Embed Size (px)
Citation preview

Using c-FOS Expression to Investigate Rapid Testosterone Effects on Neuronal Activity in Carassius auratus Preoptic Areas and Optic Tecta
Jordan Francke, 2013
It has long been thought that steroids are slow-acting molecules used to create circuits, and that they don’t have dramatic influences on rapid and dynamic processes (Steinman, 2010). It was believed that a hormone binds to receptors in the cytoplasm, and then the hormone/receptor complex migrates to the nucleus to alter gene expression (mRNA). However, a more recent, albeit widely-accepted, theory of steroid modulation is that hormones also have the capability of acting in a non-genomic, rapid fashion (Steinman, 2010). This is assumed to occur by hormones binding to receptors in the cell membrane. These receptors then activate other cellular signaling pathways (second messengers) that can lead to rapid changes in behavior. Thus, sex steroids affecting cell physiology and brain function rapidly is now accepted, but the behavioral consequences of such actions, and the social contexts in which they occur have thus far only been demonstrated in a few species (Dewing et al, 2007). The fact that sex steroids can rapidly affect cell physiology and brain function is now widely accepted. However, the behavioral consequences of such actions and the social contexts in which they occur have thus far only been demonstrated in a few species. The Thompson lab uses the Carassius auratus goldfish to explore the behavioral effects of nongenomic testerone action in social and sexual communication contexts.
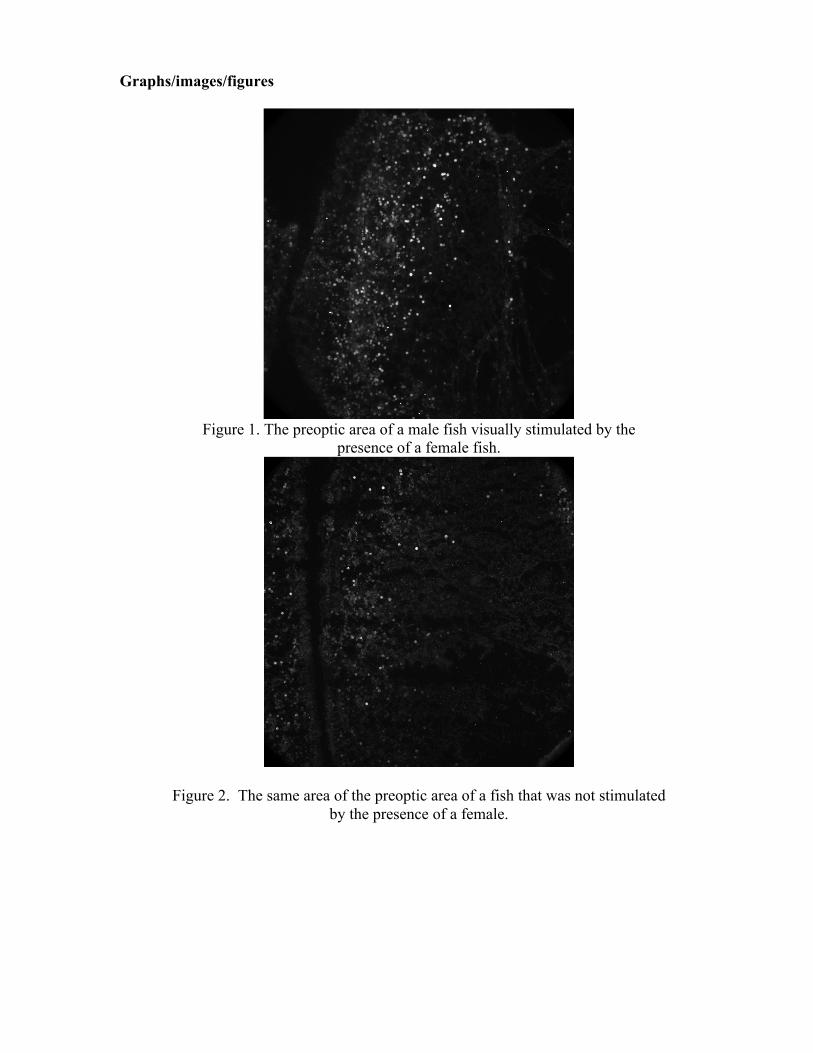
An enzyme known as aromatase is known for converting testosterone into estradiol via a process called aromatization. Previous studies in Gloria Callard’s lab demonstrated that aromatase levels are high not only in the regions of the brain known to regulate male sexual behaviors in vertebrates, most notably the preoptic area, but also in the neuronal fibers in the retina and the optic tectum. My work this summer, which will be continuing in the fall in the form of an honors project, was interested in examining how testosterone rapidly affects (via a nongenomic pathway) visual processing in the optic tectum area, as well as understanding more about the mechanism through which testosterone rapidly modulates preoptic area activity. To accomplish this, I employed two visualization methods: immunohistochemistry and in situ hybridization. The majority of this summer was spent troubleshooting which method was most effective at visualizing a protein called c-FOS, which is a documented marker for neuronal activity (as you can’t do an MRI on a fish). The fluorescent microscopy images from the immunohistochemical procedures (seen on the second page of this document) seem to show the most significant contrast between the control and a stimulated fish’s preoptic area activity. In the fall, we will likely continue utilizing this protocol, quantify the activity, and then run statistical analyses to see if the activity differences are significant.
Graphs/images/figures
Figure 1. The preoptic area of a male fish visually stimulated by the
presence of a female fish.
Figure 2. The same area of the preoptic area of a fish that was not stimulated by the presence of a female.

Faculty Mentor: Rick Thompson Funded by the Surdna Foundation Undergraduate Research Fellowship References Cornil, C. A., et al. (2006b). Estradiol rapidly activates male sexual behavior and affects brain monoamine levels in the quail brain. Behavioral Brain Research, 166, 110 – 123. Dewing, P., et al. (2007). Membrane estrogen receptor-alpha interactions with
metabotropic glutamate receptor 1a modulate female sexual receptivity in rats. Journal of Neuroscience, 27, 9294-300.
Lord, L.D., Bond, J., Thompson R.R. (2009). Rapid steroid influences on visually- guided sexual behavior in goldfish. Hormones and Behavior, 56(5), 519-526. Remage-Healey, L., & Bass, A. H. (2005). Rapid elevations in both steroid hormones
and vocal signaling during playback challenge: a field experiment in gulf toadfish. Hormones and Behavior, 47, 297-305.
Steinman, M. Q. & Trainor, B. C. (2010) Rapid Effects of Steroid Hormones on Animal Behavior. Nature Education Knowledge 1(10):1. Thompson, R.R., Mangiamele, L.A. (2012) Rapid Effects of Estradiol on Behavioral and Physiological Responses Related to Reproduction in Male Goldfish.





























