Embed Size (px)
Citation preview

Isolation of morphologically intact mitochondrial nucleoids from the
yeast, Saccharomyces cerevisiae
ISAMU MIYAKAWA, NOBUNDO SANDO
Biological Institute, i'aculty of Science, Yionaguchi i'niveisily. Yamatiuchi 753, Japan
SIIIGEYUKI KAVVANO, SOIC1II NAKAMURA and TSUNEYOSIII KUROIWA
Department of Cell Biology. Salional Institute for Basic Biology, Okazaki, 444, Japan
Summary
Mitochondrial nucleoids (mt-nucleoids) of theyeast, Saccharomyces cerevisiae, were isolatedfrom spheroplasts of stationary phase cells andtheir structure and organization were investi-gated by fluorescence microscopy, electron mi-croscopy, and biochemical techniques. Isolatedmt-nucleoids were spherical or ovoid and0-3-0-6 ion in diameter, and were about thesame size and shape as those observed in the cellby the DAPI staining technique.
Measurement of DNA content of mt-nucleoids,using a video-intensified microscope system, afterDAPI staining revealed that a mt-nucleoid inspheroplasts from stationary phase cells contains,
on average, 3-9 mtDNA molecules and an isolatedmt-nucleoid contains, on average, 3-1.
Negatively stained electron micrographsshowed that mt-nucleoids consist of particles20-50 nm in diameter. SDS-polyacrylamide gelelectrophoresis of mt-nucleoids detected 20species of polypeptides in the molecular weightrange from lOxlO3 to 70xl03. Acid-urea/SDStwo-dimensional electrophoresis of acid extractfrom mt-nucleoids indicated that a polypeptide of20xl03 is the only major polypeptide with basicproperty like histones.
Key words: yeast, Scuclioroinvces cerevisiae,mitochondrial nucleoids, DAPI, isolation.
Introduction
During the past several years considerable efforts haveproduced great progress in the study of mitochondrialgenes and their products. The mitochondrial genomeof the yeast, Saccharoinvces cerevisiae, is at presentone of the best characterized organdie genomes incukaryotic cells.
It encodes the genetic information for essentialmitochondrial components: namely, several mitochon-drial proteins, mitochondrial rRNAs and tRNAs(Dujon, 1981). These mitochondrial genomes havebeen known to be localized at the electron-transparentarea named mitochondrial nucleoids (mt-nucleoids) inthe matrix from electron microscopic examinations.But there is still little valuable information on the stateof mitochondrial genomes /'// vivo.
Protein—DNA complex has been isolated in mito-chondria of the slime mold Phxsanun polxcephaluiu
Journal of Cell Science 88. 431-439 (1<J87)I'rintuil in Grunt Britain © The Company of Biologists Limited \')S7
(Kuroiwa el al. 1976; Suzuki el al. 1982; Kuroiwa,1982), I leLa cells (Albringe/ al. 1977), Xeiiopus Uwvis(Pinon et al. 1978; Rickwood & Jurd, 1978), sea-urchinembryos (Sevaljevic el al. 1978, 1979), rat liver (VanTuyle & McPherson, 1979; Van Tuyle & Pavco, 1985)and Paraineciuiu aureha (Olszewska & Fait, 1980).DNA—protein complex has been isolated from themitochondria of the yeast, .S\ cerevisiae (Rickwood elal. 1981; Rickwood & Chambers, 1981). However,with the exception of studies on isolated mt-nuclcoidsfrom P. polxcephaluiu, which revealed that isolated mt-nucleoids have the same shape, size and DNA contentas in vivo, the question remains ambiguous as towhether isolated DNA—protein complexes correspondto the mt-nucleoids observed in the electron micro-scope and whether they retain the morphologicalintactness.
We previously reported on fusing and separatingfigures of mitochondrial nucleoids during the life cycle
431

of yeast using the fluorescence microscope and DNA-binding fluorescent dye, 4',6-diamidino-2-phenylin-dolc (DAPI) (Sando et al. 1981; Miyakawa et al.1984). By DAPI staining, mt-nucleoids in spheroplastsof stationary phase cells were clearly observed asspherical fluorescent particles dispersed in the cyto-plasm. The use of DAPI and fluorescence microscopyalso provide a useful means for detection and isolationof DNA-containing particles like mt-nucleoids.
The present paper describes the isolation of morpho-logically intact mt-nucleoids from spheroplasts andsome structural and biochemical characteristics ofisolated mt-nuclcoids.
Materials and methods
Strain and cultivation
The diploid strain G2-2 of 5. cerevisiae was culturedacrobically at 30°C to stationary phase in modified Burk-holdcr's medium supplemented with 3-6% (v/v) tomatoextract as described previously (Miyakawa et al. 1984).
Preparation of mitochondria! nucleoids (mt-nucleoids)Mitochondria and mt-nucleoids were prepared from sphero-plasts of stationary phase cells according to the methods ofSuzuki el al. (1982) with slight modification.
