Embed Size (px)
Citation preview

The RNA World, Second Edition © 1999 Cold Spring Harbor Laboratory Press 0-87969-561-7/99 637
24Dynamics of the Genetic Code
JJoohhnn FF.. AAttkkiinnssHoward Hughes Medical Institute and Department of Human GeneticsUniversity of UtahSalt Lake City, Utah 84112-5330;Department of GeneticsTrinity College, Dublin, Ireland
AAuugguusstt BBööcckkLehrstuhl für Mikrobiologie der Universität München80638 München, Germany
SSeennyyaa MMaattssuuffuujjiiDepartment of Biochemistry IIJikei University School of MedicineTokyo 105-8461, Japan
RRaayymmoonndd FF.. GGeesstteellaannddHoward Hughes Medical Institute and Department of Human GeneticsUniversity of UtahSalt Lake City, Utah 84112-5330
Shape is crucial for catalysis. In the hypothetical RNA World, the replica-tive RNAs, which constituted the hereditary information, also functionedas the “shapes” for catalysis. There was no need for decoding. When de-coding originated, presumably discrimination between alternate codingpossibilities was initially weak, but once one mode became predominant,there would have been selection to lock it in with increasing efficiency.Nontriplet translocation and nonstandard meaning of code words presum-ably generally approached a minimum compatible with speed and energyuse optimization. In the present day, nonstandard decoding alternativesgenerally just contribute to a low level of translational errors, of whichframeshifting errors (Atkins et al. 1972; Kurland 1992) are a grave type.However, some unknown proportion of genes in probably all organismshas special sites where efficient decoding alternatives are programmed intothe mRNA. The group of mechanisms involved in redirection of decodingis called “recoding” (Gesteland et al. 1992). Either the evolution of theability to perform recoding was coincident with evolution of the ability ofthe decoding apparatus to perform standard decoding, or it is a later so-

638 J.F. Atkins et al.
phistication. Of course, the answer is unknown, but the conservation of therequired mRNA signals presented below indicates that recoding has beenpart of the decoding repertoire for at least several hundred million yearsand must therefore be favored by evolution. This is clearly distinct fromthe error rate that is a trade-off between energy expenditure, speed, and ac-curacy. In the latter case, evolution has optimized the balance as a whole.With recoding, it is the specific, nonstandard decoding events themselvesthat have led to selective advantage. In addition to the evolutionary ques-tions posed by recoding, the mRNA signals involved reveal heretofore un-suspected roles of RNA sequences and structures in modern organisms.
Signals for recoding are carried in the mRNA sequence and includespecification of a site in the coding sequence and stimulatory elementsranging from simple sequences on either side of the site to far removedand complex stem-loop structures. Little is known about the involvementof other cellular components, although in one case (insertion of seleno-cysteine in prokaryotes), a special protein factor is known to be essential.
Three classes of recoding are recognized. The meaning of specificcode words can be redefined. The reading frame can be altered by ribo-somes that switch from one overlapping reading frame to another, andblocks of nucleotides can be bypassed with, or without, a change in read-ing frame (for review, see Gesteland and Atkins 1996).
Recoding is in competition with standard decoding. For instance, theprogrammed ribosomal frameshifting that occurs two-thirds of the waythrough the Escherichia coli dnaX coding sequence is 50% efficient. Halfthe ribosomes frameshift and soon terminate at a stop codon in the newframe, whereas the other half traverse the shift site with standard decod-ing and synthesize a product with an extra domain. The two products arepresent in a 1:1 ratio in DNA polymerase III (Blinkova and Walker 1990;Flower and McHenry 1990; Tsuchihashi and Kornberg 1990; Kelman andO’Donnell 1995). The signals in other cases of recoding give different setefficiencies, sometimes for viral packaging purposes (Wickner 1989;Atkins et al. 1990; Levin et al. 1993), or the process can be regulated. Thedynamic nature of recoding contrasts with the “hard-wired” derivatives ofthe universal code found in specialized niches such as mitochondria,where the meaning of a code word is altered wherever it occurs. In re-coding, the rule change is dependent on mRNA context.
REDEFINITION
Redefinition of stop codons by recoding involves a variety of stimulatorysignals. The first case of redefinition studied, the low-efficiency, but es-

Recoding 639
sential, readthrough of the UGA at the end of the phage Qβ coat proteingene (Weiner and Weber 1973; Hofstetter et al. 1974), is probably influ-enced only by the identity of a few flanking nucleotides. Tate and col-leagues have shown that the efficiency of release factor 2 function at UGAis influenced by the identity of the flanking 3� nucleotide and, to a lesserextent, the following two nucleotides (Poole et al. 1998). The identity ofcorresponding Qβ nucleotides is expected to give somewhat inefficienttermination. The identity of the last couple of amino acids encoded priorto the stop codon also influences termination efficiency (Mottagui-Tabarand Isaksson 1997). At the end of the Qβ coat protein gene, there appearsto be a trade-off between amino acids that give efficient termination and3� nucleotides that give moderately efficient readthrough, so that modestlevels of readthrough result.
In tobacco mosaic virus, the recoding signal is the identity of six nu-cleotides 3� of the redefined UAG “stop” codon (Skuzeski et al. 1991; Zer-fass and Beier 1992; Stahl et al. 1995). The counterpart for synthesis of theGag-Pol precursor of murine leukemia virus is a nearby 3� pseudoknot (Fig.1A) (ten Dam et al. 1990; Wills et al. 1991, 1994; Felsenstein and Goff1992; Feng et al. 1992). Although not yet studied, a pseudoknot may alsobe important for the putative gag-pro analog readthrough of the Dic-tyostelium retrotransposon, Skipper (Leng et al. 1998). For UAG stop codonredefinition in decoding barley yellow dwarf virus, both close and distantsignals are important (Brown et al. 1996). Given the widespread occurrenceof redefinition in plant viruses, understanding the mechanism involved isespecially interesting and worthwhile. For Drosophila trachea branchingcontroller, hdc, high-level readthrough of UAA is required, but the locationof the stimulatory signals is unknown (Steneberg et al. 1998). ForDrosophila kelch, the recoding signal is also unknown, but the efficiency ofredefinition is regulated (Robinson and Cooley 1997). For these cases of re-definition, the amino acid inserted is either known, or highly likely to be oneof the standard 20 amino acids having their own unique codon(s). However,the 21st (excluding formyl methionine) directly encoded amino acid, se-lenocysteine, is encoded only by UGA, a stop codon in the standard code.
Selenocysteine
Selenocysteine was recognized as a constituent of special proteins in 1976(Cone et al. 1976). Experiments in two different biological systems in1986 showed that this nonstandard amino acid is inserted cotranslation-ally, directed by an in-frame UGA codon in the mRNA (Chambers et al.1986; Zinoni et al. 1986). Thus, in a single mRNA, UGA can have two

640 J.F. Atkins et al.
contrasting meanings, stop or selenocysteine insertion. This is in contrastto the use of UGA as a codon for tryptophan in Mycoplasma and mito-chondria, where the new meaning is hard-wired and the stop function islost. UGA thus is a “chameleon” among codons, which raises intriguingquestions about whether the ancestral UGA encoded stop or an amino acidand how the evolution of the change of meaning occurred.
Figure 1 (A) Redefinition of the murine leukemia virus gag terminator requiredfor synthesis of the Gag-Pol precursor. The size of loop 1 and stem 2 shown havebeen determined in recent work by N.M. Wills et al. (unpubl.). (B) Model of theribosomal complex decoding UGA with selenocysteine. The model shows thequaternary complex between SelB, GTP, selenocysteyl-tRNA, and the SECIS element of the mRNA. Contact between SelB in the quaternary complex and theribosome triggers GTP hydrolysis resulting in the release of selenocysteyl-tRNA(Hüttenhofer and Böck 1998b).
A
B

Recoding 641
Distribution of Selenoproteins
Selenocysteine-containing proteins occur in all three lines of descent, but not in all organisms. Our own recent screening revealed that amongtwenty gram-positive and gram-negative bacteria, only five synthesizedselenoproteins (S. Schorling et al., unpubl.). This finding of a nonubiqui-tous distribution is supported by the recent results of whole genome se-quence analysis, which reveals a similar frequency. We were also unableto demonstrate the occurrence of selenoproteins in several plant cell cul-tures (Neuhierl and Böck 1996). Intriguingly, Mycoplasma species inwhich UGA codes only for tryptophan do not contain selenoproteins(Himmelreich et al. 1997). The numbers of selenoproteins synthesized aredifferent among organisms. They can range from just two in the case ofHaemophilus influenzae (Wilting et al. 1998) to three for E. coli (Baronand Böck 1995) to seven for Methanococcus jannaschi (Bult et al. 1996;Wilting et al. 1997) to an estimated more than thirty for mammals (Behneet al. 1996).
S and Se in Metabolism
Sulfur and selenium occur in the biosphere at a ratio between 103–105 to 1and, with the exception of one major branch point leading to the specificbiosynthesis of selenocysteine, they share the same metabolic paths. Freeselenocysteine formed via the cysteine biosynthetic enzymes (Müller et al.1997) can be aminoacylated onto tRNAs by cysteyl-tRNA synthetase andincorporated into any cysteine position of proteins (Müller et al. 1994).Low-molecular-weight selenocysteine is also the precursor for selenome-thionine (Sliwkowski and Stadtman 1986). For our discussion, one has tokeep in mind, therefore, that any cysteine in a protein is “contaminated” byselenocysteine at a ratio determined by the relative abundance of the twoelements and the biochemical S/Se discrimination capacity of the respec-tive organism. The major branch point mentioned above separates the fateof the two elements by a high-affinity metabolic route targeted to the effi-cient synthesis of selenocysteine under low trace element concentrations.
Selenocysteine Biosynthesis
The biosynthesis of selenocysteine in bacteria differs from that leading tocysteine as it takes place in a tRNA-bound state (Leinfelder et al. 1990;Forchhammer and Böck 1991). A specific tRNA (tRNASec) is chargedwith L-serine by seryl-tRNA synthetase, and the seryl moiety is convertedinto the selenocysteyl residue by selenocysteine synthase with se-lenomonophosphate (SeP) as selenium donor. SeP itself is the reaction

642 J.F. Atkins et al.
product of selenophosphate synthetase (Leinfelder et al. 1990; Ehrenreichet al. 1992; Veres et al. 1992). Genes coding for tRNASec have been iden-tified in many organisms within Bacteria, Archaea, and Eukarya, and theirproducts share a number of characteristics differentiating them from ordi-nary elongator tRNAs. In addition to the UCA anticodon, complementaryto UGA, they display sequence and architectural deviations from the con-sensus of classic elongator tRNAs (Baron et al. 1993; Sturchler et al.1993). They have a 6-bp D-stem with a 4-bp loop, an extended amino-acyl-acceptor-T-stem axis of 13 bp and a large extra arm which makesthese tRNAs the largest ones known. These sequence deviations have arole in maintaining novel tertiary interactions (Baron et al. 1993; Sturch-ler et al. 1993). tRNASec, therefore, may constitute a different evolution-ary line of elongator tRNAs. The structural differences are the basis of theadditional functions which tRNASec has compared to elongator tRNAs,namely, serving as an adapter for the biosynthesis of selenocysteine by se-lenocysteine synthase, binding in a selenocysteyl-specific manner to aspecialized elongation factor (see below), and precluding binding to elon-gation factor Tu (Baron and Böck 1991).
Biosynthesis of selenocysteine resembles that of glutamine and as-paragine, which also take place starting from a precursor in the tRNA-bound state (Ibba et al. 1997). In this context, it is intriguing that archaealgenomes sequenced thus far do not contain recognizable genes for essen-tial cysteine biosynthetic enzymes or for a cysteyl-tRNA synthetase. Thus,either the sequences of these enzymes in Archaea are highly diverged, orcysteine biosynthesis could occur from a specialized seryl-tRNA by anal-ogy with that of selenocysteyl-tRNA (Bult et al. 1996; Ibba et al. 1997;Smith et al. 1997). If so, this would add another example of aminoacyltransformations and fill an important gap in our knowledge of the connec-tions between evolution of the genetic code and amino acid biosynthesis(Wong 1975; Di Giulio 1997).
Elements Involved in Decoding UGA as Selenocysteine
Discrimination between UGA as the selenocysteine-specific codon andUGA as a stop codon is by an mRNA secondary/tertiary structure (the SECIS element) which, in bacteria, is located at the immediate 3� side ofUGA, i.e., within the reading frame. Swapping of the SECIS within bac-teria is restricted by specific interaction of SECIS with SelB (see below)and the ribosome (Tormay and Böck 1997; Wilting et al. 1998). In Eu-karya (Berry et al. 1991, 1993; Hill et al. 1993; Kollmus et al. 1996; Wal-czak et al. 1998) and Archaea (Wilting et al. 1997), the SECIS motif is po-sitioned outside the reading frame in the 3�untranslated region and acts at

Recoding 643
a distance of up to 2,700 bases. The SECIS motifs in bacteria are variablein sequence, based on the constraint of the reading frame, whereas thosefrom mammalia and Archaea possess highly conserved motifs (Fig. 2).Mammalian SECIS elements are predicted to fall into two classes, whichdiffer in the secondary structure around the conserved adenosines (Lowand Berry 1996). Selenoprotein P has one of each class in its 3� UTR, butno functional difference between the two is known (M. Berry, pers.comm.). The mammalian SECIS element can be transplanted and func-tions to cause selenocysteine insertion at UGA in a heterologous gene(Shen et al. 1993). Although the analysis is at an early stage, there appearsto be remarkable conservation between the SECIS element(s) in lowermetazoans and mammals. Caenorhabditis elegans thioredoxin reductaseis almost certainly a selenoprotein very similar to its mammalian counter-part. The 3� UTR of its mRNA shows a putative SECIS element that hasonly a single, nonsignificant, substitution from the highly conserved ver-
Figure 2 Consensus features of SECIS elements from Bacteria, Archaea, andEukarya as adopted from Hüttenhofer et al. (1996), Low and Berry (1996), Wilt-ing et al. (1997), Walczak et al. (1998), and M. Berry (pers. comm.). Consensusnucleotides are boxed and semi-conservative ones are printed in bold.

