Embed Size (px)
Citation preview

Journal of Cell Science 102, 857-865 (1992)Printed in Great Britain © The Company of Biologists Limited 1992
857
Isoprenylation of rab proteins on structurally distinct cysteine motifs
MATTHIAS PETER2, PHILIPPE CHAVRIER1, ERICH A. NIGG2 and MARINO ZERIAL1-*
' European Molecular Biology Laboratory (EMBL), Meyerhofstrasse 1, Postfach 10.2209, D-6900 Heidelberg, Germany2Swiss Institute for Experimental Cancer Research (ISREC), 155, Chemin des Boveresses, CH-1066 Epalinges, Switzerland
•Author for correspondence
Summary
rab proteins are low molecular weight GTP-bindingproteins highly related to Yptlp and Sec4p, which areinvolved in the control of secretion in yeast Saccharo-myces cerevisiae. Morphological and biochemical studieshave shown that rab proteins are membrane associatedand are localized to specific subcompartments along theexocytic and endocytic pathway. Membrane associationrequires the presence of C-tcrminal cysteine residues.The present report indicates that the structurallydistinct cysteine motifs of rab proteins are subjected toisoprenylation both in vitro and in vivo. Studies on
deletion mutants suggest that an intact C-terminal end isrequired for the association of rab proteins with themembrane and is necessary for the post-translationalmodification. Finally, we show that the isoprenoidtransferase which modifies rab termini is different fromthe enzyme which farnesylates nuclear lamins and rasproteins in vitro.
Key words: isoprenylation, rab proteins, membraneassociation.
Introduction
Several recent in vitro and in vivo studies have shownthat the superfamily of ras-related low molecular weightGTP-binding proteins includes a subset of proteinswhich are involved in the control of membrane traffic(see Bourne et al., 1990, for a review). In the yeastSaccharomyces cerevisiae, the SEC4 gene encodes aGTP-binding protein associated with both the cytoplas-mic surface of the plasma membrane and secretoryvesicles, which is involved in the regulation of vesiculartraffic from the Golgi apparatus to the plasma mem-brane (Salminen and Novick, 1987; Goud et al., 1988).The YPT1 gene product (Gallwitz et al., 1983)functions at an earlier step of the secretory pathway,from the ER (endoplasmic reticulum) to, or within, theGolgi apparatus (Segev et al., 1988; Baker et al., 1990).
Mammalian cells express a large number of lowmolecular weight GTP-binding proteins (rab proteins)highly related to Yptlp and Sec4p (Valencia et al.,1991). Owing to this sequence similarity they have beenfunctionally implicated in the regulation of membranetraffic in mammalian cells. In support of this idea, rablhas been shown to functionally replace Yptlp inSaccharomyces cerevisiae (Haubruck et al., 1989) andstudies on rab5 have indicated that this protein isinvolved in the process of early endosome fusion in vitro(Gorvel et al., 1991).
rab proteins have been found associated with specificsubcompartments along the exocytic and endocytic
pathways. rab2 was localized to an "intermediatecompartment" between the ER and the Golgi appar-atus, probably equivalent to the so-called "salvagecompartment" postulated to recycle ER-resident pro-teins to the ER (Chavrier et al., 1990a). rab6 waslocalized to the middle- and fra/w-Golgi cisternae(Goud et al., 1990). rab3a (called also smg25A) wasfound associated with specialized organelles of theregulated secretory pathway: synaptic vesicles in neur-ons and chromaffin granules in adrenal medulla(Fischer von Mollard et al., 1990; Darchen et al., 1990;Mizoguchi et al., 1990). Three other rab proteins werefound associated with compartments along the endocy-tic pathway: rab5 was detected at the cytoplasmicsurface of both the plasma membrane and earlyendosomes (Chavrier et al., 1990a), whereas rab4 wasassociated with early endosomes (Van der Sluijs et al.,1991) and rab7 with late endosomes (Chavrier et al.,1990a).
The mechanism responsible for the membrane associ-ation of these different GTP-binding proteins is not yetclear. Like all members of the ras protein superfamily,rab proteins share four highly conserved regions, whichcooperate to form the GTP-binding site, and theycontain one or two cysteine residues near their Ctermini (Valencia et al., 1991). In the case of p21H"ras,three post-translational modifications of the C-terminalcysteine motif, the so-called CAAX box (C, cysteine;A, aliphatic; X, any amino acid), have been shown tooccur and are required for membrane association:
858 M. Peter and others
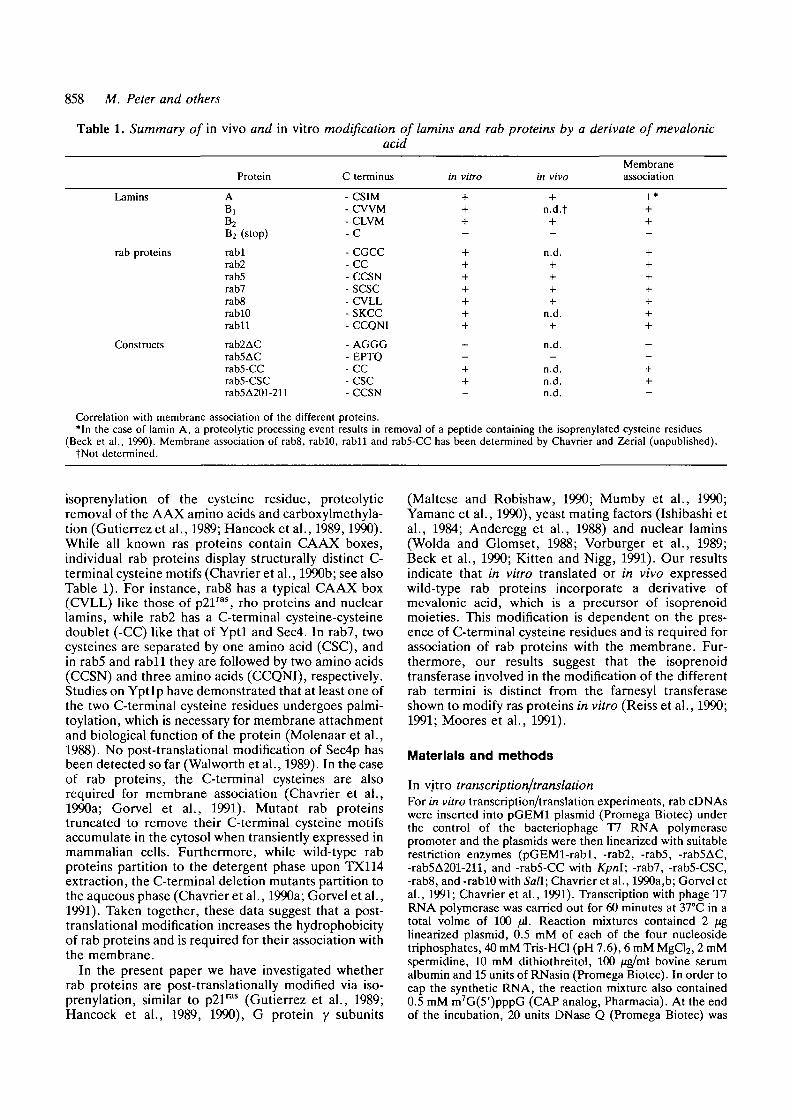
Table 1. Summary of in vivo and in vitro modification of lamins and rab proteins by a derivate of mevalonicacid
Protein C terminus in vitroMembraneassociation
Lamins
rab proteins
Constructs
ABiB2B2 (stop)
rablrab2rab5rab7rab8rablOrabll
rab2ACrab5ACrab5-CCrab5-CSCrab5A201-211
-CSIM- C W M-CLVM-C
-CGCC-CC-CCSN-SCSC- CVLL-SKCC- CCQNI
- AGGG- EPTQ- C C-CSC-CCSN
n.d.t
n.d.