Cells of about 50g wet weight were treated with 0-4 M-2-mercaptocthanol (2-ME) at 30°C in SP buffer (O8M-sorbi-tol, 25 mM-potassium phosphate buffer (pli 75) . After wash-ing with SP buffer, cells were treated with zymolyase 60000or 100T (Kirin Brewery Co., Ltd) at 30°C for l h in SPbuffer to make spheroplasts. Spheroplasts, washed twice withSP buffer, were suspended in NE1 buffer (0-3 M-sucrose,20mM-T'risTICI, pH7-6, 1 mM-EDTA and 0-4mM-spermi-dine, 7mM-2-ME, 0-4mM-phenylmethylsulphonyl fluoride(PMSF)). The suspension was homogenized in a Waringblender for 45 s at high speed and centrifuged at 950jf for5 min to remove cell debris. The supernatant was filteredthrough col fee filter paper placed between two layers of nylonsheet and centrifuged at 1600^ for 5 min. The supernatantwas then centrifuged at 2500|f for 5 min and at 12 000^for 20 min. The scdimented pellet was suspended in NE2buffer (0-5 M-sucrose, 20 mM-Tris TIC1, pH 7-6, 2mM-EDTA, 0-8niM-spermidine, 7mM-2-ME, 0-4mM-PMSF)and cv-amylase (2 mg) was added. The mixture was kept for10 min at 0°C, filtered as described above, and centrifugedtwice at 3900|f for 5 min. The supernatant was centrifuged at15 000£ for 20 min to obtain the mitochondrial pellet.
The mitochondrial pellet was thoroughly resuspended inNE2 buffer, and diluted by adding an equal volume of DLbuffer (0-25 M-sucrose, 20 mM-Tris • HC1, pH7-6, 7mM-2-ME, 0-4mM-PMSF). To lyse the mitochondrial membrane,20% Nonidet P-40 (NP-40) was dropped into the solution at4°C to a final concentration of 0-5 % with gentle stirring. Thesolution was centrifuged at 16 000^ for 10 min to remove NP-40 insoluble materials. The clear supernatant was layered on20, 40, 60% (w/v) sucrose discontinuous density gradientin the presence of 20mM-Tris- HC1, pH7-6, 1 mM-EDTA,0-6mM-spcrniidine, 7mM-2-ME, 04mM-PMSF, centrifuged
in a Hitachi RPS 27-2 swing bucket at 46 000g for 1 h. Thefractions containing mt-nucleoids equivalent to the boundarylayer between 20 % and 40 % sucrose and the boundary layerbetween 40% and 60% sucrose were collected and pooled.
Fluorescence measurement
Lysed mitochondrial solution was mixed with l.ugml"1
DAPI and the fluorescence intensity of each fraction afterdensity gradient centrifugation was measured with a Shi-madzu spectrofluorometer RF-500 with the wavelength ofexcitation at 350 nm and emission at 450 nm.
Biochemical analysis
The fractions containing mt-nucleoids were pooled andcentrifuged at 131 000g for 1 h. Pellets of isolated mitochon-dria and mt-nucleoids were suspended in 4% trichloroaceticacid (TCA) dissolved in 50% acetone-water (v/v). After 2hat 0°C, the suspension was centrifuged and the pellet waswashed with 4% TCA. Pellets were washed twice with0'25 M-perchloric acid and then extracted with 0-5M-pcr-chloric acid at 70°C for 15 min. After centrifugation, theprotein content of the pellets was determined by the methodof Lowry et al. (1951) using bovine serum albumin (BSA) asa standard. DNA concentration in the extract was deter-mined by Burton's (1956) method using calf thymus DNA asa standard. RNA concentration in the extract was determinedby the orcinol reaction (Ceriotti, 1955), using yeast RNA as astandard.
Hoechst 33258-CsCI density gradient centrifugationAnalysis of DNA by Hoechst 33258-CsCI density gradientcentrifugation was carried out by the method of Hudspeth elal. (1980). The mitochondrial and nuclear DNA bands werephotographed by illuminating the tubes with long wavelengthu.v. irradiation.
Photomicroscopy
All observations were made with an Olympus BHS-RFKepifluorescence microscope equipped with phase-contrastobjectives, mt-nucleoids were stained with DAPI as de-scribed previously (Miyakawa et al. 1984). Photographs weretaken at a magnification of X850 on 35 mm Fuji Neopan(ASA 400) or Fuji Minicopy film. The number of mt-nucleoids in the fraction was estimated from the ratio of thenumber of mt-nucleoids to the number of polystyrene latexspheres (1-09um in diameter, Dow Chemical Co., Midland,Michigan) in the mixed suspension as described by Suzuki elal. (1982).
Measurement of DNA content of mt-nucleoidsThe DNA contents of mt-nucleoids were determined bymeasuring the fluorescence intensity of each mt-nucieoidstained with DAPI, using a video-intensified microscopephoton counting system (VIMPCS) (Hamamatsu PhotonicsLtd, Hamamatsu, Japan) which was connected to an Olym-pus epifluorescence microscope BHS-RFK as describedpreviously (Kuroiwa el al. 1986). The fluorescence intensityof T4 phage after DAPI staining was used as the standard toexpress the relative fluorescence intensity.
432 /. Mivalunva et al.
Electron microscopyFor electron microscopy, mt-nucleoids were deposited on amesh-grid coated with carbon film by centrifugation (90 000 £for 10min). Negative staining of the mt-nucleoids wasperformed with 1 % sodium phosphotungstate solution neu-tralized to pi 1 7-0 by adding 5 M-NaOI I. Protease digestion ofisolated mt-nucleoids was carried out as follows. One drop ofDL buffer containing pronase E (SOjUgml"') was added tomt-nucleoids on a mesh grid and samples were incubated at20°C for 15 min. After fixation by adding 1 drop of DL buffercontaining 1 % glutaraldehyde, the grid was washed oncewith distilled water and mt-nucleoids were negativelystained. The specimens were observed with a Hitachi HU 500transmission electron microscope at an accelerating voltage of100 kV.
Acid extraction of mt-nucleoids
Acid extraction of proteins from mt-nucleoids was carried outaccording to the method of Sommcr (1978).
