Embed Size (px)
Citation preview

Isolation, Biochemical Characterization and
DNA Identification of Yoghurt Starters
Streptococcus Thermophilus & Lactobacillus
Delbrueckii ssp. Bulgaricus
ن الستربتوكوكاس والجينية لبادئات اللبالبيوكيميائية عزل وتحديد الخصائص ثيرموفيالس والالكتوباسيالس ديليبروكي تحت نوع بولجاريكاس
By
Ismail Mosbah Alquqa
Supervised by
Dr. Kamal Elkahlout
Assis. prof. of Biotechnology
Dr. Tarik Elbashiti
Assoc. prof. of Biotechnology
A thesis submitted in partial fulfillment
of the requirements for the degree of
Master of Science in Biotechnology
Apr. /2018
ـزةبغـ اإلســـــالميــةـة ـــــــــامعـالج
عمادة البحث العلمي والدراسات العليا
ـــــــــــــــومة العلـــــــــــــــــــــــليــــــك
ــةالتكنولوجيا الحيويــــــــــــماجستير
The Islamic University of Gaza
Deanship of Research & Graduate Studies
Faculty of Science
Master of Biotechnology

I
إقــــــــــــــرار
أنا الموقع أدناه مقدم الرسالة التي تحمل العنوان:
Isolation, Biochemical Characterization and DNA
Identification of Yoghurt Starters Streptococcus
Thermophilus & Lactobacillus Delbrueckii ssp.
Bulgaricus
بتوكوكاس ن الستر اللب اللبن والجينية لبادئات البيوكيميائيةعزل وتحديد الخصائص ا ثيرموفيالس والالكتوباسيالس ديليبروكي تحت نوع بولجاريكاس
د، وأن أقر بأن ما اشتملت عليه هذه الرسالة إنما هو نتاج جهدي الخاص، باستثناء ما تمت اإلشارة إليه حيثما ور
لنيل درجة أو لقب علمي أو بحثي لدى أي مؤسسة االخرين الرسالة ككل أو أي جزء منها لم يقدم من قبلهذه
تعليمية أو بحثية أخرى.
Declaration
I understand the nature of plagiarism, and I am aware of the University’s policy on
this.
The work provided in this thesis, unless otherwise referenced, is the researcher's own
work, and has not been submitted by others elsewhere for any other degree or
qualification.
إسماعيل مصباح القوقا اسم الطالب: Student's name: Ismail Mosbah Alquqa
إمساعيل القوقا التوقيع: Signature: Ismail Alquqa
Date: 20/6/2018 20/6/2018 التاريخ:

II


III
Abstract
Fermented dairy products are manufacured by specific bacterial strains which are
known as starters. Yoghurt is prepared from milk by the synergetic action of well-known
starters Streptococcus thermophilus and Lactobacillus delbrueckii ssp. bulgaricus. These
starters by lactic acid fermentation cause milk coagulation and other features, such as sourness,
aroma, and consistency. Old-style approaches, using parts of a former batch to inject a new
batch for centuries. These styles produce inconstant performance, but industrial production
needs consistency. With this perspective, this study aims to isolate starters from households of
geographical regions of the Gaza strip and to define their phonotypic and genotypic
characteristics.
M17 and MRS culture media were used to pick out streptococci and lactobacilli,
respectively from rural yoghurt samples. Under anaerobic environments, all plates incubated
at 37ºC for 3 days by anaerobic gars. Strains, which identified as L.bulgaricus and
S.thermophilus by simple physiological examinations, i.e. morphology, CO2 production from
glucose, NaCl resistance , growing at different temperatures, were identified by carbohydrate
fermentation profiles using API 50 CHL identification system and species- specific primers
by PCR using 16S rRNA sequences to confirm preliminary tests.Then, the isolated starters
injected in milk to evaluate it on yoghurt making, microbiological and biochemical changes
that take place during fermentation of milk were also studied. Generally, mixed yoghurt
culture presented superior growth, more sourness comparing with the single starter cultures.
In conclusions, genetic tools in combination with API 50 CHL were shown to be
reliable and quick approaches to identify S. thermophilus and L. delbrueckii isolates at the
subspecies level and to differentiate these species from other lactic acid bacteria without prior
preliminary isolation. The natural starters' identification is a required for the preservation and
protection of natural LAB diversity in the Gaza Strip, not only for the dairy Industry which
still imports starters abroad.
Keywords: Streptococcus thermophilus, Lactobacillus delbrueckii ssp. Bulgaricus,
Yoghurt, Starters, 16S rRNA, PCR.

IV
الملخص
لتصنيع منتجات من الحليب المخمر. الخاصة والمعروفة بإسم البادئاتالبكتيرية سالالتالتستخدم الستربتوكوكاس ثيرموفيالس لبادئاتافري ظمن الحليب بواسطة سلوك ت اللبن عادة يصنع
وخصائص البادئات إلى تخثر الحليب هذهتقود .بولجاريكاسوالالكتوباسيالس ديليبروكي تحت نوع الطرق التقليدية جزء تستخدم .الالكتيكعن طريق تخمر حمض أخرى، مثل الحامضية، النكهة والِقوام
اج المزارع إلى كفاءة متغيرة، ولكن االنتاج المصنعي يحت هذهتقود دفعة جديدة. حقنمن دفعة سابقة ل، مناطق جغرافية لقطاع غزةعزل البادئات من هذه الدراسة إلى تهدفإلى الِقوام. من هذا المنظور،
تحديد الخصائص المظهرية والجينية لها.ومن ثم الترتيب من على والالكتوباسياليالستربتوكوكاي لعزل MRS و M17األوساط الغذائية تم استخدام
أيام تحت ظروف درجة مئوية لثالث 37ة على درجة حرار جميع االطباق ضينتم تحعينات لبن ريفية. ستربتوكوكاس ثيرموفيالس أنهاالهوائية. السالالت التي ُحددت على حاويات الهوائية باستخدام
كل، النمو عند درجات االختبارات الفسيولوجية البسيطة مثل الش بواسطة والكتوباسيالس بلجاريكاس ا أيضاتم التعرف عليه ج ثاني أكسيد الكربون من الجلوكوز، مقاومتها لملح كلوريد الصوديومامختلفة، إنت
برايمرات للنوع متخصصة وبواسطة API 50 CHLبواسطة فحص تخمر الكربوهيدرات باستخدام التغيرات ةتم دراس . بعد ذلكلتأكيد الفحوصات األولية PCR تقنيةواسطة ب 16S rRNA استخدامب
ام، عالميكروبيولوجية والبيوكيميائية الحادثة أثناء تخمر الحليب المحقون بالبادئات المعزولة. بشكل مقارنة مع مزارع البادئات المفردة. أكثرمزارع البادئات المختلطة نموًا أعلى وحموضة أظهرت ت طرق موثوقة وسريعة لتحديد عزال لتصبح API 50 CHLبجانب الجينية وسائلالثبتت أأخيرًا،
كتيك ولتمييز هذه األنواع من بكتيريا حمض الال بلجاريكاس الستربتوكوكاس ثيرموفيالس والالكتوباسيالساألخرى بدون عزل تمهيدي مسبق. التحديد للبادئات الطبيعية هو احتياج ليس لمصانع األلبان فحسب،
عية في والتي ال تزال تستورد البادئات من الخارج، لكن أيضًا لحفظ التنوع لبكتيريا حمض الالكتيك الطبي قطاع غزة.
،بادئات، نلب ،ستبرتوكوكاس ثيرموفيالس، الكتوباسيالس ديليبروكي بلجاريكاس :المفتاحيةالكلمات
16S rRNA، PCR.

V

VI
Dedicated
To
My Mother
A strong and gentle soul who taught me to trust in
Allah, believe in hard work and that so much
could be done with little
My Father
For earning an honest living for us and for
supporting and encouraging me to believe in
myself
My Wife
Who leads me through the valley of darkness
with light of hope and support
The little darling, my Daughter,
" Dania "
My Beloved
Brothers and Sisters

VII
Acknowledgment
Foremost, I would like to express my sincere gratitude to my supervisor Dr. Kamal
Elkahlout for the continuous support of my MSc. study and research, for his patience,
motivation, enthusiasm, and immense knowledge. His guidance helped me in all the
time of research and writing of this thesis. I could not have imagined having a better
advisor and mentor for my Msc. study.
My sincere thanks also goes to Dr. Tarik Elbashiti, the second supervisor, for
enlightening me the first glance of research, for his Directives in hard times during my
research study, and for his all kind of help.
Also special thanks to Mr. Hussein Ajrami & Mr. Husam AboTayyem for their useful
contributions, and sharing their valuable science experiences.
Last but not the least, I would like to thank my parents for giving birth to me at the
first place and supporting me spiritually throughout my life.

VIII
contents
DECLARATION .......................................................................................................... I
ABSTRACT IN ENGLISH ......................................................................................... II
ABSTRACT IN ARABIC ......................................................................................... III
DEDICATION ............................................................................................................. V
ACKNOWLEDGMENT ........................................................................................... VI
LIST OF TABLES ....................................................................................................... X
LIST OF FIGURES .................................................................................................. XI
LIST OF ABBREVIATIONS ................................................................................. XIV
CHAPTER 1 INTRODUCTION ................................................................................. 1
1.1 Background and Context ................................................................... 2
1.2 Objectives .................................................................................... 3
1.2.1General objective: ............................................................................................ 3
1.2.2Specific objective: ............................................................................................ 3
1.3 Signification ................................................................................. 3
1.4 Limitations ................................................................................... 3
1.5 Overview of Thesis ......................................................................... 4
CHAPTER 2 LITERATURE REVIEW ..................................................................... 5
2.1 What is Yoghurt? ........................................................................... 6
2.2 The History of Yoghurt .................................................................... 6
2.3 Microorganisms used in starters for cultured dairy products ......................... 7
2.4 Yoghurt Starter Cultures ................................................................. 12
2.4.1 Streptococcus thermophilus ......................................................................... 14
2.4.2 Lactobacillus delbrueckii subsp. buglaricus ………………….…………..17
2.5 Protocooperation between S.thermophilus & L.bulgaricus ......................... 19
2.5.1 Factors of protocooperation .......................................................................... 20
2.7 classification of LAB based on by-products of suger................................ 23
2.7.1 Homofermentative LAB ............................................................................... 23
2.7.2 Heterofermentative LAB .............................................................................. 23
2.7.3 Other Gas Producing Pathways: ................................................................... 23
2.8 Types of Yoghurt ......................................................................... 24
2.9 Yoghurt Processing ....................................................................... 26

IX
2.9.1 Initial Treatment of Milk .............................................................................. 27
2.9.2 Standardization of Milk Components ........................................................... 27
2.9.2.1 Standardization of fat content: ............................................................... 27
2.9.2.2 Standardization of solid (nonfat) content: ............................................. 28
2.9.3 Addition of additives .................................................................................... 28
2.9.4 Homogenization ............................................................................................ 28
2.9.5 Heat Treatment ............................................................................................. 29
2.9.6 Fermentation Process .................................................................................... 29
2.9.7 Cooling and Storage ...................................................................................... 30
2.10 Factors affecting slow growth of starter cultures and yoghurt quality: .......... 30
2.10.1 Compounds that are naturally present in milk ............................................ 30
2.10.2 Effect of incubation temperature and inoculation rate ................................ 31
2.10.3 Mastitis milk and somatic cell count .......................................................... 31
2.10.4 Antibiotic residues ...................................................................................... 31
2.10.5 Detergent and disinfectant residues ............................................................ 32
2.10.6 Environmental pollution ............................................................................. 32
2.10.7 Bacteriophages ............................................................................................ 32
2.10.8 Bacteriocins ................................................................................................ 33
2.11 Molecular identification: ............................................................... 33
CHAPTER 3 MATERIALS AND METHODS ........................................................ 35
3.1 Materials ................................................................................... 36
3.1.1 Chemicals and Reagents ............................................................................... 36
3.1.2 Equipments ................................................................................................... 37
3.2 Methodology: .............................................................................. 37
3.2.1 Drinking yoghurt samples ............................................................................. 37
3.2.2 Isolation of lactic acid bacteria ..................................................................... 37
3.2.2.1 Selective Media and Growth Conditions .............................................. 37
3.2.3 Phenotypic identification of isolates ............................................................. 38
3.2.3.1 Gram Staining ........................................................................................ 38
3.2.3.3 Gas Production from Glucose ................................................................ 38
3.2.3.4 Growth at Different Temperatures ......................................................... 39
3.2.3.5 Growth at Different NaCl Concentrations ............................................. 39

X
3.2.3.6 Carbohydrate Fermentation profiles ...................................................... 39
3.2.3.6.1 Preparation of active cell culture free from sugar: ......................... 40
3.2.3.6.2 Combination of Active Cell Culture free from sugar and tested
sugar: .............................................................................................................. 40
3.2.4 Molecular identification for Isolated Bacteria ............................................. 40
3.2.4.1 DNA isolation ........................................................................................ 40
3.2.4.2 Primers ................................................................................................... 40
3.2.4.3 PCR conditions ...................................................................................... 41
3.2.4.4 Restriction analysis of 16S rRNA gene product ................................... 41
3.2.4.5 Separation of Amplification Products .................................................... 41
3.2.4.5.1 Preparation of Agarose Gel ............................................................. 41
3.2.4.5.2 Loading of Agarose Gel .................................................................. 42
3.2.4.5.3 Electrophoresis of the Products ...................................................... 42
3.2.5 Assessment of technological performance of strains: ................................... 42
3.2.5.1 Preparation of fermented milk ............................................................... 42
3.2.5.2 Measurement of pH and titratable acidity .............................................. 42
CHAPTER 4 RESULTS AND DISCUSSION .......................................................... 44
4.1 Isolation and Biochemical identification of S. thermophilus and L. delbrueckii
ssp. bulgaricas .......................................................................................................... 45
4.2: DNA Identification ...................................................................... 48
4.3: Assessment of technological performance of strains ............................... 50
4.3.1: Microbiological analysis .............................................................................. 50
4.3.1.1 The viable counts of starter cultures during fermentation ..................... 50
4.3.2: Biochemical analysis ................................................................................... 54
4.3.2.1: Changes in the Total acidity and pH ..................................................... 54
CHAPTER 5 CONCLUSIONS AND RECOMMENDATIONS .............................. 62
REFERENCES ........................................................................................................ 64
APPENDIXES ........................................................................................................... 72
APPENDIX A.................................................................................. 73
APPENDIX B .................................................................................. 75
APPENDIX C .................................................................................. 76

XI
LIST OF TABLES
Table (2.1): Microorganisms Used In Starter Culture For Fermented Milk Products
And Their Functions. .................................................................................................. 9
Table (2.2): Features Of Mesophilic Starters Used For Fermented Milk Products. 10
Table (2.3): Features Of Thermotolerant Starters Used For Fermented Milk
Products. ................................................................................................................... 11
Table (2.4): Required & Optional Composition Of Yoghurt Bacteria ..................... 14
Table (2.5): The Composition Of Regular, Low-Fat & Non-Fat Yoghurt. .............. 23
Table (3.1): Chemicals, Reagents And Cultures Mediums ....................................... 36
Table (3.2): Major Equipments Used In The Present Study. .................................... 37
Table (4.1): Ability Of The Tested Strains To Utilize The 49 Carbon Sources Included
In The Identification System API50 CHL. ............................................................... 47
Table (4.2): The Changes On The Viable Counts Of The Starter Culture Strains
During Fermentation Of Milk For 8h At 42ºc ........................................................... 50
Table (4.3): Effect Of Culture Composition On Acidification Characteristics Of Pure
& Mixed Cultures Grown On Milk At Different Times On 42ºc. ............................ 54
Table (4.4): Effect Of Culture Composition On Total Acidity Of Pure & Mixed
Cultures Grown On Milk At 42ºc. ............................................................................. 58
Table (A.1): MRS Medium ....................................................................................... 73
Table (A.2): M17 Medium ........................................................................................ 74

XII
LIST OF FIGURES
Figure (2.1): (A) Streptococcus thermophilus Cells Under Microscope.1000x
Magnification. (B) St Cells Observed By Scanning Electron Microscope. ............... 15
Figure (2.2): Stained Cells Of Streptococcus thermophilus Under A Light
Microscope. ................................................................................................................ 16
Figure (2.3): (A) Lactobacillus delbrueckii (Lb) Cells Under Microscope. 1000x
Magnification. (B) Lb Cells As Observed By Scanning Electron Microscopy. ........ 17
Figure (2.4): Stained Cells Of Lactobacillus delbrueckii subsp. bulgaricus Under A
Light Microscope ....................................................................................................... 18
Figure (2.5): An Approximation Of The Relative Concentration Between
L.bulgaricus And S.thermophilus From The Point Of Inoculum. ............................. 19
Figure (2.6): Scheme For Proto-Cooperation Between S.thermophilus and
L.bulgaricus ............................................................................................................... 21
Figure (2.7): Behaviour Of Single And Mixed Strain Yoghurt Cultures Propagated At
Different Temperature . ............................................................................................ 22
Figure (2.8): Behaviour Of Single And Mixed Strain Yoghurt Cultures Propagated At
40ºc. ........................................................................................................................... 22
Figure (2.9): Manufacturing Process Of Set & Stirred Yoghurt .............................. 25
Figure (2.10): Flow Chart Of Yoghurt .................................................................. 27
Figure (2.11): Scanning Electron Micrograph Illustrating (A) A Healthy
S.thermophilus Culture And (B) The Lysis Of Cells After Invasion With A Virulent
Bacteriophage. ........................................................................................................... 33
Figure (4.1): Microscopic Appearance Of Cocci And Bacilli Isolates ..................... 45
Figure (4.2): Partial Amplification Of The Methionine Biosynthesis Gene In Rural
Yoghurt Isolates. Lanes 1, 2 : Coccus B1 , Lanes 3,4 : Coccus B2, Lanes 5, 6: Coccus
B3, Lanes 7, 8 : Bacillus A1, Lanes 9, 10 : Bacillus A2, Nc: Negative Control, M: 100
Bp DNA Ladder. ........................................................................................................ 48
Figure (4.3): Amplification Of 16S rRNA Gene From Isolates That Amplified
Methionine Biosynthesis Gene Product. Lanes 1–4 : Lactobacilli A1,A2 Respectively,
Lanes 5-10: Streptococci B1,B2,B3 Respectively, M: 100 Bp DNA Ladder. .......... 49
Figure (4.4): Restriction Analysis Of 16S rRNA Gene With Eco RI. (1-4) Restriction
Analysis Of 16S rRNA Gene With Eco Ri For Lactobacilli A1,A2 Respectively (5-

XIII
10) Restriction Analysis Of 16S rRNA Gene With Eco RI For Streptococci B1,B2,B3
Respectively, M: 100 Bp DNA Ladder. .................................................................... 49
Figure (4.5): Changes In The Viable Counts Of The Starter L.Bulgaricus A1,
S.Thermophilus B1 & Combination Of L. Bulgaricus Plus S.Thermophilus (1: 1)
During Fermentation Of Raw Milk For 8h At 42ºc. .................................................. 51
Figure (4.6): Changes In The Viable Counts Of The Starter L.Bulgaricus A1,
S.Thermophilus B2 & Combination Of L.Bulgaricus Plus S.Thermophilus (1:1) During
Fermentation Of Raw Milk For 8h At 42ºc. .............................................................. 51
Figure (4.7): Changes In The Viable Counts Of The Starter L.Bulgaricus A1,
S.Thermophilus B3 & Combination Of L.Bulgaricus Plus S.Thermophilus (1:1) During
Fermentation Of Raw Milk For 8h At 42ºc. .............................................................. 52
Figure (4.8): Changes In The Viable Counts Of The Starter L.Bulgaricus A2,
S.Thermophilus B1 & Combination Of L.Bulgaricus Plus S.Thermophilus (1:1) During
Fermentation Of Raw Milk For 8h At 42ºc. .............................................................. 52
Figure (4.9): Changes In The Viable Counts Of The Starter L.Bulgaricus A2,
S.Thermophilus B2 & Combination Of L.Bulgaricus Plus S.Thermophilus (1:1) During
Fermentation Of Raw Milk For 8h At 42ºc. .............................................................. 53
Figure (4.10): Changes In The Viable Counts Of The Starter L.Bulgaricus A2,
S.Thermophilus B3 & Combination Of L.Bulgaricus Plus S.Thermophilus (1:1) During
Fermentation Of Raw Milk For 8h At 42ºc. .............................................................. 53
Figure (4.11): Changes In The PH Of The Starter L.Bulgaricus A1, S.Thermophilus
B1 & Combination Of L.Bulgaricus Plus S.Thermophilus (1:1) During Fermentation
Of Raw Milk At Different Times On 42ºc. ................................................................ 55
Figure (4.12): Changes In The PH Of The Starter L.Bulgaricus A1, S.Thermophilus
B2 & Combination Of L.Bulgaricus Plus S.Thermophilus (1:1) During Fermentation
Of Raw Milk At Different Times On 42ºc. ................................................................ 55
Figure (4.13): Changes In The PH Of The Starter L.Bulgaricus A1, S.Thermophilus
B3 & Combination Of L.Bulgaricus Plus S.Thermophilus (1:1) During Fermentation
Of Raw Milk At Different Times On 42ºc. ................................................................ 56
Figure (4.14): Changes In The PH Of The Starter L.Bulgaricus A2, S.Thermophilus
B1 & Combination Of L.Bulgaricus Plus S.Thermophilus (1:1) During Fermentation
Of Raw Milk At Different Times On 42ºc. ................................................................ 56

XIV
Figure (4.15): Changes In The PH Of The Starter L.Bulgaricus A2, S.Thermophilus
B2 & Combination Of L.Bulgaricus Plus S.Thermophilus (1:1) During Fermentation
Of Raw Milk At Different Times On 42ºc. ................................................................ 57
Figure (4.16): Changes In The PH Of The Starter L.Bulgaricus A2, S.Thermophilus
B3 & Combination Of L.Bulgaricus Plus S.Thermophilus (1:1) During Fermentation
Of Raw Milk At Different Times On 42ºc. ................................................................ 57
Figure (4.17): Changes In Total Titratable Acidity Of The Starter L.Bulgaricus A1,
S.Thermophilus B1 & Combination Of L.Bulgaricus Plus S.Thermophilus (1:1) During
Fermentation Of Raw Milk For 6h At 42ºc. .............................................................. 58
Figure (4.18): Changes In Total Titratable Acidity Of The Starter L.Bulgaricus A1,
S.Thermophilus B2 & Combination Of L.Bulgaricus Plus S.Thermophilus (1:1) During
Fermentation Of Raw Milk For 6h At 42ºc. .............................................................. 59
Figure (4.19): Changes In Total Titratable Acidity Of The Starterculture L.Bulgaricus
A1, S.Thermophilus B3 & Combination Of L.Bulgaricus Plus S.Thermophilus (1:1)
During Fermentation Of Raw Milk For 6h At 42ºc. .................................................. 59
Figure (4.20): Changes In Total Titratable Acidity Of The Starter Culture
L.Bulgaricus A2, S.Thermophilus B1 & Combination Of L.Bulgaricus Plus
S.Thermophilus (1:1) During Fermentation Of Raw Milk For 6h At 42ºc. ............... 60
Figure (4.21): Changes In Total Titratable Acidity Of The Starter L.Bulgaricus A2,
S.Thermophilus B2 & Combination Of L.Bulgaricus Plus S.Thermophilus (1:1) During
Fermentation Of Raw Milk For 6h At 42ºc. .............................................................. 60
Figure (4.22): Changes In Total Titratable Acidity Of The Starter L.Bulgaricus A2,
S.Thermophilus B3 & Combination Of L.Bulgaricus Plus S.Thermophilus (1:1) During
Fermentation Of Raw Milk For 6h At 42ºc. .............................................................. 61

XIV
List of Abbreviations
:Percentage % :microliter µl :micrometer µm :Amplified Ribosomal DNA Restriction Analysis ARDRA :Base pair Bp :colony forming unit CFU
:Celsius Co
:Deoxyribonucleic acid DNA
:Deoxynucleotide triphosphate dNTP
:Ethylene Diamide Tetra Acetic Acid EDTA
:Exopolysaccaride EPS
:Figure Fig.
:Gram G
:hour H
:Hexsose Monophosphate HMP
:International unit IU
:kilo base pair kbp
:Kilogram Kg
:liter L
:Lactobacillus L.
:Lactic acid bacteria LAB
:milligram Mg
:minute Min.
:Milliliter Ml
:de-man, Rogosa and sharpe medium MRS
:non-fat milk solids NFMS
:ribonucleic acid RNA
:round per minute rpm
:polymerase chain reaction PCR
:Streptococcus S.
:Solid- non fat SNF
:subspecies ssp.
:Tris Acetate EDTA TAE
:Tris Borate EDTA TBE :Tris-EDTA TE

1
Chapter 1
Introduction

2
Chapter 1
Introduction
1.1 Background and Context
Acidifying milk by microbial cells has been used since long time as natural protective
methods. These methods were tamed to make man-made fermented dairy products such as
yoghurt. However, with the onset of the 20th century, fermentation of milk was uncontrolled
process and the researchs were empiric and relied on trials and errors to improve it. (Caplice
& Fitzgerald, 1999).
Products of fermented milk have been enormously developed with the discovery of
lactic acid bacteria (LAB). Over the last fifty years, great efforts have been devoted to augment
our information regarding the physiology, biochemistry and molecular biology of LAB. Other
than to promote our understanding of the life of microbe, these efforts have allowed dairy
microbiologists and cheesemakers to pick out better strains and change for the better quality,
productivity, and safety of the final products (Amanze & Amanze, 2011).
Lactic acid bacteria characterizations have boosted the rational development of
mixtures of defined strains of bacteria, now widely known as starter cultures, which are
increasingly changing the undefined mixtures that were conventionally employed by the dairy
makers. This method causes it simpler to control the acid production and, somewhat, phage
invasions. Regrettably, it is also noticed that this method reduces the much desirable flavour
of fermented products that were created by undefined strains. Therefore, the study about LAB
cultures that yield desirable aroma and tastes still a major challenge for the dairy industry
(Vadeboncoeur & Moineau, 2004).
Yoghurt is uncomplicated ecological unit whose successful manufacturing depends
on correlation among two thermophilic LAB, Streptococcus thermophilus and Lactobacillus
delbreckii subsp. bulgaricus. This correlation among S. thermophilus and L. bulgaricus in milk
fermentation is known as proto-cooperation. Proto-cooperation is ground for making a
synergetic relationship among the two species (S.thermophilus and L.bulgaricus) and
collaborated metabolism with affirmative outcomes on the ultimate fermented product
(Angelov et al., 2009).

