengeti (n = 929)and (b) Ngorongoro districts (n = 155)monitored by contact tracing from January2001 to January 2007
8 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Spatiotemporal incidence patterns in hosts
Suspected rabies cases in domestic dogs were significantpredictors of those in other carnivores (pooled cases indomestic cats and wildlife P lt 0middot001 in both districts)Credibility intervals (95) of the expected frequency ofwithin- and between-species transmission events based uponthe null assumption of non-assortative transmission (derivedfrom the bootstrap analysis) are shown in Fig 7 Morewithin-species transmission than expected was estimated inboth districts (domestic dog-to-domestic dog transmissionP = 0middot052 in SD and P = 0middot008 in ND transmission withinother carnivores ie wild carnivores and domestic catsFelis catus P lt 0middot0001 in both districts) Estimated between-species transmission was less than expected (P lt 0middot03 in SD
and P lt 0middot001 in ND) except for transmission from domesticdogs to livestock These patterns were consistent and significantwhen the randomization procedure was carried out on a10 times 10 km grid while on a 5 times 5 km grid the patterns wereevident but no longer significant probably because of reducedstatistical power
The majority of human exposures to suspected rabid animalswere from domestic dogs (Table 4) Similarly domestic dogswere the predominant source of infection for livestock (Fig 7)
Discussion
Demonstrating persistence of infection is a prerequisite foridentifying reservoir(s) Domestic dogs living at high densities(within the SD) are the only species in which rabies has been
Fig 6 Phylogenetic tree of nucleoprotein gene sequences (282 bp 94 deduced amino acids nucleotide positions 1139ndash1420 on the SAD B19genome (Conzelmann et al 1990) for RABV samples from the study area compared with isolates from other areas of Africa The tree isconstructed using Bayesian phylogenetics under the transitional model of nucleotide evolution with a gamma-shaped distribution of rates acrosssites (TIM + Γ Posada amp Crandall 1998 base frequencies = 0middot3253 0middot2134 0middot2360 0middot2253 nucleotide substitution rates = 1middot0000 3middot67230middot4393 0middot4393 8middot1773 1middot0000 Γ = 0middot3390) For samples from the study area the year of collection is indicated within square bracketsPreviously published sequences are designated by strain names (Kissi et al 1995 Randall et al 2004) The tree is rooted with isolate 1500AFSdefined as outgroup representative of the lineage Africa 3 Nodal posterior probabilities gt 0middot95 are shown The scale indicates branch-lengthexpressed as the expected number of substitutions per site Species not definitively identified
Rabies reservoir dynamics in the Serengeti 9
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
recorded continuously throughout the study period whereascases in other species (inside and outside SNP) have beensporadic Although disease monitoring presumably detects alower proportion of cases in wildlife than domestic dogs itseems likely that infection in wildlife populations had it beenpresent continuously would have been detected in more recentyears where the number of samples collected and tested washigher than in previous years when rabies was recorded(Table 2 Fig 3b) These observations provide circumstantialevidence for rabies persistence in domestic dog populationsand lack of persistence in other carnivores Clusters ofwildlife cases that coincided with outbreaks in domestic dogpopulations (Figs 3b 4 amp 5) and explosive but transient(lt 2 months) epidemics previously reported in bat-eared foxes(Maas 1993) are also consistent with chains of transmissionthat are not sustained and contrast with incidence patterns inwild carnivores that are believed to maintain rabies throughindependent cycles elsewhere in Africa For example rabieswas recorded sporadically in bat-eared fox populations inSouth Africa from the 1950s to 1970 then continuouslysuggesting a shift to persistent infection (Thomson amp Meredith1993) and in Zimbabwe outbreaks in jackals although ofteninitiated by domestic dog cases appear to be sustained overseveral years (Bingham et al 1999ab)
Specific demographic attributes (high densities andorturnover) are thought to be required for sustained rabiestransmission A threshold density of gt 5 dogs kmndash2 has beensuggested for persistence of rabies in domestic dog populationsin Africa (Foggin 1988 Brooks 1990 Cleaveland amp Dye 1995Kitala et al 2001) Domestic dog populations in the studyregion have grown considerably and even in the lower-density
area (within the ND) the estimated average density hasapproached this threshold (Table 1) and in 2003 populationsizes were sufficient for a sizable epidemic to occur (Fig 5b)Rabies-control measures make it hard to determine whethercurrent densities in ND are high enough for infection to persistin the absence of vaccination but they may now be close
We detected only one canid-associated variant (Africa 1b)from a range of species in the Serengeti and were unable toreplicate East et alrsquos (2001) finding of a distinct virus variantcirculating atypically in spotted hyaenas (Lembo et al 2007)The lack of species-specific virusndashhost associations in thecanid-associated variant that we detected and the high degreeof genetic relatedness between virus samples from differentspecies indicate frequent cross-species transmission This isconsistent with results from southern Africa where phylogeneticbranches from a single geographic area include several hostspecies (Sabeta Bingham amp Nel 2003)
The statistical association between rabies cases in domesticdogs and other species also indicates cross-species transmissionUnfortunately the length of our time series and paucity ofcases detected in species other than domestic dogs precludeanalyses of lsquospill-overrsquo risk and reduce the interpretativepower of time-lagged regressions like those conducted forrabies in North America that could otherwise provide insight(Guerra et al 2003 Gordon et al 2004) In general as theamount of between-species transmission increases the moredifficult it becomes to tease apart the role of differentpotentially independent maintenance hosts We find greaterwithin-species and less between-species transmission thanwould be expected if species interacted randomly This patterncould occur as a consequence of the spatial distribution ofhost species andor preferential contact between conspecificsOur randomization indicates that large-scale patterns in thedistribution of hosts (gt 10 times 10 km) do not explain thislsquoassortativersquo transmission but we cannot make inference ona smaller scale These analyses suggest that occasionallsquospill-overrsquo of infection from domestic dog populations into
Fig 7 The proportion of estimated transmission within andbetween host types (associated with 1255 suspected rabies cases 1044in Serengeti and 211 in Ngorongoro) in Serengeti (light grey) andNgorongoro (dark grey) districts with lsquodogrsquo corresponding to domesticdogs and lsquocarnivorersquo to domestic cats and wild carnivores Credibilityintervals (95) are shown for expected transmission within andbetween host types assuming inter- and intraspecific transmissionwere equally likely and that livestock are dead-end hosts
Table 4 Human exposures to rabies as a result of bites by suspectedrabid animals in Serengeti and Ngorongoro districts (January 2002ndashDecember 2006) Data were obtained by contact tracing
SpeciesSerengeti district ()
Ngorongorodistrict ()
Domestic dogs 466 (85middot5) 84 (70middot0)Domestic cats 25 (4middot6) 6 (5middot0)Livestock 7 (1middot3) 3 (2middot5)Human 2 (0middot4) 1 (0middot8)Spotted hyaena Crocuta crocuta 4 (0middot7) 16 (13middot3)Honey badger Mellivora capensis 11 (2middot0) 6 (5middot0)African wildcat Felis lybica 0 (0) 2 (1middot7)White-tailed mongoose
Ichneumia albicauda2 (0middot4) 1 (0middot8)
Bat-eared fox Otocyon megalotis 2 (0middot4) 0 (0)Small-spotted genet Genetta genetta 1 (0middot2) 1 (0middot8)Jackal spp Canis spp 23 (4middot2) 0 (0)Leopard Panthera pardus 2 (0middot4) 0 (0)Total 545 (100) 120 (100)
10 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
alternative hosts sometimes results in short-lived chains oftransmission as contact is more common among conspecifics(Dobson 2004)
Definitive identification of infection reservoirs is extremelychallenging and rarely possible Despite intensive effortunderstanding of reservoir dynamics in many complexmulti-host systems remains incomplete (Woodroffe et al2005 Biek et al 2006 Independent Scientific Group on CattleTB 2007) Here the balance of evidence implicates domesticdogs as the only maintenance population of rabies in theSerengeti whereas other carnivores are nonessential tomaintenance However because they transmit disease totarget populations (and frequent transmission within andbetween such species may occur) they do constitute part ofthe reservoir (Table 4 Fig 1a) A single maintenance-hostspecies is consistent with broad patterns of rabies infectionobserved elsewhere (Smith 1989 Rupprecht et al 1991Wandeler et al 1994 Smith Orciari amp Yager 1995)
A lesson from this study is that even with considerable datafrom various sources we cannot exhaustively quantify thecontribution of transmission from all non-essential reservoircomponents However unravelling every thread of reservoirdynamics may not be essential Our key finding that wildlifeare not able to maintain rabies cycles or distinct geneticvariants provides new and critical information allowing rabiescontrol measures in the Serengeti to be targeted moreconfidently at domestic dog populations alone The long-term implication is that control of domestic dog rabies in theSerengeti will eliminate rabies in all other species includinghumans livestock and wildlife The absence of wildlifemaintenance hosts even in an area with such abundantwildlife raises the expectation that canine rabies eliminationis a feasible objective throughout much of Africa
Interventions to prevent transmission from domestic dogmaintenance populations would also provide convincingevidence with which to identify reservoirs Such approacheshave demonstrated that cattle were the reservoir for rinderpestafter cattle vaccinations rinderpest disappeared from wildlifeand the wildebeest Connochaetes taurinus population increasednearly ten-fold (Sinclair 1979 Plowright 1982 Dobson 1995)We are currently ring-vaccinating domestic dogs around SNPwhich should provide critical data to evaluate the impact ofalternative hosts on long-term control efforts in particular theireffects on the level of vaccination required for disease control
In conclusion resolving reservoir dynamics in any complexsystem is likely to prove challenging and our experiences inthe Serengeti suggest that consideration needs to be given toidentifying (i) the potential role of different componentsof the system (ii) the key information needed for differentdisease control strategies (iii) the epidemiological approacheslikely to yield most useful information and (iv) the range ofanalytical tools available
Acknowledgements
We are grateful to Tanzania Government ministries TANAPA TAWIRINCA Authority Tanzania Commission for Science and Technology andNational Institute for Medical Research for permissions and TANAPA and
TAWIRI Veterinary Units Viral Transmission Dynamics Project SerengetiLion and Cheetah Projects Frankfurt Zoological Society livestock field-officersof the Ministry of Water and Livestock Development in Mara and ArushaRegions M Magoto P Tiringa M Roelke-Parker R Fyumagwa H Wiickand Mwanza and Arusha Veterinary Investigation Centres for assistance withsample collection We are indebted to J Barrat AFSSA for performingdiagnostic testing and virus isolation and the staff of the Rabies Section of theCDC especially CE Rupprecht A Velasco-Villa LA Orciari IV Kuzminand M Niezgoda and PE Brandatildeo (Universidade de Satildeo Paulo) for technicalassistance with diagnostic tests and virus characterization We thankKLaurenson D Bennett J Pulliam and H Horn for valuable discussions
This work was supported by the joint National Institutes of Health (NIH)National Science Foundation (NSF) Ecology of Infectious Diseases Programunder grant no NSFDEB0225453 Opinions findings conclusions andrecommendations are those of the authors and do not necessarily reflectthe views of NIH or NSF KH was supported by NSF grant DEB-0513994the Pew Charitable Trusts award 2000-002558 to Princeton University theDepartment of Ecology and Evolution Princeton University and the HeinzFoundation TL was supported by the Royal (Dick) School of VeterinaryStudies University of Edinburgh and the University of Edinburgh DevelopmentTrust (visits to CDC) and SC by the Wellcome Trust and the Department forInternational Development Animal Health Programme