644 J.F. Atkins et al.
tebrate-like SECIS (M. Berry, pers. comm.). This SECIS, however, isvery different from those identified in Archaea.
SelB, first discovered in E. coli (Forchhammer et al. 1989), is a spe-cialized translation factor that interacts with guanine nucleotides, seleno-cysteyl-tRNA, and the SECIS element in mRNA forming a quaternarycomplex (Fig. 1B) (Heider et al. 1992; Hüttenhofer et al. 1996). Withinthis complex, SelB attains a structure suitable for interaction with the ri-bosome; as a consequence, GTP is hydrolyzed and selenocysteyl-tRNA isreleased in the proximity of the ribosomal A site (Hüttenhofer and Böck1998a,b). Despite considerable effort, a homolog to SelB has not yet beenidentified in Eukarya and Archaea.
The domains of translation factor SelB involved in selenocysteyl-tRNA binding and in mRNA binding can be separated. A 17-kD carboxy-terminal domain of SelB is responsible for binding to the SECIS element(Kromayer et al. 1996) and retains this property when separated from therest of the protein. The amino-terminal part—separated by a linker do-main—has considerable sequence similarity with EF-Tu, and can bind se-lenocysteyl-tRNA in vivo and in vitro. SelB thus is an elongation factorhomologous to EF-Tu that is tethered to the mRNA by its carboxy-termi-nal extension.
An intriguing consequence of the mechanism used for the “localized”decoding of the UGA codon is that the substrate—selenocysteyl-tRNA—is bound to the translation factor together with the mRNA. It will be veryinteresting to see the spatial relationship of codon and anticodon withinthis complex. Is there an interaction between the two nucleic acids beforethe ribosome even arrives at the UGA?
Evolution of Selenocysteine Insertion
Selective Advantage of Selenocysteine
Selenoproteins that have identified functions are enzymes with seleno-cysteine in their active site. Natural variants containing a cysteine in thisposition have been identified for many of these enzymes, showing thatselenocysteine per se, in most of the selenoproteins, does not possess anessential role. Mutational change of the selenocysteine to a cysteine alsogives variants that are active (Axley et al. 1991; Berry et al. 1992) buthave decreased overall catalytic efficiency by a factor of 300 to 400,mostly due to a reduction of the reaction velocity (Axley et al. 1991). It is clear from these studies that although selenocysteine confers a con-siderable catalytic advantage, it can be replaced by a cysteine in most enzymes.

Recoding 645
Did UGA Previously Code for Cysteine?
The discovery of UGA encoding selenocysteine raises the questionwhether UGA was originally a “sense codon” specifying selenocysteineincorporation or whether this is a “new” development selected to expandthe genetic code (Leinfelder et al. 1988). Osawa (1995) and Jukes (1990)in their “codon capture” hypothesis present a detailed pro and con dis-cussion of the two alternatives, and they point out that the evolution ofUGA from a selenocysteine to a stop codon may be very difficult toachieve. They assume that the UGN family box originally encoded bothcysteine and selenocysteine, pairing with the anticodon UCA. After du-plication of the tRNA gene, one of the siblings mutated to GCA, pairingwith UGY codons (the present cysteine codons). The other one (UCA)was “captured” by the newly evolving amino acid tryptophan, and thenchanged to CCA that only pairs with UGG (the present tryptophancodon). The only remaining function of UGA then was coding for se-lenocysteine. In this scheme, UGA as a stop codon then appeared by mu-tation of UAA (Jukes 1990).
Several of these arguments are in accord with recent biochemicalfacts. First, it was shown that cysteine and selenocysteine are equally wellaccepted by cysteinyl-tRNA synthetase and incorporated into protein(Stadtman et al. 1989; Müller et al. 1994, 1997). The previous failure todetect unspecific selenocysteine incorporation might have been due to itschemical instability or to the fact that cystathionine-β-synthase has ahigher affinity for selenocysteine than cysteinyl-tRNA-synthetase, rela-tive to the substrate cysteine. The UGY codons thus can be considered toindiscriminately code for cysteine plus its selenium analog. Second, whenthe UGA codon in the fdhF mRNA was changed to a UGY codon, seleno-cysteine was still incorporated, although to a reduced extent, since seleno-cysteyl-tRNA bound to SelB was competing with cysteyl-tRNA.EF-Tu(Baron et al. 1989). This indicates that the UGY codon can pair with UCAof the selenocysteyl-tRNA and also leads to the conclusion that the switchfrom UGN to UGY and UGA would not have been detrimental, since itinterchanged chemically very similar amino acids.
The assumption that the UGN codon family, and therefore UGA,originally coded for cysteine plus selenocysteine also supports specula-tions on why selenocysteine is incorporated at only a few specific sitesand was not maintained at “neutral” positions. It is assumed that theswitch to specific selenocysteine insertion with the exclusion of cysteineinsertion was a continuous, step by step, optimization process, which al-lowed the development of all components of the insertion machinery, SECIS, tRNASec, SelB, and the biosynthetic path.

646 J.F. Atkins et al.
A Possible Scenario
As discussed previously, selenocysteine might have been incorporated in-discriminately with cysteine, encoded by the UGN codon family and acysteine-specific tRNA (UCA). Indiscriminate incorporation at certainpositions may have conferred to that gene product a higher reactivity andthereby a selective advantage. After duplication of the anticodon and sep-aration of the UGY (cysteine) and UGR families, UGA was maintainedfor the readout of cysteine and selenocysteine and UGG was “captured”by the new amino acid tryptophan (Jukes 1990). The selective advantageforced the development of the selenium biosynthesis and insertion ma-chinery for these special positions. UGA at other positions may have beencounterselected by the high reactivity of selenol residues leading totrapped folding intermediates or by oxidative inactivation due to the ap-pearance of oxygen in the atmosphere (Leinfelder et al. 1988).
The existence of the SECIS element and of the specialized translationfactor might not have been crucial at this stage, since there was no needfor discrimination against chain termination. On the other hand, later de-velopment of the SECIS motif designated the special UGA as sense, spe-cific for selenocysteine, and forced its maintenance. “Unprotected” UGAcould disappear or gain a new function, e.g., termination. It is noteworthythat the SECIS elements of Bacteria, Archaea, and Eukarya bear no struc-tural similarity, which supports the possibility of convergent evolution. Itis also an open question whether the SECIS element within the coding sequence is original and the 3� SECIS derived from it. An argument in favor of this view is that the influence of an mRNA structure on the recoding process may be easier mechanistically if the codon is in thevicinity. However, SECIS elements within coding regions are under se-quence constraint and may not be suitable to direct the insertion of morethan one selenocysteine residue, whereas 3� SECIS elements are not un-der such constraints and have the capacity for multiple insertions (Lowand Berry 1996; Wilting et al. 1997).
The human selP gene has 10 UGA codons, and recoding is promotedby two tandem SECIS elements in the 3� UTR (Hill et al. 1993). It is notat all clear how 3� SECIS elements can promote multiple insertions, espe-cially in view of the apparent inefficiency of even single insertions (seebelow). It is difficult to imagine that a mechanism analogous to theprokaryotic example could suffice; the 3� element would need to cycleaminoacyl tRNA from the element to each UGA as ribosomes progresseddown the message.
An alternative model is that the 3� element interacts with the 5� end ofthe mRNA analogous to the well-known communication of 3� and 5� ends

Recoding 647
of eukaryotic mRNAs. Through this interaction, perhaps the initiating ri-bosome is modified so that for its transit of the mRNA, it reads each UGAcodon as selenocysteine. In this case, the 3� element would provide a ribo-some switch rather than a tRNA delivery system (Gesteland and Atkins1996; Kollmus et al. 1996). Even if triggering the switch for recoding is in-efficient, perhaps once the ribosomes are programmed for selenocysteineincorporation, the efficiency at each subsequent site would be high, and theoverall efficiency of multiple and single selenocysteine incorporations maynot be very different (Kollmus et al. 1996). However, recent experimentscaution against a simple version of this model and can more readily be ex-plained by the information being delivered to ribosomes at individual UGAcodons. In deiodinase mRNA, there is a single selenocysteine-encoding in-ternal UGA codon and a single SECIS element in the 3� UTR. Increasingthe number of SECIS elements had no effect on the efficiency of seleno-cysteine incorporation, whereas it did when the number of UGA codonswas artificially increased (S.C. Low et al., pers. comm.).
The mechanism of mammalian selenocysteine incorporation is unre-solved, particularly as to how the distinction is made between terminationand redefinition. Two additional results may be relevant. A UGA that isless than 55–110 nucleotides from a SECIS element functions as a termi-nator (Martin et al. 1996; Gu et al. 1997), and at least the second UGA inSelP mRNA may sometimes function efficiently as a terminator, sinceforeshortened forms of SelP protein are found (Himeno et al. 1996).
Efficiency
A recent study in E. coli shows that the efficiency of selenocysteine insertion is low. The normal decoding of UGA by SelB-GTP-seleno-cysteyl-tRNA was only 2% efficient compared to decoding of UCA byEF-Tu.GTP.selenocysteyl-tRNA (with a suitably mutant tRNA; S. Supp-mann et al., unpubl.). The efficiency seems limited by nonsaturatingamounts of charged tRNASec and by the kinetics of the formation and res-olution of SelB quaternary complex itself (S. Suppmann et al., unpubl.).This low efficiency in E. coli is similar to the 1–3% levels (Berry et al.1992; Kollmus et al. 1996) measured for selenocysteine insertion in mam-malia (these efficiencies are based on transient transfection experiments,but see Martin et al. 1996).
Evolution of Selenocysteine Biosynthesis
The mode of synthesis of selenocysteine is in accord with an hypothesisfor the coevolution of the genetic code and amino acid biosynthesis