3
S.thermophilus and L.bulgaricus are thermophilic LAB species extensively used for
making yoghurt that wants high temperatures. The availability of genome for these species
will give the opportunety to apply the methods of molecular biology (Vadeboncoeur &
Moineau, 2004). The microbial compositions of traditional fermented milk products have been
subjected to microbiological assays and molecular researches at genus and species level
(Angelov et al., 2009).
1.2 Objectives
1.2.1 General objective:
o Isolation, Biochemical characterization and DNA identification of yoghurt starters
Streptococcus thermophilus & Lactobacillus delbrueckii ssp. bulgaricus.
1.2.2 Specific objective:
o Identification the strains of two Yoghurt Starters Streptococcus thermophilus &
Lactobacillus delbrueckii ssp. bulgaricus from cow’s yoghurt samples that produced
by traditional ways.
o Biochemical characterization and DNA identification of isolated yoghurt starter.
o Evaluation of the isolated starters on production of yoghurt.
1.3 Signification
According to our knowledge, this is the first thesis in Gaza sheds the light on the isolation,
biochemical and molecular characterization of S.thermophilus and L. bulgaricus cultures from
fermented milk transformed into home-production yoghurt supplied from geographical
regions of the Gaza strip. We, in this study seek to select future starters of S.thermophilus &
L. bulgaricus from traditional dairy products. It is a significant way to maintain the genetic
pool of the wild cultures.
1.4 Limitations
Even though the research has achieved its aims, there were some unavoidable drawbacks.
o Time limitation for compelling the project.
o Limitation of cost that DNA identification is required.
o Delayed arrival of some materials that the project is needed.

4
1.5 Overview of Thesis
Milk is one of diets for human nourishment and yoghurt is a style of fermented milk as a
result of lactic acid strains of bacteria, L.bulgaricus and S.thermophilus.
This study aims to isolate and identify yoghurt starters with several biochemicals,
physiological and molecular features, then assess these starters on yoghurt production.
Samples of cow’s traditional yoghurt were gathered from the households of 3 geographical
areas of the Gaza strip.
Then, blends of L.bulgaricus and S.thermophilus isolates were carried out in order to
produce yoghurt. Experiments of yoghurt production were conducted by combinations oftwo
L.bulgaricus and S.thermophilus isolates and compare them with single isolates. The isolates
approximately have the primary count of 107-108 cells/ml. The sugars in yoghurt of cow milk
are fermented via the bacterial cells into lactic acid which make the properties of curd; the acid
depresses to pH of the yoghurt.

5
Chapter 2
Literature Review

6
Chapter 2
Literature Review
2.1 What is Yoghurt?
Basically yoghurt is the product of useful bacterial cells which causes milk
fermentation and turn it into coagulated, sour diet that will persist freshness longer than milk
itself and that yoghurt possesses millions of bacterial cells that are received by the human gut.
Yoghurt can be soft and runny, or dense and firm. It is a ground of numerous vital nutrients,
comprising protein, calcium, potassium, phosphorus, vitamins B2 and B12, and act as a vehicle
for immunization (Moreno et al., 2012).
2.2 The History of Yoghurt
The word “yoghurt” is origionally a Turkish word “yoğurmak” which indicates to
thicken, clot, or curdle (Donovan, 2006). Nowadays it is pronounced yogurt, yoghurt, or
yogourt, with yogurt being the most prevalent American spelling (Fisberg & Machado, 2015).
It’s possible that the emergence of yoghurt was achieved by accident in Mesopotamia
nearly 5,000 BC, when milk-producing animals were firstly tamed. The milk was likely
maintained and moved in bags constructed from the guts of these animals, the digestive juices
and bacterial cells in the guts lining cause the thickening and sourness of the milk and keep it
for long periods of time (Weerathilake et al., 2014).
Or possibly, a herdsman put milk in a pots that accomodate some types of friendly bacteria,
during a warm times of the year. When he turns back to these pots to restore his milk, he
founded that the milk had curdled into a thick consistency with agreeably tangy flavor. Then
he founded that if he left curdled milk in the vessel and added more milk he could repeat the
process with similar results (Amanze & Amanze, 2011).
There are also some records of yoghurt being used as a cleaning product and a beauty
lotion nearly 2000 BC. The sourness of the yoghurt enables to remove dirtness and corrosion,
and also allows eliminating dead cells of skin and nourishing intact cells of skin.
Yoghurt was widespread in the Greeks and Romans empires where the Greeks were the earliest
to state it in written references in 100 BC, pointing out that the use of yoghurt via barbaric
societies (Fisberg & Machado, 2015). In the Bible (Book of Job), Abraham owed his long life
and fertility to yoghurt utilization, and there is reference to the “Land of Milk and Honey,”
which many historians have interpreted to be a reference to yoghurt (Anukam & Reid, 2007;
Batmanglij, 2007; Weerathilake et al., 2014).

7
The Turks were also the first to estimate the medicinal uses of yoghurt for a many of diseases
and symptoms, such as diarrhea, cramps and to reduce the discomfort of sunburned skin
(Donovan & Shamir, 2014).
It was not until the 20th century that researchers gave an enterpretation for the health
benefits that were related to yoghurt utilization. In 1905, Stamen Grigorov, a Bulgarian student
from medicine, was the first one who discovered Bacillus bulgaricus that is still used in
yoghurt making to now. Based on Grigorov’s findings, in 1909, Yllia Metchnikoff from the
Pasteur Institution in France indicated that lactobacilli in yoghurt were related with long life
in the Bulgarian rural people (Anukam & Reid, 2007).
At the starting of the 20th century, yoghurt became familiar due to its health benefits
and it was offered in pharmacies as a medication. Yoghurt realized commercial boom when
Isaac Carasso from Spain begun making yoghurt with jams. Then, Dannon (Danone in France)
founded by Daniel Carasso, Isaac Carasso’s son, after escaping from the Nazi invasion. In
1932, in France, the first laboratory and factory for yoghurt were opened while the first
laboratory and factory in the United States were opened in 1941. (Estrada et al., 2011).
For thousands of years, Yoghurt was a popular diet in the Middle East and it has been
a necessary of the Eastern European food. It’s currently eaten all over the world as a main
meal, a snack or an ingredient in many recipes. It has achieved extensive popularity in United
States in the lastest forty or fifty years, in keeping with general tendencies toward organic,
cultured and nutrient-dense foods (Amanze & Amanze, 2011).
.
Currently, yoghurt is made by addition two bacterial cultures to pasteurized
milk: Streptococcus thermophilus and Lactobacillus bulgaricus. These bacterial cultures
give the properties of yoghurt taste and aroma, and also improve digestion and absorption of
protein, calcium, and phosphorus naturally existing in milk (Donovan, 2006). Other bacterial
cultures such as Lactobacillus acidophilus, Lactobacillus casei and Bifidobacterium bifidus
are often supplemented to yoghurt to support the immune system, lower cholesterol levels,
and keep a healthy digestive system (Weerathilake et al., 2014).
2.3 Microorganisms used in starters for cultured dairy products:
The microbial cells that used as starters for manufacture of fermented dairy products
are divided into two groups, based on the optimal temperature at which they work. The LAB
incubated at 20–30Cº are termed mesophilic starters while those incubated at temperatures
above 35ºC are termed thermophilic starters. (Vedamuthu, 2006; Hutkins, 2008; Chandan,
2014).

8
Mesophilic starters include Lactococcus lactis subspecies and Leuconostoc species.
Lactococcus lactis ssp. lactis that ferment citrate is often referred to as “Lactococcus lactis
ssp. lactis biovar diacetylactis” in the writings. The other subspecies are Lactococcus lactis
ssp. cremoris and ssp. lactis. The Leuconostoc bacterial strains usually used in dairy
fermentations in connection with lactococci are Leuconostoc lactis and Leuconostoc
mesenteroides ssp. cremoris (Chandan, 2014).
Thermophilc starter employed for fermentation of milk involves Streptococcus
thermophilus and Lactobacillus spp. Amongst the lactobacilli, two subspecies of
Lactobacillus delbruechii ssp. bulgaricus and lactis are extensively used for making fermented
dairy products. Lactobacillus acidophilus, Lactobacillus helveticus and Lactobacillus casei
ssp. casei are additional lactobacilli used in fermented milk products in combining with other
particular microorganisms. (Chandan, 2014).
Yoghurt is created by thermophilic starters, which work in cooperation with each other. In
contrast, tart cream or cultured cream is created by mesophilic starters lactococci and
leuconostocs. (Kilara & Chandan, 2013; Chandan, 2014).
Microorganisms and some of the physiological and biochemical features of starters
are brief in Tables 2.1, 2.2 and 2.3.
9
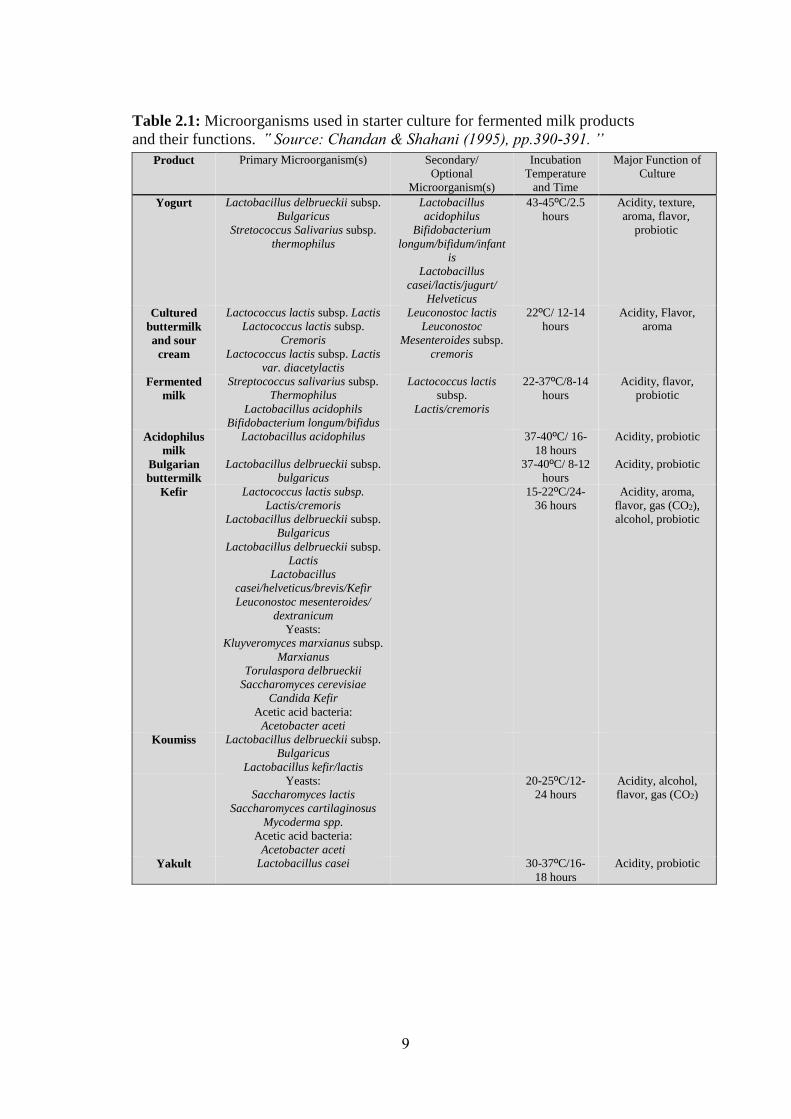
Table 2.1: Microorganisms used in starter culture for fermented milk products
and their functions. ՙՙ Source: Chandan & Shahani (1995), pp.390-391. ՚՚
Major Function of
Culture
Incubation
Temperature
and Time
Secondary/
Optional
Microorganism(s)
Primary Microorganism(s) Product
Acidity, texture,
aroma, flavor,
probiotic
43-45ºC/2.5
hours
Lactobacillus
acidophilus
Bifidobacterium
longum/bifidum/infant
is
Lactobacillus
casei/lactis/jugurt/
Helveticus
Lactobacillus delbrueckii subsp.
Bulgaricus
Stretococcus Salivarius subsp.
thermophilus
Yogurt
Acidity, Flavor,
aroma
22ºC/ 12-14
hours
Leuconostoc lactis
Leuconostoc
Mesenteroides subsp.
cremoris
Lactococcus lactis subsp. Lactis
Lactococcus lactis subsp.
Cremoris Lactococcus lactis subsp. Lactis
var. diacetylactis
Cultured
buttermilk
and sour
cream
Acidity, flavor,
probiotic
22-37ºC/8-14
hours
Lactococcus lactis
subsp.
Lactis/cremoris
Streptococcus salivarius subsp.
Thermophilus
Lactobacillus acidophils
Bifidobacterium longum/bifidus
Fermented
milk
Acidity, probiotic 37-40ºC/ 16-
18 hours
Lactobacillus acidophilus Acidophilus
milk
Acidity, probiotic 37-40ºC/ 8-12
hours
Lactobacillus delbrueckii subsp.
bulgaricus Bulgarian
buttermilk
Acidity, aroma,
flavor, gas (CO2),
alcohol, probiotic
15-22ºC/24-
36 hours
Lactococcus lactis subsp.
Lactis/cremoris
Lactobacillus delbrueckii subsp.
Bulgaricus
Lactobacillus delbrueckii subsp.
Lactis
Lactobacillus
casei/helveticus/brevis/Kefir
Leuconostoc mesenteroides/
dextranicum
Yeasts:
Kluyveromyces marxianus subsp.
Marxianus
Torulaspora delbrueckii
Saccharomyces cerevisiae
Candida Kefir
Acetic acid bacteria:
Acetobacter aceti
Kefir
Lactobacillus delbrueckii subsp.
Bulgaricus
Lactobacillus kefir/lactis
Koumiss
Acidity, alcohol,
flavor, gas (CO2)
20-25ºC/12-
24 hours
Yeasts:
Saccharomyces lactis
Saccharomyces cartilaginosus
Mycoderma spp.
Acetic acid bacteria:
Acetobacter aceti
Acidity, probiotic 30-37ºC/16-
18 hours
Lactobacillus casei Yakult
10
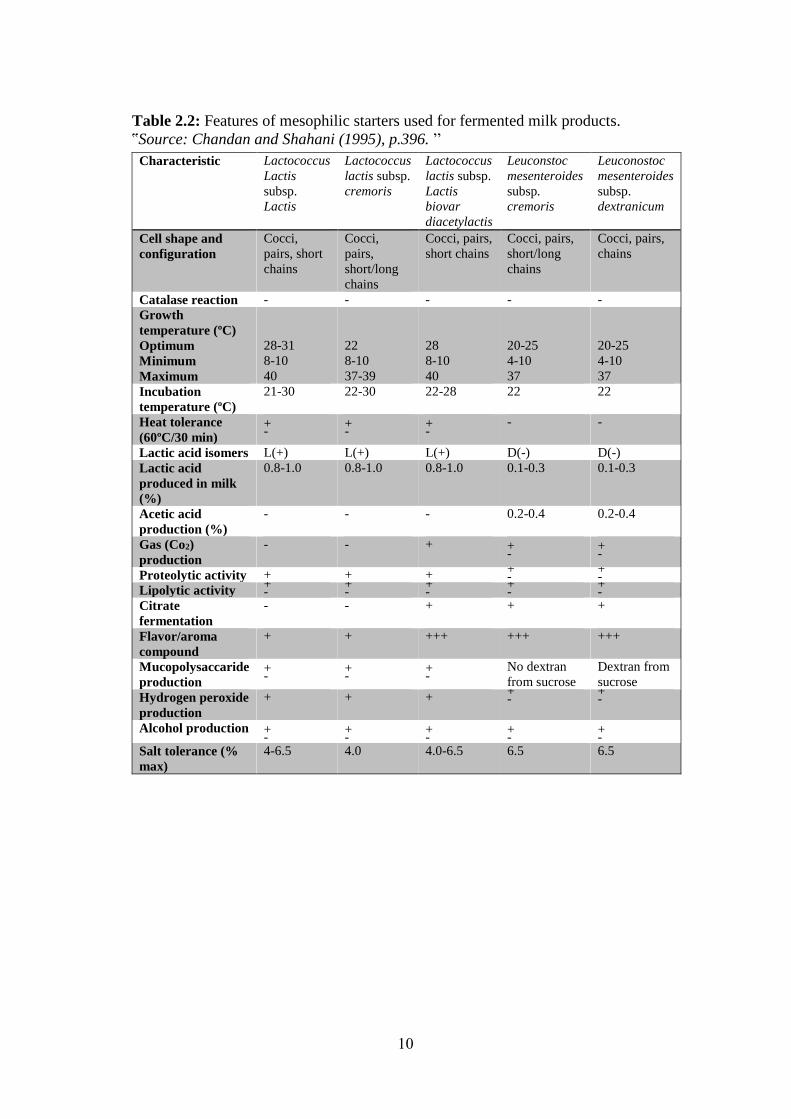
Table 2.2: Features of mesophilic starters used for fermented milk products.
ՙՙSource: Chandan and Shahani (1995), p.396. ՚՚
Leuconostoc
mesenteroides
subsp.
dextranicum
Leuconstoc
mesenteroides
subsp.
cremoris
Lactococcus
lactis subsp.
Lactis
biovar
diacetylactis
Lactococcus
lactis subsp.
cremoris
Lactococcus
Lactis
subsp.
Lactis
Characteristic
Cocci, pairs,
chains
Cocci, pairs,
short/long
chains
Cocci, pairs,
short chains
Cocci,
pairs,
short/long
chains
Cocci,
pairs, short
chains
Cell shape and
configuration
- - - - - Catalase reaction
20-25
4-10
37
20-25
4-10
37
28
8-10
40
22
8-10
37-39
28-31
8-10
40
Growth
temperature (ºC)
Optimum
Minimum
Maximum
22 22 22-28 22-30 21-30 Incubation
temperature (ºC)
- - + -
+ -
+ -
Heat tolerance
(60ºC/30 min)
D(-) D(-) L(+) L(+) L(+) Lactic acid isomers
0.1-0.3 0.1-0.3 0.8-1.0 0.8-1.0 0.8-1.0 Lactic acid
produced in milk
(%)
0.2-0.4 0.2-0.4 - - - Acetic acid
production (%) + -
+ -
+ - - Gas (Co2)
production + -
+ - + + + Proteolytic activity
+ -
+ -
+ -
+ -
+ - Lipolytic activity
+ + + - - Citrate
fermentation
+++ +++ +++ + + Flavor/aroma
compound
Dextran from
sucrose
No dextran
from sucrose
+ -
+ -
+ -
Mucopolysaccaride
production + -
+ - + + + Hydrogen peroxide
production + -
+ -
+ -
+ -
+ -
Alcohol production
6.5 6.5 4.0-6.5 4.0 4-6.5 Salt tolerance (%
max)
11
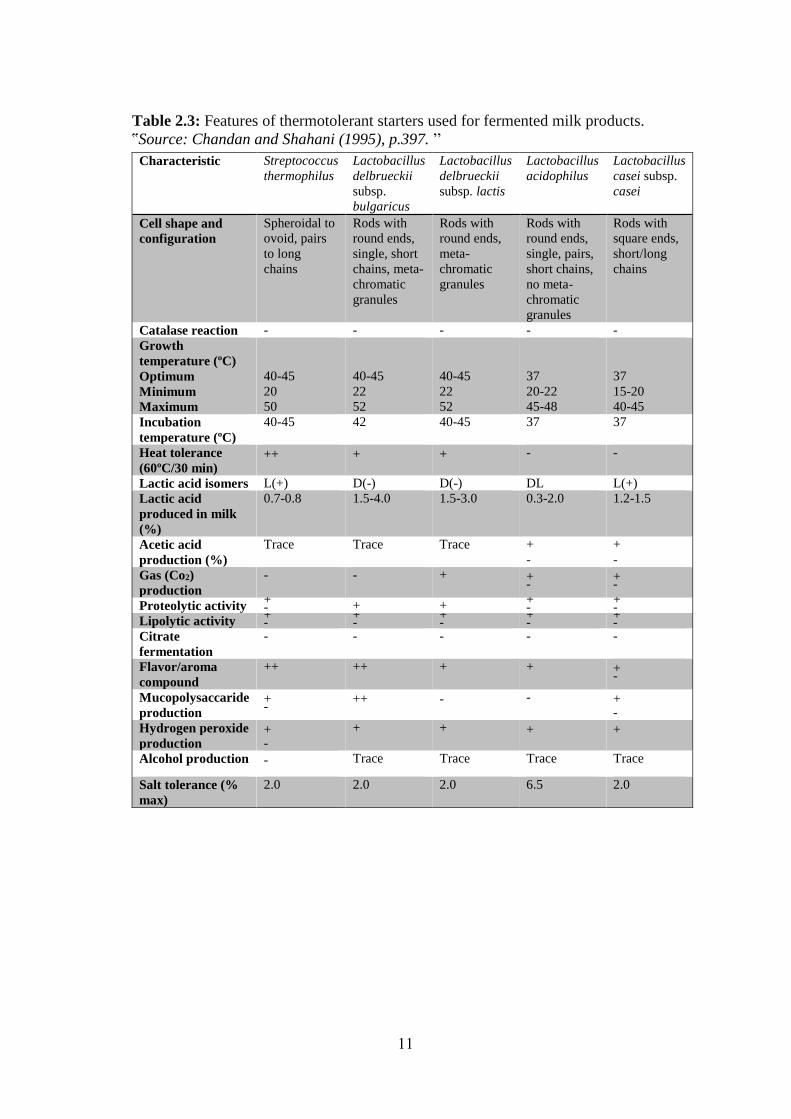
Table 2.3: Features of thermotolerant starters used for fermented milk products.
ՙՙSource: Chandan and Shahani (1995), p.397. ՚՚
Lactobacillus
casei subsp.
casei
Lactobacillus
acidophilus Lactobacillus
delbrueckii
subsp. lactis
Lactobacillus
delbrueckii
subsp.
bulgaricus
Streptococcus
thermophilus Characteristic
Rods with
square ends,
short/long
chains
Rods with
round ends,
single, pairs,
short chains,
no meta-
chromatic
granules
Rods with
round ends,
meta-
chromatic
granules
Rods with
round ends,
single, short
chains, meta-
chromatic
granules
Spheroidal to
ovoid, pairs
to long
chains
Cell shape and
configuration
- - - - - Catalase reaction
37
15-20
40-45
37
20-22
45-48
40-45
22
52
40-45
22
52
40-45
20
50
Growth
temperature (ºC)
Optimum
Minimum
Maximum
37 37 40-45 42 40-45 Incubation
temperature (ºC)
- - +
+
++ Heat tolerance
(60ºC/30 min)
L(+) DL D(-) D(-) L(+) Lactic acid isomers
1.2-1.5 0.3-2.0 1.5-3.0 1.5-4.0 0.7-0.8 Lactic acid
produced in milk
(%)
+
-
+
-
Trace Trace Trace Acetic acid
production (%) + -
+ -
+ - - Gas (Co2)
production + -
+ - + +
+ - Proteolytic activity
+ -
+ -
+ -
+ -
+ - Lipolytic activity
- - - - - Citrate
fermentation + -
+ + ++ ++ Flavor/aroma
compound + -
- -
++
+ -
Mucopolysaccaride
production +
+ + +
+ -
Hydrogen peroxide
production
Trace Trace Trace Trace - Alcohol production
2.0 6.5 2.0 2.0 2.0
Salt tolerance (%
max)

12
2.4 Yoghurt Starter Cultures:
Starter cultures or “starters” for short are food-grade that involves carefully chosen
microbial cells added to milk to make desired changes that result in the production of a specific
fermented dairy product with some desired features (Gandhi, 2006, Doyle & Meng, 2006).
One type of starters is called undefined "artisanal" cultures. It contains mixtures of
starter cells. The actual identities of the cells existing in a blended culture are often not known,
and the types of unique species may not have been characterised biochemically or
microbiologically. The ratio of different cells in a blended culture may be a variable from one
product to another. Hence, the chief disadvantage of undefined cultures is that they may
produce products of changeful quality. Additionally, fermentation ratios may differ from day
to day, influencing production programmes. In large facilities of production where accurate
programmes are necessary and regular quality of product is predicted so, undefined starters
can not be used. Defined starters as an alternative have become prevalent (Durso and Hutkins
2003; Erkuş, 2007). Defined starters have known physiological, biochemical and molecular
characterization strains, which are used as single or mix cultures. Most of the defined strains
have been isolated from wild or artisanal cultures (Hebert et al., 2000; Erkuş, 2007). They are
characterized and screened for the desired characters. Hence they give constant quality, and
softness to improve the productivity, quality and safety (Erkuş, 2007).
The whole fermented dairy product making is depends on the viability of the starter cells.
Starters not only initiate but also carry through each change needed to achieve the required
body, texture and flavour in the fermented dairy product. Moreover, starters play a preserving
role in repressing the organisms of rot, thus it raises the shelf-life. Additional vital roles related
to their protecting purpose in delaying or stopping pathogen organisms, and to the creation of
toxins in the finished fermented dairy product. Shortly, starters determine the shelf-life and
the security of fermented dairy products. In probiotic products, the supplemented cultures
provide health-promoting properties to the consumer (Chandan & O’Rell, 2006).
As mentioned, starters are composed of alive entities. Alive organisms necessitate
suitable environments to boom and achieve their tasks. Environmental conditions involve
optimal temperature ranges, suitable nutrition and pH range, lack of lethal elements or
byproducts and cautious handling procedures (Vedamuthu, 2006).
The two most starters frequently used in yoghurt making are now categorized to
Lactobacillus delbrueckii ssp. bulgaricus and Streptococcus thermophilus ssp. thermophilus,
usually shortened as L.bulgaricus and S.thermophilus, respectively. Both are Gram-positive,