References
Barrat J (1996) Simple technique for the collection and shipment of brainspecimens for rabies diagnosis Laboratory Techniques in Rabies (edsF-X Meslin MM Kaplan amp H Koprowski) pp 425ndash432 World HealthOrganization Geneva Switzerland
Barrat J Barrat MJ Picard M amp Aubert MFA (1988) Diagnosis of rabiesby cell-culture Comparative Immunology Microbiology and InfectiousDiseases 11 207ndash214
Bartlett MS (1960) Stochastic Population Models in Ecology and EpidemiologyMethuen London
Bernardi F Nadin-Davis SA Wandeler AI Armstrong J Gomes AABLima FS Nogueira FRB amp Ito FH (2005) Antigenic and geneticcharacterization of rabies viruses isolated from domestic and wild animalsof Brazil identifies the hoary fox as a rabies reservoir Journal of GeneralVirology 86 3153ndash3162
Biek R Walsh PD Leroy EM amp Real LA (2006) Recent common ancestryof Ebola Zaire virus found in a bat reservoir Public Library of SciencePathogens 2 885ndash886
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999a) Theepidemiology of rabies in Zimbabwe 2 Rabies in jackals (Canis adustus andCanis mesomelas) Onderstepoort Journal of Veterinary Research 66 11ndash23
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999b) Theepidemiology of rabies in Zimbabwe 1 Rabies in dogs (Canis familiaris)Onderstepoort Journal of Veterinary Research 66 1ndash10
Bingham J Javangwe S Sabeta CT Wandeler AI amp Nel LH (2001)Report of isolations of unusual lyssaviruses (rabies and Mokola virus)identified retrospectively from Zimbabwe Journal of the South AfricanVeterinary Association 72 92ndash94
Brooks R (1990) Survey of the dog-population of Zimbabwe and its level ofrabies vaccination Veterinary Record 127 592ndash596
Cleaveland S amp Dye C (1995) Maintenance of a microparasite infectingseveral host species rabies in the Serengeti Parasitology 111 S33ndashS47
Cleaveland S Kaare M Tiringa P Mlengeya T amp Barrat J (2003) A dograbies vaccination campaign in rural Africa impact on the incidence of dograbies and human dog-bite injuries Vaccine 21 1965ndash1973
Cleaveland S Laurenson MK amp Taylor LH (2001) Diseases of humansand their domestic mammals pathogen characteristics host range andthe risk of emergence Philosophical Transactions of the Royal Society BBiological Sciences 356 991ndash999
Cleaveland S Mlengeya T Kaare M Haydon DT Lembo T LaurensonMK amp Packer C (2007) The conservation relevance of epidemiologicalresearch into carnivore viral diseases in the Serengeti Conservation Biology21 612ndash622
Conzelmann KK Cox JH Schneider LG amp Thiel HJ (1990) Molecularcloning and complete nucleotide sequence of the attenuated rabies virusSAD B19 Virology 175 485ndash499
Daszak P Cunningham AA amp Hyatt AD (2000) Wildlife ecology ndash Emerginginfectious diseases of wildlife ndash Threats to Biodiversity and human healthScience 287 443ndash449
Dean DJ Abelseth MK amp Atanasiu P (1996) The fluorescent antibodytest Laboratory Techniques in Rabies (eds F-X Meslin MM Kaplan
Rabies reservoir dynamics in the Serengeti 11
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
amp H Koprowski) pp 88ndash95 World Health Organization GenevaSwitzerland
Dobson A (1995) The ecology and epidemiology of rinderpest virus inSerengeti and Ngorongoro Conservation Area Serengeti II DynamicsManagement and Conservation of an Ecosystem (eds ARE Sinclair ampP Arcese) pp 485ndash505 University of Chicago Press Chicago
Dobson A (2004) Population dynamics of pathogens with multiple host speciesAmerican Naturalist 164 S64ndashS78
Dobson A amp Foufopoulos J (2001) Emerging infectious pathogens of wildlifePhilosophical Transactions of the Royal Society B Biological Sciences 3561001ndash1012
Dye C (1996) Serengeti wild dogs what really happened Trends in Ecology ampEvolution 11 188ndash189
East ML Hofer H Cox JH Wulle U Wiik H amp Pitra C (2001) Regularexposure to rabies virus and lack of symptomatic disease in Serengetispotted hyenas Proceedings of the National Academy of Sciences USA 9815026ndash15031
Foggin CM (1988) Rabies and Rabies-related Viruses in Zimbabwe HistoricalVirological and Ecological Aspects PhD thesis University of ZimbabweHarare Zimbabwe
Gascoyne SC King AA Laurenson MK Borner M Schildger B ampBarrat J (1993) Aspects of rabies infection and control in the conservationof the African wild dog (Lycaon pictus) in the Serengeti region TanzaniaOnderstepoort Journal of Veterinary Research 60 415ndash420
Gordon ER Curns AT Krebs JW Rupprecht CE Real LA ampChilds JE (2004) Temporal dynamics of rabies in a wildlife host and the riskof cross-species transmission Epidemiology and Infection 132 515ndash524
Guerra MA Curns AT Rupprecht CE Hanlon CA Krebs JW ampChilds JE (2003) Skunk and raccoon rabies in the eastern United Statestemporal and spatial analysis Emerging Infectious Diseases 9 1143ndash1150
Hall TA (1999) bioedit a user-friendly biological sequence alignment editorand analysis program for Windows 9598NT Nucleic Acids SymposiumSeries 41 95ndash98
Haydon DT Cleaveland S Taylor LH amp Laurenson MK (2002) Identi-fying reservoirs of infection a conceptual and practical challenge EmergingInfectious Diseases 8 1468ndash1473
Independent Scientific Group on Cattle TB (2007) Bovine TB The ScientificEvidence Defra Publications London PB12634 httpwwwdefragovukanimalhtbisgindexhtm
Jeanmougin F Thompson JD Gouy M Higgins DG amp Gibson TJ(1998) Multiple sequence alignment with clustal-x Trends in BiochemicalSciences 23 403ndash405
Johnson N Black C Smith J Un H McElhinney LM Aylan O ampFooks AR (2003) Rabies emergence among foxes in Turkey Journal ofWildlife Diseases 39 262ndash270
King AA Meredith CD amp Thomson GR (1993) Canid and viverrid rabiesviruses in South Africa Onderstepoort Journal of Veterinary Research 60295ndash299
Kissi B Tordo N amp Bourhy H (1995) Genetic polymorphism in the rabiesvirus nucleoprotein gene Virology 209 526ndash537
Kitala PM McDermott JJ Kyule MN amp Gathuma JM (2000) Community-based active surveillance for rabies in Machakos District Kenya PreventiveVeterinary Medicine 44 73ndash85
Kitala PM McDermott J Kyule M Gathuma JM Perry B amp Wandeler A(2001) Dog ecology and demography information to support the planning ofrabies control in Machakos District Kenya Acta Tropica 78 217ndash230
Knobel DL Cleaveland S Coleman PG Fevre EM Meltzer MIMiranda MEG Shaw A Zinsstag J amp Meslin F-X (2005) Re-evaluatingthe burden of rabies in Africa and Asia Bulletin of the World HealthOrganization 83 360ndash368
Lembo T Haydon DT Velasco-Villa A Rupprecht CE Packer CBrandatildeo PE Kuzmin IV Fooks AR Barrat J amp Cleaveland S (2007)Molecular epidemiology identifies only a single rabies virus variantcirculating in complex carnivore communities of the Serengeti Proceedingsof the Royal Society B Biological Sciences 274 2123ndash2130
Maas B (1993) Behavioural Ecology and Social Organisation of the Bat-earedFox in the Serengeti National Park Tanzania University of CambridgeCambridge UK
National Bureau of Statistics Tanzania (2005) 2002 Population and HousingCensus Presidentrsquos Office Planning and Privatization Dar es SalaamTanzania
Nel JAJ (1993) The bat-eared fox-a prime candidate for rabies vectorOnderstepoort Journal of Veterinary Research 60 395ndash397
Nel LH Sabeta CT von Teichman B Jaftha JB Rupprecht CE amp
Bingham J (2005) Mongoose rabies in southern Africa a re-evaluationbased on molecular epidemiology Virus Research 109 165ndash173
Page RDM (1996) treeview an application to display phylogenetic trees onpersonal computers Computer Applications in the Biosciences 12 357ndash358
Plowright W (1982) The effects of rinderpest and rinderpest control on wildlifein Africa Symposia of the Zoological Society of London 50 1ndash28
Posada D amp Crandall KA (1998) modeltest testing the model of DNAsubstitution Bioinformatics 14 817ndash818
Randall DA Marino J Haydon DT Sillero-Zubiri C Knobel DLTallents LA Macdonald DW amp Laurenson MK (2006) An integrateddisease management strategy for the control of rabies in Ethiopian wolvesBiological Conservation 131 151ndash162
Randall DA Williams SD Kuzmin IV Rupprecht CE Tallents LATefera Z Argaw K Shiferaw F Knobel DL Sillero-Zubiri C ampLaurenson MK (2004) Rabies in endangered Ethiopian wolves EmergingInfectious Diseases 10 2214ndash2217
Rhodes CJ Atkinson RPD Anderson RM amp Macdonald DW (1998)Rabies in Zimbabwe reservoir dogs and the implications for disease controlPhilosophical Transactions of the Royal Society B Biological Sciences 353999ndash1010
Roelke-Parker ME Munson L Packer C Kock R Cleaveland SCarpenter M Obrien SJ Pospischil A HofmannLehmann R Lutz HMwamengele GLM Mgasa MN Machange GA Summers BA ampAppel MJG (1996) A canine distemper virus epidemic in Serengeti lions(Panthera leo) Nature 379 441ndash445
Ronquist F amp Huelsenbeck JP (2003) mrbayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 19 1572ndash1574
Rupprecht CE Dietzschold B Wunner WH amp Koprowsky H (1991)Antigenic relationships of lyssaviruses The Natural History of Rabies (edsGM Baer) pp 69ndash100 CRC Press Boca Raton
Sabeta CT Bingham J amp Nel LH (2003) Molecular epidemiology of canidrabies in Zimbabwe and South Africa Virus Research 91 203ndash211
Sinclair ARE (1979) The eruption of the ruminants Serengeti Dynamics ofan Ecosystem (eds ARE Sinclair amp M Norton-Griffiths) pp 82ndash103University of Chicago Press Chicago
Sinclair ARE amp Arcese P (1995) Serengeti II Dynamics Management andConservation of an Ecosystem University of Chicago Press Chicago
Smith JS (1989) Rabies virus epitopic variation use in ecological studiesAdvances in Virus Research 36 215ndash253
Smith JS Orciari LA amp Yager PA (1995) Molecular epidemiology ofrabies in the United States Seminars in Virology 6 387ndash400
Swanepoel R Barnard BJH Meredith CD Bishop GC BrucknerGK Foggin CM amp Hubschle OJB (1993) Rabies in Southern AfricaOnderstepoort Journal of Veterinary Research 60 325ndash346
Taylor LH Latham SM amp Woolhouse MEJ (2001) Risk factors forhuman disease emergence Philosophical Transactions of the Royal SocietyB Biological Sciences 356 983ndash989
Tepsumethanon V Wilde H amp Meslin FX (2005) Six criteria for rabiesdiagnosis in living dogs Journal of the Medical Association of Thailand 88419ndash422
Thomson GR amp Meredith CD (1993) Rabies in bat-eared foxes in SouthAfrica Onderstepoort Journal of Veterinary Research 60 399ndash403
Vial F Cleaveland S Rasmussen G amp Haydon DT (2006) Development ofvaccination strategies for the management of rabies in African wild dogsBiological Conservation 131 180ndash192
Wandeler AI Budde A Capt S Kappeler A amp Matter H (1988) Dogecology and dog rabies control Reviews of Infectious Diseases 10 S684ndashS688
Wandeler AI Nadin-Davis SA Tinline RR amp Rupprecht CE (1994)Rabies epidemiology some ecological and evolutionary perspectivesLyssaviruses (eds CE Rupprecht B Dietzschold amp H Koprowski)pp 297ndash324 Springer-Verlag Berlin
Woodroffe R (2001) Assessing the risks of intervention immobilizationradio-collaring and vaccination of African wild dogs Oryx 35 234ndash244
Woodroffe R Donnelly CA Johnston WT Bourne FJ Cheeseman CLClifton-Hadley RS Cox DR Gettinby G Hewinson RG Le FevreAM McInerney JP amp Morrison WI (2005) Spatial association ofMycobacterium bovis infection in cattle and badgers Meles meles Journal ofApplied Ecology 42 852ndash862
World Health Organization (1999) World Survey of Rabies No 34 for the Year1998 Geneva Switzerland
Received 14 June 2007 accepted 23 January 2008Handling Editor Simon Thirgood
12 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Supplementary material
The following supplementary material is available for thisarticle
Appendix S1 Latin names of wildlife species
Appendix S2 Supplementary details of methods
Table S1 Virus samples from the study area compared inphylogenetic analysis
Table S2 Details of all wildlife confirmed and suspect casesin SNP
Table S3 Details of confirmed wildlife cases in Serengeti andNgorongoro districts
Fig S1 Incidence of bites by domestic dogs in Serengeti andNgorongoro districts
This material is available as part of the online article fromhttpwwwblackwell-synergycomdoifull101111j1365-2664200801468x(This link will take you to the article abstract)