648 J.F. Atkins et al.
(Wong 1975, 1988). UGA belongs to the serine/cysteine codon family,and both cysteine and selenocysteine are synthesized from a serine pre-cursor. Whereas cysteine is synthesized in the low-molecular state andcharged to tRNA by a specific enzyme in Bacteria and Eukarya, biosyn-thesis of selenocysteine takes place in the tRNA-bound state. This is sim-ilar to the biosynthesis of glutaminyl-tRNA or asparaginyl-tRNA from theglutamyl or aspartyl precursors (Ibba et al. 1997) and may reflect coevo-lution as postulated by Wong (1975, 1988). On the other hand, additionalforces may also have been involved in necessitating tRNA-bound biosyn-thesis: (1) Free selenocysteine is highly toxic; (2) the development ofaminoacyl-tRNA synthetases with specific recognition of selenocysteineand cysteine may be difficult to reach in view of the known lack of dis-crimination by cysteyl-tRNA synthetase (Müller et al. 1994). With respectto our model presented above, selenocysteine biosynthesis had to switchfrom a co-synthesis via the cysteine biosynthetic path to the formation inthe tRNA-bound state. This had to occur early in the sequence of eventsdescribed, possibly after the split of the primordial UGN codon familyinto UGY and UGR.
Phylogeny of Sel Gene Products
The fact that selenoproteins occur in all three lines of descent and that se-lenocysteine is encoded in all cases by UGA supports, but by no meansproves, an early evolutionary origin. A considerable number of sel geneshave been cloned and sequenced in the past years allowing (with all reser-vations) some conclusions about relationships.
Selenocysteine Synthase. Selenocysteine synthase is a pyridoxal-phosphate-dependent enzyme. Alignment of the known sequences showsthat the enzyme belongs to the α /γ-superfamily of PLP-dependent en-zymes and that it has diverged very early from the γ-family. It is intrigu-ing that the closest relatives of selenocysteine synthase are enzymes from sulfur metabolism, namely cystathionine-γ-lyase, O-acetylhomo-serine sulfhydrylase, cystathionine-γ-synthase, and cystathionine-β-lyase(Tormay et al. 1998). Thus, selenocysteine synthase may have divergedearly, possibly from some enzyme of sulfur metabolism.
Translation Factor SelB. A dendrogram of the known SelB sequences(Hilgenfeld et al. 1996) revealed that the part of the SelB protein that ishomologous to EF-Tu displays a greater similarity in different organismsthan it does to the EF-Tu sequence from the same organism. This alsoholds for the relationship with IF-2. One can conclude that SelB belongsto an individual class of translation factors that separated very early from

Recoding 649
other factors involved in protein synthesis (Hilgenfeld et al. 1996). Un-fortunately, sequences of SelB homologs from archaeal or eukaryalspecies are not yet available for comparison.
tRNASec. The predicted tRNASec secondary and tertiary structures aremuch more conserved than the primary structure. Alignment of the se-quences shows that the sequence relationships are parallel to those de-duced from the 16S rRNA structures of the same organisms (Tormay etal. 1994; Baron and Böck 1995), although the small size of the moleculedoes not allow statistically significant conclusions
Generality of Redefinition
Sense codons can be redefined to function as start codons. GUG, UUG,and AUU specify valine, leucine, and isoleucine, respectively, when at in-ternal positions of a coding region, but when they function as an initiatorthey specify methionine (or formyl methionine in E. coli). In E. coli andits phages, this redefinition requires an appropriately positioned, preced-ing Shine-Dalgarno sequence. The process is again dynamic; for instance,in the transposon IS911, one particular AUU acts sometimes as an initia-tor and sometimes as an internal sense codon (Polard et al. 1991), and inthe RNA phage fr, a particular UUG behaves similarly (Adhin and vanDuin 1990).
The fact that the meaning of specific codons can be redefined bymRNA context raises the important possibility of specific alteration of themeaning of one internal sense codon to another. Conventional proteinchemistry could easily miss such events if their efficiency was below 10%.
An intriguing question is whether a redefinition strategy is used forthe insertion of additional amino acids beyond the encoded 21, or perhapscould be experimentally exploited for the targeted insertion of normallynonencoded amino acids.
These examples of redefinition of codon meaning all use triplettranslocation, the standard mechanism of mRNA readout. In contrast, thenext type of recoding to be considered involves altering linear readoutandx thus changing the reading frame.
REDIRECTION OF LINEAR READOUT
Frameshifting: Once-only Codon Anticodon Pairing Versus Dissociation and Re-pairing
The issue of entering and maintaining the desired reading frame musthave been a significant one for the early translation apparatus. TripletRNA:RNA interactions are inherently unstable even when the stabilizing

650 J.F. Atkins et al.
topology of an anticodon loop is involved (for review, see Grosjean andChantrenne 1980). This instability is important. The potential for cognatetRNA to dissociate at initial pairing at the A-site allows near-cognate tRNAs to dissociate and to be preferentially discarded (noncognates areless of an issue) (for review, see Yarus and Smith 1995). Dissociation atthe P-site is essential for some types of programmed frameshifting andperhaps one way of dealing with translation errors (Menninger 1977; for review, see Heurgué-Hamard et al. 1996). The weak triplet RNA:RNA in-teraction is stabilized by events at the ribosome in an active way (for re-view, see Yarus and Smith 1995), which themselves favor discrimination.tRNA design is integral to this process. However, the instability of tripletRNA:RNA interactions, which is advantageous now, must have posed aproblem for early decoding in the absence of the stabilizing role of a so-phisticated ribosome and associated factors. If more codon–anticodonbases were paired in early decoding, there is a problem in comprehendinghow decoding could have evolved to triplet codon–anticodon pairingwithout wiping out the fruits of previously selected codons. One proposedscheme for early decoding (Crick et al. 1976) was that at any one time,five codon–anticodon bases were paired, but because of a ratcheting of thetRNA (Woese 1970), only triplet “decoding” was involved. An alterna-tive, which would also not involve whole-scale scrambling of previous information, is that six codon–anticodon bases were initially involved inpairing. If this were so, a transition to triplet pairing would just result in interspersed amino acids. Another alternative is that decoding wastriplet from the start, but that stacking interactions with protoribosomalRNA stabilized the pairing (Noller et al. 1986). Whatever the explanationfor early decoding, it is highly likely that modern protein synthesis involves tRNA interactions with ribosomal components that stabilizecodon:anticodon pairing, and presumably these have a major role, director indirect (Lodmell and Dahlberg 1997), in mediating framing. Eventhough pairing is stabilized, if a tRNA anticodon dissociates from pairingwith its cognate codon within the ribosome and quickly repairs with thesame codon, this would have been undetected in the experiments per-formed to date. If this happens, one might imagine that the function ofsome ribosomal component would be to minimize this dissociation. Onthe basis of what has been found in genetic studies with the large riboso-mal protein L9 (Herbst et al. 1994; Adamski et al. 1996; C. Johnston, un-publ.), it is a candidate for having such a function in E. coli.
The discrimination at the initial selection of tRNAs at the ribosomalA site is impressive. However, this selectivity can get overwhelmed, withserious consequences for frame maintenance, if the balance of competing

Recoding 651
tRNAs is upset, especially with a small minority of tRNAs (Atkins et al.1979; Gallant and Foley 1980; Gallant and Lindsley 1993). An imbalancecan lead to acceptance of a noncognate, or near-cognate, tRNA for pair-ing of its anticodon with the codon. This can be important for frameshift-ing, even if on occasion the effect of framing is not manifest until thetRNA enters the P site (for review, see Farabaugh 1996). However, inmany cases efficient programmed frameshifting follows after selection ofthe cognate tRNA. The tRNAs that mediate frameshifting in response toan imbalance, as well as in response to other signals, are not special interms of their anticodon loop size; they have the same size as virtually alltRNAs. In general, this means that their anticodon size is also the standardthree bases. Whether the shift tRNAs, in some cases, are special in termsof their base modifications or other features (Hatfield et al. 1989; Atkinsand Gesteland 1995; Brierley et al. 1997) is currently being investigated.Following initial studies with model systems (Weiss et al. 1987), it hasbeen found that most cases of programmed frameshifting involve tRNAdissociation and triplet re-pairing in a new frame. Because of this, weakinitial pairing contributes to dissociation and so to frameshifting (Tsuchi-hashi and Brown 1992; Curran 1993).
A key ingredient for efficient programmed frameshifting is having anoverlapping codon available for re-pairing by the P-site tRNA. For +1frameshifting (quadruplet translocation) this is achieved by having thefirst base of the next zero-frame codon temporarily unoccupied. Thismeans having an empty A site. As initially found in model systems (forreview, see Gallant and Lindsley 1993), and later with yeast Ty pro-grammed frameshifting (for review, see Farabaugh 1996), having thezero-frame A-site codon as a rare codon stimulates +1 frameshifting. ThetRNAs for rare codons are themselves sparse. When one of these tRNAsis specified by the codon immediately 3� of a shift codon, the level ofaminoacylation of the tRNA becomes critical for the level of frameshift-ing. Hence, shortage of the amino acid used to charge such a sparse tRNAcan be revealed via programmed +1 frameshifting (Kawakami et al.1993). Amino acid starvation must have been a problem in ancient times,and it is common today for many bacteria. It will be interesting to see ifthe expected attendant frameshifting has regulatory significance. Variousmovable elements, including the yeast Ty elements, become more mobilewhen “hard times” are encountered. It may be of selective advantage fortheir hosts and consequently themselves if they transpose out of importantgenes whose inactivation causes hard times, or if by inserting elsewherethey provide a promoter that activates expression of desirable, but hereto-fore silent, genes. Whatever the reason, synthesis of the transposase in Ty

652 J.F. Atkins et al.
elements requires programmed +1 frameshifting that is responsive to thelevel of a particular aminoacylated, sparse tRNA. In addition, the yeastgene est3 (ever shorter telomeres 3), whose product is required for telom-erase, has similar programmed frameshifting (Morris and Lundblad1997). In contrast, the telomere-specific retrotransposon used for telo-mere maintenance in Drosophila apparently uses –1 frameshifting, and itis in the middle of its gag counterpart in distinction to the location offrameshifting in retrovirus decoding (Danilevskaya et al. 1994, 1998).
Another way to reduce competition in the A site is for the first base ofthe zero-frame codon 3� adjacent to the shift codon to be part of an effi-cient stop codon (Poole et al. 1998). This is illustrated by the programmedframeshifting required for E. coli release factor 2 expression, wherecodons 25 and 26 are CUU UGA (Craigen et al. 1985). tRNALeu pairs ini-tially with the zero-frame CUU and some of the time dissociates and re-pairs with the overlapping UUU (underlined) to cause a shift to the +1frame that encodes the rest of the release factor (Fig. 3A) (Weiss et al.1987). Since termination at UGA is specifically mediated by release fac-tor 2, low amounts of release factor 2 permit a greater chance of re-pair-ing in the +1 frame. Subsequent triplet reading of the +1 frame leads tosynthesis of functional release factor 2. The converse is also true, givingan autoregulatory circuit (Craigen and Caskey 1986).
However, stops may not only stimulate shifting frame by diminishingcompetition from an incoming tRNA. Model systems have shown thatthey stimulate –1 frameshifting where a tRNA re-pairs with a triplet thatoverlaps the previous upstream codon (Weiss et al. 1987; 1990b; Hors-field et al. 1995). This is the basis for the programmed –1 frameshiftingin decoding potato virus M that involves a single-shift tRNA (Gramstat etal. 1994), but the mechanism is unclear.
A very different way to make the last nucleotide of the previous zero-frame codon available for tRNA to re-pair with the overlapping –1 codonis for its corresponding tRNA (in the P site) to also shift –1; i.e., for tan-dem A-site and P-site –1 shifting. This was first discovered for retroviral,programmed –1 frameshifting (Jacks et al. 1988), and is a common type ofprogrammed –1 frameshifting. Since both tRNAs re-pair, the characteris-tic shift sequence for this type of frameshifting is of the general form X-XXY-YYZ. In infectious bronchitis virus it is U-UUA-AAC (Brierley etal. 1992) and in dnaX it is A AAA AAG (see Fig. 3B). There have beenseveral suggestions as to the details of re-pairing with respect to ribosomalA and P sites (Jacks et al. 1988; Weiss et al. 1989; Yelverton et al. 1994;Atkins and Gesteland 1995). However, not all programmed –1 frameshiftsare tandem shifts.

