Embed Size (px)
Citation preview

energies
Article
Isolation and Characterization of Native Microalgaefrom the Peruvian Amazon with Potential forBiodiesel Production
Marianela Cobos 1 Jae D Paredes 1 J Dylan Maddox 23 Gabriel Vargas-Arana 1Leenin Flores 4 Carla P Aguilar 4 Jorge L Marapara 5 and Juan C Castro 5
1 Laboratorio de Biotecnologiacutea y Bioenergeacutetica Universidad Cientiacutefica del Peruacute (UCP) Av AbelardoQuintildeones km 25 Iquitos 6024 Peru jparedes92hotmailcom (JDP) gvargasucpedupe (GV-A)
2 Pritzker Laboratory for Molecular Systematics and Evolution The Field Museum of Natural HistoryChicago IL 60605 USA jdylanmaddoxgmailcom
3 Environmental Sciences American Public University System Charles Town WV 25414 USA4 Laboratorio de Biotecnologiacutea Acuaacutetica Instituto del Mar del Peruacute (IMARPE) Esquina Gamarra y General
Valle SN Chucuito Callao 07021 Peru lfloresimarpegobpe (LF) caguilarimarpegobpe (CPA)5 Unidad Especializada de Biotecnologiacutea Centro de Investigaciones de Recursos Naturales de la
Amazoniacutea (CIRNA) Universidad Nacional de la Amazoniacutea Peruana (UNAP) Psje Los Paujiles SNIquitos 16024 Peru jlmdelahotmailcom
Correspondence mcobosucpedupe (MC) juanccgomezyahooes (JCC)Tel +51-065-261074 (MC) +51-065-263569 (JCC)
Academic Editor Thomas E AmidonReceived 10 December 2016 Accepted 8 February 2017 Published 14 February 2017
Abstract Biodiesel production from microalgae triacylglycerols is growing because this feedstockis a more sustainable and advantageous alternative In this study we isolated and identifiedfourteen strains of native microalgae from the Peruvian Amazon These strains showed greatheterogeneity in biomass productivity lipid productivity and lipid content and thus three ofthem (Acutodesmus obliquus Ankistrodesmus sp and Chlorella lewinii) were selected for furtherevaluation under culture of nitrogen-sufficient (+N) and nitrogen-deficient (minusN) Chu mediumNo 10 These microalgae species showed modifications in biomolecule content (protein lipidand carbohydrate) with a pronounced increase of lipids and carbohydrate and a decrease ofprotein content under stress culture Furthermore the fatty acid profile was peculiar for eachspecies and these patterns showed evident changes particularly in the proportion of saturated andmonounsaturated fatty acids The results of this research suggest that the isolated native microalgaefrom the Peruvian Amazon could be suitable candidates for biodiesel production
Keywords biodiesel fatty acid profiling lipid content oleaginous microalgae Peruvian Amazon
1 Introduction
The global increase of energy demand the depletion and increasing of the costs of fossil fuels [1]and climate change [2] are problems that require urgent solutions In order to mitigate these problemsPeru has legal dispositions (Law 28054 of the promotion of the biofuel market and their regulation withSupreme Decree No 013-2005-EM) so that the energy matrix of the country depends on renewableresources [34] These legal dispositions have increased the importation of biodiesel due to insufficientnational production which is currently obtained from plant oils such as Jatropha curcas ldquowhite pinionrdquoand Elaeis guineensis ldquopalmrdquo both cultured in large areas of the Peruvian Amazon [5]
Biodiesel obtained from crop-based plant oils however has major drawbacks These include lowyields high land and water requirements detrimental effects on food supplies and associated extensive
Energies 2017 10 224 doi103390en10020224 wwwmdpicomjournalenergies
Energies 2017 10 224 2 of 16
deforestation of the rainforest which pose threats to native biodiversity and ecosystem functionsgoods and services [6ndash8] These limitations can be overcome by the next generation of microalgae-basedbiofuels Compared to terrestrial crops the main advantages of microalgal systems are that they havea higher photon conversion efficiency can be harvested batch-wise nearly all-year-round can utilizesalt and waste water streams can couple CO2-neutral fuel production with CO2 sequestration andproduce non-toxic and highly biodegradable biofuels [7]
The isolation selection and culturing of robust oleaginous microalgae with desirable attributes(eg high triacylglycerol content appropriate fatty acid profiles high growth rate and resistance toinvasion of local microorganism) however are the principal challenges associated with microalgaebiofuel production [910] Although such microalgae strains are currently unavailable optimismpersists due to the fact that there is an abundance of species in the wild yet to be isolated [11] Indeedonly a few thousand (~50000) are conserved worldwide in microalgal culture collections [912] and ofthose only a few hundred (lt10) have been investigated for their biochemical composition [13]
Consequently the isolation selection biochemical and molecular characterization of microalgaefrom aquatic environments must be a continuous effort of screening for strains with the potentialfor biodiesel production [14] Therefore a sustainable development of microalgal-based biodieseland bioactive chemicals production in Peru requires the bioprospecting and exploitation of nativemicroalgae bioresources as the most viable option The northeastern region of Peru particularly theLoreto region contains a great variety of freshwater aquatic environments (ie rivers lagoons etc)that harbor a rich biodiversity of microalgae In this study 14 strains of native microalgae from thePeruvian Amazon were isolated identified and tested for their biomass productivity lipid productivityand total lipid content In addition the growth profile total carbohydrate total protein total lipidcontent and fatty acids profiles of three selected strains were also investigated and compared undernitrogen-sufficient and -deficient culture conditions
2 Experimental Section
21 Sample Collection and Isolation of Microalgae
The freshwater samples (100 mL) were collected from three river basins in the Loreto Regionof the Peruvian Amazon Amazon River (0341prime06prime prime S 7314prime89prime prime W) Itaya River (0343prime14prime prime S7314prime178prime prime W) and Nanay River (0342prime02prime prime S 7315prime32prime prime W) Microalgae samples (50 mL) wereenriched with 50 mL of Chu medium No 10 [15] The pH was adjusted to 75 with 1 M NaOH beforeautoclaving and the addition of microalgae The microalgal cells were maintained in Chu mediumNo 10 for four weeks in a controlled culture room at 25 plusmn 1 C with 1212-h light-dark cycles using80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 110 rpm Afterinitial cultivation of the mixed cultures unicellular microalgae were subjected to isolation by the cellwashing method [12] Briefly a 20-microL aliquot of the mixed microalgae culture was placed on a sterileglass slide under an inverted microscope to pick up a single microalgae cell with a Pasteur pipettecontaining a cotton-wool filter and connected to a flexible plastic hose Each microalgae cell was thensequentially washed in six 20-microL drops of Chu medium No 10 Subsequently the drop with a singlemicroalgae cell was transferred to a test tube with 1 mL of Chu medium No10 medium followed byinoculation onto Petri plates containing Chu medium No 10 supplemented with 15 (wv) of agarRepeated streaking on the nutrient agar plate and routine microscopic examination ensured the purityof the cultures Single individual colonies appearing on plates (~7 days) were inoculated into Chuliquid medium No 10 and grown for 14 days at the described conditions
22 Morphological Identification of Microalgae
Isolated strains were preliminarily identified using standard morphological features [16]Intracellular lipid droplets of isolated microalgae were detected by the fluorescence emitted byinteracting with Nile Red [17] Briefly 15 mL of microalgae cells culture were harvested by
Energies 2017 10 224 3 of 16
centrifugation at 1900times g for 10 min and washed two times with 1 mL of physiological saline solutionMicroalgal cell pellets were dissolved in Nile Red acetone solution (2 microgmL) and incubated for15 min at 37 C After washing once stained microalgal cells were observed by Carl Zeiss fluorescentmicroscopy and photographed with a digital camera AxioLabA1 AxioCamERc real time 5 s Imageswere obtained at a magnification of 1000times with visible light and epifluorescence (excitation 510ndash560emission 590)
23 Genomic DNA Isolation
Microalgae cells were harvested from 100 mL of liquid culture by centrifugation at 1900times g for10 min Genomic DNA was extracted using a modified version of the CTAB method as describedby Doyle and Doyle [18] Briefly algal cells were completely ground in a mortar containing 50 mgof sterilized sand and 3 mL of extraction buffer (300 mM Tris-HCl pH 80 50 mM ethylenediaminetetraacetic acid (EDTA) 2 M NaCl 2 cetyltrimethylammonium bromide 3 polyvinylpyrrolidone(MW 40000) and 2 β-mercaptoethanol) Homogenized cells were incubated at 70 C for 30 min withgentle inversion every 2 min An equal volume of phenolchloroformisoamyl alcohol (25241 vv)was then added followed by centrifugation at 20000times g The aqueous supernatant was transferred toa new microtube and an equal volume of chloroformisoamyl alcohol (241 vv) was added followedby centrifugation at 20000times g The aqueous supernatant was transferred to a new microtube andmixed with 01 volumes of 3 M sodium acetate (pH 52) and an equal volume of chilled isopropanolwas added to precipitate the DNA at 15000times g Washing was done with 70 alcohol and the air-driedDNA pellet was dissolved in 100 microL of sterilized water treated with RNase A at 40 C for 30 min andthen extracted with chloroformisoamyl alcohol and DNA precipitation with absolute ethanol Finallythe air-dried DNA pellet was dissolved in TE buffer (10 mM Tris-HCl pH 80 1 mM EDTA) and storedat minus20 C Genomic DNA quality and quantity were assessed by standard OD measurement [19]using a Nanodrop 2000 UV-Vis Spectrophotometer (Thermo Fisher Scientific Wilmington DE USA)In order to verify DNA integrity samples were resolved by standard gel electrophoresis in 12agarose gels [19] stained with ethidium bromide and visualized under ultraviolet light (SupplementaryMaterials Figure S1)
24 PCR Amplification and Sequencing of ITS2-rDNA Region
The ITS-2 rDNA gene was amplified according to Kaur et al [20] with minor modifications in20-microL reactions containing 1times PCR buffer 3 mM MgCl2 02 mM dNTPs mix 0025 UmicroL AmplitaqGold 10 microM each of the forward primer SQITS1 (5prime-GAGCATGTCTGCCTCAGC-3prime) and reverseprimer SQITS2 (5prime-GGTAGCCTTGCCTGAGC-3prime) and 20 ng of genomic DNA The general conditionsfor PCR in a thermal cycler Realplex S4 (Eppendorf Hamburg Germany) were 5 min at 94 C for thefirst cycle followed by 30 s at 94 C 30 s at 55 C and 1 min at 72 C for 35 cycles and a final 10-minextension step at 72 C The PCR gene products were separated on 2 agarose gels using 05 times TBEbuffer at 85 volts for 60 min and detected by staining with ethidium bromide (Figure S2) To purify4 microL of the PCR gene product were mixed with 2 microL of ExoSAP-IT (Affymetrix Santa Clara CAUSA) incubated at 37 C for 30 min and 80 C for 15 min and directly sequenced in both directionswith primers SQITS1 and SQITS2 following the instructions of the BigDyereg Terminator v31 CycleSequencing Kit (Applied Biosystemstrade Foster City CA USA) The sequencing products were purifiedby precipitation with ethanolEDTA denatured and resolved on a Genetic Analyzer 3130XL (AppliedBiosystemstrade Foster City CA USA) All sequences presented here were submitted to GenBank [21]
25 Phylogenetic Analysis
The ITS-2 rDNA gene sequences of the isolates were searched for homology in the National Centerfor Biotechnology Information database [22] using BLAST [23] Sequences were edited in BioEdit 725software [24] to trim the non-overlapping 5prime and 3prime regions for the purpose of restricting the multiplesequence alignment within the minimum common region Those edited sequences were aligned using
Energies 2017 10 224 4 of 16
Clustal omega [25] The unrooted phylogenetic tree was constructed using the neighbor-joining (NJ)method [26]] and Kimurarsquos two-parameter algorithm [27] as implemented within the MEGA 606software [28] The bootstrap analysis [29] was used with 1000 replications to test the relative supportfor the branches produced by NJ analysis
26 Microalgae Cultivation Biomass and Total Lipids Measurements
Two hundred milliliters Chu medium No 10 in a 500-mL Erlenmeyer flask were inoculatedwith the microalgae cells (OD680 005) and incubated at 25 plusmn 1 C using 80 microEmiddotmminus2middotsminus1 intensity ofcool-white fluorescent light and continuous agitation at 150 rpm for four weeks In this culture stage allmicroalgae strains were found in the stationary growth phase and centrifuged at 1920times g for 10 min toharvest the microalgae cells The cell pellets were rinsed with 50 mL deionized water centrifuged againdried in an oven at 70 C to a constant weight and the dry weight was determined gravimetricallywith an analytical balance Kern ABJ 220-4NM (Kern amp Sohn GmbH Balingen Germany) The algalpowders were stored at minus20 C for total lipid analysis
Total lipid was extracted using a modified protocol of Bligh and Dyer [30] For each sampleapproximately 50 mg of microalgal powder were mixed with 3 mL of chloroformmethanol (21 vv)and homogenized in an ultrasonic Branson 2510 (Branson Danbury CT USA) for 5 min The mixturewas then centrifuged at 20000times g for 5 min and the supernatant was transferred into a pre-weighedvial Cellular debris was mixed with another 2 mL of chloroformmethanol (21 vv) by vortex andthen centrifuged as described above After that the pooled organic phases were filtered throughsyringe filters of 045 microm evaporated with nitrogen gas and dried at 80 C in a vacuum ovenVacucell 55mdashComfort (MMM Muumlnchen Germany) to a constant weight The lipid content andproductivity of each sample were measured gravimetrically using a semi-micro analytical balance(Sartorius MSU225S-000-DU Foster City CA USA) and calculated as follows [31]
Total lipid ( dry weight) = WLWA times 100
where WL (g) is the weight of the total lipids extracted and WA (g) is the weight of the drymicroalgae biomass
PLipid (mgmiddotLminus1middotdminus1) = WA (g) times CLipid ()V(L) times T(d)
where PLipid is the lipid productivity CLipid is the lipid content V is the working volume and T is thecultivation time
27 Strains Culture under Nitrogen-Sufficient and Nitrogen-Deficient Medium
The selected microalgae species Acutodesmus obliquus (KP8785051) Ankistrodesmus sp(KP8785001) and Chlorella lewinii (KP8785011) (Figure S3) were grown in 500-mL Erlenmeyerflasks containing 250 mL of liquid Chu medium No 10 with nitrogen sufficient (5756 microgmL ofCa(NO3)2middot4H2O) or liquid Chu medium No 10 without a nitrogen source (deficient) for seven days ina controlled culture room at 25 plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity ofcool-white fluorescent light and continuous agitation at 150 rpm
For growth profile analysis the maximum absorption peak for each microalgae strain wasdetermined in the 600ndash800 nm range by spectrophotometric analysis using a Nanodrop 2000 UV-VisSpectrophotometer (Figure S4) Then aliquots of the microalgal cultures were evaluated every 24 h(between 8 and 9 h) at 680 nm The specific growth rate (micro) was calculated during the exponentialgrowth phase (the first three days of culture) according to the equation micro = (ln Nf minus ln Ni)(tf minus ti)where N is the optical density (OD680) at final (f) or initial (i) time (t) Previously it was determinedthat a high (R2 from 0992 to 0998) and positive correlation exists between optical density and celldensities this variable was calculated using a hemocytometer The experiments were carried out intriplicate and the data were expressed as the mean plusmn standard deviation (plusmnSD)
Energies 2017 10 224 5 of 16
28 Determination of Total Protein Total Carbohydrate Ash Content Esterification and Fatty Acids Profiling
The microalgae cultures maintained for seven days under +N and minusN Chu medium No 10were harvested by centrifugation rinsed and dried as previously described (Section 25) Fromthe microalgae powders total protein content was determined according to the HartreendashLowrymethod [32] To 5 mg of dry microalgal biomass was added 5 mL of 05 M NaOH and incubated for10 min at 80 C then centrifuged for 5 min at 14000times g Protein content in 1 mL of the supernatantwas determined by adding 05 mL of 05 M sodium carbonate 05 mL of 05 M sodium potassiumtartrate 05 M of copper sulfate and 2 mL of Folin-Ciocalteu reagent The blue complex was analyzedin a Varian Cary 50 Bio UV-Visible Spectrophotometer (Agilent Technologies Santa Clara CA USA)set at 650 nm against a calibration curve based on a known concentration of BSA as the standard
Total carbohydrate content of the microalgal biomass was determined with the phenol-sulfuricacid method [3233] For this purpose 5 mg of dry microalgae biomass were added to 5 mL of waterAfter this a 1-mL aliquot of the sample was added to 3 mL of sulfuric acid and 1 mL of 5 aqueoussolution of phenol and the mixture was stirred and incubated for 5 min at 90 C The yellow-browncomplex was analyzed by spectrophotometric analysis at 488 nm against a calibration curve based ona known concentration of glucose
Ash content was done according to the AOAC procedure [34] One hundred milligrams of drymaterial were ashed in a muffle furnace Thermolynetrade F6010 (Thermo Fisher Scientific WalthmanMA USA) at 550 C for 16 h and the content was determined gravimetrically All analyses werecarried out in triplicate and the data are expressed as the mean plusmn SD
Fatty acids methyl esters (FAME) were prepared by acid transesterification according to Ichiharaand Fukubayashi [35] The crude lipid extract (1 mg) was dissolved in 02 mL of toluene 15 mL ofmethanol and 03 mL of 08 HCl (prepared in methanolwater 8515 vv) transferred into a cappedtest tube mixed for 5 min and then incubated at 45 C for 12 h for derivatization reaction Then 2 mLof n-hexane and 2 mL of distilled water were added to the tube After vortexing the hexane phasescontaining FAMEs were transferred into a 15-mL tube and dried under a stream of nitrogen Finallythe FAMEs were redissolved in 10 microL acetonitrile and analyzed by using a gas chromatograph VarianCP-3800 GC (Agilent Technologies Santa Clara CA USA) equipped with an automated sampler andinjector a flame ionization detector and using a 30 m times 032 mm times 025 microm Stabilwaxreg capillarycolumn (Restek Bellefonte PA USA) The GC conditions were as follows injector temperature 250 Cthe column temperature gradient was 120 C for 1 min followed by an increase to 160 C at the rateof 30 Cmin 160 C for 1 min followed by an increase to 240 C at the rate of 4 Cmin and 240 Cfor 7 min detector temperature 260 C Gas pressures for He H2 and synthetic air were maintained at40 80 and 60 psi respectively The carrier gas (He) flow was maintained at 1 mLmin The fatty acidswere analyzed by comparing their retention time of the corresponding peaks with a known standardmixture of FAMEs (Nu-Check Prep Elysian MN USA) and methyl tricosanoate (Sigma-AldrichSaint Louis MO USA) was added to each sample as the internal standard All chromatograms ofthe microalgal samples were analyzed by using the Galaxietrade Chromatography Data System Version1932 (Agilent Technologies Santa Clara CA USA) All peaks spanning a peak area of more than50 units were integrated All analyses were carried out in triplicate and the data are expressed as themean plusmn SD
29 Statistical Analysis
Statistical analyses were performed using the IBM SPSS Statistics 210 software (IBM ArmonkNY USA) The growth rate biomass and lipid content data for microalgal strains were analyzedusing a one-way analysis of variance (ANOVA) and the HSD Tukey test The correlation of biomassproductivity and lipid content was analyzed using bivariate correlations and Pearsonrsquos correlationcoefficients are given with their significance levels Differences were considered significant at p lt 005All data presented here are given as the mean plusmn SD
Energies 2017 10 224 6 of 16
3 Results and Discussion
31 Isolation and Identification of the Microalgae Strains
Bioprospecting of tropical ecosystems can provide exceptional native microalgae strains withthe potential for the production of biodiesel nutraceuticals bioactive chemicals and other highvalue products [36ndash38] Furthermore native microalgae strains can have potential applications forefficient biological sequestration of CO2 wastewater treatment and other environmentally-friendlyapplications [38ndash40] Consequently isolation is a fundamental process to obtain pure cultures and isthe first phase towards the screening and selection of microalgae strains with the potential for biodieselproduction and the aforementioned applications
According to these demands in this research samples were collected from three different typesof water bodies the Amazon River (white-water type) the Itaya and Nanay Rivers both of theblack-water type The white-water rivers originate from the Andean catchments bringing high amountsof nutrient-rich sediments In contrast the black-water rivers drain geochemically-poor watershedsand are characterized by their high content in humic acids low pH and poor nutrient status [41]A total of 14 microalgal strains were isolated using the standard cell washing technique and basedon distinguishable morphological characters under light microscopic examination these strains werepreliminary ascribed to the genera Acutodesmus (Hegewald) Tsarenko 2001 Ankistrodesmus Corda1838 Chlorella Beyerinck Beijerinck 1890 Desmodesmus (Chodat) SS An T Friedl and E Hegewald1999 and Tetradesmus GM Smith 1913 (Table 1) These genera are green algae that belong todivision Chlorophyta class Chlorophyceae and orders Chlorellales and Sphaeropleales Althoughmorphological analysis is frequently used to identify microalgae it is inaccurate and very difficultfor the identification at the species level because the relationship between diagnostic morphologyand biological species boundaries are largely unknown in many micro-eukaryotes [42] Furthermoreaccording to Yu et al [31] the microalgae morphology for the same strain varies in relation to ageand culture conditions Consequently in microalgae the classic taxonomy is supported by moleculartools [42] For example the ITS2 rDNA region is thought to be an excellent marker for molecularphylogenetics studies in lower taxonomic levels due to being highly divergent and fast-evolvingwhich can discriminate among closely-related organisms which otherwise display almost identicalsequences [4344] For this reason the molecular identification based in phylogenetic analysis of ITS2rDNA gene sequences is commonly used to identify andor delineate species [204546]
Table 1 Genera GenBank accession number of ITS2 sequences biomass productivity lipid productivityand lipid content of isolated microalgae strains
Source ofCollection Strain GeneraSpecies
GenBankAccessionNumber
BiomassProductivity
(mgmiddotLminus1middotdminus1)
LipidProductivity
(mgmiddotLminus1middotdminus1)
LipidContent( dw)
Amazon River(0341prime06prime prime S7314prime89prime prime W)
AMA001 Ankistrodesmus sp KP8784961 122 plusmn 03 37 plusmn 01 307 plusmn 15AMA002 Ankistrodesmus sp KP8784971 36 plusmn 04 10 plusmn 02 277 plusmn 21AMA003 Chlorella lewinii KP8785011 223 plusmn 07 51 plusmn 05 227 plusmn 15AMA004 Acutodesmus abliquus KP8785051 316 plusmn 04 64 plusmn 07 203 plusmn 21
Itaya River(0343prime14prime prime S
7314prime178prime prime W)
ITA001 Ankistrodesmus sp KP8784991 59 plusmn 02 40 plusmn 03 437 plusmn 17ITA002 Ankistrodesmus sp KP8785001 136 plusmn 04 57 plusmn 05 306 plusmn 20ITA003 Chlorella lewinii KP8785031 129 plusmn 03 40 plusmn 05 224 plusmn 27ITA004 Chlorella lewinii KP8785041 221 plusmn 07 31 plusmn 04 215 plusmn 23ITA005 Scenedesmus regularis KP8785081 93 plusmn 04 56 plusmn 04 242 plusmn 14ITA006 Scenedesmus dimorphus KP8785091 186 plusmn 06 32 plusmn 03 260 plusmn 17ITA007 Acutodesmus abliquus KP8785101 179 plusmn 04 22 plusmn 03 156 plusmn 19
Nanay River(0342prime02prime prime S7315prime32prime prime W)
NAN001 Ankistrodesmus sp KP8784981 143 plusmn 06 28 plusmn 02 467 plusmn 15NAN002 Desmodesmus tropicus KP8785061 122 plusmn 04 23 plusmn 02 175 plusmn 11NAN003 Tetradesmus sp KP8785071 139 plusmn 04 30 plusmn 03 135 plusmn 12
Energies 2017 10 224 7 of 16
To realize phylogenetic analysis genomic DNA of each isolate strain was purified and used forPCR amplification of ITS2 rDNA The universal primers used to amplify the ITS2 rDNA successfullyamplified the desired DNA fragments (~300 bp) from all microalgal strains (Figure S2) Furtherthese amplicons were sequenced published in the NCBI databases and the corresponding GenBankaccession numbers are presented in Table 1 Figure 1 shows a neighbor-joining phylogenetic treedepicting the relatedness of the native microalgae strain isolates with selected sequences from thepublic GenBank database This phylogram contained three clusters each of which included differentmicroalgae genera For example the first cluster consisted of microalgae of the genera AcutodesmusScenedesmus Tetradesmus and Desmodesmus and six isolated strains (ITA007 AMA004 ITA006 NAN003ITA005 and NAN002) The second cluster consisted of a microalgae of the genus Ankistrodesmus andthe isolated strains AMA001 ITA001 ITA002 NAN001 and AMA002 The third one consisted of amicroalgae of the genus Chlorella and three isolated strains (ITA004 AMA003 and ITA003) Overall theidentification of native microalgae strains presented a good correspondence between morphologicalcharacters and sequence-based phylogenetic analysis Microalgae species of these genera were alsoisolated from different countries of the world such as Brazil [47] Argentina [46] United States [48]India [20] Egypt [49] Taiwan [50] South Korea [45] and China [51] Some of these microalgae generasuch as Ankistrodesmus Chlorella Desmodesmus and Scenedesmus were used in large-scale biodieselproduction using photobioreactors and raceway pond systems [5253]
Energies 2017 10 224 7 of 16
public GenBank database This phylogram contained three clusters each of which included different microalgae genera For example the first cluster consisted of microalgae of the genera Acutodesmus Scenedesmus Tetradesmus and Desmodesmus and six isolated strains (ITA007 AMA004 ITA006 NAN003 ITA005 and NAN002) The second cluster consisted of a microalgae of the genus Ankistrodesmus and the isolated strains AMA001 ITA001 ITA002 NAN001 and AMA002 The third one consisted of a microalgae of the genus Chlorella and three isolated strains (ITA004 AMA003 and ITA003) Overall the identification of native microalgae strains presented a good correspondence between morphological characters and sequence-based phylogenetic analysis Microalgae species of these genera were also isolated from different countries of the world such as Brazil [47] Argentina [46] United States [48] India [20] Egypt [49] Taiwan [50] South Korea [45] and China [51] Some of these microalgae genera such as Ankistrodesmus Chlorella Desmodesmus and Scenedesmus were used in large-scale biodiesel production using photobioreactors and raceway pond systems [5253]
Figure 1 Unrooted phylogenetic tree of isolated microalgae strains using the ITS2 sequences The tree was constructed by the neighbor-joining method and Kimurarsquos two-parameter algorithm in MEGA 60 software based on the multiple sequence alignment by Clustal Omega Bootstrap values of 1000 replicates () are shown at the branches
32 Microalgae Cultivation Biomass and Total Lipids Measurements
The isolated microalgae strains exhibited remarkable differences in productivity parameters and lipid content among individual strains in the stationary growth phase (Table 1) With regard to the biomass productivity the 14 strains showed statistically-significant differences (F = 71911 df = 13 p lt 0001) and ranged from 36 plusmn 04 mgmiddotLminus1middotdminus1 (Ankistrodesmus sp strain AMA002) to 316 plusmn 04 mgmiddotLminus1middotdminus1 (Acutodesmus obliquus strain AMA004) Similar biomass productivities were reported for other marine microalgae species [54] However other investigations show major biomass productivities (ranging
Figure 1 Unrooted phylogenetic tree of isolated microalgae strains using the ITS2 sequences The treewas constructed by the neighbor-joining method and Kimurarsquos two-parameter algorithm in MEGA60 software based on the multiple sequence alignment by Clustal Omega Bootstrap values of1000 replicates () are shown at the branches
Energies 2017 10 224 8 of 16
32 Microalgae Cultivation Biomass and Total Lipids Measurements
The isolated microalgae strains exhibited remarkable differences in productivity parametersand lipid content among individual strains in the stationary growth phase (Table 1) With regardto the biomass productivity the 14 strains showed statistically-significant differences (F = 71911df = 13 p lt 0001) and ranged from 36 plusmn 04 mgmiddotLminus1middotdminus1 (Ankistrodesmus sp strain AMA002) to316 plusmn 04 mgmiddotLminus1middotdminus1 (Acutodesmus obliquus strain AMA004) Similar biomass productivities werereported for other marine microalgae species [54] However other investigations show major biomassproductivities (ranging from 30 to 120 mgmiddotLminus1middotdminus1) in the same and other microalgae genera [455556]These discrepancies may be attributed to differences in the culture conditions in the culture mediacomposition and in the microalgae genotypes Lipid productivity also exhibited statistically-significantdifferences (F = 4774 df = 13 p lt 0001) and ranged from 10plusmn 04 mgmiddotLminus1middotdminus1 to 64 plusmn 07 mgmiddotLminus1middotdminus1
with strains AMA002 and AMA004 respectively These lipid productivities were low in comparison toother reports which reported values from 190 to 433 mgmiddotLminus1middotdminus1 [4555] Furthermore lipid contentas a percentage of the microalgal biomass dry weight showed statistically-significant differences(F = 8509 df = 13 p lt 0001) and ranged from 135 plusmn 12 (Tetradesmus sp strain NAN003) to467 plusmn 15 (Ankistrodesmus sp strain NAN001) These percentage values in total lipid contentare similar to several microalgae species of these genera isolated and characterized by otherresearchers worldwide [20454648ndash5057] Furthermore it is important to emphasize that lipid contentis dependent on the different growth phases Commonly in several microalgae taxa the lipid contentincreases gradually from logarithmic to stationary growth phases [58ndash60] For example Kaur et al [20]reported that the total lipid content of Scenedesmus sp DRLMA9 demonstrated values of 207 inthe late exponential phase and 397 in the stationary phase and Desmodesmus elegans DRLMA13displayed values of 103 in the late exponential phase and 169 in the stationary phase LikewiseChiu et al [61] found that the total lipid content of Nannochloropsis oculata was 308 in the logarithmicphase 397 in the early stationary phase and 504 in the stationary phase These changes in lipidcontent with culture age have been associated with nitrate depletion [61] which causes a channeling ofphotosynthetic ATP to carbohydrate and lipid biosynthesis [60] Finally Pearson correlation analysisof biomass productivity and lipid content was inversely related (r = minus037 p lt 001) which is similarto the results obtained by Liang et al [62] and Rodolfi et al [52] In contrast Li et al [51] andHempel et al [63] found no relationship between these parameters The negative correlation obtainedis likely due to the high metabolic cost of lipid biosynthesis [62]
33 Strains Culture under Nitrogen-Sufficient and Nitrogen-Deficient Medium
The microalgae species Acutodesmus obliquus (KP8785051) Ankistrodesmus sp (KP8785001)and Chlorella lewinii (KP8785011) were selected and further characterized because they displayedthe highest biomass and lipid productivities (Table 1) These microalgae showed noticeablevariations in the growth profile when cultured in Chu medium No 10 with nitrogen (+N) orwithout a nitrogen source (minusN) In +N medium all strains showed exponential growth butChlorella lewinii showed the highest average specific growth rates (micro = 0458 plusmn 0098 dminus1) followed byAcutodesmus obliquus (micro = 0399 plusmn 0112 dminus1) and the lowest value was produced by Ankistrodesmus sp(micro = 0265 plusmn 0013 dminus1) In contrast all cultures displayed a lag phase pattern in their growthprofile under minusN medium (Figure 2) These findings corroborate the results obtained by otherresearchers who demonstrate that the specific growth rate is directly proportional to the initialnitrate concentration [46596465] because the reduction or cessation of microalgae cell divisionis due to nitrogen limitation [66] which is a key nutrient for protein and nucleic acids de novobiosynthesis consequently causing an arrest in the early part of the microalgae cell cycle [6768]In accordance with these reports Li et al [69] showed that nitrogen deficiency induced in the greenmicroalgae Micractinium pusillum a downregulation in the expression of genes involved in cell growthand cell cycle regulation Additionally differences in growth profile and specific growth rates aredependent on several factors such as microalgae genotypes the culture medium composition pH
Energies 2017 10 224 9 of 16
nutrients concentration the cell culture phase (ie log phase stationary phase) and environmentalfactors [4549545770ndash72]Energies 2017 10 224 9 of 16
Figure 2 Growth profiles of three microalgal strains cultivated under +N (filled boxes) and minusN (empty boxes) Chu medium No 10 The culture was incubated in a controlled culture room at 25 plusmn 1 degC with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 150 rpm Experiments were carried out in triplicate
34 Total Protein Total Lipid and Total Carbohydrate Content
Biochemical parameters of the microalgae species such as total protein total lipid and total carbohydrate content exhibited differences and changes under both culture conditions (Figure 3) For example Chlorella lewinii showed the highest protein content Ankistrodesmus sp had the highest total lipid content but Acutodesmus obliquus presented the highest levels of total carbohydrate Therefore these strains can be valuable stocks for the industry because total protein and total carbohydrate are potentially byproducts of microalgal biomass processing [7374] Additionally the three strains cultured under minusN conditions showed a significant increase in total lipid content (from 237 in Acutodesmus obliquus to 667 in Ankistrodesmus sp) and total carbohydrate content (from 119 in Acutodesmus obliquus to 963 in Chlorella lewinii) but showed a decreased in total protein levels (from 242 in Acutodesmus obliquus to 545 in Chlorella lewinii) in comparison to +N cultures (Figure 3) These results are in accordance with well-documented results obtained by other researchers with several microalgae species of the genera Acutodesmus Ankistrodesmus Botryococcus Chlamydomonas Chlorella Desmodesmus Dunaliella Haematococcus Neochloris Phaeodactylum Pseudokirchneriella and Scenedesmus [465058596475ndash80] Nevertheless the accumulation of total lipid induced by nitrogen deficiency is not correlated with lipid productivity but rather is due to a decline and stopping of the microalgae specific growth rate [75] The changes in biomolecule contents suggest redirection of microalgae metabolism during nitrogen starvation such as a decrease in biosynthesisincrease in protein degradation and an increase in the biosynthesis of storage biomolecules such as triacylglycerols and starch The increase in total lipid content could be partially explained for a boost in transcript levels of genes encoding enzymes of the lipid biosynthesis pathways According to this assumption recent investigations showed that genes encoding the enzymes that catalyze the last step in the Kennedy pathway of triacylglycerol biosynthesis [81] such as glycerol-3-phosphate acyltransferase and acyl-glycerol-3-phosphate acyltransferase are upregulated in response to nitrogen starvation [648283] Furthermore the increase in total carbohydrate content can be explained by changes in the expression levels of genes involved in carbohydrate metabolism For example when the oleaginous microalgae Neochloris oleoabundans is cultured under nitrogen starvation the catabolic phase of starch specifically α-amylase which hydrolyzes starch to glucose is strongly repressed [83]
Figure 2 Growth profiles of three microalgal strains cultivated under +N (filled boxes) and minusN(empty boxes) Chu medium No 10 The culture was incubated in a controlled culture room at25 plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent lightand continuous agitation at 150 rpm Experiments were carried out in triplicate
34 Total Protein Total Lipid and Total Carbohydrate Content
Biochemical parameters of the microalgae species such as total protein total lipid and totalcarbohydrate content exhibited differences and changes under both culture conditions (Figure 3)For example Chlorella lewinii showed the highest protein content Ankistrodesmus sp had the highesttotal lipid content but Acutodesmus obliquus presented the highest levels of total carbohydrateTherefore these strains can be valuable stocks for the industry because total protein and totalcarbohydrate are potentially byproducts of microalgal biomass processing [7374] Additionallythe three strains cultured under minusN conditions showed a significant increase in total lipid content(from 237 in Acutodesmus obliquus to 667 in Ankistrodesmus sp) and total carbohydrate content(from 119 in Acutodesmus obliquus to 963 in Chlorella lewinii) but showed a decreased in totalprotein levels (from 242 in Acutodesmus obliquus to 545 in Chlorella lewinii) in comparison to +Ncultures (Figure 3) These results are in accordance with well-documented results obtained by otherresearchers with several microalgae species of the genera Acutodesmus Ankistrodesmus BotryococcusChlamydomonas Chlorella Desmodesmus Dunaliella Haematococcus Neochloris PhaeodactylumPseudokirchneriella and Scenedesmus [465058596475ndash80] Nevertheless the accumulation of totallipid induced by nitrogen deficiency is not correlated with lipid productivity but rather is due toa decline and stopping of the microalgae specific growth rate [75] The changes in biomoleculecontents suggest redirection of microalgae metabolism during nitrogen starvation such as a decreasein biosynthesisincrease in protein degradation and an increase in the biosynthesis of storagebiomolecules such as triacylglycerols and starch The increase in total lipid content could bepartially explained for a boost in transcript levels of genes encoding enzymes of the lipid biosynthesispathways According to this assumption recent investigations showed that genes encoding theenzymes that catalyze the last step in the Kennedy pathway of triacylglycerol biosynthesis [81] such asglycerol-3-phosphate acyltransferase and acyl-glycerol-3-phosphate acyltransferase are upregulated
Energies 2017 10 224 10 of 16
in response to nitrogen starvation [648283] Furthermore the increase in total carbohydrate contentcan be explained by changes in the expression levels of genes involved in carbohydrate metabolismFor example when the oleaginous microalgae Neochloris oleoabundans is cultured under nitrogenstarvation the catabolic phase of starch specifically α-amylase which hydrolyzes starch to glucoseis strongly repressed [83]Energies 2017 10 224 10 of 16
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strains cultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled culture room at 25 plusmn 1 degC with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized with respect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that each microalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majority of fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised 7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii the majority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9) acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectively Finally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic (182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in +N and minusN conditions respectively Our results agree with multiple previously-published reports in that fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent in many green microalgae species [454648ndash5055] It is notable that under nitrogen starvation culture all microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 in Ankistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid (181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findings support the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content of SFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Nevertheless there are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed a decrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increased between 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively However in the same culture condition the three microalgae species showed a decrease in polyunsaturated fatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmus spp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgae strains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFA which improves the quality properties of biodiesel such as an increase
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strainscultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled cultureroom at 25plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescentlight and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized withrespect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that eachmicroalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majorityof fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii themajority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9)acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectivelyFinally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic(182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in+N and minusN conditions respectively Our results agree with multiple previously-published reports inthat fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent inmany green microalgae species [454648ndash5055] It is notable that under nitrogen starvation cultureall microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 inAnkistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid(181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findingssupport the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content ofSFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other
Energies 2017 10 224 11 of 16
researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Neverthelessthere are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed adecrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increasedbetween 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively Howeverin the same culture condition the three microalgae species showed a decrease in polyunsaturatedfatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmusspp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgaestrains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFAwhich improves the quality properties of biodiesel such as an increase in cetane number [84ndash86]Considering the SFA and unsaturated fatty acids (UFA) components approximately 20ndash31 are SFAand 69ndash80 are UFA Hence the SFAUFA ratio in the microalgae strains evaluated ranges between025 and 045 which is similar to the reports of several microalgae species isolated in different latitudesworldwide [203148497072]
Table 2 Effect of nitrogen deficiency on fatty acid profiling in three native microalgal species from thePeruvian Amazon
Fatty AcidsAcutodesmus obliquus Ankistrodesmus sp Chlorella lewinii
+N minusN +N minusN +N minusN
140 - - 052 plusmn 002 088 plusmn 002 - 058 plusmn 001160 2724 plusmn 007 2867 plusmn 017 1868 plusmn 019 2261 plusmn 006 1964 plusmn 021 2123 plusmn 010180 184 plusmn 005 255 plusmn 051 101 plusmn 002 181 plusmn 002 371 plusmn 026 153 plusmn 015
161n-7 023 plusmn 000 025 plusmn 001 071 plusmn 003 055 plusmn 001 085 plusmn 043 038 plusmn 000181n-7 131 plusmn 002 131 plusmn 000 141 plusmn 005 094 plusmn 003 146 plusmn 006 123 plusmn 002181n-9 3213 plusmn 021 3608 plusmn 021 3699 plusmn 004 4047 plusmn 038 1371 plusmn 015 2326 plusmn 041182n-6 1732 plusmn 004 1651 plusmn 016 747 plusmn 018 656 plusmn 007 2034 plusmn 039 1957 plusmn 001183n-3 1130 plusmn 017 854 plusmn 031 1486 plusmn 004 1173 plusmn 012 1815 plusmn 022 1371 plusmn 016183n-6 101 plusmn 001 078 plusmn 002 055 plusmn 000 054 plusmn 002 124 plusmn 006 100 plusmn 020184n-3 250 plusmn 005 177 plusmn 008 631 plusmn 002 601 plusmn 005 491 plusmn 025 403 plusmn 020204n-6 - - - 084 plusmn 008 - -225n-3 - - - 062 plusmn 002 - -
Unidentified 512 plusmn 003 355 plusmn 006 1149 plusmn 004 645 plusmn 003 1599 plusmn 034 1348 plusmn 006SFA 2907 3122 2022 2531 2335 2334UFA 7093 6878 7978 7469 7665 7666
MUFA 3367 3764 3911 4195 1602 2487PUFA 3213 2759 2918 2629 4464 3831SFA +MUFA 6274 6886 5933 6726 3936 4821
It is important to highlight that the fatty acid profile determination is an essential step inthe process of the characterization of microalgae strains because it ultimately affects the qualityof the biodiesel product [8485] The quality properties of biodiesel such as high cetane numberoxidative stability and cold-flow are dependent on the hydrocarbonated chain length of saturated andunsaturated fatty acids [8587] For example biodiesel composed of a mixture of a high percentage ofSFA and MUFA is preferred for increased energy yield superior oxidative stability and higher cetanenumbers but is prone to solidify at low temperature In contrast biodiesel with high levels of PUFAhas decreased energy yield low cetane numbers and excellent cold-flow properties but insufficientoxidative stability [548588] Therefore to solve these performance problems five approaches havebeen developed such as changing the fatty acids profile by physical means genetic modification ofthe feedstock or the use of alternative feedstocks with different fatty acids profiles [89]
4 Conclusions
Fourteen microalgae were isolated from three river basins from the Peruvian Amazon and wereidentified by morphological features and molecular phylogeny approaches These strains were initially
Energies 2017 10 224 12 of 16
characterized in the stationary growth phase and exhibited significant heterogeneity in key parameterssuch as biomass productivity (36ndash316 mgmiddotLminus1middotdminus1) lipid productivity (10ndash64 mgmiddotLminus1middotdminus1) andtotal lipid content (135ndash467) Furthermore three selected strains based on elevated biomass andlipid productivities were cultured under nitrogen-sufficient and nitrogen-deficient medium to boostlipid accumulation Under these culture conditions all strains showed noticeable decreases in totalprotein content and increases in total lipid and total carbohydrate contents In addition these strainspresented modifications in the fatty acids profiles with improvement in biodiesel properties due to apredominance of SFA and MUFA which corresponds to an increase in cetane number The results ofthis research suggest that the isolated native microalgae from the Peruvian Amazon could be suitablerenewable feedstocks for the production of both biodiesel and bioethanol which could contribute tosustainable development at the local regional and national levels as a viable alternative to petroleumexploitation However it will be necessary to develop open closed or hybrid systems for large-scalebiomass production of selected native microalgae strains andor de novo microalgae strains designedto produce biofuels and with the abilities to synthesize useful bioactive compounds to be efficientlyexploited using biorefinery approaches
Supplementary MaterialsThe following are available online at wwwmdpicom1996-1073102224s1 Figure S1 Results of
electrophoretic and spectrophotometric analysis of the genomic DNA isolated from the fourteen microalgaeFigure S2 ITS2-rDNA structure (A) and amplicons obtained (B) from the microalgae isolated Figure S3Microphotographic images of the three microalgae strains analyzed under light and fluorescence stained withNile Red Figure S4 Spectrophotometric scanning for the determination of the maximum absorption peak foreach microalgae strain
Acknowledgments This research was supported by the Peruvian funding agencies Consejo Nacional de CienciaTecnologiacutea e Innovacioacuten Tecnoloacutegica (CONCYTEC) with Grant Contract No 364-CONCYTEC-OAJ and Fondospara la Innovacioacuten Ciencia y Tecnologiacutea (FINCyT) with Grant Contract No 383-PNICP-PIBA-2014
Author Contributions Marianela Cobos and Juan C Castro obtained funds for the research participated inthe study design coordinated activities and participated in the preparation of the manuscript Jae D Paredesparticipated in sample collection microalgae isolation and helped to draft the manuscript J Dylan Maddoxperformed the bioinformatics analysis and participated in the preparation of the manuscript Gabriel Vargas-Aranaand Leenin Flores participated in biochemical analysis and helped to draft the manuscript Carla P Aguilarparticipated in the study design supervised biochemical analysis and helped to draft the manuscriptJorge L Marapara participated in molecular analysis and helped to draft the manuscript
Conflicts of Interest The authors declare no conflict of interest
References
1 BP Global BP Statisical Review of World Energy 2014 Available online httpwwwbpcomcontentdambppdfEnergy-economicsstatistical-review-2014BP-statistical-review-of-world-energy-2014-full-reportpdf (accessed on 20 June 2014)
2 Pachauri RK Reisinger A Cambio Climaacutetico 2007 Informe de Siacutentesis Contribucioacuten de los Grupos de Trabajo III y III al Cuarto Informe de Evaluacioacuten del Grupo Intergubernamental de Expertos Sobre el Cambio Climaacutetico IPCCGeneva Switzerland 2007 p 104
3 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Ley de Promocioacuten del Mercado deBiocombustibles Available online httpwww2congresogobpesicrcendocbibcon4_uibdnsf9E95620CC059138105257C9E005AB2F9$FILE28054pdf (accessed on 12 February 2017)
4 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Reglamento de la Ley de Promocioacuten del Mercadode Biocombustibles Available online httpwww2osinerggobpeMarcoLegaldocrevDS-013-2005-EM-CONCORDADOpdf (accessed on 12 February 2017)
5 Areacutevalo LF Nalvarte J Torres J Ramiacuterez Y Impactos Socio-Econoacutemicos de la Produccioacuten de Biocombustiblesen la Amazoniacutea Peruana Primera SNV-WWF Lima Peruacute 2009
6 Sharma KK Schuhmann H Schenk PM High Lipid Induction in Microalgae for Biodiesel ProductionEnergies 2012 5 1532ndash1553 [CrossRef]
Energies 2017 10 224 13 of 16
7 Schenk PM Thomas-Hall SR Stephens E Marx UC Mussgnug JH Posten C Kruse O Hankamer BSecond Generation Biofuels High-Efficiency Microalgae for Biodiesel Production BioEnergy Res 2008 120ndash43 [CrossRef]
8 Foley JA Asner GP Costa MH Coe MT de Fries R Gibbs HK Howard EA Olson S Patz JRamankutty N et al Amazonia revealed Forest degradation and loss of ecosystem goods and services inthe Amazon Basin Front Ecol Environ 2007 5 25ndash32 [CrossRef]
9 Duong VT Li Y Nowak E Schenk PM Microalgae Isolation and Selection for Prospective BiodieselProduction Energies 2012 5 1835ndash1849 [CrossRef]
10 Georgianna DR Mayfield SP Exploiting diversity and synthetic biology for the production of algalbiofuels Nature 2012 488 329ndash335 [CrossRef] [PubMed]
11 Wijffels RH Barbosa MJ An Outlook on Microalgal Biofuels Science 2010 329 796ndash799 [CrossRef][PubMed]
12 Richmond A Handbook of Microalgal Culture Biotechnology and Applied Phycology Blackwell Sci Ltd HudsonNJ USA 2004
13 Menetrez MY An overview of algae biofuel production and potential environmental impactEnviron Sci Technol 2012 46 7073ndash7085 [CrossRef] [PubMed]
14 Khoo HH Sharratt PN Das P Balasubramanian RK Naraharisetti PK Shaik S Life cycle energy andCO2 analysis of microalgae-to-biodiesel Preliminary results and comparisons Bioresour Technol 2011 1025800ndash5807 [CrossRef] [PubMed]
15 Chu SP The Influence of the Mineral Composition of the Medium on the Growth of Planktonic AlgaePart I Methods and Culture Media J Ecol 1942 30 284ndash325 [CrossRef]
16 Carlos EMB Mariatildengela M Gecircneros de Algas de Aacuteguas Continentais do BrasilmdashChave Para Identificaccedilatildeo eDescriccedilotildees 2nd ed Rima Sao Carlos Brasil 2006
17 Greenspan P Mayer EP Fowler SD Nile red A selective fluorescent stain for intracellular lipid dropletsJ Cell Biol 1985 100 965ndash973 [CrossRef] [PubMed]
18 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissuePhytochem Bull 1987 19 11ndash15
19 Sambrook J Fritsch EF Maniatis T Molecular Cloning A Laboratory Manual 2nd ed Cold Spring HarborLaboratory Press New York NY USA 1989
20 Kaur S Sarkar M Srivastava RB Gogoi HK Kalita MC Fatty acid profiling and molecularcharacterization of some freshwater microalgae from India with potential for biodiesel productionNew Biotechnol 2012 29 332ndash344 [CrossRef] [PubMed]
21 Benson DA Cavanaugh M Clark K Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW GenBankNucleic Acids Res 2013 41 D36ndashD42 [CrossRef] [PubMed]
22 NCBI Resource Coordinators Database Resources of the National Center for Biotechnology InformationNucleic Acids Res 2015 43 D6ndashD17
23 Ye J McGinnis S Madden TL BLAST Improvements for better sequence analysis Nucleic Acids Res 200634 W6ndashW9 [CrossRef] [PubMed]
24 BioEdit Sequence Alignment Editor for Windows 9598NTXPVista7 Available online httpwwwmbioncsuedubioeditbioedithtml (accessed on 2 December 2015)
25 Sievers F Higgins DG Clustal omega accurate alignment of very large numbers of sequencesMethods Mol Biol (Clifton NJ) 2014 1079 105ndash116
26 Saitou N Nei M The neighbor-joining method A new method for reconstructing phylogenetic treesMol Biol Evol 1987 4 406ndash425 [PubMed]
27 Kimura M A simple method for estimating evolutionary rates of base substitutions through comparativestudies of nucleotide sequences J Mol Evol 1980 16 111ndash120 [CrossRef] [PubMed]
28 Tamura K Stecher G Peterson D Filipski A Kumar S MEGA6 Molecular Evolutionary GeneticsAnalysis version 60 Mol Biol Evol 2013 30 2725ndash2729 [CrossRef] [PubMed]
29 Felsenstein J Confidence Limits on Phylogenies An Approach Using the Bootstrap Evolution 1985 39783ndash791 [CrossRef]
30 Bligh EG Dyer WJ A Rapid Method of Total Lipid Extraction and Purification Can J Biochem Physiol1959 37 911ndash917 [CrossRef] [PubMed]
Energies 2017 10 224 14 of 16
31 Yu X Zhao P He C Li J Tang X Zhou J Huang Z Isolation of a novel strain of Monoraphidium spand characterization of its potential application as biodiesel feedstock Bioresour Technol 2012 121 256ndash262[CrossRef] [PubMed]
32 Hartree EF Determination of protein A modification of the lowry method that gives a linear photometricresponse Anal Biochem 1972 48 422ndash427 [CrossRef]
33 DuBois M Gilles KA Hamilton JK Rebers PA Smith F Colorimetric Method for Determination ofSugars and Related Substances Anal Chem 1956 28 350ndash356 [CrossRef]
34 AOAC International AOAC Official Methods of Analysis of the Association of Official Analytical Chemists (AOAC)15th ed AOAC International Washington DC USA 1990 Volume 1
35 Ichihara K Fukubayashi Y Preparation of fatty acid methyl esters for gas-liquid chromatographyJ Lipid Res 2010 51 635ndash640 [CrossRef] [PubMed]
36 Brennan L Owende P Biofuels from microalgaemdashA review of technologies for production processingand extractions of biofuels and co-products Renew Sustain Energ Rev 2010 14 557ndash577 [CrossRef]
37 Raja R Hemaiswarya S Kumar NA Sridhar S Rengasamy R A perspective on the biotechnologicalpotential of microalgae Crit Rev Microbiol 2008 34 77ndash88 [CrossRef] [PubMed]
38 Mata TM Martins AA Caetano NS Microalgae for biodiesel production and other applicationsA review Renew Sustain Energy Rev 2010 14 217ndash232 [CrossRef]
39 De Wilt A Butkovskyi A Tuantet K Leal LH Fernandes TV Langenhoff A Zeeman GMicropollutant removal in an algal treatment system fed with source separated wastewater streamsJ Hazard Mater 2015 304 84ndash92 [CrossRef] [PubMed]
40 Li T Lin G Podola B Melkonian M Continuous removal of zinc from wastewater and mine dumpleachate by a microalgal biofilm PSBR J Hazard Mater 2015 297 112ndash118 [CrossRef] [PubMed]
41 Sioli H The Amazon and its main affluents Hydrography morphology of the river courses and river typesIn Monographiae Biologicae Sioli H Ed Springer Dordrecht The Netherlands 1984 pp 127ndash165
42 Moniz MBJ Kaczmarska I Barcoding of Diatoms Nuclear Encoded ITS Revisited Protist 2010 161 7ndash34[CrossRef] [PubMed]
43 Coleman AW Is there a molecular key to the level of ldquobiological speciesrdquo in eukaryotes A DNA guideMol Phylogenet Evol 2009 50 197ndash203 [CrossRef] [PubMed]
44 Mai JC Coleman AW The Internal Transcribed Spacer 2 Exhibits a Common Secondary Structure inGreen Algae and Flowering Plants J Mol Evol 1997 44 258ndash271 [CrossRef] [PubMed]
45 Abou-Shanab RAI Matter IA Kim S-N Oh Y-K Choi J Jeon B-H Characterization andidentification of lipid-producing microalgae species isolated from a freshwater lake Biomass Bioenergy2011 35 3079ndash3085 [CrossRef]
46 Do Nascimento M Ortiz-Marquez JCF Sanchez-Rizza L Echarte MM Curatti L Bioprospectingfor fast growing and biomass characterization of oleaginous microalgae from South-Eastern Buenos AiresArgentina Bioresour Technol 2012 125 283ndash290 [CrossRef] [PubMed]
47 Araujo GS Matos LJBL Gonccedilalves LRB Fernandes FAN Farias WRL Bioprospecting foroil producing microalgal strains Evaluation of oil and biomass production for ten microalgal strainsBioresour Technol 2011 102 5248ndash5250 [CrossRef] [PubMed]
48 Zhou W Li Y Min M Hu B Chen P Ruan R Local bioprospecting for high-lipid producing microalgalstrains to be grown on concentrated municipal wastewater for biofuel production Bioresour Technol 2011102 6909ndash6919 [CrossRef] [PubMed]
49 Mahmoud EA Farahat LA Abdel Aziz ZK Fatthallah NA Salah El Din RA Evaluation of thepotential for some isolated microalgae to produce biodiesel Egypt J Pet 2015 24 97ndash101 [CrossRef]
50 Ho S Lai Y-Y Chiang C-Y Chen C-NN Chang J-S Selection of elite microalgae for biodieselproduction in tropical conditions using a standardized platform Bioresour Technol 2013 147 135ndash142[CrossRef] [PubMed]
51 Li L Cui J Liu Q Ding Y Liu J Screening and phylogenetic analysis of lipid-rich microalgae Algal Res2015 11 381ndash386 [CrossRef]
52 Rodolfi L Chini Zittelli G Bassi N Padovani G Biondi N Bonini G Tredici MR Microalgae for oilStrain selection induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactorBiotechnol Bioeng 2009 102 100ndash112 [CrossRef] [PubMed]
Energies 2017 10 224 15 of 16
53 Li P Miao X Li R Zhong J In situ biodiesel production from fast-growing and high oil content Chlorellapyrenoidosa in rice straw hydrolysate J Biomed Biotechnol 2011 2011 141207 [CrossRef] [PubMed]
54 Doan TTY Sivaloganathan B Obbard JP Screening of marine microalgae for biodiesel feedstockBiomass Bioenergy 2011 35 2534ndash2544 [CrossRef]
55 Abou-Shanab RAI Hwang J-H Cho Y Min B Jeon B-H Characterization of microalgal speciesisolated from fresh water bodies as a potential source for biodiesel production Appl Energy 2011 883300ndash3306 [CrossRef]
56 Chaichalerm S Pokethitiyook P Yuan W Meetam M Sritong K Pugkaew W Kungvansaichol KKruatrachue M Damrongphol P Culture of microalgal strains isolated from natural habitats in Thailand invarious enriched media Appl Energy 2012 89 296ndash302 [CrossRef]
57 Zhang S Liu P Yang X Hao Z Zhang L Luo N Shi J Isolation and identification by 18S rDNAsequence of high lipid potential microalgal species for fuel production in Hainan Dao Biomass Bioenergy2014 66 197ndash203 [CrossRef]
58 Chiu S-Y Kao C-Y Tsai M-T Ong S-C Chen C-H Lin C-S Lipid accumulation and CO2 utilizationof Nannochloropsis oculata in response to CO2 aeration Bioresour Technol 2009 100 833ndash838 [CrossRef][PubMed]
59 Huerlimann R de Nys R Heimann K Growth lipid content productivity and fatty acid composition oftropical microalgae for scale-up production Biotechnol Bioeng 2010 107 245ndash257 [CrossRef] [PubMed]
60 Liang Y Sarkany N Cui Y Biomass and lipid productivities of Chlorella vulgaris under autotrophicheterotrophic and mixotrophic growth conditions Biotechnol Lett 2009 31 1043ndash1049 [CrossRef] [PubMed]
61 Hempel N Petrick I Behrendt F Biomass productivity and productivity of fatty acids and amino acidsof microalgae strains as key characteristics of suitability for biodiesel production J Appl Phycol 2012 241407ndash1418 [CrossRef] [PubMed]
62 Yeesang C Cheirsilp B Effect of nitrogen salt and iron content in the growth medium and light intensityon lipid production by microalgae isolated from freshwater sources in Thailand Bioresour Technol 2011 1023034ndash3040 [CrossRef] [PubMed]
63 Msanne J Xu D Konda AR Casas-Mollano JA Awada T Cahoon EB Cerutti H Metabolic andgene expression changes triggered by nitrogen deprivation in the photoautotrophically grown microalgaeChlamydomonas reinhardtii and Coccomyxa sp C-169 Phytochemistry 2012 75 50ndash59 [CrossRef] [PubMed]
64 Lin Q Lin J Effects of nitrogen source and concentration on biomass and oil production of aScenedesmus rubescens like microalga Bioresour Technol 2011 102 1615ndash1621 [CrossRef] [PubMed]
65 Sheehan J Dunahay T Benemann J Roessler P A Look Back at the US Department of Energyrsquos AquaticSpecies ProgrammdashBiodiesel from Algae US Department of Energyrsquos Office of Fuels Development Golden COUSA 1998 p 328