Please note Blackwell Publishing is not responsible for the con-tent or functionality of any supplementary materials suppliedby the authors Any queries (other than missing material)should be directed to the corresponding author for the article

8 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Spatiotemporal incidence patterns in hosts
Suspected rabies cases in domestic dogs were significantpredictors of those in other carnivores (pooled cases indomestic cats and wildlife P lt 0middot001 in both districts)Credibility intervals (95) of the expected frequency ofwithin- and between-species transmission events based uponthe null assumption of non-assortative transmission (derivedfrom the bootstrap analysis) are shown in Fig 7 Morewithin-species transmission than expected was estimated inboth districts (domestic dog-to-domestic dog transmissionP = 0middot052 in SD and P = 0middot008 in ND transmission withinother carnivores ie wild carnivores and domestic catsFelis catus P lt 0middot0001 in both districts) Estimated between-species transmission was less than expected (P lt 0middot03 in SD
and P lt 0middot001 in ND) except for transmission from domesticdogs to livestock These patterns were consistent and significantwhen the randomization procedure was carried out on a10 times 10 km grid while on a 5 times 5 km grid the patterns wereevident but no longer significant probably because of reducedstatistical power
The majority of human exposures to suspected rabid animalswere from domestic dogs (Table 4) Similarly domestic dogswere the predominant source of infection for livestock (Fig 7)
Discussion
Demonstrating persistence of infection is a prerequisite foridentifying reservoir(s) Domestic dogs living at high densities(within the SD) are the only species in which rabies has been
Fig 6 Phylogenetic tree of nucleoprotein gene sequences (282 bp 94 deduced amino acids nucleotide positions 1139ndash1420 on the SAD B19genome (Conzelmann et al 1990) for RABV samples from the study area compared with isolates from other areas of Africa The tree isconstructed using Bayesian phylogenetics under the transitional model of nucleotide evolution with a gamma-shaped distribution of rates acrosssites (TIM + Γ Posada amp Crandall 1998 base frequencies = 0middot3253 0middot2134 0middot2360 0middot2253 nucleotide substitution rates = 1middot0000 3middot67230middot4393 0middot4393 8middot1773 1middot0000 Γ = 0middot3390) For samples from the study area the year of collection is indicated within square bracketsPreviously published sequences are designated by strain names (Kissi et al 1995 Randall et al 2004) The tree is rooted with isolate 1500AFSdefined as outgroup representative of the lineage Africa 3 Nodal posterior probabilities gt 0middot95 are shown The scale indicates branch-lengthexpressed as the expected number of substitutions per site Species not definitively identified
Rabies reservoir dynamics in the Serengeti 9
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
recorded continuously throughout the study period whereascases in other species (inside and outside SNP) have beensporadic Although disease monitoring presumably detects alower proportion of cases in wildlife than domestic dogs itseems likely that infection in wildlife populations had it beenpresent continuously would have been detected in more recentyears where the number of samples collected and tested washigher than in previous years when rabies was recorded(Table 2 Fig 3b) These observations provide circumstantialevidence for rabies persistence in domestic dog populationsand lack of persistence in other carnivores Clusters ofwildlife cases that coincided with outbreaks in domestic dogpopulations (Figs 3b 4 amp 5) and explosive but transient(lt 2 months) epidemics previously reported in bat-eared foxes(Maas 1993) are also consistent with chains of transmissionthat are not sustained and contrast with incidence patterns inwild carnivores that are believed to maintain rabies throughindependent cycles elsewhere in Africa For example rabieswas recorded sporadically in bat-eared fox populations inSouth Africa from the 1950s to 1970 then continuouslysuggesting a shift to persistent infection (Thomson amp Meredith1993) and in Zimbabwe outbreaks in jackals although ofteninitiated by domestic dog cases appear to be sustained overseveral years (Bingham et al 1999ab)
Specific demographic attributes (high densities andorturnover) are thought to be required for sustained rabiestransmission A threshold density of gt 5 dogs kmndash2 has beensuggested for persistence of rabies in domestic dog populationsin Africa (Foggin 1988 Brooks 1990 Cleaveland amp Dye 1995Kitala et al 2001) Domestic dog populations in the studyregion have grown considerably and even in the lower-density
area (within the ND) the estimated average density hasapproached this threshold (Table 1) and in 2003 populationsizes were sufficient for a sizable epidemic to occur (Fig 5b)Rabies-control measures make it hard to determine whethercurrent densities in ND are high enough for infection to persistin the absence of vaccination but they may now be close
We detected only one canid-associated variant (Africa 1b)from a range of species in the Serengeti and were unable toreplicate East et alrsquos (2001) finding of a distinct virus variantcirculating atypically in spotted hyaenas (Lembo et al 2007)The lack of species-specific virusndashhost associations in thecanid-associated variant that we detected and the high degreeof genetic relatedness between virus samples from differentspecies indicate frequent cross-species transmission This isconsistent with results from southern Africa where phylogeneticbranches from a single geographic area include several hostspecies (Sabeta Bingham amp Nel 2003)
The statistical association between rabies cases in domesticdogs and other species also indicates cross-species transmissionUnfortunately the length of our time series and paucity ofcases detected in species other than domestic dogs precludeanalyses of lsquospill-overrsquo risk and reduce the interpretativepower of time-lagged regressions like those conducted forrabies in North America that could otherwise provide insight(Guerra et al 2003 Gordon et al 2004) In general as theamount of between-species transmission increases the moredifficult it becomes to tease apart the role of differentpotentially independent maintenance hosts We find greaterwithin-species and less between-species transmission thanwould be expected if species interacted randomly This patterncould occur as a consequence of the spatial distribution ofhost species andor preferential contact between conspecificsOur randomization indicates that large-scale patterns in thedistribution of hosts (gt 10 times 10 km) do not explain thislsquoassortativersquo transmission but we cannot make inference ona smaller scale These analyses suggest that occasionallsquospill-overrsquo of infection from domestic dog populations into
Fig 7 The proportion of estimated transmission within andbetween host types (associated with 1255 suspected rabies cases 1044in Serengeti and 211 in Ngorongoro) in Serengeti (light grey) andNgorongoro (dark grey) districts with lsquodogrsquo corresponding to domesticdogs and lsquocarnivorersquo to domestic cats and wild carnivores Credibilityintervals (95) are shown for expected transmission within andbetween host types assuming inter- and intraspecific transmissionwere equally likely and that livestock are dead-end hosts
Table 4 Human exposures to rabies as a result of bites by suspectedrabid animals in Serengeti and Ngorongoro districts (January 2002ndashDecember 2006) Data were obtained by contact tracing
SpeciesSerengeti district ()
Ngorongorodistrict ()
Domestic dogs 466 (85middot5) 84 (70middot0)Domestic cats 25 (4middot6) 6 (5middot0)Livestock 7 (1middot3) 3 (2middot5)Human 2 (0middot4) 1 (0middot8)Spotted hyaena Crocuta crocuta 4 (0middot7) 16 (13middot3)Honey badger Mellivora capensis 11 (2middot0) 6 (5middot0)African wildcat Felis lybica 0 (0) 2 (1middot7)White-tailed mongoose
Ichneumia albicauda2 (0middot4) 1 (0middot8)
Bat-eared fox Otocyon megalotis 2 (0middot4) 0 (0)Small-spotted genet Genetta genetta 1 (0middot2) 1 (0middot8)Jackal spp Canis spp 23 (4middot2) 0 (0)Leopard Panthera pardus 2 (0middot4) 0 (0)Total 545 (100) 120 (100)
10 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
alternative hosts sometimes results in short-lived chains oftransmission as contact is more common among conspecifics(Dobson 2004)
Definitive identification of infection reservoirs is extremelychallenging and rarely possible Despite intensive effortunderstanding of reservoir dynamics in many complexmulti-host systems remains incomplete (Woodroffe et al2005 Biek et al 2006 Independent Scientific Group on CattleTB 2007) Here the balance of evidence implicates domesticdogs as the only maintenance population of rabies in theSerengeti whereas other carnivores are nonessential tomaintenance However because they transmit disease totarget populations (and frequent transmission within andbetween such species may occur) they do constitute part ofthe reservoir (Table 4 Fig 1a) A single maintenance-hostspecies is consistent with broad patterns of rabies infectionobserved elsewhere (Smith 1989 Rupprecht et al 1991Wandeler et al 1994 Smith Orciari amp Yager 1995)
A lesson from this study is that even with considerable datafrom various sources we cannot exhaustively quantify thecontribution of transmission from all non-essential reservoircomponents However unravelling every thread of reservoirdynamics may not be essential Our key finding that wildlifeare not able to maintain rabies cycles or distinct geneticvariants provides new and critical information allowing rabiescontrol measures in the Serengeti to be targeted moreconfidently at domestic dog populations alone The long-term implication is that control of domestic dog rabies in theSerengeti will eliminate rabies in all other species includinghumans livestock and wildlife The absence of wildlifemaintenance hosts even in an area with such abundantwildlife raises the expectation that canine rabies eliminationis a feasible objective throughout much of Africa
Interventions to prevent transmission from domestic dogmaintenance populations would also provide convincingevidence with which to identify reservoirs Such approacheshave demonstrated that cattle were the reservoir for rinderpestafter cattle vaccinations rinderpest disappeared from wildlifeand the wildebeest Connochaetes taurinus population increasednearly ten-fold (Sinclair 1979 Plowright 1982 Dobson 1995)We are currently ring-vaccinating domestic dogs around SNPwhich should provide critical data to evaluate the impact ofalternative hosts on long-term control efforts in particular theireffects on the level of vaccination required for disease control
In conclusion resolving reservoir dynamics in any complexsystem is likely to prove challenging and our experiences inthe Serengeti suggest that consideration needs to be given toidentifying (i) the potential role of different componentsof the system (ii) the key information needed for differentdisease control strategies (iii) the epidemiological approacheslikely to yield most useful information and (iv) the range ofanalytical tools available
Acknowledgements
We are grateful to Tanzania Government ministries TANAPA TAWIRINCA Authority Tanzania Commission for Science and Technology andNational Institute for Medical Research for permissions and TANAPA and
TAWIRI Veterinary Units Viral Transmission Dynamics Project SerengetiLion and Cheetah Projects Frankfurt Zoological Society livestock field-officersof the Ministry of Water and Livestock Development in Mara and ArushaRegions M Magoto P Tiringa M Roelke-Parker R Fyumagwa H Wiickand Mwanza and Arusha Veterinary Investigation Centres for assistance withsample collection We are indebted to J Barrat AFSSA for performingdiagnostic testing and virus isolation and the staff of the Rabies Section of theCDC especially CE Rupprecht A Velasco-Villa LA Orciari IV Kuzminand M Niezgoda and PE Brandatildeo (Universidade de Satildeo Paulo) for technicalassistance with diagnostic tests and virus characterization We thankKLaurenson D Bennett J Pulliam and H Horn for valuable discussions
This work was supported by the joint National Institutes of Health (NIH)National Science Foundation (NSF) Ecology of Infectious Diseases Programunder grant no NSFDEB0225453 Opinions findings conclusions andrecommendations are those of the authors and do not necessarily reflectthe views of NIH or NSF KH was supported by NSF grant DEB-0513994the Pew Charitable Trusts award 2000-002558 to Princeton University theDepartment of Ecology and Evolution Princeton University and the HeinzFoundation TL was supported by the Royal (Dick) School of VeterinaryStudies University of Edinburgh and the University of Edinburgh DevelopmentTrust (visits to CDC) and SC by the Wellcome Trust and the Department forInternational Development Animal Health Programme