+ n.d. +
n.d.
+ n.d. ++ n.d. +
n.d.
Correlation with membrane association of the different proteins.•In the case of lamin A, a proteolytic processing event results in removal of a peptide containing the isoprenylated cysteine residues
(Beck et al., 1990). Membrane association of rabS, rablO, rabll and rab5-CC has been determined by Chavrier and Zerial (unpublished).tNot determined.
isoprenylation of the cysteine residue, proteolyticremoval of the AAX amino acids and carboxylmethyla-tion (Gutierrez etal., 1989; Hancock etal., 1989,1990).While all known ras proteins contain CAAX boxes,individual rab proteins display structurally distinct C-termi nal cysteine motifs (Chavrier et al., 1990b; see alsoTable 1). For instance, rab8 has a typical CAAX box(CVLL) like those of p21ras, rho proteins and nuclearlamins, while rab2 has a C-terminal cysteine-cysteinedoublet (-CC) like that of Yptl and Sec4. In rab7, twocysteines are separated by one amino acid (CSC), andin rab5 and rabll they are followed by two amino acids(CCSN) and three amino acids (CCQNI), respectively.Studies on Yptlp have demonstrated that at least one ofthe two C-terminal cysteine residues undergoes palmi-toylation, which is necessary for membrane attachmentand biological function of the protein (Molenaar et al.,1988). No post-translational modification of Sec4p hasbeen detected so far (Walworth et al., 1989). In the caseof rab proteins, the C-terminal cysteines are alsorequired for membrane association (Chavrier et al.,1990a; Gorvel et al., 1991). Mutant rab proteinstruncated to remove their C-terminal cysteine motifsaccumulate in the cytosol when transiently expressed inmammalian cells. Furthermore, while wild-type rabproteins partition to the detergent phase upon TX114extraction, the C-terminal deletion mutants partition tothe aqueous phase (Chavrier et al., 1990a; Gorvel et al.,1991). Taken together, these data suggest that a post-translational modification increases the hydrophobicityof rab proteins and is required for their association withthe membrane.
In the present paper we have investigated whetherrab proteins are post-translationally modified via iso-prenylation, similar to p21ras (Gutierrez et al., 1989;Hancock et al., 1989, 1990), G protein y subunits
(Maltese and Robishaw, 1990; Mumby et al., 1990;Yamane et al., 1990), yeast mating factors (Ishibashi etal., 1984; Anderegg et al., 1988) and nuclear lamins(Wolda and Glomset, 1988; Vorburger et al., 1989;Beck et al., 1990; Kitten and Nigg, 1991). Our resultsindicate that in vitro translated or in vivo expressedwild-type rab proteins incorporate a derivative ofmevalonic acid, which is a precursor of isoprenoidmoieties. This modification is dependent on the pres-ence of C-terminal cysteine residues and is required forassociation of rab proteins with the membrane. Fur-thermore, our results suggest that the isoprenoidtransferase involved in the modification of the differentrab termini is distinct from the farnesyl transferaseshown to modify ras proteins in vitro (Reiss et al., 1990;1991; Mooreset al., 1991).
Materials and methods
In vitro transcription/translationFor in vitro transcription/translation experiments, rab cDNAswere inserted into pGEMl plasmid (Promega Biotec) underthe control of the bacteriophage T7 RNA polymerasepromoter and the plasmids were then linearized with suitablerestriction enzymes (pGEMl-rabl, -rab2, -rab5, -rab5AC,-rab5A201-211, and -rab5-CC with Kpnl; -rab7, -rab5-CSC,-rab8, and -rablOwith Sail; Chavrier et al., 1990a,b; Gorvel etal., 1991; Chavrier et al., 1991). Transcription with phage T7RNA polymerase was carried out for 60 minutes at 37°C in atotal volme of 100 /d. Reaction mixtures contained 2 /iglinearized plasmid, 0.5 mM of each of the four nucleosidetriphosphates, 40 mM Tris-HCl (pH 7.6), 6 mM MgCl2, 2 mMspermidine, 10 mM dithiothreitol, 100 fig/ml bovine serumalbumin and 15 units of RNasin (Promega Biotec). In order tocap the synthetic RNA, the reaction mixture also contained0.5 mM m7G(5')pppG (CAP analog, Pharmacia). At the endof the incubation, 20 units DNase Q (Promega Biotec) was

Isoprenylation of rab proteins 859
added, and the mixture was incubated for an additional 10minutes at 37°C. Then the sample was extracted withphenol/chloroform (1:1, v/v) and the RNA ethanol precipi-tated. The RNA was dissolved in 30 /A water.
In vitro translations were carried out in reticulocyte lysatefor 90 minutes according to the instructions provided by themanufacturers (Promega Biotec). Approximately 400 ng ofsynthetic RNA was used in 50 p\ reactions. Translatedproteins were labelled using either [35S]methionine or R-[2-
C]mevalonic acid lactone (56.7 mCi/mmol; Amersham,CFA.660) as described previously (Vorburger et al., 1989).For inhibition experiments, synthetic peptides (dissolved in 10mM Tris-HG, pH 7.4, 10 mM dithiothreitol) were added atfinal concentrations as indicated (0-50 /JM) or in the case ofthe rab5 C-terminal peptides, at a concentration of 10 fM-1mM. The reactions were stopped by the addition of 50 JX\sample buffer and boiling for 5 min or by adding 500 fd of theappropriate immunoprecipitation buffer (see below).