66 Vaulot D Olson RJ Merkel S Chisholm SW Cell-cycle response to nutrient starvation in twophytoplankton species Thalassiosira weissflogii and Hymenomonas carterae Mar Biol 1987 95 625ndash630[CrossRef]
67 Olson RJ Chisholm SW Effects of light and nitrogen limitation on the cell cycle of the dinoflagellateAmphidinium carteri J Plankton Res 1986 8 785ndash793 [CrossRef]
68 Li Y Fei X Deng X Novel molecular insights into nitrogen starvation-induced triacylglycerolsaccumulation revealed by differential gene expression analysis in green algae Micractinium pusillumBiomass Bioenergy 2012 42 199ndash211 [CrossRef]
69 Abdelaziz AEM Leite GB Belhaj MA Hallenbeck PC Screening microalgae native to Quebec forwastewater treatment and biodiesel production Bioresour Technol 2014 157 140ndash148 [CrossRef] [PubMed]
70 Valdez-Ojeda R Gonzaacutelez-Muntildeoz M Us-Vaacutezquez R Narvaacuteez-Zapata J Chavarria-Hernandez JCLoacutepez-Adriaacuten S Barahona-Peacuterez F Toledano-Thompson T Garduntildeo-Soloacuterzano G Escobedo-GraciaMedrano RM Characterization of five fresh water microalgae with potential for biodiesel productionAlgal Res 2015 7 33ndash44 [CrossRef]
71 Shrivastav A Mishra SK Suh WI Farooq W Moon M Kim T-H Kumar K Choi G-G Park MSYang J-W Characterization of newly isolated oleaginous microalga Monoraphidium sp for lipid productionunder different conditions Algal Res 2015 12 289ndash294 [CrossRef]
72 Yen H-W Hu I-C Chen C-Y Ho S-H Lee D-J Chang J-S Microalgae-based biorefinerymdashFrombiofuels to natural products Bioresour Technol 2013 135 166ndash174 [CrossRef] [PubMed]
Energies 2017 10 224 16 of 16
73 Li J Liu Y Cheng JJ Mos M Daroch M Biological potential of microalgae in China for biorefinery-basedproduction of biofuels and high value compounds New Biotechnol 2015 32 588ndash596 [CrossRef] [PubMed]
74 El-Sheekh M Abomohra AE-F Hanelt D Optimization of biomass and fatty acid productivity ofScenedesmus obliquus as a promising microalga for biodiesel production World J Microbiol Biotechnol 201329 915ndash922 [CrossRef] [PubMed]
75 Singh P Guldhe A Kumari S Rawat I Bux F Investigation of combined effect of nitrogen phosphorusand iron on lipid productivity of microalgae Ankistrodesmus falcatus KJ671624 using response surfacemethodology Biochem Eng J 2015 94 22ndash29 [CrossRef]
76 Kilham S Kreeger D Goulden C Lynn S Effects of nutrient limitation on biochemical constituents ofAnkistrodesmus falcatus Freshw Biol 1997 38 591ndash596 [CrossRef]
77 Breuer G Lamers PP Martens DE Draaisma RB Wijffels RH The impact of nitrogen starvation on thedynamics of triacylglycerol accumulation in nine microalgae strains Bioresour Technol 2012 124 217ndash226[CrossRef] [PubMed]
78 Breuer G de Jaeger L Artus VG Martens DE Springer J Draaisma RB Eggink G Wijffels RHLamers PP Superior triacylglycerol (TAG) accumulation in starchless mutants of Scenedesmus obliquus (II)evaluation of TAG yield and productivity in controlled photobioreactors Biotechnol Biofuels 2014 7 70[CrossRef] [PubMed]
79 Nigam S Prakash-Rai M Sharma R Effect of Nitrogen on Growth and Lipid Content ofChlorella pyrenoidosa Am J Biochem Biotechnol 2011 7 126ndash131 [CrossRef]
80 Piligaev AV Sorokina KN Bryanskaya AV Peltek SE Kolchanov NA Parmon VN Isolation ofprospective microalgal strains with high saturated fatty acid content for biofuel production Algal Res 201512 368ndash376 [CrossRef]
81 Weiss SB Kennedy EP The enzymatic synthesis of triglycerides J Am Chem Soc 1956 78 3550[CrossRef]
82 Miller R Wu G Deshpande RR Vieler A Gaumlrtner K Li X Moellering ER Zaumluner S Cornish AJLiu B et al Changes in Transcript Abundance in Chlamydomonas reinhardtii following Nitrogen DeprivationPredict Diversion of Metabolism Plant Physiol 2010 154 1737ndash1752 [CrossRef] [PubMed]
83 Rismani-Yazdi H Haznedaroglu BZ Bibby K Peccia J Transcriptome sequencing and annotation of themicroalgae Dunaliella tertiolecta Pathway description and gene discovery for production of next-generationbiofuels BMC Genom 2011 12 148 [CrossRef] [PubMed]
84 Ramos MJ Fernaacutendez CM Casas A Rodriacuteguez L Peacuterez A Influence of fatty acid composition of rawmaterials on biodiesel properties Bioresour Technol 2009 100 261ndash268 [CrossRef] [PubMed]
85 Knothe G Dependence of biodiesel fuel properties on the structure of fatty acid alkyl estersFuel Process Technol 2005 86 1059ndash1070 [CrossRef]
86 Knothe G ldquoDesignerrdquo Biodiesel Optimizing Fatty Ester Composition to Improve Fuel PropertiesEnergy Fuels 2008 22 1358ndash1364 [CrossRef]
87 Smith PC Ngothai Y Dzuy Nguyen Q OrsquoNeill BK Improving the low-temperature properties ofbiodiesel Methods and consequences Renew Energy 2010 35 1145ndash1151 [CrossRef]
88 Dunn RO Effect of antioxidants on the oxidative stability of methyl soyate (biodiesel) Fuel Process Technol2005 86 1071ndash1085 [CrossRef]
89 Knothe G Improving biodiesel fuel properties by modifying fatty ester composition Energy Environ Sci2009 2 759ndash766 [CrossRef]
copy 2017 by the authors licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)

Energies 2017 10 224 2 of 16
deforestation of the rainforest which pose threats to native biodiversity and ecosystem functionsgoods and services [6ndash8] These limitations can be overcome by the next generation of microalgae-basedbiofuels Compared to terrestrial crops the main advantages of microalgal systems are that they havea higher photon conversion efficiency can be harvested batch-wise nearly all-year-round can utilizesalt and waste water streams can couple CO2-neutral fuel production with CO2 sequestration andproduce non-toxic and highly biodegradable biofuels [7]
The isolation selection and culturing of robust oleaginous microalgae with desirable attributes(eg high triacylglycerol content appropriate fatty acid profiles high growth rate and resistance toinvasion of local microorganism) however are the principal challenges associated with microalgaebiofuel production [910] Although such microalgae strains are currently unavailable optimismpersists due to the fact that there is an abundance of species in the wild yet to be isolated [11] Indeedonly a few thousand (~50000) are conserved worldwide in microalgal culture collections [912] and ofthose only a few hundred (lt10) have been investigated for their biochemical composition [13]
Consequently the isolation selection biochemical and molecular characterization of microalgaefrom aquatic environments must be a continuous effort of screening for strains with the potentialfor biodiesel production [14] Therefore a sustainable development of microalgal-based biodieseland bioactive chemicals production in Peru requires the bioprospecting and exploitation of nativemicroalgae bioresources as the most viable option The northeastern region of Peru particularly theLoreto region contains a great variety of freshwater aquatic environments (ie rivers lagoons etc)that harbor a rich biodiversity of microalgae In this study 14 strains of native microalgae from thePeruvian Amazon were isolated identified and tested for their biomass productivity lipid productivityand total lipid content In addition the growth profile total carbohydrate total protein total lipidcontent and fatty acids profiles of three selected strains were also investigated and compared undernitrogen-sufficient and -deficient culture conditions
2 Experimental Section
21 Sample Collection and Isolation of Microalgae
The freshwater samples (100 mL) were collected from three river basins in the Loreto Regionof the Peruvian Amazon Amazon River (0341prime06prime prime S 7314prime89prime prime W) Itaya River (0343prime14prime prime S7314prime178prime prime W) and Nanay River (0342prime02prime prime S 7315prime32prime prime W) Microalgae samples (50 mL) wereenriched with 50 mL of Chu medium No 10 [15] The pH was adjusted to 75 with 1 M NaOH beforeautoclaving and the addition of microalgae The microalgal cells were maintained in Chu mediumNo 10 for four weeks in a controlled culture room at 25 plusmn 1 C with 1212-h light-dark cycles using80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 110 rpm Afterinitial cultivation of the mixed cultures unicellular microalgae were subjected to isolation by the cellwashing method [12] Briefly a 20-microL aliquot of the mixed microalgae culture was placed on a sterileglass slide under an inverted microscope to pick up a single microalgae cell with a Pasteur pipettecontaining a cotton-wool filter and connected to a flexible plastic hose Each microalgae cell was thensequentially washed in six 20-microL drops of Chu medium No 10 Subsequently the drop with a singlemicroalgae cell was transferred to a test tube with 1 mL of Chu medium No10 medium followed byinoculation onto Petri plates containing Chu medium No 10 supplemented with 15 (wv) of agarRepeated streaking on the nutrient agar plate and routine microscopic examination ensured the purityof the cultures Single individual colonies appearing on plates (~7 days) were inoculated into Chuliquid medium No 10 and grown for 14 days at the described conditions
22 Morphological Identification of Microalgae
Isolated strains were preliminarily identified using standard morphological features [16]Intracellular lipid droplets of isolated microalgae were detected by the fluorescence emitted byinteracting with Nile Red [17] Briefly 15 mL of microalgae cells culture were harvested by
Energies 2017 10 224 3 of 16
centrifugation at 1900times g for 10 min and washed two times with 1 mL of physiological saline solutionMicroalgal cell pellets were dissolved in Nile Red acetone solution (2 microgmL) and incubated for15 min at 37 C After washing once stained microalgal cells were observed by Carl Zeiss fluorescentmicroscopy and photographed with a digital camera AxioLabA1 AxioCamERc real time 5 s Imageswere obtained at a magnification of 1000times with visible light and epifluorescence (excitation 510ndash560emission 590)
23 Genomic DNA Isolation
Microalgae cells were harvested from 100 mL of liquid culture by centrifugation at 1900times g for10 min Genomic DNA was extracted using a modified version of the CTAB method as describedby Doyle and Doyle [18] Briefly algal cells were completely ground in a mortar containing 50 mgof sterilized sand and 3 mL of extraction buffer (300 mM Tris-HCl pH 80 50 mM ethylenediaminetetraacetic acid (EDTA) 2 M NaCl 2 cetyltrimethylammonium bromide 3 polyvinylpyrrolidone(MW 40000) and 2 β-mercaptoethanol) Homogenized cells were incubated at 70 C for 30 min withgentle inversion every 2 min An equal volume of phenolchloroformisoamyl alcohol (25241 vv)was then added followed by centrifugation at 20000times g The aqueous supernatant was transferred toa new microtube and an equal volume of chloroformisoamyl alcohol (241 vv) was added followedby centrifugation at 20000times g The aqueous supernatant was transferred to a new microtube andmixed with 01 volumes of 3 M sodium acetate (pH 52) and an equal volume of chilled isopropanolwas added to precipitate the DNA at 15000times g Washing was done with 70 alcohol and the air-driedDNA pellet was dissolved in 100 microL of sterilized water treated with RNase A at 40 C for 30 min andthen extracted with chloroformisoamyl alcohol and DNA precipitation with absolute ethanol Finallythe air-dried DNA pellet was dissolved in TE buffer (10 mM Tris-HCl pH 80 1 mM EDTA) and storedat minus20 C Genomic DNA quality and quantity were assessed by standard OD measurement [19]using a Nanodrop 2000 UV-Vis Spectrophotometer (Thermo Fisher Scientific Wilmington DE USA)In order to verify DNA integrity samples were resolved by standard gel electrophoresis in 12agarose gels [19] stained with ethidium bromide and visualized under ultraviolet light (SupplementaryMaterials Figure S1)
24 PCR Amplification and Sequencing of ITS2-rDNA Region
The ITS-2 rDNA gene was amplified according to Kaur et al [20] with minor modifications in20-microL reactions containing 1times PCR buffer 3 mM MgCl2 02 mM dNTPs mix 0025 UmicroL AmplitaqGold 10 microM each of the forward primer SQITS1 (5prime-GAGCATGTCTGCCTCAGC-3prime) and reverseprimer SQITS2 (5prime-GGTAGCCTTGCCTGAGC-3prime) and 20 ng of genomic DNA The general conditionsfor PCR in a thermal cycler Realplex S4 (Eppendorf Hamburg Germany) were 5 min at 94 C for thefirst cycle followed by 30 s at 94 C 30 s at 55 C and 1 min at 72 C for 35 cycles and a final 10-minextension step at 72 C The PCR gene products were separated on 2 agarose gels using 05 times TBEbuffer at 85 volts for 60 min and detected by staining with ethidium bromide (Figure S2) To purify4 microL of the PCR gene product were mixed with 2 microL of ExoSAP-IT (Affymetrix Santa Clara CAUSA) incubated at 37 C for 30 min and 80 C for 15 min and directly sequenced in both directionswith primers SQITS1 and SQITS2 following the instructions of the BigDyereg Terminator v31 CycleSequencing Kit (Applied Biosystemstrade Foster City CA USA) The sequencing products were purifiedby precipitation with ethanolEDTA denatured and resolved on a Genetic Analyzer 3130XL (AppliedBiosystemstrade Foster City CA USA) All sequences presented here were submitted to GenBank [21]
25 Phylogenetic Analysis
The ITS-2 rDNA gene sequences of the isolates were searched for homology in the National Centerfor Biotechnology Information database [22] using BLAST [23] Sequences were edited in BioEdit 725software [24] to trim the non-overlapping 5prime and 3prime regions for the purpose of restricting the multiplesequence alignment within the minimum common region Those edited sequences were aligned using
Energies 2017 10 224 4 of 16
Clustal omega [25] The unrooted phylogenetic tree was constructed using the neighbor-joining (NJ)method [26]] and Kimurarsquos two-parameter algorithm [27] as implemented within the MEGA 606software [28] The bootstrap analysis [29] was used with 1000 replications to test the relative supportfor the branches produced by NJ analysis
26 Microalgae Cultivation Biomass and Total Lipids Measurements
Two hundred milliliters Chu medium No 10 in a 500-mL Erlenmeyer flask were inoculatedwith the microalgae cells (OD680 005) and incubated at 25 plusmn 1 C using 80 microEmiddotmminus2middotsminus1 intensity ofcool-white fluorescent light and continuous agitation at 150 rpm for four weeks In this culture stage allmicroalgae strains were found in the stationary growth phase and centrifuged at 1920times g for 10 min toharvest the microalgae cells The cell pellets were rinsed with 50 mL deionized water centrifuged againdried in an oven at 70 C to a constant weight and the dry weight was determined gravimetricallywith an analytical balance Kern ABJ 220-4NM (Kern amp Sohn GmbH Balingen Germany) The algalpowders were stored at minus20 C for total lipid analysis
Total lipid was extracted using a modified protocol of Bligh and Dyer [30] For each sampleapproximately 50 mg of microalgal powder were mixed with 3 mL of chloroformmethanol (21 vv)and homogenized in an ultrasonic Branson 2510 (Branson Danbury CT USA) for 5 min The mixturewas then centrifuged at 20000times g for 5 min and the supernatant was transferred into a pre-weighedvial Cellular debris was mixed with another 2 mL of chloroformmethanol (21 vv) by vortex andthen centrifuged as described above After that the pooled organic phases were filtered throughsyringe filters of 045 microm evaporated with nitrogen gas and dried at 80 C in a vacuum ovenVacucell 55mdashComfort (MMM Muumlnchen Germany) to a constant weight The lipid content andproductivity of each sample were measured gravimetrically using a semi-micro analytical balance(Sartorius MSU225S-000-DU Foster City CA USA) and calculated as follows [31]
Total lipid ( dry weight) = WLWA times 100
where WL (g) is the weight of the total lipids extracted and WA (g) is the weight of the drymicroalgae biomass
PLipid (mgmiddotLminus1middotdminus1) = WA (g) times CLipid ()V(L) times T(d)
where PLipid is the lipid productivity CLipid is the lipid content V is the working volume and T is thecultivation time
27 Strains Culture under Nitrogen-Sufficient and Nitrogen-Deficient Medium
The selected microalgae species Acutodesmus obliquus (KP8785051) Ankistrodesmus sp(KP8785001) and Chlorella lewinii (KP8785011) (Figure S3) were grown in 500-mL Erlenmeyerflasks containing 250 mL of liquid Chu medium No 10 with nitrogen sufficient (5756 microgmL ofCa(NO3)2middot4H2O) or liquid Chu medium No 10 without a nitrogen source (deficient) for seven days ina controlled culture room at 25 plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity ofcool-white fluorescent light and continuous agitation at 150 rpm
For growth profile analysis the maximum absorption peak for each microalgae strain wasdetermined in the 600ndash800 nm range by spectrophotometric analysis using a Nanodrop 2000 UV-VisSpectrophotometer (Figure S4) Then aliquots of the microalgal cultures were evaluated every 24 h(between 8 and 9 h) at 680 nm The specific growth rate (micro) was calculated during the exponentialgrowth phase (the first three days of culture) according to the equation micro = (ln Nf minus ln Ni)(tf minus ti)where N is the optical density (OD680) at final (f) or initial (i) time (t) Previously it was determinedthat a high (R2 from 0992 to 0998) and positive correlation exists between optical density and celldensities this variable was calculated using a hemocytometer The experiments were carried out intriplicate and the data were expressed as the mean plusmn standard deviation (plusmnSD)
Energies 2017 10 224 5 of 16
28 Determination of Total Protein Total Carbohydrate Ash Content Esterification and Fatty Acids Profiling
The microalgae cultures maintained for seven days under +N and minusN Chu medium No 10were harvested by centrifugation rinsed and dried as previously described (Section 25) Fromthe microalgae powders total protein content was determined according to the HartreendashLowrymethod [32] To 5 mg of dry microalgal biomass was added 5 mL of 05 M NaOH and incubated for10 min at 80 C then centrifuged for 5 min at 14000times g Protein content in 1 mL of the supernatantwas determined by adding 05 mL of 05 M sodium carbonate 05 mL of 05 M sodium potassiumtartrate 05 M of copper sulfate and 2 mL of Folin-Ciocalteu reagent The blue complex was analyzedin a Varian Cary 50 Bio UV-Visible Spectrophotometer (Agilent Technologies Santa Clara CA USA)set at 650 nm against a calibration curve based on a known concentration of BSA as the standard
Total carbohydrate content of the microalgal biomass was determined with the phenol-sulfuricacid method [3233] For this purpose 5 mg of dry microalgae biomass were added to 5 mL of waterAfter this a 1-mL aliquot of the sample was added to 3 mL of sulfuric acid and 1 mL of 5 aqueoussolution of phenol and the mixture was stirred and incubated for 5 min at 90 C The yellow-browncomplex was analyzed by spectrophotometric analysis at 488 nm against a calibration curve based ona known concentration of glucose
Ash content was done according to the AOAC procedure [34] One hundred milligrams of drymaterial were ashed in a muffle furnace Thermolynetrade F6010 (Thermo Fisher Scientific WalthmanMA USA) at 550 C for 16 h and the content was determined gravimetrically All analyses werecarried out in triplicate and the data are expressed as the mean plusmn SD
Fatty acids methyl esters (FAME) were prepared by acid transesterification according to Ichiharaand Fukubayashi [35] The crude lipid extract (1 mg) was dissolved in 02 mL of toluene 15 mL ofmethanol and 03 mL of 08 HCl (prepared in methanolwater 8515 vv) transferred into a cappedtest tube mixed for 5 min and then incubated at 45 C for 12 h for derivatization reaction Then 2 mLof n-hexane and 2 mL of distilled water were added to the tube After vortexing the hexane phasescontaining FAMEs were transferred into a 15-mL tube and dried under a stream of nitrogen Finallythe FAMEs were redissolved in 10 microL acetonitrile and analyzed by using a gas chromatograph VarianCP-3800 GC (Agilent Technologies Santa Clara CA USA) equipped with an automated sampler andinjector a flame ionization detector and using a 30 m times 032 mm times 025 microm Stabilwaxreg capillarycolumn (Restek Bellefonte PA USA) The GC conditions were as follows injector temperature 250 Cthe column temperature gradient was 120 C for 1 min followed by an increase to 160 C at the rateof 30 Cmin 160 C for 1 min followed by an increase to 240 C at the rate of 4 Cmin and 240 Cfor 7 min detector temperature 260 C Gas pressures for He H2 and synthetic air were maintained at40 80 and 60 psi respectively The carrier gas (He) flow was maintained at 1 mLmin The fatty acidswere analyzed by comparing their retention time of the corresponding peaks with a known standardmixture of FAMEs (Nu-Check Prep Elysian MN USA) and methyl tricosanoate (Sigma-AldrichSaint Louis MO USA) was added to each sample as the internal standard All chromatograms ofthe microalgal samples were analyzed by using the Galaxietrade Chromatography Data System Version1932 (Agilent Technologies Santa Clara CA USA) All peaks spanning a peak area of more than50 units were integrated All analyses were carried out in triplicate and the data are expressed as themean plusmn SD
29 Statistical Analysis
Statistical analyses were performed using the IBM SPSS Statistics 210 software (IBM ArmonkNY USA) The growth rate biomass and lipid content data for microalgal strains were analyzedusing a one-way analysis of variance (ANOVA) and the HSD Tukey test The correlation of biomassproductivity and lipid content was analyzed using bivariate correlations and Pearsonrsquos correlationcoefficients are given with their significance levels Differences were considered significant at p lt 005All data presented here are given as the mean plusmn SD
Energies 2017 10 224 6 of 16
3 Results and Discussion
31 Isolation and Identification of the Microalgae Strains
Bioprospecting of tropical ecosystems can provide exceptional native microalgae strains withthe potential for the production of biodiesel nutraceuticals bioactive chemicals and other highvalue products [36ndash38] Furthermore native microalgae strains can have potential applications forefficient biological sequestration of CO2 wastewater treatment and other environmentally-friendlyapplications [38ndash40] Consequently isolation is a fundamental process to obtain pure cultures and isthe first phase towards the screening and selection of microalgae strains with the potential for biodieselproduction and the aforementioned applications
According to these demands in this research samples were collected from three different typesof water bodies the Amazon River (white-water type) the Itaya and Nanay Rivers both of theblack-water type The white-water rivers originate from the Andean catchments bringing high amountsof nutrient-rich sediments In contrast the black-water rivers drain geochemically-poor watershedsand are characterized by their high content in humic acids low pH and poor nutrient status [41]A total of 14 microalgal strains were isolated using the standard cell washing technique and basedon distinguishable morphological characters under light microscopic examination these strains werepreliminary ascribed to the genera Acutodesmus (Hegewald) Tsarenko 2001 Ankistrodesmus Corda1838 Chlorella Beyerinck Beijerinck 1890 Desmodesmus (Chodat) SS An T Friedl and E Hegewald1999 and Tetradesmus GM Smith 1913 (Table 1) These genera are green algae that belong todivision Chlorophyta class Chlorophyceae and orders Chlorellales and Sphaeropleales Althoughmorphological analysis is frequently used to identify microalgae it is inaccurate and very difficultfor the identification at the species level because the relationship between diagnostic morphologyand biological species boundaries are largely unknown in many micro-eukaryotes [42] Furthermoreaccording to Yu et al [31] the microalgae morphology for the same strain varies in relation to ageand culture conditions Consequently in microalgae the classic taxonomy is supported by moleculartools [42] For example the ITS2 rDNA region is thought to be an excellent marker for molecularphylogenetics studies in lower taxonomic levels due to being highly divergent and fast-evolvingwhich can discriminate among closely-related organisms which otherwise display almost identicalsequences [4344] For this reason the molecular identification based in phylogenetic analysis of ITS2rDNA gene sequences is commonly used to identify andor delineate species [204546]
Table 1 Genera GenBank accession number of ITS2 sequences biomass productivity lipid productivityand lipid content of isolated microalgae strains
Source ofCollection Strain GeneraSpecies
GenBankAccessionNumber
BiomassProductivity
(mgmiddotLminus1middotdminus1)
LipidProductivity
(mgmiddotLminus1middotdminus1)
LipidContent( dw)
Amazon River(0341prime06prime prime S7314prime89prime prime W)
AMA001 Ankistrodesmus sp KP8784961 122 plusmn 03 37 plusmn 01 307 plusmn 15AMA002 Ankistrodesmus sp KP8784971 36 plusmn 04 10 plusmn 02 277 plusmn 21AMA003 Chlorella lewinii KP8785011 223 plusmn 07 51 plusmn 05 227 plusmn 15AMA004 Acutodesmus abliquus KP8785051 316 plusmn 04 64 plusmn 07 203 plusmn 21
Itaya River(0343prime14prime prime S
7314prime178prime prime W)
ITA001 Ankistrodesmus sp KP8784991 59 plusmn 02 40 plusmn 03 437 plusmn 17ITA002 Ankistrodesmus sp KP8785001 136 plusmn 04 57 plusmn 05 306 plusmn 20ITA003 Chlorella lewinii KP8785031 129 plusmn 03 40 plusmn 05 224 plusmn 27ITA004 Chlorella lewinii KP8785041 221 plusmn 07 31 plusmn 04 215 plusmn 23ITA005 Scenedesmus regularis KP8785081 93 plusmn 04 56 plusmn 04 242 plusmn 14ITA006 Scenedesmus dimorphus KP8785091 186 plusmn 06 32 plusmn 03 260 plusmn 17ITA007 Acutodesmus abliquus KP8785101 179 plusmn 04 22 plusmn 03 156 plusmn 19
Nanay River(0342prime02prime prime S7315prime32prime prime W)
NAN001 Ankistrodesmus sp KP8784981 143 plusmn 06 28 plusmn 02 467 plusmn 15NAN002 Desmodesmus tropicus KP8785061 122 plusmn 04 23 plusmn 02 175 plusmn 11NAN003 Tetradesmus sp KP8785071 139 plusmn 04 30 plusmn 03 135 plusmn 12
Energies 2017 10 224 7 of 16
To realize phylogenetic analysis genomic DNA of each isolate strain was purified and used forPCR amplification of ITS2 rDNA The universal primers used to amplify the ITS2 rDNA successfullyamplified the desired DNA fragments (~300 bp) from all microalgal strains (Figure S2) Furtherthese amplicons were sequenced published in the NCBI databases and the corresponding GenBankaccession numbers are presented in Table 1 Figure 1 shows a neighbor-joining phylogenetic treedepicting the relatedness of the native microalgae strain isolates with selected sequences from thepublic GenBank database This phylogram contained three clusters each of which included differentmicroalgae genera For example the first cluster consisted of microalgae of the genera AcutodesmusScenedesmus Tetradesmus and Desmodesmus and six isolated strains (ITA007 AMA004 ITA006 NAN003ITA005 and NAN002) The second cluster consisted of a microalgae of the genus Ankistrodesmus andthe isolated strains AMA001 ITA001 ITA002 NAN001 and AMA002 The third one consisted of amicroalgae of the genus Chlorella and three isolated strains (ITA004 AMA003 and ITA003) Overall theidentification of native microalgae strains presented a good correspondence between morphologicalcharacters and sequence-based phylogenetic analysis Microalgae species of these genera were alsoisolated from different countries of the world such as Brazil [47] Argentina [46] United States [48]India [20] Egypt [49] Taiwan [50] South Korea [45] and China [51] Some of these microalgae generasuch as Ankistrodesmus Chlorella Desmodesmus and Scenedesmus were used in large-scale biodieselproduction using photobioreactors and raceway pond systems [5253]
Energies 2017 10 224 7 of 16
public GenBank database This phylogram contained three clusters each of which included different microalgae genera For example the first cluster consisted of microalgae of the genera Acutodesmus Scenedesmus Tetradesmus and Desmodesmus and six isolated strains (ITA007 AMA004 ITA006 NAN003 ITA005 and NAN002) The second cluster consisted of a microalgae of the genus Ankistrodesmus and the isolated strains AMA001 ITA001 ITA002 NAN001 and AMA002 The third one consisted of a microalgae of the genus Chlorella and three isolated strains (ITA004 AMA003 and ITA003) Overall the identification of native microalgae strains presented a good correspondence between morphological characters and sequence-based phylogenetic analysis Microalgae species of these genera were also isolated from different countries of the world such as Brazil [47] Argentina [46] United States [48] India [20] Egypt [49] Taiwan [50] South Korea [45] and China [51] Some of these microalgae genera such as Ankistrodesmus Chlorella Desmodesmus and Scenedesmus were used in large-scale biodiesel production using photobioreactors and raceway pond systems [5253]
Figure 1 Unrooted phylogenetic tree of isolated microalgae strains using the ITS2 sequences The tree was constructed by the neighbor-joining method and Kimurarsquos two-parameter algorithm in MEGA 60 software based on the multiple sequence alignment by Clustal Omega Bootstrap values of 1000 replicates () are shown at the branches
32 Microalgae Cultivation Biomass and Total Lipids Measurements
The isolated microalgae strains exhibited remarkable differences in productivity parameters and lipid content among individual strains in the stationary growth phase (Table 1) With regard to the biomass productivity the 14 strains showed statistically-significant differences (F = 71911 df = 13 p lt 0001) and ranged from 36 plusmn 04 mgmiddotLminus1middotdminus1 (Ankistrodesmus sp strain AMA002) to 316 plusmn 04 mgmiddotLminus1middotdminus1 (Acutodesmus obliquus strain AMA004) Similar biomass productivities were reported for other marine microalgae species [54] However other investigations show major biomass productivities (ranging
Figure 1 Unrooted phylogenetic tree of isolated microalgae strains using the ITS2 sequences The treewas constructed by the neighbor-joining method and Kimurarsquos two-parameter algorithm in MEGA60 software based on the multiple sequence alignment by Clustal Omega Bootstrap values of1000 replicates () are shown at the branches
Energies 2017 10 224 8 of 16
32 Microalgae Cultivation Biomass and Total Lipids Measurements
The isolated microalgae strains exhibited remarkable differences in productivity parametersand lipid content among individual strains in the stationary growth phase (Table 1) With regardto the biomass productivity the 14 strains showed statistically-significant differences (F = 71911df = 13 p lt 0001) and ranged from 36 plusmn 04 mgmiddotLminus1middotdminus1 (Ankistrodesmus sp strain AMA002) to316 plusmn 04 mgmiddotLminus1middotdminus1 (Acutodesmus obliquus strain AMA004) Similar biomass productivities werereported for other marine microalgae species [54] However other investigations show major biomassproductivities (ranging from 30 to 120 mgmiddotLminus1middotdminus1) in the same and other microalgae genera [455556]These discrepancies may be attributed to differences in the culture conditions in the culture mediacomposition and in the microalgae genotypes Lipid productivity also exhibited statistically-significantdifferences (F = 4774 df = 13 p lt 0001) and ranged from 10plusmn 04 mgmiddotLminus1middotdminus1 to 64 plusmn 07 mgmiddotLminus1middotdminus1
with strains AMA002 and AMA004 respectively These lipid productivities were low in comparison toother reports which reported values from 190 to 433 mgmiddotLminus1middotdminus1 [4555] Furthermore lipid contentas a percentage of the microalgal biomass dry weight showed statistically-significant differences(F = 8509 df = 13 p lt 0001) and ranged from 135 plusmn 12 (Tetradesmus sp strain NAN003) to467 plusmn 15 (Ankistrodesmus sp strain NAN001) These percentage values in total lipid contentare similar to several microalgae species of these genera isolated and characterized by otherresearchers worldwide [20454648ndash5057] Furthermore it is important to emphasize that lipid contentis dependent on the different growth phases Commonly in several microalgae taxa the lipid contentincreases gradually from logarithmic to stationary growth phases [58ndash60] For example Kaur et al [20]reported that the total lipid content of Scenedesmus sp DRLMA9 demonstrated values of 207 inthe late exponential phase and 397 in the stationary phase and Desmodesmus elegans DRLMA13displayed values of 103 in the late exponential phase and 169 in the stationary phase LikewiseChiu et al [61] found that the total lipid content of Nannochloropsis oculata was 308 in the logarithmicphase 397 in the early stationary phase and 504 in the stationary phase These changes in lipidcontent with culture age have been associated with nitrate depletion [61] which causes a channeling ofphotosynthetic ATP to carbohydrate and lipid biosynthesis [60] Finally Pearson correlation analysisof biomass productivity and lipid content was inversely related (r = minus037 p lt 001) which is similarto the results obtained by Liang et al [62] and Rodolfi et al [52] In contrast Li et al [51] andHempel et al [63] found no relationship between these parameters The negative correlation obtainedis likely due to the high metabolic cost of lipid biosynthesis [62]
33 Strains Culture under Nitrogen-Sufficient and Nitrogen-Deficient Medium
The microalgae species Acutodesmus obliquus (KP8785051) Ankistrodesmus sp (KP8785001)and Chlorella lewinii (KP8785011) were selected and further characterized because they displayedthe highest biomass and lipid productivities (Table 1) These microalgae showed noticeablevariations in the growth profile when cultured in Chu medium No 10 with nitrogen (+N) orwithout a nitrogen source (minusN) In +N medium all strains showed exponential growth butChlorella lewinii showed the highest average specific growth rates (micro = 0458 plusmn 0098 dminus1) followed byAcutodesmus obliquus (micro = 0399 plusmn 0112 dminus1) and the lowest value was produced by Ankistrodesmus sp(micro = 0265 plusmn 0013 dminus1) In contrast all cultures displayed a lag phase pattern in their growthprofile under minusN medium (Figure 2) These findings corroborate the results obtained by otherresearchers who demonstrate that the specific growth rate is directly proportional to the initialnitrate concentration [46596465] because the reduction or cessation of microalgae cell divisionis due to nitrogen limitation [66] which is a key nutrient for protein and nucleic acids de novobiosynthesis consequently causing an arrest in the early part of the microalgae cell cycle [6768]In accordance with these reports Li et al [69] showed that nitrogen deficiency induced in the greenmicroalgae Micractinium pusillum a downregulation in the expression of genes involved in cell growthand cell cycle regulation Additionally differences in growth profile and specific growth rates aredependent on several factors such as microalgae genotypes the culture medium composition pH
Energies 2017 10 224 9 of 16
nutrients concentration the cell culture phase (ie log phase stationary phase) and environmentalfactors [4549545770ndash72]Energies 2017 10 224 9 of 16
Figure 2 Growth profiles of three microalgal strains cultivated under +N (filled boxes) and minusN (empty boxes) Chu medium No 10 The culture was incubated in a controlled culture room at 25 plusmn 1 degC with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 150 rpm Experiments were carried out in triplicate
34 Total Protein Total Lipid and Total Carbohydrate Content
Biochemical parameters of the microalgae species such as total protein total lipid and total carbohydrate content exhibited differences and changes under both culture conditions (Figure 3) For example Chlorella lewinii showed the highest protein content Ankistrodesmus sp had the highest total lipid content but Acutodesmus obliquus presented the highest levels of total carbohydrate Therefore these strains can be valuable stocks for the industry because total protein and total carbohydrate are potentially byproducts of microalgal biomass processing [7374] Additionally the three strains cultured under minusN conditions showed a significant increase in total lipid content (from 237 in Acutodesmus obliquus to 667 in Ankistrodesmus sp) and total carbohydrate content (from 119 in Acutodesmus obliquus to 963 in Chlorella lewinii) but showed a decreased in total protein levels (from 242 in Acutodesmus obliquus to 545 in Chlorella lewinii) in comparison to +N cultures (Figure 3) These results are in accordance with well-documented results obtained by other researchers with several microalgae species of the genera Acutodesmus Ankistrodesmus Botryococcus Chlamydomonas Chlorella Desmodesmus Dunaliella Haematococcus Neochloris Phaeodactylum Pseudokirchneriella and Scenedesmus [465058596475ndash80] Nevertheless the accumulation of total lipid induced by nitrogen deficiency is not correlated with lipid productivity but rather is due to a decline and stopping of the microalgae specific growth rate [75] The changes in biomolecule contents suggest redirection of microalgae metabolism during nitrogen starvation such as a decrease in biosynthesisincrease in protein degradation and an increase in the biosynthesis of storage biomolecules such as triacylglycerols and starch The increase in total lipid content could be partially explained for a boost in transcript levels of genes encoding enzymes of the lipid biosynthesis pathways According to this assumption recent investigations showed that genes encoding the enzymes that catalyze the last step in the Kennedy pathway of triacylglycerol biosynthesis [81] such as glycerol-3-phosphate acyltransferase and acyl-glycerol-3-phosphate acyltransferase are upregulated in response to nitrogen starvation [648283] Furthermore the increase in total carbohydrate content can be explained by changes in the expression levels of genes involved in carbohydrate metabolism For example when the oleaginous microalgae Neochloris oleoabundans is cultured under nitrogen starvation the catabolic phase of starch specifically α-amylase which hydrolyzes starch to glucose is strongly repressed [83]
Figure 2 Growth profiles of three microalgal strains cultivated under +N (filled boxes) and minusN(empty boxes) Chu medium No 10 The culture was incubated in a controlled culture room at25 plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent lightand continuous agitation at 150 rpm Experiments were carried out in triplicate
34 Total Protein Total Lipid and Total Carbohydrate Content
Biochemical parameters of the microalgae species such as total protein total lipid and totalcarbohydrate content exhibited differences and changes under both culture conditions (Figure 3)For example Chlorella lewinii showed the highest protein content Ankistrodesmus sp had the highesttotal lipid content but Acutodesmus obliquus presented the highest levels of total carbohydrateTherefore these strains can be valuable stocks for the industry because total protein and totalcarbohydrate are potentially byproducts of microalgal biomass processing [7374] Additionallythe three strains cultured under minusN conditions showed a significant increase in total lipid content(from 237 in Acutodesmus obliquus to 667 in Ankistrodesmus sp) and total carbohydrate content(from 119 in Acutodesmus obliquus to 963 in Chlorella lewinii) but showed a decreased in totalprotein levels (from 242 in Acutodesmus obliquus to 545 in Chlorella lewinii) in comparison to +Ncultures (Figure 3) These results are in accordance with well-documented results obtained by otherresearchers with several microalgae species of the genera Acutodesmus Ankistrodesmus BotryococcusChlamydomonas Chlorella Desmodesmus Dunaliella Haematococcus Neochloris PhaeodactylumPseudokirchneriella and Scenedesmus [465058596475ndash80] Nevertheless the accumulation of totallipid induced by nitrogen deficiency is not correlated with lipid productivity but rather is due toa decline and stopping of the microalgae specific growth rate [75] The changes in biomoleculecontents suggest redirection of microalgae metabolism during nitrogen starvation such as a decreasein biosynthesisincrease in protein degradation and an increase in the biosynthesis of storagebiomolecules such as triacylglycerols and starch The increase in total lipid content could bepartially explained for a boost in transcript levels of genes encoding enzymes of the lipid biosynthesispathways According to this assumption recent investigations showed that genes encoding theenzymes that catalyze the last step in the Kennedy pathway of triacylglycerol biosynthesis [81] such asglycerol-3-phosphate acyltransferase and acyl-glycerol-3-phosphate acyltransferase are upregulated
Energies 2017 10 224 10 of 16
in response to nitrogen starvation [648283] Furthermore the increase in total carbohydrate contentcan be explained by changes in the expression levels of genes involved in carbohydrate metabolismFor example when the oleaginous microalgae Neochloris oleoabundans is cultured under nitrogenstarvation the catabolic phase of starch specifically α-amylase which hydrolyzes starch to glucoseis strongly repressed [83]Energies 2017 10 224 10 of 16
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strains cultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled culture room at 25 plusmn 1 degC with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized with respect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that each microalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majority of fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised 7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii the majority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9) acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectively Finally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic (182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in +N and minusN conditions respectively Our results agree with multiple previously-published reports in that fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent in many green microalgae species [454648ndash5055] It is notable that under nitrogen starvation culture all microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 in Ankistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid (181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findings support the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content of SFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Nevertheless there are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed a decrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increased between 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively However in the same culture condition the three microalgae species showed a decrease in polyunsaturated fatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmus spp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgae strains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFA which improves the quality properties of biodiesel such as an increase
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strainscultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled cultureroom at 25plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescentlight and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized withrespect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that eachmicroalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majorityof fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii themajority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9)acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectivelyFinally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic(182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in+N and minusN conditions respectively Our results agree with multiple previously-published reports inthat fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent inmany green microalgae species [454648ndash5055] It is notable that under nitrogen starvation cultureall microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 inAnkistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid(181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findingssupport the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content ofSFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other
Energies 2017 10 224 11 of 16
researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Neverthelessthere are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed adecrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increasedbetween 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively Howeverin the same culture condition the three microalgae species showed a decrease in polyunsaturatedfatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmusspp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgaestrains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFAwhich improves the quality properties of biodiesel such as an increase in cetane number [84ndash86]Considering the SFA and unsaturated fatty acids (UFA) components approximately 20ndash31 are SFAand 69ndash80 are UFA Hence the SFAUFA ratio in the microalgae strains evaluated ranges between025 and 045 which is similar to the reports of several microalgae species isolated in different latitudesworldwide [203148497072]
Table 2 Effect of nitrogen deficiency on fatty acid profiling in three native microalgal species from thePeruvian Amazon
Fatty AcidsAcutodesmus obliquus Ankistrodesmus sp Chlorella lewinii
+N minusN +N minusN +N minusN
140 - - 052 plusmn 002 088 plusmn 002 - 058 plusmn 001160 2724 plusmn 007 2867 plusmn 017 1868 plusmn 019 2261 plusmn 006 1964 plusmn 021 2123 plusmn 010180 184 plusmn 005 255 plusmn 051 101 plusmn 002 181 plusmn 002 371 plusmn 026 153 plusmn 015
161n-7 023 plusmn 000 025 plusmn 001 071 plusmn 003 055 plusmn 001 085 plusmn 043 038 plusmn 000181n-7 131 plusmn 002 131 plusmn 000 141 plusmn 005 094 plusmn 003 146 plusmn 006 123 plusmn 002181n-9 3213 plusmn 021 3608 plusmn 021 3699 plusmn 004 4047 plusmn 038 1371 plusmn 015 2326 plusmn 041182n-6 1732 plusmn 004 1651 plusmn 016 747 plusmn 018 656 plusmn 007 2034 plusmn 039 1957 plusmn 001183n-3 1130 plusmn 017 854 plusmn 031 1486 plusmn 004 1173 plusmn 012 1815 plusmn 022 1371 plusmn 016183n-6 101 plusmn 001 078 plusmn 002 055 plusmn 000 054 plusmn 002 124 plusmn 006 100 plusmn 020184n-3 250 plusmn 005 177 plusmn 008 631 plusmn 002 601 plusmn 005 491 plusmn 025 403 plusmn 020204n-6 - - - 084 plusmn 008 - -225n-3 - - - 062 plusmn 002 - -
Unidentified 512 plusmn 003 355 plusmn 006 1149 plusmn 004 645 plusmn 003 1599 plusmn 034 1348 plusmn 006SFA 2907 3122 2022 2531 2335 2334UFA 7093 6878 7978 7469 7665 7666
MUFA 3367 3764 3911 4195 1602 2487PUFA 3213 2759 2918 2629 4464 3831SFA +MUFA 6274 6886 5933 6726 3936 4821
It is important to highlight that the fatty acid profile determination is an essential step inthe process of the characterization of microalgae strains because it ultimately affects the qualityof the biodiesel product [8485] The quality properties of biodiesel such as high cetane numberoxidative stability and cold-flow are dependent on the hydrocarbonated chain length of saturated andunsaturated fatty acids [8587] For example biodiesel composed of a mixture of a high percentage ofSFA and MUFA is preferred for increased energy yield superior oxidative stability and higher cetanenumbers but is prone to solidify at low temperature In contrast biodiesel with high levels of PUFAhas decreased energy yield low cetane numbers and excellent cold-flow properties but insufficientoxidative stability [548588] Therefore to solve these performance problems five approaches havebeen developed such as changing the fatty acids profile by physical means genetic modification ofthe feedstock or the use of alternative feedstocks with different fatty acids profiles [89]
4 Conclusions
Fourteen microalgae were isolated from three river basins from the Peruvian Amazon and wereidentified by morphological features and molecular phylogeny approaches These strains were initially
Energies 2017 10 224 12 of 16
characterized in the stationary growth phase and exhibited significant heterogeneity in key parameterssuch as biomass productivity (36ndash316 mgmiddotLminus1middotdminus1) lipid productivity (10ndash64 mgmiddotLminus1middotdminus1) andtotal lipid content (135ndash467) Furthermore three selected strains based on elevated biomass andlipid productivities were cultured under nitrogen-sufficient and nitrogen-deficient medium to boostlipid accumulation Under these culture conditions all strains showed noticeable decreases in totalprotein content and increases in total lipid and total carbohydrate contents In addition these strainspresented modifications in the fatty acids profiles with improvement in biodiesel properties due to apredominance of SFA and MUFA which corresponds to an increase in cetane number The results ofthis research suggest that the isolated native microalgae from the Peruvian Amazon could be suitablerenewable feedstocks for the production of both biodiesel and bioethanol which could contribute tosustainable development at the local regional and national levels as a viable alternative to petroleumexploitation However it will be necessary to develop open closed or hybrid systems for large-scalebiomass production of selected native microalgae strains andor de novo microalgae strains designedto produce biofuels and with the abilities to synthesize useful bioactive compounds to be efficientlyexploited using biorefinery approaches
Supplementary MaterialsThe following are available online at wwwmdpicom1996-1073102224s1 Figure S1 Results of
electrophoretic and spectrophotometric analysis of the genomic DNA isolated from the fourteen microalgaeFigure S2 ITS2-rDNA structure (A) and amplicons obtained (B) from the microalgae isolated Figure S3Microphotographic images of the three microalgae strains analyzed under light and fluorescence stained withNile Red Figure S4 Spectrophotometric scanning for the determination of the maximum absorption peak foreach microalgae strain
Acknowledgments This research was supported by the Peruvian funding agencies Consejo Nacional de CienciaTecnologiacutea e Innovacioacuten Tecnoloacutegica (CONCYTEC) with Grant Contract No 364-CONCYTEC-OAJ and Fondospara la Innovacioacuten Ciencia y Tecnologiacutea (FINCyT) with Grant Contract No 383-PNICP-PIBA-2014
Author Contributions Marianela Cobos and Juan C Castro obtained funds for the research participated inthe study design coordinated activities and participated in the preparation of the manuscript Jae D Paredesparticipated in sample collection microalgae isolation and helped to draft the manuscript J Dylan Maddoxperformed the bioinformatics analysis and participated in the preparation of the manuscript Gabriel Vargas-Aranaand Leenin Flores participated in biochemical analysis and helped to draft the manuscript Carla P Aguilarparticipated in the study design supervised biochemical analysis and helped to draft the manuscriptJorge L Marapara participated in molecular analysis and helped to draft the manuscript
Conflicts of Interest The authors declare no conflict of interest
References
1 BP Global BP Statisical Review of World Energy 2014 Available online httpwwwbpcomcontentdambppdfEnergy-economicsstatistical-review-2014BP-statistical-review-of-world-energy-2014-full-reportpdf (accessed on 20 June 2014)
2 Pachauri RK Reisinger A Cambio Climaacutetico 2007 Informe de Siacutentesis Contribucioacuten de los Grupos de Trabajo III y III al Cuarto Informe de Evaluacioacuten del Grupo Intergubernamental de Expertos Sobre el Cambio Climaacutetico IPCCGeneva Switzerland 2007 p 104
3 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Ley de Promocioacuten del Mercado deBiocombustibles Available online httpwww2congresogobpesicrcendocbibcon4_uibdnsf9E95620CC059138105257C9E005AB2F9$FILE28054pdf (accessed on 12 February 2017)
4 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Reglamento de la Ley de Promocioacuten del Mercadode Biocombustibles Available online httpwww2osinerggobpeMarcoLegaldocrevDS-013-2005-EM-CONCORDADOpdf (accessed on 12 February 2017)
5 Areacutevalo LF Nalvarte J Torres J Ramiacuterez Y Impactos Socio-Econoacutemicos de la Produccioacuten de Biocombustiblesen la Amazoniacutea Peruana Primera SNV-WWF Lima Peruacute 2009
6 Sharma KK Schuhmann H Schenk PM High Lipid Induction in Microalgae for Biodiesel ProductionEnergies 2012 5 1532ndash1553 [CrossRef]
Energies 2017 10 224 13 of 16
7 Schenk PM Thomas-Hall SR Stephens E Marx UC Mussgnug JH Posten C Kruse O Hankamer BSecond Generation Biofuels High-Efficiency Microalgae for Biodiesel Production BioEnergy Res 2008 120ndash43 [CrossRef]
8 Foley JA Asner GP Costa MH Coe MT de Fries R Gibbs HK Howard EA Olson S Patz JRamankutty N et al Amazonia revealed Forest degradation and loss of ecosystem goods and services inthe Amazon Basin Front Ecol Environ 2007 5 25ndash32 [CrossRef]
9 Duong VT Li Y Nowak E Schenk PM Microalgae Isolation and Selection for Prospective BiodieselProduction Energies 2012 5 1835ndash1849 [CrossRef]
10 Georgianna DR Mayfield SP Exploiting diversity and synthetic biology for the production of algalbiofuels Nature 2012 488 329ndash335 [CrossRef] [PubMed]
11 Wijffels RH Barbosa MJ An Outlook on Microalgal Biofuels Science 2010 329 796ndash799 [CrossRef][PubMed]
12 Richmond A Handbook of Microalgal Culture Biotechnology and Applied Phycology Blackwell Sci Ltd HudsonNJ USA 2004
13 Menetrez MY An overview of algae biofuel production and potential environmental impactEnviron Sci Technol 2012 46 7073ndash7085 [CrossRef] [PubMed]
14 Khoo HH Sharratt PN Das P Balasubramanian RK Naraharisetti PK Shaik S Life cycle energy andCO2 analysis of microalgae-to-biodiesel Preliminary results and comparisons Bioresour Technol 2011 1025800ndash5807 [CrossRef] [PubMed]
15 Chu SP The Influence of the Mineral Composition of the Medium on the Growth of Planktonic AlgaePart I Methods and Culture Media J Ecol 1942 30 284ndash325 [CrossRef]
16 Carlos EMB Mariatildengela M Gecircneros de Algas de Aacuteguas Continentais do BrasilmdashChave Para Identificaccedilatildeo eDescriccedilotildees 2nd ed Rima Sao Carlos Brasil 2006
17 Greenspan P Mayer EP Fowler SD Nile red A selective fluorescent stain for intracellular lipid dropletsJ Cell Biol 1985 100 965ndash973 [CrossRef] [PubMed]
18 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissuePhytochem Bull 1987 19 11ndash15
19 Sambrook J Fritsch EF Maniatis T Molecular Cloning A Laboratory Manual 2nd ed Cold Spring HarborLaboratory Press New York NY USA 1989
20 Kaur S Sarkar M Srivastava RB Gogoi HK Kalita MC Fatty acid profiling and molecularcharacterization of some freshwater microalgae from India with potential for biodiesel productionNew Biotechnol 2012 29 332ndash344 [CrossRef] [PubMed]
21 Benson DA Cavanaugh M Clark K Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW GenBankNucleic Acids Res 2013 41 D36ndashD42 [CrossRef] [PubMed]
22 NCBI Resource Coordinators Database Resources of the National Center for Biotechnology InformationNucleic Acids Res 2015 43 D6ndashD17
23 Ye J McGinnis S Madden TL BLAST Improvements for better sequence analysis Nucleic Acids Res 200634 W6ndashW9 [CrossRef] [PubMed]
24 BioEdit Sequence Alignment Editor for Windows 9598NTXPVista7 Available online httpwwwmbioncsuedubioeditbioedithtml (accessed on 2 December 2015)
25 Sievers F Higgins DG Clustal omega accurate alignment of very large numbers of sequencesMethods Mol Biol (Clifton NJ) 2014 1079 105ndash116
26 Saitou N Nei M The neighbor-joining method A new method for reconstructing phylogenetic treesMol Biol Evol 1987 4 406ndash425 [PubMed]
27 Kimura M A simple method for estimating evolutionary rates of base substitutions through comparativestudies of nucleotide sequences J Mol Evol 1980 16 111ndash120 [CrossRef] [PubMed]
28 Tamura K Stecher G Peterson D Filipski A Kumar S MEGA6 Molecular Evolutionary GeneticsAnalysis version 60 Mol Biol Evol 2013 30 2725ndash2729 [CrossRef] [PubMed]
29 Felsenstein J Confidence Limits on Phylogenies An Approach Using the Bootstrap Evolution 1985 39783ndash791 [CrossRef]
30 Bligh EG Dyer WJ A Rapid Method of Total Lipid Extraction and Purification Can J Biochem Physiol1959 37 911ndash917 [CrossRef] [PubMed]
Energies 2017 10 224 14 of 16
31 Yu X Zhao P He C Li J Tang X Zhou J Huang Z Isolation of a novel strain of Monoraphidium spand characterization of its potential application as biodiesel feedstock Bioresour Technol 2012 121 256ndash262[CrossRef] [PubMed]
32 Hartree EF Determination of protein A modification of the lowry method that gives a linear photometricresponse Anal Biochem 1972 48 422ndash427 [CrossRef]
33 DuBois M Gilles KA Hamilton JK Rebers PA Smith F Colorimetric Method for Determination ofSugars and Related Substances Anal Chem 1956 28 350ndash356 [CrossRef]
34 AOAC International AOAC Official Methods of Analysis of the Association of Official Analytical Chemists (AOAC)15th ed AOAC International Washington DC USA 1990 Volume 1
35 Ichihara K Fukubayashi Y Preparation of fatty acid methyl esters for gas-liquid chromatographyJ Lipid Res 2010 51 635ndash640 [CrossRef] [PubMed]
36 Brennan L Owende P Biofuels from microalgaemdashA review of technologies for production processingand extractions of biofuels and co-products Renew Sustain Energ Rev 2010 14 557ndash577 [CrossRef]
37 Raja R Hemaiswarya S Kumar NA Sridhar S Rengasamy R A perspective on the biotechnologicalpotential of microalgae Crit Rev Microbiol 2008 34 77ndash88 [CrossRef] [PubMed]
38 Mata TM Martins AA Caetano NS Microalgae for biodiesel production and other applicationsA review Renew Sustain Energy Rev 2010 14 217ndash232 [CrossRef]
39 De Wilt A Butkovskyi A Tuantet K Leal LH Fernandes TV Langenhoff A Zeeman GMicropollutant removal in an algal treatment system fed with source separated wastewater streamsJ Hazard Mater 2015 304 84ndash92 [CrossRef] [PubMed]
40 Li T Lin G Podola B Melkonian M Continuous removal of zinc from wastewater and mine dumpleachate by a microalgal biofilm PSBR J Hazard Mater 2015 297 112ndash118 [CrossRef] [PubMed]
41 Sioli H The Amazon and its main affluents Hydrography morphology of the river courses and river typesIn Monographiae Biologicae Sioli H Ed Springer Dordrecht The Netherlands 1984 pp 127ndash165
42 Moniz MBJ Kaczmarska I Barcoding of Diatoms Nuclear Encoded ITS Revisited Protist 2010 161 7ndash34[CrossRef] [PubMed]
43 Coleman AW Is there a molecular key to the level of ldquobiological speciesrdquo in eukaryotes A DNA guideMol Phylogenet Evol 2009 50 197ndash203 [CrossRef] [PubMed]
44 Mai JC Coleman AW The Internal Transcribed Spacer 2 Exhibits a Common Secondary Structure inGreen Algae and Flowering Plants J Mol Evol 1997 44 258ndash271 [CrossRef] [PubMed]
45 Abou-Shanab RAI Matter IA Kim S-N Oh Y-K Choi J Jeon B-H Characterization andidentification of lipid-producing microalgae species isolated from a freshwater lake Biomass Bioenergy2011 35 3079ndash3085 [CrossRef]
46 Do Nascimento M Ortiz-Marquez JCF Sanchez-Rizza L Echarte MM Curatti L Bioprospectingfor fast growing and biomass characterization of oleaginous microalgae from South-Eastern Buenos AiresArgentina Bioresour Technol 2012 125 283ndash290 [CrossRef] [PubMed]
47 Araujo GS Matos LJBL Gonccedilalves LRB Fernandes FAN Farias WRL Bioprospecting foroil producing microalgal strains Evaluation of oil and biomass production for ten microalgal strainsBioresour Technol 2011 102 5248ndash5250 [CrossRef] [PubMed]
48 Zhou W Li Y Min M Hu B Chen P Ruan R Local bioprospecting for high-lipid producing microalgalstrains to be grown on concentrated municipal wastewater for biofuel production Bioresour Technol 2011102 6909ndash6919 [CrossRef] [PubMed]
49 Mahmoud EA Farahat LA Abdel Aziz ZK Fatthallah NA Salah El Din RA Evaluation of thepotential for some isolated microalgae to produce biodiesel Egypt J Pet 2015 24 97ndash101 [CrossRef]
50 Ho S Lai Y-Y Chiang C-Y Chen C-NN Chang J-S Selection of elite microalgae for biodieselproduction in tropical conditions using a standardized platform Bioresour Technol 2013 147 135ndash142[CrossRef] [PubMed]
51 Li L Cui J Liu Q Ding Y Liu J Screening and phylogenetic analysis of lipid-rich microalgae Algal Res2015 11 381ndash386 [CrossRef]
52 Rodolfi L Chini Zittelli G Bassi N Padovani G Biondi N Bonini G Tredici MR Microalgae for oilStrain selection induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactorBiotechnol Bioeng 2009 102 100ndash112 [CrossRef] [PubMed]
Energies 2017 10 224 15 of 16
53 Li P Miao X Li R Zhong J In situ biodiesel production from fast-growing and high oil content Chlorellapyrenoidosa in rice straw hydrolysate J Biomed Biotechnol 2011 2011 141207 [CrossRef] [PubMed]
54 Doan TTY Sivaloganathan B Obbard JP Screening of marine microalgae for biodiesel feedstockBiomass Bioenergy 2011 35 2534ndash2544 [CrossRef]
55 Abou-Shanab RAI Hwang J-H Cho Y Min B Jeon B-H Characterization of microalgal speciesisolated from fresh water bodies as a potential source for biodiesel production Appl Energy 2011 883300ndash3306 [CrossRef]
56 Chaichalerm S Pokethitiyook P Yuan W Meetam M Sritong K Pugkaew W Kungvansaichol KKruatrachue M Damrongphol P Culture of microalgal strains isolated from natural habitats in Thailand invarious enriched media Appl Energy 2012 89 296ndash302 [CrossRef]
57 Zhang S Liu P Yang X Hao Z Zhang L Luo N Shi J Isolation and identification by 18S rDNAsequence of high lipid potential microalgal species for fuel production in Hainan Dao Biomass Bioenergy2014 66 197ndash203 [CrossRef]
58 Chiu S-Y Kao C-Y Tsai M-T Ong S-C Chen C-H Lin C-S Lipid accumulation and CO2 utilizationof Nannochloropsis oculata in response to CO2 aeration Bioresour Technol 2009 100 833ndash838 [CrossRef][PubMed]
59 Huerlimann R de Nys R Heimann K Growth lipid content productivity and fatty acid composition oftropical microalgae for scale-up production Biotechnol Bioeng 2010 107 245ndash257 [CrossRef] [PubMed]
