Embed Size (px)
Citation preview

ISOLATION OF O-ACETYLSERINE THIOL LYASE GENOMIC DNA SEQUENCE FROM LEUCAENA LEUCOCEPHALA VIA PCR
Abirame AlP Segaran (33657)
QL Bachelor of Science with Honours
(Resource Biotechnology)L8 2015A149
2015
561

ACKNOWLEDGEMENTS
With completion of this thesis, there are many who deserve a word of appreciation. First, I
would like to express my sincere appreciation and deepest gratitude to my supervisor, Dr.
Ho Wei Seng, Faculty of Resource Science and Technology, UNIMAS, who had provided
me the opportunity to perform and complete my final year project research in Forest
Genomics and informatics Laboratory. Thanks a million for his countless advice, patience
and guidance, definitely deserves more recognition than a word of merit.
Extended assistance from the postgraduates in Forest Genomics and informatics
Laboratory is much treasured too. My truthful gratitude goes to especially, Miss Natalie
anak Gali for her guidance, advices and patience. She had dispensed wisdom on assortment
of basic molecular techniques which helped me a lot in performing the research. She spent
many hours ofmeticulous care in helping me to complete the research besides giving me
the very important knowledge on how to use th~ software and analyse my result data.
Next, I WQuld like to thank all my friends who had been with me with their constant
support throughout the progression of the thesis since day one. Last but not least, I would
like to express my gratitude to my lovely family members who always supporting me,
accommodating and understanding in every way.
Thank you all who ha&made this thesis a success.

UNIVERSITI MALAYSIA SARAWAK
Grade: __________
Please tick (-.1) Final Year Project Report [;2] Masters D PhD D
DECLARATION OF ORIGINAL WORK
This declaration is made on the ...?:.~ ........day of.J.~~~~~Y ..201~
Student's Declaration:
I _~1?~B~~f __~_lf __~~~_~_~E_~_~_~_~~~_:f:_,.J_~_~~hT_~ __~[__~~~~~_~~~ ___~_~~~~_~_~ _~_~_I~_(~~_~_~ct-~. ------- (PLEASE INDICATE STUDENT'S NAME, MATRIC NO. AND FACULTY) hereby ~ declare th;:t,t the work entitled ;I;~~l~:E~_':LM'__Q:-_~~~~~_iJ!~'--~~L~~_LU~J_~__~~tl1~~~~______~_ L~c~liIo, 'lU{Ot£.;;'~11i \lltI -PCp... . , .-------1". IS my ongmal work. I have not copIed from any other students work or from any other sources except where due reference or acknowledgement is made explicitly in the text, nor has any part been written for me by another person.
Date submitted Name of the student (Matric No.)
Supervisor's Declaration:
1-------------------------------------------------------------- (SUPERVISOR'S NAME) hereby certifies that.. the work entitled ---------------------------------------------------------.--- ---------------- (TITLE) was prepared by the above named student, and was submitted to the "FACULTY" as a * partial/full fulfillment for the conferment of --------------------------------------------------------------- ---------. (PLEASE INDICATE THE DEGREE), and the aforementioned work, to the best of my knowledge, is the said student's work.
Received for examination by: Date:_________
(Name of the supervisor
ii

I
I declare that Project/Thesis is classified as (Please tick C-J»:
D CONFIDENTIAL (Contains confidential information under the Official Secret Act 1972)* DRESTRICTED (Contains restricted information as specified by the organisation where
research was done)* DOPEN ACCESS
Validation of ProjectIThesis
I therefore duly affirm with free consent and willingly declare that this said Project/Thesis shall be placed officially in the Centre for Academic Information Services with the abiding interest and rights as follows:
• This Project/Thesis is the sole legal property of Universiti Malaysia Sarawak (UNIMAS).
• The Centre for Academic Information Services has the lawful right to make copies for the purpose of academic and research only and not for other purpose.
• The Centre for Academic Information Services has the lawful right to digitalise the content for the Local Content Database.
• The Centre for Academic Information Services has the lawful right to make copies of the Project/Thesis for academic exchange between Higher Learning Institute.
• No dispute or any claim shall arise from the student itself neither third party on this Project/Thesis once it becomes the sole property of UNIMAS.
• This ProjectlThesis or any material, data and information related to it shall not be distributed, published or disclosed to any party by the student except with UNIMAS permission.
cAr: Student signature _2_3_1_,_!_?l)_r_~___ Supervisor signature: _______
(Date) (Date)
Current Address: Lc.. £50, ;]~N PI4Yf}-(Y)f)$ TArt1AN S't}JTOSIl,
Notes: * If the ProjectlThesis is CONFIDENTIAL or RESTRICTED, please attach together as annexure a letter from the organisation with the period and reasons of confidentiality and restriction.
[The instrument is duly prepared by The Centre for Academic Information Services]
iii
I
I