Gel electrophoresis
SDS-polyacrylamide gel electrophoresis (SDS-PAGE) wascarried out by the method of Laemmli (1970). Acid-ureaelectrophoresis was carried out according to the methodof Panyim & Chalkley (1969) and further resolution wasachieved by running samples in the second dimension usingthe SDS system of Laemmli (1970). Gels were stained with0-25% Coomassie Brilliant Blue R-250 or by the silverstaining technique (Oakley et al. 1980).
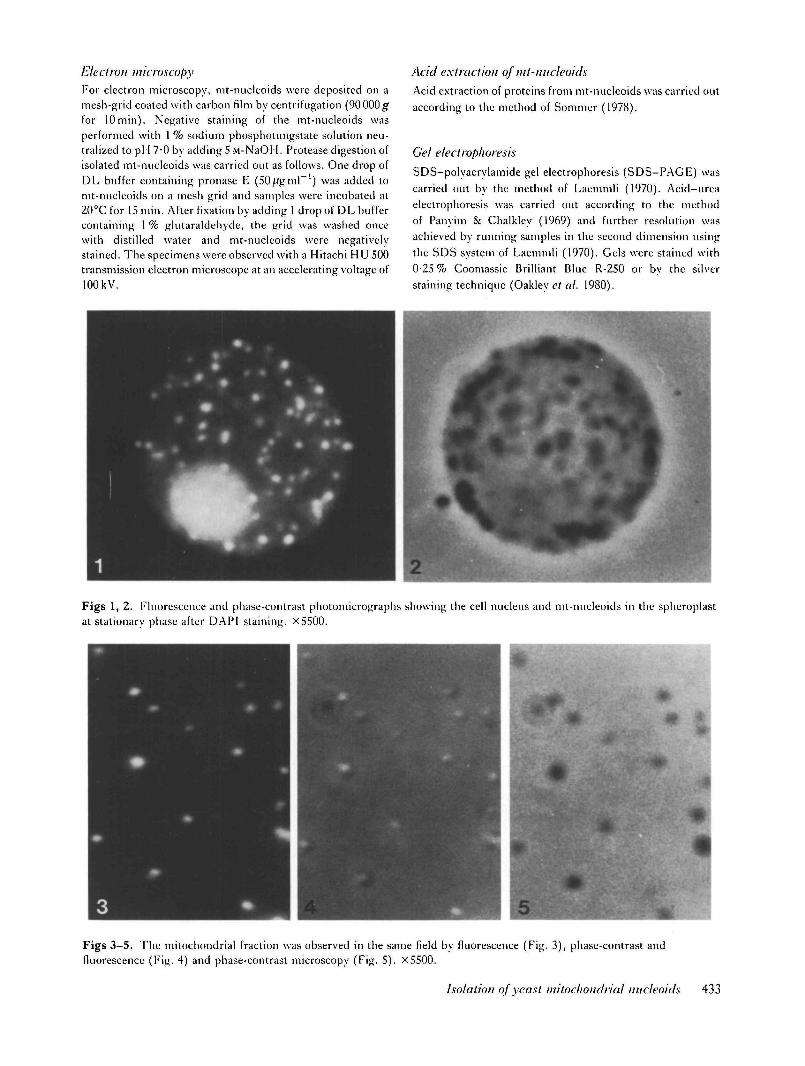
Figs 1, 2. Fluorescence and phase-contrast photomicrographs showing the cell nucleus and mt-nucleoids in the spheroplastat stationary phase after DAPI staining. X5500.
Figs 3-5. The mitochondria! fraction was observed in the same field by fluorescence (Fig. 3), phase-contrast andfluorescence (Fig. 4) and phase-contrast microscopy (Fig. 5). X5500.
Isolation of yeast mitochondria! nucleoids 433
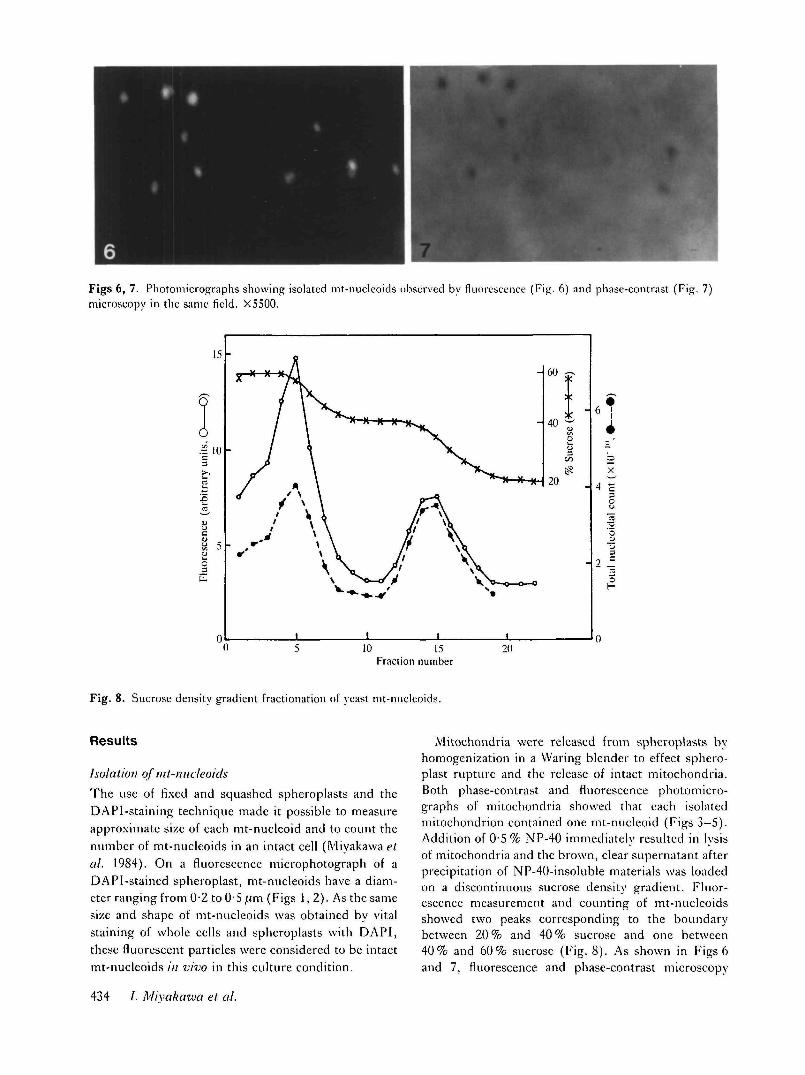
Figs 6, 7. Photomicrographs showing isolated mt-nucleoids observed by fluorescence (Fig. 6) and phase-contrast (Fig. 7)microscopy in the same field. X5500.
