Embed Size (px)
Citation preview

Research ArticlePurification and Biochemical Characterization of ThreeMyotoxins from Bothrops mattogrossensis Snake Venom withToxicity against Leishmania and Tumor Cells
Andreacutea A de Moura1 Anderson M Kayano1 George A Oliveira1 Sulamita S Setuacutebal2
Joatildeo G Ribeiro12 Neuza B Barros2 Roberto Nicolete2 Laura A Moura3 Andre L Fuly3
Auro Nomizo4 Saulo L da Silva5 Carla F C Fernandes2 Juliana P Zuliani12
Rodrigo G Staacutebeli12 Andreimar M Soares12 and Leonardo A Calderon12
1 Centro de Estudos de Biomoleculas Aplicadas a Saude CEBio Fundacao Oswaldo Cruz Fiocruz Rondoniae Departamento de Medicina Nucleo de Saude Universidade Federal de Rondonia UNIR 76812ndash245 Porto Velho RO Brazil
2 Fundacao Oswaldo Cruz Fiocruz Rondonia 76812ndash245 Porto Velho RO Brazil3 Departamento de Biologia Celular e Molecular Instituto de Biologia Universidade Federal Fluminense UFF24020-141 Niteroi RJ Brazil
4 Faculdade de Ciencias Farmaceuticas FCFRP Universidade de Sao Paulo USP 14040-903 Ribeirao Preto SP Brazil5 Universidade Federal de Sao Joao del Rei UFSJ 36420-000 Ouro Branco MG Brazil
Correspondence should be addressed toAndreimarM Soares andreimarfiocruzbr and LeonardoA Calderon calderonunirbr
Received 16 September 2013 Accepted 3 December 2013 Published 3 March 2014
Academic Editor Fernando Albericio
Copyright copy 2014 Andrea A de Moura et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Bothrops mattogrossensis snake is widely distributed throughout eastern South America and is responsible for snakebites in thisregion This paper reports the purification and biochemical characterization of three new phospholipases A
2(PLA
2s) one of
which is presumably an enzymatically active Asp49 and two are very likely enzymatically inactive Lys49 PLA2homologues
The purification was obtained after two chromatographic steps on ion exchange and reverse phase column The 2D SDS-PAGEanalysis revealed that the proteins have pI values around 10 are each made of a single chain and have molecular masses near13 kDa which was confirmed by MALDI-TOF mass spectrometry The N-terminal similarity analysis of the sequences showedthat the proteins are highly homologous with other Lys49 and Asp49 PLA
2s from Bothrops species The PLA
2s isolated were
named BmatTX-I (Lys49 PLA2-like) BmatTX-II (Lys49 PLA
2-like) and BmatTX-III (Asp49 PLA
2) The PLA
2s induced cytokine
release frommouse neutrophils and showed cytotoxicity towards JURKAT (leukemia T) and SK-BR-3 (breast adenocarcinoma) celllines and promastigote forms of Leishmania amazonensis The structural and functional elucidation of snake venoms componentsmay contribute to a better understanding of the mechanism of action of these proteins during envenomation and their potentialpharmacological and therapeutic applications
1 Introduction
Snake venoms contain a complex mixture of componentswith a wide range of biological and pharmacological activ-ities More than 90 of their dry weight is composed ofproteins including a variety of enzymes such as phospho-lipases A
2 proteases (metallo and serine) L-amino acid
oxidases esterases as well as many other nonenzymatic
proteins and peptides [1ndash3] These proteins and peptidescan be grouped into a small number of superfamilies basedon remarkable similarities in their primary secondary andtertiary structures while showing distinct pharmacologiceffects [3]
One important protein superfamily present in all snakevenoms is phospholipase A
2(PLA2 EC 3114) PLA
2s are a
class of heat-stable and highly homologous enzymes which
Hindawi Publishing CorporationBioMed Research InternationalVolume 2014 Article ID 195356 13 pageshttpdxdoiorg1011552014195356
2 BioMed Research International
catalyze the hydrolysis of the 2-acyl bond of cell membranephospholipids releasing free fatty acids such as arachidonicacid and lysophospholipids PLA
2s have been characterized
as the major component of snake venoms being responsiblefor several pathophysiological effects caused by snake enven-omation such as neurotoxic cardiotoxic myotoxic cyto-toxic hypotensive and anticoagulant activities triggering anintense inflammatory reaction with the release of cytotoxinsand eicosanoids [4ndash6] PLA
2rsquos involvement in a variety of
inflammatory diseases and accidents caused by venomousanimals has raised medical and scientific interest in thisenzyme [7 8]
Myotoxic PLA2s from the Bothrops species are composed
of approximately 110 to 135 amino acid residues and canbe divided into two groups ldquoclassicalrdquo which contain anaspartate residue at position 49 (Asp49) and catalyze esterbond hydrolysis at the glycerophospholipid sn-2 position ina Ca2+-dependent manner and ldquovariantrdquo which contain alysine residue at the same position (Lys49) This substitutionaffects the ability of these proteins to bind to Ca2+ which isan essential cofactor for catalysis leading to decreased or nocatalytic activity [1ndash5 7 9 10]
The Bothrops mattogrossensis snake belongs to the Both-rops neuwiedi complex [11]This snake is found in the easternregion of South America including Bolivia Brazil SoutheastPeru Paraguay Uruguay and Argentina [12] The presentstudy describes for the first time the isolation identificationand functional characterization of three myotoxic phospho-lipases A
2 named BmatTX-I (Lys49 PLA
2-like) BmatTX-
II (Lys49 PLA2-like) and BmatTX-III (Asp49 PLA
2) and
evaluates their activity against Leishmania and tumor cells
2 Materials and Methods
21 Venom The venom from the Bothrops mattogrossensissnake was acquired from Serpentario de Proteınas BioativasBatatais-SP Brazil This study was authorized by InstitutoBrasileiro do Meio Ambiente e dos Recursos NaturaisRenovaveismdashIBAMA Instituto Chico Mendes de Conser-vacao da BiodiversidademdashICMBio (number 27131-1) andConselho de Gestao do Patrimonio GeneticomdashCGENBrazil(number 0106272011-1)
22 Animals Swiss male mice (18ndash20 g) were used Theseanimals were housed in temperature-controlled rooms andreceived water and food ad libitum until used Animalcare was in accordance with the guidelines of the BrazilianCollege for Animal Experimentation (COBEA) and wasapproved by the Committee of Ethics on Animal Utiliza-tion in Research from the Institute of Research of Tropi-cal Pathologies (IPEPATROFIOCRUZ-Rondonia) (protocolnumber 20121)
23 Bothrops mattogrossensis Venom Fractioning Around250mg of B mattogrossensis venom was eluted in 12mL ofsterile deionized water and centrifuged at 1530timesg for 10min-utes and then submitted to cation exchange chromatographyin anAkta purifier 10 (GE-Healthcare) using aCM-Sepharose
column (27 times 300mm GE Healthcare) equilibrated with50mM ammonium bicarbonate buffer pH 80 and elutedwith a linear gradient of 0ndash100 500mM ammonium bicar-bonate pH 80 at a flow rate of 50mLminute The frac-tions were monitored at an absorbance of 280 nm collectedmanually identified lyophilized and stored at minus20∘C Allfractions were submitted to SDS-PAGE and enzymatic activ-ity analyses The isolation of the three PLA
2s was obtained
by liquid chromatography using a Discovery C18 column(25 times 46mm Supelco) equilibrated with deionized waterwith 01 trifluoroacetic acid (vv) and eluted with tenvolumes of a linear gradient of 0ndash100 acetonitrile with 01trifluoroacetic acid (vv) at a flow rate of 10mLminuteThe elution was monitored at an absorbance of 280 nmmanually collected lyophilized and stored at minus20∘C In orderto determine protein concentration the Bradford method(Bio-Rad) was used with bovine serum albumin (BSA) as astandard [14]
24 Monodimensional Electrophoresis (SDS-PAGE) Poly-acrylamide gel electrophoresis (PAGE) in the presence ofsodium dodecyl sulfate (SDS) was conducted using themethod described by Laemmli [15] The electrophoresis wascarried out at 15mA and 5W using a conventional molecularweight standard (BioLabs) and a Lys49-PLA
2homologue
isolated from B jararacussu snake venom (BthTX-I) asmarkers The gel was stained with Coomassie Brilliant Blue(CBB)G-250 and images of the gel were obtained using ImageScanner (GE Healthcare)
25 Bidimensional Electrophoresis (2D-SDS-PAGE) and pIDetermination Theprotein fractions were submitted to elec-trofocusing in 13 cm strips with pHs ranging from 3 to 10in a nonlinear form using an Ettan IPGphor 3 IEF System(GE Healthcare) At the end of the electrofocusing thestrips containing the PLA
2s were equilibrated and transferred
to a 125 polyacrylamide gel where they were separatedaccording to molecular mass The electrophoretic run wasperformed at 25mA and 100W during 55 hours and atthe end of the protein separation the gel was stained withCoomassie Brilliant Blue (CBB) G-250 and images of the gelwere captured using Image Scanner (Amershan Bioscience)
26 Mass Spectrometry (MALDI-TOF) The samples (1 120583L)were mixed with a matrix solution composed of sinapinicacid and acetonitrile with 01 trifluoroacetic solubilized ata 1 1 ratio The average mass of the protein was obtained in aMALDI-TOF system (Shimatzu Biotech) operated in linearmode using insulin (57345Da) cytochrome C (123619Da)apomyoglobin (169522Da) aldolase (392122Da) and albu-min (664300Da) as calibrators The mass spectra obtainedwere submitted to automatic baseline subtraction
27 N-Terminal Sequencing and Similarity Search The aminoterminal sequence of each isolated PLA
2previously deter-
mined by automated Edman degradation was used to searchfor sequence similarity Approximately 50120583g of each isolatedPLA2corresponding to approximately 2 nmolsmL sample
BioMed Research International 3
was submitted for N-terminal amino acid sequencing usingthe automated Edman degradation method [13] on a PPSQ-33A (Shimadzu) automatic sequencer A sequence similaritysearch and multiple sequence alignment were performed inthe SWISS-PROTTREMBL database using the programsFASTA BLAST and CLUSTALW2
28 Functional Characterization281 Phospholipase Activity with 4-nitro-3(octanoyloxy)ben-zoic (4N3OBA) Phospholipase activity of the three PLA
2s
was assayed following the protocols described by Holzer andMackessy [16] modified for 96-well plates The standardassay mixture contained 200120583L of buffer (10mM Tris-HClplus 10mM CaCl
2 and 100mM NaCl pH 80) 20 120583L of
4-nitro-3(octanoyloxy)benzoic (4N3OBA ndash Biomol EUA)20120583L of deionized water and 20120583L of PLA
2(5 120583g) After
the addition of the PLA2s the mixture was incubated for
30 minutes at 37∘C and the absorbance was determined at425 nmusing an Eon (Biotek)microplate spectrophotometerfor 3-minute intervals The enzymatic activity was expressedas the reactionrsquos initial velocity (119881
119900) calculated based on the
increase in absorbance
282 Phospholipase Activity with Fluorescent SubstratePhospholipase A
2rsquos (PLA
2) enzymatic activity was evaluated
through the hydrolysis of synthetic fluorescent phospholipidusing the fluorescent substrate Acyl 6 0 NBD phospholipidNBD-phosphatidylcholine (NBD-PC) (Avanti Polar LipidsInc Alabaster AL USA) The assay was performed in aspectrofluorometer (Shimadzu RF-5301PC software RFPC)with excitation and emission wavelengths of 460 and 534respectivelyThe enzymatic activity of each Bmattogrossensischromatographic fraction (7 12 15 16 17 and 20) wasevaluated over 250 seconds after the addition of substrate(33 120583gmL final concentration) in a reaction media con-taining 50mM Tris-HCl and 8mM CaCl
2at pH 75 at room
temperature
283 Platelet Aggregation A platelet aggregation assay wascarried out according to the process described by Fuly et al[17] with modifications Platelet aggregation was monitoredin an Aggregometer (Chrono Logmodel 490 2D HavertownUSA) using Platelet-Rich-Plasma (PRP) PRP was obtainedfrom human whole blood of health volunteers (CAAE14204413500000011) BmatTX-III (rechromatography frac-tion 15)was incubatedwith PRP for fiveminutes at 37∘Cbeingstirred constantly and then platelet aggregation was trig-gered by the addition of ADP (15120583M) or collagen (16 120583gmL)Assays were performed at 37∘C in siliconized glass cuvettesin a final volume of 300 120583L Control experiments wereperformed by adding agonists in the absence of peak 15 Onehundred percent (100) platelet aggregation was obtainedwith a supramaximal concentration of each agonist anddetermined 6 minutes after the addition of each while PRPrsquoslight transmittance showed 0 aggregation
284 Hemorrhage Activity Micewere injected intradermallyin the dorsal region with 50 120583g of crude venom dissolved
in 50120583L of physiological solution [18] Controls received50 120583L of physiological solution in identical conditions After 3hours the animals were euthanized through cervical disloca-tion and the skin was removed The hemorrhagicactivitywasexpressed as size of thehemorrhagic area on the inner surfacemeasured in mm2
285 Coagulation Activity and Minimal Coagulation Dose(MCD)Determination Coagulation activity was tested usinga methodology previously described by Gene et al [19] Inbrief 200120583L of plasma frommicewas distributed in a 96-wellplate and 10 120583L of crude venom containing different concen-trations (0312 0625 125 25 5 and 10 120583g) of proteins wasadded The plate was placed in a thermostatically controlledenvironment and the optical density was measured every 3seconds at 600 nm using an Eonmicroplate spectrophotome-ter (Biotek) in order to evaluate the smallest concentration ofvenom able to coagulate 200120583L of plasmaminute
286 Proteolytic Activity Proteolytic activitywas ascertainedusing azocasein (Sigma) as a substrate according to the pro-cedure described by Charney and Tomarelli [20] Azocaseinsolubilized in distilled water (150 120583L) was added to 7 120583L ofthe crude venom in different protein concentrations (03120625 125 25 5 and 10 120583g) and then the mixture wasincubated in a water bath at 37∘C for 1 hour The reactionstopped when 150 120583L of 20 (mv) trichloroacetic acid wasadded which was followed by incubation for 30min andcentrifugation at 10000timesg for 10 more minutesThen 100 120583Lof the supernatant was transferred to a multiple-well plateand the absorbance was measured at 440 nm using an Eonmicroplate spectrometer (Biotek) and one enzymatic unit (U)was defined as the amount of enzyme necessary to increasethe absorbance by 005
287Myotoxic Activity Myotoxic activity was determined bymeasuring the creatine kinase (CK) activity in the plasma[21] Groups of mice were injected in the gastrocnemiusmuscle with 25 120583g of isolated myotoxins diluted in 50120583L ofphosphate buffered saline (PBS) Negative controls receivedan injection of 50 120583L of PBSThree hours after the injectionsaliquots of mice blood were collected from the caudal veinin heparinized capillaries and centrifuged at 1530timesg for 20minutes Creatine kinasersquos enzymatic activity was determinedusing the CK-NAC kinetic kit (Bioclin Brazil) accordingto the manufacturerrsquos protocol Absorbance was measuredfor 3 minutes at 37∘C in a spectrophotometer at 340 nmEnzymatic activity was expressed in unitsliter (UL) andeach unit consists of the result of the phosphorylation of onenanomol of creatine per minute
288 Neutrophil Viability Neutrophils were collected 6hours after the intraperitoneal (IP) injection of 15mL of3 thioglycollate sterile solution according to the methodpreviously described by Call et al [22] The animals wereeuthanized in order to collect the cells and the peritonealcavity was washed with 3mL of PBS The predominance of
4 BioMed Research International
neutrophils in the liquid obtained was confirmed by micro-scopic analysis with glass slides stained with a panoptic dyeThe peritoneal neutrophils obtained were suspended in anRPMI culturemedium (Gibco-BRL) supplemented with gen-tamicin (100 120583gmL) L-glutamine (2mM) and 10 bovinefetal serum (SFB) in order to obtain 2times105 cells100 120583L Nextcellular viability was assayed over 1 12 and 24 h at 37∘Cand 5 CO
2in which the cells were incubated in a 96-well
plate with the previously isolated PLA2s at a concentration
of 12 120583gmL using RPMI as a negative control After thisthe samples were centrifuged and the supernatant removedCellular viability was determined using the MTT method[23]
289 Quantification of Cytokine EIA was used to evaluateIL-1120573 cytokine as described by Schumacher et al [24] Brieflythe neutrophils (2 times 105 cells200120583L) were either incubatedwith the isolated proteins at 3 6 and 12 120583gmL concentrations(experimental group) or with PMA (positive control group)or with RPMI (negative control) and kept for 12 and 24 hoursat 37∘C in a humid atmosphere with 5 CO
2 After that 96-
well plates were coatedwith 100 120583L of the capturemonoclonalantibody anti-IL-1120573 and incubated for 18 hours at 37∘C Theplate was then washed with washer buffer (PBSTween20)After that 200120583L of blocking buffer containing 5 bovineserum albumin (BSA) in PBSTween20 was added to thewells and the plates were incubated for 1 hour at 37∘CFollowing this the wells were washed and 50 120583L of eithersamples or standard were dispensed into each well and theplates were incubated for 2 hours at 37∘C After this theplate was washed and 100 120583L of detection antibody anti-IL-1120573 was added for 2 hours at 37∘C After incubation andwashing 100 120583L of streptavidin-peroxidase was added fol-lowed by incubation and addition of the substrate (100120583LmL331015840551015840-tetramethylbenzidine) Finally sulfuric acid (50120583L)was added to stop the reaction Absorbances were recordedat 540 and 450 nm and concentration of IL-1120573 was estimatedfrom standard curves prepared with recombinant cytokineThe results were expressed as pgmL of IL-1120573
2810 Antitumor Activity Cytotoxic activity of isolatedPLA2s on human T-cell leukemia (JURKAT) and breast
adenocarcinoma (SK-BR-3) lines obtained from the Amer-ican Culture Collection of Cells (ATCC American TypeCulture Collection Rockville MD USA) were investigated(Figure 6) This activity was assayed by MTT staining asdescribed by Mosmann [25] and adapted by Stabeli et al[21] Cells were dispensed in 96-well plates at a densityof 5 times 105 cellsmL After 24 h of incubation the mediumwas removed and fresh medium with or without differentconcentrations of PLA
2s (BmatTX-I BmatTX-II BmatTX-
III or methotrexate) was added to the wells and incubatedfor another 24 hours The evaluation of the cytotoxic activitywas measured in a spectrophotometer using an interferencefilter of 570 nm and expressed as a percentage
2811 Anti-Leishmania Activity Promastigote forms ofLeishmania amazonensis (IFLABR67PH8) were dis-pensed in a 96-well plate with 1 times 105 cellswell Different
Table 1 Activities induced by B mattogrossensis snake venom
Effecta ActivityPhospholipase activityb (Umg) 186405Proteolytic activityc (U10120583g) 30 plusmn 01
Hemorrhagic halod (cm2) 333 plusmn 005
Coagulation activitye (MCD 120583g) 0325aAll experiments were carried out in triplicate bActivity using NOB stainedsubstrate cOne enzymatic unit (U) was defined as the quantity of enzymeneeded to increase the absorbance by 005UA440 nm dValues 3 h afterincubation with crude venom of B mattogrossensis (50120583g) eMCD theminimum coagulation dose was the dose capable of coagulating 200120583L ofcitrated plasma in less than a minute
concentrations (312 625 125 25 50 and 100120583gmL) ofthe crude venom of B mattogrossensis and the isolatedPLA2s (BmatTX-I and BmatTX-II) were added to each well
100mgmL of pentamidine was used as a positive controlAfter an incubation period of 48 h 10 120583L of a 5mgmL MTTsolution was added Then the plates were placed in theoven at 33∘C with 5 CO
2for 4 hours of incubation after
which 50120583L of SDS (20 wv) was added Absorbance wasmonitored at 570 nm Results were expressed in toxicitypercentage following the equation 1 minus (DO sampleDOcontrol) times 100
29 Statistical Analysis Results were expressed as mean +minusstandard deviation An ANOVA test was used to evaluate thesignificance of the differences observed with 119875 value le 005considered to be significant
3 Results and Discussion
31 Crude Venom Biological Activities B mattogrossensissnake venom induced hemorrhage coagulation proteolyticand phospholipase activities in vitro (Table 1) The hemor-rhagic activity of B mattogrossensis crude venom was eval-uated based on the dimensions of the average hemorrhagichalo which was 333 cm2 This result is in agreement with theresults recently obtained with B atrox [26]
Coagulation activity was confirmed after incubation ofdifferent concentrations of B mattogrossensis crude venomwith plasma in which theminimum coagulation dose capableof promoting coagulation in less than 1min was 0325120583gof protein Analysis of the proteolytic activity of B mat-togrossensis crude venom demonstrated a concentration-dependent response (data not shown) Regarding phospho-lipase proteolytic coagulating and hemorrhagic activitiesof B mattogrossensis crude venom assays confirmed thepresence and activity of proteases and phospholipases Thepresence of metalloproteases was evidenced by the importantformation of an extensive hemorrhagic halo in vivo Thepresence of serine proteases was evidenced by coagulatingactivity present even in low concentrations of the venom(325 120583gmL)
Phospholipase activity of B mattogrossensis crude venomassayed with 4N3OBA synthetic substrate was 186405Umgmeasured by the number of moles of chromophores released
BioMed Research International 5
per minute (n∘molsmin or U) for a given quantity of proteinin milligrams (mg)
The properties of snake venom components observed inthis study are characteristic of accidents caused by snakesof the Bothrops sp genus symptoms such as pain edemahemorrhage and necrosis and additionally systemic distur-bances are characteristic and corroborate the literature thatdescribes that proteases of snake venom proteins are closelyrelated to interference in the hemostatic system promotingblood coagulation fibrinolysis and platelet aggregation [27ndash30]
32 Isolation and Biochemical Characterization of Phospholi-pases The present study showed for the first time the isola-tion of the three phospholipases A
2from B mattogrossensis
snake venom obtained by two chromatographic steps Firstionic exchange chromatography was performed on a CM-Sepharose column with an ammonium bicarbonate gradient(50 to 500mM pH 80) The elution of absorbed proteinswith a linear gradient of concentrated buffer resulted inthirteen fractions (Figure 1(a)) of which fraction nine (9) wasrelated to PLA
2s because it showed phospholipase activity
of 22109Umg on artificial substratum All fractions werelyophilized and submitted to unidimensional electrophoresisrevealing many protein bands
Fraction 9 was submitted to the second chromatographicstep in a reverse column phase on a Discovery C18 columnusing 01 Trifluoroacetic (TFA) and 999 Acetonitrile(ACN) as solvents for the separation of other venom com-ponents The elution of absorbed proteins with a lineargradient of concentrated buffer resulted in twenty-two (22)fractions (Figure 1(b)) The PLA
2s were highly purified with
approximately 40 ACN Similar results were observed inthe isolation of other PLA
2s from snake venoms in high per-
formance liquid chromatography on reverse phase columnswhere the elution profile of these proteins occurs between 30and 40 ACN [5]
The association of chromatographic techniques suchas ionic exchange and reverse phase has commonly beenused and many snake venoms have been fractioned thisway highlighting the phospholipase purification of a speciesbelonging to the old complex ldquoBothrops neuwiedirdquo as well asthe target species studied in this research Two PLA
2basic
isoforms from B (neuwiedi) pauloensis venom were purifiedby Rodrigues et al [31] using biochemical techniques similarto the ones used in this study with the combination of ionicexchange (cationic) and reverse phase chromatographies
Other PLA2s have also been purified using simplified
procedures based in CM-Sepharose andor reverse phase asfor example venomPLA
2s fromBothropsmoojeni [21 32 33]
B pirajai [34 35] B jararacussu [36 37] B alternatus [8 38]Cerastes cerastes [39] and Elaphe climacophora [40]
Fractions 13 and 14 were related to enzymatically inactivePLA2s whereas fraction 15was related to enzymatically active
PLA2s The phospholipase activity of the collected fractions
were analyzed with synthetic NOB and NBD-PC substrates(Figures 2(a) and 2(b))The amount of activity was comparedto BthTX-II a basic enzymatically active PLA
2(Asp49) and
BthTX-I a basic enzymatically inactive PLA2(Lys49) both
20
15
10
5
0
100 200 300 400
80
60
40
20
0
B (
)
(mAU
)
1
23
4
5 67 8
9
10 11 12 13
MW 02 03 04 06 08 09 10 12 13
175805846
30
2317
07
times102
(min)
Btx
(a)
100
80
60
40
20
B (
)
MW175
805846
30
2317
07
(mAU
)
times102
6
5
4
3
2
1
0
0 10 20 30 40
1
2
3 45
6
7
8
910 11
12
13
1415
16
1718
19
20 2122
(min)
07 12 1314 15 16 17 18Btx
(b)
Figure 1 Chromatographic and electrophoretic profile of Bothropsmattogrossensis venom fractioning (a) CM-Sepharose columnsequilibrated with solvent A (50mM ammonium bicarbonatepH 80) and eluted with a 0ndash100 concentration gradient of solventB (500mM ammonium bicarbonate pH 80) at a 50mLminuteflow Emphasis on peak 9 rechromatographed (b) Rechromatog-raphy of fraction 9 on Discovery C18 column equilibrated withsolvent A (01 TFA) and eluted with a concentration gradient of0ndash100 of solvent B (999 acetonitrile and 01 trifluoroaceticacid) and a 10mLmin flow Emphasized in blue are fractions 1314 and 15 characterized as phospholipases BmatTX-I BmatTX-II and BmatTX-III respectively Controls MW molecular weightstandard BTx BthTX-I a basic enzymatically inactive PLA
2(Lys49)
(10120583g) isolated from Bothrops jararacussu venom Absorbancesread at 280 nm Electrophoresis gel made with 125 (ww)acrylamidebis-acrylamide
previously isolated from Bothrops jararacussu venom [36 41]The BmatTX-III PLA
2s (Asp49) were not able to induce
platelet aggregation and did not inhibit collagen or ADPinduced platelet aggregation (data not shown)
The degree of purity of the isolated proteins was furtherdemonstrated by SDS-PAGE mass spectrometry (Figure 3)and N-terminal sequencing (Figures 4(a) and 4(b)) Thepurified PLA
2s were named BmatTX-I BmatTX-II and
6 BioMed Research International
F12
Bmat
TX-I
Bmat
TX-I
I
Bmat
TX-I
II
F16
F17
F18
F19
F20
F21
F22
BthT
X-I
BthT
X-II
0
200
400
600
800
1000Ph
osph
olip
ase a
ctiv
ity (U
mg)
(a)
F12
Bmat
TX-I
II
F16
F17
F20
0
200
400
600
800
1000
Fluo
resc
ent i
nten
sity
(b)
Control BmatTX-I BmatTX-II BmatTX-III0
1000
2000
3000
BmatTX-I (Lys49)BmatTX-II (Lys49)BmatTX-III (Asp49)
Control
CK (U
L)
25120583g50120583L
lowastlowast
lowast
(c)
Figure 2 Enzymatic activity and myotoxic activity of PLA2s isolated from the venom of B mattogrossensis (a) Phospholipase activity of the
fractions collected from the rechromatography of fraction 9 done in C18 column assayed using an NOB stained substrate This activity wasassessed through the measurement of the number of moles of the released chromophore per minute (n∘molsmin or U) per milligram ofprotein (b) Phospholipase activity of the fractions collected from the rechromatography of fraction 9 done in C18 column assayed using afluorescent substrate (c) Myotoxic activity evaluated for inoculation of PLA
2s (25 120583g50 120583L) or PBS (control) done intramuscularly in the
gastrocnemius muscle of mice After 3 hours the creatine kinase (CK) level an important marker of muscular lesion was assayed in theanimalrsquos plasma Each bar represents the average +minus SD of three independent groups lowast119875 lt 005 compared to the control F13 BmatTX-I(Lys49) F14 BmatTX-II (Lys49) F15 BmatTX-III (Asp49) Controls BthTX-I (Lys49) and BthTX-II (Asp49)
BmatTX-III They were characterized as single polypeptidechains with isoelectric points around 10 (data not shown)This result agrees with data from published literature wheremost basic PLA
2s with or without catalytic activity on
artificial substrates [32 42] show an isoelectric point between8 and 10
The average molecular mass defined by mass spectrom-etry was 13304 Da for BmatTX-I 13623Da for BmatTX-IIand 13681Da for BmatTX-III (Figure 3)Thismolecularmassis consistent with most isolated PLA
2s from snake venoms
which are around 13 to 16 kDa [27 36 43]BmatTX-I sequencing showed a lysine (Lys) at position
49 and although only the first 28 amino acid residuesof BmatTX-II have been sequenced both are highly sim-ilar to the PLA
2Lys49 homologue subgroup The N-
terminal sequence alignment of BmatTX-I and BmatTX-IIhas revealed that these basic proteins are PLA
2s similar to
homologous Lys49 and other Lys49 myotoxins from snake
venoms (Figure 4(a)) BmatTX-I showed 94 similarity withMjTX-I present in B moojeni venom 92 with BthTX-I andBOJU-I present in B jararacussu venom and 94 similaritywith MTX-II present in B brazili venom
Additionallymultiple sequence alignment of BmatTX-IIIshowed a PLA
2-Asp49 basic myotoxin with another Asp49
of the Bothrops genera (Figure 4(b)) It can be observedthat BmatTX-III presented 78 similarity with BmjeTX-Iand 75 with BmoTX-I both from B moojeni venom 71similarity with BthTX-II isolated from B jararacussu venomand 65 similarity with PrTX-III from B pirajai venom
In the analysis of the sequences of the PLA2s isolated
from B mattogrossensis in this study highly preserved con-stituent residues of the 120572-helix structure characteristic ofphospholipases were identified as well as the presence ofmany cysteine residues which suggests the existence of manydisulfide bridges important for the stabilization of PLA
2rsquos
molecular structure [21 44]
BioMed Research International 7
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1360475
1398779
(a)
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1362326
1380673
(b)
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1256219
1368171
1402625
(c)
Figure 3 Mass determination by spectrometryThe average proteinmasses of BmatTX-I (a) BmatTX-II (b) and BmatTX-III (c) wereobtained in a MALDI-TOF system operated in linear mode usingexternal standards for calibration The resulting mass spectra weresubmitted to automatic baseline subtraction
Some studies regarding the amino acid composition ofPLA2s demonstrate that these proteins are rich in basic
and hydrophobic amino acids containing three long 120572-helixes two beta sheets and a Ca2+ binding site [45ndash47]Calcium is absolutely necessary for hydrolysis thereforealmost all PLA
2s have a highly preserved region for a Ca2+
bond (XCGXGG) and a catalytic site (DXCCXXHD) [48]It is observed in BmatTX-III that the calcium binding site(27YCGWGG
32) is preserved indicating that it is a catalyt-
ically active PLA2belonging to the PLA
2Asp49 subgroup
33 Biological Activities of Phospholipases A2 The main
objective for the isolation and characterization of compo-nents of snake venom is to better understand the partic-ipation of each component in different pathophysiologicalprocesses resulting from envenomation Local lesions can beattributed to proteases phospholipase activity and hemor-rhagic factors of these venoms followed by the release ofvasoactive agents causing hemorrhaging in various organsand tissues [5 44 49ndash52]
The PLA2s isolated from B mattogrossensis venom
BmatTX-I BmatTX-II and BmatTX-III showed highmyotoxic activity (Figure 2(c)) At a concentration of25 120583g50 120583L the PLA
2s induced a significant release of CK
an important muscular lesion marker when compared to thecontrol Myotoxicity is the characteristic presented by mostbasic phospholipases A
2from snake venoms Several studies
demonstrate that the myotoxic effect begins quickly by directaction of the myotoxic PLA
2s on the plasma membrane of
muscle cells or it is mediated by metalloproteases due toconsequent degeneration and ischemia [27 28 44]
Regarding the Lys49-PLA2myotoxins it is evident that
they lyse the plasma membrane of the muscle cell infected invivo however the exact mechanism has not been describedyet Furthermore it is not known if the toxin is internalizedbefore during or after the initial lysis or if it is not internal-ized Althoughmyotoxicity can be induced by the productionof fatty acids there is a second mechanism that seems to beindependent of the enzymatic activity and is mediated bythe C-terminal region at sites 115ndash129 of the Lys49 molecules[43 53 54]
In an attempt to better understand the development ofthe inflammatory process unleashed by the protein complexpresent in snake venoms many studies have been donesuch as edema induction [55] leukocyte participation [5657] mast cells degranulation [55] participation of variouscytotoxins in the inflammatory system [58 59] participationof cyclooxygenases [60 61] and the participation of venomPLA2s in the inflammatory process [6 51 56 62]
Many studies about PLA2activity in macrophages have
already been done [8 63 64] Little is known about PLA2rsquos
effect on neutrophils however Escocard et al [65] describedan influx of inflammatory cells including many neutrophilsinto the peritoneal cavity of mice after the injection ofBothrops atrox venom The induction of reactive oxygenspecies (ROS) cytokines like IL-6 and IL-1120573 was seen inthese neutrophils These data were also observed in a studycarried out by Souza et al [26] where besides the influx ofneutrophils into the peritoneal cavity ofmice after injection ofthe venom of B atrox there was also induction of superoxideby these cells mast cell degranulation and phagocytosisby macrophages Regarding the activity of PLA
2Gambero
et al [66] have observed the ability of some myotoxins(bothropstoxin-I-II and piratoxin-I) to induce neutrophilchemotaxis in a concentration-dependent manner
In order to evaluate the activation of leukocytes thetoxicity of B mattogrossensis myotoxins on neutrophil cellswas investigated The cells were incubated with differentconcentrations (3 6 and 12 120583gmL) of BmatTX-I (Lys49)BmatTX-II (Lys49) and BmatTX-III (Asp49) myotoxinsduring 1 12 and 24 hours (data not shown)These myotoxinsdid not affect the neutrophilsrsquo viability which agrees withZuliani et al [64] showing low toxicity on thioglycollateelicited mice macrophages
Nonetheless our data showed that neutrophils incubatedwith BmatTX-I and BmatTX-II myotoxins induced therelease of IL-1120573 (Figures 5(a) and 5(b)) Moreover theseresults suggest that phospholipid hydrolysis is not essentialfor the activity observed and argue with the hyphotesis that
8 BioMed Research International
lowastlowast lowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowast lowastlowastlowastlowastlowastlowastlowastlowastlowast
1 10 20 30 40 50(Identity )
100
89
92
92
92
94
94
BmatTX-IBmatTX-IIPrtx-IBOJU-IBthTX-IMjtx-IMtx-Ii
(a)
lowastlowastlowast lowastlowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowastlowast lowastlowastlowast
1 10 20 30 40 50
(Identity )100
78
75
71
65
BmatTX-IIIBmoTX-IBmjeTX-IBthTX-IIPrTX-III
(b)
Figure 4 Comparison of the N-terminal sequence of the snakersquos PLA2s with the PLA
2s isolated from B mattogrossensis venom N-terminal
sequencing of the PLA2s obtained by Edman [13] degradation and alignment done with CLUSTALW2 expressed as of similarity (a)
BmatTX-I BmatTX-II with Lys49 residues in marked sequences (b) BmatTX-III with Asp49 residues in marked sequences Sequencesused for the alignment and their respective access numbers BthTX-I (gi51890398) MjTX-I (gi17368325) Mtx-II (gi390981003) PrTX-I (gi190016174) BOJU-I (gi209572966) BthTX-II (gi1171971) PrTX-III (gi90016174) BmjeTX-I (gi313471399) BmoTX-I (gi221272396)BthTX-I (Lys49) BthTX-II (Asp49) and BOJU-I (Lys49) isolated from the venom of B jararacussu MjTX-I (Lys49) BmoTX-I (Asp49)and BmjeTX-I (Asp49) isolated from the venom of B moojeni Mtx-II (Lys49) isolated from the venom of B brazili PrTX-I (Lys49) andPrTX-III (Asp49) isolated from the venom of B pirajai the ldquolowastrdquo was used to indicate the amino acid residues that are the same between thesequences
other molecular regions distinct from the active site may beinvolved in this effect
PLA2s are multifunctional proteins that can be used as
mediators in several areas of medicine such as in the treat-ment of rheumatoid arthritis as a new class of HIV inhibitorsby blocking the host cell invasion as a potential treatmentagainst malaria and as an antibiotic by inducing cytotoxicityvia the disruption of bacterial membranes [21 23 30 6768] A study by Costa et al [67] with PLA
2s isolated from
B brazili MTX-I and II demonstrated cytotoxic activityagainst Jurkat tumor cells as well as antimicrobial activityagainstE coli andC albicans and antiparasitic activity againstLeishmania sp
In the present study we evaluated the cytotoxic activityof PLA
2s isolated from B mattogrossensis on JURKAT (T
leukemia) and SK-BR-3 (breast adenocarcinoma) both ofwhich are human tumor cell lines Like MTX-II of B brazili[67] the cytotoxic activity of PLA
2s BmatTX-I and BmatTX-
II on JURKATcells was independent of their catalytic activitysince these are characterized as Lys49 PLA
2s and are therefore
catalytically inactive BmatTX-III characterized as Asp49despite being enzymatically active also showed a lower level oftoxicity Some authors propose that the cytotoxic activity ontumor cell lines is associated with the induction of apoptosisconsidering the fact that PLA
2promotes alterations in the
cell membrane And some studies involving Lys49 PLA2s
isolated from B asper demonstrated that the C-terminal
region comprised of amino acids 115ndash129 is concerned withthe cytotoxic and bactericidal activities of this protein [69ndash71] The same observation was made by Costa et al [67] withsynthetic peptides derived from the C-terminal portion ofMTX-I and II PLA
2s
We also evaluated the antiparasitic activity of the crudevenom and isolated PLA
2of B mattogrossensis on promastig-
ote forms of L amazonensis After the analysis it wasobserved that the crude venom of B mattogrossensis presentsincreasing toxic activity (approximately 50 to 80) againstpromastigote forms of L amazonensis after 48 h of incubation(Figure 7(a)) The PLA
2s isolated from B mattogrossensis at
100 120583gmL characterized as BmatTX-I (Lys49) and BmatTX-III (Asp49) presented toxic activity between 25 and 30respectively even with values close to those presented afterincubation of the protozoan with Pentamidine a drug usedas a positive control (Figures 7(b) and 7(c))
When compared the cytotoxicity values of PLA2s against
promastigote forms of L amazonensis show similar activitybetween the Lys49-PLA
2and the Asp49-PLA
2 Comparison
of this activity with the crude venom (70) showed thatPLA2s are responsible for almost half of the observed effect
Nonetheless notably the results suggest that other toxinspresent in the venom contribute to the parasitersquos death
Similar to the results obtained in the present studyStabeli et al [21] demonstrated that MjTX-II a Lys49-PLA
2
homologue isolated from B moojeni venom in different
BioMed Research International 9
0
500
1000
1500
2000
lowast
lowast
lowast
lowast
lowast
Control 3 6 12 3 6 12 3 6 12
ControlBmatTX-I (Lys49)
BmatTX-II (Lys49)BmatTX-III (Asp49)
BmatTX-I (120583g) BmatTX-II (120583g) BmatTX-III (120583g)
IL-1120573
(pg
mL)
(a)
Control 3 6 12 3 6 12 3 6 120
500
1000
1500
2000
ControlBmatTX-I (Lys49)
BmatTX-II (Lys49)BmatTX-III (Asp49)
lowastlowastlowast
BmatTX-I (120583g) BmatTX-II (120583g) BmatTX-III (120583g)
IL-1120573
(pg
mL)
(b)
Figure 5 Production of IL-1120573 by mice neutrophils These cells wereincubatedwith PLA
2(3 6 and 12 120583gmL) or RPMI (control) for 12
hours (a) and 24 hours (b) at 37∘C in a humid atmosphere with 5CO2 Quantification was done by ELISA as described in 29 Each
bar represents the average +minus SDof three independent groups lowast119875 lt005 compared to the control
concentrations (5 25 and 75 120583g) was effective as a parasiticideagent against Schistosomamansoni and promastigote forms ofLeishmania (L amazonensis L braziliensis L donovani andL major)
The action of the PLA2s BmatTX-I (Lys49) and
BmatTX-III (Asp49) on promastigote forms of L ama-zonensis was independent of its catalytic activity sincecatalytically inactive Lys49 myotoxin also demonstrated tox-icity against Leishmania It is believed that the observedcytotoxic activity might be related to the C-terminal regionsof these phospholipase-homologues that are able to promotea disturbance in the cellular membranes independent oftheir catalytic activity [67 72] However more studies arenecessary to define the exact mechanism of action of theseenzymes on parasites
Growing interest in the comprehension of the structureand function of snake venom components especially PLA
2
10 100 10 100 10 100 10 1000
20
40
60
80
100
BmatTX-I (Lys49)BmatTX-II (Lys49)
BmatTX-III (Asp49)Methotrexate (MTX)
Cyto
toxi
city
()
Concentration (120583gmL)
(a)
10 100 10 100 10 100 10 1000
20
40
60
80
100
BmatTX-I (Lys49)BmatTX-II (Lys49)
BmatTX-III (Asp49)Methotrexate (MTX)
Cyto
toxi
city
()
Concentration (120583gmL)
(b)
Figure 6 Antitumoral activity of PLA2s from B mattogrossensis
(a) Antitumoral activity on human acute T-cell leukemia (JURKAT)lines (b) Antitumoral activity on human breast adenocarcinoma(SK-BR-3) Different concentrations of the PLA
2s were incubated
with cell linesMethotrexate was used as the positive control Resultsare presented as mean +minus SD (119899 = 3)
contributes to a better understanding of the mechanism ofaction of their enzymatic and toxic activities It opens thepath to better understand the intoxication caused by enven-omation and the physiopathology behind its side effectsFuture studies will potentially improve serum therapy andhelp develop the pharmaceutical potential that moleculesisolated from animal venoms can have such as the PLA
2s
isolated from the B mattogrossensis venom which show anti-Leishmania and antitumor activities
4 Conclusion
In conclusion the venom of Bothrops mattogrossensis hasa qualitatively similar toxicological profile to previously
10 BioMed Research International
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineB mattogrossensis crude venom
Ant
i-leis
hman
ia ac
tivity
()
B mattogrossensis crude venom (120583g)
(a)
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineBmatTX-I (Lys49)
Ant
i-leis
hman
ia ac
tivity
()
BmatTX-I (120583g)
lowastlowast lowast lowast
(b)
Ant
i-leis
hman
ia ac
tivity
()
Pent 36 625 125 25 50 1000
20
40
60
80
100
BmatTX-III (Asp49)Pentamidine
BmatTX-III (120583g)
lowastlowastlowast
lowastlowast lowast
(c)
Figure 7 Antileishmanial activity of the crude venom of and PLA2s from B mattogrossensis The promastigote forms of L amazonensiswere
plated with 1times105 cellswellThen different concentrations of crude venom and isolated PLA2s were added to each wellThe experiment was
done in a 48 h period with the (a) crude venom and the PLA2enzymes (b) BmatTX-I and (c) BmatTX-III MTT was added and after the
incubation period at 33∘C the formazan crystal formed was dissolved in SDS Readings were done in a spectrophotometer at 570 nm Eachbar represents the average +minus SD of the three independent experimental groups sixfold total lowast119875 lt 005 compared to the control
studied snake venoms of the Bothrops sp genera despite theobservation of quantitative variations Of the three basicPLA2s from B mattogrossensis venom now isolated for the
first time two are characterized as Lys49-PLA2homologues
BmatTX-I and -II and the other as an Asp49-PLA2 named
BmatTX-III This showed high phospholipase activity ThePLA2s isolated induced myotoxic effects as well as the release
of proinflammatory cytokines by neutrophils BmatTX-I and-III PLA
2s were cytotoxic to human tumor cell lines JURKAT
and SK-BR-3 and showed activity against promastigote formsof L amazonensis
Conflict of Interests
The authors state that there is no conflict of interests
Acknowledgments
The authors are grateful to the Ministry of Science andTechnology (MCTI) Conselho Nacional de Desenvolvi-mento Cientıfico e Tecnologico (CNPq) Financiadora deEstudos e Projetos (FINEP) Fundacao de Tecnologia doAcre (FUNTACFDCT) Coordenacao de Aperfeicoamentode Nıvel Superior (CAPES)mdashProjeto NanoBiotec Redede Biodiversidade e Biotecnologia da Amazonia Legal(BIONORTECNPqMCTI) Instituto Nacional paraPesquisa Translacional em Saude e Ambiente na RegiaoAmazonica (INCT-INPeTAmCNPqMCTI) and InstitutoNacional para Pesquisa em Toxinas (INCT-Tox) andSecretary of Development of Rondonia State (SEPLANPRONEXCNPq) for financial support We are grateful to
BioMed Research International 11
Sr Guilherme G Galassi (Mantenedouro da Fauna Silvestreautorizacao IBAMA 020270089912001-95 Americana-SP)for their helpful technical collaboration in provide the snakevenom Amy Grabner provided the English editing of thepaper
References
[1] Y Angulo and B Lomonte ldquoBiochemistry and toxicology oftoxins purified from the venom of the snake Bothrops asperrdquoToxicon vol 54 no 7 pp 949ndash957 2009
[2] J J Calvete P Juarez and L Sanz ldquoSnake venomics Strategyand applicationsrdquo Journal of Mass Spectrometry vol 42 no 11pp 1405ndash1414 2007
[3] T S Kang D Georgieva N Genov et al ldquoEnzymatic toxinsfrom snake venom structural characterization and mechanismof catalysisrdquoThe FEBS Journal vol 278 no 23 pp 4544ndash45762011
[4] C ZOliveiraCaracterizacao funcional e estrutural de inibidoresde fosfolipases A
2isolados do plasma de serpente Bothrops
jararacussu [PhD thesis] Faculdade de Ciencias Farmaceuticasde Ribeirao Preto Ribeirao Preto Brazil 2009
[5] R G Stabeli R Simoes-Silva A M Kayano et al ldquoPurifi-cation of phospholipases A
2from American snake venomsrdquo
in ChromatographymdashThe Most Versatile Method of ChemicalAnalysis chapter 1 pp 1ndash34 InTech Vienna Austria 2012
[6] J P Zuliani C M Fernandes S R Zamuner J M Gutierrezand C F P Teixeira ldquoInflammatory events induced by Lys-49 and Asp-49 phospholipases A
2isolated from Bothrops asper
snake venom role of catalytic activityrdquo Toxicon vol 45 no 3pp 335ndash346 2005
[7] S Marcussi R G Stabeli N A Santos-Filho et al ldquoGenotoxiceffect of Bothrops snake venoms and isolated toxins on humanlymphocyte DNArdquo Toxicon vol 65 pp 9ndash14 2013
[8] S S Setubal A S Pontes J L Furtado et al ldquoAction oftwo phospholipases A
2purified from Bothrops alternatus snake
venom on macrophagesrdquo Biochemistry vol 78 no 2 pp 194ndash203 2013
[9] J M Gutierrez L A Ponce-Soto S Marangoni and BLomonte ldquoSystemic and local myotoxicity induced by snakevenom group II phospholipases A
2 comparison between cro-
toxin crotoxin B and a Lys49 PLA2homologuerdquo Toxicon vol
51 no 1 pp 80ndash92 2008[10] A Q Rueda I G Rodrıguez E C Ara et al ldquoBiochemical
characterization action onmacrophages and superoxide anionproduction of four basic phospholipases A
2from Panamanian
Bothrops asper snake venomrdquo BioMed Research Internationalvol 2013 Article ID 789689 9 pages 2013
[11] V X da Silva and M T Rodrigues ldquoTaxonomic revision ofthe Bothrops neuwiedi complex (Serpentes Viperidae) withdescription of a new speciesrdquo Phyllomedusa vol 7 no 1 pp 45ndash90 2008
[12] J A Campbell and W W Lamar The Venomous Reptiles of theWestern Hemisphere Comstock Ithaca NY USA 2004
[13] P Edman ldquoMethod for determination of the amino acidsequence in peptidesrdquo Acta Chemica Scandinavica vol 4 pp283ndash293 1950
[14] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[15] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[16] M Holzer and S P Mackessy ldquoAn aqueous endpoint assay ofsnake venom phospholipase A
2rdquo Toxicon vol 34 no 10 pp
1149ndash1155 1996[17] A L Fuly A L P deMiranda R B Zingali and J AGuimaraes
ldquoPurification and characterization of a phospholipase A2isoen-
zyme isolated from Lachesis muta snake venomrdquo BiochemicalPharmacology vol 63 no 9 pp 1589ndash1597 2002
[18] T Nikai N Mori M Kishida H Sugihara and A T Tu ldquoIso-lation and biochemical characterization of hemorrhagic toxinf from the venom of Crotalus atrox (Western DiamondbackRattlesnake)rdquo Archives of Biochemistry and Biophysics vol 231no 2 pp 309ndash319 1984
[19] J A Gene A Roy G Rojas J M Gutierrez and L CerdasldquoComparative study on coagulant defibrinating fibrinolyticand fibrinogenolytic activities of Costa Rican crotaline snakevenoms and their neutralization by a polyvalent antivenomrdquoToxicon vol 27 no 8 pp 841ndash848 1989
[20] M S Charney and R M Tomarelli ldquoA colorimetric method forthe determination of the proteolytic activity of duodenal juicerdquoThe Journal of Biological Chemistry vol 171 pp 501ndash505 1947
[21] R G Stabeli S F Amui C D SantrsquoAna et al ldquoBothrops moojenimyotoxin-II a Lys49-phospholipaseA
2homologue an example
of function versatility of snake venom proteinsrdquo ComparativeBiochemistry and Physiology C vol 142 no 3-4 pp 371ndash3812006
[22] D R Call J A Nemzek S J Ebong G L Bolgos D ENewcomb and D G Remick ldquoRatio of local to systemicchemokine concentrations regulates neutrophil recruitmentrdquoThe American Journal of Pathology vol 158 no 2 pp 715ndash7212001
[23] P G Roberto S Kashima S Marcussi et al ldquoCloning andidentification of a complete cDNA coding for a bactericidal andantitumoral acidic phospholipase A
2from Bothrops jararacussu
venomrdquoThe Protein Journal vol 23 no 4 pp 273ndash285 2004[24] J H Schumacher A OrsquoGarra B Shrader et al ldquoThe charac-
terization of four monoclonal antibodies specific for mouse IL-5 and development of mouse and human IL-5 enzyme-linkedimmunosorbentrdquoThe Journal of Immunology vol 141 no 5 pp1576ndash1581 1988
[25] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[26] C A T Souza A M Kayano S S Setubal et al ldquoLocal andsystemic biochemical alterations induced by Bothrops atroxsnake venom in micerdquo Journal of Venom Research vol 3 pp28ndash34 2012
[27] J M Gutierrez and B Lomonte ldquoPhospholipase A2myotoxins
from Bothrops snake venomsrdquo Toxicon vol 33 no 11 pp 1405ndash1424 1995
[28] J M Gutierrez and A Rucavado ldquoSnake venom metallopro-teinases their role in the pathogenesis of local tissue damagerdquoBiochimie vol 82 no 9-10 pp 841ndash850 2000
[29] T Matsui Y Fujimura and K Titani ldquoSnake venom proteasesaffecting hemostasis and thrombosisrdquo Biochimica et BiophysicaActa vol 1477 no 1-2 pp 146ndash156 2000
[30] A M Soares M R M Fontes and J R Giglio ldquoPhospholipaseA2myotoxins from Bothrops snake venoms structure-function
12 BioMed Research International
relationshiprdquoCurrent Organic Chemistry vol 8 no 17 pp 1677ndash1690 2004
[31] VM Rodrigues SMarcussi R S Cambraia et al ldquoBactericidaland neurotoxic activities of two myotoxic phospholipases A
2
from Bothrops neuwiedi pauloensis snake venomrdquo Toxicon vol44 no 3 pp 305ndash314 2004
[32] N A Santos-Filho L B Silveira C Z Oliveira et al ldquoAnew acidic myotoxic anti-platelet and prostaglandin I
2induc-
tor phospholipase A2isolated from Bothrops moojeni snake
venomrdquo Toxicon vol 52 no 8 pp 908ndash917 2008[33] A M Soares S H Andriao-Escarso Y Angulo et al ldquoStruc-
tural and functional characterization of myotoxin I a Lys49phospholipaseA
2homologue fromBothropsmoojeni (Caissaca)
snake venomrdquo Archives of Biochemistry and Biophysics vol 373no 1 pp 7ndash15 2000
[34] A M Soares S H Andriao-Escarso R K Bortoleto et alldquoDissociation of enzymatic and pharmacological properties ofpiratoxins-I and -III two myotoxic phospholipases A
2from
Bothrops pirajai snake venomrdquo Archives of Biochemistry andBiophysics vol 387 no 2 pp 188ndash196 2001
[35] S S Teixeira L B Silveira F M N da Silva et al ldquoMolecularcharacterization of an acidic phospholipase A
2from Bothrops
pirajai snake venom synthetic C-terminal peptide identifies itsantiplatelet regionrdquo Archives of Toxicology vol 85 no 10 pp1219ndash1233 2011