13
facultative anaerobic, non-motile and non spore-forming cells. Negative for endospore and
capsule staining, Negative with catalase and oxidase (El Bashiti, 2010).
The yoghurt culture contains S.thermophilus and L.bulgaricus. Moreover, a majority
of the yoghurt sold contains optional bacteria that have been shown to possess probiotic
features, including bifidobacteria, Lactobacillus acidophilus, Lactobacillus casei and other
Lactobacilli, which are famous as “probiotic bacteria”. Some yoghurt makers combine them
after fermentation of yoghurt, while others co-culture them with yoghurt starters (Chandan &
O’Rell, 2006).
S.thermophilus and L.bulgaricus are fairly well-matched with one another and are
synergetic for growth in milk medium. On the other hand, the optional organisms not
necessarily display compatibility with S.thermophilus and L.bulgaricus. Wise selection of
yoghurt starters and the optional organisms is basic to ensure the viability of all the organisms
in starters' components.
However, product appearances especially flavour may be considerably changed from old-style
yoghurt flavour when yoghurt is co-cultured with optional cells, particularly bifidobacteria
(Schmidt, 2004).
Generally, the yoghurt starters are responsible for the flavour and aroma creation of
yoghurt through the production of acetaldehyde, diacetyl and acetic acid through the
fermentation course. Lactic acid, non-volatile substance, donates the acidic and refreshing
flavour of yoghurt, while the volatile by-products donate the pleasant and aroma characteristic.
Regarding volatile flavor substances, acetaldehyde nearly about 90%. However,
bifidobacteria create more acetic acid than lactic acid. Hence, if they are used in the culture
make-up, the total flavor profile will develop as a result of higher acetic-acid content (Erkuş,
2007).
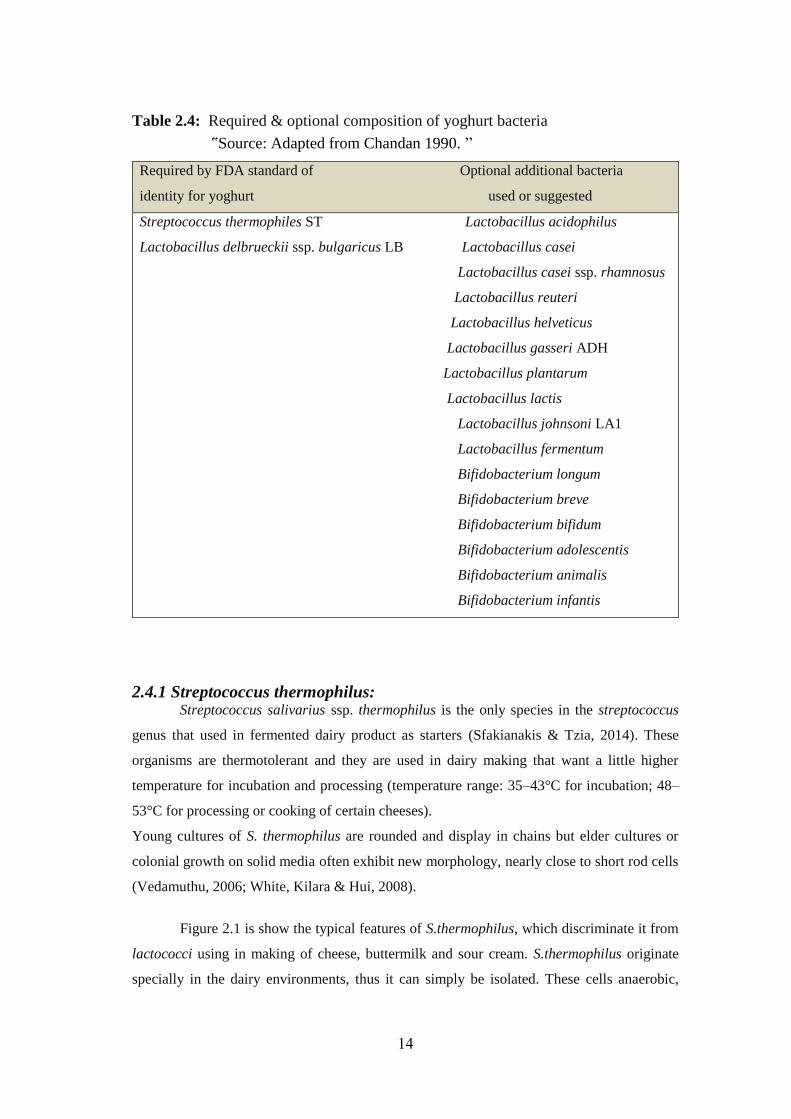
The compositions of yoghurt starters are shown in Table 2.4; also presented some
supplementary organisms found in yoghurt or yoghurt-like products marketed in various parts
of the world (Schmidt, 2004).
14
Table 2.4: Required & optional composition of yoghurt bacteria
ՙՙSource: Adapted from Chandan 1990. ՚՚
Required by FDA standard of Optional additional bacteria
identity for yoghurt used or suggested
Streptococcus thermophiles ST Lactobacillus acidophilus
Lactobacillus delbrueckii ssp. bulgaricus LB Lactobacillus casei
Lactobacillus casei ssp. rhamnosus
Lactobacillus reuteri
Lactobacillus helveticus
Lactobacillus gasseri ADH
Lactobacillus plantarum
Lactobacillus lactis
Lactobacillus johnsoni LA1
Lactobacillus fermentum
Bifidobacterium longum
Bifidobacterium breve
Bifidobacterium bifidum
Bifidobacterium adolescentis
Bifidobacterium animalis
Bifidobacterium infantis
2.4.1 Streptococcus thermophilus: Streptococcus salivarius ssp. thermophilus is the only species in the streptococcus
genus that used in fermented dairy product as starters (Sfakianakis & Tzia, 2014). These
organisms are thermotolerant and they are used in dairy making that want a little higher
temperature for incubation and processing (temperature range: 35–43°C for incubation; 48–
53°C for processing or cooking of certain cheeses).
Young cultures of S. thermophilus are rounded and display in chains but elder cultures or
colonial growth on solid media often exhibit new morphology, nearly close to short rod cells
(Vedamuthu, 2006; White, Kilara & Hui, 2008).
Figure 2.1 is show the typical features of S.thermophilus, which discriminate it from
lactococci using in making of cheese, buttermilk and sour cream. S.thermophilus originate
specially in the dairy environments, thus it can simply be isolated. These cells anaerobic,

15
nonmotile, Gram-positive and catalase-negative cells with round/ovoid forms of 0.7–0.9 μm
diameter (Erkuş, 2007). It may live at 60°C for 30min (Chandan and Nauth, 2012). It doesn't
survive at 10°C. Even though the best growing temperature for S.thermophilus at 37°C, these
strains grow well in co-operation with L.bulgaricus at 43°C (yoghurt incubation temperature).
In the course of yoghurt fermentation, S.thermophilus initially wants for a nitrogen source
which comes from the free amino acids found in the milk medium.As fermentation proceeds;
the peptides created via L.bulgaricus are digested by the peptidases of S.thermophilus to
produce free amino acids to meet its nutritive requirements (Chandan & O’Rell, 2006).
Figure 2.1: (a) Streptococcus thermophilus cells under microscope.1000x magnification. (b)
ST cells observed by scanning electron microscope.
Milk is a good media for S.thermophilus development. It can ferment glucose,
fructose, mannose, sucrose and lactose. Lactose in milk is moved through the cell membranes
of starters with the aid of galactoside permease enzymes which located within the membrane.
Then the lactose in the cells is digested by lactase or β -galactsidase enzyme. S. thermophilus
creates significant levels of lactase which stimulates the hydrolysis of lactose to glucose and
galactose. Glucose is altered to pyruvic acid by Embden–Meyerhof pathway. Pyruvic acid is
break down to lactic acid by lactic dehydrogenase. In most cultures, glucose is readily
consumed in milk medium whereas lactic acid and galactose piling up. Some cultures can
consume galactose; these show activity of galactokinase enzyme, changing galactose to
galactose-1-phosphate which is additional changed to glucose-1-phosphate or galactose-6-
phosphate and moreover metabolized to lactic acid. The produced lactic acid is L (+) lactic
acid. S.thermophilus is repressed by accumulative levels of lactic acid. At around 1.0%
concentration (pH 4.3) it's growing is stopped and cell numbers reach steadiness. At this phase

16
the fermented mass presentations S.thermophilus counts of 107-108 CFU/g (Chandan &
O’Rell, 2006).
The lactase activity of S.thermophilus has a physiological significance in aiding the
breakdown of lactose in the human gut subsequent intake of yoghurt by lactose-intolerant
persons. When milk incubated at 43°C, numerous strains seem round, happening in diploid or
in elongated chains of 10–20 cells. Most cells seem as diplococci. At high sourness levels in
milk, if the cells are elderly or if grown on solid media, S. thermophilus may display elongated
chains. When plated on solid media, S. thermophilus seems as pinpoint colonies. The
appearance of abnormal shapes gotten from liquid media is an index of stress conditions
affecting the organism, especially bacteriophage invasion and inhibitors (sanitizers,
antibiotics, cleaning compounds, etc.) in the growing medium. S. thermophilus is oversensitive
to materials of repression, especially antibiotics. It's easily repressed by 0.005 IU penicillin/ml
of milk. It must be noticed that S. thermophilus is more often invaded by phages than
L.bulgaricus (Chandan & O’Rell, 2006).
Figure 2.2: Stained cells of Streptococcus thermophilus under a light microscope.
Additionally, EPS cab produced by S. thermophilus in fermentation of yoghurt to
achieve a gelatinous form and soft consistency. The physiology and genetics of EPSs creation
by coccus are nowadays well understood. More details on the inheritances of S.thermophilus
are available in the collected works.These polymers may work as texturizers, viscosifiers, or
emulsifying agents to enhance the texture and consistency of dairy products (Vaningelgem et
al., 2004). EPSs that are formed by lactic acid bacteria have been shown to play a significant
role in the avoidance of whey separation (syneresis), a common challenging in manufacture
of yoghurt (Grobben et al., 1996; Zhang et al., 2011). In situ creation of EPSs by yoghurt
starters have been recommended to be used as a substitute to the adding of stabilizers, e.g.
animal hydrocolloids (gelatin and casein) or chemically modified plant carbohydrates (starch,
pectin) (Vaningelgem et al., 2004; Zhang et al, 2011).

17
2.4.2 Lactobacillus delbrueckii subsp. bulgaricus:
These bacteria firstly described in 1919 by Orla Jensen as thermobacterium
bulgaricum. Depended on molecular researchs, four subspecies of L.delbrueckii are
categorized as bulgaricus, leichmannii, lactis and delbrueckii. Lactobacilli generally are rod-
shaped as shown in figure 2.3. Some occur as long, slender, straight rods; others are curved;
others still are short, almost coccoid rods. A few display pleomorphic cells (Vedamuthu,
2006).
Figure 2.3: (a) Lactobacillus delbrueckii (LB) cells under microscope. 1000X
magnification. (b) LB cells as observed by scanning electron microscopy.
Lactobacilli do not ferment pentoses but ferment hexoses via HMP to lactic acid.
L.bulgaricus is positive with Gram stain, negative with catalase test and it nonmotile
bacterium. These cells are an anaerobic/aerotolerant homofermentative that yields D (−) lactic
acid and some H2O2. It may yield big amounts of lactic acid (up to 1.8%) but the strains which
produce mild acid are chosen for yoghurt making. Similar S.thermophilus, L.bulgaricus gives
lactase to breakdown lactose into glucose and galactose and glucose will metabolize to lactic
acid whereas galactose will gather in the growing media (White, Kilara & Hui, 2008). The
cells of L.bulgaricus look like slender rods with curved edges under microscope. These cells
appear individually or in the form of chains 3 or 4 short rods (0.5–0.8; 2.0–9.0 μm). In young
and vigorous conditions, the cells exhibit commonly single or in pairs. Under microscope,
newer L.bulgaricus cells don't display volutin grains (metachromatic). With increasing age
(20–24h), these cells will extend and the volutin grains become more obvious. Abundant
grains occur in nutritious stress in the rods. L.bulgaricus cells have a greater resistance to
antibiotics than S.thermophilus cells but repressed at 0.3–0.6 IU penicillin/ml of milk
(Vedamuthu, 2006).L.bulgaricus is an oversensetive cells. Absence of certain critical nutrients

18
and minerals in the growth media impacts on the integrity of these bacterial cells and these
cells display unusual morphology under nutritive stress. Furthermore, the preparations of rod
cultures commercially are enormously challenging task due to oversensitivity for
environmental and nutritional conditions (temperature- pH-controlling, rejection of air)
associated with harvesting and preservation processes (Vedamuthu, 2006).
The ideal growing temperature of L.bulgaricus is 45°C, but 42–43°C is utilized to
accommodate the lower ideal growing temperature of S.thermophilus in yoghurt making.
Glucose, lactose, fructose are consumed via L.bulgaricus. Also, galactose is consumed to yield
as high as D (−) lactic acid 1.8% in some cultures. It’s resists low pH much better than
S.thermophilus. Not like S.thermophilus, L.bulgaricus can convert casein (β -casein, specially)
to peptides, via proteinase enzymes bounded in cell walls. L.bulgaricus must depend on
S.thermophilus which has an active peptidase action to convert the resulting peptides to free
amino acids (White, Kilara & Hui, 2008).
Figure 2.4: Stained cells of Lactobacillus delbrueckii ssp. bulgaricus under a light
microscope.
2.5 Protocooperation between S.thermophilus & L.bulgaricus
Lactic acid bacteria are oversensitive cells and because of milk scarcity in necessary
nutriments, their grown is often limited in it. Thus successful fermentation of milk depends
mostly on the interaction between S.thermophilus and L.bulgaricus. Because both types of
cells are able to propagate alone in milk, this indirect affirmative relation is called
protocooperation. This relation often has a useful influence on growing of cells and on the
creation of lactic acid and aroma compounds (Courtin & Rul, 2004; Erkmen & Bozoglu,
2016). Some of the effectors of this relationship have been well-known and result from the
metabolic actions of the two bacteria (Angelov et al., 2009).
The grouping of the two bacterial partners is called “rod–coccus”. Rod–coccus mixtures show
a synergistic evolution response in milk. Strains of the bacterial cells should be combined with
precaution after carry out trial with numerous mixes to make the most of the synergistic effect
(Vedamuthu, 2006).
19
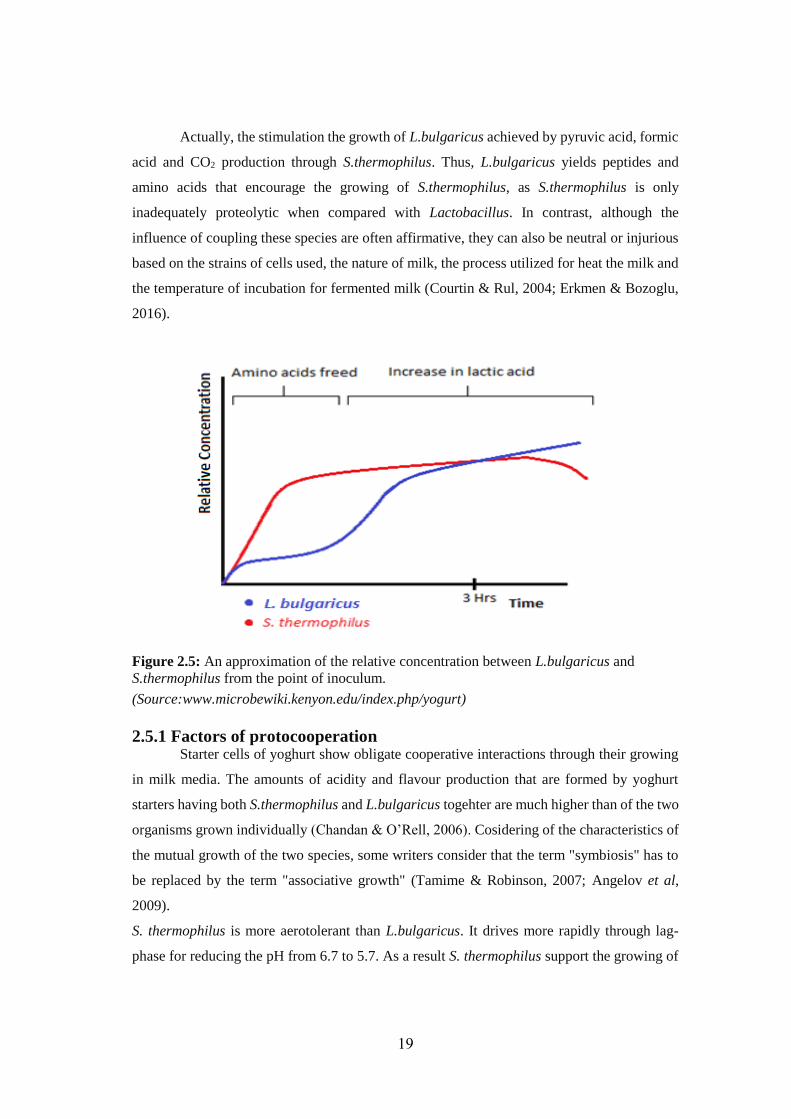
Actually, the stimulation the growth of L.bulgaricus achieved by pyruvic acid, formic
acid and CO2 production through S.thermophilus. Thus, L.bulgaricus yields peptides and
amino acids that encourage the growing of S.thermophilus, as S.thermophilus is only
inadequately proteolytic when compared with Lactobacillus. In contrast, although the
influence of coupling these species are often affirmative, they can also be neutral or injurious
based on the strains of cells used, the nature of milk, the process utilized for heat the milk and
the temperature of incubation for fermented milk (Courtin & Rul, 2004; Erkmen & Bozoglu,
2016).
Figure 2.5: An approximation of the relative concentration between L.bulgaricus and
S.thermophilus from the point of inoculum.
(Source:www.microbewiki.kenyon.edu/index.php/yogurt)
2.5.1 Factors of protocooperation Starter cells of yoghurt show obligate cooperative interactions through their growing
in milk media. The amounts of acidity and flavour production that are formed by yoghurt
starters having both S.thermophilus and L.bulgaricus togehter are much higher than of the two
organisms grown individually (Chandan & O’Rell, 2006). Cosidering of the characteristics of
the mutual growth of the two species, some writers consider that the term "symbiosis" has to
be replaced by the term "associative growth" (Tamime & Robinson, 2007; Angelov et al,
2009).
S. thermophilus is more aerotolerant than L.bulgaricus. It drives more rapidly through lag-
phase for reducing the pH from 6.7 to 5.7. As a result S. thermophilus support the growing of

20
L.bulgaricus primarily by generating lactic acid and formic acid (Kosikowski & Mistry, 1977;
Tamime & Robinson, 2007, Angelov et al., 2009).
Throughout the incubation at early stages, S.thermophilus propagates more rapidly
and be more numerous than L.bulgaricus by 3–4 to 1. However, at pH 5.0 in the later stages,
S. thermophilus growing slow down owing to contrary effect of acid development and the
number of L.bulgaricus approaches gradually to the population of S.thermophilus. Thus,
accomplishing of the production of acid happens in the initial stage of incubation in the main
by S. thermophilus, and in the following stage basically by L.bulgaricus (Chandan & O’Rell,
2006). S.thermophilus assimilates O2 in the milk more rapidly thus forming favourable
environments for growth of L.bulgaricus (Kosikowski & Mistry, 1977; Tamime & Robinson,
2007, Angelov et al., 2009).
Corresponding to some authors, S. thermophilus yields large amount of CO2 which is
not a product of lactose fermentation because LAB forming that kind are homo-fermentative
(Driessen et al, 1982; Angelov et al., 2009). Creation of CO2 is due to the production of urease
enzyme by S.thermophilus, which breakdowns milk urea to CO2 and NH3, ammonia acts as a
weak buffer (Chandan & O’Rell, 2006; Angelov et al., 2009). Collected works are comprised
also the theory which states that the CO2 is produced by S.thermophilus during the hydrolysis
of urea in the milk encourages the growth of L.bulgaricus when grown together (Driessen et
al., 1982; Louaileche et al., 1993; Louailèche et al.,1996; Tamime & Robinson, 2007).
S.thermophilus promotes the growth of L.bulgaricus in milk in numerous ways: a decline of
the pH, change of the redox potential, consuming of dissolved O2 and forming CO2, as
presented in fig.2.6; S.thermophilus generates the required anaerobic environments.

21
Figure 2.6: Scheme for proto-cooperation between S.thermophilus & L.bulgaricus
In the milk, the conjoint growing of two forms is specified by the metabolism of them.
For example, the lactose turns to glucose and galactose and those are used in different ways
by these two species of the mixed culture. The streptococcus and lactobacillus digested the
glucose and turn this into D (-) lactic acid. Not like glucose, which is assimilated by the two
species, the galactose consumed by S.thermophilus via Leloir' metabolic pathway and turning
this into lactic acid and CO2. This confirms the anaerobic conditions for lactobacillus growth
(Angelov et al., 2009).
As it's known that some strains of S.thermophilus don't have urease enzyme and can't
produce carbon dioxide. Hence the Leloir' metabolic pathway is a probable way to realize the
anaerobic environments, which is considered as a good clarification of protocooperation in the
lack of urease enzymes. (Angelov et al., 2009). L.bulgaricus yields short peptides and amino
acids that promote the evolution of S.thermophilus (Tammam et al., 2000). Histidine,
threonine, and Valine are the most important amino acids required for the conjoint growth of
both strains (Tamime & Robinson, 2007). S.thermophilus often has low proteolytic activity
due to shortage of protease in the cell wall for these strains. So, their growing is limited because
of deficiency of an adequate amount of amino acids and peptides in the milk wanted for these
starins. In contrast, L.bulgaricus is able to produce amino acids and small peptides from the
casein of milk because of these strains have protease activity in the cell walls. This conjoint
growing to both species leads to more formation of effective acid, more concentration of final
cells and it in addition rises specific growing speed comparing with monocultures from both
species (Driessen et al, 1982; Rajagopal & Sandine, 1990, Chandan & O’Rell, 2006; Angelov
et al., 2009).
22
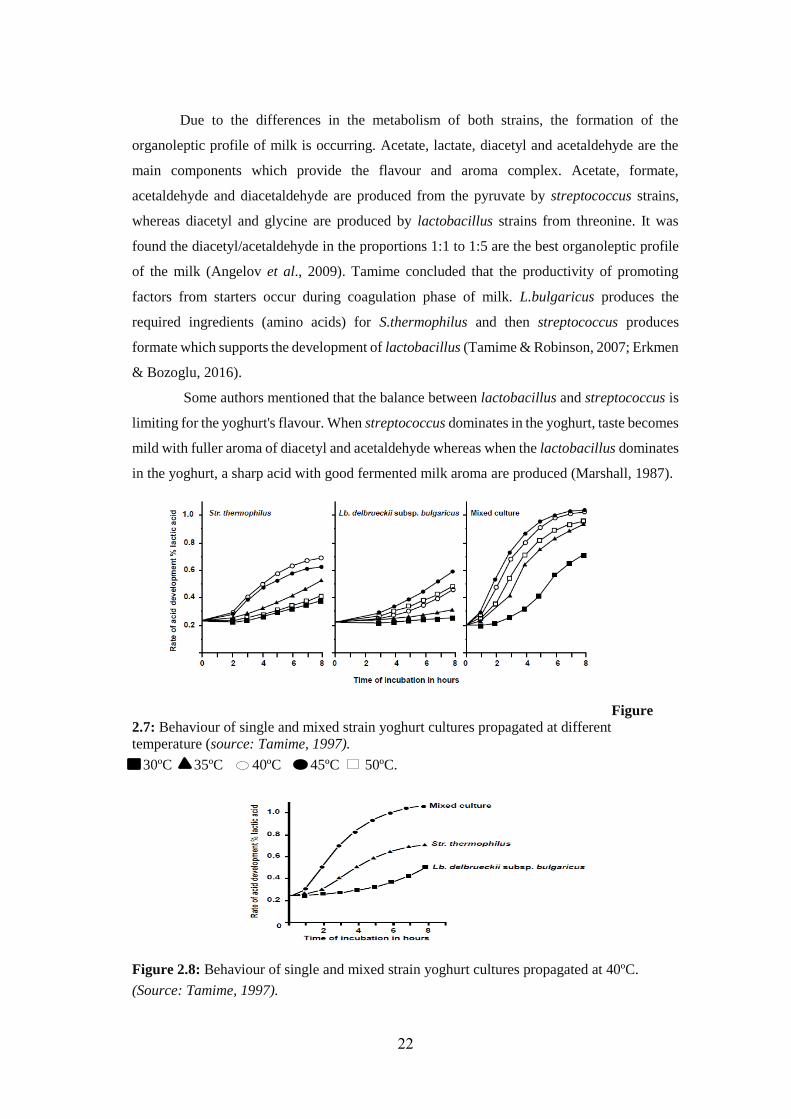
Due to the differences in the metabolism of both strains, the formation of the
organoleptic profile of milk is occurring. Acetate, lactate, diacetyl and acetaldehyde are the
main components which provide the flavour and aroma complex. Acetate, formate,
acetaldehyde and diacetaldehyde are produced from the pyruvate by streptococcus strains,
whereas diacetyl and glycine are produced by lactobacillus strains from threonine. It was
found the diacetyl/acetaldehyde in the proportions 1:1 to 1:5 are the best organoleptic profile
of the milk (Angelov et al., 2009). Tamime concluded that the productivity of promoting
factors from starters occur during coagulation phase of milk. L.bulgaricus produces the
required ingredients (amino acids) for S.thermophilus and then streptococcus produces
formate which supports the development of lactobacillus (Tamime & Robinson, 2007; Erkmen
& Bozoglu, 2016).
Some authors mentioned that the balance between lactobacillus and streptococcus is
limiting for the yoghurt's flavour. When streptococcus dominates in the yoghurt, taste becomes
mild with fuller aroma of diacetyl and acetaldehyde whereas when the lactobacillus dominates
in the yoghurt, a sharp acid with good fermented milk aroma are produced (Marshall, 1987).
Figure
2.7: Behaviour of single and mixed strain yoghurt cultures propagated at different
temperature (source: Tamime, 1997).
30ºC 35ºC 40ºC 45ºC 50ºC.
Figure 2.8: Behaviour of single and mixed strain yoghurt cultures propagated at 40ºC. (Source: Tamime, 1997).
5