10 15Fraction number
20
Fig. 8. Sucrose density gradient fractionation of yeast mt-nucleoids.
Results
Isolation of mt-nucleoids
The use of fixed and squashed spheroplasts and theDAPI-staining technique made it possible to measureapproximate size of each mt-nucleoid and to count thenumber of mt-nucleoids in an intact cell (Miyakawa <?/at. 1984). On a fluorescence microphotograph of aDAPI-stained spheroplast, mt-nucleoids have a diam-eter ranging from 0-2 to OS ;tm (Figs 1,2). As the samesize and shape of mt-nucleoids was obtained by vitalstaining of whole cells and spheroplasts with DAPI,these fluorescent particles were considered to be intactmt-nucleoids /;/ vivo in this culture condition.
Mitochondria were released from spheroplasts byhomogenization in a Waring blender to effect sphero-plast rupture and the release of intact mitochondria.Both phase-contrast and fluorescence photomicro-graphs of mitochondria showed that each isolatedmitochondrion contained one mt-nucleoid (Figs 3-5).Addition of 0-5 % NP-40 immediately resulted in lysisof mitochondria and the brown, clear supernatant afterprecipitation of NP-40-insoluble materials was loadedon a discontinuous sucrose density gradient. Fluor-escence measurement and counting of mt-nucleoidsshowed two peaks corresponding to the boundarybetween 20 % and 40 % sucrose and one between40% and 60% sucrose (Fig. 8). As shown in Figs 6and 7, fluorescence and phase-contrast microscopy
434 /. Mivakawa et al.
mtDNA-
nDNA-
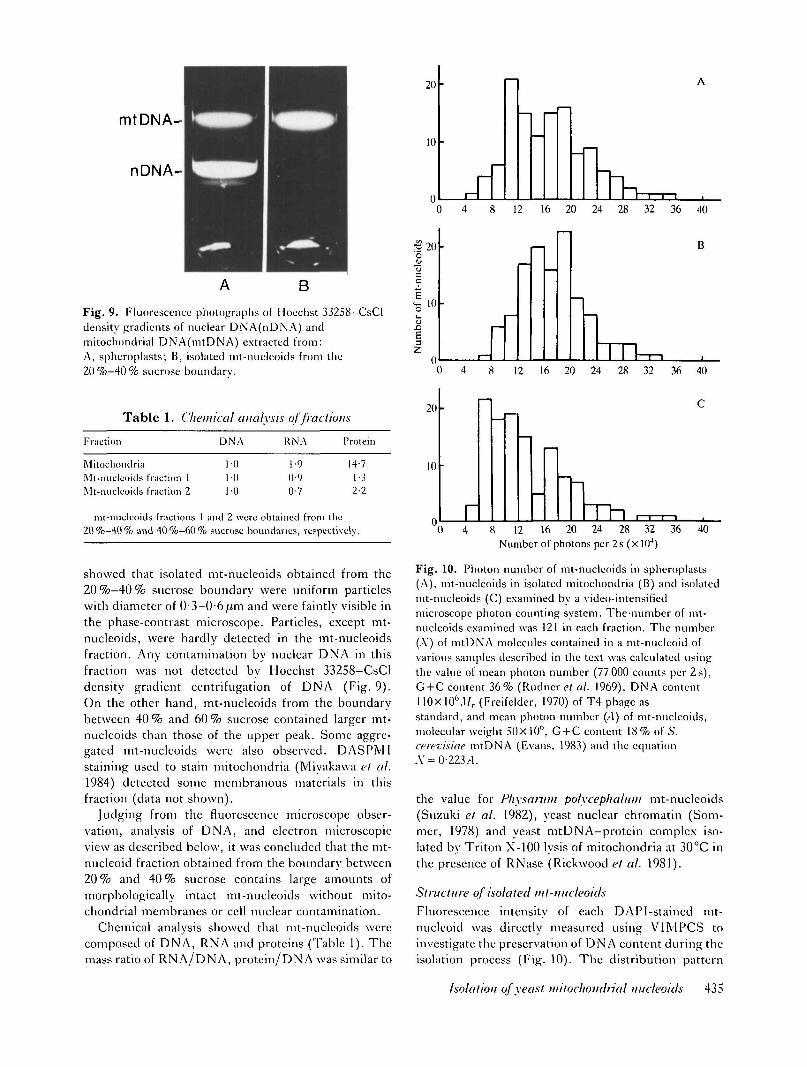
Fig. 9. Fluorescence photographs of I loechst 33258-CsCldensity gradients of nuclear DNA(nDNA) andmitochondrial DNA(mtDNA) extracted from:A, spheroplasts; B, isolated mt-nucleoids from the20 %-40 % sucrose boundary.