60 Liang Y Sarkany N Cui Y Biomass and lipid productivities of Chlorella vulgaris under autotrophicheterotrophic and mixotrophic growth conditions Biotechnol Lett 2009 31 1043ndash1049 [CrossRef] [PubMed]
61 Hempel N Petrick I Behrendt F Biomass productivity and productivity of fatty acids and amino acidsof microalgae strains as key characteristics of suitability for biodiesel production J Appl Phycol 2012 241407ndash1418 [CrossRef] [PubMed]
62 Yeesang C Cheirsilp B Effect of nitrogen salt and iron content in the growth medium and light intensityon lipid production by microalgae isolated from freshwater sources in Thailand Bioresour Technol 2011 1023034ndash3040 [CrossRef] [PubMed]
63 Msanne J Xu D Konda AR Casas-Mollano JA Awada T Cahoon EB Cerutti H Metabolic andgene expression changes triggered by nitrogen deprivation in the photoautotrophically grown microalgaeChlamydomonas reinhardtii and Coccomyxa sp C-169 Phytochemistry 2012 75 50ndash59 [CrossRef] [PubMed]
64 Lin Q Lin J Effects of nitrogen source and concentration on biomass and oil production of aScenedesmus rubescens like microalga Bioresour Technol 2011 102 1615ndash1621 [CrossRef] [PubMed]
65 Sheehan J Dunahay T Benemann J Roessler P A Look Back at the US Department of Energyrsquos AquaticSpecies ProgrammdashBiodiesel from Algae US Department of Energyrsquos Office of Fuels Development Golden COUSA 1998 p 328
66 Vaulot D Olson RJ Merkel S Chisholm SW Cell-cycle response to nutrient starvation in twophytoplankton species Thalassiosira weissflogii and Hymenomonas carterae Mar Biol 1987 95 625ndash630[CrossRef]
67 Olson RJ Chisholm SW Effects of light and nitrogen limitation on the cell cycle of the dinoflagellateAmphidinium carteri J Plankton Res 1986 8 785ndash793 [CrossRef]
68 Li Y Fei X Deng X Novel molecular insights into nitrogen starvation-induced triacylglycerolsaccumulation revealed by differential gene expression analysis in green algae Micractinium pusillumBiomass Bioenergy 2012 42 199ndash211 [CrossRef]
69 Abdelaziz AEM Leite GB Belhaj MA Hallenbeck PC Screening microalgae native to Quebec forwastewater treatment and biodiesel production Bioresour Technol 2014 157 140ndash148 [CrossRef] [PubMed]
70 Valdez-Ojeda R Gonzaacutelez-Muntildeoz M Us-Vaacutezquez R Narvaacuteez-Zapata J Chavarria-Hernandez JCLoacutepez-Adriaacuten S Barahona-Peacuterez F Toledano-Thompson T Garduntildeo-Soloacuterzano G Escobedo-GraciaMedrano RM Characterization of five fresh water microalgae with potential for biodiesel productionAlgal Res 2015 7 33ndash44 [CrossRef]
71 Shrivastav A Mishra SK Suh WI Farooq W Moon M Kim T-H Kumar K Choi G-G Park MSYang J-W Characterization of newly isolated oleaginous microalga Monoraphidium sp for lipid productionunder different conditions Algal Res 2015 12 289ndash294 [CrossRef]
72 Yen H-W Hu I-C Chen C-Y Ho S-H Lee D-J Chang J-S Microalgae-based biorefinerymdashFrombiofuels to natural products Bioresour Technol 2013 135 166ndash174 [CrossRef] [PubMed]
Energies 2017 10 224 16 of 16
73 Li J Liu Y Cheng JJ Mos M Daroch M Biological potential of microalgae in China for biorefinery-basedproduction of biofuels and high value compounds New Biotechnol 2015 32 588ndash596 [CrossRef] [PubMed]
74 El-Sheekh M Abomohra AE-F Hanelt D Optimization of biomass and fatty acid productivity ofScenedesmus obliquus as a promising microalga for biodiesel production World J Microbiol Biotechnol 201329 915ndash922 [CrossRef] [PubMed]
75 Singh P Guldhe A Kumari S Rawat I Bux F Investigation of combined effect of nitrogen phosphorusand iron on lipid productivity of microalgae Ankistrodesmus falcatus KJ671624 using response surfacemethodology Biochem Eng J 2015 94 22ndash29 [CrossRef]
76 Kilham S Kreeger D Goulden C Lynn S Effects of nutrient limitation on biochemical constituents ofAnkistrodesmus falcatus Freshw Biol 1997 38 591ndash596 [CrossRef]
77 Breuer G Lamers PP Martens DE Draaisma RB Wijffels RH The impact of nitrogen starvation on thedynamics of triacylglycerol accumulation in nine microalgae strains Bioresour Technol 2012 124 217ndash226[CrossRef] [PubMed]
78 Breuer G de Jaeger L Artus VG Martens DE Springer J Draaisma RB Eggink G Wijffels RHLamers PP Superior triacylglycerol (TAG) accumulation in starchless mutants of Scenedesmus obliquus (II)evaluation of TAG yield and productivity in controlled photobioreactors Biotechnol Biofuels 2014 7 70[CrossRef] [PubMed]
79 Nigam S Prakash-Rai M Sharma R Effect of Nitrogen on Growth and Lipid Content ofChlorella pyrenoidosa Am J Biochem Biotechnol 2011 7 126ndash131 [CrossRef]
80 Piligaev AV Sorokina KN Bryanskaya AV Peltek SE Kolchanov NA Parmon VN Isolation ofprospective microalgal strains with high saturated fatty acid content for biofuel production Algal Res 201512 368ndash376 [CrossRef]
81 Weiss SB Kennedy EP The enzymatic synthesis of triglycerides J Am Chem Soc 1956 78 3550[CrossRef]
82 Miller R Wu G Deshpande RR Vieler A Gaumlrtner K Li X Moellering ER Zaumluner S Cornish AJLiu B et al Changes in Transcript Abundance in Chlamydomonas reinhardtii following Nitrogen DeprivationPredict Diversion of Metabolism Plant Physiol 2010 154 1737ndash1752 [CrossRef] [PubMed]
83 Rismani-Yazdi H Haznedaroglu BZ Bibby K Peccia J Transcriptome sequencing and annotation of themicroalgae Dunaliella tertiolecta Pathway description and gene discovery for production of next-generationbiofuels BMC Genom 2011 12 148 [CrossRef] [PubMed]
84 Ramos MJ Fernaacutendez CM Casas A Rodriacuteguez L Peacuterez A Influence of fatty acid composition of rawmaterials on biodiesel properties Bioresour Technol 2009 100 261ndash268 [CrossRef] [PubMed]
85 Knothe G Dependence of biodiesel fuel properties on the structure of fatty acid alkyl estersFuel Process Technol 2005 86 1059ndash1070 [CrossRef]
86 Knothe G ldquoDesignerrdquo Biodiesel Optimizing Fatty Ester Composition to Improve Fuel PropertiesEnergy Fuels 2008 22 1358ndash1364 [CrossRef]
87 Smith PC Ngothai Y Dzuy Nguyen Q OrsquoNeill BK Improving the low-temperature properties ofbiodiesel Methods and consequences Renew Energy 2010 35 1145ndash1151 [CrossRef]
88 Dunn RO Effect of antioxidants on the oxidative stability of methyl soyate (biodiesel) Fuel Process Technol2005 86 1071ndash1085 [CrossRef]
89 Knothe G Improving biodiesel fuel properties by modifying fatty ester composition Energy Environ Sci2009 2 759ndash766 [CrossRef]
copy 2017 by the authors licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)

Energies 2017 10 224 3 of 16
centrifugation at 1900times g for 10 min and washed two times with 1 mL of physiological saline solutionMicroalgal cell pellets were dissolved in Nile Red acetone solution (2 microgmL) and incubated for15 min at 37 C After washing once stained microalgal cells were observed by Carl Zeiss fluorescentmicroscopy and photographed with a digital camera AxioLabA1 AxioCamERc real time 5 s Imageswere obtained at a magnification of 1000times with visible light and epifluorescence (excitation 510ndash560emission 590)
23 Genomic DNA Isolation
Microalgae cells were harvested from 100 mL of liquid culture by centrifugation at 1900times g for10 min Genomic DNA was extracted using a modified version of the CTAB method as describedby Doyle and Doyle [18] Briefly algal cells were completely ground in a mortar containing 50 mgof sterilized sand and 3 mL of extraction buffer (300 mM Tris-HCl pH 80 50 mM ethylenediaminetetraacetic acid (EDTA) 2 M NaCl 2 cetyltrimethylammonium bromide 3 polyvinylpyrrolidone(MW 40000) and 2 β-mercaptoethanol) Homogenized cells were incubated at 70 C for 30 min withgentle inversion every 2 min An equal volume of phenolchloroformisoamyl alcohol (25241 vv)was then added followed by centrifugation at 20000times g The aqueous supernatant was transferred toa new microtube and an equal volume of chloroformisoamyl alcohol (241 vv) was added followedby centrifugation at 20000times g The aqueous supernatant was transferred to a new microtube andmixed with 01 volumes of 3 M sodium acetate (pH 52) and an equal volume of chilled isopropanolwas added to precipitate the DNA at 15000times g Washing was done with 70 alcohol and the air-driedDNA pellet was dissolved in 100 microL of sterilized water treated with RNase A at 40 C for 30 min andthen extracted with chloroformisoamyl alcohol and DNA precipitation with absolute ethanol Finallythe air-dried DNA pellet was dissolved in TE buffer (10 mM Tris-HCl pH 80 1 mM EDTA) and storedat minus20 C Genomic DNA quality and quantity were assessed by standard OD measurement [19]using a Nanodrop 2000 UV-Vis Spectrophotometer (Thermo Fisher Scientific Wilmington DE USA)In order to verify DNA integrity samples were resolved by standard gel electrophoresis in 12agarose gels [19] stained with ethidium bromide and visualized under ultraviolet light (SupplementaryMaterials Figure S1)
24 PCR Amplification and Sequencing of ITS2-rDNA Region
The ITS-2 rDNA gene was amplified according to Kaur et al [20] with minor modifications in20-microL reactions containing 1times PCR buffer 3 mM MgCl2 02 mM dNTPs mix 0025 UmicroL AmplitaqGold 10 microM each of the forward primer SQITS1 (5prime-GAGCATGTCTGCCTCAGC-3prime) and reverseprimer SQITS2 (5prime-GGTAGCCTTGCCTGAGC-3prime) and 20 ng of genomic DNA The general conditionsfor PCR in a thermal cycler Realplex S4 (Eppendorf Hamburg Germany) were 5 min at 94 C for thefirst cycle followed by 30 s at 94 C 30 s at 55 C and 1 min at 72 C for 35 cycles and a final 10-minextension step at 72 C The PCR gene products were separated on 2 agarose gels using 05 times TBEbuffer at 85 volts for 60 min and detected by staining with ethidium bromide (Figure S2) To purify4 microL of the PCR gene product were mixed with 2 microL of ExoSAP-IT (Affymetrix Santa Clara CAUSA) incubated at 37 C for 30 min and 80 C for 15 min and directly sequenced in both directionswith primers SQITS1 and SQITS2 following the instructions of the BigDyereg Terminator v31 CycleSequencing Kit (Applied Biosystemstrade Foster City CA USA) The sequencing products were purifiedby precipitation with ethanolEDTA denatured and resolved on a Genetic Analyzer 3130XL (AppliedBiosystemstrade Foster City CA USA) All sequences presented here were submitted to GenBank [21]
25 Phylogenetic Analysis
The ITS-2 rDNA gene sequences of the isolates were searched for homology in the National Centerfor Biotechnology Information database [22] using BLAST [23] Sequences were edited in BioEdit 725software [24] to trim the non-overlapping 5prime and 3prime regions for the purpose of restricting the multiplesequence alignment within the minimum common region Those edited sequences were aligned using
Energies 2017 10 224 4 of 16
Clustal omega [25] The unrooted phylogenetic tree was constructed using the neighbor-joining (NJ)method [26]] and Kimurarsquos two-parameter algorithm [27] as implemented within the MEGA 606software [28] The bootstrap analysis [29] was used with 1000 replications to test the relative supportfor the branches produced by NJ analysis
26 Microalgae Cultivation Biomass and Total Lipids Measurements
Two hundred milliliters Chu medium No 10 in a 500-mL Erlenmeyer flask were inoculatedwith the microalgae cells (OD680 005) and incubated at 25 plusmn 1 C using 80 microEmiddotmminus2middotsminus1 intensity ofcool-white fluorescent light and continuous agitation at 150 rpm for four weeks In this culture stage allmicroalgae strains were found in the stationary growth phase and centrifuged at 1920times g for 10 min toharvest the microalgae cells The cell pellets were rinsed with 50 mL deionized water centrifuged againdried in an oven at 70 C to a constant weight and the dry weight was determined gravimetricallywith an analytical balance Kern ABJ 220-4NM (Kern amp Sohn GmbH Balingen Germany) The algalpowders were stored at minus20 C for total lipid analysis
Total lipid was extracted using a modified protocol of Bligh and Dyer [30] For each sampleapproximately 50 mg of microalgal powder were mixed with 3 mL of chloroformmethanol (21 vv)and homogenized in an ultrasonic Branson 2510 (Branson Danbury CT USA) for 5 min The mixturewas then centrifuged at 20000times g for 5 min and the supernatant was transferred into a pre-weighedvial Cellular debris was mixed with another 2 mL of chloroformmethanol (21 vv) by vortex andthen centrifuged as described above After that the pooled organic phases were filtered throughsyringe filters of 045 microm evaporated with nitrogen gas and dried at 80 C in a vacuum ovenVacucell 55mdashComfort (MMM Muumlnchen Germany) to a constant weight The lipid content andproductivity of each sample were measured gravimetrically using a semi-micro analytical balance(Sartorius MSU225S-000-DU Foster City CA USA) and calculated as follows [31]
Total lipid ( dry weight) = WLWA times 100
where WL (g) is the weight of the total lipids extracted and WA (g) is the weight of the drymicroalgae biomass
PLipid (mgmiddotLminus1middotdminus1) = WA (g) times CLipid ()V(L) times T(d)
where PLipid is the lipid productivity CLipid is the lipid content V is the working volume and T is thecultivation time
27 Strains Culture under Nitrogen-Sufficient and Nitrogen-Deficient Medium
The selected microalgae species Acutodesmus obliquus (KP8785051) Ankistrodesmus sp(KP8785001) and Chlorella lewinii (KP8785011) (Figure S3) were grown in 500-mL Erlenmeyerflasks containing 250 mL of liquid Chu medium No 10 with nitrogen sufficient (5756 microgmL ofCa(NO3)2middot4H2O) or liquid Chu medium No 10 without a nitrogen source (deficient) for seven days ina controlled culture room at 25 plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity ofcool-white fluorescent light and continuous agitation at 150 rpm
For growth profile analysis the maximum absorption peak for each microalgae strain wasdetermined in the 600ndash800 nm range by spectrophotometric analysis using a Nanodrop 2000 UV-VisSpectrophotometer (Figure S4) Then aliquots of the microalgal cultures were evaluated every 24 h(between 8 and 9 h) at 680 nm The specific growth rate (micro) was calculated during the exponentialgrowth phase (the first three days of culture) according to the equation micro = (ln Nf minus ln Ni)(tf minus ti)where N is the optical density (OD680) at final (f) or initial (i) time (t) Previously it was determinedthat a high (R2 from 0992 to 0998) and positive correlation exists between optical density and celldensities this variable was calculated using a hemocytometer The experiments were carried out intriplicate and the data were expressed as the mean plusmn standard deviation (plusmnSD)
Energies 2017 10 224 5 of 16
28 Determination of Total Protein Total Carbohydrate Ash Content Esterification and Fatty Acids Profiling
The microalgae cultures maintained for seven days under +N and minusN Chu medium No 10were harvested by centrifugation rinsed and dried as previously described (Section 25) Fromthe microalgae powders total protein content was determined according to the HartreendashLowrymethod [32] To 5 mg of dry microalgal biomass was added 5 mL of 05 M NaOH and incubated for10 min at 80 C then centrifuged for 5 min at 14000times g Protein content in 1 mL of the supernatantwas determined by adding 05 mL of 05 M sodium carbonate 05 mL of 05 M sodium potassiumtartrate 05 M of copper sulfate and 2 mL of Folin-Ciocalteu reagent The blue complex was analyzedin a Varian Cary 50 Bio UV-Visible Spectrophotometer (Agilent Technologies Santa Clara CA USA)set at 650 nm against a calibration curve based on a known concentration of BSA as the standard
Total carbohydrate content of the microalgal biomass was determined with the phenol-sulfuricacid method [3233] For this purpose 5 mg of dry microalgae biomass were added to 5 mL of waterAfter this a 1-mL aliquot of the sample was added to 3 mL of sulfuric acid and 1 mL of 5 aqueoussolution of phenol and the mixture was stirred and incubated for 5 min at 90 C The yellow-browncomplex was analyzed by spectrophotometric analysis at 488 nm against a calibration curve based ona known concentration of glucose
Ash content was done according to the AOAC procedure [34] One hundred milligrams of drymaterial were ashed in a muffle furnace Thermolynetrade F6010 (Thermo Fisher Scientific WalthmanMA USA) at 550 C for 16 h and the content was determined gravimetrically All analyses werecarried out in triplicate and the data are expressed as the mean plusmn SD
Fatty acids methyl esters (FAME) were prepared by acid transesterification according to Ichiharaand Fukubayashi [35] The crude lipid extract (1 mg) was dissolved in 02 mL of toluene 15 mL ofmethanol and 03 mL of 08 HCl (prepared in methanolwater 8515 vv) transferred into a cappedtest tube mixed for 5 min and then incubated at 45 C for 12 h for derivatization reaction Then 2 mLof n-hexane and 2 mL of distilled water were added to the tube After vortexing the hexane phasescontaining FAMEs were transferred into a 15-mL tube and dried under a stream of nitrogen Finallythe FAMEs were redissolved in 10 microL acetonitrile and analyzed by using a gas chromatograph VarianCP-3800 GC (Agilent Technologies Santa Clara CA USA) equipped with an automated sampler andinjector a flame ionization detector and using a 30 m times 032 mm times 025 microm Stabilwaxreg capillarycolumn (Restek Bellefonte PA USA) The GC conditions were as follows injector temperature 250 Cthe column temperature gradient was 120 C for 1 min followed by an increase to 160 C at the rateof 30 Cmin 160 C for 1 min followed by an increase to 240 C at the rate of 4 Cmin and 240 Cfor 7 min detector temperature 260 C Gas pressures for He H2 and synthetic air were maintained at40 80 and 60 psi respectively The carrier gas (He) flow was maintained at 1 mLmin The fatty acidswere analyzed by comparing their retention time of the corresponding peaks with a known standardmixture of FAMEs (Nu-Check Prep Elysian MN USA) and methyl tricosanoate (Sigma-AldrichSaint Louis MO USA) was added to each sample as the internal standard All chromatograms ofthe microalgal samples were analyzed by using the Galaxietrade Chromatography Data System Version1932 (Agilent Technologies Santa Clara CA USA) All peaks spanning a peak area of more than50 units were integrated All analyses were carried out in triplicate and the data are expressed as themean plusmn SD
29 Statistical Analysis
Statistical analyses were performed using the IBM SPSS Statistics 210 software (IBM ArmonkNY USA) The growth rate biomass and lipid content data for microalgal strains were analyzedusing a one-way analysis of variance (ANOVA) and the HSD Tukey test The correlation of biomassproductivity and lipid content was analyzed using bivariate correlations and Pearsonrsquos correlationcoefficients are given with their significance levels Differences were considered significant at p lt 005All data presented here are given as the mean plusmn SD
Energies 2017 10 224 6 of 16
3 Results and Discussion
31 Isolation and Identification of the Microalgae Strains
Bioprospecting of tropical ecosystems can provide exceptional native microalgae strains withthe potential for the production of biodiesel nutraceuticals bioactive chemicals and other highvalue products [36ndash38] Furthermore native microalgae strains can have potential applications forefficient biological sequestration of CO2 wastewater treatment and other environmentally-friendlyapplications [38ndash40] Consequently isolation is a fundamental process to obtain pure cultures and isthe first phase towards the screening and selection of microalgae strains with the potential for biodieselproduction and the aforementioned applications
According to these demands in this research samples were collected from three different typesof water bodies the Amazon River (white-water type) the Itaya and Nanay Rivers both of theblack-water type The white-water rivers originate from the Andean catchments bringing high amountsof nutrient-rich sediments In contrast the black-water rivers drain geochemically-poor watershedsand are characterized by their high content in humic acids low pH and poor nutrient status [41]A total of 14 microalgal strains were isolated using the standard cell washing technique and basedon distinguishable morphological characters under light microscopic examination these strains werepreliminary ascribed to the genera Acutodesmus (Hegewald) Tsarenko 2001 Ankistrodesmus Corda1838 Chlorella Beyerinck Beijerinck 1890 Desmodesmus (Chodat) SS An T Friedl and E Hegewald1999 and Tetradesmus GM Smith 1913 (Table 1) These genera are green algae that belong todivision Chlorophyta class Chlorophyceae and orders Chlorellales and Sphaeropleales Althoughmorphological analysis is frequently used to identify microalgae it is inaccurate and very difficultfor the identification at the species level because the relationship between diagnostic morphologyand biological species boundaries are largely unknown in many micro-eukaryotes [42] Furthermoreaccording to Yu et al [31] the microalgae morphology for the same strain varies in relation to ageand culture conditions Consequently in microalgae the classic taxonomy is supported by moleculartools [42] For example the ITS2 rDNA region is thought to be an excellent marker for molecularphylogenetics studies in lower taxonomic levels due to being highly divergent and fast-evolvingwhich can discriminate among closely-related organisms which otherwise display almost identicalsequences [4344] For this reason the molecular identification based in phylogenetic analysis of ITS2rDNA gene sequences is commonly used to identify andor delineate species [204546]
Table 1 Genera GenBank accession number of ITS2 sequences biomass productivity lipid productivityand lipid content of isolated microalgae strains
Source ofCollection Strain GeneraSpecies
GenBankAccessionNumber
BiomassProductivity
(mgmiddotLminus1middotdminus1)
LipidProductivity
(mgmiddotLminus1middotdminus1)
LipidContent( dw)
Amazon River(0341prime06prime prime S7314prime89prime prime W)
AMA001 Ankistrodesmus sp KP8784961 122 plusmn 03 37 plusmn 01 307 plusmn 15AMA002 Ankistrodesmus sp KP8784971 36 plusmn 04 10 plusmn 02 277 plusmn 21AMA003 Chlorella lewinii KP8785011 223 plusmn 07 51 plusmn 05 227 plusmn 15AMA004 Acutodesmus abliquus KP8785051 316 plusmn 04 64 plusmn 07 203 plusmn 21
Itaya River(0343prime14prime prime S
7314prime178prime prime W)
ITA001 Ankistrodesmus sp KP8784991 59 plusmn 02 40 plusmn 03 437 plusmn 17ITA002 Ankistrodesmus sp KP8785001 136 plusmn 04 57 plusmn 05 306 plusmn 20ITA003 Chlorella lewinii KP8785031 129 plusmn 03 40 plusmn 05 224 plusmn 27ITA004 Chlorella lewinii KP8785041 221 plusmn 07 31 plusmn 04 215 plusmn 23ITA005 Scenedesmus regularis KP8785081 93 plusmn 04 56 plusmn 04 242 plusmn 14ITA006 Scenedesmus dimorphus KP8785091 186 plusmn 06 32 plusmn 03 260 plusmn 17ITA007 Acutodesmus abliquus KP8785101 179 plusmn 04 22 plusmn 03 156 plusmn 19
Nanay River(0342prime02prime prime S7315prime32prime prime W)
NAN001 Ankistrodesmus sp KP8784981 143 plusmn 06 28 plusmn 02 467 plusmn 15NAN002 Desmodesmus tropicus KP8785061 122 plusmn 04 23 plusmn 02 175 plusmn 11NAN003 Tetradesmus sp KP8785071 139 plusmn 04 30 plusmn 03 135 plusmn 12
Energies 2017 10 224 7 of 16
To realize phylogenetic analysis genomic DNA of each isolate strain was purified and used forPCR amplification of ITS2 rDNA The universal primers used to amplify the ITS2 rDNA successfullyamplified the desired DNA fragments (~300 bp) from all microalgal strains (Figure S2) Furtherthese amplicons were sequenced published in the NCBI databases and the corresponding GenBankaccession numbers are presented in Table 1 Figure 1 shows a neighbor-joining phylogenetic treedepicting the relatedness of the native microalgae strain isolates with selected sequences from thepublic GenBank database This phylogram contained three clusters each of which included differentmicroalgae genera For example the first cluster consisted of microalgae of the genera AcutodesmusScenedesmus Tetradesmus and Desmodesmus and six isolated strains (ITA007 AMA004 ITA006 NAN003ITA005 and NAN002) The second cluster consisted of a microalgae of the genus Ankistrodesmus andthe isolated strains AMA001 ITA001 ITA002 NAN001 and AMA002 The third one consisted of amicroalgae of the genus Chlorella and three isolated strains (ITA004 AMA003 and ITA003) Overall theidentification of native microalgae strains presented a good correspondence between morphologicalcharacters and sequence-based phylogenetic analysis Microalgae species of these genera were alsoisolated from different countries of the world such as Brazil [47] Argentina [46] United States [48]India [20] Egypt [49] Taiwan [50] South Korea [45] and China [51] Some of these microalgae generasuch as Ankistrodesmus Chlorella Desmodesmus and Scenedesmus were used in large-scale biodieselproduction using photobioreactors and raceway pond systems [5253]
Energies 2017 10 224 7 of 16
public GenBank database This phylogram contained three clusters each of which included different microalgae genera For example the first cluster consisted of microalgae of the genera Acutodesmus Scenedesmus Tetradesmus and Desmodesmus and six isolated strains (ITA007 AMA004 ITA006 NAN003 ITA005 and NAN002) The second cluster consisted of a microalgae of the genus Ankistrodesmus and the isolated strains AMA001 ITA001 ITA002 NAN001 and AMA002 The third one consisted of a microalgae of the genus Chlorella and three isolated strains (ITA004 AMA003 and ITA003) Overall the identification of native microalgae strains presented a good correspondence between morphological characters and sequence-based phylogenetic analysis Microalgae species of these genera were also isolated from different countries of the world such as Brazil [47] Argentina [46] United States [48] India [20] Egypt [49] Taiwan [50] South Korea [45] and China [51] Some of these microalgae genera such as Ankistrodesmus Chlorella Desmodesmus and Scenedesmus were used in large-scale biodiesel production using photobioreactors and raceway pond systems [5253]
Figure 1 Unrooted phylogenetic tree of isolated microalgae strains using the ITS2 sequences The tree was constructed by the neighbor-joining method and Kimurarsquos two-parameter algorithm in MEGA 60 software based on the multiple sequence alignment by Clustal Omega Bootstrap values of 1000 replicates () are shown at the branches
32 Microalgae Cultivation Biomass and Total Lipids Measurements
The isolated microalgae strains exhibited remarkable differences in productivity parameters and lipid content among individual strains in the stationary growth phase (Table 1) With regard to the biomass productivity the 14 strains showed statistically-significant differences (F = 71911 df = 13 p lt 0001) and ranged from 36 plusmn 04 mgmiddotLminus1middotdminus1 (Ankistrodesmus sp strain AMA002) to 316 plusmn 04 mgmiddotLminus1middotdminus1 (Acutodesmus obliquus strain AMA004) Similar biomass productivities were reported for other marine microalgae species [54] However other investigations show major biomass productivities (ranging
Figure 1 Unrooted phylogenetic tree of isolated microalgae strains using the ITS2 sequences The treewas constructed by the neighbor-joining method and Kimurarsquos two-parameter algorithm in MEGA60 software based on the multiple sequence alignment by Clustal Omega Bootstrap values of1000 replicates () are shown at the branches
Energies 2017 10 224 8 of 16
32 Microalgae Cultivation Biomass and Total Lipids Measurements
The isolated microalgae strains exhibited remarkable differences in productivity parametersand lipid content among individual strains in the stationary growth phase (Table 1) With regardto the biomass productivity the 14 strains showed statistically-significant differences (F = 71911df = 13 p lt 0001) and ranged from 36 plusmn 04 mgmiddotLminus1middotdminus1 (Ankistrodesmus sp strain AMA002) to316 plusmn 04 mgmiddotLminus1middotdminus1 (Acutodesmus obliquus strain AMA004) Similar biomass productivities werereported for other marine microalgae species [54] However other investigations show major biomassproductivities (ranging from 30 to 120 mgmiddotLminus1middotdminus1) in the same and other microalgae genera [455556]These discrepancies may be attributed to differences in the culture conditions in the culture mediacomposition and in the microalgae genotypes Lipid productivity also exhibited statistically-significantdifferences (F = 4774 df = 13 p lt 0001) and ranged from 10plusmn 04 mgmiddotLminus1middotdminus1 to 64 plusmn 07 mgmiddotLminus1middotdminus1
with strains AMA002 and AMA004 respectively These lipid productivities were low in comparison toother reports which reported values from 190 to 433 mgmiddotLminus1middotdminus1 [4555] Furthermore lipid contentas a percentage of the microalgal biomass dry weight showed statistically-significant differences(F = 8509 df = 13 p lt 0001) and ranged from 135 plusmn 12 (Tetradesmus sp strain NAN003) to467 plusmn 15 (Ankistrodesmus sp strain NAN001) These percentage values in total lipid contentare similar to several microalgae species of these genera isolated and characterized by otherresearchers worldwide [20454648ndash5057] Furthermore it is important to emphasize that lipid contentis dependent on the different growth phases Commonly in several microalgae taxa the lipid contentincreases gradually from logarithmic to stationary growth phases [58ndash60] For example Kaur et al [20]reported that the total lipid content of Scenedesmus sp DRLMA9 demonstrated values of 207 inthe late exponential phase and 397 in the stationary phase and Desmodesmus elegans DRLMA13displayed values of 103 in the late exponential phase and 169 in the stationary phase LikewiseChiu et al [61] found that the total lipid content of Nannochloropsis oculata was 308 in the logarithmicphase 397 in the early stationary phase and 504 in the stationary phase These changes in lipidcontent with culture age have been associated with nitrate depletion [61] which causes a channeling ofphotosynthetic ATP to carbohydrate and lipid biosynthesis [60] Finally Pearson correlation analysisof biomass productivity and lipid content was inversely related (r = minus037 p lt 001) which is similarto the results obtained by Liang et al [62] and Rodolfi et al [52] In contrast Li et al [51] andHempel et al [63] found no relationship between these parameters The negative correlation obtainedis likely due to the high metabolic cost of lipid biosynthesis [62]
33 Strains Culture under Nitrogen-Sufficient and Nitrogen-Deficient Medium
The microalgae species Acutodesmus obliquus (KP8785051) Ankistrodesmus sp (KP8785001)and Chlorella lewinii (KP8785011) were selected and further characterized because they displayedthe highest biomass and lipid productivities (Table 1) These microalgae showed noticeablevariations in the growth profile when cultured in Chu medium No 10 with nitrogen (+N) orwithout a nitrogen source (minusN) In +N medium all strains showed exponential growth butChlorella lewinii showed the highest average specific growth rates (micro = 0458 plusmn 0098 dminus1) followed byAcutodesmus obliquus (micro = 0399 plusmn 0112 dminus1) and the lowest value was produced by Ankistrodesmus sp(micro = 0265 plusmn 0013 dminus1) In contrast all cultures displayed a lag phase pattern in their growthprofile under minusN medium (Figure 2) These findings corroborate the results obtained by otherresearchers who demonstrate that the specific growth rate is directly proportional to the initialnitrate concentration [46596465] because the reduction or cessation of microalgae cell divisionis due to nitrogen limitation [66] which is a key nutrient for protein and nucleic acids de novobiosynthesis consequently causing an arrest in the early part of the microalgae cell cycle [6768]In accordance with these reports Li et al [69] showed that nitrogen deficiency induced in the greenmicroalgae Micractinium pusillum a downregulation in the expression of genes involved in cell growthand cell cycle regulation Additionally differences in growth profile and specific growth rates aredependent on several factors such as microalgae genotypes the culture medium composition pH
Energies 2017 10 224 9 of 16
nutrients concentration the cell culture phase (ie log phase stationary phase) and environmentalfactors [4549545770ndash72]Energies 2017 10 224 9 of 16
Figure 2 Growth profiles of three microalgal strains cultivated under +N (filled boxes) and minusN (empty boxes) Chu medium No 10 The culture was incubated in a controlled culture room at 25 plusmn 1 degC with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 150 rpm Experiments were carried out in triplicate
34 Total Protein Total Lipid and Total Carbohydrate Content
Biochemical parameters of the microalgae species such as total protein total lipid and total carbohydrate content exhibited differences and changes under both culture conditions (Figure 3) For example Chlorella lewinii showed the highest protein content Ankistrodesmus sp had the highest total lipid content but Acutodesmus obliquus presented the highest levels of total carbohydrate Therefore these strains can be valuable stocks for the industry because total protein and total carbohydrate are potentially byproducts of microalgal biomass processing [7374] Additionally the three strains cultured under minusN conditions showed a significant increase in total lipid content (from 237 in Acutodesmus obliquus to 667 in Ankistrodesmus sp) and total carbohydrate content (from 119 in Acutodesmus obliquus to 963 in Chlorella lewinii) but showed a decreased in total protein levels (from 242 in Acutodesmus obliquus to 545 in Chlorella lewinii) in comparison to +N cultures (Figure 3) These results are in accordance with well-documented results obtained by other researchers with several microalgae species of the genera Acutodesmus Ankistrodesmus Botryococcus Chlamydomonas Chlorella Desmodesmus Dunaliella Haematococcus Neochloris Phaeodactylum Pseudokirchneriella and Scenedesmus [465058596475ndash80] Nevertheless the accumulation of total lipid induced by nitrogen deficiency is not correlated with lipid productivity but rather is due to a decline and stopping of the microalgae specific growth rate [75] The changes in biomolecule contents suggest redirection of microalgae metabolism during nitrogen starvation such as a decrease in biosynthesisincrease in protein degradation and an increase in the biosynthesis of storage biomolecules such as triacylglycerols and starch The increase in total lipid content could be partially explained for a boost in transcript levels of genes encoding enzymes of the lipid biosynthesis pathways According to this assumption recent investigations showed that genes encoding the enzymes that catalyze the last step in the Kennedy pathway of triacylglycerol biosynthesis [81] such as glycerol-3-phosphate acyltransferase and acyl-glycerol-3-phosphate acyltransferase are upregulated in response to nitrogen starvation [648283] Furthermore the increase in total carbohydrate content can be explained by changes in the expression levels of genes involved in carbohydrate metabolism For example when the oleaginous microalgae Neochloris oleoabundans is cultured under nitrogen starvation the catabolic phase of starch specifically α-amylase which hydrolyzes starch to glucose is strongly repressed [83]
Figure 2 Growth profiles of three microalgal strains cultivated under +N (filled boxes) and minusN(empty boxes) Chu medium No 10 The culture was incubated in a controlled culture room at25 plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent lightand continuous agitation at 150 rpm Experiments were carried out in triplicate
34 Total Protein Total Lipid and Total Carbohydrate Content
Biochemical parameters of the microalgae species such as total protein total lipid and totalcarbohydrate content exhibited differences and changes under both culture conditions (Figure 3)For example Chlorella lewinii showed the highest protein content Ankistrodesmus sp had the highesttotal lipid content but Acutodesmus obliquus presented the highest levels of total carbohydrateTherefore these strains can be valuable stocks for the industry because total protein and totalcarbohydrate are potentially byproducts of microalgal biomass processing [7374] Additionallythe three strains cultured under minusN conditions showed a significant increase in total lipid content(from 237 in Acutodesmus obliquus to 667 in Ankistrodesmus sp) and total carbohydrate content(from 119 in Acutodesmus obliquus to 963 in Chlorella lewinii) but showed a decreased in totalprotein levels (from 242 in Acutodesmus obliquus to 545 in Chlorella lewinii) in comparison to +Ncultures (Figure 3) These results are in accordance with well-documented results obtained by otherresearchers with several microalgae species of the genera Acutodesmus Ankistrodesmus BotryococcusChlamydomonas Chlorella Desmodesmus Dunaliella Haematococcus Neochloris PhaeodactylumPseudokirchneriella and Scenedesmus [465058596475ndash80] Nevertheless the accumulation of totallipid induced by nitrogen deficiency is not correlated with lipid productivity but rather is due toa decline and stopping of the microalgae specific growth rate [75] The changes in biomoleculecontents suggest redirection of microalgae metabolism during nitrogen starvation such as a decreasein biosynthesisincrease in protein degradation and an increase in the biosynthesis of storagebiomolecules such as triacylglycerols and starch The increase in total lipid content could bepartially explained for a boost in transcript levels of genes encoding enzymes of the lipid biosynthesispathways According to this assumption recent investigations showed that genes encoding theenzymes that catalyze the last step in the Kennedy pathway of triacylglycerol biosynthesis [81] such asglycerol-3-phosphate acyltransferase and acyl-glycerol-3-phosphate acyltransferase are upregulated
Energies 2017 10 224 10 of 16
in response to nitrogen starvation [648283] Furthermore the increase in total carbohydrate contentcan be explained by changes in the expression levels of genes involved in carbohydrate metabolismFor example when the oleaginous microalgae Neochloris oleoabundans is cultured under nitrogenstarvation the catabolic phase of starch specifically α-amylase which hydrolyzes starch to glucoseis strongly repressed [83]Energies 2017 10 224 10 of 16
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strains cultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled culture room at 25 plusmn 1 degC with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized with respect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that each microalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majority of fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised 7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii the majority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9) acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectively Finally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic (182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in +N and minusN conditions respectively Our results agree with multiple previously-published reports in that fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent in many green microalgae species [454648ndash5055] It is notable that under nitrogen starvation culture all microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 in Ankistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid (181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findings support the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content of SFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Nevertheless there are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed a decrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increased between 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively However in the same culture condition the three microalgae species showed a decrease in polyunsaturated fatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmus spp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgae strains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFA which improves the quality properties of biodiesel such as an increase
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strainscultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled cultureroom at 25plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescentlight and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized withrespect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that eachmicroalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majorityof fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii themajority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9)acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectivelyFinally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic(182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in+N and minusN conditions respectively Our results agree with multiple previously-published reports inthat fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent inmany green microalgae species [454648ndash5055] It is notable that under nitrogen starvation cultureall microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 inAnkistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid(181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findingssupport the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content ofSFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other
Energies 2017 10 224 11 of 16
researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Neverthelessthere are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed adecrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increasedbetween 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively Howeverin the same culture condition the three microalgae species showed a decrease in polyunsaturatedfatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmusspp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgaestrains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFAwhich improves the quality properties of biodiesel such as an increase in cetane number [84ndash86]Considering the SFA and unsaturated fatty acids (UFA) components approximately 20ndash31 are SFAand 69ndash80 are UFA Hence the SFAUFA ratio in the microalgae strains evaluated ranges between025 and 045 which is similar to the reports of several microalgae species isolated in different latitudesworldwide [203148497072]
Table 2 Effect of nitrogen deficiency on fatty acid profiling in three native microalgal species from thePeruvian Amazon
Fatty AcidsAcutodesmus obliquus Ankistrodesmus sp Chlorella lewinii
+N minusN +N minusN +N minusN
140 - - 052 plusmn 002 088 plusmn 002 - 058 plusmn 001160 2724 plusmn 007 2867 plusmn 017 1868 plusmn 019 2261 plusmn 006 1964 plusmn 021 2123 plusmn 010180 184 plusmn 005 255 plusmn 051 101 plusmn 002 181 plusmn 002 371 plusmn 026 153 plusmn 015
161n-7 023 plusmn 000 025 plusmn 001 071 plusmn 003 055 plusmn 001 085 plusmn 043 038 plusmn 000181n-7 131 plusmn 002 131 plusmn 000 141 plusmn 005 094 plusmn 003 146 plusmn 006 123 plusmn 002181n-9 3213 plusmn 021 3608 plusmn 021 3699 plusmn 004 4047 plusmn 038 1371 plusmn 015 2326 plusmn 041182n-6 1732 plusmn 004 1651 plusmn 016 747 plusmn 018 656 plusmn 007 2034 plusmn 039 1957 plusmn 001183n-3 1130 plusmn 017 854 plusmn 031 1486 plusmn 004 1173 plusmn 012 1815 plusmn 022 1371 plusmn 016183n-6 101 plusmn 001 078 plusmn 002 055 plusmn 000 054 plusmn 002 124 plusmn 006 100 plusmn 020184n-3 250 plusmn 005 177 plusmn 008 631 plusmn 002 601 plusmn 005 491 plusmn 025 403 plusmn 020204n-6 - - - 084 plusmn 008 - -225n-3 - - - 062 plusmn 002 - -
Unidentified 512 plusmn 003 355 plusmn 006 1149 plusmn 004 645 plusmn 003 1599 plusmn 034 1348 plusmn 006SFA 2907 3122 2022 2531 2335 2334UFA 7093 6878 7978 7469 7665 7666
MUFA 3367 3764 3911 4195 1602 2487PUFA 3213 2759 2918 2629 4464 3831SFA +MUFA 6274 6886 5933 6726 3936 4821
It is important to highlight that the fatty acid profile determination is an essential step inthe process of the characterization of microalgae strains because it ultimately affects the qualityof the biodiesel product [8485] The quality properties of biodiesel such as high cetane numberoxidative stability and cold-flow are dependent on the hydrocarbonated chain length of saturated andunsaturated fatty acids [8587] For example biodiesel composed of a mixture of a high percentage ofSFA and MUFA is preferred for increased energy yield superior oxidative stability and higher cetanenumbers but is prone to solidify at low temperature In contrast biodiesel with high levels of PUFAhas decreased energy yield low cetane numbers and excellent cold-flow properties but insufficientoxidative stability [548588] Therefore to solve these performance problems five approaches havebeen developed such as changing the fatty acids profile by physical means genetic modification ofthe feedstock or the use of alternative feedstocks with different fatty acids profiles [89]
4 Conclusions
Fourteen microalgae were isolated from three river basins from the Peruvian Amazon and wereidentified by morphological features and molecular phylogeny approaches These strains were initially
Energies 2017 10 224 12 of 16
characterized in the stationary growth phase and exhibited significant heterogeneity in key parameterssuch as biomass productivity (36ndash316 mgmiddotLminus1middotdminus1) lipid productivity (10ndash64 mgmiddotLminus1middotdminus1) andtotal lipid content (135ndash467) Furthermore three selected strains based on elevated biomass andlipid productivities were cultured under nitrogen-sufficient and nitrogen-deficient medium to boostlipid accumulation Under these culture conditions all strains showed noticeable decreases in totalprotein content and increases in total lipid and total carbohydrate contents In addition these strainspresented modifications in the fatty acids profiles with improvement in biodiesel properties due to apredominance of SFA and MUFA which corresponds to an increase in cetane number The results ofthis research suggest that the isolated native microalgae from the Peruvian Amazon could be suitablerenewable feedstocks for the production of both biodiesel and bioethanol which could contribute tosustainable development at the local regional and national levels as a viable alternative to petroleumexploitation However it will be necessary to develop open closed or hybrid systems for large-scalebiomass production of selected native microalgae strains andor de novo microalgae strains designedto produce biofuels and with the abilities to synthesize useful bioactive compounds to be efficientlyexploited using biorefinery approaches
Supplementary MaterialsThe following are available online at wwwmdpicom1996-1073102224s1 Figure S1 Results of
electrophoretic and spectrophotometric analysis of the genomic DNA isolated from the fourteen microalgaeFigure S2 ITS2-rDNA structure (A) and amplicons obtained (B) from the microalgae isolated Figure S3Microphotographic images of the three microalgae strains analyzed under light and fluorescence stained withNile Red Figure S4 Spectrophotometric scanning for the determination of the maximum absorption peak foreach microalgae strain
Acknowledgments This research was supported by the Peruvian funding agencies Consejo Nacional de CienciaTecnologiacutea e Innovacioacuten Tecnoloacutegica (CONCYTEC) with Grant Contract No 364-CONCYTEC-OAJ and Fondospara la Innovacioacuten Ciencia y Tecnologiacutea (FINCyT) with Grant Contract No 383-PNICP-PIBA-2014
Author Contributions Marianela Cobos and Juan C Castro obtained funds for the research participated inthe study design coordinated activities and participated in the preparation of the manuscript Jae D Paredesparticipated in sample collection microalgae isolation and helped to draft the manuscript J Dylan Maddoxperformed the bioinformatics analysis and participated in the preparation of the manuscript Gabriel Vargas-Aranaand Leenin Flores participated in biochemical analysis and helped to draft the manuscript Carla P Aguilarparticipated in the study design supervised biochemical analysis and helped to draft the manuscriptJorge L Marapara participated in molecular analysis and helped to draft the manuscript
Conflicts of Interest The authors declare no conflict of interest
References
1 BP Global BP Statisical Review of World Energy 2014 Available online httpwwwbpcomcontentdambppdfEnergy-economicsstatistical-review-2014BP-statistical-review-of-world-energy-2014-full-reportpdf (accessed on 20 June 2014)
2 Pachauri RK Reisinger A Cambio Climaacutetico 2007 Informe de Siacutentesis Contribucioacuten de los Grupos de Trabajo III y III al Cuarto Informe de Evaluacioacuten del Grupo Intergubernamental de Expertos Sobre el Cambio Climaacutetico IPCCGeneva Switzerland 2007 p 104
3 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Ley de Promocioacuten del Mercado deBiocombustibles Available online httpwww2congresogobpesicrcendocbibcon4_uibdnsf9E95620CC059138105257C9E005AB2F9$FILE28054pdf (accessed on 12 February 2017)
4 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Reglamento de la Ley de Promocioacuten del Mercadode Biocombustibles Available online httpwww2osinerggobpeMarcoLegaldocrevDS-013-2005-EM-CONCORDADOpdf (accessed on 12 February 2017)
5 Areacutevalo LF Nalvarte J Torres J Ramiacuterez Y Impactos Socio-Econoacutemicos de la Produccioacuten de Biocombustiblesen la Amazoniacutea Peruana Primera SNV-WWF Lima Peruacute 2009
6 Sharma KK Schuhmann H Schenk PM High Lipid Induction in Microalgae for Biodiesel ProductionEnergies 2012 5 1532ndash1553 [CrossRef]
Energies 2017 10 224 13 of 16
7 Schenk PM Thomas-Hall SR Stephens E Marx UC Mussgnug JH Posten C Kruse O Hankamer BSecond Generation Biofuels High-Efficiency Microalgae for Biodiesel Production BioEnergy Res 2008 120ndash43 [CrossRef]
8 Foley JA Asner GP Costa MH Coe MT de Fries R Gibbs HK Howard EA Olson S Patz JRamankutty N et al Amazonia revealed Forest degradation and loss of ecosystem goods and services inthe Amazon Basin Front Ecol Environ 2007 5 25ndash32 [CrossRef]
9 Duong VT Li Y Nowak E Schenk PM Microalgae Isolation and Selection for Prospective BiodieselProduction Energies 2012 5 1835ndash1849 [CrossRef]
10 Georgianna DR Mayfield SP Exploiting diversity and synthetic biology for the production of algalbiofuels Nature 2012 488 329ndash335 [CrossRef] [PubMed]
11 Wijffels RH Barbosa MJ An Outlook on Microalgal Biofuels Science 2010 329 796ndash799 [CrossRef][PubMed]
12 Richmond A Handbook of Microalgal Culture Biotechnology and Applied Phycology Blackwell Sci Ltd HudsonNJ USA 2004
13 Menetrez MY An overview of algae biofuel production and potential environmental impactEnviron Sci Technol 2012 46 7073ndash7085 [CrossRef] [PubMed]
14 Khoo HH Sharratt PN Das P Balasubramanian RK Naraharisetti PK Shaik S Life cycle energy andCO2 analysis of microalgae-to-biodiesel Preliminary results and comparisons Bioresour Technol 2011 1025800ndash5807 [CrossRef] [PubMed]
15 Chu SP The Influence of the Mineral Composition of the Medium on the Growth of Planktonic AlgaePart I Methods and Culture Media J Ecol 1942 30 284ndash325 [CrossRef]
16 Carlos EMB Mariatildengela M Gecircneros de Algas de Aacuteguas Continentais do BrasilmdashChave Para Identificaccedilatildeo eDescriccedilotildees 2nd ed Rima Sao Carlos Brasil 2006
17 Greenspan P Mayer EP Fowler SD Nile red A selective fluorescent stain for intracellular lipid dropletsJ Cell Biol 1985 100 965ndash973 [CrossRef] [PubMed]
18 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissuePhytochem Bull 1987 19 11ndash15
19 Sambrook J Fritsch EF Maniatis T Molecular Cloning A Laboratory Manual 2nd ed Cold Spring HarborLaboratory Press New York NY USA 1989
20 Kaur S Sarkar M Srivastava RB Gogoi HK Kalita MC Fatty acid profiling and molecularcharacterization of some freshwater microalgae from India with potential for biodiesel productionNew Biotechnol 2012 29 332ndash344 [CrossRef] [PubMed]
21 Benson DA Cavanaugh M Clark K Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW GenBankNucleic Acids Res 2013 41 D36ndashD42 [CrossRef] [PubMed]
22 NCBI Resource Coordinators Database Resources of the National Center for Biotechnology InformationNucleic Acids Res 2015 43 D6ndashD17
23 Ye J McGinnis S Madden TL BLAST Improvements for better sequence analysis Nucleic Acids Res 200634 W6ndashW9 [CrossRef] [PubMed]
24 BioEdit Sequence Alignment Editor for Windows 9598NTXPVista7 Available online httpwwwmbioncsuedubioeditbioedithtml (accessed on 2 December 2015)
25 Sievers F Higgins DG Clustal omega accurate alignment of very large numbers of sequencesMethods Mol Biol (Clifton NJ) 2014 1079 105ndash116
26 Saitou N Nei M The neighbor-joining method A new method for reconstructing phylogenetic treesMol Biol Evol 1987 4 406ndash425 [PubMed]
27 Kimura M A simple method for estimating evolutionary rates of base substitutions through comparativestudies of nucleotide sequences J Mol Evol 1980 16 111ndash120 [CrossRef] [PubMed]
28 Tamura K Stecher G Peterson D Filipski A Kumar S MEGA6 Molecular Evolutionary GeneticsAnalysis version 60 Mol Biol Evol 2013 30 2725ndash2729 [CrossRef] [PubMed]
29 Felsenstein J Confidence Limits on Phylogenies An Approach Using the Bootstrap Evolution 1985 39783ndash791 [CrossRef]
30 Bligh EG Dyer WJ A Rapid Method of Total Lipid Extraction and Purification Can J Biochem Physiol1959 37 911ndash917 [CrossRef] [PubMed]
Energies 2017 10 224 14 of 16
31 Yu X Zhao P He C Li J Tang X Zhou J Huang Z Isolation of a novel strain of Monoraphidium spand characterization of its potential application as biodiesel feedstock Bioresour Technol 2012 121 256ndash262[CrossRef] [PubMed]
32 Hartree EF Determination of protein A modification of the lowry method that gives a linear photometricresponse Anal Biochem 1972 48 422ndash427 [CrossRef]
33 DuBois M Gilles KA Hamilton JK Rebers PA Smith F Colorimetric Method for Determination ofSugars and Related Substances Anal Chem 1956 28 350ndash356 [CrossRef]
34 AOAC International AOAC Official Methods of Analysis of the Association of Official Analytical Chemists (AOAC)15th ed AOAC International Washington DC USA 1990 Volume 1
35 Ichihara K Fukubayashi Y Preparation of fatty acid methyl esters for gas-liquid chromatographyJ Lipid Res 2010 51 635ndash640 [CrossRef] [PubMed]
36 Brennan L Owende P Biofuels from microalgaemdashA review of technologies for production processingand extractions of biofuels and co-products Renew Sustain Energ Rev 2010 14 557ndash577 [CrossRef]
37 Raja R Hemaiswarya S Kumar NA Sridhar S Rengasamy R A perspective on the biotechnologicalpotential of microalgae Crit Rev Microbiol 2008 34 77ndash88 [CrossRef] [PubMed]
38 Mata TM Martins AA Caetano NS Microalgae for biodiesel production and other applicationsA review Renew Sustain Energy Rev 2010 14 217ndash232 [CrossRef]
39 De Wilt A Butkovskyi A Tuantet K Leal LH Fernandes TV Langenhoff A Zeeman GMicropollutant removal in an algal treatment system fed with source separated wastewater streamsJ Hazard Mater 2015 304 84ndash92 [CrossRef] [PubMed]
40 Li T Lin G Podola B Melkonian M Continuous removal of zinc from wastewater and mine dumpleachate by a microalgal biofilm PSBR J Hazard Mater 2015 297 112ndash118 [CrossRef] [PubMed]
41 Sioli H The Amazon and its main affluents Hydrography morphology of the river courses and river typesIn Monographiae Biologicae Sioli H Ed Springer Dordrecht The Netherlands 1984 pp 127ndash165
42 Moniz MBJ Kaczmarska I Barcoding of Diatoms Nuclear Encoded ITS Revisited Protist 2010 161 7ndash34[CrossRef] [PubMed]
43 Coleman AW Is there a molecular key to the level of ldquobiological speciesrdquo in eukaryotes A DNA guideMol Phylogenet Evol 2009 50 197ndash203 [CrossRef] [PubMed]
44 Mai JC Coleman AW The Internal Transcribed Spacer 2 Exhibits a Common Secondary Structure inGreen Algae and Flowering Plants J Mol Evol 1997 44 258ndash271 [CrossRef] [PubMed]
45 Abou-Shanab RAI Matter IA Kim S-N Oh Y-K Choi J Jeon B-H Characterization andidentification of lipid-producing microalgae species isolated from a freshwater lake Biomass Bioenergy2011 35 3079ndash3085 [CrossRef]
46 Do Nascimento M Ortiz-Marquez JCF Sanchez-Rizza L Echarte MM Curatti L Bioprospectingfor fast growing and biomass characterization of oleaginous microalgae from South-Eastern Buenos AiresArgentina Bioresour Technol 2012 125 283ndash290 [CrossRef] [PubMed]
47 Araujo GS Matos LJBL Gonccedilalves LRB Fernandes FAN Farias WRL Bioprospecting foroil producing microalgal strains Evaluation of oil and biomass production for ten microalgal strainsBioresour Technol 2011 102 5248ndash5250 [CrossRef] [PubMed]
48 Zhou W Li Y Min M Hu B Chen P Ruan R Local bioprospecting for high-lipid producing microalgalstrains to be grown on concentrated municipal wastewater for biofuel production Bioresour Technol 2011102 6909ndash6919 [CrossRef] [PubMed]
49 Mahmoud EA Farahat LA Abdel Aziz ZK Fatthallah NA Salah El Din RA Evaluation of thepotential for some isolated microalgae to produce biodiesel Egypt J Pet 2015 24 97ndash101 [CrossRef]
50 Ho S Lai Y-Y Chiang C-Y Chen C-NN Chang J-S Selection of elite microalgae for biodieselproduction in tropical conditions using a standardized platform Bioresour Technol 2013 147 135ndash142[CrossRef] [PubMed]
51 Li L Cui J Liu Q Ding Y Liu J Screening and phylogenetic analysis of lipid-rich microalgae Algal Res2015 11 381ndash386 [CrossRef]
52 Rodolfi L Chini Zittelli G Bassi N Padovani G Biondi N Bonini G Tredici MR Microalgae for oilStrain selection induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactorBiotechnol Bioeng 2009 102 100ndash112 [CrossRef] [PubMed]
Energies 2017 10 224 15 of 16
53 Li P Miao X Li R Zhong J In situ biodiesel production from fast-growing and high oil content Chlorellapyrenoidosa in rice straw hydrolysate J Biomed Biotechnol 2011 2011 141207 [CrossRef] [PubMed]
54 Doan TTY Sivaloganathan B Obbard JP Screening of marine microalgae for biodiesel feedstockBiomass Bioenergy 2011 35 2534ndash2544 [CrossRef]
55 Abou-Shanab RAI Hwang J-H Cho Y Min B Jeon B-H Characterization of microalgal speciesisolated from fresh water bodies as a potential source for biodiesel production Appl Energy 2011 883300ndash3306 [CrossRef]
56 Chaichalerm S Pokethitiyook P Yuan W Meetam M Sritong K Pugkaew W Kungvansaichol KKruatrachue M Damrongphol P Culture of microalgal strains isolated from natural habitats in Thailand invarious enriched media Appl Energy 2012 89 296ndash302 [CrossRef]
57 Zhang S Liu P Yang X Hao Z Zhang L Luo N Shi J Isolation and identification by 18S rDNAsequence of high lipid potential microalgal species for fuel production in Hainan Dao Biomass Bioenergy2014 66 197ndash203 [CrossRef]
58 Chiu S-Y Kao C-Y Tsai M-T Ong S-C Chen C-H Lin C-S Lipid accumulation and CO2 utilizationof Nannochloropsis oculata in response to CO2 aeration Bioresour Technol 2009 100 833ndash838 [CrossRef][PubMed]
59 Huerlimann R de Nys R Heimann K Growth lipid content productivity and fatty acid composition oftropical microalgae for scale-up production Biotechnol Bioeng 2010 107 245ndash257 [CrossRef] [PubMed]
60 Liang Y Sarkany N Cui Y Biomass and lipid productivities of Chlorella vulgaris under autotrophicheterotrophic and mixotrophic growth conditions Biotechnol Lett 2009 31 1043ndash1049 [CrossRef] [PubMed]
61 Hempel N Petrick I Behrendt F Biomass productivity and productivity of fatty acids and amino acidsof microalgae strains as key characteristics of suitability for biodiesel production J Appl Phycol 2012 241407ndash1418 [CrossRef] [PubMed]
62 Yeesang C Cheirsilp B Effect of nitrogen salt and iron content in the growth medium and light intensityon lipid production by microalgae isolated from freshwater sources in Thailand Bioresour Technol 2011 1023034ndash3040 [CrossRef] [PubMed]
63 Msanne J Xu D Konda AR Casas-Mollano JA Awada T Cahoon EB Cerutti H Metabolic andgene expression changes triggered by nitrogen deprivation in the photoautotrophically grown microalgaeChlamydomonas reinhardtii and Coccomyxa sp C-169 Phytochemistry 2012 75 50ndash59 [CrossRef] [PubMed]
64 Lin Q Lin J Effects of nitrogen source and concentration on biomass and oil production of aScenedesmus rubescens like microalga Bioresour Technol 2011 102 1615ndash1621 [CrossRef] [PubMed]
65 Sheehan J Dunahay T Benemann J Roessler P A Look Back at the US Department of Energyrsquos AquaticSpecies ProgrammdashBiodiesel from Algae US Department of Energyrsquos Office of Fuels Development Golden COUSA 1998 p 328
66 Vaulot D Olson RJ Merkel S Chisholm SW Cell-cycle response to nutrient starvation in twophytoplankton species Thalassiosira weissflogii and Hymenomonas carterae Mar Biol 1987 95 625ndash630[CrossRef]
67 Olson RJ Chisholm SW Effects of light and nitrogen limitation on the cell cycle of the dinoflagellateAmphidinium carteri J Plankton Res 1986 8 785ndash793 [CrossRef]
68 Li Y Fei X Deng X Novel molecular insights into nitrogen starvation-induced triacylglycerolsaccumulation revealed by differential gene expression analysis in green algae Micractinium pusillumBiomass Bioenergy 2012 42 199ndash211 [CrossRef]
69 Abdelaziz AEM Leite GB Belhaj MA Hallenbeck PC Screening microalgae native to Quebec forwastewater treatment and biodiesel production Bioresour Technol 2014 157 140ndash148 [CrossRef] [PubMed]
70 Valdez-Ojeda R Gonzaacutelez-Muntildeoz M Us-Vaacutezquez R Narvaacuteez-Zapata J Chavarria-Hernandez JCLoacutepez-Adriaacuten S Barahona-Peacuterez F Toledano-Thompson T Garduntildeo-Soloacuterzano G Escobedo-GraciaMedrano RM Characterization of five fresh water microalgae with potential for biodiesel productionAlgal Res 2015 7 33ndash44 [CrossRef]
71 Shrivastav A Mishra SK Suh WI Farooq W Moon M Kim T-H Kumar K Choi G-G Park MSYang J-W Characterization of newly isolated oleaginous microalga Monoraphidium sp for lipid productionunder different conditions Algal Res 2015 12 289ndash294 [CrossRef]
72 Yen H-W Hu I-C Chen C-Y Ho S-H Lee D-J Chang J-S Microalgae-based biorefinerymdashFrombiofuels to natural products Bioresour Technol 2013 135 166ndash174 [CrossRef] [PubMed]