23
2.7 classification of LAB based on by-products of suger
Lactic acid bacteria may be classed depending on their by-products of carbohydrates
fermentation as homofermentative or heterofermentative.
2.7.1 Homofermentative lactic acid bacteria
Lactic acid is the main by-product which produces from fermentation of glucose.
Lactococcus ssp. are considered one of examples of homofermentative lactic acid bacteria
which are used in applications of dairy starters where the quick formation of lactic acid and
decline in the pH are desirable. L.bulgaricus, L.acidophilus, Streptococcus thermophilus and
thermophilic cultures that might be used in cheese manufacturing (e.g., L.helveticus) are other
examples of homofermentative LAB. Other Streptococcus spp., Enterococcus, Pediococcus
and Aerococcus are additional homofermentative cocci that might be originate in milk and
dairy products, but they are seldom used as starter cultures.
2.7.2 Heterofermentative lactic acid bacteria
Lactic acid, ethanol/acetic acid and carbon dioxide are by-products which produce
from fermentation of glucose. In general, the detection of gas (e.g. CO2) is used to examine
the heterofermentative cultures. Heterofermentative lactic acid bacteria are seldom used as
dairy starters. If significant numbers are allowed to be grown, they can cause faults related to
their acidity and production of CO2, such as slots in hard cheeses or swollen the packaging of
other dairy products. Leuconostoc spp. (Gram-positive cocci) and L.brevis, L.fermentum, L.
reuteri (Gram-positive rods) are considered examples of heterofermentative lactic acid
bacteria.Other Lactobacillus species comprise L.plantarum, L.casei and L.curvatus are
considered facultatively heterofermentative, this mean they will make CO2 and other by-
products only under certain environments or from limited substrates.
2.7.3 Other Gas Producing Pathways:
Other substrates comprising citrate, gluconate and certain amino acids are used to
produce gas by other lactic acid bacteria. Some citrate-fermentors are used in certain dairy
products to make flavour as diacetyl. Manufacturing products such as buttermilk, sour cream
and cultured butter are often occurring by Leuconostoc mesenteroides ssp. cremoris and
Lactococcus lactis ssp. lactis biovar diacetylactis. These organisms can also happen as wild
contaminants causing defects in dairy products wherever some flavours and gas are not
desired.
24
2.8 Types of Yogurt
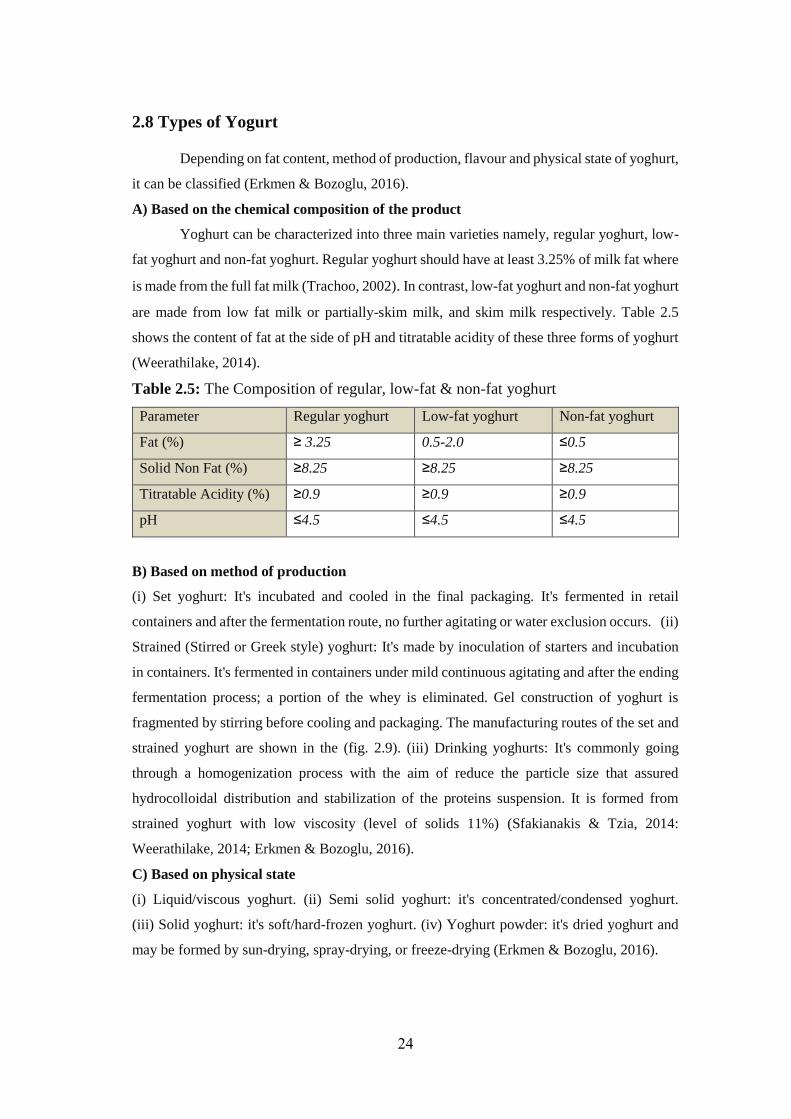
Depending on fat content, method of production, flavour and physical state of yoghurt,
it can be classified (Erkmen & Bozoglu, 2016).
A) Based on the chemical composition of the product
Yoghurt can be characterized into three main varieties namely, regular yoghurt, low-
fat yoghurt and non-fat yoghurt. Regular yoghurt should have at least 3.25% of milk fat where
is made from the full fat milk (Trachoo, 2002). In contrast, low-fat yoghurt and non-fat yoghurt
are made from low fat milk or partially-skim milk, and skim milk respectively. Table 2.5
shows the content of fat at the side of pH and titratable acidity of these three forms of yoghurt
(Weerathilake, 2014).
Table 2.5: The Composition of regular, low-fat & non-fat yoghurt
Parameter Regular yoghurt Low-fat yoghurt Non-fat yoghurt
Fat (%) ≥ 3.25 0.5-2.0 ≤0.5
Solid Non Fat (%) ≥8.25 ≥8.25 ≥8.25
Titratable Acidity (%) ≥0.9 ≥0.9 ≥0.9
pH ≤4.5 ≤4.5 ≤4.5
B) Based on method of production
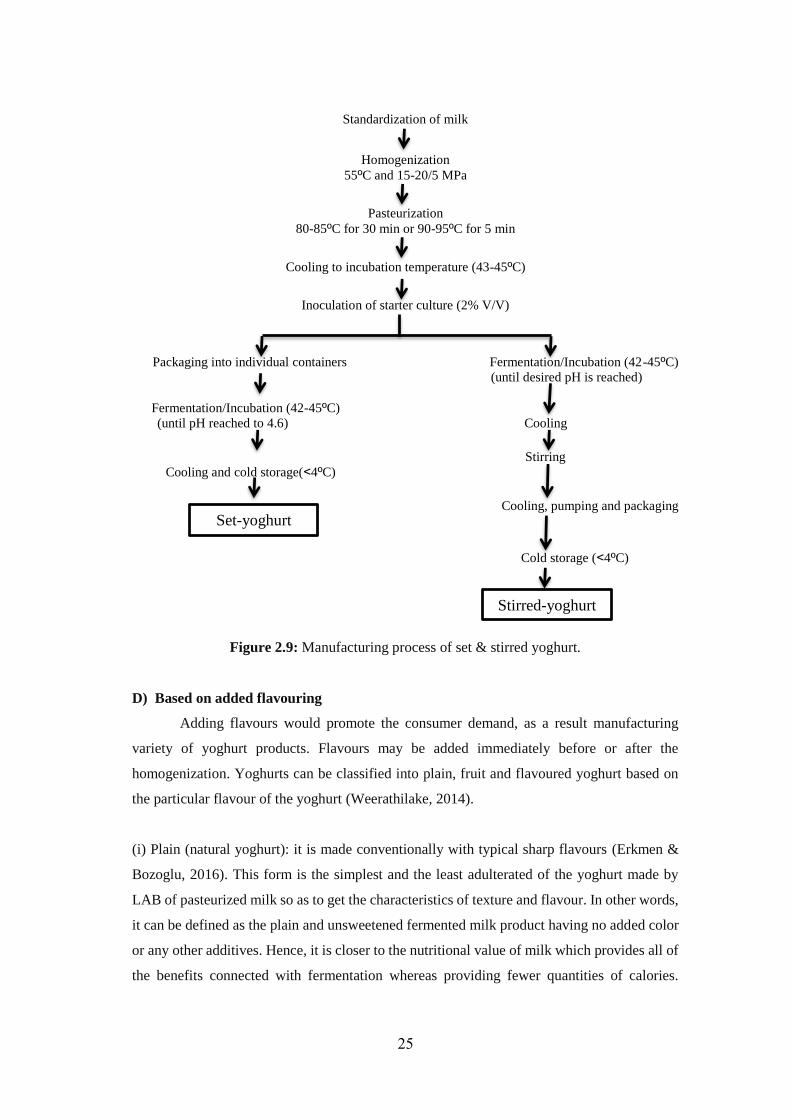
(i) Set yoghurt: It's incubated and cooled in the final packaging. It's fermented in retail
containers and after the fermentation route, no further agitating or water exclusion occurs. (ii)
Strained (Stirred or Greek style) yoghurt: It's made by inoculation of starters and incubation
in containers. It's fermented in containers under mild continuous agitating and after the ending
fermentation process; a portion of the whey is eliminated. Gel construction of yoghurt is
fragmented by stirring before cooling and packaging. The manufacturing routes of the set and
strained yoghurt are shown in the (fig. 2.9). (iii) Drinking yoghurts: It's commonly going
through a homogenization process with the aim of reduce the particle size that assured
hydrocolloidal distribution and stabilization of the proteins suspension. It is formed from
strained yoghurt with low viscosity (level of solids 11%) (Sfakianakis & Tzia, 2014:
Weerathilake, 2014; Erkmen & Bozoglu, 2016).
C) Based on physical state
(i) Liquid/viscous yoghurt. (ii) Semi solid yoghurt: it's concentrated/condensed yoghurt.
(iii) Solid yoghurt: it's soft/hard-frozen yoghurt. (iv) Yoghurt powder: it's dried yoghurt and
may be formed by sun-drying, spray-drying, or freeze-drying (Erkmen & Bozoglu, 2016).
25
Standardization of milk
Homogenization
55ºC and 15-20/5 MPa
Pasteurization
80-85ºC for 30 min or 90-95ºC for 5 min
Cooling to incubation temperature (43-45ºC)
Inoculation of starter culture (2% V/V)
Packaging into individual containers Fermentation/Incubation (42-45ºC)
(until desired pH is reached)
Fermentation/Incubation (42-45ºC)
Cooling (until pH reached to 4.6)
Stirring
cold storage(˂4ºC) Cooling and
Cooling, pumping and packaging
Cold storage (˂4ºC)
Figure 2.9: Manufacturing process of set & stirred yoghurt.
D) Based on added flavouring
Adding flavours would promote the consumer demand, as a result manufacturing
variety of yoghurt products. Flavours may be added immediately before or after the
homogenization. Yoghurts can be classified into plain, fruit and flavoured yoghurt based on
the particular flavour of the yoghurt (Weerathilake, 2014).
(i) Plain (natural yoghurt): it is made conventionally with typical sharp flavours (Erkmen &
Bozoglu, 2016). This form is the simplest and the least adulterated of the yoghurt made by
LAB of pasteurized milk so as to get the characteristics of texture and flavour. In other words,
it can be defined as the plain and unsweetened fermented milk product having no added color
or any other additives. Hence, it is closer to the nutritional value of milk which provides all of
the benefits connected with fermentation whereas providing fewer quantities of calories.
Stirred-yoghurt
Set-yoghurt

26
Additionally, plain yoghurt provides the pure yoghurt flavour and holds the richest calcium
content among the yoghurt products (Weerathilake, 2014).
(ii) Fruit yoghurt: it's formed by adding fruit such as strawberry, grape, apple, banana, etc.
(Erkmen & Bozoglu, 2016). There are two types of fruit yoghurts: The first one has the fruits
set at the bottom of the packaging (sundae-style yoghurt) whereas the second one has the fruit
homogeneously distributed within the yoghurt itself (Swiss-style yoghurt). Fruit pieces or pulp
that produces diverse flavours are added at production stage (Weerathilake, 2014, Erkmen &
Bozoglu, 2016).
(iii) Flavoured yoghurt: it's formed by adding synthetic flavouring, colouring and sweetening
agents to yoghurt (Erkmen & Bozoglu, 2016). Generally, flavours are added to yoghurt during
production stage and the additions of flavours lead to not only a wide array of tastes but also
increase sweetness of the product (Weerathilake, 2014).
In some countries a layer of fruit syrup on the upper of set yoghurt is a common
substitute. In other countries thicker stirred yoghurt is desired. This can be prepared by
addition dried skimmed milk (at around 50g/l) to the milk before pasteurising. Using additional
thickeners such as starch and pectin is as well possible, but generally unnecessary (Sfakianakis
& Tzia, 2014).
2.9 Yoghurt Processing
Yoghurt making starts from milking of the animal then passes through several
processes and the last step is the packaging of the final product (Sfakianakis & Tzia, 2014).
Yoghurt is basically made from bovine milk and milk from other animals is employed for
yoghurt making also. Yoghurt resulting from non-bovine species tends to be different in
several sensory and physicochemical features due to differencies in the composition of milk.
For instance, yoghurt resulting from milk with high content of fat (e.g., sheep, goat, and
buffalo) have creamier comparing with milk having lower content of fat (e.g., bovine, mare,
and ass). Thus, the species of the milk-producing animal meaningfully affect on the features
of the formed yoghurt (Tamime & Robinson, 2007; Sfakianakis & Tzia, 2014). Yoghurt
processing comprises following steps: 1-Standardization 2-pasteurization 3-homogenization
of milk 4-inoculation with starters 5-adding of flavours (optional) 6-incubation 7-cooling 8-
packaging (Fig. 2.7) (Erkmen & Bozoglu, 2016).

27
Figure 2.10: Flow Chart of yoghurt (Source: Erkmen & Bozoglu, 2016).
2.9.1 Initial Treatment of Milk
In the manufacturing of dairy products, the raw milk undergoes into clarification via
centrifugation to eliminate somatic cells and any other solid contaminations. Next,
thermalization (mild heating) is done at temperature ranged from 60–69 °C for 20–30 s and
aims to kill a lot of vegetative germs and the fractional inhibition of some enzymes. then, the
milk is cooled at <5 °C or inoculated with LAB or other cells to regulate the development of
the psychrotrophic bacterial cells (Tamime & Robinson, 2007).
2.9.2 Milk components standardization:
The standardization of milk refers to the adjustment of fat and solid-non-fat content (SNF).
2.9.2.1 Fat content standardization:
Fat content of bovine milk ranged from 3.2%–4.2%. The fat content of the milk is
adjusted to range from <0.5% for skim milk, 1.5%–2% for semi-fat milk, 3.5% for full fat milk
(Sfakianakis & Tzia, 2014). According to consumer demands, the fat content varieties from
0.1%–10% (Erkmen & Bozoglu, 2016).
For the reason that the fat content of the milk plays essential role about the yoghurt
features, so the standardization is an important process. The increase in fat content of milk
leads to an increase in the consistency and viscosity of yoghurt. Too, the content fat in milk
Milk+ supplements
Homogenization
(at 100-200 atm. and 50-60ºC)
Pasteurization
(at 90-95ºC for 5-10min)
Fermentation
(at 43ºC for 3 to 6h)
Cooling (to 29ºC)
Packaging and storage
Starter culture
Cooling (to 43ºC)

28
affects on the maximum rate of pH decline and pH lag phase through yoghurt making
(Soukoulis et al.; 2007; Sfakianakis & Tzia, 2014).
Practically, achieving the considered fat level, either the adding of skim milk, milk fat or the
separation of fat from milk by centrifugation and blending milk fat with skimmed milk is
carried out (Tamime & Robinson, 2007).
2.9.2.2 Solid (nonfat) content standardization:
The SNF constituents of milk primarily are composed of lactose, protein and minerals;
SNF content differs from 11% to 14% in the milk whereas the SNF of yoghurt ranged from
9% to 16%. In some cases, the SNF content of milk intended for yoghurt making is improved
by makers so as to achieve the preferred characteristics of the coagulum; greater SNF level in
yoghurt, greater resulting viscosity and firmness. In some countries, adding native milk
constituents such as powdered milk (skim or full fat), whey protein concentrates or powdered
casein is allowed to yoghurt and fermented dairy products. It's relatively common in yoghurt
production to reach at the desired SNF content thus leads to increase in firmness and stability.
It should be noted that the fat and SNF content of milk have an effect on the fermentation
process. Particularly, the interactions of SNF content of milk and incubation temperature play
a significant role in the duration of the fermentation process; increasing SNF increases the
duration of the fermentation process (Kristo et al., 2003; Sfakianakis & Tzia, 2014).
2.9.3 Addition of additives:
The following additives can be used in yoghurt making (50 mg /kg
in final product):
sorbic acid and its salts (K+, Na+, Ca+2), sulfur dioxide and benzoic acid proposed by food and
agricultural organization (FAO) and world health organization (WHO). Gelatin, starch and
agar also can be used as stabilizer (0.1–0.5%) and supplemented into yoghurt to enhance gel
structure, creaminess, viscosity, firmness, water-holding capacity and protein stability
(Erkmen & Bozoglu, 2016).
2.9.4 Homogenization:
Increases in viscosity and coagulation steadiness of the yoghurt occurred by
homogenization of the milk (Erkmen & Bozoglu, 2016). It leads to reduce the globules sizes
of fat into 1-2μm. It also stops the separation of fat during production. Thus; it enhances the
incorporation of other materials as powdered skim milk. After homogeneousness, membranes
of the fat globules are destroyed then lipase enzymes attack these globules. Therefore, to
prevent lipolysis, heat treatment of milk should occur directly after homogenization (Erkuş,
2007).

29
2.9.5 Heat Treatment: Pasteurized milk treated thermally either by passing it through plate heat exchanger at 90-
95°C for 5-10 min or in process vessel at 80-85°C for 30 min (Erkuş, 2007). Heat treatment
has several functions in milk as follows:
Partially hydrolysing and denaturing casein, liberating glycopeptide from κ-casein,
causing interchain cross-linkage (increases in micelle size), deactivating
immunoglobulins and making active SH group.
Increasing the hydrophilic properties of casein and forming stable coagulum.
Deactivating protease, lipase and microbial cells.
Denaturing proteins, creating peptides and flavour compounds.
Leading to form lactones and ketones (flavour) compounds.
Destructing some water-soluble vitamins as C, B1, B6, B12 and folic acid (Erkmen &
Bozoglu, 2016).
2.9.6 Fermentation Process
Treated milk is cooled to 43°C to inoculate starter cultures (Erkmen & Bozoglu,
2016). The amount of inoculation may vary from 0.5-5 % but 2% is the optimal value. Yoghurt
starters are S.thermophilus and L.bulgaricus. Additional Lactobacilli ssp are also utilized such
L.lactis, L.casei, L.jugurticus and L.helveticus. Generally, S.thermophilus and L.bulgaricus
are produced as blends in (1:1) ratio for inoculation and they are propagated together at 43°C.
The starters are mixed with milk by stirring. Inoculated milk are incubated at 43°C for 3- 6 h
until reach to the wanted acid. The lactic acid percentage of good ended yoghurt is about 0.85–
0.90 % (pH 4.2) (Erkuş, 2007).
During the fermentation process, cocci grow initially followed by bacilli with the purpose of
the counts of two strains become nearly equal after around 3h. At pH 4.2–4.4, S.thermophilus
tends to be repressed whereas L.bulgaricus can resist the pH in the range 3.5–3.8. Resulting
biological and physical modifications happen in milk throughout fermentation as following:
(i)The starters consume lactose in milk to create lactic acid and energy.
(ii) Casein micelles destabilized by lactic acid.
(iii) Aggregation for casein micelles and partially combine once reaching pH to the isoelectric
point 4.6–4.7.
(iv) α-Lactoalbumin and β-lactoglobulin interact with κ-casein and keeps the micelles from
complete disruption (Erkmen & Bozoglu, 2016).

30
2.9.7 Cooling and Storage:
The yoghurt is cooled to around 5°C after the pH reached 4.7–4.3. This stops the
growth and metabolism of the starters and stops the increase in sourness. Cooling may be in
one or two phases. One-phase cooling comprises the quick decrease of the coagulum
temperature to less than 10°C whereas the fermentation process is stopped and leads to produce
low viscosity of yoghurt. Two-phase cooling starts with rapid decrease in temperature to less
than 20°C and then slowly reaching to 4°C (storage temperature) leading to increase in
viscosity and restricted syneresis for yoghurt. This is fairly common in the yoghurt making,
particularly when fruits are added (Tamime & Robinson, 2007; Sfakianakis & Tzia, 2014).
Yoghurt are kept at 4°C for 7-14 day. Cooling decreases, the metabolic activity of the starters
thus controls the acidity of the yoghurt. Fresh yoghurt contains around 109
cell/gram, which
may decline to 106
cell/gram throughout storage. Bacilli generally drop more rapidly than cocci
and L.bulgaricus is more sensitive than S.thermophilus to low temperature (Erkmen &
Bozoglu, 2016).
2.10 Factors affecting slow growth of starter cultures and Yoghurt
quality:
To get a high quality of yoghurt with desired flavour, aroma, viscosity, consistency
and appearance, free from synersis and have long duration,there are numerous factors should
be controlled throughout the processing of yoghurt.The factors that affect on the quality of
yoghurt are standardization of milk, additives which are used to enhance the texture and
viscosity of yoghurt, homogenization, treatment by heating, starter strains, invasion of phages,
and inhibitory substances as disinfectants, detergents and antibiotics (Erkmen & Bozoglu ,
2016).
2.10.1 Compounds that are naturally present in milk
Numerous antimicrobial systems occur in milk to protect the suckling animals from
infections and diseases. Lactenins, the inhibitory complexes, are heat sensitive and they are
damaged when heating the milk at 68–74°C. Peroxidase system, another bactericidal
component found naturally in milk, comprises lactoperoxidase /thiocyanate/hydrogen
peroxide [LP/SCN- /H2O2] shortened as LPS.The inhibitory compounds in this systems are the
outcome of oxidation reactions where in the presence of H2O2, the LP stimulates the oxidation
of thiocynate to non-inhibitory compounds (SO42-, CO2 and NH3) next more oxidation to form
intermediate inhibitory substances as hypothiocyanate or higher oxyacids. In the presence of
some reducing substances (e.g. cysteine and dithionite), the inhibition became reversible. Most

31
starters are resistant to LP systems but some lactic cultures may cause sensitive mutants.
Otherwise, continual proliferation of starters in sterilized milk may affect on the sensitivity of
the cells to the LP system. Adding peroxidase or reducing agents like cysteine and dithionite
into sterilized milk are a preventative measure. LP systems deactivated by heating milk at
85°C for 16 sec. Heat treatment of yoghurt milk at 85°C for 30min or at 90–95°C for 5–10min
are adequate to destroy the natural inhibitors .Other inhibitory systems which must be taken
in consideration are: (a) bacterial agglutinin, causing agglutination of the starters, can affect
on the growth and metabolic activity (b) certain forms of forage (i.e; musty silage, turnips,
vetch), resulting in a milk containing inhibitory substances, can decline the amount of acid
producing by starters even after heating the milk for 15 min at 90°C (Tamime & Robinson,
1999).
2.10.2 Inoculation rate and Incubation temperature
The optimal rate for acid development is obtained within a very short period when the
starters are incubated at 40–50°C. The growth manners of S. thermophilus and L.bulgaricus
(single or blended cultures) have been shown in Fig. 2.7 but in industrial conditions, yoghurt
is manufactured over a short or long period using fermentation temperatures at 30°C or 45°C,
respectively. In the previous method of production, a reduced rate of acid development
becomes unavoidable although this effect is controlled by processing conditions, thus the
quality of the end product can be influenced. The inoculation rate may also affect on the rate
of acid development during production of yoghurt. Imprecise rates of starters' addition to the
milk can influence on the acid development of S.thermophilus and L.bulgaricus (Tamime &
Robinson, 1999).
2.10.3 Somatic cell count and Mastitis milk
35% of decline in the action of yoghurt starter caused by the large numbers of somatic
cells in milk but heating the milk to 90°C for 20 min or boiling for 2 min deactivates the cells
absolutely (Tamime & Robinson, 1999).
2.10.4 Antibiotic residues
Mastitis can influence on the quality and yield of milk. Antibiotic therapy, particularly
intra mammary injection, is extensively used to treat of mastitis in the mammal animals. The
most common antibiotics used are penicillin, neomycin, streptomycin, tetracycline,
chloramphenicol, erythromycin, sulfonamide, spiramycin, ampicillin and cloxacillin. These
antibiotics are concentrated in the udder cells then progressively diffuse in milk and pollute it.