Table 1. Chemical analysis of fractions
Fraction
MitochondriaMt-nueleoidsMt-nucleoids
nit-nuclcou
fraction 1fraction 2
Is20%-40% and
fractions 140 %-60 %
DNA
1-01-01-0
and 2 were
1-90-90-7
obtainedsucrose boundaries,
\ Protein
14-1-2-
from therespectively.
732
showed that isolated mt-nucleoids obtained from the20%—40% sucrose boundary were uniform particleswith diameter of 0-3-0-6,um and were faintly visible inthe phase-contrast microscope. Particles, except mt-nucleoids, were hardly detected in the mt-nucleoidsfraction. Any contamination by nuclear DNA in thisfraction was not detected by Hoechst 33258-CsCldensity gradient centrifugation of DNA (Fig. 9).On the other hand, mt-nucleoids from the boundarybetween 40 % and 60 % sucrose contained larger mt-nucleoids than those of the upper peak. Some aggre-gated mt-nucleoids were also observed. DASPM1staining used to stain mitochondria (Miyakawa et al.1984) detected some membranous materials in thisfraction (data not shown).
Judging from the fluorescence microscope obser-vation, analysis of DNA, and electron microscopicview as described below, it was concluded that the mt-nucleoid fraction obtained from the boundary between20% and 40% sucrose contains large amounts ofmorphologically intact mt-nucleoids without mito-chondrial membranes or cell nuclear contamination.
Chemical analysis showed that mt-nucleoids werecomposed of DNA, RNA and proteins (Table 1). Themass ratio of RNA/DNA, protein/DNA was similar to
20
10
0 4 12 16 20 24 28 32 36 40
^ 2 0o
10
B I 7TU-.0 4 8 12 16 20 24 28 32 36 40
20
10
0 4 8 12 16 20 24 28 32 36 40Number of photons per 2 s (x 104)
Fig. 10. Photon number of mt-nucleoids in spheroplasts(A), mt-nucleoids in isolated mitochondria (B) and isolatedmt-nucleoids (C) examined by a video-intensifiedmicroscope photon counting system. Thenumber of mt-nucleoids examined was 121 in each fraction. The number(A) of mtDNA molecules contained in a mt-nucleoid ofvarious samples described in the text was calculated usingthe value of mean photon number (77 000 counts per 2 s),G + C content 36% (Rudnere/ al. 1969), DNA content110xl0°.Ur (Freifelder, 1970) of T4 phage asstandard, and mean photon number (A) of mt-nuclcoids,molecular weight 50X 10°, G + C content 18 % of .S'.cerevisiae mtDNA (Evans, 1983) and the equationA = 0-223.4.
the value for Physaniin polycephaltmi mt-nucleoids(Suzuki et al. 1982), yeast nuclear chromatin (Som-mer, 1978) and yeast mtDNA-protein complex iso-lated by Triton X-100 lysis of mitochondria at 30cC inthe presence of RNase (Rickwood et al. 1981).
Structure of isolated mt-imcleotdsFluorescence intensity of each DAPI-staincd mt-nucleoid was directly measured using VIM PCS toinvestigate the preservation of DNA content during theisolation process (Fig. 10). The distribution pattern
Isolation of yeast mitochondrial micleoids 435
12
13
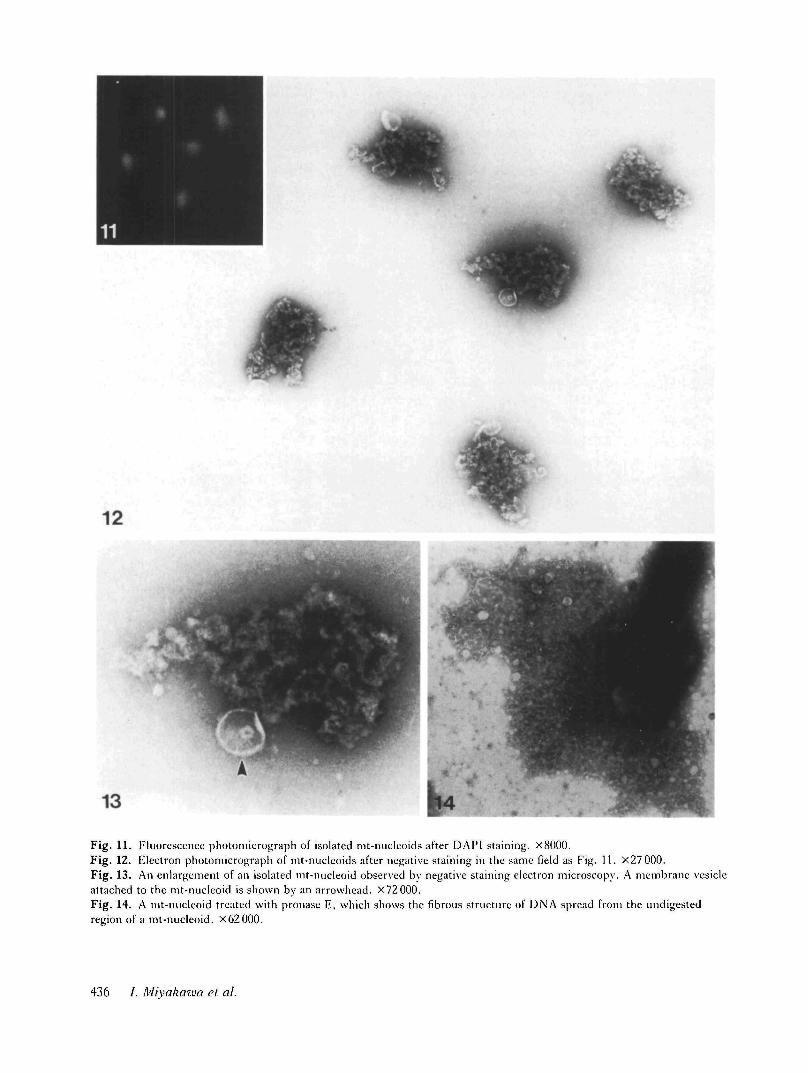
Fig. 11. Fluorescence photomicrograph of isolated mt-nuclcoids after DAPI staining. X8000.Fig. 12. Electron photomicrograph of mt-nucleoids after negative staining in the same field as Fig. 11. X27 000.Fig. 13. An enlargement of an isolated mt-nucleoid observed by negative staining electron microscopy. A membrane vesicleattached to the mt-nucleoid is shown by an arrowhead. X72000.Fig. 14. A mt-nucleoid treated with pronase E, which shows the fibrous structure of DNA spread from the undigestedregion of a mt-nucleoid. X62000.
