Embed Size (px)
Citation preview

Tumor and Stem Cell Biology
JAK-STATBlockade Inhibits Tumor Initiation andClonogenicRecovery of Prostate Cancer Stem-like Cells
Paula Kroon1, Paul A. Berry1, Michael J. Stower3, Greta Rodrigues4, Vincent M. Mann5, Matthew Simms2,5,Deepak Bhasin6, Somsundaram Chettiar6, Chenglong Li6, Pui-Kai Li6, Norman J. Maitland1, andAnne T. Collins1
AbstractInterleukin (IL)-6 overexpression and constitutive STAT3 activation occur inmany cancers, including prostate
cancer. However, their contribution to prostate stem and progenitor cells has not been explored. In this study,we show that stem-like cells from patients with prostate cancer secrete higher levels of IL-6 than theircounterparts in non-neoplastic prostate. Tumor grade did not influence the levels of expression or secretion.Stem-like and progenitor cells expressed the IL-6 receptor gp80 with concomitant expression of pSTAT3.Blockade of activated STAT3, by either anti-IL-6 antibody siltuximab (CNTO 328) or LLL12, a specific pSTAT3inhibitor, suppressed the clonogenicity of the stem-like cells in patients with high-grade disease. In a murinexenograft model used to determine the in vivo effects of pSTAT3 suppression, LLL12 treatment effectivelyabolished outgrowth of a patient-derived castrate-resistant tumor. Our results indicate that the most primitivecells in prostate cancer require pSTAT3 for survival, rationalizing STAT3 as a therapeutic target to treat advancedprostate cancer. Cancer Res; 73(16); 5288–98. �2013 AACR.
IntroductionProstate cancer is the most common cancer in men (1).
Although the disease often responds initially to androgen-ablation therapy, the cancer recurs in the majority of patients.For these patients with castration-resistant disease, few ther-apeutic options exist, thus novel therapies are urgentlyrequired. There is accumulating evidence that the inflamma-tory cytokine interleukin (IL)-6 is involved in the etiology ofprostate cancer. IL-6 serum levels are elevated in patients withadvanced prostate cancer (2), and levels correlate with tumorburden and poor prognosis (3, 4). The ligand and receptor areexpressed in both premalignant andmalignant prostatic tissue(5, 6) suggesting that the pathway is constitutively active.Accordingly, inhibition of receptor binding using an antibodyspecific for the ligand-specific subunit gp80 causes regressionof prostate tumors in vivo (7, 8), showing the potential of IL-6/IL-6 receptor interactions as a cancer therapy. IL-6 receptors
(IL-6R) are composed of an IL-6–specific receptor subunit(gp80) and a signal transducer, gp130 (9). IL-6 uses Janus-activated kinase (JAK)-STAT as mediators of signal transduc-tion (10), primarily STAT3 in prostate cancer (11). Constitu-tively activated STAT3 has been found inmany types of cancer,including leukemia, (12) squamous cell carcinoma of the headand neck (HNSCC; ref. 13), multiple myeloma (14), breast (15,16), and prostate cancer (11). Introduction of an activatingmutant of STAT3 is sufficient to induce transformation ofimmortalized cells (17), suggesting that the JAK-STAT signalingpathway, through STAT3 activation, is sufficient in mediatingtumorigenesis. More recent studies have suggested that STAT3activation is important for the tumorigenic ability of cancerstem cells in glioblastoma (18), lung (19), and colon cancer (20).
Cancer stem cells have been isolated from a wide range ofcancers including leukemia (21), breast (22), brain (23, 24), lung(25), colon (26), pancreas (27), and prostate (28). In prostatecancer, cells with a basal phenotype, CD44þ/a2b1hi/CD133þ
(28), CD49fhiTrop2hi (29, 30), and CD44þCD49fhiTrop2þ (31)have many of the properties of cancer stem cells. Gene expres-sion profiling of CD44þ/a2b1hi/CD133þ cancer cells, fromprimary cultures, revealed significant over-representation ofseveral components of the JAK-STAT signaling pathway (32)providing further evidence of the role of this signaling pathwayin prostate cancer.
Current studies on the prostate have failed to account for theheterogeneity at the cellular level as well as the differencesbetween patients. The aimof this studywas to examine the roleof IL-6 on the fate of stem-like cells derived from patients withprostate cancer. Here, we show that the most undifferentiatedcells in cancers express elevated levels of IL-6 and the IL-6–specific receptor. We also show that the JAK-STAT signaling
Authors' Affiliations: 1Yorkshire Cancer Research Unit, Department ofBiology; 2Hull York Medical School, University of York; 3Department ofUrology, York Hospital NHS Trust, York; 4Department of Pathology, HullRoyal Infirmary, Hull; 5Department of Urology, Castle Hill Hospital (Hull &East Yorkshire Hospitals NHS Trust), Cottingham, United Kingdom; and6Division of Medicinal Chemistry and Pharmacognosy, College of Phar-macy, The Ohio State University, Columbus, Ohio
Note: Supplementary data for this article are available at Cancer ResearchOnline (http://cancerres.aacrjournals.org/).
Corresponding Author: Anne T. Collins, Yorkshire Cancer ResearchUnit, Department of Biology, University of York, Heslington, York YO105DD, United Kingdom. Phone: 44-01904-328708; Fax: 44-01904-328710;E-mail: [email protected]
doi: 10.1158/0008-5472.CAN-13-0874
�2013 American Association for Cancer Research.
CancerResearch
Cancer Res; 73(16) August 15, 20135288
on April 25, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2013; DOI: 10.1158/0008-5472.CAN-13-0874
pathway is constitutively active in themost primitive cells, andlastly that inhibition of the pathway significantly decreasescolony-forming ability in vitro and tumor-initiation in immu-nocompromised mice.
Materials and MethodsJAK-STAT inhibitors and CNTO 328The pan-JAK inhibitor 2-tert-butyl-9-fluoro-3,6-dihydro-7H-
benz[h]-imidaz[4,5-f]isoquinoline-7-one, pyridine 6 (P6) wasobtained from Calbiochem. The monoclonal antibody CNTO328 (siltuximab) was obtained from Johnson & Johnson. CNTO328 is a chimeric (murine-human) IgG1 monoclonal antibodyagainst IL-6, which blocks ligandbinding. LLL12wasdevelopedas previously described (33). LLL12 binds directly to thephosphoryl tyrosine 705 (pTyr705)–binding site of the STAT3monomer.
Tissue collection, isolation, and culture of tumor cellsHuman prostate tissue was obtained with patients' consent
and full ethical approval from patients undergoing radicalprostatectomy and channel transurethral resection (TURP)for prostate cancer and from patients undergoing TURP of theprostate for benign prostatic hyperplasia (BPH; Table 1). Gradeand stage of tumor were confirmed by histologic examination,by an uropathologist, of representative fragments. Epithelialcultures were prepared and characterized as described previ-ously (34). Cell cultures were maintained in stem cell media(SCM) consisting of keratinocyte growth medium supple-mented with (EGF, bovine pituitary extract; Invitrogen Ltd.)2 ng/mL stem cell factor [First Link (UK) Ltd.], 100 ng/mLcholera toxin (Sigma-Aldrich Company Ltd.), and 1 ng/mLgranulocyte macrophage colony-stimulating factor [GM-CSF;First Link (UK) Ltd.]. Cells were cultured in the presence ofirradiated (60 Gy) STO (mouse embryonic fibroblasts) cells.After expansion, a2b1hi/CD133þ (stem-like), a2b1hi/CD133�
(transit-amplifying), and a2b1low (committed basal) cells wereisolated by magnetic activated cell sorting (MACS) (MiltenyiBiotec), as described previously (35).
Generation of xenografts and isolation of tumor cellsAll animal work was approved by the University of York
(York, United Kingdom) Animal Procedures and Ethics Com-mittee and conducted under a United Kingdom Home OfficeLicense. Human prostatic tissue was obtained, with informedconsent, from an 83-year-old patient undergoing palliativechannel TURP for advanced prostate cancer. At diagnosis, thetumor was graded Gleason 4þ5 and the patient was placedon androgen-ablation therapy. At xenotransplantation, thepatient was castrate-resistant.The fresh sample was cut into 5-mm sections, mixed with
10% Matrigel at 4�C (BD Biosciences), and immediatelyimplanted into the subcutaneous tissues of 2 BALB/c/Rag2�/�gC�/� mice. This strain was chosen as human cellscan be engrafted with high efficiency, as the mice are deficientin T, B, and natural killer cells (36). Mice were bred in ourfacility and were housed in individually ventilated cages. Oncetumors reached 1.5 cm3, which was considered a humaneendpoint, the mice were sacrificed and the tumors were either
reimplanted into further mice or the tissue was digested forfurther experiments. Tumor latency was initially 93 days, butreduced to 45 days with serial transplantation. Tomaintain thetumor xenograft as "near-patient," tumors were reestablishedfrom frozen cells after five passages in mice.