32
Subsequently, the milk shouldn't be got from animals' treated with antibiotic within 72h. Slight
amounts of antibiotic can influence on the viability of the starters. The main outcome of
antibiotic remains in yoghurt milk causing collapse in the cooperative growth among
S.thermophilus and L.bulgaricus or a slowdown in the acid development rate (i.e. longer
processing time) and this can lead to syneresis or wheying-off. Antibiotic residues in milk and
other dairy products are also unsafe on public health (Tamime & Robinson, 1999 & Erkmen
& Bozoglu, 2016).
2.10.5 Disinfectant and detergent residues
Disinfectant and detergent are extensively used in the dairy industry to disinfect and
clean dairy apparatus. Those are chlorine compounds, quaternary ammonium compounds,
iodophore, ampholytes, and alkaline. Some dairy manufacturers add biocidal compounds
(e.g. H2O2) to milk to enhance and maintain the quality. This later method is not
recommended for public health reasons and the existence of such compounds in milk can
adversely influence or completely deactivates the growth of starters (Tamime & Robinson,
1999 & Erkmen & Bozoglu, 2016).
2.10.6 Environmental pollution
Cases of insecticide residues in milk are stated and this incidence may be due either
to post-milking contamination or by fodder sprayed with insecticide gave as feeding for
cattle. Heating the milk contaminated with pesticide and the fermentation process
contributed towards the degradation of pesticides (Tamime & Robinson, 1999).
2.10.7 Bacteriophage
Bacteriophage are viruses which can invade and damage the yoghurt starters and cause
failure in lactic acid production lead to poor coagulation of the processed milk. Until recently,
phages of mesophilic LAB which are used in cheese manufacture occupy the most attention.
However, the phage problems of yoghurt starters become more common and they cause
serious economic losses because of the vast increase in the production of yoghurt around the
world (Erkuş, 2007). Phage in yoghurt ordinarily represses S.thermophilus. Phages are
removed by heating of milk at 85°C for 20min (Erkmen & Bozoglu, 2016).

33
Figure 11: Scanning electron micrograph illustrating (A) a healthy S.thermophilus culture
and (B) the lysis of cells after invasion with a virulent bacteriophage (Source: Tamime &
Robinson, 1999).
2.10.8 Bacteriocins
Antibacterial materials are formed by a wide range of bacteria including dairy
starter's starins (Tamime & Robinson, 1999).
2.11 Molecular identification:
Old-style identification techniques based on physiological characteristics, besides
innovative phenotypic methods, such as cell wall composition analysis, whole-cell protein
fingerprinting and fatty acid analysis, often have need of considerable times and expertises,
and the outcomes are not always acceptable, mainly in cases of phylogenetically closely
related species ( Papadelli et al., 2003 ; Dellaglio & Felis, 2005 ; Tabasco et al., 2007 ),
Therefore, a clear identification into species may sometimes be very hard (Giraffa et al.,
1998).
Recently, investigations have focused on applying molecular techniques for reliable
differentiation of LAB and a variety of phenotypic and genotypic approaches is now
obtainable (Drake et al. 1996; Khaled et al. 1997; Quere et al. 1997; Tilsala-Timisjarvi and
Alatossava 1997; Giraffa et al., 1998; Papadelli et al., 2003).
Gathering data related to the sequences of the 16S rRNA, 23S rRNA, the 16S–23S
rRNA intergenic spacer regions or genes encoding enzymes have allowed identification of
LAB using specific oligonucleotides as probes or in PCR assays (Ward et al., 1998; Flint et
al., 1999; Deasy et al., 2000; Igarashi et al., 2001; Fortina et al., 2001; Chagnaud et al., 2001;
Papadelli et al., 2003).

34
To attain a quick and reliable identification of species, PCR- based approaches using
species-specific primers targeting the 16S rRNA gene sequence diversity have become very
popular (Coeuret et al., 2003; Tabasco et al., 2007). 16S ribosomal DNA is a part of the
ribosomal RNA- a 1542 nucleotide long component of the small prokaryotic ribosomal subunit
(30S). 16S rRNA sequence is provide precise basis for phylogenetic studies (Kermanshahi &
Peymanfar, 2012) as it is highly conserved between different species of bacteria. DNA
primers, for use in the PCR, can be designed to the highly-conserved sequences on the 16S
rRNA, which often flank hypervariable regions that can provide species-specific signature
sequences useful for identification.
Detecting and identifying various species of LAB with rapid means is considered as a
major importance among others in order to control the quality of dairy products, in particular,
when blended cultures are present and to monitor fermentation processes (Papadelli et al.,
2003).

35
Chapter 3
Materials and Methods

36
Chapter 3
Materials and Methods
3.1 Materials
3.1.1 Chemicals and Reagents
Chemicals, cultures medium and reagents used in this study are shown in Table 3.1
Table (3.1): Chemicals, reagents and cultures mediums that were used in this work
Reagents & Cultures Media Manufacture Country
Agar HiMedia India
Yeast extract HiMedia India
Tween 80 HiMedia India
Meat Extract HiMedia India
D(+) Glucose HiMedia India
Sodium Acetate HiMedia India
D(+) Lactose HiMedia India
Triammonium citrate HiMedia India
Trisodium Citrate Dihydrate HiMedia India
O2.7H4MgSO HiMedia India
2MgCl HiMedia India
O2.4H4MnSO HiMedia India
NaCl HiMedia India
4HPO2K HiMedia India
NaOH HiMedia India
Glycerol HiMedia India
Skim milk HiMedia India
Bromocresol purple Sigma-Aldrich USA
Hydrogen peroxide (30%) HiMedia India
Gas generating kit Oxoid UK
Gram stain kit HiMedia India
API 50 CH fermentation assays BioMerieux France
Phenolphthalein HiMedia India
Agarose HiMedia India
Promega wizard Genomic DNA Purification
Kit
Promega USA
Primers Invitrogen USA
PCR Master Mix 2X Kit Invitrogen USA
DNA Ladder 100bp Invitrogen USA
Eco RI enzyme Invitrogen USA
37
3.1.2 Equipments
The major equipments used are listed in Table 3.2.
Table (3.2): Major Equipments used in the present study.
Instrument Manufacturer Country
Centrifugation DRE USA
Shaker Grant UK
Incubator IKS international USA
PCR BioRad USA
Gel Electrophoresis Biometra Germany
Light microscopic Olympus Japan
Anaerobic jars Oxoid UK
3.2 Methodology:
3.2.1 Traditional yoghurt samples
Six samples of cow’s traditional yoghurt samples were gathered from the households
of 3 geographical areas of the Gaza strip (Rafah, Khanyonis and Gaza cities). The samples
gathered in sterile cups and saved cool until they could be taken to the laboratory, where
they kept at 4ºC for further use.
3.2.2 Isolation of starter cultures
Ten grams of homogenized samples are diluted in 90 ml sterile saline solution (0.85%
NaCl) followed by making 7- fold serial dilution for isolation.
3.2.2.1 Selective Media and Growth Conditions
One in ten milliliter from each dilution subcultured twice in M17 and MRS plates used
for cocci and bacilli respectively. Avoiding yeast contaminants, the media complemented
with 100mgl-1 of cycloheximide antibiotic before being incubated at 42ºC for 2-3 days. The
MRS and M17 plates incubated anaerobically using the gas pack system to provide the
optimum gas for L.bulgaricus and S.thermophilus respectively. Colonies randomly
selected, then streak plate method was used to get purified, next kept at 4ºC for MRS and
M17 agar plates and at -20ºC for M17 and MRS broths supplemented by 20% glycerol as
a cryoprotective agent for further use (Mathara et al.,2004).

38
3.2.3 Phenotypic identification of isolates
Initially, all isolates examined for Gram stain and catalase activity (Harrigan and McCance,
1976). Cultures that gave Gram positive and catalase negative reactions lastly used for further
identification (Sharpe et al., 1979).
3.2.3.1 Gram Staining
The Gram status of the isolated bacteria were determined by the microscopic
examination of Gram-stained isolates. Ten μl of cell suspension was pipetted to a microscopic
slide and they were Gram stained after drying and fixation by exposure to a flame. The main
steps in Gram Staining procedure were as follows:
• Crystal violet staining for 1 min then removing the excess stain by washing under tap water.
• Staining with Gram’s iodine solution for 1 min then washing under tap water.
• Fixation the slide with decolourizer for 15sec then washing under tap water.
• Counter staining with safranin for 30 s then washing under tap water.
• Drying the slide then examine it under microscope to observe bacterial cells.
Gram-positive bacteria Deep violet to blue gram-negative bacteria Pink to red
3.2.3.2 Catalase Test
Catalase enzyme splits H2O2 into H2O and O2. Both L.bulgaricus and S.thermophilus
negative with catalase test.Small amount of culture placed onto a clean slide. Add a few drops
of 3% H2O2 onto the smear.
Positive result Rapid evolution of O2 (bubbling)
Negative result No bubbles / only a few scattered bubbles
3.2.3.3 Gas Production from Glucose
It's a major criterion for identifying the nature of isolated strains about homofermentative
or heterofermentative. Broth media of MRS or M17 containing inverted Durham tubes are
used for evaluation of gas production. Bubbles trapped within the Durham tube indicate the
production of gas. Even a single bubble is significant and means evidence of gas production
(Forbes et al., 2002). No bubbles within the Durham tube indicate a non-gas producing
organism (Mac Faddin, 1976).

39
Fifty μl of reactivated cultures are inoculated into 5 ml MRS containing Durham tubes
and incubated at 42°C for 5 days.
Homofermentative Accumulation of gas in the Durham tubes
Heterofermentative No gas in the Durham tubes
3.2.3.4 Growth at Different Temperatures
The criterion which is commonly used for the classification of bacilli is the Growing at
15°C and 45 °C, where L.bulgaricus can't grow at 15°C but can grow at 45°C. For cocci
isolates, growing at 10°C and 45°C are used. S. thermophilus can't grow at 10°C but can grow
at 45°C.To determine the growing of isolates at given temperatures, MRS and M17 media
containing bromocresol purple are used. Colour change 5+indicate lactic acid production and
cell growth (Hammes and Vogel 1995).
Fifty microliter of reactivated cultures are inoculated into 5 ml test media at 42ºC for
5 days (Erkuş, 2007).
Positive result Colour change from purple to yellow
Negative result No change in colour
3.2.3.5 Growth at Different NaCL Concentrations
The tolerance of isolates at different levels of salts is also evaluated (Ali, 2011).
L.bulgaricus and S.thermophilus are oversensitive to NaCl. S. thermophilus does't grows even
at 2% NaCl, but there is no data available for L.bulgaricus. The most commonly used NaCl
concentration for identification of bacilli are 4% and 6.5%. Therefore, the growth at 2% and
4% for cocci isolates and the growth at 4% and 6.5% for bacilli isolates are used in NaCl test
medium (Appendix B).
Fifty microliter of reactivated cultures are inoculated into 5ml NaCl test media at 42ºC
for 5 days (Erkuş, 2007).
Positive result Colour change from purple to yellow
Negative result No change in colour
3.2.3.6 Carbohydrate Fermentation profiles
Carbohydrate fermentation tests aims to determe the ability of organisms to ferment a
specific suger. It can be used to differentiate among bacterial groups or species. It detected by
the colour change of a pH indicator as forming acid products. Colour change only happens

40
when enough acid products have been produced by fermentation of the suger to low pH (6.8
or less). This test achieve by two steps as follows:
3.2.3.6.1 Preparation of active cell culture free from sugar:
• Ten ml MRS and M17 media are used for overnight activation of isolates at 37°C.
• Five ml of each bacterial culture pelleted by Centrifugation for 10 min at 10.000 rpm then
wash the pellet twice with sterile physiological saline (0.9%) (Tanigawa and Watanabe, 2011).
3.2.3.6.2 Combinations of active cell culture free from sugar and tested sugar:
An API 50 CH fermentation assay is a ready-to-use medium which allows the fermentation
of the 49 carbohydrates. The main steps this assay were as follows:
Transferring 100 µl of suspension into each of the 50 wells on the API 50 CH strips
except the first strip served as a control well.
Covering wells with parafin oil to affect anaerobiosis.
Incubating the strips at 42°C for 48h and monitoring the colour changes.
Positive result Colour change from purple "bromocresol purple indicator" to yellow.
For the esculin (tube no. 25), a change in colour from purple to black.
Negative result No change in colour.
The biochemical profile obtained for the strain after the final reading can be identified using
ABIS online for bacterial identification.
3.2.4 Molecular identification for isolated bacteria
3.2.4.1 DNA isolation
Ten ml MRS and M17 media are used for overnight activation of isolates at 37°C.
Harvest the isolates at 6.000 rpm for 5 min then remove the supernatant.
Genomic DNA of the isolated bacteria were extracted by Genomic DNA Purification
Kit Brand WISARD Promega® for isolation of Gram positive bacteria follow the
manufacturer instructions that presented in appendix C.
The extracted DNA was stored at –20 °C until use as a template for PCR.
3.2.4.2 Primers
cysmet2F (forward): 5′ GGAACCTGAAGGCTCAAT 3′
cysmet2R (reverse): 5′ GTCAACCACGGTAAAGGTC 3′
Primers for the methionine biosynthesis gene

41
9699–9700 primers are used for the amplification of 16S rRNA gene.
9699 5′ ATCCGAGCTCAGAGTTTGATCCTGGC 3′
9700 5′ TCAGGTCGACGCTACCTTGTTACGAC 3′
Primers for the amplification of 16S rRNA gene
3.2.4.3 PCR conditions
PCR amplifications are made on thermal cycler (BioRad, USA). For the analysis of
methionine biosynthesis gene, PCR conditions and reaction mixture was as follows:
Step 1: 94 ºC for 2 min (initial denaturation)
Step 2: 94 ºC for 30sec (denaturation)
Step 3: 54 ºC for 40 sec (annealing) 45 cycles
Step4: 72 ºC for 45 sec (elongation)
Step 1: 72 ºC for 10 min (final annealing)
The total reaction volume of PCR was 30μl and the amount of DNA was 1µl.
For 16S rRNA gene, PCR conditions and reaction mixture were as follows:
Step 1: 95 ºC for 2 min (initial denaturation)
Step 2: 95ºC for 1min (denaturation)
Step 3: 56 ºC for 1min (annealing) 35 cycles
Step4: 72 ºC for 1min (elongation)
Step 1: 72 ºC for 10 min (final annealing)
The total reaction volume of PCR was 30μl and the amount of DNA was 1µl.
3.2.4.4 Restriction analysis of 16S rRNA gene product
By Eco RI as the endonuclease enzyme, the restriction of 16S rRNA gene product
carried out. The reaction mixture contained 1x of the restriction enzyme buffer, 10 µl of PCR
product and 0.5 µl Eco RI (Delley & Germond, 2002).
3.2.4.5 Separation of Amplification Products
3.2.4.5.1 Preparation of Agarose Gel:
Dissolve 1.5 % agarose in 100µl 1x TAE buffer by boiling (Appendix C). It's cooled
to 45º C. Add 15µl ethidium bromide solution (Appendix C) with stirring. Pour the agarose
gel in casting stand then place the combs (Markiewicz et al., 2008).

42
3.2.4.5.2 Loading of Agarose Gel:
5µl of PCR products are mixed with 2μl of gel loading dye. Start from the second
well to load the samples into the wells. In the first well, load DNA ladder of 100 bp which is
used as marker (DNA molecular weight).
3.2.4.5.3 Electrophoresis of the Products:
PCR products are electrophoresed at 80V for 1h. Visualize the Amplified products in
a gel documentation system. Images can capture in a photodocumentation device. The
presence 1500-2000bp DNA fragment sizes point to that the amplification is achieved.
3.2.5 Assessment of technological performance of strains:
3.2.5.1 Preparation of fermented milk
Five liter pasteurized milk used for yoghurt production and milk powder with adding
(60g/L).
The mix was heated at 80ºC for 15 min in water bath, homogenized and then rapidly
cooled to 45ºC.
One % inocula (106 - 107 cfu/ml) of isolated yoghurt starters reactivated by incubation
in 10% (w/v) sterilized reconstituted skimmed milk and incubated at 37ºC for 18-24h
before experimentation involving pasteurized milk as the growth medium.
Each milk was inoculated with 5% (106 - 107 cfu/ml) of S.thermophilus, L.bulgaricus
and mixed yoghurt culture (1:1) with stirring after inoculation and incubated at 43oC
in a shaker water-bath for 8h.
Fiftty ml of samples were taken in aseptic sterile cups for microbiological and
biochemical tests every 1½-2hrs. The experiment was repeated 3 times.
3.2.5.2 Measurement of pH and titratable acidity
To measure the pH: Inserting the pH probe directly into a homogenized sample of the
fermented milk to determine pH.
Rinsing the electrode between samples with D.W and drying with tissue.
To determine the titratable acidity (lactic acid %): Titrating 1 ml of homogenized
fermented milk by 0.1 N NaOH using phenolphthalein indicator (0.1%, 3drops) with
continuously stirring until the indicator change to a definite pink colour lasting for 30
sec.
Recording the volume of NaOH required to neutralize the acidof yoghurt and calculate
the titratable acids using the following formula:

43
LA% = 10∗V NaOH ∗ 0.009∗0.1
𝑊∗ 100%
Where 10= Dilution factor; W=Weight of sample for titration; V NaOH= Volume of
NaOH used to neutralize the lactic acid; 0.1= Normality of NaOH (Arslan & Bayrakci,
2016).

44
Chapter 4
Results and Discussion

45
Chapter 4
Results and Discussion
4.1. Isolation and Biochemical identification of S. thermophilus and L. delbrueckii ssp.
bulgaricus:
In total six of cow’s traditional samples of home made yoghurt were used for the
isolation of yoghurt starters bacteria. Under anaerobic conditions at 42°C, bacterial cells were
isolated with MRS and M17 agar media for 3 days. M17 was used in order to isolate of S.
thermophilus and MRS was used in order to isolate of L. delbrueckii ssp. bulgaricus.
Has been observed under light microscope that the isolated strains gave blue- purple colour;
hence they were Gram positive bacteria. The morphology of the isolates coming from M17
plates were cocci with round or ovoid shape and it's appeared predominantly as pairs or
forming chains while the morphology of the isolates coming from MRS plates were long
bacilli with curved ends and it's appeared mostly as single or a chain of 3-4 cells. Light
microscopic images of cocci and bacilli isolates were given in Figure 4.1.
Figure 4.1: Microscopic appearance of cocci and bacilli isolates

46
The isolates were examined for catalase activity. Has been observed that all isolates
give a negative result with this test (no bubbles).
Additionally, when subjected to examine the gas production from glucose
fermentation and the test tubes left for 5 days, it was no observed gas accumulation in test
tubes. This means that tha isolates behave as homofermentative when ferment glucose.
With respect to the ability of isolates on growth at different concentrations of NaCl;
no record shows the ability of S.thermophilus growth even at 2% NaCl concentration but for
the isolates of bacilli, the most considerably used NaCl concentration was 4% and 6.5% for
identification. Bacilli isolates were sensitive to 4% and 6.5% NaCl concentration.
The ability of isolates on growth at different temperatures is another criterion for
classification. The ability of cocci isolates on growth at 10°C and 45°C were examined. S.
thermophilus were not able to grow at 10°C, but it grew well at 45°C. L. bulgaricus can't grow
at 15°C, but can grow at 45°C. Any that the isolates of cocci didn't show their ability to grow
at 10°C, also the isolates of bacilli didn't show their ability to grow at 15°C.
The worthiest examination to identify the differences among strains is carbohydrate
fermentation using API 50 CHL assays which allow the fermentation of 49 different sugars as
shown in table 4.1. The attained profile for the strains after the final reading can be identified
using ABIS online for bacterial identification.
After processing of the obtained results with software ABIS online, the strains
Lactobacillus and Streptococcus isolates were identified as Lactobacillus dekbrueckii ssp.
bulgaricus and Streptococcus thermophilus strains with high percentage of reliability.
According to MRS isolates, glucose, fructose, lactose and mannose were affirmative
with these isolates but with respect to S. thermophilus, the gathered data from different
studies show the variability of S. thermophilus for fermentation of sugars. It was noticed the
agreement all the scientists about affirmative fermentation of glucose, lactose and fructose
for S. thermophilus. In contrast mannose, galactose, sucrose, maltose, melibiose and
raffinose fermentations were variable. Consequently, in the estimation of data, affirmative
strains with glucose, lactose and fructose were specified to be S. thermophilus.
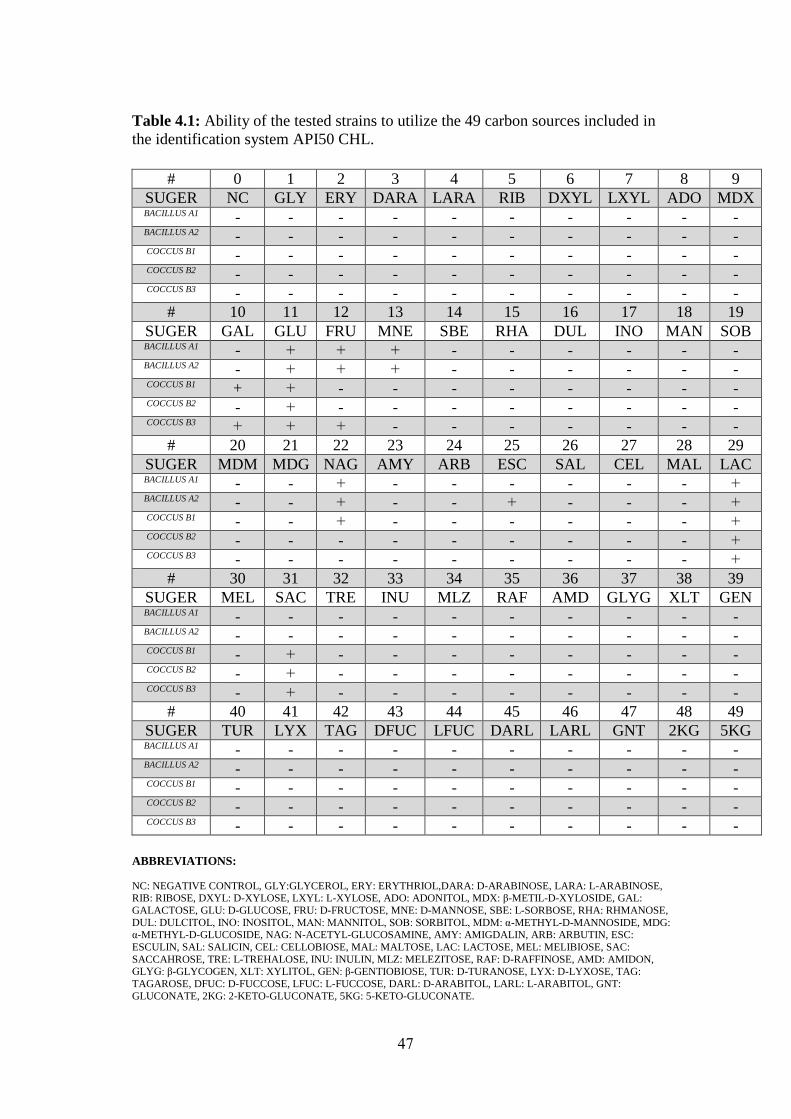
47
Table 4.1: Ability of the tested strains to utilize the 49 carbon sources included in
the identification system API50 CHL.
9 8 7 6 5 4 3 2 1 0 #
MDX ADO LXYL DXYL RIB LARA DARA ERY GLY NC SUGER
- - - - - - - - - - BACILLUS A1
- - - - - - - - - - BACILLUS A2
- - - - - - - - - - COCCUS B1
- - - - - - - - - - COCCUS B2
- - - - - - - - - - COCCUS B3
19 18 17 16 15 14 13 12 11 10 #
SOB MAN INO DUL RHA SBE MNE FRU GLU GAL SUGER
- - - - - - + + + - BACILLUS A1
- - - - - - + + + - BACILLUS A2
- - - - - - - - + + COCCUS B1
- - - - - - - - + - COCCUS B2
- - - - - - - + + + COCCUS B3
29 28 27 26 25 24 23 22 21 20 #
LAC MAL CEL SAL ESC ARB AMY NAG MDG MDM SUGER
+ - - - - - - + - - BACILLUS A1
+ - - - + - - + - - BACILLUS A2
+ - - - - - - + - - COCCUS B1
+ - - - - - - - - - COCCUS B2
+ - - - - - - - - - COCCUS B3
39 38 37 36 35 34 33 32 31 30 #
GEN XLT GLYG AMD RAF MLZ INU TRE SAC MEL SUGER
- - - - - - - - - - BACILLUS A1
- - - - - - - - - - BACILLUS A2
- - - - - - - - + - COCCUS B1
- - - - - - - - + - COCCUS B2
- - - - - - - - + - COCCUS B3
49 48 47 46 45 44 43 42 41 40 #
5KG 2KG GNT LARL DARL LFUC DFUC TAG LYX TUR SUGER
- - - - - - - - - - BACILLUS A1
- - - - - - - - - - BACILLUS A2
- - - - - - - - - - COCCUS B1
- - - - - - - - - - COCCUS B2
- - - - - - - - - - COCCUS B3
ABBREVIATIONS:
NC: NEGATIVE CONTROL, GLY:GLYCEROL, ERY: ERYTHRIOL,DARA: D-ARABINOSE, LARA: L-ARABINOSE, RIB: RIBOSE, DXYL: D-XYLOSE, LXYL: L-XYLOSE, ADO: ADONITOL, MDX: β-METIL-D-XYLOSIDE, GAL:
GALACTOSE, GLU: D-GLUCOSE, FRU: D-FRUCTOSE, MNE: D-MANNOSE, SBE: L-SORBOSE, RHA: RHMANOSE,
DUL: DULCITOL, INO: INOSITOL, MAN: MANNITOL, SOB: SORBITOL, MDM: α-METHYL-D-MANNOSIDE, MDG: α-METHYL-D-GLUCOSIDE, NAG: N-ACETYL-GLUCOSAMINE, AMY: AMIGDALIN, ARB: ARBUTIN, ESC:
ESCULIN, SAL: SALICIN, CEL: CELLOBIOSE, MAL: MALTOSE, LAC: LACTOSE, MEL: MELIBIOSE, SAC:
SACCAHROSE, TRE: L-TREHALOSE, INU: INULIN, MLZ: MELEZITOSE, RAF: D-RAFFINOSE, AMD: AMIDON,
GLYG: β-GLYCOGEN, XLT: XYLITOL, GEN: β-GENTIOBIOSE, TUR: D-TURANOSE, LYX: D-LYXOSE, TAG:
TAGAROSE, DFUC: D-FUCCOSE, LFUC: L-FUCCOSE, DARL: D-ARABITOL, LARL: L-ARABITOL, GNT:
GLUCONATE, 2KG: 2-KETO-GLUCONATE, 5KG: 5-KETO-GLUCONATE.