Pusat Khic1 mat l 1aklumat Akadcl. " UNlVERSfTI MALAY fA SAW WA"
TABLE OF CONTENTS
Acknowledgement
Declaration
Table of Contents
List of Abbreviations
List of Figures
List of Tables
Abstract
Introduction
Literature Review
2.1 Selection of Species Studied
2.1.1 Family Fabaceae
2.1.2 Genus
2.1.3 Species - Leucaena Leucocephala
2.2 O-acetylserine (thiol) lyase
2.3 Mimosine
Materials and Methods
3.1 DNA Extraction and Purification
3.1.1 Plant Materials
3.1.2 Chemicals and Reagents
" 3.1.3 DNA Extraction and Purification Prot@col
3.2 Primer Design
3.3 PCR Amplification
3.5 DNA Sequencing and Data Analysis
Results and Discussion
4.1 DNA Isolation and Purification
4.2 Polymerase Chain Reaction (PCR)
4.3 DNA Sequencing and Data Analysis
iv
I
II
IV
VI
VII
VIII
1
2
5
5
5
6
8
9
12
12
12
12
14
14
15
16
18
20

Conclusion and Recommendation 23
References 24
v

LIST OF ABBREVIATIONS
BLAST Basic Local Alignment Search Tool
CIA Chlorofonn-Isoamyl Alcohol
CTAB Cetyltrimethylammonium Bromide
DHP 3 -h ydrox y-4( 1 H)-pyridone
DNA Deoxyribonucleic Acid
EB Extraction buffer
EDTA Ethylenediamine tetraacetic acid
HP . 3-hydroxy-4-pyridone
NCBI National Centre for Biotechnology Infonnation
OAS O-acetyl-L-serine
OAS-TL O-acetylserine (thiol) lyase
PCR Polymerase Chain Reaction
PVP Polyvinylpyrrolidone
SAT Serine acetyl transferase
vi

LIST OF FIGURES
2.1 Map showing the distribution of Leucaena, about 2393 herbarium
specimen collection sites in South Mexico with verifiable localities. 6
2.2 Biosynthetic pathways of mimosine relative to the formation of cysteine.
SA T catalyses the activation of serine to the intermediate molecule, OAS in
the cysteine synthase complex. Formation ofmimosine is catalysed by
homodimers of OAS-TL in the presence of the appropriate substrates. 9
2.3 (a) Chemical structure of mimosine. 11
2.3 (b) Transformation of mimosine into dihydroxypyridine by the rumen
of microorganisms. 11
4.1 (a) Electrophoresis of DNA sample of un purified genomic DNA. 16
4.1 (b) Electrophoresis of DNA sample of purified genomic DNA. 16
4.2 Agarose gel electrophoresis of PCR protiucts of OASTL amplified using
different T a °C. 20
4.3 Graphic display of BLAST output for similarity searching ofOASTL of
Leucaena leucocephala. 21
vii

LIST OF TABLES
Table 3.3 (a) Primer (OASTL) 14
Table 3.3 (b) PCR reaction mixture 15
Table 3.3 (c) PCR reaction cycles, temperatures and durations 15
Table 4.3 BLAST search result for amplified 1667 bp of OASTL DNA
fragment. 21
viii