Energies 2017 10 224 16 of 16
73 Li J Liu Y Cheng JJ Mos M Daroch M Biological potential of microalgae in China for biorefinery-basedproduction of biofuels and high value compounds New Biotechnol 2015 32 588ndash596 [CrossRef] [PubMed]
74 El-Sheekh M Abomohra AE-F Hanelt D Optimization of biomass and fatty acid productivity ofScenedesmus obliquus as a promising microalga for biodiesel production World J Microbiol Biotechnol 201329 915ndash922 [CrossRef] [PubMed]
75 Singh P Guldhe A Kumari S Rawat I Bux F Investigation of combined effect of nitrogen phosphorusand iron on lipid productivity of microalgae Ankistrodesmus falcatus KJ671624 using response surfacemethodology Biochem Eng J 2015 94 22ndash29 [CrossRef]
76 Kilham S Kreeger D Goulden C Lynn S Effects of nutrient limitation on biochemical constituents ofAnkistrodesmus falcatus Freshw Biol 1997 38 591ndash596 [CrossRef]
77 Breuer G Lamers PP Martens DE Draaisma RB Wijffels RH The impact of nitrogen starvation on thedynamics of triacylglycerol accumulation in nine microalgae strains Bioresour Technol 2012 124 217ndash226[CrossRef] [PubMed]
78 Breuer G de Jaeger L Artus VG Martens DE Springer J Draaisma RB Eggink G Wijffels RHLamers PP Superior triacylglycerol (TAG) accumulation in starchless mutants of Scenedesmus obliquus (II)evaluation of TAG yield and productivity in controlled photobioreactors Biotechnol Biofuels 2014 7 70[CrossRef] [PubMed]
79 Nigam S Prakash-Rai M Sharma R Effect of Nitrogen on Growth and Lipid Content ofChlorella pyrenoidosa Am J Biochem Biotechnol 2011 7 126ndash131 [CrossRef]
80 Piligaev AV Sorokina KN Bryanskaya AV Peltek SE Kolchanov NA Parmon VN Isolation ofprospective microalgal strains with high saturated fatty acid content for biofuel production Algal Res 201512 368ndash376 [CrossRef]
81 Weiss SB Kennedy EP The enzymatic synthesis of triglycerides J Am Chem Soc 1956 78 3550[CrossRef]
82 Miller R Wu G Deshpande RR Vieler A Gaumlrtner K Li X Moellering ER Zaumluner S Cornish AJLiu B et al Changes in Transcript Abundance in Chlamydomonas reinhardtii following Nitrogen DeprivationPredict Diversion of Metabolism Plant Physiol 2010 154 1737ndash1752 [CrossRef] [PubMed]
83 Rismani-Yazdi H Haznedaroglu BZ Bibby K Peccia J Transcriptome sequencing and annotation of themicroalgae Dunaliella tertiolecta Pathway description and gene discovery for production of next-generationbiofuels BMC Genom 2011 12 148 [CrossRef] [PubMed]
84 Ramos MJ Fernaacutendez CM Casas A Rodriacuteguez L Peacuterez A Influence of fatty acid composition of rawmaterials on biodiesel properties Bioresour Technol 2009 100 261ndash268 [CrossRef] [PubMed]
85 Knothe G Dependence of biodiesel fuel properties on the structure of fatty acid alkyl estersFuel Process Technol 2005 86 1059ndash1070 [CrossRef]
86 Knothe G ldquoDesignerrdquo Biodiesel Optimizing Fatty Ester Composition to Improve Fuel PropertiesEnergy Fuels 2008 22 1358ndash1364 [CrossRef]
87 Smith PC Ngothai Y Dzuy Nguyen Q OrsquoNeill BK Improving the low-temperature properties ofbiodiesel Methods and consequences Renew Energy 2010 35 1145ndash1151 [CrossRef]
88 Dunn RO Effect of antioxidants on the oxidative stability of methyl soyate (biodiesel) Fuel Process Technol2005 86 1071ndash1085 [CrossRef]
89 Knothe G Improving biodiesel fuel properties by modifying fatty ester composition Energy Environ Sci2009 2 759ndash766 [CrossRef]
copy 2017 by the authors licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)

Energies 2017 10 224 4 of 16
Clustal omega [25] The unrooted phylogenetic tree was constructed using the neighbor-joining (NJ)method [26]] and Kimurarsquos two-parameter algorithm [27] as implemented within the MEGA 606software [28] The bootstrap analysis [29] was used with 1000 replications to test the relative supportfor the branches produced by NJ analysis
26 Microalgae Cultivation Biomass and Total Lipids Measurements
Two hundred milliliters Chu medium No 10 in a 500-mL Erlenmeyer flask were inoculatedwith the microalgae cells (OD680 005) and incubated at 25 plusmn 1 C using 80 microEmiddotmminus2middotsminus1 intensity ofcool-white fluorescent light and continuous agitation at 150 rpm for four weeks In this culture stage allmicroalgae strains were found in the stationary growth phase and centrifuged at 1920times g for 10 min toharvest the microalgae cells The cell pellets were rinsed with 50 mL deionized water centrifuged againdried in an oven at 70 C to a constant weight and the dry weight was determined gravimetricallywith an analytical balance Kern ABJ 220-4NM (Kern amp Sohn GmbH Balingen Germany) The algalpowders were stored at minus20 C for total lipid analysis
Total lipid was extracted using a modified protocol of Bligh and Dyer [30] For each sampleapproximately 50 mg of microalgal powder were mixed with 3 mL of chloroformmethanol (21 vv)and homogenized in an ultrasonic Branson 2510 (Branson Danbury CT USA) for 5 min The mixturewas then centrifuged at 20000times g for 5 min and the supernatant was transferred into a pre-weighedvial Cellular debris was mixed with another 2 mL of chloroformmethanol (21 vv) by vortex andthen centrifuged as described above After that the pooled organic phases were filtered throughsyringe filters of 045 microm evaporated with nitrogen gas and dried at 80 C in a vacuum ovenVacucell 55mdashComfort (MMM Muumlnchen Germany) to a constant weight The lipid content andproductivity of each sample were measured gravimetrically using a semi-micro analytical balance(Sartorius MSU225S-000-DU Foster City CA USA) and calculated as follows [31]
Total lipid ( dry weight) = WLWA times 100
where WL (g) is the weight of the total lipids extracted and WA (g) is the weight of the drymicroalgae biomass
PLipid (mgmiddotLminus1middotdminus1) = WA (g) times CLipid ()V(L) times T(d)
where PLipid is the lipid productivity CLipid is the lipid content V is the working volume and T is thecultivation time
27 Strains Culture under Nitrogen-Sufficient and Nitrogen-Deficient Medium
The selected microalgae species Acutodesmus obliquus (KP8785051) Ankistrodesmus sp(KP8785001) and Chlorella lewinii (KP8785011) (Figure S3) were grown in 500-mL Erlenmeyerflasks containing 250 mL of liquid Chu medium No 10 with nitrogen sufficient (5756 microgmL ofCa(NO3)2middot4H2O) or liquid Chu medium No 10 without a nitrogen source (deficient) for seven days ina controlled culture room at 25 plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity ofcool-white fluorescent light and continuous agitation at 150 rpm
For growth profile analysis the maximum absorption peak for each microalgae strain wasdetermined in the 600ndash800 nm range by spectrophotometric analysis using a Nanodrop 2000 UV-VisSpectrophotometer (Figure S4) Then aliquots of the microalgal cultures were evaluated every 24 h(between 8 and 9 h) at 680 nm The specific growth rate (micro) was calculated during the exponentialgrowth phase (the first three days of culture) according to the equation micro = (ln Nf minus ln Ni)(tf minus ti)where N is the optical density (OD680) at final (f) or initial (i) time (t) Previously it was determinedthat a high (R2 from 0992 to 0998) and positive correlation exists between optical density and celldensities this variable was calculated using a hemocytometer The experiments were carried out intriplicate and the data were expressed as the mean plusmn standard deviation (plusmnSD)
Energies 2017 10 224 5 of 16
28 Determination of Total Protein Total Carbohydrate Ash Content Esterification and Fatty Acids Profiling
The microalgae cultures maintained for seven days under +N and minusN Chu medium No 10were harvested by centrifugation rinsed and dried as previously described (Section 25) Fromthe microalgae powders total protein content was determined according to the HartreendashLowrymethod [32] To 5 mg of dry microalgal biomass was added 5 mL of 05 M NaOH and incubated for10 min at 80 C then centrifuged for 5 min at 14000times g Protein content in 1 mL of the supernatantwas determined by adding 05 mL of 05 M sodium carbonate 05 mL of 05 M sodium potassiumtartrate 05 M of copper sulfate and 2 mL of Folin-Ciocalteu reagent The blue complex was analyzedin a Varian Cary 50 Bio UV-Visible Spectrophotometer (Agilent Technologies Santa Clara CA USA)set at 650 nm against a calibration curve based on a known concentration of BSA as the standard
Total carbohydrate content of the microalgal biomass was determined with the phenol-sulfuricacid method [3233] For this purpose 5 mg of dry microalgae biomass were added to 5 mL of waterAfter this a 1-mL aliquot of the sample was added to 3 mL of sulfuric acid and 1 mL of 5 aqueoussolution of phenol and the mixture was stirred and incubated for 5 min at 90 C The yellow-browncomplex was analyzed by spectrophotometric analysis at 488 nm against a calibration curve based ona known concentration of glucose
Ash content was done according to the AOAC procedure [34] One hundred milligrams of drymaterial were ashed in a muffle furnace Thermolynetrade F6010 (Thermo Fisher Scientific WalthmanMA USA) at 550 C for 16 h and the content was determined gravimetrically All analyses werecarried out in triplicate and the data are expressed as the mean plusmn SD
Fatty acids methyl esters (FAME) were prepared by acid transesterification according to Ichiharaand Fukubayashi [35] The crude lipid extract (1 mg) was dissolved in 02 mL of toluene 15 mL ofmethanol and 03 mL of 08 HCl (prepared in methanolwater 8515 vv) transferred into a cappedtest tube mixed for 5 min and then incubated at 45 C for 12 h for derivatization reaction Then 2 mLof n-hexane and 2 mL of distilled water were added to the tube After vortexing the hexane phasescontaining FAMEs were transferred into a 15-mL tube and dried under a stream of nitrogen Finallythe FAMEs were redissolved in 10 microL acetonitrile and analyzed by using a gas chromatograph VarianCP-3800 GC (Agilent Technologies Santa Clara CA USA) equipped with an automated sampler andinjector a flame ionization detector and using a 30 m times 032 mm times 025 microm Stabilwaxreg capillarycolumn (Restek Bellefonte PA USA) The GC conditions were as follows injector temperature 250 Cthe column temperature gradient was 120 C for 1 min followed by an increase to 160 C at the rateof 30 Cmin 160 C for 1 min followed by an increase to 240 C at the rate of 4 Cmin and 240 Cfor 7 min detector temperature 260 C Gas pressures for He H2 and synthetic air were maintained at40 80 and 60 psi respectively The carrier gas (He) flow was maintained at 1 mLmin The fatty acidswere analyzed by comparing their retention time of the corresponding peaks with a known standardmixture of FAMEs (Nu-Check Prep Elysian MN USA) and methyl tricosanoate (Sigma-AldrichSaint Louis MO USA) was added to each sample as the internal standard All chromatograms ofthe microalgal samples were analyzed by using the Galaxietrade Chromatography Data System Version1932 (Agilent Technologies Santa Clara CA USA) All peaks spanning a peak area of more than50 units were integrated All analyses were carried out in triplicate and the data are expressed as themean plusmn SD
29 Statistical Analysis
Statistical analyses were performed using the IBM SPSS Statistics 210 software (IBM ArmonkNY USA) The growth rate biomass and lipid content data for microalgal strains were analyzedusing a one-way analysis of variance (ANOVA) and the HSD Tukey test The correlation of biomassproductivity and lipid content was analyzed using bivariate correlations and Pearsonrsquos correlationcoefficients are given with their significance levels Differences were considered significant at p lt 005All data presented here are given as the mean plusmn SD
Energies 2017 10 224 6 of 16
3 Results and Discussion
31 Isolation and Identification of the Microalgae Strains
Bioprospecting of tropical ecosystems can provide exceptional native microalgae strains withthe potential for the production of biodiesel nutraceuticals bioactive chemicals and other highvalue products [36ndash38] Furthermore native microalgae strains can have potential applications forefficient biological sequestration of CO2 wastewater treatment and other environmentally-friendlyapplications [38ndash40] Consequently isolation is a fundamental process to obtain pure cultures and isthe first phase towards the screening and selection of microalgae strains with the potential for biodieselproduction and the aforementioned applications
According to these demands in this research samples were collected from three different typesof water bodies the Amazon River (white-water type) the Itaya and Nanay Rivers both of theblack-water type The white-water rivers originate from the Andean catchments bringing high amountsof nutrient-rich sediments In contrast the black-water rivers drain geochemically-poor watershedsand are characterized by their high content in humic acids low pH and poor nutrient status [41]A total of 14 microalgal strains were isolated using the standard cell washing technique and basedon distinguishable morphological characters under light microscopic examination these strains werepreliminary ascribed to the genera Acutodesmus (Hegewald) Tsarenko 2001 Ankistrodesmus Corda1838 Chlorella Beyerinck Beijerinck 1890 Desmodesmus (Chodat) SS An T Friedl and E Hegewald1999 and Tetradesmus GM Smith 1913 (Table 1) These genera are green algae that belong todivision Chlorophyta class Chlorophyceae and orders Chlorellales and Sphaeropleales Althoughmorphological analysis is frequently used to identify microalgae it is inaccurate and very difficultfor the identification at the species level because the relationship between diagnostic morphologyand biological species boundaries are largely unknown in many micro-eukaryotes [42] Furthermoreaccording to Yu et al [31] the microalgae morphology for the same strain varies in relation to ageand culture conditions Consequently in microalgae the classic taxonomy is supported by moleculartools [42] For example the ITS2 rDNA region is thought to be an excellent marker for molecularphylogenetics studies in lower taxonomic levels due to being highly divergent and fast-evolvingwhich can discriminate among closely-related organisms which otherwise display almost identicalsequences [4344] For this reason the molecular identification based in phylogenetic analysis of ITS2rDNA gene sequences is commonly used to identify andor delineate species [204546]
Table 1 Genera GenBank accession number of ITS2 sequences biomass productivity lipid productivityand lipid content of isolated microalgae strains
Source ofCollection Strain GeneraSpecies
GenBankAccessionNumber
BiomassProductivity
(mgmiddotLminus1middotdminus1)
LipidProductivity
(mgmiddotLminus1middotdminus1)
LipidContent( dw)
Amazon River(0341prime06prime prime S7314prime89prime prime W)
AMA001 Ankistrodesmus sp KP8784961 122 plusmn 03 37 plusmn 01 307 plusmn 15AMA002 Ankistrodesmus sp KP8784971 36 plusmn 04 10 plusmn 02 277 plusmn 21AMA003 Chlorella lewinii KP8785011 223 plusmn 07 51 plusmn 05 227 plusmn 15AMA004 Acutodesmus abliquus KP8785051 316 plusmn 04 64 plusmn 07 203 plusmn 21
Itaya River(0343prime14prime prime S
7314prime178prime prime W)
ITA001 Ankistrodesmus sp KP8784991 59 plusmn 02 40 plusmn 03 437 plusmn 17ITA002 Ankistrodesmus sp KP8785001 136 plusmn 04 57 plusmn 05 306 plusmn 20ITA003 Chlorella lewinii KP8785031 129 plusmn 03 40 plusmn 05 224 plusmn 27ITA004 Chlorella lewinii KP8785041 221 plusmn 07 31 plusmn 04 215 plusmn 23ITA005 Scenedesmus regularis KP8785081 93 plusmn 04 56 plusmn 04 242 plusmn 14ITA006 Scenedesmus dimorphus KP8785091 186 plusmn 06 32 plusmn 03 260 plusmn 17ITA007 Acutodesmus abliquus KP8785101 179 plusmn 04 22 plusmn 03 156 plusmn 19
Nanay River(0342prime02prime prime S7315prime32prime prime W)
NAN001 Ankistrodesmus sp KP8784981 143 plusmn 06 28 plusmn 02 467 plusmn 15NAN002 Desmodesmus tropicus KP8785061 122 plusmn 04 23 plusmn 02 175 plusmn 11NAN003 Tetradesmus sp KP8785071 139 plusmn 04 30 plusmn 03 135 plusmn 12
Energies 2017 10 224 7 of 16
To realize phylogenetic analysis genomic DNA of each isolate strain was purified and used forPCR amplification of ITS2 rDNA The universal primers used to amplify the ITS2 rDNA successfullyamplified the desired DNA fragments (~300 bp) from all microalgal strains (Figure S2) Furtherthese amplicons were sequenced published in the NCBI databases and the corresponding GenBankaccession numbers are presented in Table 1 Figure 1 shows a neighbor-joining phylogenetic treedepicting the relatedness of the native microalgae strain isolates with selected sequences from thepublic GenBank database This phylogram contained three clusters each of which included differentmicroalgae genera For example the first cluster consisted of microalgae of the genera AcutodesmusScenedesmus Tetradesmus and Desmodesmus and six isolated strains (ITA007 AMA004 ITA006 NAN003ITA005 and NAN002) The second cluster consisted of a microalgae of the genus Ankistrodesmus andthe isolated strains AMA001 ITA001 ITA002 NAN001 and AMA002 The third one consisted of amicroalgae of the genus Chlorella and three isolated strains (ITA004 AMA003 and ITA003) Overall theidentification of native microalgae strains presented a good correspondence between morphologicalcharacters and sequence-based phylogenetic analysis Microalgae species of these genera were alsoisolated from different countries of the world such as Brazil [47] Argentina [46] United States [48]India [20] Egypt [49] Taiwan [50] South Korea [45] and China [51] Some of these microalgae generasuch as Ankistrodesmus Chlorella Desmodesmus and Scenedesmus were used in large-scale biodieselproduction using photobioreactors and raceway pond systems [5253]
Energies 2017 10 224 7 of 16
public GenBank database This phylogram contained three clusters each of which included different microalgae genera For example the first cluster consisted of microalgae of the genera Acutodesmus Scenedesmus Tetradesmus and Desmodesmus and six isolated strains (ITA007 AMA004 ITA006 NAN003 ITA005 and NAN002) The second cluster consisted of a microalgae of the genus Ankistrodesmus and the isolated strains AMA001 ITA001 ITA002 NAN001 and AMA002 The third one consisted of a microalgae of the genus Chlorella and three isolated strains (ITA004 AMA003 and ITA003) Overall the identification of native microalgae strains presented a good correspondence between morphological characters and sequence-based phylogenetic analysis Microalgae species of these genera were also isolated from different countries of the world such as Brazil [47] Argentina [46] United States [48] India [20] Egypt [49] Taiwan [50] South Korea [45] and China [51] Some of these microalgae genera such as Ankistrodesmus Chlorella Desmodesmus and Scenedesmus were used in large-scale biodiesel production using photobioreactors and raceway pond systems [5253]
Figure 1 Unrooted phylogenetic tree of isolated microalgae strains using the ITS2 sequences The tree was constructed by the neighbor-joining method and Kimurarsquos two-parameter algorithm in MEGA 60 software based on the multiple sequence alignment by Clustal Omega Bootstrap values of 1000 replicates () are shown at the branches
32 Microalgae Cultivation Biomass and Total Lipids Measurements
The isolated microalgae strains exhibited remarkable differences in productivity parameters and lipid content among individual strains in the stationary growth phase (Table 1) With regard to the biomass productivity the 14 strains showed statistically-significant differences (F = 71911 df = 13 p lt 0001) and ranged from 36 plusmn 04 mgmiddotLminus1middotdminus1 (Ankistrodesmus sp strain AMA002) to 316 plusmn 04 mgmiddotLminus1middotdminus1 (Acutodesmus obliquus strain AMA004) Similar biomass productivities were reported for other marine microalgae species [54] However other investigations show major biomass productivities (ranging
Figure 1 Unrooted phylogenetic tree of isolated microalgae strains using the ITS2 sequences The treewas constructed by the neighbor-joining method and Kimurarsquos two-parameter algorithm in MEGA60 software based on the multiple sequence alignment by Clustal Omega Bootstrap values of1000 replicates () are shown at the branches
Energies 2017 10 224 8 of 16
32 Microalgae Cultivation Biomass and Total Lipids Measurements
The isolated microalgae strains exhibited remarkable differences in productivity parametersand lipid content among individual strains in the stationary growth phase (Table 1) With regardto the biomass productivity the 14 strains showed statistically-significant differences (F = 71911df = 13 p lt 0001) and ranged from 36 plusmn 04 mgmiddotLminus1middotdminus1 (Ankistrodesmus sp strain AMA002) to316 plusmn 04 mgmiddotLminus1middotdminus1 (Acutodesmus obliquus strain AMA004) Similar biomass productivities werereported for other marine microalgae species [54] However other investigations show major biomassproductivities (ranging from 30 to 120 mgmiddotLminus1middotdminus1) in the same and other microalgae genera [455556]These discrepancies may be attributed to differences in the culture conditions in the culture mediacomposition and in the microalgae genotypes Lipid productivity also exhibited statistically-significantdifferences (F = 4774 df = 13 p lt 0001) and ranged from 10plusmn 04 mgmiddotLminus1middotdminus1 to 64 plusmn 07 mgmiddotLminus1middotdminus1
with strains AMA002 and AMA004 respectively These lipid productivities were low in comparison toother reports which reported values from 190 to 433 mgmiddotLminus1middotdminus1 [4555] Furthermore lipid contentas a percentage of the microalgal biomass dry weight showed statistically-significant differences(F = 8509 df = 13 p lt 0001) and ranged from 135 plusmn 12 (Tetradesmus sp strain NAN003) to467 plusmn 15 (Ankistrodesmus sp strain NAN001) These percentage values in total lipid contentare similar to several microalgae species of these genera isolated and characterized by otherresearchers worldwide [20454648ndash5057] Furthermore it is important to emphasize that lipid contentis dependent on the different growth phases Commonly in several microalgae taxa the lipid contentincreases gradually from logarithmic to stationary growth phases [58ndash60] For example Kaur et al [20]reported that the total lipid content of Scenedesmus sp DRLMA9 demonstrated values of 207 inthe late exponential phase and 397 in the stationary phase and Desmodesmus elegans DRLMA13displayed values of 103 in the late exponential phase and 169 in the stationary phase LikewiseChiu et al [61] found that the total lipid content of Nannochloropsis oculata was 308 in the logarithmicphase 397 in the early stationary phase and 504 in the stationary phase These changes in lipidcontent with culture age have been associated with nitrate depletion [61] which causes a channeling ofphotosynthetic ATP to carbohydrate and lipid biosynthesis [60] Finally Pearson correlation analysisof biomass productivity and lipid content was inversely related (r = minus037 p lt 001) which is similarto the results obtained by Liang et al [62] and Rodolfi et al [52] In contrast Li et al [51] andHempel et al [63] found no relationship between these parameters The negative correlation obtainedis likely due to the high metabolic cost of lipid biosynthesis [62]
33 Strains Culture under Nitrogen-Sufficient and Nitrogen-Deficient Medium
The microalgae species Acutodesmus obliquus (KP8785051) Ankistrodesmus sp (KP8785001)and Chlorella lewinii (KP8785011) were selected and further characterized because they displayedthe highest biomass and lipid productivities (Table 1) These microalgae showed noticeablevariations in the growth profile when cultured in Chu medium No 10 with nitrogen (+N) orwithout a nitrogen source (minusN) In +N medium all strains showed exponential growth butChlorella lewinii showed the highest average specific growth rates (micro = 0458 plusmn 0098 dminus1) followed byAcutodesmus obliquus (micro = 0399 plusmn 0112 dminus1) and the lowest value was produced by Ankistrodesmus sp(micro = 0265 plusmn 0013 dminus1) In contrast all cultures displayed a lag phase pattern in their growthprofile under minusN medium (Figure 2) These findings corroborate the results obtained by otherresearchers who demonstrate that the specific growth rate is directly proportional to the initialnitrate concentration [46596465] because the reduction or cessation of microalgae cell divisionis due to nitrogen limitation [66] which is a key nutrient for protein and nucleic acids de novobiosynthesis consequently causing an arrest in the early part of the microalgae cell cycle [6768]In accordance with these reports Li et al [69] showed that nitrogen deficiency induced in the greenmicroalgae Micractinium pusillum a downregulation in the expression of genes involved in cell growthand cell cycle regulation Additionally differences in growth profile and specific growth rates aredependent on several factors such as microalgae genotypes the culture medium composition pH
Energies 2017 10 224 9 of 16
nutrients concentration the cell culture phase (ie log phase stationary phase) and environmentalfactors [4549545770ndash72]Energies 2017 10 224 9 of 16
Figure 2 Growth profiles of three microalgal strains cultivated under +N (filled boxes) and minusN (empty boxes) Chu medium No 10 The culture was incubated in a controlled culture room at 25 plusmn 1 degC with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 150 rpm Experiments were carried out in triplicate
34 Total Protein Total Lipid and Total Carbohydrate Content
Biochemical parameters of the microalgae species such as total protein total lipid and total carbohydrate content exhibited differences and changes under both culture conditions (Figure 3) For example Chlorella lewinii showed the highest protein content Ankistrodesmus sp had the highest total lipid content but Acutodesmus obliquus presented the highest levels of total carbohydrate Therefore these strains can be valuable stocks for the industry because total protein and total carbohydrate are potentially byproducts of microalgal biomass processing [7374] Additionally the three strains cultured under minusN conditions showed a significant increase in total lipid content (from 237 in Acutodesmus obliquus to 667 in Ankistrodesmus sp) and total carbohydrate content (from 119 in Acutodesmus obliquus to 963 in Chlorella lewinii) but showed a decreased in total protein levels (from 242 in Acutodesmus obliquus to 545 in Chlorella lewinii) in comparison to +N cultures (Figure 3) These results are in accordance with well-documented results obtained by other researchers with several microalgae species of the genera Acutodesmus Ankistrodesmus Botryococcus Chlamydomonas Chlorella Desmodesmus Dunaliella Haematococcus Neochloris Phaeodactylum Pseudokirchneriella and Scenedesmus [465058596475ndash80] Nevertheless the accumulation of total lipid induced by nitrogen deficiency is not correlated with lipid productivity but rather is due to a decline and stopping of the microalgae specific growth rate [75] The changes in biomolecule contents suggest redirection of microalgae metabolism during nitrogen starvation such as a decrease in biosynthesisincrease in protein degradation and an increase in the biosynthesis of storage biomolecules such as triacylglycerols and starch The increase in total lipid content could be partially explained for a boost in transcript levels of genes encoding enzymes of the lipid biosynthesis pathways According to this assumption recent investigations showed that genes encoding the enzymes that catalyze the last step in the Kennedy pathway of triacylglycerol biosynthesis [81] such as glycerol-3-phosphate acyltransferase and acyl-glycerol-3-phosphate acyltransferase are upregulated in response to nitrogen starvation [648283] Furthermore the increase in total carbohydrate content can be explained by changes in the expression levels of genes involved in carbohydrate metabolism For example when the oleaginous microalgae Neochloris oleoabundans is cultured under nitrogen starvation the catabolic phase of starch specifically α-amylase which hydrolyzes starch to glucose is strongly repressed [83]
Figure 2 Growth profiles of three microalgal strains cultivated under +N (filled boxes) and minusN(empty boxes) Chu medium No 10 The culture was incubated in a controlled culture room at25 plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent lightand continuous agitation at 150 rpm Experiments were carried out in triplicate
34 Total Protein Total Lipid and Total Carbohydrate Content
Biochemical parameters of the microalgae species such as total protein total lipid and totalcarbohydrate content exhibited differences and changes under both culture conditions (Figure 3)For example Chlorella lewinii showed the highest protein content Ankistrodesmus sp had the highesttotal lipid content but Acutodesmus obliquus presented the highest levels of total carbohydrateTherefore these strains can be valuable stocks for the industry because total protein and totalcarbohydrate are potentially byproducts of microalgal biomass processing [7374] Additionallythe three strains cultured under minusN conditions showed a significant increase in total lipid content(from 237 in Acutodesmus obliquus to 667 in Ankistrodesmus sp) and total carbohydrate content(from 119 in Acutodesmus obliquus to 963 in Chlorella lewinii) but showed a decreased in totalprotein levels (from 242 in Acutodesmus obliquus to 545 in Chlorella lewinii) in comparison to +Ncultures (Figure 3) These results are in accordance with well-documented results obtained by otherresearchers with several microalgae species of the genera Acutodesmus Ankistrodesmus BotryococcusChlamydomonas Chlorella Desmodesmus Dunaliella Haematococcus Neochloris PhaeodactylumPseudokirchneriella and Scenedesmus [465058596475ndash80] Nevertheless the accumulation of totallipid induced by nitrogen deficiency is not correlated with lipid productivity but rather is due toa decline and stopping of the microalgae specific growth rate [75] The changes in biomoleculecontents suggest redirection of microalgae metabolism during nitrogen starvation such as a decreasein biosynthesisincrease in protein degradation and an increase in the biosynthesis of storagebiomolecules such as triacylglycerols and starch The increase in total lipid content could bepartially explained for a boost in transcript levels of genes encoding enzymes of the lipid biosynthesispathways According to this assumption recent investigations showed that genes encoding theenzymes that catalyze the last step in the Kennedy pathway of triacylglycerol biosynthesis [81] such asglycerol-3-phosphate acyltransferase and acyl-glycerol-3-phosphate acyltransferase are upregulated
Energies 2017 10 224 10 of 16
in response to nitrogen starvation [648283] Furthermore the increase in total carbohydrate contentcan be explained by changes in the expression levels of genes involved in carbohydrate metabolismFor example when the oleaginous microalgae Neochloris oleoabundans is cultured under nitrogenstarvation the catabolic phase of starch specifically α-amylase which hydrolyzes starch to glucoseis strongly repressed [83]Energies 2017 10 224 10 of 16
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strains cultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled culture room at 25 plusmn 1 degC with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized with respect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that each microalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majority of fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised 7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii the majority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9) acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectively Finally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic (182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in +N and minusN conditions respectively Our results agree with multiple previously-published reports in that fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent in many green microalgae species [454648ndash5055] It is notable that under nitrogen starvation culture all microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 in Ankistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid (181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findings support the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content of SFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Nevertheless there are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed a decrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increased between 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively However in the same culture condition the three microalgae species showed a decrease in polyunsaturated fatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmus spp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgae strains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFA which improves the quality properties of biodiesel such as an increase
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strainscultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled cultureroom at 25plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescentlight and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized withrespect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that eachmicroalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majorityof fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii themajority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9)acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectivelyFinally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic(182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in+N and minusN conditions respectively Our results agree with multiple previously-published reports inthat fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent inmany green microalgae species [454648ndash5055] It is notable that under nitrogen starvation cultureall microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 inAnkistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid(181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findingssupport the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content ofSFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other
Energies 2017 10 224 11 of 16
researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Neverthelessthere are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed adecrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increasedbetween 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively Howeverin the same culture condition the three microalgae species showed a decrease in polyunsaturatedfatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmusspp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgaestrains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFAwhich improves the quality properties of biodiesel such as an increase in cetane number [84ndash86]Considering the SFA and unsaturated fatty acids (UFA) components approximately 20ndash31 are SFAand 69ndash80 are UFA Hence the SFAUFA ratio in the microalgae strains evaluated ranges between025 and 045 which is similar to the reports of several microalgae species isolated in different latitudesworldwide [203148497072]
Table 2 Effect of nitrogen deficiency on fatty acid profiling in three native microalgal species from thePeruvian Amazon
Fatty AcidsAcutodesmus obliquus Ankistrodesmus sp Chlorella lewinii
+N minusN +N minusN +N minusN
140 - - 052 plusmn 002 088 plusmn 002 - 058 plusmn 001160 2724 plusmn 007 2867 plusmn 017 1868 plusmn 019 2261 plusmn 006 1964 plusmn 021 2123 plusmn 010180 184 plusmn 005 255 plusmn 051 101 plusmn 002 181 plusmn 002 371 plusmn 026 153 plusmn 015
161n-7 023 plusmn 000 025 plusmn 001 071 plusmn 003 055 plusmn 001 085 plusmn 043 038 plusmn 000181n-7 131 plusmn 002 131 plusmn 000 141 plusmn 005 094 plusmn 003 146 plusmn 006 123 plusmn 002181n-9 3213 plusmn 021 3608 plusmn 021 3699 plusmn 004 4047 plusmn 038 1371 plusmn 015 2326 plusmn 041182n-6 1732 plusmn 004 1651 plusmn 016 747 plusmn 018 656 plusmn 007 2034 plusmn 039 1957 plusmn 001183n-3 1130 plusmn 017 854 plusmn 031 1486 plusmn 004 1173 plusmn 012 1815 plusmn 022 1371 plusmn 016183n-6 101 plusmn 001 078 plusmn 002 055 plusmn 000 054 plusmn 002 124 plusmn 006 100 plusmn 020184n-3 250 plusmn 005 177 plusmn 008 631 plusmn 002 601 plusmn 005 491 plusmn 025 403 plusmn 020204n-6 - - - 084 plusmn 008 - -225n-3 - - - 062 plusmn 002 - -
Unidentified 512 plusmn 003 355 plusmn 006 1149 plusmn 004 645 plusmn 003 1599 plusmn 034 1348 plusmn 006SFA 2907 3122 2022 2531 2335 2334UFA 7093 6878 7978 7469 7665 7666
MUFA 3367 3764 3911 4195 1602 2487PUFA 3213 2759 2918 2629 4464 3831SFA +MUFA 6274 6886 5933 6726 3936 4821
It is important to highlight that the fatty acid profile determination is an essential step inthe process of the characterization of microalgae strains because it ultimately affects the qualityof the biodiesel product [8485] The quality properties of biodiesel such as high cetane numberoxidative stability and cold-flow are dependent on the hydrocarbonated chain length of saturated andunsaturated fatty acids [8587] For example biodiesel composed of a mixture of a high percentage ofSFA and MUFA is preferred for increased energy yield superior oxidative stability and higher cetanenumbers but is prone to solidify at low temperature In contrast biodiesel with high levels of PUFAhas decreased energy yield low cetane numbers and excellent cold-flow properties but insufficientoxidative stability [548588] Therefore to solve these performance problems five approaches havebeen developed such as changing the fatty acids profile by physical means genetic modification ofthe feedstock or the use of alternative feedstocks with different fatty acids profiles [89]
4 Conclusions
Fourteen microalgae were isolated from three river basins from the Peruvian Amazon and wereidentified by morphological features and molecular phylogeny approaches These strains were initially
Energies 2017 10 224 12 of 16
characterized in the stationary growth phase and exhibited significant heterogeneity in key parameterssuch as biomass productivity (36ndash316 mgmiddotLminus1middotdminus1) lipid productivity (10ndash64 mgmiddotLminus1middotdminus1) andtotal lipid content (135ndash467) Furthermore three selected strains based on elevated biomass andlipid productivities were cultured under nitrogen-sufficient and nitrogen-deficient medium to boostlipid accumulation Under these culture conditions all strains showed noticeable decreases in totalprotein content and increases in total lipid and total carbohydrate contents In addition these strainspresented modifications in the fatty acids profiles with improvement in biodiesel properties due to apredominance of SFA and MUFA which corresponds to an increase in cetane number The results ofthis research suggest that the isolated native microalgae from the Peruvian Amazon could be suitablerenewable feedstocks for the production of both biodiesel and bioethanol which could contribute tosustainable development at the local regional and national levels as a viable alternative to petroleumexploitation However it will be necessary to develop open closed or hybrid systems for large-scalebiomass production of selected native microalgae strains andor de novo microalgae strains designedto produce biofuels and with the abilities to synthesize useful bioactive compounds to be efficientlyexploited using biorefinery approaches
Supplementary MaterialsThe following are available online at wwwmdpicom1996-1073102224s1 Figure S1 Results of
electrophoretic and spectrophotometric analysis of the genomic DNA isolated from the fourteen microalgaeFigure S2 ITS2-rDNA structure (A) and amplicons obtained (B) from the microalgae isolated Figure S3Microphotographic images of the three microalgae strains analyzed under light and fluorescence stained withNile Red Figure S4 Spectrophotometric scanning for the determination of the maximum absorption peak foreach microalgae strain
Acknowledgments This research was supported by the Peruvian funding agencies Consejo Nacional de CienciaTecnologiacutea e Innovacioacuten Tecnoloacutegica (CONCYTEC) with Grant Contract No 364-CONCYTEC-OAJ and Fondospara la Innovacioacuten Ciencia y Tecnologiacutea (FINCyT) with Grant Contract No 383-PNICP-PIBA-2014
Author Contributions Marianela Cobos and Juan C Castro obtained funds for the research participated inthe study design coordinated activities and participated in the preparation of the manuscript Jae D Paredesparticipated in sample collection microalgae isolation and helped to draft the manuscript J Dylan Maddoxperformed the bioinformatics analysis and participated in the preparation of the manuscript Gabriel Vargas-Aranaand Leenin Flores participated in biochemical analysis and helped to draft the manuscript Carla P Aguilarparticipated in the study design supervised biochemical analysis and helped to draft the manuscriptJorge L Marapara participated in molecular analysis and helped to draft the manuscript
Conflicts of Interest The authors declare no conflict of interest
References
1 BP Global BP Statisical Review of World Energy 2014 Available online httpwwwbpcomcontentdambppdfEnergy-economicsstatistical-review-2014BP-statistical-review-of-world-energy-2014-full-reportpdf (accessed on 20 June 2014)
2 Pachauri RK Reisinger A Cambio Climaacutetico 2007 Informe de Siacutentesis Contribucioacuten de los Grupos de Trabajo III y III al Cuarto Informe de Evaluacioacuten del Grupo Intergubernamental de Expertos Sobre el Cambio Climaacutetico IPCCGeneva Switzerland 2007 p 104
3 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Ley de Promocioacuten del Mercado deBiocombustibles Available online httpwww2congresogobpesicrcendocbibcon4_uibdnsf9E95620CC059138105257C9E005AB2F9$FILE28054pdf (accessed on 12 February 2017)
4 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Reglamento de la Ley de Promocioacuten del Mercadode Biocombustibles Available online httpwww2osinerggobpeMarcoLegaldocrevDS-013-2005-EM-CONCORDADOpdf (accessed on 12 February 2017)
5 Areacutevalo LF Nalvarte J Torres J Ramiacuterez Y Impactos Socio-Econoacutemicos de la Produccioacuten de Biocombustiblesen la Amazoniacutea Peruana Primera SNV-WWF Lima Peruacute 2009
6 Sharma KK Schuhmann H Schenk PM High Lipid Induction in Microalgae for Biodiesel ProductionEnergies 2012 5 1532ndash1553 [CrossRef]
Energies 2017 10 224 13 of 16
7 Schenk PM Thomas-Hall SR Stephens E Marx UC Mussgnug JH Posten C Kruse O Hankamer BSecond Generation Biofuels High-Efficiency Microalgae for Biodiesel Production BioEnergy Res 2008 120ndash43 [CrossRef]
8 Foley JA Asner GP Costa MH Coe MT de Fries R Gibbs HK Howard EA Olson S Patz JRamankutty N et al Amazonia revealed Forest degradation and loss of ecosystem goods and services inthe Amazon Basin Front Ecol Environ 2007 5 25ndash32 [CrossRef]
9 Duong VT Li Y Nowak E Schenk PM Microalgae Isolation and Selection for Prospective BiodieselProduction Energies 2012 5 1835ndash1849 [CrossRef]
10 Georgianna DR Mayfield SP Exploiting diversity and synthetic biology for the production of algalbiofuels Nature 2012 488 329ndash335 [CrossRef] [PubMed]
11 Wijffels RH Barbosa MJ An Outlook on Microalgal Biofuels Science 2010 329 796ndash799 [CrossRef][PubMed]
12 Richmond A Handbook of Microalgal Culture Biotechnology and Applied Phycology Blackwell Sci Ltd HudsonNJ USA 2004
13 Menetrez MY An overview of algae biofuel production and potential environmental impactEnviron Sci Technol 2012 46 7073ndash7085 [CrossRef] [PubMed]
14 Khoo HH Sharratt PN Das P Balasubramanian RK Naraharisetti PK Shaik S Life cycle energy andCO2 analysis of microalgae-to-biodiesel Preliminary results and comparisons Bioresour Technol 2011 1025800ndash5807 [CrossRef] [PubMed]
15 Chu SP The Influence of the Mineral Composition of the Medium on the Growth of Planktonic AlgaePart I Methods and Culture Media J Ecol 1942 30 284ndash325 [CrossRef]
16 Carlos EMB Mariatildengela M Gecircneros de Algas de Aacuteguas Continentais do BrasilmdashChave Para Identificaccedilatildeo eDescriccedilotildees 2nd ed Rima Sao Carlos Brasil 2006
17 Greenspan P Mayer EP Fowler SD Nile red A selective fluorescent stain for intracellular lipid dropletsJ Cell Biol 1985 100 965ndash973 [CrossRef] [PubMed]
18 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissuePhytochem Bull 1987 19 11ndash15
19 Sambrook J Fritsch EF Maniatis T Molecular Cloning A Laboratory Manual 2nd ed Cold Spring HarborLaboratory Press New York NY USA 1989
20 Kaur S Sarkar M Srivastava RB Gogoi HK Kalita MC Fatty acid profiling and molecularcharacterization of some freshwater microalgae from India with potential for biodiesel productionNew Biotechnol 2012 29 332ndash344 [CrossRef] [PubMed]
21 Benson DA Cavanaugh M Clark K Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW GenBankNucleic Acids Res 2013 41 D36ndashD42 [CrossRef] [PubMed]
22 NCBI Resource Coordinators Database Resources of the National Center for Biotechnology InformationNucleic Acids Res 2015 43 D6ndashD17
23 Ye J McGinnis S Madden TL BLAST Improvements for better sequence analysis Nucleic Acids Res 200634 W6ndashW9 [CrossRef] [PubMed]
24 BioEdit Sequence Alignment Editor for Windows 9598NTXPVista7 Available online httpwwwmbioncsuedubioeditbioedithtml (accessed on 2 December 2015)
25 Sievers F Higgins DG Clustal omega accurate alignment of very large numbers of sequencesMethods Mol Biol (Clifton NJ) 2014 1079 105ndash116
26 Saitou N Nei M The neighbor-joining method A new method for reconstructing phylogenetic treesMol Biol Evol 1987 4 406ndash425 [PubMed]
27 Kimura M A simple method for estimating evolutionary rates of base substitutions through comparativestudies of nucleotide sequences J Mol Evol 1980 16 111ndash120 [CrossRef] [PubMed]
28 Tamura K Stecher G Peterson D Filipski A Kumar S MEGA6 Molecular Evolutionary GeneticsAnalysis version 60 Mol Biol Evol 2013 30 2725ndash2729 [CrossRef] [PubMed]
29 Felsenstein J Confidence Limits on Phylogenies An Approach Using the Bootstrap Evolution 1985 39783ndash791 [CrossRef]
30 Bligh EG Dyer WJ A Rapid Method of Total Lipid Extraction and Purification Can J Biochem Physiol1959 37 911ndash917 [CrossRef] [PubMed]
Energies 2017 10 224 14 of 16
31 Yu X Zhao P He C Li J Tang X Zhou J Huang Z Isolation of a novel strain of Monoraphidium spand characterization of its potential application as biodiesel feedstock Bioresour Technol 2012 121 256ndash262[CrossRef] [PubMed]
32 Hartree EF Determination of protein A modification of the lowry method that gives a linear photometricresponse Anal Biochem 1972 48 422ndash427 [CrossRef]
33 DuBois M Gilles KA Hamilton JK Rebers PA Smith F Colorimetric Method for Determination ofSugars and Related Substances Anal Chem 1956 28 350ndash356 [CrossRef]
34 AOAC International AOAC Official Methods of Analysis of the Association of Official Analytical Chemists (AOAC)15th ed AOAC International Washington DC USA 1990 Volume 1
35 Ichihara K Fukubayashi Y Preparation of fatty acid methyl esters for gas-liquid chromatographyJ Lipid Res 2010 51 635ndash640 [CrossRef] [PubMed]
36 Brennan L Owende P Biofuels from microalgaemdashA review of technologies for production processingand extractions of biofuels and co-products Renew Sustain Energ Rev 2010 14 557ndash577 [CrossRef]
37 Raja R Hemaiswarya S Kumar NA Sridhar S Rengasamy R A perspective on the biotechnologicalpotential of microalgae Crit Rev Microbiol 2008 34 77ndash88 [CrossRef] [PubMed]
38 Mata TM Martins AA Caetano NS Microalgae for biodiesel production and other applicationsA review Renew Sustain Energy Rev 2010 14 217ndash232 [CrossRef]
39 De Wilt A Butkovskyi A Tuantet K Leal LH Fernandes TV Langenhoff A Zeeman GMicropollutant removal in an algal treatment system fed with source separated wastewater streamsJ Hazard Mater 2015 304 84ndash92 [CrossRef] [PubMed]
40 Li T Lin G Podola B Melkonian M Continuous removal of zinc from wastewater and mine dumpleachate by a microalgal biofilm PSBR J Hazard Mater 2015 297 112ndash118 [CrossRef] [PubMed]
41 Sioli H The Amazon and its main affluents Hydrography morphology of the river courses and river typesIn Monographiae Biologicae Sioli H Ed Springer Dordrecht The Netherlands 1984 pp 127ndash165
42 Moniz MBJ Kaczmarska I Barcoding of Diatoms Nuclear Encoded ITS Revisited Protist 2010 161 7ndash34[CrossRef] [PubMed]
43 Coleman AW Is there a molecular key to the level of ldquobiological speciesrdquo in eukaryotes A DNA guideMol Phylogenet Evol 2009 50 197ndash203 [CrossRef] [PubMed]
44 Mai JC Coleman AW The Internal Transcribed Spacer 2 Exhibits a Common Secondary Structure inGreen Algae and Flowering Plants J Mol Evol 1997 44 258ndash271 [CrossRef] [PubMed]
45 Abou-Shanab RAI Matter IA Kim S-N Oh Y-K Choi J Jeon B-H Characterization andidentification of lipid-producing microalgae species isolated from a freshwater lake Biomass Bioenergy2011 35 3079ndash3085 [CrossRef]
46 Do Nascimento M Ortiz-Marquez JCF Sanchez-Rizza L Echarte MM Curatti L Bioprospectingfor fast growing and biomass characterization of oleaginous microalgae from South-Eastern Buenos AiresArgentina Bioresour Technol 2012 125 283ndash290 [CrossRef] [PubMed]
47 Araujo GS Matos LJBL Gonccedilalves LRB Fernandes FAN Farias WRL Bioprospecting foroil producing microalgal strains Evaluation of oil and biomass production for ten microalgal strainsBioresour Technol 2011 102 5248ndash5250 [CrossRef] [PubMed]
48 Zhou W Li Y Min M Hu B Chen P Ruan R Local bioprospecting for high-lipid producing microalgalstrains to be grown on concentrated municipal wastewater for biofuel production Bioresour Technol 2011102 6909ndash6919 [CrossRef] [PubMed]
49 Mahmoud EA Farahat LA Abdel Aziz ZK Fatthallah NA Salah El Din RA Evaluation of thepotential for some isolated microalgae to produce biodiesel Egypt J Pet 2015 24 97ndash101 [CrossRef]
50 Ho S Lai Y-Y Chiang C-Y Chen C-NN Chang J-S Selection of elite microalgae for biodieselproduction in tropical conditions using a standardized platform Bioresour Technol 2013 147 135ndash142[CrossRef] [PubMed]
51 Li L Cui J Liu Q Ding Y Liu J Screening and phylogenetic analysis of lipid-rich microalgae Algal Res2015 11 381ndash386 [CrossRef]
52 Rodolfi L Chini Zittelli G Bassi N Padovani G Biondi N Bonini G Tredici MR Microalgae for oilStrain selection induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactorBiotechnol Bioeng 2009 102 100ndash112 [CrossRef] [PubMed]
Energies 2017 10 224 15 of 16
53 Li P Miao X Li R Zhong J In situ biodiesel production from fast-growing and high oil content Chlorellapyrenoidosa in rice straw hydrolysate J Biomed Biotechnol 2011 2011 141207 [CrossRef] [PubMed]
54 Doan TTY Sivaloganathan B Obbard JP Screening of marine microalgae for biodiesel feedstockBiomass Bioenergy 2011 35 2534ndash2544 [CrossRef]
55 Abou-Shanab RAI Hwang J-H Cho Y Min B Jeon B-H Characterization of microalgal speciesisolated from fresh water bodies as a potential source for biodiesel production Appl Energy 2011 883300ndash3306 [CrossRef]
56 Chaichalerm S Pokethitiyook P Yuan W Meetam M Sritong K Pugkaew W Kungvansaichol KKruatrachue M Damrongphol P Culture of microalgal strains isolated from natural habitats in Thailand invarious enriched media Appl Energy 2012 89 296ndash302 [CrossRef]
57 Zhang S Liu P Yang X Hao Z Zhang L Luo N Shi J Isolation and identification by 18S rDNAsequence of high lipid potential microalgal species for fuel production in Hainan Dao Biomass Bioenergy2014 66 197ndash203 [CrossRef]
58 Chiu S-Y Kao C-Y Tsai M-T Ong S-C Chen C-H Lin C-S Lipid accumulation and CO2 utilizationof Nannochloropsis oculata in response to CO2 aeration Bioresour Technol 2009 100 833ndash838 [CrossRef][PubMed]
59 Huerlimann R de Nys R Heimann K Growth lipid content productivity and fatty acid composition oftropical microalgae for scale-up production Biotechnol Bioeng 2010 107 245ndash257 [CrossRef] [PubMed]
60 Liang Y Sarkany N Cui Y Biomass and lipid productivities of Chlorella vulgaris under autotrophicheterotrophic and mixotrophic growth conditions Biotechnol Lett 2009 31 1043ndash1049 [CrossRef] [PubMed]
61 Hempel N Petrick I Behrendt F Biomass productivity and productivity of fatty acids and amino acidsof microalgae strains as key characteristics of suitability for biodiesel production J Appl Phycol 2012 241407ndash1418 [CrossRef] [PubMed]
62 Yeesang C Cheirsilp B Effect of nitrogen salt and iron content in the growth medium and light intensityon lipid production by microalgae isolated from freshwater sources in Thailand Bioresour Technol 2011 1023034ndash3040 [CrossRef] [PubMed]
63 Msanne J Xu D Konda AR Casas-Mollano JA Awada T Cahoon EB Cerutti H Metabolic andgene expression changes triggered by nitrogen deprivation in the photoautotrophically grown microalgaeChlamydomonas reinhardtii and Coccomyxa sp C-169 Phytochemistry 2012 75 50ndash59 [CrossRef] [PubMed]
64 Lin Q Lin J Effects of nitrogen source and concentration on biomass and oil production of aScenedesmus rubescens like microalga Bioresour Technol 2011 102 1615ndash1621 [CrossRef] [PubMed]
65 Sheehan J Dunahay T Benemann J Roessler P A Look Back at the US Department of Energyrsquos AquaticSpecies ProgrammdashBiodiesel from Algae US Department of Energyrsquos Office of Fuels Development Golden COUSA 1998 p 328
66 Vaulot D Olson RJ Merkel S Chisholm SW Cell-cycle response to nutrient starvation in twophytoplankton species Thalassiosira weissflogii and Hymenomonas carterae Mar Biol 1987 95 625ndash630[CrossRef]
67 Olson RJ Chisholm SW Effects of light and nitrogen limitation on the cell cycle of the dinoflagellateAmphidinium carteri J Plankton Res 1986 8 785ndash793 [CrossRef]
68 Li Y Fei X Deng X Novel molecular insights into nitrogen starvation-induced triacylglycerolsaccumulation revealed by differential gene expression analysis in green algae Micractinium pusillumBiomass Bioenergy 2012 42 199ndash211 [CrossRef]
69 Abdelaziz AEM Leite GB Belhaj MA Hallenbeck PC Screening microalgae native to Quebec forwastewater treatment and biodiesel production Bioresour Technol 2014 157 140ndash148 [CrossRef] [PubMed]
70 Valdez-Ojeda R Gonzaacutelez-Muntildeoz M Us-Vaacutezquez R Narvaacuteez-Zapata J Chavarria-Hernandez JCLoacutepez-Adriaacuten S Barahona-Peacuterez F Toledano-Thompson T Garduntildeo-Soloacuterzano G Escobedo-GraciaMedrano RM Characterization of five fresh water microalgae with potential for biodiesel productionAlgal Res 2015 7 33ndash44 [CrossRef]
71 Shrivastav A Mishra SK Suh WI Farooq W Moon M Kim T-H Kumar K Choi G-G Park MSYang J-W Characterization of newly isolated oleaginous microalga Monoraphidium sp for lipid productionunder different conditions Algal Res 2015 12 289ndash294 [CrossRef]
72 Yen H-W Hu I-C Chen C-Y Ho S-H Lee D-J Chang J-S Microalgae-based biorefinerymdashFrombiofuels to natural products Bioresour Technol 2013 135 166ndash174 [CrossRef] [PubMed]
Energies 2017 10 224 16 of 16
73 Li J Liu Y Cheng JJ Mos M Daroch M Biological potential of microalgae in China for biorefinery-basedproduction of biofuels and high value compounds New Biotechnol 2015 32 588ndash596 [CrossRef] [PubMed]
74 El-Sheekh M Abomohra AE-F Hanelt D Optimization of biomass and fatty acid productivity ofScenedesmus obliquus as a promising microalga for biodiesel production World J Microbiol Biotechnol 201329 915ndash922 [CrossRef] [PubMed]
75 Singh P Guldhe A Kumari S Rawat I Bux F Investigation of combined effect of nitrogen phosphorusand iron on lipid productivity of microalgae Ankistrodesmus falcatus KJ671624 using response surfacemethodology Biochem Eng J 2015 94 22ndash29 [CrossRef]
76 Kilham S Kreeger D Goulden C Lynn S Effects of nutrient limitation on biochemical constituents ofAnkistrodesmus falcatus Freshw Biol 1997 38 591ndash596 [CrossRef]
77 Breuer G Lamers PP Martens DE Draaisma RB Wijffels RH The impact of nitrogen starvation on thedynamics of triacylglycerol accumulation in nine microalgae strains Bioresour Technol 2012 124 217ndash226[CrossRef] [PubMed]
78 Breuer G de Jaeger L Artus VG Martens DE Springer J Draaisma RB Eggink G Wijffels RHLamers PP Superior triacylglycerol (TAG) accumulation in starchless mutants of Scenedesmus obliquus (II)evaluation of TAG yield and productivity in controlled photobioreactors Biotechnol Biofuels 2014 7 70[CrossRef] [PubMed]
79 Nigam S Prakash-Rai M Sharma R Effect of Nitrogen on Growth and Lipid Content ofChlorella pyrenoidosa Am J Biochem Biotechnol 2011 7 126ndash131 [CrossRef]
80 Piligaev AV Sorokina KN Bryanskaya AV Peltek SE Kolchanov NA Parmon VN Isolation ofprospective microalgal strains with high saturated fatty acid content for biofuel production Algal Res 201512 368ndash376 [CrossRef]
81 Weiss SB Kennedy EP The enzymatic synthesis of triglycerides J Am Chem Soc 1956 78 3550[CrossRef]
82 Miller R Wu G Deshpande RR Vieler A Gaumlrtner K Li X Moellering ER Zaumluner S Cornish AJLiu B et al Changes in Transcript Abundance in Chlamydomonas reinhardtii following Nitrogen DeprivationPredict Diversion of Metabolism Plant Physiol 2010 154 1737ndash1752 [CrossRef] [PubMed]
83 Rismani-Yazdi H Haznedaroglu BZ Bibby K Peccia J Transcriptome sequencing and annotation of themicroalgae Dunaliella tertiolecta Pathway description and gene discovery for production of next-generationbiofuels BMC Genom 2011 12 148 [CrossRef] [PubMed]
84 Ramos MJ Fernaacutendez CM Casas A Rodriacuteguez L Peacuterez A Influence of fatty acid composition of rawmaterials on biodiesel properties Bioresour Technol 2009 100 261ndash268 [CrossRef] [PubMed]
85 Knothe G Dependence of biodiesel fuel properties on the structure of fatty acid alkyl estersFuel Process Technol 2005 86 1059ndash1070 [CrossRef]
86 Knothe G ldquoDesignerrdquo Biodiesel Optimizing Fatty Ester Composition to Improve Fuel PropertiesEnergy Fuels 2008 22 1358ndash1364 [CrossRef]
87 Smith PC Ngothai Y Dzuy Nguyen Q OrsquoNeill BK Improving the low-temperature properties ofbiodiesel Methods and consequences Renew Energy 2010 35 1145ndash1151 [CrossRef]
88 Dunn RO Effect of antioxidants on the oxidative stability of methyl soyate (biodiesel) Fuel Process Technol2005 86 1071ndash1085 [CrossRef]
89 Knothe G Improving biodiesel fuel properties by modifying fatty ester composition Energy Environ Sci2009 2 759ndash766 [CrossRef]
copy 2017 by the authors licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)

Energies 2017 10 224 5 of 16
28 Determination of Total Protein Total Carbohydrate Ash Content Esterification and Fatty Acids Profiling
The microalgae cultures maintained for seven days under +N and minusN Chu medium No 10were harvested by centrifugation rinsed and dried as previously described (Section 25) Fromthe microalgae powders total protein content was determined according to the HartreendashLowrymethod [32] To 5 mg of dry microalgal biomass was added 5 mL of 05 M NaOH and incubated for10 min at 80 C then centrifuged for 5 min at 14000times g Protein content in 1 mL of the supernatantwas determined by adding 05 mL of 05 M sodium carbonate 05 mL of 05 M sodium potassiumtartrate 05 M of copper sulfate and 2 mL of Folin-Ciocalteu reagent The blue complex was analyzedin a Varian Cary 50 Bio UV-Visible Spectrophotometer (Agilent Technologies Santa Clara CA USA)set at 650 nm against a calibration curve based on a known concentration of BSA as the standard
Total carbohydrate content of the microalgal biomass was determined with the phenol-sulfuricacid method [3233] For this purpose 5 mg of dry microalgae biomass were added to 5 mL of waterAfter this a 1-mL aliquot of the sample was added to 3 mL of sulfuric acid and 1 mL of 5 aqueoussolution of phenol and the mixture was stirred and incubated for 5 min at 90 C The yellow-browncomplex was analyzed by spectrophotometric analysis at 488 nm against a calibration curve based ona known concentration of glucose
Ash content was done according to the AOAC procedure [34] One hundred milligrams of drymaterial were ashed in a muffle furnace Thermolynetrade F6010 (Thermo Fisher Scientific WalthmanMA USA) at 550 C for 16 h and the content was determined gravimetrically All analyses werecarried out in triplicate and the data are expressed as the mean plusmn SD
Fatty acids methyl esters (FAME) were prepared by acid transesterification according to Ichiharaand Fukubayashi [35] The crude lipid extract (1 mg) was dissolved in 02 mL of toluene 15 mL ofmethanol and 03 mL of 08 HCl (prepared in methanolwater 8515 vv) transferred into a cappedtest tube mixed for 5 min and then incubated at 45 C for 12 h for derivatization reaction Then 2 mLof n-hexane and 2 mL of distilled water were added to the tube After vortexing the hexane phasescontaining FAMEs were transferred into a 15-mL tube and dried under a stream of nitrogen Finallythe FAMEs were redissolved in 10 microL acetonitrile and analyzed by using a gas chromatograph VarianCP-3800 GC (Agilent Technologies Santa Clara CA USA) equipped with an automated sampler andinjector a flame ionization detector and using a 30 m times 032 mm times 025 microm Stabilwaxreg capillarycolumn (Restek Bellefonte PA USA) The GC conditions were as follows injector temperature 250 Cthe column temperature gradient was 120 C for 1 min followed by an increase to 160 C at the rateof 30 Cmin 160 C for 1 min followed by an increase to 240 C at the rate of 4 Cmin and 240 Cfor 7 min detector temperature 260 C Gas pressures for He H2 and synthetic air were maintained at40 80 and 60 psi respectively The carrier gas (He) flow was maintained at 1 mLmin The fatty acidswere analyzed by comparing their retention time of the corresponding peaks with a known standardmixture of FAMEs (Nu-Check Prep Elysian MN USA) and methyl tricosanoate (Sigma-AldrichSaint Louis MO USA) was added to each sample as the internal standard All chromatograms ofthe microalgal samples were analyzed by using the Galaxietrade Chromatography Data System Version1932 (Agilent Technologies Santa Clara CA USA) All peaks spanning a peak area of more than50 units were integrated All analyses were carried out in triplicate and the data are expressed as themean plusmn SD