48
4.2: DNA Identification
Molecular approaches are proposed to emphasis the former results provided from the
physiological and biochemical assays to identify and diagnose Streptococci and Lactobacilli
isolates using PCR method and to achieve selective identification for proposed isolates. This
is realized by methionine biosynthesis gene and primers (cysmet2F and cysmet2R) to
identify the isolated strains of yoghurt samples. PCR method that used to amplify these
genes offer partial amplification of the methionine biosynthesis gene at 700 bp product in all
isolates as expected (Fig 4.2).
1kb
0.5kb
Figure 4.2: Partial amplification of the methionine biosynthesis gene in rural yoghurt isolates. Lanes
1, 2: Coccus B1, lanes 3,4: Coccus B2, Lanes 5, 6: Coccus B3, Lanes 7, 8: Bacillus A1, Lanes 9, 10:
Bacillus A2, NC: Negative control (no DNA control), M: 100 bp DNA ladder.
And to discriminate of lactobacilli from streptococci, this is realized by ARDRA
using endonucleases Eco RI. When we amplifying the 16S rRNA genes, the amplified
product was nearly 1500bp in all proposed isolates of lactobacilli and streptococci (Fig 4.3).
Then, the entire amplified product digested by Eco RI restriction enzyme. Two sizes of
lactobacilli were around 700,850bp whereas the two sizes of streptococci were around
650,850bp.
This is remarkable that supposed lactobacilli produced a different profile than that of
supposed streptococci when performing restriction analysis (Fig 4.4 a-b) and these result
agree with those obtained by Delley & Germond (2002).
NC 1 2 3 4 5 6 7
1 M 2 3 4 6 5 10 9 8
8
7
9 10 M
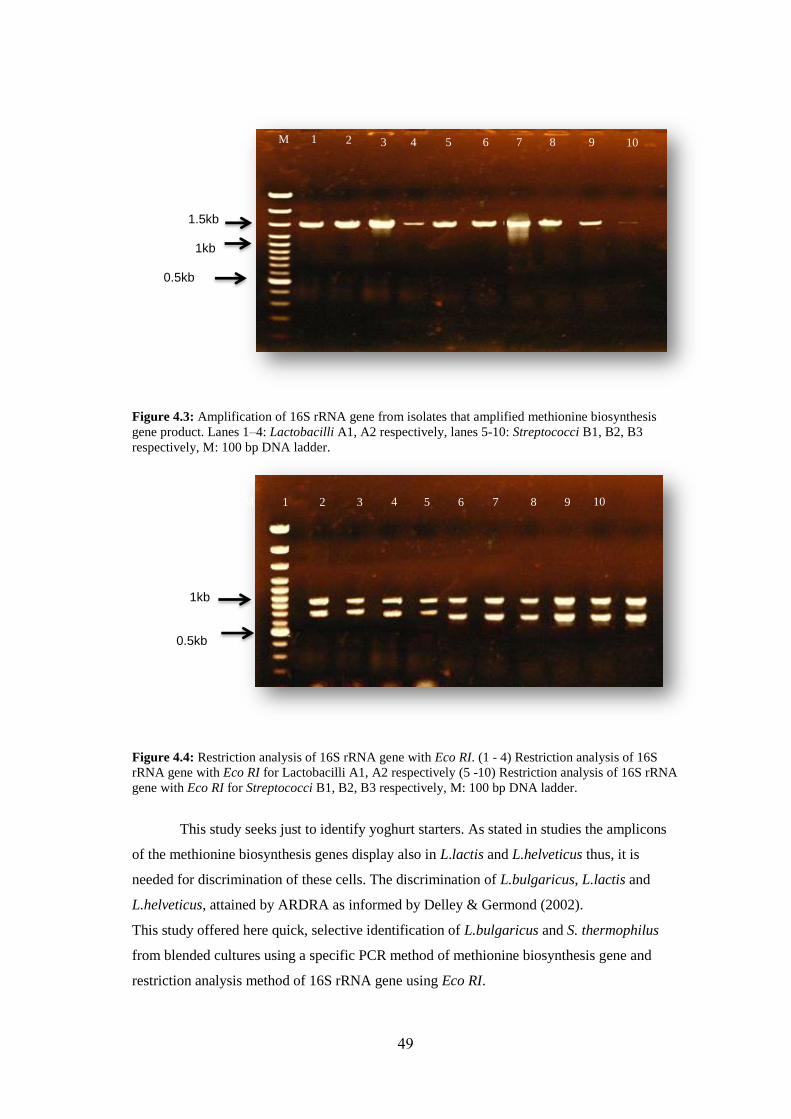
49
1.5kb
1kb 0.5kb
Figure 4.3: Amplification of 16S rRNA gene from isolates that amplified methionine biosynthesis
gene product. Lanes 1–4: Lactobacilli A1, A2 respectively, lanes 5-10: Streptococci B1, B2, B3
respectively, M: 100 bp DNA ladder.
1kb 0.5kb
Figure 4.4: Restriction analysis of 16S rRNA gene with Eco RI. (1 - 4) Restriction analysis of 16S
rRNA gene with Eco RI for Lactobacilli A1, A2 respectively (5 -10) Restriction analysis of 16S rRNA
gene with Eco RI for Streptococci B1, B2, B3 respectively, M: 100 bp DNA ladder.
This study seeks just to identify yoghurt starters. As stated in studies the amplicons
of the methionine biosynthesis genes display also in L.lactis and L.helveticus thus, it is
needed for discrimination of these cells. The discrimination of L.bulgaricus, L.lactis and
L.helveticus, attained by ARDRA as informed by Delley & Germond (2002).
This study offered here quick, selective identification of L.bulgaricus and S. thermophilus
from blended cultures using a specific PCR method of methionine biosynthesis gene and
restriction analysis method of 16S rRNA gene using Eco RI.
2
M
3
1
3
4
2
5
4
5
6
6
7
8
8
7
9
9
10
10 1
M
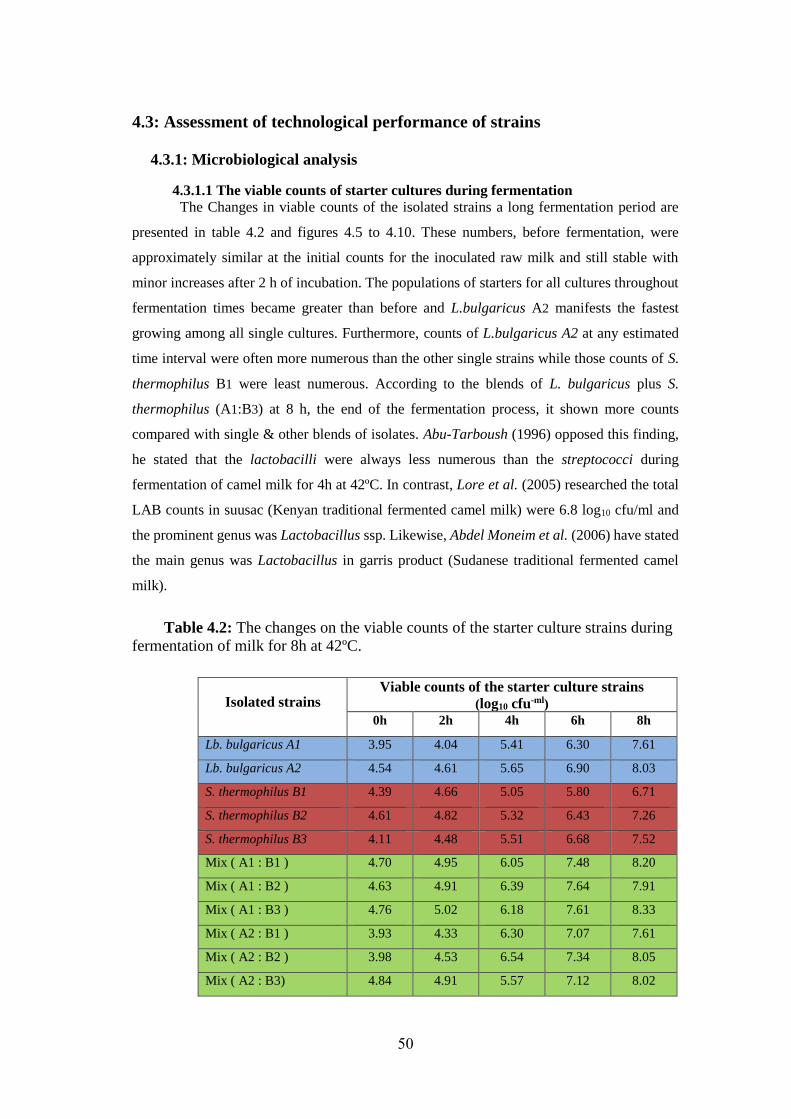
50
4.3: Assessment of technological performance of strains
4.3.1: Microbiological analysis
4.3.1.1 The viable counts of starter cultures during fermentation
The Changes in viable counts of the isolated strains a long fermentation period are
presented in table 4.2 and figures 4.5 to 4.10. These numbers, before fermentation, were
approximately similar at the initial counts for the inoculated raw milk and still stable with
minor increases after 2 h of incubation. The populations of starters for all cultures throughout
fermentation times became greater than before and L.bulgaricus A2 manifests the fastest
growing among all single cultures. Furthermore, counts of L.bulgaricus A2 at any estimated
time interval were often more numerous than the other single strains while those counts of S.
thermophilus B1 were least numerous. According to the blends of L. bulgaricus plus S.
thermophilus (A1:B3) at 8 h, the end of the fermentation process, it shown more counts
compared with single & other blends of isolates. Abu-Tarboush (1996) opposed this finding,
he stated that the lactobacilli were always less numerous than the streptococci during
fermentation of camel milk for 4h at 42ºC. In contrast, Lore et al. (2005) researched the total
LAB counts in suusac (Kenyan traditional fermented camel milk) were 6.8 log10 cfu/ml and
the prominent genus was Lactobacillus ssp. Likewise, Abdel Moneim et al. (2006) have stated
the main genus was Lactobacillus in garris product (Sudanese traditional fermented camel
milk).
Table 4.2: The changes on the viable counts of the starter culture strains during
fermentation of milk for 8h at 42ºC.
Isolated strains Viable counts of the starter culture strains
(log10 cfu-ml)
0h 2h 4h 6h 8h
Lb. bulgaricus A1 3.95 4.04 5.41 6.30 7.61
Lb. bulgaricus A2 4.54 4.61 5.65 6.90 8.03
S. thermophilus B1 4.39 4.66 5.05 5.80 6.71
S. thermophilus B2 4.61 4.82 5.32 6.43 7.26
S. thermophilus B3 4.11 4.48 5.51 6.68 7.52
Mix ( A1 : B1 ) 4.70 4.95 6.05 7.48 8.20
Mix ( A1 : B2 ) 4.63 4.91 6.39 7.64 7.91
Mix ( A1 : B3 ) 4.76 5.02 6.18 7.61 8.33
Mix ( A2 : B1 ) 3.93 4.33 6.30 7.07 7.61
Mix ( A2 : B2 ) 3.98 4.53 6.54 7.34 8.05
Mix ( A2 : B3) 4.84 4.91 5.57 7.12 8.02
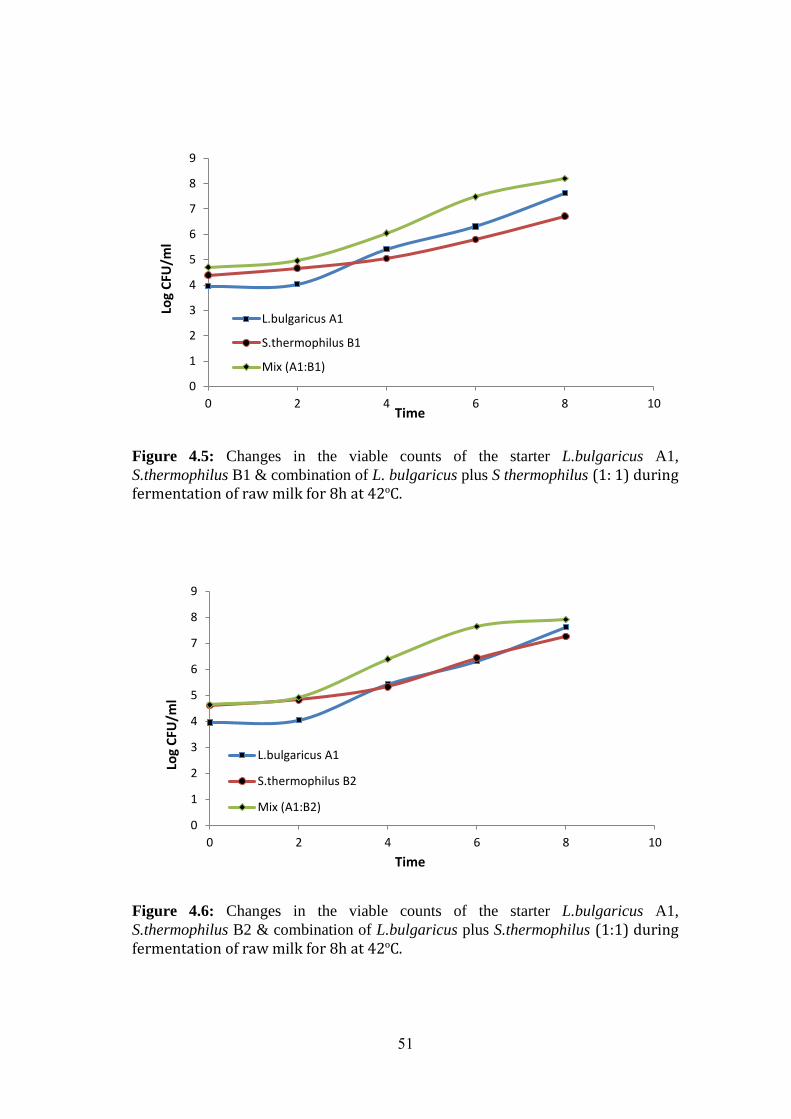
51
Figure 4.5: Changes in the viable counts of the starter L.bulgaricus A1,
S.thermophilus B1 & combination of L. bulgaricus plus S thermophilus (1: 1) during fermentation of raw milk for 8h at 42ºC.
Figure 4.6: Changes in the viable counts of the starter L.bulgaricus A1,
S.thermophilus B2 & combination of L.bulgaricus plus S.thermophilus (1:1) during fermentation of raw milk for 8h at 42ºC.
0
1
2
3
4
5
6
7
8
9
0 2 4 6 8 10
L.bulgaricus A1
S.thermophilus B1
Mix (A1:B1)
Log
CFU
/ml
Time
0
1
2
3
4
5
6
7
8
9
0 2 4 6 8 10
L.bulgaricus A1
S.thermophilus B2
Mix (A1:B2)
Log
CFU
/ml
Time
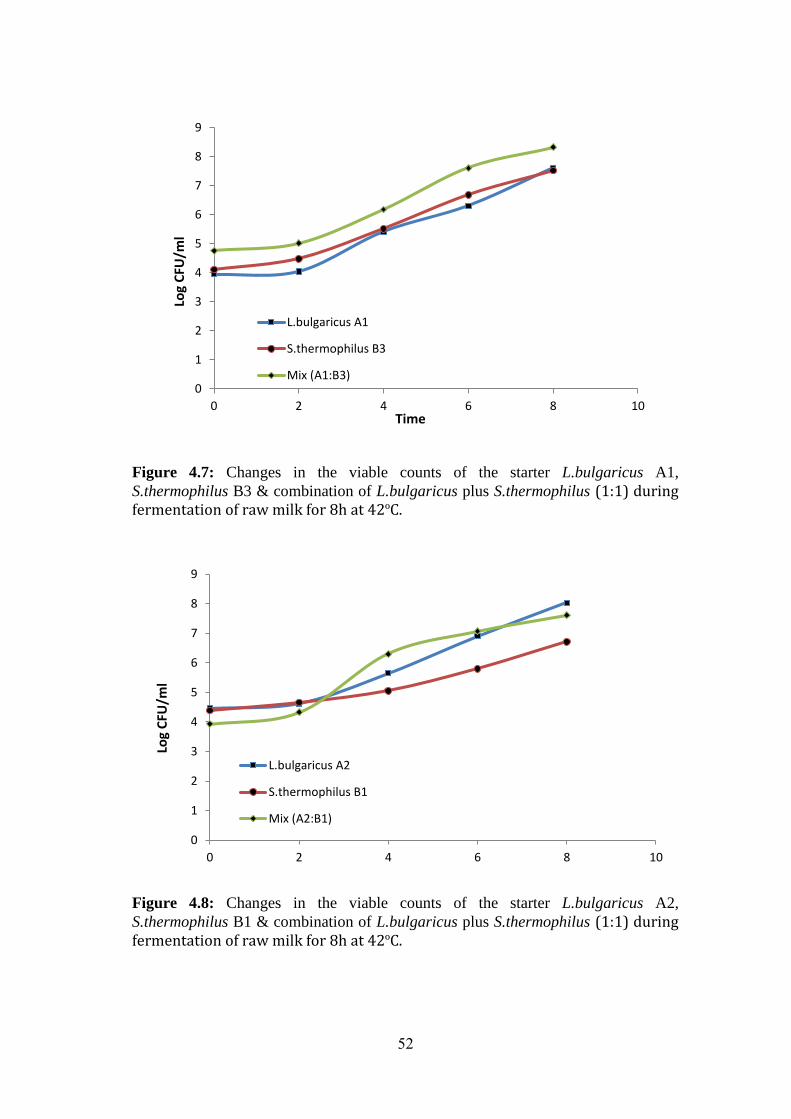
52
Figure 4.7: Changes in the viable counts of the starter L.bulgaricus A1,
S.thermophilus B3 & combination of L.bulgaricus plus S.thermophilus (1:1) during fermentation of raw milk for 8h at 42ºC.
Figure 4.8: Changes in the viable counts of the starter L.bulgaricus A2,
S.thermophilus B1 & combination of L.bulgaricus plus S.thermophilus (1:1) during fermentation of raw milk for 8h at 42ºC.
0
1
2
3
4
5
6
7
8
9
0 2 4 6 8 10
L.bulgaricus A1
S.thermophilus B3
Mix (A1:B3)
Log
CFU
/ml
Time
0
1
2
3
4
5
6
7
8
9
0 2 4 6 8 10
L.bulgaricus A2
S.thermophilus B1
Mix (A2:B1)
Log
CFU
/ml
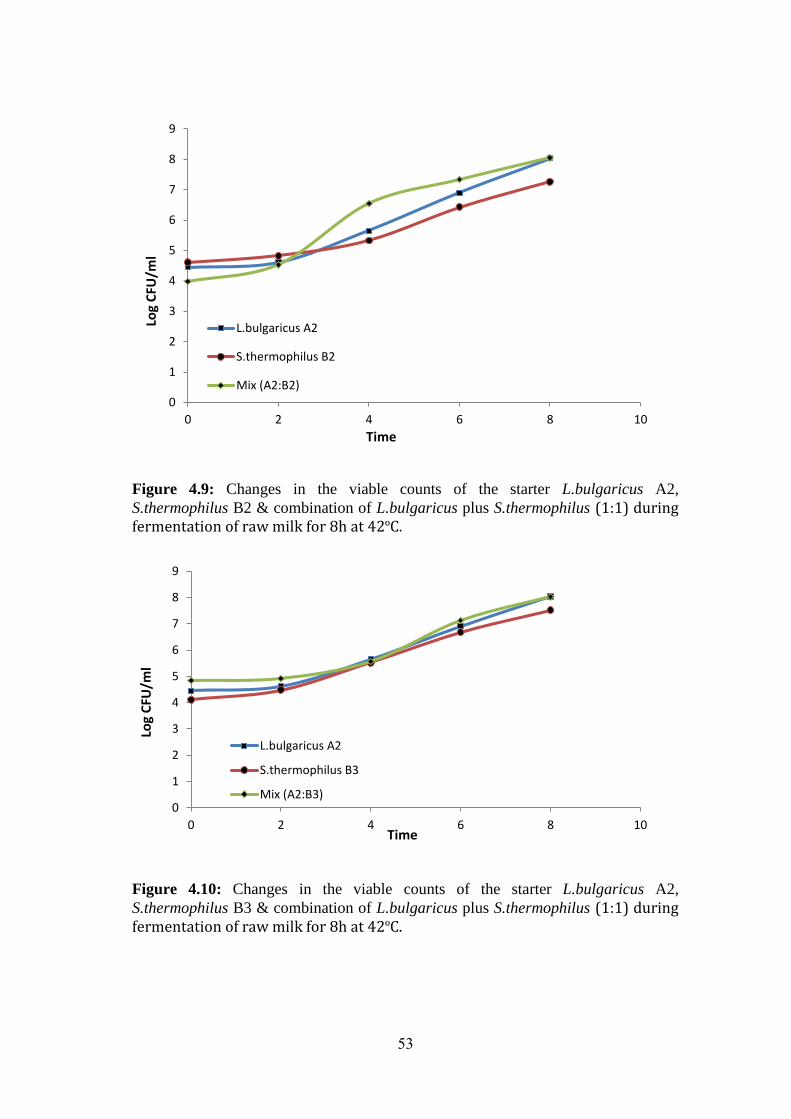
53
Figure 4.9: Changes in the viable counts of the starter L.bulgaricus A2,
S.thermophilus B2 & combination of L.bulgaricus plus S.thermophilus (1:1) during fermentation of raw milk for 8h at 42ºC.
Figure 4.10: Changes in the viable counts of the starter L.bulgaricus A2,
S.thermophilus B3 & combination of L.bulgaricus plus S.thermophilus (1:1) during fermentation of raw milk for 8h at 42ºC.
0
1
2
3
4
5
6
7
8
9
0 2 4 6 8 10
L.bulgaricus A2
S.thermophilus B2
Mix (A2:B2)
Log
CFU
/ml
Time
0
1
2
3
4
5
6
7
8
9
0 2 4 6 8 10
L.bulgaricus A2
S.thermophilus B3
Mix (A2:B3)
Log
CFU
/ml
Time
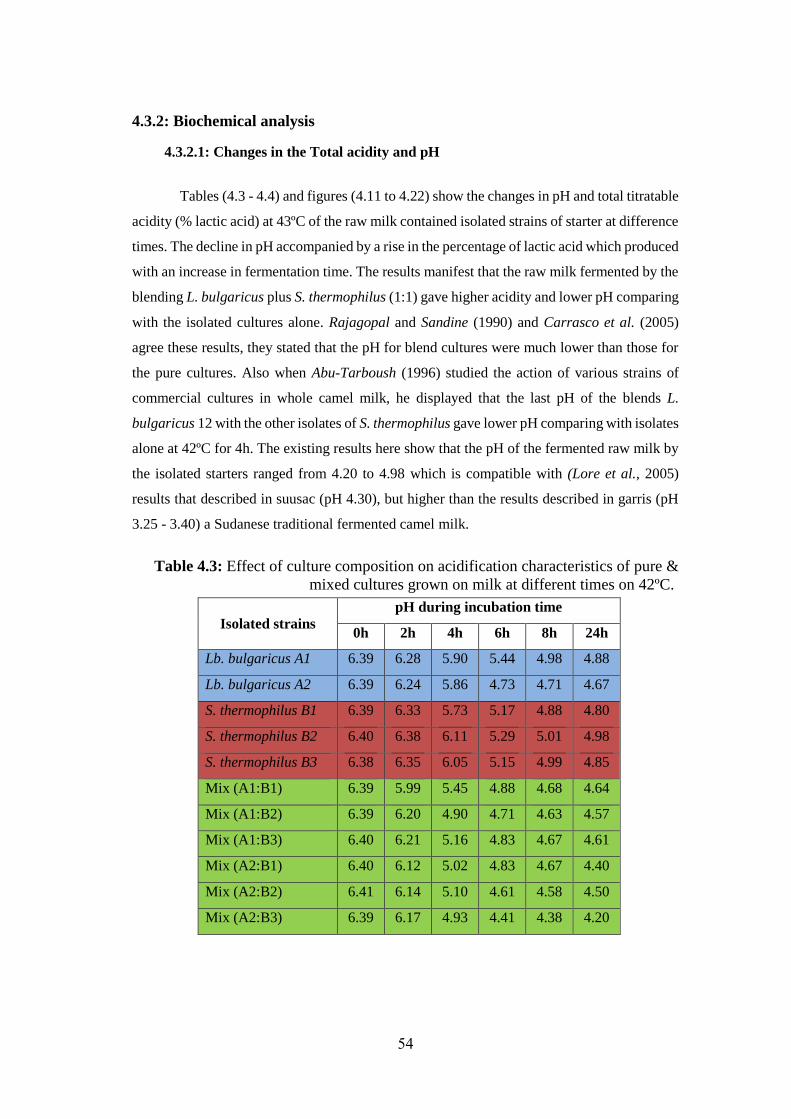
54
4.3.2: Biochemical analysis
4.3.2.1: Changes in the Total acidity and pH
Tables (4.3 - 4.4) and figures (4.11 to 4.22) show the changes in pH and total titratable
acidity (% lactic acid) at 43ºC of the raw milk contained isolated strains of starter at difference
times. The decline in pH accompanied by a rise in the percentage of lactic acid which produced
with an increase in fermentation time. The results manifest that the raw milk fermented by the
blending L. bulgaricus plus S. thermophilus (1:1) gave higher acidity and lower pH comparing
with the isolated cultures alone. Rajagopal and Sandine (1990) and Carrasco et al. (2005)
agree these results, they stated that the pH for blend cultures were much lower than those for
the pure cultures. Also when Abu-Tarboush (1996) studied the action of various strains of
commercial cultures in whole camel milk, he displayed that the last pH of the blends L.
bulgaricus 12 with the other isolates of S. thermophilus gave lower pH comparing with isolates
alone at 42ºC for 4h. The existing results here show that the pH of the fermented raw milk by
the isolated starters ranged from 4.20 to 4.98 which is compatible with (Lore et al., 2005)
results that described in suusac (pH 4.30), but higher than the results described in garris (pH
3.25 - 3.40) a Sudanese traditional fermented camel milk.
Table 4.3: Effect of culture composition on acidification characteristics of pure &
mixed cultures grown on milk at different times on 42ºC.
Isolated strains
pH during incubation time
0h 2h 4h 6h 8h 24h
Lb. bulgaricus A1 6.39 6.28 5.90 5.44 4.98 4.88
Lb. bulgaricus A2 6.39 6.24 5.86 4.73 4.71 4.67
S. thermophilus B1 6.39 6.33 5.73 5.17 4.88 4.80
S. thermophilus B2 6.40 6.38 6.11 5.29 5.01 4.98
S. thermophilus B3 6.38 6.35 6.05 5.15 4.99 4.85
Mix (A1:B1) 6.39 5.99 5.45 4.88 4.68 4.64
Mix (A1:B2) 6.39 6.20 4.90 4.71 4.63 4.57
Mix (A1:B3) 6.40 6.21 5.16 4.83 4.67 4.61
Mix (A2:B1) 6.40 6.12 5.02 4.83 4.67 4.40
Mix (A2:B2) 6.41 6.14 5.10 4.61 4.58 4.50
Mix (A2:B3) 6.39 6.17 4.93 4.41 4.38 4.20
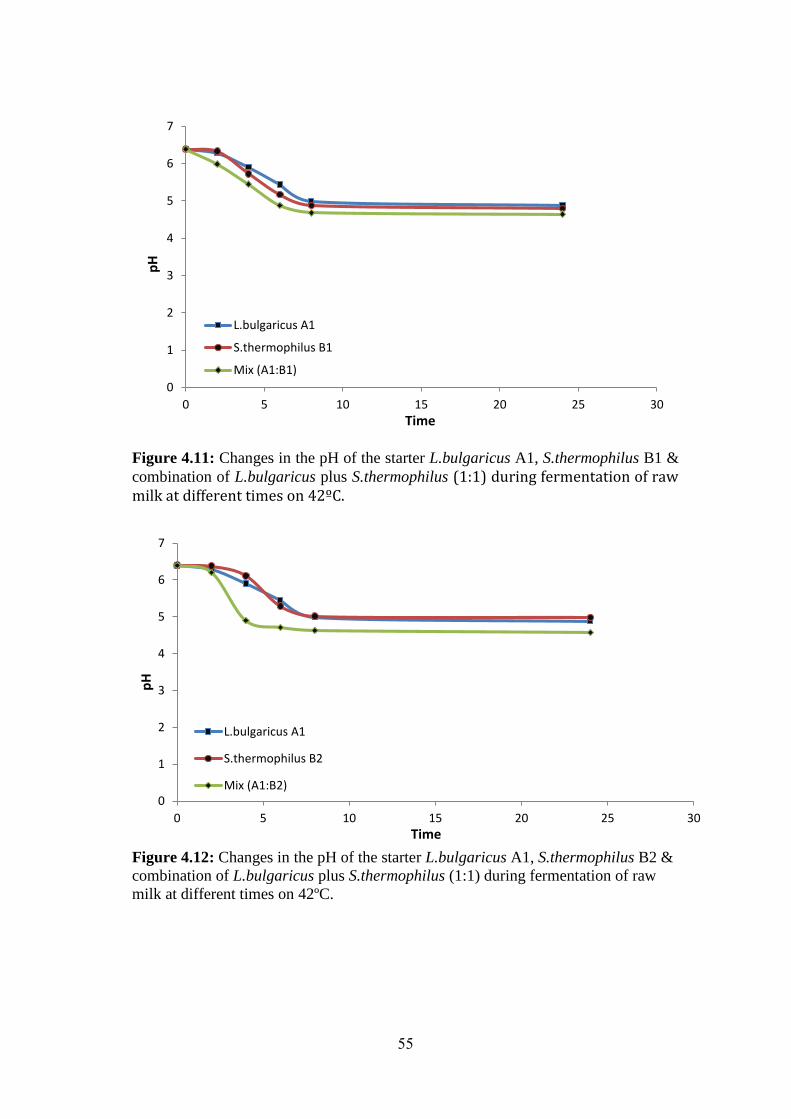
55
Figure 4.11: Changes in the pH of the starter L.bulgaricus A1, S.thermophilus B1 &
combination of L.bulgaricus plus S.thermophilus (1:1) during fermentation of raw milk at different times on 42ºC.
Figure 4.12: Changes in the pH of the starter L.bulgaricus A1, S.thermophilus B2 &
combination of L.bulgaricus plus S.thermophilus (1:1) during fermentation of raw
milk at different times on 42ºC.
0
1
2
3
4
5
6
7
0 5 10 15 20 25 30
L.bulgaricus A1
S.thermophilus B1
Mix (A1:B1)
pH
Time
0
1
2
3
4
5
6
7
0 5 10 15 20 25 30
L.bulgaricus A1
S.thermophilus B2
Mix (A1:B2)
pH
Time
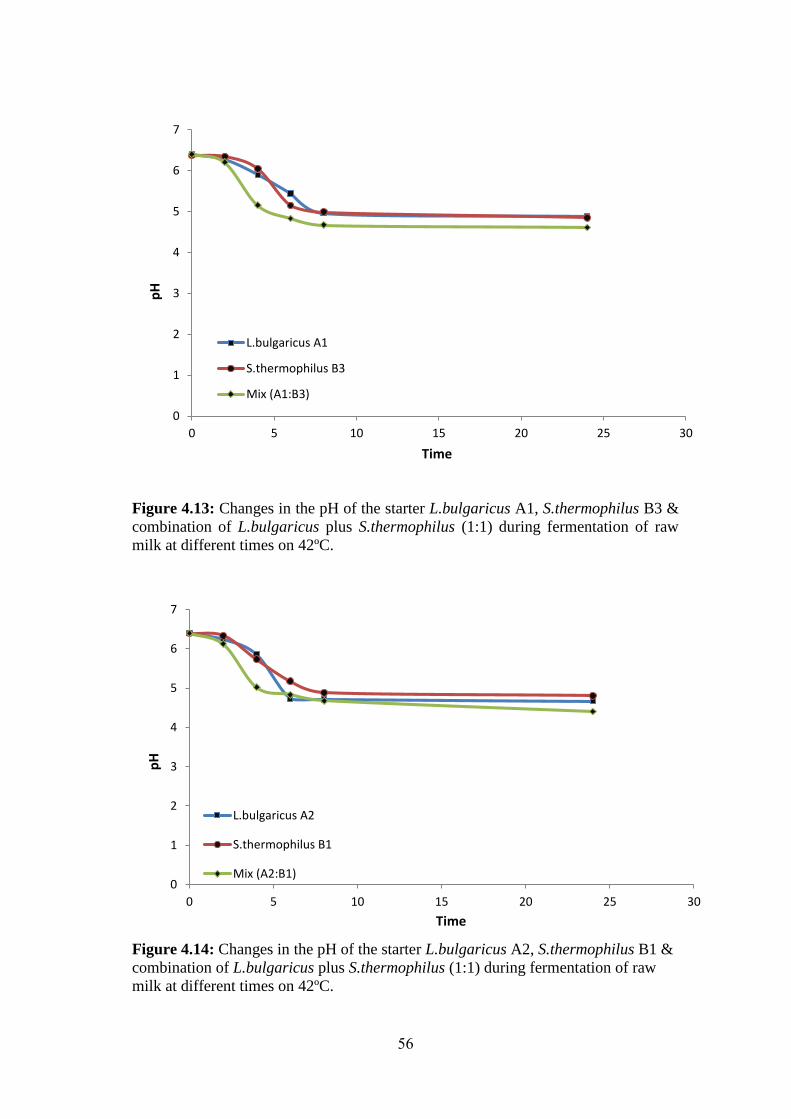
56
Figure 4.13: Changes in the pH of the starter L.bulgaricus A1, S.thermophilus B3 &
combination of L.bulgaricus plus S.thermophilus (1:1) during fermentation of raw
milk at different times on 42ºC.
Figure 4.14: Changes in the pH of the starter L.bulgaricus A2, S.thermophilus B1 &
combination of L.bulgaricus plus S.thermophilus (1:1) during fermentation of raw
milk at different times on 42ºC.
0
1
2
3
4
5
6
7
0 5 10 15 20 25 30
L.bulgaricus A1
S.thermophilus B3
Mix (A1:B3)
pH
Time
0
1
2
3
4
5
6
7
0 5 10 15 20 25 30
L.bulgaricus A2
S.thermophilus B1
Mix (A2:B1)
pH
Time
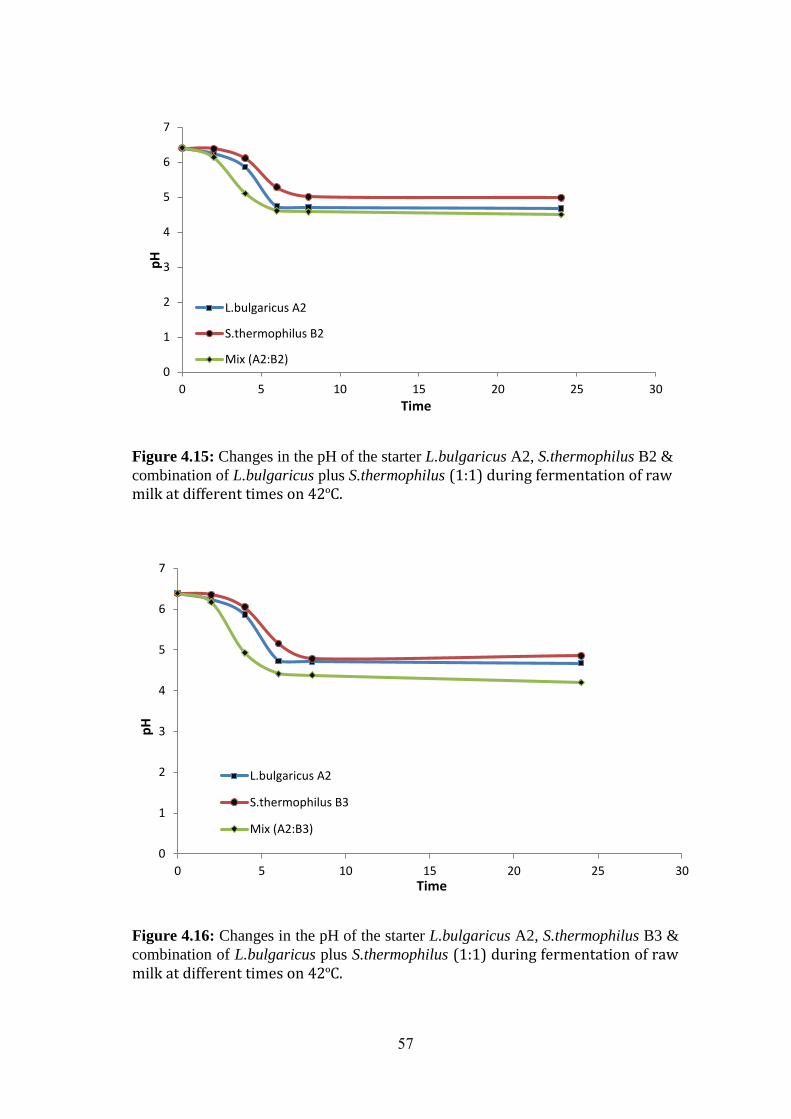
57
Figure 4.15: Changes in the pH of the starter L.bulgaricus A2, S.thermophilus B2 &
combination of L.bulgaricus plus S.thermophilus (1:1) during fermentation of raw milk at different times on 42ºC.
Figure 4.16: Changes in the pH of the starter L.bulgaricus A2, S.thermophilus B3 &
combination of L.bulgaricus plus S.thermophilus (1:1) during fermentation of raw milk at different times on 42ºC.
0
1
2
3
4
5
6
7
0 5 10 15 20 25 30
L.bulgaricus A2
S.thermophilus B2
Mix (A2:B2)
pH
Time
0
1
2
3
4
5
6
7
0 5 10 15 20 25 30
L.bulgaricus A2
S.thermophilus B3
Mix (A2:B3)
pH
Time
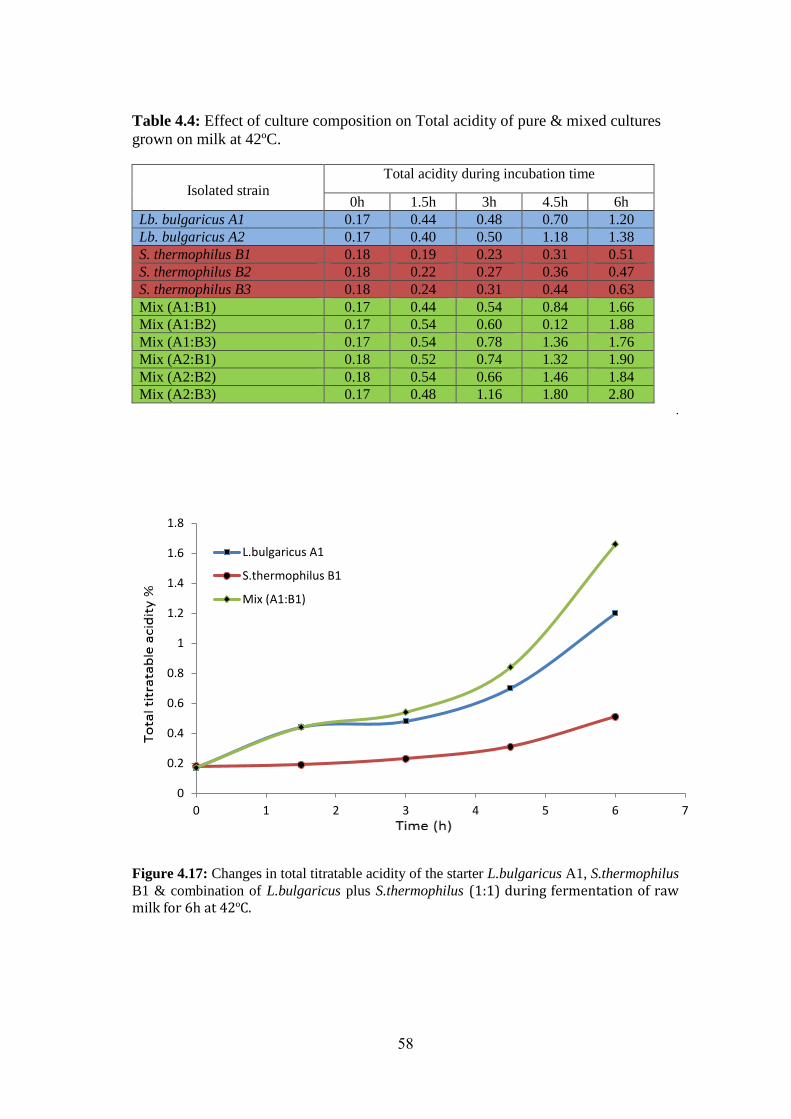
58
Table 4.4: Effect of culture composition on Total acidity of pure & mixed cultures
grown on milk at 42ºC.
Total acidity during incubation time
Isolated strain 6h 4.5h 3h 1.5h 0h
1.20 0.70 0.48 0.44 0.17 Lb. bulgaricus A1
1.38 1.18 0.50 0.40 0.17 Lb. bulgaricus A2
0.51 0.31 0.23 0.19 0.18 S. thermophilus B1
0.47 0.36 0.27 0.22 0.18 S. thermophilus B2
0.63 0.44 0.31 0.24 0.18 S. thermophilus B3
1.66 0.84 0.54 0.44 0.17 Mix (A1:B1)
1.88 0.12 0.60 0.54 0.17 Mix (A1:B2)
1.76 1.36 0.78 0.54 0.17 Mix (A1:B3)
1.90 1.32 0.74 0.52 0.18 Mix (A2:B1)
1.84 1.46 0.66 0.54 0.18 Mix (A2:B2)
2.80 1.80 1.16 0.48 0.17 Mix (A2:B3) .
Figure 4.17: Changes in total titratable acidity of the starter L.bulgaricus A1, S.thermophilus
B1 & combination of L.bulgaricus plus S.thermophilus (1:1) during fermentation of raw milk for 6h at 42ºC.
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
0 1 2 3 4 5 6 7
L.bulgaricus A1
S.thermophilus B1
Mix (A1:B1)
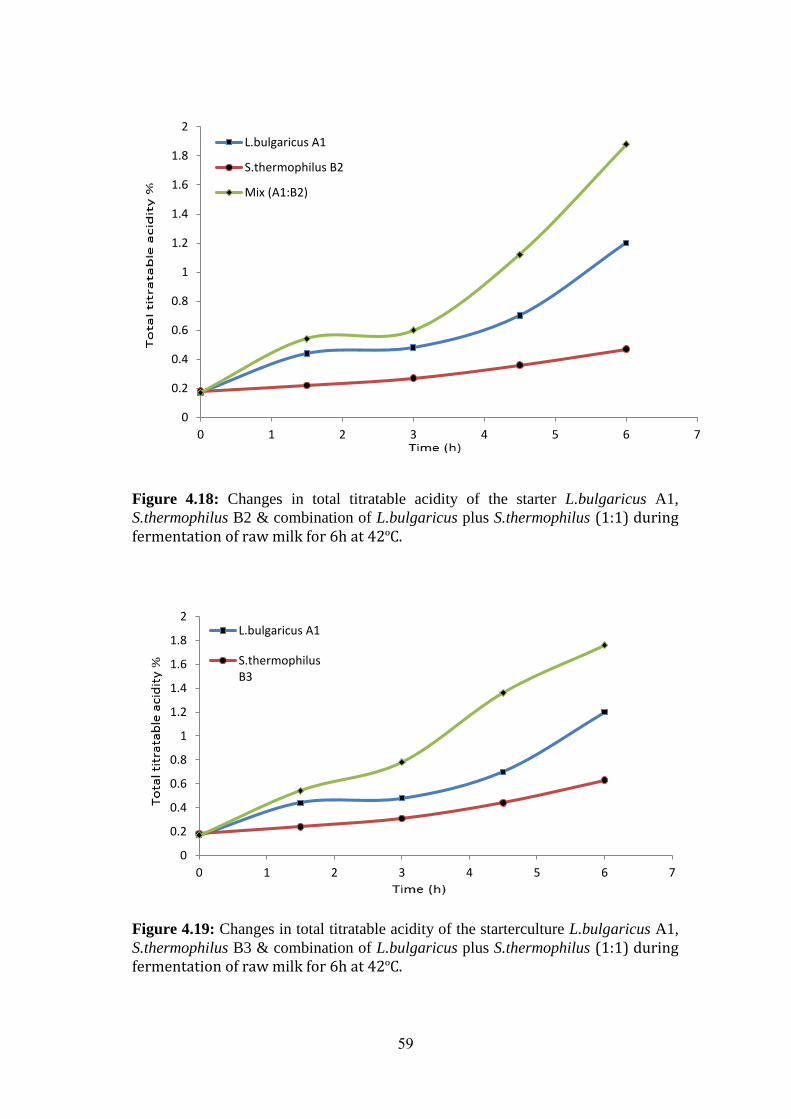
59
Figure 4.18: Changes in total titratable acidity of the starter L.bulgaricus A1,
S.thermophilus B2 & combination of L.bulgaricus plus S.thermophilus (1:1) during fermentation of raw milk for 6h at 42ºC.
Figure 4.19: Changes in total titratable acidity of the starterculture L.bulgaricus A1,
S.thermophilus B3 & combination of L.bulgaricus plus S.thermophilus (1:1) during fermentation of raw milk for 6h at 42ºC.
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
0 1 2 3 4 5 6 7
L.bulgaricus A1
S.thermophilus B2
Mix (A1:B2)
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
0 1 2 3 4 5 6 7
L.bulgaricus A1
S.thermophilusB3
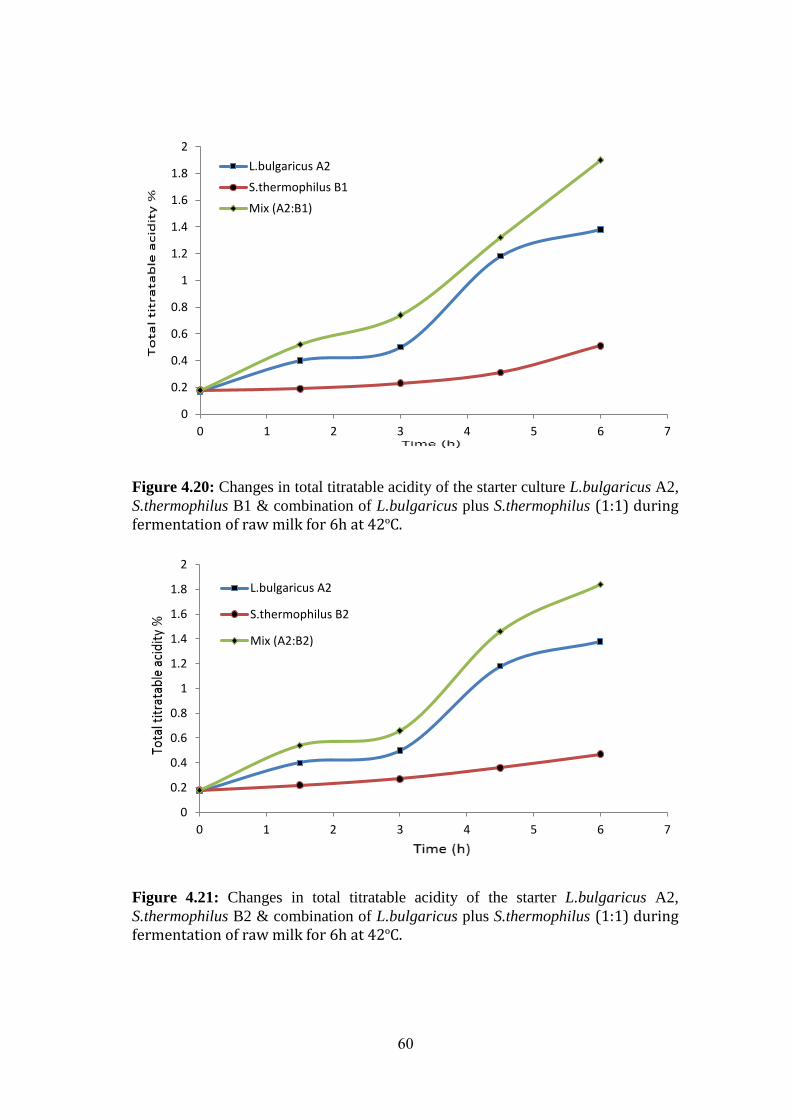
60
Figure 4.20: Changes in total titratable acidity of the starter culture L.bulgaricus A2,
S.thermophilus B1 & combination of L.bulgaricus plus S.thermophilus (1:1) during fermentation of raw milk for 6h at 42ºC.
Figure 4.21: Changes in total titratable acidity of the starter L.bulgaricus A2,
S.thermophilus B2 & combination of L.bulgaricus plus S.thermophilus (1:1) during fermentation of raw milk for 6h at 42ºC.
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
0 1 2 3 4 5 6 7
L.bulgaricus A2
S.thermophilus B1
Mix (A2:B1)
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
0 1 2 3 4 5 6 7
L.bulgaricus A2
S.thermophilus B2
Mix (A2:B2)
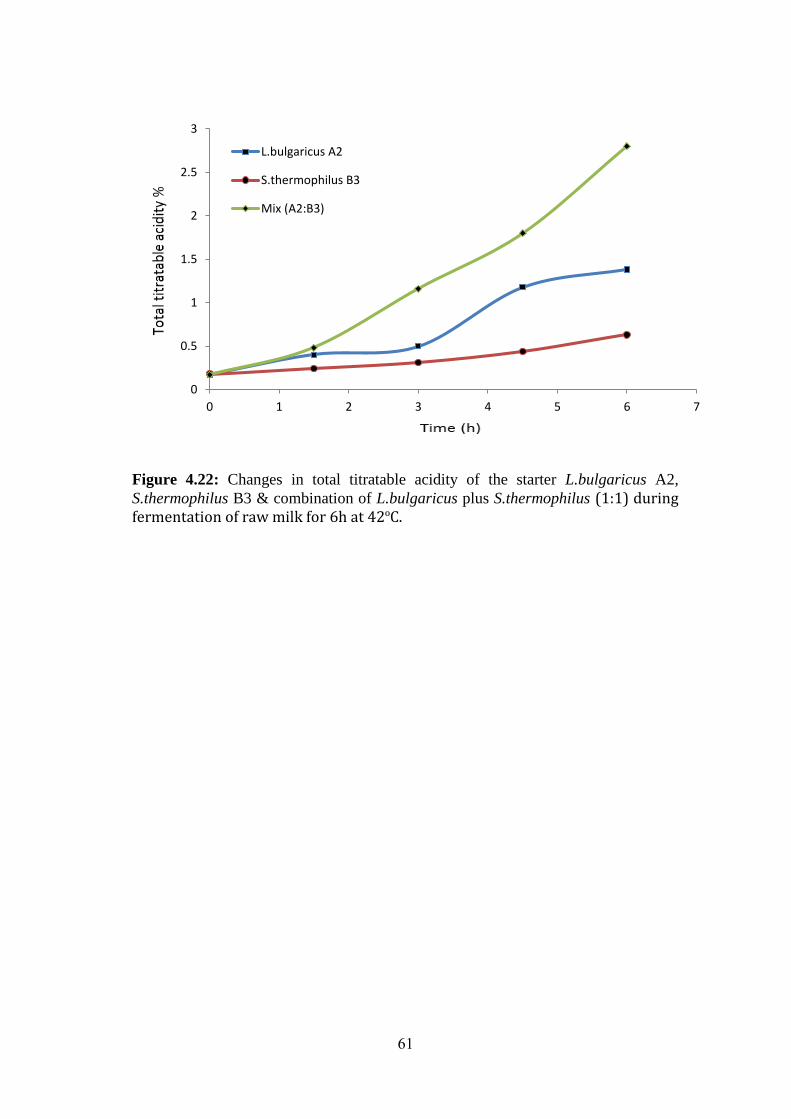
61
Figure 4.22: Changes in total titratable acidity of the starter L.bulgaricus A2,
S.thermophilus B3 & combination of L.bulgaricus plus S.thermophilus (1:1) during fermentation of raw milk for 6h at 42ºC.
0
0.5
1
1.5
2
2.5
3
0 1 2 3 4 5 6 7
L.bulgaricus A2
S.thermophilus B3
Mix (A2:B3)