Isolation of O-acetylserine (thiol) lyase (OAS-TL) Genomic DNA Sequence from Leucaena leucocephala via PCR
Abirame AlP Segaran
Resource Biotechnology Prograllll)1e Faculty of Resource Science and Technology
University Malaysia Sarawak
ABSTRACT
Leucaena leucocephala is widely used as a fodder for livestock animals due to its high nutritious value compared to other tropical plants. However, quality of the plant product as a livestock fodder became questionable due to the toxic effect by its degradation product, mimosine (b-N-[3-hydroxy-4-pyridone]-aaminopropionic acid) to herbivores upon ingestion. Studies suggest that inactivation or reduction of these substances will improve the quality ofL. leucocephala in animal feed production. Most of the studies have been done yet focus on reducing these substances by the use of chemicals such as FeCl3 and HCI or by the degradation enzymes extracted from ruminant microorganisms. On the other hand, the enzyme 0acetylserine thiollyase which catalyzes the synthesis of mimosine is crucial in studies to reduce mimosine content. In this study, the leaf ofLeucaena leucocephala was used to extract the total genomic DNA. Forward and reverse primers were designed based on the O-acetylserine thiollyase mRNA sequences obtained from the NCBI database. Polymerase Chain Reaction (PCR) was performed based on the standard PCR method to amplify O-acetylserine thiollyase DNA fragment. The DNA fragment of about 1.7 kb was sent for sequencing.
Key words: O-acetylserine thiollyase, OASTL gene, Leucaena leucocephala, PCR
ABSTRAK
Leucaena leucocephala digunakan secara meluas sebagai makanan bagi haiwan ternakan kerana nilai berkhasiat yang tinggi berbanding dengan tumbuhan tropika lain. Walau bagaimanapun, kualiti produk tumbuhan sebagai makanan ternakan dipersoalkan kerana kesan toksik daripada produk degradasi. mimosine (b-N-{3-hydroxy-4-pyridoneJ-a-aminopropionic asid) kepada haiwan herbivor. Kajian menunjukkan bahawa inaktivasi atau pengurangan bahan-bahan ini akan meningkatkan kualiti L. leucocephala dalam pengeluaran makanan haiwan. Kebanyakan kajian yang dijalankan hanya memberi tumpuan kepada mengurangkan bahan-bahan ini dengan menggunakan bahan kimia seperti FeCl3 dan HCI atau dengan enzim degradasi yang diekstrak daripada mikroorganisma ruminan. Sebaliknya. enzim 0acetylserine thiollyase·Yang memangkinkan sintesis mimosine adalah penting dalam kajian untuk mengurangkan kandungan mimosine. Dalam kajian ini. daun Petai belalang telah digunakan untuk mengekstrak DNA genomik. Primers telah direka berdasarkan urutan mRNA O-acetylserine thiollyase yang diperolehi dari NCB!. Polymerase Chain Reaction (PCR) dilakukan berdasarkan kaedah PCR standard untuk mengamplifikasi O-acetylserine thiollyase DNA/ragment. Fragmentasi DNA bersaiz kira-kira 1. 7 kb telah dihantar untuk sequencing.
Kata kunci: O-acetylserine thiollyase, OASTL gene, Leucaena leucocephala, PCR
1

INTRODUCTION
Leucaena leucocephala which is locally known as petai belalang is a perennial non-
climbing, non-spiny shrub or tree. It is originated from tropical America; two of the three
subspecies now have a pan-tropical distribution facilitated by its use as a fodder, wood
source and reclamation species. Described as the 'alfalfa of the tropics', it is considered a
versatile and widely used tree legume in the tropics for various purposes. Moreover, it is
considered as a weed in more than 25 countries around the globe (Walton, 2003).
Wood products manufacturers in Malaysia are now declining the good quality
timber supply due to deforestation especially rubber woods considering it's extended uses
as a raw material to many wood based industries. Therefore, wood industry sector in
Malaysia is now looking forward to other fast growing species to sustain the demand for a
new raw material. As one of the fast growing tropical species Petai belalang (Leucaena
leucocephala) has the potential to be developed as a raw material. The Malaysian
Agricultural Research Development Institute (MARDI) has used this tree for multipurpose.
For example, it has been used for shade and wind protection in a variety of crops,
especially during earlygrowth and food for some animals (Rahman et al., 2012).
L. leucocephala (leucaena) is fast-growing, nitrogen fixing leguminous tree that is
widely distributed in tropical and subtropical countries. Leucaena can grow under different
soil and environmental conditions, such as alkaline and arid regions, and it is highly
resistant to insects and diseases. In many countries, leucaena is used as fuelwood, and for I
2