29 Statistical Analysis
Statistical analyses were performed using the IBM SPSS Statistics 210 software (IBM ArmonkNY USA) The growth rate biomass and lipid content data for microalgal strains were analyzedusing a one-way analysis of variance (ANOVA) and the HSD Tukey test The correlation of biomassproductivity and lipid content was analyzed using bivariate correlations and Pearsonrsquos correlationcoefficients are given with their significance levels Differences were considered significant at p lt 005All data presented here are given as the mean plusmn SD
Energies 2017 10 224 6 of 16
3 Results and Discussion
31 Isolation and Identification of the Microalgae Strains
Bioprospecting of tropical ecosystems can provide exceptional native microalgae strains withthe potential for the production of biodiesel nutraceuticals bioactive chemicals and other highvalue products [36ndash38] Furthermore native microalgae strains can have potential applications forefficient biological sequestration of CO2 wastewater treatment and other environmentally-friendlyapplications [38ndash40] Consequently isolation is a fundamental process to obtain pure cultures and isthe first phase towards the screening and selection of microalgae strains with the potential for biodieselproduction and the aforementioned applications
According to these demands in this research samples were collected from three different typesof water bodies the Amazon River (white-water type) the Itaya and Nanay Rivers both of theblack-water type The white-water rivers originate from the Andean catchments bringing high amountsof nutrient-rich sediments In contrast the black-water rivers drain geochemically-poor watershedsand are characterized by their high content in humic acids low pH and poor nutrient status [41]A total of 14 microalgal strains were isolated using the standard cell washing technique and basedon distinguishable morphological characters under light microscopic examination these strains werepreliminary ascribed to the genera Acutodesmus (Hegewald) Tsarenko 2001 Ankistrodesmus Corda1838 Chlorella Beyerinck Beijerinck 1890 Desmodesmus (Chodat) SS An T Friedl and E Hegewald1999 and Tetradesmus GM Smith 1913 (Table 1) These genera are green algae that belong todivision Chlorophyta class Chlorophyceae and orders Chlorellales and Sphaeropleales Althoughmorphological analysis is frequently used to identify microalgae it is inaccurate and very difficultfor the identification at the species level because the relationship between diagnostic morphologyand biological species boundaries are largely unknown in many micro-eukaryotes [42] Furthermoreaccording to Yu et al [31] the microalgae morphology for the same strain varies in relation to ageand culture conditions Consequently in microalgae the classic taxonomy is supported by moleculartools [42] For example the ITS2 rDNA region is thought to be an excellent marker for molecularphylogenetics studies in lower taxonomic levels due to being highly divergent and fast-evolvingwhich can discriminate among closely-related organisms which otherwise display almost identicalsequences [4344] For this reason the molecular identification based in phylogenetic analysis of ITS2rDNA gene sequences is commonly used to identify andor delineate species [204546]
Table 1 Genera GenBank accession number of ITS2 sequences biomass productivity lipid productivityand lipid content of isolated microalgae strains
Source ofCollection Strain GeneraSpecies
GenBankAccessionNumber
BiomassProductivity
(mgmiddotLminus1middotdminus1)
LipidProductivity
(mgmiddotLminus1middotdminus1)
LipidContent( dw)
Amazon River(0341prime06prime prime S7314prime89prime prime W)
AMA001 Ankistrodesmus sp KP8784961 122 plusmn 03 37 plusmn 01 307 plusmn 15AMA002 Ankistrodesmus sp KP8784971 36 plusmn 04 10 plusmn 02 277 plusmn 21AMA003 Chlorella lewinii KP8785011 223 plusmn 07 51 plusmn 05 227 plusmn 15AMA004 Acutodesmus abliquus KP8785051 316 plusmn 04 64 plusmn 07 203 plusmn 21
Itaya River(0343prime14prime prime S
7314prime178prime prime W)
ITA001 Ankistrodesmus sp KP8784991 59 plusmn 02 40 plusmn 03 437 plusmn 17ITA002 Ankistrodesmus sp KP8785001 136 plusmn 04 57 plusmn 05 306 plusmn 20ITA003 Chlorella lewinii KP8785031 129 plusmn 03 40 plusmn 05 224 plusmn 27ITA004 Chlorella lewinii KP8785041 221 plusmn 07 31 plusmn 04 215 plusmn 23ITA005 Scenedesmus regularis KP8785081 93 plusmn 04 56 plusmn 04 242 plusmn 14ITA006 Scenedesmus dimorphus KP8785091 186 plusmn 06 32 plusmn 03 260 plusmn 17ITA007 Acutodesmus abliquus KP8785101 179 plusmn 04 22 plusmn 03 156 plusmn 19
Nanay River(0342prime02prime prime S7315prime32prime prime W)
NAN001 Ankistrodesmus sp KP8784981 143 plusmn 06 28 plusmn 02 467 plusmn 15NAN002 Desmodesmus tropicus KP8785061 122 plusmn 04 23 plusmn 02 175 plusmn 11NAN003 Tetradesmus sp KP8785071 139 plusmn 04 30 plusmn 03 135 plusmn 12
Energies 2017 10 224 7 of 16
To realize phylogenetic analysis genomic DNA of each isolate strain was purified and used forPCR amplification of ITS2 rDNA The universal primers used to amplify the ITS2 rDNA successfullyamplified the desired DNA fragments (~300 bp) from all microalgal strains (Figure S2) Furtherthese amplicons were sequenced published in the NCBI databases and the corresponding GenBankaccession numbers are presented in Table 1 Figure 1 shows a neighbor-joining phylogenetic treedepicting the relatedness of the native microalgae strain isolates with selected sequences from thepublic GenBank database This phylogram contained three clusters each of which included differentmicroalgae genera For example the first cluster consisted of microalgae of the genera AcutodesmusScenedesmus Tetradesmus and Desmodesmus and six isolated strains (ITA007 AMA004 ITA006 NAN003ITA005 and NAN002) The second cluster consisted of a microalgae of the genus Ankistrodesmus andthe isolated strains AMA001 ITA001 ITA002 NAN001 and AMA002 The third one consisted of amicroalgae of the genus Chlorella and three isolated strains (ITA004 AMA003 and ITA003) Overall theidentification of native microalgae strains presented a good correspondence between morphologicalcharacters and sequence-based phylogenetic analysis Microalgae species of these genera were alsoisolated from different countries of the world such as Brazil [47] Argentina [46] United States [48]India [20] Egypt [49] Taiwan [50] South Korea [45] and China [51] Some of these microalgae generasuch as Ankistrodesmus Chlorella Desmodesmus and Scenedesmus were used in large-scale biodieselproduction using photobioreactors and raceway pond systems [5253]
Energies 2017 10 224 7 of 16
public GenBank database This phylogram contained three clusters each of which included different microalgae genera For example the first cluster consisted of microalgae of the genera Acutodesmus Scenedesmus Tetradesmus and Desmodesmus and six isolated strains (ITA007 AMA004 ITA006 NAN003 ITA005 and NAN002) The second cluster consisted of a microalgae of the genus Ankistrodesmus and the isolated strains AMA001 ITA001 ITA002 NAN001 and AMA002 The third one consisted of a microalgae of the genus Chlorella and three isolated strains (ITA004 AMA003 and ITA003) Overall the identification of native microalgae strains presented a good correspondence between morphological characters and sequence-based phylogenetic analysis Microalgae species of these genera were also isolated from different countries of the world such as Brazil [47] Argentina [46] United States [48] India [20] Egypt [49] Taiwan [50] South Korea [45] and China [51] Some of these microalgae genera such as Ankistrodesmus Chlorella Desmodesmus and Scenedesmus were used in large-scale biodiesel production using photobioreactors and raceway pond systems [5253]
Figure 1 Unrooted phylogenetic tree of isolated microalgae strains using the ITS2 sequences The tree was constructed by the neighbor-joining method and Kimurarsquos two-parameter algorithm in MEGA 60 software based on the multiple sequence alignment by Clustal Omega Bootstrap values of 1000 replicates () are shown at the branches
32 Microalgae Cultivation Biomass and Total Lipids Measurements
The isolated microalgae strains exhibited remarkable differences in productivity parameters and lipid content among individual strains in the stationary growth phase (Table 1) With regard to the biomass productivity the 14 strains showed statistically-significant differences (F = 71911 df = 13 p lt 0001) and ranged from 36 plusmn 04 mgmiddotLminus1middotdminus1 (Ankistrodesmus sp strain AMA002) to 316 plusmn 04 mgmiddotLminus1middotdminus1 (Acutodesmus obliquus strain AMA004) Similar biomass productivities were reported for other marine microalgae species [54] However other investigations show major biomass productivities (ranging
Figure 1 Unrooted phylogenetic tree of isolated microalgae strains using the ITS2 sequences The treewas constructed by the neighbor-joining method and Kimurarsquos two-parameter algorithm in MEGA60 software based on the multiple sequence alignment by Clustal Omega Bootstrap values of1000 replicates () are shown at the branches
Energies 2017 10 224 8 of 16
32 Microalgae Cultivation Biomass and Total Lipids Measurements
The isolated microalgae strains exhibited remarkable differences in productivity parametersand lipid content among individual strains in the stationary growth phase (Table 1) With regardto the biomass productivity the 14 strains showed statistically-significant differences (F = 71911df = 13 p lt 0001) and ranged from 36 plusmn 04 mgmiddotLminus1middotdminus1 (Ankistrodesmus sp strain AMA002) to316 plusmn 04 mgmiddotLminus1middotdminus1 (Acutodesmus obliquus strain AMA004) Similar biomass productivities werereported for other marine microalgae species [54] However other investigations show major biomassproductivities (ranging from 30 to 120 mgmiddotLminus1middotdminus1) in the same and other microalgae genera [455556]These discrepancies may be attributed to differences in the culture conditions in the culture mediacomposition and in the microalgae genotypes Lipid productivity also exhibited statistically-significantdifferences (F = 4774 df = 13 p lt 0001) and ranged from 10plusmn 04 mgmiddotLminus1middotdminus1 to 64 plusmn 07 mgmiddotLminus1middotdminus1
with strains AMA002 and AMA004 respectively These lipid productivities were low in comparison toother reports which reported values from 190 to 433 mgmiddotLminus1middotdminus1 [4555] Furthermore lipid contentas a percentage of the microalgal biomass dry weight showed statistically-significant differences(F = 8509 df = 13 p lt 0001) and ranged from 135 plusmn 12 (Tetradesmus sp strain NAN003) to467 plusmn 15 (Ankistrodesmus sp strain NAN001) These percentage values in total lipid contentare similar to several microalgae species of these genera isolated and characterized by otherresearchers worldwide [20454648ndash5057] Furthermore it is important to emphasize that lipid contentis dependent on the different growth phases Commonly in several microalgae taxa the lipid contentincreases gradually from logarithmic to stationary growth phases [58ndash60] For example Kaur et al [20]reported that the total lipid content of Scenedesmus sp DRLMA9 demonstrated values of 207 inthe late exponential phase and 397 in the stationary phase and Desmodesmus elegans DRLMA13displayed values of 103 in the late exponential phase and 169 in the stationary phase LikewiseChiu et al [61] found that the total lipid content of Nannochloropsis oculata was 308 in the logarithmicphase 397 in the early stationary phase and 504 in the stationary phase These changes in lipidcontent with culture age have been associated with nitrate depletion [61] which causes a channeling ofphotosynthetic ATP to carbohydrate and lipid biosynthesis [60] Finally Pearson correlation analysisof biomass productivity and lipid content was inversely related (r = minus037 p lt 001) which is similarto the results obtained by Liang et al [62] and Rodolfi et al [52] In contrast Li et al [51] andHempel et al [63] found no relationship between these parameters The negative correlation obtainedis likely due to the high metabolic cost of lipid biosynthesis [62]
33 Strains Culture under Nitrogen-Sufficient and Nitrogen-Deficient Medium
The microalgae species Acutodesmus obliquus (KP8785051) Ankistrodesmus sp (KP8785001)and Chlorella lewinii (KP8785011) were selected and further characterized because they displayedthe highest biomass and lipid productivities (Table 1) These microalgae showed noticeablevariations in the growth profile when cultured in Chu medium No 10 with nitrogen (+N) orwithout a nitrogen source (minusN) In +N medium all strains showed exponential growth butChlorella lewinii showed the highest average specific growth rates (micro = 0458 plusmn 0098 dminus1) followed byAcutodesmus obliquus (micro = 0399 plusmn 0112 dminus1) and the lowest value was produced by Ankistrodesmus sp(micro = 0265 plusmn 0013 dminus1) In contrast all cultures displayed a lag phase pattern in their growthprofile under minusN medium (Figure 2) These findings corroborate the results obtained by otherresearchers who demonstrate that the specific growth rate is directly proportional to the initialnitrate concentration [46596465] because the reduction or cessation of microalgae cell divisionis due to nitrogen limitation [66] which is a key nutrient for protein and nucleic acids de novobiosynthesis consequently causing an arrest in the early part of the microalgae cell cycle [6768]In accordance with these reports Li et al [69] showed that nitrogen deficiency induced in the greenmicroalgae Micractinium pusillum a downregulation in the expression of genes involved in cell growthand cell cycle regulation Additionally differences in growth profile and specific growth rates aredependent on several factors such as microalgae genotypes the culture medium composition pH
Energies 2017 10 224 9 of 16
nutrients concentration the cell culture phase (ie log phase stationary phase) and environmentalfactors [4549545770ndash72]Energies 2017 10 224 9 of 16
Figure 2 Growth profiles of three microalgal strains cultivated under +N (filled boxes) and minusN (empty boxes) Chu medium No 10 The culture was incubated in a controlled culture room at 25 plusmn 1 degC with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 150 rpm Experiments were carried out in triplicate
34 Total Protein Total Lipid and Total Carbohydrate Content
Biochemical parameters of the microalgae species such as total protein total lipid and total carbohydrate content exhibited differences and changes under both culture conditions (Figure 3) For example Chlorella lewinii showed the highest protein content Ankistrodesmus sp had the highest total lipid content but Acutodesmus obliquus presented the highest levels of total carbohydrate Therefore these strains can be valuable stocks for the industry because total protein and total carbohydrate are potentially byproducts of microalgal biomass processing [7374] Additionally the three strains cultured under minusN conditions showed a significant increase in total lipid content (from 237 in Acutodesmus obliquus to 667 in Ankistrodesmus sp) and total carbohydrate content (from 119 in Acutodesmus obliquus to 963 in Chlorella lewinii) but showed a decreased in total protein levels (from 242 in Acutodesmus obliquus to 545 in Chlorella lewinii) in comparison to +N cultures (Figure 3) These results are in accordance with well-documented results obtained by other researchers with several microalgae species of the genera Acutodesmus Ankistrodesmus Botryococcus Chlamydomonas Chlorella Desmodesmus Dunaliella Haematococcus Neochloris Phaeodactylum Pseudokirchneriella and Scenedesmus [465058596475ndash80] Nevertheless the accumulation of total lipid induced by nitrogen deficiency is not correlated with lipid productivity but rather is due to a decline and stopping of the microalgae specific growth rate [75] The changes in biomolecule contents suggest redirection of microalgae metabolism during nitrogen starvation such as a decrease in biosynthesisincrease in protein degradation and an increase in the biosynthesis of storage biomolecules such as triacylglycerols and starch The increase in total lipid content could be partially explained for a boost in transcript levels of genes encoding enzymes of the lipid biosynthesis pathways According to this assumption recent investigations showed that genes encoding the enzymes that catalyze the last step in the Kennedy pathway of triacylglycerol biosynthesis [81] such as glycerol-3-phosphate acyltransferase and acyl-glycerol-3-phosphate acyltransferase are upregulated in response to nitrogen starvation [648283] Furthermore the increase in total carbohydrate content can be explained by changes in the expression levels of genes involved in carbohydrate metabolism For example when the oleaginous microalgae Neochloris oleoabundans is cultured under nitrogen starvation the catabolic phase of starch specifically α-amylase which hydrolyzes starch to glucose is strongly repressed [83]
Figure 2 Growth profiles of three microalgal strains cultivated under +N (filled boxes) and minusN(empty boxes) Chu medium No 10 The culture was incubated in a controlled culture room at25 plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent lightand continuous agitation at 150 rpm Experiments were carried out in triplicate
34 Total Protein Total Lipid and Total Carbohydrate Content
Biochemical parameters of the microalgae species such as total protein total lipid and totalcarbohydrate content exhibited differences and changes under both culture conditions (Figure 3)For example Chlorella lewinii showed the highest protein content Ankistrodesmus sp had the highesttotal lipid content but Acutodesmus obliquus presented the highest levels of total carbohydrateTherefore these strains can be valuable stocks for the industry because total protein and totalcarbohydrate are potentially byproducts of microalgal biomass processing [7374] Additionallythe three strains cultured under minusN conditions showed a significant increase in total lipid content(from 237 in Acutodesmus obliquus to 667 in Ankistrodesmus sp) and total carbohydrate content(from 119 in Acutodesmus obliquus to 963 in Chlorella lewinii) but showed a decreased in totalprotein levels (from 242 in Acutodesmus obliquus to 545 in Chlorella lewinii) in comparison to +Ncultures (Figure 3) These results are in accordance with well-documented results obtained by otherresearchers with several microalgae species of the genera Acutodesmus Ankistrodesmus BotryococcusChlamydomonas Chlorella Desmodesmus Dunaliella Haematococcus Neochloris PhaeodactylumPseudokirchneriella and Scenedesmus [465058596475ndash80] Nevertheless the accumulation of totallipid induced by nitrogen deficiency is not correlated with lipid productivity but rather is due toa decline and stopping of the microalgae specific growth rate [75] The changes in biomoleculecontents suggest redirection of microalgae metabolism during nitrogen starvation such as a decreasein biosynthesisincrease in protein degradation and an increase in the biosynthesis of storagebiomolecules such as triacylglycerols and starch The increase in total lipid content could bepartially explained for a boost in transcript levels of genes encoding enzymes of the lipid biosynthesispathways According to this assumption recent investigations showed that genes encoding theenzymes that catalyze the last step in the Kennedy pathway of triacylglycerol biosynthesis [81] such asglycerol-3-phosphate acyltransferase and acyl-glycerol-3-phosphate acyltransferase are upregulated
Energies 2017 10 224 10 of 16
in response to nitrogen starvation [648283] Furthermore the increase in total carbohydrate contentcan be explained by changes in the expression levels of genes involved in carbohydrate metabolismFor example when the oleaginous microalgae Neochloris oleoabundans is cultured under nitrogenstarvation the catabolic phase of starch specifically α-amylase which hydrolyzes starch to glucoseis strongly repressed [83]Energies 2017 10 224 10 of 16
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strains cultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled culture room at 25 plusmn 1 degC with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized with respect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that each microalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majority of fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised 7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii the majority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9) acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectively Finally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic (182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in +N and minusN conditions respectively Our results agree with multiple previously-published reports in that fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent in many green microalgae species [454648ndash5055] It is notable that under nitrogen starvation culture all microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 in Ankistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid (181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findings support the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content of SFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Nevertheless there are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed a decrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increased between 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively However in the same culture condition the three microalgae species showed a decrease in polyunsaturated fatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmus spp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgae strains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFA which improves the quality properties of biodiesel such as an increase
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strainscultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled cultureroom at 25plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescentlight and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized withrespect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that eachmicroalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majorityof fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii themajority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9)acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectivelyFinally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic(182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in+N and minusN conditions respectively Our results agree with multiple previously-published reports inthat fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent inmany green microalgae species [454648ndash5055] It is notable that under nitrogen starvation cultureall microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 inAnkistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid(181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findingssupport the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content ofSFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other
Energies 2017 10 224 11 of 16
researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Neverthelessthere are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed adecrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increasedbetween 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively Howeverin the same culture condition the three microalgae species showed a decrease in polyunsaturatedfatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmusspp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgaestrains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFAwhich improves the quality properties of biodiesel such as an increase in cetane number [84ndash86]Considering the SFA and unsaturated fatty acids (UFA) components approximately 20ndash31 are SFAand 69ndash80 are UFA Hence the SFAUFA ratio in the microalgae strains evaluated ranges between025 and 045 which is similar to the reports of several microalgae species isolated in different latitudesworldwide [203148497072]
Table 2 Effect of nitrogen deficiency on fatty acid profiling in three native microalgal species from thePeruvian Amazon
Fatty AcidsAcutodesmus obliquus Ankistrodesmus sp Chlorella lewinii
+N minusN +N minusN +N minusN
140 - - 052 plusmn 002 088 plusmn 002 - 058 plusmn 001160 2724 plusmn 007 2867 plusmn 017 1868 plusmn 019 2261 plusmn 006 1964 plusmn 021 2123 plusmn 010180 184 plusmn 005 255 plusmn 051 101 plusmn 002 181 plusmn 002 371 plusmn 026 153 plusmn 015
161n-7 023 plusmn 000 025 plusmn 001 071 plusmn 003 055 plusmn 001 085 plusmn 043 038 plusmn 000181n-7 131 plusmn 002 131 plusmn 000 141 plusmn 005 094 plusmn 003 146 plusmn 006 123 plusmn 002181n-9 3213 plusmn 021 3608 plusmn 021 3699 plusmn 004 4047 plusmn 038 1371 plusmn 015 2326 plusmn 041182n-6 1732 plusmn 004 1651 plusmn 016 747 plusmn 018 656 plusmn 007 2034 plusmn 039 1957 plusmn 001183n-3 1130 plusmn 017 854 plusmn 031 1486 plusmn 004 1173 plusmn 012 1815 plusmn 022 1371 plusmn 016183n-6 101 plusmn 001 078 plusmn 002 055 plusmn 000 054 plusmn 002 124 plusmn 006 100 plusmn 020184n-3 250 plusmn 005 177 plusmn 008 631 plusmn 002 601 plusmn 005 491 plusmn 025 403 plusmn 020204n-6 - - - 084 plusmn 008 - -225n-3 - - - 062 plusmn 002 - -
Unidentified 512 plusmn 003 355 plusmn 006 1149 plusmn 004 645 plusmn 003 1599 plusmn 034 1348 plusmn 006SFA 2907 3122 2022 2531 2335 2334UFA 7093 6878 7978 7469 7665 7666
MUFA 3367 3764 3911 4195 1602 2487PUFA 3213 2759 2918 2629 4464 3831SFA +MUFA 6274 6886 5933 6726 3936 4821
It is important to highlight that the fatty acid profile determination is an essential step inthe process of the characterization of microalgae strains because it ultimately affects the qualityof the biodiesel product [8485] The quality properties of biodiesel such as high cetane numberoxidative stability and cold-flow are dependent on the hydrocarbonated chain length of saturated andunsaturated fatty acids [8587] For example biodiesel composed of a mixture of a high percentage ofSFA and MUFA is preferred for increased energy yield superior oxidative stability and higher cetanenumbers but is prone to solidify at low temperature In contrast biodiesel with high levels of PUFAhas decreased energy yield low cetane numbers and excellent cold-flow properties but insufficientoxidative stability [548588] Therefore to solve these performance problems five approaches havebeen developed such as changing the fatty acids profile by physical means genetic modification ofthe feedstock or the use of alternative feedstocks with different fatty acids profiles [89]
4 Conclusions
Fourteen microalgae were isolated from three river basins from the Peruvian Amazon and wereidentified by morphological features and molecular phylogeny approaches These strains were initially
Energies 2017 10 224 12 of 16
characterized in the stationary growth phase and exhibited significant heterogeneity in key parameterssuch as biomass productivity (36ndash316 mgmiddotLminus1middotdminus1) lipid productivity (10ndash64 mgmiddotLminus1middotdminus1) andtotal lipid content (135ndash467) Furthermore three selected strains based on elevated biomass andlipid productivities were cultured under nitrogen-sufficient and nitrogen-deficient medium to boostlipid accumulation Under these culture conditions all strains showed noticeable decreases in totalprotein content and increases in total lipid and total carbohydrate contents In addition these strainspresented modifications in the fatty acids profiles with improvement in biodiesel properties due to apredominance of SFA and MUFA which corresponds to an increase in cetane number The results ofthis research suggest that the isolated native microalgae from the Peruvian Amazon could be suitablerenewable feedstocks for the production of both biodiesel and bioethanol which could contribute tosustainable development at the local regional and national levels as a viable alternative to petroleumexploitation However it will be necessary to develop open closed or hybrid systems for large-scalebiomass production of selected native microalgae strains andor de novo microalgae strains designedto produce biofuels and with the abilities to synthesize useful bioactive compounds to be efficientlyexploited using biorefinery approaches
Supplementary MaterialsThe following are available online at wwwmdpicom1996-1073102224s1 Figure S1 Results of
electrophoretic and spectrophotometric analysis of the genomic DNA isolated from the fourteen microalgaeFigure S2 ITS2-rDNA structure (A) and amplicons obtained (B) from the microalgae isolated Figure S3Microphotographic images of the three microalgae strains analyzed under light and fluorescence stained withNile Red Figure S4 Spectrophotometric scanning for the determination of the maximum absorption peak foreach microalgae strain
Acknowledgments This research was supported by the Peruvian funding agencies Consejo Nacional de CienciaTecnologiacutea e Innovacioacuten Tecnoloacutegica (CONCYTEC) with Grant Contract No 364-CONCYTEC-OAJ and Fondospara la Innovacioacuten Ciencia y Tecnologiacutea (FINCyT) with Grant Contract No 383-PNICP-PIBA-2014
Author Contributions Marianela Cobos and Juan C Castro obtained funds for the research participated inthe study design coordinated activities and participated in the preparation of the manuscript Jae D Paredesparticipated in sample collection microalgae isolation and helped to draft the manuscript J Dylan Maddoxperformed the bioinformatics analysis and participated in the preparation of the manuscript Gabriel Vargas-Aranaand Leenin Flores participated in biochemical analysis and helped to draft the manuscript Carla P Aguilarparticipated in the study design supervised biochemical analysis and helped to draft the manuscriptJorge L Marapara participated in molecular analysis and helped to draft the manuscript
Conflicts of Interest The authors declare no conflict of interest
References
1 BP Global BP Statisical Review of World Energy 2014 Available online httpwwwbpcomcontentdambppdfEnergy-economicsstatistical-review-2014BP-statistical-review-of-world-energy-2014-full-reportpdf (accessed on 20 June 2014)
2 Pachauri RK Reisinger A Cambio Climaacutetico 2007 Informe de Siacutentesis Contribucioacuten de los Grupos de Trabajo III y III al Cuarto Informe de Evaluacioacuten del Grupo Intergubernamental de Expertos Sobre el Cambio Climaacutetico IPCCGeneva Switzerland 2007 p 104
3 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Ley de Promocioacuten del Mercado deBiocombustibles Available online httpwww2congresogobpesicrcendocbibcon4_uibdnsf9E95620CC059138105257C9E005AB2F9$FILE28054pdf (accessed on 12 February 2017)
4 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Reglamento de la Ley de Promocioacuten del Mercadode Biocombustibles Available online httpwww2osinerggobpeMarcoLegaldocrevDS-013-2005-EM-CONCORDADOpdf (accessed on 12 February 2017)
5 Areacutevalo LF Nalvarte J Torres J Ramiacuterez Y Impactos Socio-Econoacutemicos de la Produccioacuten de Biocombustiblesen la Amazoniacutea Peruana Primera SNV-WWF Lima Peruacute 2009
6 Sharma KK Schuhmann H Schenk PM High Lipid Induction in Microalgae for Biodiesel ProductionEnergies 2012 5 1532ndash1553 [CrossRef]
Energies 2017 10 224 13 of 16
7 Schenk PM Thomas-Hall SR Stephens E Marx UC Mussgnug JH Posten C Kruse O Hankamer BSecond Generation Biofuels High-Efficiency Microalgae for Biodiesel Production BioEnergy Res 2008 120ndash43 [CrossRef]
8 Foley JA Asner GP Costa MH Coe MT de Fries R Gibbs HK Howard EA Olson S Patz JRamankutty N et al Amazonia revealed Forest degradation and loss of ecosystem goods and services inthe Amazon Basin Front Ecol Environ 2007 5 25ndash32 [CrossRef]
9 Duong VT Li Y Nowak E Schenk PM Microalgae Isolation and Selection for Prospective BiodieselProduction Energies 2012 5 1835ndash1849 [CrossRef]
10 Georgianna DR Mayfield SP Exploiting diversity and synthetic biology for the production of algalbiofuels Nature 2012 488 329ndash335 [CrossRef] [PubMed]
11 Wijffels RH Barbosa MJ An Outlook on Microalgal Biofuels Science 2010 329 796ndash799 [CrossRef][PubMed]
12 Richmond A Handbook of Microalgal Culture Biotechnology and Applied Phycology Blackwell Sci Ltd HudsonNJ USA 2004
13 Menetrez MY An overview of algae biofuel production and potential environmental impactEnviron Sci Technol 2012 46 7073ndash7085 [CrossRef] [PubMed]
14 Khoo HH Sharratt PN Das P Balasubramanian RK Naraharisetti PK Shaik S Life cycle energy andCO2 analysis of microalgae-to-biodiesel Preliminary results and comparisons Bioresour Technol 2011 1025800ndash5807 [CrossRef] [PubMed]
15 Chu SP The Influence of the Mineral Composition of the Medium on the Growth of Planktonic AlgaePart I Methods and Culture Media J Ecol 1942 30 284ndash325 [CrossRef]
16 Carlos EMB Mariatildengela M Gecircneros de Algas de Aacuteguas Continentais do BrasilmdashChave Para Identificaccedilatildeo eDescriccedilotildees 2nd ed Rima Sao Carlos Brasil 2006
17 Greenspan P Mayer EP Fowler SD Nile red A selective fluorescent stain for intracellular lipid dropletsJ Cell Biol 1985 100 965ndash973 [CrossRef] [PubMed]
18 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissuePhytochem Bull 1987 19 11ndash15
19 Sambrook J Fritsch EF Maniatis T Molecular Cloning A Laboratory Manual 2nd ed Cold Spring HarborLaboratory Press New York NY USA 1989
20 Kaur S Sarkar M Srivastava RB Gogoi HK Kalita MC Fatty acid profiling and molecularcharacterization of some freshwater microalgae from India with potential for biodiesel productionNew Biotechnol 2012 29 332ndash344 [CrossRef] [PubMed]
21 Benson DA Cavanaugh M Clark K Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW GenBankNucleic Acids Res 2013 41 D36ndashD42 [CrossRef] [PubMed]
22 NCBI Resource Coordinators Database Resources of the National Center for Biotechnology InformationNucleic Acids Res 2015 43 D6ndashD17
23 Ye J McGinnis S Madden TL BLAST Improvements for better sequence analysis Nucleic Acids Res 200634 W6ndashW9 [CrossRef] [PubMed]
24 BioEdit Sequence Alignment Editor for Windows 9598NTXPVista7 Available online httpwwwmbioncsuedubioeditbioedithtml (accessed on 2 December 2015)
25 Sievers F Higgins DG Clustal omega accurate alignment of very large numbers of sequencesMethods Mol Biol (Clifton NJ) 2014 1079 105ndash116
26 Saitou N Nei M The neighbor-joining method A new method for reconstructing phylogenetic treesMol Biol Evol 1987 4 406ndash425 [PubMed]
27 Kimura M A simple method for estimating evolutionary rates of base substitutions through comparativestudies of nucleotide sequences J Mol Evol 1980 16 111ndash120 [CrossRef] [PubMed]
28 Tamura K Stecher G Peterson D Filipski A Kumar S MEGA6 Molecular Evolutionary GeneticsAnalysis version 60 Mol Biol Evol 2013 30 2725ndash2729 [CrossRef] [PubMed]
29 Felsenstein J Confidence Limits on Phylogenies An Approach Using the Bootstrap Evolution 1985 39783ndash791 [CrossRef]
30 Bligh EG Dyer WJ A Rapid Method of Total Lipid Extraction and Purification Can J Biochem Physiol1959 37 911ndash917 [CrossRef] [PubMed]
Energies 2017 10 224 14 of 16
31 Yu X Zhao P He C Li J Tang X Zhou J Huang Z Isolation of a novel strain of Monoraphidium spand characterization of its potential application as biodiesel feedstock Bioresour Technol 2012 121 256ndash262[CrossRef] [PubMed]
32 Hartree EF Determination of protein A modification of the lowry method that gives a linear photometricresponse Anal Biochem 1972 48 422ndash427 [CrossRef]
33 DuBois M Gilles KA Hamilton JK Rebers PA Smith F Colorimetric Method for Determination ofSugars and Related Substances Anal Chem 1956 28 350ndash356 [CrossRef]
34 AOAC International AOAC Official Methods of Analysis of the Association of Official Analytical Chemists (AOAC)15th ed AOAC International Washington DC USA 1990 Volume 1
35 Ichihara K Fukubayashi Y Preparation of fatty acid methyl esters for gas-liquid chromatographyJ Lipid Res 2010 51 635ndash640 [CrossRef] [PubMed]
36 Brennan L Owende P Biofuels from microalgaemdashA review of technologies for production processingand extractions of biofuels and co-products Renew Sustain Energ Rev 2010 14 557ndash577 [CrossRef]
37 Raja R Hemaiswarya S Kumar NA Sridhar S Rengasamy R A perspective on the biotechnologicalpotential of microalgae Crit Rev Microbiol 2008 34 77ndash88 [CrossRef] [PubMed]
38 Mata TM Martins AA Caetano NS Microalgae for biodiesel production and other applicationsA review Renew Sustain Energy Rev 2010 14 217ndash232 [CrossRef]
39 De Wilt A Butkovskyi A Tuantet K Leal LH Fernandes TV Langenhoff A Zeeman GMicropollutant removal in an algal treatment system fed with source separated wastewater streamsJ Hazard Mater 2015 304 84ndash92 [CrossRef] [PubMed]
40 Li T Lin G Podola B Melkonian M Continuous removal of zinc from wastewater and mine dumpleachate by a microalgal biofilm PSBR J Hazard Mater 2015 297 112ndash118 [CrossRef] [PubMed]
41 Sioli H The Amazon and its main affluents Hydrography morphology of the river courses and river typesIn Monographiae Biologicae Sioli H Ed Springer Dordrecht The Netherlands 1984 pp 127ndash165
42 Moniz MBJ Kaczmarska I Barcoding of Diatoms Nuclear Encoded ITS Revisited Protist 2010 161 7ndash34[CrossRef] [PubMed]
43 Coleman AW Is there a molecular key to the level of ldquobiological speciesrdquo in eukaryotes A DNA guideMol Phylogenet Evol 2009 50 197ndash203 [CrossRef] [PubMed]
44 Mai JC Coleman AW The Internal Transcribed Spacer 2 Exhibits a Common Secondary Structure inGreen Algae and Flowering Plants J Mol Evol 1997 44 258ndash271 [CrossRef] [PubMed]
45 Abou-Shanab RAI Matter IA Kim S-N Oh Y-K Choi J Jeon B-H Characterization andidentification of lipid-producing microalgae species isolated from a freshwater lake Biomass Bioenergy2011 35 3079ndash3085 [CrossRef]
46 Do Nascimento M Ortiz-Marquez JCF Sanchez-Rizza L Echarte MM Curatti L Bioprospectingfor fast growing and biomass characterization of oleaginous microalgae from South-Eastern Buenos AiresArgentina Bioresour Technol 2012 125 283ndash290 [CrossRef] [PubMed]
47 Araujo GS Matos LJBL Gonccedilalves LRB Fernandes FAN Farias WRL Bioprospecting foroil producing microalgal strains Evaluation of oil and biomass production for ten microalgal strainsBioresour Technol 2011 102 5248ndash5250 [CrossRef] [PubMed]
48 Zhou W Li Y Min M Hu B Chen P Ruan R Local bioprospecting for high-lipid producing microalgalstrains to be grown on concentrated municipal wastewater for biofuel production Bioresour Technol 2011102 6909ndash6919 [CrossRef] [PubMed]
49 Mahmoud EA Farahat LA Abdel Aziz ZK Fatthallah NA Salah El Din RA Evaluation of thepotential for some isolated microalgae to produce biodiesel Egypt J Pet 2015 24 97ndash101 [CrossRef]
50 Ho S Lai Y-Y Chiang C-Y Chen C-NN Chang J-S Selection of elite microalgae for biodieselproduction in tropical conditions using a standardized platform Bioresour Technol 2013 147 135ndash142[CrossRef] [PubMed]
51 Li L Cui J Liu Q Ding Y Liu J Screening and phylogenetic analysis of lipid-rich microalgae Algal Res2015 11 381ndash386 [CrossRef]
52 Rodolfi L Chini Zittelli G Bassi N Padovani G Biondi N Bonini G Tredici MR Microalgae for oilStrain selection induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactorBiotechnol Bioeng 2009 102 100ndash112 [CrossRef] [PubMed]
Energies 2017 10 224 15 of 16
53 Li P Miao X Li R Zhong J In situ biodiesel production from fast-growing and high oil content Chlorellapyrenoidosa in rice straw hydrolysate J Biomed Biotechnol 2011 2011 141207 [CrossRef] [PubMed]
54 Doan TTY Sivaloganathan B Obbard JP Screening of marine microalgae for biodiesel feedstockBiomass Bioenergy 2011 35 2534ndash2544 [CrossRef]
55 Abou-Shanab RAI Hwang J-H Cho Y Min B Jeon B-H Characterization of microalgal speciesisolated from fresh water bodies as a potential source for biodiesel production Appl Energy 2011 883300ndash3306 [CrossRef]
56 Chaichalerm S Pokethitiyook P Yuan W Meetam M Sritong K Pugkaew W Kungvansaichol KKruatrachue M Damrongphol P Culture of microalgal strains isolated from natural habitats in Thailand invarious enriched media Appl Energy 2012 89 296ndash302 [CrossRef]
57 Zhang S Liu P Yang X Hao Z Zhang L Luo N Shi J Isolation and identification by 18S rDNAsequence of high lipid potential microalgal species for fuel production in Hainan Dao Biomass Bioenergy2014 66 197ndash203 [CrossRef]
58 Chiu S-Y Kao C-Y Tsai M-T Ong S-C Chen C-H Lin C-S Lipid accumulation and CO2 utilizationof Nannochloropsis oculata in response to CO2 aeration Bioresour Technol 2009 100 833ndash838 [CrossRef][PubMed]
59 Huerlimann R de Nys R Heimann K Growth lipid content productivity and fatty acid composition oftropical microalgae for scale-up production Biotechnol Bioeng 2010 107 245ndash257 [CrossRef] [PubMed]
60 Liang Y Sarkany N Cui Y Biomass and lipid productivities of Chlorella vulgaris under autotrophicheterotrophic and mixotrophic growth conditions Biotechnol Lett 2009 31 1043ndash1049 [CrossRef] [PubMed]
61 Hempel N Petrick I Behrendt F Biomass productivity and productivity of fatty acids and amino acidsof microalgae strains as key characteristics of suitability for biodiesel production J Appl Phycol 2012 241407ndash1418 [CrossRef] [PubMed]
62 Yeesang C Cheirsilp B Effect of nitrogen salt and iron content in the growth medium and light intensityon lipid production by microalgae isolated from freshwater sources in Thailand Bioresour Technol 2011 1023034ndash3040 [CrossRef] [PubMed]
63 Msanne J Xu D Konda AR Casas-Mollano JA Awada T Cahoon EB Cerutti H Metabolic andgene expression changes triggered by nitrogen deprivation in the photoautotrophically grown microalgaeChlamydomonas reinhardtii and Coccomyxa sp C-169 Phytochemistry 2012 75 50ndash59 [CrossRef] [PubMed]
64 Lin Q Lin J Effects of nitrogen source and concentration on biomass and oil production of aScenedesmus rubescens like microalga Bioresour Technol 2011 102 1615ndash1621 [CrossRef] [PubMed]
65 Sheehan J Dunahay T Benemann J Roessler P A Look Back at the US Department of Energyrsquos AquaticSpecies ProgrammdashBiodiesel from Algae US Department of Energyrsquos Office of Fuels Development Golden COUSA 1998 p 328
66 Vaulot D Olson RJ Merkel S Chisholm SW Cell-cycle response to nutrient starvation in twophytoplankton species Thalassiosira weissflogii and Hymenomonas carterae Mar Biol 1987 95 625ndash630[CrossRef]
67 Olson RJ Chisholm SW Effects of light and nitrogen limitation on the cell cycle of the dinoflagellateAmphidinium carteri J Plankton Res 1986 8 785ndash793 [CrossRef]
68 Li Y Fei X Deng X Novel molecular insights into nitrogen starvation-induced triacylglycerolsaccumulation revealed by differential gene expression analysis in green algae Micractinium pusillumBiomass Bioenergy 2012 42 199ndash211 [CrossRef]
69 Abdelaziz AEM Leite GB Belhaj MA Hallenbeck PC Screening microalgae native to Quebec forwastewater treatment and biodiesel production Bioresour Technol 2014 157 140ndash148 [CrossRef] [PubMed]
70 Valdez-Ojeda R Gonzaacutelez-Muntildeoz M Us-Vaacutezquez R Narvaacuteez-Zapata J Chavarria-Hernandez JCLoacutepez-Adriaacuten S Barahona-Peacuterez F Toledano-Thompson T Garduntildeo-Soloacuterzano G Escobedo-GraciaMedrano RM Characterization of five fresh water microalgae with potential for biodiesel productionAlgal Res 2015 7 33ndash44 [CrossRef]
71 Shrivastav A Mishra SK Suh WI Farooq W Moon M Kim T-H Kumar K Choi G-G Park MSYang J-W Characterization of newly isolated oleaginous microalga Monoraphidium sp for lipid productionunder different conditions Algal Res 2015 12 289ndash294 [CrossRef]
72 Yen H-W Hu I-C Chen C-Y Ho S-H Lee D-J Chang J-S Microalgae-based biorefinerymdashFrombiofuels to natural products Bioresour Technol 2013 135 166ndash174 [CrossRef] [PubMed]
Energies 2017 10 224 16 of 16
73 Li J Liu Y Cheng JJ Mos M Daroch M Biological potential of microalgae in China for biorefinery-basedproduction of biofuels and high value compounds New Biotechnol 2015 32 588ndash596 [CrossRef] [PubMed]
74 El-Sheekh M Abomohra AE-F Hanelt D Optimization of biomass and fatty acid productivity ofScenedesmus obliquus as a promising microalga for biodiesel production World J Microbiol Biotechnol 201329 915ndash922 [CrossRef] [PubMed]
75 Singh P Guldhe A Kumari S Rawat I Bux F Investigation of combined effect of nitrogen phosphorusand iron on lipid productivity of microalgae Ankistrodesmus falcatus KJ671624 using response surfacemethodology Biochem Eng J 2015 94 22ndash29 [CrossRef]
76 Kilham S Kreeger D Goulden C Lynn S Effects of nutrient limitation on biochemical constituents ofAnkistrodesmus falcatus Freshw Biol 1997 38 591ndash596 [CrossRef]
77 Breuer G Lamers PP Martens DE Draaisma RB Wijffels RH The impact of nitrogen starvation on thedynamics of triacylglycerol accumulation in nine microalgae strains Bioresour Technol 2012 124 217ndash226[CrossRef] [PubMed]
78 Breuer G de Jaeger L Artus VG Martens DE Springer J Draaisma RB Eggink G Wijffels RHLamers PP Superior triacylglycerol (TAG) accumulation in starchless mutants of Scenedesmus obliquus (II)evaluation of TAG yield and productivity in controlled photobioreactors Biotechnol Biofuels 2014 7 70[CrossRef] [PubMed]
79 Nigam S Prakash-Rai M Sharma R Effect of Nitrogen on Growth and Lipid Content ofChlorella pyrenoidosa Am J Biochem Biotechnol 2011 7 126ndash131 [CrossRef]
80 Piligaev AV Sorokina KN Bryanskaya AV Peltek SE Kolchanov NA Parmon VN Isolation ofprospective microalgal strains with high saturated fatty acid content for biofuel production Algal Res 201512 368ndash376 [CrossRef]
81 Weiss SB Kennedy EP The enzymatic synthesis of triglycerides J Am Chem Soc 1956 78 3550[CrossRef]
82 Miller R Wu G Deshpande RR Vieler A Gaumlrtner K Li X Moellering ER Zaumluner S Cornish AJLiu B et al Changes in Transcript Abundance in Chlamydomonas reinhardtii following Nitrogen DeprivationPredict Diversion of Metabolism Plant Physiol 2010 154 1737ndash1752 [CrossRef] [PubMed]
83 Rismani-Yazdi H Haznedaroglu BZ Bibby K Peccia J Transcriptome sequencing and annotation of themicroalgae Dunaliella tertiolecta Pathway description and gene discovery for production of next-generationbiofuels BMC Genom 2011 12 148 [CrossRef] [PubMed]
84 Ramos MJ Fernaacutendez CM Casas A Rodriacuteguez L Peacuterez A Influence of fatty acid composition of rawmaterials on biodiesel properties Bioresour Technol 2009 100 261ndash268 [CrossRef] [PubMed]
85 Knothe G Dependence of biodiesel fuel properties on the structure of fatty acid alkyl estersFuel Process Technol 2005 86 1059ndash1070 [CrossRef]
86 Knothe G ldquoDesignerrdquo Biodiesel Optimizing Fatty Ester Composition to Improve Fuel PropertiesEnergy Fuels 2008 22 1358ndash1364 [CrossRef]
87 Smith PC Ngothai Y Dzuy Nguyen Q OrsquoNeill BK Improving the low-temperature properties ofbiodiesel Methods and consequences Renew Energy 2010 35 1145ndash1151 [CrossRef]
88 Dunn RO Effect of antioxidants on the oxidative stability of methyl soyate (biodiesel) Fuel Process Technol2005 86 1071ndash1085 [CrossRef]
89 Knothe G Improving biodiesel fuel properties by modifying fatty ester composition Energy Environ Sci2009 2 759ndash766 [CrossRef]
copy 2017 by the authors licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)

Energies 2017 10 224 6 of 16
3 Results and Discussion
31 Isolation and Identification of the Microalgae Strains
Bioprospecting of tropical ecosystems can provide exceptional native microalgae strains withthe potential for the production of biodiesel nutraceuticals bioactive chemicals and other highvalue products [36ndash38] Furthermore native microalgae strains can have potential applications forefficient biological sequestration of CO2 wastewater treatment and other environmentally-friendlyapplications [38ndash40] Consequently isolation is a fundamental process to obtain pure cultures and isthe first phase towards the screening and selection of microalgae strains with the potential for biodieselproduction and the aforementioned applications
According to these demands in this research samples were collected from three different typesof water bodies the Amazon River (white-water type) the Itaya and Nanay Rivers both of theblack-water type The white-water rivers originate from the Andean catchments bringing high amountsof nutrient-rich sediments In contrast the black-water rivers drain geochemically-poor watershedsand are characterized by their high content in humic acids low pH and poor nutrient status [41]A total of 14 microalgal strains were isolated using the standard cell washing technique and basedon distinguishable morphological characters under light microscopic examination these strains werepreliminary ascribed to the genera Acutodesmus (Hegewald) Tsarenko 2001 Ankistrodesmus Corda1838 Chlorella Beyerinck Beijerinck 1890 Desmodesmus (Chodat) SS An T Friedl and E Hegewald1999 and Tetradesmus GM Smith 1913 (Table 1) These genera are green algae that belong todivision Chlorophyta class Chlorophyceae and orders Chlorellales and Sphaeropleales Althoughmorphological analysis is frequently used to identify microalgae it is inaccurate and very difficultfor the identification at the species level because the relationship between diagnostic morphologyand biological species boundaries are largely unknown in many micro-eukaryotes [42] Furthermoreaccording to Yu et al [31] the microalgae morphology for the same strain varies in relation to ageand culture conditions Consequently in microalgae the classic taxonomy is supported by moleculartools [42] For example the ITS2 rDNA region is thought to be an excellent marker for molecularphylogenetics studies in lower taxonomic levels due to being highly divergent and fast-evolvingwhich can discriminate among closely-related organisms which otherwise display almost identicalsequences [4344] For this reason the molecular identification based in phylogenetic analysis of ITS2rDNA gene sequences is commonly used to identify andor delineate species [204546]
Table 1 Genera GenBank accession number of ITS2 sequences biomass productivity lipid productivityand lipid content of isolated microalgae strains
Source ofCollection Strain GeneraSpecies
GenBankAccessionNumber
BiomassProductivity
(mgmiddotLminus1middotdminus1)
LipidProductivity
(mgmiddotLminus1middotdminus1)
LipidContent( dw)
Amazon River(0341prime06prime prime S7314prime89prime prime W)
AMA001 Ankistrodesmus sp KP8784961 122 plusmn 03 37 plusmn 01 307 plusmn 15AMA002 Ankistrodesmus sp KP8784971 36 plusmn 04 10 plusmn 02 277 plusmn 21AMA003 Chlorella lewinii KP8785011 223 plusmn 07 51 plusmn 05 227 plusmn 15AMA004 Acutodesmus abliquus KP8785051 316 plusmn 04 64 plusmn 07 203 plusmn 21
Itaya River(0343prime14prime prime S
7314prime178prime prime W)
ITA001 Ankistrodesmus sp KP8784991 59 plusmn 02 40 plusmn 03 437 plusmn 17ITA002 Ankistrodesmus sp KP8785001 136 plusmn 04 57 plusmn 05 306 plusmn 20ITA003 Chlorella lewinii KP8785031 129 plusmn 03 40 plusmn 05 224 plusmn 27ITA004 Chlorella lewinii KP8785041 221 plusmn 07 31 plusmn 04 215 plusmn 23ITA005 Scenedesmus regularis KP8785081 93 plusmn 04 56 plusmn 04 242 plusmn 14ITA006 Scenedesmus dimorphus KP8785091 186 plusmn 06 32 plusmn 03 260 plusmn 17ITA007 Acutodesmus abliquus KP8785101 179 plusmn 04 22 plusmn 03 156 plusmn 19
Nanay River(0342prime02prime prime S7315prime32prime prime W)
NAN001 Ankistrodesmus sp KP8784981 143 plusmn 06 28 plusmn 02 467 plusmn 15NAN002 Desmodesmus tropicus KP8785061 122 plusmn 04 23 plusmn 02 175 plusmn 11NAN003 Tetradesmus sp KP8785071 139 plusmn 04 30 plusmn 03 135 plusmn 12
Energies 2017 10 224 7 of 16
To realize phylogenetic analysis genomic DNA of each isolate strain was purified and used forPCR amplification of ITS2 rDNA The universal primers used to amplify the ITS2 rDNA successfullyamplified the desired DNA fragments (~300 bp) from all microalgal strains (Figure S2) Furtherthese amplicons were sequenced published in the NCBI databases and the corresponding GenBankaccession numbers are presented in Table 1 Figure 1 shows a neighbor-joining phylogenetic treedepicting the relatedness of the native microalgae strain isolates with selected sequences from thepublic GenBank database This phylogram contained three clusters each of which included differentmicroalgae genera For example the first cluster consisted of microalgae of the genera AcutodesmusScenedesmus Tetradesmus and Desmodesmus and six isolated strains (ITA007 AMA004 ITA006 NAN003ITA005 and NAN002) The second cluster consisted of a microalgae of the genus Ankistrodesmus andthe isolated strains AMA001 ITA001 ITA002 NAN001 and AMA002 The third one consisted of amicroalgae of the genus Chlorella and three isolated strains (ITA004 AMA003 and ITA003) Overall theidentification of native microalgae strains presented a good correspondence between morphologicalcharacters and sequence-based phylogenetic analysis Microalgae species of these genera were alsoisolated from different countries of the world such as Brazil [47] Argentina [46] United States [48]India [20] Egypt [49] Taiwan [50] South Korea [45] and China [51] Some of these microalgae generasuch as Ankistrodesmus Chlorella Desmodesmus and Scenedesmus were used in large-scale biodieselproduction using photobioreactors and raceway pond systems [5253]
Energies 2017 10 224 7 of 16
public GenBank database This phylogram contained three clusters each of which included different microalgae genera For example the first cluster consisted of microalgae of the genera Acutodesmus Scenedesmus Tetradesmus and Desmodesmus and six isolated strains (ITA007 AMA004 ITA006 NAN003 ITA005 and NAN002) The second cluster consisted of a microalgae of the genus Ankistrodesmus and the isolated strains AMA001 ITA001 ITA002 NAN001 and AMA002 The third one consisted of a microalgae of the genus Chlorella and three isolated strains (ITA004 AMA003 and ITA003) Overall the identification of native microalgae strains presented a good correspondence between morphological characters and sequence-based phylogenetic analysis Microalgae species of these genera were also isolated from different countries of the world such as Brazil [47] Argentina [46] United States [48] India [20] Egypt [49] Taiwan [50] South Korea [45] and China [51] Some of these microalgae genera such as Ankistrodesmus Chlorella Desmodesmus and Scenedesmus were used in large-scale biodiesel production using photobioreactors and raceway pond systems [5253]
Figure 1 Unrooted phylogenetic tree of isolated microalgae strains using the ITS2 sequences The tree was constructed by the neighbor-joining method and Kimurarsquos two-parameter algorithm in MEGA 60 software based on the multiple sequence alignment by Clustal Omega Bootstrap values of 1000 replicates () are shown at the branches
32 Microalgae Cultivation Biomass and Total Lipids Measurements
The isolated microalgae strains exhibited remarkable differences in productivity parameters and lipid content among individual strains in the stationary growth phase (Table 1) With regard to the biomass productivity the 14 strains showed statistically-significant differences (F = 71911 df = 13 p lt 0001) and ranged from 36 plusmn 04 mgmiddotLminus1middotdminus1 (Ankistrodesmus sp strain AMA002) to 316 plusmn 04 mgmiddotLminus1middotdminus1 (Acutodesmus obliquus strain AMA004) Similar biomass productivities were reported for other marine microalgae species [54] However other investigations show major biomass productivities (ranging
Figure 1 Unrooted phylogenetic tree of isolated microalgae strains using the ITS2 sequences The treewas constructed by the neighbor-joining method and Kimurarsquos two-parameter algorithm in MEGA60 software based on the multiple sequence alignment by Clustal Omega Bootstrap values of1000 replicates () are shown at the branches
Energies 2017 10 224 8 of 16
32 Microalgae Cultivation Biomass and Total Lipids Measurements
The isolated microalgae strains exhibited remarkable differences in productivity parametersand lipid content among individual strains in the stationary growth phase (Table 1) With regardto the biomass productivity the 14 strains showed statistically-significant differences (F = 71911df = 13 p lt 0001) and ranged from 36 plusmn 04 mgmiddotLminus1middotdminus1 (Ankistrodesmus sp strain AMA002) to316 plusmn 04 mgmiddotLminus1middotdminus1 (Acutodesmus obliquus strain AMA004) Similar biomass productivities werereported for other marine microalgae species [54] However other investigations show major biomassproductivities (ranging from 30 to 120 mgmiddotLminus1middotdminus1) in the same and other microalgae genera [455556]These discrepancies may be attributed to differences in the culture conditions in the culture mediacomposition and in the microalgae genotypes Lipid productivity also exhibited statistically-significantdifferences (F = 4774 df = 13 p lt 0001) and ranged from 10plusmn 04 mgmiddotLminus1middotdminus1 to 64 plusmn 07 mgmiddotLminus1middotdminus1
with strains AMA002 and AMA004 respectively These lipid productivities were low in comparison toother reports which reported values from 190 to 433 mgmiddotLminus1middotdminus1 [4555] Furthermore lipid contentas a percentage of the microalgal biomass dry weight showed statistically-significant differences(F = 8509 df = 13 p lt 0001) and ranged from 135 plusmn 12 (Tetradesmus sp strain NAN003) to467 plusmn 15 (Ankistrodesmus sp strain NAN001) These percentage values in total lipid contentare similar to several microalgae species of these genera isolated and characterized by otherresearchers worldwide [20454648ndash5057] Furthermore it is important to emphasize that lipid contentis dependent on the different growth phases Commonly in several microalgae taxa the lipid contentincreases gradually from logarithmic to stationary growth phases [58ndash60] For example Kaur et al [20]reported that the total lipid content of Scenedesmus sp DRLMA9 demonstrated values of 207 inthe late exponential phase and 397 in the stationary phase and Desmodesmus elegans DRLMA13displayed values of 103 in the late exponential phase and 169 in the stationary phase LikewiseChiu et al [61] found that the total lipid content of Nannochloropsis oculata was 308 in the logarithmicphase 397 in the early stationary phase and 504 in the stationary phase These changes in lipidcontent with culture age have been associated with nitrate depletion [61] which causes a channeling ofphotosynthetic ATP to carbohydrate and lipid biosynthesis [60] Finally Pearson correlation analysisof biomass productivity and lipid content was inversely related (r = minus037 p lt 001) which is similarto the results obtained by Liang et al [62] and Rodolfi et al [52] In contrast Li et al [51] andHempel et al [63] found no relationship between these parameters The negative correlation obtainedis likely due to the high metabolic cost of lipid biosynthesis [62]
33 Strains Culture under Nitrogen-Sufficient and Nitrogen-Deficient Medium
The microalgae species Acutodesmus obliquus (KP8785051) Ankistrodesmus sp (KP8785001)and Chlorella lewinii (KP8785011) were selected and further characterized because they displayedthe highest biomass and lipid productivities (Table 1) These microalgae showed noticeablevariations in the growth profile when cultured in Chu medium No 10 with nitrogen (+N) orwithout a nitrogen source (minusN) In +N medium all strains showed exponential growth butChlorella lewinii showed the highest average specific growth rates (micro = 0458 plusmn 0098 dminus1) followed byAcutodesmus obliquus (micro = 0399 plusmn 0112 dminus1) and the lowest value was produced by Ankistrodesmus sp(micro = 0265 plusmn 0013 dminus1) In contrast all cultures displayed a lag phase pattern in their growthprofile under minusN medium (Figure 2) These findings corroborate the results obtained by otherresearchers who demonstrate that the specific growth rate is directly proportional to the initialnitrate concentration [46596465] because the reduction or cessation of microalgae cell divisionis due to nitrogen limitation [66] which is a key nutrient for protein and nucleic acids de novobiosynthesis consequently causing an arrest in the early part of the microalgae cell cycle [6768]In accordance with these reports Li et al [69] showed that nitrogen deficiency induced in the greenmicroalgae Micractinium pusillum a downregulation in the expression of genes involved in cell growthand cell cycle regulation Additionally differences in growth profile and specific growth rates aredependent on several factors such as microalgae genotypes the culture medium composition pH
Energies 2017 10 224 9 of 16
nutrients concentration the cell culture phase (ie log phase stationary phase) and environmentalfactors [4549545770ndash72]Energies 2017 10 224 9 of 16
Figure 2 Growth profiles of three microalgal strains cultivated under +N (filled boxes) and minusN (empty boxes) Chu medium No 10 The culture was incubated in a controlled culture room at 25 plusmn 1 degC with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 150 rpm Experiments were carried out in triplicate
34 Total Protein Total Lipid and Total Carbohydrate Content