Recoding 653
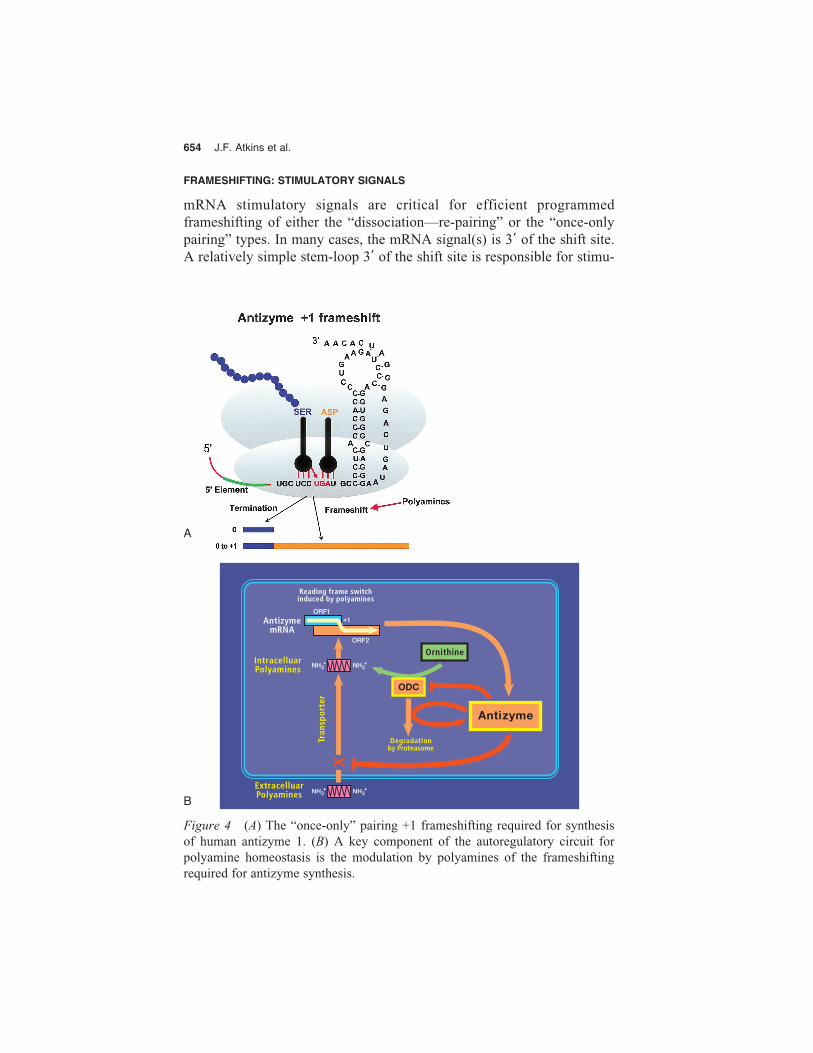
Not all programmed frameshifting involves dissociation and re-pair-ing in an overlapping frame. In yeast Ty3 and mammalian antizyme +1frameshifting, the evidence points to “once-only” pairing so that the firstbase of the next zero-frame codon is somehow unavailable for pairingwith an incoming tRNA (Fig. 4A) (Farabaugh et al. 1993; Matsufuji et al.1995). This means that finding potential frameshift sequences by lookingfor overlapping cognate codons will miss some examples.
Figure 3 (A) The obligatory regulated +1 frameshifting required for synthesis ofE. coli polypeptide chain release factor 2. (B) The tandem codon –1 frameshift re-quired for synthesis of the γ subunit of E. coli DNA polymerase III.
A
B
654 J.F. Atkins et al.
FRAMESHIFTING: STIMULATORY SIGNALS
mRNA stimulatory signals are critical for efficient programmedframeshifting of either the “dissociation—re-pairing” or the “once-onlypairing” types. In many cases, the mRNA signal(s) is 3� of the shift site.A relatively simple stem-loop 3� of the shift site is responsible for stimu-
Figure 4 (A) The “once-only” pairing +1 frameshifting required for synthesis of human antizyme 1. (B) A key component of the autoregulatory circuit for polyamine homeostasis is the modulation by polyamines of the frameshifting required for antizyme synthesis.
A
B

Recoding 655
lating the programmed frameshifting in decoding HIV gag-pol (Parkin etal. 1992; Bidou et al. 1997) and E. coli dnaX (Fig. 3B) (Larsen et al.1997), whereas a complicated stem-loop is utilized in the bacterial trans-posable element, IS911 (Polard et al. 1991). In the case of dnaX, theframeshift efficiency is directly proportional to the predicted stability ofthe stem-loop structure.
The stimulatory mRNA structure is often a pseudoknot located within8 bases 3� of the shift site (Brierley et al. 1989; ten Dam et al. 1990). Ex-amples are found in Coronaviruses (Brierley et al. 1991), retroviruses, thedouble-stranded RNA virus L-A of yeast (Dinman and Wickner 1992; Tuet al. 1992), and mammalian antizyme (Matsufuji et al. 1995, Fig. 4).With mouse mammary tumor virus gag-pro frameshifting, a wedge baseat the junction of the two pseudoknot stems that keeps them from stackingcoaxially was shown to be important for stimulation (Chen et al. 1996).However, there is controversy as to the generality of this conformation (Duet al. 1997; Sung and Kang 1998). A number of these structures are likelyto interact directly with the oncoming ribosome to influence frameshift-ing, but it is possible that some extraribosomal factors are involved inother cases.
Tantalizingly, distant sequences in the 3� UTR are important for pro-grammed frameshifting in decoding barley yellow dwarf luteovirus(Miller et al. 1997), and phage T7 gene 10 (Condron et al. 1991). PhageT7 RNA polymerase transcribes faster than E. coli ribosomes translateand faster than E. coli RNA polymerase transcribes, so that distant 3� se-quences will be transcribed before ribosomes reach the shift site. The pos-sible importance of coupling of replication and translation in some RNAanimal viruses also needs investigation (Lewis and Matsui 1996).
In the few cases tested (Tu et al. 1992; Somogyi et al. 1993), pseudo-knots cause pausing of ribosomes that may be necessary but not sufficientfor recoding. Insertion of a stem-loop with at least equal predicted stabil-ity to a pseudoknot does not lead to frameshifting, even though it stillcauses pausing, albeit less effectively (Somogyi et al. 1993).
Stimulatory signals 5� of shift sites are also found. In one case, a directinteraction with ribosomal RNA of the translocating ribosome has beencharacterized. Three bases 5� of the shift site in the E. coli release factor2 coding sequence, there is a Shine-Dalgarno sequence which is importantfor the +1 frameshifting by ribosomes that initiated 25 codons upstreamof the shift site (Fig. 3A) (Weiss et al. 1987; Curran and Yarus 1988). Theanti-Shine-Dalgarno sequence near the 3� end of 16S rRNA in translocat-ing ribosomes pairs with its mRNA complement and stimulatesframeshifting (Weiss et al. 1988). Similar mRNA:16S rRNA pairing is

656 J.F. Atkins et al.
important for the programmed –1 frameshifting in E. coli dnaX decoding,but here the Shine-Dalgarno sequence is 10 bases 5� of the shift site (Fig.3B) (Larsen et al. 1994). Spacing of the Shine-Dalgarno sequence influ-ences directionality of the shift at the slippery sequence. Perhaps tensionin the short region of 16S rRNA between the anti-Shine-Dalgarno se-quence and the part of the 16S rRNA at the decoding site upsets the fram-ing mechanism. Since Shine-Dalgarno interactions between mRNA andrRNA were discovered for initiation before they were found to be utilizedby translocating ribosomes, one tends unconsciously to think that theyfirst arose for initiation, but of course we don’t know which came first.
Although Shine-Dalgarno interactions are not used for initiation by eu-karyotic ribosomes, it is much too soon to write off the possibility that, atleast for translocating ribosomes and programmed frameshifting, sometype of mRNA–rRNA interaction may be involved. One place to startlooking is at the 5� signal for mammalian antizyme programmed +1frameshifting (Fig. 4A) (Matsufuji et al. 1995; Ivanov et al. 1998a,c; S.Matsufuji, in prep.). It is also possible that interactions of ribosomal RNAin translating ribosomes with mRNA sequences are not confined just tomRNA sequences 5� of the shift site. One candidate for such an interac-tion is the sequence 3� of the Ty3 shift site (Farabaugh et al. 1993).
Ancient Programmed Frameshifting
Most of the known or suspected cases of programmed frameshifting andcodon redefinition, other than selenocysteine, are in viruses or transposableelements. Frameshifting is rampant in the expression of plant virus genesand probably also for bacterial insertion sequences of the IS3 family(Chandler and Fayet 1993; Ohtsubo and Sekine 1996), where approxi-mately 60 cases are suspected (O. Fayet, pers. comm.). It is also found inthe expression of quite a number of animal viruses, especially retroviruses,and also their retrotransposon counterparts. Inferring the evolutionary rela-tionships of the recoding involved in these cases is at an early stage. Ourcomments on this topic, other than selenocysteine discussed above, will berestricted to the programmed frameshifting used in the expression of twononmobile chromosomal genes. The first example is the autoregulatoryframeshifting involved in decoding the bacterial gene for release factor 2which, as described above, mediates termination at UGA (Fig. 3A).
The early evidence that the release factor 2 programmed frameshift-ing signals were highly similar among divergent bacteria came from a se-quence comparison of the shift signals from Bacillus subtilis and E. coli.The 12 nucleotides known to be important for the autoregulatory frame-

Recoding 657
shifting are identical (Pel et al. 1992). A recent analysis of the sequencesfrom 20 bacteria, several of them even more distant than B. subtilis isfrom E. coli, has led to the inference that this frameshift mechanism waspresent in the common ancient ancestor of a large group of divergent bac-teria but was subsequently lost in three independent lineages (Persson andAtkins 1998).
The second case is the +1 frameshifting in decoding antizyme genes.The protein antizyme governs the intracellular level of polyamines bynegatively impacting the intracellular synthesis, and extracellular uptake,of polyamines. It binds to, and inactivates, ornithine decarboxylase, whichcatalyzes the first step of the synthesis of polyamines and also inhibits the polyamine transporter (Fig. 4B). As discovered by Matsufuji and col-leagues (for review, see Gesteland et al. 1992), the programmed frame-shifting required for the synthesis of antizyme is in turn regulated bypolyamines, thus completing an autoregulatory circuit. Following on fromthe original identification of a gene in rats (Miyazaki et al. 1992), a genefor antizyme has been detected in other mammals (Tewari et al. 1994;Kankare et al. 1997; Nilsson et al. 1997), in fowl (Drozdowski et al.1998), in zebra fish (T. Saito et al.; I.P. Ivanov et al., both unpublished),in Xenopus (Ichiba et al. 1995), in Drosophila melanogaster (Ivanov et al. 1998c), in Schizosaccharomyces pombe yeast, and in C. elegans (I.P.Ivanov, unpubl.). When a cassette containing the mammalian antizymeshift site and recoding signal is introduced into the budding yeast, Sac-charomyces cerevisiae, high levels of frameshifting to the +1 frame occurat the shift site. However, the product has an extra amino acid as the ri-bosomes shift –2 instead of +1 and the utilization of the recoding signalsis very different from what it is in mammals (Matsufuji et al. 1996). Incontrast, the same mammalian shift cassette directs mammalian-like +1shifting in the fission yeast, S. pombe (Ivanov et al. 1998b).
Recently, a second antizyme gene has been identified in mammals, andits product, antizyme 2, is distinct from the previously known mammalianantizyme 1 (Ivanov et al. 1998a). Two antizymes are now also known inzebra fish (termed Short and Long to avoid implying correspondence withthe respective mammalian antizymes 1 and 2) (T. Saito et al., unpubl.).Despite substantial divergence of overall nucleotide sequence, the UGAstop codon of ORF1—the first nucleotide of which is part of the shift site(Rom and Kahana 1994; Matsufuji et al. 1995)—and 16 out of 18 nu-cleotides immediately 5� of it are identical from Drosophila antizymemRNA to mammalian antizymes 1 and 2 mRNAs (Ivanov et al. 1998a,c).This sequence includes much of the 5� element discussed above, whichacts in an unknown manner to stimulate frameshifting (S. Matsufuji,