436 /. Mivakawa el al.

of fluorescence intensity per nucleoid in isolatedmitochondria coincided well with the pattern of fluor-escence intensity of nucleoids in spheroplasts. How-ever, the distribution of isolated nucleoids shifted tothe smaller side as a whole, indicating that selectiveisolation of smaller nucleoids might occur in thefraction. DNA content of a mt-nucleoid was calculatedfrom photon numbers on the basis of G + C content ofmtDNA and T4 phage used as standard, becauseDAPI has specific affinity to A-T base-pair (William-son & Fennell, 1975; Kapuscinski & Skoczylas, 1977;Lin el al. 1977; Kapuscinski & Szer, 1979). Theaverage number of mtDNA molecules in an mt-nucleoid was estimated as 3-9 in spheroplasts, 4-0 inisolated mitochondria and 3-1 in isolated mt-nucleoids,respectively.
In order to observe the fine structure of the isolatedmt-nucleoids, DAPI-stained mt-nucleoids depositedon a mesh-grid were observed with the fluorescencemicroscope and subsequently observed in the electronmicroscope in the same field after negative staining(Figs 11 and 12). Electron microscopic observationclearly showed that the mt-nucleoids have a sphericalor ovoid configuration 0-4-0-6/im in diameter intowhich the chromatin-like structure is three-dimen-sionally folded. An enlarged figure of one of the mt-nucleoids is shown in Fig. 13. mt-nucleoids consistedof particles measuring 20—50 nm in diameter and insome mt-nucleoids one or two membrane vesicles wereobserved to attach to the core portion of foldedchromatin. Pronase E digestion disrupted the chroma-tin-like structure of the mt-nucleoid and a DNA-like
94-
67
43- T~
30-i20-
OT94-§67-
4 3 -
3 0 -
2 0 -14 —
Fig. 16. Acid—urea/SDS two-dimensional PAGE of acidextract from mt-nucleoids. The first dimensional acid-ureagel is shown at the top of a two-dimensional SDS gel.Direction of electrophoresis is indicated by an arrow. A20xl03;l/ r polvpeptide is indicated by an arrowhead. Gelswere stained with Coomassie Brilliant Blue.
thin filament (less than 7 nm in diameter) appearedradiating from the undigested region of the mt-nucleoid (Fig. 14). Apart from pronase E, the nuc-leoids were completely dispersed by treatment withNaOH, HC1, DNase and SDS, tended to expand whenheat treated or exposed to high KG or NaCl concen-trations and were resistant to RNase.
Niicleoid-oiganizing proteins
SDS—PAGE of mt-nucleoids was performed to investi-gate proteins which organize mtDNA into nucleoids(Fig. 15). . The mt-nucleoid fraction from the20 %—40 % sucrose boundary contained at least 20species of polvpeptides mainly in the molecular weightrange from 10 to 70(X 103). As Rickwood el al. (1981)reported, we also found several polvpeptides above70K (K = 103/l/r), but these polvpeptides were quanti-tatively minor components of nucleoids. To search forbasic, histone-like proteins, acid extract from nucleoidswas analysed by acid—urea/SDS two-dimensional elec-trophoresis (Fig. 16). The results revealed only onemajor polvpeptide of 20K, which suggested that the20K polvpeptide plays an important role in packing ofmtDNA similar to nuclear histones.
Discussion
14- ^LFig. 15. Analysis of mt-nucleoid proteins by SDS-PAGE.Molecular weight standard is indicated to the left of a gel(xlCT3).
In the electron microscope mtDNA appears as tinyelectron-dense spots, filaments, or bundles of filamentsin regions of lower electron density in the mitochon-drial matrix. This depends on the fixation method usedand it has been assumed that the almost complete
Isolation of yeast mitochondrial nucleoids 437

absence of associated protein is responsible for theclumping of DNA (Stevens, 1981). On the other hand,Rickwood el til. (1981) isolated the DNA-proteincomplex from mitochondria of the yeast, Saccham-myces cerevisiae and detected four species of proteinsassociated with mt-nucleoids. In the electron micro-scope the isolated DNA-protein complexes appearedas various sized aggregates composed of spheroidalbodies. However, it was not certain whether DNA-protein complexes remained intact.
The purpose of this study was to isolate morphologi-cally intact mt-nucleoids without disrupting theirnative structure and to investigate the organization.For this purpose, DAPI seems a useful tool formonitoring the preservation of size, shape and (es-pecially combined with VIM PCS) DNA content ofnucleoids during the isolation process.