Biochemical parameters of the microalgae species such as total protein total lipid and total carbohydrate content exhibited differences and changes under both culture conditions (Figure 3) For example Chlorella lewinii showed the highest protein content Ankistrodesmus sp had the highest total lipid content but Acutodesmus obliquus presented the highest levels of total carbohydrate Therefore these strains can be valuable stocks for the industry because total protein and total carbohydrate are potentially byproducts of microalgal biomass processing [7374] Additionally the three strains cultured under minusN conditions showed a significant increase in total lipid content (from 237 in Acutodesmus obliquus to 667 in Ankistrodesmus sp) and total carbohydrate content (from 119 in Acutodesmus obliquus to 963 in Chlorella lewinii) but showed a decreased in total protein levels (from 242 in Acutodesmus obliquus to 545 in Chlorella lewinii) in comparison to +N cultures (Figure 3) These results are in accordance with well-documented results obtained by other researchers with several microalgae species of the genera Acutodesmus Ankistrodesmus Botryococcus Chlamydomonas Chlorella Desmodesmus Dunaliella Haematococcus Neochloris Phaeodactylum Pseudokirchneriella and Scenedesmus [465058596475ndash80] Nevertheless the accumulation of total lipid induced by nitrogen deficiency is not correlated with lipid productivity but rather is due to a decline and stopping of the microalgae specific growth rate [75] The changes in biomolecule contents suggest redirection of microalgae metabolism during nitrogen starvation such as a decrease in biosynthesisincrease in protein degradation and an increase in the biosynthesis of storage biomolecules such as triacylglycerols and starch The increase in total lipid content could be partially explained for a boost in transcript levels of genes encoding enzymes of the lipid biosynthesis pathways According to this assumption recent investigations showed that genes encoding the enzymes that catalyze the last step in the Kennedy pathway of triacylglycerol biosynthesis [81] such as glycerol-3-phosphate acyltransferase and acyl-glycerol-3-phosphate acyltransferase are upregulated in response to nitrogen starvation [648283] Furthermore the increase in total carbohydrate content can be explained by changes in the expression levels of genes involved in carbohydrate metabolism For example when the oleaginous microalgae Neochloris oleoabundans is cultured under nitrogen starvation the catabolic phase of starch specifically α-amylase which hydrolyzes starch to glucose is strongly repressed [83]
Figure 2 Growth profiles of three microalgal strains cultivated under +N (filled boxes) and minusN(empty boxes) Chu medium No 10 The culture was incubated in a controlled culture room at25 plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent lightand continuous agitation at 150 rpm Experiments were carried out in triplicate
34 Total Protein Total Lipid and Total Carbohydrate Content
Biochemical parameters of the microalgae species such as total protein total lipid and totalcarbohydrate content exhibited differences and changes under both culture conditions (Figure 3)For example Chlorella lewinii showed the highest protein content Ankistrodesmus sp had the highesttotal lipid content but Acutodesmus obliquus presented the highest levels of total carbohydrateTherefore these strains can be valuable stocks for the industry because total protein and totalcarbohydrate are potentially byproducts of microalgal biomass processing [7374] Additionallythe three strains cultured under minusN conditions showed a significant increase in total lipid content(from 237 in Acutodesmus obliquus to 667 in Ankistrodesmus sp) and total carbohydrate content(from 119 in Acutodesmus obliquus to 963 in Chlorella lewinii) but showed a decreased in totalprotein levels (from 242 in Acutodesmus obliquus to 545 in Chlorella lewinii) in comparison to +Ncultures (Figure 3) These results are in accordance with well-documented results obtained by otherresearchers with several microalgae species of the genera Acutodesmus Ankistrodesmus BotryococcusChlamydomonas Chlorella Desmodesmus Dunaliella Haematococcus Neochloris PhaeodactylumPseudokirchneriella and Scenedesmus [465058596475ndash80] Nevertheless the accumulation of totallipid induced by nitrogen deficiency is not correlated with lipid productivity but rather is due toa decline and stopping of the microalgae specific growth rate [75] The changes in biomoleculecontents suggest redirection of microalgae metabolism during nitrogen starvation such as a decreasein biosynthesisincrease in protein degradation and an increase in the biosynthesis of storagebiomolecules such as triacylglycerols and starch The increase in total lipid content could bepartially explained for a boost in transcript levels of genes encoding enzymes of the lipid biosynthesispathways According to this assumption recent investigations showed that genes encoding theenzymes that catalyze the last step in the Kennedy pathway of triacylglycerol biosynthesis [81] such asglycerol-3-phosphate acyltransferase and acyl-glycerol-3-phosphate acyltransferase are upregulated
Energies 2017 10 224 10 of 16
in response to nitrogen starvation [648283] Furthermore the increase in total carbohydrate contentcan be explained by changes in the expression levels of genes involved in carbohydrate metabolismFor example when the oleaginous microalgae Neochloris oleoabundans is cultured under nitrogenstarvation the catabolic phase of starch specifically α-amylase which hydrolyzes starch to glucoseis strongly repressed [83]Energies 2017 10 224 10 of 16
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strains cultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled culture room at 25 plusmn 1 degC with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized with respect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that each microalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majority of fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised 7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii the majority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9) acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectively Finally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic (182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in +N and minusN conditions respectively Our results agree with multiple previously-published reports in that fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent in many green microalgae species [454648ndash5055] It is notable that under nitrogen starvation culture all microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 in Ankistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid (181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findings support the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content of SFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Nevertheless there are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed a decrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increased between 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively However in the same culture condition the three microalgae species showed a decrease in polyunsaturated fatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmus spp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgae strains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFA which improves the quality properties of biodiesel such as an increase
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strainscultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled cultureroom at 25plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescentlight and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized withrespect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that eachmicroalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majorityof fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii themajority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9)acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectivelyFinally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic(182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in+N and minusN conditions respectively Our results agree with multiple previously-published reports inthat fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent inmany green microalgae species [454648ndash5055] It is notable that under nitrogen starvation cultureall microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 inAnkistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid(181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findingssupport the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content ofSFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other
Energies 2017 10 224 11 of 16
researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Neverthelessthere are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed adecrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increasedbetween 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively Howeverin the same culture condition the three microalgae species showed a decrease in polyunsaturatedfatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmusspp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgaestrains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFAwhich improves the quality properties of biodiesel such as an increase in cetane number [84ndash86]Considering the SFA and unsaturated fatty acids (UFA) components approximately 20ndash31 are SFAand 69ndash80 are UFA Hence the SFAUFA ratio in the microalgae strains evaluated ranges between025 and 045 which is similar to the reports of several microalgae species isolated in different latitudesworldwide [203148497072]
Table 2 Effect of nitrogen deficiency on fatty acid profiling in three native microalgal species from thePeruvian Amazon
Fatty AcidsAcutodesmus obliquus Ankistrodesmus sp Chlorella lewinii
+N minusN +N minusN +N minusN
140 - - 052 plusmn 002 088 plusmn 002 - 058 plusmn 001160 2724 plusmn 007 2867 plusmn 017 1868 plusmn 019 2261 plusmn 006 1964 plusmn 021 2123 plusmn 010180 184 plusmn 005 255 plusmn 051 101 plusmn 002 181 plusmn 002 371 plusmn 026 153 plusmn 015
161n-7 023 plusmn 000 025 plusmn 001 071 plusmn 003 055 plusmn 001 085 plusmn 043 038 plusmn 000181n-7 131 plusmn 002 131 plusmn 000 141 plusmn 005 094 plusmn 003 146 plusmn 006 123 plusmn 002181n-9 3213 plusmn 021 3608 plusmn 021 3699 plusmn 004 4047 plusmn 038 1371 plusmn 015 2326 plusmn 041182n-6 1732 plusmn 004 1651 plusmn 016 747 plusmn 018 656 plusmn 007 2034 plusmn 039 1957 plusmn 001183n-3 1130 plusmn 017 854 plusmn 031 1486 plusmn 004 1173 plusmn 012 1815 plusmn 022 1371 plusmn 016183n-6 101 plusmn 001 078 plusmn 002 055 plusmn 000 054 plusmn 002 124 plusmn 006 100 plusmn 020184n-3 250 plusmn 005 177 plusmn 008 631 plusmn 002 601 plusmn 005 491 plusmn 025 403 plusmn 020204n-6 - - - 084 plusmn 008 - -225n-3 - - - 062 plusmn 002 - -
Unidentified 512 plusmn 003 355 plusmn 006 1149 plusmn 004 645 plusmn 003 1599 plusmn 034 1348 plusmn 006SFA 2907 3122 2022 2531 2335 2334UFA 7093 6878 7978 7469 7665 7666
MUFA 3367 3764 3911 4195 1602 2487PUFA 3213 2759 2918 2629 4464 3831SFA +MUFA 6274 6886 5933 6726 3936 4821
It is important to highlight that the fatty acid profile determination is an essential step inthe process of the characterization of microalgae strains because it ultimately affects the qualityof the biodiesel product [8485] The quality properties of biodiesel such as high cetane numberoxidative stability and cold-flow are dependent on the hydrocarbonated chain length of saturated andunsaturated fatty acids [8587] For example biodiesel composed of a mixture of a high percentage ofSFA and MUFA is preferred for increased energy yield superior oxidative stability and higher cetanenumbers but is prone to solidify at low temperature In contrast biodiesel with high levels of PUFAhas decreased energy yield low cetane numbers and excellent cold-flow properties but insufficientoxidative stability [548588] Therefore to solve these performance problems five approaches havebeen developed such as changing the fatty acids profile by physical means genetic modification ofthe feedstock or the use of alternative feedstocks with different fatty acids profiles [89]
4 Conclusions
Fourteen microalgae were isolated from three river basins from the Peruvian Amazon and wereidentified by morphological features and molecular phylogeny approaches These strains were initially
Energies 2017 10 224 12 of 16
characterized in the stationary growth phase and exhibited significant heterogeneity in key parameterssuch as biomass productivity (36ndash316 mgmiddotLminus1middotdminus1) lipid productivity (10ndash64 mgmiddotLminus1middotdminus1) andtotal lipid content (135ndash467) Furthermore three selected strains based on elevated biomass andlipid productivities were cultured under nitrogen-sufficient and nitrogen-deficient medium to boostlipid accumulation Under these culture conditions all strains showed noticeable decreases in totalprotein content and increases in total lipid and total carbohydrate contents In addition these strainspresented modifications in the fatty acids profiles with improvement in biodiesel properties due to apredominance of SFA and MUFA which corresponds to an increase in cetane number The results ofthis research suggest that the isolated native microalgae from the Peruvian Amazon could be suitablerenewable feedstocks for the production of both biodiesel and bioethanol which could contribute tosustainable development at the local regional and national levels as a viable alternative to petroleumexploitation However it will be necessary to develop open closed or hybrid systems for large-scalebiomass production of selected native microalgae strains andor de novo microalgae strains designedto produce biofuels and with the abilities to synthesize useful bioactive compounds to be efficientlyexploited using biorefinery approaches
Supplementary MaterialsThe following are available online at wwwmdpicom1996-1073102224s1 Figure S1 Results of
electrophoretic and spectrophotometric analysis of the genomic DNA isolated from the fourteen microalgaeFigure S2 ITS2-rDNA structure (A) and amplicons obtained (B) from the microalgae isolated Figure S3Microphotographic images of the three microalgae strains analyzed under light and fluorescence stained withNile Red Figure S4 Spectrophotometric scanning for the determination of the maximum absorption peak foreach microalgae strain
Acknowledgments This research was supported by the Peruvian funding agencies Consejo Nacional de CienciaTecnologiacutea e Innovacioacuten Tecnoloacutegica (CONCYTEC) with Grant Contract No 364-CONCYTEC-OAJ and Fondospara la Innovacioacuten Ciencia y Tecnologiacutea (FINCyT) with Grant Contract No 383-PNICP-PIBA-2014
Author Contributions Marianela Cobos and Juan C Castro obtained funds for the research participated inthe study design coordinated activities and participated in the preparation of the manuscript Jae D Paredesparticipated in sample collection microalgae isolation and helped to draft the manuscript J Dylan Maddoxperformed the bioinformatics analysis and participated in the preparation of the manuscript Gabriel Vargas-Aranaand Leenin Flores participated in biochemical analysis and helped to draft the manuscript Carla P Aguilarparticipated in the study design supervised biochemical analysis and helped to draft the manuscriptJorge L Marapara participated in molecular analysis and helped to draft the manuscript
Conflicts of Interest The authors declare no conflict of interest
References
1 BP Global BP Statisical Review of World Energy 2014 Available online httpwwwbpcomcontentdambppdfEnergy-economicsstatistical-review-2014BP-statistical-review-of-world-energy-2014-full-reportpdf (accessed on 20 June 2014)
2 Pachauri RK Reisinger A Cambio Climaacutetico 2007 Informe de Siacutentesis Contribucioacuten de los Grupos de Trabajo III y III al Cuarto Informe de Evaluacioacuten del Grupo Intergubernamental de Expertos Sobre el Cambio Climaacutetico IPCCGeneva Switzerland 2007 p 104
3 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Ley de Promocioacuten del Mercado deBiocombustibles Available online httpwww2congresogobpesicrcendocbibcon4_uibdnsf9E95620CC059138105257C9E005AB2F9$FILE28054pdf (accessed on 12 February 2017)
4 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Reglamento de la Ley de Promocioacuten del Mercadode Biocombustibles Available online httpwww2osinerggobpeMarcoLegaldocrevDS-013-2005-EM-CONCORDADOpdf (accessed on 12 February 2017)
5 Areacutevalo LF Nalvarte J Torres J Ramiacuterez Y Impactos Socio-Econoacutemicos de la Produccioacuten de Biocombustiblesen la Amazoniacutea Peruana Primera SNV-WWF Lima Peruacute 2009
6 Sharma KK Schuhmann H Schenk PM High Lipid Induction in Microalgae for Biodiesel ProductionEnergies 2012 5 1532ndash1553 [CrossRef]
Energies 2017 10 224 13 of 16
7 Schenk PM Thomas-Hall SR Stephens E Marx UC Mussgnug JH Posten C Kruse O Hankamer BSecond Generation Biofuels High-Efficiency Microalgae for Biodiesel Production BioEnergy Res 2008 120ndash43 [CrossRef]
8 Foley JA Asner GP Costa MH Coe MT de Fries R Gibbs HK Howard EA Olson S Patz JRamankutty N et al Amazonia revealed Forest degradation and loss of ecosystem goods and services inthe Amazon Basin Front Ecol Environ 2007 5 25ndash32 [CrossRef]
9 Duong VT Li Y Nowak E Schenk PM Microalgae Isolation and Selection for Prospective BiodieselProduction Energies 2012 5 1835ndash1849 [CrossRef]
10 Georgianna DR Mayfield SP Exploiting diversity and synthetic biology for the production of algalbiofuels Nature 2012 488 329ndash335 [CrossRef] [PubMed]
11 Wijffels RH Barbosa MJ An Outlook on Microalgal Biofuels Science 2010 329 796ndash799 [CrossRef][PubMed]
12 Richmond A Handbook of Microalgal Culture Biotechnology and Applied Phycology Blackwell Sci Ltd HudsonNJ USA 2004
13 Menetrez MY An overview of algae biofuel production and potential environmental impactEnviron Sci Technol 2012 46 7073ndash7085 [CrossRef] [PubMed]
14 Khoo HH Sharratt PN Das P Balasubramanian RK Naraharisetti PK Shaik S Life cycle energy andCO2 analysis of microalgae-to-biodiesel Preliminary results and comparisons Bioresour Technol 2011 1025800ndash5807 [CrossRef] [PubMed]
15 Chu SP The Influence of the Mineral Composition of the Medium on the Growth of Planktonic AlgaePart I Methods and Culture Media J Ecol 1942 30 284ndash325 [CrossRef]
16 Carlos EMB Mariatildengela M Gecircneros de Algas de Aacuteguas Continentais do BrasilmdashChave Para Identificaccedilatildeo eDescriccedilotildees 2nd ed Rima Sao Carlos Brasil 2006
17 Greenspan P Mayer EP Fowler SD Nile red A selective fluorescent stain for intracellular lipid dropletsJ Cell Biol 1985 100 965ndash973 [CrossRef] [PubMed]
18 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissuePhytochem Bull 1987 19 11ndash15
19 Sambrook J Fritsch EF Maniatis T Molecular Cloning A Laboratory Manual 2nd ed Cold Spring HarborLaboratory Press New York NY USA 1989
20 Kaur S Sarkar M Srivastava RB Gogoi HK Kalita MC Fatty acid profiling and molecularcharacterization of some freshwater microalgae from India with potential for biodiesel productionNew Biotechnol 2012 29 332ndash344 [CrossRef] [PubMed]
21 Benson DA Cavanaugh M Clark K Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW GenBankNucleic Acids Res 2013 41 D36ndashD42 [CrossRef] [PubMed]
22 NCBI Resource Coordinators Database Resources of the National Center for Biotechnology InformationNucleic Acids Res 2015 43 D6ndashD17
23 Ye J McGinnis S Madden TL BLAST Improvements for better sequence analysis Nucleic Acids Res 200634 W6ndashW9 [CrossRef] [PubMed]
24 BioEdit Sequence Alignment Editor for Windows 9598NTXPVista7 Available online httpwwwmbioncsuedubioeditbioedithtml (accessed on 2 December 2015)
25 Sievers F Higgins DG Clustal omega accurate alignment of very large numbers of sequencesMethods Mol Biol (Clifton NJ) 2014 1079 105ndash116
26 Saitou N Nei M The neighbor-joining method A new method for reconstructing phylogenetic treesMol Biol Evol 1987 4 406ndash425 [PubMed]
27 Kimura M A simple method for estimating evolutionary rates of base substitutions through comparativestudies of nucleotide sequences J Mol Evol 1980 16 111ndash120 [CrossRef] [PubMed]
28 Tamura K Stecher G Peterson D Filipski A Kumar S MEGA6 Molecular Evolutionary GeneticsAnalysis version 60 Mol Biol Evol 2013 30 2725ndash2729 [CrossRef] [PubMed]
29 Felsenstein J Confidence Limits on Phylogenies An Approach Using the Bootstrap Evolution 1985 39783ndash791 [CrossRef]
30 Bligh EG Dyer WJ A Rapid Method of Total Lipid Extraction and Purification Can J Biochem Physiol1959 37 911ndash917 [CrossRef] [PubMed]
Energies 2017 10 224 14 of 16
31 Yu X Zhao P He C Li J Tang X Zhou J Huang Z Isolation of a novel strain of Monoraphidium spand characterization of its potential application as biodiesel feedstock Bioresour Technol 2012 121 256ndash262[CrossRef] [PubMed]
32 Hartree EF Determination of protein A modification of the lowry method that gives a linear photometricresponse Anal Biochem 1972 48 422ndash427 [CrossRef]
33 DuBois M Gilles KA Hamilton JK Rebers PA Smith F Colorimetric Method for Determination ofSugars and Related Substances Anal Chem 1956 28 350ndash356 [CrossRef]
34 AOAC International AOAC Official Methods of Analysis of the Association of Official Analytical Chemists (AOAC)15th ed AOAC International Washington DC USA 1990 Volume 1
35 Ichihara K Fukubayashi Y Preparation of fatty acid methyl esters for gas-liquid chromatographyJ Lipid Res 2010 51 635ndash640 [CrossRef] [PubMed]
36 Brennan L Owende P Biofuels from microalgaemdashA review of technologies for production processingand extractions of biofuels and co-products Renew Sustain Energ Rev 2010 14 557ndash577 [CrossRef]
37 Raja R Hemaiswarya S Kumar NA Sridhar S Rengasamy R A perspective on the biotechnologicalpotential of microalgae Crit Rev Microbiol 2008 34 77ndash88 [CrossRef] [PubMed]
38 Mata TM Martins AA Caetano NS Microalgae for biodiesel production and other applicationsA review Renew Sustain Energy Rev 2010 14 217ndash232 [CrossRef]
39 De Wilt A Butkovskyi A Tuantet K Leal LH Fernandes TV Langenhoff A Zeeman GMicropollutant removal in an algal treatment system fed with source separated wastewater streamsJ Hazard Mater 2015 304 84ndash92 [CrossRef] [PubMed]
40 Li T Lin G Podola B Melkonian M Continuous removal of zinc from wastewater and mine dumpleachate by a microalgal biofilm PSBR J Hazard Mater 2015 297 112ndash118 [CrossRef] [PubMed]
41 Sioli H The Amazon and its main affluents Hydrography morphology of the river courses and river typesIn Monographiae Biologicae Sioli H Ed Springer Dordrecht The Netherlands 1984 pp 127ndash165
42 Moniz MBJ Kaczmarska I Barcoding of Diatoms Nuclear Encoded ITS Revisited Protist 2010 161 7ndash34[CrossRef] [PubMed]
43 Coleman AW Is there a molecular key to the level of ldquobiological speciesrdquo in eukaryotes A DNA guideMol Phylogenet Evol 2009 50 197ndash203 [CrossRef] [PubMed]
44 Mai JC Coleman AW The Internal Transcribed Spacer 2 Exhibits a Common Secondary Structure inGreen Algae and Flowering Plants J Mol Evol 1997 44 258ndash271 [CrossRef] [PubMed]
45 Abou-Shanab RAI Matter IA Kim S-N Oh Y-K Choi J Jeon B-H Characterization andidentification of lipid-producing microalgae species isolated from a freshwater lake Biomass Bioenergy2011 35 3079ndash3085 [CrossRef]
46 Do Nascimento M Ortiz-Marquez JCF Sanchez-Rizza L Echarte MM Curatti L Bioprospectingfor fast growing and biomass characterization of oleaginous microalgae from South-Eastern Buenos AiresArgentina Bioresour Technol 2012 125 283ndash290 [CrossRef] [PubMed]
47 Araujo GS Matos LJBL Gonccedilalves LRB Fernandes FAN Farias WRL Bioprospecting foroil producing microalgal strains Evaluation of oil and biomass production for ten microalgal strainsBioresour Technol 2011 102 5248ndash5250 [CrossRef] [PubMed]
48 Zhou W Li Y Min M Hu B Chen P Ruan R Local bioprospecting for high-lipid producing microalgalstrains to be grown on concentrated municipal wastewater for biofuel production Bioresour Technol 2011102 6909ndash6919 [CrossRef] [PubMed]
49 Mahmoud EA Farahat LA Abdel Aziz ZK Fatthallah NA Salah El Din RA Evaluation of thepotential for some isolated microalgae to produce biodiesel Egypt J Pet 2015 24 97ndash101 [CrossRef]
50 Ho S Lai Y-Y Chiang C-Y Chen C-NN Chang J-S Selection of elite microalgae for biodieselproduction in tropical conditions using a standardized platform Bioresour Technol 2013 147 135ndash142[CrossRef] [PubMed]
51 Li L Cui J Liu Q Ding Y Liu J Screening and phylogenetic analysis of lipid-rich microalgae Algal Res2015 11 381ndash386 [CrossRef]
52 Rodolfi L Chini Zittelli G Bassi N Padovani G Biondi N Bonini G Tredici MR Microalgae for oilStrain selection induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactorBiotechnol Bioeng 2009 102 100ndash112 [CrossRef] [PubMed]
Energies 2017 10 224 15 of 16
53 Li P Miao X Li R Zhong J In situ biodiesel production from fast-growing and high oil content Chlorellapyrenoidosa in rice straw hydrolysate J Biomed Biotechnol 2011 2011 141207 [CrossRef] [PubMed]
54 Doan TTY Sivaloganathan B Obbard JP Screening of marine microalgae for biodiesel feedstockBiomass Bioenergy 2011 35 2534ndash2544 [CrossRef]
55 Abou-Shanab RAI Hwang J-H Cho Y Min B Jeon B-H Characterization of microalgal speciesisolated from fresh water bodies as a potential source for biodiesel production Appl Energy 2011 883300ndash3306 [CrossRef]
56 Chaichalerm S Pokethitiyook P Yuan W Meetam M Sritong K Pugkaew W Kungvansaichol KKruatrachue M Damrongphol P Culture of microalgal strains isolated from natural habitats in Thailand invarious enriched media Appl Energy 2012 89 296ndash302 [CrossRef]
57 Zhang S Liu P Yang X Hao Z Zhang L Luo N Shi J Isolation and identification by 18S rDNAsequence of high lipid potential microalgal species for fuel production in Hainan Dao Biomass Bioenergy2014 66 197ndash203 [CrossRef]
58 Chiu S-Y Kao C-Y Tsai M-T Ong S-C Chen C-H Lin C-S Lipid accumulation and CO2 utilizationof Nannochloropsis oculata in response to CO2 aeration Bioresour Technol 2009 100 833ndash838 [CrossRef][PubMed]
59 Huerlimann R de Nys R Heimann K Growth lipid content productivity and fatty acid composition oftropical microalgae for scale-up production Biotechnol Bioeng 2010 107 245ndash257 [CrossRef] [PubMed]
60 Liang Y Sarkany N Cui Y Biomass and lipid productivities of Chlorella vulgaris under autotrophicheterotrophic and mixotrophic growth conditions Biotechnol Lett 2009 31 1043ndash1049 [CrossRef] [PubMed]
61 Hempel N Petrick I Behrendt F Biomass productivity and productivity of fatty acids and amino acidsof microalgae strains as key characteristics of suitability for biodiesel production J Appl Phycol 2012 241407ndash1418 [CrossRef] [PubMed]
62 Yeesang C Cheirsilp B Effect of nitrogen salt and iron content in the growth medium and light intensityon lipid production by microalgae isolated from freshwater sources in Thailand Bioresour Technol 2011 1023034ndash3040 [CrossRef] [PubMed]
63 Msanne J Xu D Konda AR Casas-Mollano JA Awada T Cahoon EB Cerutti H Metabolic andgene expression changes triggered by nitrogen deprivation in the photoautotrophically grown microalgaeChlamydomonas reinhardtii and Coccomyxa sp C-169 Phytochemistry 2012 75 50ndash59 [CrossRef] [PubMed]
64 Lin Q Lin J Effects of nitrogen source and concentration on biomass and oil production of aScenedesmus rubescens like microalga Bioresour Technol 2011 102 1615ndash1621 [CrossRef] [PubMed]
65 Sheehan J Dunahay T Benemann J Roessler P A Look Back at the US Department of Energyrsquos AquaticSpecies ProgrammdashBiodiesel from Algae US Department of Energyrsquos Office of Fuels Development Golden COUSA 1998 p 328
66 Vaulot D Olson RJ Merkel S Chisholm SW Cell-cycle response to nutrient starvation in twophytoplankton species Thalassiosira weissflogii and Hymenomonas carterae Mar Biol 1987 95 625ndash630[CrossRef]
67 Olson RJ Chisholm SW Effects of light and nitrogen limitation on the cell cycle of the dinoflagellateAmphidinium carteri J Plankton Res 1986 8 785ndash793 [CrossRef]
68 Li Y Fei X Deng X Novel molecular insights into nitrogen starvation-induced triacylglycerolsaccumulation revealed by differential gene expression analysis in green algae Micractinium pusillumBiomass Bioenergy 2012 42 199ndash211 [CrossRef]
69 Abdelaziz AEM Leite GB Belhaj MA Hallenbeck PC Screening microalgae native to Quebec forwastewater treatment and biodiesel production Bioresour Technol 2014 157 140ndash148 [CrossRef] [PubMed]
70 Valdez-Ojeda R Gonzaacutelez-Muntildeoz M Us-Vaacutezquez R Narvaacuteez-Zapata J Chavarria-Hernandez JCLoacutepez-Adriaacuten S Barahona-Peacuterez F Toledano-Thompson T Garduntildeo-Soloacuterzano G Escobedo-GraciaMedrano RM Characterization of five fresh water microalgae with potential for biodiesel productionAlgal Res 2015 7 33ndash44 [CrossRef]
71 Shrivastav A Mishra SK Suh WI Farooq W Moon M Kim T-H Kumar K Choi G-G Park MSYang J-W Characterization of newly isolated oleaginous microalga Monoraphidium sp for lipid productionunder different conditions Algal Res 2015 12 289ndash294 [CrossRef]
72 Yen H-W Hu I-C Chen C-Y Ho S-H Lee D-J Chang J-S Microalgae-based biorefinerymdashFrombiofuels to natural products Bioresour Technol 2013 135 166ndash174 [CrossRef] [PubMed]
Energies 2017 10 224 16 of 16
73 Li J Liu Y Cheng JJ Mos M Daroch M Biological potential of microalgae in China for biorefinery-basedproduction of biofuels and high value compounds New Biotechnol 2015 32 588ndash596 [CrossRef] [PubMed]
74 El-Sheekh M Abomohra AE-F Hanelt D Optimization of biomass and fatty acid productivity ofScenedesmus obliquus as a promising microalga for biodiesel production World J Microbiol Biotechnol 201329 915ndash922 [CrossRef] [PubMed]
75 Singh P Guldhe A Kumari S Rawat I Bux F Investigation of combined effect of nitrogen phosphorusand iron on lipid productivity of microalgae Ankistrodesmus falcatus KJ671624 using response surfacemethodology Biochem Eng J 2015 94 22ndash29 [CrossRef]
76 Kilham S Kreeger D Goulden C Lynn S Effects of nutrient limitation on biochemical constituents ofAnkistrodesmus falcatus Freshw Biol 1997 38 591ndash596 [CrossRef]
77 Breuer G Lamers PP Martens DE Draaisma RB Wijffels RH The impact of nitrogen starvation on thedynamics of triacylglycerol accumulation in nine microalgae strains Bioresour Technol 2012 124 217ndash226[CrossRef] [PubMed]
78 Breuer G de Jaeger L Artus VG Martens DE Springer J Draaisma RB Eggink G Wijffels RHLamers PP Superior triacylglycerol (TAG) accumulation in starchless mutants of Scenedesmus obliquus (II)evaluation of TAG yield and productivity in controlled photobioreactors Biotechnol Biofuels 2014 7 70[CrossRef] [PubMed]
79 Nigam S Prakash-Rai M Sharma R Effect of Nitrogen on Growth and Lipid Content ofChlorella pyrenoidosa Am J Biochem Biotechnol 2011 7 126ndash131 [CrossRef]
80 Piligaev AV Sorokina KN Bryanskaya AV Peltek SE Kolchanov NA Parmon VN Isolation ofprospective microalgal strains with high saturated fatty acid content for biofuel production Algal Res 201512 368ndash376 [CrossRef]
81 Weiss SB Kennedy EP The enzymatic synthesis of triglycerides J Am Chem Soc 1956 78 3550[CrossRef]
82 Miller R Wu G Deshpande RR Vieler A Gaumlrtner K Li X Moellering ER Zaumluner S Cornish AJLiu B et al Changes in Transcript Abundance in Chlamydomonas reinhardtii following Nitrogen DeprivationPredict Diversion of Metabolism Plant Physiol 2010 154 1737ndash1752 [CrossRef] [PubMed]
83 Rismani-Yazdi H Haznedaroglu BZ Bibby K Peccia J Transcriptome sequencing and annotation of themicroalgae Dunaliella tertiolecta Pathway description and gene discovery for production of next-generationbiofuels BMC Genom 2011 12 148 [CrossRef] [PubMed]
84 Ramos MJ Fernaacutendez CM Casas A Rodriacuteguez L Peacuterez A Influence of fatty acid composition of rawmaterials on biodiesel properties Bioresour Technol 2009 100 261ndash268 [CrossRef] [PubMed]
85 Knothe G Dependence of biodiesel fuel properties on the structure of fatty acid alkyl estersFuel Process Technol 2005 86 1059ndash1070 [CrossRef]
86 Knothe G ldquoDesignerrdquo Biodiesel Optimizing Fatty Ester Composition to Improve Fuel PropertiesEnergy Fuels 2008 22 1358ndash1364 [CrossRef]
87 Smith PC Ngothai Y Dzuy Nguyen Q OrsquoNeill BK Improving the low-temperature properties ofbiodiesel Methods and consequences Renew Energy 2010 35 1145ndash1151 [CrossRef]
88 Dunn RO Effect of antioxidants on the oxidative stability of methyl soyate (biodiesel) Fuel Process Technol2005 86 1071ndash1085 [CrossRef]
89 Knothe G Improving biodiesel fuel properties by modifying fatty ester composition Energy Environ Sci2009 2 759ndash766 [CrossRef]
copy 2017 by the authors licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)

Energies 2017 10 224 7 of 16
To realize phylogenetic analysis genomic DNA of each isolate strain was purified and used forPCR amplification of ITS2 rDNA The universal primers used to amplify the ITS2 rDNA successfullyamplified the desired DNA fragments (~300 bp) from all microalgal strains (Figure S2) Furtherthese amplicons were sequenced published in the NCBI databases and the corresponding GenBankaccession numbers are presented in Table 1 Figure 1 shows a neighbor-joining phylogenetic treedepicting the relatedness of the native microalgae strain isolates with selected sequences from thepublic GenBank database This phylogram contained three clusters each of which included differentmicroalgae genera For example the first cluster consisted of microalgae of the genera AcutodesmusScenedesmus Tetradesmus and Desmodesmus and six isolated strains (ITA007 AMA004 ITA006 NAN003ITA005 and NAN002) The second cluster consisted of a microalgae of the genus Ankistrodesmus andthe isolated strains AMA001 ITA001 ITA002 NAN001 and AMA002 The third one consisted of amicroalgae of the genus Chlorella and three isolated strains (ITA004 AMA003 and ITA003) Overall theidentification of native microalgae strains presented a good correspondence between morphologicalcharacters and sequence-based phylogenetic analysis Microalgae species of these genera were alsoisolated from different countries of the world such as Brazil [47] Argentina [46] United States [48]India [20] Egypt [49] Taiwan [50] South Korea [45] and China [51] Some of these microalgae generasuch as Ankistrodesmus Chlorella Desmodesmus and Scenedesmus were used in large-scale biodieselproduction using photobioreactors and raceway pond systems [5253]
Energies 2017 10 224 7 of 16
public GenBank database This phylogram contained three clusters each of which included different microalgae genera For example the first cluster consisted of microalgae of the genera Acutodesmus Scenedesmus Tetradesmus and Desmodesmus and six isolated strains (ITA007 AMA004 ITA006 NAN003 ITA005 and NAN002) The second cluster consisted of a microalgae of the genus Ankistrodesmus and the isolated strains AMA001 ITA001 ITA002 NAN001 and AMA002 The third one consisted of a microalgae of the genus Chlorella and three isolated strains (ITA004 AMA003 and ITA003) Overall the identification of native microalgae strains presented a good correspondence between morphological characters and sequence-based phylogenetic analysis Microalgae species of these genera were also isolated from different countries of the world such as Brazil [47] Argentina [46] United States [48] India [20] Egypt [49] Taiwan [50] South Korea [45] and China [51] Some of these microalgae genera such as Ankistrodesmus Chlorella Desmodesmus and Scenedesmus were used in large-scale biodiesel production using photobioreactors and raceway pond systems [5253]
Figure 1 Unrooted phylogenetic tree of isolated microalgae strains using the ITS2 sequences The tree was constructed by the neighbor-joining method and Kimurarsquos two-parameter algorithm in MEGA 60 software based on the multiple sequence alignment by Clustal Omega Bootstrap values of 1000 replicates () are shown at the branches
32 Microalgae Cultivation Biomass and Total Lipids Measurements
The isolated microalgae strains exhibited remarkable differences in productivity parameters and lipid content among individual strains in the stationary growth phase (Table 1) With regard to the biomass productivity the 14 strains showed statistically-significant differences (F = 71911 df = 13 p lt 0001) and ranged from 36 plusmn 04 mgmiddotLminus1middotdminus1 (Ankistrodesmus sp strain AMA002) to 316 plusmn 04 mgmiddotLminus1middotdminus1 (Acutodesmus obliquus strain AMA004) Similar biomass productivities were reported for other marine microalgae species [54] However other investigations show major biomass productivities (ranging
Figure 1 Unrooted phylogenetic tree of isolated microalgae strains using the ITS2 sequences The treewas constructed by the neighbor-joining method and Kimurarsquos two-parameter algorithm in MEGA60 software based on the multiple sequence alignment by Clustal Omega Bootstrap values of1000 replicates () are shown at the branches
Energies 2017 10 224 8 of 16
32 Microalgae Cultivation Biomass and Total Lipids Measurements
The isolated microalgae strains exhibited remarkable differences in productivity parametersand lipid content among individual strains in the stationary growth phase (Table 1) With regardto the biomass productivity the 14 strains showed statistically-significant differences (F = 71911df = 13 p lt 0001) and ranged from 36 plusmn 04 mgmiddotLminus1middotdminus1 (Ankistrodesmus sp strain AMA002) to316 plusmn 04 mgmiddotLminus1middotdminus1 (Acutodesmus obliquus strain AMA004) Similar biomass productivities werereported for other marine microalgae species [54] However other investigations show major biomassproductivities (ranging from 30 to 120 mgmiddotLminus1middotdminus1) in the same and other microalgae genera [455556]These discrepancies may be attributed to differences in the culture conditions in the culture mediacomposition and in the microalgae genotypes Lipid productivity also exhibited statistically-significantdifferences (F = 4774 df = 13 p lt 0001) and ranged from 10plusmn 04 mgmiddotLminus1middotdminus1 to 64 plusmn 07 mgmiddotLminus1middotdminus1
with strains AMA002 and AMA004 respectively These lipid productivities were low in comparison toother reports which reported values from 190 to 433 mgmiddotLminus1middotdminus1 [4555] Furthermore lipid contentas a percentage of the microalgal biomass dry weight showed statistically-significant differences(F = 8509 df = 13 p lt 0001) and ranged from 135 plusmn 12 (Tetradesmus sp strain NAN003) to467 plusmn 15 (Ankistrodesmus sp strain NAN001) These percentage values in total lipid contentare similar to several microalgae species of these genera isolated and characterized by otherresearchers worldwide [20454648ndash5057] Furthermore it is important to emphasize that lipid contentis dependent on the different growth phases Commonly in several microalgae taxa the lipid contentincreases gradually from logarithmic to stationary growth phases [58ndash60] For example Kaur et al [20]reported that the total lipid content of Scenedesmus sp DRLMA9 demonstrated values of 207 inthe late exponential phase and 397 in the stationary phase and Desmodesmus elegans DRLMA13displayed values of 103 in the late exponential phase and 169 in the stationary phase LikewiseChiu et al [61] found that the total lipid content of Nannochloropsis oculata was 308 in the logarithmicphase 397 in the early stationary phase and 504 in the stationary phase These changes in lipidcontent with culture age have been associated with nitrate depletion [61] which causes a channeling ofphotosynthetic ATP to carbohydrate and lipid biosynthesis [60] Finally Pearson correlation analysisof biomass productivity and lipid content was inversely related (r = minus037 p lt 001) which is similarto the results obtained by Liang et al [62] and Rodolfi et al [52] In contrast Li et al [51] andHempel et al [63] found no relationship between these parameters The negative correlation obtainedis likely due to the high metabolic cost of lipid biosynthesis [62]
33 Strains Culture under Nitrogen-Sufficient and Nitrogen-Deficient Medium
The microalgae species Acutodesmus obliquus (KP8785051) Ankistrodesmus sp (KP8785001)and Chlorella lewinii (KP8785011) were selected and further characterized because they displayedthe highest biomass and lipid productivities (Table 1) These microalgae showed noticeablevariations in the growth profile when cultured in Chu medium No 10 with nitrogen (+N) orwithout a nitrogen source (minusN) In +N medium all strains showed exponential growth butChlorella lewinii showed the highest average specific growth rates (micro = 0458 plusmn 0098 dminus1) followed byAcutodesmus obliquus (micro = 0399 plusmn 0112 dminus1) and the lowest value was produced by Ankistrodesmus sp(micro = 0265 plusmn 0013 dminus1) In contrast all cultures displayed a lag phase pattern in their growthprofile under minusN medium (Figure 2) These findings corroborate the results obtained by otherresearchers who demonstrate that the specific growth rate is directly proportional to the initialnitrate concentration [46596465] because the reduction or cessation of microalgae cell divisionis due to nitrogen limitation [66] which is a key nutrient for protein and nucleic acids de novobiosynthesis consequently causing an arrest in the early part of the microalgae cell cycle [6768]In accordance with these reports Li et al [69] showed that nitrogen deficiency induced in the greenmicroalgae Micractinium pusillum a downregulation in the expression of genes involved in cell growthand cell cycle regulation Additionally differences in growth profile and specific growth rates aredependent on several factors such as microalgae genotypes the culture medium composition pH
Energies 2017 10 224 9 of 16
nutrients concentration the cell culture phase (ie log phase stationary phase) and environmentalfactors [4549545770ndash72]Energies 2017 10 224 9 of 16
Figure 2 Growth profiles of three microalgal strains cultivated under +N (filled boxes) and minusN (empty boxes) Chu medium No 10 The culture was incubated in a controlled culture room at 25 plusmn 1 degC with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 150 rpm Experiments were carried out in triplicate
34 Total Protein Total Lipid and Total Carbohydrate Content
Biochemical parameters of the microalgae species such as total protein total lipid and total carbohydrate content exhibited differences and changes under both culture conditions (Figure 3) For example Chlorella lewinii showed the highest protein content Ankistrodesmus sp had the highest total lipid content but Acutodesmus obliquus presented the highest levels of total carbohydrate Therefore these strains can be valuable stocks for the industry because total protein and total carbohydrate are potentially byproducts of microalgal biomass processing [7374] Additionally the three strains cultured under minusN conditions showed a significant increase in total lipid content (from 237 in Acutodesmus obliquus to 667 in Ankistrodesmus sp) and total carbohydrate content (from 119 in Acutodesmus obliquus to 963 in Chlorella lewinii) but showed a decreased in total protein levels (from 242 in Acutodesmus obliquus to 545 in Chlorella lewinii) in comparison to +N cultures (Figure 3) These results are in accordance with well-documented results obtained by other researchers with several microalgae species of the genera Acutodesmus Ankistrodesmus Botryococcus Chlamydomonas Chlorella Desmodesmus Dunaliella Haematococcus Neochloris Phaeodactylum Pseudokirchneriella and Scenedesmus [465058596475ndash80] Nevertheless the accumulation of total lipid induced by nitrogen deficiency is not correlated with lipid productivity but rather is due to a decline and stopping of the microalgae specific growth rate [75] The changes in biomolecule contents suggest redirection of microalgae metabolism during nitrogen starvation such as a decrease in biosynthesisincrease in protein degradation and an increase in the biosynthesis of storage biomolecules such as triacylglycerols and starch The increase in total lipid content could be partially explained for a boost in transcript levels of genes encoding enzymes of the lipid biosynthesis pathways According to this assumption recent investigations showed that genes encoding the enzymes that catalyze the last step in the Kennedy pathway of triacylglycerol biosynthesis [81] such as glycerol-3-phosphate acyltransferase and acyl-glycerol-3-phosphate acyltransferase are upregulated in response to nitrogen starvation [648283] Furthermore the increase in total carbohydrate content can be explained by changes in the expression levels of genes involved in carbohydrate metabolism For example when the oleaginous microalgae Neochloris oleoabundans is cultured under nitrogen starvation the catabolic phase of starch specifically α-amylase which hydrolyzes starch to glucose is strongly repressed [83]
Figure 2 Growth profiles of three microalgal strains cultivated under +N (filled boxes) and minusN(empty boxes) Chu medium No 10 The culture was incubated in a controlled culture room at25 plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent lightand continuous agitation at 150 rpm Experiments were carried out in triplicate
34 Total Protein Total Lipid and Total Carbohydrate Content
Biochemical parameters of the microalgae species such as total protein total lipid and totalcarbohydrate content exhibited differences and changes under both culture conditions (Figure 3)For example Chlorella lewinii showed the highest protein content Ankistrodesmus sp had the highesttotal lipid content but Acutodesmus obliquus presented the highest levels of total carbohydrateTherefore these strains can be valuable stocks for the industry because total protein and totalcarbohydrate are potentially byproducts of microalgal biomass processing [7374] Additionallythe three strains cultured under minusN conditions showed a significant increase in total lipid content(from 237 in Acutodesmus obliquus to 667 in Ankistrodesmus sp) and total carbohydrate content(from 119 in Acutodesmus obliquus to 963 in Chlorella lewinii) but showed a decreased in totalprotein levels (from 242 in Acutodesmus obliquus to 545 in Chlorella lewinii) in comparison to +Ncultures (Figure 3) These results are in accordance with well-documented results obtained by otherresearchers with several microalgae species of the genera Acutodesmus Ankistrodesmus BotryococcusChlamydomonas Chlorella Desmodesmus Dunaliella Haematococcus Neochloris PhaeodactylumPseudokirchneriella and Scenedesmus [465058596475ndash80] Nevertheless the accumulation of totallipid induced by nitrogen deficiency is not correlated with lipid productivity but rather is due toa decline and stopping of the microalgae specific growth rate [75] The changes in biomoleculecontents suggest redirection of microalgae metabolism during nitrogen starvation such as a decreasein biosynthesisincrease in protein degradation and an increase in the biosynthesis of storagebiomolecules such as triacylglycerols and starch The increase in total lipid content could bepartially explained for a boost in transcript levels of genes encoding enzymes of the lipid biosynthesispathways According to this assumption recent investigations showed that genes encoding theenzymes that catalyze the last step in the Kennedy pathway of triacylglycerol biosynthesis [81] such asglycerol-3-phosphate acyltransferase and acyl-glycerol-3-phosphate acyltransferase are upregulated
Energies 2017 10 224 10 of 16
in response to nitrogen starvation [648283] Furthermore the increase in total carbohydrate contentcan be explained by changes in the expression levels of genes involved in carbohydrate metabolismFor example when the oleaginous microalgae Neochloris oleoabundans is cultured under nitrogenstarvation the catabolic phase of starch specifically α-amylase which hydrolyzes starch to glucoseis strongly repressed [83]Energies 2017 10 224 10 of 16
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strains cultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled culture room at 25 plusmn 1 degC with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized with respect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that each microalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majority of fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised 7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii the majority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9) acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectively Finally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic (182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in +N and minusN conditions respectively Our results agree with multiple previously-published reports in that fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent in many green microalgae species [454648ndash5055] It is notable that under nitrogen starvation culture all microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 in Ankistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid (181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findings support the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content of SFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Nevertheless there are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed a decrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increased between 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively However in the same culture condition the three microalgae species showed a decrease in polyunsaturated fatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmus spp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgae strains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFA which improves the quality properties of biodiesel such as an increase
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strainscultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled cultureroom at 25plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescentlight and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized withrespect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that eachmicroalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majorityof fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii themajority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9)acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectivelyFinally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic(182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in+N and minusN conditions respectively Our results agree with multiple previously-published reports inthat fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent inmany green microalgae species [454648ndash5055] It is notable that under nitrogen starvation cultureall microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 inAnkistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid(181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findingssupport the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content ofSFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other
Energies 2017 10 224 11 of 16
researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Neverthelessthere are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed adecrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increasedbetween 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively Howeverin the same culture condition the three microalgae species showed a decrease in polyunsaturatedfatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmusspp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgaestrains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFAwhich improves the quality properties of biodiesel such as an increase in cetane number [84ndash86]Considering the SFA and unsaturated fatty acids (UFA) components approximately 20ndash31 are SFAand 69ndash80 are UFA Hence the SFAUFA ratio in the microalgae strains evaluated ranges between025 and 045 which is similar to the reports of several microalgae species isolated in different latitudesworldwide [203148497072]
Table 2 Effect of nitrogen deficiency on fatty acid profiling in three native microalgal species from thePeruvian Amazon
Fatty AcidsAcutodesmus obliquus Ankistrodesmus sp Chlorella lewinii
+N minusN +N minusN +N minusN
140 - - 052 plusmn 002 088 plusmn 002 - 058 plusmn 001160 2724 plusmn 007 2867 plusmn 017 1868 plusmn 019 2261 plusmn 006 1964 plusmn 021 2123 plusmn 010180 184 plusmn 005 255 plusmn 051 101 plusmn 002 181 plusmn 002 371 plusmn 026 153 plusmn 015
161n-7 023 plusmn 000 025 plusmn 001 071 plusmn 003 055 plusmn 001 085 plusmn 043 038 plusmn 000181n-7 131 plusmn 002 131 plusmn 000 141 plusmn 005 094 plusmn 003 146 plusmn 006 123 plusmn 002181n-9 3213 plusmn 021 3608 plusmn 021 3699 plusmn 004 4047 plusmn 038 1371 plusmn 015 2326 plusmn 041182n-6 1732 plusmn 004 1651 plusmn 016 747 plusmn 018 656 plusmn 007 2034 plusmn 039 1957 plusmn 001183n-3 1130 plusmn 017 854 plusmn 031 1486 plusmn 004 1173 plusmn 012 1815 plusmn 022 1371 plusmn 016183n-6 101 plusmn 001 078 plusmn 002 055 plusmn 000 054 plusmn 002 124 plusmn 006 100 plusmn 020184n-3 250 plusmn 005 177 plusmn 008 631 plusmn 002 601 plusmn 005 491 plusmn 025 403 plusmn 020204n-6 - - - 084 plusmn 008 - -225n-3 - - - 062 plusmn 002 - -
Unidentified 512 plusmn 003 355 plusmn 006 1149 plusmn 004 645 plusmn 003 1599 plusmn 034 1348 plusmn 006SFA 2907 3122 2022 2531 2335 2334UFA 7093 6878 7978 7469 7665 7666
MUFA 3367 3764 3911 4195 1602 2487PUFA 3213 2759 2918 2629 4464 3831SFA +MUFA 6274 6886 5933 6726 3936 4821
It is important to highlight that the fatty acid profile determination is an essential step inthe process of the characterization of microalgae strains because it ultimately affects the qualityof the biodiesel product [8485] The quality properties of biodiesel such as high cetane numberoxidative stability and cold-flow are dependent on the hydrocarbonated chain length of saturated andunsaturated fatty acids [8587] For example biodiesel composed of a mixture of a high percentage ofSFA and MUFA is preferred for increased energy yield superior oxidative stability and higher cetanenumbers but is prone to solidify at low temperature In contrast biodiesel with high levels of PUFAhas decreased energy yield low cetane numbers and excellent cold-flow properties but insufficientoxidative stability [548588] Therefore to solve these performance problems five approaches havebeen developed such as changing the fatty acids profile by physical means genetic modification ofthe feedstock or the use of alternative feedstocks with different fatty acids profiles [89]
4 Conclusions
Fourteen microalgae were isolated from three river basins from the Peruvian Amazon and wereidentified by morphological features and molecular phylogeny approaches These strains were initially
Energies 2017 10 224 12 of 16
characterized in the stationary growth phase and exhibited significant heterogeneity in key parameterssuch as biomass productivity (36ndash316 mgmiddotLminus1middotdminus1) lipid productivity (10ndash64 mgmiddotLminus1middotdminus1) andtotal lipid content (135ndash467) Furthermore three selected strains based on elevated biomass andlipid productivities were cultured under nitrogen-sufficient and nitrogen-deficient medium to boostlipid accumulation Under these culture conditions all strains showed noticeable decreases in totalprotein content and increases in total lipid and total carbohydrate contents In addition these strainspresented modifications in the fatty acids profiles with improvement in biodiesel properties due to apredominance of SFA and MUFA which corresponds to an increase in cetane number The results ofthis research suggest that the isolated native microalgae from the Peruvian Amazon could be suitablerenewable feedstocks for the production of both biodiesel and bioethanol which could contribute tosustainable development at the local regional and national levels as a viable alternative to petroleumexploitation However it will be necessary to develop open closed or hybrid systems for large-scalebiomass production of selected native microalgae strains andor de novo microalgae strains designedto produce biofuels and with the abilities to synthesize useful bioactive compounds to be efficientlyexploited using biorefinery approaches
Supplementary MaterialsThe following are available online at wwwmdpicom1996-1073102224s1 Figure S1 Results of
electrophoretic and spectrophotometric analysis of the genomic DNA isolated from the fourteen microalgaeFigure S2 ITS2-rDNA structure (A) and amplicons obtained (B) from the microalgae isolated Figure S3Microphotographic images of the three microalgae strains analyzed under light and fluorescence stained withNile Red Figure S4 Spectrophotometric scanning for the determination of the maximum absorption peak foreach microalgae strain
Acknowledgments This research was supported by the Peruvian funding agencies Consejo Nacional de CienciaTecnologiacutea e Innovacioacuten Tecnoloacutegica (CONCYTEC) with Grant Contract No 364-CONCYTEC-OAJ and Fondospara la Innovacioacuten Ciencia y Tecnologiacutea (FINCyT) with Grant Contract No 383-PNICP-PIBA-2014
Author Contributions Marianela Cobos and Juan C Castro obtained funds for the research participated inthe study design coordinated activities and participated in the preparation of the manuscript Jae D Paredesparticipated in sample collection microalgae isolation and helped to draft the manuscript J Dylan Maddoxperformed the bioinformatics analysis and participated in the preparation of the manuscript Gabriel Vargas-Aranaand Leenin Flores participated in biochemical analysis and helped to draft the manuscript Carla P Aguilarparticipated in the study design supervised biochemical analysis and helped to draft the manuscriptJorge L Marapara participated in molecular analysis and helped to draft the manuscript
Conflicts of Interest The authors declare no conflict of interest
References
1 BP Global BP Statisical Review of World Energy 2014 Available online httpwwwbpcomcontentdambppdfEnergy-economicsstatistical-review-2014BP-statistical-review-of-world-energy-2014-full-reportpdf (accessed on 20 June 2014)
2 Pachauri RK Reisinger A Cambio Climaacutetico 2007 Informe de Siacutentesis Contribucioacuten de los Grupos de Trabajo III y III al Cuarto Informe de Evaluacioacuten del Grupo Intergubernamental de Expertos Sobre el Cambio Climaacutetico IPCCGeneva Switzerland 2007 p 104
3 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Ley de Promocioacuten del Mercado deBiocombustibles Available online httpwww2congresogobpesicrcendocbibcon4_uibdnsf9E95620CC059138105257C9E005AB2F9$FILE28054pdf (accessed on 12 February 2017)
4 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Reglamento de la Ley de Promocioacuten del Mercadode Biocombustibles Available online httpwww2osinerggobpeMarcoLegaldocrevDS-013-2005-EM-CONCORDADOpdf (accessed on 12 February 2017)
5 Areacutevalo LF Nalvarte J Torres J Ramiacuterez Y Impactos Socio-Econoacutemicos de la Produccioacuten de Biocombustiblesen la Amazoniacutea Peruana Primera SNV-WWF Lima Peruacute 2009
6 Sharma KK Schuhmann H Schenk PM High Lipid Induction in Microalgae for Biodiesel ProductionEnergies 2012 5 1532ndash1553 [CrossRef]
Energies 2017 10 224 13 of 16
7 Schenk PM Thomas-Hall SR Stephens E Marx UC Mussgnug JH Posten C Kruse O Hankamer BSecond Generation Biofuels High-Efficiency Microalgae for Biodiesel Production BioEnergy Res 2008 120ndash43 [CrossRef]
8 Foley JA Asner GP Costa MH Coe MT de Fries R Gibbs HK Howard EA Olson S Patz JRamankutty N et al Amazonia revealed Forest degradation and loss of ecosystem goods and services inthe Amazon Basin Front Ecol Environ 2007 5 25ndash32 [CrossRef]
9 Duong VT Li Y Nowak E Schenk PM Microalgae Isolation and Selection for Prospective BiodieselProduction Energies 2012 5 1835ndash1849 [CrossRef]
10 Georgianna DR Mayfield SP Exploiting diversity and synthetic biology for the production of algalbiofuels Nature 2012 488 329ndash335 [CrossRef] [PubMed]
11 Wijffels RH Barbosa MJ An Outlook on Microalgal Biofuels Science 2010 329 796ndash799 [CrossRef][PubMed]
12 Richmond A Handbook of Microalgal Culture Biotechnology and Applied Phycology Blackwell Sci Ltd HudsonNJ USA 2004
13 Menetrez MY An overview of algae biofuel production and potential environmental impactEnviron Sci Technol 2012 46 7073ndash7085 [CrossRef] [PubMed]
14 Khoo HH Sharratt PN Das P Balasubramanian RK Naraharisetti PK Shaik S Life cycle energy andCO2 analysis of microalgae-to-biodiesel Preliminary results and comparisons Bioresour Technol 2011 1025800ndash5807 [CrossRef] [PubMed]
15 Chu SP The Influence of the Mineral Composition of the Medium on the Growth of Planktonic AlgaePart I Methods and Culture Media J Ecol 1942 30 284ndash325 [CrossRef]
16 Carlos EMB Mariatildengela M Gecircneros de Algas de Aacuteguas Continentais do BrasilmdashChave Para Identificaccedilatildeo eDescriccedilotildees 2nd ed Rima Sao Carlos Brasil 2006
17 Greenspan P Mayer EP Fowler SD Nile red A selective fluorescent stain for intracellular lipid dropletsJ Cell Biol 1985 100 965ndash973 [CrossRef] [PubMed]
18 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissuePhytochem Bull 1987 19 11ndash15
19 Sambrook J Fritsch EF Maniatis T Molecular Cloning A Laboratory Manual 2nd ed Cold Spring HarborLaboratory Press New York NY USA 1989
20 Kaur S Sarkar M Srivastava RB Gogoi HK Kalita MC Fatty acid profiling and molecularcharacterization of some freshwater microalgae from India with potential for biodiesel productionNew Biotechnol 2012 29 332ndash344 [CrossRef] [PubMed]
21 Benson DA Cavanaugh M Clark K Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW GenBankNucleic Acids Res 2013 41 D36ndashD42 [CrossRef] [PubMed]
22 NCBI Resource Coordinators Database Resources of the National Center for Biotechnology InformationNucleic Acids Res 2015 43 D6ndashD17
23 Ye J McGinnis S Madden TL BLAST Improvements for better sequence analysis Nucleic Acids Res 200634 W6ndashW9 [CrossRef] [PubMed]
24 BioEdit Sequence Alignment Editor for Windows 9598NTXPVista7 Available online httpwwwmbioncsuedubioeditbioedithtml (accessed on 2 December 2015)
25 Sievers F Higgins DG Clustal omega accurate alignment of very large numbers of sequencesMethods Mol Biol (Clifton NJ) 2014 1079 105ndash116
26 Saitou N Nei M The neighbor-joining method A new method for reconstructing phylogenetic treesMol Biol Evol 1987 4 406ndash425 [PubMed]