658 J.F. Atkins et al.
unpubl.). The sequences of the stems of the stimulatory pseudoknot, 3� ofthe shift site, are highly conserved between mammalian antizymes 1 and 2,but the loop sequences have diverged (Ivanov et al. 1998a). A flanking 3�
pseudoknot is not apparent in Drosophila by sequence inspection, but thereis some sequence conservation with its mammalian counterparts in this re-gion. It seems safe to discount convergent evolution in the case of an-tizyme and deduce that the shift signals have been used for efficient regu-lated frameshifting for hundreds of millions of years. As suggested by A.E.Dahlberg (pers. comm.), perhaps polyamines played a crucial role with pri-mordial ribosomal RNA and, subsequently, ribosomal proteins displacedsome of these roles. This raises the question of whether the sensing ofpolyamine levels by modern ribosomes is an evolutionary remnant.
SUBVERSION OF CONTIGUITY
Bypassing
As described above, codon–anticodon dissociation can lead to the anti-codon re-pairing to an overlapping triplet resulting in frameshifting. How-ever, the re-pairing can be elsewhere on the mRNA leading to bypassingof mRNA sequences. This was initially discovered with low efficiency(ca. 1%) to nearby sequences in special model systems (Weiss et al. 1987;O’Connor et al. 1989). However, with phage T4 gene 60 decoding, by-passing of 50 bases occurs with an efficiency of 50% from a so-called“take-off” codon to a “landing site” (Fig. 5A) (Huang et al. 1988; Weisset al. 1990a; Maldonado and Herr 1998). The mechanism of this bypassinvolves 70S ribosome complexes, with peptidyl tRNA scanning the gapregion to find the landing site (F. Adamski et al., unpubl.). Part of thenascent peptide, still within the ribosome, is important for this bypassing(Weiss et al. 1990a). The nascent peptide is cross-linkable to 50S subunitcomponents (Choi and Brimacombe 1998) and appears flexible, perhapspartly folded, in an exit tunnel in that subunit. However, it is also cross-linkable to the 30S subunit, close to the decoding site. At least some of itsrole in bypassing may be mediated by direct contacts with the decodingarea of the 30S subunit or with the tRNA–mRNA complex (Choi et al.1998). In addition to the nascent peptide, a short stem-loop within the cod-ing gap is important for bypassing. However, without these two specialfeatures exhibited by gene 60, efficient bypassing can occur over shorterdistances if the codon following the take-off site is a rare codon and itscognate aminoacylated tRNA is limiting (J. Gallant and D. Lindsley, pers.comm.). The above-described translational bypassing is quite distinct

Recoding 659
from the shunting of 40S ribosomal subunits to another site within the 5untranslated regions of cauliflower mosaic virus and adenovirus mRNAs.Here the intervening sequence is not traversed; rather, specific structuresappear to pass the ribosomal subunit from one site to the other (Fütterer etal. 1993; Yueh and Schneider 1996; Hemmings-Mieszczak et al. 1997).
A
B
Figure 5 (A) The recoding signal for the 50% efficient translational bypass of50 nucleotides in decoding phage T4 gene 60. (B) Rescue of ribosomes stalled atthe end of bacterial mRNAs lacking a terminator and degradation of the aberrantprotein product, utilizes tmRNA which functions both as a tRNA and mRNA.

660 J.F. Atkins et al.
Clearly, if bypassing without special signals were rampant, decodingwould be chaotic. Nonetheless, the current translational mechanism is ca-pable of carrying out noncontiguous decoding of a message.
AN INTRAMOLECULAR RIBOSOME?
The primary (modern) function of mRNA is to be a linear tape, feedingthrough the ribosome readout machine. This linear property is, of course,a direct reflection of the information style of its DNA origins. However,as we have seen, mRNA sequences have additional roles in mRNA de-coding. Do these observations provide an insight into primitive decodingearly in evolution?
It is difficult to imagine how the early decoding apparatus could as-semble amino acid chains according to a nucleic acid code. A major dif-ficulty must have been maintaining high enough local concentrations ofthe reactants to drive reactions. However, the number of diffusible reac-tants might have been minimized by combined functions within a multi-functional mRNA molecule. Any such scheme would require evolution ofcomplex RNA molecules, which seems counterintuitive. The trade-off be-tween dealing with many reacting molecules and constructing a complexmolecule may have been tilted toward the latter by evolution of RNA li-gation activity.
The modern translation apparatus assembles the amino acid chain bysequential passage of aminoacyl tRNAs through A and P sites on the ri-bosome, as dictated by codon sequence in the mRNA. The success of invitro protein synthesis experiments makes us think of the decoding appa-ratus operating in a soluble soup with a diffusible flow of substrate tRNAsinto the ribosomal A site, where incorrect molecules are rejected until thecorrect one is identified. However, there is ample evidence that substratesand factors are not just free-floating, but rather are harbored in a ribosomemegacomplex (Stapulionis et al. 1997; Kruse et al. 1998). Within thiscomplex, aminoacyl-tRNAs are tested at the A site, but the volume avail-able for free diffusion must be greatly reduced, aiding the reaction rate.The discharged tRNAs are likely recycled within the complex by residentacylating enzyme. The implication is that small molecules—ATP andamino acids—flow into the complex and the newly synthesized polypep-tide chain emerges.
Could the megacomplex of current ribosomes be suggestive of an ear-lier strategy to deal with the substrate concentration problem in early evo-lution of a translation system? The ultimate megacomplex would havemany functions and substrates in one molecule to maximize the number

Recoding 661
of unimolecular reactions. The multiple capabilities of RNA are intrigu-ing for thinking about a primitive, unimolecular decoding complex in theabsence of proteins.
First, tmRNA combines mRNA and tRNA functions in one molecule(Fig. 5B) (Tu et al. 1995; Keiler et al. 1996; Himeno et al. 1997), henceits name “tmRNA” (Jentsch 1996; Atkins and Gesteland 1996). This 363-nucleotide RNA has 3� and 5� ends that come together to form a partialtRNA-like structure that can be charged with alanine (Komine et al. 1994;Ushida et al. 1994; Felden et al. 1998). It can access a ribosomal A sitethat has no tRNA and no mRNA codon, such as at the end of an mRNAwith no stop codon. The alanine is donated to the growing peptide chainin the P site just as if tmRNA was an ordinary tRNA. Then, remarkably,the ribosome reads out 10 codons from an internal part of tmRNA, addingan 11-amino-acid tag to the growing chain, targeting it for degradation(Keiler et al. 1996). tmRNA has a very complex shape with at least 4pseudoknots (Williams and Bartel 1996; Felden et al. 1997) that must provide the conditions necessary for the tRNA ends to be in the P site andthe internal coding sequence to be in the mRNA track for the A site. Al-though this interesting mechanism employs modern-day ribosomes, itdoes encourage thoughts about one molecule having multiple functionsinvolved in decoding.
Second, peptide-bond catalysis by RNA is clearly possible (Zhang andCech 1997), and it seems likely that the peptidyl transfer function of mod-ern ribosomes is affected by ribosomal RNA (Nitta et al. 1998; see Chap-ter 8). We could imagine that at early times the catalytic center for peptide-bond formation and the informational sequence (mRNA) could be in oneRNA molecule; each mRNA would need to have its own, resident cat-alytic center.
Third, we know that folded RNA structures within an mRNA can par-ticipate in bringing an appropriate substrate aminoacyl tRNA into the ri-bosome for recognition of its codon, as exemplified by selenocysteine in-sertion, in E. coli. In this case, a downstream stem-loop structure (withinthe coding sequence) tethers the tRNASec (via a special EF-Tu protein) inorder to deliver the tRNA to the waiting UGA codon in the ribosomal Asite. Could we imagine that the primitive mRNA catalytic center moleculesuggested above might also have an amino acid delivery system?
There are intriguing suggestions that the amino acid acceptor branch oftRNA (acceptor arm plus TψC arm) originated independently from theanticodon branch (anticodon arm plus DHU arm) (see Chapters 3 and 8)and a reason for the origin of the acceptor branch independent of its rolein protein synthesis has been proposed (see Chapter 3). Perhaps the anti-

662 J.F. Atkins et al.
codon branch of tRNAs originated in folded structures in internal regionsof primordial mRNAs that folded back and paired with “codons” in thesame mRNA. One proto-anticodon branch would need to be capable offorming for each of a limited number of amino acids. How could pairingof the proto-anticodon trigger delivery of an amino acid on a proto-acceptor branch that is not contiguous with the proto-anticodon branch?Two alternatives can be considered. One is that a stereochemical, foldedRNA pocket containing an amino acid is delivered by a structurally con-tiguous proto-anticodon branch due to an association between the two. Ifso, the pocket could hold the now-positioned amino acid at the catalyticsite until the next amino acid is delivered. The amino acid-specific foldcould then bind another amino acid, ready for delivery when its codonwas required again. Thus, a series of “fingers” with bound amino acidscould play back on coding sequences within one molecule to decode a partof the RNA sequence. In this scenario, activation of the amino acid takesplace at the catalytic center. Alternatively, if a 3� end is involved in pri-mordial aminoacylation, it could act repetitively to deliver amino acids tothe catalytic site. In this scenario, the amino acid could not be held at thecatalytic site by the delivery system and might be held by the catalytic siteuntil the 3� end delivered the subsequent amino acid. Perhaps pairing of aparticular anticodon branch with its codon influences, by way of tertiaryinteractions, the identity of the amino acid aminoacylated to the 3� end.(For a discussion of self-aminoacylation, see Chapter 7.) In either case, aseach amino acid reaches the catalytic center, a peptide bond needs to formwith the growing chain. If amino acids are delivered by folded internalpockets, then subsequent evolution of 3� end aminoacylation is a big step.However, synthesis of proteins by the internal delivery system would pro-vide a different milieu for the subsequent but parallel development of the3� end delivery system.
By this imaginary scheme a single RNA molecule is mRNA, peptidebond catalyst, and “tRNA” that acts as an amino acid collection and de-livery system. Many interactions would be intramolecular; the diffusion-limited reactions would be the amino acids finding their binding pockets.Another possibility, maybe a step further in evolution, might be a two-component system, with the mRNA molecule separate from a “primitiveribosome” that had the peptide bond catalyst, the tRNAs, and the deliverysystem in one molecule. This “ribosome” could then act on a variety ofmRNAs, much like the modern ribosome megacomplex. These scenariosare admittedly farfetched and do not deal with a number of crucial issues.However, they may be illustrative of ways to think about multifunctionalRNA “mega” molecules.

Recoding 663
PERSPECTIVE
With a more complete understanding of decoding and recoding, it may bepossible to consider engineering organisms with an expanded repertoireof coding capacities to include nontraditional amino acids. The challengesare clearly formidable, but there is already an impressive start by specificmanipulation of an aminoacyl tRNA synthetase (Liu et al. 1997).
Early decoding likely yielded many products from a single coding se-quence, because of randomness in the mechanism. Presumably, the dif-fering specificities of these products gave some molecules with activitiesthat provided a survival advantage. As sophistication of the decoding sys-tem evolved, the repertoire of products from a single coding sequencemust have become more limited, eventually reaching the current codingrules. Although these rules, in general, result in one protein product permRNA, recoding examples tell us that there has been coevolution of spe-cific mechanisms to produce more than one product. The big unknown ishow many coding sequences have themselves evolved to take advantageof the diversity of expression offered by recoding. Might there be manymessages where 5% of the ribosomes bypass the terminator, readingcodons in what is normally considered to be the 3�UTR? How commonlydoes decoding of mRNAs involve frameshifting akin to mammalian an-tizyme? Could it be that, in some mRNAs, ribosomes bypass codons byscanning from one codon to the next cognate one, as seen in T4 gene 60?Each of these acts would result in a different protein with perhaps a newor additional function. Although current methods for protein analysis arevery powerful, rarely would protein variants at the level of 5% be discov-ered, except by a fortuitous observation or by serious digging. This alsoholds true for posttranslational modifications. Again, we know a greatdeal about some proteins, but we have little appreciation for the overallpicture. A serious attack on the “proteome” is needed to begin to under-stand the full diversity of the products of genes.
ACKNOWLEDGMENTS
We thank Marla Berry for characteristically helpful comments and sharingunpublished data. J.F.A. is supported by National Institutes of Health grantRO1-GM48152 and Japan Health Science Foundation grant K-1005; A.B.by Deutsche Forschungsgemeinschaft and Fonds der Chemischen Indus-trie; S.M. by Grants-in-Aid from the Ministry of Education, Science andCulture in Japan, and the Human Frontier Sciences Program; and R.F.G. isan investigator of the Howard Hughes Medical Institute.