Antibodies and immunoprecipitationsThe monoclonal antibody (E3) against lamin B2 as well asanti-rab2, -rab5 and -rab7 rabbit antisera have been charac-terized previously (Lehner et al., 1986; Chavrier et al.,1990a). Rabbit anti-rab8 and anti-rablO sera were raisedagainst a MSIl-rab8 fusion protein as previously described(Chavrier et al., 1990a) and a peptide derived from the C-terminal region of rab 10 (KTPVKEPNSENVDIS), respect-ively. A 100 /A sample of this rabbit sera was incubatedovernight at 4°C with 100 /A Protein A-Sepharose (Pierce)with gentle agitation. Then the serum was removed and thebeads were washed three times with NP40 buffer (0.5% NP40,50 mM Tris-HCI, pH 7.5,150 mM NaCl, 2 mM EDTA, 1 mMPMSF (phenylmethylsulfonyl fluoride, 1% Aprotinin). Forimmunoprecipitation of the rab proteins, 500 /A NP40 buffer(see above) was added to each in vitro translation reaction. Toremove insoluble material, the lysates were centrifuged for 10minutes at 17,000g. Then, 40 jA antibody bound to Protein A-Sepharose (see above) was added to the supernatant and themixture was incubated overnight at 4°C with gentle agitation.The Sepharose beads were then washed three times with ice-cold NP40 buffer, once with high salt buffer (50 mM Tris-HCI,pH 7.5,100 mM NaCl, 0.5% Triton X-100) and once with PBS(137 mM NaCl, 2.7 mM KC1, 8.1 mM Na2HPO4, 1.5 mMKH2HPO4, pH 7.2). The immunoprecipitated proteins werereleased with 50 (i\ sample buffer (0.01% Bromophenol blue,60 mM Tris-HCI, pH 6.8,10% glycerol, 2% SDS, 5% (v/v) 0-mercaptoethanol) and boiled for 5 minutes. Lamin B2 wasimmunoprecipitated basically as described previously (Kittenand Nigg, 1991). Briefly, 500 [A of RIPA buffer (1% Triton X-100, 1% sodium deoxycholate, 0.1% SDS, 150 mM NaCl, 20mM Tris-HCI, pH 7.2, 1 mM PMSF, 1% Aprotinin) wasadded to the translation reaction and insoluble material wasremoved by centrifugation as described above. The super-natant was then incubated for one hour on ice with 2 \A E3ascites fluid. Immuncomplexes were isolated by centrifu-gation after incubation for one hour at 4°C with goat anti-mouse antibodies coupled to Sepharose (Sigma). The Sepha-rose beads were washed as above and the immunoprecipitatedlamin B2 was released from the Sepharose beads by boiling for5 minutes in 50 \A sample buffer. The immunoprecipitatedproteins were analysed by SDS-PAGE and fluorography.
Cell culture, metabolic labeling and transfectionConditions for BHK21 cells culture, infection with the T7RNA polymerase recombinant vaccinia virus and transfectionwere as previously described (Chavrier et al., 1990a). Twohours after transfection cells were washed twice in MEM
methionine-free medium and incubated for 4 hours in thesame medium containing 100 juCi/ml [35S]methionine (Amer-sham) in the presence or absence of 30 pM Mevinolin (giftfrom Dr A. W. Alberts, Merck, Sharp & Dohme, Rehway,NJ). For [14C]isoprenoid labelling, cells were incubated 2hours after transfection in GMEM medium containing 5%foetal calf serum, 15 /iCi/ml [14C]mevalonic acid lactone(Amersham) for 9 hours. Cells were lysed in 1% Triton X-100, 50 mM Tris-HCI, pH 7.5,150 mM NaCl, 2 mM EDTA, 20jug/ml PMSF. Post-nuclear supernatants were adjusted to 2%Triton X-100, 0.4% SDS and rab proteins immunoprecipi-tated as described above.
Results
Lamin and rab proteins translated in reticulocytelysate specifically incorporate a derivative of[14C]mevalonic acidIn previous studies it was shown that Cys597 located inthe CAAX box of chicken lamin B2 is specificallyisoprenylated in vivo and that this modification isrequired for proper association of lamin B2 with theinner nuclear membrane (Kitten and Nigg, 1991). Asdiscovered by Vorburger et al. (1989) isoprenylationoccurs also in vitro, following translation of lamin B2 ina reticulocyte lysate (Fig. 1A, lane 2). In contrast, amutant form of lamin B2, which lacks the three terminalamino acids of the CAAX box (598 STOP codonmutant called B2 stop), is neither isoprenylated (Fig.1A, lane 4) nor associated with the nuclear envelope(Kitten and Nigg, 1991).
Since these results suggested that the in vivospecificity of isoprenylation reactions is faithfullyreflected in the reticulocyte lysate, we proceeded to testwhether members of the rab protein family, which carrydifferent C-terminal motifs, would also be modified by amevalonic acid derivative in vitro. For this purpose,various rab proteins were in vitro translated in reticulo-cyte lysate in the presence of either [35S]methionine or[ CJmevalonic acid, then immunoprecipitated usingspecific anti-rab antibodies (Chavrier et al., 1990a) andanalysed after SDS-PAGE and autoradiography. Bothrab2 and rab5 proteins are labelled in the presence of[14C]mevalonic acid (Fig. IB, lanes 6 and 10). Incontrast, similar to the lamin B2598 STOP mutant, thein vitro translated deletion mutants of rab2 (rab2 A) andrab5 (rab5A) proteins (Fig. IB, lanes 7 and 11), wherethe C-terminal Cys residues have been deleted (Chav-rier et al., 1990a; Gorvel et al., 1991), are not modifiedby an isoprene residue (Fig. IB, lanes 8 and 12). Thisresult strongly suggests that the C-terminal cysteineresidues are required for modification by mevalonicacid. The fact that the C-terminal motifs of rab2(GGCC) and rab5 (CCSN) do not fit the classicalCAAX motif indicates that other types of C-terminalmotifs are substrates for isoprenylation in the reticulo-cyte lysate.