27 Kimura M A simple method for estimating evolutionary rates of base substitutions through comparativestudies of nucleotide sequences J Mol Evol 1980 16 111ndash120 [CrossRef] [PubMed]
28 Tamura K Stecher G Peterson D Filipski A Kumar S MEGA6 Molecular Evolutionary GeneticsAnalysis version 60 Mol Biol Evol 2013 30 2725ndash2729 [CrossRef] [PubMed]
29 Felsenstein J Confidence Limits on Phylogenies An Approach Using the Bootstrap Evolution 1985 39783ndash791 [CrossRef]
30 Bligh EG Dyer WJ A Rapid Method of Total Lipid Extraction and Purification Can J Biochem Physiol1959 37 911ndash917 [CrossRef] [PubMed]
Energies 2017 10 224 14 of 16
31 Yu X Zhao P He C Li J Tang X Zhou J Huang Z Isolation of a novel strain of Monoraphidium spand characterization of its potential application as biodiesel feedstock Bioresour Technol 2012 121 256ndash262[CrossRef] [PubMed]
32 Hartree EF Determination of protein A modification of the lowry method that gives a linear photometricresponse Anal Biochem 1972 48 422ndash427 [CrossRef]
33 DuBois M Gilles KA Hamilton JK Rebers PA Smith F Colorimetric Method for Determination ofSugars and Related Substances Anal Chem 1956 28 350ndash356 [CrossRef]
34 AOAC International AOAC Official Methods of Analysis of the Association of Official Analytical Chemists (AOAC)15th ed AOAC International Washington DC USA 1990 Volume 1
35 Ichihara K Fukubayashi Y Preparation of fatty acid methyl esters for gas-liquid chromatographyJ Lipid Res 2010 51 635ndash640 [CrossRef] [PubMed]
36 Brennan L Owende P Biofuels from microalgaemdashA review of technologies for production processingand extractions of biofuels and co-products Renew Sustain Energ Rev 2010 14 557ndash577 [CrossRef]
37 Raja R Hemaiswarya S Kumar NA Sridhar S Rengasamy R A perspective on the biotechnologicalpotential of microalgae Crit Rev Microbiol 2008 34 77ndash88 [CrossRef] [PubMed]
38 Mata TM Martins AA Caetano NS Microalgae for biodiesel production and other applicationsA review Renew Sustain Energy Rev 2010 14 217ndash232 [CrossRef]
39 De Wilt A Butkovskyi A Tuantet K Leal LH Fernandes TV Langenhoff A Zeeman GMicropollutant removal in an algal treatment system fed with source separated wastewater streamsJ Hazard Mater 2015 304 84ndash92 [CrossRef] [PubMed]
40 Li T Lin G Podola B Melkonian M Continuous removal of zinc from wastewater and mine dumpleachate by a microalgal biofilm PSBR J Hazard Mater 2015 297 112ndash118 [CrossRef] [PubMed]
41 Sioli H The Amazon and its main affluents Hydrography morphology of the river courses and river typesIn Monographiae Biologicae Sioli H Ed Springer Dordrecht The Netherlands 1984 pp 127ndash165
42 Moniz MBJ Kaczmarska I Barcoding of Diatoms Nuclear Encoded ITS Revisited Protist 2010 161 7ndash34[CrossRef] [PubMed]
43 Coleman AW Is there a molecular key to the level of ldquobiological speciesrdquo in eukaryotes A DNA guideMol Phylogenet Evol 2009 50 197ndash203 [CrossRef] [PubMed]
44 Mai JC Coleman AW The Internal Transcribed Spacer 2 Exhibits a Common Secondary Structure inGreen Algae and Flowering Plants J Mol Evol 1997 44 258ndash271 [CrossRef] [PubMed]
45 Abou-Shanab RAI Matter IA Kim S-N Oh Y-K Choi J Jeon B-H Characterization andidentification of lipid-producing microalgae species isolated from a freshwater lake Biomass Bioenergy2011 35 3079ndash3085 [CrossRef]
46 Do Nascimento M Ortiz-Marquez JCF Sanchez-Rizza L Echarte MM Curatti L Bioprospectingfor fast growing and biomass characterization of oleaginous microalgae from South-Eastern Buenos AiresArgentina Bioresour Technol 2012 125 283ndash290 [CrossRef] [PubMed]
47 Araujo GS Matos LJBL Gonccedilalves LRB Fernandes FAN Farias WRL Bioprospecting foroil producing microalgal strains Evaluation of oil and biomass production for ten microalgal strainsBioresour Technol 2011 102 5248ndash5250 [CrossRef] [PubMed]
48 Zhou W Li Y Min M Hu B Chen P Ruan R Local bioprospecting for high-lipid producing microalgalstrains to be grown on concentrated municipal wastewater for biofuel production Bioresour Technol 2011102 6909ndash6919 [CrossRef] [PubMed]
49 Mahmoud EA Farahat LA Abdel Aziz ZK Fatthallah NA Salah El Din RA Evaluation of thepotential for some isolated microalgae to produce biodiesel Egypt J Pet 2015 24 97ndash101 [CrossRef]
50 Ho S Lai Y-Y Chiang C-Y Chen C-NN Chang J-S Selection of elite microalgae for biodieselproduction in tropical conditions using a standardized platform Bioresour Technol 2013 147 135ndash142[CrossRef] [PubMed]
51 Li L Cui J Liu Q Ding Y Liu J Screening and phylogenetic analysis of lipid-rich microalgae Algal Res2015 11 381ndash386 [CrossRef]
52 Rodolfi L Chini Zittelli G Bassi N Padovani G Biondi N Bonini G Tredici MR Microalgae for oilStrain selection induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactorBiotechnol Bioeng 2009 102 100ndash112 [CrossRef] [PubMed]
Energies 2017 10 224 15 of 16
53 Li P Miao X Li R Zhong J In situ biodiesel production from fast-growing and high oil content Chlorellapyrenoidosa in rice straw hydrolysate J Biomed Biotechnol 2011 2011 141207 [CrossRef] [PubMed]
54 Doan TTY Sivaloganathan B Obbard JP Screening of marine microalgae for biodiesel feedstockBiomass Bioenergy 2011 35 2534ndash2544 [CrossRef]
55 Abou-Shanab RAI Hwang J-H Cho Y Min B Jeon B-H Characterization of microalgal speciesisolated from fresh water bodies as a potential source for biodiesel production Appl Energy 2011 883300ndash3306 [CrossRef]
56 Chaichalerm S Pokethitiyook P Yuan W Meetam M Sritong K Pugkaew W Kungvansaichol KKruatrachue M Damrongphol P Culture of microalgal strains isolated from natural habitats in Thailand invarious enriched media Appl Energy 2012 89 296ndash302 [CrossRef]
57 Zhang S Liu P Yang X Hao Z Zhang L Luo N Shi J Isolation and identification by 18S rDNAsequence of high lipid potential microalgal species for fuel production in Hainan Dao Biomass Bioenergy2014 66 197ndash203 [CrossRef]
58 Chiu S-Y Kao C-Y Tsai M-T Ong S-C Chen C-H Lin C-S Lipid accumulation and CO2 utilizationof Nannochloropsis oculata in response to CO2 aeration Bioresour Technol 2009 100 833ndash838 [CrossRef][PubMed]
59 Huerlimann R de Nys R Heimann K Growth lipid content productivity and fatty acid composition oftropical microalgae for scale-up production Biotechnol Bioeng 2010 107 245ndash257 [CrossRef] [PubMed]
60 Liang Y Sarkany N Cui Y Biomass and lipid productivities of Chlorella vulgaris under autotrophicheterotrophic and mixotrophic growth conditions Biotechnol Lett 2009 31 1043ndash1049 [CrossRef] [PubMed]
61 Hempel N Petrick I Behrendt F Biomass productivity and productivity of fatty acids and amino acidsof microalgae strains as key characteristics of suitability for biodiesel production J Appl Phycol 2012 241407ndash1418 [CrossRef] [PubMed]
62 Yeesang C Cheirsilp B Effect of nitrogen salt and iron content in the growth medium and light intensityon lipid production by microalgae isolated from freshwater sources in Thailand Bioresour Technol 2011 1023034ndash3040 [CrossRef] [PubMed]
63 Msanne J Xu D Konda AR Casas-Mollano JA Awada T Cahoon EB Cerutti H Metabolic andgene expression changes triggered by nitrogen deprivation in the photoautotrophically grown microalgaeChlamydomonas reinhardtii and Coccomyxa sp C-169 Phytochemistry 2012 75 50ndash59 [CrossRef] [PubMed]
64 Lin Q Lin J Effects of nitrogen source and concentration on biomass and oil production of aScenedesmus rubescens like microalga Bioresour Technol 2011 102 1615ndash1621 [CrossRef] [PubMed]
65 Sheehan J Dunahay T Benemann J Roessler P A Look Back at the US Department of Energyrsquos AquaticSpecies ProgrammdashBiodiesel from Algae US Department of Energyrsquos Office of Fuels Development Golden COUSA 1998 p 328
66 Vaulot D Olson RJ Merkel S Chisholm SW Cell-cycle response to nutrient starvation in twophytoplankton species Thalassiosira weissflogii and Hymenomonas carterae Mar Biol 1987 95 625ndash630[CrossRef]
67 Olson RJ Chisholm SW Effects of light and nitrogen limitation on the cell cycle of the dinoflagellateAmphidinium carteri J Plankton Res 1986 8 785ndash793 [CrossRef]
68 Li Y Fei X Deng X Novel molecular insights into nitrogen starvation-induced triacylglycerolsaccumulation revealed by differential gene expression analysis in green algae Micractinium pusillumBiomass Bioenergy 2012 42 199ndash211 [CrossRef]
69 Abdelaziz AEM Leite GB Belhaj MA Hallenbeck PC Screening microalgae native to Quebec forwastewater treatment and biodiesel production Bioresour Technol 2014 157 140ndash148 [CrossRef] [PubMed]
70 Valdez-Ojeda R Gonzaacutelez-Muntildeoz M Us-Vaacutezquez R Narvaacuteez-Zapata J Chavarria-Hernandez JCLoacutepez-Adriaacuten S Barahona-Peacuterez F Toledano-Thompson T Garduntildeo-Soloacuterzano G Escobedo-GraciaMedrano RM Characterization of five fresh water microalgae with potential for biodiesel productionAlgal Res 2015 7 33ndash44 [CrossRef]
71 Shrivastav A Mishra SK Suh WI Farooq W Moon M Kim T-H Kumar K Choi G-G Park MSYang J-W Characterization of newly isolated oleaginous microalga Monoraphidium sp for lipid productionunder different conditions Algal Res 2015 12 289ndash294 [CrossRef]
72 Yen H-W Hu I-C Chen C-Y Ho S-H Lee D-J Chang J-S Microalgae-based biorefinerymdashFrombiofuels to natural products Bioresour Technol 2013 135 166ndash174 [CrossRef] [PubMed]
Energies 2017 10 224 16 of 16
73 Li J Liu Y Cheng JJ Mos M Daroch M Biological potential of microalgae in China for biorefinery-basedproduction of biofuels and high value compounds New Biotechnol 2015 32 588ndash596 [CrossRef] [PubMed]
74 El-Sheekh M Abomohra AE-F Hanelt D Optimization of biomass and fatty acid productivity ofScenedesmus obliquus as a promising microalga for biodiesel production World J Microbiol Biotechnol 201329 915ndash922 [CrossRef] [PubMed]
75 Singh P Guldhe A Kumari S Rawat I Bux F Investigation of combined effect of nitrogen phosphorusand iron on lipid productivity of microalgae Ankistrodesmus falcatus KJ671624 using response surfacemethodology Biochem Eng J 2015 94 22ndash29 [CrossRef]
76 Kilham S Kreeger D Goulden C Lynn S Effects of nutrient limitation on biochemical constituents ofAnkistrodesmus falcatus Freshw Biol 1997 38 591ndash596 [CrossRef]
77 Breuer G Lamers PP Martens DE Draaisma RB Wijffels RH The impact of nitrogen starvation on thedynamics of triacylglycerol accumulation in nine microalgae strains Bioresour Technol 2012 124 217ndash226[CrossRef] [PubMed]
78 Breuer G de Jaeger L Artus VG Martens DE Springer J Draaisma RB Eggink G Wijffels RHLamers PP Superior triacylglycerol (TAG) accumulation in starchless mutants of Scenedesmus obliquus (II)evaluation of TAG yield and productivity in controlled photobioreactors Biotechnol Biofuels 2014 7 70[CrossRef] [PubMed]
79 Nigam S Prakash-Rai M Sharma R Effect of Nitrogen on Growth and Lipid Content ofChlorella pyrenoidosa Am J Biochem Biotechnol 2011 7 126ndash131 [CrossRef]
80 Piligaev AV Sorokina KN Bryanskaya AV Peltek SE Kolchanov NA Parmon VN Isolation ofprospective microalgal strains with high saturated fatty acid content for biofuel production Algal Res 201512 368ndash376 [CrossRef]
81 Weiss SB Kennedy EP The enzymatic synthesis of triglycerides J Am Chem Soc 1956 78 3550[CrossRef]
82 Miller R Wu G Deshpande RR Vieler A Gaumlrtner K Li X Moellering ER Zaumluner S Cornish AJLiu B et al Changes in Transcript Abundance in Chlamydomonas reinhardtii following Nitrogen DeprivationPredict Diversion of Metabolism Plant Physiol 2010 154 1737ndash1752 [CrossRef] [PubMed]
83 Rismani-Yazdi H Haznedaroglu BZ Bibby K Peccia J Transcriptome sequencing and annotation of themicroalgae Dunaliella tertiolecta Pathway description and gene discovery for production of next-generationbiofuels BMC Genom 2011 12 148 [CrossRef] [PubMed]
84 Ramos MJ Fernaacutendez CM Casas A Rodriacuteguez L Peacuterez A Influence of fatty acid composition of rawmaterials on biodiesel properties Bioresour Technol 2009 100 261ndash268 [CrossRef] [PubMed]
85 Knothe G Dependence of biodiesel fuel properties on the structure of fatty acid alkyl estersFuel Process Technol 2005 86 1059ndash1070 [CrossRef]
86 Knothe G ldquoDesignerrdquo Biodiesel Optimizing Fatty Ester Composition to Improve Fuel PropertiesEnergy Fuels 2008 22 1358ndash1364 [CrossRef]
87 Smith PC Ngothai Y Dzuy Nguyen Q OrsquoNeill BK Improving the low-temperature properties ofbiodiesel Methods and consequences Renew Energy 2010 35 1145ndash1151 [CrossRef]
88 Dunn RO Effect of antioxidants on the oxidative stability of methyl soyate (biodiesel) Fuel Process Technol2005 86 1071ndash1085 [CrossRef]
89 Knothe G Improving biodiesel fuel properties by modifying fatty ester composition Energy Environ Sci2009 2 759ndash766 [CrossRef]
copy 2017 by the authors licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)

Energies 2017 10 224 8 of 16
32 Microalgae Cultivation Biomass and Total Lipids Measurements
The isolated microalgae strains exhibited remarkable differences in productivity parametersand lipid content among individual strains in the stationary growth phase (Table 1) With regardto the biomass productivity the 14 strains showed statistically-significant differences (F = 71911df = 13 p lt 0001) and ranged from 36 plusmn 04 mgmiddotLminus1middotdminus1 (Ankistrodesmus sp strain AMA002) to316 plusmn 04 mgmiddotLminus1middotdminus1 (Acutodesmus obliquus strain AMA004) Similar biomass productivities werereported for other marine microalgae species [54] However other investigations show major biomassproductivities (ranging from 30 to 120 mgmiddotLminus1middotdminus1) in the same and other microalgae genera [455556]These discrepancies may be attributed to differences in the culture conditions in the culture mediacomposition and in the microalgae genotypes Lipid productivity also exhibited statistically-significantdifferences (F = 4774 df = 13 p lt 0001) and ranged from 10plusmn 04 mgmiddotLminus1middotdminus1 to 64 plusmn 07 mgmiddotLminus1middotdminus1
with strains AMA002 and AMA004 respectively These lipid productivities were low in comparison toother reports which reported values from 190 to 433 mgmiddotLminus1middotdminus1 [4555] Furthermore lipid contentas a percentage of the microalgal biomass dry weight showed statistically-significant differences(F = 8509 df = 13 p lt 0001) and ranged from 135 plusmn 12 (Tetradesmus sp strain NAN003) to467 plusmn 15 (Ankistrodesmus sp strain NAN001) These percentage values in total lipid contentare similar to several microalgae species of these genera isolated and characterized by otherresearchers worldwide [20454648ndash5057] Furthermore it is important to emphasize that lipid contentis dependent on the different growth phases Commonly in several microalgae taxa the lipid contentincreases gradually from logarithmic to stationary growth phases [58ndash60] For example Kaur et al [20]reported that the total lipid content of Scenedesmus sp DRLMA9 demonstrated values of 207 inthe late exponential phase and 397 in the stationary phase and Desmodesmus elegans DRLMA13displayed values of 103 in the late exponential phase and 169 in the stationary phase LikewiseChiu et al [61] found that the total lipid content of Nannochloropsis oculata was 308 in the logarithmicphase 397 in the early stationary phase and 504 in the stationary phase These changes in lipidcontent with culture age have been associated with nitrate depletion [61] which causes a channeling ofphotosynthetic ATP to carbohydrate and lipid biosynthesis [60] Finally Pearson correlation analysisof biomass productivity and lipid content was inversely related (r = minus037 p lt 001) which is similarto the results obtained by Liang et al [62] and Rodolfi et al [52] In contrast Li et al [51] andHempel et al [63] found no relationship between these parameters The negative correlation obtainedis likely due to the high metabolic cost of lipid biosynthesis [62]
33 Strains Culture under Nitrogen-Sufficient and Nitrogen-Deficient Medium
The microalgae species Acutodesmus obliquus (KP8785051) Ankistrodesmus sp (KP8785001)and Chlorella lewinii (KP8785011) were selected and further characterized because they displayedthe highest biomass and lipid productivities (Table 1) These microalgae showed noticeablevariations in the growth profile when cultured in Chu medium No 10 with nitrogen (+N) orwithout a nitrogen source (minusN) In +N medium all strains showed exponential growth butChlorella lewinii showed the highest average specific growth rates (micro = 0458 plusmn 0098 dminus1) followed byAcutodesmus obliquus (micro = 0399 plusmn 0112 dminus1) and the lowest value was produced by Ankistrodesmus sp(micro = 0265 plusmn 0013 dminus1) In contrast all cultures displayed a lag phase pattern in their growthprofile under minusN medium (Figure 2) These findings corroborate the results obtained by otherresearchers who demonstrate that the specific growth rate is directly proportional to the initialnitrate concentration [46596465] because the reduction or cessation of microalgae cell divisionis due to nitrogen limitation [66] which is a key nutrient for protein and nucleic acids de novobiosynthesis consequently causing an arrest in the early part of the microalgae cell cycle [6768]In accordance with these reports Li et al [69] showed that nitrogen deficiency induced in the greenmicroalgae Micractinium pusillum a downregulation in the expression of genes involved in cell growthand cell cycle regulation Additionally differences in growth profile and specific growth rates aredependent on several factors such as microalgae genotypes the culture medium composition pH
Energies 2017 10 224 9 of 16
nutrients concentration the cell culture phase (ie log phase stationary phase) and environmentalfactors [4549545770ndash72]Energies 2017 10 224 9 of 16
Figure 2 Growth profiles of three microalgal strains cultivated under +N (filled boxes) and minusN (empty boxes) Chu medium No 10 The culture was incubated in a controlled culture room at 25 plusmn 1 degC with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 150 rpm Experiments were carried out in triplicate
34 Total Protein Total Lipid and Total Carbohydrate Content
Biochemical parameters of the microalgae species such as total protein total lipid and total carbohydrate content exhibited differences and changes under both culture conditions (Figure 3) For example Chlorella lewinii showed the highest protein content Ankistrodesmus sp had the highest total lipid content but Acutodesmus obliquus presented the highest levels of total carbohydrate Therefore these strains can be valuable stocks for the industry because total protein and total carbohydrate are potentially byproducts of microalgal biomass processing [7374] Additionally the three strains cultured under minusN conditions showed a significant increase in total lipid content (from 237 in Acutodesmus obliquus to 667 in Ankistrodesmus sp) and total carbohydrate content (from 119 in Acutodesmus obliquus to 963 in Chlorella lewinii) but showed a decreased in total protein levels (from 242 in Acutodesmus obliquus to 545 in Chlorella lewinii) in comparison to +N cultures (Figure 3) These results are in accordance with well-documented results obtained by other researchers with several microalgae species of the genera Acutodesmus Ankistrodesmus Botryococcus Chlamydomonas Chlorella Desmodesmus Dunaliella Haematococcus Neochloris Phaeodactylum Pseudokirchneriella and Scenedesmus [465058596475ndash80] Nevertheless the accumulation of total lipid induced by nitrogen deficiency is not correlated with lipid productivity but rather is due to a decline and stopping of the microalgae specific growth rate [75] The changes in biomolecule contents suggest redirection of microalgae metabolism during nitrogen starvation such as a decrease in biosynthesisincrease in protein degradation and an increase in the biosynthesis of storage biomolecules such as triacylglycerols and starch The increase in total lipid content could be partially explained for a boost in transcript levels of genes encoding enzymes of the lipid biosynthesis pathways According to this assumption recent investigations showed that genes encoding the enzymes that catalyze the last step in the Kennedy pathway of triacylglycerol biosynthesis [81] such as glycerol-3-phosphate acyltransferase and acyl-glycerol-3-phosphate acyltransferase are upregulated in response to nitrogen starvation [648283] Furthermore the increase in total carbohydrate content can be explained by changes in the expression levels of genes involved in carbohydrate metabolism For example when the oleaginous microalgae Neochloris oleoabundans is cultured under nitrogen starvation the catabolic phase of starch specifically α-amylase which hydrolyzes starch to glucose is strongly repressed [83]
Figure 2 Growth profiles of three microalgal strains cultivated under +N (filled boxes) and minusN(empty boxes) Chu medium No 10 The culture was incubated in a controlled culture room at25 plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent lightand continuous agitation at 150 rpm Experiments were carried out in triplicate
34 Total Protein Total Lipid and Total Carbohydrate Content
Biochemical parameters of the microalgae species such as total protein total lipid and totalcarbohydrate content exhibited differences and changes under both culture conditions (Figure 3)For example Chlorella lewinii showed the highest protein content Ankistrodesmus sp had the highesttotal lipid content but Acutodesmus obliquus presented the highest levels of total carbohydrateTherefore these strains can be valuable stocks for the industry because total protein and totalcarbohydrate are potentially byproducts of microalgal biomass processing [7374] Additionallythe three strains cultured under minusN conditions showed a significant increase in total lipid content(from 237 in Acutodesmus obliquus to 667 in Ankistrodesmus sp) and total carbohydrate content(from 119 in Acutodesmus obliquus to 963 in Chlorella lewinii) but showed a decreased in totalprotein levels (from 242 in Acutodesmus obliquus to 545 in Chlorella lewinii) in comparison to +Ncultures (Figure 3) These results are in accordance with well-documented results obtained by otherresearchers with several microalgae species of the genera Acutodesmus Ankistrodesmus BotryococcusChlamydomonas Chlorella Desmodesmus Dunaliella Haematococcus Neochloris PhaeodactylumPseudokirchneriella and Scenedesmus [465058596475ndash80] Nevertheless the accumulation of totallipid induced by nitrogen deficiency is not correlated with lipid productivity but rather is due toa decline and stopping of the microalgae specific growth rate [75] The changes in biomoleculecontents suggest redirection of microalgae metabolism during nitrogen starvation such as a decreasein biosynthesisincrease in protein degradation and an increase in the biosynthesis of storagebiomolecules such as triacylglycerols and starch The increase in total lipid content could bepartially explained for a boost in transcript levels of genes encoding enzymes of the lipid biosynthesispathways According to this assumption recent investigations showed that genes encoding theenzymes that catalyze the last step in the Kennedy pathway of triacylglycerol biosynthesis [81] such asglycerol-3-phosphate acyltransferase and acyl-glycerol-3-phosphate acyltransferase are upregulated
Energies 2017 10 224 10 of 16
in response to nitrogen starvation [648283] Furthermore the increase in total carbohydrate contentcan be explained by changes in the expression levels of genes involved in carbohydrate metabolismFor example when the oleaginous microalgae Neochloris oleoabundans is cultured under nitrogenstarvation the catabolic phase of starch specifically α-amylase which hydrolyzes starch to glucoseis strongly repressed [83]Energies 2017 10 224 10 of 16
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strains cultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled culture room at 25 plusmn 1 degC with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized with respect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that each microalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majority of fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised 7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii the majority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9) acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectively Finally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic (182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in +N and minusN conditions respectively Our results agree with multiple previously-published reports in that fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent in many green microalgae species [454648ndash5055] It is notable that under nitrogen starvation culture all microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 in Ankistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid (181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findings support the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content of SFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Nevertheless there are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed a decrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increased between 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively However in the same culture condition the three microalgae species showed a decrease in polyunsaturated fatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmus spp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgae strains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFA which improves the quality properties of biodiesel such as an increase
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strainscultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled cultureroom at 25plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescentlight and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized withrespect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that eachmicroalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majorityof fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii themajority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9)acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectivelyFinally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic(182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in+N and minusN conditions respectively Our results agree with multiple previously-published reports inthat fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent inmany green microalgae species [454648ndash5055] It is notable that under nitrogen starvation cultureall microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 inAnkistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid(181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findingssupport the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content ofSFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other
Energies 2017 10 224 11 of 16
researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Neverthelessthere are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed adecrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increasedbetween 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively Howeverin the same culture condition the three microalgae species showed a decrease in polyunsaturatedfatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmusspp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgaestrains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFAwhich improves the quality properties of biodiesel such as an increase in cetane number [84ndash86]Considering the SFA and unsaturated fatty acids (UFA) components approximately 20ndash31 are SFAand 69ndash80 are UFA Hence the SFAUFA ratio in the microalgae strains evaluated ranges between025 and 045 which is similar to the reports of several microalgae species isolated in different latitudesworldwide [203148497072]
Table 2 Effect of nitrogen deficiency on fatty acid profiling in three native microalgal species from thePeruvian Amazon
Fatty AcidsAcutodesmus obliquus Ankistrodesmus sp Chlorella lewinii
+N minusN +N minusN +N minusN
140 - - 052 plusmn 002 088 plusmn 002 - 058 plusmn 001160 2724 plusmn 007 2867 plusmn 017 1868 plusmn 019 2261 plusmn 006 1964 plusmn 021 2123 plusmn 010180 184 plusmn 005 255 plusmn 051 101 plusmn 002 181 plusmn 002 371 plusmn 026 153 plusmn 015
161n-7 023 plusmn 000 025 plusmn 001 071 plusmn 003 055 plusmn 001 085 plusmn 043 038 plusmn 000181n-7 131 plusmn 002 131 plusmn 000 141 plusmn 005 094 plusmn 003 146 plusmn 006 123 plusmn 002181n-9 3213 plusmn 021 3608 plusmn 021 3699 plusmn 004 4047 plusmn 038 1371 plusmn 015 2326 plusmn 041182n-6 1732 plusmn 004 1651 plusmn 016 747 plusmn 018 656 plusmn 007 2034 plusmn 039 1957 plusmn 001183n-3 1130 plusmn 017 854 plusmn 031 1486 plusmn 004 1173 plusmn 012 1815 plusmn 022 1371 plusmn 016183n-6 101 plusmn 001 078 plusmn 002 055 plusmn 000 054 plusmn 002 124 plusmn 006 100 plusmn 020184n-3 250 plusmn 005 177 plusmn 008 631 plusmn 002 601 plusmn 005 491 plusmn 025 403 plusmn 020204n-6 - - - 084 plusmn 008 - -225n-3 - - - 062 plusmn 002 - -
Unidentified 512 plusmn 003 355 plusmn 006 1149 plusmn 004 645 plusmn 003 1599 plusmn 034 1348 plusmn 006SFA 2907 3122 2022 2531 2335 2334UFA 7093 6878 7978 7469 7665 7666
MUFA 3367 3764 3911 4195 1602 2487PUFA 3213 2759 2918 2629 4464 3831SFA +MUFA 6274 6886 5933 6726 3936 4821
It is important to highlight that the fatty acid profile determination is an essential step inthe process of the characterization of microalgae strains because it ultimately affects the qualityof the biodiesel product [8485] The quality properties of biodiesel such as high cetane numberoxidative stability and cold-flow are dependent on the hydrocarbonated chain length of saturated andunsaturated fatty acids [8587] For example biodiesel composed of a mixture of a high percentage ofSFA and MUFA is preferred for increased energy yield superior oxidative stability and higher cetanenumbers but is prone to solidify at low temperature In contrast biodiesel with high levels of PUFAhas decreased energy yield low cetane numbers and excellent cold-flow properties but insufficientoxidative stability [548588] Therefore to solve these performance problems five approaches havebeen developed such as changing the fatty acids profile by physical means genetic modification ofthe feedstock or the use of alternative feedstocks with different fatty acids profiles [89]
4 Conclusions
Fourteen microalgae were isolated from three river basins from the Peruvian Amazon and wereidentified by morphological features and molecular phylogeny approaches These strains were initially
Energies 2017 10 224 12 of 16
characterized in the stationary growth phase and exhibited significant heterogeneity in key parameterssuch as biomass productivity (36ndash316 mgmiddotLminus1middotdminus1) lipid productivity (10ndash64 mgmiddotLminus1middotdminus1) andtotal lipid content (135ndash467) Furthermore three selected strains based on elevated biomass andlipid productivities were cultured under nitrogen-sufficient and nitrogen-deficient medium to boostlipid accumulation Under these culture conditions all strains showed noticeable decreases in totalprotein content and increases in total lipid and total carbohydrate contents In addition these strainspresented modifications in the fatty acids profiles with improvement in biodiesel properties due to apredominance of SFA and MUFA which corresponds to an increase in cetane number The results ofthis research suggest that the isolated native microalgae from the Peruvian Amazon could be suitablerenewable feedstocks for the production of both biodiesel and bioethanol which could contribute tosustainable development at the local regional and national levels as a viable alternative to petroleumexploitation However it will be necessary to develop open closed or hybrid systems for large-scalebiomass production of selected native microalgae strains andor de novo microalgae strains designedto produce biofuels and with the abilities to synthesize useful bioactive compounds to be efficientlyexploited using biorefinery approaches
Supplementary MaterialsThe following are available online at wwwmdpicom1996-1073102224s1 Figure S1 Results of
electrophoretic and spectrophotometric analysis of the genomic DNA isolated from the fourteen microalgaeFigure S2 ITS2-rDNA structure (A) and amplicons obtained (B) from the microalgae isolated Figure S3Microphotographic images of the three microalgae strains analyzed under light and fluorescence stained withNile Red Figure S4 Spectrophotometric scanning for the determination of the maximum absorption peak foreach microalgae strain
Acknowledgments This research was supported by the Peruvian funding agencies Consejo Nacional de CienciaTecnologiacutea e Innovacioacuten Tecnoloacutegica (CONCYTEC) with Grant Contract No 364-CONCYTEC-OAJ and Fondospara la Innovacioacuten Ciencia y Tecnologiacutea (FINCyT) with Grant Contract No 383-PNICP-PIBA-2014
Author Contributions Marianela Cobos and Juan C Castro obtained funds for the research participated inthe study design coordinated activities and participated in the preparation of the manuscript Jae D Paredesparticipated in sample collection microalgae isolation and helped to draft the manuscript J Dylan Maddoxperformed the bioinformatics analysis and participated in the preparation of the manuscript Gabriel Vargas-Aranaand Leenin Flores participated in biochemical analysis and helped to draft the manuscript Carla P Aguilarparticipated in the study design supervised biochemical analysis and helped to draft the manuscriptJorge L Marapara participated in molecular analysis and helped to draft the manuscript
Conflicts of Interest The authors declare no conflict of interest
References
1 BP Global BP Statisical Review of World Energy 2014 Available online httpwwwbpcomcontentdambppdfEnergy-economicsstatistical-review-2014BP-statistical-review-of-world-energy-2014-full-reportpdf (accessed on 20 June 2014)
2 Pachauri RK Reisinger A Cambio Climaacutetico 2007 Informe de Siacutentesis Contribucioacuten de los Grupos de Trabajo III y III al Cuarto Informe de Evaluacioacuten del Grupo Intergubernamental de Expertos Sobre el Cambio Climaacutetico IPCCGeneva Switzerland 2007 p 104
3 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Ley de Promocioacuten del Mercado deBiocombustibles Available online httpwww2congresogobpesicrcendocbibcon4_uibdnsf9E95620CC059138105257C9E005AB2F9$FILE28054pdf (accessed on 12 February 2017)
4 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Reglamento de la Ley de Promocioacuten del Mercadode Biocombustibles Available online httpwww2osinerggobpeMarcoLegaldocrevDS-013-2005-EM-CONCORDADOpdf (accessed on 12 February 2017)
5 Areacutevalo LF Nalvarte J Torres J Ramiacuterez Y Impactos Socio-Econoacutemicos de la Produccioacuten de Biocombustiblesen la Amazoniacutea Peruana Primera SNV-WWF Lima Peruacute 2009
6 Sharma KK Schuhmann H Schenk PM High Lipid Induction in Microalgae for Biodiesel ProductionEnergies 2012 5 1532ndash1553 [CrossRef]
Energies 2017 10 224 13 of 16
7 Schenk PM Thomas-Hall SR Stephens E Marx UC Mussgnug JH Posten C Kruse O Hankamer BSecond Generation Biofuels High-Efficiency Microalgae for Biodiesel Production BioEnergy Res 2008 120ndash43 [CrossRef]
8 Foley JA Asner GP Costa MH Coe MT de Fries R Gibbs HK Howard EA Olson S Patz JRamankutty N et al Amazonia revealed Forest degradation and loss of ecosystem goods and services inthe Amazon Basin Front Ecol Environ 2007 5 25ndash32 [CrossRef]
9 Duong VT Li Y Nowak E Schenk PM Microalgae Isolation and Selection for Prospective BiodieselProduction Energies 2012 5 1835ndash1849 [CrossRef]
10 Georgianna DR Mayfield SP Exploiting diversity and synthetic biology for the production of algalbiofuels Nature 2012 488 329ndash335 [CrossRef] [PubMed]
11 Wijffels RH Barbosa MJ An Outlook on Microalgal Biofuels Science 2010 329 796ndash799 [CrossRef][PubMed]
12 Richmond A Handbook of Microalgal Culture Biotechnology and Applied Phycology Blackwell Sci Ltd HudsonNJ USA 2004
13 Menetrez MY An overview of algae biofuel production and potential environmental impactEnviron Sci Technol 2012 46 7073ndash7085 [CrossRef] [PubMed]
14 Khoo HH Sharratt PN Das P Balasubramanian RK Naraharisetti PK Shaik S Life cycle energy andCO2 analysis of microalgae-to-biodiesel Preliminary results and comparisons Bioresour Technol 2011 1025800ndash5807 [CrossRef] [PubMed]
15 Chu SP The Influence of the Mineral Composition of the Medium on the Growth of Planktonic AlgaePart I Methods and Culture Media J Ecol 1942 30 284ndash325 [CrossRef]
16 Carlos EMB Mariatildengela M Gecircneros de Algas de Aacuteguas Continentais do BrasilmdashChave Para Identificaccedilatildeo eDescriccedilotildees 2nd ed Rima Sao Carlos Brasil 2006
17 Greenspan P Mayer EP Fowler SD Nile red A selective fluorescent stain for intracellular lipid dropletsJ Cell Biol 1985 100 965ndash973 [CrossRef] [PubMed]
18 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissuePhytochem Bull 1987 19 11ndash15
19 Sambrook J Fritsch EF Maniatis T Molecular Cloning A Laboratory Manual 2nd ed Cold Spring HarborLaboratory Press New York NY USA 1989
20 Kaur S Sarkar M Srivastava RB Gogoi HK Kalita MC Fatty acid profiling and molecularcharacterization of some freshwater microalgae from India with potential for biodiesel productionNew Biotechnol 2012 29 332ndash344 [CrossRef] [PubMed]
21 Benson DA Cavanaugh M Clark K Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW GenBankNucleic Acids Res 2013 41 D36ndashD42 [CrossRef] [PubMed]
22 NCBI Resource Coordinators Database Resources of the National Center for Biotechnology InformationNucleic Acids Res 2015 43 D6ndashD17
23 Ye J McGinnis S Madden TL BLAST Improvements for better sequence analysis Nucleic Acids Res 200634 W6ndashW9 [CrossRef] [PubMed]
24 BioEdit Sequence Alignment Editor for Windows 9598NTXPVista7 Available online httpwwwmbioncsuedubioeditbioedithtml (accessed on 2 December 2015)
25 Sievers F Higgins DG Clustal omega accurate alignment of very large numbers of sequencesMethods Mol Biol (Clifton NJ) 2014 1079 105ndash116
26 Saitou N Nei M The neighbor-joining method A new method for reconstructing phylogenetic treesMol Biol Evol 1987 4 406ndash425 [PubMed]
27 Kimura M A simple method for estimating evolutionary rates of base substitutions through comparativestudies of nucleotide sequences J Mol Evol 1980 16 111ndash120 [CrossRef] [PubMed]
28 Tamura K Stecher G Peterson D Filipski A Kumar S MEGA6 Molecular Evolutionary GeneticsAnalysis version 60 Mol Biol Evol 2013 30 2725ndash2729 [CrossRef] [PubMed]
29 Felsenstein J Confidence Limits on Phylogenies An Approach Using the Bootstrap Evolution 1985 39783ndash791 [CrossRef]
30 Bligh EG Dyer WJ A Rapid Method of Total Lipid Extraction and Purification Can J Biochem Physiol1959 37 911ndash917 [CrossRef] [PubMed]
Energies 2017 10 224 14 of 16
31 Yu X Zhao P He C Li J Tang X Zhou J Huang Z Isolation of a novel strain of Monoraphidium spand characterization of its potential application as biodiesel feedstock Bioresour Technol 2012 121 256ndash262[CrossRef] [PubMed]
32 Hartree EF Determination of protein A modification of the lowry method that gives a linear photometricresponse Anal Biochem 1972 48 422ndash427 [CrossRef]
33 DuBois M Gilles KA Hamilton JK Rebers PA Smith F Colorimetric Method for Determination ofSugars and Related Substances Anal Chem 1956 28 350ndash356 [CrossRef]
34 AOAC International AOAC Official Methods of Analysis of the Association of Official Analytical Chemists (AOAC)15th ed AOAC International Washington DC USA 1990 Volume 1
35 Ichihara K Fukubayashi Y Preparation of fatty acid methyl esters for gas-liquid chromatographyJ Lipid Res 2010 51 635ndash640 [CrossRef] [PubMed]
36 Brennan L Owende P Biofuels from microalgaemdashA review of technologies for production processingand extractions of biofuels and co-products Renew Sustain Energ Rev 2010 14 557ndash577 [CrossRef]
37 Raja R Hemaiswarya S Kumar NA Sridhar S Rengasamy R A perspective on the biotechnologicalpotential of microalgae Crit Rev Microbiol 2008 34 77ndash88 [CrossRef] [PubMed]
38 Mata TM Martins AA Caetano NS Microalgae for biodiesel production and other applicationsA review Renew Sustain Energy Rev 2010 14 217ndash232 [CrossRef]
39 De Wilt A Butkovskyi A Tuantet K Leal LH Fernandes TV Langenhoff A Zeeman GMicropollutant removal in an algal treatment system fed with source separated wastewater streamsJ Hazard Mater 2015 304 84ndash92 [CrossRef] [PubMed]
40 Li T Lin G Podola B Melkonian M Continuous removal of zinc from wastewater and mine dumpleachate by a microalgal biofilm PSBR J Hazard Mater 2015 297 112ndash118 [CrossRef] [PubMed]
41 Sioli H The Amazon and its main affluents Hydrography morphology of the river courses and river typesIn Monographiae Biologicae Sioli H Ed Springer Dordrecht The Netherlands 1984 pp 127ndash165
42 Moniz MBJ Kaczmarska I Barcoding of Diatoms Nuclear Encoded ITS Revisited Protist 2010 161 7ndash34[CrossRef] [PubMed]
43 Coleman AW Is there a molecular key to the level of ldquobiological speciesrdquo in eukaryotes A DNA guideMol Phylogenet Evol 2009 50 197ndash203 [CrossRef] [PubMed]
44 Mai JC Coleman AW The Internal Transcribed Spacer 2 Exhibits a Common Secondary Structure inGreen Algae and Flowering Plants J Mol Evol 1997 44 258ndash271 [CrossRef] [PubMed]
45 Abou-Shanab RAI Matter IA Kim S-N Oh Y-K Choi J Jeon B-H Characterization andidentification of lipid-producing microalgae species isolated from a freshwater lake Biomass Bioenergy2011 35 3079ndash3085 [CrossRef]
46 Do Nascimento M Ortiz-Marquez JCF Sanchez-Rizza L Echarte MM Curatti L Bioprospectingfor fast growing and biomass characterization of oleaginous microalgae from South-Eastern Buenos AiresArgentina Bioresour Technol 2012 125 283ndash290 [CrossRef] [PubMed]
47 Araujo GS Matos LJBL Gonccedilalves LRB Fernandes FAN Farias WRL Bioprospecting foroil producing microalgal strains Evaluation of oil and biomass production for ten microalgal strainsBioresour Technol 2011 102 5248ndash5250 [CrossRef] [PubMed]
48 Zhou W Li Y Min M Hu B Chen P Ruan R Local bioprospecting for high-lipid producing microalgalstrains to be grown on concentrated municipal wastewater for biofuel production Bioresour Technol 2011102 6909ndash6919 [CrossRef] [PubMed]
49 Mahmoud EA Farahat LA Abdel Aziz ZK Fatthallah NA Salah El Din RA Evaluation of thepotential for some isolated microalgae to produce biodiesel Egypt J Pet 2015 24 97ndash101 [CrossRef]
50 Ho S Lai Y-Y Chiang C-Y Chen C-NN Chang J-S Selection of elite microalgae for biodieselproduction in tropical conditions using a standardized platform Bioresour Technol 2013 147 135ndash142[CrossRef] [PubMed]
51 Li L Cui J Liu Q Ding Y Liu J Screening and phylogenetic analysis of lipid-rich microalgae Algal Res2015 11 381ndash386 [CrossRef]
52 Rodolfi L Chini Zittelli G Bassi N Padovani G Biondi N Bonini G Tredici MR Microalgae for oilStrain selection induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactorBiotechnol Bioeng 2009 102 100ndash112 [CrossRef] [PubMed]
Energies 2017 10 224 15 of 16
53 Li P Miao X Li R Zhong J In situ biodiesel production from fast-growing and high oil content Chlorellapyrenoidosa in rice straw hydrolysate J Biomed Biotechnol 2011 2011 141207 [CrossRef] [PubMed]
54 Doan TTY Sivaloganathan B Obbard JP Screening of marine microalgae for biodiesel feedstockBiomass Bioenergy 2011 35 2534ndash2544 [CrossRef]
55 Abou-Shanab RAI Hwang J-H Cho Y Min B Jeon B-H Characterization of microalgal speciesisolated from fresh water bodies as a potential source for biodiesel production Appl Energy 2011 883300ndash3306 [CrossRef]
56 Chaichalerm S Pokethitiyook P Yuan W Meetam M Sritong K Pugkaew W Kungvansaichol KKruatrachue M Damrongphol P Culture of microalgal strains isolated from natural habitats in Thailand invarious enriched media Appl Energy 2012 89 296ndash302 [CrossRef]
57 Zhang S Liu P Yang X Hao Z Zhang L Luo N Shi J Isolation and identification by 18S rDNAsequence of high lipid potential microalgal species for fuel production in Hainan Dao Biomass Bioenergy2014 66 197ndash203 [CrossRef]
58 Chiu S-Y Kao C-Y Tsai M-T Ong S-C Chen C-H Lin C-S Lipid accumulation and CO2 utilizationof Nannochloropsis oculata in response to CO2 aeration Bioresour Technol 2009 100 833ndash838 [CrossRef][PubMed]
59 Huerlimann R de Nys R Heimann K Growth lipid content productivity and fatty acid composition oftropical microalgae for scale-up production Biotechnol Bioeng 2010 107 245ndash257 [CrossRef] [PubMed]
60 Liang Y Sarkany N Cui Y Biomass and lipid productivities of Chlorella vulgaris under autotrophicheterotrophic and mixotrophic growth conditions Biotechnol Lett 2009 31 1043ndash1049 [CrossRef] [PubMed]
61 Hempel N Petrick I Behrendt F Biomass productivity and productivity of fatty acids and amino acidsof microalgae strains as key characteristics of suitability for biodiesel production J Appl Phycol 2012 241407ndash1418 [CrossRef] [PubMed]
62 Yeesang C Cheirsilp B Effect of nitrogen salt and iron content in the growth medium and light intensityon lipid production by microalgae isolated from freshwater sources in Thailand Bioresour Technol 2011 1023034ndash3040 [CrossRef] [PubMed]
63 Msanne J Xu D Konda AR Casas-Mollano JA Awada T Cahoon EB Cerutti H Metabolic andgene expression changes triggered by nitrogen deprivation in the photoautotrophically grown microalgaeChlamydomonas reinhardtii and Coccomyxa sp C-169 Phytochemistry 2012 75 50ndash59 [CrossRef] [PubMed]
64 Lin Q Lin J Effects of nitrogen source and concentration on biomass and oil production of aScenedesmus rubescens like microalga Bioresour Technol 2011 102 1615ndash1621 [CrossRef] [PubMed]
65 Sheehan J Dunahay T Benemann J Roessler P A Look Back at the US Department of Energyrsquos AquaticSpecies ProgrammdashBiodiesel from Algae US Department of Energyrsquos Office of Fuels Development Golden COUSA 1998 p 328
66 Vaulot D Olson RJ Merkel S Chisholm SW Cell-cycle response to nutrient starvation in twophytoplankton species Thalassiosira weissflogii and Hymenomonas carterae Mar Biol 1987 95 625ndash630[CrossRef]
67 Olson RJ Chisholm SW Effects of light and nitrogen limitation on the cell cycle of the dinoflagellateAmphidinium carteri J Plankton Res 1986 8 785ndash793 [CrossRef]
68 Li Y Fei X Deng X Novel molecular insights into nitrogen starvation-induced triacylglycerolsaccumulation revealed by differential gene expression analysis in green algae Micractinium pusillumBiomass Bioenergy 2012 42 199ndash211 [CrossRef]
69 Abdelaziz AEM Leite GB Belhaj MA Hallenbeck PC Screening microalgae native to Quebec forwastewater treatment and biodiesel production Bioresour Technol 2014 157 140ndash148 [CrossRef] [PubMed]
70 Valdez-Ojeda R Gonzaacutelez-Muntildeoz M Us-Vaacutezquez R Narvaacuteez-Zapata J Chavarria-Hernandez JCLoacutepez-Adriaacuten S Barahona-Peacuterez F Toledano-Thompson T Garduntildeo-Soloacuterzano G Escobedo-GraciaMedrano RM Characterization of five fresh water microalgae with potential for biodiesel productionAlgal Res 2015 7 33ndash44 [CrossRef]
71 Shrivastav A Mishra SK Suh WI Farooq W Moon M Kim T-H Kumar K Choi G-G Park MSYang J-W Characterization of newly isolated oleaginous microalga Monoraphidium sp for lipid productionunder different conditions Algal Res 2015 12 289ndash294 [CrossRef]
72 Yen H-W Hu I-C Chen C-Y Ho S-H Lee D-J Chang J-S Microalgae-based biorefinerymdashFrombiofuels to natural products Bioresour Technol 2013 135 166ndash174 [CrossRef] [PubMed]
Energies 2017 10 224 16 of 16
73 Li J Liu Y Cheng JJ Mos M Daroch M Biological potential of microalgae in China for biorefinery-basedproduction of biofuels and high value compounds New Biotechnol 2015 32 588ndash596 [CrossRef] [PubMed]
74 El-Sheekh M Abomohra AE-F Hanelt D Optimization of biomass and fatty acid productivity ofScenedesmus obliquus as a promising microalga for biodiesel production World J Microbiol Biotechnol 201329 915ndash922 [CrossRef] [PubMed]
75 Singh P Guldhe A Kumari S Rawat I Bux F Investigation of combined effect of nitrogen phosphorusand iron on lipid productivity of microalgae Ankistrodesmus falcatus KJ671624 using response surfacemethodology Biochem Eng J 2015 94 22ndash29 [CrossRef]
76 Kilham S Kreeger D Goulden C Lynn S Effects of nutrient limitation on biochemical constituents ofAnkistrodesmus falcatus Freshw Biol 1997 38 591ndash596 [CrossRef]
77 Breuer G Lamers PP Martens DE Draaisma RB Wijffels RH The impact of nitrogen starvation on thedynamics of triacylglycerol accumulation in nine microalgae strains Bioresour Technol 2012 124 217ndash226[CrossRef] [PubMed]
78 Breuer G de Jaeger L Artus VG Martens DE Springer J Draaisma RB Eggink G Wijffels RHLamers PP Superior triacylglycerol (TAG) accumulation in starchless mutants of Scenedesmus obliquus (II)evaluation of TAG yield and productivity in controlled photobioreactors Biotechnol Biofuels 2014 7 70[CrossRef] [PubMed]
79 Nigam S Prakash-Rai M Sharma R Effect of Nitrogen on Growth and Lipid Content ofChlorella pyrenoidosa Am J Biochem Biotechnol 2011 7 126ndash131 [CrossRef]
80 Piligaev AV Sorokina KN Bryanskaya AV Peltek SE Kolchanov NA Parmon VN Isolation ofprospective microalgal strains with high saturated fatty acid content for biofuel production Algal Res 201512 368ndash376 [CrossRef]
81 Weiss SB Kennedy EP The enzymatic synthesis of triglycerides J Am Chem Soc 1956 78 3550[CrossRef]
82 Miller R Wu G Deshpande RR Vieler A Gaumlrtner K Li X Moellering ER Zaumluner S Cornish AJLiu B et al Changes in Transcript Abundance in Chlamydomonas reinhardtii following Nitrogen DeprivationPredict Diversion of Metabolism Plant Physiol 2010 154 1737ndash1752 [CrossRef] [PubMed]
83 Rismani-Yazdi H Haznedaroglu BZ Bibby K Peccia J Transcriptome sequencing and annotation of themicroalgae Dunaliella tertiolecta Pathway description and gene discovery for production of next-generationbiofuels BMC Genom 2011 12 148 [CrossRef] [PubMed]
84 Ramos MJ Fernaacutendez CM Casas A Rodriacuteguez L Peacuterez A Influence of fatty acid composition of rawmaterials on biodiesel properties Bioresour Technol 2009 100 261ndash268 [CrossRef] [PubMed]
85 Knothe G Dependence of biodiesel fuel properties on the structure of fatty acid alkyl estersFuel Process Technol 2005 86 1059ndash1070 [CrossRef]
86 Knothe G ldquoDesignerrdquo Biodiesel Optimizing Fatty Ester Composition to Improve Fuel PropertiesEnergy Fuels 2008 22 1358ndash1364 [CrossRef]
87 Smith PC Ngothai Y Dzuy Nguyen Q OrsquoNeill BK Improving the low-temperature properties ofbiodiesel Methods and consequences Renew Energy 2010 35 1145ndash1151 [CrossRef]
88 Dunn RO Effect of antioxidants on the oxidative stability of methyl soyate (biodiesel) Fuel Process Technol2005 86 1071ndash1085 [CrossRef]
89 Knothe G Improving biodiesel fuel properties by modifying fatty ester composition Energy Environ Sci2009 2 759ndash766 [CrossRef]
copy 2017 by the authors licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
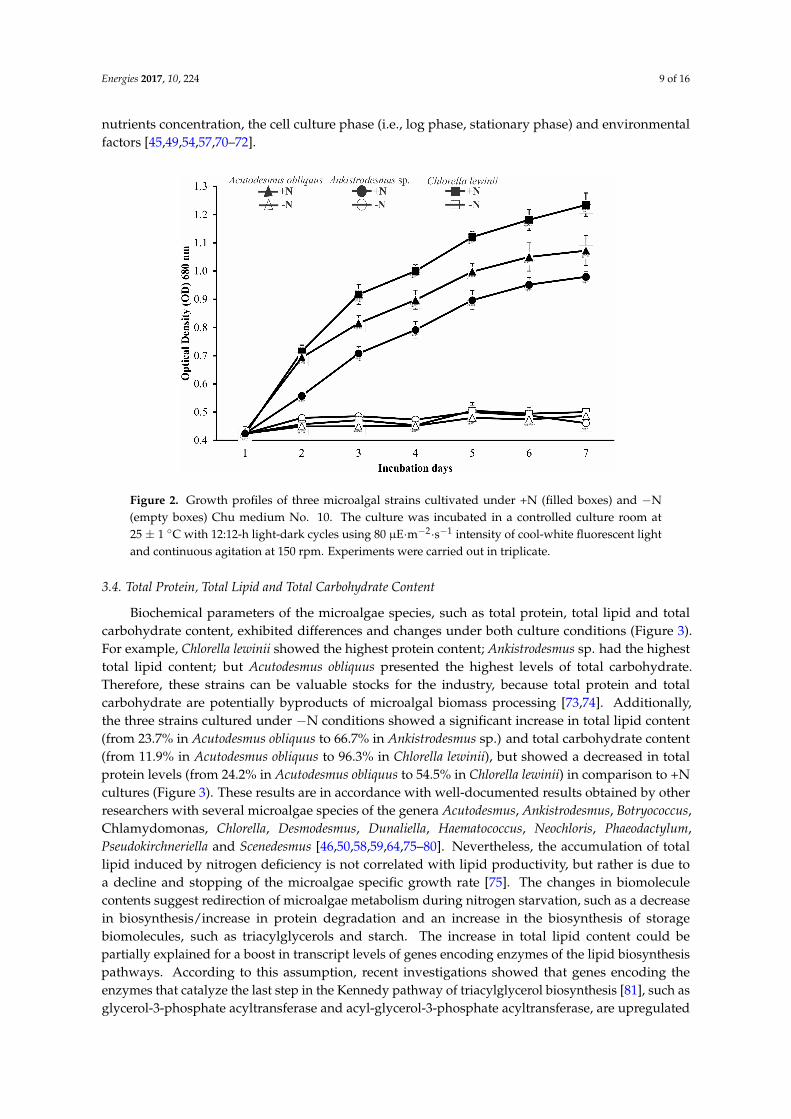
Energies 2017 10 224 9 of 16
nutrients concentration the cell culture phase (ie log phase stationary phase) and environmentalfactors [4549545770ndash72]Energies 2017 10 224 9 of 16
Figure 2 Growth profiles of three microalgal strains cultivated under +N (filled boxes) and minusN (empty boxes) Chu medium No 10 The culture was incubated in a controlled culture room at 25 plusmn 1 degC with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 150 rpm Experiments were carried out in triplicate
34 Total Protein Total Lipid and Total Carbohydrate Content
Biochemical parameters of the microalgae species such as total protein total lipid and total carbohydrate content exhibited differences and changes under both culture conditions (Figure 3) For example Chlorella lewinii showed the highest protein content Ankistrodesmus sp had the highest total lipid content but Acutodesmus obliquus presented the highest levels of total carbohydrate Therefore these strains can be valuable stocks for the industry because total protein and total carbohydrate are potentially byproducts of microalgal biomass processing [7374] Additionally the three strains cultured under minusN conditions showed a significant increase in total lipid content (from 237 in Acutodesmus obliquus to 667 in Ankistrodesmus sp) and total carbohydrate content (from 119 in Acutodesmus obliquus to 963 in Chlorella lewinii) but showed a decreased in total protein levels (from 242 in Acutodesmus obliquus to 545 in Chlorella lewinii) in comparison to +N cultures (Figure 3) These results are in accordance with well-documented results obtained by other researchers with several microalgae species of the genera Acutodesmus Ankistrodesmus Botryococcus Chlamydomonas Chlorella Desmodesmus Dunaliella Haematococcus Neochloris Phaeodactylum Pseudokirchneriella and Scenedesmus [465058596475ndash80] Nevertheless the accumulation of total lipid induced by nitrogen deficiency is not correlated with lipid productivity but rather is due to a decline and stopping of the microalgae specific growth rate [75] The changes in biomolecule contents suggest redirection of microalgae metabolism during nitrogen starvation such as a decrease in biosynthesisincrease in protein degradation and an increase in the biosynthesis of storage biomolecules such as triacylglycerols and starch The increase in total lipid content could be partially explained for a boost in transcript levels of genes encoding enzymes of the lipid biosynthesis pathways According to this assumption recent investigations showed that genes encoding the enzymes that catalyze the last step in the Kennedy pathway of triacylglycerol biosynthesis [81] such as glycerol-3-phosphate acyltransferase and acyl-glycerol-3-phosphate acyltransferase are upregulated in response to nitrogen starvation [648283] Furthermore the increase in total carbohydrate content can be explained by changes in the expression levels of genes involved in carbohydrate metabolism For example when the oleaginous microalgae Neochloris oleoabundans is cultured under nitrogen starvation the catabolic phase of starch specifically α-amylase which hydrolyzes starch to glucose is strongly repressed [83]
Figure 2 Growth profiles of three microalgal strains cultivated under +N (filled boxes) and minusN(empty boxes) Chu medium No 10 The culture was incubated in a controlled culture room at25 plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent lightand continuous agitation at 150 rpm Experiments were carried out in triplicate
34 Total Protein Total Lipid and Total Carbohydrate Content
Biochemical parameters of the microalgae species such as total protein total lipid and totalcarbohydrate content exhibited differences and changes under both culture conditions (Figure 3)For example Chlorella lewinii showed the highest protein content Ankistrodesmus sp had the highesttotal lipid content but Acutodesmus obliquus presented the highest levels of total carbohydrateTherefore these strains can be valuable stocks for the industry because total protein and totalcarbohydrate are potentially byproducts of microalgal biomass processing [7374] Additionallythe three strains cultured under minusN conditions showed a significant increase in total lipid content(from 237 in Acutodesmus obliquus to 667 in Ankistrodesmus sp) and total carbohydrate content(from 119 in Acutodesmus obliquus to 963 in Chlorella lewinii) but showed a decreased in totalprotein levels (from 242 in Acutodesmus obliquus to 545 in Chlorella lewinii) in comparison to +Ncultures (Figure 3) These results are in accordance with well-documented results obtained by otherresearchers with several microalgae species of the genera Acutodesmus Ankistrodesmus BotryococcusChlamydomonas Chlorella Desmodesmus Dunaliella Haematococcus Neochloris PhaeodactylumPseudokirchneriella and Scenedesmus [465058596475ndash80] Nevertheless the accumulation of totallipid induced by nitrogen deficiency is not correlated with lipid productivity but rather is due toa decline and stopping of the microalgae specific growth rate [75] The changes in biomoleculecontents suggest redirection of microalgae metabolism during nitrogen starvation such as a decreasein biosynthesisincrease in protein degradation and an increase in the biosynthesis of storagebiomolecules such as triacylglycerols and starch The increase in total lipid content could bepartially explained for a boost in transcript levels of genes encoding enzymes of the lipid biosynthesispathways According to this assumption recent investigations showed that genes encoding theenzymes that catalyze the last step in the Kennedy pathway of triacylglycerol biosynthesis [81] such asglycerol-3-phosphate acyltransferase and acyl-glycerol-3-phosphate acyltransferase are upregulated
Energies 2017 10 224 10 of 16
in response to nitrogen starvation [648283] Furthermore the increase in total carbohydrate contentcan be explained by changes in the expression levels of genes involved in carbohydrate metabolismFor example when the oleaginous microalgae Neochloris oleoabundans is cultured under nitrogenstarvation the catabolic phase of starch specifically α-amylase which hydrolyzes starch to glucoseis strongly repressed [83]Energies 2017 10 224 10 of 16
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strains cultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled culture room at 25 plusmn 1 degC with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized with respect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that each microalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majority of fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised 7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii the majority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9) acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectively Finally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic (182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in +N and minusN conditions respectively Our results agree with multiple previously-published reports in that fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent in many green microalgae species [454648ndash5055] It is notable that under nitrogen starvation culture all microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 in Ankistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid (181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findings support the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content of SFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Nevertheless there are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed a decrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increased between 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively However in the same culture condition the three microalgae species showed a decrease in polyunsaturated fatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmus spp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgae strains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFA which improves the quality properties of biodiesel such as an increase
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strainscultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled cultureroom at 25plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescentlight and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized withrespect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that eachmicroalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majorityof fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii themajority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9)acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectivelyFinally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic(182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in+N and minusN conditions respectively Our results agree with multiple previously-published reports inthat fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent inmany green microalgae species [454648ndash5055] It is notable that under nitrogen starvation cultureall microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 inAnkistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid(181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findingssupport the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content ofSFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other
Energies 2017 10 224 11 of 16
researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Neverthelessthere are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed adecrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increasedbetween 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively Howeverin the same culture condition the three microalgae species showed a decrease in polyunsaturatedfatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmusspp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgaestrains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFAwhich improves the quality properties of biodiesel such as an increase in cetane number [84ndash86]Considering the SFA and unsaturated fatty acids (UFA) components approximately 20ndash31 are SFAand 69ndash80 are UFA Hence the SFAUFA ratio in the microalgae strains evaluated ranges between025 and 045 which is similar to the reports of several microalgae species isolated in different latitudesworldwide [203148497072]
Table 2 Effect of nitrogen deficiency on fatty acid profiling in three native microalgal species from thePeruvian Amazon
Fatty AcidsAcutodesmus obliquus Ankistrodesmus sp Chlorella lewinii
+N minusN +N minusN +N minusN
140 - - 052 plusmn 002 088 plusmn 002 - 058 plusmn 001160 2724 plusmn 007 2867 plusmn 017 1868 plusmn 019 2261 plusmn 006 1964 plusmn 021 2123 plusmn 010180 184 plusmn 005 255 plusmn 051 101 plusmn 002 181 plusmn 002 371 plusmn 026 153 plusmn 015