664 J.F. Atkins et al.
REFERENCES
Adamski F.M., Atkins J.F., and Gesteland R.F. 1996. Ribosomal protein L9 interactionswith 23 S rRNA: The use of a translational bypass assay to study the effect of aminoacid substitutions. J. Mol. Biol. 261: 357–371.
Adhin, M.R. and van Duin, J. 1990. Scanning model for translational reinitiation in eu-bacteria. J. Mol. Biol. 213: 811–818.
Atkins J.F. and Gesteland R.F. 1995. Discontinuous triplet decoding with or without re-pairing by peptidyl tRNA. In tRNA: Structure, biosynthesis and function (ed. D. Sölland U.L. RajBhandary) pp. 471–490. ASM Press, Washington, D.C.
———. 1996. A case for trans translation. Nature 379: 769–770.Atkins J.F., Elseviers D., and Gorini L. 1972. Low activity of β-galactosidase in frameshift
mutants of Escherichia coli. Proc. Natl. Acad. Sci. 69: 1192–1195. Atkins J.F., Weiss R.B., and Gesteland R.F. 1990. Ribosome gymnastics—Degree of dif-
ficulty 9.5, style 10.0. Cell 62: 413–423.Atkins J.F., Gesteland R.F., Reid B.R., and Anderson C.W. 1979. Normal tRNAs promote
ribosomal frameshifting. Cell 18: 1119–1131. Axley M.J, Böck A., and Stadtman T.C. 1991. Catalytic properties of an Escherichia coli
formate dehydrogenase mutant in which sulfur replaces selenium. Proc. Natl. Acad.Sci. 88: 8450–8454.
Baron C. and Böck A. 1991. The length of the aminoacyl acceptor stem of the selenocys-teine-specific tRNASec of Escherichia coli is the determinant for binding to elongationfactor SELB or Tu. J. Biol. Chem. 266: 20375–20379.
———. 1995. The selenocysteine-inserting tRNA species: Structure and function. IntRNA: Structure, biosynthesis and function (ed. D. Söll and U.L. RajBhandary), pp.529–544. ASM Press, Washington, D.C.
Baron C., Heider J., and Böck A. 1989. Mutagenesis of selC, the gene for the selenocys-teine-inserting tRNA species in E. coli: Effects on in vivo function. Nucleic Acids Res.18: 6761–6766.
Baron C., Westhof E., Böck A., and Giege R. 1993. Solution structure of selenocysteine-inserting tRNASec from Escherichia coli. Comparison with canonical tRNASec. J. Mol.Biol. 231: 274–292.
Behne D., Kyriakopoeulos A., Weiss-Nowak C., Kalcklösch M., Westphal C., and Gess-ner H. 1996. Newly found selenium-containing proteins in the tissues of the rat. Biol.Trace Elem. Res. 55: 99–110.
Berry M.J., Banu L., Harney J.W., and Larsen P.R. 1993. Functional characterization ofthe eukaryotic SECIS elements which direct selenocysteine insertion at UGA codons.EMBO J. 12: 3315–3322.
Berry M.J., Harney J.W., Ohama T., and Hatfield D.L. 1994. Selenocysteine insertion or termination: Factors affecting UGA codon fate and complementary anticodon:codon mutations. Nucleic Acids Res. 22: 3753–3759.
Berry M.J., Mai A.L., Kieffer J.D., Harney J.W., and Larsen P.R. 1992. Substitution of cysteine for selenocysteine in type I iodothyronine deiodinase reduces the cata-lytic efficiency of the protein but enhances its translation. Endocrinology 131: 1848–1852.
Berry M.J., Banu L., Chen Y., Mandel S.J., Kieffer J.D., Harney J.W., and Larsen P.R.1991. Recognition of UGA as a selenocysteine codon in type I deiodinase requires sequences in the 3� untranslated region. Nature 353: 273–276.

Recoding 665
Bidou L., Stahl G., Grima B., Liu H., Cassan M., and Rousset J.-P. 1997. In vivo HIV-1frameshifting efficiency is directly related to the stability of the stem-loop stimulatorysignal. RNA 3: 1153–1158.
Blinkowa A.L. and Walker J.R. 1990. Programmed ribosomal frameshifting generates theEscherichia coli DNA polymerase III γ subunit from within the τ subunit readingframe. Nucleic Acids Res. 18: 1725–1729.
Brierley I., Digard P., and Inglis S.C. 1989. Characterization of an efficient Coronavirusframeshifting signal: Requirement for an RNA pseudoknot. Cell 57: 537–547.
Brierley I., Jenner A.J., and Inglis S.C. 1992. Mutational analysis of the “slippery-sequence”component of a Coronavirus ribosomal frameshifting signal. J. Mol. Biol. 227: 463–479.
Brierley I., Meredith M.R., Bloys A.J., and Hagervall T.G. 1997. Expression of a Corona-virus ribosomal frameshift signal in Escherichia coli: Influence of tRNA anticodonmodification on frameshifting. J. Mol. Biol. 271: 1–14.
Brierley I., Rolley N.J., Jenner A.J., and Inglis S.J. 1991. Mutational analysis of the RNApseudoknot component of a Coronavirus frameshifting signal. J. Mol. Biol. 220:889–902.
Brown C.M., Dinesh-Kumar S.P., and Miller A. 1996. Local and distant sequences are re-quired for efficient read-through of the barley yellow virus-PAV coat protein gene stopcodon. J. Virol. 70: 5884–5892.
Bult C.J., White O., Olsen G.J., Zhou L., Fleischmann R.D., Sutton G.G., Blake J.A.,FitzGerald L.M., Clayton R.A., Gocayne J.D et al. 1996. Complete genome sequenceof the methanogenic archaeon, Methanococcus jannaschii. Science 273: 1058–1072.
Chambers I., Frampton J., Goldfarb P, Affara N., McBain W., and Harrison P.R. 1986.The structure of the mouse glutathione peroxidase gene: The selenocysteine in the ac-tive site is encoded by the “termination” codon TGA. EMBO J. 5: 1221–1227.
Chandler M. and Fayet O. 1993. Translational frameshifting in the control of transpositionin bacteria. Mol. Microbiol. 7: 497–503.
Chen X., Kang H., Shen L.X., Chamorro M., Varmus H.E., and Tinoco I, Jr. 1996. A char-acteristic bent conformation of RNA pseudoknots promotes –1 frameshifting duringtranslation of retroviral RNA. J. Mol. Biol. 260: 479–483.
Choi K.M. and Brimacombe R. 1998. The path of the growing peptide chain through the23S rRNA in the 50S ribosomal subunit; a comparative cross-linking study with threedifferent peptide families. Nucleic Acids Res. 26: 887–895.
Choi K.M., Atkins J.M., Gesteland R.F., and Brimacombe R. 1998. Flexibility of thenascent polypeptide chain within the ribosome. Contacts from the peptide N-terminusto a specific region of the 30S subunit. Eur. J. Biochem. 255: 409–413.
Condron B.G., Gesteland R.F., and Atkins J.F. 1991. An analysis of sequences stimulat-ing frameshifting in the decoding of gene 10 of bacteriophage T7. Nucleic Acids Res.19: 5607–5612.
Cone J.E., del Rio M., Davis J.N., and Stadtman T.C. 1976. Chemical characterization ofthe selenoprotein component of clostridial glycine reductase: Identification of seleno-cysteine as the organoselenium moiety. Proc. Natl. Acad. Sci. 73: 2659–2663.
Craigen W.J. and Caskey C.T. 1986. Expression of peptide chain release factor 2 requireshigh-efficiency frameshift. Nature 322: 273–275.
Craigen W.J., Cook R.G., Tate W.P., and Caskey C.T. 1985. Bacterial peptide chain re-lease factors: Conserved primary structure and possible frameshift regulation of releasefactor 2. Proc. Natl. Acad. Sci. 82: 3616–3620.

666 J.F. Atkins et al.
Crick F.H.C., Brenner S., Klug A., and Pieczenik G. 1976. A speculation on the origin ofprotein synthesis. Origins Life 7: 389–397.
Curran J.F. 1993. Analysis of effects of tRNA: Message stability on frameshift frequencyat the Escherichia coli RF2 programmed frameshift site. Nucleic Acids Res. 21: 1837–1843.
Curran J.F. and Yarus M. 1988. Use of tRNA suppressors to probe regulation of Esch-erichia coli release factor 2. J. Mol. Biol. 203: 75–83.
Danilvskaya O., Slot F., Pavlova M., and Pardue M.-L. 1994. Structure of the DrosophilaHeT-A transposon: A retrotransposon-like element forming telomeres. Chromosoma103: 215–224.
Danilevskaya O.N., Tan C., Wong J., Alibhai M., and Pardue M.-L. 1998. Unusual fea-tures of the Drosophila melanogaster telomere transposable element HeT-A are con-served in Drosophila yakuba telomere elements. Proc. Natl. Acad. Sci. 95: 3770–3775.
Di Guilio M. 1997. The origin of the genetic code. Trends Biochem. Sci. 22: 49.Dinman J.D. and Wickner R.B. 1992. Ribosomal frameshifting efficiency and gag/gag-pol
ratio are critical for yeast M1 double-stranded RNA virus propagation. J. Virol. 66:3669–3676.
Drozdowski B., Gong T.W.-L., and Lomax M.I. 1998. The chicken cDNA for ornithinedecarboxylase antizyme. Biochim. Biophys. Acta 1396: 21–26.
Du Z., Holland J.A., Hansen M.R., Giedroc D.P., and Hoffman D.W. 1997. Base-pairingswithin the RNA pseudoknot associated with the simian retrovirus-1 gag-pro frameshiftsite. J. Mol. Biol. 270: 464–470.
Ehrenreich A., Forchhammer K., Tormay P., Vepreck B., and Böck A. 1992. Selenopro-tein synthesis in E. coli. Purification and characterization of the enzyme catalyzing se-lenium activation. Eur. J. Biochem. 206: 767–773.
Farabaugh P.J. 1996. Programmed translational frameshifting. Microbiol. Rev. 60: 103–134.
Farabaugh P.J., Zhao H., and Vimaladithan A. 1993. A novel programmed frameshift ex-presses the POL3 gene of retrotransposon Ty3 of yeast: Frameshifting without tRNAslippage. Cell 74: 93–103.
Felden B., Himeno H., Muto A., McCutcheon J.P., Atkins J.F., and Gesteland R.F. 1997.Probing the structure of the Escherichia coli 10Sa (tmRNA). RNA 3: 89–104.
Felden B., Hanawa K., Atkins J.F., Himeno H., Muto A., Gesteland R.F., McCloskey J.A.,and Crain P.F. 1998. Presence and location of modified nucleotides in E. coli tmRNA:Structural mimicry with tRNA acceptor branches. EMBO J. 17: 3188–3196.
Felsenstein K.M. and Goff S.P. 1992. Mutational analysis of the gag-pol junction ofMoloney murine leukemia virus: Requirements for expression of the gag-pol fusionprotein. J. Virol. 66: 6601–6608.
Feng Y., Yuan H., Rein A., and Levin J.G. 1992. Bipartite signal for read-through sup-pression in murine leukemia virus mRNA: An eight nucleotide purine-rich sequenceimmediately downstream of the gag termination codon followed by an RNA pseudo-knot. J. Virol. 66: 5127–5132.
Flower A.M. and McHenry C.S. 1990. The γ subunit of DNA polymerase III holoenzymeof Escherichia coli is produced by ribosomal frameshifting. Proc. Natl. Acad. Sci. 87:3713–3717.
Forchhammer K. and Böck A. 1991. Selenocysteine synthase from Escherichia coli. Anal-ysis of the reaction sequence. J. Biol. Chem. 266: 6324–6328.