To obtain highly pure mitochondria from yeast,combined methods of repeated differential centrifu-gations and density gradient eentrifugation have beenwidely used, and DNase I treatment of mitochondriahas also been generally carried out to remove contami-nated cell nuclear DNA. However, in this study theprolonged isolation process had to be avoided to protectmt-nucleoids from proteolytic and nucleolytic diges-tion, which cause the disruption of the intact mor-phology of mt-nucleoids, and to obtain a good yield. Inthe present work, we found it possible to isolate mt-nucleoids with high purity without the time-consumingdensity gradient eentrifugation and DNase I digestionof mitochondria. In fact, the mitochondrial fractioncontained some membranous structures besides mito-chondria. Analysis of DNA by Hoechst-CsCl densitygradient also revealed contamination of nuclearchromatin of about 20% of the total DNA in themitochondrial fraction. However, it was possible toremove almost all cell nuclear chromatin and NP-40-insoluble membranous material from the NP-40 lysateof mitochondria as an NP-40-insoluble fraction. Theremaining contamination cosedimented with aggre-gates of mt-nucleoids in the fraction at the 40 %-60 %sucrose boundary. As a result, the fraction from the20%-40% sucrose boundary contained highly puremt-nucleoid particles as seen in Figs 11 and 12, wheninvestigated by fluorescence and electron microscopy.Analysis by a CsCl density gradient always showedabove 98 % purity of mtDNA in total DNA as seen inFig. 9. The rapid method described here enabled usto isolate morphologically intact mt-nucleoids fromspheroplasts within 5h, although we might selectsomewhat smaller mt-nucleoids during the isolationprocess. Analysis of nucleoid-organizing proteins pro-duced quite different results from those of Rickwood elal. (1981) who detected only four major polypeptides(68K, 80K, 2 species of 95K) using a [35S]sulphate-labelling technique. Although our results also detected
several polypeptides of high molecular weight above70K, these polypeptides were minor components ofmt-nucleoids. The discrepancy might be partly causedby the difference in the method for detection ofpolypeptides. In the case of yeast, it has been claimedthat a basic histone-like protein named HM proteinwith molecular weight of 20K is present in mitochon-dria (Caron el al. 1979; Certa et al. 1984). Weconfirmed that the 20K protein, which may be identicalto HM protein, is really one of the major components ofnucleoids with the most basic property. At present, thefunctions of almost all other polypeptides detected inmt-nucleoids still remain to be determined. Rickwood& Chambers (1981) have reported that the significantfraction of the mtDNA of S. cerevisiae is organizedinto a nucleosome-like structure. Our preliminary workwith DNA-cellulose affinity chromatography indicatesthat at least five major species of polypeptides includingthe 20K polypeptide have the ability to bind to double-stranded DNA. Further investigation of isolated mt-nucleoids would bring us new information about how75 kb circular mtDNAs are compactly folded.
We wish to thank Dr T. Suzuki of Nagoya UniversitySchool of Medicine for his valuable advice. This work wassupported in part by Grants-in-Aid for Scientific Researchnos 60740383 and 60304008 from the Ministry of Education,Science and Culture of Japan. This work was carried outunder the N1BB Cooperative Research Program(1981-1986).
References
ALBRING, M., GRIFFITH, J. & ATTARDI, G. (1977).
Association of a protein structure of probable membranederivation with HeLa cell mitochondrial DNA near itsorigin of replication. Proc. naln. Acad. Sci. U.S.A. 74,1348-1352.
BURTON, K. (1956). A study of the conditions andmechanism of the diphenylamine reaction for thecolorimetric estimation of deoxvribonucleic acid.Biochem. J. 62, 315-323.
CARON, F., JACQ, C. & ROUVIERE-YANIV, J. (1979).
Characterization of a histone-like protein extracted fromyeast mitochondria. Proc. naln. Acad. Sci. L.S.A. 76,4265-4269.
CERIOTTI, G. (1955). Determination of nucleic acids inanimal tissues. .7- biol. Chew. 241, 59-70.
CERTA, U., COLAVITO-SHEPANSKI, M. & GRUNSTEIN, M.
(1984). Yeast may not contain historic Ml : the onlyknown 'histone HI-like' protein in Saccharoinvcescerevisiae is a mitochondrial protein. Xncl. Acid Res. 12,7975-7985.
DUJON, B. (1981). Mitochondrial genetics and functions.In The Molecular Biology of the Yeast Saccharoniyces.Life Cycle and Inheritance (ed. J. N. Strathern, E. W.Jones & J. R. Broach), pp. 59-96. New York: ColdSpring Harbor Laboratory.
438 /. Mivakaiva el al.

EVANS, I. II. (1983). Molecular genetic aspects of yeastmitochondria. In Yeast Genetics. Fundamental andApplied Aspects (ed. J. F. T. Spencer, D. M. Spencer &A. R. W. Smith), pp. 269-370. New York: Springer-Verlag.
FREIFELDER, D. (1970). Molecular weights of coliphagesand coliphage DNA. IV. Molecular weights of DNAfrom bacteriophage T4, T5, T7 and the general problemof determination of M.Jf. inolec. Biol. 54, 567-577.
IIUDSPETH, M. E. S., SHUMARD, D. S., TATTI, K. M. &
GROSSMAN, L. I. (1980). Rapid purification of yeast(Saccharoinyces cerevisiae) mitochondrial DNA in highyield. Biochim. biophys. Ada 610, 221-228.
KAPUSCINSKI, J. & SKOCZYLAS, B. (1977). Simple andrapid fluorimetric methods for DNA microassay. Analyt.Biochem. 83, 252-257.
KAPUSCINSKI, J. & SZER, VV. (1979). Interactions of 4',6-diamidine-2-phcnyhndole with synthetic polynucleotides.Sucl. Acid Res. 6, 3519-3534.