[36] S H Andriao-Escarso A M Soares V M Rodrigues et alldquoMyotoxic phospholipases A
2in Bothrops snake venoms effect
of chemical modifications on the enzymatic and pharmacolog-ical properties of bothropstoxins from Bothrops jararacussurdquoBiochimie vol 82 no 8 pp 755ndash763 2000
[37] S H Andriao-Escarso A M Soares M R M Fontes et alldquoStructural and functional characterization of an acidic plateletaggregation inhibitor and hypotensive phospholipase A
2from
Bothrops jararacussu snake venomrdquo Biochemical Pharmacologyvol 64 no 4 pp 723ndash732 2002
[38] M E G Denegri O C Acosta S Huancahuire-Vega et alldquoIsolation and functional characterization of a new acidic PLA
2
Ba SpII RP4 of the Bothrops alternatus snake venom fromArgentinardquo Toxicon vol 56 no 1 pp 64ndash74 2010
[39] R Zouari-Kessentini J Luis A Karray et al ldquoTwo purified andcharacterized phospholipases A
2from Cerastes cerastes venom
that inhibit cancerous cell adhesion and migrationrdquo Toxiconvol 53 no 4 pp 444ndash453 2009
[40] R Shirai M Toriba K Hayashi K Ikeda and S Inoue ldquoIden-tification and characterization of phospholipase A
2inhibitors
from the serum of the Japanese rat snake Elaphe climacophorardquoToxicon vol 53 no 6 pp 685ndash692 2009
[41] M I Homsi-Brandenburgo L S Queiroz H Santo-Neto LRodrigues-Simoni and J R Giglio ldquoFractionation of Bothropsjararacussu snake venom partial chemical characterization andbiological activity of bothropstoxinrdquo Toxicon vol 26 no 7 pp615ndash627 1988
[42] G P Fernandez A Segura M Herrera et al ldquoNeutralizationof Bothrops mattogrossensis snake venom from Bolivia exper-imental evaluation of llama and donkey antivenoms producedby caprylic acid precipitationrdquoToxicon vol 55 no 2-3 pp 642ndash645 2010
[43] C L Ownby H S S de Araujo S P White and J E FletcherldquoLysine 49 phospholipase A
2proteinsrdquo Toxicon vol 37 no 3
pp 411ndash445 1999[44] A M Soares W P Sestito S Marcussi et al ldquoAlkylation of
myotoxic phospholipases A2in Bothrops moojeni venom a
promising approach to an enhanced antivenom productionrdquoInternational Journal of Biochemistry amp Cell Biology vol 36 no2 pp 258ndash270 2004
[45] B Lomonte J M Gutierrez M F Furtado et al ldquoIsolationof basic myotoxins from Bothrops moojeni and Bothrops atroxsnake venomsrdquo Toxicon vol 28 no 10 pp 1137ndash1146 1990
[46] L C Mancuso M M Correa C A Vieira et al ldquoFractionationof Bothrops pirajai snake venom isolation and characterizationof piratoxin-I a new myotoxic proteinrdquo Toxicon vol 33 no 5pp 615ndash626 1995
[47] H S Selistre L S Queiroz O A B Cunha G E P deSouza and J R Giglio ldquoIsolation and characterization ofhemorrhagic myonecrotic and edema-inducing toxins fromBothrops insularis (jararaca ilhoa) snake venomrdquo Toxicon vol28 no 3 pp 261ndash273 1990
[48] E A Dennis J Cao Y-H Hsu V Magrioti and G KokotosldquoPhospholipaseA
2enzymes physical structure biological func-
tion disease implication chemical inhibition and therapeuticinterventionrdquo Chemical Reviews vol 111 no 10 pp 6130ndash61852011
[49] A G T Cardoso Acao do veneno de Bothrops moojeni e suafracao L-aminoacido oxidase submetida ao tratamento comraios gama de 60CO em Leishmania spp [MS thesis] IPEN-CNENSP Sao Paulo Brazil 1999
[50] MGallacci andWLGCavalcante ldquoUnderstanding the in vitroneuromuscular activity of snake venomLys49 phospholipaseA
2
homologuesrdquo Toxicon vol 55 no 1 pp 1ndash11 2010[51] S S Teixeira Caracterizacao funcional e estrutural de uma
fosfolipase A2acida isolada do veneno de Bothrops pirajai
[MS thesis] Faculdade de Ciencias Farmaceuticas de RibeiraoPretoUSP Ribeirao Preto Brazil 2009
[52] E Valentin and G Lambeau ldquoIncreasing molecular diversityof secreted phospholipases A
2and their receptors and binding
proteinsrdquo Biochimica et Biophysica Acta vol 1488 no 1-2 pp59ndash70 2000
[53] J M Gutierrez and B Lomonte ldquoPhospholipases A2from
Viperidae snake venoms how do they induce skeletal muscledamagerdquo Acta Chimica Slovenica vol 58 no 4 pp 647ndash6592011
[54] J M Gutierrez C L Ownby and G V Odell ldquoIsolation of amyotoxin from Bothrops asper venom partial characterizationand action on skeletal musclerdquo Toxicon vol 22 no 1 pp 115ndash128 1984
[55] N G Nascimento M C Sampaio R A Olivo and C TeixeiraldquoContribution of mast cells to the oedema induced by Bothropsmoojeni snake venom and a pharmacological assessment of theinflammatory mediators involvedrdquo Toxicon vol 55 no 2-3 pp343ndash352 2010
[56] C F P Teixeira E C T Landucci E Antunes M Chacurand Y Cury ldquoInflammatory effects of snake venom myotoxicphospholipases A
2rdquo Toxicon vol 42 no 8 pp 947ndash962 2003
[57] S R Zamuner and C F P Teixeira ldquoCell adhesion moleculesinvolved in the leukocyte recruitment induced by venom of thesnake Bothrops jararacardquoMediators of Inflammation vol 11 no6 pp 351ndash357 2002
[58] F Chaves C F P Teixeira and J M Gutierrez ldquoRole ofTNF-120572 IL-1120573 and IL-6 in the local tissue damage induced byBothrops asper snake venom an experimental assessment inmicerdquo Toxicon vol 45 no 2 pp 171ndash178 2005
BioMed Research International 13
[59] C D F P Teixeira C M Fernandes J P Zuliani and SF Zamuner ldquoInflammatory effects of snake venom metallo-proteinasesrdquo Memorias do Instituto Oswaldo Cruz vol 100supplement 1 pp 181ndash184 2005
[60] R D A Olivo C F P Teixeira J LWallace J M Gutierrez andS R Zamuner ldquoRole of cyclooxygenases in oedema-formingactivity of bothropic venomsrdquo Toxicon vol 49 no 5 pp 670ndash677 2007
[61] L B Silveira D P Marchi-Salvador N A Santos-Filho et alldquoIsolation and expression of a hypotensive and anti-plateletacidic phospholipase A
2from Bothrops moojeni snake venomrdquo
Journal of Pharmaceutical and Biomedical Analysis vol 73 pp35ndash43 2013
[62] A Rucavado T Escalante C F P Teixeira C M Fernandes CDıaz and J M Gutierrez ldquoIncrements in cytokines and matrixmetalloproteinases in skeletal muscle after injection of tissue-damaging toxins from the venom of the snake Bothrops asperrdquoMediators of Inflammation vol 11 no 2 pp 121ndash128 2002
[63] E Leiguez J P Zuliani A M Cianciarullo C M Fernandes JM Gutierrez and C Teixeira ldquoA group IIA-secreted phospho-lipase A
2from snake venom induces lipid body formation in
macrophages the roles of intracellular phospholipases A2and
distinct signaling pathwaysrdquo Journal of Leukocyte Biology vol90 no 1 pp 155ndash166 2011
[64] J P Zuliani J M Gutierrez L L Casais e Silva S C SampaioB Lomonte and C D F P Teixeira ldquoActivation of cellularfunctions in macrophages by venom secretory Asp-49 and Lys-49 phospholipasesA
2rdquoToxicon vol 46 no 5 pp 523ndash532 2005
[65] R D C M Escocard M M Kanashiro J H Petretski et alldquoNeutrophils regulate the expression of cytokines chemokinesand nitric oxide synthasenitric oxide in mice injected withBothrops atrox venomrdquo Immunobiology vol 211 no 1-2 pp 37ndash46 2006
[66] A Gambero E C T Landucci M H Toyama et al ldquoHumanneutrophil migration in vitro induced by secretory phospholi-pases A
2 a role for cell surface glycosaminoglycansrdquo Biochemi-
cal Pharmacology vol 63 no 1 pp 65ndash72 2002[67] F L S Costa R S Rodrigues L F M Izidoro et al ldquoBio-
chemical and functional properties of a thrombin-like enzymeisolated from Bothrops pauloensis snake venomrdquo Toxicon vol54 no 6 pp 725ndash735 2009
[68] J C Villalobos R Mora B Lomonte J M Gutierrez andY Angulo ldquoCytotoxicity induced in myotubes by a Lys49phospholipase A
2homologue from the venom of the snake
Bothrops asper evidence of rapid plasma membrane damageand a dual role for extracellular calciumrdquo Toxicology in Vitrovol 21 no 8 pp 1382ndash1389 2007
[69] B S Cummings J McHowat and R G Schnellmann ldquoPhos-pholipase A
2s in cell injury and deathrdquoThe Journal of Pharma-
cology and Experimental Therapeutics vol 294 no 3 pp 793ndash799 2000
[70] S R Panini L Yang A E Rusinol M S Sinensky J VBonventre and C C Leslie ldquoArachidonate metabolism andthe signaling pathway of induction of apoptosis by oxidizedLDLoxysterolrdquo Journal of Lipid Research vol 42 no 10 pp1678ndash1686 2001
[71] L Paramo B Lomonte J Pizarro-Cerda J-A Bengoechea J-P Gorvel and E Moreno ldquoBactericidal activity of Lys49 andAsp49 myotoxic phospholipases A
2from Bothrops asper snake
venom synthetic Lys49myotoxin II-(115-129)-peptide identifiesits bactericidal regionrdquo European Journal of Biochemistry vol253 no 2 pp 452ndash461 1998
[72] B Lomonte Y Angulo and E Moreno ldquoSynthetic peptidesderived from the C-terminal region of Lys49 phospholipaseA2homologues from viperidae snake venoms biomimetic
activities and potential applicationsrdquo Current PharmaceuticalDesign vol 16 no 28 pp 3224ndash3230 2010
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

2 BioMed Research International
catalyze the hydrolysis of the 2-acyl bond of cell membranephospholipids releasing free fatty acids such as arachidonicacid and lysophospholipids PLA
2s have been characterized
as the major component of snake venoms being responsiblefor several pathophysiological effects caused by snake enven-omation such as neurotoxic cardiotoxic myotoxic cyto-toxic hypotensive and anticoagulant activities triggering anintense inflammatory reaction with the release of cytotoxinsand eicosanoids [4ndash6] PLA
2rsquos involvement in a variety of
inflammatory diseases and accidents caused by venomousanimals has raised medical and scientific interest in thisenzyme [7 8]
Myotoxic PLA2s from the Bothrops species are composed
of approximately 110 to 135 amino acid residues and canbe divided into two groups ldquoclassicalrdquo which contain anaspartate residue at position 49 (Asp49) and catalyze esterbond hydrolysis at the glycerophospholipid sn-2 position ina Ca2+-dependent manner and ldquovariantrdquo which contain alysine residue at the same position (Lys49) This substitutionaffects the ability of these proteins to bind to Ca2+ which isan essential cofactor for catalysis leading to decreased or nocatalytic activity [1ndash5 7 9 10]
The Bothrops mattogrossensis snake belongs to the Both-rops neuwiedi complex [11]This snake is found in the easternregion of South America including Bolivia Brazil SoutheastPeru Paraguay Uruguay and Argentina [12] The presentstudy describes for the first time the isolation identificationand functional characterization of three myotoxic phospho-lipases A
2 named BmatTX-I (Lys49 PLA
2-like) BmatTX-
II (Lys49 PLA2-like) and BmatTX-III (Asp49 PLA
2) and
evaluates their activity against Leishmania and tumor cells
2 Materials and Methods
21 Venom The venom from the Bothrops mattogrossensissnake was acquired from Serpentario de Proteınas BioativasBatatais-SP Brazil This study was authorized by InstitutoBrasileiro do Meio Ambiente e dos Recursos NaturaisRenovaveismdashIBAMA Instituto Chico Mendes de Conser-vacao da BiodiversidademdashICMBio (number 27131-1) andConselho de Gestao do Patrimonio GeneticomdashCGENBrazil(number 0106272011-1)
22 Animals Swiss male mice (18ndash20 g) were used Theseanimals were housed in temperature-controlled rooms andreceived water and food ad libitum until used Animalcare was in accordance with the guidelines of the BrazilianCollege for Animal Experimentation (COBEA) and wasapproved by the Committee of Ethics on Animal Utiliza-tion in Research from the Institute of Research of Tropi-cal Pathologies (IPEPATROFIOCRUZ-Rondonia) (protocolnumber 20121)
23 Bothrops mattogrossensis Venom Fractioning Around250mg of B mattogrossensis venom was eluted in 12mL ofsterile deionized water and centrifuged at 1530timesg for 10min-utes and then submitted to cation exchange chromatographyin anAkta purifier 10 (GE-Healthcare) using aCM-Sepharose
column (27 times 300mm GE Healthcare) equilibrated with50mM ammonium bicarbonate buffer pH 80 and elutedwith a linear gradient of 0ndash100 500mM ammonium bicar-bonate pH 80 at a flow rate of 50mLminute The frac-tions were monitored at an absorbance of 280 nm collectedmanually identified lyophilized and stored at minus20∘C Allfractions were submitted to SDS-PAGE and enzymatic activ-ity analyses The isolation of the three PLA
2s was obtained
by liquid chromatography using a Discovery C18 column(25 times 46mm Supelco) equilibrated with deionized waterwith 01 trifluoroacetic acid (vv) and eluted with tenvolumes of a linear gradient of 0ndash100 acetonitrile with 01trifluoroacetic acid (vv) at a flow rate of 10mLminuteThe elution was monitored at an absorbance of 280 nmmanually collected lyophilized and stored at minus20∘C In orderto determine protein concentration the Bradford method(Bio-Rad) was used with bovine serum albumin (BSA) as astandard [14]
24 Monodimensional Electrophoresis (SDS-PAGE) Poly-acrylamide gel electrophoresis (PAGE) in the presence ofsodium dodecyl sulfate (SDS) was conducted using themethod described by Laemmli [15] The electrophoresis wascarried out at 15mA and 5W using a conventional molecularweight standard (BioLabs) and a Lys49-PLA
2homologue
isolated from B jararacussu snake venom (BthTX-I) asmarkers The gel was stained with Coomassie Brilliant Blue(CBB)G-250 and images of the gel were obtained using ImageScanner (GE Healthcare)
25 Bidimensional Electrophoresis (2D-SDS-PAGE) and pIDetermination Theprotein fractions were submitted to elec-trofocusing in 13 cm strips with pHs ranging from 3 to 10in a nonlinear form using an Ettan IPGphor 3 IEF System(GE Healthcare) At the end of the electrofocusing thestrips containing the PLA
2s were equilibrated and transferred
to a 125 polyacrylamide gel where they were separatedaccording to molecular mass The electrophoretic run wasperformed at 25mA and 100W during 55 hours and atthe end of the protein separation the gel was stained withCoomassie Brilliant Blue (CBB) G-250 and images of the gelwere captured using Image Scanner (Amershan Bioscience)
26 Mass Spectrometry (MALDI-TOF) The samples (1 120583L)were mixed with a matrix solution composed of sinapinicacid and acetonitrile with 01 trifluoroacetic solubilized ata 1 1 ratio The average mass of the protein was obtained in aMALDI-TOF system (Shimatzu Biotech) operated in linearmode using insulin (57345Da) cytochrome C (123619Da)apomyoglobin (169522Da) aldolase (392122Da) and albu-min (664300Da) as calibrators The mass spectra obtainedwere submitted to automatic baseline subtraction
27 N-Terminal Sequencing and Similarity Search The aminoterminal sequence of each isolated PLA
2previously deter-
mined by automated Edman degradation was used to searchfor sequence similarity Approximately 50120583g of each isolatedPLA2corresponding to approximately 2 nmolsmL sample
BioMed Research International 3
was submitted for N-terminal amino acid sequencing usingthe automated Edman degradation method [13] on a PPSQ-33A (Shimadzu) automatic sequencer A sequence similaritysearch and multiple sequence alignment were performed inthe SWISS-PROTTREMBL database using the programsFASTA BLAST and CLUSTALW2
28 Functional Characterization281 Phospholipase Activity with 4-nitro-3(octanoyloxy)ben-zoic (4N3OBA) Phospholipase activity of the three PLA
2s
was assayed following the protocols described by Holzer andMackessy [16] modified for 96-well plates The standardassay mixture contained 200120583L of buffer (10mM Tris-HClplus 10mM CaCl
2 and 100mM NaCl pH 80) 20 120583L of
4-nitro-3(octanoyloxy)benzoic (4N3OBA ndash Biomol EUA)20120583L of deionized water and 20120583L of PLA
2(5 120583g) After
the addition of the PLA2s the mixture was incubated for
30 minutes at 37∘C and the absorbance was determined at425 nmusing an Eon (Biotek)microplate spectrophotometerfor 3-minute intervals The enzymatic activity was expressedas the reactionrsquos initial velocity (119881
119900) calculated based on the
increase in absorbance
282 Phospholipase Activity with Fluorescent SubstratePhospholipase A
2rsquos (PLA
2) enzymatic activity was evaluated
through the hydrolysis of synthetic fluorescent phospholipidusing the fluorescent substrate Acyl 6 0 NBD phospholipidNBD-phosphatidylcholine (NBD-PC) (Avanti Polar LipidsInc Alabaster AL USA) The assay was performed in aspectrofluorometer (Shimadzu RF-5301PC software RFPC)with excitation and emission wavelengths of 460 and 534respectivelyThe enzymatic activity of each Bmattogrossensischromatographic fraction (7 12 15 16 17 and 20) wasevaluated over 250 seconds after the addition of substrate(33 120583gmL final concentration) in a reaction media con-taining 50mM Tris-HCl and 8mM CaCl
2at pH 75 at room
temperature
283 Platelet Aggregation A platelet aggregation assay wascarried out according to the process described by Fuly et al[17] with modifications Platelet aggregation was monitoredin an Aggregometer (Chrono Logmodel 490 2D HavertownUSA) using Platelet-Rich-Plasma (PRP) PRP was obtainedfrom human whole blood of health volunteers (CAAE14204413500000011) BmatTX-III (rechromatography frac-tion 15)was incubatedwith PRP for fiveminutes at 37∘Cbeingstirred constantly and then platelet aggregation was trig-gered by the addition of ADP (15120583M) or collagen (16 120583gmL)Assays were performed at 37∘C in siliconized glass cuvettesin a final volume of 300 120583L Control experiments wereperformed by adding agonists in the absence of peak 15 Onehundred percent (100) platelet aggregation was obtainedwith a supramaximal concentration of each agonist anddetermined 6 minutes after the addition of each while PRPrsquoslight transmittance showed 0 aggregation
284 Hemorrhage Activity Micewere injected intradermallyin the dorsal region with 50 120583g of crude venom dissolved
in 50120583L of physiological solution [18] Controls received50 120583L of physiological solution in identical conditions After 3hours the animals were euthanized through cervical disloca-tion and the skin was removed The hemorrhagicactivitywasexpressed as size of thehemorrhagic area on the inner surfacemeasured in mm2
285 Coagulation Activity and Minimal Coagulation Dose(MCD)Determination Coagulation activity was tested usinga methodology previously described by Gene et al [19] Inbrief 200120583L of plasma frommicewas distributed in a 96-wellplate and 10 120583L of crude venom containing different concen-trations (0312 0625 125 25 5 and 10 120583g) of proteins wasadded The plate was placed in a thermostatically controlledenvironment and the optical density was measured every 3seconds at 600 nm using an Eonmicroplate spectrophotome-ter (Biotek) in order to evaluate the smallest concentration ofvenom able to coagulate 200120583L of plasmaminute
286 Proteolytic Activity Proteolytic activitywas ascertainedusing azocasein (Sigma) as a substrate according to the pro-cedure described by Charney and Tomarelli [20] Azocaseinsolubilized in distilled water (150 120583L) was added to 7 120583L ofthe crude venom in different protein concentrations (03120625 125 25 5 and 10 120583g) and then the mixture wasincubated in a water bath at 37∘C for 1 hour The reactionstopped when 150 120583L of 20 (mv) trichloroacetic acid wasadded which was followed by incubation for 30min andcentrifugation at 10000timesg for 10 more minutesThen 100 120583Lof the supernatant was transferred to a multiple-well plateand the absorbance was measured at 440 nm using an Eonmicroplate spectrometer (Biotek) and one enzymatic unit (U)was defined as the amount of enzyme necessary to increasethe absorbance by 005
287Myotoxic Activity Myotoxic activity was determined bymeasuring the creatine kinase (CK) activity in the plasma[21] Groups of mice were injected in the gastrocnemiusmuscle with 25 120583g of isolated myotoxins diluted in 50120583L ofphosphate buffered saline (PBS) Negative controls receivedan injection of 50 120583L of PBSThree hours after the injectionsaliquots of mice blood were collected from the caudal veinin heparinized capillaries and centrifuged at 1530timesg for 20minutes Creatine kinasersquos enzymatic activity was determinedusing the CK-NAC kinetic kit (Bioclin Brazil) accordingto the manufacturerrsquos protocol Absorbance was measuredfor 3 minutes at 37∘C in a spectrophotometer at 340 nmEnzymatic activity was expressed in unitsliter (UL) andeach unit consists of the result of the phosphorylation of onenanomol of creatine per minute
288 Neutrophil Viability Neutrophils were collected 6hours after the intraperitoneal (IP) injection of 15mL of3 thioglycollate sterile solution according to the methodpreviously described by Call et al [22] The animals wereeuthanized in order to collect the cells and the peritonealcavity was washed with 3mL of PBS The predominance of
4 BioMed Research International
neutrophils in the liquid obtained was confirmed by micro-scopic analysis with glass slides stained with a panoptic dyeThe peritoneal neutrophils obtained were suspended in anRPMI culturemedium (Gibco-BRL) supplemented with gen-tamicin (100 120583gmL) L-glutamine (2mM) and 10 bovinefetal serum (SFB) in order to obtain 2times105 cells100 120583L Nextcellular viability was assayed over 1 12 and 24 h at 37∘Cand 5 CO
2in which the cells were incubated in a 96-well
plate with the previously isolated PLA2s at a concentration
of 12 120583gmL using RPMI as a negative control After thisthe samples were centrifuged and the supernatant removedCellular viability was determined using the MTT method[23]
289 Quantification of Cytokine EIA was used to evaluateIL-1120573 cytokine as described by Schumacher et al [24] Brieflythe neutrophils (2 times 105 cells200120583L) were either incubatedwith the isolated proteins at 3 6 and 12 120583gmL concentrations(experimental group) or with PMA (positive control group)or with RPMI (negative control) and kept for 12 and 24 hoursat 37∘C in a humid atmosphere with 5 CO
2 After that 96-
well plates were coatedwith 100 120583L of the capturemonoclonalantibody anti-IL-1120573 and incubated for 18 hours at 37∘C Theplate was then washed with washer buffer (PBSTween20)After that 200120583L of blocking buffer containing 5 bovineserum albumin (BSA) in PBSTween20 was added to thewells and the plates were incubated for 1 hour at 37∘CFollowing this the wells were washed and 50 120583L of eithersamples or standard were dispensed into each well and theplates were incubated for 2 hours at 37∘C After this theplate was washed and 100 120583L of detection antibody anti-IL-1120573 was added for 2 hours at 37∘C After incubation andwashing 100 120583L of streptavidin-peroxidase was added fol-lowed by incubation and addition of the substrate (100120583LmL331015840551015840-tetramethylbenzidine) Finally sulfuric acid (50120583L)was added to stop the reaction Absorbances were recordedat 540 and 450 nm and concentration of IL-1120573 was estimatedfrom standard curves prepared with recombinant cytokineThe results were expressed as pgmL of IL-1120573
2810 Antitumor Activity Cytotoxic activity of isolatedPLA2s on human T-cell leukemia (JURKAT) and breast
adenocarcinoma (SK-BR-3) lines obtained from the Amer-ican Culture Collection of Cells (ATCC American TypeCulture Collection Rockville MD USA) were investigated(Figure 6) This activity was assayed by MTT staining asdescribed by Mosmann [25] and adapted by Stabeli et al[21] Cells were dispensed in 96-well plates at a densityof 5 times 105 cellsmL After 24 h of incubation the mediumwas removed and fresh medium with or without differentconcentrations of PLA
2s (BmatTX-I BmatTX-II BmatTX-
III or methotrexate) was added to the wells and incubatedfor another 24 hours The evaluation of the cytotoxic activitywas measured in a spectrophotometer using an interferencefilter of 570 nm and expressed as a percentage
2811 Anti-Leishmania Activity Promastigote forms ofLeishmania amazonensis (IFLABR67PH8) were dis-pensed in a 96-well plate with 1 times 105 cellswell Different
Table 1 Activities induced by B mattogrossensis snake venom
Effecta ActivityPhospholipase activityb (Umg) 186405Proteolytic activityc (U10120583g) 30 plusmn 01
Hemorrhagic halod (cm2) 333 plusmn 005
Coagulation activitye (MCD 120583g) 0325aAll experiments were carried out in triplicate bActivity using NOB stainedsubstrate cOne enzymatic unit (U) was defined as the quantity of enzymeneeded to increase the absorbance by 005UA440 nm dValues 3 h afterincubation with crude venom of B mattogrossensis (50120583g) eMCD theminimum coagulation dose was the dose capable of coagulating 200120583L ofcitrated plasma in less than a minute
concentrations (312 625 125 25 50 and 100120583gmL) ofthe crude venom of B mattogrossensis and the isolatedPLA2s (BmatTX-I and BmatTX-II) were added to each well
100mgmL of pentamidine was used as a positive controlAfter an incubation period of 48 h 10 120583L of a 5mgmL MTTsolution was added Then the plates were placed in theoven at 33∘C with 5 CO
2for 4 hours of incubation after
which 50120583L of SDS (20 wv) was added Absorbance wasmonitored at 570 nm Results were expressed in toxicitypercentage following the equation 1 minus (DO sampleDOcontrol) times 100
29 Statistical Analysis Results were expressed as mean +minusstandard deviation An ANOVA test was used to evaluate thesignificance of the differences observed with 119875 value le 005considered to be significant
3 Results and Discussion
31 Crude Venom Biological Activities B mattogrossensissnake venom induced hemorrhage coagulation proteolyticand phospholipase activities in vitro (Table 1) The hemor-rhagic activity of B mattogrossensis crude venom was eval-uated based on the dimensions of the average hemorrhagichalo which was 333 cm2 This result is in agreement with theresults recently obtained with B atrox [26]
Coagulation activity was confirmed after incubation ofdifferent concentrations of B mattogrossensis crude venomwith plasma in which theminimum coagulation dose capableof promoting coagulation in less than 1min was 0325120583gof protein Analysis of the proteolytic activity of B mat-togrossensis crude venom demonstrated a concentration-dependent response (data not shown) Regarding phospho-lipase proteolytic coagulating and hemorrhagic activitiesof B mattogrossensis crude venom assays confirmed thepresence and activity of proteases and phospholipases Thepresence of metalloproteases was evidenced by the importantformation of an extensive hemorrhagic halo in vivo Thepresence of serine proteases was evidenced by coagulatingactivity present even in low concentrations of the venom(325 120583gmL)
Phospholipase activity of B mattogrossensis crude venomassayed with 4N3OBA synthetic substrate was 186405Umgmeasured by the number of moles of chromophores released
BioMed Research International 5
per minute (n∘molsmin or U) for a given quantity of proteinin milligrams (mg)
The properties of snake venom components observed inthis study are characteristic of accidents caused by snakesof the Bothrops sp genus symptoms such as pain edemahemorrhage and necrosis and additionally systemic distur-bances are characteristic and corroborate the literature thatdescribes that proteases of snake venom proteins are closelyrelated to interference in the hemostatic system promotingblood coagulation fibrinolysis and platelet aggregation [27ndash30]
32 Isolation and Biochemical Characterization of Phospholi-pases The present study showed for the first time the isola-tion of the three phospholipases A
2from B mattogrossensis
snake venom obtained by two chromatographic steps Firstionic exchange chromatography was performed on a CM-Sepharose column with an ammonium bicarbonate gradient(50 to 500mM pH 80) The elution of absorbed proteinswith a linear gradient of concentrated buffer resulted inthirteen fractions (Figure 1(a)) of which fraction nine (9) wasrelated to PLA
2s because it showed phospholipase activity
of 22109Umg on artificial substratum All fractions werelyophilized and submitted to unidimensional electrophoresisrevealing many protein bands
Fraction 9 was submitted to the second chromatographicstep in a reverse column phase on a Discovery C18 columnusing 01 Trifluoroacetic (TFA) and 999 Acetonitrile(ACN) as solvents for the separation of other venom com-ponents The elution of absorbed proteins with a lineargradient of concentrated buffer resulted in twenty-two (22)fractions (Figure 1(b)) The PLA
2s were highly purified with
approximately 40 ACN Similar results were observed inthe isolation of other PLA
2s from snake venoms in high per-
formance liquid chromatography on reverse phase columnswhere the elution profile of these proteins occurs between 30and 40 ACN [5]
The association of chromatographic techniques suchas ionic exchange and reverse phase has commonly beenused and many snake venoms have been fractioned thisway highlighting the phospholipase purification of a speciesbelonging to the old complex ldquoBothrops neuwiedirdquo as well asthe target species studied in this research Two PLA
2basic
isoforms from B (neuwiedi) pauloensis venom were purifiedby Rodrigues et al [31] using biochemical techniques similarto the ones used in this study with the combination of ionicexchange (cationic) and reverse phase chromatographies
Other PLA2s have also been purified using simplified
procedures based in CM-Sepharose andor reverse phase asfor example venomPLA
2s fromBothropsmoojeni [21 32 33]
B pirajai [34 35] B jararacussu [36 37] B alternatus [8 38]Cerastes cerastes [39] and Elaphe climacophora [40]
Fractions 13 and 14 were related to enzymatically inactivePLA2s whereas fraction 15was related to enzymatically active
PLA2s The phospholipase activity of the collected fractions
were analyzed with synthetic NOB and NBD-PC substrates(Figures 2(a) and 2(b))The amount of activity was comparedto BthTX-II a basic enzymatically active PLA
2(Asp49) and
BthTX-I a basic enzymatically inactive PLA2(Lys49) both
20
15
10
5
0
100 200 300 400
80
60
40
20
0
B (
)
(mAU
)
1
23
4
5 67 8
9
10 11 12 13
MW 02 03 04 06 08 09 10 12 13
175805846
30
2317
07
times102
(min)
Btx
(a)
100
80
60
40
20
B (
)
MW175
805846
30
2317
07
(mAU
)
times102
6
5
4
3
2
1
0
0 10 20 30 40
1
2
3 45
6
7
8
910 11
12
13
1415
16
1718
19
20 2122
(min)
07 12 1314 15 16 17 18Btx
(b)
Figure 1 Chromatographic and electrophoretic profile of Bothropsmattogrossensis venom fractioning (a) CM-Sepharose columnsequilibrated with solvent A (50mM ammonium bicarbonatepH 80) and eluted with a 0ndash100 concentration gradient of solventB (500mM ammonium bicarbonate pH 80) at a 50mLminuteflow Emphasis on peak 9 rechromatographed (b) Rechromatog-raphy of fraction 9 on Discovery C18 column equilibrated withsolvent A (01 TFA) and eluted with a concentration gradient of0ndash100 of solvent B (999 acetonitrile and 01 trifluoroaceticacid) and a 10mLmin flow Emphasized in blue are fractions 1314 and 15 characterized as phospholipases BmatTX-I BmatTX-II and BmatTX-III respectively Controls MW molecular weightstandard BTx BthTX-I a basic enzymatically inactive PLA
2(Lys49)
(10120583g) isolated from Bothrops jararacussu venom Absorbancesread at 280 nm Electrophoresis gel made with 125 (ww)acrylamidebis-acrylamide
previously isolated from Bothrops jararacussu venom [36 41]The BmatTX-III PLA
2s (Asp49) were not able to induce
platelet aggregation and did not inhibit collagen or ADPinduced platelet aggregation (data not shown)
The degree of purity of the isolated proteins was furtherdemonstrated by SDS-PAGE mass spectrometry (Figure 3)and N-terminal sequencing (Figures 4(a) and 4(b)) Thepurified PLA
2s were named BmatTX-I BmatTX-II and
6 BioMed Research International
F12
Bmat
TX-I
Bmat
TX-I
I
Bmat
TX-I
II
F16
F17
F18
F19
F20
F21
F22
BthT
X-I
BthT
X-II
0
200
400
600
800
1000Ph
osph
olip
ase a
ctiv
ity (U
mg)
(a)
F12
Bmat
TX-I
II
F16
F17
F20
0
200
400
600
800
1000
Fluo
resc
ent i
nten
sity
(b)
Control BmatTX-I BmatTX-II BmatTX-III0
1000
2000
3000
BmatTX-I (Lys49)BmatTX-II (Lys49)BmatTX-III (Asp49)
Control
CK (U
L)
25120583g50120583L
lowastlowast
lowast
(c)
Figure 2 Enzymatic activity and myotoxic activity of PLA2s isolated from the venom of B mattogrossensis (a) Phospholipase activity of the
fractions collected from the rechromatography of fraction 9 done in C18 column assayed using an NOB stained substrate This activity wasassessed through the measurement of the number of moles of the released chromophore per minute (n∘molsmin or U) per milligram ofprotein (b) Phospholipase activity of the fractions collected from the rechromatography of fraction 9 done in C18 column assayed using afluorescent substrate (c) Myotoxic activity evaluated for inoculation of PLA
2s (25 120583g50 120583L) or PBS (control) done intramuscularly in the
gastrocnemius muscle of mice After 3 hours the creatine kinase (CK) level an important marker of muscular lesion was assayed in theanimalrsquos plasma Each bar represents the average +minus SD of three independent groups lowast119875 lt 005 compared to the control F13 BmatTX-I(Lys49) F14 BmatTX-II (Lys49) F15 BmatTX-III (Asp49) Controls BthTX-I (Lys49) and BthTX-II (Asp49)
BmatTX-III They were characterized as single polypeptidechains with isoelectric points around 10 (data not shown)This result agrees with data from published literature wheremost basic PLA
2s with or without catalytic activity on
artificial substrates [32 42] show an isoelectric point between8 and 10
The average molecular mass defined by mass spectrom-etry was 13304 Da for BmatTX-I 13623Da for BmatTX-IIand 13681Da for BmatTX-III (Figure 3)Thismolecularmassis consistent with most isolated PLA
2s from snake venoms
which are around 13 to 16 kDa [27 36 43]BmatTX-I sequencing showed a lysine (Lys) at position
49 and although only the first 28 amino acid residuesof BmatTX-II have been sequenced both are highly sim-ilar to the PLA
2Lys49 homologue subgroup The N-
terminal sequence alignment of BmatTX-I and BmatTX-IIhas revealed that these basic proteins are PLA
2s similar to
homologous Lys49 and other Lys49 myotoxins from snake
venoms (Figure 4(a)) BmatTX-I showed 94 similarity withMjTX-I present in B moojeni venom 92 with BthTX-I andBOJU-I present in B jararacussu venom and 94 similaritywith MTX-II present in B brazili venom
Additionallymultiple sequence alignment of BmatTX-IIIshowed a PLA
2-Asp49 basic myotoxin with another Asp49
of the Bothrops genera (Figure 4(b)) It can be observedthat BmatTX-III presented 78 similarity with BmjeTX-Iand 75 with BmoTX-I both from B moojeni venom 71similarity with BthTX-II isolated from B jararacussu venomand 65 similarity with PrTX-III from B pirajai venom
In the analysis of the sequences of the PLA2s isolated
from B mattogrossensis in this study highly preserved con-stituent residues of the 120572-helix structure characteristic ofphospholipases were identified as well as the presence ofmany cysteine residues which suggests the existence of manydisulfide bridges important for the stabilization of PLA
2rsquos
molecular structure [21 44]
BioMed Research International 7
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1360475
1398779
(a)
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1362326
1380673
(b)
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1256219
1368171
1402625
(c)
Figure 3 Mass determination by spectrometryThe average proteinmasses of BmatTX-I (a) BmatTX-II (b) and BmatTX-III (c) wereobtained in a MALDI-TOF system operated in linear mode usingexternal standards for calibration The resulting mass spectra weresubmitted to automatic baseline subtraction
Some studies regarding the amino acid composition ofPLA2s demonstrate that these proteins are rich in basic
and hydrophobic amino acids containing three long 120572-helixes two beta sheets and a Ca2+ binding site [45ndash47]Calcium is absolutely necessary for hydrolysis thereforealmost all PLA
2s have a highly preserved region for a Ca2+
bond (XCGXGG) and a catalytic site (DXCCXXHD) [48]It is observed in BmatTX-III that the calcium binding site(27YCGWGG
32) is preserved indicating that it is a catalyt-
ically active PLA2belonging to the PLA
2Asp49 subgroup
33 Biological Activities of Phospholipases A2 The main
objective for the isolation and characterization of compo-nents of snake venom is to better understand the partic-ipation of each component in different pathophysiologicalprocesses resulting from envenomation Local lesions can beattributed to proteases phospholipase activity and hemor-rhagic factors of these venoms followed by the release ofvasoactive agents causing hemorrhaging in various organsand tissues [5 44 49ndash52]
The PLA2s isolated from B mattogrossensis venom
BmatTX-I BmatTX-II and BmatTX-III showed highmyotoxic activity (Figure 2(c)) At a concentration of25 120583g50 120583L the PLA
2s induced a significant release of CK
an important muscular lesion marker when compared to thecontrol Myotoxicity is the characteristic presented by mostbasic phospholipases A
2from snake venoms Several studies
demonstrate that the myotoxic effect begins quickly by directaction of the myotoxic PLA
2s on the plasma membrane of
muscle cells or it is mediated by metalloproteases due toconsequent degeneration and ischemia [27 28 44]
Regarding the Lys49-PLA2myotoxins it is evident that
they lyse the plasma membrane of the muscle cell infected invivo however the exact mechanism has not been describedyet Furthermore it is not known if the toxin is internalizedbefore during or after the initial lysis or if it is not internal-ized Althoughmyotoxicity can be induced by the productionof fatty acids there is a second mechanism that seems to beindependent of the enzymatic activity and is mediated bythe C-terminal region at sites 115ndash129 of the Lys49 molecules[43 53 54]
In an attempt to better understand the development ofthe inflammatory process unleashed by the protein complexpresent in snake venoms many studies have been donesuch as edema induction [55] leukocyte participation [5657] mast cells degranulation [55] participation of variouscytotoxins in the inflammatory system [58 59] participationof cyclooxygenases [60 61] and the participation of venomPLA2s in the inflammatory process [6 51 56 62]
Many studies about PLA2activity in macrophages have
already been done [8 63 64] Little is known about PLA2rsquos
effect on neutrophils however Escocard et al [65] describedan influx of inflammatory cells including many neutrophilsinto the peritoneal cavity of mice after the injection ofBothrops atrox venom The induction of reactive oxygenspecies (ROS) cytokines like IL-6 and IL-1120573 was seen inthese neutrophils These data were also observed in a studycarried out by Souza et al [26] where besides the influx ofneutrophils into the peritoneal cavity ofmice after injection ofthe venom of B atrox there was also induction of superoxideby these cells mast cell degranulation and phagocytosisby macrophages Regarding the activity of PLA
2Gambero
et al [66] have observed the ability of some myotoxins(bothropstoxin-I-II and piratoxin-I) to induce neutrophilchemotaxis in a concentration-dependent manner
In order to evaluate the activation of leukocytes thetoxicity of B mattogrossensis myotoxins on neutrophil cellswas investigated The cells were incubated with differentconcentrations (3 6 and 12 120583gmL) of BmatTX-I (Lys49)BmatTX-II (Lys49) and BmatTX-III (Asp49) myotoxinsduring 1 12 and 24 hours (data not shown)These myotoxinsdid not affect the neutrophilsrsquo viability which agrees withZuliani et al [64] showing low toxicity on thioglycollateelicited mice macrophages
Nonetheless our data showed that neutrophils incubatedwith BmatTX-I and BmatTX-II myotoxins induced therelease of IL-1120573 (Figures 5(a) and 5(b)) Moreover theseresults suggest that phospholipid hydrolysis is not essentialfor the activity observed and argue with the hyphotesis that
8 BioMed Research International
lowastlowast lowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowast lowastlowastlowastlowastlowastlowastlowastlowastlowast
1 10 20 30 40 50(Identity )
100
89
92
92
92
94
94
BmatTX-IBmatTX-IIPrtx-IBOJU-IBthTX-IMjtx-IMtx-Ii
(a)
lowastlowastlowast lowastlowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowastlowast lowastlowastlowast
1 10 20 30 40 50
(Identity )100
78
75
71
65
BmatTX-IIIBmoTX-IBmjeTX-IBthTX-IIPrTX-III
(b)
Figure 4 Comparison of the N-terminal sequence of the snakersquos PLA2s with the PLA
2s isolated from B mattogrossensis venom N-terminal
sequencing of the PLA2s obtained by Edman [13] degradation and alignment done with CLUSTALW2 expressed as of similarity (a)
BmatTX-I BmatTX-II with Lys49 residues in marked sequences (b) BmatTX-III with Asp49 residues in marked sequences Sequencesused for the alignment and their respective access numbers BthTX-I (gi51890398) MjTX-I (gi17368325) Mtx-II (gi390981003) PrTX-I (gi190016174) BOJU-I (gi209572966) BthTX-II (gi1171971) PrTX-III (gi90016174) BmjeTX-I (gi313471399) BmoTX-I (gi221272396)BthTX-I (Lys49) BthTX-II (Asp49) and BOJU-I (Lys49) isolated from the venom of B jararacussu MjTX-I (Lys49) BmoTX-I (Asp49)and BmjeTX-I (Asp49) isolated from the venom of B moojeni Mtx-II (Lys49) isolated from the venom of B brazili PrTX-I (Lys49) andPrTX-III (Asp49) isolated from the venom of B pirajai the ldquolowastrdquo was used to indicate the amino acid residues that are the same between thesequences
other molecular regions distinct from the active site may beinvolved in this effect
PLA2s are multifunctional proteins that can be used as
mediators in several areas of medicine such as in the treat-ment of rheumatoid arthritis as a new class of HIV inhibitorsby blocking the host cell invasion as a potential treatmentagainst malaria and as an antibiotic by inducing cytotoxicityvia the disruption of bacterial membranes [21 23 30 6768] A study by Costa et al [67] with PLA
2s isolated from
B brazili MTX-I and II demonstrated cytotoxic activityagainst Jurkat tumor cells as well as antimicrobial activityagainstE coli andC albicans and antiparasitic activity againstLeishmania sp
In the present study we evaluated the cytotoxic activityof PLA
2s isolated from B mattogrossensis on JURKAT (T
leukemia) and SK-BR-3 (breast adenocarcinoma) both ofwhich are human tumor cell lines Like MTX-II of B brazili[67] the cytotoxic activity of PLA
2s BmatTX-I and BmatTX-
II on JURKATcells was independent of their catalytic activitysince these are characterized as Lys49 PLA
2s and are therefore
catalytically inactive BmatTX-III characterized as Asp49despite being enzymatically active also showed a lower level oftoxicity Some authors propose that the cytotoxic activity ontumor cell lines is associated with the induction of apoptosisconsidering the fact that PLA
2promotes alterations in the
cell membrane And some studies involving Lys49 PLA2s
isolated from B asper demonstrated that the C-terminal
region comprised of amino acids 115ndash129 is concerned withthe cytotoxic and bactericidal activities of this protein [69ndash71] The same observation was made by Costa et al [67] withsynthetic peptides derived from the C-terminal portion ofMTX-I and II PLA
2s
We also evaluated the antiparasitic activity of the crudevenom and isolated PLA
2of B mattogrossensis on promastig-
ote forms of L amazonensis After the analysis it wasobserved that the crude venom of B mattogrossensis presentsincreasing toxic activity (approximately 50 to 80) againstpromastigote forms of L amazonensis after 48 h of incubation(Figure 7(a)) The PLA
2s isolated from B mattogrossensis at
100 120583gmL characterized as BmatTX-I (Lys49) and BmatTX-III (Asp49) presented toxic activity between 25 and 30respectively even with values close to those presented afterincubation of the protozoan with Pentamidine a drug usedas a positive control (Figures 7(b) and 7(c))
When compared the cytotoxicity values of PLA2s against
promastigote forms of L amazonensis show similar activitybetween the Lys49-PLA
2and the Asp49-PLA
2 Comparison
of this activity with the crude venom (70) showed thatPLA2s are responsible for almost half of the observed effect
Nonetheless notably the results suggest that other toxinspresent in the venom contribute to the parasitersquos death
Similar to the results obtained in the present studyStabeli et al [21] demonstrated that MjTX-II a Lys49-PLA
2
homologue isolated from B moojeni venom in different
BioMed Research International 9
0
500
1000
1500
2000
lowast
lowast
lowast
lowast
lowast
Control 3 6 12 3 6 12 3 6 12
ControlBmatTX-I (Lys49)
BmatTX-II (Lys49)BmatTX-III (Asp49)
BmatTX-I (120583g) BmatTX-II (120583g) BmatTX-III (120583g)
IL-1120573
(pg
mL)
(a)
Control 3 6 12 3 6 12 3 6 120
500
1000
1500
2000
ControlBmatTX-I (Lys49)
BmatTX-II (Lys49)BmatTX-III (Asp49)
lowastlowastlowast
BmatTX-I (120583g) BmatTX-II (120583g) BmatTX-III (120583g)
IL-1120573
(pg
mL)
(b)
Figure 5 Production of IL-1120573 by mice neutrophils These cells wereincubatedwith PLA
2(3 6 and 12 120583gmL) or RPMI (control) for 12
hours (a) and 24 hours (b) at 37∘C in a humid atmosphere with 5CO2 Quantification was done by ELISA as described in 29 Each
bar represents the average +minus SDof three independent groups lowast119875 lt005 compared to the control
concentrations (5 25 and 75 120583g) was effective as a parasiticideagent against Schistosomamansoni and promastigote forms ofLeishmania (L amazonensis L braziliensis L donovani andL major)
The action of the PLA2s BmatTX-I (Lys49) and
BmatTX-III (Asp49) on promastigote forms of L ama-zonensis was independent of its catalytic activity sincecatalytically inactive Lys49 myotoxin also demonstrated tox-icity against Leishmania It is believed that the observedcytotoxic activity might be related to the C-terminal regionsof these phospholipase-homologues that are able to promotea disturbance in the cellular membranes independent oftheir catalytic activity [67 72] However more studies arenecessary to define the exact mechanism of action of theseenzymes on parasites
Growing interest in the comprehension of the structureand function of snake venom components especially PLA
2
10 100 10 100 10 100 10 1000
20
40
60
80
100
BmatTX-I (Lys49)BmatTX-II (Lys49)
BmatTX-III (Asp49)Methotrexate (MTX)
Cyto
toxi
city
()
Concentration (120583gmL)
(a)
10 100 10 100 10 100 10 1000
20
40
60
80
100
BmatTX-I (Lys49)BmatTX-II (Lys49)
BmatTX-III (Asp49)Methotrexate (MTX)
Cyto
toxi
city
()
Concentration (120583gmL)
(b)
Figure 6 Antitumoral activity of PLA2s from B mattogrossensis
(a) Antitumoral activity on human acute T-cell leukemia (JURKAT)lines (b) Antitumoral activity on human breast adenocarcinoma(SK-BR-3) Different concentrations of the PLA
2s were incubated
with cell linesMethotrexate was used as the positive control Resultsare presented as mean +minus SD (119899 = 3)
contributes to a better understanding of the mechanism ofaction of their enzymatic and toxic activities It opens thepath to better understand the intoxication caused by enven-omation and the physiopathology behind its side effectsFuture studies will potentially improve serum therapy andhelp develop the pharmaceutical potential that moleculesisolated from animal venoms can have such as the PLA
2s
isolated from the B mattogrossensis venom which show anti-Leishmania and antitumor activities
4 Conclusion
In conclusion the venom of Bothrops mattogrossensis hasa qualitatively similar toxicological profile to previously
10 BioMed Research International
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineB mattogrossensis crude venom
Ant
i-leis
hman
ia ac
tivity
()
B mattogrossensis crude venom (120583g)
(a)
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineBmatTX-I (Lys49)
Ant
i-leis
hman
ia ac
tivity
()
BmatTX-I (120583g)
lowastlowast lowast lowast
(b)
Ant
i-leis
hman
ia ac
tivity
()
Pent 36 625 125 25 50 1000
20
40
60
80
100
BmatTX-III (Asp49)Pentamidine
BmatTX-III (120583g)
lowastlowastlowast
lowastlowast lowast
(c)
Figure 7 Antileishmanial activity of the crude venom of and PLA2s from B mattogrossensis The promastigote forms of L amazonensiswere
plated with 1times105 cellswellThen different concentrations of crude venom and isolated PLA2s were added to each wellThe experiment was
done in a 48 h period with the (a) crude venom and the PLA2enzymes (b) BmatTX-I and (c) BmatTX-III MTT was added and after the
incubation period at 33∘C the formazan crystal formed was dissolved in SDS Readings were done in a spectrophotometer at 570 nm Eachbar represents the average +minus SD of the three independent experimental groups sixfold total lowast119875 lt 005 compared to the control
studied snake venoms of the Bothrops sp genera despite theobservation of quantitative variations Of the three basicPLA2s from B mattogrossensis venom now isolated for the
first time two are characterized as Lys49-PLA2homologues
BmatTX-I and -II and the other as an Asp49-PLA2 named
BmatTX-III This showed high phospholipase activity ThePLA2s isolated induced myotoxic effects as well as the release
of proinflammatory cytokines by neutrophils BmatTX-I and-III PLA
2s were cytotoxic to human tumor cell lines JURKAT
and SK-BR-3 and showed activity against promastigote formsof L amazonensis
Conflict of Interests
The authors state that there is no conflict of interests
Acknowledgments
The authors are grateful to the Ministry of Science andTechnology (MCTI) Conselho Nacional de Desenvolvi-mento Cientıfico e Tecnologico (CNPq) Financiadora deEstudos e Projetos (FINEP) Fundacao de Tecnologia doAcre (FUNTACFDCT) Coordenacao de Aperfeicoamentode Nıvel Superior (CAPES)mdashProjeto NanoBiotec Redede Biodiversidade e Biotecnologia da Amazonia Legal(BIONORTECNPqMCTI) Instituto Nacional paraPesquisa Translacional em Saude e Ambiente na RegiaoAmazonica (INCT-INPeTAmCNPqMCTI) and InstitutoNacional para Pesquisa em Toxinas (INCT-Tox) andSecretary of Development of Rondonia State (SEPLANPRONEXCNPq) for financial support We are grateful to
BioMed Research International 11
Sr Guilherme G Galassi (Mantenedouro da Fauna Silvestreautorizacao IBAMA 020270089912001-95 Americana-SP)for their helpful technical collaboration in provide the snakevenom Amy Grabner provided the English editing of thepaper
References
[1] Y Angulo and B Lomonte ldquoBiochemistry and toxicology oftoxins purified from the venom of the snake Bothrops asperrdquoToxicon vol 54 no 7 pp 949ndash957 2009
[2] J J Calvete P Juarez and L Sanz ldquoSnake venomics Strategyand applicationsrdquo Journal of Mass Spectrometry vol 42 no 11pp 1405ndash1414 2007
[3] T S Kang D Georgieva N Genov et al ldquoEnzymatic toxinsfrom snake venom structural characterization and mechanismof catalysisrdquoThe FEBS Journal vol 278 no 23 pp 4544ndash45762011
[4] C ZOliveiraCaracterizacao funcional e estrutural de inibidoresde fosfolipases A
2isolados do plasma de serpente Bothrops
jararacussu [PhD thesis] Faculdade de Ciencias Farmaceuticasde Ribeirao Preto Ribeirao Preto Brazil 2009
[5] R G Stabeli R Simoes-Silva A M Kayano et al ldquoPurifi-cation of phospholipases A
2from American snake venomsrdquo
in ChromatographymdashThe Most Versatile Method of ChemicalAnalysis chapter 1 pp 1ndash34 InTech Vienna Austria 2012
[6] J P Zuliani C M Fernandes S R Zamuner J M Gutierrezand C F P Teixeira ldquoInflammatory events induced by Lys-49 and Asp-49 phospholipases A
2isolated from Bothrops asper
snake venom role of catalytic activityrdquo Toxicon vol 45 no 3pp 335ndash346 2005
[7] S Marcussi R G Stabeli N A Santos-Filho et al ldquoGenotoxiceffect of Bothrops snake venoms and isolated toxins on humanlymphocyte DNArdquo Toxicon vol 65 pp 9ndash14 2013
[8] S S Setubal A S Pontes J L Furtado et al ldquoAction oftwo phospholipases A
2purified from Bothrops alternatus snake
venom on macrophagesrdquo Biochemistry vol 78 no 2 pp 194ndash203 2013
[9] J M Gutierrez L A Ponce-Soto S Marangoni and BLomonte ldquoSystemic and local myotoxicity induced by snakevenom group II phospholipases A
2 comparison between cro-
toxin crotoxin B and a Lys49 PLA2homologuerdquo Toxicon vol
51 no 1 pp 80ndash92 2008[10] A Q Rueda I G Rodrıguez E C Ara et al ldquoBiochemical