Recoding 667
Forchhammer K., Leinfelder W., and Böck A. 1989. Identification of a novel translationfactor necessary for the incorporation of selenocysteine into proteins. Nature 342:453–456.
Fütterer J., Kiss-László Z., and Hohn T. 1993. Nonlinear ribosome migration oncauliflower mosaic virus 35S RNA. Cell 73: 789–802.
Gallant J. and Foley D. 1980. On the causes and prevention of mistranslation. In Ribo-somes: Structure, function and genetics (ed. G. Chambliss et al.), pp. 615–638. Uni-versity Park Press, Baltimore.
Gallant J. and Lindsley D. 1993. Ribosome frameshifting at hungry codons: Sequencerules, directional specificity and possible relationship to mobile element behaviour.Biochem. Soc. Trans. 21: 817–821.
Gesteland R.F. and Atkins J.F. 1996. Recoding: Dynamic reprogramming of translation.Annu. Rev. Biochem. 65: 741–768.
Gesteland R.F., Weiss R.B., and Atkins J.F. 1992. Recoding: Reprogrammed genetic de-coding. Science 257: 1640–1641.
Gramstat A. Prüfer D., and Rohde W. 1994. The nucleic acid-binding zinc finger proteinof potato virus M is translated by internal initiation as well as by ribosomal frameshift-ing involving a shifty stop codon and a novel mechanism of P-site slippage. NucleicAcids Res. 22: 3911–3917.
Grosjean H. and Chantrenne H. 1980. On codon-anticodon interactions. Mol. Biol. Bio-chem. Biophys. 32: 347–367.
Gu Q.-P., Beilstein M.A., Vendeland S.C., Lugade A., Ream W., and Whanger P.D. 1997.Conserved features of selenocysteine insertion sequence (SECIS) elements in seleno-protein W cDNAs from five species. Gene 193: 187–196.
Hatfield D., Feng Y.X., Lee, B.J., Rein A., Levin J.G., and Oroszlan S. 1989. Chromato-graphic analysis of the aminoacyl tRNAs which are required for translation of codonsat and around the ribosomal frameshift sites of HIV, HTLV-1, and BLV. Virology 173:736–742.
Heider J., Baron C., and Böck A. 1992. Coding from a distance: Dissection of the mRNAdeterminants required for the incorporation of selenocysteine into proteins. EMBO J.11: 3759–3766.
Hemmings-Mieszczak M., Steger G., and Hohn T. 1997. Alternative structures of thecauliflower mosaic virus 35S RNA leader: Implications for viral expression and repli-cation. J. Mol. Biol. 267: 1075–1088.
Herbst K.L., Nichols L.M., Gesteland R.F., and Weiss R.B. 1994. A mutation in riboso-mal protein L9 affects ribosomal hopping during translation of gene 60 from bacterio-phage T4. Proc. Natl. Acad. Sci. 91: 12525–12529.
Heurgué-Hamard V., Mora L., Guarneros G., and Buckingham R.H. 1996. The growth de-fect in Escherichia coli deficient in peptidyl-tRNA hydrolyase is due to starvation forLys-tRNALys . EMBO J. 15: 2826–2833.
Hilgenfeld R., Wilting R., and Böck A. 1996. Structural model for the selenocysteine-specific elongation factor SelB. Biochimie 78: 971–978.
Hill K.E., Lloyd R.S., and Burk R.F. 1993. Conserved nucleotide sequences in the openreading frame and 3� untranslated region of selenoprotein P mRNA. Proc. Natl. Acad.Sci. 90: 537–541.
Himeno S., Chittum H.S., and Burk R.F. 1996. Isoforms of selenoprotein P in rat plasma.J. Biol. Chem. 271: 15769–15775.

668 J.F. Atkins et al.
Himeno H., Sato M., Tadaki T., Fukushima M., Ushida C., and Muto A. 1997. In vitrotrans translation mediated by alanine-charged 10Sa RNA. J. Mol. Biol. 268: 803–808.
Himmelreich R., Plagens H., Hilbert H., Reiner B., and Herrmann R. 1997. Comparativeanalysis of the genomes of the bacteria Mycoplasma pneumoniae and Mycoplasmagenitalium. Nucleic Acids Res. 25: 701–712.
Hofstetter H., Monstein H.-J., and Weissman C. 1974. The readthrough protein A1 is es-sential for the formation of viable Qβ particles. Biochim. Biophys. Acta 374: 238–251.
Horsfield J.A., Wilson D.N., Mannering S.A., Adamski F.M., and Tate W.P. 1995.Prokaryotic ribosomes recode the HIV-1 gag-pol–1 frameshift sequence by an E/P sitepost-translocation simultaneous slippage mechanism. Nucleic Acids Res. 23:1487–1494.
Huang W.M., Ao S.-Z., Casjens S., Orlandi R., Zeikus R., Weiss R., Winge D., and FangM. 1988. A persistent untranslated sequence within bacteriophage T4 DNA topoisom-erase gene 60. Science 239: 1005–1012.
Hüttenhofer A. and Böck A. 1998a. RNA structures involved in selenoprotein synthesis.In RNA structure and function (ed. R. Simons and M. Grunberg-Manago), pp.603–639. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
———. 1998b. Selenocysteine-inserting RNA elements mediate GTP hydrolysis by elon-gation factor SelB. Biochemistry 37: 885–890.
Hüttenhofer A., Heider J., and Böck A. 1996. Interaction of the Escherichia coli fdhFmRNA hairpin promoting selenocysteine incorporation with the ribosome. NucleicAcids Res. 20: 3903–3910.
Ibba M., Curnow A.W., and Söll D. 1997. Aminoacyl-tRNA synthesis: Divergent routesto a common goal. Trends Biochem. Sci. 22: 39–42.
Ichiba T., Matsufuji S., Miyazaki Y., and Hayashi S. 1995. Nucleotide sequence of or-nithine decarboxylase antizyme cDNA from Xenopus laevis. Biochim. Biophys. Acta1262: 83–86.
Ivanov I.P., Gesteland R.F., and Atkins J.F. 1998a. A second mammalian antizyme: Con-servation of programmed ribosomal frameshifting. Genomics (in press).
Ivanov I.P., Gesteland R.F., Matsufuji S., and Atkins J.F. 1998b. Programmed frameshift-ing in the synthesis of mammalian antizyme is +1 in mammals, predominantly +1 infission yeast but –2 in budding yeast. RNA (in press).
Ivanov I.P., Simin K., Letsou, A., Atkins J.F., and Gesteland R.F. 1998c. The Drosophilagene for antizyme requires ribosomal frameshifting for expression and contains an intronic gene for snRNP Sm D3 on the opposite strand. Mol. Cell. Biol. 18: 1553–1561.
Jacks T., Madhani T.H.D., Masiarz F.R., and Varmus H.E. 1988. Signals for ribosomalframeshifting in the Rous sarcoma virus gag-pol region. Cell 55: 447–458.
Jentsch S. 1996. When proteins receive deadly messages at birth. Science 271: 955–956.Jukes T.H. 1990. Genetic code 1990. Outlook. Experientia 46: 1149–1157.Kankare K., Uusi-Oukari M., and Jänne O.A. 1997. Structure, organization and expression
of the mouse ornithine decarboxylase antizyme gene. Biochem. J. 324: 807–813. Kawakami K., Pande S., Faiola B., Moore D.P., Boeke J.D., Farabaugh P.J., Strathern
J.N., Nakamura Y., and Garfinkel D.J. 1993. A rare tRNA-Arg(CCU) that regulatesTy1 element ribosomal frameshifting is essential for Ty1 retrotransposition in Saccha-romyces cerevisiae. Genetics 135: 309–320.
Keiler K.C., Waller P.R.H., and Sauer R.T. 1996. Role of a peptide tagging system in degra-dation of proteins synthesized from damaged messenger RNA. Science 271: 990–993.

Recoding 669
Kelman Z. and O’Donnell M. 1995. DNA polymerase III: Structure and function of a chro-mosomal replicating machine. Annu. Rev. Biochem. 64: 171–200.
Kollmus H., Flohe L., and McCarthy J.E.G. 1996. Analysis of eukaryotic mRNA structuredirecting cotranslational incorporation of selenocysteine. Nucleic Acids Res. 24:1195–1201.
Komine Y., Kitabatake M., Yokogawa T., Nishikawa K., and Inokuchi H. 1994. A tRNA-like structure is present in 10Sa RNA, a small stable RNA from Escherichia coli. Proc.Natl. Acad. Sci. 91: 9223–9227.
Kromayer M., Wilting R., Tormay P., and Böck A. 1996. Domain structure of the prokary-otic selenocysteine-specific elongation factor SELB. J. Mol. Biol. 262: 413–420.
Kruse C., Grünweller A., Willkomm D.K., Pfeiffer T., Hartmann R.K., and Müller P.K.1998. tRNA is entrapped in similar, but distinct, nuclear and cytoplasmic ribonucleo-protein complexes, both of which contain vigilin and elongation factor 1α. Biochem. J.329: 615–621.
Kurland C.G. 1992. Translational accuracy and the fitness of bacteria. Annu. Rev. Genet.26: 29–50.
Larsen B., Gesteland R.F., and Atkins J.F. 1997. Structural probing and mutagenic analy-sis of the stem-loop required for E. coli dnaX ribosomal frameshifting: Programmed ef-ficiency of 50%. J. Mol. Biol. 271: 47–60.
Larsen B., Wills N.M., Gesteland R.F., and Atkins J.F. 1994. rRNA-mRNA base pairingstimulates a programmed –1 ribosomal frameshift. J. Bacteriol. 176: 6842–6851.
Leinfelder W., Zehelein E., Mandrand-Berthelot M.A., and Böck A. 1988. Gene for anovel tRNA species that accepts L-serine and cotranslationally inserts selenocysteine.Nature 331: 723–725.
Leinfelder W., Forchhammer K., Veprek B., Zehelein E., and Böck A. 1990. In vitro syn-thesis of selenocysteyl-tRNAUCA from seryl-tRNAUCA: Involvement and characteriza-tion of the selD gene product. Proc. Natl. Acad. Sci. 87: 543–547.
Leng P., Klatte D.H., Schumann G., Boeke J.D., and Steck T.L. 1998. Skipper, an LTRretrotransposon of Dictyostelium. Nucleic Acids Res. 26: 2008–2015.
Levin M.E., Hendrix R.W., and Casjens S.R. 1993. A programmed translational frameshiftis required for the synthesis of a bacteriophage λ tail assembly protein. J. Mol. Biol.234: 124–139.
Lewis T.L. and Matsui S.M. 1996. Astrovirus ribosomal frameshifting in an infection-transfection transient expression system. J. Virol. 70: 2869–2875.
Liu D.R., Magliery T.J., Pastrnak M., and Schultz P.G. 1997. Engineering a tRNA andaminoacyl-tRNA synthetase for the site-specific incorporation of unnatural aminoacids into proteins in vivo. Proc. Natl. Acad. Sci. 94: 10092–10097.
Lodmell J.S. and Dahlberg A.E. 1997. A conformational switch in Escherichia coli 16Sribosomal RNA during decoding of messenger RNA. Science 277: 1262–1267.
Low S.C. and Berry M.J. 1996. Knowing when not to stop: Selenocysteine incorporationin eukaryotes. Trends Biochem. Sci. 21: 203–208.
Maldonado R. and Herr A.J. 1998. The efficiency of T4 gene 60 translational bypassing.J. Bacteriol. 180: 1822–1830.
Martin G.W., Harney J.W., and Berry M.J. 1996. Selenocysteine incorporation in eukary-otes: Insights into mechanism and efficiency from sequence, structure, and spacingproximity studies of the type I deiodinase SECIS element. RNA 2: 171–182.
Matsufuji S., Matsufuji T., Wills N.M., Gesteland R.F., and Atkins J.F. 1996. Reading twobases twice: Mammalian antizyme frameshifting in yeast. EMBO J. 15: 1360–1370.

670 J.F. Atkins et al.
Matsufuji S., Matsufuji T., Miyazaki Y., Atkins J.F., Gesteland R.F., and Hayashi S. 1995.Autoregulatory frameshifting in decoding mammalian ornithine decarboxylase an-tizyme. Cell 80: 51–60.
Menninger J.R. 1977. Ribosome editing and the error catastrophe hypothesis of cellularaging. Mech. Ageing Dev. 6: 131–142.
Miller W.A., Brown C.M., and Wang S. 1997. New punctuation for the genetic code: Lu-teovirus gene expression. Semin. Virol. 8: 3–13.
Miyazaki Y., Matsufuji S., and Hayashi S. 1992. Cloning and characterization of a rat geneencoding ornithine decarboxylase antizyme. Gene 113: 191–197.
Morris D.K. and Lundblad V. 1997. Programmed translational frameshifting in a gene re-quired for yeast telomere replication. Curr. Biol. 7: 969–976.
Mottagui-Tabar S., and Isaksson L.A. 1997. Only the last amino acids in the nascent peptideinfluence translation termination in Escherichia coli genes. FEBS Lett. 414: 165–170.
Müller S., Heider J., and Böck A. 1997. The path of unspecific incorporation of seleniumin Escherichia coli. Arch. Microbiol. 168: 421–427.
Müller S., Senn H., Gsell B., Vetter W., Baron C., and Böck A. 1994. The formation ofdiselenide bridges in proteins by incorporation of selenocysteine residues: Biosynthe-sis and characterization of (Se2)-thioredoxin. Biochemistry 33: 3404–3412.
Neuhierl B. and Böck A. 1996. On the mechanism of selenium tolerance in selenium-accumulating plants. Purification and properties of a specific selenocysteine methy-transferase from cultured cells of Astragalus bisulcatus. Eur. J. Biochem. 239: 235–238.
Nilsson J., Koskiniemi S., Persson K., Grahn B., and Holm I. 1997. Polyamines regulateboth transcription and translation of the gene encoding ornithine decarboxylase an-tizyme in mouse. Eur. J. Biochem. 250: 223–231.
Nitta I., Ueda T., and Watanabe K. 1998. Possible involvement of Escherichia coli 23S ri-bosomal RNA in peptide bond formation. RNA 4: 257–267.
Noller H.F., Asire M., Barta A., Douthwaite S., Goldstein T., Gutell R.R., Moazed D.,Normanly J., Prince J.B., Stern S., Triman K., Turner S., van Stolk B., Wheaton V.,Weiser B., and Woese C.R. 1986. Studies on the structure and function of ribosomalRNA. In Structure, function and genetics of ribosomes (ed. B. Hardesty and G.Kramer), pp. 143–163. Springer-Verlag, New York.
O’Connor M., Gesteland R.F., and Atkins J.F. 1989. tRNA hopping: Enhancement by anexpanded anticodon. EMBO J. 8: 4315–4323.
Ohtsubo E. and Sekine Y. 1996. Bacterial insertion sequences. Curr. Top. Microbiol. Im-munol. 204: 1–26.
Osawa S. 1995. Evolution of the genetic code. Oxford University Press, New York.Parkin N.T., Chamorro M., and Varmus H.E. 1992. Human immunodeficiency virus type
1 gag-pol frameshifting is dependent on downstream mRNA secondary structure:Demonstration by expression in vivo. J. Virol. 66: 5147–5151.
Pel H.J., Rep M., and Grivell L.A. 1992. Sequence comparison of new prokaryotic and mi-tochondrial members of the polypeptide chain release factor family predicts a five-domain model for release factor structure. Nucleic Acids Res. 20: 4423–4428.
Persson B.C. and Atkins J.F. 1998. Does the disparate occurrence of the autoregulatoryprogrammed frameshifting in decoding the release factor 2 gene reflect an ancient ori-gin with loss in independent lineages? J. Bacteriol. 180: 3462–3466.
Polard P., Prère M.F., Chandler M., and Fayet O. 1991. Programmed translationalframeshifting and initiation at an AUU codon in gene expression of bacterial insertionsequence IS911. J. Mol. Biol. 222: 465–477.