References
Barrat J (1996) Simple technique for the collection and shipment of brainspecimens for rabies diagnosis Laboratory Techniques in Rabies (edsF-X Meslin MM Kaplan amp H Koprowski) pp 425ndash432 World HealthOrganization Geneva Switzerland
Barrat J Barrat MJ Picard M amp Aubert MFA (1988) Diagnosis of rabiesby cell-culture Comparative Immunology Microbiology and InfectiousDiseases 11 207ndash214
Bartlett MS (1960) Stochastic Population Models in Ecology and EpidemiologyMethuen London
Bernardi F Nadin-Davis SA Wandeler AI Armstrong J Gomes AABLima FS Nogueira FRB amp Ito FH (2005) Antigenic and geneticcharacterization of rabies viruses isolated from domestic and wild animalsof Brazil identifies the hoary fox as a rabies reservoir Journal of GeneralVirology 86 3153ndash3162
Biek R Walsh PD Leroy EM amp Real LA (2006) Recent common ancestryof Ebola Zaire virus found in a bat reservoir Public Library of SciencePathogens 2 885ndash886
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999a) Theepidemiology of rabies in Zimbabwe 2 Rabies in jackals (Canis adustus andCanis mesomelas) Onderstepoort Journal of Veterinary Research 66 11ndash23
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999b) Theepidemiology of rabies in Zimbabwe 1 Rabies in dogs (Canis familiaris)Onderstepoort Journal of Veterinary Research 66 1ndash10
Bingham J Javangwe S Sabeta CT Wandeler AI amp Nel LH (2001)Report of isolations of unusual lyssaviruses (rabies and Mokola virus)identified retrospectively from Zimbabwe Journal of the South AfricanVeterinary Association 72 92ndash94
Brooks R (1990) Survey of the dog-population of Zimbabwe and its level ofrabies vaccination Veterinary Record 127 592ndash596
Cleaveland S amp Dye C (1995) Maintenance of a microparasite infectingseveral host species rabies in the Serengeti Parasitology 111 S33ndashS47
Cleaveland S Kaare M Tiringa P Mlengeya T amp Barrat J (2003) A dograbies vaccination campaign in rural Africa impact on the incidence of dograbies and human dog-bite injuries Vaccine 21 1965ndash1973
Cleaveland S Laurenson MK amp Taylor LH (2001) Diseases of humansand their domestic mammals pathogen characteristics host range andthe risk of emergence Philosophical Transactions of the Royal Society BBiological Sciences 356 991ndash999
Cleaveland S Mlengeya T Kaare M Haydon DT Lembo T LaurensonMK amp Packer C (2007) The conservation relevance of epidemiologicalresearch into carnivore viral diseases in the Serengeti Conservation Biology21 612ndash622
Conzelmann KK Cox JH Schneider LG amp Thiel HJ (1990) Molecularcloning and complete nucleotide sequence of the attenuated rabies virusSAD B19 Virology 175 485ndash499
Daszak P Cunningham AA amp Hyatt AD (2000) Wildlife ecology ndash Emerginginfectious diseases of wildlife ndash Threats to Biodiversity and human healthScience 287 443ndash449
Dean DJ Abelseth MK amp Atanasiu P (1996) The fluorescent antibodytest Laboratory Techniques in Rabies (eds F-X Meslin MM Kaplan
Rabies reservoir dynamics in the Serengeti 11
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
amp H Koprowski) pp 88ndash95 World Health Organization GenevaSwitzerland
Dobson A (1995) The ecology and epidemiology of rinderpest virus inSerengeti and Ngorongoro Conservation Area Serengeti II DynamicsManagement and Conservation of an Ecosystem (eds ARE Sinclair ampP Arcese) pp 485ndash505 University of Chicago Press Chicago
Dobson A (2004) Population dynamics of pathogens with multiple host speciesAmerican Naturalist 164 S64ndashS78
Dobson A amp Foufopoulos J (2001) Emerging infectious pathogens of wildlifePhilosophical Transactions of the Royal Society B Biological Sciences 3561001ndash1012
Dye C (1996) Serengeti wild dogs what really happened Trends in Ecology ampEvolution 11 188ndash189
East ML Hofer H Cox JH Wulle U Wiik H amp Pitra C (2001) Regularexposure to rabies virus and lack of symptomatic disease in Serengetispotted hyenas Proceedings of the National Academy of Sciences USA 9815026ndash15031
Foggin CM (1988) Rabies and Rabies-related Viruses in Zimbabwe HistoricalVirological and Ecological Aspects PhD thesis University of ZimbabweHarare Zimbabwe
Gascoyne SC King AA Laurenson MK Borner M Schildger B ampBarrat J (1993) Aspects of rabies infection and control in the conservationof the African wild dog (Lycaon pictus) in the Serengeti region TanzaniaOnderstepoort Journal of Veterinary Research 60 415ndash420
Gordon ER Curns AT Krebs JW Rupprecht CE Real LA ampChilds JE (2004) Temporal dynamics of rabies in a wildlife host and the riskof cross-species transmission Epidemiology and Infection 132 515ndash524
Guerra MA Curns AT Rupprecht CE Hanlon CA Krebs JW ampChilds JE (2003) Skunk and raccoon rabies in the eastern United Statestemporal and spatial analysis Emerging Infectious Diseases 9 1143ndash1150
Hall TA (1999) bioedit a user-friendly biological sequence alignment editorand analysis program for Windows 9598NT Nucleic Acids SymposiumSeries 41 95ndash98
Haydon DT Cleaveland S Taylor LH amp Laurenson MK (2002) Identi-fying reservoirs of infection a conceptual and practical challenge EmergingInfectious Diseases 8 1468ndash1473
Independent Scientific Group on Cattle TB (2007) Bovine TB The ScientificEvidence Defra Publications London PB12634 httpwwwdefragovukanimalhtbisgindexhtm
Jeanmougin F Thompson JD Gouy M Higgins DG amp Gibson TJ(1998) Multiple sequence alignment with clustal-x Trends in BiochemicalSciences 23 403ndash405
Johnson N Black C Smith J Un H McElhinney LM Aylan O ampFooks AR (2003) Rabies emergence among foxes in Turkey Journal ofWildlife Diseases 39 262ndash270
King AA Meredith CD amp Thomson GR (1993) Canid and viverrid rabiesviruses in South Africa Onderstepoort Journal of Veterinary Research 60295ndash299
Kissi B Tordo N amp Bourhy H (1995) Genetic polymorphism in the rabiesvirus nucleoprotein gene Virology 209 526ndash537
Kitala PM McDermott JJ Kyule MN amp Gathuma JM (2000) Community-based active surveillance for rabies in Machakos District Kenya PreventiveVeterinary Medicine 44 73ndash85
Kitala PM McDermott J Kyule M Gathuma JM Perry B amp Wandeler A(2001) Dog ecology and demography information to support the planning ofrabies control in Machakos District Kenya Acta Tropica 78 217ndash230
Knobel DL Cleaveland S Coleman PG Fevre EM Meltzer MIMiranda MEG Shaw A Zinsstag J amp Meslin F-X (2005) Re-evaluatingthe burden of rabies in Africa and Asia Bulletin of the World HealthOrganization 83 360ndash368
Lembo T Haydon DT Velasco-Villa A Rupprecht CE Packer CBrandatildeo PE Kuzmin IV Fooks AR Barrat J amp Cleaveland S (2007)Molecular epidemiology identifies only a single rabies virus variantcirculating in complex carnivore communities of the Serengeti Proceedingsof the Royal Society B Biological Sciences 274 2123ndash2130
Maas B (1993) Behavioural Ecology and Social Organisation of the Bat-earedFox in the Serengeti National Park Tanzania University of CambridgeCambridge UK
National Bureau of Statistics Tanzania (2005) 2002 Population and HousingCensus Presidentrsquos Office Planning and Privatization Dar es SalaamTanzania
Nel JAJ (1993) The bat-eared fox-a prime candidate for rabies vectorOnderstepoort Journal of Veterinary Research 60 395ndash397
Nel LH Sabeta CT von Teichman B Jaftha JB Rupprecht CE amp
Bingham J (2005) Mongoose rabies in southern Africa a re-evaluationbased on molecular epidemiology Virus Research 109 165ndash173
Page RDM (1996) treeview an application to display phylogenetic trees onpersonal computers Computer Applications in the Biosciences 12 357ndash358
Plowright W (1982) The effects of rinderpest and rinderpest control on wildlifein Africa Symposia of the Zoological Society of London 50 1ndash28
Posada D amp Crandall KA (1998) modeltest testing the model of DNAsubstitution Bioinformatics 14 817ndash818
Randall DA Marino J Haydon DT Sillero-Zubiri C Knobel DLTallents LA Macdonald DW amp Laurenson MK (2006) An integrateddisease management strategy for the control of rabies in Ethiopian wolvesBiological Conservation 131 151ndash162
Randall DA Williams SD Kuzmin IV Rupprecht CE Tallents LATefera Z Argaw K Shiferaw F Knobel DL Sillero-Zubiri C ampLaurenson MK (2004) Rabies in endangered Ethiopian wolves EmergingInfectious Diseases 10 2214ndash2217
Rhodes CJ Atkinson RPD Anderson RM amp Macdonald DW (1998)Rabies in Zimbabwe reservoir dogs and the implications for disease controlPhilosophical Transactions of the Royal Society B Biological Sciences 353999ndash1010
Roelke-Parker ME Munson L Packer C Kock R Cleaveland SCarpenter M Obrien SJ Pospischil A HofmannLehmann R Lutz HMwamengele GLM Mgasa MN Machange GA Summers BA ampAppel MJG (1996) A canine distemper virus epidemic in Serengeti lions(Panthera leo) Nature 379 441ndash445
Ronquist F amp Huelsenbeck JP (2003) mrbayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 19 1572ndash1574
Rupprecht CE Dietzschold B Wunner WH amp Koprowsky H (1991)Antigenic relationships of lyssaviruses The Natural History of Rabies (edsGM Baer) pp 69ndash100 CRC Press Boca Raton
Sabeta CT Bingham J amp Nel LH (2003) Molecular epidemiology of canidrabies in Zimbabwe and South Africa Virus Research 91 203ndash211
Sinclair ARE (1979) The eruption of the ruminants Serengeti Dynamics ofan Ecosystem (eds ARE Sinclair amp M Norton-Griffiths) pp 82ndash103University of Chicago Press Chicago
Sinclair ARE amp Arcese P (1995) Serengeti II Dynamics Management andConservation of an Ecosystem University of Chicago Press Chicago
Smith JS (1989) Rabies virus epitopic variation use in ecological studiesAdvances in Virus Research 36 215ndash253
Smith JS Orciari LA amp Yager PA (1995) Molecular epidemiology ofrabies in the United States Seminars in Virology 6 387ndash400
Swanepoel R Barnard BJH Meredith CD Bishop GC BrucknerGK Foggin CM amp Hubschle OJB (1993) Rabies in Southern AfricaOnderstepoort Journal of Veterinary Research 60 325ndash346
Taylor LH Latham SM amp Woolhouse MEJ (2001) Risk factors forhuman disease emergence Philosophical Transactions of the Royal SocietyB Biological Sciences 356 983ndash989
Tepsumethanon V Wilde H amp Meslin FX (2005) Six criteria for rabiesdiagnosis in living dogs Journal of the Medical Association of Thailand 88419ndash422
Thomson GR amp Meredith CD (1993) Rabies in bat-eared foxes in SouthAfrica Onderstepoort Journal of Veterinary Research 60 399ndash403
Vial F Cleaveland S Rasmussen G amp Haydon DT (2006) Development ofvaccination strategies for the management of rabies in African wild dogsBiological Conservation 131 180ndash192
Wandeler AI Budde A Capt S Kappeler A amp Matter H (1988) Dogecology and dog rabies control Reviews of Infectious Diseases 10 S684ndashS688
Wandeler AI Nadin-Davis SA Tinline RR amp Rupprecht CE (1994)Rabies epidemiology some ecological and evolutionary perspectivesLyssaviruses (eds CE Rupprecht B Dietzschold amp H Koprowski)pp 297ndash324 Springer-Verlag Berlin
Woodroffe R (2001) Assessing the risks of intervention immobilizationradio-collaring and vaccination of African wild dogs Oryx 35 234ndash244
Woodroffe R Donnelly CA Johnston WT Bourne FJ Cheeseman CLClifton-Hadley RS Cox DR Gettinby G Hewinson RG Le FevreAM McInerney JP amp Morrison WI (2005) Spatial association ofMycobacterium bovis infection in cattle and badgers Meles meles Journal ofApplied Ecology 42 852ndash862
World Health Organization (1999) World Survey of Rabies No 34 for the Year1998 Geneva Switzerland
Received 14 June 2007 accepted 23 January 2008Handling Editor Simon Thirgood
12 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Supplementary material
The following supplementary material is available for thisarticle
Appendix S1 Latin names of wildlife species
Appendix S2 Supplementary details of methods
Table S1 Virus samples from the study area compared inphylogenetic analysis
Table S2 Details of all wildlife confirmed and suspect casesin SNP
Table S3 Details of confirmed wildlife cases in Serengeti andNgorongoro districts
Fig S1 Incidence of bites by domestic dogs in Serengeti andNgorongoro districts
This material is available as part of the online article fromhttpwwwblackwell-synergycomdoifull101111j1365-2664200801468x(This link will take you to the article abstract)