To generate single cells from xenografts, tumors wereminced into small cubes (3 mm3) and incubated in RPMI-1640 (Invitrogen) containing 5% fetal calf serum (FCS; Invitro-gen) and collagenase type I at 200 IU/mL (Lorne Laboratories)for 20 hours at 37�C. Cells were washed in Dulbecco "A" PBS(Oxoid Ltd.) and further disrupted by trituration through a 21Gblunt needle (Scientific Laboratory Supplies Ltd.). The cellsuspension was then incubated in 0.05% trypsin/EDTA for
Table 1. Clinical and pathologic characteristicsof patient population
Samplenumber Age, y Stage
Gleasonscore
Hormonestatus
1 59 — 8/9 Naïve2 63 — 8 Naïve3 65 — 8 Naïve4 68 — 8 Naïve5 64 — 6 Naïve6 79 — 6 Naïve7 56 — 6 Naïve8 47 — 6 Naïve9 70 — 6 Naïve10 62 — 7 Naïve11 57 — 9 Naïve12 — T3a 9 Sensitive13 64 — 7 Sensitive14 81 — 9 Castrate-Resistant15 72 — 8 Castrate-Resistant16 85 — 9 Castrate-Resistant17 54 — 9 Castrate-Resistant18 80 — 10 Castrate-Resistant19 68 — — Naïve20 59 — 6 Naïve21 — T3a 7 Naïve22 65 T2c 7 Naïve23 — T3a 9 Naïve24 — T3a 7 Naïve25 — T2c 7 Naïve26 — T2b 7 Naïve27 — T2c 7 Naïve28 — T3a 7 Naïve29 68 — 7 Naïve30 64 Tx 7 Naïve31 — T2c 9 Naïve32 — Tx 7 Sensitive33 61 — 9 Castrate-resistant34 56 — 9 Sensitive35 — T3 9 Castrate-resistant36 — T3a 8 Naïve37 58 T2c 7 Naïve
STAT3 Blockade Reduces Cancer Stem Cell Frequency
www.aacrjournals.org Cancer Res; 73(16) August 15, 2013 5289
on April 25, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2013; DOI: 10.1158/0008-5472.CAN-13-0874

30 minutes at 37�C, passed through a 70-mm cell strainer and aFicoll gradient (Ficoll-Paque Plus; GEHealthcare Life Sciences)to further enrich for viable cells. Mouse lineageþ and endo-thelial cells (CD31þ) were depleted by indirectly labeling withbiotinylated anti-lineageþ antibody cocktail (Miltenyi Biotec)andCD31 (ER-MP12; 1:20 dilution; AbD Serotec) for 10minutesat 4�C. Anti-biotin antibodies conjugated to Microbeads wereused as secondary labeling reagent and Lin�/CD31� cells werecollected according to the manufacturer's instructions (Milte-nyi Biotec).
Quantitative real-time PCRTotal RNA was extracted from sorted a2b1hi/CD133þ,
a2b1hi/CD133�, and a2b1low cells, derived from malignant andnonmalignant primary cultures, using Qiagen RNease mini-columns, according to the manufacturer's protocol. RNA wasreverse transcribed, using random hexamers (Invitrogen) andreverse transcriptase (SuperScript II; Invitrogen). Real-timePCR (RT-PCR) was conducted using TaqMan gene expressionpresynthesized reagents andmastermix (Applied Biosystems).Reactions were prepared following the manufacturer's proto-col. All reactions were carried out in triplicate on 96-well PCRplates (ABI PRISM; Applied Biosystems) in an ABI PRISM 7000sequence detection system (Applied Biosystems). Standardthermal cycling conditions included a hot start of 5 minutesat 50�C and 10 minutes at 95�C, followed by up to 50 cycles of95�C for 15 seconds and 60�C for 1 minute. Data analysis wasconducted using Microsoft Excel software. Expression valuesare presented relative to the endogenous control gene, RPLP0.
IL-6 ELISASorted a2b1hi/CD133þ, a2b1hi/CD133�, and a2b1low cells,
from malignant and nonmalignant prostate primary cultures,were plated (in triplicate) onto 96-well collagen-coated plates(BD-Biocoat; BD Biosciences) at a density of 1 � 103 cells,together with 4 � 104 irradiated STO cells. Conditioned medi-umwas collected over 48 hours and was immediately stored at�20�C. The concentrations of secreted IL-6 in the conditionedmedia were measured by using the R&D Systems' QuantikineELISA Kit, according to the manufacturer's instructions. Brief-ly, conditioned media or IL-6 standards were incubated for 2hours at room temperature in 96-well polystyrenemicroplates,coated with a mouse monoclonal antibody against IL-6. Afterwashing, an antibody against IL-6 conjugated to alkalinephosphatase was added and incubated for 2 hours at roomtemperature. A further wash step was conducted, substrateand amplifier was added, and the plate was read at 485 nm.
Cell-based ELISA for detection of STAT3A cell-based ELISA was used to measure the levels of
phosphorylated STAT3 (Tyr705) and total STAT3 simulta-neously in primary prostatic cell cultures (R&D Systems),according to the manufacturer's protocol. Briefly, primaryprostate cells (1 � 104) were plated overnight, in triplicate,onto a black collagen-I–coated 96-well plate, washed, and fixedwith 4%paraformaldehyde (PFA; w/v). Cellswere subsequentlyincubated with primary antibodies followed by horseradishperoxidase (HRP)- and alkaline phosphatase (AP)-conjugated
secondary antibodies. Relative fluorescence units (RFU) ofphosphorylated STAT3 and total STAT3 were then measured.
Flow cytometryLin�/CD31� tumor cells (isolated from xenografts) were
incubated with antibodies to CD44 (clone C26; BD Pharmin-gen) and CD24 (clone 32D12; Miltenyi Biotec), or an isotypecontol (Miltenyi Biotec), for 10 minutes at 4�C. Dead cells wereexcluded using SYTOX Blue (Invitrogen).
For the detection of pSTAT3 and androgen receptor, Lin�
/CD31� cells were incubatedwith Live/Dead stain (Invitrogen)before labeling with cell surface CD44 and CD24 (as above).Cells were then fixed with 1.5% formaldehyde (v/v) for 5minutes at room temperature, permeabilized with ice-coldmethanol for 10 minutes at 4�C, and finally incubated withanti-phospho-STAT3 antibody [Tyr705 (1:10); Cell SignalingTechnology] or androgen receptor (Clone 441; Santa CruzBiotechnology, Inc.), for 30 minutes at 4�C. All cells wereanalyzed on a CyAn ADP flow cytometer (DakoCytomation)and data processed using Summit v4.3 software (BeckmanCoulter Ltd.).
Histochemistry, immunohistochemistry, andimmunocytochemistry
Histochemistry was conducted on formalin-fixed, paraffin-embedded mouse xenografts. Sections were stained withhematoxylin and eosin and classified according to Gleasongrading (37). Immunohistochemistry was conducted on for-malin-fixed, paraffin-embedded normal adult prostate. Anti-gen retrieval with citric acid (pH 6.0), at 97�C for 30 minutes,was followed by incubation with antibodies to IL-6R (rabbitpolyclonal C-20; 1:100; Santa Cruz Biotechnology, Inc.), CK5(mouse monoclonal XM26; 1:100; Vector Laboratories Ltd.), oran isotype control. For the detection of IL-6R, anti-rabbitconjugated to Alexa Fluor 488 was used and anti-mouse AlexaFluor 586 was used for the detection of CK5 (Molecular Probes;Invitrogen). Sections were counterstainedwith 40,6-diamidino-2-phenylindole (DAPI) and mounted in Vectashield (VectorLaboratories Ltd.) and visualized using a Zeiss LSM510 metaconfocal microscope.
Immunocytochemistry was conducted on sorted a2b1hi/CD133þ, a2b1hi/CD133�, and a2b1low primary cells. The cellisolates were allowed to adhere for 2 to 3 hours, onto collagen-coated slides, blocked with 10% serum and incubated withprimary antibody against IL-6R (rabbit polyclonal C-20; 1:100)or an isotype control for 1 hour. Cells were then fixed with 4%PFA, washed twice with 50 mmol/L NH4Cl, and visualizedusing anti-rabbit Alexa Fluor 488 and DAPI.