To determine whether other rab proteins are simi-larly modified we extended our analysis to rab7 (Fig. 1,lanes 13 and 14) and rab8 (Fig. 1, lanes 15 and 16).Whereas rab8 bears a "CAAX"-motif at its carboxy

860 M. Peter and others
B2 (wt) B2 (stop) rab 2 rab 2A rab 5 rab 5A rab7 rab 8
Met MVA Met MVA Met MVA Met MVA Met MVA Met MVA Met MVA Met MVA
B ,C
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16Fig. 1. Specific in vitro processing of lamin B2, rab2, rab5, rab7 and rab8. Lamin B2 and rab proteins were in vitrotranslated in reticulocyte lysates, in the presence of either [35S]methionine (lanes 1, 3, 5, 7, 9, 11, 13 and 15) or R-[l4C]mevalonic acid lactone (lanes 2, 4, 6, 8, 10, 12, 14 and 16). Labelled proteins were immunoprecipitated and analysedby SDS-PAGE and fluorography. (A) Lanes 1 and 2, lamin B2; lanes 3 and 4, lamin B2 STOP mutant. (B) Lanes 5 and 6,rab2; lanes 7 and 8, rab2 C-terminal deletion; lanes 9 and 10, rab5; lanes 11 and 12, rab5 C-terminal deletion. (C) Lanes13 and 14, rab7; lanes 15 and 16, rab8. Note that none of the deletion mutant proteins is labelled by [14C]mevalonic acid.
terminus (CVLL), rab7 has yet a different terminus ofthe "CXC" type (Chavrier et al., 1990b). Clearly, bothproteins incorporate mevalonic acid label (Fig. 1, lane14 and 16). In addition we found that rabl (GGCC),rablO (SKCC), both ending with a cysteine doublet,and rabll (CCQNI) are isoprenylated in vitro (data notshown). Taken together, these data demonstrate thatrab proteins with different types of C-terminal motifsare isoprenylated in vitro.
We next determined whether mutations within or inthe vicinity of the C-terminal motifs affect isoprenyla-tion. For this purpose we constructed rab5 mutantproteins that carry either a deletion of the two C-terminal amino acid residues (rab5-CC) or the cysteinemotif replaced with that from rab7 (rab5-CSC). Asshown in Fig. 2, both in vitro translated mutant proteinsincorporate [14C]mevalonic acid (lanes 4 and 5).However, a deletion mutant of rab5 lacking ten aminoacids immediately upstream from the C-terminalCCSN-motif (rab5A201-211), is not labelled in thepresence of [14C]mevalonic acid (Fig. 2, lane 6). Thesedata indicate that while the presence of a carboxy-terminal cysteine motif is necessary for isoprenylation,this alone is not sufficient, since deletion of upstreamsequences abolishes isoprenylation.
rab proteins are isoprenylated in vivoHaving established that different rab proteins areisoprenylated in vitro, it was important to confirm thatthey are similarly modified in vivo. For this purpose,rab2, rab5, rab5AC, rab7, rab8 and rabll proteins were
transiently expressed in BHK21 cells, using the recom-binant vaccinia T7 RNA polymerase system (Fuerst etal., 1986). BHK21 cells infected with this virus werethen transfected with plasmids containing the variousrab inserts under the control of the T7 promoter. Twohours after transfection, cells were incubated for eitherfour or nine hours in medium containing [35S]methio-nine or [14C]mevalonic acid, respectively. Then, theywere lysed in NP40 buffer, and the different rabproteins were immunoprecipitated using anti-rab-specific antibodies. Immunoprecipitated rab5, rab2 andrab7 proteins labelled with [ Sjmethionine migrated asclear doublets on SDS-polyacrylamide gels (Fig. 3).When cells were labelled with [35S]methionine in thepresence of Mevinolin, an inhibitor of the isoprenoidbiosynthesis pathway, only the upper band of thedoublet was visible. This behaviour is very similar to theone previously observed with chicken lamin B2 (Kittenand Nigg, 1991; see below) and suggests that the lowerband corresponds to the isoprenylated form of rabproteins. In agreement with this interpretation, allimmunoprecipitated rab proteins were also labelledwith [14C]mevalonic acid (Fig. 3) and co-migrated withthe lower band of the doublet. In contrast, rab5AClacking the two C-terminal cysteine residues (Fig. 3,lane 6) and the internal deletion mutant rab5A201-211(not shown) are not modified in vivo and do not shift inelectrophoretic mobility upon Mevinolin treatment.These results demonstrate that rab2, rab5, rab7, rab8 aswell as rabll, are modified by a derivative of mevalonicacid not only in vitro, but also in vivo, and we conclude

Met MVA
oonQ
OIf)uin
i
inS3
uuinn
uoin m
CO (0 CO (0 CO CQ
Isoprenylation of rab proteins 861
Fig. 2. In vitro processing of different rab5 mutantproteins. In vitro transcribed RNAs were translated inreticolocyte lysates in the presence of either[35S]methionine (lanes 1, 2 and 3) or R-[14C]mevalonic acidlactone (lanes 4, 5 and 6) as described in the legend to Fig.1. The labelled proteins were immunoprecipitated andanalysed after SDS-PAGE and fluorography. The followingtruncated versions of rab5 were analysed. Lanes 1 and 4,rab5-CC; lanes 2 and 5, rab5-CSC; lanes 3 and 6, rab5deletion 201-211. Note that the ten amino acid deletion ofrab5 which leaves the C-terminal CCSN motif abolishesisoprenylation of rab5, despite the presence of an intact C-terminal motif.
1 2 3 4 5 6
that all three different C-terminal motifs (-CC, -CXC,-CCXX) of rab proteins are subject to isoprenylation invivo (see also Table 1).
Isoprenylation of the different rab proteins is notinhibited by a synthetic lamin CSIM peptideAddition of increasing amounts of a CSIM peptide,synthesized to mimic the C terminus of chicken lamin A(Peter et al., 1989), to in vitro translated chicken[ S]labelled lamin B2 quantitatively shifted this proteinto its slower migrating non-modified form (Fig. 4A).Under the same conditions, the peptide inhibited[14C]mevalonate labelling of chicken lamin B2 (Fig. 4B,lane 9). Similar peptides, when used at a comparableconcentration (4 /iM), have been shown to inhibitpurified farnesyltransferase, in good agreement withtheir inhibition constants (Reiss et al., 1990).
Having established that the CSIM peptide specificallyinhibits isoprenylation of chicken lamin B2, we nextasked whether synthetic peptides mimicking the C-terminal motifs of rab2 (GGCC), rab5 (CCSN) or rab7
rabS
35 c 14,
MtvtooUn + -
rabSAC
35 s l 4 c• - I I
- +
rab2
35 r
rab7 rab8 rabll
351 14, 1 4 1 4
S i -
I8 0 10 11 12 13 15 16 17 18
Fig. 3. In vivo labelling of rab2, rab5, rab7, rab8 and rabll proteins by [14C]mevalonate. BHK21 cells were infected withthe T7 RNA polymerase recombinant virus and transfected with the appropriate plasmids as previously described (Chavrieret al., 1990a). Cells were then labelled in the presence of [35S]methionine (lanes 1-2, 4-5, 7-8, 10-11, 13-14 and 16-17) or[14C]mevalonic acid lactone (3, 6, 9, 12, 15 and 18), in the absence (—) or the presence (+) of mevinolin. Cells were lysedand the rab5 (lanes 1-3), rab5AC (4-6), rab2 (7-9), rab7 (10-12), rab8 (13-15), and rabll (16-18) proteins wereimmunoprecipitated. Samples were analysed by 10% SDS-PAGE and fluorography.