producing pulp and charcoal. Leucaena foliage is especially useful as a fodder for livestock
animals, since it has high nutritional value, comparable to alfalfa. Moreover, the protein
content of leucaena (15-18%) is much higher compared to tropical grasses and cereal
straws (3-10%) making it a valuable protein supplement to low quality forage diets. The
foliage of leucaena is highly digestible and palatable, and a rich source of macro and
micro-elements. In spite of its high nutritious value, leucaena has an undesirable feature; it
produces the toxic free ammo acid mimosine (b-N-[3-hydroxy-4-pyridone]-a
aminopropionic acid). L-mimosine, fi-(3-hydroxy-4-pyridon-1-yl)-L-alanine and its
degradation product 3-hydroxy-4-pyridone (HP) are thyrotoxic amino acid where they
have toxic effect that can cause loss of hair in growing animals (Jkegami et al., 1990).
Therefore, leucaena can be fed to animals only in limited amounts. Mimosine is produced
by all species in the genera Leucaena and Mimosa. In the young leaves, pods, seeds, shoots
and stems, which are the parts of the plant used in animal feed, mimosine concentration
can be as high as 5% on a dry weight basis (JUDe & Borthakur, 2010).
It is important to study the gene that transcribes the synthesis of mimosine to
produce mimosine free in order to improve the quality of livestock feed products.
Unfortunately, not many studies have been carried out at present to reduce the mimosine
content in leucaena species. So far, most of the work on mimosine that has been done is
restricted to the reduction of this compound from Leucaena by the use of chemicals such
as FeC13 and Hel or by the degradation enzymes extracted from ruminant microorganisms
(Xuan et al., 2006).
3

The objective of this study was to isolate and characterise the O-acetylserine (thiol)
lyase genomic DNA sequence from Leucaena leucocephala using peR method. The .0
acetyl serine (thiol) lyase catalyses the synthesis of mimosine in Leucaena leucocephala.
4

Pusat Khirlmat Maklumat Ak d " , "~ " a eo! ' I. YsrA SARA\ '
LITERATURE REVIEW
2.1 Selection of Species Studied
2.1.1 Family Fabaceae
Under the order of Fabales, the Fabaceae or Leguminosae is the species-rich third largest
flowering plant family and extensively distributed worldwide. Fabaceae are also
commonly known with other names like Leguminosae, legume family, legumes, bean and
pea family. There are about 720 genera in the Fabaceae family and it is further divided into
three subfamilies: Caesalpinioideae, Mimosoideae, and Faboideae (Wojciechowski, 2003).
The three subfamilies are recognized based on particular characteristics such as its flower,
size, symmetry, aestivation of petals, sepals (united or free), stamen number and
heteromorphy, pollen (single or polyads), even the presence of a pleurogram, embryo
radicle shape, leaf complexity, and presence of root nodules. These distinguishable
characteristics led to the inspection that the Mimosoideae and Papilionoideae are unique
and distinct lineages in the family which arose independently within a paraphyletic "basal"
caesalpinioid assemblage. The Dimorphandra group of tribe Caesalpinieae and papilionoid
tribe Swartzieae were considered to be respectively transitional groups between them
(Herendeen & Dilcher, 1990).
2.1.2 Genus
Leucaena (Leguminosae - Mimosoideae) is a small genus of seventeen diploid and five
tetraploid species, six infraspecific taxa, and two named hybrids. They are available from
small to medium-sized trees that grow in seasonally dry tropical forests of Mexico, Central
America and northern South America. The economic importance of Leucaena species, and
5