KUROIWA, T. (1982). Mitochondrial nuclei. Int. Rev.Cylol. 75, 1-59.
KUROIWA, T., KAWANO, S. & HIZUME, M. (1976). A
method of isolation of mitochondrial nucleoid ofPhvsarum polvcephalum and evidence for the presence ofa basic protein. F.xpl Cell Res. 97, 435-440.
KUROIWA, T., MIYAMURA, S., KAWANO, S., HIZUME, M.,
TOH-E, A., MIYAKAWA, I. & SANDO, N. (1986).
Cytological characterization of NOR in the bivalent ofSaccharoinyces cerevisiae. Fxpl. Cell Res. 165, 199-206.
LAEMMU, U. K. (1970). Cleavage of structural proteinsduring assembly of the head of bacteriophage T4.Salure, hind. Ill, 680-685.
LIN, M. S., COMINGS, D. E. & ALFI, O. S. (1977). Opticalstudies of the interaction of 4',6-diamidino-2-phenylindole with DNA and metaphase chromosomes.Chromosoma 60, 15-25.
LOWRY, O. II., ROSEBROUGH, N. J., FARR, A. L. &
RANDALL, R. J. (1951). Protein measurement with theFolin phenol reagent. J. biol. Chem. 193, 265-275.
MIYAKAWA, I., Aoi, II., SANDO, N. & KUROIWA, T.
(1984). Fluorescence microscopic studies ofmitochondrial nucleoids during meiosis and sporulationin the yeast, Saccharoinyces cerevisiae. J. Cell Sci. 66,21-38."
OAKLEY, B. R., KIRSCH, D. R. & MORRIS, N. R. (1980). A
simplified ultrasensitive silver stain. Analvt. Biochem.105, 361-363.
OLSZEWSKA, E. & T A I T , A. (1980). Mitochondrialchromatin in i'arainecium aurelia. Molec. gen. Genet.178, 453-457.
PANYIM, S. & CHALKLEY, R. (1969). High resolutionacrylamide gel electrophoresis of histories. Arch.Biochem. Biophys. 130, 337-346.
PINON, II., BARAT, M., TOURTE, M., DUFRESNE, C. &
MOUNOLOU, J. C. (1978). Evidence for mitochondrial
chromosomes in Senopns laevis oocytcs. Chromosoma65, 383-389.
RICKWOOD, D. & JURD, R. D. (1978). A general methodfor the isolation and partial characterization ofmitochondrial nucleoids by centrifugation in metrizamidegradients. Biochem. Soc. Trans. 6, 266-268.
RICKWOOD, D., CHAMBERS, J. A. A. & BARAT, M. (1981).
Isolation and preliminary characterisation of DNA—protein complexes from the mitochondria ofSaccharoinyces cerevisiae. Fxpl Cell Res. 133, 1-13.
RICKWOOD, D. & CHAMBERS, J. A. A. (1981). Evidence forprotected region of DNA in the mitochondrial nucleoidof Saccharoinyces cerevisiae. FFMS Microbiol. letters12, 187-190.
RUDNER, R., KARKAS, J. D. & CHARGAFF, E. (1969).
Separation of microbial dcoxyribonucleic acids intocomplementary strands. Proc. natn. Acad. Sci. I .S.A.63, 152-159. "
SANDO, N., MIYAKAWA, I., NISHIBAYASHI, S. & KUROIWA,
T. (1981). Arrangement of mitochondrial nucleoidsduring life cycle of Saccharoinyces cerevisiae. J. gen.appl. Microbiol., Tokyo 27, 511-516.
SEVALJEVIC, L., PETROVIC, S. & RICKWOOD, D. (1978).
Isolation and partial characterization of a mitochondrialdeoxyribonucleic acid-protein complex from sea urchinembryos. Molec. cell. Biochem. 21, 139-143.
SEVALJEVIC, L., RICKWOOD, D. & TOMOVIC, M. (1979).
Comparative characterization of mitochondrial nucleoidsand of nuclear chromatin of sea urchin embryos. Molec.cell. Biochem. 23, 53—61.
SOMMER, A. (1978). Yeast chromatin: Search for historicHI. Molec. gen. Genet. 161, 323-331.
STEVENS, B. J. (1981). Mitochondrial structure. In TheMolecular Biology of the Yeast Saccharoinyces. Life Cycleand Inheritance (ed. J. N. Strathern, E. W. Jones & J.R. Broach), pp. 471-504. New York: Cold SpringHarbor Laboratory.
SUZUKI, T., KAWANO, S. & KUROIWA, T. (1982).
Structure of three-dimensionally rod-shapedmitochondrial nucleoids isolated from the slime mouldPhysarum polycephalum. J. Cell Sci. 58, 241-261.
VAN TUYLE, G. C. & MCPHERSON, M. L. (1979). A
compact form of rat liver mitochondria DNA stabilizedby bound proteins. J. biol. Chem. 254, 6044-6053.
VAN TUYLE, G. C. & PAVCO, P. A. (1985). The rat livermitochondria DNA-protein complex: Displaced singlestrands of replicative intermediates are protein coated..7.Cell Biol. 100, 251-257.
WILLIAMSON, D. H. & FENNELL, D. J. (1975). The use of
fluorescent DNA-binding agent for detecting andseparating yeast mitochondrial DNA. In Methods in CellBiology, vol. 12 (ed. D. M. Prescott), pp. 335-351. NewYork: Academic Press.
(Received 30 March 1987 - Accepted 10 August 1987)
Isolation of yeast mitochondrial nucleoids 439






