Please note Blackwell Publishing is not responsible for the con-tent or functionality of any supplementary materials suppliedby the authors Any queries (other than missing material)should be directed to the corresponding author for the article

Rabies reservoir dynamics in the Serengeti 9
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
recorded continuously throughout the study period whereascases in other species (inside and outside SNP) have beensporadic Although disease monitoring presumably detects alower proportion of cases in wildlife than domestic dogs itseems likely that infection in wildlife populations had it beenpresent continuously would have been detected in more recentyears where the number of samples collected and tested washigher than in previous years when rabies was recorded(Table 2 Fig 3b) These observations provide circumstantialevidence for rabies persistence in domestic dog populationsand lack of persistence in other carnivores Clusters ofwildlife cases that coincided with outbreaks in domestic dogpopulations (Figs 3b 4 amp 5) and explosive but transient(lt 2 months) epidemics previously reported in bat-eared foxes(Maas 1993) are also consistent with chains of transmissionthat are not sustained and contrast with incidence patterns inwild carnivores that are believed to maintain rabies throughindependent cycles elsewhere in Africa For example rabieswas recorded sporadically in bat-eared fox populations inSouth Africa from the 1950s to 1970 then continuouslysuggesting a shift to persistent infection (Thomson amp Meredith1993) and in Zimbabwe outbreaks in jackals although ofteninitiated by domestic dog cases appear to be sustained overseveral years (Bingham et al 1999ab)
Specific demographic attributes (high densities andorturnover) are thought to be required for sustained rabiestransmission A threshold density of gt 5 dogs kmndash2 has beensuggested for persistence of rabies in domestic dog populationsin Africa (Foggin 1988 Brooks 1990 Cleaveland amp Dye 1995Kitala et al 2001) Domestic dog populations in the studyregion have grown considerably and even in the lower-density
area (within the ND) the estimated average density hasapproached this threshold (Table 1) and in 2003 populationsizes were sufficient for a sizable epidemic to occur (Fig 5b)Rabies-control measures make it hard to determine whethercurrent densities in ND are high enough for infection to persistin the absence of vaccination but they may now be close
We detected only one canid-associated variant (Africa 1b)from a range of species in the Serengeti and were unable toreplicate East et alrsquos (2001) finding of a distinct virus variantcirculating atypically in spotted hyaenas (Lembo et al 2007)The lack of species-specific virusndashhost associations in thecanid-associated variant that we detected and the high degreeof genetic relatedness between virus samples from differentspecies indicate frequent cross-species transmission This isconsistent with results from southern Africa where phylogeneticbranches from a single geographic area include several hostspecies (Sabeta Bingham amp Nel 2003)
The statistical association between rabies cases in domesticdogs and other species also indicates cross-species transmissionUnfortunately the length of our time series and paucity ofcases detected in species other than domestic dogs precludeanalyses of lsquospill-overrsquo risk and reduce the interpretativepower of time-lagged regressions like those conducted forrabies in North America that could otherwise provide insight(Guerra et al 2003 Gordon et al 2004) In general as theamount of between-species transmission increases the moredifficult it becomes to tease apart the role of differentpotentially independent maintenance hosts We find greaterwithin-species and less between-species transmission thanwould be expected if species interacted randomly This patterncould occur as a consequence of the spatial distribution ofhost species andor preferential contact between conspecificsOur randomization indicates that large-scale patterns in thedistribution of hosts (gt 10 times 10 km) do not explain thislsquoassortativersquo transmission but we cannot make inference ona smaller scale These analyses suggest that occasionallsquospill-overrsquo of infection from domestic dog populations into
Fig 7 The proportion of estimated transmission within andbetween host types (associated with 1255 suspected rabies cases 1044in Serengeti and 211 in Ngorongoro) in Serengeti (light grey) andNgorongoro (dark grey) districts with lsquodogrsquo corresponding to domesticdogs and lsquocarnivorersquo to domestic cats and wild carnivores Credibilityintervals (95) are shown for expected transmission within andbetween host types assuming inter- and intraspecific transmissionwere equally likely and that livestock are dead-end hosts
Table 4 Human exposures to rabies as a result of bites by suspectedrabid animals in Serengeti and Ngorongoro districts (January 2002ndashDecember 2006) Data were obtained by contact tracing
SpeciesSerengeti district ()
Ngorongorodistrict ()
Domestic dogs 466 (85middot5) 84 (70middot0)Domestic cats 25 (4middot6) 6 (5middot0)Livestock 7 (1middot3) 3 (2middot5)Human 2 (0middot4) 1 (0middot8)Spotted hyaena Crocuta crocuta 4 (0middot7) 16 (13middot3)Honey badger Mellivora capensis 11 (2middot0) 6 (5middot0)African wildcat Felis lybica 0 (0) 2 (1middot7)White-tailed mongoose
Ichneumia albicauda2 (0middot4) 1 (0middot8)
Bat-eared fox Otocyon megalotis 2 (0middot4) 0 (0)Small-spotted genet Genetta genetta 1 (0middot2) 1 (0middot8)Jackal spp Canis spp 23 (4middot2) 0 (0)Leopard Panthera pardus 2 (0middot4) 0 (0)Total 545 (100) 120 (100)
10 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
alternative hosts sometimes results in short-lived chains oftransmission as contact is more common among conspecifics(Dobson 2004)
Definitive identification of infection reservoirs is extremelychallenging and rarely possible Despite intensive effortunderstanding of reservoir dynamics in many complexmulti-host systems remains incomplete (Woodroffe et al2005 Biek et al 2006 Independent Scientific Group on CattleTB 2007) Here the balance of evidence implicates domesticdogs as the only maintenance population of rabies in theSerengeti whereas other carnivores are nonessential tomaintenance However because they transmit disease totarget populations (and frequent transmission within andbetween such species may occur) they do constitute part ofthe reservoir (Table 4 Fig 1a) A single maintenance-hostspecies is consistent with broad patterns of rabies infectionobserved elsewhere (Smith 1989 Rupprecht et al 1991Wandeler et al 1994 Smith Orciari amp Yager 1995)
A lesson from this study is that even with considerable datafrom various sources we cannot exhaustively quantify thecontribution of transmission from all non-essential reservoircomponents However unravelling every thread of reservoirdynamics may not be essential Our key finding that wildlifeare not able to maintain rabies cycles or distinct geneticvariants provides new and critical information allowing rabiescontrol measures in the Serengeti to be targeted moreconfidently at domestic dog populations alone The long-term implication is that control of domestic dog rabies in theSerengeti will eliminate rabies in all other species includinghumans livestock and wildlife The absence of wildlifemaintenance hosts even in an area with such abundantwildlife raises the expectation that canine rabies eliminationis a feasible objective throughout much of Africa
Interventions to prevent transmission from domestic dogmaintenance populations would also provide convincingevidence with which to identify reservoirs Such approacheshave demonstrated that cattle were the reservoir for rinderpestafter cattle vaccinations rinderpest disappeared from wildlifeand the wildebeest Connochaetes taurinus population increasednearly ten-fold (Sinclair 1979 Plowright 1982 Dobson 1995)We are currently ring-vaccinating domestic dogs around SNPwhich should provide critical data to evaluate the impact ofalternative hosts on long-term control efforts in particular theireffects on the level of vaccination required for disease control
In conclusion resolving reservoir dynamics in any complexsystem is likely to prove challenging and our experiences inthe Serengeti suggest that consideration needs to be given toidentifying (i) the potential role of different componentsof the system (ii) the key information needed for differentdisease control strategies (iii) the epidemiological approacheslikely to yield most useful information and (iv) the range ofanalytical tools available
Acknowledgements
We are grateful to Tanzania Government ministries TANAPA TAWIRINCA Authority Tanzania Commission for Science and Technology andNational Institute for Medical Research for permissions and TANAPA and
TAWIRI Veterinary Units Viral Transmission Dynamics Project SerengetiLion and Cheetah Projects Frankfurt Zoological Society livestock field-officersof the Ministry of Water and Livestock Development in Mara and ArushaRegions M Magoto P Tiringa M Roelke-Parker R Fyumagwa H Wiickand Mwanza and Arusha Veterinary Investigation Centres for assistance withsample collection We are indebted to J Barrat AFSSA for performingdiagnostic testing and virus isolation and the staff of the Rabies Section of theCDC especially CE Rupprecht A Velasco-Villa LA Orciari IV Kuzminand M Niezgoda and PE Brandatildeo (Universidade de Satildeo Paulo) for technicalassistance with diagnostic tests and virus characterization We thankKLaurenson D Bennett J Pulliam and H Horn for valuable discussions
This work was supported by the joint National Institutes of Health (NIH)National Science Foundation (NSF) Ecology of Infectious Diseases Programunder grant no NSFDEB0225453 Opinions findings conclusions andrecommendations are those of the authors and do not necessarily reflectthe views of NIH or NSF KH was supported by NSF grant DEB-0513994the Pew Charitable Trusts award 2000-002558 to Princeton University theDepartment of Ecology and Evolution Princeton University and the HeinzFoundation TL was supported by the Royal (Dick) School of VeterinaryStudies University of Edinburgh and the University of Edinburgh DevelopmentTrust (visits to CDC) and SC by the Wellcome Trust and the Department forInternational Development Animal Health Programme
References
Barrat J (1996) Simple technique for the collection and shipment of brainspecimens for rabies diagnosis Laboratory Techniques in Rabies (edsF-X Meslin MM Kaplan amp H Koprowski) pp 425ndash432 World HealthOrganization Geneva Switzerland
Barrat J Barrat MJ Picard M amp Aubert MFA (1988) Diagnosis of rabiesby cell-culture Comparative Immunology Microbiology and InfectiousDiseases 11 207ndash214
Bartlett MS (1960) Stochastic Population Models in Ecology and EpidemiologyMethuen London
Bernardi F Nadin-Davis SA Wandeler AI Armstrong J Gomes AABLima FS Nogueira FRB amp Ito FH (2005) Antigenic and geneticcharacterization of rabies viruses isolated from domestic and wild animalsof Brazil identifies the hoary fox as a rabies reservoir Journal of GeneralVirology 86 3153ndash3162
Biek R Walsh PD Leroy EM amp Real LA (2006) Recent common ancestryof Ebola Zaire virus found in a bat reservoir Public Library of SciencePathogens 2 885ndash886
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999a) Theepidemiology of rabies in Zimbabwe 2 Rabies in jackals (Canis adustus andCanis mesomelas) Onderstepoort Journal of Veterinary Research 66 11ndash23
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999b) Theepidemiology of rabies in Zimbabwe 1 Rabies in dogs (Canis familiaris)Onderstepoort Journal of Veterinary Research 66 1ndash10
Bingham J Javangwe S Sabeta CT Wandeler AI amp Nel LH (2001)Report of isolations of unusual lyssaviruses (rabies and Mokola virus)identified retrospectively from Zimbabwe Journal of the South AfricanVeterinary Association 72 92ndash94
Brooks R (1990) Survey of the dog-population of Zimbabwe and its level ofrabies vaccination Veterinary Record 127 592ndash596
Cleaveland S amp Dye C (1995) Maintenance of a microparasite infectingseveral host species rabies in the Serengeti Parasitology 111 S33ndashS47
Cleaveland S Kaare M Tiringa P Mlengeya T amp Barrat J (2003) A dograbies vaccination campaign in rural Africa impact on the incidence of dograbies and human dog-bite injuries Vaccine 21 1965ndash1973
Cleaveland S Laurenson MK amp Taylor LH (2001) Diseases of humansand their domestic mammals pathogen characteristics host range andthe risk of emergence Philosophical Transactions of the Royal Society BBiological Sciences 356 991ndash999