- --- - - -- -- -------- - -
especially L. leucocephala for the production of livestock fodder, green manure, small
wood products and for soil conservation is well known and widely acknowledged.
Map shOVlfing the distribution of ~coeno. Data points ....e 2393
herbarium specimen collection sites vwtt:h veriftable localities.
FIGURE 2.1 Map showing the distribution of Leucaena, about 2393 herbarium specimen collection sites in South Mexico with verifiable localities (Retrieved from: http://herbaria. plants. ox.ac. uk/bol/ content! groups/leucaena/images/research. pdf
2.1.3 Species - Leucaena /eucocepha/a
L. leucocephala and L. glabrata are the two subspecies of L. leucocephala. The
distinguishable characteristics between these two subspecies are L. leucocephala IS a
relatively smaH and much-branched tree with younger stems covered by dense fine
greyish-coloured hairs while L. glabrata is a rather large and sparsely-branched tree with
hairless younger stems. L. leucocephala is native of Yucatan Peninsula and the Isthmus of
Tehuantepec in southern Mexico, and is widely distributed throughout the tropics. The
origin of the species from South Mexico and the northern part of Central America is
6

extended for the most part of Southeast Asia, except Laos after it has been introduced
throughout the tropics and has become grow wild in many places. For examgle, in the 16th
century, this species is introduced into the Philippines as a feed for ruminant livestock
before it is spread throughout Asia-Pacific area.
Its leaves structure found to be bipinnately compound with 10-20 pairs of leaflets
on each pinnae. It has white flowers in round heads which are 2-2.5 cm in diameter. The
"giant" forms of the species are distinguished from L. diversifolia by two 1-2 mm wide
round glands on the leaf stalk at the first and last pair of pinnae of most leaves.
Several types exist from shrubs to small or medium sized trees. "Giant" types may
reach 20 m in height. Bark is smooth, grey tq brown with small tan spots. Leaves alternate,
evenly bipinnate and 10-20 cm long with 4-10 cm long pinnae. Each pinnae has 10-20
pairs of oblong or lanceolate leaflets that are 8-15 mm long and 3 mm wide which folds up
in the night. Leaf base is sub-equal or oblique. The yellowish-white flowers are grouped in
round flower heads that 2- 2.5 cm in diameter. Pods are 10-20 em long and 1.5 to 2 cm
wide, flat and pointed in both ends which tum to brown colour when ripe.
L. leucocephala is a tropical lowland pioneer species which has ability of fast
growing, competitive and thrives in full sunlight and survives with little water. However, it
grows best with 1,000-3,000 mm evenly distributed annual rain, neutral to alkaline soils
and temperatures between 22-30°C. Although some provenances are adapted to higher
elevations, most forms thrive better below 500 m elevation. The prevalent infestation of
psyllids has been a main restriction in exploitation of this species (Devi, 2013).
7

There are various benefits from this speCIes which can be exploited such as
reforestation, erosion control and soil improvement. Its wood can be used for light
construction, poles, pit props, pulp, furniture, flooring and fuel wood while the other green
parts are used as fodder and green manure.
2.2 O-acetylserine (thiol) lyase
O-acetylserine (thiol) lyase (OAS-TL) is an enzyme from the family of ~-replacement
enzymes that require pyridoxal phosphate (PLP) as its cofactor. Multiple copies of OAS
TL found in plants which are located in the cytosol, chloroplast, and mitochondria.
Mimosine formation is a two-step process. The two steps of biosynthesis of mimosine are
identical to the pathway of the cysteine synthesis. The first step in cysteine synthesis is the
formation of O-acetylserine (OAS) from acetyl-CoA and serine which is catalyzed by
serine acetyltransferase (SAT). Whereas OAS-TL involve in the second step of the
process where it conduct the catalytic activity of the formation of cysteine by replacing
acetate with sulphide in the intermediate molecule, ~AS. On the other hand, a heterocyclic
ring of 3-hydroxy-4-pyridone (3H4P) is used in place of sulphide for mimosine formation,
the formation ofa ~-substituted alanine (Yafuso et at., 2014).
Study about the formation of L-mimosine from 3,4-dihydroxypyridine (DHP) and
O-acetyl-L-serine (OAS) has been conducted by Murakoshi using cell-free extracts of
Leucaena seedlings (Murakoshi et at., 1984). According to Murakoshi (1984), mimosine is
synthesized by a reaction mechanism comparable to the biosynthesis of cysteine in that
both use O-acetyl-L-serine as the donor of the alanyl group. In fact, it was shown in L.
8