Western blot analysis for detection of phosphorylatedSTAT3
Cell lysates were prepared, from primary prostate cultures,by adding lysis buffer (CytoBuster Protein Extraction Reagent;Novagen;Merck Chemicals Ltd.) to culture plates for 5minutesat room temperature. Complete protease inhibitor (EDTA-freeProtease Inhibitor Cocktail Tablet; Roche) and phosphataseinhibitors (Phosphatase Inhibitor Cocktail; Sigma) were thenadded. Equal amounts of proteinwere resolved on a 10% to 12%
Kroon et al.
Cancer Res; 73(16) August 15, 2013 Cancer Research5290
on April 25, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2013; DOI: 10.1158/0008-5472.CAN-13-0874

SDS-PAGE gel and transferred to polyvinylidene difluoride(PVDF) Immobilon-Pmembrane (Millipore). Membranes wereblocked with 5% semidried milk for 1 hour and then incubatedwith primary antibodies against phospho-STAT3 (Tyr705;1:500), STAT3 (1:1,000; Cell Signaling Technology), and b-actin(1:10,000; Sigma). Primary antibodies were incubated over-night at 4�C or 1 hour at room temperature followed byincubation with goat anti-mouse immunoglobulin G (IgG)immunoglobulins-HRP or goat anti-rabbit IgG immunoglobu-lins-HRP secondary antibodies. Membranes were visualizedusing BM Chemiluminescence Western Blotting Substrate(POD; Roche) and enhanced chemiluminescence (ECL) Hyper-film (GE Healthcare Life Sciences).
Clonogenic recoveryPrimary prostate cultures were treated with either 5 mmol/L
P6 for 16 hours, 10 mg/mL CNTO 328 for 6 days, or 1 mmol/LLLL12 for 24 hours. CD133þ (stem-like) and CD133� (progen-itor) cells were then isolated, counted, and plated on to 35-mmcollagen-coated plates (BD Biocoat; BD biosciences) at a celldensity of 100 cells per well in the presence of irradiated STOfeeders. Colonies were subsequently scored if they containedmore than 32 cells (5 population doublings) usually between 14to 28 days after treatment.
Tumor initiationMouse depleted tumor cells (Lin�/CD31�) were treated
overnight with 0.5 to 10 mmol/L LLL12 or vehicle control. Thecells were counted and injected subcutaneously into 6- to 8-week-old male Rag2�/�gC�/� mice at limiting dilutions,together with 2 � 105 irradiated STO feeder cells in Matrigel.
The mice were monitored until the tumors reached 1.5 cm3 orup to 120 days.
Statistical analysisStudent t test and Kaplan–Meier survival analyses were
calculated using Sigma Plot. The log-rank test was used tocompare Kaplan–Meier curves. The x2 tests (to determinetumor initiation frequencies and pairwise tests) were calcu-lated using ELDA software (38).
ResultsIL-6 expression is highest in undifferentiated, hormone-na€�ve, cancer cells
Primary cultures were generated from tumor tissues of 11patients with prostate cancer at Gleason grade 6 and above(who had not undergone treatment before biopsy; samples 1–11; Table 1) and 7 patients who had undergone androgen-ablation therapy (samples 12–18; Table 1). Primary cultureswere also generated from 11 patients with the benign condi-tion, BPH.
Quantitative RT-PCR (qRT-PCR) analysis revealed that themore undifferentiated, stem-like (CD44þ/a2b1hi/CD133þ) andtransit-amplifying cells (CD44þ/a2b1hi/CD133�) from tumorbiopsies expressed 7- and 6-fold higher levels of IL-6 than themore differentiated (committed basal; CD44þ, a2b1low) cells(Fig. 1A). Because of patient variability, this difference in IL-6expression did not reach statistical significance. The increaseof IL-6 expression in the stem-like and transit-amplifyingpopulation was not observed in patients with either benigndisease or those who had undergone androgen-ablation ther-apy. Within the group of patient who had undergone androgen
BPHA B
SC TA CB SC TA CB SC TA
Se
cre
ted
IL
-6 (
pg
/mL
)
No
rma
lize
d m
RN
A leve
ls
CB SC TA
10
9
8
7
6
5
4
3
2
1
0
150
130
110
90
40
30
20
10
0CB SC TA CB SC TA CB
HNPC HT BPH HNPC HT
Figure 1. IL-6 is highly expressed within the stem-like cells in prostate cancer. A, qRT-PCR analysis of IL-6 on selected cell populations derived frompatients with BPH (n ¼ 5), hormone-naïve (HNPC; samples 1–4), and hormone-sensitive and castrate-resistant prostate cancer (HT; samples 12–16).The results are normalized to the housekeeping gene RPLP0 and the fold difference is relative to the averageCT-value of the committed basal population. B,ELISA for secreted IL-6. The concentration of IL-6was determined in conditionedmediumcollected over 48 hours from1,000 preselected stem-like cells (SC),transit-amplifying (TA), and committed basal (CB) cells. Conditioned medium was collected from BPH (n ¼ 6), HNPC (samples 5–11), and HT patients(samples 12, 14, 17, and 18). Individual samples are represented by different symbols and the bar depicts the average within each group. Statistical analysiswas conducted using the Student t test; �, P < 0.05; ��, P < 0.005.
STAT3 Blockade Reduces Cancer Stem Cell Frequency
www.aacrjournals.org Cancer Res; 73(16) August 15, 2013 5291
on April 25, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2013; DOI: 10.1158/0008-5472.CAN-13-0874

ablation, the levels of IL-6 increased with the differentiation ofthe epithelial cells.
We then assessed the levels of IL-6 secreted from sorted cellpopulations over 48 hours in culture (Fig. 1B). Similarly,significantly higher levels of IL-6 (P < 0.05) were secreted fromthe stem-like population (from patients with cancer) com-pared with those with benign disease and the more differen-tiated cells. In contrast, cultures from patients with benigndisease secreted more IL-6 from the transit-amplifying popu-lation, compared with either stem-like cells or cell populationscommitted to differentiation (P < 0.05).
These data indicate that IL-6 expression is significantlyhigher in more stem-like cells from patients with prostatecancer, but only in those patients who had not undergoneandrogen-ablation therapy, whereas secreted levels of IL-6were significantly higher in patients with cancer, irrespectiveof treatment. These data also show that tumor grade did notinfluence the levels of expression or secretion.
Stem-like cells express the IL-6RWe next looked at the expression of IL-6R (gp80) using a
combination of immunohistochemistry and immunofluores-cence of sorted cell populations (Fig. 2). Analysis of prostatetissue sections showed strong expression within the luminalcells, withweak to absent expressionwithin the basal layer (Fig.2). However, closer examination, at higher magnification,revealed that rare cells within the basal layer, marked withCK5 (a basal cell marker), coexpressed the receptor (Fig. 2A,inset, arrows). To confirm whether those rare cells were thestem-like population, basal cells (derived from 2 patients withBPH and 1 patient with cancer) were sorted into three popula-tions (stem-like cells, transit-amplifying, and committed bas-al), and subsequently stained with antibodies to gp80 Fig. 2B).
We observed that the majority of stem-like cells expressed thereceptor, but the number of cells expressing IL-6R decreasedwith differentiation of the epithelial cells. These results indi-cate that the JAK-STAT signaling pathway could be constitu-tively active (within the stem-like population) through IL-6 andits receptor subunit (gp80).