862 M. Peter and others
CSIM LSPTR
ft ml*
GGCC CSIM
8 8
B
rab 2 rab 5 rab 7 rab 8
10 10 10 10
8 9
GGCC CCSN SCSC CSIM
10 50 0 10 50 0 10 50 10
6 8 9 10
Fig. 4. The synthetic peptide CSIM specifically abolishesmodification of lamin B^. Synthetic lamin B2 RNA wastranslated in reticulocyte lysates in the presence of varioustetrapeptides at final concentrations between 0 and 50 /̂ M(as indicated). Proteins labelled with [35S]methionine (Aand C) were analysed directly, whereas proteins modifiedby R-[14C]mevalonic acid (B) were concentrated byimmunoprecipitation. Samples were analysed by 8% SDS-PAGE and fluorography. The following peptides wereused: (A) lanes 1-4, CSIM; lane 5, LSPTR. (B) Lane 8,GGCC; lane 9, CSIM. (C) Lanes 1-3, GGCC; lanes 4-6,CCSN; lanes 7-9, SCSC; lane 10, CSIM. Note that incontrast to the lamin A-CSIM peptide, none of the testedrab peptides inhibits the shift of lamin B2 to the fastermigrating form.
(SCSC) are also able to inhibit lamin B2 isoprenylation.As shown in Fig. 4C none of these peptides (up toconcentrations of 50 ;uM) significantly interfered withthe conversion of lamin B2 to the faster migrating form.
1«2 215PKNEPQNPG AN SARGRG VDLTEPTQPTR SQCCSN
CCSNPTRSQCCSN
LTEPTQPTRSQCCSNLTEPTQPTRSQ
PKNEPQNPG ANSARGR
1S2-2 152 12-2 15207-215201-215201-2111*2-197
>
8
Fig. 5. The lamin CSIM-peptide does not inhibit themodification of rab2, rab5, rab7 and rab8. rab2 (lanes 1and 2), rab5 (lanes 3 and 4), rab7 (lanes 4 and 5) and rab8proteins (lanes 6 and 8) were translated in reticulocytelysates in the presence (lanes 2, 4, 6 and 8) or absence ofCSIM peptide (lanes 1, 3, 5 and 7) at a final concentrationof 10 /*M. The R-[14C]mevalonic acid-labelled rab proteinswere then concentrated by immunoprecipitation andvisualized by SDS-PAGE and fluorography.
Conversely, lamin CSIM peptide (at 10 /M) did notinterfere with the incorporation of [14C]mevalonic acidinto the C-terminal motif of in vitro translated rab2,rab5, rab7 and rab8 proteins (Fig. 5). Also, none of thetetrapeptides synthesized according to the C-terminalmotif of rab proteins was able to inhibit labelling of rabproteins or lamin B2 by [14C]mevalonic acid (not shownand Fig. 6). We then investigated whether differentpeptides derived from the C-terminal sequence of rab5could inhibit isoprenylation of in vitro translated rab5.Fig. 6A displays the amino acid sequence of thepeptides used in this experiment. Peptides correspond-ing to the last 4 (212-215), 9 (207-215) and 15 (201-215)C-terminal amino acids of rab5 did not affect labellingof rab5 by [14C]mevalonic acid even at high concen-tration (1 mM). Likewise, no effect on isoprenylationwas caused by peptide 201-211, which corresponds to
Fig. 6. Inhibition of rab5isoprenylation by rab5 C-terminal peptides. In vitrotranscribed rab5 RNAs wastranslated in reticolocytelysates in the presence ofvarious peptides derived fromthe C-terminal region of rab5(A) at a concentration of 1mM or in the presence of 100fjM recombinant rab5 purifiedfrom E. coli. In vitrotranslated proteins werelabelled with either 35S-methionine (B) or R-[14C]mevalonic acid lactone(C). The labelled proteinswere immunoprecipitated andanalysed after SDS-PAGE andfluorography.

Isoprenylation of rab proteins 863
the deleted region in the unmodified rab5A201-211mutant. In contrast, peptide 182-215, corresponding tothe last 34 amino acids of rab5, caused a 53% inhibitionof [I4C]mevalonic acid incorporation at 1 mM concen-tration, while peptide 182-197 had no effect. Isopreny-lation was completely abolished when in vitro trans-lation of rab5 was carried out in the presence of 100 fjmrecombinant rab5 purified from Escherichia coli.Although we cannot rigorously exclude the possibilitythat the peptides are rapidly degraded in the reticulo-cyte lysates, these results suggest that the enzymeresponsible for isoprenylation of rab proteins is notaffected by short (4 amino acids) peptides. Only apeptide derived from the last C-terminal 34 amino acidof rab5 had a partial inhibitory effect at a concentration100-fold higher than the one used for peptides inhibitingthe farnesyltransferase. The most likely interpretationof these findings is that the enzyme which catalyses theattachment of an isoprene moiety to rab proteins isdistinct from the farnesyltransferase modifying theCAAX box of nuclear lamins and ras proteins (Reiss etal., 1990, 1991; Seabra et al., 1991).
Discussion
rab proteins are isoprenylated in vitro and in vivoYeast mating factors, ras proteins, G-protein y-subunitsand nuclear lamins are post-translationally modified byaddition of thioether-linked prenyl groups (reviewed byGlomset et al, 1990; Rine and Kim, 1990; Maltese,1990; Spiegel et al., 1991). The modification occurs onthe CAAX motif at the C-terminal end of the protein.For instance, in the case of ras proteins, first a farnesylmoiety is added to the Cys residue, then the last threeamino acids are removed and, finally, the resulting C-terminal isoprenylated Cys residue is carboxylmeth-ylated (Hancock et al., 1989, 1990). With the exceptionof rab8, none of the rab proteins identified so far has aconventional CAAX box, although they contain twocysteines in their C termini. Instead, the C-terminalcysteine motifs of rab proteins are strikingly hetero-geneous in sequence (Chavrier et al., 1990b; Table 1).Like 5. cerevisiae Yptl, Ypt3 and Sec4, mammalianrabl, rab2, rab9 and rablO contain a cysteine doublet atthe C terminus (-CC). In contrast, mammalian rab3,rab4, rab6 and rab7 proteins contain a C-terminal CXCmotif, rab5 contains a CCXX motif, and rabll containsa CCXXX motif.