leucocephala the enzymatic activities catalyzjng the synthesis of mimosine and cysteine
reside in a single enzyme, O-acetylserine thiollyase, known as cysteine synthase.
.. SAT+
Coli'0)-<. A 0
Serine AcetylCoA
Mimosine
Figure 2.2 shows biosynthetic pathways of mimosine relative to the formation of cysteine. SAT catalyses the activation of serine to the intermediate molecule, OAS in the cysteine synthase complex. Formation of mimosine is catalysed by homodimers
ofOAS-TL in the presence of the appropriate substrates (Yafuso et al. , 2014).
Based on the study conducted by Wirtz (2004), the genes encoding isoforms of
OAS-TL proteins are expressed more or less ubiquitously in all plant organ cell types
analysed so far with little variation of contents of RNA, protein, and extractable enzyme
activity in response to external factors. OAS-TL activity has not only been found In
plastids as the site of sulphate reduction, but also in the cytosol and mitochondria.
2.3 Mimosine
Mimosine is a major chemical compound present in all plant parts of Mimosaceae, which
includes Leucaena (Leucaena leucocephala) while the seeds and foliage of leguminous
trees and shrubs of the genera Leucaena and Mimosa contribute the largest quantity of
mimosine. Mimosine [B-N-(3-hydroxy-4-pyridone)-a-aminopropionic acid] is a non
9

protein ammo acid and is structurally comparable with dihydroxyphenylalanine as it
contain a 3-hydroxy-4-pyridone ring instead of a 3,4-dihydroxyphenyl ring.
The quantity of mimosine in the young leaves and mature seeds was the greatest,
2.66 and 2.38% of dry weight, respectively, while the quantity in the root xylems and
xylems was the lowest: 0.18 and 0.11 % of dry weight, respectively. Younger plant parts of
Leucaena contained greater amounts of mimosine than the mature parts. The seeds, stems,
pods, and leaf tissues of different Leucaena species have been shown to contain mimosine
in various amounts. Seeds contain 4 to 5% mimosine on a dry-weight basis. The mimosine
contents of different parts of the shoot vary from 1 to 12%; old stems contain the smallest
and growing tips contain the largest amounts. Leucaena root also contains 1 to 1.5%
mimosine.
Both mimosine and 3-hydroxy-4(fH)-pyridone (DHP), the degradation product of
mimosine are toxic upon ingestion by herbivores. Thus, their inactivation or reduction
would improve the quality of the plant product as livestock fodder (Chanchay, 2009). The
toxicity ofmimosine is attributable by the presence of -OH and -0 in the pyridine ring and
known to suppress iron-containing enzymes and compete with tyrosine. The characteristic
activity of growth inhibitory properties of mimosine is a hydroxyl group a to the oxo .' function of the pyridone ring. The location of the' amino acid side chain seems to be less
critical and an isomer. The synthesis of two mimosine isomers with the position of the a
hydroxy-oxo function in the pyridine ring of mimosine was at least as active in vitro and in
vivo as the natural amino acid. The constituent properties of the a-hydroxy-oxo group are
involved in the biological activity of mimosine and other systems and may play a key
factor in growth suppression. The structure of the heterocyclic ring in mimosine is possible
10
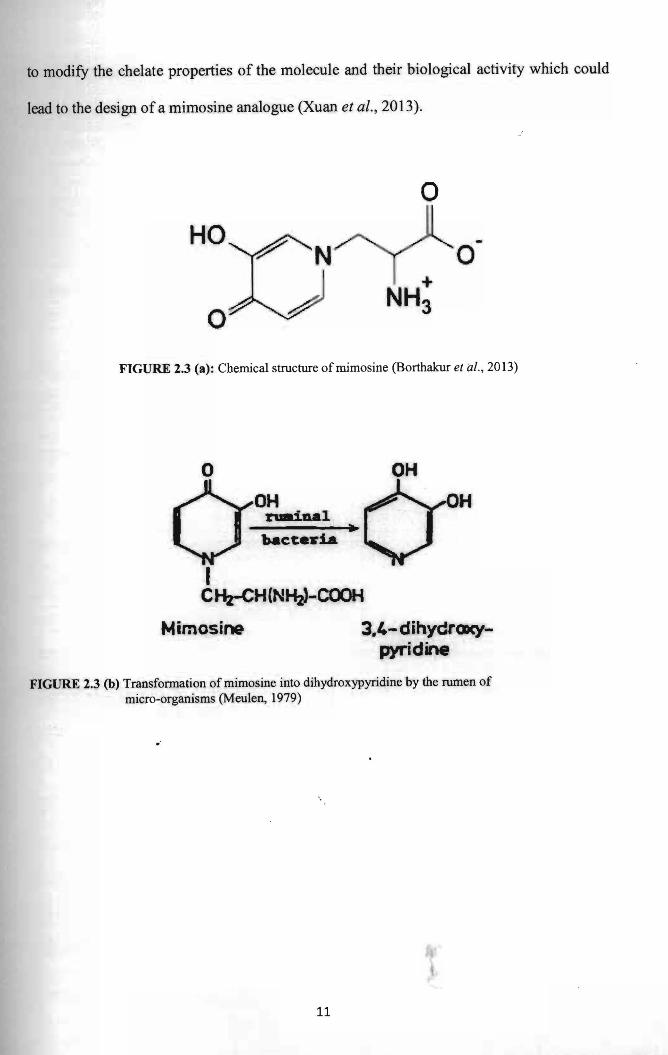
to modify the chelate properties of the molecule and their biological activity which could
lead to the design of a mimosine analogue (Xuan et al., 2013).
o HO
N o
o FIGURE 2.3 (a): Chemical structure ofmimosine (Borthakur et a/. , 2013)
o OH
OH nainal
H
I CHz-CH(NHV-COOH
Mimosine 3.4-dihydrCDCypyridine
FIGURE 2.3 (b) Transfonnation ofmimosine into dihydroxypyridine by the rumen of micro-organisms (Meulen, 1979)
11