62
Chapter 5
Conclusions and
Recommendations

63
Chapter 5
Conclusions and Recommendations
Two isolates of L.delbrueckii ssp. bulgaricus and S.thermophilus are selected to
produce starters. These starters with low cost can be used in production of yoghurt at small
and large scale.
The overall data proved the need to conjoint phenotypic and molecular methods for
identifying natural yoghurt starters L.bulgaricus and S.thermophilus which are often
imprecisely identified by phenotypic and biochemical tests only. PCR-ARDRA is
recommended as the tool of choice for identification of homofermentative thermophilic dairy
species or subspecies. The distinguish between isolated cultures by PCR-ARDRA can be used
in combination along with API 50 CHL.
After testing and assessing the isolated strains on yoghurt production, the data
indicated that certain microbial and biochemical changes happen during fermentation of milk.
The results showed that microorganisms differed in growth, acid production and titratable
acidity. Generally, mixed yoghurt culture manifest superior growth, acid production and
titratable acidity than single starter cultures.
Ultimately, Identification the starters of yoghurt, in fact, is the first step for using these
starters in dairy industry. Thus, it should also be screened for their technological parameters.
Technological parameters basically include acidifying activity, aroma compounds production
particularly acetaldehyde and diacetyl, EPS production and resist to bacteriophage invasion.
Determination of such characteristics would be helpful for industrial applications in the future.
Analysis of yoghurt starters are a complicated process and the use of various
techniques, especially molecular-based phenotypic or genomic techniques, are recommended.