In this study we demonstrate that not only rab8(displaying a CVLL motifs), but also rab2, rab5, rab7and rabll proteins (having no C-terminal CAAX box),are isoprenylated both in vitro and in vivo. We alsoshow that these modifications occur on the C-terminalcysteine motifs, since deletion mutants lacking the C-terminal cysteines cannot be isoprenylated. Based onpeptide inhibition experiments, our results furthersuggest that the enzyme catalysing the modification ofthe rab proteins analysed in this study is distinct fromthe farnesyltransferase which has been found previously
to modify the CAAX motifs of p21ras (Reiss et al., 1990;Manne et al., 1990; Reiss et al., 1991; Seabra et al.,1991; Moores et al., 1991). rab8 is the only rab proteincontaining a CVLL C-terminal motif. The lack ofinhibition of rab8 isoprenylation by the CSIM peptide isconsistent with the finding that the CXXL C-terminalmotif undergoes a geranylgeranyl modification (Mal-tese and Robishaw, 1990; Mumby et al., 1990; Yamaneet al., 1990; Kawata et al., 1990; Reiss et al., 1991;Seabra et al., 1991; Yamane et al., 1991; Moores et al.,1991). Studies using purified farnesyltransferase re-vealed that tetrapeptides corresponding to the Ctermini of known farnesylated proteins efficientlyinhibited the enzyme, while tetrapeptides correspond-ing to the C terminus of geranylgeranyl-modified G-protein y-subunits (Maltese and Robishaw, 1990;Mumby et al., 1990; Yamane et al., 1990) did notcompete in the farnesyl-transfer assay (Reiss et al.,1991). Our results are in excellent agreement withrecent data from other laboratories. In particular,rabla, rablb, rab2, rab3a, rab5, rab6 and Yptl proteinshave been found to be modified by a geranylgeranylisoprene (Farnsworth et al., 1991; Khosravi-Far et al.,1991; Moores et al., 1991; Kinsella and Maltese, 1992).More specifically, evidence has been obtained for themodification of rab proteins by a geranylgeranyltransferase distinct from the one which modifiesproteins with a CXXL C-terminal motif (Moores et al.,1991; Horiuchi et al., 1991; Kinsella and Maltese, 1992).
Our data also indicate that, at least in the case ofrab5, the C-terminal cysteine motifs can be experimen-tally interchanged between different rab proteinswithout dramatic effects on subsequent isoprenylation.In particular, conversion of the CCXX motif of rab5 to-CC or to -CXC did not abolish isoprenylation. Incontrast, our data suggest that sequences upstream ofthe C-terminal cysteine motif are important for modifi-cation of rab proteins. Specifically, a deletion of 10amino acids immediately upstream of the C-terminalCCSN motif of rab5 (rab5A201-211) did preventisoprenylation (this study) as well as membrane associ-ation (Chavrier et al., 1991), indicating that a C-terminal cysteine motif is required but not sufficient forisoprenylation. One possible explanation is that thismutant protein folds in a conformation which preventsrecognition of the C-terminal motif by the isoprenoidtransferase. Alternatively, the deleted region mightcontain (or be part of) a recognition sequence for thetransferase. The latter interpretation is in agreementwith the finding that isoprenylation of rab5 is notabolished in the presence of a peptide corresponding tothe cysteine motif of rab5 alone but is, at least partially,inhibited by a peptide corresponding to the C-terminal34 amino acids of rab5. This partial inhibition effectmight be explained by assuming either that only a smallproportion of the peptide has the proper conformationto be recognized and modified by the isoprenoydtransferase or that other sequences upstream from thelast C-terminal 34 amino acid residues of rab5 areinvolved in this recognition event. Further work will berequired to distinguish between these possibilities.

864 M. Peter and others
Isoprenylatlon of rab proteins is required formembrane associationrab proteins were found associated with membranes ofdifferent compartments along the endocytic or se-cretory pathway, but the mechanisms mediating thismembrane association remain to be determined. In thecase of the nuclear lamins and pll™* proteins, theimportance of the CAAX box modifications for mem-brane association has been well documented (Krohne etal., 1989; Holtz et al., 1989; Beck et al, 1990; Kitten andNigg, 1991; Gutierrez et al., 1989; Hancock et al., 1989,1990). Specifically, expression of mutated forms ofchicken lamin B2, which are neither isoprenylated norcarboxylmethylated, accumulate in the nucleus, but areno longer capable of interacting with the inner nuclearmembrane (Kitten and Nigg, 1991). Similarly, we foundthat the deletion mutants rab2A or rab5A lacking the C-terminal motifs (see above) do not interact with theirrespective target membranes (Chavrier et al., 1990a)and that rab5A is functionally inactive (Gorvel et al.,1991). The results presented in this paper indicate thatthese mutant proteins are not isoprenylated, suggestingthat membrane association of rab proteins depends onthis post-translational modification. The isoprenoidgroup might serve to anchor the protein in the lipidbilayer and/or mediate the interaction with membraneand cytosolic proteins (Maltese and Robishaw, 1990;Mumby et al., 1990; Yamane et al., 1990; Araki et al.,1991). Moreover, carboxylmethylation of the isopreny-lated C-terminal cysteine residues was shown toincrease binding of p21ras to microsomal membranes invitro (Hancock et al., 1991). Interestingly, the rabprotein smgp25A/rab3a was recently shown to becarboxylmethylated (Farnsworth et al., 1991). Thisobservation, in analogy to ras proteins, raises thepossibility that carboxylmethylation contributes tomembrane association of rab proteins.
In the case of ras proteins, additional palmitoylationor the presence of a stretch of basic residues were foundto be necessary for plasma membrane localization(Hancock et al., 1989, 1990). By determining thesubcellular localization of several hybrid proteinsbetween rab2, rab5 and rab7, we have recently shownthat the hypervariable C-terminal domain determinesthe specific intracellular location of rab proteins(Chavrier et al., 1991). The C-terminal cysteine motifs,although required, have been shown to play no role inthe specificity of this process. From these results itappears that membrane localization of rab proteins is amulti-step process involving common post-translationalmodifications (isoprenylation, carboxylmethylation,proteolytic processing), as well as specific sequencerecognition by poorly characterized targetting mechan-isms.
We thank J. Gruenberg for critical reading of themanuscript, Dominique Nalis for peptide synthesis and M.Stapleton for help in some experiments. P.C was supported byan EMBL fellowship.
References
Anderegg, R. J., Beta, R., Carr, S. A., Crabb, J. W. and Duntze, W.(1988). Structure of Saccharomyces cerevisiae mating hormone a-factor. / . Biol. Chem. 263, 18,236-18,240.