Cleaveland S Mlengeya T Kaare M Haydon DT Lembo T LaurensonMK amp Packer C (2007) The conservation relevance of epidemiologicalresearch into carnivore viral diseases in the Serengeti Conservation Biology21 612ndash622
Conzelmann KK Cox JH Schneider LG amp Thiel HJ (1990) Molecularcloning and complete nucleotide sequence of the attenuated rabies virusSAD B19 Virology 175 485ndash499
Daszak P Cunningham AA amp Hyatt AD (2000) Wildlife ecology ndash Emerginginfectious diseases of wildlife ndash Threats to Biodiversity and human healthScience 287 443ndash449
Dean DJ Abelseth MK amp Atanasiu P (1996) The fluorescent antibodytest Laboratory Techniques in Rabies (eds F-X Meslin MM Kaplan
Rabies reservoir dynamics in the Serengeti 11
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
amp H Koprowski) pp 88ndash95 World Health Organization GenevaSwitzerland
Dobson A (1995) The ecology and epidemiology of rinderpest virus inSerengeti and Ngorongoro Conservation Area Serengeti II DynamicsManagement and Conservation of an Ecosystem (eds ARE Sinclair ampP Arcese) pp 485ndash505 University of Chicago Press Chicago
Dobson A (2004) Population dynamics of pathogens with multiple host speciesAmerican Naturalist 164 S64ndashS78
Dobson A amp Foufopoulos J (2001) Emerging infectious pathogens of wildlifePhilosophical Transactions of the Royal Society B Biological Sciences 3561001ndash1012
Dye C (1996) Serengeti wild dogs what really happened Trends in Ecology ampEvolution 11 188ndash189
East ML Hofer H Cox JH Wulle U Wiik H amp Pitra C (2001) Regularexposure to rabies virus and lack of symptomatic disease in Serengetispotted hyenas Proceedings of the National Academy of Sciences USA 9815026ndash15031
Foggin CM (1988) Rabies and Rabies-related Viruses in Zimbabwe HistoricalVirological and Ecological Aspects PhD thesis University of ZimbabweHarare Zimbabwe
Gascoyne SC King AA Laurenson MK Borner M Schildger B ampBarrat J (1993) Aspects of rabies infection and control in the conservationof the African wild dog (Lycaon pictus) in the Serengeti region TanzaniaOnderstepoort Journal of Veterinary Research 60 415ndash420
Gordon ER Curns AT Krebs JW Rupprecht CE Real LA ampChilds JE (2004) Temporal dynamics of rabies in a wildlife host and the riskof cross-species transmission Epidemiology and Infection 132 515ndash524
Guerra MA Curns AT Rupprecht CE Hanlon CA Krebs JW ampChilds JE (2003) Skunk and raccoon rabies in the eastern United Statestemporal and spatial analysis Emerging Infectious Diseases 9 1143ndash1150
Hall TA (1999) bioedit a user-friendly biological sequence alignment editorand analysis program for Windows 9598NT Nucleic Acids SymposiumSeries 41 95ndash98
Haydon DT Cleaveland S Taylor LH amp Laurenson MK (2002) Identi-fying reservoirs of infection a conceptual and practical challenge EmergingInfectious Diseases 8 1468ndash1473
Independent Scientific Group on Cattle TB (2007) Bovine TB The ScientificEvidence Defra Publications London PB12634 httpwwwdefragovukanimalhtbisgindexhtm
Jeanmougin F Thompson JD Gouy M Higgins DG amp Gibson TJ(1998) Multiple sequence alignment with clustal-x Trends in BiochemicalSciences 23 403ndash405
Johnson N Black C Smith J Un H McElhinney LM Aylan O ampFooks AR (2003) Rabies emergence among foxes in Turkey Journal ofWildlife Diseases 39 262ndash270
King AA Meredith CD amp Thomson GR (1993) Canid and viverrid rabiesviruses in South Africa Onderstepoort Journal of Veterinary Research 60295ndash299
Kissi B Tordo N amp Bourhy H (1995) Genetic polymorphism in the rabiesvirus nucleoprotein gene Virology 209 526ndash537
Kitala PM McDermott JJ Kyule MN amp Gathuma JM (2000) Community-based active surveillance for rabies in Machakos District Kenya PreventiveVeterinary Medicine 44 73ndash85
Kitala PM McDermott J Kyule M Gathuma JM Perry B amp Wandeler A(2001) Dog ecology and demography information to support the planning ofrabies control in Machakos District Kenya Acta Tropica 78 217ndash230
Knobel DL Cleaveland S Coleman PG Fevre EM Meltzer MIMiranda MEG Shaw A Zinsstag J amp Meslin F-X (2005) Re-evaluatingthe burden of rabies in Africa and Asia Bulletin of the World HealthOrganization 83 360ndash368
Lembo T Haydon DT Velasco-Villa A Rupprecht CE Packer CBrandatildeo PE Kuzmin IV Fooks AR Barrat J amp Cleaveland S (2007)Molecular epidemiology identifies only a single rabies virus variantcirculating in complex carnivore communities of the Serengeti Proceedingsof the Royal Society B Biological Sciences 274 2123ndash2130
Maas B (1993) Behavioural Ecology and Social Organisation of the Bat-earedFox in the Serengeti National Park Tanzania University of CambridgeCambridge UK
National Bureau of Statistics Tanzania (2005) 2002 Population and HousingCensus Presidentrsquos Office Planning and Privatization Dar es SalaamTanzania
Nel JAJ (1993) The bat-eared fox-a prime candidate for rabies vectorOnderstepoort Journal of Veterinary Research 60 395ndash397
Nel LH Sabeta CT von Teichman B Jaftha JB Rupprecht CE amp
Bingham J (2005) Mongoose rabies in southern Africa a re-evaluationbased on molecular epidemiology Virus Research 109 165ndash173
Page RDM (1996) treeview an application to display phylogenetic trees onpersonal computers Computer Applications in the Biosciences 12 357ndash358
Plowright W (1982) The effects of rinderpest and rinderpest control on wildlifein Africa Symposia of the Zoological Society of London 50 1ndash28
Posada D amp Crandall KA (1998) modeltest testing the model of DNAsubstitution Bioinformatics 14 817ndash818
Randall DA Marino J Haydon DT Sillero-Zubiri C Knobel DLTallents LA Macdonald DW amp Laurenson MK (2006) An integrateddisease management strategy for the control of rabies in Ethiopian wolvesBiological Conservation 131 151ndash162
Randall DA Williams SD Kuzmin IV Rupprecht CE Tallents LATefera Z Argaw K Shiferaw F Knobel DL Sillero-Zubiri C ampLaurenson MK (2004) Rabies in endangered Ethiopian wolves EmergingInfectious Diseases 10 2214ndash2217
Rhodes CJ Atkinson RPD Anderson RM amp Macdonald DW (1998)Rabies in Zimbabwe reservoir dogs and the implications for disease controlPhilosophical Transactions of the Royal Society B Biological Sciences 353999ndash1010
Roelke-Parker ME Munson L Packer C Kock R Cleaveland SCarpenter M Obrien SJ Pospischil A HofmannLehmann R Lutz HMwamengele GLM Mgasa MN Machange GA Summers BA ampAppel MJG (1996) A canine distemper virus epidemic in Serengeti lions(Panthera leo) Nature 379 441ndash445
Ronquist F amp Huelsenbeck JP (2003) mrbayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 19 1572ndash1574
Rupprecht CE Dietzschold B Wunner WH amp Koprowsky H (1991)Antigenic relationships of lyssaviruses The Natural History of Rabies (edsGM Baer) pp 69ndash100 CRC Press Boca Raton
Sabeta CT Bingham J amp Nel LH (2003) Molecular epidemiology of canidrabies in Zimbabwe and South Africa Virus Research 91 203ndash211
Sinclair ARE (1979) The eruption of the ruminants Serengeti Dynamics ofan Ecosystem (eds ARE Sinclair amp M Norton-Griffiths) pp 82ndash103University of Chicago Press Chicago
Sinclair ARE amp Arcese P (1995) Serengeti II Dynamics Management andConservation of an Ecosystem University of Chicago Press Chicago
Smith JS (1989) Rabies virus epitopic variation use in ecological studiesAdvances in Virus Research 36 215ndash253
Smith JS Orciari LA amp Yager PA (1995) Molecular epidemiology ofrabies in the United States Seminars in Virology 6 387ndash400
Swanepoel R Barnard BJH Meredith CD Bishop GC BrucknerGK Foggin CM amp Hubschle OJB (1993) Rabies in Southern AfricaOnderstepoort Journal of Veterinary Research 60 325ndash346
Taylor LH Latham SM amp Woolhouse MEJ (2001) Risk factors forhuman disease emergence Philosophical Transactions of the Royal SocietyB Biological Sciences 356 983ndash989
Tepsumethanon V Wilde H amp Meslin FX (2005) Six criteria for rabiesdiagnosis in living dogs Journal of the Medical Association of Thailand 88419ndash422
Thomson GR amp Meredith CD (1993) Rabies in bat-eared foxes in SouthAfrica Onderstepoort Journal of Veterinary Research 60 399ndash403
Vial F Cleaveland S Rasmussen G amp Haydon DT (2006) Development ofvaccination strategies for the management of rabies in African wild dogsBiological Conservation 131 180ndash192
Wandeler AI Budde A Capt S Kappeler A amp Matter H (1988) Dogecology and dog rabies control Reviews of Infectious Diseases 10 S684ndashS688
Wandeler AI Nadin-Davis SA Tinline RR amp Rupprecht CE (1994)Rabies epidemiology some ecological and evolutionary perspectivesLyssaviruses (eds CE Rupprecht B Dietzschold amp H Koprowski)pp 297ndash324 Springer-Verlag Berlin
Woodroffe R (2001) Assessing the risks of intervention immobilizationradio-collaring and vaccination of African wild dogs Oryx 35 234ndash244
Woodroffe R Donnelly CA Johnston WT Bourne FJ Cheeseman CLClifton-Hadley RS Cox DR Gettinby G Hewinson RG Le FevreAM McInerney JP amp Morrison WI (2005) Spatial association ofMycobacterium bovis infection in cattle and badgers Meles meles Journal ofApplied Ecology 42 852ndash862
World Health Organization (1999) World Survey of Rabies No 34 for the Year1998 Geneva Switzerland
Received 14 June 2007 accepted 23 January 2008Handling Editor Simon Thirgood
12 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Supplementary material
The following supplementary material is available for thisarticle
Appendix S1 Latin names of wildlife species
Appendix S2 Supplementary details of methods
Table S1 Virus samples from the study area compared inphylogenetic analysis
Table S2 Details of all wildlife confirmed and suspect casesin SNP
Table S3 Details of confirmed wildlife cases in Serengeti andNgorongoro districts
Fig S1 Incidence of bites by domestic dogs in Serengeti andNgorongoro districts
This material is available as part of the online article fromhttpwwwblackwell-synergycomdoifull101111j1365-2664200801468x(This link will take you to the article abstract)
Please note Blackwell Publishing is not responsible for the con-tent or functionality of any supplementary materials suppliedby the authors Any queries (other than missing material)should be directed to the corresponding author for the article

10 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
alternative hosts sometimes results in short-lived chains oftransmission as contact is more common among conspecifics(Dobson 2004)
Definitive identification of infection reservoirs is extremelychallenging and rarely possible Despite intensive effortunderstanding of reservoir dynamics in many complexmulti-host systems remains incomplete (Woodroffe et al2005 Biek et al 2006 Independent Scientific Group on CattleTB 2007) Here the balance of evidence implicates domesticdogs as the only maintenance population of rabies in theSerengeti whereas other carnivores are nonessential tomaintenance However because they transmit disease totarget populations (and frequent transmission within andbetween such species may occur) they do constitute part ofthe reservoir (Table 4 Fig 1a) A single maintenance-hostspecies is consistent with broad patterns of rabies infectionobserved elsewhere (Smith 1989 Rupprecht et al 1991Wandeler et al 1994 Smith Orciari amp Yager 1995)
A lesson from this study is that even with considerable datafrom various sources we cannot exhaustively quantify thecontribution of transmission from all non-essential reservoircomponents However unravelling every thread of reservoirdynamics may not be essential Our key finding that wildlifeare not able to maintain rabies cycles or distinct geneticvariants provides new and critical information allowing rabiescontrol measures in the Serengeti to be targeted moreconfidently at domestic dog populations alone The long-term implication is that control of domestic dog rabies in theSerengeti will eliminate rabies in all other species includinghumans livestock and wildlife The absence of wildlifemaintenance hosts even in an area with such abundantwildlife raises the expectation that canine rabies eliminationis a feasible objective throughout much of Africa
Interventions to prevent transmission from domestic dogmaintenance populations would also provide convincingevidence with which to identify reservoirs Such approacheshave demonstrated that cattle were the reservoir for rinderpestafter cattle vaccinations rinderpest disappeared from wildlifeand the wildebeest Connochaetes taurinus population increasednearly ten-fold (Sinclair 1979 Plowright 1982 Dobson 1995)We are currently ring-vaccinating domestic dogs around SNPwhich should provide critical data to evaluate the impact ofalternative hosts on long-term control efforts in particular theireffects on the level of vaccination required for disease control