Activation of STAT3 is mediated through IL-6To investigate whether the JAK-STAT signaling pathway is
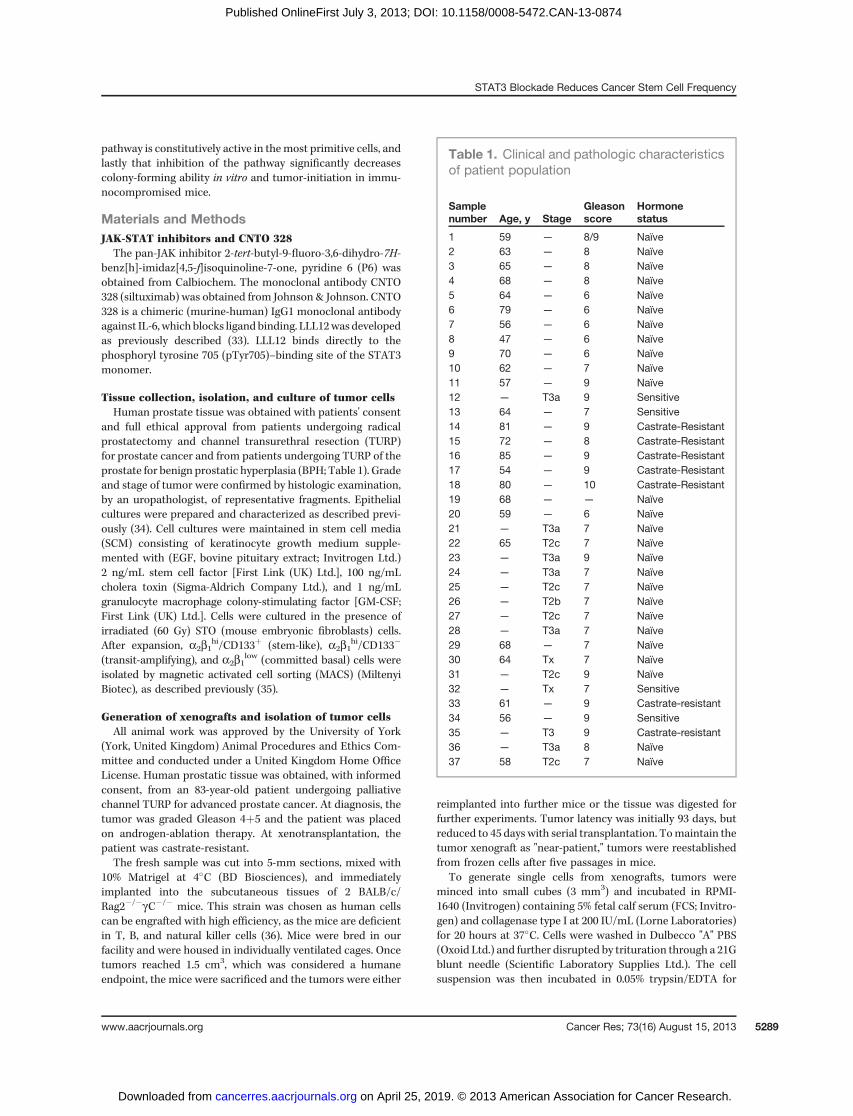
regulated by IL-6 in prostate cancer, we examined the relativelevels of activated STAT3 (rSTAT3-tyrosine 705). Primarycultures were generated from 2 patients with BPH and 5patients with prostate cancer (samples 4, 15, 19–21). Westernblot analysis results revealed that the level of activated STAT3was variable, but pSTAT3 was present in both benign diseaseand cancer (Fig. 3A). Of note, 3 of 5 cancers examined had highlevels ofrSTAT3; all three had aGleason score between 7 and 9.We next investigated whether we could abrogate activation ofSTAT3 using the chemical inhibitor P6. P6 is a specific pan-JAKinhibitor that has been reported to act within the low nano-molar range (IC50, 1–15 nmol/L; ref. 39). Downregulation ofpSTAT3 was observed, in all primary cultures examined, andwas optimal at 5 mmol/L after 16 hours treatment (samples 17,22, and 23; Fig. 3B). Using P6 as a positive control for pSTAT3abrogation, we next treated primary cells [from patients withhormone-na€�ve prostate cancer (HNPC)] with a neutralizingantibody to IL-6, CNTO 328 (siltuximab), to determinewhetherpSTAT3 was similarly downregulated. Siltuximab was effectiveat blocking activation of STAT3, but only after 6 days oftreatment (P < 0.05; samples 22–24; Fig. 3C). There was vari-ability between samples, most likely due to the differing levelsof endogenous IL-6 secretion between individuals (Fig. 1).Steiner and colleagues similarly observed downregulation of
Cancer
Benign
Benign
AStem-like TA
70 µmol/L 70 µmol/L
CBB
TA CBStem-like
Perc
enta
ge o
f IL
–6R
positiv
e c
ells
IL-6R expression in primary prostate cells100
90
80
70
60
50
40
30
20
10
0
20 µmol/L
20 µmol/L
20 µmol/L
Figure 2. IL-6R is expressed by luminal cells and rare stem-like cells. A, immunofluorescence of a paraffin section of normal prostate stained with antibodiesto CK5 (red), a basal marker, and IL-6R (green). The nuclei were counterstained with DAPI (blue). B, IL-6R staining of selected cells [sorted cells, transit-amplifying (TA), and committed basal (CB)]. The percentage of cells of each phenotype expressing the receptor was quantified by counting 10 random fields.Three patient samples (2 BPH and 1 cancer; sample 10) were analyzed and images were taken by confocal microscopy at �63 magnification.
Kroon et al.
Cancer Res; 73(16) August 15, 2013 Cancer Research5292
on April 25, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2013; DOI: 10.1158/0008-5472.CAN-13-0874
pSTAT3 levels after 6 days treatmentwith 10mg/mL siltuximab(40).These results confirmed that pSTAT3 is constitutively active
and activation of the pathway is mediated through IL-6.
Blockadeof STAT3activation suppresses colony-formingabilityWenextwanted to establishwhether prostate stem-like cells
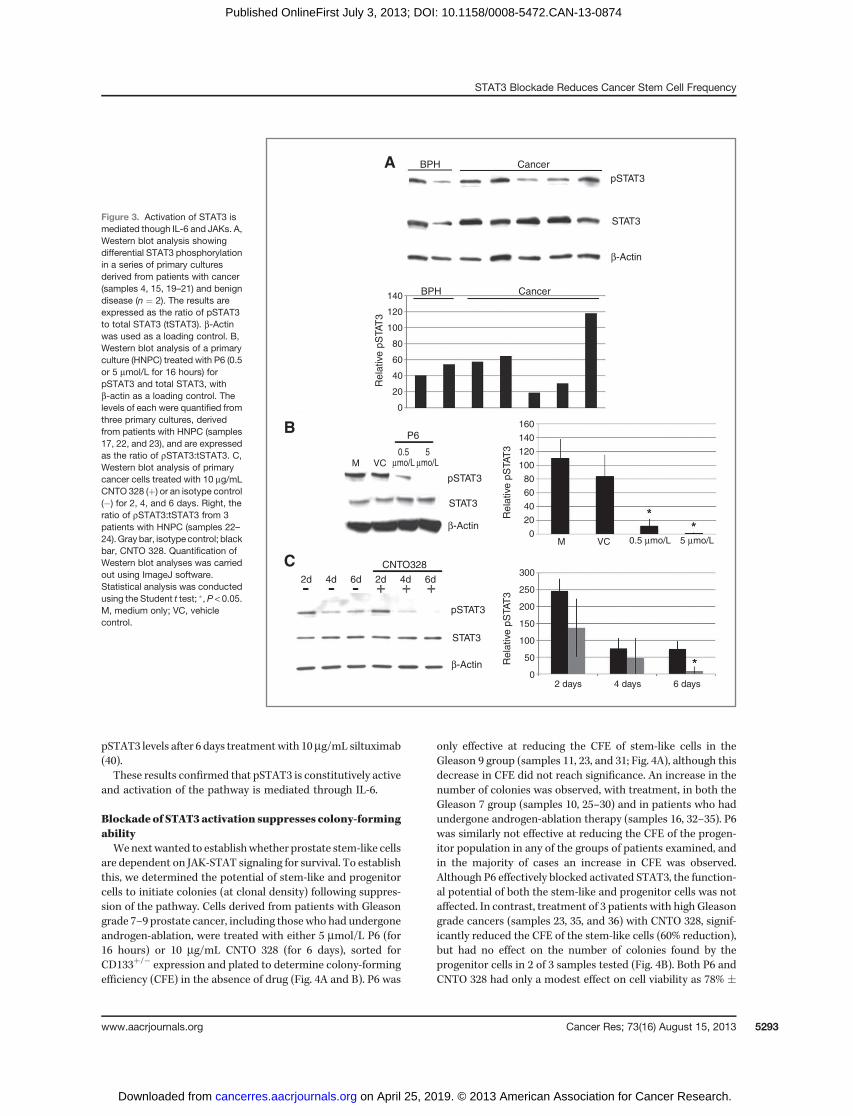
are dependent on JAK-STAT signaling for survival. To establishthis, we determined the potential of stem-like and progenitorcells to initiate colonies (at clonal density) following suppres-sion of the pathway. Cells derived from patients with Gleasongrade 7–9 prostate cancer, including those who had undergoneandrogen-ablation, were treated with either 5 mmol/L P6 (for16 hours) or 10 mg/mL CNTO 328 (for 6 days), sorted forCD133þ/� expression and plated to determine colony-formingefficiency (CFE) in the absence of drug (Fig. 4A and B). P6 was
only effective at reducing the CFE of stem-like cells in theGleason 9 group (samples 11, 23, and 31; Fig. 4A), although thisdecrease in CFE did not reach significance. An increase in thenumber of colonies was observed, with treatment, in both theGleason 7 group (samples 10, 25–30) and in patients who hadundergone androgen-ablation therapy (samples 16, 32–35). P6was similarly not effective at reducing the CFE of the progen-itor population in any of the groups of patients examined, andin the majority of cases an increase in CFE was observed.Although P6 effectively blocked activated STAT3, the function-al potential of both the stem-like and progenitor cells was notaffected. In contrast, treatment of 3 patients with high Gleasongrade cancers (samples 23, 35, and 36) with CNTO 328, signif-icantly reduced the CFE of the stem-like cells (60% reduction),but had no effect on the number of colonies found by theprogenitor cells in 2 of 3 samples tested (Fig. 4B). Both P6 andCNTO 328 had only a modest effect on cell viability as 78% �
BPHA
B
C
Cancer
BPH140
120
100
80
60
40
20
0
M VC
P6
CNTO328
160
140
120
100
80
60
40
20
0
300
250
200
150
100
50
0
Rela
tive
pS
TAT
3
Re
lative
pS
TAT
3R
ela
tive
pS
TAT
3
Cancer
pSTAT3
STAT3
β-Actin
pSTAT3
2d 4d 6d 2d 4d 6d
STAT3
β-Actin
0.5μmo/L
5μmo/L
M
2 days 4 days 6 days
VC 0.5 μmo/L 5 μmo/L
pSTAT3
STAT3
β-Actin