Araki, S., Kaibuchi, K., Sasaki, T., Hata, Y. and Takai, Y. (1991).Role of the C-terminal region of smg p25A in its interaction withmembranes and the GDP/GTP exchange protein. Mol. Cell. Biol.11, 1438-1447.
Baker, D., Wuestehube, L., Schekman, R., Botstein, D. and Segev, N.(1990). GTP-binding protein Yptl and Ca2+ functionindependently in a cell free protein transport reaction. Proc. Nat.Acad. Sci. USA 87, 355-359.
Beck, L. A., Hosick, T. J. and Sinensky, M. (1990). Isoprenylation isrequired for the processing of the lamin A precursor. / . Cell Biol.110, 1489-1499.
Bourne, H. R., Sanders, D. A. and McCormick, F. (1990). TheGTPase superfamily: conserved structure and molecularmechanism. Nature 348, 125-132.
Chavrier, P., Gorvel, J.-P., Stelzer, H., Simons, K., Gruenberg, J.and Zerial, M. (1991). Hypervariable C-terminal domain of rabproteins acts as a targeting signal. Nature 353, 769-772.
Chavrier, P., Parton, R. G., Hauri, H. P., Simons, K. and Zerial, M.(1990a). Localization of low molecular weight GTP-bindingproteins to exocytic and endocytic compartments. Cell 62, 317-329.
Chavrier, P., Vingron, M., Sander, C , Simons, K. and Zerial, M.(1990b). Molecular cloning of YPTl/SEC4-related cDNAs from anepithelial cell line. Mol. Cell. Biol. 10, 6578-6585.
Darchen, F., Zahraoui, A., Hammel, F., Montells, M.-P., Tavitian,A. and Scherman, D. (1990). Association of the GTP-bindingprotein Rab3A with bovine adrenal chromaffin granules. Proc.Nat. Acad. Sci. USA 87, 5692-5696.
Farnsworth, C. C , Kawata, M., Yoshida, Y., Takai, Y., Gelb, M. H.and Gtomset, J. A. (1991). C terminus of the small GTP-bindingprotein smg p25a contains two geranylgeranylated cysteine residuesand a methyl ester. Proc. Nat. Acad. Sci. USA 88, 6196-6200.
Fischer von Mollard, G., Mignery, G. A., Baumert, M., Perin, M. S.,Hanson, T. J., Burger, P. M., Jahn, R. and SUdhof, T. C. (1990).Rab3 is a small GTP-binding protein exclusively localized tosynaptic vesicles. Proc. Nat. Acad. Sci. USA 87, 1988-1992.
Fuerst, T. T., Niles, E. G., Studier, F. W. and Moss, B. (1986).Eukaryotic transient-expression system based on recombinantvaccinia virus that synthesizes bacteriophage T7 RNA polymerase.Proc. Nat. Acad. Sci. USA 83, 8122-8126.
Gallwitz, D., Donatfa, C. and Sander, C. (1983). A yeast geneencoding a protein homologous to the human c-has/bas proto-oncogene product. Nature 306, 704-707.
Gtomset, J. A., Gelb, M. H. and Farnsworth, C. C. (1990). Prenylproteins in eukaryotic cells: a new type of membrane anchor.Trends Biochem Sci. 15, 139-142.
Gorvel, J. P., Chavrier, P., Zerial, M. and Gruenberg, J. (1991). rab5controls early endosome fusion in vitro. Cell 64, 915-925.
Goud, B., Salmlnen, A., Walworth, N. C. and Novkk, P. J. (1988). AGTP-binding protein required for secretion rapidly associates withsecretory vesicles and the plasma membrane in yeast. Cell 53, 753-768.
Goud, B., Zahraoui, A., Tavitian, A. and Saraste, J. (1990). SmallGTP binding protein associated with Golgi cisternae. Nature 345,553-556.
Gutierrez, L., Magee, A. I., Marshall, C. J. and Hancock, J. F.(1989). Post-translational processing of p21ras is two-step andinvolves carboxy-methylation and carboxy-terminal proteolysis.EMBO J. 8, 1093-1098.
Hancock, J. F., Cadwallader, K. and Marshall, C. J. (1991).Methylation and proteolysis are essential for efficient membranebinding of prenylated p21K-n u ( B ) . EMBO J. 10, 641-646.
Hancock, J. F. , Magee, A. I., Childs, J. E. and Marshall, C. J. (1989).All ras proteins are polyisoprenylated but only some arepalmitoylated. Cell 57, 1167-1177.
Hancock, J. F., Paterson, H. and Marshall, C. J. (1990). A polybasicdomain or palmitoylation is required in addition to the CAAXmotif to localize p21ras to the plasma membrane. Cell 63,133-139.
Haubruck, H., Prange, R., Vorgias, C. and Gallwitz, D. (1989). The

Isoprenylation of rab proteins 865
ras-related mouse yptl protein can functionally replace the YPT1gene product in yeast. EMBO J. 8, 1427-1432.
Holtz, D., Tanaka, R. A., Hartwig, J. and McKeon, F. (1989). TheCaaX motif of lamin A functions in conjunction with the nuclearlocalization signal to target assembly to the nuclear envelope. Cell59, 969-977.
Horiuchi, H., Kawata, M., Katayama, M., Yoshida, Y., Musha, T.,Ando, S. and Takai, Y. (1991). A novel prenyltransferase for asmall GTP-binding protein having a C-terminal Cys-Ala-Cysstructure. J. Biol. Chem. 266, 16981-16984.
lshibashl, Y., Sakagami, Y., Isogal, A. and Suzuki, A. (1984).Structures of Tremerogens A-9291-I and A-9291-VIII: peptidyl sexhormones of Tremella brasiliensis. Biochemistry 23, 1399-1404.
Kawata, M., Farnsworth, C. C , Yoshida, Y., Gelb, M. H., Glomset,J. A. and Takai, Y. (1990). Posttranslationally processed structureof the human platelet protein smg p21B: evidence forgeranylgeranylation and carboxyl methylation of the C-terminalcysteine. Proc. Nat. Acad. Sci. USA 87, 8960-8964.
Khosravi-Far, R., Lutz, R. J., Cox, A. D., Conroy, L., Bourne, J. R.,Sinenski, M., Balch, W. E., Buss, J. E. and Der, C. J. (1991).Isoprenoid modification of rab proteins terminating in CC or CXCmotifs. Proc. Nat. Acad. Sci. USA 88, 6264-6268.