64
References

65
The References List
Abdel Moneim ES, Abdalla, AI and Ahmed, EE (2006). Chemical and microbiological
quality of Garris, Sudanese fermented camel's milk product. International Journal of
Food Science Technology, (41): 321-328.
Abu-Tarboush HM (1996). Comparison of associative growth of yogurt starter in whole
milk from camels and cows. Journal of Dairy Science, (79): 366-371.
Ali, A. A. (2011). Isolation and Identification of lactic acid bacteria isolated from
traditional drinking yoghurt in Khartoum State, Sudan. Current Research in
Bacteriology, 4(1), 16-22.
Amanze, K. O., & Amanze, J. O. (2011). Quality evaluation of yoghurt from cowmilk,
soymilk and cow/soymilk. Journal of Research and National Development, 9(2), 44-
47.
Angelov, M., Kostov, G., Simova, E., Beshkova, D., & Koprinkova-Hristova, P. (2009).
Proto-cooperation factors in yogurt starter cultures. Revue Genie Industruiel , (3), 4-
12.
Angelov, M., Kostov, G., Simova, E., Beshkova, D., & Koprinkova-Hristova, P. (2009).
Oxygen influence in the mutual metabolism of S. thermophilus and Lb. bulgaricus in
yogurt starter cultures. Revue Genie Industruiel , (3), 13-23.
Anukam, K. C., & Reid, G. (2007). Probiotics: 100 years (1907-2007) after Elie
Metchnikoff’s observation. Communicating current research and educational topics
and trends in applied microbiology, 1, 466-474.
Arslan, S., & Bayrakci, S. (2016). Physicochemical, functional, and sensory properties of
yogurts containing persimmon. Turkish Journal of Agriculture and Forestry, 40(1),
68-74.
Batmanglij, N. (2007). A taste of Persia: an introduction to Persian cooking. IB Tauris.
Caplice, E., & Fitzgerald, G. F. (1999). Food fermentations: role of microorganisms in
food production and preservation. International journal of food microbiology, 50(1),
131-149.
Carrasco MS, Scarinci EH, Simonetta AC (2005). Associative growth of lactic acid
bacteria for cheese starter: Acidifying and proteolytic activities and redox potential
development. Journal of Food and Agriculture Environment, 3(2): 116-119
Chagnaud, P., Machinis, K., Coutte, L. A., Marecat, A., & Mercenier, A. (2001). Rapid
PCR-based procedure to identify lactic acid bacteria: application to six common
Lactobacillus species. Journal of Microbiological Methods, 44(2), 139-148.

66
Chandan, R. C. (2014). 18 Dairy–Fermented Products. Food Processing: Principles and
Applications, Seconed Edition, 405-436.
Chandan, R. C., & O’Rell, K. R. (2006). Principles of yogurt processing (pp. 195-197).
Oxford, England: Blackwell Publishing.
Chandan, R. C., & Nauth, K. R. (2012). Chapter 12: Yogurt. YH Hui (Administrative
Associate Ed), EO Evranuz (Associate Ed), RC Chandan, L Cocolin, EH Drosinos, L
Goddick, A Rodrigues, F Toldra (Eds), Handbook of Animal-Based Food and
Beverage Technology. CRC Press, Boca Raton, FL, 213-233.
Chandan, R. C., & Shahani, K. M. (1995). Other fermented dairy products. Biotechnology
Set, Second Edition, (9), 385-418.
Coeuret, V., Dubernet, S., Bernardeau, M., Gueguen, M., & Vernoux, J. P. (2003).
Isolation, characterisation and identification of lactobacilli focusing mainly on cheeses
and other dairy products. Le Lait, 83(4), 269-306.
Courtin, P., & Rul, F. (2004). Interactions between microorganisms in a simple ecosystem:
yogurt bacteria as a study model. Le Lait, 84(1-2), 125-134.
Deasy, B. M., Rea, M. C., Fitzgerald, G. F., Cogan, T. M., & Beresford, T. P. (2000). A
rapid PCR based method to distinguish between Lactococcus and
Enterococcus. Systematic and Applied Microbiology, 23(4), 510-522.
Dellaglio, F., & Felis, G. E. (2005). Taxonomy of lactobacilli and
bifidobacteria. Probiotics and prebiotics: scientific aspects, 25-50. Edited by G.W.
Tannock, Caister Academic press, Norfolk, UK.
Delley, M., & Germond, J. E. (2002). Differentiation of Lactobacillus helveticus,
Lactobacillus delbrueckii subsp bulgaricus, subsp lactis and subsp delbrueckii Using
Physiological and Genetic Tools and Reclassification of some Strains from the ATCC
Collection. Systematic and applied microbiology, 25(2), 228-231.
Donovan, M. (2006). On food and cooking: the science and lore of the kitchen.
Gastronomica, 6(4),117.
Donovan, S. M., & Shamir, R. (2014). Introduction to the yogurt in nutrition initiative and
the First Global Summit on the health effects of yogurt. The American journal of
clinical nutrition, 99(5), 1209S-1211S.
Doyle, M. P., & Meng, J. (2006). Bacteria in food and beverage production. In The
prokaryotes (pp. 797-811). Springer New York.
Drake, M., Small, C. L., Spence, K. D., & Swanson, B. G. (1996). Rapid detection and
identification of Lactobacillus spp. in dairy products by using the polymerase chain
reaction. Journal of Food Protection, 59(10), 1031-1036.

67
Driessen, F. M., Kingma, F., & Stadhouders, J. (1982). Evidence that Lactobacillus
bulgaricus in yogurt is stimulated by carbon dioxide produced by Streptococcus
thermophilus. Netherlands Milk and Dairy Journal (Netherlands), (36),135-144.
Durso, L., Hutkins, R., 2003. “Starter Cultures”, in Encyclopedia of Food Science and
Nutrition, edited by B. Caballero, L. Trugo, P. Finglas (Academic Press, United
Kingdom), pp. 5583-5593.
El Bashiti, T. (2010). Production of Yogurt by locally isolated starters: Streptococcus
thermophilus and Lactobacillus bulgaricus. Journal of Al Azhar University–Gaza, 12,
56-58.
Erkmen, O., & Bozoglu, T. F. (2016). Food Microbiology: Principles into Practice. John
Wiley & Sons.
Erkuş, O. (2007). Isolation, phenotypic and genotypic charecterization of yoghurt starter
bacteris (Master’s Thesis, Izmir Institute of Technology).
Estrada, A., Coates Estrada, R., Meritt Jr, D., Bojic Bultrini, D., Klein, A. M., Steffan
Dewenter, I., ... & Argenti, O. (2011). World livestock 2011: livestock in food
security (No. FAO 363.8 W927 2011). FAO, Roma (Italia).
Fisberg, M., & Machado, R. (2015). History of yogurt and current patterns of
consumption. Nutrition reviews, 73(suppl 1), 4-7.
Flint, S. H., Ward, L. J., & Brooks, J. D. (1999). Streptococcus waius sp. nov., a
thermophilic streptococcus from a biofilm. International Journal of Systematic and
Evolutionary Microbiology, 49(2), 759-767.
Forbes, B. A., Sahm, D. F., Weissfeld, A. S., Scott, E. G., & Bailley, W. R. (2002).
Diagnostic microbiology. Bailey & Scott1s Diagnostic Microbiology, 11(1), 11-14.
Fortina, M. G., Ricci, G., Mora, D., Parini, C., & Manachini, P. L. (2001). Specific
identification of Lactobacillus helveticus by PCR with pepC, pepN and htrA targeted
primers. FEMS microbiology letters, 198(1), 85-89.
Gandhi, D. N. (2006). Food and Industrial Microbiology: Microbiology of Fermented
Dairy Products. Principal Scientist Dairy Microbiology Division National Dairy
Research Institute, Karnal.
Giraffa, G., De Vecchi, P., & Rossetti, L. (1998). Note: identification of Lactobacillus
delbrueckii subspecies bulgaricus and subspecies lactis dairy isolates by amplified
rDNA restriction analysis. Journal of Applied Microbiology, 85(5), 918-918.
Grobben, G. J., Smith, M. R., Sikkema, J., & De Bont, J. A. M. (1996). Influence of
fructose and glucose on the production of exopolysaccharides and the activities of
enzymes involved in the sugar metabolism and the synthesis of sugar nucleotides in

68
Lactobacillus delbrueckii subsp. bulgaricus NCFB 2772. Applied Microbiology and
Biotechnology, 46(3), 279-284.
Hammes, W. P., & Vogel, R. F. (1995). The genus lactobacillus. In The genera of lactic
acid bacteria (pp. 19-54). Springer, Boston, MA.
Harrigan, W. F., & McCance, M. E. (1976). Laboratory methods in food and dairy
microbiology. Academic Press Inc.(London) Ltd.
Hébert, E. M., Raya, R. R., Tailliez, P., & de Giori, G. S. (2000). Characterization of
natural isolates of Lactobacillus strains to be used as starter cultures in dairy
fermentation. International journal of food microbiology, 59(1), 19-27.
https:// www.microbewiki.kenyon.edu/index.php/yogurt.
Hutkins, R. W. (2008). Microbiology and technology of fermented foods (Vol. 22). John
Wiley & Sons.
Igarashi, T., Yamamoto, A., & Goto, N. (1996). Direct detection of Streptococcus mutans
in human dental plaque by polymerase chain reaction. Oral microbiology and
immunology, 11(5), 294-298.
Kermanshahi, R. K., & Peymanfar, S. (2012). Isolation and identification of lactobacilli
from cheese, yoghurt and silage by 16S rDNA gene and study of bacteriocin and
biosurfactant production. Jundishapur journal of Microbiology, 5(4), 528-532.
Khaled, A. D., Neilan, B. A., Henriksson, A., & Conway, P. L. (1997). Identification and
phylogenetic analysis of Lactobacillus using multiplex RAPD-PCR. FEMS
microbiology letters, 153(1), 191-197.
Kilara, A., & Chandan, R. C. (2013). Greek‐style yogurt and related
products. Manufacturing Yogurt and Fermented Milks, 297-318.
Kosikowski, F., & Mistry, V. V. (1977). Cheese and fermented milk foods.
Kristo, E., Biliaderis, C. G., & Tzanetakis, N. (2003). Modelling of the acidification
process and rheological properties of milk fermented with a yogurt starter culture using
response surface methodology. Food Chemistry, 83(3), 437-446.
Lore TA, Mbugua SK, Wangoh J (2005). Enumeration and identification of microflora in
Suusac, a Kenyan traditional fermented camel milk product. Lebensmittel-
Wisseschaft Technol. 38(2): 125-130.
Louailèche, H., Bracquart, P., Guimont, C., & Linden, G. (1996). Carbon dioxide fixation
by cells and cell-free extracts of. Journal of dairy research, 63, 321-325.

69
Louaileche, H., Bracquart, P., Saulnier, F., Desmazeaud, M., & Linden, G. (1993). Carbon
dioxide effects on the growth and metabolites of morphological variants of
Streptococcus thermophilus. Journal of dairy science, 76(12), 3683-3689.
Mac Faddin, J. F. (1976). Biochemical tests for identification of medical bacteria.
Williams & Wilkins Co.
Markiewicz, L. H., Wasilewska, E., & Bielecka, M. (2008). Identification of Lactobacillus
strains present in fermented dairy products and their differentiation using molecular
methods. Polish Journal of Food and Nutrition Sciences, 58(2).
Marshall, V. M. (1987). Lactic acid bacteria: starters for flavour. FEMS Microbiology
Reviews, 3(3), 327-336.
Mathara, J. M., Schillinger, U., Kutima, P. M., Mbugua, S. K., & Holzapfel, W. H. (2004).
Isolation, identification and characterisation of the dominant microorganisms of kule
naoto: The Maasai traditional fermented milk in Kenya. International journal of food
microbiology, 94(3), 269-278.
Mirghani AA (1994). Micrbiological and biochemical properties of fermented camel milk
(gariss). M.Sc. Thesis. University of Khartoum. Sudan.
Moreno, A. L., Cervera, R. P., Ortega, A. R., Díaz, M. J., Baladia, E., Basulto, J., ... &
Rodríguez, R. E. (2012). [Scientific evidence about the role of yogurt and other
fermented milks in the healthy diet for the Spanish population]. Nutricion
hospitalaria, 28(6), 2039-2089.
Papadelli, M., Manolopoulou, E., Kalantzopoulos, G., & Tsakalidou, E. (2003). Rapid
detection and identification of Streptococcus macedonicus by species-specific PCR
and DNA hybridisation. International journal of food microbiology, 81(3), 231-239.
Quere, F., Deschamps, A., & Urdaci, M. C. (1997). DNA probe and PCR-specific reaction
for Lactobacillus plantarum. Journal of Applied Microbiology, 82(6), 783-790.
Rajagopal, S. N., & Sandine, W. E. (1990). Associative growth and proteolysis of
Streptococcus thermophilus and Lactobacillus bulgaricus in skim milk. Journal of
dairy science, 73(4), 894-899.
Schmidt, K. A. (2004). Dairy: Ice cream. Food processing–Principles and applications,
287-296.
Sfakianakis, P., & Tzia, C. (2014). Conventional and innovative processing of milk for
yogurt manufacture; development of texture and flavor: a review. Foods, 3(1), 176-
193.
Sharpe, M. E., Fryer, T. F., & Smith, D. G. (1979). Identification of the lactic acid
bacteria. Identification methods for microbiologists, 157, 233-259.

70
Soukoulis, C., Panagiotidis, P., Koureli, R., & Tzia, C. (2007). Industrial yogurt
manufacture: monitoring of fermentation process and improvement of final product
quality. Journal of dairy science, 90(6), 2641-2654.
Tabasco, R., Paarup, T., Janer, C., Peláez, C., & Requena, T. (2007). Selective
enumeration and identification of mixed cultures of Streptococcus thermophilus,
Lactobacillus delbrueckii subsp. bulgaricus, L. acidophilus, L. paracasei subsp.
paracasei and Bifidobacterium lactis in fermented milk. International Dairy
Journal, 17(9), 1107-1114.
Tamime, A. Y. (1997). Some aspects of the production of yoghurt and condensed
yoghurt (unpublished Doctoral dissertation, University of Reading).
Tamime, A. Y., & Robinson, R. K. (1999). Microbiology of yoghurt and “bio” starter
cultures. Yoghurt science and technology, 2, 389-431.
Tamime, A. Y., & Robinson, R. K. (2007). Ch. 2, Background to manufacturing
practice. Tamime and Robinson's yoghurt: science and technology, (Ed. 3), 13-161.
Tammam, J. D., Williams, A. G., Noble, J., & Lloyd, D. (2000). Amino acid fermentation
in non‐starter Lactobacillus spp. isolated from Cheddar cheese. Letters in applied
microbiology, 30(5), 370-374.
Tanigawa, K., & Watanabe, K. (2011). Multilocus sequence typing reveals a novel
subspeciation of Lactobacillus delbrueckii. Microbiology, 157(3), 727-738.
Tilsala-Timisjärvi, A., & Alatossava, T. (1997). Development of oligonucleotide primers
from the 16S-23S rRNA intergenic sequences for identifying different dairy and
probiotic lactic acid bacteria by PCR. International journal of food
microbiology, 35(1), 49-56.
Trachoo, N. (2002). Yogurt: The fermented milk. Songklanakarin J. Sci. Technol, 24(4),
726-736.
Vadeboncoeur, C., & Moineau, S. (2004). The relevance of genetic analysis to dairy
bacteria: building upon our heritage. Microbial cell factories, 3(1), 15.
Vaningelgem, F., Zamfir, M., Adriany, T., & De Vuyst, L. (2004). Fermentation
conditions affecting the bacterial growth and exopolysaccharide production by
Streptococcus thermophilus ST 111 in milk‐based medium. Journal of applied
microbiology, 97(6), 1257-1273.
Vaningelgem, F., Zamfir, M., Mozzi, F., Adriany, T., Vancanneyt, M., Swings, J., & De
Vuyst, L. (2004). Biodiversity of exopolysaccharides produced by Streptococcus
thermophilus strains is reflected in their production and their molecular and functional
characteristics. Applied and environmental microbiology, 70(2), 900-912.

71
Vedamuthu, E. R. (2006). Starter cultures for yogurt and fermented milks. Manufacturing
yogurt and fermented milks, 89.
Ward, L. J., Brown, J. C., & Davey, G. P. (1998). Two methods for the genetic
differentiation of Lactococcus lactis ssp. lactis and cremoris based on differences in
the 16S rRNA gene sequence. FEMS microbiology letters, 166(1), 15-20.
Weerathilake, W. A. D. V., Rasika, D. M. D., Ruwanmali, J. K. U., & Munasinghe, M. A.
D. D. (2014). The evolution, processing, varieties, and health benefits of
yoghurt. International Journal of Scientific and Research Publications, 4(4), 1-10.
White, C. H., Kilara, A., & Hui, Y. H. (2008). Manufacturing yogurt and fermented milks.
R. C. Chandan (Ed.). John Wiley & Sons.
Zhang, T., Zhang, C., Li, S., Zhang, Y., & Yang, Z. (2011). Growth and
exopolysaccharide production by Streptococcus thermophilus ST1 in skim
milk. Brazilian Journal of Microbiology, 42(4), 1470-1478.

72
Appendixes

73
APPENDIX A
Recipes for Culture Media and Biochemical Tests
Table (A.1): MRS medium
g/l Ingredients
10.0 Peptone
10.0 Meat extract
5.0 Yeast Extract
20.0 D(-) Glucose
1ml Tween 80
2 K2HPO4
5.0 Sodium acetate
2.0 Triammonium citrate
0.2 MgSO4.7H2O
0.05 MnSO4.4H2O
Modified MRS Broth For Testing The Growth At Different NaCl
Concentrations
0.04 Bromocresol purple
20,40,65 for the concentration of 2%,
4% and 6.5%
NaCl
Modified MRS Broth For Testing The Growth At Different Temperatures
0.04 Bromocresol purple
MRS Agar
15 Agar
Final pH: 6.3
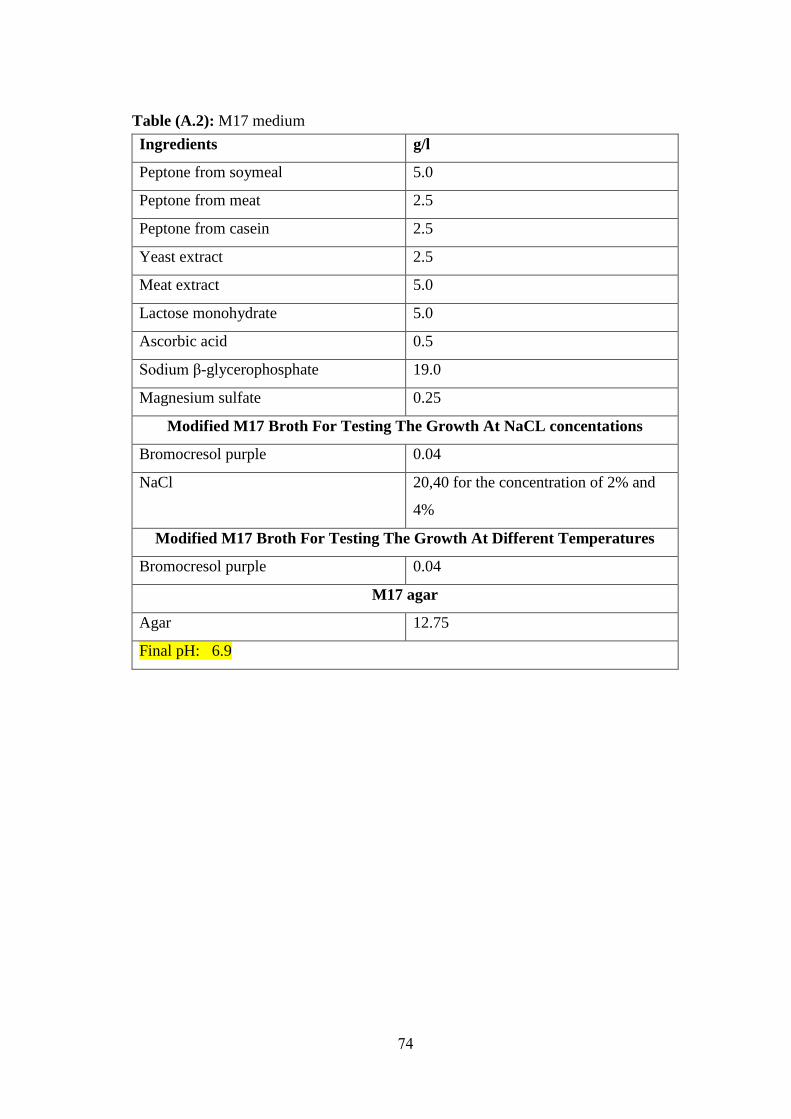
74
Table (A.2): M17 medium
g/l Ingredients
5.0 Peptone from soymeal
2.5 Peptone from meat
2.5 Peptone from casein
2.5 Yeast extract
5.0 Meat extract
5.0 Lactose monohydrate
0.5 Ascorbic acid
19.0 Sodium β-glycerophosphate
0.25 Magnesium sulfate
Modified M17 Broth For Testing The Growth At NaCL concentations
0.04 Bromocresol purple
20,40 for the concentration of 2% and
4%
NaCl
Modified M17 Broth For Testing The Growth At Different Temperatures
0.04 Bromocresol purple
M17 agar
12.75 Agar
Final pH: 6.9

75
APPENDIX B Isolation of Genomic DNA from Gram positive and Gram negative bacteria

76
APPENDIX C
C1: Buffers and Stock Solutions for Molecular Characterization
1X TAE
20ml of 50 X TAE buffer was taken and the volume was adjusted to 1 L with
deionized water. The 1x working solution was 40 mM Tris-acetate/1 mM EDTA.
Ethidium Bromide Stock Solution (10mg/ml)
0.5g ethidium bromide was dissolved in 50ml deionized water and the solution
was stored in dark bottle at room temperature.
C2: PCR recipes
PCR Mixture
Mg free Taq DNA polymerase buffer 5μl
MgCl2 (25Mm) 3μl
Sterile deionized water 32μl
Oligo forward 10 picomole/μl 1μl
Oligo reverse 10 picomole/μl 1μl
dNTP (2 mM each) 5μl
dNTP (10X)
Twenty microliters of each 100mM dATP, dCTP, dGTP, and dTTP are taken and mixed in
an eppendorf tube. 920μl of sterile deionized water was added to a final concentration of
2mM. Solution was mixed gently and stored at -20°C.
C3: Restriction Enzymes Used for Molecular Characterization
Restriction Enzyme Reaction Mixture
Restriction enzyme buffer 5μl
Sterile deionized water 34.5μl
Restriction Enzyme 0.5μl (from 5U)
DNA 10μl