characterization action onmacrophages and superoxide anionproduction of four basic phospholipases A
2from Panamanian
Bothrops asper snake venomrdquo BioMed Research Internationalvol 2013 Article ID 789689 9 pages 2013
[11] V X da Silva and M T Rodrigues ldquoTaxonomic revision ofthe Bothrops neuwiedi complex (Serpentes Viperidae) withdescription of a new speciesrdquo Phyllomedusa vol 7 no 1 pp 45ndash90 2008
[12] J A Campbell and W W Lamar The Venomous Reptiles of theWestern Hemisphere Comstock Ithaca NY USA 2004
[13] P Edman ldquoMethod for determination of the amino acidsequence in peptidesrdquo Acta Chemica Scandinavica vol 4 pp283ndash293 1950
[14] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[15] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[16] M Holzer and S P Mackessy ldquoAn aqueous endpoint assay ofsnake venom phospholipase A
2rdquo Toxicon vol 34 no 10 pp
1149ndash1155 1996[17] A L Fuly A L P deMiranda R B Zingali and J AGuimaraes
ldquoPurification and characterization of a phospholipase A2isoen-
zyme isolated from Lachesis muta snake venomrdquo BiochemicalPharmacology vol 63 no 9 pp 1589ndash1597 2002
[18] T Nikai N Mori M Kishida H Sugihara and A T Tu ldquoIso-lation and biochemical characterization of hemorrhagic toxinf from the venom of Crotalus atrox (Western DiamondbackRattlesnake)rdquo Archives of Biochemistry and Biophysics vol 231no 2 pp 309ndash319 1984
[19] J A Gene A Roy G Rojas J M Gutierrez and L CerdasldquoComparative study on coagulant defibrinating fibrinolyticand fibrinogenolytic activities of Costa Rican crotaline snakevenoms and their neutralization by a polyvalent antivenomrdquoToxicon vol 27 no 8 pp 841ndash848 1989
[20] M S Charney and R M Tomarelli ldquoA colorimetric method forthe determination of the proteolytic activity of duodenal juicerdquoThe Journal of Biological Chemistry vol 171 pp 501ndash505 1947
[21] R G Stabeli S F Amui C D SantrsquoAna et al ldquoBothrops moojenimyotoxin-II a Lys49-phospholipaseA
2homologue an example
of function versatility of snake venom proteinsrdquo ComparativeBiochemistry and Physiology C vol 142 no 3-4 pp 371ndash3812006
[22] D R Call J A Nemzek S J Ebong G L Bolgos D ENewcomb and D G Remick ldquoRatio of local to systemicchemokine concentrations regulates neutrophil recruitmentrdquoThe American Journal of Pathology vol 158 no 2 pp 715ndash7212001
[23] P G Roberto S Kashima S Marcussi et al ldquoCloning andidentification of a complete cDNA coding for a bactericidal andantitumoral acidic phospholipase A
2from Bothrops jararacussu
venomrdquoThe Protein Journal vol 23 no 4 pp 273ndash285 2004[24] J H Schumacher A OrsquoGarra B Shrader et al ldquoThe charac-
terization of four monoclonal antibodies specific for mouse IL-5 and development of mouse and human IL-5 enzyme-linkedimmunosorbentrdquoThe Journal of Immunology vol 141 no 5 pp1576ndash1581 1988
[25] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[26] C A T Souza A M Kayano S S Setubal et al ldquoLocal andsystemic biochemical alterations induced by Bothrops atroxsnake venom in micerdquo Journal of Venom Research vol 3 pp28ndash34 2012
[27] J M Gutierrez and B Lomonte ldquoPhospholipase A2myotoxins
from Bothrops snake venomsrdquo Toxicon vol 33 no 11 pp 1405ndash1424 1995
[28] J M Gutierrez and A Rucavado ldquoSnake venom metallopro-teinases their role in the pathogenesis of local tissue damagerdquoBiochimie vol 82 no 9-10 pp 841ndash850 2000
[29] T Matsui Y Fujimura and K Titani ldquoSnake venom proteasesaffecting hemostasis and thrombosisrdquo Biochimica et BiophysicaActa vol 1477 no 1-2 pp 146ndash156 2000
[30] A M Soares M R M Fontes and J R Giglio ldquoPhospholipaseA2myotoxins from Bothrops snake venoms structure-function
12 BioMed Research International
relationshiprdquoCurrent Organic Chemistry vol 8 no 17 pp 1677ndash1690 2004
[31] VM Rodrigues SMarcussi R S Cambraia et al ldquoBactericidaland neurotoxic activities of two myotoxic phospholipases A
2
from Bothrops neuwiedi pauloensis snake venomrdquo Toxicon vol44 no 3 pp 305ndash314 2004
[32] N A Santos-Filho L B Silveira C Z Oliveira et al ldquoAnew acidic myotoxic anti-platelet and prostaglandin I
2induc-
tor phospholipase A2isolated from Bothrops moojeni snake
venomrdquo Toxicon vol 52 no 8 pp 908ndash917 2008[33] A M Soares S H Andriao-Escarso Y Angulo et al ldquoStruc-
tural and functional characterization of myotoxin I a Lys49phospholipaseA
2homologue fromBothropsmoojeni (Caissaca)
snake venomrdquo Archives of Biochemistry and Biophysics vol 373no 1 pp 7ndash15 2000
[34] A M Soares S H Andriao-Escarso R K Bortoleto et alldquoDissociation of enzymatic and pharmacological properties ofpiratoxins-I and -III two myotoxic phospholipases A
2from
Bothrops pirajai snake venomrdquo Archives of Biochemistry andBiophysics vol 387 no 2 pp 188ndash196 2001
[35] S S Teixeira L B Silveira F M N da Silva et al ldquoMolecularcharacterization of an acidic phospholipase A
2from Bothrops
pirajai snake venom synthetic C-terminal peptide identifies itsantiplatelet regionrdquo Archives of Toxicology vol 85 no 10 pp1219ndash1233 2011
[36] S H Andriao-Escarso A M Soares V M Rodrigues et alldquoMyotoxic phospholipases A
2in Bothrops snake venoms effect
of chemical modifications on the enzymatic and pharmacolog-ical properties of bothropstoxins from Bothrops jararacussurdquoBiochimie vol 82 no 8 pp 755ndash763 2000
[37] S H Andriao-Escarso A M Soares M R M Fontes et alldquoStructural and functional characterization of an acidic plateletaggregation inhibitor and hypotensive phospholipase A
2from
Bothrops jararacussu snake venomrdquo Biochemical Pharmacologyvol 64 no 4 pp 723ndash732 2002
[38] M E G Denegri O C Acosta S Huancahuire-Vega et alldquoIsolation and functional characterization of a new acidic PLA
2
Ba SpII RP4 of the Bothrops alternatus snake venom fromArgentinardquo Toxicon vol 56 no 1 pp 64ndash74 2010
[39] R Zouari-Kessentini J Luis A Karray et al ldquoTwo purified andcharacterized phospholipases A
2from Cerastes cerastes venom
that inhibit cancerous cell adhesion and migrationrdquo Toxiconvol 53 no 4 pp 444ndash453 2009
[40] R Shirai M Toriba K Hayashi K Ikeda and S Inoue ldquoIden-tification and characterization of phospholipase A
2inhibitors
from the serum of the Japanese rat snake Elaphe climacophorardquoToxicon vol 53 no 6 pp 685ndash692 2009
[41] M I Homsi-Brandenburgo L S Queiroz H Santo-Neto LRodrigues-Simoni and J R Giglio ldquoFractionation of Bothropsjararacussu snake venom partial chemical characterization andbiological activity of bothropstoxinrdquo Toxicon vol 26 no 7 pp615ndash627 1988
[42] G P Fernandez A Segura M Herrera et al ldquoNeutralizationof Bothrops mattogrossensis snake venom from Bolivia exper-imental evaluation of llama and donkey antivenoms producedby caprylic acid precipitationrdquoToxicon vol 55 no 2-3 pp 642ndash645 2010
[43] C L Ownby H S S de Araujo S P White and J E FletcherldquoLysine 49 phospholipase A
2proteinsrdquo Toxicon vol 37 no 3
pp 411ndash445 1999[44] A M Soares W P Sestito S Marcussi et al ldquoAlkylation of
myotoxic phospholipases A2in Bothrops moojeni venom a
promising approach to an enhanced antivenom productionrdquoInternational Journal of Biochemistry amp Cell Biology vol 36 no2 pp 258ndash270 2004
[45] B Lomonte J M Gutierrez M F Furtado et al ldquoIsolationof basic myotoxins from Bothrops moojeni and Bothrops atroxsnake venomsrdquo Toxicon vol 28 no 10 pp 1137ndash1146 1990
[46] L C Mancuso M M Correa C A Vieira et al ldquoFractionationof Bothrops pirajai snake venom isolation and characterizationof piratoxin-I a new myotoxic proteinrdquo Toxicon vol 33 no 5pp 615ndash626 1995
[47] H S Selistre L S Queiroz O A B Cunha G E P deSouza and J R Giglio ldquoIsolation and characterization ofhemorrhagic myonecrotic and edema-inducing toxins fromBothrops insularis (jararaca ilhoa) snake venomrdquo Toxicon vol28 no 3 pp 261ndash273 1990
[48] E A Dennis J Cao Y-H Hsu V Magrioti and G KokotosldquoPhospholipaseA
2enzymes physical structure biological func-
tion disease implication chemical inhibition and therapeuticinterventionrdquo Chemical Reviews vol 111 no 10 pp 6130ndash61852011
[49] A G T Cardoso Acao do veneno de Bothrops moojeni e suafracao L-aminoacido oxidase submetida ao tratamento comraios gama de 60CO em Leishmania spp [MS thesis] IPEN-CNENSP Sao Paulo Brazil 1999
[50] MGallacci andWLGCavalcante ldquoUnderstanding the in vitroneuromuscular activity of snake venomLys49 phospholipaseA
2
homologuesrdquo Toxicon vol 55 no 1 pp 1ndash11 2010[51] S S Teixeira Caracterizacao funcional e estrutural de uma
fosfolipase A2acida isolada do veneno de Bothrops pirajai
[MS thesis] Faculdade de Ciencias Farmaceuticas de RibeiraoPretoUSP Ribeirao Preto Brazil 2009
[52] E Valentin and G Lambeau ldquoIncreasing molecular diversityof secreted phospholipases A
2and their receptors and binding
proteinsrdquo Biochimica et Biophysica Acta vol 1488 no 1-2 pp59ndash70 2000
[53] J M Gutierrez and B Lomonte ldquoPhospholipases A2from
Viperidae snake venoms how do they induce skeletal muscledamagerdquo Acta Chimica Slovenica vol 58 no 4 pp 647ndash6592011
[54] J M Gutierrez C L Ownby and G V Odell ldquoIsolation of amyotoxin from Bothrops asper venom partial characterizationand action on skeletal musclerdquo Toxicon vol 22 no 1 pp 115ndash128 1984
[55] N G Nascimento M C Sampaio R A Olivo and C TeixeiraldquoContribution of mast cells to the oedema induced by Bothropsmoojeni snake venom and a pharmacological assessment of theinflammatory mediators involvedrdquo Toxicon vol 55 no 2-3 pp343ndash352 2010
[56] C F P Teixeira E C T Landucci E Antunes M Chacurand Y Cury ldquoInflammatory effects of snake venom myotoxicphospholipases A
2rdquo Toxicon vol 42 no 8 pp 947ndash962 2003
[57] S R Zamuner and C F P Teixeira ldquoCell adhesion moleculesinvolved in the leukocyte recruitment induced by venom of thesnake Bothrops jararacardquoMediators of Inflammation vol 11 no6 pp 351ndash357 2002
[58] F Chaves C F P Teixeira and J M Gutierrez ldquoRole ofTNF-120572 IL-1120573 and IL-6 in the local tissue damage induced byBothrops asper snake venom an experimental assessment inmicerdquo Toxicon vol 45 no 2 pp 171ndash178 2005
BioMed Research International 13
[59] C D F P Teixeira C M Fernandes J P Zuliani and SF Zamuner ldquoInflammatory effects of snake venom metallo-proteinasesrdquo Memorias do Instituto Oswaldo Cruz vol 100supplement 1 pp 181ndash184 2005
[60] R D A Olivo C F P Teixeira J LWallace J M Gutierrez andS R Zamuner ldquoRole of cyclooxygenases in oedema-formingactivity of bothropic venomsrdquo Toxicon vol 49 no 5 pp 670ndash677 2007
[61] L B Silveira D P Marchi-Salvador N A Santos-Filho et alldquoIsolation and expression of a hypotensive and anti-plateletacidic phospholipase A
2from Bothrops moojeni snake venomrdquo
Journal of Pharmaceutical and Biomedical Analysis vol 73 pp35ndash43 2013
[62] A Rucavado T Escalante C F P Teixeira C M Fernandes CDıaz and J M Gutierrez ldquoIncrements in cytokines and matrixmetalloproteinases in skeletal muscle after injection of tissue-damaging toxins from the venom of the snake Bothrops asperrdquoMediators of Inflammation vol 11 no 2 pp 121ndash128 2002
[63] E Leiguez J P Zuliani A M Cianciarullo C M Fernandes JM Gutierrez and C Teixeira ldquoA group IIA-secreted phospho-lipase A
2from snake venom induces lipid body formation in
macrophages the roles of intracellular phospholipases A2and
distinct signaling pathwaysrdquo Journal of Leukocyte Biology vol90 no 1 pp 155ndash166 2011
[64] J P Zuliani J M Gutierrez L L Casais e Silva S C SampaioB Lomonte and C D F P Teixeira ldquoActivation of cellularfunctions in macrophages by venom secretory Asp-49 and Lys-49 phospholipasesA
2rdquoToxicon vol 46 no 5 pp 523ndash532 2005
[65] R D C M Escocard M M Kanashiro J H Petretski et alldquoNeutrophils regulate the expression of cytokines chemokinesand nitric oxide synthasenitric oxide in mice injected withBothrops atrox venomrdquo Immunobiology vol 211 no 1-2 pp 37ndash46 2006
[66] A Gambero E C T Landucci M H Toyama et al ldquoHumanneutrophil migration in vitro induced by secretory phospholi-pases A
2 a role for cell surface glycosaminoglycansrdquo Biochemi-
cal Pharmacology vol 63 no 1 pp 65ndash72 2002[67] F L S Costa R S Rodrigues L F M Izidoro et al ldquoBio-
chemical and functional properties of a thrombin-like enzymeisolated from Bothrops pauloensis snake venomrdquo Toxicon vol54 no 6 pp 725ndash735 2009
[68] J C Villalobos R Mora B Lomonte J M Gutierrez andY Angulo ldquoCytotoxicity induced in myotubes by a Lys49phospholipase A
2homologue from the venom of the snake
Bothrops asper evidence of rapid plasma membrane damageand a dual role for extracellular calciumrdquo Toxicology in Vitrovol 21 no 8 pp 1382ndash1389 2007
[69] B S Cummings J McHowat and R G Schnellmann ldquoPhos-pholipase A
2s in cell injury and deathrdquoThe Journal of Pharma-
cology and Experimental Therapeutics vol 294 no 3 pp 793ndash799 2000
[70] S R Panini L Yang A E Rusinol M S Sinensky J VBonventre and C C Leslie ldquoArachidonate metabolism andthe signaling pathway of induction of apoptosis by oxidizedLDLoxysterolrdquo Journal of Lipid Research vol 42 no 10 pp1678ndash1686 2001
[71] L Paramo B Lomonte J Pizarro-Cerda J-A Bengoechea J-P Gorvel and E Moreno ldquoBactericidal activity of Lys49 andAsp49 myotoxic phospholipases A
2from Bothrops asper snake
venom synthetic Lys49myotoxin II-(115-129)-peptide identifiesits bactericidal regionrdquo European Journal of Biochemistry vol253 no 2 pp 452ndash461 1998
[72] B Lomonte Y Angulo and E Moreno ldquoSynthetic peptidesderived from the C-terminal region of Lys49 phospholipaseA2homologues from viperidae snake venoms biomimetic
activities and potential applicationsrdquo Current PharmaceuticalDesign vol 16 no 28 pp 3224ndash3230 2010
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

BioMed Research International 3
was submitted for N-terminal amino acid sequencing usingthe automated Edman degradation method [13] on a PPSQ-33A (Shimadzu) automatic sequencer A sequence similaritysearch and multiple sequence alignment were performed inthe SWISS-PROTTREMBL database using the programsFASTA BLAST and CLUSTALW2
28 Functional Characterization281 Phospholipase Activity with 4-nitro-3(octanoyloxy)ben-zoic (4N3OBA) Phospholipase activity of the three PLA
2s
was assayed following the protocols described by Holzer andMackessy [16] modified for 96-well plates The standardassay mixture contained 200120583L of buffer (10mM Tris-HClplus 10mM CaCl
2 and 100mM NaCl pH 80) 20 120583L of
4-nitro-3(octanoyloxy)benzoic (4N3OBA ndash Biomol EUA)20120583L of deionized water and 20120583L of PLA
2(5 120583g) After
the addition of the PLA2s the mixture was incubated for
30 minutes at 37∘C and the absorbance was determined at425 nmusing an Eon (Biotek)microplate spectrophotometerfor 3-minute intervals The enzymatic activity was expressedas the reactionrsquos initial velocity (119881
119900) calculated based on the
increase in absorbance
282 Phospholipase Activity with Fluorescent SubstratePhospholipase A
2rsquos (PLA
2) enzymatic activity was evaluated
through the hydrolysis of synthetic fluorescent phospholipidusing the fluorescent substrate Acyl 6 0 NBD phospholipidNBD-phosphatidylcholine (NBD-PC) (Avanti Polar LipidsInc Alabaster AL USA) The assay was performed in aspectrofluorometer (Shimadzu RF-5301PC software RFPC)with excitation and emission wavelengths of 460 and 534respectivelyThe enzymatic activity of each Bmattogrossensischromatographic fraction (7 12 15 16 17 and 20) wasevaluated over 250 seconds after the addition of substrate(33 120583gmL final concentration) in a reaction media con-taining 50mM Tris-HCl and 8mM CaCl
2at pH 75 at room
temperature
283 Platelet Aggregation A platelet aggregation assay wascarried out according to the process described by Fuly et al[17] with modifications Platelet aggregation was monitoredin an Aggregometer (Chrono Logmodel 490 2D HavertownUSA) using Platelet-Rich-Plasma (PRP) PRP was obtainedfrom human whole blood of health volunteers (CAAE14204413500000011) BmatTX-III (rechromatography frac-tion 15)was incubatedwith PRP for fiveminutes at 37∘Cbeingstirred constantly and then platelet aggregation was trig-gered by the addition of ADP (15120583M) or collagen (16 120583gmL)Assays were performed at 37∘C in siliconized glass cuvettesin a final volume of 300 120583L Control experiments wereperformed by adding agonists in the absence of peak 15 Onehundred percent (100) platelet aggregation was obtainedwith a supramaximal concentration of each agonist anddetermined 6 minutes after the addition of each while PRPrsquoslight transmittance showed 0 aggregation
284 Hemorrhage Activity Micewere injected intradermallyin the dorsal region with 50 120583g of crude venom dissolved
in 50120583L of physiological solution [18] Controls received50 120583L of physiological solution in identical conditions After 3hours the animals were euthanized through cervical disloca-tion and the skin was removed The hemorrhagicactivitywasexpressed as size of thehemorrhagic area on the inner surfacemeasured in mm2
285 Coagulation Activity and Minimal Coagulation Dose(MCD)Determination Coagulation activity was tested usinga methodology previously described by Gene et al [19] Inbrief 200120583L of plasma frommicewas distributed in a 96-wellplate and 10 120583L of crude venom containing different concen-trations (0312 0625 125 25 5 and 10 120583g) of proteins wasadded The plate was placed in a thermostatically controlledenvironment and the optical density was measured every 3seconds at 600 nm using an Eonmicroplate spectrophotome-ter (Biotek) in order to evaluate the smallest concentration ofvenom able to coagulate 200120583L of plasmaminute
286 Proteolytic Activity Proteolytic activitywas ascertainedusing azocasein (Sigma) as a substrate according to the pro-cedure described by Charney and Tomarelli [20] Azocaseinsolubilized in distilled water (150 120583L) was added to 7 120583L ofthe crude venom in different protein concentrations (03120625 125 25 5 and 10 120583g) and then the mixture wasincubated in a water bath at 37∘C for 1 hour The reactionstopped when 150 120583L of 20 (mv) trichloroacetic acid wasadded which was followed by incubation for 30min andcentrifugation at 10000timesg for 10 more minutesThen 100 120583Lof the supernatant was transferred to a multiple-well plateand the absorbance was measured at 440 nm using an Eonmicroplate spectrometer (Biotek) and one enzymatic unit (U)was defined as the amount of enzyme necessary to increasethe absorbance by 005
287Myotoxic Activity Myotoxic activity was determined bymeasuring the creatine kinase (CK) activity in the plasma[21] Groups of mice were injected in the gastrocnemiusmuscle with 25 120583g of isolated myotoxins diluted in 50120583L ofphosphate buffered saline (PBS) Negative controls receivedan injection of 50 120583L of PBSThree hours after the injectionsaliquots of mice blood were collected from the caudal veinin heparinized capillaries and centrifuged at 1530timesg for 20minutes Creatine kinasersquos enzymatic activity was determinedusing the CK-NAC kinetic kit (Bioclin Brazil) accordingto the manufacturerrsquos protocol Absorbance was measuredfor 3 minutes at 37∘C in a spectrophotometer at 340 nmEnzymatic activity was expressed in unitsliter (UL) andeach unit consists of the result of the phosphorylation of onenanomol of creatine per minute
288 Neutrophil Viability Neutrophils were collected 6hours after the intraperitoneal (IP) injection of 15mL of3 thioglycollate sterile solution according to the methodpreviously described by Call et al [22] The animals wereeuthanized in order to collect the cells and the peritonealcavity was washed with 3mL of PBS The predominance of
4 BioMed Research International
neutrophils in the liquid obtained was confirmed by micro-scopic analysis with glass slides stained with a panoptic dyeThe peritoneal neutrophils obtained were suspended in anRPMI culturemedium (Gibco-BRL) supplemented with gen-tamicin (100 120583gmL) L-glutamine (2mM) and 10 bovinefetal serum (SFB) in order to obtain 2times105 cells100 120583L Nextcellular viability was assayed over 1 12 and 24 h at 37∘Cand 5 CO
2in which the cells were incubated in a 96-well
plate with the previously isolated PLA2s at a concentration
of 12 120583gmL using RPMI as a negative control After thisthe samples were centrifuged and the supernatant removedCellular viability was determined using the MTT method[23]
289 Quantification of Cytokine EIA was used to evaluateIL-1120573 cytokine as described by Schumacher et al [24] Brieflythe neutrophils (2 times 105 cells200120583L) were either incubatedwith the isolated proteins at 3 6 and 12 120583gmL concentrations(experimental group) or with PMA (positive control group)or with RPMI (negative control) and kept for 12 and 24 hoursat 37∘C in a humid atmosphere with 5 CO
2 After that 96-
well plates were coatedwith 100 120583L of the capturemonoclonalantibody anti-IL-1120573 and incubated for 18 hours at 37∘C Theplate was then washed with washer buffer (PBSTween20)After that 200120583L of blocking buffer containing 5 bovineserum albumin (BSA) in PBSTween20 was added to thewells and the plates were incubated for 1 hour at 37∘CFollowing this the wells were washed and 50 120583L of eithersamples or standard were dispensed into each well and theplates were incubated for 2 hours at 37∘C After this theplate was washed and 100 120583L of detection antibody anti-IL-1120573 was added for 2 hours at 37∘C After incubation andwashing 100 120583L of streptavidin-peroxidase was added fol-lowed by incubation and addition of the substrate (100120583LmL331015840551015840-tetramethylbenzidine) Finally sulfuric acid (50120583L)was added to stop the reaction Absorbances were recordedat 540 and 450 nm and concentration of IL-1120573 was estimatedfrom standard curves prepared with recombinant cytokineThe results were expressed as pgmL of IL-1120573
2810 Antitumor Activity Cytotoxic activity of isolatedPLA2s on human T-cell leukemia (JURKAT) and breast
adenocarcinoma (SK-BR-3) lines obtained from the Amer-ican Culture Collection of Cells (ATCC American TypeCulture Collection Rockville MD USA) were investigated(Figure 6) This activity was assayed by MTT staining asdescribed by Mosmann [25] and adapted by Stabeli et al[21] Cells were dispensed in 96-well plates at a densityof 5 times 105 cellsmL After 24 h of incubation the mediumwas removed and fresh medium with or without differentconcentrations of PLA
2s (BmatTX-I BmatTX-II BmatTX-
III or methotrexate) was added to the wells and incubatedfor another 24 hours The evaluation of the cytotoxic activitywas measured in a spectrophotometer using an interferencefilter of 570 nm and expressed as a percentage
2811 Anti-Leishmania Activity Promastigote forms ofLeishmania amazonensis (IFLABR67PH8) were dis-pensed in a 96-well plate with 1 times 105 cellswell Different
Table 1 Activities induced by B mattogrossensis snake venom
Effecta ActivityPhospholipase activityb (Umg) 186405Proteolytic activityc (U10120583g) 30 plusmn 01
Hemorrhagic halod (cm2) 333 plusmn 005
Coagulation activitye (MCD 120583g) 0325aAll experiments were carried out in triplicate bActivity using NOB stainedsubstrate cOne enzymatic unit (U) was defined as the quantity of enzymeneeded to increase the absorbance by 005UA440 nm dValues 3 h afterincubation with crude venom of B mattogrossensis (50120583g) eMCD theminimum coagulation dose was the dose capable of coagulating 200120583L ofcitrated plasma in less than a minute
concentrations (312 625 125 25 50 and 100120583gmL) ofthe crude venom of B mattogrossensis and the isolatedPLA2s (BmatTX-I and BmatTX-II) were added to each well
100mgmL of pentamidine was used as a positive controlAfter an incubation period of 48 h 10 120583L of a 5mgmL MTTsolution was added Then the plates were placed in theoven at 33∘C with 5 CO
2for 4 hours of incubation after
which 50120583L of SDS (20 wv) was added Absorbance wasmonitored at 570 nm Results were expressed in toxicitypercentage following the equation 1 minus (DO sampleDOcontrol) times 100
29 Statistical Analysis Results were expressed as mean +minusstandard deviation An ANOVA test was used to evaluate thesignificance of the differences observed with 119875 value le 005considered to be significant
3 Results and Discussion
31 Crude Venom Biological Activities B mattogrossensissnake venom induced hemorrhage coagulation proteolyticand phospholipase activities in vitro (Table 1) The hemor-rhagic activity of B mattogrossensis crude venom was eval-uated based on the dimensions of the average hemorrhagichalo which was 333 cm2 This result is in agreement with theresults recently obtained with B atrox [26]
Coagulation activity was confirmed after incubation ofdifferent concentrations of B mattogrossensis crude venomwith plasma in which theminimum coagulation dose capableof promoting coagulation in less than 1min was 0325120583gof protein Analysis of the proteolytic activity of B mat-togrossensis crude venom demonstrated a concentration-dependent response (data not shown) Regarding phospho-lipase proteolytic coagulating and hemorrhagic activitiesof B mattogrossensis crude venom assays confirmed thepresence and activity of proteases and phospholipases Thepresence of metalloproteases was evidenced by the importantformation of an extensive hemorrhagic halo in vivo Thepresence of serine proteases was evidenced by coagulatingactivity present even in low concentrations of the venom(325 120583gmL)
Phospholipase activity of B mattogrossensis crude venomassayed with 4N3OBA synthetic substrate was 186405Umgmeasured by the number of moles of chromophores released
BioMed Research International 5
per minute (n∘molsmin or U) for a given quantity of proteinin milligrams (mg)
The properties of snake venom components observed inthis study are characteristic of accidents caused by snakesof the Bothrops sp genus symptoms such as pain edemahemorrhage and necrosis and additionally systemic distur-bances are characteristic and corroborate the literature thatdescribes that proteases of snake venom proteins are closelyrelated to interference in the hemostatic system promotingblood coagulation fibrinolysis and platelet aggregation [27ndash30]
32 Isolation and Biochemical Characterization of Phospholi-pases The present study showed for the first time the isola-tion of the three phospholipases A
2from B mattogrossensis
snake venom obtained by two chromatographic steps Firstionic exchange chromatography was performed on a CM-Sepharose column with an ammonium bicarbonate gradient(50 to 500mM pH 80) The elution of absorbed proteinswith a linear gradient of concentrated buffer resulted inthirteen fractions (Figure 1(a)) of which fraction nine (9) wasrelated to PLA
2s because it showed phospholipase activity
of 22109Umg on artificial substratum All fractions werelyophilized and submitted to unidimensional electrophoresisrevealing many protein bands
Fraction 9 was submitted to the second chromatographicstep in a reverse column phase on a Discovery C18 columnusing 01 Trifluoroacetic (TFA) and 999 Acetonitrile(ACN) as solvents for the separation of other venom com-ponents The elution of absorbed proteins with a lineargradient of concentrated buffer resulted in twenty-two (22)fractions (Figure 1(b)) The PLA
2s were highly purified with
approximately 40 ACN Similar results were observed inthe isolation of other PLA
2s from snake venoms in high per-
formance liquid chromatography on reverse phase columnswhere the elution profile of these proteins occurs between 30and 40 ACN [5]
The association of chromatographic techniques suchas ionic exchange and reverse phase has commonly beenused and many snake venoms have been fractioned thisway highlighting the phospholipase purification of a speciesbelonging to the old complex ldquoBothrops neuwiedirdquo as well asthe target species studied in this research Two PLA
2basic
isoforms from B (neuwiedi) pauloensis venom were purifiedby Rodrigues et al [31] using biochemical techniques similarto the ones used in this study with the combination of ionicexchange (cationic) and reverse phase chromatographies
Other PLA2s have also been purified using simplified
procedures based in CM-Sepharose andor reverse phase asfor example venomPLA
2s fromBothropsmoojeni [21 32 33]
B pirajai [34 35] B jararacussu [36 37] B alternatus [8 38]Cerastes cerastes [39] and Elaphe climacophora [40]
Fractions 13 and 14 were related to enzymatically inactivePLA2s whereas fraction 15was related to enzymatically active
PLA2s The phospholipase activity of the collected fractions
were analyzed with synthetic NOB and NBD-PC substrates(Figures 2(a) and 2(b))The amount of activity was comparedto BthTX-II a basic enzymatically active PLA
2(Asp49) and
BthTX-I a basic enzymatically inactive PLA2(Lys49) both
20
15
10
5
0
100 200 300 400
80
60
40
20
0
B (
)
(mAU
)
1
23
4
5 67 8
9
10 11 12 13
MW 02 03 04 06 08 09 10 12 13
175805846
30
2317
07
times102
(min)
Btx
(a)
100
80
60
40
20
B (
)
MW175
805846
30
2317
07
(mAU
)
times102
6
5
4
3
2
1
0
0 10 20 30 40
1
2
3 45
6
7
8
910 11
12
13
1415
16
1718
19
20 2122
(min)
07 12 1314 15 16 17 18Btx
(b)
Figure 1 Chromatographic and electrophoretic profile of Bothropsmattogrossensis venom fractioning (a) CM-Sepharose columnsequilibrated with solvent A (50mM ammonium bicarbonatepH 80) and eluted with a 0ndash100 concentration gradient of solventB (500mM ammonium bicarbonate pH 80) at a 50mLminuteflow Emphasis on peak 9 rechromatographed (b) Rechromatog-raphy of fraction 9 on Discovery C18 column equilibrated withsolvent A (01 TFA) and eluted with a concentration gradient of0ndash100 of solvent B (999 acetonitrile and 01 trifluoroaceticacid) and a 10mLmin flow Emphasized in blue are fractions 1314 and 15 characterized as phospholipases BmatTX-I BmatTX-II and BmatTX-III respectively Controls MW molecular weightstandard BTx BthTX-I a basic enzymatically inactive PLA
2(Lys49)
(10120583g) isolated from Bothrops jararacussu venom Absorbancesread at 280 nm Electrophoresis gel made with 125 (ww)acrylamidebis-acrylamide
previously isolated from Bothrops jararacussu venom [36 41]The BmatTX-III PLA
2s (Asp49) were not able to induce
platelet aggregation and did not inhibit collagen or ADPinduced platelet aggregation (data not shown)
The degree of purity of the isolated proteins was furtherdemonstrated by SDS-PAGE mass spectrometry (Figure 3)and N-terminal sequencing (Figures 4(a) and 4(b)) Thepurified PLA
2s were named BmatTX-I BmatTX-II and
6 BioMed Research International
F12
Bmat
TX-I
Bmat
TX-I
I
Bmat
TX-I
II
F16
F17
F18
F19
F20
F21
F22
BthT
X-I
BthT
X-II
0
200
400
600
800
1000Ph
osph
olip
ase a
ctiv
ity (U
mg)
(a)
F12
Bmat
TX-I
II
F16
F17
F20
0
200
400
600
800
1000
Fluo
resc
ent i
nten
sity
(b)
Control BmatTX-I BmatTX-II BmatTX-III0
1000
2000
3000
BmatTX-I (Lys49)BmatTX-II (Lys49)BmatTX-III (Asp49)
Control
CK (U
L)
25120583g50120583L
lowastlowast
lowast
(c)
Figure 2 Enzymatic activity and myotoxic activity of PLA2s isolated from the venom of B mattogrossensis (a) Phospholipase activity of the
fractions collected from the rechromatography of fraction 9 done in C18 column assayed using an NOB stained substrate This activity wasassessed through the measurement of the number of moles of the released chromophore per minute (n∘molsmin or U) per milligram ofprotein (b) Phospholipase activity of the fractions collected from the rechromatography of fraction 9 done in C18 column assayed using afluorescent substrate (c) Myotoxic activity evaluated for inoculation of PLA
2s (25 120583g50 120583L) or PBS (control) done intramuscularly in the
gastrocnemius muscle of mice After 3 hours the creatine kinase (CK) level an important marker of muscular lesion was assayed in theanimalrsquos plasma Each bar represents the average +minus SD of three independent groups lowast119875 lt 005 compared to the control F13 BmatTX-I(Lys49) F14 BmatTX-II (Lys49) F15 BmatTX-III (Asp49) Controls BthTX-I (Lys49) and BthTX-II (Asp49)
BmatTX-III They were characterized as single polypeptidechains with isoelectric points around 10 (data not shown)This result agrees with data from published literature wheremost basic PLA
2s with or without catalytic activity on
artificial substrates [32 42] show an isoelectric point between8 and 10
The average molecular mass defined by mass spectrom-etry was 13304 Da for BmatTX-I 13623Da for BmatTX-IIand 13681Da for BmatTX-III (Figure 3)Thismolecularmassis consistent with most isolated PLA
2s from snake venoms
which are around 13 to 16 kDa [27 36 43]BmatTX-I sequencing showed a lysine (Lys) at position
49 and although only the first 28 amino acid residuesof BmatTX-II have been sequenced both are highly sim-ilar to the PLA
2Lys49 homologue subgroup The N-
terminal sequence alignment of BmatTX-I and BmatTX-IIhas revealed that these basic proteins are PLA
2s similar to
homologous Lys49 and other Lys49 myotoxins from snake
venoms (Figure 4(a)) BmatTX-I showed 94 similarity withMjTX-I present in B moojeni venom 92 with BthTX-I andBOJU-I present in B jararacussu venom and 94 similaritywith MTX-II present in B brazili venom
Additionallymultiple sequence alignment of BmatTX-IIIshowed a PLA
2-Asp49 basic myotoxin with another Asp49
of the Bothrops genera (Figure 4(b)) It can be observedthat BmatTX-III presented 78 similarity with BmjeTX-Iand 75 with BmoTX-I both from B moojeni venom 71similarity with BthTX-II isolated from B jararacussu venomand 65 similarity with PrTX-III from B pirajai venom
In the analysis of the sequences of the PLA2s isolated
from B mattogrossensis in this study highly preserved con-stituent residues of the 120572-helix structure characteristic ofphospholipases were identified as well as the presence ofmany cysteine residues which suggests the existence of manydisulfide bridges important for the stabilization of PLA
2rsquos
molecular structure [21 44]
BioMed Research International 7
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1360475
1398779
(a)
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1362326
1380673
(b)
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1256219
1368171
1402625
(c)
Figure 3 Mass determination by spectrometryThe average proteinmasses of BmatTX-I (a) BmatTX-II (b) and BmatTX-III (c) wereobtained in a MALDI-TOF system operated in linear mode usingexternal standards for calibration The resulting mass spectra weresubmitted to automatic baseline subtraction
Some studies regarding the amino acid composition ofPLA2s demonstrate that these proteins are rich in basic
and hydrophobic amino acids containing three long 120572-helixes two beta sheets and a Ca2+ binding site [45ndash47]Calcium is absolutely necessary for hydrolysis thereforealmost all PLA
2s have a highly preserved region for a Ca2+
bond (XCGXGG) and a catalytic site (DXCCXXHD) [48]It is observed in BmatTX-III that the calcium binding site(27YCGWGG
32) is preserved indicating that it is a catalyt-
ically active PLA2belonging to the PLA
2Asp49 subgroup
33 Biological Activities of Phospholipases A2 The main
objective for the isolation and characterization of compo-nents of snake venom is to better understand the partic-ipation of each component in different pathophysiologicalprocesses resulting from envenomation Local lesions can beattributed to proteases phospholipase activity and hemor-rhagic factors of these venoms followed by the release ofvasoactive agents causing hemorrhaging in various organsand tissues [5 44 49ndash52]
The PLA2s isolated from B mattogrossensis venom
BmatTX-I BmatTX-II and BmatTX-III showed highmyotoxic activity (Figure 2(c)) At a concentration of25 120583g50 120583L the PLA
2s induced a significant release of CK
an important muscular lesion marker when compared to thecontrol Myotoxicity is the characteristic presented by mostbasic phospholipases A
2from snake venoms Several studies
demonstrate that the myotoxic effect begins quickly by directaction of the myotoxic PLA
2s on the plasma membrane of
muscle cells or it is mediated by metalloproteases due toconsequent degeneration and ischemia [27 28 44]
Regarding the Lys49-PLA2myotoxins it is evident that
they lyse the plasma membrane of the muscle cell infected invivo however the exact mechanism has not been describedyet Furthermore it is not known if the toxin is internalizedbefore during or after the initial lysis or if it is not internal-ized Althoughmyotoxicity can be induced by the productionof fatty acids there is a second mechanism that seems to beindependent of the enzymatic activity and is mediated bythe C-terminal region at sites 115ndash129 of the Lys49 molecules[43 53 54]
In an attempt to better understand the development ofthe inflammatory process unleashed by the protein complexpresent in snake venoms many studies have been donesuch as edema induction [55] leukocyte participation [5657] mast cells degranulation [55] participation of variouscytotoxins in the inflammatory system [58 59] participationof cyclooxygenases [60 61] and the participation of venomPLA2s in the inflammatory process [6 51 56 62]
Many studies about PLA2activity in macrophages have
already been done [8 63 64] Little is known about PLA2rsquos
effect on neutrophils however Escocard et al [65] describedan influx of inflammatory cells including many neutrophilsinto the peritoneal cavity of mice after the injection ofBothrops atrox venom The induction of reactive oxygenspecies (ROS) cytokines like IL-6 and IL-1120573 was seen inthese neutrophils These data were also observed in a studycarried out by Souza et al [26] where besides the influx ofneutrophils into the peritoneal cavity ofmice after injection ofthe venom of B atrox there was also induction of superoxideby these cells mast cell degranulation and phagocytosisby macrophages Regarding the activity of PLA
2Gambero
et al [66] have observed the ability of some myotoxins(bothropstoxin-I-II and piratoxin-I) to induce neutrophilchemotaxis in a concentration-dependent manner
In order to evaluate the activation of leukocytes thetoxicity of B mattogrossensis myotoxins on neutrophil cellswas investigated The cells were incubated with differentconcentrations (3 6 and 12 120583gmL) of BmatTX-I (Lys49)BmatTX-II (Lys49) and BmatTX-III (Asp49) myotoxinsduring 1 12 and 24 hours (data not shown)These myotoxinsdid not affect the neutrophilsrsquo viability which agrees withZuliani et al [64] showing low toxicity on thioglycollateelicited mice macrophages
Nonetheless our data showed that neutrophils incubatedwith BmatTX-I and BmatTX-II myotoxins induced therelease of IL-1120573 (Figures 5(a) and 5(b)) Moreover theseresults suggest that phospholipid hydrolysis is not essentialfor the activity observed and argue with the hyphotesis that
8 BioMed Research International
lowastlowast lowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowast lowastlowastlowastlowastlowastlowastlowastlowastlowast
1 10 20 30 40 50(Identity )
100
89
92
92
92
94
94
BmatTX-IBmatTX-IIPrtx-IBOJU-IBthTX-IMjtx-IMtx-Ii
(a)
lowastlowastlowast lowastlowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowastlowast lowastlowastlowast
1 10 20 30 40 50
(Identity )100
78
75
71
65
BmatTX-IIIBmoTX-IBmjeTX-IBthTX-IIPrTX-III
(b)
Figure 4 Comparison of the N-terminal sequence of the snakersquos PLA2s with the PLA
2s isolated from B mattogrossensis venom N-terminal
sequencing of the PLA2s obtained by Edman [13] degradation and alignment done with CLUSTALW2 expressed as of similarity (a)
BmatTX-I BmatTX-II with Lys49 residues in marked sequences (b) BmatTX-III with Asp49 residues in marked sequences Sequencesused for the alignment and their respective access numbers BthTX-I (gi51890398) MjTX-I (gi17368325) Mtx-II (gi390981003) PrTX-I (gi190016174) BOJU-I (gi209572966) BthTX-II (gi1171971) PrTX-III (gi90016174) BmjeTX-I (gi313471399) BmoTX-I (gi221272396)BthTX-I (Lys49) BthTX-II (Asp49) and BOJU-I (Lys49) isolated from the venom of B jararacussu MjTX-I (Lys49) BmoTX-I (Asp49)and BmjeTX-I (Asp49) isolated from the venom of B moojeni Mtx-II (Lys49) isolated from the venom of B brazili PrTX-I (Lys49) andPrTX-III (Asp49) isolated from the venom of B pirajai the ldquolowastrdquo was used to indicate the amino acid residues that are the same between thesequences
other molecular regions distinct from the active site may beinvolved in this effect
PLA2s are multifunctional proteins that can be used as
mediators in several areas of medicine such as in the treat-ment of rheumatoid arthritis as a new class of HIV inhibitorsby blocking the host cell invasion as a potential treatmentagainst malaria and as an antibiotic by inducing cytotoxicityvia the disruption of bacterial membranes [21 23 30 6768] A study by Costa et al [67] with PLA
2s isolated from
B brazili MTX-I and II demonstrated cytotoxic activityagainst Jurkat tumor cells as well as antimicrobial activityagainstE coli andC albicans and antiparasitic activity againstLeishmania sp
In the present study we evaluated the cytotoxic activityof PLA
2s isolated from B mattogrossensis on JURKAT (T
leukemia) and SK-BR-3 (breast adenocarcinoma) both ofwhich are human tumor cell lines Like MTX-II of B brazili[67] the cytotoxic activity of PLA
2s BmatTX-I and BmatTX-
II on JURKATcells was independent of their catalytic activitysince these are characterized as Lys49 PLA
2s and are therefore
catalytically inactive BmatTX-III characterized as Asp49despite being enzymatically active also showed a lower level oftoxicity Some authors propose that the cytotoxic activity ontumor cell lines is associated with the induction of apoptosisconsidering the fact that PLA
2promotes alterations in the
cell membrane And some studies involving Lys49 PLA2s
isolated from B asper demonstrated that the C-terminal
region comprised of amino acids 115ndash129 is concerned withthe cytotoxic and bactericidal activities of this protein [69ndash71] The same observation was made by Costa et al [67] withsynthetic peptides derived from the C-terminal portion ofMTX-I and II PLA
2s
We also evaluated the antiparasitic activity of the crudevenom and isolated PLA
2of B mattogrossensis on promastig-
ote forms of L amazonensis After the analysis it wasobserved that the crude venom of B mattogrossensis presentsincreasing toxic activity (approximately 50 to 80) againstpromastigote forms of L amazonensis after 48 h of incubation(Figure 7(a)) The PLA
2s isolated from B mattogrossensis at
100 120583gmL characterized as BmatTX-I (Lys49) and BmatTX-III (Asp49) presented toxic activity between 25 and 30respectively even with values close to those presented afterincubation of the protozoan with Pentamidine a drug usedas a positive control (Figures 7(b) and 7(c))
When compared the cytotoxicity values of PLA2s against
promastigote forms of L amazonensis show similar activitybetween the Lys49-PLA
2and the Asp49-PLA
2 Comparison
of this activity with the crude venom (70) showed thatPLA2s are responsible for almost half of the observed effect
Nonetheless notably the results suggest that other toxinspresent in the venom contribute to the parasitersquos death
Similar to the results obtained in the present studyStabeli et al [21] demonstrated that MjTX-II a Lys49-PLA
2
homologue isolated from B moojeni venom in different
BioMed Research International 9
0
500
1000
1500
2000
lowast
lowast
lowast
lowast
lowast
Control 3 6 12 3 6 12 3 6 12
ControlBmatTX-I (Lys49)
BmatTX-II (Lys49)BmatTX-III (Asp49)
BmatTX-I (120583g) BmatTX-II (120583g) BmatTX-III (120583g)
IL-1120573
(pg
mL)
(a)
Control 3 6 12 3 6 12 3 6 120
500
1000
1500
2000
ControlBmatTX-I (Lys49)
BmatTX-II (Lys49)BmatTX-III (Asp49)
lowastlowastlowast
BmatTX-I (120583g) BmatTX-II (120583g) BmatTX-III (120583g)
IL-1120573
(pg
mL)
(b)
Figure 5 Production of IL-1120573 by mice neutrophils These cells wereincubatedwith PLA
2(3 6 and 12 120583gmL) or RPMI (control) for 12
hours (a) and 24 hours (b) at 37∘C in a humid atmosphere with 5CO2 Quantification was done by ELISA as described in 29 Each
bar represents the average +minus SDof three independent groups lowast119875 lt005 compared to the control
concentrations (5 25 and 75 120583g) was effective as a parasiticideagent against Schistosomamansoni and promastigote forms ofLeishmania (L amazonensis L braziliensis L donovani andL major)
The action of the PLA2s BmatTX-I (Lys49) and
BmatTX-III (Asp49) on promastigote forms of L ama-zonensis was independent of its catalytic activity sincecatalytically inactive Lys49 myotoxin also demonstrated tox-icity against Leishmania It is believed that the observedcytotoxic activity might be related to the C-terminal regionsof these phospholipase-homologues that are able to promotea disturbance in the cellular membranes independent oftheir catalytic activity [67 72] However more studies arenecessary to define the exact mechanism of action of theseenzymes on parasites
Growing interest in the comprehension of the structureand function of snake venom components especially PLA
2
10 100 10 100 10 100 10 1000
20
40
60
80
100
BmatTX-I (Lys49)BmatTX-II (Lys49)
BmatTX-III (Asp49)Methotrexate (MTX)
Cyto
toxi
city
()
Concentration (120583gmL)
(a)
10 100 10 100 10 100 10 1000
20
40
60
80
100
BmatTX-I (Lys49)BmatTX-II (Lys49)
BmatTX-III (Asp49)Methotrexate (MTX)
Cyto
toxi
city
()
Concentration (120583gmL)
(b)
Figure 6 Antitumoral activity of PLA2s from B mattogrossensis
(a) Antitumoral activity on human acute T-cell leukemia (JURKAT)lines (b) Antitumoral activity on human breast adenocarcinoma(SK-BR-3) Different concentrations of the PLA
2s were incubated
with cell linesMethotrexate was used as the positive control Resultsare presented as mean +minus SD (119899 = 3)
contributes to a better understanding of the mechanism ofaction of their enzymatic and toxic activities It opens thepath to better understand the intoxication caused by enven-omation and the physiopathology behind its side effectsFuture studies will potentially improve serum therapy andhelp develop the pharmaceutical potential that moleculesisolated from animal venoms can have such as the PLA
2s
isolated from the B mattogrossensis venom which show anti-Leishmania and antitumor activities
4 Conclusion
In conclusion the venom of Bothrops mattogrossensis hasa qualitatively similar toxicological profile to previously
10 BioMed Research International
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineB mattogrossensis crude venom
Ant
i-leis
hman
ia ac
tivity
()
B mattogrossensis crude venom (120583g)
(a)
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineBmatTX-I (Lys49)
Ant
i-leis
hman
ia ac
tivity
()
BmatTX-I (120583g)
lowastlowast lowast lowast
(b)
Ant
i-leis
hman
ia ac
tivity
()
Pent 36 625 125 25 50 1000
20
40
60
80
100
BmatTX-III (Asp49)Pentamidine
BmatTX-III (120583g)
lowastlowastlowast
lowastlowast lowast
(c)
Figure 7 Antileishmanial activity of the crude venom of and PLA2s from B mattogrossensis The promastigote forms of L amazonensiswere
plated with 1times105 cellswellThen different concentrations of crude venom and isolated PLA2s were added to each wellThe experiment was
done in a 48 h period with the (a) crude venom and the PLA2enzymes (b) BmatTX-I and (c) BmatTX-III MTT was added and after the
incubation period at 33∘C the formazan crystal formed was dissolved in SDS Readings were done in a spectrophotometer at 570 nm Eachbar represents the average +minus SD of the three independent experimental groups sixfold total lowast119875 lt 005 compared to the control
studied snake venoms of the Bothrops sp genera despite theobservation of quantitative variations Of the three basicPLA2s from B mattogrossensis venom now isolated for the
first time two are characterized as Lys49-PLA2homologues
BmatTX-I and -II and the other as an Asp49-PLA2 named
BmatTX-III This showed high phospholipase activity ThePLA2s isolated induced myotoxic effects as well as the release
of proinflammatory cytokines by neutrophils BmatTX-I and-III PLA
2s were cytotoxic to human tumor cell lines JURKAT
and SK-BR-3 and showed activity against promastigote formsof L amazonensis
Conflict of Interests
The authors state that there is no conflict of interests
Acknowledgments
The authors are grateful to the Ministry of Science andTechnology (MCTI) Conselho Nacional de Desenvolvi-mento Cientıfico e Tecnologico (CNPq) Financiadora deEstudos e Projetos (FINEP) Fundacao de Tecnologia doAcre (FUNTACFDCT) Coordenacao de Aperfeicoamentode Nıvel Superior (CAPES)mdashProjeto NanoBiotec Redede Biodiversidade e Biotecnologia da Amazonia Legal(BIONORTECNPqMCTI) Instituto Nacional paraPesquisa Translacional em Saude e Ambiente na RegiaoAmazonica (INCT-INPeTAmCNPqMCTI) and InstitutoNacional para Pesquisa em Toxinas (INCT-Tox) andSecretary of Development of Rondonia State (SEPLANPRONEXCNPq) for financial support We are grateful to
BioMed Research International 11
Sr Guilherme G Galassi (Mantenedouro da Fauna Silvestreautorizacao IBAMA 020270089912001-95 Americana-SP)for their helpful technical collaboration in provide the snakevenom Amy Grabner provided the English editing of thepaper
References
[1] Y Angulo and B Lomonte ldquoBiochemistry and toxicology oftoxins purified from the venom of the snake Bothrops asperrdquoToxicon vol 54 no 7 pp 949ndash957 2009
[2] J J Calvete P Juarez and L Sanz ldquoSnake venomics Strategyand applicationsrdquo Journal of Mass Spectrometry vol 42 no 11pp 1405ndash1414 2007
[3] T S Kang D Georgieva N Genov et al ldquoEnzymatic toxinsfrom snake venom structural characterization and mechanismof catalysisrdquoThe FEBS Journal vol 278 no 23 pp 4544ndash45762011
[4] C ZOliveiraCaracterizacao funcional e estrutural de inibidoresde fosfolipases A
2isolados do plasma de serpente Bothrops
jararacussu [PhD thesis] Faculdade de Ciencias Farmaceuticasde Ribeirao Preto Ribeirao Preto Brazil 2009
[5] R G Stabeli R Simoes-Silva A M Kayano et al ldquoPurifi-cation of phospholipases A
2from American snake venomsrdquo
in ChromatographymdashThe Most Versatile Method of ChemicalAnalysis chapter 1 pp 1ndash34 InTech Vienna Austria 2012
[6] J P Zuliani C M Fernandes S R Zamuner J M Gutierrezand C F P Teixeira ldquoInflammatory events induced by Lys-49 and Asp-49 phospholipases A
2isolated from Bothrops asper
snake venom role of catalytic activityrdquo Toxicon vol 45 no 3pp 335ndash346 2005
[7] S Marcussi R G Stabeli N A Santos-Filho et al ldquoGenotoxiceffect of Bothrops snake venoms and isolated toxins on humanlymphocyte DNArdquo Toxicon vol 65 pp 9ndash14 2013
[8] S S Setubal A S Pontes J L Furtado et al ldquoAction oftwo phospholipases A
2purified from Bothrops alternatus snake
venom on macrophagesrdquo Biochemistry vol 78 no 2 pp 194ndash203 2013
[9] J M Gutierrez L A Ponce-Soto S Marangoni and BLomonte ldquoSystemic and local myotoxicity induced by snakevenom group II phospholipases A
2 comparison between cro-
toxin crotoxin B and a Lys49 PLA2homologuerdquo Toxicon vol
51 no 1 pp 80ndash92 2008[10] A Q Rueda I G Rodrıguez E C Ara et al ldquoBiochemical
characterization action onmacrophages and superoxide anionproduction of four basic phospholipases A
2from Panamanian
Bothrops asper snake venomrdquo BioMed Research Internationalvol 2013 Article ID 789689 9 pages 2013
[11] V X da Silva and M T Rodrigues ldquoTaxonomic revision ofthe Bothrops neuwiedi complex (Serpentes Viperidae) withdescription of a new speciesrdquo Phyllomedusa vol 7 no 1 pp 45ndash90 2008
[12] J A Campbell and W W Lamar The Venomous Reptiles of theWestern Hemisphere Comstock Ithaca NY USA 2004
[13] P Edman ldquoMethod for determination of the amino acidsequence in peptidesrdquo Acta Chemica Scandinavica vol 4 pp283ndash293 1950