In conclusion resolving reservoir dynamics in any complexsystem is likely to prove challenging and our experiences inthe Serengeti suggest that consideration needs to be given toidentifying (i) the potential role of different componentsof the system (ii) the key information needed for differentdisease control strategies (iii) the epidemiological approacheslikely to yield most useful information and (iv) the range ofanalytical tools available
Acknowledgements
We are grateful to Tanzania Government ministries TANAPA TAWIRINCA Authority Tanzania Commission for Science and Technology andNational Institute for Medical Research for permissions and TANAPA and
TAWIRI Veterinary Units Viral Transmission Dynamics Project SerengetiLion and Cheetah Projects Frankfurt Zoological Society livestock field-officersof the Ministry of Water and Livestock Development in Mara and ArushaRegions M Magoto P Tiringa M Roelke-Parker R Fyumagwa H Wiickand Mwanza and Arusha Veterinary Investigation Centres for assistance withsample collection We are indebted to J Barrat AFSSA for performingdiagnostic testing and virus isolation and the staff of the Rabies Section of theCDC especially CE Rupprecht A Velasco-Villa LA Orciari IV Kuzminand M Niezgoda and PE Brandatildeo (Universidade de Satildeo Paulo) for technicalassistance with diagnostic tests and virus characterization We thankKLaurenson D Bennett J Pulliam and H Horn for valuable discussions
This work was supported by the joint National Institutes of Health (NIH)National Science Foundation (NSF) Ecology of Infectious Diseases Programunder grant no NSFDEB0225453 Opinions findings conclusions andrecommendations are those of the authors and do not necessarily reflectthe views of NIH or NSF KH was supported by NSF grant DEB-0513994the Pew Charitable Trusts award 2000-002558 to Princeton University theDepartment of Ecology and Evolution Princeton University and the HeinzFoundation TL was supported by the Royal (Dick) School of VeterinaryStudies University of Edinburgh and the University of Edinburgh DevelopmentTrust (visits to CDC) and SC by the Wellcome Trust and the Department forInternational Development Animal Health Programme
References
Barrat J (1996) Simple technique for the collection and shipment of brainspecimens for rabies diagnosis Laboratory Techniques in Rabies (edsF-X Meslin MM Kaplan amp H Koprowski) pp 425ndash432 World HealthOrganization Geneva Switzerland
Barrat J Barrat MJ Picard M amp Aubert MFA (1988) Diagnosis of rabiesby cell-culture Comparative Immunology Microbiology and InfectiousDiseases 11 207ndash214
Bartlett MS (1960) Stochastic Population Models in Ecology and EpidemiologyMethuen London
Bernardi F Nadin-Davis SA Wandeler AI Armstrong J Gomes AABLima FS Nogueira FRB amp Ito FH (2005) Antigenic and geneticcharacterization of rabies viruses isolated from domestic and wild animalsof Brazil identifies the hoary fox as a rabies reservoir Journal of GeneralVirology 86 3153ndash3162
Biek R Walsh PD Leroy EM amp Real LA (2006) Recent common ancestryof Ebola Zaire virus found in a bat reservoir Public Library of SciencePathogens 2 885ndash886
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999a) Theepidemiology of rabies in Zimbabwe 2 Rabies in jackals (Canis adustus andCanis mesomelas) Onderstepoort Journal of Veterinary Research 66 11ndash23
Bingham J Foggin CM Wandeler AI amp Hill FWG (1999b) Theepidemiology of rabies in Zimbabwe 1 Rabies in dogs (Canis familiaris)Onderstepoort Journal of Veterinary Research 66 1ndash10
Bingham J Javangwe S Sabeta CT Wandeler AI amp Nel LH (2001)Report of isolations of unusual lyssaviruses (rabies and Mokola virus)identified retrospectively from Zimbabwe Journal of the South AfricanVeterinary Association 72 92ndash94
Brooks R (1990) Survey of the dog-population of Zimbabwe and its level ofrabies vaccination Veterinary Record 127 592ndash596
Cleaveland S amp Dye C (1995) Maintenance of a microparasite infectingseveral host species rabies in the Serengeti Parasitology 111 S33ndashS47
Cleaveland S Kaare M Tiringa P Mlengeya T amp Barrat J (2003) A dograbies vaccination campaign in rural Africa impact on the incidence of dograbies and human dog-bite injuries Vaccine 21 1965ndash1973
Cleaveland S Laurenson MK amp Taylor LH (2001) Diseases of humansand their domestic mammals pathogen characteristics host range andthe risk of emergence Philosophical Transactions of the Royal Society BBiological Sciences 356 991ndash999
Cleaveland S Mlengeya T Kaare M Haydon DT Lembo T LaurensonMK amp Packer C (2007) The conservation relevance of epidemiologicalresearch into carnivore viral diseases in the Serengeti Conservation Biology21 612ndash622
Conzelmann KK Cox JH Schneider LG amp Thiel HJ (1990) Molecularcloning and complete nucleotide sequence of the attenuated rabies virusSAD B19 Virology 175 485ndash499
Daszak P Cunningham AA amp Hyatt AD (2000) Wildlife ecology ndash Emerginginfectious diseases of wildlife ndash Threats to Biodiversity and human healthScience 287 443ndash449
Dean DJ Abelseth MK amp Atanasiu P (1996) The fluorescent antibodytest Laboratory Techniques in Rabies (eds F-X Meslin MM Kaplan
Rabies reservoir dynamics in the Serengeti 11
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
amp H Koprowski) pp 88ndash95 World Health Organization GenevaSwitzerland
Dobson A (1995) The ecology and epidemiology of rinderpest virus inSerengeti and Ngorongoro Conservation Area Serengeti II DynamicsManagement and Conservation of an Ecosystem (eds ARE Sinclair ampP Arcese) pp 485ndash505 University of Chicago Press Chicago
Dobson A (2004) Population dynamics of pathogens with multiple host speciesAmerican Naturalist 164 S64ndashS78
Dobson A amp Foufopoulos J (2001) Emerging infectious pathogens of wildlifePhilosophical Transactions of the Royal Society B Biological Sciences 3561001ndash1012
Dye C (1996) Serengeti wild dogs what really happened Trends in Ecology ampEvolution 11 188ndash189
East ML Hofer H Cox JH Wulle U Wiik H amp Pitra C (2001) Regularexposure to rabies virus and lack of symptomatic disease in Serengetispotted hyenas Proceedings of the National Academy of Sciences USA 9815026ndash15031
Foggin CM (1988) Rabies and Rabies-related Viruses in Zimbabwe HistoricalVirological and Ecological Aspects PhD thesis University of ZimbabweHarare Zimbabwe
Gascoyne SC King AA Laurenson MK Borner M Schildger B ampBarrat J (1993) Aspects of rabies infection and control in the conservationof the African wild dog (Lycaon pictus) in the Serengeti region TanzaniaOnderstepoort Journal of Veterinary Research 60 415ndash420
Gordon ER Curns AT Krebs JW Rupprecht CE Real LA ampChilds JE (2004) Temporal dynamics of rabies in a wildlife host and the riskof cross-species transmission Epidemiology and Infection 132 515ndash524
Guerra MA Curns AT Rupprecht CE Hanlon CA Krebs JW ampChilds JE (2003) Skunk and raccoon rabies in the eastern United Statestemporal and spatial analysis Emerging Infectious Diseases 9 1143ndash1150
Hall TA (1999) bioedit a user-friendly biological sequence alignment editorand analysis program for Windows 9598NT Nucleic Acids SymposiumSeries 41 95ndash98
Haydon DT Cleaveland S Taylor LH amp Laurenson MK (2002) Identi-fying reservoirs of infection a conceptual and practical challenge EmergingInfectious Diseases 8 1468ndash1473
Independent Scientific Group on Cattle TB (2007) Bovine TB The ScientificEvidence Defra Publications London PB12634 httpwwwdefragovukanimalhtbisgindexhtm
Jeanmougin F Thompson JD Gouy M Higgins DG amp Gibson TJ(1998) Multiple sequence alignment with clustal-x Trends in BiochemicalSciences 23 403ndash405
Johnson N Black C Smith J Un H McElhinney LM Aylan O ampFooks AR (2003) Rabies emergence among foxes in Turkey Journal ofWildlife Diseases 39 262ndash270
King AA Meredith CD amp Thomson GR (1993) Canid and viverrid rabiesviruses in South Africa Onderstepoort Journal of Veterinary Research 60295ndash299
Kissi B Tordo N amp Bourhy H (1995) Genetic polymorphism in the rabiesvirus nucleoprotein gene Virology 209 526ndash537
Kitala PM McDermott JJ Kyule MN amp Gathuma JM (2000) Community-based active surveillance for rabies in Machakos District Kenya PreventiveVeterinary Medicine 44 73ndash85
Kitala PM McDermott J Kyule M Gathuma JM Perry B amp Wandeler A(2001) Dog ecology and demography information to support the planning ofrabies control in Machakos District Kenya Acta Tropica 78 217ndash230
Knobel DL Cleaveland S Coleman PG Fevre EM Meltzer MIMiranda MEG Shaw A Zinsstag J amp Meslin F-X (2005) Re-evaluatingthe burden of rabies in Africa and Asia Bulletin of the World HealthOrganization 83 360ndash368
Lembo T Haydon DT Velasco-Villa A Rupprecht CE Packer CBrandatildeo PE Kuzmin IV Fooks AR Barrat J amp Cleaveland S (2007)Molecular epidemiology identifies only a single rabies virus variantcirculating in complex carnivore communities of the Serengeti Proceedingsof the Royal Society B Biological Sciences 274 2123ndash2130
Maas B (1993) Behavioural Ecology and Social Organisation of the Bat-earedFox in the Serengeti National Park Tanzania University of CambridgeCambridge UK
National Bureau of Statistics Tanzania (2005) 2002 Population and HousingCensus Presidentrsquos Office Planning and Privatization Dar es SalaamTanzania
Nel JAJ (1993) The bat-eared fox-a prime candidate for rabies vectorOnderstepoort Journal of Veterinary Research 60 395ndash397
Nel LH Sabeta CT von Teichman B Jaftha JB Rupprecht CE amp
Bingham J (2005) Mongoose rabies in southern Africa a re-evaluationbased on molecular epidemiology Virus Research 109 165ndash173
Page RDM (1996) treeview an application to display phylogenetic trees onpersonal computers Computer Applications in the Biosciences 12 357ndash358
Plowright W (1982) The effects of rinderpest and rinderpest control on wildlifein Africa Symposia of the Zoological Society of London 50 1ndash28
Posada D amp Crandall KA (1998) modeltest testing the model of DNAsubstitution Bioinformatics 14 817ndash818
Randall DA Marino J Haydon DT Sillero-Zubiri C Knobel DLTallents LA Macdonald DW amp Laurenson MK (2006) An integrateddisease management strategy for the control of rabies in Ethiopian wolvesBiological Conservation 131 151ndash162
Randall DA Williams SD Kuzmin IV Rupprecht CE Tallents LATefera Z Argaw K Shiferaw F Knobel DL Sillero-Zubiri C ampLaurenson MK (2004) Rabies in endangered Ethiopian wolves EmergingInfectious Diseases 10 2214ndash2217
Rhodes CJ Atkinson RPD Anderson RM amp Macdonald DW (1998)Rabies in Zimbabwe reservoir dogs and the implications for disease controlPhilosophical Transactions of the Royal Society B Biological Sciences 353999ndash1010
Roelke-Parker ME Munson L Packer C Kock R Cleaveland SCarpenter M Obrien SJ Pospischil A HofmannLehmann R Lutz HMwamengele GLM Mgasa MN Machange GA Summers BA ampAppel MJG (1996) A canine distemper virus epidemic in Serengeti lions(Panthera leo) Nature 379 441ndash445
Ronquist F amp Huelsenbeck JP (2003) mrbayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 19 1572ndash1574
Rupprecht CE Dietzschold B Wunner WH amp Koprowsky H (1991)Antigenic relationships of lyssaviruses The Natural History of Rabies (edsGM Baer) pp 69ndash100 CRC Press Boca Raton
Sabeta CT Bingham J amp Nel LH (2003) Molecular epidemiology of canidrabies in Zimbabwe and South Africa Virus Research 91 203ndash211
Sinclair ARE (1979) The eruption of the ruminants Serengeti Dynamics ofan Ecosystem (eds ARE Sinclair amp M Norton-Griffiths) pp 82ndash103University of Chicago Press Chicago
Sinclair ARE amp Arcese P (1995) Serengeti II Dynamics Management andConservation of an Ecosystem University of Chicago Press Chicago
Smith JS (1989) Rabies virus epitopic variation use in ecological studiesAdvances in Virus Research 36 215ndash253
Smith JS Orciari LA amp Yager PA (1995) Molecular epidemiology ofrabies in the United States Seminars in Virology 6 387ndash400
Swanepoel R Barnard BJH Meredith CD Bishop GC BrucknerGK Foggin CM amp Hubschle OJB (1993) Rabies in Southern AfricaOnderstepoort Journal of Veterinary Research 60 325ndash346
Taylor LH Latham SM amp Woolhouse MEJ (2001) Risk factors forhuman disease emergence Philosophical Transactions of the Royal SocietyB Biological Sciences 356 983ndash989
Tepsumethanon V Wilde H amp Meslin FX (2005) Six criteria for rabiesdiagnosis in living dogs Journal of the Medical Association of Thailand 88419ndash422
Thomson GR amp Meredith CD (1993) Rabies in bat-eared foxes in SouthAfrica Onderstepoort Journal of Veterinary Research 60 399ndash403