Figure 3. Activation of STAT3 ismediated though IL-6 and JAKs. A,Western blot analysis showingdifferential STAT3 phosphorylationin a series of primary culturesderived from patients with cancer(samples 4, 15, 19–21) and benigndisease (n ¼ 2). The results areexpressed as the ratio of pSTAT3to total STAT3 (tSTAT3). b-Actinwas used as a loading control. B,Western blot analysis of a primaryculture (HNPC) treated with P6 (0.5or 5 mmol/L for 16 hours) forpSTAT3 and total STAT3, withb-actin as a loading control. Thelevels of each were quantified fromthree primary cultures, derivedfrom patients with HNPC (samples17, 22, and 23), and are expressedas the ratio of rSTAT3:tSTAT3. C,Western blot analysis of primarycancer cells treated with 10 mg/mLCNTO 328 (þ) or an isotype control(�) for 2, 4, and 6 days. Right, theratio of rSTAT3:tSTAT3 from 3patients with HNPC (samples 22–24).Gray bar, isotype control; blackbar, CNTO 328. Quantification ofWestern blot analyses was carriedout using ImageJ software.Statistical analysis was conductedusing the Student t test; �, P < 0.05.M, medium only; VC, vehiclecontrol.
STAT3 Blockade Reduces Cancer Stem Cell Frequency
www.aacrjournals.org Cancer Res; 73(16) August 15, 2013 5293
on April 25, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2013; DOI: 10.1158/0008-5472.CAN-13-0874

15.9% of cells were viable after 16 hours in the presence of P6and 91% � 9.5% of cells were viable in the presence of CNTO328 (results not shown). These results show that IL-6 isrequired for survival of stem-like cells in prostate cancer, butthat the JAK-STATpathwaymay not be involved as P6 had littleeffect on reducing colony-forming activity. To address this, wetreated cells with a novel pSTAT3 inhibitor, LLL12, which bindsspecifically to the phosphoryl tyrosine 705 (pTyr705)–bindingsite of the STAT3 monomer (41). Treatment with LLL12prevented phosphorylation of STAT3 at a concentration of 1mmol/L. P6 was used as a positive control and similarlysuppressed pSTAT3 (sample 37; Fig. 4C). We then determinedthe functional effect of suppressing pSTAT3 following treat-ment with LLL12. Primary cultures from high Gleason gradecancers (samples 23, 33, 35, and 36) were treated with 1mmol/LLLL12 for 24 hours. We observed that, unlike CNT0328 and P6,LLL12 treatment significantly affected cell viability. The num-
ber of cells remaining after treatment was approximately 40%� 26% (results not shown). The viable cells were then plated todetermine whether CFE was similarly affected. Unlike treat-ment with P6, we found a significant and marked reduction inthe number of colonies in all patients tested (n¼ 4) and in bothpopulations (Fig. 4D). These data suggest that targeting con-stitutive STAT3 in prostate cancer maybe an attractive ther-apeutic option.
STAT3 blockade abolishes tumor initiationTo provide further preclinical evidence, a murine xenograft
model was used to determine the in vivo effects of pSTAT3suppression. This xenograft was recently derived, in our lab-oratory, from a castrate-resistant patient (Supplementary Fig.S1). pSTAT3 status was determined (by flow cytometry) ondissociated tumor cells [depleted of mouse lineageþ hemato-poietic and endothelial cells (Lin�/CD31�)]. The majority of
Gleason 7A B
C D
Gleason 9
Stem-like ProgenitorStem-like Progenitor Stem-like Progenitor Stem-like Progenitor
Stem-like Progenitor
HT
Re
lative
CF
E (
%)
Re
lative
CF
E (
%)
1.8
1.6
1.4
1.2
1
0.8
0.6
0.4
0.2
0
Rela
tive
CF
E (
%)
Rela
tive
pS
TAT
3
1.8
1.6
1.4
1.2
1
0.8
0.6
0.4
0.2
0
60
50
40
30
20
10
0
7
6
5
4
3
2
1
0
M VC P6
10μmo/L
5μmo/L
1μmo/L
M VC P6
LLL12
10μmo/L
5μmo/L
1μmo/L
pSTAT3
STAT3
β-Actin
Figure 4. STAT3 blockade suppresses colony-forming ability. Primary prostate cells derived from patients with either Gleason grade 7HNPC (samples 10, 25–30), Gleason 9HNPC (samples 11, 23, and 31), or HT [which included hormone-sensitive and castrate-resistant patients (samples 16, 32–35)] were treated for16 hours with 5 mmol/L P6 (A) or 10 mg/mL CNTO 328 for 6 days (samples 23, 35, and 36; B). CD133þ (stem-like) and CD133� (progenitor) cells weresubsequently isolated and plated (at clonal density) in the presence of irradiated STO feeder cells to determine CFE. Colonies were scored if they hadundergone five ormore population doublings (32 cells), approximately 14 days after plating. The results are expressed as relative to theCFE of the nontreatedcells (1%). Each circle represents an individual patient and the bar is the average of each group. C, Western blot analysis of a representative primary culture(sample 37) treated overnight with increasing concentrations of LLL12. D, relative CFE following treatment of stem-like and progenitor cells with 1 mmol/LLLL12 for 24 hours. Primary cultures were derived from patients with Gleason 8/9 disease and included patients who had undergone androgen-ablationtherapy (samples 23, 33, 35, and 36). Statistical analyses was conducted using the Student t test; �, P < 0.05.
Kroon et al.