[14] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[15] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[16] M Holzer and S P Mackessy ldquoAn aqueous endpoint assay ofsnake venom phospholipase A
2rdquo Toxicon vol 34 no 10 pp
1149ndash1155 1996[17] A L Fuly A L P deMiranda R B Zingali and J AGuimaraes
ldquoPurification and characterization of a phospholipase A2isoen-
zyme isolated from Lachesis muta snake venomrdquo BiochemicalPharmacology vol 63 no 9 pp 1589ndash1597 2002
[18] T Nikai N Mori M Kishida H Sugihara and A T Tu ldquoIso-lation and biochemical characterization of hemorrhagic toxinf from the venom of Crotalus atrox (Western DiamondbackRattlesnake)rdquo Archives of Biochemistry and Biophysics vol 231no 2 pp 309ndash319 1984
[19] J A Gene A Roy G Rojas J M Gutierrez and L CerdasldquoComparative study on coagulant defibrinating fibrinolyticand fibrinogenolytic activities of Costa Rican crotaline snakevenoms and their neutralization by a polyvalent antivenomrdquoToxicon vol 27 no 8 pp 841ndash848 1989
[20] M S Charney and R M Tomarelli ldquoA colorimetric method forthe determination of the proteolytic activity of duodenal juicerdquoThe Journal of Biological Chemistry vol 171 pp 501ndash505 1947
[21] R G Stabeli S F Amui C D SantrsquoAna et al ldquoBothrops moojenimyotoxin-II a Lys49-phospholipaseA
2homologue an example
of function versatility of snake venom proteinsrdquo ComparativeBiochemistry and Physiology C vol 142 no 3-4 pp 371ndash3812006
[22] D R Call J A Nemzek S J Ebong G L Bolgos D ENewcomb and D G Remick ldquoRatio of local to systemicchemokine concentrations regulates neutrophil recruitmentrdquoThe American Journal of Pathology vol 158 no 2 pp 715ndash7212001
[23] P G Roberto S Kashima S Marcussi et al ldquoCloning andidentification of a complete cDNA coding for a bactericidal andantitumoral acidic phospholipase A
2from Bothrops jararacussu
venomrdquoThe Protein Journal vol 23 no 4 pp 273ndash285 2004[24] J H Schumacher A OrsquoGarra B Shrader et al ldquoThe charac-
terization of four monoclonal antibodies specific for mouse IL-5 and development of mouse and human IL-5 enzyme-linkedimmunosorbentrdquoThe Journal of Immunology vol 141 no 5 pp1576ndash1581 1988
[25] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[26] C A T Souza A M Kayano S S Setubal et al ldquoLocal andsystemic biochemical alterations induced by Bothrops atroxsnake venom in micerdquo Journal of Venom Research vol 3 pp28ndash34 2012
[27] J M Gutierrez and B Lomonte ldquoPhospholipase A2myotoxins
from Bothrops snake venomsrdquo Toxicon vol 33 no 11 pp 1405ndash1424 1995
[28] J M Gutierrez and A Rucavado ldquoSnake venom metallopro-teinases their role in the pathogenesis of local tissue damagerdquoBiochimie vol 82 no 9-10 pp 841ndash850 2000
[29] T Matsui Y Fujimura and K Titani ldquoSnake venom proteasesaffecting hemostasis and thrombosisrdquo Biochimica et BiophysicaActa vol 1477 no 1-2 pp 146ndash156 2000
[30] A M Soares M R M Fontes and J R Giglio ldquoPhospholipaseA2myotoxins from Bothrops snake venoms structure-function
12 BioMed Research International
relationshiprdquoCurrent Organic Chemistry vol 8 no 17 pp 1677ndash1690 2004
[31] VM Rodrigues SMarcussi R S Cambraia et al ldquoBactericidaland neurotoxic activities of two myotoxic phospholipases A
2
from Bothrops neuwiedi pauloensis snake venomrdquo Toxicon vol44 no 3 pp 305ndash314 2004
[32] N A Santos-Filho L B Silveira C Z Oliveira et al ldquoAnew acidic myotoxic anti-platelet and prostaglandin I
2induc-
tor phospholipase A2isolated from Bothrops moojeni snake
venomrdquo Toxicon vol 52 no 8 pp 908ndash917 2008[33] A M Soares S H Andriao-Escarso Y Angulo et al ldquoStruc-
tural and functional characterization of myotoxin I a Lys49phospholipaseA
2homologue fromBothropsmoojeni (Caissaca)
snake venomrdquo Archives of Biochemistry and Biophysics vol 373no 1 pp 7ndash15 2000
[34] A M Soares S H Andriao-Escarso R K Bortoleto et alldquoDissociation of enzymatic and pharmacological properties ofpiratoxins-I and -III two myotoxic phospholipases A
2from
Bothrops pirajai snake venomrdquo Archives of Biochemistry andBiophysics vol 387 no 2 pp 188ndash196 2001
[35] S S Teixeira L B Silveira F M N da Silva et al ldquoMolecularcharacterization of an acidic phospholipase A
2from Bothrops
pirajai snake venom synthetic C-terminal peptide identifies itsantiplatelet regionrdquo Archives of Toxicology vol 85 no 10 pp1219ndash1233 2011
[36] S H Andriao-Escarso A M Soares V M Rodrigues et alldquoMyotoxic phospholipases A
2in Bothrops snake venoms effect
of chemical modifications on the enzymatic and pharmacolog-ical properties of bothropstoxins from Bothrops jararacussurdquoBiochimie vol 82 no 8 pp 755ndash763 2000
[37] S H Andriao-Escarso A M Soares M R M Fontes et alldquoStructural and functional characterization of an acidic plateletaggregation inhibitor and hypotensive phospholipase A
2from
Bothrops jararacussu snake venomrdquo Biochemical Pharmacologyvol 64 no 4 pp 723ndash732 2002
[38] M E G Denegri O C Acosta S Huancahuire-Vega et alldquoIsolation and functional characterization of a new acidic PLA
2
Ba SpII RP4 of the Bothrops alternatus snake venom fromArgentinardquo Toxicon vol 56 no 1 pp 64ndash74 2010
[39] R Zouari-Kessentini J Luis A Karray et al ldquoTwo purified andcharacterized phospholipases A
2from Cerastes cerastes venom
that inhibit cancerous cell adhesion and migrationrdquo Toxiconvol 53 no 4 pp 444ndash453 2009
[40] R Shirai M Toriba K Hayashi K Ikeda and S Inoue ldquoIden-tification and characterization of phospholipase A
2inhibitors
from the serum of the Japanese rat snake Elaphe climacophorardquoToxicon vol 53 no 6 pp 685ndash692 2009
[41] M I Homsi-Brandenburgo L S Queiroz H Santo-Neto LRodrigues-Simoni and J R Giglio ldquoFractionation of Bothropsjararacussu snake venom partial chemical characterization andbiological activity of bothropstoxinrdquo Toxicon vol 26 no 7 pp615ndash627 1988
[42] G P Fernandez A Segura M Herrera et al ldquoNeutralizationof Bothrops mattogrossensis snake venom from Bolivia exper-imental evaluation of llama and donkey antivenoms producedby caprylic acid precipitationrdquoToxicon vol 55 no 2-3 pp 642ndash645 2010
[43] C L Ownby H S S de Araujo S P White and J E FletcherldquoLysine 49 phospholipase A
2proteinsrdquo Toxicon vol 37 no 3
pp 411ndash445 1999[44] A M Soares W P Sestito S Marcussi et al ldquoAlkylation of
myotoxic phospholipases A2in Bothrops moojeni venom a
promising approach to an enhanced antivenom productionrdquoInternational Journal of Biochemistry amp Cell Biology vol 36 no2 pp 258ndash270 2004
[45] B Lomonte J M Gutierrez M F Furtado et al ldquoIsolationof basic myotoxins from Bothrops moojeni and Bothrops atroxsnake venomsrdquo Toxicon vol 28 no 10 pp 1137ndash1146 1990
[46] L C Mancuso M M Correa C A Vieira et al ldquoFractionationof Bothrops pirajai snake venom isolation and characterizationof piratoxin-I a new myotoxic proteinrdquo Toxicon vol 33 no 5pp 615ndash626 1995
[47] H S Selistre L S Queiroz O A B Cunha G E P deSouza and J R Giglio ldquoIsolation and characterization ofhemorrhagic myonecrotic and edema-inducing toxins fromBothrops insularis (jararaca ilhoa) snake venomrdquo Toxicon vol28 no 3 pp 261ndash273 1990
[48] E A Dennis J Cao Y-H Hsu V Magrioti and G KokotosldquoPhospholipaseA
2enzymes physical structure biological func-
tion disease implication chemical inhibition and therapeuticinterventionrdquo Chemical Reviews vol 111 no 10 pp 6130ndash61852011
[49] A G T Cardoso Acao do veneno de Bothrops moojeni e suafracao L-aminoacido oxidase submetida ao tratamento comraios gama de 60CO em Leishmania spp [MS thesis] IPEN-CNENSP Sao Paulo Brazil 1999
[50] MGallacci andWLGCavalcante ldquoUnderstanding the in vitroneuromuscular activity of snake venomLys49 phospholipaseA
2
homologuesrdquo Toxicon vol 55 no 1 pp 1ndash11 2010[51] S S Teixeira Caracterizacao funcional e estrutural de uma
fosfolipase A2acida isolada do veneno de Bothrops pirajai
[MS thesis] Faculdade de Ciencias Farmaceuticas de RibeiraoPretoUSP Ribeirao Preto Brazil 2009
[52] E Valentin and G Lambeau ldquoIncreasing molecular diversityof secreted phospholipases A
2and their receptors and binding
proteinsrdquo Biochimica et Biophysica Acta vol 1488 no 1-2 pp59ndash70 2000
[53] J M Gutierrez and B Lomonte ldquoPhospholipases A2from
Viperidae snake venoms how do they induce skeletal muscledamagerdquo Acta Chimica Slovenica vol 58 no 4 pp 647ndash6592011
[54] J M Gutierrez C L Ownby and G V Odell ldquoIsolation of amyotoxin from Bothrops asper venom partial characterizationand action on skeletal musclerdquo Toxicon vol 22 no 1 pp 115ndash128 1984
[55] N G Nascimento M C Sampaio R A Olivo and C TeixeiraldquoContribution of mast cells to the oedema induced by Bothropsmoojeni snake venom and a pharmacological assessment of theinflammatory mediators involvedrdquo Toxicon vol 55 no 2-3 pp343ndash352 2010
[56] C F P Teixeira E C T Landucci E Antunes M Chacurand Y Cury ldquoInflammatory effects of snake venom myotoxicphospholipases A
2rdquo Toxicon vol 42 no 8 pp 947ndash962 2003
[57] S R Zamuner and C F P Teixeira ldquoCell adhesion moleculesinvolved in the leukocyte recruitment induced by venom of thesnake Bothrops jararacardquoMediators of Inflammation vol 11 no6 pp 351ndash357 2002
[58] F Chaves C F P Teixeira and J M Gutierrez ldquoRole ofTNF-120572 IL-1120573 and IL-6 in the local tissue damage induced byBothrops asper snake venom an experimental assessment inmicerdquo Toxicon vol 45 no 2 pp 171ndash178 2005
BioMed Research International 13
[59] C D F P Teixeira C M Fernandes J P Zuliani and SF Zamuner ldquoInflammatory effects of snake venom metallo-proteinasesrdquo Memorias do Instituto Oswaldo Cruz vol 100supplement 1 pp 181ndash184 2005
[60] R D A Olivo C F P Teixeira J LWallace J M Gutierrez andS R Zamuner ldquoRole of cyclooxygenases in oedema-formingactivity of bothropic venomsrdquo Toxicon vol 49 no 5 pp 670ndash677 2007
[61] L B Silveira D P Marchi-Salvador N A Santos-Filho et alldquoIsolation and expression of a hypotensive and anti-plateletacidic phospholipase A
2from Bothrops moojeni snake venomrdquo
Journal of Pharmaceutical and Biomedical Analysis vol 73 pp35ndash43 2013
[62] A Rucavado T Escalante C F P Teixeira C M Fernandes CDıaz and J M Gutierrez ldquoIncrements in cytokines and matrixmetalloproteinases in skeletal muscle after injection of tissue-damaging toxins from the venom of the snake Bothrops asperrdquoMediators of Inflammation vol 11 no 2 pp 121ndash128 2002
[63] E Leiguez J P Zuliani A M Cianciarullo C M Fernandes JM Gutierrez and C Teixeira ldquoA group IIA-secreted phospho-lipase A
2from snake venom induces lipid body formation in
macrophages the roles of intracellular phospholipases A2and
distinct signaling pathwaysrdquo Journal of Leukocyte Biology vol90 no 1 pp 155ndash166 2011
[64] J P Zuliani J M Gutierrez L L Casais e Silva S C SampaioB Lomonte and C D F P Teixeira ldquoActivation of cellularfunctions in macrophages by venom secretory Asp-49 and Lys-49 phospholipasesA
2rdquoToxicon vol 46 no 5 pp 523ndash532 2005
[65] R D C M Escocard M M Kanashiro J H Petretski et alldquoNeutrophils regulate the expression of cytokines chemokinesand nitric oxide synthasenitric oxide in mice injected withBothrops atrox venomrdquo Immunobiology vol 211 no 1-2 pp 37ndash46 2006
[66] A Gambero E C T Landucci M H Toyama et al ldquoHumanneutrophil migration in vitro induced by secretory phospholi-pases A
2 a role for cell surface glycosaminoglycansrdquo Biochemi-
cal Pharmacology vol 63 no 1 pp 65ndash72 2002[67] F L S Costa R S Rodrigues L F M Izidoro et al ldquoBio-
chemical and functional properties of a thrombin-like enzymeisolated from Bothrops pauloensis snake venomrdquo Toxicon vol54 no 6 pp 725ndash735 2009
[68] J C Villalobos R Mora B Lomonte J M Gutierrez andY Angulo ldquoCytotoxicity induced in myotubes by a Lys49phospholipase A
2homologue from the venom of the snake
Bothrops asper evidence of rapid plasma membrane damageand a dual role for extracellular calciumrdquo Toxicology in Vitrovol 21 no 8 pp 1382ndash1389 2007
[69] B S Cummings J McHowat and R G Schnellmann ldquoPhos-pholipase A
2s in cell injury and deathrdquoThe Journal of Pharma-
cology and Experimental Therapeutics vol 294 no 3 pp 793ndash799 2000
[70] S R Panini L Yang A E Rusinol M S Sinensky J VBonventre and C C Leslie ldquoArachidonate metabolism andthe signaling pathway of induction of apoptosis by oxidizedLDLoxysterolrdquo Journal of Lipid Research vol 42 no 10 pp1678ndash1686 2001
[71] L Paramo B Lomonte J Pizarro-Cerda J-A Bengoechea J-P Gorvel and E Moreno ldquoBactericidal activity of Lys49 andAsp49 myotoxic phospholipases A
2from Bothrops asper snake
venom synthetic Lys49myotoxin II-(115-129)-peptide identifiesits bactericidal regionrdquo European Journal of Biochemistry vol253 no 2 pp 452ndash461 1998
[72] B Lomonte Y Angulo and E Moreno ldquoSynthetic peptidesderived from the C-terminal region of Lys49 phospholipaseA2homologues from viperidae snake venoms biomimetic
activities and potential applicationsrdquo Current PharmaceuticalDesign vol 16 no 28 pp 3224ndash3230 2010
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

4 BioMed Research International
neutrophils in the liquid obtained was confirmed by micro-scopic analysis with glass slides stained with a panoptic dyeThe peritoneal neutrophils obtained were suspended in anRPMI culturemedium (Gibco-BRL) supplemented with gen-tamicin (100 120583gmL) L-glutamine (2mM) and 10 bovinefetal serum (SFB) in order to obtain 2times105 cells100 120583L Nextcellular viability was assayed over 1 12 and 24 h at 37∘Cand 5 CO
2in which the cells were incubated in a 96-well
plate with the previously isolated PLA2s at a concentration
of 12 120583gmL using RPMI as a negative control After thisthe samples were centrifuged and the supernatant removedCellular viability was determined using the MTT method[23]
289 Quantification of Cytokine EIA was used to evaluateIL-1120573 cytokine as described by Schumacher et al [24] Brieflythe neutrophils (2 times 105 cells200120583L) were either incubatedwith the isolated proteins at 3 6 and 12 120583gmL concentrations(experimental group) or with PMA (positive control group)or with RPMI (negative control) and kept for 12 and 24 hoursat 37∘C in a humid atmosphere with 5 CO
2 After that 96-
well plates were coatedwith 100 120583L of the capturemonoclonalantibody anti-IL-1120573 and incubated for 18 hours at 37∘C Theplate was then washed with washer buffer (PBSTween20)After that 200120583L of blocking buffer containing 5 bovineserum albumin (BSA) in PBSTween20 was added to thewells and the plates were incubated for 1 hour at 37∘CFollowing this the wells were washed and 50 120583L of eithersamples or standard were dispensed into each well and theplates were incubated for 2 hours at 37∘C After this theplate was washed and 100 120583L of detection antibody anti-IL-1120573 was added for 2 hours at 37∘C After incubation andwashing 100 120583L of streptavidin-peroxidase was added fol-lowed by incubation and addition of the substrate (100120583LmL331015840551015840-tetramethylbenzidine) Finally sulfuric acid (50120583L)was added to stop the reaction Absorbances were recordedat 540 and 450 nm and concentration of IL-1120573 was estimatedfrom standard curves prepared with recombinant cytokineThe results were expressed as pgmL of IL-1120573
2810 Antitumor Activity Cytotoxic activity of isolatedPLA2s on human T-cell leukemia (JURKAT) and breast
adenocarcinoma (SK-BR-3) lines obtained from the Amer-ican Culture Collection of Cells (ATCC American TypeCulture Collection Rockville MD USA) were investigated(Figure 6) This activity was assayed by MTT staining asdescribed by Mosmann [25] and adapted by Stabeli et al[21] Cells were dispensed in 96-well plates at a densityof 5 times 105 cellsmL After 24 h of incubation the mediumwas removed and fresh medium with or without differentconcentrations of PLA
2s (BmatTX-I BmatTX-II BmatTX-
III or methotrexate) was added to the wells and incubatedfor another 24 hours The evaluation of the cytotoxic activitywas measured in a spectrophotometer using an interferencefilter of 570 nm and expressed as a percentage
2811 Anti-Leishmania Activity Promastigote forms ofLeishmania amazonensis (IFLABR67PH8) were dis-pensed in a 96-well plate with 1 times 105 cellswell Different
Table 1 Activities induced by B mattogrossensis snake venom
Effecta ActivityPhospholipase activityb (Umg) 186405Proteolytic activityc (U10120583g) 30 plusmn 01
Hemorrhagic halod (cm2) 333 plusmn 005
Coagulation activitye (MCD 120583g) 0325aAll experiments were carried out in triplicate bActivity using NOB stainedsubstrate cOne enzymatic unit (U) was defined as the quantity of enzymeneeded to increase the absorbance by 005UA440 nm dValues 3 h afterincubation with crude venom of B mattogrossensis (50120583g) eMCD theminimum coagulation dose was the dose capable of coagulating 200120583L ofcitrated plasma in less than a minute
concentrations (312 625 125 25 50 and 100120583gmL) ofthe crude venom of B mattogrossensis and the isolatedPLA2s (BmatTX-I and BmatTX-II) were added to each well
100mgmL of pentamidine was used as a positive controlAfter an incubation period of 48 h 10 120583L of a 5mgmL MTTsolution was added Then the plates were placed in theoven at 33∘C with 5 CO
2for 4 hours of incubation after
which 50120583L of SDS (20 wv) was added Absorbance wasmonitored at 570 nm Results were expressed in toxicitypercentage following the equation 1 minus (DO sampleDOcontrol) times 100
29 Statistical Analysis Results were expressed as mean +minusstandard deviation An ANOVA test was used to evaluate thesignificance of the differences observed with 119875 value le 005considered to be significant
3 Results and Discussion
31 Crude Venom Biological Activities B mattogrossensissnake venom induced hemorrhage coagulation proteolyticand phospholipase activities in vitro (Table 1) The hemor-rhagic activity of B mattogrossensis crude venom was eval-uated based on the dimensions of the average hemorrhagichalo which was 333 cm2 This result is in agreement with theresults recently obtained with B atrox [26]
Coagulation activity was confirmed after incubation ofdifferent concentrations of B mattogrossensis crude venomwith plasma in which theminimum coagulation dose capableof promoting coagulation in less than 1min was 0325120583gof protein Analysis of the proteolytic activity of B mat-togrossensis crude venom demonstrated a concentration-dependent response (data not shown) Regarding phospho-lipase proteolytic coagulating and hemorrhagic activitiesof B mattogrossensis crude venom assays confirmed thepresence and activity of proteases and phospholipases Thepresence of metalloproteases was evidenced by the importantformation of an extensive hemorrhagic halo in vivo Thepresence of serine proteases was evidenced by coagulatingactivity present even in low concentrations of the venom(325 120583gmL)
Phospholipase activity of B mattogrossensis crude venomassayed with 4N3OBA synthetic substrate was 186405Umgmeasured by the number of moles of chromophores released
BioMed Research International 5
per minute (n∘molsmin or U) for a given quantity of proteinin milligrams (mg)
The properties of snake venom components observed inthis study are characteristic of accidents caused by snakesof the Bothrops sp genus symptoms such as pain edemahemorrhage and necrosis and additionally systemic distur-bances are characteristic and corroborate the literature thatdescribes that proteases of snake venom proteins are closelyrelated to interference in the hemostatic system promotingblood coagulation fibrinolysis and platelet aggregation [27ndash30]
32 Isolation and Biochemical Characterization of Phospholi-pases The present study showed for the first time the isola-tion of the three phospholipases A
2from B mattogrossensis
snake venom obtained by two chromatographic steps Firstionic exchange chromatography was performed on a CM-Sepharose column with an ammonium bicarbonate gradient(50 to 500mM pH 80) The elution of absorbed proteinswith a linear gradient of concentrated buffer resulted inthirteen fractions (Figure 1(a)) of which fraction nine (9) wasrelated to PLA
2s because it showed phospholipase activity
of 22109Umg on artificial substratum All fractions werelyophilized and submitted to unidimensional electrophoresisrevealing many protein bands
Fraction 9 was submitted to the second chromatographicstep in a reverse column phase on a Discovery C18 columnusing 01 Trifluoroacetic (TFA) and 999 Acetonitrile(ACN) as solvents for the separation of other venom com-ponents The elution of absorbed proteins with a lineargradient of concentrated buffer resulted in twenty-two (22)fractions (Figure 1(b)) The PLA
2s were highly purified with
approximately 40 ACN Similar results were observed inthe isolation of other PLA
2s from snake venoms in high per-
formance liquid chromatography on reverse phase columnswhere the elution profile of these proteins occurs between 30and 40 ACN [5]
The association of chromatographic techniques suchas ionic exchange and reverse phase has commonly beenused and many snake venoms have been fractioned thisway highlighting the phospholipase purification of a speciesbelonging to the old complex ldquoBothrops neuwiedirdquo as well asthe target species studied in this research Two PLA
2basic
isoforms from B (neuwiedi) pauloensis venom were purifiedby Rodrigues et al [31] using biochemical techniques similarto the ones used in this study with the combination of ionicexchange (cationic) and reverse phase chromatographies
Other PLA2s have also been purified using simplified
procedures based in CM-Sepharose andor reverse phase asfor example venomPLA
2s fromBothropsmoojeni [21 32 33]
B pirajai [34 35] B jararacussu [36 37] B alternatus [8 38]Cerastes cerastes [39] and Elaphe climacophora [40]
Fractions 13 and 14 were related to enzymatically inactivePLA2s whereas fraction 15was related to enzymatically active
PLA2s The phospholipase activity of the collected fractions
were analyzed with synthetic NOB and NBD-PC substrates(Figures 2(a) and 2(b))The amount of activity was comparedto BthTX-II a basic enzymatically active PLA
2(Asp49) and
BthTX-I a basic enzymatically inactive PLA2(Lys49) both
20
15
10
5
0
100 200 300 400
80
60
40
20
0
B (
)
(mAU
)
1
23
4
5 67 8
9
10 11 12 13
MW 02 03 04 06 08 09 10 12 13
175805846
30
2317
07
times102
(min)
Btx
(a)
100
80
60
40
20
B (
)
MW175
805846
30
2317
07
(mAU
)
times102
6
5
4
3
2
1
0
0 10 20 30 40
1
2
3 45
6
7
8
910 11
12
13
1415
16
1718
19
20 2122
(min)
07 12 1314 15 16 17 18Btx
(b)
Figure 1 Chromatographic and electrophoretic profile of Bothropsmattogrossensis venom fractioning (a) CM-Sepharose columnsequilibrated with solvent A (50mM ammonium bicarbonatepH 80) and eluted with a 0ndash100 concentration gradient of solventB (500mM ammonium bicarbonate pH 80) at a 50mLminuteflow Emphasis on peak 9 rechromatographed (b) Rechromatog-raphy of fraction 9 on Discovery C18 column equilibrated withsolvent A (01 TFA) and eluted with a concentration gradient of0ndash100 of solvent B (999 acetonitrile and 01 trifluoroaceticacid) and a 10mLmin flow Emphasized in blue are fractions 1314 and 15 characterized as phospholipases BmatTX-I BmatTX-II and BmatTX-III respectively Controls MW molecular weightstandard BTx BthTX-I a basic enzymatically inactive PLA
2(Lys49)
(10120583g) isolated from Bothrops jararacussu venom Absorbancesread at 280 nm Electrophoresis gel made with 125 (ww)acrylamidebis-acrylamide
previously isolated from Bothrops jararacussu venom [36 41]The BmatTX-III PLA
2s (Asp49) were not able to induce
platelet aggregation and did not inhibit collagen or ADPinduced platelet aggregation (data not shown)
The degree of purity of the isolated proteins was furtherdemonstrated by SDS-PAGE mass spectrometry (Figure 3)and N-terminal sequencing (Figures 4(a) and 4(b)) Thepurified PLA
2s were named BmatTX-I BmatTX-II and
6 BioMed Research International
F12
Bmat
TX-I
Bmat
TX-I
I
Bmat
TX-I
II
F16
F17
F18
F19
F20
F21
F22
BthT
X-I
BthT
X-II
0
200
400
600
800
1000Ph
osph
olip
ase a
ctiv
ity (U
mg)
(a)
F12
Bmat
TX-I
II
F16
F17
F20
0
200
400
600
800
1000
Fluo
resc
ent i
nten
sity
(b)
Control BmatTX-I BmatTX-II BmatTX-III0
1000
2000
3000
BmatTX-I (Lys49)BmatTX-II (Lys49)BmatTX-III (Asp49)
Control
CK (U
L)
25120583g50120583L
lowastlowast
lowast
(c)
Figure 2 Enzymatic activity and myotoxic activity of PLA2s isolated from the venom of B mattogrossensis (a) Phospholipase activity of the
fractions collected from the rechromatography of fraction 9 done in C18 column assayed using an NOB stained substrate This activity wasassessed through the measurement of the number of moles of the released chromophore per minute (n∘molsmin or U) per milligram ofprotein (b) Phospholipase activity of the fractions collected from the rechromatography of fraction 9 done in C18 column assayed using afluorescent substrate (c) Myotoxic activity evaluated for inoculation of PLA
2s (25 120583g50 120583L) or PBS (control) done intramuscularly in the
gastrocnemius muscle of mice After 3 hours the creatine kinase (CK) level an important marker of muscular lesion was assayed in theanimalrsquos plasma Each bar represents the average +minus SD of three independent groups lowast119875 lt 005 compared to the control F13 BmatTX-I(Lys49) F14 BmatTX-II (Lys49) F15 BmatTX-III (Asp49) Controls BthTX-I (Lys49) and BthTX-II (Asp49)
BmatTX-III They were characterized as single polypeptidechains with isoelectric points around 10 (data not shown)This result agrees with data from published literature wheremost basic PLA
2s with or without catalytic activity on
artificial substrates [32 42] show an isoelectric point between8 and 10
The average molecular mass defined by mass spectrom-etry was 13304 Da for BmatTX-I 13623Da for BmatTX-IIand 13681Da for BmatTX-III (Figure 3)Thismolecularmassis consistent with most isolated PLA
2s from snake venoms
which are around 13 to 16 kDa [27 36 43]BmatTX-I sequencing showed a lysine (Lys) at position
49 and although only the first 28 amino acid residuesof BmatTX-II have been sequenced both are highly sim-ilar to the PLA
2Lys49 homologue subgroup The N-
terminal sequence alignment of BmatTX-I and BmatTX-IIhas revealed that these basic proteins are PLA
2s similar to
homologous Lys49 and other Lys49 myotoxins from snake
venoms (Figure 4(a)) BmatTX-I showed 94 similarity withMjTX-I present in B moojeni venom 92 with BthTX-I andBOJU-I present in B jararacussu venom and 94 similaritywith MTX-II present in B brazili venom
Additionallymultiple sequence alignment of BmatTX-IIIshowed a PLA
2-Asp49 basic myotoxin with another Asp49
of the Bothrops genera (Figure 4(b)) It can be observedthat BmatTX-III presented 78 similarity with BmjeTX-Iand 75 with BmoTX-I both from B moojeni venom 71similarity with BthTX-II isolated from B jararacussu venomand 65 similarity with PrTX-III from B pirajai venom
In the analysis of the sequences of the PLA2s isolated
from B mattogrossensis in this study highly preserved con-stituent residues of the 120572-helix structure characteristic ofphospholipases were identified as well as the presence ofmany cysteine residues which suggests the existence of manydisulfide bridges important for the stabilization of PLA
2rsquos
molecular structure [21 44]
BioMed Research International 7
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1360475
1398779
(a)
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1362326
1380673
(b)
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1256219
1368171
1402625
(c)
Figure 3 Mass determination by spectrometryThe average proteinmasses of BmatTX-I (a) BmatTX-II (b) and BmatTX-III (c) wereobtained in a MALDI-TOF system operated in linear mode usingexternal standards for calibration The resulting mass spectra weresubmitted to automatic baseline subtraction
Some studies regarding the amino acid composition ofPLA2s demonstrate that these proteins are rich in basic
and hydrophobic amino acids containing three long 120572-helixes two beta sheets and a Ca2+ binding site [45ndash47]Calcium is absolutely necessary for hydrolysis thereforealmost all PLA
2s have a highly preserved region for a Ca2+
bond (XCGXGG) and a catalytic site (DXCCXXHD) [48]It is observed in BmatTX-III that the calcium binding site(27YCGWGG
32) is preserved indicating that it is a catalyt-
ically active PLA2belonging to the PLA
2Asp49 subgroup
33 Biological Activities of Phospholipases A2 The main
objective for the isolation and characterization of compo-nents of snake venom is to better understand the partic-ipation of each component in different pathophysiologicalprocesses resulting from envenomation Local lesions can beattributed to proteases phospholipase activity and hemor-rhagic factors of these venoms followed by the release ofvasoactive agents causing hemorrhaging in various organsand tissues [5 44 49ndash52]
The PLA2s isolated from B mattogrossensis venom
BmatTX-I BmatTX-II and BmatTX-III showed highmyotoxic activity (Figure 2(c)) At a concentration of25 120583g50 120583L the PLA
2s induced a significant release of CK
an important muscular lesion marker when compared to thecontrol Myotoxicity is the characteristic presented by mostbasic phospholipases A
2from snake venoms Several studies
demonstrate that the myotoxic effect begins quickly by directaction of the myotoxic PLA
2s on the plasma membrane of
muscle cells or it is mediated by metalloproteases due toconsequent degeneration and ischemia [27 28 44]
Regarding the Lys49-PLA2myotoxins it is evident that
they lyse the plasma membrane of the muscle cell infected invivo however the exact mechanism has not been describedyet Furthermore it is not known if the toxin is internalizedbefore during or after the initial lysis or if it is not internal-ized Althoughmyotoxicity can be induced by the productionof fatty acids there is a second mechanism that seems to beindependent of the enzymatic activity and is mediated bythe C-terminal region at sites 115ndash129 of the Lys49 molecules[43 53 54]
In an attempt to better understand the development ofthe inflammatory process unleashed by the protein complexpresent in snake venoms many studies have been donesuch as edema induction [55] leukocyte participation [5657] mast cells degranulation [55] participation of variouscytotoxins in the inflammatory system [58 59] participationof cyclooxygenases [60 61] and the participation of venomPLA2s in the inflammatory process [6 51 56 62]
Many studies about PLA2activity in macrophages have
already been done [8 63 64] Little is known about PLA2rsquos
effect on neutrophils however Escocard et al [65] describedan influx of inflammatory cells including many neutrophilsinto the peritoneal cavity of mice after the injection ofBothrops atrox venom The induction of reactive oxygenspecies (ROS) cytokines like IL-6 and IL-1120573 was seen inthese neutrophils These data were also observed in a studycarried out by Souza et al [26] where besides the influx ofneutrophils into the peritoneal cavity ofmice after injection ofthe venom of B atrox there was also induction of superoxideby these cells mast cell degranulation and phagocytosisby macrophages Regarding the activity of PLA
2Gambero
et al [66] have observed the ability of some myotoxins(bothropstoxin-I-II and piratoxin-I) to induce neutrophilchemotaxis in a concentration-dependent manner
In order to evaluate the activation of leukocytes thetoxicity of B mattogrossensis myotoxins on neutrophil cellswas investigated The cells were incubated with differentconcentrations (3 6 and 12 120583gmL) of BmatTX-I (Lys49)BmatTX-II (Lys49) and BmatTX-III (Asp49) myotoxinsduring 1 12 and 24 hours (data not shown)These myotoxinsdid not affect the neutrophilsrsquo viability which agrees withZuliani et al [64] showing low toxicity on thioglycollateelicited mice macrophages
Nonetheless our data showed that neutrophils incubatedwith BmatTX-I and BmatTX-II myotoxins induced therelease of IL-1120573 (Figures 5(a) and 5(b)) Moreover theseresults suggest that phospholipid hydrolysis is not essentialfor the activity observed and argue with the hyphotesis that
8 BioMed Research International
lowastlowast lowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowast lowastlowastlowastlowastlowastlowastlowastlowastlowast
1 10 20 30 40 50(Identity )
100
89
92
92
92
94
94
BmatTX-IBmatTX-IIPrtx-IBOJU-IBthTX-IMjtx-IMtx-Ii
(a)
lowastlowastlowast lowastlowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowastlowast lowastlowastlowast
1 10 20 30 40 50
(Identity )100
78
75
71
65
BmatTX-IIIBmoTX-IBmjeTX-IBthTX-IIPrTX-III
(b)
Figure 4 Comparison of the N-terminal sequence of the snakersquos PLA2s with the PLA
2s isolated from B mattogrossensis venom N-terminal
sequencing of the PLA2s obtained by Edman [13] degradation and alignment done with CLUSTALW2 expressed as of similarity (a)
BmatTX-I BmatTX-II with Lys49 residues in marked sequences (b) BmatTX-III with Asp49 residues in marked sequences Sequencesused for the alignment and their respective access numbers BthTX-I (gi51890398) MjTX-I (gi17368325) Mtx-II (gi390981003) PrTX-I (gi190016174) BOJU-I (gi209572966) BthTX-II (gi1171971) PrTX-III (gi90016174) BmjeTX-I (gi313471399) BmoTX-I (gi221272396)BthTX-I (Lys49) BthTX-II (Asp49) and BOJU-I (Lys49) isolated from the venom of B jararacussu MjTX-I (Lys49) BmoTX-I (Asp49)and BmjeTX-I (Asp49) isolated from the venom of B moojeni Mtx-II (Lys49) isolated from the venom of B brazili PrTX-I (Lys49) andPrTX-III (Asp49) isolated from the venom of B pirajai the ldquolowastrdquo was used to indicate the amino acid residues that are the same between thesequences
other molecular regions distinct from the active site may beinvolved in this effect
PLA2s are multifunctional proteins that can be used as
mediators in several areas of medicine such as in the treat-ment of rheumatoid arthritis as a new class of HIV inhibitorsby blocking the host cell invasion as a potential treatmentagainst malaria and as an antibiotic by inducing cytotoxicityvia the disruption of bacterial membranes [21 23 30 6768] A study by Costa et al [67] with PLA
2s isolated from
B brazili MTX-I and II demonstrated cytotoxic activityagainst Jurkat tumor cells as well as antimicrobial activityagainstE coli andC albicans and antiparasitic activity againstLeishmania sp
In the present study we evaluated the cytotoxic activityof PLA
2s isolated from B mattogrossensis on JURKAT (T
leukemia) and SK-BR-3 (breast adenocarcinoma) both ofwhich are human tumor cell lines Like MTX-II of B brazili[67] the cytotoxic activity of PLA
2s BmatTX-I and BmatTX-
II on JURKATcells was independent of their catalytic activitysince these are characterized as Lys49 PLA
2s and are therefore
catalytically inactive BmatTX-III characterized as Asp49despite being enzymatically active also showed a lower level oftoxicity Some authors propose that the cytotoxic activity ontumor cell lines is associated with the induction of apoptosisconsidering the fact that PLA
2promotes alterations in the
cell membrane And some studies involving Lys49 PLA2s
isolated from B asper demonstrated that the C-terminal
region comprised of amino acids 115ndash129 is concerned withthe cytotoxic and bactericidal activities of this protein [69ndash71] The same observation was made by Costa et al [67] withsynthetic peptides derived from the C-terminal portion ofMTX-I and II PLA
2s
We also evaluated the antiparasitic activity of the crudevenom and isolated PLA
2of B mattogrossensis on promastig-
ote forms of L amazonensis After the analysis it wasobserved that the crude venom of B mattogrossensis presentsincreasing toxic activity (approximately 50 to 80) againstpromastigote forms of L amazonensis after 48 h of incubation(Figure 7(a)) The PLA
2s isolated from B mattogrossensis at
100 120583gmL characterized as BmatTX-I (Lys49) and BmatTX-III (Asp49) presented toxic activity between 25 and 30respectively even with values close to those presented afterincubation of the protozoan with Pentamidine a drug usedas a positive control (Figures 7(b) and 7(c))
When compared the cytotoxicity values of PLA2s against
promastigote forms of L amazonensis show similar activitybetween the Lys49-PLA
2and the Asp49-PLA
2 Comparison
of this activity with the crude venom (70) showed thatPLA2s are responsible for almost half of the observed effect
Nonetheless notably the results suggest that other toxinspresent in the venom contribute to the parasitersquos death
Similar to the results obtained in the present studyStabeli et al [21] demonstrated that MjTX-II a Lys49-PLA
2
homologue isolated from B moojeni venom in different
BioMed Research International 9
0
500
1000
1500
2000
lowast
lowast
lowast
lowast
lowast
Control 3 6 12 3 6 12 3 6 12
ControlBmatTX-I (Lys49)
BmatTX-II (Lys49)BmatTX-III (Asp49)
BmatTX-I (120583g) BmatTX-II (120583g) BmatTX-III (120583g)
IL-1120573
(pg
mL)
(a)
Control 3 6 12 3 6 12 3 6 120
500
1000
1500
2000
ControlBmatTX-I (Lys49)
BmatTX-II (Lys49)BmatTX-III (Asp49)
lowastlowastlowast
BmatTX-I (120583g) BmatTX-II (120583g) BmatTX-III (120583g)
IL-1120573
(pg
mL)
(b)
Figure 5 Production of IL-1120573 by mice neutrophils These cells wereincubatedwith PLA
2(3 6 and 12 120583gmL) or RPMI (control) for 12
hours (a) and 24 hours (b) at 37∘C in a humid atmosphere with 5CO2 Quantification was done by ELISA as described in 29 Each
bar represents the average +minus SDof three independent groups lowast119875 lt005 compared to the control
concentrations (5 25 and 75 120583g) was effective as a parasiticideagent against Schistosomamansoni and promastigote forms ofLeishmania (L amazonensis L braziliensis L donovani andL major)
The action of the PLA2s BmatTX-I (Lys49) and
BmatTX-III (Asp49) on promastigote forms of L ama-zonensis was independent of its catalytic activity sincecatalytically inactive Lys49 myotoxin also demonstrated tox-icity against Leishmania It is believed that the observedcytotoxic activity might be related to the C-terminal regionsof these phospholipase-homologues that are able to promotea disturbance in the cellular membranes independent oftheir catalytic activity [67 72] However more studies arenecessary to define the exact mechanism of action of theseenzymes on parasites
Growing interest in the comprehension of the structureand function of snake venom components especially PLA
2
10 100 10 100 10 100 10 1000
20
40
60
80
100
BmatTX-I (Lys49)BmatTX-II (Lys49)
BmatTX-III (Asp49)Methotrexate (MTX)
Cyto
toxi
city
()
Concentration (120583gmL)
(a)
10 100 10 100 10 100 10 1000
20
40
60
80
100
BmatTX-I (Lys49)BmatTX-II (Lys49)
BmatTX-III (Asp49)Methotrexate (MTX)
Cyto
toxi
city
()
Concentration (120583gmL)
(b)
Figure 6 Antitumoral activity of PLA2s from B mattogrossensis
(a) Antitumoral activity on human acute T-cell leukemia (JURKAT)lines (b) Antitumoral activity on human breast adenocarcinoma(SK-BR-3) Different concentrations of the PLA
2s were incubated
with cell linesMethotrexate was used as the positive control Resultsare presented as mean +minus SD (119899 = 3)
contributes to a better understanding of the mechanism ofaction of their enzymatic and toxic activities It opens thepath to better understand the intoxication caused by enven-omation and the physiopathology behind its side effectsFuture studies will potentially improve serum therapy andhelp develop the pharmaceutical potential that moleculesisolated from animal venoms can have such as the PLA
2s
isolated from the B mattogrossensis venom which show anti-Leishmania and antitumor activities
4 Conclusion
In conclusion the venom of Bothrops mattogrossensis hasa qualitatively similar toxicological profile to previously
10 BioMed Research International
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineB mattogrossensis crude venom
Ant
i-leis
hman
ia ac
tivity
()
B mattogrossensis crude venom (120583g)
(a)
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineBmatTX-I (Lys49)
Ant
i-leis
hman
ia ac
tivity
()
BmatTX-I (120583g)
lowastlowast lowast lowast
(b)
Ant
i-leis
hman
ia ac
tivity
()
Pent 36 625 125 25 50 1000
20
40
60
80
100
BmatTX-III (Asp49)Pentamidine
BmatTX-III (120583g)
lowastlowastlowast
lowastlowast lowast
(c)
Figure 7 Antileishmanial activity of the crude venom of and PLA2s from B mattogrossensis The promastigote forms of L amazonensiswere
plated with 1times105 cellswellThen different concentrations of crude venom and isolated PLA2s were added to each wellThe experiment was
done in a 48 h period with the (a) crude venom and the PLA2enzymes (b) BmatTX-I and (c) BmatTX-III MTT was added and after the
incubation period at 33∘C the formazan crystal formed was dissolved in SDS Readings were done in a spectrophotometer at 570 nm Eachbar represents the average +minus SD of the three independent experimental groups sixfold total lowast119875 lt 005 compared to the control
studied snake venoms of the Bothrops sp genera despite theobservation of quantitative variations Of the three basicPLA2s from B mattogrossensis venom now isolated for the
first time two are characterized as Lys49-PLA2homologues
BmatTX-I and -II and the other as an Asp49-PLA2 named
BmatTX-III This showed high phospholipase activity ThePLA2s isolated induced myotoxic effects as well as the release
of proinflammatory cytokines by neutrophils BmatTX-I and-III PLA
2s were cytotoxic to human tumor cell lines JURKAT
and SK-BR-3 and showed activity against promastigote formsof L amazonensis
Conflict of Interests
The authors state that there is no conflict of interests
Acknowledgments
The authors are grateful to the Ministry of Science andTechnology (MCTI) Conselho Nacional de Desenvolvi-mento Cientıfico e Tecnologico (CNPq) Financiadora deEstudos e Projetos (FINEP) Fundacao de Tecnologia doAcre (FUNTACFDCT) Coordenacao de Aperfeicoamentode Nıvel Superior (CAPES)mdashProjeto NanoBiotec Redede Biodiversidade e Biotecnologia da Amazonia Legal(BIONORTECNPqMCTI) Instituto Nacional paraPesquisa Translacional em Saude e Ambiente na RegiaoAmazonica (INCT-INPeTAmCNPqMCTI) and InstitutoNacional para Pesquisa em Toxinas (INCT-Tox) andSecretary of Development of Rondonia State (SEPLANPRONEXCNPq) for financial support We are grateful to
BioMed Research International 11
Sr Guilherme G Galassi (Mantenedouro da Fauna Silvestreautorizacao IBAMA 020270089912001-95 Americana-SP)for their helpful technical collaboration in provide the snakevenom Amy Grabner provided the English editing of thepaper
References
[1] Y Angulo and B Lomonte ldquoBiochemistry and toxicology oftoxins purified from the venom of the snake Bothrops asperrdquoToxicon vol 54 no 7 pp 949ndash957 2009
[2] J J Calvete P Juarez and L Sanz ldquoSnake venomics Strategyand applicationsrdquo Journal of Mass Spectrometry vol 42 no 11pp 1405ndash1414 2007
[3] T S Kang D Georgieva N Genov et al ldquoEnzymatic toxinsfrom snake venom structural characterization and mechanismof catalysisrdquoThe FEBS Journal vol 278 no 23 pp 4544ndash45762011
[4] C ZOliveiraCaracterizacao funcional e estrutural de inibidoresde fosfolipases A
2isolados do plasma de serpente Bothrops
jararacussu [PhD thesis] Faculdade de Ciencias Farmaceuticasde Ribeirao Preto Ribeirao Preto Brazil 2009
[5] R G Stabeli R Simoes-Silva A M Kayano et al ldquoPurifi-cation of phospholipases A
2from American snake venomsrdquo
in ChromatographymdashThe Most Versatile Method of ChemicalAnalysis chapter 1 pp 1ndash34 InTech Vienna Austria 2012
[6] J P Zuliani C M Fernandes S R Zamuner J M Gutierrezand C F P Teixeira ldquoInflammatory events induced by Lys-49 and Asp-49 phospholipases A
2isolated from Bothrops asper
snake venom role of catalytic activityrdquo Toxicon vol 45 no 3pp 335ndash346 2005
[7] S Marcussi R G Stabeli N A Santos-Filho et al ldquoGenotoxiceffect of Bothrops snake venoms and isolated toxins on humanlymphocyte DNArdquo Toxicon vol 65 pp 9ndash14 2013
[8] S S Setubal A S Pontes J L Furtado et al ldquoAction oftwo phospholipases A
2purified from Bothrops alternatus snake
venom on macrophagesrdquo Biochemistry vol 78 no 2 pp 194ndash203 2013
[9] J M Gutierrez L A Ponce-Soto S Marangoni and BLomonte ldquoSystemic and local myotoxicity induced by snakevenom group II phospholipases A
2 comparison between cro-
toxin crotoxin B and a Lys49 PLA2homologuerdquo Toxicon vol
51 no 1 pp 80ndash92 2008[10] A Q Rueda I G Rodrıguez E C Ara et al ldquoBiochemical
characterization action onmacrophages and superoxide anionproduction of four basic phospholipases A
2from Panamanian
Bothrops asper snake venomrdquo BioMed Research Internationalvol 2013 Article ID 789689 9 pages 2013
[11] V X da Silva and M T Rodrigues ldquoTaxonomic revision ofthe Bothrops neuwiedi complex (Serpentes Viperidae) withdescription of a new speciesrdquo Phyllomedusa vol 7 no 1 pp 45ndash90 2008
[12] J A Campbell and W W Lamar The Venomous Reptiles of theWestern Hemisphere Comstock Ithaca NY USA 2004
[13] P Edman ldquoMethod for determination of the amino acidsequence in peptidesrdquo Acta Chemica Scandinavica vol 4 pp283ndash293 1950
[14] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[15] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[16] M Holzer and S P Mackessy ldquoAn aqueous endpoint assay ofsnake venom phospholipase A
2rdquo Toxicon vol 34 no 10 pp
1149ndash1155 1996[17] A L Fuly A L P deMiranda R B Zingali and J AGuimaraes
ldquoPurification and characterization of a phospholipase A2isoen-
zyme isolated from Lachesis muta snake venomrdquo BiochemicalPharmacology vol 63 no 9 pp 1589ndash1597 2002
[18] T Nikai N Mori M Kishida H Sugihara and A T Tu ldquoIso-lation and biochemical characterization of hemorrhagic toxinf from the venom of Crotalus atrox (Western DiamondbackRattlesnake)rdquo Archives of Biochemistry and Biophysics vol 231no 2 pp 309ndash319 1984
[19] J A Gene A Roy G Rojas J M Gutierrez and L CerdasldquoComparative study on coagulant defibrinating fibrinolyticand fibrinogenolytic activities of Costa Rican crotaline snakevenoms and their neutralization by a polyvalent antivenomrdquoToxicon vol 27 no 8 pp 841ndash848 1989
[20] M S Charney and R M Tomarelli ldquoA colorimetric method forthe determination of the proteolytic activity of duodenal juicerdquoThe Journal of Biological Chemistry vol 171 pp 501ndash505 1947
[21] R G Stabeli S F Amui C D SantrsquoAna et al ldquoBothrops moojenimyotoxin-II a Lys49-phospholipaseA
2homologue an example
of function versatility of snake venom proteinsrdquo ComparativeBiochemistry and Physiology C vol 142 no 3-4 pp 371ndash3812006
[22] D R Call J A Nemzek S J Ebong G L Bolgos D ENewcomb and D G Remick ldquoRatio of local to systemicchemokine concentrations regulates neutrophil recruitmentrdquoThe American Journal of Pathology vol 158 no 2 pp 715ndash7212001
[23] P G Roberto S Kashima S Marcussi et al ldquoCloning andidentification of a complete cDNA coding for a bactericidal andantitumoral acidic phospholipase A
2from Bothrops jararacussu
venomrdquoThe Protein Journal vol 23 no 4 pp 273ndash285 2004[24] J H Schumacher A OrsquoGarra B Shrader et al ldquoThe charac-
terization of four monoclonal antibodies specific for mouse IL-5 and development of mouse and human IL-5 enzyme-linkedimmunosorbentrdquoThe Journal of Immunology vol 141 no 5 pp1576ndash1581 1988
[25] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[26] C A T Souza A M Kayano S S Setubal et al ldquoLocal andsystemic biochemical alterations induced by Bothrops atroxsnake venom in micerdquo Journal of Venom Research vol 3 pp28ndash34 2012
[27] J M Gutierrez and B Lomonte ldquoPhospholipase A2myotoxins
from Bothrops snake venomsrdquo Toxicon vol 33 no 11 pp 1405ndash1424 1995
[28] J M Gutierrez and A Rucavado ldquoSnake venom metallopro-teinases their role in the pathogenesis of local tissue damagerdquoBiochimie vol 82 no 9-10 pp 841ndash850 2000
[29] T Matsui Y Fujimura and K Titani ldquoSnake venom proteasesaffecting hemostasis and thrombosisrdquo Biochimica et BiophysicaActa vol 1477 no 1-2 pp 146ndash156 2000
[30] A M Soares M R M Fontes and J R Giglio ldquoPhospholipaseA2myotoxins from Bothrops snake venoms structure-function
12 BioMed Research International
relationshiprdquoCurrent Organic Chemistry vol 8 no 17 pp 1677ndash1690 2004
[31] VM Rodrigues SMarcussi R S Cambraia et al ldquoBactericidaland neurotoxic activities of two myotoxic phospholipases A
2
from Bothrops neuwiedi pauloensis snake venomrdquo Toxicon vol44 no 3 pp 305ndash314 2004
[32] N A Santos-Filho L B Silveira C Z Oliveira et al ldquoAnew acidic myotoxic anti-platelet and prostaglandin I
2induc-
tor phospholipase A2isolated from Bothrops moojeni snake
venomrdquo Toxicon vol 52 no 8 pp 908ndash917 2008[33] A M Soares S H Andriao-Escarso Y Angulo et al ldquoStruc-
tural and functional characterization of myotoxin I a Lys49phospholipaseA
2homologue fromBothropsmoojeni (Caissaca)
snake venomrdquo Archives of Biochemistry and Biophysics vol 373no 1 pp 7ndash15 2000
[34] A M Soares S H Andriao-Escarso R K Bortoleto et alldquoDissociation of enzymatic and pharmacological properties ofpiratoxins-I and -III two myotoxic phospholipases A
2from
Bothrops pirajai snake venomrdquo Archives of Biochemistry andBiophysics vol 387 no 2 pp 188ndash196 2001
[35] S S Teixeira L B Silveira F M N da Silva et al ldquoMolecularcharacterization of an acidic phospholipase A
2from Bothrops
pirajai snake venom synthetic C-terminal peptide identifies itsantiplatelet regionrdquo Archives of Toxicology vol 85 no 10 pp1219ndash1233 2011
[36] S H Andriao-Escarso A M Soares V M Rodrigues et alldquoMyotoxic phospholipases A
2in Bothrops snake venoms effect
of chemical modifications on the enzymatic and pharmacolog-ical properties of bothropstoxins from Bothrops jararacussurdquoBiochimie vol 82 no 8 pp 755ndash763 2000
[37] S H Andriao-Escarso A M Soares M R M Fontes et alldquoStructural and functional characterization of an acidic plateletaggregation inhibitor and hypotensive phospholipase A
2from
Bothrops jararacussu snake venomrdquo Biochemical Pharmacologyvol 64 no 4 pp 723ndash732 2002
[38] M E G Denegri O C Acosta S Huancahuire-Vega et alldquoIsolation and functional characterization of a new acidic PLA
2
Ba SpII RP4 of the Bothrops alternatus snake venom fromArgentinardquo Toxicon vol 56 no 1 pp 64ndash74 2010
[39] R Zouari-Kessentini J Luis A Karray et al ldquoTwo purified andcharacterized phospholipases A
2from Cerastes cerastes venom
that inhibit cancerous cell adhesion and migrationrdquo Toxiconvol 53 no 4 pp 444ndash453 2009
[40] R Shirai M Toriba K Hayashi K Ikeda and S Inoue ldquoIden-tification and characterization of phospholipase A
2inhibitors
from the serum of the Japanese rat snake Elaphe climacophorardquoToxicon vol 53 no 6 pp 685ndash692 2009
[41] M I Homsi-Brandenburgo L S Queiroz H Santo-Neto LRodrigues-Simoni and J R Giglio ldquoFractionation of Bothropsjararacussu snake venom partial chemical characterization andbiological activity of bothropstoxinrdquo Toxicon vol 26 no 7 pp615ndash627 1988
[42] G P Fernandez A Segura M Herrera et al ldquoNeutralizationof Bothrops mattogrossensis snake venom from Bolivia exper-imental evaluation of llama and donkey antivenoms producedby caprylic acid precipitationrdquoToxicon vol 55 no 2-3 pp 642ndash645 2010
[43] C L Ownby H S S de Araujo S P White and J E FletcherldquoLysine 49 phospholipase A
2proteinsrdquo Toxicon vol 37 no 3
pp 411ndash445 1999[44] A M Soares W P Sestito S Marcussi et al ldquoAlkylation of
myotoxic phospholipases A2in Bothrops moojeni venom a
promising approach to an enhanced antivenom productionrdquoInternational Journal of Biochemistry amp Cell Biology vol 36 no2 pp 258ndash270 2004
[45] B Lomonte J M Gutierrez M F Furtado et al ldquoIsolationof basic myotoxins from Bothrops moojeni and Bothrops atroxsnake venomsrdquo Toxicon vol 28 no 10 pp 1137ndash1146 1990