Vial F Cleaveland S Rasmussen G amp Haydon DT (2006) Development ofvaccination strategies for the management of rabies in African wild dogsBiological Conservation 131 180ndash192
Wandeler AI Budde A Capt S Kappeler A amp Matter H (1988) Dogecology and dog rabies control Reviews of Infectious Diseases 10 S684ndashS688
Wandeler AI Nadin-Davis SA Tinline RR amp Rupprecht CE (1994)Rabies epidemiology some ecological and evolutionary perspectivesLyssaviruses (eds CE Rupprecht B Dietzschold amp H Koprowski)pp 297ndash324 Springer-Verlag Berlin
Woodroffe R (2001) Assessing the risks of intervention immobilizationradio-collaring and vaccination of African wild dogs Oryx 35 234ndash244
Woodroffe R Donnelly CA Johnston WT Bourne FJ Cheeseman CLClifton-Hadley RS Cox DR Gettinby G Hewinson RG Le FevreAM McInerney JP amp Morrison WI (2005) Spatial association ofMycobacterium bovis infection in cattle and badgers Meles meles Journal ofApplied Ecology 42 852ndash862
World Health Organization (1999) World Survey of Rabies No 34 for the Year1998 Geneva Switzerland
Received 14 June 2007 accepted 23 January 2008Handling Editor Simon Thirgood
12 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Supplementary material
The following supplementary material is available for thisarticle
Appendix S1 Latin names of wildlife species
Appendix S2 Supplementary details of methods
Table S1 Virus samples from the study area compared inphylogenetic analysis
Table S2 Details of all wildlife confirmed and suspect casesin SNP
Table S3 Details of confirmed wildlife cases in Serengeti andNgorongoro districts
Fig S1 Incidence of bites by domestic dogs in Serengeti andNgorongoro districts
This material is available as part of the online article fromhttpwwwblackwell-synergycomdoifull101111j1365-2664200801468x(This link will take you to the article abstract)
Please note Blackwell Publishing is not responsible for the con-tent or functionality of any supplementary materials suppliedby the authors Any queries (other than missing material)should be directed to the corresponding author for the article

Rabies reservoir dynamics in the Serengeti 11
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
amp H Koprowski) pp 88ndash95 World Health Organization GenevaSwitzerland
Dobson A (1995) The ecology and epidemiology of rinderpest virus inSerengeti and Ngorongoro Conservation Area Serengeti II DynamicsManagement and Conservation of an Ecosystem (eds ARE Sinclair ampP Arcese) pp 485ndash505 University of Chicago Press Chicago
Dobson A (2004) Population dynamics of pathogens with multiple host speciesAmerican Naturalist 164 S64ndashS78
Dobson A amp Foufopoulos J (2001) Emerging infectious pathogens of wildlifePhilosophical Transactions of the Royal Society B Biological Sciences 3561001ndash1012
Dye C (1996) Serengeti wild dogs what really happened Trends in Ecology ampEvolution 11 188ndash189
East ML Hofer H Cox JH Wulle U Wiik H amp Pitra C (2001) Regularexposure to rabies virus and lack of symptomatic disease in Serengetispotted hyenas Proceedings of the National Academy of Sciences USA 9815026ndash15031
Foggin CM (1988) Rabies and Rabies-related Viruses in Zimbabwe HistoricalVirological and Ecological Aspects PhD thesis University of ZimbabweHarare Zimbabwe
Gascoyne SC King AA Laurenson MK Borner M Schildger B ampBarrat J (1993) Aspects of rabies infection and control in the conservationof the African wild dog (Lycaon pictus) in the Serengeti region TanzaniaOnderstepoort Journal of Veterinary Research 60 415ndash420
Gordon ER Curns AT Krebs JW Rupprecht CE Real LA ampChilds JE (2004) Temporal dynamics of rabies in a wildlife host and the riskof cross-species transmission Epidemiology and Infection 132 515ndash524
Guerra MA Curns AT Rupprecht CE Hanlon CA Krebs JW ampChilds JE (2003) Skunk and raccoon rabies in the eastern United Statestemporal and spatial analysis Emerging Infectious Diseases 9 1143ndash1150
Hall TA (1999) bioedit a user-friendly biological sequence alignment editorand analysis program for Windows 9598NT Nucleic Acids SymposiumSeries 41 95ndash98
Haydon DT Cleaveland S Taylor LH amp Laurenson MK (2002) Identi-fying reservoirs of infection a conceptual and practical challenge EmergingInfectious Diseases 8 1468ndash1473
Independent Scientific Group on Cattle TB (2007) Bovine TB The ScientificEvidence Defra Publications London PB12634 httpwwwdefragovukanimalhtbisgindexhtm
Jeanmougin F Thompson JD Gouy M Higgins DG amp Gibson TJ(1998) Multiple sequence alignment with clustal-x Trends in BiochemicalSciences 23 403ndash405
Johnson N Black C Smith J Un H McElhinney LM Aylan O ampFooks AR (2003) Rabies emergence among foxes in Turkey Journal ofWildlife Diseases 39 262ndash270
King AA Meredith CD amp Thomson GR (1993) Canid and viverrid rabiesviruses in South Africa Onderstepoort Journal of Veterinary Research 60295ndash299
Kissi B Tordo N amp Bourhy H (1995) Genetic polymorphism in the rabiesvirus nucleoprotein gene Virology 209 526ndash537
Kitala PM McDermott JJ Kyule MN amp Gathuma JM (2000) Community-based active surveillance for rabies in Machakos District Kenya PreventiveVeterinary Medicine 44 73ndash85
Kitala PM McDermott J Kyule M Gathuma JM Perry B amp Wandeler A(2001) Dog ecology and demography information to support the planning ofrabies control in Machakos District Kenya Acta Tropica 78 217ndash230
Knobel DL Cleaveland S Coleman PG Fevre EM Meltzer MIMiranda MEG Shaw A Zinsstag J amp Meslin F-X (2005) Re-evaluatingthe burden of rabies in Africa and Asia Bulletin of the World HealthOrganization 83 360ndash368
Lembo T Haydon DT Velasco-Villa A Rupprecht CE Packer CBrandatildeo PE Kuzmin IV Fooks AR Barrat J amp Cleaveland S (2007)Molecular epidemiology identifies only a single rabies virus variantcirculating in complex carnivore communities of the Serengeti Proceedingsof the Royal Society B Biological Sciences 274 2123ndash2130
Maas B (1993) Behavioural Ecology and Social Organisation of the Bat-earedFox in the Serengeti National Park Tanzania University of CambridgeCambridge UK
National Bureau of Statistics Tanzania (2005) 2002 Population and HousingCensus Presidentrsquos Office Planning and Privatization Dar es SalaamTanzania
Nel JAJ (1993) The bat-eared fox-a prime candidate for rabies vectorOnderstepoort Journal of Veterinary Research 60 395ndash397
Nel LH Sabeta CT von Teichman B Jaftha JB Rupprecht CE amp
Bingham J (2005) Mongoose rabies in southern Africa a re-evaluationbased on molecular epidemiology Virus Research 109 165ndash173
Page RDM (1996) treeview an application to display phylogenetic trees onpersonal computers Computer Applications in the Biosciences 12 357ndash358
Plowright W (1982) The effects of rinderpest and rinderpest control on wildlifein Africa Symposia of the Zoological Society of London 50 1ndash28
Posada D amp Crandall KA (1998) modeltest testing the model of DNAsubstitution Bioinformatics 14 817ndash818
Randall DA Marino J Haydon DT Sillero-Zubiri C Knobel DLTallents LA Macdonald DW amp Laurenson MK (2006) An integrateddisease management strategy for the control of rabies in Ethiopian wolvesBiological Conservation 131 151ndash162
Randall DA Williams SD Kuzmin IV Rupprecht CE Tallents LATefera Z Argaw K Shiferaw F Knobel DL Sillero-Zubiri C ampLaurenson MK (2004) Rabies in endangered Ethiopian wolves EmergingInfectious Diseases 10 2214ndash2217
Rhodes CJ Atkinson RPD Anderson RM amp Macdonald DW (1998)Rabies in Zimbabwe reservoir dogs and the implications for disease controlPhilosophical Transactions of the Royal Society B Biological Sciences 353999ndash1010
Roelke-Parker ME Munson L Packer C Kock R Cleaveland SCarpenter M Obrien SJ Pospischil A HofmannLehmann R Lutz HMwamengele GLM Mgasa MN Machange GA Summers BA ampAppel MJG (1996) A canine distemper virus epidemic in Serengeti lions(Panthera leo) Nature 379 441ndash445
Ronquist F amp Huelsenbeck JP (2003) mrbayes 3 Bayesian phylogeneticinference under mixed models Bioinformatics 19 1572ndash1574
Rupprecht CE Dietzschold B Wunner WH amp Koprowsky H (1991)Antigenic relationships of lyssaviruses The Natural History of Rabies (edsGM Baer) pp 69ndash100 CRC Press Boca Raton
Sabeta CT Bingham J amp Nel LH (2003) Molecular epidemiology of canidrabies in Zimbabwe and South Africa Virus Research 91 203ndash211
Sinclair ARE (1979) The eruption of the ruminants Serengeti Dynamics ofan Ecosystem (eds ARE Sinclair amp M Norton-Griffiths) pp 82ndash103University of Chicago Press Chicago
Sinclair ARE amp Arcese P (1995) Serengeti II Dynamics Management andConservation of an Ecosystem University of Chicago Press Chicago
Smith JS (1989) Rabies virus epitopic variation use in ecological studiesAdvances in Virus Research 36 215ndash253
Smith JS Orciari LA amp Yager PA (1995) Molecular epidemiology ofrabies in the United States Seminars in Virology 6 387ndash400
Swanepoel R Barnard BJH Meredith CD Bishop GC BrucknerGK Foggin CM amp Hubschle OJB (1993) Rabies in Southern AfricaOnderstepoort Journal of Veterinary Research 60 325ndash346
Taylor LH Latham SM amp Woolhouse MEJ (2001) Risk factors forhuman disease emergence Philosophical Transactions of the Royal SocietyB Biological Sciences 356 983ndash989
Tepsumethanon V Wilde H amp Meslin FX (2005) Six criteria for rabiesdiagnosis in living dogs Journal of the Medical Association of Thailand 88419ndash422
Thomson GR amp Meredith CD (1993) Rabies in bat-eared foxes in SouthAfrica Onderstepoort Journal of Veterinary Research 60 399ndash403
Vial F Cleaveland S Rasmussen G amp Haydon DT (2006) Development ofvaccination strategies for the management of rabies in African wild dogsBiological Conservation 131 180ndash192
Wandeler AI Budde A Capt S Kappeler A amp Matter H (1988) Dogecology and dog rabies control Reviews of Infectious Diseases 10 S684ndashS688
Wandeler AI Nadin-Davis SA Tinline RR amp Rupprecht CE (1994)Rabies epidemiology some ecological and evolutionary perspectivesLyssaviruses (eds CE Rupprecht B Dietzschold amp H Koprowski)pp 297ndash324 Springer-Verlag Berlin
Woodroffe R (2001) Assessing the risks of intervention immobilizationradio-collaring and vaccination of African wild dogs Oryx 35 234ndash244
Woodroffe R Donnelly CA Johnston WT Bourne FJ Cheeseman CLClifton-Hadley RS Cox DR Gettinby G Hewinson RG Le FevreAM McInerney JP amp Morrison WI (2005) Spatial association ofMycobacterium bovis infection in cattle and badgers Meles meles Journal ofApplied Ecology 42 852ndash862
World Health Organization (1999) World Survey of Rabies No 34 for the Year1998 Geneva Switzerland
Received 14 June 2007 accepted 23 January 2008Handling Editor Simon Thirgood
12 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Supplementary material
The following supplementary material is available for thisarticle
Appendix S1 Latin names of wildlife species
Appendix S2 Supplementary details of methods
Table S1 Virus samples from the study area compared inphylogenetic analysis
Table S2 Details of all wildlife confirmed and suspect casesin SNP
Table S3 Details of confirmed wildlife cases in Serengeti andNgorongoro districts
Fig S1 Incidence of bites by domestic dogs in Serengeti andNgorongoro districts
This material is available as part of the online article fromhttpwwwblackwell-synergycomdoifull101111j1365-2664200801468x(This link will take you to the article abstract)
Please note Blackwell Publishing is not responsible for the con-tent or functionality of any supplementary materials suppliedby the authors Any queries (other than missing material)should be directed to the corresponding author for the article

12 T Lembo et al
copy 2008 The Authors Journal compilation copy 2008 British Ecological Society Journal of Applied Ecology
Supplementary material
The following supplementary material is available for thisarticle
Appendix S1 Latin names of wildlife species
Appendix S2 Supplementary details of methods
Table S1 Virus samples from the study area compared inphylogenetic analysis
Table S2 Details of all wildlife confirmed and suspect casesin SNP
Table S3 Details of confirmed wildlife cases in Serengeti andNgorongoro districts
Fig S1 Incidence of bites by domestic dogs in Serengeti andNgorongoro districts
This material is available as part of the online article fromhttpwwwblackwell-synergycomdoifull101111j1365-2664200801468x(This link will take you to the article abstract)
Please note Blackwell Publishing is not responsible for the con-tent or functionality of any supplementary materials suppliedby the authors Any queries (other than missing material)should be directed to the corresponding author for the article