Kinsella, B. T. and Maltese, W. A. (1992). Rab GTP-binding proteinswith three different carboxyl-terminal cysteine motifs are modifiedin vivo by 20-carbon isoprenoids. J. Biol. Chem. 267, 3940-3945.
Kitten, G. T. and Nigg, E. A. (1991). The CaaX motif is required forisoprenylation, carboxyl methylation and nuclear membraneassociation of lamin B2. J. Cell Biol. 113, 13-23.
Krohne, G., Waizenegger, I. and Hoger, T. H. (1989). The conservedcarboxy-terminal cysteine of nuclear lamins is essential for laminassociation with the nuclear envelope. J. Cell Biol. 109, 2003-2011.
Lehner, C. V., Kurer, V., Eppenberger, H. M. and Nlgg, E. A.(1986). The nuclear lamin protein family in higher vertebrates:identification of quantitatively minor lamin proteins by monoclonalantibodies. J. Biol. Chem. 261, 13,293-13,301.
Maltese, W. A. (1990). Posttranslational modification of proteins byisoprenoids in mammalian cells. FASEB J. 4, 3319-3328.
Maltese, W. A. and Robishaw, J. D. (1990). Isoprenylation of C-terminal cysteine in a G-protein y subunit. J. Biol. Chem. 265,18,071-18,074.
Manne, V., Roberts, D., Tobin, A., O'Rourke, E., De Vigilio, M.,Meyers, C , Ahmed, N., Kurz, B., Resh, M., Kung, H.-F. andBar back), M. (1990). Identification and preliminarycharacterization of protein-cysteine farnesyltransferase. Proc. Nat.Acad. Sci. USA 87, 7541-7545.
Mizoguchi, A., Kim, S., Ueda, T., Kikuchi, A., Yorifuji, H.,Hirokawa, N. and Takai, Y. (1990). Localization and subcellulardistribution of smg p25A, a ras p21-like GTP-binding protein, in ratbrain. J. Biol. Chem. 265, 11,872-11,879.
Molenaar, C. M. T., Prange, R. and Gallwitz, D. (1988). A carboxyl-terminal cysteine residue is required for palmitic acid binding andbiological activity of the ras-related yeast YPT1 protein. EMBO J.7, 971-976.
Moores, S. L., Schaber, M. D., Mosser, S. D., Rands, E., O'Hara, M.B., Garsky, V. M., MarshaU, M. S., PompUano, D. L. and Gibbs, J.B. (1991). Sequence dependence of protein isoprenylation. J. Biol.Chem. 266, 14,603-14,610.
Mumby, S. M., Casey, P. J., Oilman, A. G., Gutowski, S. and
Stemweis, P. C. (1990). Protein y subunits contain a 20-carbonisoprenoid. Proc. Nat. Acad. Sci. USA 87, 5873-5877.
Peter, M., Kitten, G. T., Lehner, C. F., Vorburger, K., Bailer, S. M.,Maridor, G. and Nigg, E. A. (1989). Cloning and sequencing ofcDNA clones encoding chicken lamins a and Bl and comparison ofthe primary structures of vertebrate A- and B-type lamins. J. Mol.Biol. 208, 393-404.
Reiss, Y., Goldstein, J., Seabra, M. C , Casey, P. J. and Brown, M. S.(1990). Inhibition of purified p21ras farnesyhprotein transferase byCys-AAX tetrapetides. Cell 62, 81-88.
Reiss, Y., Stradley, S. J., Gierasch, L. M., Brown, M. S. andGoldstein, J. L. (1991). Sequence requirement for peptiderecognition by rat brain p21ras protein farnesyltransferase. Proc.Nat. Acad. Sci. USA 88, 732-736.
Rine, J. and Kim, S.-H. (1990). A role for isoprenoid lipids in thelocalization and function of an oncoprotein. New Biol. 2, 219-226.
Salminen, A. and Novkk, P. J. (1987). A r&j-like protein is requiredfor a post-Golgi event in yeast scretion. Cell 49, 527-538.
Seabra, M. C , Reiss, Y., Casey, P. J., Brown, M. S. andGoldstein, J. L. (1991). Protein Farnesyltransferase andgeranylgeranyltransferase share a common or subunit. Cell 65, 429-434.
Segev, N., Mulholland, J. and Botstein, D. (1988). The yeast GTP-binding YPT1 protein and a mammalian counterpart are associatedwith the secretion machinery. Cell 52, 915-924.
Spiegel, A. M., Backlund, P. S. Jr, Butrynski, J. E., Jones, T. L. Z.and Simonds, W. F. (1991). The G protein connection: molecularbasis of membrane association. Trends Biochem. Sci. 16, 338-341.
Valencia, A., Chardin, P., Wittinghofer, A. and Sander, C. (1991).The ras protein family: evolutionary tree and role of conservedamino acids. Biochemistry 30, 4637-4648.
Van der Shnjs, P., Hull, M., Zahraoui, A., Tavitian, A., Goud, B. andMellman, I. (1991). The small GTP-binding protein rab4 isassociated with early endosomes. Proc. Nat. Acad. Sci. USA 88,6313-6317.
Vorburger, K., Kitten, G. T. and Nigg, E. A. (1989). Modification ofnuclear lamin proteins by a mevalonic acid derivative occurs inreticulocyte lysates and requires the cysteine residue of the C-terminal CXXM motif. EMBO J. 8, 4007-4013.
Walworth, N. C , Goud, B., Kastan KabceneU, A. and Novick, P. J.(1989). Mutational analysis of SEC4 suggests a cyclical mechanismfor the regulation of vesicular traffic. EMBO J. 8, 1685-1693.
Wolda, S. L. and Glomset, J. A. (1988). Evidence for modification oflamin B by a product of mevalonic acid. J. Biol. Chem. 263, 5997-6000.
Yamane, H. K., Farnsworth, C. C , Xie, H., Evans, T., Howald, W.N., Fung, B. K.-K., Clarke, S., Gelb, M. H. and Glomset, J. A.(1990). Brain G protein y subunits contain an a\\-trans-geranylgeranyl-cysteine methyl ester at their carboxyl termini.Proc. Nat. Acad. Sci. USA 87, 5868-5872.
Yamane, H. K., Farnsworth, C. C , Xle, H., Evans, T., Howald, W.N., Gelb, M. H., Glomset, J. A., Clarke, S. and Fung, B. K.-K.(1991). Membrane-binding domain of the small G protein G25Kcontains an S-(all-fra/is-geranylgeranyl)cysteine methyl ester at itscarboxyl terminus. Proc. Nat. Acad. Sci. USA 88, 286-290.
(Received 26 November 1991 - Accepted, in revised form,8 May 1992)






