[46] L C Mancuso M M Correa C A Vieira et al ldquoFractionationof Bothrops pirajai snake venom isolation and characterizationof piratoxin-I a new myotoxic proteinrdquo Toxicon vol 33 no 5pp 615ndash626 1995
[47] H S Selistre L S Queiroz O A B Cunha G E P deSouza and J R Giglio ldquoIsolation and characterization ofhemorrhagic myonecrotic and edema-inducing toxins fromBothrops insularis (jararaca ilhoa) snake venomrdquo Toxicon vol28 no 3 pp 261ndash273 1990
[48] E A Dennis J Cao Y-H Hsu V Magrioti and G KokotosldquoPhospholipaseA
2enzymes physical structure biological func-
tion disease implication chemical inhibition and therapeuticinterventionrdquo Chemical Reviews vol 111 no 10 pp 6130ndash61852011
[49] A G T Cardoso Acao do veneno de Bothrops moojeni e suafracao L-aminoacido oxidase submetida ao tratamento comraios gama de 60CO em Leishmania spp [MS thesis] IPEN-CNENSP Sao Paulo Brazil 1999
[50] MGallacci andWLGCavalcante ldquoUnderstanding the in vitroneuromuscular activity of snake venomLys49 phospholipaseA
2
homologuesrdquo Toxicon vol 55 no 1 pp 1ndash11 2010[51] S S Teixeira Caracterizacao funcional e estrutural de uma
fosfolipase A2acida isolada do veneno de Bothrops pirajai
[MS thesis] Faculdade de Ciencias Farmaceuticas de RibeiraoPretoUSP Ribeirao Preto Brazil 2009
[52] E Valentin and G Lambeau ldquoIncreasing molecular diversityof secreted phospholipases A
2and their receptors and binding
proteinsrdquo Biochimica et Biophysica Acta vol 1488 no 1-2 pp59ndash70 2000
[53] J M Gutierrez and B Lomonte ldquoPhospholipases A2from
Viperidae snake venoms how do they induce skeletal muscledamagerdquo Acta Chimica Slovenica vol 58 no 4 pp 647ndash6592011
[54] J M Gutierrez C L Ownby and G V Odell ldquoIsolation of amyotoxin from Bothrops asper venom partial characterizationand action on skeletal musclerdquo Toxicon vol 22 no 1 pp 115ndash128 1984
[55] N G Nascimento M C Sampaio R A Olivo and C TeixeiraldquoContribution of mast cells to the oedema induced by Bothropsmoojeni snake venom and a pharmacological assessment of theinflammatory mediators involvedrdquo Toxicon vol 55 no 2-3 pp343ndash352 2010
[56] C F P Teixeira E C T Landucci E Antunes M Chacurand Y Cury ldquoInflammatory effects of snake venom myotoxicphospholipases A
2rdquo Toxicon vol 42 no 8 pp 947ndash962 2003
[57] S R Zamuner and C F P Teixeira ldquoCell adhesion moleculesinvolved in the leukocyte recruitment induced by venom of thesnake Bothrops jararacardquoMediators of Inflammation vol 11 no6 pp 351ndash357 2002
[58] F Chaves C F P Teixeira and J M Gutierrez ldquoRole ofTNF-120572 IL-1120573 and IL-6 in the local tissue damage induced byBothrops asper snake venom an experimental assessment inmicerdquo Toxicon vol 45 no 2 pp 171ndash178 2005
BioMed Research International 13
[59] C D F P Teixeira C M Fernandes J P Zuliani and SF Zamuner ldquoInflammatory effects of snake venom metallo-proteinasesrdquo Memorias do Instituto Oswaldo Cruz vol 100supplement 1 pp 181ndash184 2005
[60] R D A Olivo C F P Teixeira J LWallace J M Gutierrez andS R Zamuner ldquoRole of cyclooxygenases in oedema-formingactivity of bothropic venomsrdquo Toxicon vol 49 no 5 pp 670ndash677 2007
[61] L B Silveira D P Marchi-Salvador N A Santos-Filho et alldquoIsolation and expression of a hypotensive and anti-plateletacidic phospholipase A
2from Bothrops moojeni snake venomrdquo
Journal of Pharmaceutical and Biomedical Analysis vol 73 pp35ndash43 2013
[62] A Rucavado T Escalante C F P Teixeira C M Fernandes CDıaz and J M Gutierrez ldquoIncrements in cytokines and matrixmetalloproteinases in skeletal muscle after injection of tissue-damaging toxins from the venom of the snake Bothrops asperrdquoMediators of Inflammation vol 11 no 2 pp 121ndash128 2002
[63] E Leiguez J P Zuliani A M Cianciarullo C M Fernandes JM Gutierrez and C Teixeira ldquoA group IIA-secreted phospho-lipase A
2from snake venom induces lipid body formation in
macrophages the roles of intracellular phospholipases A2and
distinct signaling pathwaysrdquo Journal of Leukocyte Biology vol90 no 1 pp 155ndash166 2011
[64] J P Zuliani J M Gutierrez L L Casais e Silva S C SampaioB Lomonte and C D F P Teixeira ldquoActivation of cellularfunctions in macrophages by venom secretory Asp-49 and Lys-49 phospholipasesA
2rdquoToxicon vol 46 no 5 pp 523ndash532 2005
[65] R D C M Escocard M M Kanashiro J H Petretski et alldquoNeutrophils regulate the expression of cytokines chemokinesand nitric oxide synthasenitric oxide in mice injected withBothrops atrox venomrdquo Immunobiology vol 211 no 1-2 pp 37ndash46 2006
[66] A Gambero E C T Landucci M H Toyama et al ldquoHumanneutrophil migration in vitro induced by secretory phospholi-pases A
2 a role for cell surface glycosaminoglycansrdquo Biochemi-
cal Pharmacology vol 63 no 1 pp 65ndash72 2002[67] F L S Costa R S Rodrigues L F M Izidoro et al ldquoBio-
chemical and functional properties of a thrombin-like enzymeisolated from Bothrops pauloensis snake venomrdquo Toxicon vol54 no 6 pp 725ndash735 2009
[68] J C Villalobos R Mora B Lomonte J M Gutierrez andY Angulo ldquoCytotoxicity induced in myotubes by a Lys49phospholipase A
2homologue from the venom of the snake
Bothrops asper evidence of rapid plasma membrane damageand a dual role for extracellular calciumrdquo Toxicology in Vitrovol 21 no 8 pp 1382ndash1389 2007
[69] B S Cummings J McHowat and R G Schnellmann ldquoPhos-pholipase A
2s in cell injury and deathrdquoThe Journal of Pharma-
cology and Experimental Therapeutics vol 294 no 3 pp 793ndash799 2000
[70] S R Panini L Yang A E Rusinol M S Sinensky J VBonventre and C C Leslie ldquoArachidonate metabolism andthe signaling pathway of induction of apoptosis by oxidizedLDLoxysterolrdquo Journal of Lipid Research vol 42 no 10 pp1678ndash1686 2001
[71] L Paramo B Lomonte J Pizarro-Cerda J-A Bengoechea J-P Gorvel and E Moreno ldquoBactericidal activity of Lys49 andAsp49 myotoxic phospholipases A
2from Bothrops asper snake
venom synthetic Lys49myotoxin II-(115-129)-peptide identifiesits bactericidal regionrdquo European Journal of Biochemistry vol253 no 2 pp 452ndash461 1998
[72] B Lomonte Y Angulo and E Moreno ldquoSynthetic peptidesderived from the C-terminal region of Lys49 phospholipaseA2homologues from viperidae snake venoms biomimetic
activities and potential applicationsrdquo Current PharmaceuticalDesign vol 16 no 28 pp 3224ndash3230 2010
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

BioMed Research International 5
per minute (n∘molsmin or U) for a given quantity of proteinin milligrams (mg)
The properties of snake venom components observed inthis study are characteristic of accidents caused by snakesof the Bothrops sp genus symptoms such as pain edemahemorrhage and necrosis and additionally systemic distur-bances are characteristic and corroborate the literature thatdescribes that proteases of snake venom proteins are closelyrelated to interference in the hemostatic system promotingblood coagulation fibrinolysis and platelet aggregation [27ndash30]
32 Isolation and Biochemical Characterization of Phospholi-pases The present study showed for the first time the isola-tion of the three phospholipases A
2from B mattogrossensis
snake venom obtained by two chromatographic steps Firstionic exchange chromatography was performed on a CM-Sepharose column with an ammonium bicarbonate gradient(50 to 500mM pH 80) The elution of absorbed proteinswith a linear gradient of concentrated buffer resulted inthirteen fractions (Figure 1(a)) of which fraction nine (9) wasrelated to PLA
2s because it showed phospholipase activity
of 22109Umg on artificial substratum All fractions werelyophilized and submitted to unidimensional electrophoresisrevealing many protein bands
Fraction 9 was submitted to the second chromatographicstep in a reverse column phase on a Discovery C18 columnusing 01 Trifluoroacetic (TFA) and 999 Acetonitrile(ACN) as solvents for the separation of other venom com-ponents The elution of absorbed proteins with a lineargradient of concentrated buffer resulted in twenty-two (22)fractions (Figure 1(b)) The PLA
2s were highly purified with
approximately 40 ACN Similar results were observed inthe isolation of other PLA
2s from snake venoms in high per-
formance liquid chromatography on reverse phase columnswhere the elution profile of these proteins occurs between 30and 40 ACN [5]
The association of chromatographic techniques suchas ionic exchange and reverse phase has commonly beenused and many snake venoms have been fractioned thisway highlighting the phospholipase purification of a speciesbelonging to the old complex ldquoBothrops neuwiedirdquo as well asthe target species studied in this research Two PLA
2basic
isoforms from B (neuwiedi) pauloensis venom were purifiedby Rodrigues et al [31] using biochemical techniques similarto the ones used in this study with the combination of ionicexchange (cationic) and reverse phase chromatographies
Other PLA2s have also been purified using simplified
procedures based in CM-Sepharose andor reverse phase asfor example venomPLA
2s fromBothropsmoojeni [21 32 33]
B pirajai [34 35] B jararacussu [36 37] B alternatus [8 38]Cerastes cerastes [39] and Elaphe climacophora [40]
Fractions 13 and 14 were related to enzymatically inactivePLA2s whereas fraction 15was related to enzymatically active
PLA2s The phospholipase activity of the collected fractions
were analyzed with synthetic NOB and NBD-PC substrates(Figures 2(a) and 2(b))The amount of activity was comparedto BthTX-II a basic enzymatically active PLA
2(Asp49) and
BthTX-I a basic enzymatically inactive PLA2(Lys49) both
20
15
10
5
0
100 200 300 400
80
60
40
20
0
B (
)
(mAU
)
1
23
4
5 67 8
9
10 11 12 13
MW 02 03 04 06 08 09 10 12 13
175805846
30
2317
07
times102
(min)
Btx
(a)
100
80
60
40
20
B (
)
MW175
805846
30
2317
07
(mAU
)
times102
6
5
4
3
2
1
0
0 10 20 30 40
1
2
3 45
6
7
8
910 11
12
13
1415
16
1718
19
20 2122
(min)
07 12 1314 15 16 17 18Btx
(b)
Figure 1 Chromatographic and electrophoretic profile of Bothropsmattogrossensis venom fractioning (a) CM-Sepharose columnsequilibrated with solvent A (50mM ammonium bicarbonatepH 80) and eluted with a 0ndash100 concentration gradient of solventB (500mM ammonium bicarbonate pH 80) at a 50mLminuteflow Emphasis on peak 9 rechromatographed (b) Rechromatog-raphy of fraction 9 on Discovery C18 column equilibrated withsolvent A (01 TFA) and eluted with a concentration gradient of0ndash100 of solvent B (999 acetonitrile and 01 trifluoroaceticacid) and a 10mLmin flow Emphasized in blue are fractions 1314 and 15 characterized as phospholipases BmatTX-I BmatTX-II and BmatTX-III respectively Controls MW molecular weightstandard BTx BthTX-I a basic enzymatically inactive PLA
2(Lys49)
(10120583g) isolated from Bothrops jararacussu venom Absorbancesread at 280 nm Electrophoresis gel made with 125 (ww)acrylamidebis-acrylamide
previously isolated from Bothrops jararacussu venom [36 41]The BmatTX-III PLA
2s (Asp49) were not able to induce
platelet aggregation and did not inhibit collagen or ADPinduced platelet aggregation (data not shown)
The degree of purity of the isolated proteins was furtherdemonstrated by SDS-PAGE mass spectrometry (Figure 3)and N-terminal sequencing (Figures 4(a) and 4(b)) Thepurified PLA
2s were named BmatTX-I BmatTX-II and
6 BioMed Research International
F12
Bmat
TX-I
Bmat
TX-I
I
Bmat
TX-I
II
F16
F17
F18
F19
F20
F21
F22
BthT
X-I
BthT
X-II
0
200
400
600
800
1000Ph
osph
olip
ase a
ctiv
ity (U
mg)
(a)
F12
Bmat
TX-I
II
F16
F17
F20
0
200
400
600
800
1000
Fluo
resc
ent i
nten
sity
(b)
Control BmatTX-I BmatTX-II BmatTX-III0
1000
2000
3000
BmatTX-I (Lys49)BmatTX-II (Lys49)BmatTX-III (Asp49)
Control
CK (U
L)
25120583g50120583L
lowastlowast
lowast
(c)
Figure 2 Enzymatic activity and myotoxic activity of PLA2s isolated from the venom of B mattogrossensis (a) Phospholipase activity of the
fractions collected from the rechromatography of fraction 9 done in C18 column assayed using an NOB stained substrate This activity wasassessed through the measurement of the number of moles of the released chromophore per minute (n∘molsmin or U) per milligram ofprotein (b) Phospholipase activity of the fractions collected from the rechromatography of fraction 9 done in C18 column assayed using afluorescent substrate (c) Myotoxic activity evaluated for inoculation of PLA
2s (25 120583g50 120583L) or PBS (control) done intramuscularly in the
gastrocnemius muscle of mice After 3 hours the creatine kinase (CK) level an important marker of muscular lesion was assayed in theanimalrsquos plasma Each bar represents the average +minus SD of three independent groups lowast119875 lt 005 compared to the control F13 BmatTX-I(Lys49) F14 BmatTX-II (Lys49) F15 BmatTX-III (Asp49) Controls BthTX-I (Lys49) and BthTX-II (Asp49)
BmatTX-III They were characterized as single polypeptidechains with isoelectric points around 10 (data not shown)This result agrees with data from published literature wheremost basic PLA
2s with or without catalytic activity on
artificial substrates [32 42] show an isoelectric point between8 and 10
The average molecular mass defined by mass spectrom-etry was 13304 Da for BmatTX-I 13623Da for BmatTX-IIand 13681Da for BmatTX-III (Figure 3)Thismolecularmassis consistent with most isolated PLA
2s from snake venoms
which are around 13 to 16 kDa [27 36 43]BmatTX-I sequencing showed a lysine (Lys) at position
49 and although only the first 28 amino acid residuesof BmatTX-II have been sequenced both are highly sim-ilar to the PLA
2Lys49 homologue subgroup The N-
terminal sequence alignment of BmatTX-I and BmatTX-IIhas revealed that these basic proteins are PLA
2s similar to
homologous Lys49 and other Lys49 myotoxins from snake
venoms (Figure 4(a)) BmatTX-I showed 94 similarity withMjTX-I present in B moojeni venom 92 with BthTX-I andBOJU-I present in B jararacussu venom and 94 similaritywith MTX-II present in B brazili venom
Additionallymultiple sequence alignment of BmatTX-IIIshowed a PLA
2-Asp49 basic myotoxin with another Asp49
of the Bothrops genera (Figure 4(b)) It can be observedthat BmatTX-III presented 78 similarity with BmjeTX-Iand 75 with BmoTX-I both from B moojeni venom 71similarity with BthTX-II isolated from B jararacussu venomand 65 similarity with PrTX-III from B pirajai venom
In the analysis of the sequences of the PLA2s isolated
from B mattogrossensis in this study highly preserved con-stituent residues of the 120572-helix structure characteristic ofphospholipases were identified as well as the presence ofmany cysteine residues which suggests the existence of manydisulfide bridges important for the stabilization of PLA
2rsquos
molecular structure [21 44]
BioMed Research International 7
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1360475
1398779
(a)
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1362326
1380673
(b)
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1256219
1368171
1402625
(c)
Figure 3 Mass determination by spectrometryThe average proteinmasses of BmatTX-I (a) BmatTX-II (b) and BmatTX-III (c) wereobtained in a MALDI-TOF system operated in linear mode usingexternal standards for calibration The resulting mass spectra weresubmitted to automatic baseline subtraction
Some studies regarding the amino acid composition ofPLA2s demonstrate that these proteins are rich in basic
and hydrophobic amino acids containing three long 120572-helixes two beta sheets and a Ca2+ binding site [45ndash47]Calcium is absolutely necessary for hydrolysis thereforealmost all PLA
2s have a highly preserved region for a Ca2+
bond (XCGXGG) and a catalytic site (DXCCXXHD) [48]It is observed in BmatTX-III that the calcium binding site(27YCGWGG
32) is preserved indicating that it is a catalyt-
ically active PLA2belonging to the PLA
2Asp49 subgroup
33 Biological Activities of Phospholipases A2 The main
objective for the isolation and characterization of compo-nents of snake venom is to better understand the partic-ipation of each component in different pathophysiologicalprocesses resulting from envenomation Local lesions can beattributed to proteases phospholipase activity and hemor-rhagic factors of these venoms followed by the release ofvasoactive agents causing hemorrhaging in various organsand tissues [5 44 49ndash52]
The PLA2s isolated from B mattogrossensis venom
BmatTX-I BmatTX-II and BmatTX-III showed highmyotoxic activity (Figure 2(c)) At a concentration of25 120583g50 120583L the PLA
2s induced a significant release of CK
an important muscular lesion marker when compared to thecontrol Myotoxicity is the characteristic presented by mostbasic phospholipases A
2from snake venoms Several studies
demonstrate that the myotoxic effect begins quickly by directaction of the myotoxic PLA
2s on the plasma membrane of
muscle cells or it is mediated by metalloproteases due toconsequent degeneration and ischemia [27 28 44]
Regarding the Lys49-PLA2myotoxins it is evident that
they lyse the plasma membrane of the muscle cell infected invivo however the exact mechanism has not been describedyet Furthermore it is not known if the toxin is internalizedbefore during or after the initial lysis or if it is not internal-ized Althoughmyotoxicity can be induced by the productionof fatty acids there is a second mechanism that seems to beindependent of the enzymatic activity and is mediated bythe C-terminal region at sites 115ndash129 of the Lys49 molecules[43 53 54]
In an attempt to better understand the development ofthe inflammatory process unleashed by the protein complexpresent in snake venoms many studies have been donesuch as edema induction [55] leukocyte participation [5657] mast cells degranulation [55] participation of variouscytotoxins in the inflammatory system [58 59] participationof cyclooxygenases [60 61] and the participation of venomPLA2s in the inflammatory process [6 51 56 62]
Many studies about PLA2activity in macrophages have
already been done [8 63 64] Little is known about PLA2rsquos
effect on neutrophils however Escocard et al [65] describedan influx of inflammatory cells including many neutrophilsinto the peritoneal cavity of mice after the injection ofBothrops atrox venom The induction of reactive oxygenspecies (ROS) cytokines like IL-6 and IL-1120573 was seen inthese neutrophils These data were also observed in a studycarried out by Souza et al [26] where besides the influx ofneutrophils into the peritoneal cavity ofmice after injection ofthe venom of B atrox there was also induction of superoxideby these cells mast cell degranulation and phagocytosisby macrophages Regarding the activity of PLA
2Gambero
et al [66] have observed the ability of some myotoxins(bothropstoxin-I-II and piratoxin-I) to induce neutrophilchemotaxis in a concentration-dependent manner
In order to evaluate the activation of leukocytes thetoxicity of B mattogrossensis myotoxins on neutrophil cellswas investigated The cells were incubated with differentconcentrations (3 6 and 12 120583gmL) of BmatTX-I (Lys49)BmatTX-II (Lys49) and BmatTX-III (Asp49) myotoxinsduring 1 12 and 24 hours (data not shown)These myotoxinsdid not affect the neutrophilsrsquo viability which agrees withZuliani et al [64] showing low toxicity on thioglycollateelicited mice macrophages
Nonetheless our data showed that neutrophils incubatedwith BmatTX-I and BmatTX-II myotoxins induced therelease of IL-1120573 (Figures 5(a) and 5(b)) Moreover theseresults suggest that phospholipid hydrolysis is not essentialfor the activity observed and argue with the hyphotesis that
8 BioMed Research International
lowastlowast lowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowast lowastlowastlowastlowastlowastlowastlowastlowastlowast
1 10 20 30 40 50(Identity )
100
89
92
92
92
94
94
BmatTX-IBmatTX-IIPrtx-IBOJU-IBthTX-IMjtx-IMtx-Ii
(a)
lowastlowastlowast lowastlowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowastlowast lowastlowastlowast
1 10 20 30 40 50
(Identity )100
78
75
71
65
BmatTX-IIIBmoTX-IBmjeTX-IBthTX-IIPrTX-III
(b)
Figure 4 Comparison of the N-terminal sequence of the snakersquos PLA2s with the PLA
2s isolated from B mattogrossensis venom N-terminal
sequencing of the PLA2s obtained by Edman [13] degradation and alignment done with CLUSTALW2 expressed as of similarity (a)
BmatTX-I BmatTX-II with Lys49 residues in marked sequences (b) BmatTX-III with Asp49 residues in marked sequences Sequencesused for the alignment and their respective access numbers BthTX-I (gi51890398) MjTX-I (gi17368325) Mtx-II (gi390981003) PrTX-I (gi190016174) BOJU-I (gi209572966) BthTX-II (gi1171971) PrTX-III (gi90016174) BmjeTX-I (gi313471399) BmoTX-I (gi221272396)BthTX-I (Lys49) BthTX-II (Asp49) and BOJU-I (Lys49) isolated from the venom of B jararacussu MjTX-I (Lys49) BmoTX-I (Asp49)and BmjeTX-I (Asp49) isolated from the venom of B moojeni Mtx-II (Lys49) isolated from the venom of B brazili PrTX-I (Lys49) andPrTX-III (Asp49) isolated from the venom of B pirajai the ldquolowastrdquo was used to indicate the amino acid residues that are the same between thesequences
other molecular regions distinct from the active site may beinvolved in this effect
PLA2s are multifunctional proteins that can be used as
mediators in several areas of medicine such as in the treat-ment of rheumatoid arthritis as a new class of HIV inhibitorsby blocking the host cell invasion as a potential treatmentagainst malaria and as an antibiotic by inducing cytotoxicityvia the disruption of bacterial membranes [21 23 30 6768] A study by Costa et al [67] with PLA
2s isolated from
B brazili MTX-I and II demonstrated cytotoxic activityagainst Jurkat tumor cells as well as antimicrobial activityagainstE coli andC albicans and antiparasitic activity againstLeishmania sp
In the present study we evaluated the cytotoxic activityof PLA
2s isolated from B mattogrossensis on JURKAT (T
leukemia) and SK-BR-3 (breast adenocarcinoma) both ofwhich are human tumor cell lines Like MTX-II of B brazili[67] the cytotoxic activity of PLA
2s BmatTX-I and BmatTX-
II on JURKATcells was independent of their catalytic activitysince these are characterized as Lys49 PLA
2s and are therefore
catalytically inactive BmatTX-III characterized as Asp49despite being enzymatically active also showed a lower level oftoxicity Some authors propose that the cytotoxic activity ontumor cell lines is associated with the induction of apoptosisconsidering the fact that PLA
2promotes alterations in the
cell membrane And some studies involving Lys49 PLA2s
isolated from B asper demonstrated that the C-terminal
region comprised of amino acids 115ndash129 is concerned withthe cytotoxic and bactericidal activities of this protein [69ndash71] The same observation was made by Costa et al [67] withsynthetic peptides derived from the C-terminal portion ofMTX-I and II PLA
2s
We also evaluated the antiparasitic activity of the crudevenom and isolated PLA
2of B mattogrossensis on promastig-
ote forms of L amazonensis After the analysis it wasobserved that the crude venom of B mattogrossensis presentsincreasing toxic activity (approximately 50 to 80) againstpromastigote forms of L amazonensis after 48 h of incubation(Figure 7(a)) The PLA
2s isolated from B mattogrossensis at
100 120583gmL characterized as BmatTX-I (Lys49) and BmatTX-III (Asp49) presented toxic activity between 25 and 30respectively even with values close to those presented afterincubation of the protozoan with Pentamidine a drug usedas a positive control (Figures 7(b) and 7(c))
When compared the cytotoxicity values of PLA2s against
promastigote forms of L amazonensis show similar activitybetween the Lys49-PLA
2and the Asp49-PLA
2 Comparison
of this activity with the crude venom (70) showed thatPLA2s are responsible for almost half of the observed effect
Nonetheless notably the results suggest that other toxinspresent in the venom contribute to the parasitersquos death
Similar to the results obtained in the present studyStabeli et al [21] demonstrated that MjTX-II a Lys49-PLA
2
homologue isolated from B moojeni venom in different
BioMed Research International 9
0
500
1000
1500
2000
lowast
lowast
lowast
lowast
lowast
Control 3 6 12 3 6 12 3 6 12
ControlBmatTX-I (Lys49)
BmatTX-II (Lys49)BmatTX-III (Asp49)
BmatTX-I (120583g) BmatTX-II (120583g) BmatTX-III (120583g)
IL-1120573
(pg
mL)
(a)
Control 3 6 12 3 6 12 3 6 120
500
1000
1500
2000
ControlBmatTX-I (Lys49)
BmatTX-II (Lys49)BmatTX-III (Asp49)
lowastlowastlowast
BmatTX-I (120583g) BmatTX-II (120583g) BmatTX-III (120583g)
IL-1120573
(pg
mL)
(b)
Figure 5 Production of IL-1120573 by mice neutrophils These cells wereincubatedwith PLA
2(3 6 and 12 120583gmL) or RPMI (control) for 12
hours (a) and 24 hours (b) at 37∘C in a humid atmosphere with 5CO2 Quantification was done by ELISA as described in 29 Each
bar represents the average +minus SDof three independent groups lowast119875 lt005 compared to the control
concentrations (5 25 and 75 120583g) was effective as a parasiticideagent against Schistosomamansoni and promastigote forms ofLeishmania (L amazonensis L braziliensis L donovani andL major)
The action of the PLA2s BmatTX-I (Lys49) and
BmatTX-III (Asp49) on promastigote forms of L ama-zonensis was independent of its catalytic activity sincecatalytically inactive Lys49 myotoxin also demonstrated tox-icity against Leishmania It is believed that the observedcytotoxic activity might be related to the C-terminal regionsof these phospholipase-homologues that are able to promotea disturbance in the cellular membranes independent oftheir catalytic activity [67 72] However more studies arenecessary to define the exact mechanism of action of theseenzymes on parasites
Growing interest in the comprehension of the structureand function of snake venom components especially PLA
2
10 100 10 100 10 100 10 1000
20
40
60
80
100
BmatTX-I (Lys49)BmatTX-II (Lys49)
BmatTX-III (Asp49)Methotrexate (MTX)
Cyto
toxi
city
()
Concentration (120583gmL)
(a)
10 100 10 100 10 100 10 1000
20
40
60
80
100
BmatTX-I (Lys49)BmatTX-II (Lys49)
BmatTX-III (Asp49)Methotrexate (MTX)
Cyto
toxi
city
()
Concentration (120583gmL)
(b)
Figure 6 Antitumoral activity of PLA2s from B mattogrossensis
(a) Antitumoral activity on human acute T-cell leukemia (JURKAT)lines (b) Antitumoral activity on human breast adenocarcinoma(SK-BR-3) Different concentrations of the PLA
2s were incubated
with cell linesMethotrexate was used as the positive control Resultsare presented as mean +minus SD (119899 = 3)
contributes to a better understanding of the mechanism ofaction of their enzymatic and toxic activities It opens thepath to better understand the intoxication caused by enven-omation and the physiopathology behind its side effectsFuture studies will potentially improve serum therapy andhelp develop the pharmaceutical potential that moleculesisolated from animal venoms can have such as the PLA
2s
isolated from the B mattogrossensis venom which show anti-Leishmania and antitumor activities
4 Conclusion
In conclusion the venom of Bothrops mattogrossensis hasa qualitatively similar toxicological profile to previously
10 BioMed Research International
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineB mattogrossensis crude venom
Ant
i-leis
hman
ia ac
tivity
()
B mattogrossensis crude venom (120583g)
(a)
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineBmatTX-I (Lys49)
Ant
i-leis
hman
ia ac
tivity
()
BmatTX-I (120583g)
lowastlowast lowast lowast
(b)
Ant
i-leis
hman
ia ac
tivity
()
Pent 36 625 125 25 50 1000
20
40
60
80
100
BmatTX-III (Asp49)Pentamidine
BmatTX-III (120583g)
lowastlowastlowast
lowastlowast lowast
(c)
Figure 7 Antileishmanial activity of the crude venom of and PLA2s from B mattogrossensis The promastigote forms of L amazonensiswere
plated with 1times105 cellswellThen different concentrations of crude venom and isolated PLA2s were added to each wellThe experiment was
done in a 48 h period with the (a) crude venom and the PLA2enzymes (b) BmatTX-I and (c) BmatTX-III MTT was added and after the
incubation period at 33∘C the formazan crystal formed was dissolved in SDS Readings were done in a spectrophotometer at 570 nm Eachbar represents the average +minus SD of the three independent experimental groups sixfold total lowast119875 lt 005 compared to the control
studied snake venoms of the Bothrops sp genera despite theobservation of quantitative variations Of the three basicPLA2s from B mattogrossensis venom now isolated for the
first time two are characterized as Lys49-PLA2homologues
BmatTX-I and -II and the other as an Asp49-PLA2 named
BmatTX-III This showed high phospholipase activity ThePLA2s isolated induced myotoxic effects as well as the release
of proinflammatory cytokines by neutrophils BmatTX-I and-III PLA
2s were cytotoxic to human tumor cell lines JURKAT
and SK-BR-3 and showed activity against promastigote formsof L amazonensis
Conflict of Interests
The authors state that there is no conflict of interests
Acknowledgments
The authors are grateful to the Ministry of Science andTechnology (MCTI) Conselho Nacional de Desenvolvi-mento Cientıfico e Tecnologico (CNPq) Financiadora deEstudos e Projetos (FINEP) Fundacao de Tecnologia doAcre (FUNTACFDCT) Coordenacao de Aperfeicoamentode Nıvel Superior (CAPES)mdashProjeto NanoBiotec Redede Biodiversidade e Biotecnologia da Amazonia Legal(BIONORTECNPqMCTI) Instituto Nacional paraPesquisa Translacional em Saude e Ambiente na RegiaoAmazonica (INCT-INPeTAmCNPqMCTI) and InstitutoNacional para Pesquisa em Toxinas (INCT-Tox) andSecretary of Development of Rondonia State (SEPLANPRONEXCNPq) for financial support We are grateful to
BioMed Research International 11
Sr Guilherme G Galassi (Mantenedouro da Fauna Silvestreautorizacao IBAMA 020270089912001-95 Americana-SP)for their helpful technical collaboration in provide the snakevenom Amy Grabner provided the English editing of thepaper
References
[1] Y Angulo and B Lomonte ldquoBiochemistry and toxicology oftoxins purified from the venom of the snake Bothrops asperrdquoToxicon vol 54 no 7 pp 949ndash957 2009
[2] J J Calvete P Juarez and L Sanz ldquoSnake venomics Strategyand applicationsrdquo Journal of Mass Spectrometry vol 42 no 11pp 1405ndash1414 2007
[3] T S Kang D Georgieva N Genov et al ldquoEnzymatic toxinsfrom snake venom structural characterization and mechanismof catalysisrdquoThe FEBS Journal vol 278 no 23 pp 4544ndash45762011
[4] C ZOliveiraCaracterizacao funcional e estrutural de inibidoresde fosfolipases A
2isolados do plasma de serpente Bothrops
jararacussu [PhD thesis] Faculdade de Ciencias Farmaceuticasde Ribeirao Preto Ribeirao Preto Brazil 2009
[5] R G Stabeli R Simoes-Silva A M Kayano et al ldquoPurifi-cation of phospholipases A
2from American snake venomsrdquo
in ChromatographymdashThe Most Versatile Method of ChemicalAnalysis chapter 1 pp 1ndash34 InTech Vienna Austria 2012
[6] J P Zuliani C M Fernandes S R Zamuner J M Gutierrezand C F P Teixeira ldquoInflammatory events induced by Lys-49 and Asp-49 phospholipases A
2isolated from Bothrops asper
snake venom role of catalytic activityrdquo Toxicon vol 45 no 3pp 335ndash346 2005
[7] S Marcussi R G Stabeli N A Santos-Filho et al ldquoGenotoxiceffect of Bothrops snake venoms and isolated toxins on humanlymphocyte DNArdquo Toxicon vol 65 pp 9ndash14 2013
[8] S S Setubal A S Pontes J L Furtado et al ldquoAction oftwo phospholipases A
2purified from Bothrops alternatus snake
venom on macrophagesrdquo Biochemistry vol 78 no 2 pp 194ndash203 2013
[9] J M Gutierrez L A Ponce-Soto S Marangoni and BLomonte ldquoSystemic and local myotoxicity induced by snakevenom group II phospholipases A
2 comparison between cro-
toxin crotoxin B and a Lys49 PLA2homologuerdquo Toxicon vol
51 no 1 pp 80ndash92 2008[10] A Q Rueda I G Rodrıguez E C Ara et al ldquoBiochemical
characterization action onmacrophages and superoxide anionproduction of four basic phospholipases A
2from Panamanian
Bothrops asper snake venomrdquo BioMed Research Internationalvol 2013 Article ID 789689 9 pages 2013
[11] V X da Silva and M T Rodrigues ldquoTaxonomic revision ofthe Bothrops neuwiedi complex (Serpentes Viperidae) withdescription of a new speciesrdquo Phyllomedusa vol 7 no 1 pp 45ndash90 2008
[12] J A Campbell and W W Lamar The Venomous Reptiles of theWestern Hemisphere Comstock Ithaca NY USA 2004
[13] P Edman ldquoMethod for determination of the amino acidsequence in peptidesrdquo Acta Chemica Scandinavica vol 4 pp283ndash293 1950
[14] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[15] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[16] M Holzer and S P Mackessy ldquoAn aqueous endpoint assay ofsnake venom phospholipase A
2rdquo Toxicon vol 34 no 10 pp
1149ndash1155 1996[17] A L Fuly A L P deMiranda R B Zingali and J AGuimaraes
ldquoPurification and characterization of a phospholipase A2isoen-
zyme isolated from Lachesis muta snake venomrdquo BiochemicalPharmacology vol 63 no 9 pp 1589ndash1597 2002
[18] T Nikai N Mori M Kishida H Sugihara and A T Tu ldquoIso-lation and biochemical characterization of hemorrhagic toxinf from the venom of Crotalus atrox (Western DiamondbackRattlesnake)rdquo Archives of Biochemistry and Biophysics vol 231no 2 pp 309ndash319 1984
[19] J A Gene A Roy G Rojas J M Gutierrez and L CerdasldquoComparative study on coagulant defibrinating fibrinolyticand fibrinogenolytic activities of Costa Rican crotaline snakevenoms and their neutralization by a polyvalent antivenomrdquoToxicon vol 27 no 8 pp 841ndash848 1989
[20] M S Charney and R M Tomarelli ldquoA colorimetric method forthe determination of the proteolytic activity of duodenal juicerdquoThe Journal of Biological Chemistry vol 171 pp 501ndash505 1947
[21] R G Stabeli S F Amui C D SantrsquoAna et al ldquoBothrops moojenimyotoxin-II a Lys49-phospholipaseA
2homologue an example
of function versatility of snake venom proteinsrdquo ComparativeBiochemistry and Physiology C vol 142 no 3-4 pp 371ndash3812006
[22] D R Call J A Nemzek S J Ebong G L Bolgos D ENewcomb and D G Remick ldquoRatio of local to systemicchemokine concentrations regulates neutrophil recruitmentrdquoThe American Journal of Pathology vol 158 no 2 pp 715ndash7212001
[23] P G Roberto S Kashima S Marcussi et al ldquoCloning andidentification of a complete cDNA coding for a bactericidal andantitumoral acidic phospholipase A
2from Bothrops jararacussu
venomrdquoThe Protein Journal vol 23 no 4 pp 273ndash285 2004[24] J H Schumacher A OrsquoGarra B Shrader et al ldquoThe charac-
terization of four monoclonal antibodies specific for mouse IL-5 and development of mouse and human IL-5 enzyme-linkedimmunosorbentrdquoThe Journal of Immunology vol 141 no 5 pp1576ndash1581 1988
[25] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[26] C A T Souza A M Kayano S S Setubal et al ldquoLocal andsystemic biochemical alterations induced by Bothrops atroxsnake venom in micerdquo Journal of Venom Research vol 3 pp28ndash34 2012
[27] J M Gutierrez and B Lomonte ldquoPhospholipase A2myotoxins
from Bothrops snake venomsrdquo Toxicon vol 33 no 11 pp 1405ndash1424 1995
[28] J M Gutierrez and A Rucavado ldquoSnake venom metallopro-teinases their role in the pathogenesis of local tissue damagerdquoBiochimie vol 82 no 9-10 pp 841ndash850 2000
[29] T Matsui Y Fujimura and K Titani ldquoSnake venom proteasesaffecting hemostasis and thrombosisrdquo Biochimica et BiophysicaActa vol 1477 no 1-2 pp 146ndash156 2000
[30] A M Soares M R M Fontes and J R Giglio ldquoPhospholipaseA2myotoxins from Bothrops snake venoms structure-function
12 BioMed Research International
relationshiprdquoCurrent Organic Chemistry vol 8 no 17 pp 1677ndash1690 2004
[31] VM Rodrigues SMarcussi R S Cambraia et al ldquoBactericidaland neurotoxic activities of two myotoxic phospholipases A
2
from Bothrops neuwiedi pauloensis snake venomrdquo Toxicon vol44 no 3 pp 305ndash314 2004
[32] N A Santos-Filho L B Silveira C Z Oliveira et al ldquoAnew acidic myotoxic anti-platelet and prostaglandin I
2induc-
tor phospholipase A2isolated from Bothrops moojeni snake
venomrdquo Toxicon vol 52 no 8 pp 908ndash917 2008[33] A M Soares S H Andriao-Escarso Y Angulo et al ldquoStruc-
tural and functional characterization of myotoxin I a Lys49phospholipaseA
2homologue fromBothropsmoojeni (Caissaca)
snake venomrdquo Archives of Biochemistry and Biophysics vol 373no 1 pp 7ndash15 2000
[34] A M Soares S H Andriao-Escarso R K Bortoleto et alldquoDissociation of enzymatic and pharmacological properties ofpiratoxins-I and -III two myotoxic phospholipases A
2from
Bothrops pirajai snake venomrdquo Archives of Biochemistry andBiophysics vol 387 no 2 pp 188ndash196 2001
[35] S S Teixeira L B Silveira F M N da Silva et al ldquoMolecularcharacterization of an acidic phospholipase A
2from Bothrops
pirajai snake venom synthetic C-terminal peptide identifies itsantiplatelet regionrdquo Archives of Toxicology vol 85 no 10 pp1219ndash1233 2011
[36] S H Andriao-Escarso A M Soares V M Rodrigues et alldquoMyotoxic phospholipases A
2in Bothrops snake venoms effect
of chemical modifications on the enzymatic and pharmacolog-ical properties of bothropstoxins from Bothrops jararacussurdquoBiochimie vol 82 no 8 pp 755ndash763 2000
[37] S H Andriao-Escarso A M Soares M R M Fontes et alldquoStructural and functional characterization of an acidic plateletaggregation inhibitor and hypotensive phospholipase A
2from
Bothrops jararacussu snake venomrdquo Biochemical Pharmacologyvol 64 no 4 pp 723ndash732 2002
[38] M E G Denegri O C Acosta S Huancahuire-Vega et alldquoIsolation and functional characterization of a new acidic PLA
2
Ba SpII RP4 of the Bothrops alternatus snake venom fromArgentinardquo Toxicon vol 56 no 1 pp 64ndash74 2010
[39] R Zouari-Kessentini J Luis A Karray et al ldquoTwo purified andcharacterized phospholipases A
2from Cerastes cerastes venom
that inhibit cancerous cell adhesion and migrationrdquo Toxiconvol 53 no 4 pp 444ndash453 2009
[40] R Shirai M Toriba K Hayashi K Ikeda and S Inoue ldquoIden-tification and characterization of phospholipase A
2inhibitors
from the serum of the Japanese rat snake Elaphe climacophorardquoToxicon vol 53 no 6 pp 685ndash692 2009
[41] M I Homsi-Brandenburgo L S Queiroz H Santo-Neto LRodrigues-Simoni and J R Giglio ldquoFractionation of Bothropsjararacussu snake venom partial chemical characterization andbiological activity of bothropstoxinrdquo Toxicon vol 26 no 7 pp615ndash627 1988
[42] G P Fernandez A Segura M Herrera et al ldquoNeutralizationof Bothrops mattogrossensis snake venom from Bolivia exper-imental evaluation of llama and donkey antivenoms producedby caprylic acid precipitationrdquoToxicon vol 55 no 2-3 pp 642ndash645 2010
[43] C L Ownby H S S de Araujo S P White and J E FletcherldquoLysine 49 phospholipase A
2proteinsrdquo Toxicon vol 37 no 3
pp 411ndash445 1999[44] A M Soares W P Sestito S Marcussi et al ldquoAlkylation of
myotoxic phospholipases A2in Bothrops moojeni venom a
promising approach to an enhanced antivenom productionrdquoInternational Journal of Biochemistry amp Cell Biology vol 36 no2 pp 258ndash270 2004
[45] B Lomonte J M Gutierrez M F Furtado et al ldquoIsolationof basic myotoxins from Bothrops moojeni and Bothrops atroxsnake venomsrdquo Toxicon vol 28 no 10 pp 1137ndash1146 1990
[46] L C Mancuso M M Correa C A Vieira et al ldquoFractionationof Bothrops pirajai snake venom isolation and characterizationof piratoxin-I a new myotoxic proteinrdquo Toxicon vol 33 no 5pp 615ndash626 1995
[47] H S Selistre L S Queiroz O A B Cunha G E P deSouza and J R Giglio ldquoIsolation and characterization ofhemorrhagic myonecrotic and edema-inducing toxins fromBothrops insularis (jararaca ilhoa) snake venomrdquo Toxicon vol28 no 3 pp 261ndash273 1990
[48] E A Dennis J Cao Y-H Hsu V Magrioti and G KokotosldquoPhospholipaseA
2enzymes physical structure biological func-
tion disease implication chemical inhibition and therapeuticinterventionrdquo Chemical Reviews vol 111 no 10 pp 6130ndash61852011
[49] A G T Cardoso Acao do veneno de Bothrops moojeni e suafracao L-aminoacido oxidase submetida ao tratamento comraios gama de 60CO em Leishmania spp [MS thesis] IPEN-CNENSP Sao Paulo Brazil 1999
[50] MGallacci andWLGCavalcante ldquoUnderstanding the in vitroneuromuscular activity of snake venomLys49 phospholipaseA
2
homologuesrdquo Toxicon vol 55 no 1 pp 1ndash11 2010[51] S S Teixeira Caracterizacao funcional e estrutural de uma
fosfolipase A2acida isolada do veneno de Bothrops pirajai
[MS thesis] Faculdade de Ciencias Farmaceuticas de RibeiraoPretoUSP Ribeirao Preto Brazil 2009
[52] E Valentin and G Lambeau ldquoIncreasing molecular diversityof secreted phospholipases A
2and their receptors and binding
proteinsrdquo Biochimica et Biophysica Acta vol 1488 no 1-2 pp59ndash70 2000
[53] J M Gutierrez and B Lomonte ldquoPhospholipases A2from
Viperidae snake venoms how do they induce skeletal muscledamagerdquo Acta Chimica Slovenica vol 58 no 4 pp 647ndash6592011
[54] J M Gutierrez C L Ownby and G V Odell ldquoIsolation of amyotoxin from Bothrops asper venom partial characterizationand action on skeletal musclerdquo Toxicon vol 22 no 1 pp 115ndash128 1984
[55] N G Nascimento M C Sampaio R A Olivo and C TeixeiraldquoContribution of mast cells to the oedema induced by Bothropsmoojeni snake venom and a pharmacological assessment of theinflammatory mediators involvedrdquo Toxicon vol 55 no 2-3 pp343ndash352 2010
[56] C F P Teixeira E C T Landucci E Antunes M Chacurand Y Cury ldquoInflammatory effects of snake venom myotoxicphospholipases A
2rdquo Toxicon vol 42 no 8 pp 947ndash962 2003
[57] S R Zamuner and C F P Teixeira ldquoCell adhesion moleculesinvolved in the leukocyte recruitment induced by venom of thesnake Bothrops jararacardquoMediators of Inflammation vol 11 no6 pp 351ndash357 2002
[58] F Chaves C F P Teixeira and J M Gutierrez ldquoRole ofTNF-120572 IL-1120573 and IL-6 in the local tissue damage induced byBothrops asper snake venom an experimental assessment inmicerdquo Toxicon vol 45 no 2 pp 171ndash178 2005
BioMed Research International 13
[59] C D F P Teixeira C M Fernandes J P Zuliani and SF Zamuner ldquoInflammatory effects of snake venom metallo-proteinasesrdquo Memorias do Instituto Oswaldo Cruz vol 100supplement 1 pp 181ndash184 2005
[60] R D A Olivo C F P Teixeira J LWallace J M Gutierrez andS R Zamuner ldquoRole of cyclooxygenases in oedema-formingactivity of bothropic venomsrdquo Toxicon vol 49 no 5 pp 670ndash677 2007
[61] L B Silveira D P Marchi-Salvador N A Santos-Filho et alldquoIsolation and expression of a hypotensive and anti-plateletacidic phospholipase A
2from Bothrops moojeni snake venomrdquo
Journal of Pharmaceutical and Biomedical Analysis vol 73 pp35ndash43 2013
[62] A Rucavado T Escalante C F P Teixeira C M Fernandes CDıaz and J M Gutierrez ldquoIncrements in cytokines and matrixmetalloproteinases in skeletal muscle after injection of tissue-damaging toxins from the venom of the snake Bothrops asperrdquoMediators of Inflammation vol 11 no 2 pp 121ndash128 2002
[63] E Leiguez J P Zuliani A M Cianciarullo C M Fernandes JM Gutierrez and C Teixeira ldquoA group IIA-secreted phospho-lipase A
2from snake venom induces lipid body formation in
macrophages the roles of intracellular phospholipases A2and
distinct signaling pathwaysrdquo Journal of Leukocyte Biology vol90 no 1 pp 155ndash166 2011
[64] J P Zuliani J M Gutierrez L L Casais e Silva S C SampaioB Lomonte and C D F P Teixeira ldquoActivation of cellularfunctions in macrophages by venom secretory Asp-49 and Lys-49 phospholipasesA
2rdquoToxicon vol 46 no 5 pp 523ndash532 2005
[65] R D C M Escocard M M Kanashiro J H Petretski et alldquoNeutrophils regulate the expression of cytokines chemokinesand nitric oxide synthasenitric oxide in mice injected withBothrops atrox venomrdquo Immunobiology vol 211 no 1-2 pp 37ndash46 2006
[66] A Gambero E C T Landucci M H Toyama et al ldquoHumanneutrophil migration in vitro induced by secretory phospholi-pases A
2 a role for cell surface glycosaminoglycansrdquo Biochemi-
cal Pharmacology vol 63 no 1 pp 65ndash72 2002[67] F L S Costa R S Rodrigues L F M Izidoro et al ldquoBio-
chemical and functional properties of a thrombin-like enzymeisolated from Bothrops pauloensis snake venomrdquo Toxicon vol54 no 6 pp 725ndash735 2009
[68] J C Villalobos R Mora B Lomonte J M Gutierrez andY Angulo ldquoCytotoxicity induced in myotubes by a Lys49phospholipase A
2homologue from the venom of the snake
Bothrops asper evidence of rapid plasma membrane damageand a dual role for extracellular calciumrdquo Toxicology in Vitrovol 21 no 8 pp 1382ndash1389 2007
[69] B S Cummings J McHowat and R G Schnellmann ldquoPhos-pholipase A
2s in cell injury and deathrdquoThe Journal of Pharma-
cology and Experimental Therapeutics vol 294 no 3 pp 793ndash799 2000
[70] S R Panini L Yang A E Rusinol M S Sinensky J VBonventre and C C Leslie ldquoArachidonate metabolism andthe signaling pathway of induction of apoptosis by oxidizedLDLoxysterolrdquo Journal of Lipid Research vol 42 no 10 pp1678ndash1686 2001
[71] L Paramo B Lomonte J Pizarro-Cerda J-A Bengoechea J-P Gorvel and E Moreno ldquoBactericidal activity of Lys49 andAsp49 myotoxic phospholipases A
2from Bothrops asper snake
venom synthetic Lys49myotoxin II-(115-129)-peptide identifiesits bactericidal regionrdquo European Journal of Biochemistry vol253 no 2 pp 452ndash461 1998
[72] B Lomonte Y Angulo and E Moreno ldquoSynthetic peptidesderived from the C-terminal region of Lys49 phospholipaseA2homologues from viperidae snake venoms biomimetic
activities and potential applicationsrdquo Current PharmaceuticalDesign vol 16 no 28 pp 3224ndash3230 2010
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

6 BioMed Research International
F12
Bmat
TX-I
Bmat
TX-I
I
Bmat
TX-I
II
F16
F17
F18
F19
F20
F21
F22
BthT
X-I
BthT
X-II
0
200
400
600
800
1000Ph
osph
olip
ase a
ctiv
ity (U
mg)
(a)
F12
Bmat
TX-I
II
F16
F17
F20
0
200
400
600
800
1000
Fluo
resc
ent i
nten
sity
(b)
Control BmatTX-I BmatTX-II BmatTX-III0
1000
2000
3000
BmatTX-I (Lys49)BmatTX-II (Lys49)BmatTX-III (Asp49)
Control
CK (U
L)
25120583g50120583L
lowastlowast
lowast
(c)
Figure 2 Enzymatic activity and myotoxic activity of PLA2s isolated from the venom of B mattogrossensis (a) Phospholipase activity of the
fractions collected from the rechromatography of fraction 9 done in C18 column assayed using an NOB stained substrate This activity wasassessed through the measurement of the number of moles of the released chromophore per minute (n∘molsmin or U) per milligram ofprotein (b) Phospholipase activity of the fractions collected from the rechromatography of fraction 9 done in C18 column assayed using afluorescent substrate (c) Myotoxic activity evaluated for inoculation of PLA
2s (25 120583g50 120583L) or PBS (control) done intramuscularly in the
gastrocnemius muscle of mice After 3 hours the creatine kinase (CK) level an important marker of muscular lesion was assayed in theanimalrsquos plasma Each bar represents the average +minus SD of three independent groups lowast119875 lt 005 compared to the control F13 BmatTX-I(Lys49) F14 BmatTX-II (Lys49) F15 BmatTX-III (Asp49) Controls BthTX-I (Lys49) and BthTX-II (Asp49)
BmatTX-III They were characterized as single polypeptidechains with isoelectric points around 10 (data not shown)This result agrees with data from published literature wheremost basic PLA
2s with or without catalytic activity on
artificial substrates [32 42] show an isoelectric point between8 and 10
The average molecular mass defined by mass spectrom-etry was 13304 Da for BmatTX-I 13623Da for BmatTX-IIand 13681Da for BmatTX-III (Figure 3)Thismolecularmassis consistent with most isolated PLA
2s from snake venoms
which are around 13 to 16 kDa [27 36 43]BmatTX-I sequencing showed a lysine (Lys) at position
49 and although only the first 28 amino acid residuesof BmatTX-II have been sequenced both are highly sim-ilar to the PLA
2Lys49 homologue subgroup The N-
terminal sequence alignment of BmatTX-I and BmatTX-IIhas revealed that these basic proteins are PLA
2s similar to
homologous Lys49 and other Lys49 myotoxins from snake
venoms (Figure 4(a)) BmatTX-I showed 94 similarity withMjTX-I present in B moojeni venom 92 with BthTX-I andBOJU-I present in B jararacussu venom and 94 similaritywith MTX-II present in B brazili venom
Additionallymultiple sequence alignment of BmatTX-IIIshowed a PLA
2-Asp49 basic myotoxin with another Asp49
of the Bothrops genera (Figure 4(b)) It can be observedthat BmatTX-III presented 78 similarity with BmjeTX-Iand 75 with BmoTX-I both from B moojeni venom 71similarity with BthTX-II isolated from B jararacussu venomand 65 similarity with PrTX-III from B pirajai venom
In the analysis of the sequences of the PLA2s isolated
from B mattogrossensis in this study highly preserved con-stituent residues of the 120572-helix structure characteristic ofphospholipases were identified as well as the presence ofmany cysteine residues which suggests the existence of manydisulfide bridges important for the stabilization of PLA
2rsquos
molecular structure [21 44]
BioMed Research International 7
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1360475
1398779
(a)
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1362326
1380673
(b)
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1256219
1368171
1402625
(c)
Figure 3 Mass determination by spectrometryThe average proteinmasses of BmatTX-I (a) BmatTX-II (b) and BmatTX-III (c) wereobtained in a MALDI-TOF system operated in linear mode usingexternal standards for calibration The resulting mass spectra weresubmitted to automatic baseline subtraction
Some studies regarding the amino acid composition ofPLA2s demonstrate that these proteins are rich in basic
and hydrophobic amino acids containing three long 120572-helixes two beta sheets and a Ca2+ binding site [45ndash47]Calcium is absolutely necessary for hydrolysis thereforealmost all PLA
2s have a highly preserved region for a Ca2+
bond (XCGXGG) and a catalytic site (DXCCXXHD) [48]It is observed in BmatTX-III that the calcium binding site(27YCGWGG
32) is preserved indicating that it is a catalyt-
ically active PLA2belonging to the PLA
2Asp49 subgroup
33 Biological Activities of Phospholipases A2 The main
objective for the isolation and characterization of compo-nents of snake venom is to better understand the partic-ipation of each component in different pathophysiologicalprocesses resulting from envenomation Local lesions can beattributed to proteases phospholipase activity and hemor-rhagic factors of these venoms followed by the release ofvasoactive agents causing hemorrhaging in various organsand tissues [5 44 49ndash52]
The PLA2s isolated from B mattogrossensis venom
BmatTX-I BmatTX-II and BmatTX-III showed highmyotoxic activity (Figure 2(c)) At a concentration of25 120583g50 120583L the PLA
2s induced a significant release of CK
an important muscular lesion marker when compared to thecontrol Myotoxicity is the characteristic presented by mostbasic phospholipases A
2from snake venoms Several studies
demonstrate that the myotoxic effect begins quickly by directaction of the myotoxic PLA
2s on the plasma membrane of
muscle cells or it is mediated by metalloproteases due toconsequent degeneration and ischemia [27 28 44]
Regarding the Lys49-PLA2myotoxins it is evident that
they lyse the plasma membrane of the muscle cell infected invivo however the exact mechanism has not been describedyet Furthermore it is not known if the toxin is internalizedbefore during or after the initial lysis or if it is not internal-ized Althoughmyotoxicity can be induced by the productionof fatty acids there is a second mechanism that seems to beindependent of the enzymatic activity and is mediated bythe C-terminal region at sites 115ndash129 of the Lys49 molecules[43 53 54]