Recoding 671
Poole E.S., Major L.L., Mannering S.A., and Tate W.P. 1998. Translation termination inEscherichia coli: Three bases following the stop codon crosslink to release factor 2 andaffect the decoding efficiency of UGA-containing signals. Nucleic Acids Res. 26:954–960.
Robinson D.N. and Cooley L. 1997. Examination of the function of two kelch proteinsgenerated by stop codon suppression. Development 124: 1405–1417.
Rom E. and Kahana C. 1994. Polyamines regulate the expression of ornithine decarboxy-lase antizyme in vitro by inducing ribosomal frame-shifting (correction, p. 9195). Proc.Natl. Acad. Sci. 91: 3959–3963.
Shen Q., Chu F.F., and Newburger P.E. 1993. Sequences in the 3�-untranslated region ofthe human cellular glutathione peroxidase gene are necessary and sufficient for se-lenocysteine incorporation at the UGA codon. J. Biol. Chem. 268: 11463–11469.
Skuzeski J.M., Nichols L.M., Gesteland R.F., and Atkins J.F. 1991. The signal for a leakyUAG stop codon in several plant viruses includes the two downstream codons. J. Mol.Biol. 218: 365–373.
Sliwkowski M.X. and Stadtman T.C. 1985. Incorporation and distribution of selenium intothiolase from Clostridium kluyveri. J. Biol. Chem. 260: 3140–3144.
Smith D.R., Doucette-Stamm L.A., Deloughery C., Lee H., Dubois J., Aldredge T.,Bashirzadeh R., Blakely D., Cook R., Gilbert K., Harrison D., Hoang L., Keagle P.,Lumm W., Pothier B., Qiu D., Spadafora R., Vicaire R., Wang Y., Wierzbowski J.,Gibson R., Jiwani N., Caruso A., Bush D., Reeve J.N. et al. 1997. Complete genomesequence of Methanobacterium thermoautotrophicum ∆H: Functional analysis andcomparative genomics. J. Bacteriol. 179: 7135–7155.
Somogyi P., Jenner A.J., Brierley I., and Inglis S.C. 1993. Ribosomal pausing duringtranslation of an RNA pseudoknot. Mol. Cell. Biol. 13: 6931–6940.
Stadtman T.C., Davis J.N., Zehelin E., and Böck A. 1989. Biochemical and genetic anal-ysis of Salmonella typhimurium and Escherichia coli mutants defective in specific incorporation of selenium into formate dehydrogenase and tRNAs. Biofactors 2:35–44.
Stahl G., Bidou L., Rousset J.-P., and Cassan M. 1995. Versatile vectors to study recod-ing: Conservation of rules between yeast and mammalian cells. Nucleic Acids Res. 23:1557–1560.
Stapulionis R., Kolli S., and Deutscher M.P. 1997. Efficient mammalian protein synthesisrequires an intact F-actin system. J. Biol. Chem. 272: 24980–24986.
Steneberg P., Englund C., Kronhamn J., Weaver T.A., and Samakovlis C. 1998. Transla-tional readthrough in the hdc mRNA generates a novel branching inhibitor in theDrosophila trachea. Genes Dev. 12: 956–967.
Sturchler C., Westhof E., Carbon P., and Krol A. 1993. Unique secondary and tertiarystructural features of the eukaryotic selenocysteine-tRNASec. Nucleic Acids Res. 21:1073–1079.
Sung D. and Kang H. 1998. Mutational analysis of the RNA pseudoknot involved in efficient ribosomal frameshifting in simian retrovirus-1. Nucleic Acids Res. 26:1369–1372.
ten Dam E.B., Pleij C.W.A., and Bosch L. 1990. RNA pseudoknots; translationalframeshifting and readthrough on viral RNAs. Virus Genes 4: 121–136.
Tewari D.S., Qian Y., Thornton R.D., Pieringer J., Taub R., Mochan E., and Tewari M.1994. Molecular cloning and sequencing of a human cDNA encoding ornithine decar-boxylase antizyme. Biochim. Biophys. Acta 1209: 293–295.

672 J.F. Atkins et al.
Tormay P. and Böck A. 1997. Barriers to heterologous expression of a selenoprotein genein bacteria. J. Bacteriol. 179: 576–582.
Tormay P., Wilting R., Heider J., and Böck A. 1994. Genes coding for the selenocysteine-inserting tRNA species from Desulfomicrobium baculatum and Clostridium ther-moaceticum: Structural and evolutionary implications. J. Bacteriol. 176: 1268–1274.
Tormay P., Wilting R., Lottspeich F., Mehta P. Christen P., and Böck A. 1998. Bacterialselenocysteine synthase: Enzyme properties. Eur. J. Biochem. 254: 655–661.
Tsuchihashi Z. and Brown, P.O. 1992. Sequence requirements for efficient translationalframeshifting in the Escherichia coli dnaX gene and the role of an unstable interactionbetween tRNALys and an AAG lysine codon. Genes Dev. 6: 511–519.
Tsuchihashi Z. and Kornberg A. 1990. Translational frameshifting generates the γ subunitof DNA polymerase III holoenzyme. Proc. Natl. Acad. Sci. 87: 2516–2520.
Tu C., Tzeng T.-H., and Bruenn J.A. 1992. Ribosomal movement impeded at a pseudo-knot required for frameshifting. Proc. Natl. Acad. Sci. 89: 8636–8640.
Tu G.F., Reid G.E., Zhang J.G., Moritz R.L., and Simpson R.J. 1995. C-terminal exten-sion of truncated recombinant proteins in Escherichia coli with 10Sa RNA decapep-tide. J. Biol. Chem. 270: 9322–9326.
Ushida C., Himeno H., Watanabe T., and Muto A. 1994. tRNA-like structures in 10SaRNAs of Mycoplasma capricolum and Bacillus subtilis. Nucleic Acids Res. 22:3392–3396.
Veres Z., Tsai L., Scholz T.D., Polotino M., Balaban R.S., and Stadtman T.C. 1992. Syn-thesis of 5-methylaminomethyl-2–selenouridine in tRNAs: 31P NMR studies show thelabile selenium donor synthesized by the selD gene product contains selenium bondedto phosphorus. Proc. Natl. Acad. Sci. 89: 2975–2979.
Walczak R., Carbon P., and Krol A. 1998. An essential non-Watson-Crick base pair mo-tif in 3�UTR to mediate selenoprotein translation. RNA 4: 74–84.
Weiner A.M. and Weber K. 1973. A single UGA functions as a natural termination signalin the coliphage Qβ coat protein cistron. J. Mol. Biol. 80: 837–855.
Weiss R.B., Huang W.M., and Dunn D.M. 1990a. A nascent peptide is required for ribo-somal bypass of the coding gap in bacteriophage T4 gene 60. Cell 62: 117–126.
Weiss R.B., Dunn D.M., Atkins J.F., and Gesteland R.F. 1987. Slippery run, shifty stops,backward steps, and forward hops: –2, –1, +1, +2, +5 and +6 ribosomal frameshifting.Cold Spring Harbor Symp. Quant. Biol. 52: 687–693.
———. 1990b. Ribosomal frameshifting from –2 to +50 nucleotides. Prog. Nucleic AcidRes. Mol. Biol. 39: 159–183.
Weiss R.B., Dunn D.M., Dahlberg A.E., Atkins J.F., and Gesteland R.F. 1988. Readingframe switch caused by base-pair formation between the 3� end of 16S rRNA and themRNA during elongation of protein synthesis in Escherichia coli. EMBO J. 7:1503–1507.
Weiss R.B., Dunn D.M., Shuh M., Atkins J.F., and Gesteland R.F. 1989. E. coli ribosomesre-phase on retroviral frameshift signals at rates ranging from 2 to 50 percent. NewBiol. 1: 159–169.
Wickner R.B. 1989. Yeast virology. FASEB J. 3: 2257–2265.Williams K.P. and Bartel D.P. 1996. Phylogenetic analysis of tmRNA secondary struc-
tures. RNA 2: 1306–1310. Wills N.M., Gesteland R.F., and Atkins J.F. 1991. Evidence that a downstream pseudo-
knot is required for translational read-through of the Moloney murine leukemia virusgag stop codon. Proc. Natl. Acad. Sci. 88: 6991–6995.

Recoding 673
———. 1994. Pseudoknot-dependent read-through of retroviral gag termination codons:Importance of sequences in the spacer and loop 2. EMBO J. 13: 4137–4144.
Wilting R., Vamvakidou K., and Böck A. 1998. Functional expression in Escherichia coliof the Haemophilus influenzae gene coding for selenocysteine-containing selenophos-phate synthetase. Arch. Microbiol. 169: 71–75.
Wilting R., Schorling S., Persson B.C., and Böck A. 1997. Selenoprotein synthesis in ar-chaea: Identification of an mRNA element of Methanococcus jannaschii probably di-recting selenocysteine insertion. J. Mol. Biol. 266: 637–641.
Woese C.R. 1970. Molecular mechanics of translocation: A reciprocating ratchet mecha-nism. Nature 226: 817–820.
Wong J.T.-F. 1975. A co-evolution theory of the genetic code. Proc. Natl. Acad. Sci. 72:1909–1912.
———. 1988. Evolution of the genetic code. Microbiol. Sci. 5: 174–181.Yarus M. and Smith D. 1995. tRNA on the ribosome. In tRNA: Structure, biosynthesis and
function (ed. D. Söll and U.L. RajBhandary), pp. 443–469. ASM Press, Washington,D.C.
Yelverton E., Lindsley D., Yamauchi P., and Gallant J. 1994. The function of a ribosomalframeshifting signal from human immunodeficiency virus-1 in Escherichia coli. Mol.Microbiol. 11: 303–313.
Yueh A. and Schneider R.J. 1996. Selective translation initiation by ribosome jumping inadenovirus-infected and heat-shocked cells. Genes Dev. 10: 1557–1567.
Zerfass K. and Beier H. 1992. Pseudouridine in the anticodon GψA of plant cytoplasmictRNATyr is required for UAG and UAA suppression in the TMV-specific context. Nu-cleic Acids Res. 20: 5911–5918.
Zhang B. and Cech T.R. 1997. Peptide bond formation by in vitro selected ribozymes. Na-ture 390: 96–100.
Zinoni F., Birkmann A., Stadtman T.C., and Böck A. 1986. Nucleotide sequence and ex-pression of the selenocysteine-containing polypeptide of formate-dehydrogenase (formate-hydrogen-lyase-linked) from Escherichia coli. Proc. Natl. Acad. Sci. 83:4560–4564.






