MATERIALS AND METHODS
3.1 DNA Extraction and Purification
3.1.1 Plant Materials
Young leaves sample of Leucaena leucocephala were obtained at Forest Genomics &
Informatics Laboratory (/GiL) of Faculty ofResource Science and Technology, UNIMAS.
3.1.2 Chemicals and Reagents
List of solutions to be prepared: Liquid nitrogen, 1 M Tris HCl (pH8.0), 0.5 M EDT A
(pH8.0), CTAB extraction buffer (EB), CIA: Chloroform - isoamyl alcohol (24:1),
isopropanol, wash buffer and TE buffer (pH8.0).
3.1.3 DNA Extraction and Purification Protocol
A total of 120 III of 2% ~-mercaptoethanol was added into 6 ml of 2X CT AB buffer. The
mixture was preheated in a 60°C water bath. 1 g of leaf tissue was grinded in a pre-chilled
pestle and mortar into fine powder. Liquid nitrogen was added while grinding the leaf
tissue as it is necessary to avoid thaw. The plant powder was transferred to a 15/50 ml snap
cap Falcon tube containing 6 ml CT AB buffer and incubated in a water-bath for 2 hours at .'
60°C. The tube was inverted occasionally. The slurry was allowed to cool to room
temperature. An equal volume of chloroform-isoamyl alcohol (CIA) (24: 1) was added and
mixed gently for 15 minutes. The mixture was centrifuged at 4,000 rpm for 15 minutes at
room temperature. The aqueous phase was transferred to a clean 15/50 ml snap cap Falcon
tube and re-extracted with equal volume of CIA. The solution was mixed gently for 15
minutes, centrifuged for 15 minutes at 4,000 rpm. The aqueous phase was transferred again
12