In an attempt to better understand the development ofthe inflammatory process unleashed by the protein complexpresent in snake venoms many studies have been donesuch as edema induction [55] leukocyte participation [5657] mast cells degranulation [55] participation of variouscytotoxins in the inflammatory system [58 59] participationof cyclooxygenases [60 61] and the participation of venomPLA2s in the inflammatory process [6 51 56 62]
Many studies about PLA2activity in macrophages have
already been done [8 63 64] Little is known about PLA2rsquos
effect on neutrophils however Escocard et al [65] describedan influx of inflammatory cells including many neutrophilsinto the peritoneal cavity of mice after the injection ofBothrops atrox venom The induction of reactive oxygenspecies (ROS) cytokines like IL-6 and IL-1120573 was seen inthese neutrophils These data were also observed in a studycarried out by Souza et al [26] where besides the influx ofneutrophils into the peritoneal cavity ofmice after injection ofthe venom of B atrox there was also induction of superoxideby these cells mast cell degranulation and phagocytosisby macrophages Regarding the activity of PLA
2Gambero
et al [66] have observed the ability of some myotoxins(bothropstoxin-I-II and piratoxin-I) to induce neutrophilchemotaxis in a concentration-dependent manner
In order to evaluate the activation of leukocytes thetoxicity of B mattogrossensis myotoxins on neutrophil cellswas investigated The cells were incubated with differentconcentrations (3 6 and 12 120583gmL) of BmatTX-I (Lys49)BmatTX-II (Lys49) and BmatTX-III (Asp49) myotoxinsduring 1 12 and 24 hours (data not shown)These myotoxinsdid not affect the neutrophilsrsquo viability which agrees withZuliani et al [64] showing low toxicity on thioglycollateelicited mice macrophages
Nonetheless our data showed that neutrophils incubatedwith BmatTX-I and BmatTX-II myotoxins induced therelease of IL-1120573 (Figures 5(a) and 5(b)) Moreover theseresults suggest that phospholipid hydrolysis is not essentialfor the activity observed and argue with the hyphotesis that
8 BioMed Research International
lowastlowast lowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowast lowastlowastlowastlowastlowastlowastlowastlowastlowast
1 10 20 30 40 50(Identity )
100
89
92
92
92
94
94
BmatTX-IBmatTX-IIPrtx-IBOJU-IBthTX-IMjtx-IMtx-Ii
(a)
lowastlowastlowast lowastlowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowastlowast lowastlowastlowast
1 10 20 30 40 50
(Identity )100
78
75
71
65
BmatTX-IIIBmoTX-IBmjeTX-IBthTX-IIPrTX-III
(b)
Figure 4 Comparison of the N-terminal sequence of the snakersquos PLA2s with the PLA
2s isolated from B mattogrossensis venom N-terminal
sequencing of the PLA2s obtained by Edman [13] degradation and alignment done with CLUSTALW2 expressed as of similarity (a)
BmatTX-I BmatTX-II with Lys49 residues in marked sequences (b) BmatTX-III with Asp49 residues in marked sequences Sequencesused for the alignment and their respective access numbers BthTX-I (gi51890398) MjTX-I (gi17368325) Mtx-II (gi390981003) PrTX-I (gi190016174) BOJU-I (gi209572966) BthTX-II (gi1171971) PrTX-III (gi90016174) BmjeTX-I (gi313471399) BmoTX-I (gi221272396)BthTX-I (Lys49) BthTX-II (Asp49) and BOJU-I (Lys49) isolated from the venom of B jararacussu MjTX-I (Lys49) BmoTX-I (Asp49)and BmjeTX-I (Asp49) isolated from the venom of B moojeni Mtx-II (Lys49) isolated from the venom of B brazili PrTX-I (Lys49) andPrTX-III (Asp49) isolated from the venom of B pirajai the ldquolowastrdquo was used to indicate the amino acid residues that are the same between thesequences
other molecular regions distinct from the active site may beinvolved in this effect
PLA2s are multifunctional proteins that can be used as
mediators in several areas of medicine such as in the treat-ment of rheumatoid arthritis as a new class of HIV inhibitorsby blocking the host cell invasion as a potential treatmentagainst malaria and as an antibiotic by inducing cytotoxicityvia the disruption of bacterial membranes [21 23 30 6768] A study by Costa et al [67] with PLA
2s isolated from
B brazili MTX-I and II demonstrated cytotoxic activityagainst Jurkat tumor cells as well as antimicrobial activityagainstE coli andC albicans and antiparasitic activity againstLeishmania sp
In the present study we evaluated the cytotoxic activityof PLA
2s isolated from B mattogrossensis on JURKAT (T
leukemia) and SK-BR-3 (breast adenocarcinoma) both ofwhich are human tumor cell lines Like MTX-II of B brazili[67] the cytotoxic activity of PLA
2s BmatTX-I and BmatTX-
II on JURKATcells was independent of their catalytic activitysince these are characterized as Lys49 PLA
2s and are therefore
catalytically inactive BmatTX-III characterized as Asp49despite being enzymatically active also showed a lower level oftoxicity Some authors propose that the cytotoxic activity ontumor cell lines is associated with the induction of apoptosisconsidering the fact that PLA
2promotes alterations in the
cell membrane And some studies involving Lys49 PLA2s
isolated from B asper demonstrated that the C-terminal
region comprised of amino acids 115ndash129 is concerned withthe cytotoxic and bactericidal activities of this protein [69ndash71] The same observation was made by Costa et al [67] withsynthetic peptides derived from the C-terminal portion ofMTX-I and II PLA
2s
We also evaluated the antiparasitic activity of the crudevenom and isolated PLA
2of B mattogrossensis on promastig-
ote forms of L amazonensis After the analysis it wasobserved that the crude venom of B mattogrossensis presentsincreasing toxic activity (approximately 50 to 80) againstpromastigote forms of L amazonensis after 48 h of incubation(Figure 7(a)) The PLA
2s isolated from B mattogrossensis at
100 120583gmL characterized as BmatTX-I (Lys49) and BmatTX-III (Asp49) presented toxic activity between 25 and 30respectively even with values close to those presented afterincubation of the protozoan with Pentamidine a drug usedas a positive control (Figures 7(b) and 7(c))
When compared the cytotoxicity values of PLA2s against
promastigote forms of L amazonensis show similar activitybetween the Lys49-PLA
2and the Asp49-PLA
2 Comparison
of this activity with the crude venom (70) showed thatPLA2s are responsible for almost half of the observed effect
Nonetheless notably the results suggest that other toxinspresent in the venom contribute to the parasitersquos death
Similar to the results obtained in the present studyStabeli et al [21] demonstrated that MjTX-II a Lys49-PLA
2
homologue isolated from B moojeni venom in different
BioMed Research International 9
0
500
1000
1500
2000
lowast
lowast
lowast
lowast
lowast
Control 3 6 12 3 6 12 3 6 12
ControlBmatTX-I (Lys49)
BmatTX-II (Lys49)BmatTX-III (Asp49)
BmatTX-I (120583g) BmatTX-II (120583g) BmatTX-III (120583g)
IL-1120573
(pg
mL)
(a)
Control 3 6 12 3 6 12 3 6 120
500
1000
1500
2000
ControlBmatTX-I (Lys49)
BmatTX-II (Lys49)BmatTX-III (Asp49)
lowastlowastlowast
BmatTX-I (120583g) BmatTX-II (120583g) BmatTX-III (120583g)
IL-1120573
(pg
mL)
(b)
Figure 5 Production of IL-1120573 by mice neutrophils These cells wereincubatedwith PLA
2(3 6 and 12 120583gmL) or RPMI (control) for 12
hours (a) and 24 hours (b) at 37∘C in a humid atmosphere with 5CO2 Quantification was done by ELISA as described in 29 Each
bar represents the average +minus SDof three independent groups lowast119875 lt005 compared to the control
concentrations (5 25 and 75 120583g) was effective as a parasiticideagent against Schistosomamansoni and promastigote forms ofLeishmania (L amazonensis L braziliensis L donovani andL major)
The action of the PLA2s BmatTX-I (Lys49) and
BmatTX-III (Asp49) on promastigote forms of L ama-zonensis was independent of its catalytic activity sincecatalytically inactive Lys49 myotoxin also demonstrated tox-icity against Leishmania It is believed that the observedcytotoxic activity might be related to the C-terminal regionsof these phospholipase-homologues that are able to promotea disturbance in the cellular membranes independent oftheir catalytic activity [67 72] However more studies arenecessary to define the exact mechanism of action of theseenzymes on parasites
Growing interest in the comprehension of the structureand function of snake venom components especially PLA
2
10 100 10 100 10 100 10 1000
20
40
60
80
100
BmatTX-I (Lys49)BmatTX-II (Lys49)
BmatTX-III (Asp49)Methotrexate (MTX)
Cyto
toxi
city
()
Concentration (120583gmL)
(a)
10 100 10 100 10 100 10 1000
20
40
60
80
100
BmatTX-I (Lys49)BmatTX-II (Lys49)
BmatTX-III (Asp49)Methotrexate (MTX)
Cyto
toxi
city
()
Concentration (120583gmL)
(b)
Figure 6 Antitumoral activity of PLA2s from B mattogrossensis
(a) Antitumoral activity on human acute T-cell leukemia (JURKAT)lines (b) Antitumoral activity on human breast adenocarcinoma(SK-BR-3) Different concentrations of the PLA
2s were incubated
with cell linesMethotrexate was used as the positive control Resultsare presented as mean +minus SD (119899 = 3)
contributes to a better understanding of the mechanism ofaction of their enzymatic and toxic activities It opens thepath to better understand the intoxication caused by enven-omation and the physiopathology behind its side effectsFuture studies will potentially improve serum therapy andhelp develop the pharmaceutical potential that moleculesisolated from animal venoms can have such as the PLA
2s
isolated from the B mattogrossensis venom which show anti-Leishmania and antitumor activities
4 Conclusion
In conclusion the venom of Bothrops mattogrossensis hasa qualitatively similar toxicological profile to previously
10 BioMed Research International
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineB mattogrossensis crude venom
Ant
i-leis
hman
ia ac
tivity
()
B mattogrossensis crude venom (120583g)
(a)
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineBmatTX-I (Lys49)
Ant
i-leis
hman
ia ac
tivity
()
BmatTX-I (120583g)
lowastlowast lowast lowast
(b)
Ant
i-leis
hman
ia ac
tivity
()
Pent 36 625 125 25 50 1000
20
40
60
80
100
BmatTX-III (Asp49)Pentamidine
BmatTX-III (120583g)
lowastlowastlowast
lowastlowast lowast
(c)
Figure 7 Antileishmanial activity of the crude venom of and PLA2s from B mattogrossensis The promastigote forms of L amazonensiswere
plated with 1times105 cellswellThen different concentrations of crude venom and isolated PLA2s were added to each wellThe experiment was
done in a 48 h period with the (a) crude venom and the PLA2enzymes (b) BmatTX-I and (c) BmatTX-III MTT was added and after the
incubation period at 33∘C the formazan crystal formed was dissolved in SDS Readings were done in a spectrophotometer at 570 nm Eachbar represents the average +minus SD of the three independent experimental groups sixfold total lowast119875 lt 005 compared to the control
studied snake venoms of the Bothrops sp genera despite theobservation of quantitative variations Of the three basicPLA2s from B mattogrossensis venom now isolated for the
first time two are characterized as Lys49-PLA2homologues
BmatTX-I and -II and the other as an Asp49-PLA2 named
BmatTX-III This showed high phospholipase activity ThePLA2s isolated induced myotoxic effects as well as the release
of proinflammatory cytokines by neutrophils BmatTX-I and-III PLA
2s were cytotoxic to human tumor cell lines JURKAT
and SK-BR-3 and showed activity against promastigote formsof L amazonensis
Conflict of Interests
The authors state that there is no conflict of interests
Acknowledgments
The authors are grateful to the Ministry of Science andTechnology (MCTI) Conselho Nacional de Desenvolvi-mento Cientıfico e Tecnologico (CNPq) Financiadora deEstudos e Projetos (FINEP) Fundacao de Tecnologia doAcre (FUNTACFDCT) Coordenacao de Aperfeicoamentode Nıvel Superior (CAPES)mdashProjeto NanoBiotec Redede Biodiversidade e Biotecnologia da Amazonia Legal(BIONORTECNPqMCTI) Instituto Nacional paraPesquisa Translacional em Saude e Ambiente na RegiaoAmazonica (INCT-INPeTAmCNPqMCTI) and InstitutoNacional para Pesquisa em Toxinas (INCT-Tox) andSecretary of Development of Rondonia State (SEPLANPRONEXCNPq) for financial support We are grateful to
BioMed Research International 11
Sr Guilherme G Galassi (Mantenedouro da Fauna Silvestreautorizacao IBAMA 020270089912001-95 Americana-SP)for their helpful technical collaboration in provide the snakevenom Amy Grabner provided the English editing of thepaper
References
[1] Y Angulo and B Lomonte ldquoBiochemistry and toxicology oftoxins purified from the venom of the snake Bothrops asperrdquoToxicon vol 54 no 7 pp 949ndash957 2009
[2] J J Calvete P Juarez and L Sanz ldquoSnake venomics Strategyand applicationsrdquo Journal of Mass Spectrometry vol 42 no 11pp 1405ndash1414 2007
[3] T S Kang D Georgieva N Genov et al ldquoEnzymatic toxinsfrom snake venom structural characterization and mechanismof catalysisrdquoThe FEBS Journal vol 278 no 23 pp 4544ndash45762011
[4] C ZOliveiraCaracterizacao funcional e estrutural de inibidoresde fosfolipases A
2isolados do plasma de serpente Bothrops
jararacussu [PhD thesis] Faculdade de Ciencias Farmaceuticasde Ribeirao Preto Ribeirao Preto Brazil 2009
[5] R G Stabeli R Simoes-Silva A M Kayano et al ldquoPurifi-cation of phospholipases A
2from American snake venomsrdquo
in ChromatographymdashThe Most Versatile Method of ChemicalAnalysis chapter 1 pp 1ndash34 InTech Vienna Austria 2012
[6] J P Zuliani C M Fernandes S R Zamuner J M Gutierrezand C F P Teixeira ldquoInflammatory events induced by Lys-49 and Asp-49 phospholipases A
2isolated from Bothrops asper
snake venom role of catalytic activityrdquo Toxicon vol 45 no 3pp 335ndash346 2005
[7] S Marcussi R G Stabeli N A Santos-Filho et al ldquoGenotoxiceffect of Bothrops snake venoms and isolated toxins on humanlymphocyte DNArdquo Toxicon vol 65 pp 9ndash14 2013
[8] S S Setubal A S Pontes J L Furtado et al ldquoAction oftwo phospholipases A
2purified from Bothrops alternatus snake
venom on macrophagesrdquo Biochemistry vol 78 no 2 pp 194ndash203 2013
[9] J M Gutierrez L A Ponce-Soto S Marangoni and BLomonte ldquoSystemic and local myotoxicity induced by snakevenom group II phospholipases A
2 comparison between cro-
toxin crotoxin B and a Lys49 PLA2homologuerdquo Toxicon vol
51 no 1 pp 80ndash92 2008[10] A Q Rueda I G Rodrıguez E C Ara et al ldquoBiochemical
characterization action onmacrophages and superoxide anionproduction of four basic phospholipases A
2from Panamanian
Bothrops asper snake venomrdquo BioMed Research Internationalvol 2013 Article ID 789689 9 pages 2013
[11] V X da Silva and M T Rodrigues ldquoTaxonomic revision ofthe Bothrops neuwiedi complex (Serpentes Viperidae) withdescription of a new speciesrdquo Phyllomedusa vol 7 no 1 pp 45ndash90 2008
[12] J A Campbell and W W Lamar The Venomous Reptiles of theWestern Hemisphere Comstock Ithaca NY USA 2004
[13] P Edman ldquoMethod for determination of the amino acidsequence in peptidesrdquo Acta Chemica Scandinavica vol 4 pp283ndash293 1950
[14] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[15] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[16] M Holzer and S P Mackessy ldquoAn aqueous endpoint assay ofsnake venom phospholipase A
2rdquo Toxicon vol 34 no 10 pp
1149ndash1155 1996[17] A L Fuly A L P deMiranda R B Zingali and J AGuimaraes
ldquoPurification and characterization of a phospholipase A2isoen-
zyme isolated from Lachesis muta snake venomrdquo BiochemicalPharmacology vol 63 no 9 pp 1589ndash1597 2002
[18] T Nikai N Mori M Kishida H Sugihara and A T Tu ldquoIso-lation and biochemical characterization of hemorrhagic toxinf from the venom of Crotalus atrox (Western DiamondbackRattlesnake)rdquo Archives of Biochemistry and Biophysics vol 231no 2 pp 309ndash319 1984
[19] J A Gene A Roy G Rojas J M Gutierrez and L CerdasldquoComparative study on coagulant defibrinating fibrinolyticand fibrinogenolytic activities of Costa Rican crotaline snakevenoms and their neutralization by a polyvalent antivenomrdquoToxicon vol 27 no 8 pp 841ndash848 1989
[20] M S Charney and R M Tomarelli ldquoA colorimetric method forthe determination of the proteolytic activity of duodenal juicerdquoThe Journal of Biological Chemistry vol 171 pp 501ndash505 1947
[21] R G Stabeli S F Amui C D SantrsquoAna et al ldquoBothrops moojenimyotoxin-II a Lys49-phospholipaseA
2homologue an example
of function versatility of snake venom proteinsrdquo ComparativeBiochemistry and Physiology C vol 142 no 3-4 pp 371ndash3812006
[22] D R Call J A Nemzek S J Ebong G L Bolgos D ENewcomb and D G Remick ldquoRatio of local to systemicchemokine concentrations regulates neutrophil recruitmentrdquoThe American Journal of Pathology vol 158 no 2 pp 715ndash7212001
[23] P G Roberto S Kashima S Marcussi et al ldquoCloning andidentification of a complete cDNA coding for a bactericidal andantitumoral acidic phospholipase A
2from Bothrops jararacussu
venomrdquoThe Protein Journal vol 23 no 4 pp 273ndash285 2004[24] J H Schumacher A OrsquoGarra B Shrader et al ldquoThe charac-
terization of four monoclonal antibodies specific for mouse IL-5 and development of mouse and human IL-5 enzyme-linkedimmunosorbentrdquoThe Journal of Immunology vol 141 no 5 pp1576ndash1581 1988
[25] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[26] C A T Souza A M Kayano S S Setubal et al ldquoLocal andsystemic biochemical alterations induced by Bothrops atroxsnake venom in micerdquo Journal of Venom Research vol 3 pp28ndash34 2012
[27] J M Gutierrez and B Lomonte ldquoPhospholipase A2myotoxins
from Bothrops snake venomsrdquo Toxicon vol 33 no 11 pp 1405ndash1424 1995
[28] J M Gutierrez and A Rucavado ldquoSnake venom metallopro-teinases their role in the pathogenesis of local tissue damagerdquoBiochimie vol 82 no 9-10 pp 841ndash850 2000
[29] T Matsui Y Fujimura and K Titani ldquoSnake venom proteasesaffecting hemostasis and thrombosisrdquo Biochimica et BiophysicaActa vol 1477 no 1-2 pp 146ndash156 2000
[30] A M Soares M R M Fontes and J R Giglio ldquoPhospholipaseA2myotoxins from Bothrops snake venoms structure-function
12 BioMed Research International
relationshiprdquoCurrent Organic Chemistry vol 8 no 17 pp 1677ndash1690 2004
[31] VM Rodrigues SMarcussi R S Cambraia et al ldquoBactericidaland neurotoxic activities of two myotoxic phospholipases A
2
from Bothrops neuwiedi pauloensis snake venomrdquo Toxicon vol44 no 3 pp 305ndash314 2004
[32] N A Santos-Filho L B Silveira C Z Oliveira et al ldquoAnew acidic myotoxic anti-platelet and prostaglandin I
2induc-
tor phospholipase A2isolated from Bothrops moojeni snake
venomrdquo Toxicon vol 52 no 8 pp 908ndash917 2008[33] A M Soares S H Andriao-Escarso Y Angulo et al ldquoStruc-
tural and functional characterization of myotoxin I a Lys49phospholipaseA
2homologue fromBothropsmoojeni (Caissaca)
snake venomrdquo Archives of Biochemistry and Biophysics vol 373no 1 pp 7ndash15 2000
[34] A M Soares S H Andriao-Escarso R K Bortoleto et alldquoDissociation of enzymatic and pharmacological properties ofpiratoxins-I and -III two myotoxic phospholipases A
2from
Bothrops pirajai snake venomrdquo Archives of Biochemistry andBiophysics vol 387 no 2 pp 188ndash196 2001
[35] S S Teixeira L B Silveira F M N da Silva et al ldquoMolecularcharacterization of an acidic phospholipase A
2from Bothrops
pirajai snake venom synthetic C-terminal peptide identifies itsantiplatelet regionrdquo Archives of Toxicology vol 85 no 10 pp1219ndash1233 2011
[36] S H Andriao-Escarso A M Soares V M Rodrigues et alldquoMyotoxic phospholipases A
2in Bothrops snake venoms effect
of chemical modifications on the enzymatic and pharmacolog-ical properties of bothropstoxins from Bothrops jararacussurdquoBiochimie vol 82 no 8 pp 755ndash763 2000
[37] S H Andriao-Escarso A M Soares M R M Fontes et alldquoStructural and functional characterization of an acidic plateletaggregation inhibitor and hypotensive phospholipase A
2from
Bothrops jararacussu snake venomrdquo Biochemical Pharmacologyvol 64 no 4 pp 723ndash732 2002
[38] M E G Denegri O C Acosta S Huancahuire-Vega et alldquoIsolation and functional characterization of a new acidic PLA
2
Ba SpII RP4 of the Bothrops alternatus snake venom fromArgentinardquo Toxicon vol 56 no 1 pp 64ndash74 2010
[39] R Zouari-Kessentini J Luis A Karray et al ldquoTwo purified andcharacterized phospholipases A
2from Cerastes cerastes venom
that inhibit cancerous cell adhesion and migrationrdquo Toxiconvol 53 no 4 pp 444ndash453 2009
[40] R Shirai M Toriba K Hayashi K Ikeda and S Inoue ldquoIden-tification and characterization of phospholipase A
2inhibitors
from the serum of the Japanese rat snake Elaphe climacophorardquoToxicon vol 53 no 6 pp 685ndash692 2009
[41] M I Homsi-Brandenburgo L S Queiroz H Santo-Neto LRodrigues-Simoni and J R Giglio ldquoFractionation of Bothropsjararacussu snake venom partial chemical characterization andbiological activity of bothropstoxinrdquo Toxicon vol 26 no 7 pp615ndash627 1988
[42] G P Fernandez A Segura M Herrera et al ldquoNeutralizationof Bothrops mattogrossensis snake venom from Bolivia exper-imental evaluation of llama and donkey antivenoms producedby caprylic acid precipitationrdquoToxicon vol 55 no 2-3 pp 642ndash645 2010
[43] C L Ownby H S S de Araujo S P White and J E FletcherldquoLysine 49 phospholipase A
2proteinsrdquo Toxicon vol 37 no 3
pp 411ndash445 1999[44] A M Soares W P Sestito S Marcussi et al ldquoAlkylation of
myotoxic phospholipases A2in Bothrops moojeni venom a
promising approach to an enhanced antivenom productionrdquoInternational Journal of Biochemistry amp Cell Biology vol 36 no2 pp 258ndash270 2004
[45] B Lomonte J M Gutierrez M F Furtado et al ldquoIsolationof basic myotoxins from Bothrops moojeni and Bothrops atroxsnake venomsrdquo Toxicon vol 28 no 10 pp 1137ndash1146 1990
[46] L C Mancuso M M Correa C A Vieira et al ldquoFractionationof Bothrops pirajai snake venom isolation and characterizationof piratoxin-I a new myotoxic proteinrdquo Toxicon vol 33 no 5pp 615ndash626 1995
[47] H S Selistre L S Queiroz O A B Cunha G E P deSouza and J R Giglio ldquoIsolation and characterization ofhemorrhagic myonecrotic and edema-inducing toxins fromBothrops insularis (jararaca ilhoa) snake venomrdquo Toxicon vol28 no 3 pp 261ndash273 1990
[48] E A Dennis J Cao Y-H Hsu V Magrioti and G KokotosldquoPhospholipaseA
2enzymes physical structure biological func-
tion disease implication chemical inhibition and therapeuticinterventionrdquo Chemical Reviews vol 111 no 10 pp 6130ndash61852011
[49] A G T Cardoso Acao do veneno de Bothrops moojeni e suafracao L-aminoacido oxidase submetida ao tratamento comraios gama de 60CO em Leishmania spp [MS thesis] IPEN-CNENSP Sao Paulo Brazil 1999
[50] MGallacci andWLGCavalcante ldquoUnderstanding the in vitroneuromuscular activity of snake venomLys49 phospholipaseA
2
homologuesrdquo Toxicon vol 55 no 1 pp 1ndash11 2010[51] S S Teixeira Caracterizacao funcional e estrutural de uma
fosfolipase A2acida isolada do veneno de Bothrops pirajai
[MS thesis] Faculdade de Ciencias Farmaceuticas de RibeiraoPretoUSP Ribeirao Preto Brazil 2009
[52] E Valentin and G Lambeau ldquoIncreasing molecular diversityof secreted phospholipases A
2and their receptors and binding
proteinsrdquo Biochimica et Biophysica Acta vol 1488 no 1-2 pp59ndash70 2000
[53] J M Gutierrez and B Lomonte ldquoPhospholipases A2from
Viperidae snake venoms how do they induce skeletal muscledamagerdquo Acta Chimica Slovenica vol 58 no 4 pp 647ndash6592011
[54] J M Gutierrez C L Ownby and G V Odell ldquoIsolation of amyotoxin from Bothrops asper venom partial characterizationand action on skeletal musclerdquo Toxicon vol 22 no 1 pp 115ndash128 1984
[55] N G Nascimento M C Sampaio R A Olivo and C TeixeiraldquoContribution of mast cells to the oedema induced by Bothropsmoojeni snake venom and a pharmacological assessment of theinflammatory mediators involvedrdquo Toxicon vol 55 no 2-3 pp343ndash352 2010
[56] C F P Teixeira E C T Landucci E Antunes M Chacurand Y Cury ldquoInflammatory effects of snake venom myotoxicphospholipases A
2rdquo Toxicon vol 42 no 8 pp 947ndash962 2003
[57] S R Zamuner and C F P Teixeira ldquoCell adhesion moleculesinvolved in the leukocyte recruitment induced by venom of thesnake Bothrops jararacardquoMediators of Inflammation vol 11 no6 pp 351ndash357 2002
[58] F Chaves C F P Teixeira and J M Gutierrez ldquoRole ofTNF-120572 IL-1120573 and IL-6 in the local tissue damage induced byBothrops asper snake venom an experimental assessment inmicerdquo Toxicon vol 45 no 2 pp 171ndash178 2005
BioMed Research International 13
[59] C D F P Teixeira C M Fernandes J P Zuliani and SF Zamuner ldquoInflammatory effects of snake venom metallo-proteinasesrdquo Memorias do Instituto Oswaldo Cruz vol 100supplement 1 pp 181ndash184 2005
[60] R D A Olivo C F P Teixeira J LWallace J M Gutierrez andS R Zamuner ldquoRole of cyclooxygenases in oedema-formingactivity of bothropic venomsrdquo Toxicon vol 49 no 5 pp 670ndash677 2007
[61] L B Silveira D P Marchi-Salvador N A Santos-Filho et alldquoIsolation and expression of a hypotensive and anti-plateletacidic phospholipase A
2from Bothrops moojeni snake venomrdquo
Journal of Pharmaceutical and Biomedical Analysis vol 73 pp35ndash43 2013
[62] A Rucavado T Escalante C F P Teixeira C M Fernandes CDıaz and J M Gutierrez ldquoIncrements in cytokines and matrixmetalloproteinases in skeletal muscle after injection of tissue-damaging toxins from the venom of the snake Bothrops asperrdquoMediators of Inflammation vol 11 no 2 pp 121ndash128 2002
[63] E Leiguez J P Zuliani A M Cianciarullo C M Fernandes JM Gutierrez and C Teixeira ldquoA group IIA-secreted phospho-lipase A
2from snake venom induces lipid body formation in
macrophages the roles of intracellular phospholipases A2and
distinct signaling pathwaysrdquo Journal of Leukocyte Biology vol90 no 1 pp 155ndash166 2011
[64] J P Zuliani J M Gutierrez L L Casais e Silva S C SampaioB Lomonte and C D F P Teixeira ldquoActivation of cellularfunctions in macrophages by venom secretory Asp-49 and Lys-49 phospholipasesA
2rdquoToxicon vol 46 no 5 pp 523ndash532 2005
[65] R D C M Escocard M M Kanashiro J H Petretski et alldquoNeutrophils regulate the expression of cytokines chemokinesand nitric oxide synthasenitric oxide in mice injected withBothrops atrox venomrdquo Immunobiology vol 211 no 1-2 pp 37ndash46 2006
[66] A Gambero E C T Landucci M H Toyama et al ldquoHumanneutrophil migration in vitro induced by secretory phospholi-pases A
2 a role for cell surface glycosaminoglycansrdquo Biochemi-
cal Pharmacology vol 63 no 1 pp 65ndash72 2002[67] F L S Costa R S Rodrigues L F M Izidoro et al ldquoBio-
chemical and functional properties of a thrombin-like enzymeisolated from Bothrops pauloensis snake venomrdquo Toxicon vol54 no 6 pp 725ndash735 2009
[68] J C Villalobos R Mora B Lomonte J M Gutierrez andY Angulo ldquoCytotoxicity induced in myotubes by a Lys49phospholipase A
2homologue from the venom of the snake
Bothrops asper evidence of rapid plasma membrane damageand a dual role for extracellular calciumrdquo Toxicology in Vitrovol 21 no 8 pp 1382ndash1389 2007
[69] B S Cummings J McHowat and R G Schnellmann ldquoPhos-pholipase A
2s in cell injury and deathrdquoThe Journal of Pharma-
cology and Experimental Therapeutics vol 294 no 3 pp 793ndash799 2000
[70] S R Panini L Yang A E Rusinol M S Sinensky J VBonventre and C C Leslie ldquoArachidonate metabolism andthe signaling pathway of induction of apoptosis by oxidizedLDLoxysterolrdquo Journal of Lipid Research vol 42 no 10 pp1678ndash1686 2001
[71] L Paramo B Lomonte J Pizarro-Cerda J-A Bengoechea J-P Gorvel and E Moreno ldquoBactericidal activity of Lys49 andAsp49 myotoxic phospholipases A
2from Bothrops asper snake
venom synthetic Lys49myotoxin II-(115-129)-peptide identifiesits bactericidal regionrdquo European Journal of Biochemistry vol253 no 2 pp 452ndash461 1998
[72] B Lomonte Y Angulo and E Moreno ldquoSynthetic peptidesderived from the C-terminal region of Lys49 phospholipaseA2homologues from viperidae snake venoms biomimetic
activities and potential applicationsrdquo Current PharmaceuticalDesign vol 16 no 28 pp 3224ndash3230 2010
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
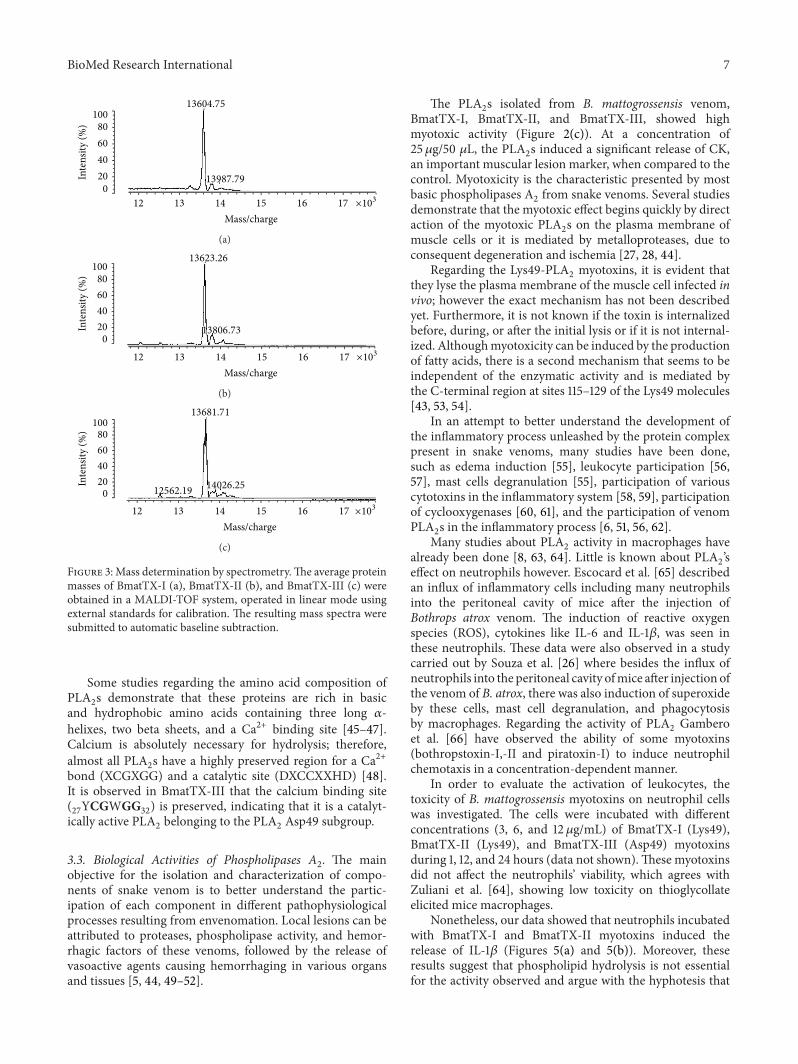
BioMed Research International 7
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1360475
1398779
(a)
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1362326
1380673
(b)
10080
60
40
200
12 13 14 15 16 17 times103
Masscharge
Inte
nsity
()
1256219
1368171
1402625
(c)
Figure 3 Mass determination by spectrometryThe average proteinmasses of BmatTX-I (a) BmatTX-II (b) and BmatTX-III (c) wereobtained in a MALDI-TOF system operated in linear mode usingexternal standards for calibration The resulting mass spectra weresubmitted to automatic baseline subtraction
Some studies regarding the amino acid composition ofPLA2s demonstrate that these proteins are rich in basic
and hydrophobic amino acids containing three long 120572-helixes two beta sheets and a Ca2+ binding site [45ndash47]Calcium is absolutely necessary for hydrolysis thereforealmost all PLA
2s have a highly preserved region for a Ca2+
bond (XCGXGG) and a catalytic site (DXCCXXHD) [48]It is observed in BmatTX-III that the calcium binding site(27YCGWGG
32) is preserved indicating that it is a catalyt-
ically active PLA2belonging to the PLA
2Asp49 subgroup
33 Biological Activities of Phospholipases A2 The main
objective for the isolation and characterization of compo-nents of snake venom is to better understand the partic-ipation of each component in different pathophysiologicalprocesses resulting from envenomation Local lesions can beattributed to proteases phospholipase activity and hemor-rhagic factors of these venoms followed by the release ofvasoactive agents causing hemorrhaging in various organsand tissues [5 44 49ndash52]
The PLA2s isolated from B mattogrossensis venom
BmatTX-I BmatTX-II and BmatTX-III showed highmyotoxic activity (Figure 2(c)) At a concentration of25 120583g50 120583L the PLA
2s induced a significant release of CK
an important muscular lesion marker when compared to thecontrol Myotoxicity is the characteristic presented by mostbasic phospholipases A
2from snake venoms Several studies
demonstrate that the myotoxic effect begins quickly by directaction of the myotoxic PLA
2s on the plasma membrane of
muscle cells or it is mediated by metalloproteases due toconsequent degeneration and ischemia [27 28 44]
Regarding the Lys49-PLA2myotoxins it is evident that
they lyse the plasma membrane of the muscle cell infected invivo however the exact mechanism has not been describedyet Furthermore it is not known if the toxin is internalizedbefore during or after the initial lysis or if it is not internal-ized Althoughmyotoxicity can be induced by the productionof fatty acids there is a second mechanism that seems to beindependent of the enzymatic activity and is mediated bythe C-terminal region at sites 115ndash129 of the Lys49 molecules[43 53 54]
In an attempt to better understand the development ofthe inflammatory process unleashed by the protein complexpresent in snake venoms many studies have been donesuch as edema induction [55] leukocyte participation [5657] mast cells degranulation [55] participation of variouscytotoxins in the inflammatory system [58 59] participationof cyclooxygenases [60 61] and the participation of venomPLA2s in the inflammatory process [6 51 56 62]
Many studies about PLA2activity in macrophages have
already been done [8 63 64] Little is known about PLA2rsquos
effect on neutrophils however Escocard et al [65] describedan influx of inflammatory cells including many neutrophilsinto the peritoneal cavity of mice after the injection ofBothrops atrox venom The induction of reactive oxygenspecies (ROS) cytokines like IL-6 and IL-1120573 was seen inthese neutrophils These data were also observed in a studycarried out by Souza et al [26] where besides the influx ofneutrophils into the peritoneal cavity ofmice after injection ofthe venom of B atrox there was also induction of superoxideby these cells mast cell degranulation and phagocytosisby macrophages Regarding the activity of PLA
2Gambero
et al [66] have observed the ability of some myotoxins(bothropstoxin-I-II and piratoxin-I) to induce neutrophilchemotaxis in a concentration-dependent manner
In order to evaluate the activation of leukocytes thetoxicity of B mattogrossensis myotoxins on neutrophil cellswas investigated The cells were incubated with differentconcentrations (3 6 and 12 120583gmL) of BmatTX-I (Lys49)BmatTX-II (Lys49) and BmatTX-III (Asp49) myotoxinsduring 1 12 and 24 hours (data not shown)These myotoxinsdid not affect the neutrophilsrsquo viability which agrees withZuliani et al [64] showing low toxicity on thioglycollateelicited mice macrophages
Nonetheless our data showed that neutrophils incubatedwith BmatTX-I and BmatTX-II myotoxins induced therelease of IL-1120573 (Figures 5(a) and 5(b)) Moreover theseresults suggest that phospholipid hydrolysis is not essentialfor the activity observed and argue with the hyphotesis that
8 BioMed Research International
lowastlowast lowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowast lowastlowastlowastlowastlowastlowastlowastlowastlowast
1 10 20 30 40 50(Identity )
100
89
92
92
92
94
94
BmatTX-IBmatTX-IIPrtx-IBOJU-IBthTX-IMjtx-IMtx-Ii
(a)
lowastlowastlowast lowastlowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowastlowast lowastlowastlowast
1 10 20 30 40 50
(Identity )100
78
75
71
65
BmatTX-IIIBmoTX-IBmjeTX-IBthTX-IIPrTX-III
(b)
Figure 4 Comparison of the N-terminal sequence of the snakersquos PLA2s with the PLA
2s isolated from B mattogrossensis venom N-terminal
sequencing of the PLA2s obtained by Edman [13] degradation and alignment done with CLUSTALW2 expressed as of similarity (a)
BmatTX-I BmatTX-II with Lys49 residues in marked sequences (b) BmatTX-III with Asp49 residues in marked sequences Sequencesused for the alignment and their respective access numbers BthTX-I (gi51890398) MjTX-I (gi17368325) Mtx-II (gi390981003) PrTX-I (gi190016174) BOJU-I (gi209572966) BthTX-II (gi1171971) PrTX-III (gi90016174) BmjeTX-I (gi313471399) BmoTX-I (gi221272396)BthTX-I (Lys49) BthTX-II (Asp49) and BOJU-I (Lys49) isolated from the venom of B jararacussu MjTX-I (Lys49) BmoTX-I (Asp49)and BmjeTX-I (Asp49) isolated from the venom of B moojeni Mtx-II (Lys49) isolated from the venom of B brazili PrTX-I (Lys49) andPrTX-III (Asp49) isolated from the venom of B pirajai the ldquolowastrdquo was used to indicate the amino acid residues that are the same between thesequences
other molecular regions distinct from the active site may beinvolved in this effect
PLA2s are multifunctional proteins that can be used as
mediators in several areas of medicine such as in the treat-ment of rheumatoid arthritis as a new class of HIV inhibitorsby blocking the host cell invasion as a potential treatmentagainst malaria and as an antibiotic by inducing cytotoxicityvia the disruption of bacterial membranes [21 23 30 6768] A study by Costa et al [67] with PLA
2s isolated from
B brazili MTX-I and II demonstrated cytotoxic activityagainst Jurkat tumor cells as well as antimicrobial activityagainstE coli andC albicans and antiparasitic activity againstLeishmania sp
In the present study we evaluated the cytotoxic activityof PLA
2s isolated from B mattogrossensis on JURKAT (T
leukemia) and SK-BR-3 (breast adenocarcinoma) both ofwhich are human tumor cell lines Like MTX-II of B brazili[67] the cytotoxic activity of PLA
2s BmatTX-I and BmatTX-
II on JURKATcells was independent of their catalytic activitysince these are characterized as Lys49 PLA
2s and are therefore
catalytically inactive BmatTX-III characterized as Asp49despite being enzymatically active also showed a lower level oftoxicity Some authors propose that the cytotoxic activity ontumor cell lines is associated with the induction of apoptosisconsidering the fact that PLA
2promotes alterations in the
cell membrane And some studies involving Lys49 PLA2s
isolated from B asper demonstrated that the C-terminal
region comprised of amino acids 115ndash129 is concerned withthe cytotoxic and bactericidal activities of this protein [69ndash71] The same observation was made by Costa et al [67] withsynthetic peptides derived from the C-terminal portion ofMTX-I and II PLA
2s
We also evaluated the antiparasitic activity of the crudevenom and isolated PLA
2of B mattogrossensis on promastig-
ote forms of L amazonensis After the analysis it wasobserved that the crude venom of B mattogrossensis presentsincreasing toxic activity (approximately 50 to 80) againstpromastigote forms of L amazonensis after 48 h of incubation(Figure 7(a)) The PLA
2s isolated from B mattogrossensis at
100 120583gmL characterized as BmatTX-I (Lys49) and BmatTX-III (Asp49) presented toxic activity between 25 and 30respectively even with values close to those presented afterincubation of the protozoan with Pentamidine a drug usedas a positive control (Figures 7(b) and 7(c))
When compared the cytotoxicity values of PLA2s against
promastigote forms of L amazonensis show similar activitybetween the Lys49-PLA
2and the Asp49-PLA
2 Comparison
of this activity with the crude venom (70) showed thatPLA2s are responsible for almost half of the observed effect
Nonetheless notably the results suggest that other toxinspresent in the venom contribute to the parasitersquos death
Similar to the results obtained in the present studyStabeli et al [21] demonstrated that MjTX-II a Lys49-PLA
2
homologue isolated from B moojeni venom in different
BioMed Research International 9
0
500
1000
1500
2000
lowast
lowast
lowast
lowast
lowast
Control 3 6 12 3 6 12 3 6 12
ControlBmatTX-I (Lys49)
BmatTX-II (Lys49)BmatTX-III (Asp49)
BmatTX-I (120583g) BmatTX-II (120583g) BmatTX-III (120583g)
IL-1120573
(pg
mL)
(a)
Control 3 6 12 3 6 12 3 6 120
500
1000
1500
2000
ControlBmatTX-I (Lys49)
BmatTX-II (Lys49)BmatTX-III (Asp49)
lowastlowastlowast
BmatTX-I (120583g) BmatTX-II (120583g) BmatTX-III (120583g)
IL-1120573
(pg
mL)
(b)
Figure 5 Production of IL-1120573 by mice neutrophils These cells wereincubatedwith PLA
2(3 6 and 12 120583gmL) or RPMI (control) for 12
hours (a) and 24 hours (b) at 37∘C in a humid atmosphere with 5CO2 Quantification was done by ELISA as described in 29 Each
bar represents the average +minus SDof three independent groups lowast119875 lt005 compared to the control
concentrations (5 25 and 75 120583g) was effective as a parasiticideagent against Schistosomamansoni and promastigote forms ofLeishmania (L amazonensis L braziliensis L donovani andL major)
The action of the PLA2s BmatTX-I (Lys49) and
BmatTX-III (Asp49) on promastigote forms of L ama-zonensis was independent of its catalytic activity sincecatalytically inactive Lys49 myotoxin also demonstrated tox-icity against Leishmania It is believed that the observedcytotoxic activity might be related to the C-terminal regionsof these phospholipase-homologues that are able to promotea disturbance in the cellular membranes independent oftheir catalytic activity [67 72] However more studies arenecessary to define the exact mechanism of action of theseenzymes on parasites
Growing interest in the comprehension of the structureand function of snake venom components especially PLA
2
10 100 10 100 10 100 10 1000
20
40
60
80
100
BmatTX-I (Lys49)BmatTX-II (Lys49)
BmatTX-III (Asp49)Methotrexate (MTX)
Cyto
toxi
city
()
Concentration (120583gmL)
(a)
10 100 10 100 10 100 10 1000
20
40
60
80
100
BmatTX-I (Lys49)BmatTX-II (Lys49)
BmatTX-III (Asp49)Methotrexate (MTX)
Cyto
toxi
city
()
Concentration (120583gmL)
(b)
Figure 6 Antitumoral activity of PLA2s from B mattogrossensis
(a) Antitumoral activity on human acute T-cell leukemia (JURKAT)lines (b) Antitumoral activity on human breast adenocarcinoma(SK-BR-3) Different concentrations of the PLA
2s were incubated
with cell linesMethotrexate was used as the positive control Resultsare presented as mean +minus SD (119899 = 3)
contributes to a better understanding of the mechanism ofaction of their enzymatic and toxic activities It opens thepath to better understand the intoxication caused by enven-omation and the physiopathology behind its side effectsFuture studies will potentially improve serum therapy andhelp develop the pharmaceutical potential that moleculesisolated from animal venoms can have such as the PLA
2s
isolated from the B mattogrossensis venom which show anti-Leishmania and antitumor activities
4 Conclusion
In conclusion the venom of Bothrops mattogrossensis hasa qualitatively similar toxicological profile to previously
10 BioMed Research International
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineB mattogrossensis crude venom
Ant
i-leis
hman
ia ac
tivity
()
B mattogrossensis crude venom (120583g)
(a)
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineBmatTX-I (Lys49)
Ant
i-leis
hman
ia ac
tivity
()
BmatTX-I (120583g)
lowastlowast lowast lowast
(b)
Ant
i-leis
hman
ia ac
tivity
()
Pent 36 625 125 25 50 1000
20
40
60
80
100
BmatTX-III (Asp49)Pentamidine
BmatTX-III (120583g)
lowastlowastlowast
lowastlowast lowast
(c)
Figure 7 Antileishmanial activity of the crude venom of and PLA2s from B mattogrossensis The promastigote forms of L amazonensiswere
plated with 1times105 cellswellThen different concentrations of crude venom and isolated PLA2s were added to each wellThe experiment was
done in a 48 h period with the (a) crude venom and the PLA2enzymes (b) BmatTX-I and (c) BmatTX-III MTT was added and after the
incubation period at 33∘C the formazan crystal formed was dissolved in SDS Readings were done in a spectrophotometer at 570 nm Eachbar represents the average +minus SD of the three independent experimental groups sixfold total lowast119875 lt 005 compared to the control
studied snake venoms of the Bothrops sp genera despite theobservation of quantitative variations Of the three basicPLA2s from B mattogrossensis venom now isolated for the
first time two are characterized as Lys49-PLA2homologues
BmatTX-I and -II and the other as an Asp49-PLA2 named
BmatTX-III This showed high phospholipase activity ThePLA2s isolated induced myotoxic effects as well as the release
of proinflammatory cytokines by neutrophils BmatTX-I and-III PLA
2s were cytotoxic to human tumor cell lines JURKAT
and SK-BR-3 and showed activity against promastigote formsof L amazonensis
Conflict of Interests
The authors state that there is no conflict of interests
Acknowledgments
The authors are grateful to the Ministry of Science andTechnology (MCTI) Conselho Nacional de Desenvolvi-mento Cientıfico e Tecnologico (CNPq) Financiadora deEstudos e Projetos (FINEP) Fundacao de Tecnologia doAcre (FUNTACFDCT) Coordenacao de Aperfeicoamentode Nıvel Superior (CAPES)mdashProjeto NanoBiotec Redede Biodiversidade e Biotecnologia da Amazonia Legal(BIONORTECNPqMCTI) Instituto Nacional paraPesquisa Translacional em Saude e Ambiente na RegiaoAmazonica (INCT-INPeTAmCNPqMCTI) and InstitutoNacional para Pesquisa em Toxinas (INCT-Tox) andSecretary of Development of Rondonia State (SEPLANPRONEXCNPq) for financial support We are grateful to
BioMed Research International 11
Sr Guilherme G Galassi (Mantenedouro da Fauna Silvestreautorizacao IBAMA 020270089912001-95 Americana-SP)for their helpful technical collaboration in provide the snakevenom Amy Grabner provided the English editing of thepaper
References
[1] Y Angulo and B Lomonte ldquoBiochemistry and toxicology oftoxins purified from the venom of the snake Bothrops asperrdquoToxicon vol 54 no 7 pp 949ndash957 2009
[2] J J Calvete P Juarez and L Sanz ldquoSnake venomics Strategyand applicationsrdquo Journal of Mass Spectrometry vol 42 no 11pp 1405ndash1414 2007
[3] T S Kang D Georgieva N Genov et al ldquoEnzymatic toxinsfrom snake venom structural characterization and mechanismof catalysisrdquoThe FEBS Journal vol 278 no 23 pp 4544ndash45762011
[4] C ZOliveiraCaracterizacao funcional e estrutural de inibidoresde fosfolipases A
2isolados do plasma de serpente Bothrops
jararacussu [PhD thesis] Faculdade de Ciencias Farmaceuticasde Ribeirao Preto Ribeirao Preto Brazil 2009
[5] R G Stabeli R Simoes-Silva A M Kayano et al ldquoPurifi-cation of phospholipases A
2from American snake venomsrdquo
in ChromatographymdashThe Most Versatile Method of ChemicalAnalysis chapter 1 pp 1ndash34 InTech Vienna Austria 2012
[6] J P Zuliani C M Fernandes S R Zamuner J M Gutierrezand C F P Teixeira ldquoInflammatory events induced by Lys-49 and Asp-49 phospholipases A
2isolated from Bothrops asper
snake venom role of catalytic activityrdquo Toxicon vol 45 no 3pp 335ndash346 2005
[7] S Marcussi R G Stabeli N A Santos-Filho et al ldquoGenotoxiceffect of Bothrops snake venoms and isolated toxins on humanlymphocyte DNArdquo Toxicon vol 65 pp 9ndash14 2013
[8] S S Setubal A S Pontes J L Furtado et al ldquoAction oftwo phospholipases A
2purified from Bothrops alternatus snake
venom on macrophagesrdquo Biochemistry vol 78 no 2 pp 194ndash203 2013
[9] J M Gutierrez L A Ponce-Soto S Marangoni and BLomonte ldquoSystemic and local myotoxicity induced by snakevenom group II phospholipases A
2 comparison between cro-
toxin crotoxin B and a Lys49 PLA2homologuerdquo Toxicon vol
51 no 1 pp 80ndash92 2008[10] A Q Rueda I G Rodrıguez E C Ara et al ldquoBiochemical
characterization action onmacrophages and superoxide anionproduction of four basic phospholipases A
2from Panamanian
Bothrops asper snake venomrdquo BioMed Research Internationalvol 2013 Article ID 789689 9 pages 2013
[11] V X da Silva and M T Rodrigues ldquoTaxonomic revision ofthe Bothrops neuwiedi complex (Serpentes Viperidae) withdescription of a new speciesrdquo Phyllomedusa vol 7 no 1 pp 45ndash90 2008
[12] J A Campbell and W W Lamar The Venomous Reptiles of theWestern Hemisphere Comstock Ithaca NY USA 2004
[13] P Edman ldquoMethod for determination of the amino acidsequence in peptidesrdquo Acta Chemica Scandinavica vol 4 pp283ndash293 1950
[14] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[15] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[16] M Holzer and S P Mackessy ldquoAn aqueous endpoint assay ofsnake venom phospholipase A
2rdquo Toxicon vol 34 no 10 pp
1149ndash1155 1996[17] A L Fuly A L P deMiranda R B Zingali and J AGuimaraes
ldquoPurification and characterization of a phospholipase A2isoen-
zyme isolated from Lachesis muta snake venomrdquo BiochemicalPharmacology vol 63 no 9 pp 1589ndash1597 2002
[18] T Nikai N Mori M Kishida H Sugihara and A T Tu ldquoIso-lation and biochemical characterization of hemorrhagic toxinf from the venom of Crotalus atrox (Western DiamondbackRattlesnake)rdquo Archives of Biochemistry and Biophysics vol 231no 2 pp 309ndash319 1984
[19] J A Gene A Roy G Rojas J M Gutierrez and L CerdasldquoComparative study on coagulant defibrinating fibrinolyticand fibrinogenolytic activities of Costa Rican crotaline snakevenoms and their neutralization by a polyvalent antivenomrdquoToxicon vol 27 no 8 pp 841ndash848 1989
[20] M S Charney and R M Tomarelli ldquoA colorimetric method forthe determination of the proteolytic activity of duodenal juicerdquoThe Journal of Biological Chemistry vol 171 pp 501ndash505 1947
[21] R G Stabeli S F Amui C D SantrsquoAna et al ldquoBothrops moojenimyotoxin-II a Lys49-phospholipaseA
2homologue an example
of function versatility of snake venom proteinsrdquo ComparativeBiochemistry and Physiology C vol 142 no 3-4 pp 371ndash3812006
[22] D R Call J A Nemzek S J Ebong G L Bolgos D ENewcomb and D G Remick ldquoRatio of local to systemicchemokine concentrations regulates neutrophil recruitmentrdquoThe American Journal of Pathology vol 158 no 2 pp 715ndash7212001
[23] P G Roberto S Kashima S Marcussi et al ldquoCloning andidentification of a complete cDNA coding for a bactericidal andantitumoral acidic phospholipase A
2from Bothrops jararacussu
venomrdquoThe Protein Journal vol 23 no 4 pp 273ndash285 2004[24] J H Schumacher A OrsquoGarra B Shrader et al ldquoThe charac-
terization of four monoclonal antibodies specific for mouse IL-5 and development of mouse and human IL-5 enzyme-linkedimmunosorbentrdquoThe Journal of Immunology vol 141 no 5 pp1576ndash1581 1988
[25] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[26] C A T Souza A M Kayano S S Setubal et al ldquoLocal andsystemic biochemical alterations induced by Bothrops atroxsnake venom in micerdquo Journal of Venom Research vol 3 pp28ndash34 2012
[27] J M Gutierrez and B Lomonte ldquoPhospholipase A2myotoxins
from Bothrops snake venomsrdquo Toxicon vol 33 no 11 pp 1405ndash1424 1995
[28] J M Gutierrez and A Rucavado ldquoSnake venom metallopro-teinases their role in the pathogenesis of local tissue damagerdquoBiochimie vol 82 no 9-10 pp 841ndash850 2000
[29] T Matsui Y Fujimura and K Titani ldquoSnake venom proteasesaffecting hemostasis and thrombosisrdquo Biochimica et BiophysicaActa vol 1477 no 1-2 pp 146ndash156 2000
[30] A M Soares M R M Fontes and J R Giglio ldquoPhospholipaseA2myotoxins from Bothrops snake venoms structure-function
12 BioMed Research International
relationshiprdquoCurrent Organic Chemistry vol 8 no 17 pp 1677ndash1690 2004
[31] VM Rodrigues SMarcussi R S Cambraia et al ldquoBactericidaland neurotoxic activities of two myotoxic phospholipases A
2
from Bothrops neuwiedi pauloensis snake venomrdquo Toxicon vol44 no 3 pp 305ndash314 2004
[32] N A Santos-Filho L B Silveira C Z Oliveira et al ldquoAnew acidic myotoxic anti-platelet and prostaglandin I
2induc-
tor phospholipase A2isolated from Bothrops moojeni snake
venomrdquo Toxicon vol 52 no 8 pp 908ndash917 2008[33] A M Soares S H Andriao-Escarso Y Angulo et al ldquoStruc-
tural and functional characterization of myotoxin I a Lys49phospholipaseA
2homologue fromBothropsmoojeni (Caissaca)
snake venomrdquo Archives of Biochemistry and Biophysics vol 373no 1 pp 7ndash15 2000
[34] A M Soares S H Andriao-Escarso R K Bortoleto et alldquoDissociation of enzymatic and pharmacological properties ofpiratoxins-I and -III two myotoxic phospholipases A
2from
Bothrops pirajai snake venomrdquo Archives of Biochemistry andBiophysics vol 387 no 2 pp 188ndash196 2001
[35] S S Teixeira L B Silveira F M N da Silva et al ldquoMolecularcharacterization of an acidic phospholipase A
2from Bothrops
pirajai snake venom synthetic C-terminal peptide identifies itsantiplatelet regionrdquo Archives of Toxicology vol 85 no 10 pp1219ndash1233 2011
[36] S H Andriao-Escarso A M Soares V M Rodrigues et alldquoMyotoxic phospholipases A
2in Bothrops snake venoms effect
of chemical modifications on the enzymatic and pharmacolog-ical properties of bothropstoxins from Bothrops jararacussurdquoBiochimie vol 82 no 8 pp 755ndash763 2000
[37] S H Andriao-Escarso A M Soares M R M Fontes et alldquoStructural and functional characterization of an acidic plateletaggregation inhibitor and hypotensive phospholipase A
2from
Bothrops jararacussu snake venomrdquo Biochemical Pharmacologyvol 64 no 4 pp 723ndash732 2002
[38] M E G Denegri O C Acosta S Huancahuire-Vega et alldquoIsolation and functional characterization of a new acidic PLA
2
Ba SpII RP4 of the Bothrops alternatus snake venom fromArgentinardquo Toxicon vol 56 no 1 pp 64ndash74 2010
[39] R Zouari-Kessentini J Luis A Karray et al ldquoTwo purified andcharacterized phospholipases A
2from Cerastes cerastes venom
that inhibit cancerous cell adhesion and migrationrdquo Toxiconvol 53 no 4 pp 444ndash453 2009
[40] R Shirai M Toriba K Hayashi K Ikeda and S Inoue ldquoIden-tification and characterization of phospholipase A
2inhibitors
from the serum of the Japanese rat snake Elaphe climacophorardquoToxicon vol 53 no 6 pp 685ndash692 2009
[41] M I Homsi-Brandenburgo L S Queiroz H Santo-Neto LRodrigues-Simoni and J R Giglio ldquoFractionation of Bothropsjararacussu snake venom partial chemical characterization andbiological activity of bothropstoxinrdquo Toxicon vol 26 no 7 pp615ndash627 1988