Cancer Res; 73(16) August 15, 2013 Cancer Research5294
on April 25, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2013; DOI: 10.1158/0008-5472.CAN-13-0874
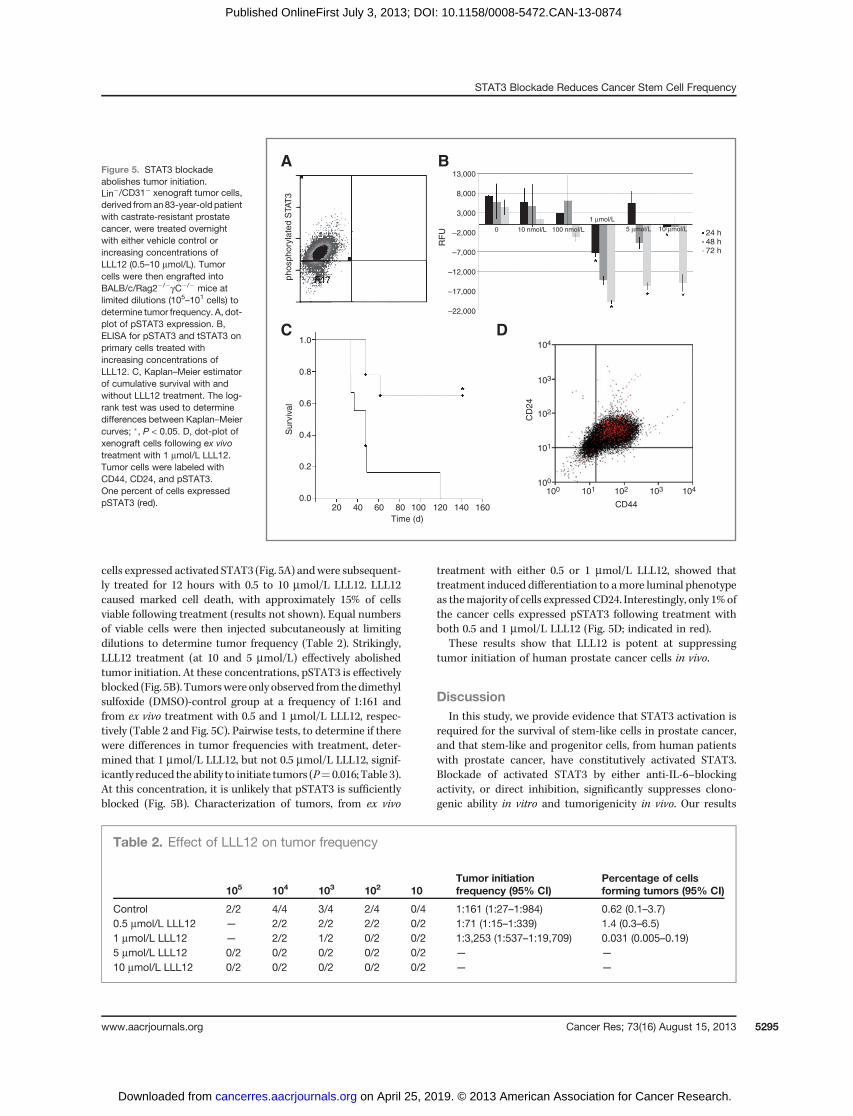
cells expressed activated STAT3 (Fig. 5A) andwere subsequent-ly treated for 12 hours with 0.5 to 10 mmol/L LLL12. LLL12caused marked cell death, with approximately 15% of cellsviable following treatment (results not shown). Equal numbersof viable cells were then injected subcutaneously at limitingdilutions to determine tumor frequency (Table 2). Strikingly,LLL12 treatment (at 10 and 5 mmol/L) effectively abolishedtumor initiation. At these concentrations, pSTAT3 is effectivelyblocked (Fig. 5B). Tumorswereonly observed fromthedimethylsulfoxide (DMSO)-control group at a frequency of 1:161 andfrom ex vivo treatment with 0.5 and 1 mmol/L LLL12, respec-tively (Table 2 and Fig. 5C). Pairwise tests, to determine if therewere differences in tumor frequencies with treatment, deter-mined that 1 mmol/L LLL12, but not 0.5 mmol/L LLL12, signif-icantly reduced theability to initiate tumors (P¼ 0.016; Table 3).At this concentration, it is unlikely that pSTAT3 is sufficientlyblocked (Fig. 5B). Characterization of tumors, from ex vivo
treatment with either 0.5 or 1 mmol/L LLL12, showed thattreatment induced differentiation to amore luminal phenotypeas themajority of cells expressed CD24. Interestingly, only 1% ofthe cancer cells expressed pSTAT3 following treatment withboth 0.5 and 1 mmol/L LLL12 (Fig. 5D; indicated in red).
These results show that LLL12 is potent at suppressingtumor initiation of human prostate cancer cells in vivo.
DiscussionIn this study, we provide evidence that STAT3 activation is
required for the survival of stem-like cells in prostate cancer,and that stem-like and progenitor cells, from human patientswith prostate cancer, have constitutively activated STAT3.Blockade of activated STAT3 by either anti-IL-6–blockingactivity, or direct inhibition, significantly suppresses clono-genic ability in vitro and tumorigenicity in vivo. Our results
1.0
0.8
0.6
0.4
0.2
0.0
A B
C D
13,000
8,000
3,000
–2,000
–7,000
–12,000
–17,000
–22,000
24 h
104
103
102
101
100
104103102101100
48 h72 h
0 10 nmol/L
1 μmol/L
5 μmol/L 10 μmol/L100 nmol/L
Surv
ival
phosphory
late
d S
TAT
3
RF
U
20 40 60 80 100 120 140 160
Time (d)
CD44
CD
24
Figure 5. STAT3 blockadeabolishes tumor initiation.Lin�/CD31� xenograft tumor cells,derived froman83-year-oldpatientwith castrate-resistant prostatecancer, were treated overnightwith either vehicle control orincreasing concentrations ofLLL12 (0.5–10 mmol/L). Tumorcells were then engrafted intoBALB/c/Rag2�/�gC�/� mice atlimited dilutions (105–101 cells) todetermine tumor frequency. A, dot-plot of pSTAT3 expression. B,ELISA for pSTAT3 and tSTAT3 onprimary cells treated withincreasing concentrations ofLLL12. C, Kaplan–Meier estimatorof cumulative survival with andwithout LLL12 treatment. The log-rank test was used to determinedifferences between Kaplan–Meiercurves; �, P < 0.05. D, dot-plot ofxenograft cells following ex vivotreatment with 1 mmol/L LLL12.Tumor cells were labeled withCD44, CD24, and pSTAT3.One percent of cells expressedpSTAT3 (red).
Table 2. Effect of LLL12 on tumor frequency
105 104 103 102 10Tumor initiationfrequency (95% CI)
Percentage of cellsforming tumors (95% CI)
Control 2/2 4/4 3/4 2/4 0/4 1:161 (1:27–1:984) 0.62 (0.1–3.7)0.5 mmol/L LLL12 — 2/2 2/2 2/2 0/2 1:71 (1:15–1:339) 1.4 (0.3–6.5)1 mmol/L LLL12 — 2/2 1/2 0/2 0/2 1:3,253 (1:537–1:19,709) 0.031 (0.005–0.19)5 mmol/L LLL12 0/2 0/2 0/2 0/2 0/2 — —
10 mmol/L LLL12 0/2 0/2 0/2 0/2 0/2 — —
STAT3 Blockade Reduces Cancer Stem Cell Frequency
www.aacrjournals.org Cancer Res; 73(16) August 15, 2013 5295
on April 25, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2013; DOI: 10.1158/0008-5472.CAN-13-0874

show that blockade of the JAK-STAT signaling pathway repre-sents an important therapeutic approach for prostate cancer.
Activation of STAT3 has been shown to occur throughbinding of the IL-6 family of cytokines to the gp130 receptorin leukemia (12), HNSCC (13), multiple myeloma (14), breast(15, 16), and prostate cancer (11). Furthermore, high circulat-ing levels of IL-6 have been found in patients with advancedcancer, including prostate cancer (2–4). More recently, amicroarray gene expression analysis of selected cell popula-tions (including stem-like cells from prostate cancer biopsies)found over-representation of several components of the JAK-STAT signaling cascade, including IL-6 (32). The results of thisstudy support those findings, as the highest levels of IL-6 wereobserved in cancer stem-like and progenitor cells. We alsofound that IL-6 levels did not correlate with tumor grade ordisease progression, in contrast to reports that IL-6 levels aresignificantly greater in patients with castrate-resistant pros-tate cancer than in those with hormone-sensitive disease(6, 42). It is proposed that this increase is due to the abolishedsuppression of IL-6 expression by androgens (43). As immunecells are likely to make a significant contribution to circulatingIL-6 levels, it is impossible to compare thisfindingwith those inourmodel.Moreover, primary cultures aremostly composed ofbasal-like cells that do not express active androgen receptor,and will therefore be most likely indirectly influenced byandrogens (28).
Our results indicate that the JAK-STAT signaling pathway isconstitutively active in the stem-like population in prostatecancer as IL-6 and the gp80 receptor were both expressed. Thiswas confirmed by detection of activated STAT3, in all samplesanalyzed. We also established that signaling is mediatedthrough IL-6 in prostate as a blocking antibody against IL-6(CNTO 328) abolished STAT3 activation. The consequences ofSTAT3 blockade included the suppression of colony-formingability, suggesting that cancer stem-like and progenitor cellsrequire pSTAT3 for survival. We found that the small-moleculeinhibitor, LLL12, was more potent at suppressing colonyformation, than either P6 or CNTO 328. Despite blockade ofactivated STAT3, P6 was relatively ineffective at suppressingcolony-forming activity. Indeed, we observed an expansion incolony numbers (in the majority of patients treated with P6).Although LLL12 treatment significantly reduced cell viability,we found that P6 had little effect on viability. Pedranzini andcolleagues showed that P6 was only effective at reducing cellproliferation of myeloma cell lines after 3 days treatment (44).Although we observed a similarly modest reduction in cellviability with CNTO 328 treatment, colony recovery of thestem-like cells was significantly reduced, suggesting that thispopulation is more dependent on IL-6 for survival. However,
direct blockade of pSTAT3, with LLL12, effectively suppressedcolony formation of both stem and progenitor populations (inpatients with high-grade cancer and advanced disease). Ourdata are supported by others in glioma and colon cancer, whopropose that STAT3 is a survival factor for cancer stem-likecells (20, 45). Although the effect with LLL12 was consistent, alarger study should be undertaken to confirm our results.