to a clean snap cap 15/50 ml Falcon tube. 2/3 volume of cold isopropanol (-20°C) was
added, gently mixed, and kept at -20°C for overnight. The nucleic acid was collected by
centrifuge at 1,500 rpm for 15 minutes at room temperature. The supernatant was poured
off. 1 ml of wash buffer was added and washed for 1 hour. The pellet was transferred
together with wash buffer to a clean 1.5 ml microcentrifuge tube and centrifuged at 13,000
rpm for 10 minutes. The supernatant was poured off and the pellet was air dried.
DNA purification was conducted using Wizard® Genomic DNA Purification Kit
(Prom ega, USA). Total volume of DNA was to up to 500 ml. 3 III of RNAse was added
and mixed. The mixture was incubated at 37°C for 1 hour. The mixture was allowed to
cool down to room temperature before proceeding with next step. 20 J.1l of protein
precipitation solution was added and vortex vigorously at high speed for 20 seconds. The
solution was centrifuged at 13,000 rpm for 15 minutes following vortexed to obtain pellet.
The supernatant was transferred into new 1.5 ml of microcentrifuge tube. Equal volume of
cold isopropanol of -20°C was added into the tube. The solution was mixed by inverting
the tube and stored at -20 °C overnight. The mixture was centrifuged again the next day at
13,000 for 15 minutes to throwaway excess isopropanol. The supernatant was discarded
and 1 ml of 70% EtOH of -20°C was added. The solution was inverted several times for 1
hour. The solution·' was centrifuged at 13,000 rpm, for 15 minutes. The supernatant was
poured off and air dried the pellet at room temperature. 50 J.1l of ultrapure H20 was added
to resuspend the pellet. The DNA solution was mixed well and stored at -20°C. 5 III of the
DNA suspension was run on 0.8% agarose gel for DNA quantification.
13

3.2 Primer Design
The nucleotide sequences of O-acetylserine (thiol) lyase from closely related species were
determined from the Genebank database. The sequence alignment of O-acetylserine (thiol)
lyase with different species was done using CLC Sequence Viewer software for the
determination of conserved region. The parameters that were used for sequence alignment
was the default parameters set in that programme.
Primers were designed based on the conserved region by using the Primer 3.0
software. The parameters of the primer design such as target sequences, primer length,
melting temperature and GC content were adjusted. 20 base pairs of forward and reverse
primers were designed and sent for synthesis.
3.3 peR Amplification
From the database results obtained, PCR method was used to detect gene of O-acetylserine
(thiol) lyase with the designed primer. Amplification was carried out by using the forward
and reverse primers that were designed from cDNA sequence of OASTL (KF754356.l) in
cell cytosol from Leucaena leucocephala. The details of the primers were described in the
table below.
Table 3.3 (a): Primer (OASTL)
Primer OASTL Forward Sequence 5' TATGGAACCCCTCTCTAGTG 3' Reverse Sequence 5'CACCAATCCCATAAATCCCA3' Forward Tm 52.7°C Reverse Tm 52.4 °C Expected Product Size 569 bp
14
Later, the PCR product was examined on 1.5 % agarose gel and 100bp DNA ladder
(Promega, USA). The amplification of OASTL was performed in Mastercycler Gradient
Thennal Cycler (Eppendorf; Germany) and the profile was set as Table 3':3 (b) and (c)
shown below.
Table 3.3 (b): PCR reaction mixture
Reagent Stock concentration Final concentration Final volume ddH20 PCR buffer 10 x 1 x 2.5 III dNTP 2.0mM 0.2mM 2.0 III Forward primer 2.5 pmol/1l1 5 pmol 2.0 III Reverse primer 2.5 pmol/1l1 5 pmol 2.0 III Template DNA lOng/ill 20 ng/1l1 2.0 III Taq DNA polymerase 0.5 U/lll 1.0 UIIlI 2.0 III Total
Table 3.3 (c): PCR reaction cycles, temperatures and durations
Stage Temperature Duration Cycles Initial denaturation 95°C 2 min 1 Denaturation 94°C 45 s 35 Annealing 59°C 45 s 35 Extension 72°C 1 min 35 Final extension 72 °c 10 min 1
The PCR products were checked using electrophoresis in a 1.5% agarose gel. The
agarose gel was stained with EtBr and visualized under UV.
3.4 DNA Sequencing and Data Analysis
DNA fragment obtained from the amplification was sent for sequencing later. The
sequencing result was viewed using Chromas Lite software version 7.5. NCBI Blast was
used for comparison of nucleotide gene sequences in data analysis.
15




![Suppressed Methionine γ-Lyase Expression Causes … · Suppressed Methionine g-Lyase Expression Causes Hyperaccumulation of S-Methylmethionine in Soybean Seeds1[OPEN] Takuya Teshima,a](https://img.pdfslide.us/doc/110x75/5f752fe516889f57ef3206fe/suppressed-methionine-lyase-expression-causes-suppressed-methionine-g-lyase-expression.jpg)
