[42] G P Fernandez A Segura M Herrera et al ldquoNeutralizationof Bothrops mattogrossensis snake venom from Bolivia exper-imental evaluation of llama and donkey antivenoms producedby caprylic acid precipitationrdquoToxicon vol 55 no 2-3 pp 642ndash645 2010
[43] C L Ownby H S S de Araujo S P White and J E FletcherldquoLysine 49 phospholipase A
2proteinsrdquo Toxicon vol 37 no 3
pp 411ndash445 1999[44] A M Soares W P Sestito S Marcussi et al ldquoAlkylation of
myotoxic phospholipases A2in Bothrops moojeni venom a
promising approach to an enhanced antivenom productionrdquoInternational Journal of Biochemistry amp Cell Biology vol 36 no2 pp 258ndash270 2004
[45] B Lomonte J M Gutierrez M F Furtado et al ldquoIsolationof basic myotoxins from Bothrops moojeni and Bothrops atroxsnake venomsrdquo Toxicon vol 28 no 10 pp 1137ndash1146 1990
[46] L C Mancuso M M Correa C A Vieira et al ldquoFractionationof Bothrops pirajai snake venom isolation and characterizationof piratoxin-I a new myotoxic proteinrdquo Toxicon vol 33 no 5pp 615ndash626 1995
[47] H S Selistre L S Queiroz O A B Cunha G E P deSouza and J R Giglio ldquoIsolation and characterization ofhemorrhagic myonecrotic and edema-inducing toxins fromBothrops insularis (jararaca ilhoa) snake venomrdquo Toxicon vol28 no 3 pp 261ndash273 1990
[48] E A Dennis J Cao Y-H Hsu V Magrioti and G KokotosldquoPhospholipaseA
2enzymes physical structure biological func-
tion disease implication chemical inhibition and therapeuticinterventionrdquo Chemical Reviews vol 111 no 10 pp 6130ndash61852011
[49] A G T Cardoso Acao do veneno de Bothrops moojeni e suafracao L-aminoacido oxidase submetida ao tratamento comraios gama de 60CO em Leishmania spp [MS thesis] IPEN-CNENSP Sao Paulo Brazil 1999
[50] MGallacci andWLGCavalcante ldquoUnderstanding the in vitroneuromuscular activity of snake venomLys49 phospholipaseA
2
homologuesrdquo Toxicon vol 55 no 1 pp 1ndash11 2010[51] S S Teixeira Caracterizacao funcional e estrutural de uma
fosfolipase A2acida isolada do veneno de Bothrops pirajai
[MS thesis] Faculdade de Ciencias Farmaceuticas de RibeiraoPretoUSP Ribeirao Preto Brazil 2009
[52] E Valentin and G Lambeau ldquoIncreasing molecular diversityof secreted phospholipases A
2and their receptors and binding
proteinsrdquo Biochimica et Biophysica Acta vol 1488 no 1-2 pp59ndash70 2000
[53] J M Gutierrez and B Lomonte ldquoPhospholipases A2from
Viperidae snake venoms how do they induce skeletal muscledamagerdquo Acta Chimica Slovenica vol 58 no 4 pp 647ndash6592011
[54] J M Gutierrez C L Ownby and G V Odell ldquoIsolation of amyotoxin from Bothrops asper venom partial characterizationand action on skeletal musclerdquo Toxicon vol 22 no 1 pp 115ndash128 1984
[55] N G Nascimento M C Sampaio R A Olivo and C TeixeiraldquoContribution of mast cells to the oedema induced by Bothropsmoojeni snake venom and a pharmacological assessment of theinflammatory mediators involvedrdquo Toxicon vol 55 no 2-3 pp343ndash352 2010
[56] C F P Teixeira E C T Landucci E Antunes M Chacurand Y Cury ldquoInflammatory effects of snake venom myotoxicphospholipases A
2rdquo Toxicon vol 42 no 8 pp 947ndash962 2003
[57] S R Zamuner and C F P Teixeira ldquoCell adhesion moleculesinvolved in the leukocyte recruitment induced by venom of thesnake Bothrops jararacardquoMediators of Inflammation vol 11 no6 pp 351ndash357 2002
[58] F Chaves C F P Teixeira and J M Gutierrez ldquoRole ofTNF-120572 IL-1120573 and IL-6 in the local tissue damage induced byBothrops asper snake venom an experimental assessment inmicerdquo Toxicon vol 45 no 2 pp 171ndash178 2005
BioMed Research International 13
[59] C D F P Teixeira C M Fernandes J P Zuliani and SF Zamuner ldquoInflammatory effects of snake venom metallo-proteinasesrdquo Memorias do Instituto Oswaldo Cruz vol 100supplement 1 pp 181ndash184 2005
[60] R D A Olivo C F P Teixeira J LWallace J M Gutierrez andS R Zamuner ldquoRole of cyclooxygenases in oedema-formingactivity of bothropic venomsrdquo Toxicon vol 49 no 5 pp 670ndash677 2007
[61] L B Silveira D P Marchi-Salvador N A Santos-Filho et alldquoIsolation and expression of a hypotensive and anti-plateletacidic phospholipase A
2from Bothrops moojeni snake venomrdquo
Journal of Pharmaceutical and Biomedical Analysis vol 73 pp35ndash43 2013
[62] A Rucavado T Escalante C F P Teixeira C M Fernandes CDıaz and J M Gutierrez ldquoIncrements in cytokines and matrixmetalloproteinases in skeletal muscle after injection of tissue-damaging toxins from the venom of the snake Bothrops asperrdquoMediators of Inflammation vol 11 no 2 pp 121ndash128 2002
[63] E Leiguez J P Zuliani A M Cianciarullo C M Fernandes JM Gutierrez and C Teixeira ldquoA group IIA-secreted phospho-lipase A
2from snake venom induces lipid body formation in
macrophages the roles of intracellular phospholipases A2and
distinct signaling pathwaysrdquo Journal of Leukocyte Biology vol90 no 1 pp 155ndash166 2011
[64] J P Zuliani J M Gutierrez L L Casais e Silva S C SampaioB Lomonte and C D F P Teixeira ldquoActivation of cellularfunctions in macrophages by venom secretory Asp-49 and Lys-49 phospholipasesA
2rdquoToxicon vol 46 no 5 pp 523ndash532 2005
[65] R D C M Escocard M M Kanashiro J H Petretski et alldquoNeutrophils regulate the expression of cytokines chemokinesand nitric oxide synthasenitric oxide in mice injected withBothrops atrox venomrdquo Immunobiology vol 211 no 1-2 pp 37ndash46 2006
[66] A Gambero E C T Landucci M H Toyama et al ldquoHumanneutrophil migration in vitro induced by secretory phospholi-pases A
2 a role for cell surface glycosaminoglycansrdquo Biochemi-
cal Pharmacology vol 63 no 1 pp 65ndash72 2002[67] F L S Costa R S Rodrigues L F M Izidoro et al ldquoBio-
chemical and functional properties of a thrombin-like enzymeisolated from Bothrops pauloensis snake venomrdquo Toxicon vol54 no 6 pp 725ndash735 2009
[68] J C Villalobos R Mora B Lomonte J M Gutierrez andY Angulo ldquoCytotoxicity induced in myotubes by a Lys49phospholipase A
2homologue from the venom of the snake
Bothrops asper evidence of rapid plasma membrane damageand a dual role for extracellular calciumrdquo Toxicology in Vitrovol 21 no 8 pp 1382ndash1389 2007
[69] B S Cummings J McHowat and R G Schnellmann ldquoPhos-pholipase A
2s in cell injury and deathrdquoThe Journal of Pharma-
cology and Experimental Therapeutics vol 294 no 3 pp 793ndash799 2000
[70] S R Panini L Yang A E Rusinol M S Sinensky J VBonventre and C C Leslie ldquoArachidonate metabolism andthe signaling pathway of induction of apoptosis by oxidizedLDLoxysterolrdquo Journal of Lipid Research vol 42 no 10 pp1678ndash1686 2001
[71] L Paramo B Lomonte J Pizarro-Cerda J-A Bengoechea J-P Gorvel and E Moreno ldquoBactericidal activity of Lys49 andAsp49 myotoxic phospholipases A
2from Bothrops asper snake
venom synthetic Lys49myotoxin II-(115-129)-peptide identifiesits bactericidal regionrdquo European Journal of Biochemistry vol253 no 2 pp 452ndash461 1998
[72] B Lomonte Y Angulo and E Moreno ldquoSynthetic peptidesderived from the C-terminal region of Lys49 phospholipaseA2homologues from viperidae snake venoms biomimetic
activities and potential applicationsrdquo Current PharmaceuticalDesign vol 16 no 28 pp 3224ndash3230 2010
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

8 BioMed Research International
lowastlowast lowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowastlowast lowastlowastlowastlowastlowastlowastlowastlowastlowast
1 10 20 30 40 50(Identity )
100
89
92
92
92
94
94
BmatTX-IBmatTX-IIPrtx-IBOJU-IBthTX-IMjtx-IMtx-Ii
(a)
lowastlowastlowast lowastlowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowastlowast lowastlowastlowast
1 10 20 30 40 50
(Identity )100
78
75
71
65
BmatTX-IIIBmoTX-IBmjeTX-IBthTX-IIPrTX-III
(b)
Figure 4 Comparison of the N-terminal sequence of the snakersquos PLA2s with the PLA
2s isolated from B mattogrossensis venom N-terminal
sequencing of the PLA2s obtained by Edman [13] degradation and alignment done with CLUSTALW2 expressed as of similarity (a)
BmatTX-I BmatTX-II with Lys49 residues in marked sequences (b) BmatTX-III with Asp49 residues in marked sequences Sequencesused for the alignment and their respective access numbers BthTX-I (gi51890398) MjTX-I (gi17368325) Mtx-II (gi390981003) PrTX-I (gi190016174) BOJU-I (gi209572966) BthTX-II (gi1171971) PrTX-III (gi90016174) BmjeTX-I (gi313471399) BmoTX-I (gi221272396)BthTX-I (Lys49) BthTX-II (Asp49) and BOJU-I (Lys49) isolated from the venom of B jararacussu MjTX-I (Lys49) BmoTX-I (Asp49)and BmjeTX-I (Asp49) isolated from the venom of B moojeni Mtx-II (Lys49) isolated from the venom of B brazili PrTX-I (Lys49) andPrTX-III (Asp49) isolated from the venom of B pirajai the ldquolowastrdquo was used to indicate the amino acid residues that are the same between thesequences
other molecular regions distinct from the active site may beinvolved in this effect
PLA2s are multifunctional proteins that can be used as
mediators in several areas of medicine such as in the treat-ment of rheumatoid arthritis as a new class of HIV inhibitorsby blocking the host cell invasion as a potential treatmentagainst malaria and as an antibiotic by inducing cytotoxicityvia the disruption of bacterial membranes [21 23 30 6768] A study by Costa et al [67] with PLA
2s isolated from
B brazili MTX-I and II demonstrated cytotoxic activityagainst Jurkat tumor cells as well as antimicrobial activityagainstE coli andC albicans and antiparasitic activity againstLeishmania sp
In the present study we evaluated the cytotoxic activityof PLA
2s isolated from B mattogrossensis on JURKAT (T
leukemia) and SK-BR-3 (breast adenocarcinoma) both ofwhich are human tumor cell lines Like MTX-II of B brazili[67] the cytotoxic activity of PLA
2s BmatTX-I and BmatTX-
II on JURKATcells was independent of their catalytic activitysince these are characterized as Lys49 PLA
2s and are therefore
catalytically inactive BmatTX-III characterized as Asp49despite being enzymatically active also showed a lower level oftoxicity Some authors propose that the cytotoxic activity ontumor cell lines is associated with the induction of apoptosisconsidering the fact that PLA
2promotes alterations in the
cell membrane And some studies involving Lys49 PLA2s
isolated from B asper demonstrated that the C-terminal
region comprised of amino acids 115ndash129 is concerned withthe cytotoxic and bactericidal activities of this protein [69ndash71] The same observation was made by Costa et al [67] withsynthetic peptides derived from the C-terminal portion ofMTX-I and II PLA
2s
We also evaluated the antiparasitic activity of the crudevenom and isolated PLA
2of B mattogrossensis on promastig-
ote forms of L amazonensis After the analysis it wasobserved that the crude venom of B mattogrossensis presentsincreasing toxic activity (approximately 50 to 80) againstpromastigote forms of L amazonensis after 48 h of incubation(Figure 7(a)) The PLA
2s isolated from B mattogrossensis at
100 120583gmL characterized as BmatTX-I (Lys49) and BmatTX-III (Asp49) presented toxic activity between 25 and 30respectively even with values close to those presented afterincubation of the protozoan with Pentamidine a drug usedas a positive control (Figures 7(b) and 7(c))
When compared the cytotoxicity values of PLA2s against
promastigote forms of L amazonensis show similar activitybetween the Lys49-PLA
2and the Asp49-PLA
2 Comparison
of this activity with the crude venom (70) showed thatPLA2s are responsible for almost half of the observed effect
Nonetheless notably the results suggest that other toxinspresent in the venom contribute to the parasitersquos death
Similar to the results obtained in the present studyStabeli et al [21] demonstrated that MjTX-II a Lys49-PLA
2
homologue isolated from B moojeni venom in different
BioMed Research International 9
0
500
1000
1500
2000
lowast
lowast
lowast
lowast
lowast
Control 3 6 12 3 6 12 3 6 12
ControlBmatTX-I (Lys49)
BmatTX-II (Lys49)BmatTX-III (Asp49)
BmatTX-I (120583g) BmatTX-II (120583g) BmatTX-III (120583g)
IL-1120573
(pg
mL)
(a)
Control 3 6 12 3 6 12 3 6 120
500
1000
1500
2000
ControlBmatTX-I (Lys49)
BmatTX-II (Lys49)BmatTX-III (Asp49)
lowastlowastlowast
BmatTX-I (120583g) BmatTX-II (120583g) BmatTX-III (120583g)
IL-1120573
(pg
mL)
(b)
Figure 5 Production of IL-1120573 by mice neutrophils These cells wereincubatedwith PLA
2(3 6 and 12 120583gmL) or RPMI (control) for 12
hours (a) and 24 hours (b) at 37∘C in a humid atmosphere with 5CO2 Quantification was done by ELISA as described in 29 Each
bar represents the average +minus SDof three independent groups lowast119875 lt005 compared to the control
concentrations (5 25 and 75 120583g) was effective as a parasiticideagent against Schistosomamansoni and promastigote forms ofLeishmania (L amazonensis L braziliensis L donovani andL major)
The action of the PLA2s BmatTX-I (Lys49) and
BmatTX-III (Asp49) on promastigote forms of L ama-zonensis was independent of its catalytic activity sincecatalytically inactive Lys49 myotoxin also demonstrated tox-icity against Leishmania It is believed that the observedcytotoxic activity might be related to the C-terminal regionsof these phospholipase-homologues that are able to promotea disturbance in the cellular membranes independent oftheir catalytic activity [67 72] However more studies arenecessary to define the exact mechanism of action of theseenzymes on parasites
Growing interest in the comprehension of the structureand function of snake venom components especially PLA
2
10 100 10 100 10 100 10 1000
20
40
60
80
100
BmatTX-I (Lys49)BmatTX-II (Lys49)
BmatTX-III (Asp49)Methotrexate (MTX)
Cyto
toxi
city
()
Concentration (120583gmL)
(a)
10 100 10 100 10 100 10 1000
20
40
60
80
100
BmatTX-I (Lys49)BmatTX-II (Lys49)
BmatTX-III (Asp49)Methotrexate (MTX)
Cyto
toxi
city
()
Concentration (120583gmL)
(b)
Figure 6 Antitumoral activity of PLA2s from B mattogrossensis
(a) Antitumoral activity on human acute T-cell leukemia (JURKAT)lines (b) Antitumoral activity on human breast adenocarcinoma(SK-BR-3) Different concentrations of the PLA
2s were incubated
with cell linesMethotrexate was used as the positive control Resultsare presented as mean +minus SD (119899 = 3)
contributes to a better understanding of the mechanism ofaction of their enzymatic and toxic activities It opens thepath to better understand the intoxication caused by enven-omation and the physiopathology behind its side effectsFuture studies will potentially improve serum therapy andhelp develop the pharmaceutical potential that moleculesisolated from animal venoms can have such as the PLA
2s
isolated from the B mattogrossensis venom which show anti-Leishmania and antitumor activities
4 Conclusion
In conclusion the venom of Bothrops mattogrossensis hasa qualitatively similar toxicological profile to previously
10 BioMed Research International
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineB mattogrossensis crude venom
Ant
i-leis
hman
ia ac
tivity
()
B mattogrossensis crude venom (120583g)
(a)
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineBmatTX-I (Lys49)
Ant
i-leis
hman
ia ac
tivity
()
BmatTX-I (120583g)
lowastlowast lowast lowast
(b)
Ant
i-leis
hman
ia ac
tivity
()
Pent 36 625 125 25 50 1000
20
40
60
80
100
BmatTX-III (Asp49)Pentamidine
BmatTX-III (120583g)
lowastlowastlowast
lowastlowast lowast
(c)
Figure 7 Antileishmanial activity of the crude venom of and PLA2s from B mattogrossensis The promastigote forms of L amazonensiswere
plated with 1times105 cellswellThen different concentrations of crude venom and isolated PLA2s were added to each wellThe experiment was
done in a 48 h period with the (a) crude venom and the PLA2enzymes (b) BmatTX-I and (c) BmatTX-III MTT was added and after the
incubation period at 33∘C the formazan crystal formed was dissolved in SDS Readings were done in a spectrophotometer at 570 nm Eachbar represents the average +minus SD of the three independent experimental groups sixfold total lowast119875 lt 005 compared to the control
studied snake venoms of the Bothrops sp genera despite theobservation of quantitative variations Of the three basicPLA2s from B mattogrossensis venom now isolated for the
first time two are characterized as Lys49-PLA2homologues
BmatTX-I and -II and the other as an Asp49-PLA2 named
BmatTX-III This showed high phospholipase activity ThePLA2s isolated induced myotoxic effects as well as the release
of proinflammatory cytokines by neutrophils BmatTX-I and-III PLA
2s were cytotoxic to human tumor cell lines JURKAT
and SK-BR-3 and showed activity against promastigote formsof L amazonensis
Conflict of Interests
The authors state that there is no conflict of interests
Acknowledgments
The authors are grateful to the Ministry of Science andTechnology (MCTI) Conselho Nacional de Desenvolvi-mento Cientıfico e Tecnologico (CNPq) Financiadora deEstudos e Projetos (FINEP) Fundacao de Tecnologia doAcre (FUNTACFDCT) Coordenacao de Aperfeicoamentode Nıvel Superior (CAPES)mdashProjeto NanoBiotec Redede Biodiversidade e Biotecnologia da Amazonia Legal(BIONORTECNPqMCTI) Instituto Nacional paraPesquisa Translacional em Saude e Ambiente na RegiaoAmazonica (INCT-INPeTAmCNPqMCTI) and InstitutoNacional para Pesquisa em Toxinas (INCT-Tox) andSecretary of Development of Rondonia State (SEPLANPRONEXCNPq) for financial support We are grateful to
BioMed Research International 11
Sr Guilherme G Galassi (Mantenedouro da Fauna Silvestreautorizacao IBAMA 020270089912001-95 Americana-SP)for their helpful technical collaboration in provide the snakevenom Amy Grabner provided the English editing of thepaper
References
[1] Y Angulo and B Lomonte ldquoBiochemistry and toxicology oftoxins purified from the venom of the snake Bothrops asperrdquoToxicon vol 54 no 7 pp 949ndash957 2009
[2] J J Calvete P Juarez and L Sanz ldquoSnake venomics Strategyand applicationsrdquo Journal of Mass Spectrometry vol 42 no 11pp 1405ndash1414 2007
[3] T S Kang D Georgieva N Genov et al ldquoEnzymatic toxinsfrom snake venom structural characterization and mechanismof catalysisrdquoThe FEBS Journal vol 278 no 23 pp 4544ndash45762011
[4] C ZOliveiraCaracterizacao funcional e estrutural de inibidoresde fosfolipases A
2isolados do plasma de serpente Bothrops
jararacussu [PhD thesis] Faculdade de Ciencias Farmaceuticasde Ribeirao Preto Ribeirao Preto Brazil 2009
[5] R G Stabeli R Simoes-Silva A M Kayano et al ldquoPurifi-cation of phospholipases A
2from American snake venomsrdquo
in ChromatographymdashThe Most Versatile Method of ChemicalAnalysis chapter 1 pp 1ndash34 InTech Vienna Austria 2012
[6] J P Zuliani C M Fernandes S R Zamuner J M Gutierrezand C F P Teixeira ldquoInflammatory events induced by Lys-49 and Asp-49 phospholipases A
2isolated from Bothrops asper
snake venom role of catalytic activityrdquo Toxicon vol 45 no 3pp 335ndash346 2005
[7] S Marcussi R G Stabeli N A Santos-Filho et al ldquoGenotoxiceffect of Bothrops snake venoms and isolated toxins on humanlymphocyte DNArdquo Toxicon vol 65 pp 9ndash14 2013
[8] S S Setubal A S Pontes J L Furtado et al ldquoAction oftwo phospholipases A
2purified from Bothrops alternatus snake
venom on macrophagesrdquo Biochemistry vol 78 no 2 pp 194ndash203 2013
[9] J M Gutierrez L A Ponce-Soto S Marangoni and BLomonte ldquoSystemic and local myotoxicity induced by snakevenom group II phospholipases A
2 comparison between cro-
toxin crotoxin B and a Lys49 PLA2homologuerdquo Toxicon vol
51 no 1 pp 80ndash92 2008[10] A Q Rueda I G Rodrıguez E C Ara et al ldquoBiochemical
characterization action onmacrophages and superoxide anionproduction of four basic phospholipases A
2from Panamanian
Bothrops asper snake venomrdquo BioMed Research Internationalvol 2013 Article ID 789689 9 pages 2013
[11] V X da Silva and M T Rodrigues ldquoTaxonomic revision ofthe Bothrops neuwiedi complex (Serpentes Viperidae) withdescription of a new speciesrdquo Phyllomedusa vol 7 no 1 pp 45ndash90 2008
[12] J A Campbell and W W Lamar The Venomous Reptiles of theWestern Hemisphere Comstock Ithaca NY USA 2004
[13] P Edman ldquoMethod for determination of the amino acidsequence in peptidesrdquo Acta Chemica Scandinavica vol 4 pp283ndash293 1950
[14] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[15] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[16] M Holzer and S P Mackessy ldquoAn aqueous endpoint assay ofsnake venom phospholipase A
2rdquo Toxicon vol 34 no 10 pp
1149ndash1155 1996[17] A L Fuly A L P deMiranda R B Zingali and J AGuimaraes
ldquoPurification and characterization of a phospholipase A2isoen-
zyme isolated from Lachesis muta snake venomrdquo BiochemicalPharmacology vol 63 no 9 pp 1589ndash1597 2002
[18] T Nikai N Mori M Kishida H Sugihara and A T Tu ldquoIso-lation and biochemical characterization of hemorrhagic toxinf from the venom of Crotalus atrox (Western DiamondbackRattlesnake)rdquo Archives of Biochemistry and Biophysics vol 231no 2 pp 309ndash319 1984
[19] J A Gene A Roy G Rojas J M Gutierrez and L CerdasldquoComparative study on coagulant defibrinating fibrinolyticand fibrinogenolytic activities of Costa Rican crotaline snakevenoms and their neutralization by a polyvalent antivenomrdquoToxicon vol 27 no 8 pp 841ndash848 1989
[20] M S Charney and R M Tomarelli ldquoA colorimetric method forthe determination of the proteolytic activity of duodenal juicerdquoThe Journal of Biological Chemistry vol 171 pp 501ndash505 1947
[21] R G Stabeli S F Amui C D SantrsquoAna et al ldquoBothrops moojenimyotoxin-II a Lys49-phospholipaseA
2homologue an example
of function versatility of snake venom proteinsrdquo ComparativeBiochemistry and Physiology C vol 142 no 3-4 pp 371ndash3812006
[22] D R Call J A Nemzek S J Ebong G L Bolgos D ENewcomb and D G Remick ldquoRatio of local to systemicchemokine concentrations regulates neutrophil recruitmentrdquoThe American Journal of Pathology vol 158 no 2 pp 715ndash7212001
[23] P G Roberto S Kashima S Marcussi et al ldquoCloning andidentification of a complete cDNA coding for a bactericidal andantitumoral acidic phospholipase A
2from Bothrops jararacussu
venomrdquoThe Protein Journal vol 23 no 4 pp 273ndash285 2004[24] J H Schumacher A OrsquoGarra B Shrader et al ldquoThe charac-
terization of four monoclonal antibodies specific for mouse IL-5 and development of mouse and human IL-5 enzyme-linkedimmunosorbentrdquoThe Journal of Immunology vol 141 no 5 pp1576ndash1581 1988
[25] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[26] C A T Souza A M Kayano S S Setubal et al ldquoLocal andsystemic biochemical alterations induced by Bothrops atroxsnake venom in micerdquo Journal of Venom Research vol 3 pp28ndash34 2012
[27] J M Gutierrez and B Lomonte ldquoPhospholipase A2myotoxins
from Bothrops snake venomsrdquo Toxicon vol 33 no 11 pp 1405ndash1424 1995
[28] J M Gutierrez and A Rucavado ldquoSnake venom metallopro-teinases their role in the pathogenesis of local tissue damagerdquoBiochimie vol 82 no 9-10 pp 841ndash850 2000
[29] T Matsui Y Fujimura and K Titani ldquoSnake venom proteasesaffecting hemostasis and thrombosisrdquo Biochimica et BiophysicaActa vol 1477 no 1-2 pp 146ndash156 2000
[30] A M Soares M R M Fontes and J R Giglio ldquoPhospholipaseA2myotoxins from Bothrops snake venoms structure-function
12 BioMed Research International
relationshiprdquoCurrent Organic Chemistry vol 8 no 17 pp 1677ndash1690 2004
[31] VM Rodrigues SMarcussi R S Cambraia et al ldquoBactericidaland neurotoxic activities of two myotoxic phospholipases A
2
from Bothrops neuwiedi pauloensis snake venomrdquo Toxicon vol44 no 3 pp 305ndash314 2004
[32] N A Santos-Filho L B Silveira C Z Oliveira et al ldquoAnew acidic myotoxic anti-platelet and prostaglandin I
2induc-
tor phospholipase A2isolated from Bothrops moojeni snake
venomrdquo Toxicon vol 52 no 8 pp 908ndash917 2008[33] A M Soares S H Andriao-Escarso Y Angulo et al ldquoStruc-
tural and functional characterization of myotoxin I a Lys49phospholipaseA
2homologue fromBothropsmoojeni (Caissaca)
snake venomrdquo Archives of Biochemistry and Biophysics vol 373no 1 pp 7ndash15 2000
[34] A M Soares S H Andriao-Escarso R K Bortoleto et alldquoDissociation of enzymatic and pharmacological properties ofpiratoxins-I and -III two myotoxic phospholipases A
2from
Bothrops pirajai snake venomrdquo Archives of Biochemistry andBiophysics vol 387 no 2 pp 188ndash196 2001
[35] S S Teixeira L B Silveira F M N da Silva et al ldquoMolecularcharacterization of an acidic phospholipase A
2from Bothrops
pirajai snake venom synthetic C-terminal peptide identifies itsantiplatelet regionrdquo Archives of Toxicology vol 85 no 10 pp1219ndash1233 2011
[36] S H Andriao-Escarso A M Soares V M Rodrigues et alldquoMyotoxic phospholipases A
2in Bothrops snake venoms effect
of chemical modifications on the enzymatic and pharmacolog-ical properties of bothropstoxins from Bothrops jararacussurdquoBiochimie vol 82 no 8 pp 755ndash763 2000
[37] S H Andriao-Escarso A M Soares M R M Fontes et alldquoStructural and functional characterization of an acidic plateletaggregation inhibitor and hypotensive phospholipase A
2from
Bothrops jararacussu snake venomrdquo Biochemical Pharmacologyvol 64 no 4 pp 723ndash732 2002
[38] M E G Denegri O C Acosta S Huancahuire-Vega et alldquoIsolation and functional characterization of a new acidic PLA
2
Ba SpII RP4 of the Bothrops alternatus snake venom fromArgentinardquo Toxicon vol 56 no 1 pp 64ndash74 2010
[39] R Zouari-Kessentini J Luis A Karray et al ldquoTwo purified andcharacterized phospholipases A
2from Cerastes cerastes venom
that inhibit cancerous cell adhesion and migrationrdquo Toxiconvol 53 no 4 pp 444ndash453 2009
[40] R Shirai M Toriba K Hayashi K Ikeda and S Inoue ldquoIden-tification and characterization of phospholipase A
2inhibitors
from the serum of the Japanese rat snake Elaphe climacophorardquoToxicon vol 53 no 6 pp 685ndash692 2009
[41] M I Homsi-Brandenburgo L S Queiroz H Santo-Neto LRodrigues-Simoni and J R Giglio ldquoFractionation of Bothropsjararacussu snake venom partial chemical characterization andbiological activity of bothropstoxinrdquo Toxicon vol 26 no 7 pp615ndash627 1988
[42] G P Fernandez A Segura M Herrera et al ldquoNeutralizationof Bothrops mattogrossensis snake venom from Bolivia exper-imental evaluation of llama and donkey antivenoms producedby caprylic acid precipitationrdquoToxicon vol 55 no 2-3 pp 642ndash645 2010
[43] C L Ownby H S S de Araujo S P White and J E FletcherldquoLysine 49 phospholipase A
2proteinsrdquo Toxicon vol 37 no 3
pp 411ndash445 1999[44] A M Soares W P Sestito S Marcussi et al ldquoAlkylation of
myotoxic phospholipases A2in Bothrops moojeni venom a
promising approach to an enhanced antivenom productionrdquoInternational Journal of Biochemistry amp Cell Biology vol 36 no2 pp 258ndash270 2004
[45] B Lomonte J M Gutierrez M F Furtado et al ldquoIsolationof basic myotoxins from Bothrops moojeni and Bothrops atroxsnake venomsrdquo Toxicon vol 28 no 10 pp 1137ndash1146 1990
[46] L C Mancuso M M Correa C A Vieira et al ldquoFractionationof Bothrops pirajai snake venom isolation and characterizationof piratoxin-I a new myotoxic proteinrdquo Toxicon vol 33 no 5pp 615ndash626 1995
[47] H S Selistre L S Queiroz O A B Cunha G E P deSouza and J R Giglio ldquoIsolation and characterization ofhemorrhagic myonecrotic and edema-inducing toxins fromBothrops insularis (jararaca ilhoa) snake venomrdquo Toxicon vol28 no 3 pp 261ndash273 1990
[48] E A Dennis J Cao Y-H Hsu V Magrioti and G KokotosldquoPhospholipaseA
2enzymes physical structure biological func-
tion disease implication chemical inhibition and therapeuticinterventionrdquo Chemical Reviews vol 111 no 10 pp 6130ndash61852011
[49] A G T Cardoso Acao do veneno de Bothrops moojeni e suafracao L-aminoacido oxidase submetida ao tratamento comraios gama de 60CO em Leishmania spp [MS thesis] IPEN-CNENSP Sao Paulo Brazil 1999
[50] MGallacci andWLGCavalcante ldquoUnderstanding the in vitroneuromuscular activity of snake venomLys49 phospholipaseA
2
homologuesrdquo Toxicon vol 55 no 1 pp 1ndash11 2010[51] S S Teixeira Caracterizacao funcional e estrutural de uma
fosfolipase A2acida isolada do veneno de Bothrops pirajai
[MS thesis] Faculdade de Ciencias Farmaceuticas de RibeiraoPretoUSP Ribeirao Preto Brazil 2009
[52] E Valentin and G Lambeau ldquoIncreasing molecular diversityof secreted phospholipases A
2and their receptors and binding
proteinsrdquo Biochimica et Biophysica Acta vol 1488 no 1-2 pp59ndash70 2000
[53] J M Gutierrez and B Lomonte ldquoPhospholipases A2from
Viperidae snake venoms how do they induce skeletal muscledamagerdquo Acta Chimica Slovenica vol 58 no 4 pp 647ndash6592011
[54] J M Gutierrez C L Ownby and G V Odell ldquoIsolation of amyotoxin from Bothrops asper venom partial characterizationand action on skeletal musclerdquo Toxicon vol 22 no 1 pp 115ndash128 1984
[55] N G Nascimento M C Sampaio R A Olivo and C TeixeiraldquoContribution of mast cells to the oedema induced by Bothropsmoojeni snake venom and a pharmacological assessment of theinflammatory mediators involvedrdquo Toxicon vol 55 no 2-3 pp343ndash352 2010
[56] C F P Teixeira E C T Landucci E Antunes M Chacurand Y Cury ldquoInflammatory effects of snake venom myotoxicphospholipases A
2rdquo Toxicon vol 42 no 8 pp 947ndash962 2003
[57] S R Zamuner and C F P Teixeira ldquoCell adhesion moleculesinvolved in the leukocyte recruitment induced by venom of thesnake Bothrops jararacardquoMediators of Inflammation vol 11 no6 pp 351ndash357 2002
[58] F Chaves C F P Teixeira and J M Gutierrez ldquoRole ofTNF-120572 IL-1120573 and IL-6 in the local tissue damage induced byBothrops asper snake venom an experimental assessment inmicerdquo Toxicon vol 45 no 2 pp 171ndash178 2005
BioMed Research International 13
[59] C D F P Teixeira C M Fernandes J P Zuliani and SF Zamuner ldquoInflammatory effects of snake venom metallo-proteinasesrdquo Memorias do Instituto Oswaldo Cruz vol 100supplement 1 pp 181ndash184 2005
[60] R D A Olivo C F P Teixeira J LWallace J M Gutierrez andS R Zamuner ldquoRole of cyclooxygenases in oedema-formingactivity of bothropic venomsrdquo Toxicon vol 49 no 5 pp 670ndash677 2007
[61] L B Silveira D P Marchi-Salvador N A Santos-Filho et alldquoIsolation and expression of a hypotensive and anti-plateletacidic phospholipase A
2from Bothrops moojeni snake venomrdquo
Journal of Pharmaceutical and Biomedical Analysis vol 73 pp35ndash43 2013
[62] A Rucavado T Escalante C F P Teixeira C M Fernandes CDıaz and J M Gutierrez ldquoIncrements in cytokines and matrixmetalloproteinases in skeletal muscle after injection of tissue-damaging toxins from the venom of the snake Bothrops asperrdquoMediators of Inflammation vol 11 no 2 pp 121ndash128 2002
[63] E Leiguez J P Zuliani A M Cianciarullo C M Fernandes JM Gutierrez and C Teixeira ldquoA group IIA-secreted phospho-lipase A
2from snake venom induces lipid body formation in
macrophages the roles of intracellular phospholipases A2and
distinct signaling pathwaysrdquo Journal of Leukocyte Biology vol90 no 1 pp 155ndash166 2011
[64] J P Zuliani J M Gutierrez L L Casais e Silva S C SampaioB Lomonte and C D F P Teixeira ldquoActivation of cellularfunctions in macrophages by venom secretory Asp-49 and Lys-49 phospholipasesA
2rdquoToxicon vol 46 no 5 pp 523ndash532 2005
[65] R D C M Escocard M M Kanashiro J H Petretski et alldquoNeutrophils regulate the expression of cytokines chemokinesand nitric oxide synthasenitric oxide in mice injected withBothrops atrox venomrdquo Immunobiology vol 211 no 1-2 pp 37ndash46 2006
[66] A Gambero E C T Landucci M H Toyama et al ldquoHumanneutrophil migration in vitro induced by secretory phospholi-pases A
2 a role for cell surface glycosaminoglycansrdquo Biochemi-
cal Pharmacology vol 63 no 1 pp 65ndash72 2002[67] F L S Costa R S Rodrigues L F M Izidoro et al ldquoBio-
chemical and functional properties of a thrombin-like enzymeisolated from Bothrops pauloensis snake venomrdquo Toxicon vol54 no 6 pp 725ndash735 2009
[68] J C Villalobos R Mora B Lomonte J M Gutierrez andY Angulo ldquoCytotoxicity induced in myotubes by a Lys49phospholipase A
2homologue from the venom of the snake
Bothrops asper evidence of rapid plasma membrane damageand a dual role for extracellular calciumrdquo Toxicology in Vitrovol 21 no 8 pp 1382ndash1389 2007
[69] B S Cummings J McHowat and R G Schnellmann ldquoPhos-pholipase A
2s in cell injury and deathrdquoThe Journal of Pharma-
cology and Experimental Therapeutics vol 294 no 3 pp 793ndash799 2000
[70] S R Panini L Yang A E Rusinol M S Sinensky J VBonventre and C C Leslie ldquoArachidonate metabolism andthe signaling pathway of induction of apoptosis by oxidizedLDLoxysterolrdquo Journal of Lipid Research vol 42 no 10 pp1678ndash1686 2001
[71] L Paramo B Lomonte J Pizarro-Cerda J-A Bengoechea J-P Gorvel and E Moreno ldquoBactericidal activity of Lys49 andAsp49 myotoxic phospholipases A
2from Bothrops asper snake
venom synthetic Lys49myotoxin II-(115-129)-peptide identifiesits bactericidal regionrdquo European Journal of Biochemistry vol253 no 2 pp 452ndash461 1998
[72] B Lomonte Y Angulo and E Moreno ldquoSynthetic peptidesderived from the C-terminal region of Lys49 phospholipaseA2homologues from viperidae snake venoms biomimetic
activities and potential applicationsrdquo Current PharmaceuticalDesign vol 16 no 28 pp 3224ndash3230 2010
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

BioMed Research International 9
0
500
1000
1500
2000
lowast
lowast
lowast
lowast
lowast
Control 3 6 12 3 6 12 3 6 12
ControlBmatTX-I (Lys49)
BmatTX-II (Lys49)BmatTX-III (Asp49)
BmatTX-I (120583g) BmatTX-II (120583g) BmatTX-III (120583g)
IL-1120573
(pg
mL)
(a)
Control 3 6 12 3 6 12 3 6 120
500
1000
1500
2000
ControlBmatTX-I (Lys49)
BmatTX-II (Lys49)BmatTX-III (Asp49)
lowastlowastlowast
BmatTX-I (120583g) BmatTX-II (120583g) BmatTX-III (120583g)
IL-1120573
(pg
mL)
(b)
Figure 5 Production of IL-1120573 by mice neutrophils These cells wereincubatedwith PLA
2(3 6 and 12 120583gmL) or RPMI (control) for 12
hours (a) and 24 hours (b) at 37∘C in a humid atmosphere with 5CO2 Quantification was done by ELISA as described in 29 Each
bar represents the average +minus SDof three independent groups lowast119875 lt005 compared to the control
concentrations (5 25 and 75 120583g) was effective as a parasiticideagent against Schistosomamansoni and promastigote forms ofLeishmania (L amazonensis L braziliensis L donovani andL major)
The action of the PLA2s BmatTX-I (Lys49) and
BmatTX-III (Asp49) on promastigote forms of L ama-zonensis was independent of its catalytic activity sincecatalytically inactive Lys49 myotoxin also demonstrated tox-icity against Leishmania It is believed that the observedcytotoxic activity might be related to the C-terminal regionsof these phospholipase-homologues that are able to promotea disturbance in the cellular membranes independent oftheir catalytic activity [67 72] However more studies arenecessary to define the exact mechanism of action of theseenzymes on parasites
Growing interest in the comprehension of the structureand function of snake venom components especially PLA
2
10 100 10 100 10 100 10 1000
20
40
60
80
100
BmatTX-I (Lys49)BmatTX-II (Lys49)
BmatTX-III (Asp49)Methotrexate (MTX)
Cyto
toxi
city
()
Concentration (120583gmL)
(a)
10 100 10 100 10 100 10 1000
20
40
60
80
100
BmatTX-I (Lys49)BmatTX-II (Lys49)
BmatTX-III (Asp49)Methotrexate (MTX)
Cyto
toxi
city
()
Concentration (120583gmL)
(b)
Figure 6 Antitumoral activity of PLA2s from B mattogrossensis
(a) Antitumoral activity on human acute T-cell leukemia (JURKAT)lines (b) Antitumoral activity on human breast adenocarcinoma(SK-BR-3) Different concentrations of the PLA
2s were incubated
with cell linesMethotrexate was used as the positive control Resultsare presented as mean +minus SD (119899 = 3)
contributes to a better understanding of the mechanism ofaction of their enzymatic and toxic activities It opens thepath to better understand the intoxication caused by enven-omation and the physiopathology behind its side effectsFuture studies will potentially improve serum therapy andhelp develop the pharmaceutical potential that moleculesisolated from animal venoms can have such as the PLA
2s
isolated from the B mattogrossensis venom which show anti-Leishmania and antitumor activities
4 Conclusion
In conclusion the venom of Bothrops mattogrossensis hasa qualitatively similar toxicological profile to previously
10 BioMed Research International
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineB mattogrossensis crude venom
Ant
i-leis
hman
ia ac
tivity
()
B mattogrossensis crude venom (120583g)
(a)
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineBmatTX-I (Lys49)
Ant
i-leis
hman
ia ac
tivity
()
BmatTX-I (120583g)
lowastlowast lowast lowast
(b)
Ant
i-leis
hman
ia ac
tivity
()
Pent 36 625 125 25 50 1000
20
40
60
80
100
BmatTX-III (Asp49)Pentamidine
BmatTX-III (120583g)
lowastlowastlowast
lowastlowast lowast
(c)
Figure 7 Antileishmanial activity of the crude venom of and PLA2s from B mattogrossensis The promastigote forms of L amazonensiswere
plated with 1times105 cellswellThen different concentrations of crude venom and isolated PLA2s were added to each wellThe experiment was
done in a 48 h period with the (a) crude venom and the PLA2enzymes (b) BmatTX-I and (c) BmatTX-III MTT was added and after the
incubation period at 33∘C the formazan crystal formed was dissolved in SDS Readings were done in a spectrophotometer at 570 nm Eachbar represents the average +minus SD of the three independent experimental groups sixfold total lowast119875 lt 005 compared to the control
studied snake venoms of the Bothrops sp genera despite theobservation of quantitative variations Of the three basicPLA2s from B mattogrossensis venom now isolated for the
first time two are characterized as Lys49-PLA2homologues
BmatTX-I and -II and the other as an Asp49-PLA2 named
BmatTX-III This showed high phospholipase activity ThePLA2s isolated induced myotoxic effects as well as the release
of proinflammatory cytokines by neutrophils BmatTX-I and-III PLA
2s were cytotoxic to human tumor cell lines JURKAT
and SK-BR-3 and showed activity against promastigote formsof L amazonensis
Conflict of Interests
The authors state that there is no conflict of interests
Acknowledgments
The authors are grateful to the Ministry of Science andTechnology (MCTI) Conselho Nacional de Desenvolvi-mento Cientıfico e Tecnologico (CNPq) Financiadora deEstudos e Projetos (FINEP) Fundacao de Tecnologia doAcre (FUNTACFDCT) Coordenacao de Aperfeicoamentode Nıvel Superior (CAPES)mdashProjeto NanoBiotec Redede Biodiversidade e Biotecnologia da Amazonia Legal(BIONORTECNPqMCTI) Instituto Nacional paraPesquisa Translacional em Saude e Ambiente na RegiaoAmazonica (INCT-INPeTAmCNPqMCTI) and InstitutoNacional para Pesquisa em Toxinas (INCT-Tox) andSecretary of Development of Rondonia State (SEPLANPRONEXCNPq) for financial support We are grateful to
BioMed Research International 11
Sr Guilherme G Galassi (Mantenedouro da Fauna Silvestreautorizacao IBAMA 020270089912001-95 Americana-SP)for their helpful technical collaboration in provide the snakevenom Amy Grabner provided the English editing of thepaper
References
[1] Y Angulo and B Lomonte ldquoBiochemistry and toxicology oftoxins purified from the venom of the snake Bothrops asperrdquoToxicon vol 54 no 7 pp 949ndash957 2009
[2] J J Calvete P Juarez and L Sanz ldquoSnake venomics Strategyand applicationsrdquo Journal of Mass Spectrometry vol 42 no 11pp 1405ndash1414 2007
[3] T S Kang D Georgieva N Genov et al ldquoEnzymatic toxinsfrom snake venom structural characterization and mechanismof catalysisrdquoThe FEBS Journal vol 278 no 23 pp 4544ndash45762011
[4] C ZOliveiraCaracterizacao funcional e estrutural de inibidoresde fosfolipases A
2isolados do plasma de serpente Bothrops
jararacussu [PhD thesis] Faculdade de Ciencias Farmaceuticasde Ribeirao Preto Ribeirao Preto Brazil 2009
[5] R G Stabeli R Simoes-Silva A M Kayano et al ldquoPurifi-cation of phospholipases A
2from American snake venomsrdquo
in ChromatographymdashThe Most Versatile Method of ChemicalAnalysis chapter 1 pp 1ndash34 InTech Vienna Austria 2012
[6] J P Zuliani C M Fernandes S R Zamuner J M Gutierrezand C F P Teixeira ldquoInflammatory events induced by Lys-49 and Asp-49 phospholipases A
2isolated from Bothrops asper
snake venom role of catalytic activityrdquo Toxicon vol 45 no 3pp 335ndash346 2005
[7] S Marcussi R G Stabeli N A Santos-Filho et al ldquoGenotoxiceffect of Bothrops snake venoms and isolated toxins on humanlymphocyte DNArdquo Toxicon vol 65 pp 9ndash14 2013
[8] S S Setubal A S Pontes J L Furtado et al ldquoAction oftwo phospholipases A
2purified from Bothrops alternatus snake
venom on macrophagesrdquo Biochemistry vol 78 no 2 pp 194ndash203 2013
[9] J M Gutierrez L A Ponce-Soto S Marangoni and BLomonte ldquoSystemic and local myotoxicity induced by snakevenom group II phospholipases A
2 comparison between cro-
toxin crotoxin B and a Lys49 PLA2homologuerdquo Toxicon vol
51 no 1 pp 80ndash92 2008[10] A Q Rueda I G Rodrıguez E C Ara et al ldquoBiochemical
characterization action onmacrophages and superoxide anionproduction of four basic phospholipases A
2from Panamanian
Bothrops asper snake venomrdquo BioMed Research Internationalvol 2013 Article ID 789689 9 pages 2013
[11] V X da Silva and M T Rodrigues ldquoTaxonomic revision ofthe Bothrops neuwiedi complex (Serpentes Viperidae) withdescription of a new speciesrdquo Phyllomedusa vol 7 no 1 pp 45ndash90 2008
[12] J A Campbell and W W Lamar The Venomous Reptiles of theWestern Hemisphere Comstock Ithaca NY USA 2004
[13] P Edman ldquoMethod for determination of the amino acidsequence in peptidesrdquo Acta Chemica Scandinavica vol 4 pp283ndash293 1950
[14] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[15] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[16] M Holzer and S P Mackessy ldquoAn aqueous endpoint assay ofsnake venom phospholipase A
2rdquo Toxicon vol 34 no 10 pp
1149ndash1155 1996[17] A L Fuly A L P deMiranda R B Zingali and J AGuimaraes
ldquoPurification and characterization of a phospholipase A2isoen-
zyme isolated from Lachesis muta snake venomrdquo BiochemicalPharmacology vol 63 no 9 pp 1589ndash1597 2002
[18] T Nikai N Mori M Kishida H Sugihara and A T Tu ldquoIso-lation and biochemical characterization of hemorrhagic toxinf from the venom of Crotalus atrox (Western DiamondbackRattlesnake)rdquo Archives of Biochemistry and Biophysics vol 231no 2 pp 309ndash319 1984
[19] J A Gene A Roy G Rojas J M Gutierrez and L CerdasldquoComparative study on coagulant defibrinating fibrinolyticand fibrinogenolytic activities of Costa Rican crotaline snakevenoms and their neutralization by a polyvalent antivenomrdquoToxicon vol 27 no 8 pp 841ndash848 1989
[20] M S Charney and R M Tomarelli ldquoA colorimetric method forthe determination of the proteolytic activity of duodenal juicerdquoThe Journal of Biological Chemistry vol 171 pp 501ndash505 1947
[21] R G Stabeli S F Amui C D SantrsquoAna et al ldquoBothrops moojenimyotoxin-II a Lys49-phospholipaseA
2homologue an example
of function versatility of snake venom proteinsrdquo ComparativeBiochemistry and Physiology C vol 142 no 3-4 pp 371ndash3812006
[22] D R Call J A Nemzek S J Ebong G L Bolgos D ENewcomb and D G Remick ldquoRatio of local to systemicchemokine concentrations regulates neutrophil recruitmentrdquoThe American Journal of Pathology vol 158 no 2 pp 715ndash7212001
[23] P G Roberto S Kashima S Marcussi et al ldquoCloning andidentification of a complete cDNA coding for a bactericidal andantitumoral acidic phospholipase A
2from Bothrops jararacussu
venomrdquoThe Protein Journal vol 23 no 4 pp 273ndash285 2004[24] J H Schumacher A OrsquoGarra B Shrader et al ldquoThe charac-
terization of four monoclonal antibodies specific for mouse IL-5 and development of mouse and human IL-5 enzyme-linkedimmunosorbentrdquoThe Journal of Immunology vol 141 no 5 pp1576ndash1581 1988
[25] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[26] C A T Souza A M Kayano S S Setubal et al ldquoLocal andsystemic biochemical alterations induced by Bothrops atroxsnake venom in micerdquo Journal of Venom Research vol 3 pp28ndash34 2012
[27] J M Gutierrez and B Lomonte ldquoPhospholipase A2myotoxins
from Bothrops snake venomsrdquo Toxicon vol 33 no 11 pp 1405ndash1424 1995
[28] J M Gutierrez and A Rucavado ldquoSnake venom metallopro-teinases their role in the pathogenesis of local tissue damagerdquoBiochimie vol 82 no 9-10 pp 841ndash850 2000
[29] T Matsui Y Fujimura and K Titani ldquoSnake venom proteasesaffecting hemostasis and thrombosisrdquo Biochimica et BiophysicaActa vol 1477 no 1-2 pp 146ndash156 2000
[30] A M Soares M R M Fontes and J R Giglio ldquoPhospholipaseA2myotoxins from Bothrops snake venoms structure-function
12 BioMed Research International
relationshiprdquoCurrent Organic Chemistry vol 8 no 17 pp 1677ndash1690 2004
[31] VM Rodrigues SMarcussi R S Cambraia et al ldquoBactericidaland neurotoxic activities of two myotoxic phospholipases A
2
from Bothrops neuwiedi pauloensis snake venomrdquo Toxicon vol44 no 3 pp 305ndash314 2004
[32] N A Santos-Filho L B Silveira C Z Oliveira et al ldquoAnew acidic myotoxic anti-platelet and prostaglandin I
2induc-
tor phospholipase A2isolated from Bothrops moojeni snake
venomrdquo Toxicon vol 52 no 8 pp 908ndash917 2008[33] A M Soares S H Andriao-Escarso Y Angulo et al ldquoStruc-
tural and functional characterization of myotoxin I a Lys49phospholipaseA
2homologue fromBothropsmoojeni (Caissaca)
snake venomrdquo Archives of Biochemistry and Biophysics vol 373no 1 pp 7ndash15 2000
[34] A M Soares S H Andriao-Escarso R K Bortoleto et alldquoDissociation of enzymatic and pharmacological properties ofpiratoxins-I and -III two myotoxic phospholipases A
2from
Bothrops pirajai snake venomrdquo Archives of Biochemistry andBiophysics vol 387 no 2 pp 188ndash196 2001
[35] S S Teixeira L B Silveira F M N da Silva et al ldquoMolecularcharacterization of an acidic phospholipase A
2from Bothrops
pirajai snake venom synthetic C-terminal peptide identifies itsantiplatelet regionrdquo Archives of Toxicology vol 85 no 10 pp1219ndash1233 2011
[36] S H Andriao-Escarso A M Soares V M Rodrigues et alldquoMyotoxic phospholipases A
2in Bothrops snake venoms effect
of chemical modifications on the enzymatic and pharmacolog-ical properties of bothropstoxins from Bothrops jararacussurdquoBiochimie vol 82 no 8 pp 755ndash763 2000
[37] S H Andriao-Escarso A M Soares M R M Fontes et alldquoStructural and functional characterization of an acidic plateletaggregation inhibitor and hypotensive phospholipase A
2from
Bothrops jararacussu snake venomrdquo Biochemical Pharmacologyvol 64 no 4 pp 723ndash732 2002
[38] M E G Denegri O C Acosta S Huancahuire-Vega et alldquoIsolation and functional characterization of a new acidic PLA
2
Ba SpII RP4 of the Bothrops alternatus snake venom fromArgentinardquo Toxicon vol 56 no 1 pp 64ndash74 2010
[39] R Zouari-Kessentini J Luis A Karray et al ldquoTwo purified andcharacterized phospholipases A
2from Cerastes cerastes venom
that inhibit cancerous cell adhesion and migrationrdquo Toxiconvol 53 no 4 pp 444ndash453 2009
[40] R Shirai M Toriba K Hayashi K Ikeda and S Inoue ldquoIden-tification and characterization of phospholipase A
2inhibitors
from the serum of the Japanese rat snake Elaphe climacophorardquoToxicon vol 53 no 6 pp 685ndash692 2009
[41] M I Homsi-Brandenburgo L S Queiroz H Santo-Neto LRodrigues-Simoni and J R Giglio ldquoFractionation of Bothropsjararacussu snake venom partial chemical characterization andbiological activity of bothropstoxinrdquo Toxicon vol 26 no 7 pp615ndash627 1988
[42] G P Fernandez A Segura M Herrera et al ldquoNeutralizationof Bothrops mattogrossensis snake venom from Bolivia exper-imental evaluation of llama and donkey antivenoms producedby caprylic acid precipitationrdquoToxicon vol 55 no 2-3 pp 642ndash645 2010
[43] C L Ownby H S S de Araujo S P White and J E FletcherldquoLysine 49 phospholipase A
2proteinsrdquo Toxicon vol 37 no 3
pp 411ndash445 1999[44] A M Soares W P Sestito S Marcussi et al ldquoAlkylation of
myotoxic phospholipases A2in Bothrops moojeni venom a
promising approach to an enhanced antivenom productionrdquoInternational Journal of Biochemistry amp Cell Biology vol 36 no2 pp 258ndash270 2004
[45] B Lomonte J M Gutierrez M F Furtado et al ldquoIsolationof basic myotoxins from Bothrops moojeni and Bothrops atroxsnake venomsrdquo Toxicon vol 28 no 10 pp 1137ndash1146 1990
[46] L C Mancuso M M Correa C A Vieira et al ldquoFractionationof Bothrops pirajai snake venom isolation and characterizationof piratoxin-I a new myotoxic proteinrdquo Toxicon vol 33 no 5pp 615ndash626 1995
[47] H S Selistre L S Queiroz O A B Cunha G E P deSouza and J R Giglio ldquoIsolation and characterization ofhemorrhagic myonecrotic and edema-inducing toxins fromBothrops insularis (jararaca ilhoa) snake venomrdquo Toxicon vol28 no 3 pp 261ndash273 1990
[48] E A Dennis J Cao Y-H Hsu V Magrioti and G KokotosldquoPhospholipaseA
2enzymes physical structure biological func-
tion disease implication chemical inhibition and therapeuticinterventionrdquo Chemical Reviews vol 111 no 10 pp 6130ndash61852011
[49] A G T Cardoso Acao do veneno de Bothrops moojeni e suafracao L-aminoacido oxidase submetida ao tratamento comraios gama de 60CO em Leishmania spp [MS thesis] IPEN-CNENSP Sao Paulo Brazil 1999
[50] MGallacci andWLGCavalcante ldquoUnderstanding the in vitroneuromuscular activity of snake venomLys49 phospholipaseA
2
homologuesrdquo Toxicon vol 55 no 1 pp 1ndash11 2010[51] S S Teixeira Caracterizacao funcional e estrutural de uma
fosfolipase A2acida isolada do veneno de Bothrops pirajai
[MS thesis] Faculdade de Ciencias Farmaceuticas de RibeiraoPretoUSP Ribeirao Preto Brazil 2009
[52] E Valentin and G Lambeau ldquoIncreasing molecular diversityof secreted phospholipases A
2and their receptors and binding
proteinsrdquo Biochimica et Biophysica Acta vol 1488 no 1-2 pp59ndash70 2000
[53] J M Gutierrez and B Lomonte ldquoPhospholipases A2from
Viperidae snake venoms how do they induce skeletal muscledamagerdquo Acta Chimica Slovenica vol 58 no 4 pp 647ndash6592011
[54] J M Gutierrez C L Ownby and G V Odell ldquoIsolation of amyotoxin from Bothrops asper venom partial characterizationand action on skeletal musclerdquo Toxicon vol 22 no 1 pp 115ndash128 1984
[55] N G Nascimento M C Sampaio R A Olivo and C TeixeiraldquoContribution of mast cells to the oedema induced by Bothropsmoojeni snake venom and a pharmacological assessment of theinflammatory mediators involvedrdquo Toxicon vol 55 no 2-3 pp343ndash352 2010
[56] C F P Teixeira E C T Landucci E Antunes M Chacurand Y Cury ldquoInflammatory effects of snake venom myotoxicphospholipases A
2rdquo Toxicon vol 42 no 8 pp 947ndash962 2003
[57] S R Zamuner and C F P Teixeira ldquoCell adhesion moleculesinvolved in the leukocyte recruitment induced by venom of thesnake Bothrops jararacardquoMediators of Inflammation vol 11 no6 pp 351ndash357 2002
[58] F Chaves C F P Teixeira and J M Gutierrez ldquoRole ofTNF-120572 IL-1120573 and IL-6 in the local tissue damage induced byBothrops asper snake venom an experimental assessment inmicerdquo Toxicon vol 45 no 2 pp 171ndash178 2005
BioMed Research International 13
[59] C D F P Teixeira C M Fernandes J P Zuliani and SF Zamuner ldquoInflammatory effects of snake venom metallo-proteinasesrdquo Memorias do Instituto Oswaldo Cruz vol 100supplement 1 pp 181ndash184 2005
[60] R D A Olivo C F P Teixeira J LWallace J M Gutierrez andS R Zamuner ldquoRole of cyclooxygenases in oedema-formingactivity of bothropic venomsrdquo Toxicon vol 49 no 5 pp 670ndash677 2007