Siltuximab (CNTO 328) had little effect on survival in recentphase II trials for chemotherapy-treated patients with castra-tion-resistant prostate cancer (46, 47). A dramatic increase inplasma IL-6 was observed following treatment with siltuximab(46) although C-reactive protein (a marker of systemic inflam-mation) declined after treatment. It is possible that theobserved increase in plasma IL-6 was due to a feedback loopin which blockade of pSTAT3 resulted in a concomitantincrease in IL-6 levels. Unfortunately, this did not result inincreased survival, suggesting that either pSTAT3 is notrequired for survival or that there is heterogeneity in expres-sion (of rSTAT3, IL-6, or IL-6R) between patients and thedifferent populations of cancer cells, as shown in our study andby others (5).
Finally, a murine xenograft model was used to determinethe in vivo effects of pSTAT3 suppression. The xenograft tumorwas derived from a patient with castrate-resistant disease andwas chosen for this study as LLL12 was effective at reducingcolonies from high-grade, advanced tumors, and we observedthat the majority of cells expressed activated STAT3. Ourresults confirm the in vitro data, as tumor initiation wasabolished at concentrations of LLL12 that effectively blockedactivated STAT3 and supports findings that STAT3 is a survivalfactor in a number of solid tumors. These data also show thatthe phenotype of the tumor-initiating cell is reminiscent of anintermediate cell as both CD44 and androgen receptor isexpressed. Further work has to be undertaken on sorted cellpopulations to confirm this. We also found that tumorsgenerated following ex vivo treatment with either 0.5 or 1mmol/L LLL12 were largely differentiated as the majority ofcells expressed CD24 and lacked expression of pSTAT3. Wepropose that those tumors would not effectively serially trans-plant and would be more amenable to combination therapy.
In conclusion, we propose that targeting the JAK-STATpathway is worthy of consideration for advanced prostatecancer. Despite the disappointing results with siltuximab, theevidence from this study and others supports further investi-gation. It is vital, however, when designing clinical trials thatpatients are stratified for their likely response to STAT3blockade.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: P. Kroon, C. Li, P.-K. Li, N.J. Maitland, A.T. CollinsDevelopment of methodology: P. Kroon, C. Li, A.T. CollinsAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): P. Kroon, P.A. Berry, M.J. Stower, G. Rodrigues, V.M.Mann, M. Simms, D. Bhasin, S. ChettiarAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): P. Kroon, N.J. Maitland, A.T. Collins
Table 3. Pairwise test to determine differencesin tumor frequencies
c2 Pairwise test Control
0.5 mmol/L LLL12 0.5221 mmol/L LLL12 0.016
Kroon et al.
Cancer Res; 73(16) August 15, 2013 Cancer Research5296
on April 25, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2013; DOI: 10.1158/0008-5472.CAN-13-0874

Writing, review, and/or revisionof themanuscript:P. Kroon, V.M.Mann,M.Simms, S. Chettiar, C. Li, N.J. Maitland, A.T. CollinsAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): P. Kroon, P.A. Berry, S. Chettiar, N.J.MaitlandStudy supervision: N.J. Maitland, A.T. Collins
AcknowledgmentsThe authors thank Johnson & Johnson (Janssen R&D) for providing siltux-
imab and Mrs. Catherine Hyde for technical support.
Grant SupportThis research was supported by Yorkshire Cancer Research (Harrogate,
United Kingdom).The costs of publication of this article were defrayed in part by the payment of
page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received March 28, 2013; revised May 28, 2013; accepted May 29, 2013;published OnlineFirst July 3, 2013.
References1. Jemal A, Murray T, Ward E, Samuels A, Tiwari RC, Ghafoor A, et al.
Cancer statistics, 2005. CA Cancer J Clin 2005;55:10–30.2. Twillie DA, EisenbergerMA, Carducci MA, HseihWS, KimWY, Simons
JW. Interleukin-6: a candidate mediator of human prostate cancermorbidity. Urology 1995;45:542–9.
3. Adler HL, McCurdy MA, Kattan MW, Timme TL, Scardino PT, Thomp-son TC. Elevated levels of circulating interleukin-6 and transforminggrowth factor-beta1 in patients with metastatic prostatic carcinoma.J Urol 1999;161:182–7.
4. Nakashima J, Tachibana M, Horiguchi Y, Oya M, Ohigashi T, AsakuraH, et al. Serum interleukin 6 as a prognostic factor in patients withprostate cancer. Clin Cancer Res 2000;6:2702–6.
5. Hobisch A, Rogatsch H, Hittmair A, Fuchs D, Bartsch G Jr, Klocker H,et al. Immunohistochemical localization of interleukin-6 and its recep-tor in benign, premalignant and malignant prostate tissue. J Pathol2000;191:239–44.
6. Giri D, OzenM, IttmannM. Interleukin-6 is an autocrine growth factor inhuman prostate cancer. Am J Pathol 2001;159:2159–65.
7. Cavarretta IT,NeuwirtH,UntergasserG,MoserPL, ZakiMH,SteinerH,et al. The antiapoptotic effect of IL-6 autocrine loop in a cellular modelof advanced prostate cancer is mediated by Mcl-1. Oncogene 2007;26:2822–32.
8. Smith PC, Hobisch A, Lin DL, Culig Z, Keller ET. Interleukin-6 andprostate cancer progression. Cytokine Growth Factor Rev 2001;12:33–40.
9. Heinrich PC, Behrmann I, Muller-Newen G, Schaper F, Graeve L.Interleukin-6-type cytokine signalling through the gp130/Jak/STATpathway. Biochem J 1998;334(Pt 2):297–314.
10. Schindler C, Darnell JE Jr. Transcriptional responses to polypeptideligands: the JAK-STAT pathway. Annu Rev Biochem 1995;64:621–51.
11. Dhir R, Ni Z, Lou W, DeMiguel F, Grandis JR, Gao AC. Stat3 activationin prostatic carcinomas. Prostate 2002;51:241–6.
12. Gouilleux-Gruart V, Gouilleux F, Desaint C, Claisse JF, Capiod JC,Delobel J, et al. STAT-related transcription factors are constitutivelyactivated in peripheral blood cells from acute leukemia patients. Blood1996;87:1692–7.
13. Grandis JR, Drenning SD, Zeng Q, Watkins SC, Melhem MF, Endo S,et al. Constitutive activation of Stat3 signaling abrogates apoptosis insquamous cell carcinogenesis in vivo. Proc Natl Acad Sci U S A2000;97:4227–32.
14. Quintanilla-Martinez L, Kremer M, Specht K, Calzada-Wack J,Nathrath M, Schaich R, et al. Analysis of signal transducer andactivator of transcription 3 (Stat 3) pathway in multiple myeloma: Stat3 activation and cyclin D1 dysregulation aremutually exclusive events.Am J Pathol 2003;162:1449–61.
15. Garcia R, Yu CL, Hudnall A, Catlett R, Nelson KL, Smithgall T, et al.Constitutive activation of Stat3 in fibroblasts transformed by diverseoncoproteins and in breast carcinoma cells. Cell Growth Differ 1997;8:1267–76.
16. Watson CJ, Miller WR. Elevated levels of members of the STAT familyof transcription factors in breast carcinoma nuclear extracts. Br JCancer 1995;71:840–4.
17. Bromberg JF, Wrzeszczynska MH, Devgan G, Zhao Y, Pestell RG,Albanese C, et al. Stat3 as an oncogene. Cell 1999;98:295–303.
18. Sherry MM, Reeves A, Wu JK, Cochran BH. STAT3 is required forproliferation and maintenance of multipotency in glioblastoma stemcells. Stem Cells 2009;27:2383–92.
19. Hsu HS, Huang PI, Chang YL, Tzao C, Chen YW, Shih HC, et al.Cucurbitacin i inhibits tumorigenic ability and enhances radiochemo-sensitivity in nonsmall cell lung cancer-derived CD133-positive cells.Cancer 2011;117:2970–85.
20. Lin L, Liu A, Peng Z, Lin HJ, Li PK, Li C, et al. STAT3 is necessary forproliferation and survival in colon cancer-initiating cells. Cancer Res2011;71:7226–37.