[61] L B Silveira D P Marchi-Salvador N A Santos-Filho et alldquoIsolation and expression of a hypotensive and anti-plateletacidic phospholipase A
2from Bothrops moojeni snake venomrdquo
Journal of Pharmaceutical and Biomedical Analysis vol 73 pp35ndash43 2013
[62] A Rucavado T Escalante C F P Teixeira C M Fernandes CDıaz and J M Gutierrez ldquoIncrements in cytokines and matrixmetalloproteinases in skeletal muscle after injection of tissue-damaging toxins from the venom of the snake Bothrops asperrdquoMediators of Inflammation vol 11 no 2 pp 121ndash128 2002
[63] E Leiguez J P Zuliani A M Cianciarullo C M Fernandes JM Gutierrez and C Teixeira ldquoA group IIA-secreted phospho-lipase A
2from snake venom induces lipid body formation in
macrophages the roles of intracellular phospholipases A2and
distinct signaling pathwaysrdquo Journal of Leukocyte Biology vol90 no 1 pp 155ndash166 2011
[64] J P Zuliani J M Gutierrez L L Casais e Silva S C SampaioB Lomonte and C D F P Teixeira ldquoActivation of cellularfunctions in macrophages by venom secretory Asp-49 and Lys-49 phospholipasesA
2rdquoToxicon vol 46 no 5 pp 523ndash532 2005
[65] R D C M Escocard M M Kanashiro J H Petretski et alldquoNeutrophils regulate the expression of cytokines chemokinesand nitric oxide synthasenitric oxide in mice injected withBothrops atrox venomrdquo Immunobiology vol 211 no 1-2 pp 37ndash46 2006
[66] A Gambero E C T Landucci M H Toyama et al ldquoHumanneutrophil migration in vitro induced by secretory phospholi-pases A
2 a role for cell surface glycosaminoglycansrdquo Biochemi-
cal Pharmacology vol 63 no 1 pp 65ndash72 2002[67] F L S Costa R S Rodrigues L F M Izidoro et al ldquoBio-
chemical and functional properties of a thrombin-like enzymeisolated from Bothrops pauloensis snake venomrdquo Toxicon vol54 no 6 pp 725ndash735 2009
[68] J C Villalobos R Mora B Lomonte J M Gutierrez andY Angulo ldquoCytotoxicity induced in myotubes by a Lys49phospholipase A
2homologue from the venom of the snake
Bothrops asper evidence of rapid plasma membrane damageand a dual role for extracellular calciumrdquo Toxicology in Vitrovol 21 no 8 pp 1382ndash1389 2007
[69] B S Cummings J McHowat and R G Schnellmann ldquoPhos-pholipase A
2s in cell injury and deathrdquoThe Journal of Pharma-
cology and Experimental Therapeutics vol 294 no 3 pp 793ndash799 2000
[70] S R Panini L Yang A E Rusinol M S Sinensky J VBonventre and C C Leslie ldquoArachidonate metabolism andthe signaling pathway of induction of apoptosis by oxidizedLDLoxysterolrdquo Journal of Lipid Research vol 42 no 10 pp1678ndash1686 2001
[71] L Paramo B Lomonte J Pizarro-Cerda J-A Bengoechea J-P Gorvel and E Moreno ldquoBactericidal activity of Lys49 andAsp49 myotoxic phospholipases A
2from Bothrops asper snake
venom synthetic Lys49myotoxin II-(115-129)-peptide identifiesits bactericidal regionrdquo European Journal of Biochemistry vol253 no 2 pp 452ndash461 1998
[72] B Lomonte Y Angulo and E Moreno ldquoSynthetic peptidesderived from the C-terminal region of Lys49 phospholipaseA2homologues from viperidae snake venoms biomimetic
activities and potential applicationsrdquo Current PharmaceuticalDesign vol 16 no 28 pp 3224ndash3230 2010
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

10 BioMed Research International
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineB mattogrossensis crude venom
Ant
i-leis
hman
ia ac
tivity
()
B mattogrossensis crude venom (120583g)
(a)
Pent 36 625 125 25 50 1000
20
40
60
80
100
PentamidineBmatTX-I (Lys49)
Ant
i-leis
hman
ia ac
tivity
()
BmatTX-I (120583g)
lowastlowast lowast lowast
(b)
Ant
i-leis
hman
ia ac
tivity
()
Pent 36 625 125 25 50 1000
20
40
60
80
100
BmatTX-III (Asp49)Pentamidine
BmatTX-III (120583g)
lowastlowastlowast
lowastlowast lowast
(c)
Figure 7 Antileishmanial activity of the crude venom of and PLA2s from B mattogrossensis The promastigote forms of L amazonensiswere
plated with 1times105 cellswellThen different concentrations of crude venom and isolated PLA2s were added to each wellThe experiment was
done in a 48 h period with the (a) crude venom and the PLA2enzymes (b) BmatTX-I and (c) BmatTX-III MTT was added and after the
incubation period at 33∘C the formazan crystal formed was dissolved in SDS Readings were done in a spectrophotometer at 570 nm Eachbar represents the average +minus SD of the three independent experimental groups sixfold total lowast119875 lt 005 compared to the control
studied snake venoms of the Bothrops sp genera despite theobservation of quantitative variations Of the three basicPLA2s from B mattogrossensis venom now isolated for the
first time two are characterized as Lys49-PLA2homologues
BmatTX-I and -II and the other as an Asp49-PLA2 named
BmatTX-III This showed high phospholipase activity ThePLA2s isolated induced myotoxic effects as well as the release
of proinflammatory cytokines by neutrophils BmatTX-I and-III PLA
2s were cytotoxic to human tumor cell lines JURKAT
and SK-BR-3 and showed activity against promastigote formsof L amazonensis
Conflict of Interests
The authors state that there is no conflict of interests
Acknowledgments
The authors are grateful to the Ministry of Science andTechnology (MCTI) Conselho Nacional de Desenvolvi-mento Cientıfico e Tecnologico (CNPq) Financiadora deEstudos e Projetos (FINEP) Fundacao de Tecnologia doAcre (FUNTACFDCT) Coordenacao de Aperfeicoamentode Nıvel Superior (CAPES)mdashProjeto NanoBiotec Redede Biodiversidade e Biotecnologia da Amazonia Legal(BIONORTECNPqMCTI) Instituto Nacional paraPesquisa Translacional em Saude e Ambiente na RegiaoAmazonica (INCT-INPeTAmCNPqMCTI) and InstitutoNacional para Pesquisa em Toxinas (INCT-Tox) andSecretary of Development of Rondonia State (SEPLANPRONEXCNPq) for financial support We are grateful to
BioMed Research International 11
Sr Guilherme G Galassi (Mantenedouro da Fauna Silvestreautorizacao IBAMA 020270089912001-95 Americana-SP)for their helpful technical collaboration in provide the snakevenom Amy Grabner provided the English editing of thepaper
References
[1] Y Angulo and B Lomonte ldquoBiochemistry and toxicology oftoxins purified from the venom of the snake Bothrops asperrdquoToxicon vol 54 no 7 pp 949ndash957 2009
[2] J J Calvete P Juarez and L Sanz ldquoSnake venomics Strategyand applicationsrdquo Journal of Mass Spectrometry vol 42 no 11pp 1405ndash1414 2007
[3] T S Kang D Georgieva N Genov et al ldquoEnzymatic toxinsfrom snake venom structural characterization and mechanismof catalysisrdquoThe FEBS Journal vol 278 no 23 pp 4544ndash45762011
[4] C ZOliveiraCaracterizacao funcional e estrutural de inibidoresde fosfolipases A
2isolados do plasma de serpente Bothrops
jararacussu [PhD thesis] Faculdade de Ciencias Farmaceuticasde Ribeirao Preto Ribeirao Preto Brazil 2009
[5] R G Stabeli R Simoes-Silva A M Kayano et al ldquoPurifi-cation of phospholipases A
2from American snake venomsrdquo
in ChromatographymdashThe Most Versatile Method of ChemicalAnalysis chapter 1 pp 1ndash34 InTech Vienna Austria 2012
[6] J P Zuliani C M Fernandes S R Zamuner J M Gutierrezand C F P Teixeira ldquoInflammatory events induced by Lys-49 and Asp-49 phospholipases A
2isolated from Bothrops asper
snake venom role of catalytic activityrdquo Toxicon vol 45 no 3pp 335ndash346 2005
[7] S Marcussi R G Stabeli N A Santos-Filho et al ldquoGenotoxiceffect of Bothrops snake venoms and isolated toxins on humanlymphocyte DNArdquo Toxicon vol 65 pp 9ndash14 2013
[8] S S Setubal A S Pontes J L Furtado et al ldquoAction oftwo phospholipases A
2purified from Bothrops alternatus snake
venom on macrophagesrdquo Biochemistry vol 78 no 2 pp 194ndash203 2013
[9] J M Gutierrez L A Ponce-Soto S Marangoni and BLomonte ldquoSystemic and local myotoxicity induced by snakevenom group II phospholipases A
2 comparison between cro-
toxin crotoxin B and a Lys49 PLA2homologuerdquo Toxicon vol
51 no 1 pp 80ndash92 2008[10] A Q Rueda I G Rodrıguez E C Ara et al ldquoBiochemical
characterization action onmacrophages and superoxide anionproduction of four basic phospholipases A
2from Panamanian
Bothrops asper snake venomrdquo BioMed Research Internationalvol 2013 Article ID 789689 9 pages 2013
[11] V X da Silva and M T Rodrigues ldquoTaxonomic revision ofthe Bothrops neuwiedi complex (Serpentes Viperidae) withdescription of a new speciesrdquo Phyllomedusa vol 7 no 1 pp 45ndash90 2008
[12] J A Campbell and W W Lamar The Venomous Reptiles of theWestern Hemisphere Comstock Ithaca NY USA 2004
[13] P Edman ldquoMethod for determination of the amino acidsequence in peptidesrdquo Acta Chemica Scandinavica vol 4 pp283ndash293 1950
[14] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[15] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[16] M Holzer and S P Mackessy ldquoAn aqueous endpoint assay ofsnake venom phospholipase A
2rdquo Toxicon vol 34 no 10 pp
1149ndash1155 1996[17] A L Fuly A L P deMiranda R B Zingali and J AGuimaraes
ldquoPurification and characterization of a phospholipase A2isoen-
zyme isolated from Lachesis muta snake venomrdquo BiochemicalPharmacology vol 63 no 9 pp 1589ndash1597 2002
[18] T Nikai N Mori M Kishida H Sugihara and A T Tu ldquoIso-lation and biochemical characterization of hemorrhagic toxinf from the venom of Crotalus atrox (Western DiamondbackRattlesnake)rdquo Archives of Biochemistry and Biophysics vol 231no 2 pp 309ndash319 1984
[19] J A Gene A Roy G Rojas J M Gutierrez and L CerdasldquoComparative study on coagulant defibrinating fibrinolyticand fibrinogenolytic activities of Costa Rican crotaline snakevenoms and their neutralization by a polyvalent antivenomrdquoToxicon vol 27 no 8 pp 841ndash848 1989
[20] M S Charney and R M Tomarelli ldquoA colorimetric method forthe determination of the proteolytic activity of duodenal juicerdquoThe Journal of Biological Chemistry vol 171 pp 501ndash505 1947
[21] R G Stabeli S F Amui C D SantrsquoAna et al ldquoBothrops moojenimyotoxin-II a Lys49-phospholipaseA
2homologue an example
of function versatility of snake venom proteinsrdquo ComparativeBiochemistry and Physiology C vol 142 no 3-4 pp 371ndash3812006
[22] D R Call J A Nemzek S J Ebong G L Bolgos D ENewcomb and D G Remick ldquoRatio of local to systemicchemokine concentrations regulates neutrophil recruitmentrdquoThe American Journal of Pathology vol 158 no 2 pp 715ndash7212001
[23] P G Roberto S Kashima S Marcussi et al ldquoCloning andidentification of a complete cDNA coding for a bactericidal andantitumoral acidic phospholipase A
2from Bothrops jararacussu
venomrdquoThe Protein Journal vol 23 no 4 pp 273ndash285 2004[24] J H Schumacher A OrsquoGarra B Shrader et al ldquoThe charac-
terization of four monoclonal antibodies specific for mouse IL-5 and development of mouse and human IL-5 enzyme-linkedimmunosorbentrdquoThe Journal of Immunology vol 141 no 5 pp1576ndash1581 1988
[25] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[26] C A T Souza A M Kayano S S Setubal et al ldquoLocal andsystemic biochemical alterations induced by Bothrops atroxsnake venom in micerdquo Journal of Venom Research vol 3 pp28ndash34 2012
[27] J M Gutierrez and B Lomonte ldquoPhospholipase A2myotoxins
from Bothrops snake venomsrdquo Toxicon vol 33 no 11 pp 1405ndash1424 1995
[28] J M Gutierrez and A Rucavado ldquoSnake venom metallopro-teinases their role in the pathogenesis of local tissue damagerdquoBiochimie vol 82 no 9-10 pp 841ndash850 2000
[29] T Matsui Y Fujimura and K Titani ldquoSnake venom proteasesaffecting hemostasis and thrombosisrdquo Biochimica et BiophysicaActa vol 1477 no 1-2 pp 146ndash156 2000
[30] A M Soares M R M Fontes and J R Giglio ldquoPhospholipaseA2myotoxins from Bothrops snake venoms structure-function
12 BioMed Research International
relationshiprdquoCurrent Organic Chemistry vol 8 no 17 pp 1677ndash1690 2004
[31] VM Rodrigues SMarcussi R S Cambraia et al ldquoBactericidaland neurotoxic activities of two myotoxic phospholipases A
2
from Bothrops neuwiedi pauloensis snake venomrdquo Toxicon vol44 no 3 pp 305ndash314 2004
[32] N A Santos-Filho L B Silveira C Z Oliveira et al ldquoAnew acidic myotoxic anti-platelet and prostaglandin I
2induc-
tor phospholipase A2isolated from Bothrops moojeni snake
venomrdquo Toxicon vol 52 no 8 pp 908ndash917 2008[33] A M Soares S H Andriao-Escarso Y Angulo et al ldquoStruc-
tural and functional characterization of myotoxin I a Lys49phospholipaseA
2homologue fromBothropsmoojeni (Caissaca)
snake venomrdquo Archives of Biochemistry and Biophysics vol 373no 1 pp 7ndash15 2000
[34] A M Soares S H Andriao-Escarso R K Bortoleto et alldquoDissociation of enzymatic and pharmacological properties ofpiratoxins-I and -III two myotoxic phospholipases A
2from
Bothrops pirajai snake venomrdquo Archives of Biochemistry andBiophysics vol 387 no 2 pp 188ndash196 2001
[35] S S Teixeira L B Silveira F M N da Silva et al ldquoMolecularcharacterization of an acidic phospholipase A
2from Bothrops
pirajai snake venom synthetic C-terminal peptide identifies itsantiplatelet regionrdquo Archives of Toxicology vol 85 no 10 pp1219ndash1233 2011
[36] S H Andriao-Escarso A M Soares V M Rodrigues et alldquoMyotoxic phospholipases A
2in Bothrops snake venoms effect
of chemical modifications on the enzymatic and pharmacolog-ical properties of bothropstoxins from Bothrops jararacussurdquoBiochimie vol 82 no 8 pp 755ndash763 2000
[37] S H Andriao-Escarso A M Soares M R M Fontes et alldquoStructural and functional characterization of an acidic plateletaggregation inhibitor and hypotensive phospholipase A
2from
Bothrops jararacussu snake venomrdquo Biochemical Pharmacologyvol 64 no 4 pp 723ndash732 2002
[38] M E G Denegri O C Acosta S Huancahuire-Vega et alldquoIsolation and functional characterization of a new acidic PLA
2
Ba SpII RP4 of the Bothrops alternatus snake venom fromArgentinardquo Toxicon vol 56 no 1 pp 64ndash74 2010
[39] R Zouari-Kessentini J Luis A Karray et al ldquoTwo purified andcharacterized phospholipases A
2from Cerastes cerastes venom
that inhibit cancerous cell adhesion and migrationrdquo Toxiconvol 53 no 4 pp 444ndash453 2009
[40] R Shirai M Toriba K Hayashi K Ikeda and S Inoue ldquoIden-tification and characterization of phospholipase A
2inhibitors
from the serum of the Japanese rat snake Elaphe climacophorardquoToxicon vol 53 no 6 pp 685ndash692 2009
[41] M I Homsi-Brandenburgo L S Queiroz H Santo-Neto LRodrigues-Simoni and J R Giglio ldquoFractionation of Bothropsjararacussu snake venom partial chemical characterization andbiological activity of bothropstoxinrdquo Toxicon vol 26 no 7 pp615ndash627 1988
[42] G P Fernandez A Segura M Herrera et al ldquoNeutralizationof Bothrops mattogrossensis snake venom from Bolivia exper-imental evaluation of llama and donkey antivenoms producedby caprylic acid precipitationrdquoToxicon vol 55 no 2-3 pp 642ndash645 2010
[43] C L Ownby H S S de Araujo S P White and J E FletcherldquoLysine 49 phospholipase A
2proteinsrdquo Toxicon vol 37 no 3
pp 411ndash445 1999[44] A M Soares W P Sestito S Marcussi et al ldquoAlkylation of
myotoxic phospholipases A2in Bothrops moojeni venom a
promising approach to an enhanced antivenom productionrdquoInternational Journal of Biochemistry amp Cell Biology vol 36 no2 pp 258ndash270 2004
[45] B Lomonte J M Gutierrez M F Furtado et al ldquoIsolationof basic myotoxins from Bothrops moojeni and Bothrops atroxsnake venomsrdquo Toxicon vol 28 no 10 pp 1137ndash1146 1990
[46] L C Mancuso M M Correa C A Vieira et al ldquoFractionationof Bothrops pirajai snake venom isolation and characterizationof piratoxin-I a new myotoxic proteinrdquo Toxicon vol 33 no 5pp 615ndash626 1995
[47] H S Selistre L S Queiroz O A B Cunha G E P deSouza and J R Giglio ldquoIsolation and characterization ofhemorrhagic myonecrotic and edema-inducing toxins fromBothrops insularis (jararaca ilhoa) snake venomrdquo Toxicon vol28 no 3 pp 261ndash273 1990
[48] E A Dennis J Cao Y-H Hsu V Magrioti and G KokotosldquoPhospholipaseA
2enzymes physical structure biological func-
tion disease implication chemical inhibition and therapeuticinterventionrdquo Chemical Reviews vol 111 no 10 pp 6130ndash61852011
[49] A G T Cardoso Acao do veneno de Bothrops moojeni e suafracao L-aminoacido oxidase submetida ao tratamento comraios gama de 60CO em Leishmania spp [MS thesis] IPEN-CNENSP Sao Paulo Brazil 1999
[50] MGallacci andWLGCavalcante ldquoUnderstanding the in vitroneuromuscular activity of snake venomLys49 phospholipaseA
2
homologuesrdquo Toxicon vol 55 no 1 pp 1ndash11 2010[51] S S Teixeira Caracterizacao funcional e estrutural de uma
fosfolipase A2acida isolada do veneno de Bothrops pirajai
[MS thesis] Faculdade de Ciencias Farmaceuticas de RibeiraoPretoUSP Ribeirao Preto Brazil 2009
[52] E Valentin and G Lambeau ldquoIncreasing molecular diversityof secreted phospholipases A
2and their receptors and binding
proteinsrdquo Biochimica et Biophysica Acta vol 1488 no 1-2 pp59ndash70 2000
[53] J M Gutierrez and B Lomonte ldquoPhospholipases A2from
Viperidae snake venoms how do they induce skeletal muscledamagerdquo Acta Chimica Slovenica vol 58 no 4 pp 647ndash6592011
[54] J M Gutierrez C L Ownby and G V Odell ldquoIsolation of amyotoxin from Bothrops asper venom partial characterizationand action on skeletal musclerdquo Toxicon vol 22 no 1 pp 115ndash128 1984
[55] N G Nascimento M C Sampaio R A Olivo and C TeixeiraldquoContribution of mast cells to the oedema induced by Bothropsmoojeni snake venom and a pharmacological assessment of theinflammatory mediators involvedrdquo Toxicon vol 55 no 2-3 pp343ndash352 2010
[56] C F P Teixeira E C T Landucci E Antunes M Chacurand Y Cury ldquoInflammatory effects of snake venom myotoxicphospholipases A
2rdquo Toxicon vol 42 no 8 pp 947ndash962 2003
[57] S R Zamuner and C F P Teixeira ldquoCell adhesion moleculesinvolved in the leukocyte recruitment induced by venom of thesnake Bothrops jararacardquoMediators of Inflammation vol 11 no6 pp 351ndash357 2002
[58] F Chaves C F P Teixeira and J M Gutierrez ldquoRole ofTNF-120572 IL-1120573 and IL-6 in the local tissue damage induced byBothrops asper snake venom an experimental assessment inmicerdquo Toxicon vol 45 no 2 pp 171ndash178 2005
BioMed Research International 13
[59] C D F P Teixeira C M Fernandes J P Zuliani and SF Zamuner ldquoInflammatory effects of snake venom metallo-proteinasesrdquo Memorias do Instituto Oswaldo Cruz vol 100supplement 1 pp 181ndash184 2005
[60] R D A Olivo C F P Teixeira J LWallace J M Gutierrez andS R Zamuner ldquoRole of cyclooxygenases in oedema-formingactivity of bothropic venomsrdquo Toxicon vol 49 no 5 pp 670ndash677 2007
[61] L B Silveira D P Marchi-Salvador N A Santos-Filho et alldquoIsolation and expression of a hypotensive and anti-plateletacidic phospholipase A
2from Bothrops moojeni snake venomrdquo
Journal of Pharmaceutical and Biomedical Analysis vol 73 pp35ndash43 2013
[62] A Rucavado T Escalante C F P Teixeira C M Fernandes CDıaz and J M Gutierrez ldquoIncrements in cytokines and matrixmetalloproteinases in skeletal muscle after injection of tissue-damaging toxins from the venom of the snake Bothrops asperrdquoMediators of Inflammation vol 11 no 2 pp 121ndash128 2002
[63] E Leiguez J P Zuliani A M Cianciarullo C M Fernandes JM Gutierrez and C Teixeira ldquoA group IIA-secreted phospho-lipase A
2from snake venom induces lipid body formation in
macrophages the roles of intracellular phospholipases A2and
distinct signaling pathwaysrdquo Journal of Leukocyte Biology vol90 no 1 pp 155ndash166 2011
[64] J P Zuliani J M Gutierrez L L Casais e Silva S C SampaioB Lomonte and C D F P Teixeira ldquoActivation of cellularfunctions in macrophages by venom secretory Asp-49 and Lys-49 phospholipasesA
2rdquoToxicon vol 46 no 5 pp 523ndash532 2005
[65] R D C M Escocard M M Kanashiro J H Petretski et alldquoNeutrophils regulate the expression of cytokines chemokinesand nitric oxide synthasenitric oxide in mice injected withBothrops atrox venomrdquo Immunobiology vol 211 no 1-2 pp 37ndash46 2006
[66] A Gambero E C T Landucci M H Toyama et al ldquoHumanneutrophil migration in vitro induced by secretory phospholi-pases A
2 a role for cell surface glycosaminoglycansrdquo Biochemi-
cal Pharmacology vol 63 no 1 pp 65ndash72 2002[67] F L S Costa R S Rodrigues L F M Izidoro et al ldquoBio-
chemical and functional properties of a thrombin-like enzymeisolated from Bothrops pauloensis snake venomrdquo Toxicon vol54 no 6 pp 725ndash735 2009
[68] J C Villalobos R Mora B Lomonte J M Gutierrez andY Angulo ldquoCytotoxicity induced in myotubes by a Lys49phospholipase A
2homologue from the venom of the snake
Bothrops asper evidence of rapid plasma membrane damageand a dual role for extracellular calciumrdquo Toxicology in Vitrovol 21 no 8 pp 1382ndash1389 2007
[69] B S Cummings J McHowat and R G Schnellmann ldquoPhos-pholipase A
2s in cell injury and deathrdquoThe Journal of Pharma-
cology and Experimental Therapeutics vol 294 no 3 pp 793ndash799 2000
[70] S R Panini L Yang A E Rusinol M S Sinensky J VBonventre and C C Leslie ldquoArachidonate metabolism andthe signaling pathway of induction of apoptosis by oxidizedLDLoxysterolrdquo Journal of Lipid Research vol 42 no 10 pp1678ndash1686 2001
[71] L Paramo B Lomonte J Pizarro-Cerda J-A Bengoechea J-P Gorvel and E Moreno ldquoBactericidal activity of Lys49 andAsp49 myotoxic phospholipases A
2from Bothrops asper snake
venom synthetic Lys49myotoxin II-(115-129)-peptide identifiesits bactericidal regionrdquo European Journal of Biochemistry vol253 no 2 pp 452ndash461 1998
[72] B Lomonte Y Angulo and E Moreno ldquoSynthetic peptidesderived from the C-terminal region of Lys49 phospholipaseA2homologues from viperidae snake venoms biomimetic
activities and potential applicationsrdquo Current PharmaceuticalDesign vol 16 no 28 pp 3224ndash3230 2010
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

BioMed Research International 11
Sr Guilherme G Galassi (Mantenedouro da Fauna Silvestreautorizacao IBAMA 020270089912001-95 Americana-SP)for their helpful technical collaboration in provide the snakevenom Amy Grabner provided the English editing of thepaper
References
[1] Y Angulo and B Lomonte ldquoBiochemistry and toxicology oftoxins purified from the venom of the snake Bothrops asperrdquoToxicon vol 54 no 7 pp 949ndash957 2009
[2] J J Calvete P Juarez and L Sanz ldquoSnake venomics Strategyand applicationsrdquo Journal of Mass Spectrometry vol 42 no 11pp 1405ndash1414 2007
[3] T S Kang D Georgieva N Genov et al ldquoEnzymatic toxinsfrom snake venom structural characterization and mechanismof catalysisrdquoThe FEBS Journal vol 278 no 23 pp 4544ndash45762011
[4] C ZOliveiraCaracterizacao funcional e estrutural de inibidoresde fosfolipases A
2isolados do plasma de serpente Bothrops
jararacussu [PhD thesis] Faculdade de Ciencias Farmaceuticasde Ribeirao Preto Ribeirao Preto Brazil 2009
[5] R G Stabeli R Simoes-Silva A M Kayano et al ldquoPurifi-cation of phospholipases A
2from American snake venomsrdquo
in ChromatographymdashThe Most Versatile Method of ChemicalAnalysis chapter 1 pp 1ndash34 InTech Vienna Austria 2012
[6] J P Zuliani C M Fernandes S R Zamuner J M Gutierrezand C F P Teixeira ldquoInflammatory events induced by Lys-49 and Asp-49 phospholipases A
2isolated from Bothrops asper
snake venom role of catalytic activityrdquo Toxicon vol 45 no 3pp 335ndash346 2005
[7] S Marcussi R G Stabeli N A Santos-Filho et al ldquoGenotoxiceffect of Bothrops snake venoms and isolated toxins on humanlymphocyte DNArdquo Toxicon vol 65 pp 9ndash14 2013
[8] S S Setubal A S Pontes J L Furtado et al ldquoAction oftwo phospholipases A
2purified from Bothrops alternatus snake
venom on macrophagesrdquo Biochemistry vol 78 no 2 pp 194ndash203 2013
[9] J M Gutierrez L A Ponce-Soto S Marangoni and BLomonte ldquoSystemic and local myotoxicity induced by snakevenom group II phospholipases A
2 comparison between cro-
toxin crotoxin B and a Lys49 PLA2homologuerdquo Toxicon vol
51 no 1 pp 80ndash92 2008[10] A Q Rueda I G Rodrıguez E C Ara et al ldquoBiochemical
characterization action onmacrophages and superoxide anionproduction of four basic phospholipases A
2from Panamanian
Bothrops asper snake venomrdquo BioMed Research Internationalvol 2013 Article ID 789689 9 pages 2013
[11] V X da Silva and M T Rodrigues ldquoTaxonomic revision ofthe Bothrops neuwiedi complex (Serpentes Viperidae) withdescription of a new speciesrdquo Phyllomedusa vol 7 no 1 pp 45ndash90 2008
[12] J A Campbell and W W Lamar The Venomous Reptiles of theWestern Hemisphere Comstock Ithaca NY USA 2004
[13] P Edman ldquoMethod for determination of the amino acidsequence in peptidesrdquo Acta Chemica Scandinavica vol 4 pp283ndash293 1950
[14] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[15] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[16] M Holzer and S P Mackessy ldquoAn aqueous endpoint assay ofsnake venom phospholipase A
2rdquo Toxicon vol 34 no 10 pp
1149ndash1155 1996[17] A L Fuly A L P deMiranda R B Zingali and J AGuimaraes
ldquoPurification and characterization of a phospholipase A2isoen-
zyme isolated from Lachesis muta snake venomrdquo BiochemicalPharmacology vol 63 no 9 pp 1589ndash1597 2002
[18] T Nikai N Mori M Kishida H Sugihara and A T Tu ldquoIso-lation and biochemical characterization of hemorrhagic toxinf from the venom of Crotalus atrox (Western DiamondbackRattlesnake)rdquo Archives of Biochemistry and Biophysics vol 231no 2 pp 309ndash319 1984
[19] J A Gene A Roy G Rojas J M Gutierrez and L CerdasldquoComparative study on coagulant defibrinating fibrinolyticand fibrinogenolytic activities of Costa Rican crotaline snakevenoms and their neutralization by a polyvalent antivenomrdquoToxicon vol 27 no 8 pp 841ndash848 1989
[20] M S Charney and R M Tomarelli ldquoA colorimetric method forthe determination of the proteolytic activity of duodenal juicerdquoThe Journal of Biological Chemistry vol 171 pp 501ndash505 1947
[21] R G Stabeli S F Amui C D SantrsquoAna et al ldquoBothrops moojenimyotoxin-II a Lys49-phospholipaseA
2homologue an example
of function versatility of snake venom proteinsrdquo ComparativeBiochemistry and Physiology C vol 142 no 3-4 pp 371ndash3812006
[22] D R Call J A Nemzek S J Ebong G L Bolgos D ENewcomb and D G Remick ldquoRatio of local to systemicchemokine concentrations regulates neutrophil recruitmentrdquoThe American Journal of Pathology vol 158 no 2 pp 715ndash7212001
[23] P G Roberto S Kashima S Marcussi et al ldquoCloning andidentification of a complete cDNA coding for a bactericidal andantitumoral acidic phospholipase A
2from Bothrops jararacussu
venomrdquoThe Protein Journal vol 23 no 4 pp 273ndash285 2004[24] J H Schumacher A OrsquoGarra B Shrader et al ldquoThe charac-
terization of four monoclonal antibodies specific for mouse IL-5 and development of mouse and human IL-5 enzyme-linkedimmunosorbentrdquoThe Journal of Immunology vol 141 no 5 pp1576ndash1581 1988
[25] T Mosmann ldquoRapid colorimetric assay for cellular growth andsurvival application to proliferation and cytotoxicity assaysrdquoJournal of Immunological Methods vol 65 no 1-2 pp 55ndash631983
[26] C A T Souza A M Kayano S S Setubal et al ldquoLocal andsystemic biochemical alterations induced by Bothrops atroxsnake venom in micerdquo Journal of Venom Research vol 3 pp28ndash34 2012
[27] J M Gutierrez and B Lomonte ldquoPhospholipase A2myotoxins
from Bothrops snake venomsrdquo Toxicon vol 33 no 11 pp 1405ndash1424 1995
[28] J M Gutierrez and A Rucavado ldquoSnake venom metallopro-teinases their role in the pathogenesis of local tissue damagerdquoBiochimie vol 82 no 9-10 pp 841ndash850 2000
[29] T Matsui Y Fujimura and K Titani ldquoSnake venom proteasesaffecting hemostasis and thrombosisrdquo Biochimica et BiophysicaActa vol 1477 no 1-2 pp 146ndash156 2000
[30] A M Soares M R M Fontes and J R Giglio ldquoPhospholipaseA2myotoxins from Bothrops snake venoms structure-function
12 BioMed Research International
relationshiprdquoCurrent Organic Chemistry vol 8 no 17 pp 1677ndash1690 2004
[31] VM Rodrigues SMarcussi R S Cambraia et al ldquoBactericidaland neurotoxic activities of two myotoxic phospholipases A
2
from Bothrops neuwiedi pauloensis snake venomrdquo Toxicon vol44 no 3 pp 305ndash314 2004
[32] N A Santos-Filho L B Silveira C Z Oliveira et al ldquoAnew acidic myotoxic anti-platelet and prostaglandin I
2induc-
tor phospholipase A2isolated from Bothrops moojeni snake
venomrdquo Toxicon vol 52 no 8 pp 908ndash917 2008[33] A M Soares S H Andriao-Escarso Y Angulo et al ldquoStruc-
tural and functional characterization of myotoxin I a Lys49phospholipaseA
2homologue fromBothropsmoojeni (Caissaca)
snake venomrdquo Archives of Biochemistry and Biophysics vol 373no 1 pp 7ndash15 2000
[34] A M Soares S H Andriao-Escarso R K Bortoleto et alldquoDissociation of enzymatic and pharmacological properties ofpiratoxins-I and -III two myotoxic phospholipases A
2from
Bothrops pirajai snake venomrdquo Archives of Biochemistry andBiophysics vol 387 no 2 pp 188ndash196 2001
[35] S S Teixeira L B Silveira F M N da Silva et al ldquoMolecularcharacterization of an acidic phospholipase A
2from Bothrops
pirajai snake venom synthetic C-terminal peptide identifies itsantiplatelet regionrdquo Archives of Toxicology vol 85 no 10 pp1219ndash1233 2011
[36] S H Andriao-Escarso A M Soares V M Rodrigues et alldquoMyotoxic phospholipases A
2in Bothrops snake venoms effect
of chemical modifications on the enzymatic and pharmacolog-ical properties of bothropstoxins from Bothrops jararacussurdquoBiochimie vol 82 no 8 pp 755ndash763 2000
[37] S H Andriao-Escarso A M Soares M R M Fontes et alldquoStructural and functional characterization of an acidic plateletaggregation inhibitor and hypotensive phospholipase A
2from
Bothrops jararacussu snake venomrdquo Biochemical Pharmacologyvol 64 no 4 pp 723ndash732 2002
[38] M E G Denegri O C Acosta S Huancahuire-Vega et alldquoIsolation and functional characterization of a new acidic PLA
2
Ba SpII RP4 of the Bothrops alternatus snake venom fromArgentinardquo Toxicon vol 56 no 1 pp 64ndash74 2010
[39] R Zouari-Kessentini J Luis A Karray et al ldquoTwo purified andcharacterized phospholipases A
2from Cerastes cerastes venom
that inhibit cancerous cell adhesion and migrationrdquo Toxiconvol 53 no 4 pp 444ndash453 2009
[40] R Shirai M Toriba K Hayashi K Ikeda and S Inoue ldquoIden-tification and characterization of phospholipase A
2inhibitors
from the serum of the Japanese rat snake Elaphe climacophorardquoToxicon vol 53 no 6 pp 685ndash692 2009
[41] M I Homsi-Brandenburgo L S Queiroz H Santo-Neto LRodrigues-Simoni and J R Giglio ldquoFractionation of Bothropsjararacussu snake venom partial chemical characterization andbiological activity of bothropstoxinrdquo Toxicon vol 26 no 7 pp615ndash627 1988
[42] G P Fernandez A Segura M Herrera et al ldquoNeutralizationof Bothrops mattogrossensis snake venom from Bolivia exper-imental evaluation of llama and donkey antivenoms producedby caprylic acid precipitationrdquoToxicon vol 55 no 2-3 pp 642ndash645 2010
[43] C L Ownby H S S de Araujo S P White and J E FletcherldquoLysine 49 phospholipase A
2proteinsrdquo Toxicon vol 37 no 3
pp 411ndash445 1999[44] A M Soares W P Sestito S Marcussi et al ldquoAlkylation of
myotoxic phospholipases A2in Bothrops moojeni venom a
promising approach to an enhanced antivenom productionrdquoInternational Journal of Biochemistry amp Cell Biology vol 36 no2 pp 258ndash270 2004
[45] B Lomonte J M Gutierrez M F Furtado et al ldquoIsolationof basic myotoxins from Bothrops moojeni and Bothrops atroxsnake venomsrdquo Toxicon vol 28 no 10 pp 1137ndash1146 1990
[46] L C Mancuso M M Correa C A Vieira et al ldquoFractionationof Bothrops pirajai snake venom isolation and characterizationof piratoxin-I a new myotoxic proteinrdquo Toxicon vol 33 no 5pp 615ndash626 1995
[47] H S Selistre L S Queiroz O A B Cunha G E P deSouza and J R Giglio ldquoIsolation and characterization ofhemorrhagic myonecrotic and edema-inducing toxins fromBothrops insularis (jararaca ilhoa) snake venomrdquo Toxicon vol28 no 3 pp 261ndash273 1990
[48] E A Dennis J Cao Y-H Hsu V Magrioti and G KokotosldquoPhospholipaseA
2enzymes physical structure biological func-
tion disease implication chemical inhibition and therapeuticinterventionrdquo Chemical Reviews vol 111 no 10 pp 6130ndash61852011
[49] A G T Cardoso Acao do veneno de Bothrops moojeni e suafracao L-aminoacido oxidase submetida ao tratamento comraios gama de 60CO em Leishmania spp [MS thesis] IPEN-CNENSP Sao Paulo Brazil 1999
[50] MGallacci andWLGCavalcante ldquoUnderstanding the in vitroneuromuscular activity of snake venomLys49 phospholipaseA
2
homologuesrdquo Toxicon vol 55 no 1 pp 1ndash11 2010[51] S S Teixeira Caracterizacao funcional e estrutural de uma
fosfolipase A2acida isolada do veneno de Bothrops pirajai
[MS thesis] Faculdade de Ciencias Farmaceuticas de RibeiraoPretoUSP Ribeirao Preto Brazil 2009
[52] E Valentin and G Lambeau ldquoIncreasing molecular diversityof secreted phospholipases A
2and their receptors and binding
proteinsrdquo Biochimica et Biophysica Acta vol 1488 no 1-2 pp59ndash70 2000
[53] J M Gutierrez and B Lomonte ldquoPhospholipases A2from
Viperidae snake venoms how do they induce skeletal muscledamagerdquo Acta Chimica Slovenica vol 58 no 4 pp 647ndash6592011
[54] J M Gutierrez C L Ownby and G V Odell ldquoIsolation of amyotoxin from Bothrops asper venom partial characterizationand action on skeletal musclerdquo Toxicon vol 22 no 1 pp 115ndash128 1984
[55] N G Nascimento M C Sampaio R A Olivo and C TeixeiraldquoContribution of mast cells to the oedema induced by Bothropsmoojeni snake venom and a pharmacological assessment of theinflammatory mediators involvedrdquo Toxicon vol 55 no 2-3 pp343ndash352 2010
[56] C F P Teixeira E C T Landucci E Antunes M Chacurand Y Cury ldquoInflammatory effects of snake venom myotoxicphospholipases A
2rdquo Toxicon vol 42 no 8 pp 947ndash962 2003
[57] S R Zamuner and C F P Teixeira ldquoCell adhesion moleculesinvolved in the leukocyte recruitment induced by venom of thesnake Bothrops jararacardquoMediators of Inflammation vol 11 no6 pp 351ndash357 2002
[58] F Chaves C F P Teixeira and J M Gutierrez ldquoRole ofTNF-120572 IL-1120573 and IL-6 in the local tissue damage induced byBothrops asper snake venom an experimental assessment inmicerdquo Toxicon vol 45 no 2 pp 171ndash178 2005
BioMed Research International 13
[59] C D F P Teixeira C M Fernandes J P Zuliani and SF Zamuner ldquoInflammatory effects of snake venom metallo-proteinasesrdquo Memorias do Instituto Oswaldo Cruz vol 100supplement 1 pp 181ndash184 2005
[60] R D A Olivo C F P Teixeira J LWallace J M Gutierrez andS R Zamuner ldquoRole of cyclooxygenases in oedema-formingactivity of bothropic venomsrdquo Toxicon vol 49 no 5 pp 670ndash677 2007
[61] L B Silveira D P Marchi-Salvador N A Santos-Filho et alldquoIsolation and expression of a hypotensive and anti-plateletacidic phospholipase A
2from Bothrops moojeni snake venomrdquo
Journal of Pharmaceutical and Biomedical Analysis vol 73 pp35ndash43 2013
[62] A Rucavado T Escalante C F P Teixeira C M Fernandes CDıaz and J M Gutierrez ldquoIncrements in cytokines and matrixmetalloproteinases in skeletal muscle after injection of tissue-damaging toxins from the venom of the snake Bothrops asperrdquoMediators of Inflammation vol 11 no 2 pp 121ndash128 2002
[63] E Leiguez J P Zuliani A M Cianciarullo C M Fernandes JM Gutierrez and C Teixeira ldquoA group IIA-secreted phospho-lipase A
2from snake venom induces lipid body formation in
macrophages the roles of intracellular phospholipases A2and
distinct signaling pathwaysrdquo Journal of Leukocyte Biology vol90 no 1 pp 155ndash166 2011
[64] J P Zuliani J M Gutierrez L L Casais e Silva S C SampaioB Lomonte and C D F P Teixeira ldquoActivation of cellularfunctions in macrophages by venom secretory Asp-49 and Lys-49 phospholipasesA
2rdquoToxicon vol 46 no 5 pp 523ndash532 2005
[65] R D C M Escocard M M Kanashiro J H Petretski et alldquoNeutrophils regulate the expression of cytokines chemokinesand nitric oxide synthasenitric oxide in mice injected withBothrops atrox venomrdquo Immunobiology vol 211 no 1-2 pp 37ndash46 2006
[66] A Gambero E C T Landucci M H Toyama et al ldquoHumanneutrophil migration in vitro induced by secretory phospholi-pases A
2 a role for cell surface glycosaminoglycansrdquo Biochemi-
cal Pharmacology vol 63 no 1 pp 65ndash72 2002[67] F L S Costa R S Rodrigues L F M Izidoro et al ldquoBio-
chemical and functional properties of a thrombin-like enzymeisolated from Bothrops pauloensis snake venomrdquo Toxicon vol54 no 6 pp 725ndash735 2009
[68] J C Villalobos R Mora B Lomonte J M Gutierrez andY Angulo ldquoCytotoxicity induced in myotubes by a Lys49phospholipase A
2homologue from the venom of the snake
Bothrops asper evidence of rapid plasma membrane damageand a dual role for extracellular calciumrdquo Toxicology in Vitrovol 21 no 8 pp 1382ndash1389 2007
[69] B S Cummings J McHowat and R G Schnellmann ldquoPhos-pholipase A
2s in cell injury and deathrdquoThe Journal of Pharma-
cology and Experimental Therapeutics vol 294 no 3 pp 793ndash799 2000
[70] S R Panini L Yang A E Rusinol M S Sinensky J VBonventre and C C Leslie ldquoArachidonate metabolism andthe signaling pathway of induction of apoptosis by oxidizedLDLoxysterolrdquo Journal of Lipid Research vol 42 no 10 pp1678ndash1686 2001
[71] L Paramo B Lomonte J Pizarro-Cerda J-A Bengoechea J-P Gorvel and E Moreno ldquoBactericidal activity of Lys49 andAsp49 myotoxic phospholipases A
2from Bothrops asper snake
venom synthetic Lys49myotoxin II-(115-129)-peptide identifiesits bactericidal regionrdquo European Journal of Biochemistry vol253 no 2 pp 452ndash461 1998
[72] B Lomonte Y Angulo and E Moreno ldquoSynthetic peptidesderived from the C-terminal region of Lys49 phospholipaseA2homologues from viperidae snake venoms biomimetic
activities and potential applicationsrdquo Current PharmaceuticalDesign vol 16 no 28 pp 3224ndash3230 2010
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

12 BioMed Research International
relationshiprdquoCurrent Organic Chemistry vol 8 no 17 pp 1677ndash1690 2004
[31] VM Rodrigues SMarcussi R S Cambraia et al ldquoBactericidaland neurotoxic activities of two myotoxic phospholipases A
2
from Bothrops neuwiedi pauloensis snake venomrdquo Toxicon vol44 no 3 pp 305ndash314 2004
[32] N A Santos-Filho L B Silveira C Z Oliveira et al ldquoAnew acidic myotoxic anti-platelet and prostaglandin I
2induc-
tor phospholipase A2isolated from Bothrops moojeni snake
venomrdquo Toxicon vol 52 no 8 pp 908ndash917 2008[33] A M Soares S H Andriao-Escarso Y Angulo et al ldquoStruc-
tural and functional characterization of myotoxin I a Lys49phospholipaseA
2homologue fromBothropsmoojeni (Caissaca)
snake venomrdquo Archives of Biochemistry and Biophysics vol 373no 1 pp 7ndash15 2000
[34] A M Soares S H Andriao-Escarso R K Bortoleto et alldquoDissociation of enzymatic and pharmacological properties ofpiratoxins-I and -III two myotoxic phospholipases A
2from
Bothrops pirajai snake venomrdquo Archives of Biochemistry andBiophysics vol 387 no 2 pp 188ndash196 2001
[35] S S Teixeira L B Silveira F M N da Silva et al ldquoMolecularcharacterization of an acidic phospholipase A
2from Bothrops
pirajai snake venom synthetic C-terminal peptide identifies itsantiplatelet regionrdquo Archives of Toxicology vol 85 no 10 pp1219ndash1233 2011
[36] S H Andriao-Escarso A M Soares V M Rodrigues et alldquoMyotoxic phospholipases A
2in Bothrops snake venoms effect
of chemical modifications on the enzymatic and pharmacolog-ical properties of bothropstoxins from Bothrops jararacussurdquoBiochimie vol 82 no 8 pp 755ndash763 2000
[37] S H Andriao-Escarso A M Soares M R M Fontes et alldquoStructural and functional characterization of an acidic plateletaggregation inhibitor and hypotensive phospholipase A
2from
Bothrops jararacussu snake venomrdquo Biochemical Pharmacologyvol 64 no 4 pp 723ndash732 2002
[38] M E G Denegri O C Acosta S Huancahuire-Vega et alldquoIsolation and functional characterization of a new acidic PLA
2
Ba SpII RP4 of the Bothrops alternatus snake venom fromArgentinardquo Toxicon vol 56 no 1 pp 64ndash74 2010
[39] R Zouari-Kessentini J Luis A Karray et al ldquoTwo purified andcharacterized phospholipases A
2from Cerastes cerastes venom
that inhibit cancerous cell adhesion and migrationrdquo Toxiconvol 53 no 4 pp 444ndash453 2009
[40] R Shirai M Toriba K Hayashi K Ikeda and S Inoue ldquoIden-tification and characterization of phospholipase A
2inhibitors
from the serum of the Japanese rat snake Elaphe climacophorardquoToxicon vol 53 no 6 pp 685ndash692 2009
[41] M I Homsi-Brandenburgo L S Queiroz H Santo-Neto LRodrigues-Simoni and J R Giglio ldquoFractionation of Bothropsjararacussu snake venom partial chemical characterization andbiological activity of bothropstoxinrdquo Toxicon vol 26 no 7 pp615ndash627 1988
[42] G P Fernandez A Segura M Herrera et al ldquoNeutralizationof Bothrops mattogrossensis snake venom from Bolivia exper-imental evaluation of llama and donkey antivenoms producedby caprylic acid precipitationrdquoToxicon vol 55 no 2-3 pp 642ndash645 2010
[43] C L Ownby H S S de Araujo S P White and J E FletcherldquoLysine 49 phospholipase A
2proteinsrdquo Toxicon vol 37 no 3
pp 411ndash445 1999[44] A M Soares W P Sestito S Marcussi et al ldquoAlkylation of
myotoxic phospholipases A2in Bothrops moojeni venom a
promising approach to an enhanced antivenom productionrdquoInternational Journal of Biochemistry amp Cell Biology vol 36 no2 pp 258ndash270 2004
[45] B Lomonte J M Gutierrez M F Furtado et al ldquoIsolationof basic myotoxins from Bothrops moojeni and Bothrops atroxsnake venomsrdquo Toxicon vol 28 no 10 pp 1137ndash1146 1990
[46] L C Mancuso M M Correa C A Vieira et al ldquoFractionationof Bothrops pirajai snake venom isolation and characterizationof piratoxin-I a new myotoxic proteinrdquo Toxicon vol 33 no 5pp 615ndash626 1995
[47] H S Selistre L S Queiroz O A B Cunha G E P deSouza and J R Giglio ldquoIsolation and characterization ofhemorrhagic myonecrotic and edema-inducing toxins fromBothrops insularis (jararaca ilhoa) snake venomrdquo Toxicon vol28 no 3 pp 261ndash273 1990
[48] E A Dennis J Cao Y-H Hsu V Magrioti and G KokotosldquoPhospholipaseA
2enzymes physical structure biological func-
tion disease implication chemical inhibition and therapeuticinterventionrdquo Chemical Reviews vol 111 no 10 pp 6130ndash61852011
[49] A G T Cardoso Acao do veneno de Bothrops moojeni e suafracao L-aminoacido oxidase submetida ao tratamento comraios gama de 60CO em Leishmania spp [MS thesis] IPEN-CNENSP Sao Paulo Brazil 1999
[50] MGallacci andWLGCavalcante ldquoUnderstanding the in vitroneuromuscular activity of snake venomLys49 phospholipaseA
2
homologuesrdquo Toxicon vol 55 no 1 pp 1ndash11 2010[51] S S Teixeira Caracterizacao funcional e estrutural de uma
fosfolipase A2acida isolada do veneno de Bothrops pirajai
[MS thesis] Faculdade de Ciencias Farmaceuticas de RibeiraoPretoUSP Ribeirao Preto Brazil 2009
[52] E Valentin and G Lambeau ldquoIncreasing molecular diversityof secreted phospholipases A
2and their receptors and binding
proteinsrdquo Biochimica et Biophysica Acta vol 1488 no 1-2 pp59ndash70 2000
[53] J M Gutierrez and B Lomonte ldquoPhospholipases A2from
Viperidae snake venoms how do they induce skeletal muscledamagerdquo Acta Chimica Slovenica vol 58 no 4 pp 647ndash6592011
[54] J M Gutierrez C L Ownby and G V Odell ldquoIsolation of amyotoxin from Bothrops asper venom partial characterizationand action on skeletal musclerdquo Toxicon vol 22 no 1 pp 115ndash128 1984
[55] N G Nascimento M C Sampaio R A Olivo and C TeixeiraldquoContribution of mast cells to the oedema induced by Bothropsmoojeni snake venom and a pharmacological assessment of theinflammatory mediators involvedrdquo Toxicon vol 55 no 2-3 pp343ndash352 2010
[56] C F P Teixeira E C T Landucci E Antunes M Chacurand Y Cury ldquoInflammatory effects of snake venom myotoxicphospholipases A
2rdquo Toxicon vol 42 no 8 pp 947ndash962 2003
[57] S R Zamuner and C F P Teixeira ldquoCell adhesion moleculesinvolved in the leukocyte recruitment induced by venom of thesnake Bothrops jararacardquoMediators of Inflammation vol 11 no6 pp 351ndash357 2002
[58] F Chaves C F P Teixeira and J M Gutierrez ldquoRole ofTNF-120572 IL-1120573 and IL-6 in the local tissue damage induced byBothrops asper snake venom an experimental assessment inmicerdquo Toxicon vol 45 no 2 pp 171ndash178 2005
BioMed Research International 13
[59] C D F P Teixeira C M Fernandes J P Zuliani and SF Zamuner ldquoInflammatory effects of snake venom metallo-proteinasesrdquo Memorias do Instituto Oswaldo Cruz vol 100supplement 1 pp 181ndash184 2005
[60] R D A Olivo C F P Teixeira J LWallace J M Gutierrez andS R Zamuner ldquoRole of cyclooxygenases in oedema-formingactivity of bothropic venomsrdquo Toxicon vol 49 no 5 pp 670ndash677 2007
[61] L B Silveira D P Marchi-Salvador N A Santos-Filho et alldquoIsolation and expression of a hypotensive and anti-plateletacidic phospholipase A
2from Bothrops moojeni snake venomrdquo
Journal of Pharmaceutical and Biomedical Analysis vol 73 pp35ndash43 2013
[62] A Rucavado T Escalante C F P Teixeira C M Fernandes CDıaz and J M Gutierrez ldquoIncrements in cytokines and matrixmetalloproteinases in skeletal muscle after injection of tissue-damaging toxins from the venom of the snake Bothrops asperrdquoMediators of Inflammation vol 11 no 2 pp 121ndash128 2002
[63] E Leiguez J P Zuliani A M Cianciarullo C M Fernandes JM Gutierrez and C Teixeira ldquoA group IIA-secreted phospho-lipase A
2from snake venom induces lipid body formation in
macrophages the roles of intracellular phospholipases A2and
distinct signaling pathwaysrdquo Journal of Leukocyte Biology vol90 no 1 pp 155ndash166 2011
[64] J P Zuliani J M Gutierrez L L Casais e Silva S C SampaioB Lomonte and C D F P Teixeira ldquoActivation of cellularfunctions in macrophages by venom secretory Asp-49 and Lys-49 phospholipasesA
2rdquoToxicon vol 46 no 5 pp 523ndash532 2005
[65] R D C M Escocard M M Kanashiro J H Petretski et alldquoNeutrophils regulate the expression of cytokines chemokinesand nitric oxide synthasenitric oxide in mice injected withBothrops atrox venomrdquo Immunobiology vol 211 no 1-2 pp 37ndash46 2006
[66] A Gambero E C T Landucci M H Toyama et al ldquoHumanneutrophil migration in vitro induced by secretory phospholi-pases A
2 a role for cell surface glycosaminoglycansrdquo Biochemi-
cal Pharmacology vol 63 no 1 pp 65ndash72 2002[67] F L S Costa R S Rodrigues L F M Izidoro et al ldquoBio-
chemical and functional properties of a thrombin-like enzymeisolated from Bothrops pauloensis snake venomrdquo Toxicon vol54 no 6 pp 725ndash735 2009
[68] J C Villalobos R Mora B Lomonte J M Gutierrez andY Angulo ldquoCytotoxicity induced in myotubes by a Lys49phospholipase A
2homologue from the venom of the snake
Bothrops asper evidence of rapid plasma membrane damageand a dual role for extracellular calciumrdquo Toxicology in Vitrovol 21 no 8 pp 1382ndash1389 2007
[69] B S Cummings J McHowat and R G Schnellmann ldquoPhos-pholipase A
2s in cell injury and deathrdquoThe Journal of Pharma-
cology and Experimental Therapeutics vol 294 no 3 pp 793ndash799 2000
[70] S R Panini L Yang A E Rusinol M S Sinensky J VBonventre and C C Leslie ldquoArachidonate metabolism andthe signaling pathway of induction of apoptosis by oxidizedLDLoxysterolrdquo Journal of Lipid Research vol 42 no 10 pp1678ndash1686 2001
[71] L Paramo B Lomonte J Pizarro-Cerda J-A Bengoechea J-P Gorvel and E Moreno ldquoBactericidal activity of Lys49 andAsp49 myotoxic phospholipases A
2from Bothrops asper snake
venom synthetic Lys49myotoxin II-(115-129)-peptide identifiesits bactericidal regionrdquo European Journal of Biochemistry vol253 no 2 pp 452ndash461 1998
[72] B Lomonte Y Angulo and E Moreno ldquoSynthetic peptidesderived from the C-terminal region of Lys49 phospholipaseA2homologues from viperidae snake venoms biomimetic
activities and potential applicationsrdquo Current PharmaceuticalDesign vol 16 no 28 pp 3224ndash3230 2010
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

BioMed Research International 13
[59] C D F P Teixeira C M Fernandes J P Zuliani and SF Zamuner ldquoInflammatory effects of snake venom metallo-proteinasesrdquo Memorias do Instituto Oswaldo Cruz vol 100supplement 1 pp 181ndash184 2005
[60] R D A Olivo C F P Teixeira J LWallace J M Gutierrez andS R Zamuner ldquoRole of cyclooxygenases in oedema-formingactivity of bothropic venomsrdquo Toxicon vol 49 no 5 pp 670ndash677 2007
[61] L B Silveira D P Marchi-Salvador N A Santos-Filho et alldquoIsolation and expression of a hypotensive and anti-plateletacidic phospholipase A
2from Bothrops moojeni snake venomrdquo
Journal of Pharmaceutical and Biomedical Analysis vol 73 pp35ndash43 2013
[62] A Rucavado T Escalante C F P Teixeira C M Fernandes CDıaz and J M Gutierrez ldquoIncrements in cytokines and matrixmetalloproteinases in skeletal muscle after injection of tissue-damaging toxins from the venom of the snake Bothrops asperrdquoMediators of Inflammation vol 11 no 2 pp 121ndash128 2002
[63] E Leiguez J P Zuliani A M Cianciarullo C M Fernandes JM Gutierrez and C Teixeira ldquoA group IIA-secreted phospho-lipase A
2from snake venom induces lipid body formation in
macrophages the roles of intracellular phospholipases A2and
distinct signaling pathwaysrdquo Journal of Leukocyte Biology vol90 no 1 pp 155ndash166 2011
[64] J P Zuliani J M Gutierrez L L Casais e Silva S C SampaioB Lomonte and C D F P Teixeira ldquoActivation of cellularfunctions in macrophages by venom secretory Asp-49 and Lys-49 phospholipasesA
2rdquoToxicon vol 46 no 5 pp 523ndash532 2005
[65] R D C M Escocard M M Kanashiro J H Petretski et alldquoNeutrophils regulate the expression of cytokines chemokinesand nitric oxide synthasenitric oxide in mice injected withBothrops atrox venomrdquo Immunobiology vol 211 no 1-2 pp 37ndash46 2006
[66] A Gambero E C T Landucci M H Toyama et al ldquoHumanneutrophil migration in vitro induced by secretory phospholi-pases A
2 a role for cell surface glycosaminoglycansrdquo Biochemi-
cal Pharmacology vol 63 no 1 pp 65ndash72 2002[67] F L S Costa R S Rodrigues L F M Izidoro et al ldquoBio-
chemical and functional properties of a thrombin-like enzymeisolated from Bothrops pauloensis snake venomrdquo Toxicon vol54 no 6 pp 725ndash735 2009
[68] J C Villalobos R Mora B Lomonte J M Gutierrez andY Angulo ldquoCytotoxicity induced in myotubes by a Lys49phospholipase A
2homologue from the venom of the snake
Bothrops asper evidence of rapid plasma membrane damageand a dual role for extracellular calciumrdquo Toxicology in Vitrovol 21 no 8 pp 1382ndash1389 2007
[69] B S Cummings J McHowat and R G Schnellmann ldquoPhos-pholipase A
2s in cell injury and deathrdquoThe Journal of Pharma-
cology and Experimental Therapeutics vol 294 no 3 pp 793ndash799 2000
[70] S R Panini L Yang A E Rusinol M S Sinensky J VBonventre and C C Leslie ldquoArachidonate metabolism andthe signaling pathway of induction of apoptosis by oxidizedLDLoxysterolrdquo Journal of Lipid Research vol 42 no 10 pp1678ndash1686 2001
[71] L Paramo B Lomonte J Pizarro-Cerda J-A Bengoechea J-P Gorvel and E Moreno ldquoBactericidal activity of Lys49 andAsp49 myotoxic phospholipases A
2from Bothrops asper snake
venom synthetic Lys49myotoxin II-(115-129)-peptide identifiesits bactericidal regionrdquo European Journal of Biochemistry vol253 no 2 pp 452ndash461 1998
[72] B Lomonte Y Angulo and E Moreno ldquoSynthetic peptidesderived from the C-terminal region of Lys49 phospholipaseA2homologues from viperidae snake venoms biomimetic
activities and potential applicationsrdquo Current PharmaceuticalDesign vol 16 no 28 pp 3224ndash3230 2010
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of





