21. Bonnet D, Dick JE. Human acute myeloid leukemia is organized as ahierarchy that originates from a primitive hematopoietic cell. Nat Med1997;3:730–7.
22. Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke MF.Prospective identification of tumorigenic breast cancer cells. Proc NatlAcad Sci U S A 2003;100:3983–8.
23. Singh SK, Clarke ID, Terasaki M, Bonn VE, Hawkins C, Squire J, et al.Identification of a cancer stem cell in human brain tumors. Cancer Res2003;63:5821–8.
24. Singh SK, Hawkins C, Clarke ID, Squire JA, Bayani J, Hide T, et al.Identification of human brain tumour initiating cells. Nature 2004;432:396–401.
25. Eramo A, Lotti F, Sette G, Pilozzi E, Biffoni M, Di Virgilio A, et al.Identification and expansion of the tumorigenic lung cancer stem cellpopulation. Cell Death Differ 2008;15:504–14.
26. O'Brien CA, Pollett A, Gallinger S, Dick JE. A human colon cancer cellcapable of initiating tumour growth in immunodeficient mice. Nature2007;445:106–10.
27. Li C, Heidt DG, Dalerba P, Burant CF, Zhang L, Adsay V, et al.Identification of pancreatic cancer stem cells. Cancer Res 2007;67:1030–7.
28. Collins AT, Berry PA, Hyde C, Stower MJ, Maitland NJ. Prospectiveidentification of tumorigenic prostate cancer stem cells. Cancer Res2005;65:10946–51.
29. Goldstein AS, Huang J, Guo C, Garraway IP, Witte ON. Identificationof a cell of origin for human prostate cancer. Science 2010;329:568–71.
30. Lawson DA, Zong Y, Memarzadeh S, Xin L, Huang J, Witte ON. Basalepithelial stem cells are efficient targets for prostate cancer initiation.Proc Natl Acad Sci U S A 2010;107:2610–5.
31. Garraway IP, Sun W, Tran CP, Perner S, Zhang B, Goldstein AS, et al.Human prostate sphere-forming cells represent a subset of basalepithelial cells capable of glandular regeneration in vivo. Prostate2010;70:491–501.
32. BirnieR,BryceSD,RoomeC,Dussupt V,DroopA, LangSH, et al. Geneexpression profiling of humanprostate cancer stemcells reveals a pro-inflammatory phenotype and the importance of extracellular matrixinteractions. Genome Biol 2008;9:R83.
33. Lin L, Benson DM Jr, DeAngelis S, Bakan CE, Li PK, Li C, et al. A smallmolecule, LLL12 inhibits constitutive STAT3 and IL-6-induced STAT3signaling and exhibits potent growth suppressive activity in humanmultiple myeloma cells. Int J Cancer 2012;130:1459–69.
34. Collins AT,Habib FK,MaitlandNJ, Neal DE. Identification and isolationof human prostate epithelial stem cells based on alpha(2)beta(1)-integrin expression. J Cell Sci 2001;114:3865–72.
35. Richardson GD, Robson CN, Lang SH, Neal DE, Maitland NJ, CollinsAT. CD133, a novel marker for human prostatic epithelial stem cells.J Cell Sci 2004;117:3539–45.
36. BrehmMA, Mangada J, Markees TG, Pearson T, Daniels KA, ThornleyTB, et al. Rapid quantification of naive alloreactive T cells by TNF-alpha
STAT3 Blockade Reduces Cancer Stem Cell Frequency
www.aacrjournals.org Cancer Res; 73(16) August 15, 2013 5297
on April 25, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2013; DOI: 10.1158/0008-5472.CAN-13-0874

production and correlation with allograft rejection in mice. Blood2007;109:819–26.
37. GleasonDF.Histologic grading of prostate cancer: a perspective.HumPathol 1992;23:273–9.
38. Hu Y, Smyth GK. ELDA: extreme limiting dilution analysis for compar-ing depleted and enriched populations in stem cell and other assays.J Immunol Methods 2009;347:70–8.
39. Thompson JE, Cubbon RM, Cummings RT, Wicker LS, Frankshun R,Cunningham BR, et al. Photochemical preparation of a pyridonecontaining tetracycle: a Jak protein kinase inhibitor. BioorgMedChemLett 2002;12:1219–23.
40. Steiner H, Cavarretta IT, Moser PL, Berger AP, Bektic J, Dietrich H,et al. Regulation of growth of prostate cancer cells selected in thepresence of interleukin-6 by the anti-interleukin-6 antibodyCNTO328.Prostate 2006;66:1744–52.
41. Lin L, Hutzen B, Li PK, Ball S, Zuo M, DeAngelis S, et al. A novel smallmolecule, LLL12, inhibits STAT3 phosphorylation and activities andexhibits potent growth-suppressive activity in human cancer cells.Neoplasia 2010;12:39–50.
42. DrachenbergDE, Elgamal AA,RowbothamR,PetersonM,MurphyGP.Circulating levels of interleukin-6 in patients with hormone refractoryprostate cancer. Prostate 1999;41:127–33.
43. Bellido T, Jilka RL, Boyce BF, Girasole G, Broxmeyer H, Dalrymple SA,et al. Regulation of interleukin-6, osteoclastogenesis, and bone massby androgens. The role of the androgen receptor. JClin Invest 1995;95:2886–95.
44. Pedranzini L, DechowT,BerishajM,ComenzoR, ZhouP, Azare J, et al.Pyridone 6, a pan-Janus-activated kinase inhibitor, induces growthinhibition of multiple myeloma cells. Cancer Res 2006;66:9714–21.
45. WangH, Lathia JD,WuQ,Wang J, Li Z, Heddleston JM, et al. Targetinginterleukin 6 signaling suppresses glioma stem cell survival and tumorgrowth. Stem Cells 2009;27:2393–404.
46. Dorff TB, Goldman B, Pinski JK, Mack PC, Lara PN Jr, Van Veld-huizen PJ Jr, et al. Clinical and correlative results of SWOG S0354: aphase II trial of CNTO328 (siltuximab), a monoclonal antibodyagainst interleukin-6, in chemotherapy-pretreated patients withcastration-resistant prostate cancer. Clin Cancer Res 2010;16:3028–34.
47. Fizazi K, De Bono JS, Flechon A, Heidenreich A, Voog E, DavisNB, et al. Randomised phase II study of siltuximab (CNTO 328), ananti-IL-6 monoclonal antibody, in combination with mitoxantrone/prednisone versus mitoxantrone/prednisone alone in metastaticcastration-resistant prostate cancer. Eur J Cancer 2012;48:85–93.
Kroon et al.
Cancer Res; 73(16) August 15, 2013 Cancer Research5298
on April 25, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2013; DOI: 10.1158/0008-5472.CAN-13-0874

2013;73:5288-5298. Published OnlineFirst July 3, 2013.Cancer Res Paula Kroon, Paul A. Berry, Michael J. Stower, et al. Recovery of Prostate Cancer Stem-like CellsJAK-STAT Blockade Inhibits Tumor Initiation and Clonogenic
Updated version
10.1158/0008-5472.CAN-13-0874doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2013/07/09/0008-5472.CAN-13-0874.DC1
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/73/16/5288.full#ref-list-1
This article cites 47 articles, 17 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/73/16/5288.full#related-urls
This article has been cited by 9 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/73/16/5288To request permission to re-use all or part of this article, use this link
on April 25, 2019. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst July 3, 2013; DOI: 10.1158/0008-5472.CAN-13-0874















![No. 19-5288 IN THE UNITED STATES COURT OF ...[ORAL ARGUMENT SCHEDULED JAN. 3, 2020] No. 19-5288 IN THE UNITED STATES COURT OF APPEALS FOR THE DISTRICT OF COLUMBIA CIRCUIT _____ IN](https://img.pdfslide.us/doc/110x75/5e49e0a352d22845e84e5ddc/no-19-5288-in-the-united-states-court-of-oral-argument-scheduled-jan-3-2020.jpg)


![PhysicaA Stochasticresonanceinperiodicpotentialsdriven ...5284 K.Liu,Y.Jin/PhysicaA392(2013)5283–5288 potentialsystems[19–21].Amongthem,KimandSung[19]foundtheresonancebehaviorinthefrequency](https://img.pdfslide.us/doc/110x75/5ed1eeb4ae118d4f2114c3db/physicaa-stochasticresonanceinperiodicpotentialsdriven-5284-kliuyjinphysicaa39220135283a5288.jpg)