161n-7 023 plusmn 000 025 plusmn 001 071 plusmn 003 055 plusmn 001 085 plusmn 043 038 plusmn 000181n-7 131 plusmn 002 131 plusmn 000 141 plusmn 005 094 plusmn 003 146 plusmn 006 123 plusmn 002181n-9 3213 plusmn 021 3608 plusmn 021 3699 plusmn 004 4047 plusmn 038 1371 plusmn 015 2326 plusmn 041182n-6 1732 plusmn 004 1651 plusmn 016 747 plusmn 018 656 plusmn 007 2034 plusmn 039 1957 plusmn 001183n-3 1130 plusmn 017 854 plusmn 031 1486 plusmn 004 1173 plusmn 012 1815 plusmn 022 1371 plusmn 016183n-6 101 plusmn 001 078 plusmn 002 055 plusmn 000 054 plusmn 002 124 plusmn 006 100 plusmn 020184n-3 250 plusmn 005 177 plusmn 008 631 plusmn 002 601 plusmn 005 491 plusmn 025 403 plusmn 020204n-6 - - - 084 plusmn 008 - -225n-3 - - - 062 plusmn 002 - -
Unidentified 512 plusmn 003 355 plusmn 006 1149 plusmn 004 645 plusmn 003 1599 plusmn 034 1348 plusmn 006SFA 2907 3122 2022 2531 2335 2334UFA 7093 6878 7978 7469 7665 7666
MUFA 3367 3764 3911 4195 1602 2487PUFA 3213 2759 2918 2629 4464 3831SFA +MUFA 6274 6886 5933 6726 3936 4821
It is important to highlight that the fatty acid profile determination is an essential step inthe process of the characterization of microalgae strains because it ultimately affects the qualityof the biodiesel product [8485] The quality properties of biodiesel such as high cetane numberoxidative stability and cold-flow are dependent on the hydrocarbonated chain length of saturated andunsaturated fatty acids [8587] For example biodiesel composed of a mixture of a high percentage ofSFA and MUFA is preferred for increased energy yield superior oxidative stability and higher cetanenumbers but is prone to solidify at low temperature In contrast biodiesel with high levels of PUFAhas decreased energy yield low cetane numbers and excellent cold-flow properties but insufficientoxidative stability [548588] Therefore to solve these performance problems five approaches havebeen developed such as changing the fatty acids profile by physical means genetic modification ofthe feedstock or the use of alternative feedstocks with different fatty acids profiles [89]
4 Conclusions
Fourteen microalgae were isolated from three river basins from the Peruvian Amazon and wereidentified by morphological features and molecular phylogeny approaches These strains were initially
Energies 2017 10 224 12 of 16
characterized in the stationary growth phase and exhibited significant heterogeneity in key parameterssuch as biomass productivity (36ndash316 mgmiddotLminus1middotdminus1) lipid productivity (10ndash64 mgmiddotLminus1middotdminus1) andtotal lipid content (135ndash467) Furthermore three selected strains based on elevated biomass andlipid productivities were cultured under nitrogen-sufficient and nitrogen-deficient medium to boostlipid accumulation Under these culture conditions all strains showed noticeable decreases in totalprotein content and increases in total lipid and total carbohydrate contents In addition these strainspresented modifications in the fatty acids profiles with improvement in biodiesel properties due to apredominance of SFA and MUFA which corresponds to an increase in cetane number The results ofthis research suggest that the isolated native microalgae from the Peruvian Amazon could be suitablerenewable feedstocks for the production of both biodiesel and bioethanol which could contribute tosustainable development at the local regional and national levels as a viable alternative to petroleumexploitation However it will be necessary to develop open closed or hybrid systems for large-scalebiomass production of selected native microalgae strains andor de novo microalgae strains designedto produce biofuels and with the abilities to synthesize useful bioactive compounds to be efficientlyexploited using biorefinery approaches
Supplementary MaterialsThe following are available online at wwwmdpicom1996-1073102224s1 Figure S1 Results of
electrophoretic and spectrophotometric analysis of the genomic DNA isolated from the fourteen microalgaeFigure S2 ITS2-rDNA structure (A) and amplicons obtained (B) from the microalgae isolated Figure S3Microphotographic images of the three microalgae strains analyzed under light and fluorescence stained withNile Red Figure S4 Spectrophotometric scanning for the determination of the maximum absorption peak foreach microalgae strain
Acknowledgments This research was supported by the Peruvian funding agencies Consejo Nacional de CienciaTecnologiacutea e Innovacioacuten Tecnoloacutegica (CONCYTEC) with Grant Contract No 364-CONCYTEC-OAJ and Fondospara la Innovacioacuten Ciencia y Tecnologiacutea (FINCyT) with Grant Contract No 383-PNICP-PIBA-2014
Author Contributions Marianela Cobos and Juan C Castro obtained funds for the research participated inthe study design coordinated activities and participated in the preparation of the manuscript Jae D Paredesparticipated in sample collection microalgae isolation and helped to draft the manuscript J Dylan Maddoxperformed the bioinformatics analysis and participated in the preparation of the manuscript Gabriel Vargas-Aranaand Leenin Flores participated in biochemical analysis and helped to draft the manuscript Carla P Aguilarparticipated in the study design supervised biochemical analysis and helped to draft the manuscriptJorge L Marapara participated in molecular analysis and helped to draft the manuscript
Conflicts of Interest The authors declare no conflict of interest
References
1 BP Global BP Statisical Review of World Energy 2014 Available online httpwwwbpcomcontentdambppdfEnergy-economicsstatistical-review-2014BP-statistical-review-of-world-energy-2014-full-reportpdf (accessed on 20 June 2014)
2 Pachauri RK Reisinger A Cambio Climaacutetico 2007 Informe de Siacutentesis Contribucioacuten de los Grupos de Trabajo III y III al Cuarto Informe de Evaluacioacuten del Grupo Intergubernamental de Expertos Sobre el Cambio Climaacutetico IPCCGeneva Switzerland 2007 p 104
3 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Ley de Promocioacuten del Mercado deBiocombustibles Available online httpwww2congresogobpesicrcendocbibcon4_uibdnsf9E95620CC059138105257C9E005AB2F9$FILE28054pdf (accessed on 12 February 2017)
4 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Reglamento de la Ley de Promocioacuten del Mercadode Biocombustibles Available online httpwww2osinerggobpeMarcoLegaldocrevDS-013-2005-EM-CONCORDADOpdf (accessed on 12 February 2017)
5 Areacutevalo LF Nalvarte J Torres J Ramiacuterez Y Impactos Socio-Econoacutemicos de la Produccioacuten de Biocombustiblesen la Amazoniacutea Peruana Primera SNV-WWF Lima Peruacute 2009
6 Sharma KK Schuhmann H Schenk PM High Lipid Induction in Microalgae for Biodiesel ProductionEnergies 2012 5 1532ndash1553 [CrossRef]
Energies 2017 10 224 13 of 16
7 Schenk PM Thomas-Hall SR Stephens E Marx UC Mussgnug JH Posten C Kruse O Hankamer BSecond Generation Biofuels High-Efficiency Microalgae for Biodiesel Production BioEnergy Res 2008 120ndash43 [CrossRef]
8 Foley JA Asner GP Costa MH Coe MT de Fries R Gibbs HK Howard EA Olson S Patz JRamankutty N et al Amazonia revealed Forest degradation and loss of ecosystem goods and services inthe Amazon Basin Front Ecol Environ 2007 5 25ndash32 [CrossRef]
9 Duong VT Li Y Nowak E Schenk PM Microalgae Isolation and Selection for Prospective BiodieselProduction Energies 2012 5 1835ndash1849 [CrossRef]
10 Georgianna DR Mayfield SP Exploiting diversity and synthetic biology for the production of algalbiofuels Nature 2012 488 329ndash335 [CrossRef] [PubMed]
11 Wijffels RH Barbosa MJ An Outlook on Microalgal Biofuels Science 2010 329 796ndash799 [CrossRef][PubMed]
12 Richmond A Handbook of Microalgal Culture Biotechnology and Applied Phycology Blackwell Sci Ltd HudsonNJ USA 2004
13 Menetrez MY An overview of algae biofuel production and potential environmental impactEnviron Sci Technol 2012 46 7073ndash7085 [CrossRef] [PubMed]
14 Khoo HH Sharratt PN Das P Balasubramanian RK Naraharisetti PK Shaik S Life cycle energy andCO2 analysis of microalgae-to-biodiesel Preliminary results and comparisons Bioresour Technol 2011 1025800ndash5807 [CrossRef] [PubMed]
15 Chu SP The Influence of the Mineral Composition of the Medium on the Growth of Planktonic AlgaePart I Methods and Culture Media J Ecol 1942 30 284ndash325 [CrossRef]
16 Carlos EMB Mariatildengela M Gecircneros de Algas de Aacuteguas Continentais do BrasilmdashChave Para Identificaccedilatildeo eDescriccedilotildees 2nd ed Rima Sao Carlos Brasil 2006
17 Greenspan P Mayer EP Fowler SD Nile red A selective fluorescent stain for intracellular lipid dropletsJ Cell Biol 1985 100 965ndash973 [CrossRef] [PubMed]
18 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissuePhytochem Bull 1987 19 11ndash15
19 Sambrook J Fritsch EF Maniatis T Molecular Cloning A Laboratory Manual 2nd ed Cold Spring HarborLaboratory Press New York NY USA 1989
20 Kaur S Sarkar M Srivastava RB Gogoi HK Kalita MC Fatty acid profiling and molecularcharacterization of some freshwater microalgae from India with potential for biodiesel productionNew Biotechnol 2012 29 332ndash344 [CrossRef] [PubMed]
21 Benson DA Cavanaugh M Clark K Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW GenBankNucleic Acids Res 2013 41 D36ndashD42 [CrossRef] [PubMed]
22 NCBI Resource Coordinators Database Resources of the National Center for Biotechnology InformationNucleic Acids Res 2015 43 D6ndashD17
23 Ye J McGinnis S Madden TL BLAST Improvements for better sequence analysis Nucleic Acids Res 200634 W6ndashW9 [CrossRef] [PubMed]
24 BioEdit Sequence Alignment Editor for Windows 9598NTXPVista7 Available online httpwwwmbioncsuedubioeditbioedithtml (accessed on 2 December 2015)
25 Sievers F Higgins DG Clustal omega accurate alignment of very large numbers of sequencesMethods Mol Biol (Clifton NJ) 2014 1079 105ndash116
26 Saitou N Nei M The neighbor-joining method A new method for reconstructing phylogenetic treesMol Biol Evol 1987 4 406ndash425 [PubMed]
27 Kimura M A simple method for estimating evolutionary rates of base substitutions through comparativestudies of nucleotide sequences J Mol Evol 1980 16 111ndash120 [CrossRef] [PubMed]
28 Tamura K Stecher G Peterson D Filipski A Kumar S MEGA6 Molecular Evolutionary GeneticsAnalysis version 60 Mol Biol Evol 2013 30 2725ndash2729 [CrossRef] [PubMed]
29 Felsenstein J Confidence Limits on Phylogenies An Approach Using the Bootstrap Evolution 1985 39783ndash791 [CrossRef]
30 Bligh EG Dyer WJ A Rapid Method of Total Lipid Extraction and Purification Can J Biochem Physiol1959 37 911ndash917 [CrossRef] [PubMed]
Energies 2017 10 224 14 of 16
31 Yu X Zhao P He C Li J Tang X Zhou J Huang Z Isolation of a novel strain of Monoraphidium spand characterization of its potential application as biodiesel feedstock Bioresour Technol 2012 121 256ndash262[CrossRef] [PubMed]
32 Hartree EF Determination of protein A modification of the lowry method that gives a linear photometricresponse Anal Biochem 1972 48 422ndash427 [CrossRef]
33 DuBois M Gilles KA Hamilton JK Rebers PA Smith F Colorimetric Method for Determination ofSugars and Related Substances Anal Chem 1956 28 350ndash356 [CrossRef]
34 AOAC International AOAC Official Methods of Analysis of the Association of Official Analytical Chemists (AOAC)15th ed AOAC International Washington DC USA 1990 Volume 1
35 Ichihara K Fukubayashi Y Preparation of fatty acid methyl esters for gas-liquid chromatographyJ Lipid Res 2010 51 635ndash640 [CrossRef] [PubMed]
36 Brennan L Owende P Biofuels from microalgaemdashA review of technologies for production processingand extractions of biofuels and co-products Renew Sustain Energ Rev 2010 14 557ndash577 [CrossRef]
37 Raja R Hemaiswarya S Kumar NA Sridhar S Rengasamy R A perspective on the biotechnologicalpotential of microalgae Crit Rev Microbiol 2008 34 77ndash88 [CrossRef] [PubMed]
38 Mata TM Martins AA Caetano NS Microalgae for biodiesel production and other applicationsA review Renew Sustain Energy Rev 2010 14 217ndash232 [CrossRef]
39 De Wilt A Butkovskyi A Tuantet K Leal LH Fernandes TV Langenhoff A Zeeman GMicropollutant removal in an algal treatment system fed with source separated wastewater streamsJ Hazard Mater 2015 304 84ndash92 [CrossRef] [PubMed]
40 Li T Lin G Podola B Melkonian M Continuous removal of zinc from wastewater and mine dumpleachate by a microalgal biofilm PSBR J Hazard Mater 2015 297 112ndash118 [CrossRef] [PubMed]
41 Sioli H The Amazon and its main affluents Hydrography morphology of the river courses and river typesIn Monographiae Biologicae Sioli H Ed Springer Dordrecht The Netherlands 1984 pp 127ndash165
42 Moniz MBJ Kaczmarska I Barcoding of Diatoms Nuclear Encoded ITS Revisited Protist 2010 161 7ndash34[CrossRef] [PubMed]
43 Coleman AW Is there a molecular key to the level of ldquobiological speciesrdquo in eukaryotes A DNA guideMol Phylogenet Evol 2009 50 197ndash203 [CrossRef] [PubMed]
44 Mai JC Coleman AW The Internal Transcribed Spacer 2 Exhibits a Common Secondary Structure inGreen Algae and Flowering Plants J Mol Evol 1997 44 258ndash271 [CrossRef] [PubMed]
45 Abou-Shanab RAI Matter IA Kim S-N Oh Y-K Choi J Jeon B-H Characterization andidentification of lipid-producing microalgae species isolated from a freshwater lake Biomass Bioenergy2011 35 3079ndash3085 [CrossRef]
46 Do Nascimento M Ortiz-Marquez JCF Sanchez-Rizza L Echarte MM Curatti L Bioprospectingfor fast growing and biomass characterization of oleaginous microalgae from South-Eastern Buenos AiresArgentina Bioresour Technol 2012 125 283ndash290 [CrossRef] [PubMed]
47 Araujo GS Matos LJBL Gonccedilalves LRB Fernandes FAN Farias WRL Bioprospecting foroil producing microalgal strains Evaluation of oil and biomass production for ten microalgal strainsBioresour Technol 2011 102 5248ndash5250 [CrossRef] [PubMed]
48 Zhou W Li Y Min M Hu B Chen P Ruan R Local bioprospecting for high-lipid producing microalgalstrains to be grown on concentrated municipal wastewater for biofuel production Bioresour Technol 2011102 6909ndash6919 [CrossRef] [PubMed]
49 Mahmoud EA Farahat LA Abdel Aziz ZK Fatthallah NA Salah El Din RA Evaluation of thepotential for some isolated microalgae to produce biodiesel Egypt J Pet 2015 24 97ndash101 [CrossRef]
50 Ho S Lai Y-Y Chiang C-Y Chen C-NN Chang J-S Selection of elite microalgae for biodieselproduction in tropical conditions using a standardized platform Bioresour Technol 2013 147 135ndash142[CrossRef] [PubMed]
51 Li L Cui J Liu Q Ding Y Liu J Screening and phylogenetic analysis of lipid-rich microalgae Algal Res2015 11 381ndash386 [CrossRef]
52 Rodolfi L Chini Zittelli G Bassi N Padovani G Biondi N Bonini G Tredici MR Microalgae for oilStrain selection induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactorBiotechnol Bioeng 2009 102 100ndash112 [CrossRef] [PubMed]
Energies 2017 10 224 15 of 16
53 Li P Miao X Li R Zhong J In situ biodiesel production from fast-growing and high oil content Chlorellapyrenoidosa in rice straw hydrolysate J Biomed Biotechnol 2011 2011 141207 [CrossRef] [PubMed]
54 Doan TTY Sivaloganathan B Obbard JP Screening of marine microalgae for biodiesel feedstockBiomass Bioenergy 2011 35 2534ndash2544 [CrossRef]
55 Abou-Shanab RAI Hwang J-H Cho Y Min B Jeon B-H Characterization of microalgal speciesisolated from fresh water bodies as a potential source for biodiesel production Appl Energy 2011 883300ndash3306 [CrossRef]
56 Chaichalerm S Pokethitiyook P Yuan W Meetam M Sritong K Pugkaew W Kungvansaichol KKruatrachue M Damrongphol P Culture of microalgal strains isolated from natural habitats in Thailand invarious enriched media Appl Energy 2012 89 296ndash302 [CrossRef]
57 Zhang S Liu P Yang X Hao Z Zhang L Luo N Shi J Isolation and identification by 18S rDNAsequence of high lipid potential microalgal species for fuel production in Hainan Dao Biomass Bioenergy2014 66 197ndash203 [CrossRef]
58 Chiu S-Y Kao C-Y Tsai M-T Ong S-C Chen C-H Lin C-S Lipid accumulation and CO2 utilizationof Nannochloropsis oculata in response to CO2 aeration Bioresour Technol 2009 100 833ndash838 [CrossRef][PubMed]
59 Huerlimann R de Nys R Heimann K Growth lipid content productivity and fatty acid composition oftropical microalgae for scale-up production Biotechnol Bioeng 2010 107 245ndash257 [CrossRef] [PubMed]
60 Liang Y Sarkany N Cui Y Biomass and lipid productivities of Chlorella vulgaris under autotrophicheterotrophic and mixotrophic growth conditions Biotechnol Lett 2009 31 1043ndash1049 [CrossRef] [PubMed]
61 Hempel N Petrick I Behrendt F Biomass productivity and productivity of fatty acids and amino acidsof microalgae strains as key characteristics of suitability for biodiesel production J Appl Phycol 2012 241407ndash1418 [CrossRef] [PubMed]
62 Yeesang C Cheirsilp B Effect of nitrogen salt and iron content in the growth medium and light intensityon lipid production by microalgae isolated from freshwater sources in Thailand Bioresour Technol 2011 1023034ndash3040 [CrossRef] [PubMed]
63 Msanne J Xu D Konda AR Casas-Mollano JA Awada T Cahoon EB Cerutti H Metabolic andgene expression changes triggered by nitrogen deprivation in the photoautotrophically grown microalgaeChlamydomonas reinhardtii and Coccomyxa sp C-169 Phytochemistry 2012 75 50ndash59 [CrossRef] [PubMed]
64 Lin Q Lin J Effects of nitrogen source and concentration on biomass and oil production of aScenedesmus rubescens like microalga Bioresour Technol 2011 102 1615ndash1621 [CrossRef] [PubMed]
65 Sheehan J Dunahay T Benemann J Roessler P A Look Back at the US Department of Energyrsquos AquaticSpecies ProgrammdashBiodiesel from Algae US Department of Energyrsquos Office of Fuels Development Golden COUSA 1998 p 328
66 Vaulot D Olson RJ Merkel S Chisholm SW Cell-cycle response to nutrient starvation in twophytoplankton species Thalassiosira weissflogii and Hymenomonas carterae Mar Biol 1987 95 625ndash630[CrossRef]
67 Olson RJ Chisholm SW Effects of light and nitrogen limitation on the cell cycle of the dinoflagellateAmphidinium carteri J Plankton Res 1986 8 785ndash793 [CrossRef]
68 Li Y Fei X Deng X Novel molecular insights into nitrogen starvation-induced triacylglycerolsaccumulation revealed by differential gene expression analysis in green algae Micractinium pusillumBiomass Bioenergy 2012 42 199ndash211 [CrossRef]
69 Abdelaziz AEM Leite GB Belhaj MA Hallenbeck PC Screening microalgae native to Quebec forwastewater treatment and biodiesel production Bioresour Technol 2014 157 140ndash148 [CrossRef] [PubMed]
70 Valdez-Ojeda R Gonzaacutelez-Muntildeoz M Us-Vaacutezquez R Narvaacuteez-Zapata J Chavarria-Hernandez JCLoacutepez-Adriaacuten S Barahona-Peacuterez F Toledano-Thompson T Garduntildeo-Soloacuterzano G Escobedo-GraciaMedrano RM Characterization of five fresh water microalgae with potential for biodiesel productionAlgal Res 2015 7 33ndash44 [CrossRef]
71 Shrivastav A Mishra SK Suh WI Farooq W Moon M Kim T-H Kumar K Choi G-G Park MSYang J-W Characterization of newly isolated oleaginous microalga Monoraphidium sp for lipid productionunder different conditions Algal Res 2015 12 289ndash294 [CrossRef]
72 Yen H-W Hu I-C Chen C-Y Ho S-H Lee D-J Chang J-S Microalgae-based biorefinerymdashFrombiofuels to natural products Bioresour Technol 2013 135 166ndash174 [CrossRef] [PubMed]
Energies 2017 10 224 16 of 16
73 Li J Liu Y Cheng JJ Mos M Daroch M Biological potential of microalgae in China for biorefinery-basedproduction of biofuels and high value compounds New Biotechnol 2015 32 588ndash596 [CrossRef] [PubMed]
74 El-Sheekh M Abomohra AE-F Hanelt D Optimization of biomass and fatty acid productivity ofScenedesmus obliquus as a promising microalga for biodiesel production World J Microbiol Biotechnol 201329 915ndash922 [CrossRef] [PubMed]
75 Singh P Guldhe A Kumari S Rawat I Bux F Investigation of combined effect of nitrogen phosphorusand iron on lipid productivity of microalgae Ankistrodesmus falcatus KJ671624 using response surfacemethodology Biochem Eng J 2015 94 22ndash29 [CrossRef]
76 Kilham S Kreeger D Goulden C Lynn S Effects of nutrient limitation on biochemical constituents ofAnkistrodesmus falcatus Freshw Biol 1997 38 591ndash596 [CrossRef]
77 Breuer G Lamers PP Martens DE Draaisma RB Wijffels RH The impact of nitrogen starvation on thedynamics of triacylglycerol accumulation in nine microalgae strains Bioresour Technol 2012 124 217ndash226[CrossRef] [PubMed]
78 Breuer G de Jaeger L Artus VG Martens DE Springer J Draaisma RB Eggink G Wijffels RHLamers PP Superior triacylglycerol (TAG) accumulation in starchless mutants of Scenedesmus obliquus (II)evaluation of TAG yield and productivity in controlled photobioreactors Biotechnol Biofuels 2014 7 70[CrossRef] [PubMed]
79 Nigam S Prakash-Rai M Sharma R Effect of Nitrogen on Growth and Lipid Content ofChlorella pyrenoidosa Am J Biochem Biotechnol 2011 7 126ndash131 [CrossRef]
80 Piligaev AV Sorokina KN Bryanskaya AV Peltek SE Kolchanov NA Parmon VN Isolation ofprospective microalgal strains with high saturated fatty acid content for biofuel production Algal Res 201512 368ndash376 [CrossRef]
81 Weiss SB Kennedy EP The enzymatic synthesis of triglycerides J Am Chem Soc 1956 78 3550[CrossRef]
82 Miller R Wu G Deshpande RR Vieler A Gaumlrtner K Li X Moellering ER Zaumluner S Cornish AJLiu B et al Changes in Transcript Abundance in Chlamydomonas reinhardtii following Nitrogen DeprivationPredict Diversion of Metabolism Plant Physiol 2010 154 1737ndash1752 [CrossRef] [PubMed]
83 Rismani-Yazdi H Haznedaroglu BZ Bibby K Peccia J Transcriptome sequencing and annotation of themicroalgae Dunaliella tertiolecta Pathway description and gene discovery for production of next-generationbiofuels BMC Genom 2011 12 148 [CrossRef] [PubMed]
84 Ramos MJ Fernaacutendez CM Casas A Rodriacuteguez L Peacuterez A Influence of fatty acid composition of rawmaterials on biodiesel properties Bioresour Technol 2009 100 261ndash268 [CrossRef] [PubMed]
85 Knothe G Dependence of biodiesel fuel properties on the structure of fatty acid alkyl estersFuel Process Technol 2005 86 1059ndash1070 [CrossRef]
86 Knothe G ldquoDesignerrdquo Biodiesel Optimizing Fatty Ester Composition to Improve Fuel PropertiesEnergy Fuels 2008 22 1358ndash1364 [CrossRef]
87 Smith PC Ngothai Y Dzuy Nguyen Q OrsquoNeill BK Improving the low-temperature properties ofbiodiesel Methods and consequences Renew Energy 2010 35 1145ndash1151 [CrossRef]
88 Dunn RO Effect of antioxidants on the oxidative stability of methyl soyate (biodiesel) Fuel Process Technol2005 86 1071ndash1085 [CrossRef]
89 Knothe G Improving biodiesel fuel properties by modifying fatty ester composition Energy Environ Sci2009 2 759ndash766 [CrossRef]
copy 2017 by the authors licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
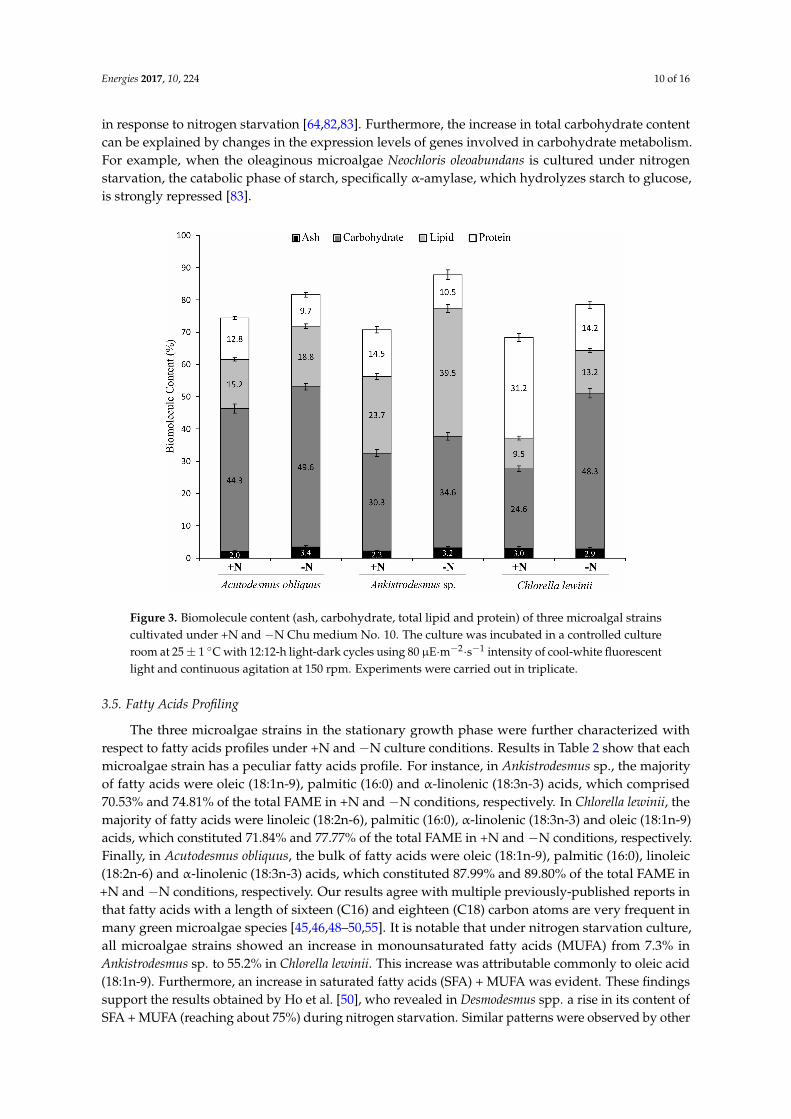
Energies 2017 10 224 10 of 16
in response to nitrogen starvation [648283] Furthermore the increase in total carbohydrate contentcan be explained by changes in the expression levels of genes involved in carbohydrate metabolismFor example when the oleaginous microalgae Neochloris oleoabundans is cultured under nitrogenstarvation the catabolic phase of starch specifically α-amylase which hydrolyzes starch to glucoseis strongly repressed [83]Energies 2017 10 224 10 of 16
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strains cultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled culture room at 25 plusmn 1 degC with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescent light and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized with respect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that each microalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majority of fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised 7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii the majority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9) acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectively Finally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic (182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in +N and minusN conditions respectively Our results agree with multiple previously-published reports in that fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent in many green microalgae species [454648ndash5055] It is notable that under nitrogen starvation culture all microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 in Ankistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid (181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findings support the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content of SFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Nevertheless there are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed a decrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increased between 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively However in the same culture condition the three microalgae species showed a decrease in polyunsaturated fatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmus spp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgae strains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFA which improves the quality properties of biodiesel such as an increase
Figure 3 Biomolecule content (ash carbohydrate total lipid and protein) of three microalgal strainscultivated under +N and minusN Chu medium No 10 The culture was incubated in a controlled cultureroom at 25plusmn 1 C with 1212-h light-dark cycles using 80 microEmiddotmminus2middotsminus1 intensity of cool-white fluorescentlight and continuous agitation at 150 rpm Experiments were carried out in triplicate
35 Fatty Acids Profiling
The three microalgae strains in the stationary growth phase were further characterized withrespect to fatty acids profiles under +N and minusN culture conditions Results in Table 2 show that eachmicroalgae strain has a peculiar fatty acids profile For instance in Ankistrodesmus sp the majorityof fatty acids were oleic (181n-9) palmitic (160) and α-linolenic (183n-3) acids which comprised7053 and 7481 of the total FAME in +N and minusN conditions respectively In Chlorella lewinii themajority of fatty acids were linoleic (182n-6) palmitic (160) α-linolenic (183n-3) and oleic (181n-9)acids which constituted 7184 and 7777 of the total FAME in +N and minusN conditions respectivelyFinally in Acutodesmus obliquus the bulk of fatty acids were oleic (181n-9) palmitic (160) linoleic(182n-6) and α-linolenic (183n-3) acids which constituted 8799 and 8980 of the total FAME in+N and minusN conditions respectively Our results agree with multiple previously-published reports inthat fatty acids with a length of sixteen (C16) and eighteen (C18) carbon atoms are very frequent inmany green microalgae species [454648ndash5055] It is notable that under nitrogen starvation cultureall microalgae strains showed an increase in monounsaturated fatty acids (MUFA) from 73 inAnkistrodesmus sp to 552 in Chlorella lewinii This increase was attributable commonly to oleic acid(181n-9) Furthermore an increase in saturated fatty acids (SFA) + MUFA was evident These findingssupport the results obtained by Ho et al [50] who revealed in Desmodesmus spp a rise in its content ofSFA + MUFA (reaching about 75) during nitrogen starvation Similar patterns were observed by other
Energies 2017 10 224 11 of 16
researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Neverthelessthere are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed adecrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increasedbetween 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively Howeverin the same culture condition the three microalgae species showed a decrease in polyunsaturatedfatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmusspp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgaestrains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFAwhich improves the quality properties of biodiesel such as an increase in cetane number [84ndash86]Considering the SFA and unsaturated fatty acids (UFA) components approximately 20ndash31 are SFAand 69ndash80 are UFA Hence the SFAUFA ratio in the microalgae strains evaluated ranges between025 and 045 which is similar to the reports of several microalgae species isolated in different latitudesworldwide [203148497072]
Table 2 Effect of nitrogen deficiency on fatty acid profiling in three native microalgal species from thePeruvian Amazon
Fatty AcidsAcutodesmus obliquus Ankistrodesmus sp Chlorella lewinii
+N minusN +N minusN +N minusN
140 - - 052 plusmn 002 088 plusmn 002 - 058 plusmn 001160 2724 plusmn 007 2867 plusmn 017 1868 plusmn 019 2261 plusmn 006 1964 plusmn 021 2123 plusmn 010180 184 plusmn 005 255 plusmn 051 101 plusmn 002 181 plusmn 002 371 plusmn 026 153 plusmn 015
161n-7 023 plusmn 000 025 plusmn 001 071 plusmn 003 055 plusmn 001 085 plusmn 043 038 plusmn 000181n-7 131 plusmn 002 131 plusmn 000 141 plusmn 005 094 plusmn 003 146 plusmn 006 123 plusmn 002181n-9 3213 plusmn 021 3608 plusmn 021 3699 plusmn 004 4047 plusmn 038 1371 plusmn 015 2326 plusmn 041182n-6 1732 plusmn 004 1651 plusmn 016 747 plusmn 018 656 plusmn 007 2034 plusmn 039 1957 plusmn 001183n-3 1130 plusmn 017 854 plusmn 031 1486 plusmn 004 1173 plusmn 012 1815 plusmn 022 1371 plusmn 016183n-6 101 plusmn 001 078 plusmn 002 055 plusmn 000 054 plusmn 002 124 plusmn 006 100 plusmn 020184n-3 250 plusmn 005 177 plusmn 008 631 plusmn 002 601 plusmn 005 491 plusmn 025 403 plusmn 020204n-6 - - - 084 plusmn 008 - -225n-3 - - - 062 plusmn 002 - -
Unidentified 512 plusmn 003 355 plusmn 006 1149 plusmn 004 645 plusmn 003 1599 plusmn 034 1348 plusmn 006SFA 2907 3122 2022 2531 2335 2334UFA 7093 6878 7978 7469 7665 7666
MUFA 3367 3764 3911 4195 1602 2487PUFA 3213 2759 2918 2629 4464 3831SFA +MUFA 6274 6886 5933 6726 3936 4821
It is important to highlight that the fatty acid profile determination is an essential step inthe process of the characterization of microalgae strains because it ultimately affects the qualityof the biodiesel product [8485] The quality properties of biodiesel such as high cetane numberoxidative stability and cold-flow are dependent on the hydrocarbonated chain length of saturated andunsaturated fatty acids [8587] For example biodiesel composed of a mixture of a high percentage ofSFA and MUFA is preferred for increased energy yield superior oxidative stability and higher cetanenumbers but is prone to solidify at low temperature In contrast biodiesel with high levels of PUFAhas decreased energy yield low cetane numbers and excellent cold-flow properties but insufficientoxidative stability [548588] Therefore to solve these performance problems five approaches havebeen developed such as changing the fatty acids profile by physical means genetic modification ofthe feedstock or the use of alternative feedstocks with different fatty acids profiles [89]
4 Conclusions
Fourteen microalgae were isolated from three river basins from the Peruvian Amazon and wereidentified by morphological features and molecular phylogeny approaches These strains were initially
Energies 2017 10 224 12 of 16
characterized in the stationary growth phase and exhibited significant heterogeneity in key parameterssuch as biomass productivity (36ndash316 mgmiddotLminus1middotdminus1) lipid productivity (10ndash64 mgmiddotLminus1middotdminus1) andtotal lipid content (135ndash467) Furthermore three selected strains based on elevated biomass andlipid productivities were cultured under nitrogen-sufficient and nitrogen-deficient medium to boostlipid accumulation Under these culture conditions all strains showed noticeable decreases in totalprotein content and increases in total lipid and total carbohydrate contents In addition these strainspresented modifications in the fatty acids profiles with improvement in biodiesel properties due to apredominance of SFA and MUFA which corresponds to an increase in cetane number The results ofthis research suggest that the isolated native microalgae from the Peruvian Amazon could be suitablerenewable feedstocks for the production of both biodiesel and bioethanol which could contribute tosustainable development at the local regional and national levels as a viable alternative to petroleumexploitation However it will be necessary to develop open closed or hybrid systems for large-scalebiomass production of selected native microalgae strains andor de novo microalgae strains designedto produce biofuels and with the abilities to synthesize useful bioactive compounds to be efficientlyexploited using biorefinery approaches
Supplementary MaterialsThe following are available online at wwwmdpicom1996-1073102224s1 Figure S1 Results of
electrophoretic and spectrophotometric analysis of the genomic DNA isolated from the fourteen microalgaeFigure S2 ITS2-rDNA structure (A) and amplicons obtained (B) from the microalgae isolated Figure S3Microphotographic images of the three microalgae strains analyzed under light and fluorescence stained withNile Red Figure S4 Spectrophotometric scanning for the determination of the maximum absorption peak foreach microalgae strain
Acknowledgments This research was supported by the Peruvian funding agencies Consejo Nacional de CienciaTecnologiacutea e Innovacioacuten Tecnoloacutegica (CONCYTEC) with Grant Contract No 364-CONCYTEC-OAJ and Fondospara la Innovacioacuten Ciencia y Tecnologiacutea (FINCyT) with Grant Contract No 383-PNICP-PIBA-2014
Author Contributions Marianela Cobos and Juan C Castro obtained funds for the research participated inthe study design coordinated activities and participated in the preparation of the manuscript Jae D Paredesparticipated in sample collection microalgae isolation and helped to draft the manuscript J Dylan Maddoxperformed the bioinformatics analysis and participated in the preparation of the manuscript Gabriel Vargas-Aranaand Leenin Flores participated in biochemical analysis and helped to draft the manuscript Carla P Aguilarparticipated in the study design supervised biochemical analysis and helped to draft the manuscriptJorge L Marapara participated in molecular analysis and helped to draft the manuscript
Conflicts of Interest The authors declare no conflict of interest
References
1 BP Global BP Statisical Review of World Energy 2014 Available online httpwwwbpcomcontentdambppdfEnergy-economicsstatistical-review-2014BP-statistical-review-of-world-energy-2014-full-reportpdf (accessed on 20 June 2014)
2 Pachauri RK Reisinger A Cambio Climaacutetico 2007 Informe de Siacutentesis Contribucioacuten de los Grupos de Trabajo III y III al Cuarto Informe de Evaluacioacuten del Grupo Intergubernamental de Expertos Sobre el Cambio Climaacutetico IPCCGeneva Switzerland 2007 p 104
3 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Ley de Promocioacuten del Mercado deBiocombustibles Available online httpwww2congresogobpesicrcendocbibcon4_uibdnsf9E95620CC059138105257C9E005AB2F9$FILE28054pdf (accessed on 12 February 2017)
4 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Reglamento de la Ley de Promocioacuten del Mercadode Biocombustibles Available online httpwww2osinerggobpeMarcoLegaldocrevDS-013-2005-EM-CONCORDADOpdf (accessed on 12 February 2017)
5 Areacutevalo LF Nalvarte J Torres J Ramiacuterez Y Impactos Socio-Econoacutemicos de la Produccioacuten de Biocombustiblesen la Amazoniacutea Peruana Primera SNV-WWF Lima Peruacute 2009
6 Sharma KK Schuhmann H Schenk PM High Lipid Induction in Microalgae for Biodiesel ProductionEnergies 2012 5 1532ndash1553 [CrossRef]
Energies 2017 10 224 13 of 16
7 Schenk PM Thomas-Hall SR Stephens E Marx UC Mussgnug JH Posten C Kruse O Hankamer BSecond Generation Biofuels High-Efficiency Microalgae for Biodiesel Production BioEnergy Res 2008 120ndash43 [CrossRef]
8 Foley JA Asner GP Costa MH Coe MT de Fries R Gibbs HK Howard EA Olson S Patz JRamankutty N et al Amazonia revealed Forest degradation and loss of ecosystem goods and services inthe Amazon Basin Front Ecol Environ 2007 5 25ndash32 [CrossRef]
9 Duong VT Li Y Nowak E Schenk PM Microalgae Isolation and Selection for Prospective BiodieselProduction Energies 2012 5 1835ndash1849 [CrossRef]
10 Georgianna DR Mayfield SP Exploiting diversity and synthetic biology for the production of algalbiofuels Nature 2012 488 329ndash335 [CrossRef] [PubMed]
11 Wijffels RH Barbosa MJ An Outlook on Microalgal Biofuels Science 2010 329 796ndash799 [CrossRef][PubMed]
12 Richmond A Handbook of Microalgal Culture Biotechnology and Applied Phycology Blackwell Sci Ltd HudsonNJ USA 2004
13 Menetrez MY An overview of algae biofuel production and potential environmental impactEnviron Sci Technol 2012 46 7073ndash7085 [CrossRef] [PubMed]
14 Khoo HH Sharratt PN Das P Balasubramanian RK Naraharisetti PK Shaik S Life cycle energy andCO2 analysis of microalgae-to-biodiesel Preliminary results and comparisons Bioresour Technol 2011 1025800ndash5807 [CrossRef] [PubMed]
15 Chu SP The Influence of the Mineral Composition of the Medium on the Growth of Planktonic AlgaePart I Methods and Culture Media J Ecol 1942 30 284ndash325 [CrossRef]
16 Carlos EMB Mariatildengela M Gecircneros de Algas de Aacuteguas Continentais do BrasilmdashChave Para Identificaccedilatildeo eDescriccedilotildees 2nd ed Rima Sao Carlos Brasil 2006
17 Greenspan P Mayer EP Fowler SD Nile red A selective fluorescent stain for intracellular lipid dropletsJ Cell Biol 1985 100 965ndash973 [CrossRef] [PubMed]
18 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissuePhytochem Bull 1987 19 11ndash15
19 Sambrook J Fritsch EF Maniatis T Molecular Cloning A Laboratory Manual 2nd ed Cold Spring HarborLaboratory Press New York NY USA 1989
20 Kaur S Sarkar M Srivastava RB Gogoi HK Kalita MC Fatty acid profiling and molecularcharacterization of some freshwater microalgae from India with potential for biodiesel productionNew Biotechnol 2012 29 332ndash344 [CrossRef] [PubMed]
21 Benson DA Cavanaugh M Clark K Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW GenBankNucleic Acids Res 2013 41 D36ndashD42 [CrossRef] [PubMed]
22 NCBI Resource Coordinators Database Resources of the National Center for Biotechnology InformationNucleic Acids Res 2015 43 D6ndashD17
23 Ye J McGinnis S Madden TL BLAST Improvements for better sequence analysis Nucleic Acids Res 200634 W6ndashW9 [CrossRef] [PubMed]
24 BioEdit Sequence Alignment Editor for Windows 9598NTXPVista7 Available online httpwwwmbioncsuedubioeditbioedithtml (accessed on 2 December 2015)
25 Sievers F Higgins DG Clustal omega accurate alignment of very large numbers of sequencesMethods Mol Biol (Clifton NJ) 2014 1079 105ndash116
26 Saitou N Nei M The neighbor-joining method A new method for reconstructing phylogenetic treesMol Biol Evol 1987 4 406ndash425 [PubMed]
27 Kimura M A simple method for estimating evolutionary rates of base substitutions through comparativestudies of nucleotide sequences J Mol Evol 1980 16 111ndash120 [CrossRef] [PubMed]
28 Tamura K Stecher G Peterson D Filipski A Kumar S MEGA6 Molecular Evolutionary GeneticsAnalysis version 60 Mol Biol Evol 2013 30 2725ndash2729 [CrossRef] [PubMed]
29 Felsenstein J Confidence Limits on Phylogenies An Approach Using the Bootstrap Evolution 1985 39783ndash791 [CrossRef]
30 Bligh EG Dyer WJ A Rapid Method of Total Lipid Extraction and Purification Can J Biochem Physiol1959 37 911ndash917 [CrossRef] [PubMed]
Energies 2017 10 224 14 of 16
31 Yu X Zhao P He C Li J Tang X Zhou J Huang Z Isolation of a novel strain of Monoraphidium spand characterization of its potential application as biodiesel feedstock Bioresour Technol 2012 121 256ndash262[CrossRef] [PubMed]
32 Hartree EF Determination of protein A modification of the lowry method that gives a linear photometricresponse Anal Biochem 1972 48 422ndash427 [CrossRef]
33 DuBois M Gilles KA Hamilton JK Rebers PA Smith F Colorimetric Method for Determination ofSugars and Related Substances Anal Chem 1956 28 350ndash356 [CrossRef]
34 AOAC International AOAC Official Methods of Analysis of the Association of Official Analytical Chemists (AOAC)15th ed AOAC International Washington DC USA 1990 Volume 1
35 Ichihara K Fukubayashi Y Preparation of fatty acid methyl esters for gas-liquid chromatographyJ Lipid Res 2010 51 635ndash640 [CrossRef] [PubMed]
36 Brennan L Owende P Biofuels from microalgaemdashA review of technologies for production processingand extractions of biofuels and co-products Renew Sustain Energ Rev 2010 14 557ndash577 [CrossRef]
37 Raja R Hemaiswarya S Kumar NA Sridhar S Rengasamy R A perspective on the biotechnologicalpotential of microalgae Crit Rev Microbiol 2008 34 77ndash88 [CrossRef] [PubMed]
38 Mata TM Martins AA Caetano NS Microalgae for biodiesel production and other applicationsA review Renew Sustain Energy Rev 2010 14 217ndash232 [CrossRef]
39 De Wilt A Butkovskyi A Tuantet K Leal LH Fernandes TV Langenhoff A Zeeman GMicropollutant removal in an algal treatment system fed with source separated wastewater streamsJ Hazard Mater 2015 304 84ndash92 [CrossRef] [PubMed]
40 Li T Lin G Podola B Melkonian M Continuous removal of zinc from wastewater and mine dumpleachate by a microalgal biofilm PSBR J Hazard Mater 2015 297 112ndash118 [CrossRef] [PubMed]
41 Sioli H The Amazon and its main affluents Hydrography morphology of the river courses and river typesIn Monographiae Biologicae Sioli H Ed Springer Dordrecht The Netherlands 1984 pp 127ndash165
42 Moniz MBJ Kaczmarska I Barcoding of Diatoms Nuclear Encoded ITS Revisited Protist 2010 161 7ndash34[CrossRef] [PubMed]
43 Coleman AW Is there a molecular key to the level of ldquobiological speciesrdquo in eukaryotes A DNA guideMol Phylogenet Evol 2009 50 197ndash203 [CrossRef] [PubMed]
44 Mai JC Coleman AW The Internal Transcribed Spacer 2 Exhibits a Common Secondary Structure inGreen Algae and Flowering Plants J Mol Evol 1997 44 258ndash271 [CrossRef] [PubMed]
45 Abou-Shanab RAI Matter IA Kim S-N Oh Y-K Choi J Jeon B-H Characterization andidentification of lipid-producing microalgae species isolated from a freshwater lake Biomass Bioenergy2011 35 3079ndash3085 [CrossRef]
46 Do Nascimento M Ortiz-Marquez JCF Sanchez-Rizza L Echarte MM Curatti L Bioprospectingfor fast growing and biomass characterization of oleaginous microalgae from South-Eastern Buenos AiresArgentina Bioresour Technol 2012 125 283ndash290 [CrossRef] [PubMed]
47 Araujo GS Matos LJBL Gonccedilalves LRB Fernandes FAN Farias WRL Bioprospecting foroil producing microalgal strains Evaluation of oil and biomass production for ten microalgal strainsBioresour Technol 2011 102 5248ndash5250 [CrossRef] [PubMed]
48 Zhou W Li Y Min M Hu B Chen P Ruan R Local bioprospecting for high-lipid producing microalgalstrains to be grown on concentrated municipal wastewater for biofuel production Bioresour Technol 2011102 6909ndash6919 [CrossRef] [PubMed]
49 Mahmoud EA Farahat LA Abdel Aziz ZK Fatthallah NA Salah El Din RA Evaluation of thepotential for some isolated microalgae to produce biodiesel Egypt J Pet 2015 24 97ndash101 [CrossRef]
50 Ho S Lai Y-Y Chiang C-Y Chen C-NN Chang J-S Selection of elite microalgae for biodieselproduction in tropical conditions using a standardized platform Bioresour Technol 2013 147 135ndash142[CrossRef] [PubMed]
51 Li L Cui J Liu Q Ding Y Liu J Screening and phylogenetic analysis of lipid-rich microalgae Algal Res2015 11 381ndash386 [CrossRef]
52 Rodolfi L Chini Zittelli G Bassi N Padovani G Biondi N Bonini G Tredici MR Microalgae for oilStrain selection induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactorBiotechnol Bioeng 2009 102 100ndash112 [CrossRef] [PubMed]
Energies 2017 10 224 15 of 16
53 Li P Miao X Li R Zhong J In situ biodiesel production from fast-growing and high oil content Chlorellapyrenoidosa in rice straw hydrolysate J Biomed Biotechnol 2011 2011 141207 [CrossRef] [PubMed]
54 Doan TTY Sivaloganathan B Obbard JP Screening of marine microalgae for biodiesel feedstockBiomass Bioenergy 2011 35 2534ndash2544 [CrossRef]
55 Abou-Shanab RAI Hwang J-H Cho Y Min B Jeon B-H Characterization of microalgal speciesisolated from fresh water bodies as a potential source for biodiesel production Appl Energy 2011 883300ndash3306 [CrossRef]
56 Chaichalerm S Pokethitiyook P Yuan W Meetam M Sritong K Pugkaew W Kungvansaichol KKruatrachue M Damrongphol P Culture of microalgal strains isolated from natural habitats in Thailand invarious enriched media Appl Energy 2012 89 296ndash302 [CrossRef]
57 Zhang S Liu P Yang X Hao Z Zhang L Luo N Shi J Isolation and identification by 18S rDNAsequence of high lipid potential microalgal species for fuel production in Hainan Dao Biomass Bioenergy2014 66 197ndash203 [CrossRef]
58 Chiu S-Y Kao C-Y Tsai M-T Ong S-C Chen C-H Lin C-S Lipid accumulation and CO2 utilizationof Nannochloropsis oculata in response to CO2 aeration Bioresour Technol 2009 100 833ndash838 [CrossRef][PubMed]
59 Huerlimann R de Nys R Heimann K Growth lipid content productivity and fatty acid composition oftropical microalgae for scale-up production Biotechnol Bioeng 2010 107 245ndash257 [CrossRef] [PubMed]
60 Liang Y Sarkany N Cui Y Biomass and lipid productivities of Chlorella vulgaris under autotrophicheterotrophic and mixotrophic growth conditions Biotechnol Lett 2009 31 1043ndash1049 [CrossRef] [PubMed]
61 Hempel N Petrick I Behrendt F Biomass productivity and productivity of fatty acids and amino acidsof microalgae strains as key characteristics of suitability for biodiesel production J Appl Phycol 2012 241407ndash1418 [CrossRef] [PubMed]
62 Yeesang C Cheirsilp B Effect of nitrogen salt and iron content in the growth medium and light intensityon lipid production by microalgae isolated from freshwater sources in Thailand Bioresour Technol 2011 1023034ndash3040 [CrossRef] [PubMed]
63 Msanne J Xu D Konda AR Casas-Mollano JA Awada T Cahoon EB Cerutti H Metabolic andgene expression changes triggered by nitrogen deprivation in the photoautotrophically grown microalgaeChlamydomonas reinhardtii and Coccomyxa sp C-169 Phytochemistry 2012 75 50ndash59 [CrossRef] [PubMed]
64 Lin Q Lin J Effects of nitrogen source and concentration on biomass and oil production of aScenedesmus rubescens like microalga Bioresour Technol 2011 102 1615ndash1621 [CrossRef] [PubMed]
65 Sheehan J Dunahay T Benemann J Roessler P A Look Back at the US Department of Energyrsquos AquaticSpecies ProgrammdashBiodiesel from Algae US Department of Energyrsquos Office of Fuels Development Golden COUSA 1998 p 328
66 Vaulot D Olson RJ Merkel S Chisholm SW Cell-cycle response to nutrient starvation in twophytoplankton species Thalassiosira weissflogii and Hymenomonas carterae Mar Biol 1987 95 625ndash630[CrossRef]
67 Olson RJ Chisholm SW Effects of light and nitrogen limitation on the cell cycle of the dinoflagellateAmphidinium carteri J Plankton Res 1986 8 785ndash793 [CrossRef]
68 Li Y Fei X Deng X Novel molecular insights into nitrogen starvation-induced triacylglycerolsaccumulation revealed by differential gene expression analysis in green algae Micractinium pusillumBiomass Bioenergy 2012 42 199ndash211 [CrossRef]
69 Abdelaziz AEM Leite GB Belhaj MA Hallenbeck PC Screening microalgae native to Quebec forwastewater treatment and biodiesel production Bioresour Technol 2014 157 140ndash148 [CrossRef] [PubMed]
70 Valdez-Ojeda R Gonzaacutelez-Muntildeoz M Us-Vaacutezquez R Narvaacuteez-Zapata J Chavarria-Hernandez JCLoacutepez-Adriaacuten S Barahona-Peacuterez F Toledano-Thompson T Garduntildeo-Soloacuterzano G Escobedo-GraciaMedrano RM Characterization of five fresh water microalgae with potential for biodiesel productionAlgal Res 2015 7 33ndash44 [CrossRef]
71 Shrivastav A Mishra SK Suh WI Farooq W Moon M Kim T-H Kumar K Choi G-G Park MSYang J-W Characterization of newly isolated oleaginous microalga Monoraphidium sp for lipid productionunder different conditions Algal Res 2015 12 289ndash294 [CrossRef]
72 Yen H-W Hu I-C Chen C-Y Ho S-H Lee D-J Chang J-S Microalgae-based biorefinerymdashFrombiofuels to natural products Bioresour Technol 2013 135 166ndash174 [CrossRef] [PubMed]
Energies 2017 10 224 16 of 16
73 Li J Liu Y Cheng JJ Mos M Daroch M Biological potential of microalgae in China for biorefinery-basedproduction of biofuels and high value compounds New Biotechnol 2015 32 588ndash596 [CrossRef] [PubMed]
74 El-Sheekh M Abomohra AE-F Hanelt D Optimization of biomass and fatty acid productivity ofScenedesmus obliquus as a promising microalga for biodiesel production World J Microbiol Biotechnol 201329 915ndash922 [CrossRef] [PubMed]
75 Singh P Guldhe A Kumari S Rawat I Bux F Investigation of combined effect of nitrogen phosphorusand iron on lipid productivity of microalgae Ankistrodesmus falcatus KJ671624 using response surfacemethodology Biochem Eng J 2015 94 22ndash29 [CrossRef]
76 Kilham S Kreeger D Goulden C Lynn S Effects of nutrient limitation on biochemical constituents ofAnkistrodesmus falcatus Freshw Biol 1997 38 591ndash596 [CrossRef]
77 Breuer G Lamers PP Martens DE Draaisma RB Wijffels RH The impact of nitrogen starvation on thedynamics of triacylglycerol accumulation in nine microalgae strains Bioresour Technol 2012 124 217ndash226[CrossRef] [PubMed]
78 Breuer G de Jaeger L Artus VG Martens DE Springer J Draaisma RB Eggink G Wijffels RHLamers PP Superior triacylglycerol (TAG) accumulation in starchless mutants of Scenedesmus obliquus (II)evaluation of TAG yield and productivity in controlled photobioreactors Biotechnol Biofuels 2014 7 70[CrossRef] [PubMed]
79 Nigam S Prakash-Rai M Sharma R Effect of Nitrogen on Growth and Lipid Content ofChlorella pyrenoidosa Am J Biochem Biotechnol 2011 7 126ndash131 [CrossRef]
80 Piligaev AV Sorokina KN Bryanskaya AV Peltek SE Kolchanov NA Parmon VN Isolation ofprospective microalgal strains with high saturated fatty acid content for biofuel production Algal Res 201512 368ndash376 [CrossRef]
81 Weiss SB Kennedy EP The enzymatic synthesis of triglycerides J Am Chem Soc 1956 78 3550[CrossRef]
82 Miller R Wu G Deshpande RR Vieler A Gaumlrtner K Li X Moellering ER Zaumluner S Cornish AJLiu B et al Changes in Transcript Abundance in Chlamydomonas reinhardtii following Nitrogen DeprivationPredict Diversion of Metabolism Plant Physiol 2010 154 1737ndash1752 [CrossRef] [PubMed]
83 Rismani-Yazdi H Haznedaroglu BZ Bibby K Peccia J Transcriptome sequencing and annotation of themicroalgae Dunaliella tertiolecta Pathway description and gene discovery for production of next-generationbiofuels BMC Genom 2011 12 148 [CrossRef] [PubMed]
84 Ramos MJ Fernaacutendez CM Casas A Rodriacuteguez L Peacuterez A Influence of fatty acid composition of rawmaterials on biodiesel properties Bioresour Technol 2009 100 261ndash268 [CrossRef] [PubMed]
85 Knothe G Dependence of biodiesel fuel properties on the structure of fatty acid alkyl estersFuel Process Technol 2005 86 1059ndash1070 [CrossRef]
86 Knothe G ldquoDesignerrdquo Biodiesel Optimizing Fatty Ester Composition to Improve Fuel PropertiesEnergy Fuels 2008 22 1358ndash1364 [CrossRef]
87 Smith PC Ngothai Y Dzuy Nguyen Q OrsquoNeill BK Improving the low-temperature properties ofbiodiesel Methods and consequences Renew Energy 2010 35 1145ndash1151 [CrossRef]
88 Dunn RO Effect of antioxidants on the oxidative stability of methyl soyate (biodiesel) Fuel Process Technol2005 86 1071ndash1085 [CrossRef]
89 Knothe G Improving biodiesel fuel properties by modifying fatty ester composition Energy Environ Sci2009 2 759ndash766 [CrossRef]
copy 2017 by the authors licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)
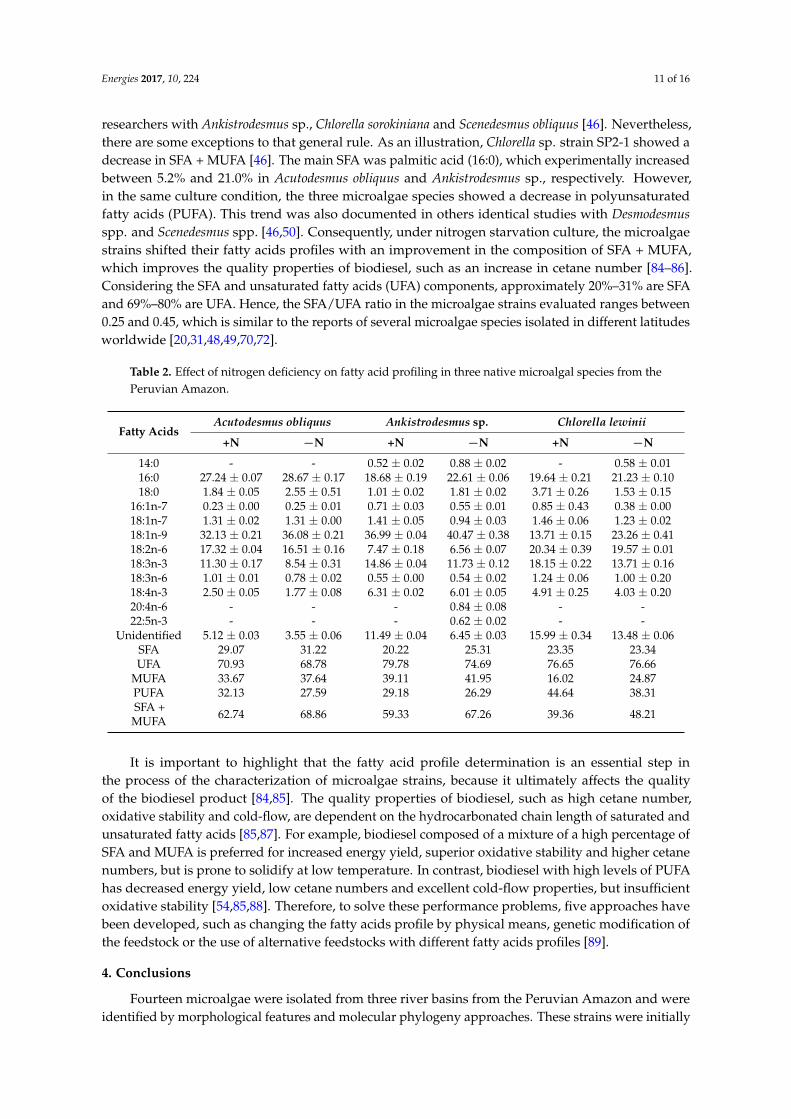
Energies 2017 10 224 11 of 16
researchers with Ankistrodesmus sp Chlorella sorokiniana and Scenedesmus obliquus [46] Neverthelessthere are some exceptions to that general rule As an illustration Chlorella sp strain SP2-1 showed adecrease in SFA + MUFA [46] The main SFA was palmitic acid (160) which experimentally increasedbetween 52 and 210 in Acutodesmus obliquus and Ankistrodesmus sp respectively Howeverin the same culture condition the three microalgae species showed a decrease in polyunsaturatedfatty acids (PUFA) This trend was also documented in others identical studies with Desmodesmusspp and Scenedesmus spp [4650] Consequently under nitrogen starvation culture the microalgaestrains shifted their fatty acids profiles with an improvement in the composition of SFA + MUFAwhich improves the quality properties of biodiesel such as an increase in cetane number [84ndash86]Considering the SFA and unsaturated fatty acids (UFA) components approximately 20ndash31 are SFAand 69ndash80 are UFA Hence the SFAUFA ratio in the microalgae strains evaluated ranges between025 and 045 which is similar to the reports of several microalgae species isolated in different latitudesworldwide [203148497072]
Table 2 Effect of nitrogen deficiency on fatty acid profiling in three native microalgal species from thePeruvian Amazon
Fatty AcidsAcutodesmus obliquus Ankistrodesmus sp Chlorella lewinii
+N minusN +N minusN +N minusN
140 - - 052 plusmn 002 088 plusmn 002 - 058 plusmn 001160 2724 plusmn 007 2867 plusmn 017 1868 plusmn 019 2261 plusmn 006 1964 plusmn 021 2123 plusmn 010180 184 plusmn 005 255 plusmn 051 101 plusmn 002 181 plusmn 002 371 plusmn 026 153 plusmn 015
161n-7 023 plusmn 000 025 plusmn 001 071 plusmn 003 055 plusmn 001 085 plusmn 043 038 plusmn 000181n-7 131 plusmn 002 131 plusmn 000 141 plusmn 005 094 plusmn 003 146 plusmn 006 123 plusmn 002181n-9 3213 plusmn 021 3608 plusmn 021 3699 plusmn 004 4047 plusmn 038 1371 plusmn 015 2326 plusmn 041182n-6 1732 plusmn 004 1651 plusmn 016 747 plusmn 018 656 plusmn 007 2034 plusmn 039 1957 plusmn 001183n-3 1130 plusmn 017 854 plusmn 031 1486 plusmn 004 1173 plusmn 012 1815 plusmn 022 1371 plusmn 016183n-6 101 plusmn 001 078 plusmn 002 055 plusmn 000 054 plusmn 002 124 plusmn 006 100 plusmn 020184n-3 250 plusmn 005 177 plusmn 008 631 plusmn 002 601 plusmn 005 491 plusmn 025 403 plusmn 020204n-6 - - - 084 plusmn 008 - -225n-3 - - - 062 plusmn 002 - -
Unidentified 512 plusmn 003 355 plusmn 006 1149 plusmn 004 645 plusmn 003 1599 plusmn 034 1348 plusmn 006SFA 2907 3122 2022 2531 2335 2334UFA 7093 6878 7978 7469 7665 7666
MUFA 3367 3764 3911 4195 1602 2487PUFA 3213 2759 2918 2629 4464 3831SFA +MUFA 6274 6886 5933 6726 3936 4821
It is important to highlight that the fatty acid profile determination is an essential step inthe process of the characterization of microalgae strains because it ultimately affects the qualityof the biodiesel product [8485] The quality properties of biodiesel such as high cetane numberoxidative stability and cold-flow are dependent on the hydrocarbonated chain length of saturated andunsaturated fatty acids [8587] For example biodiesel composed of a mixture of a high percentage ofSFA and MUFA is preferred for increased energy yield superior oxidative stability and higher cetanenumbers but is prone to solidify at low temperature In contrast biodiesel with high levels of PUFAhas decreased energy yield low cetane numbers and excellent cold-flow properties but insufficientoxidative stability [548588] Therefore to solve these performance problems five approaches havebeen developed such as changing the fatty acids profile by physical means genetic modification ofthe feedstock or the use of alternative feedstocks with different fatty acids profiles [89]
4 Conclusions
Fourteen microalgae were isolated from three river basins from the Peruvian Amazon and wereidentified by morphological features and molecular phylogeny approaches These strains were initially
Energies 2017 10 224 12 of 16
characterized in the stationary growth phase and exhibited significant heterogeneity in key parameterssuch as biomass productivity (36ndash316 mgmiddotLminus1middotdminus1) lipid productivity (10ndash64 mgmiddotLminus1middotdminus1) andtotal lipid content (135ndash467) Furthermore three selected strains based on elevated biomass andlipid productivities were cultured under nitrogen-sufficient and nitrogen-deficient medium to boostlipid accumulation Under these culture conditions all strains showed noticeable decreases in totalprotein content and increases in total lipid and total carbohydrate contents In addition these strainspresented modifications in the fatty acids profiles with improvement in biodiesel properties due to apredominance of SFA and MUFA which corresponds to an increase in cetane number The results ofthis research suggest that the isolated native microalgae from the Peruvian Amazon could be suitablerenewable feedstocks for the production of both biodiesel and bioethanol which could contribute tosustainable development at the local regional and national levels as a viable alternative to petroleumexploitation However it will be necessary to develop open closed or hybrid systems for large-scalebiomass production of selected native microalgae strains andor de novo microalgae strains designedto produce biofuels and with the abilities to synthesize useful bioactive compounds to be efficientlyexploited using biorefinery approaches
Supplementary MaterialsThe following are available online at wwwmdpicom1996-1073102224s1 Figure S1 Results of
electrophoretic and spectrophotometric analysis of the genomic DNA isolated from the fourteen microalgaeFigure S2 ITS2-rDNA structure (A) and amplicons obtained (B) from the microalgae isolated Figure S3Microphotographic images of the three microalgae strains analyzed under light and fluorescence stained withNile Red Figure S4 Spectrophotometric scanning for the determination of the maximum absorption peak foreach microalgae strain
Acknowledgments This research was supported by the Peruvian funding agencies Consejo Nacional de CienciaTecnologiacutea e Innovacioacuten Tecnoloacutegica (CONCYTEC) with Grant Contract No 364-CONCYTEC-OAJ and Fondospara la Innovacioacuten Ciencia y Tecnologiacutea (FINCyT) with Grant Contract No 383-PNICP-PIBA-2014
Author Contributions Marianela Cobos and Juan C Castro obtained funds for the research participated inthe study design coordinated activities and participated in the preparation of the manuscript Jae D Paredesparticipated in sample collection microalgae isolation and helped to draft the manuscript J Dylan Maddoxperformed the bioinformatics analysis and participated in the preparation of the manuscript Gabriel Vargas-Aranaand Leenin Flores participated in biochemical analysis and helped to draft the manuscript Carla P Aguilarparticipated in the study design supervised biochemical analysis and helped to draft the manuscriptJorge L Marapara participated in molecular analysis and helped to draft the manuscript
Conflicts of Interest The authors declare no conflict of interest
References
1 BP Global BP Statisical Review of World Energy 2014 Available online httpwwwbpcomcontentdambppdfEnergy-economicsstatistical-review-2014BP-statistical-review-of-world-energy-2014-full-reportpdf (accessed on 20 June 2014)
2 Pachauri RK Reisinger A Cambio Climaacutetico 2007 Informe de Siacutentesis Contribucioacuten de los Grupos de Trabajo III y III al Cuarto Informe de Evaluacioacuten del Grupo Intergubernamental de Expertos Sobre el Cambio Climaacutetico IPCCGeneva Switzerland 2007 p 104
3 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Ley de Promocioacuten del Mercado deBiocombustibles Available online httpwww2congresogobpesicrcendocbibcon4_uibdnsf9E95620CC059138105257C9E005AB2F9$FILE28054pdf (accessed on 12 February 2017)
4 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Reglamento de la Ley de Promocioacuten del Mercadode Biocombustibles Available online httpwww2osinerggobpeMarcoLegaldocrevDS-013-2005-EM-CONCORDADOpdf (accessed on 12 February 2017)
5 Areacutevalo LF Nalvarte J Torres J Ramiacuterez Y Impactos Socio-Econoacutemicos de la Produccioacuten de Biocombustiblesen la Amazoniacutea Peruana Primera SNV-WWF Lima Peruacute 2009
6 Sharma KK Schuhmann H Schenk PM High Lipid Induction in Microalgae for Biodiesel ProductionEnergies 2012 5 1532ndash1553 [CrossRef]
Energies 2017 10 224 13 of 16
7 Schenk PM Thomas-Hall SR Stephens E Marx UC Mussgnug JH Posten C Kruse O Hankamer BSecond Generation Biofuels High-Efficiency Microalgae for Biodiesel Production BioEnergy Res 2008 120ndash43 [CrossRef]
8 Foley JA Asner GP Costa MH Coe MT de Fries R Gibbs HK Howard EA Olson S Patz JRamankutty N et al Amazonia revealed Forest degradation and loss of ecosystem goods and services inthe Amazon Basin Front Ecol Environ 2007 5 25ndash32 [CrossRef]
9 Duong VT Li Y Nowak E Schenk PM Microalgae Isolation and Selection for Prospective BiodieselProduction Energies 2012 5 1835ndash1849 [CrossRef]
10 Georgianna DR Mayfield SP Exploiting diversity and synthetic biology for the production of algalbiofuels Nature 2012 488 329ndash335 [CrossRef] [PubMed]
11 Wijffels RH Barbosa MJ An Outlook on Microalgal Biofuels Science 2010 329 796ndash799 [CrossRef][PubMed]
12 Richmond A Handbook of Microalgal Culture Biotechnology and Applied Phycology Blackwell Sci Ltd HudsonNJ USA 2004
13 Menetrez MY An overview of algae biofuel production and potential environmental impactEnviron Sci Technol 2012 46 7073ndash7085 [CrossRef] [PubMed]
14 Khoo HH Sharratt PN Das P Balasubramanian RK Naraharisetti PK Shaik S Life cycle energy andCO2 analysis of microalgae-to-biodiesel Preliminary results and comparisons Bioresour Technol 2011 1025800ndash5807 [CrossRef] [PubMed]
15 Chu SP The Influence of the Mineral Composition of the Medium on the Growth of Planktonic AlgaePart I Methods and Culture Media J Ecol 1942 30 284ndash325 [CrossRef]
16 Carlos EMB Mariatildengela M Gecircneros de Algas de Aacuteguas Continentais do BrasilmdashChave Para Identificaccedilatildeo eDescriccedilotildees 2nd ed Rima Sao Carlos Brasil 2006
17 Greenspan P Mayer EP Fowler SD Nile red A selective fluorescent stain for intracellular lipid dropletsJ Cell Biol 1985 100 965ndash973 [CrossRef] [PubMed]
18 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissuePhytochem Bull 1987 19 11ndash15
19 Sambrook J Fritsch EF Maniatis T Molecular Cloning A Laboratory Manual 2nd ed Cold Spring HarborLaboratory Press New York NY USA 1989
20 Kaur S Sarkar M Srivastava RB Gogoi HK Kalita MC Fatty acid profiling and molecularcharacterization of some freshwater microalgae from India with potential for biodiesel productionNew Biotechnol 2012 29 332ndash344 [CrossRef] [PubMed]
21 Benson DA Cavanaugh M Clark K Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW GenBankNucleic Acids Res 2013 41 D36ndashD42 [CrossRef] [PubMed]
22 NCBI Resource Coordinators Database Resources of the National Center for Biotechnology InformationNucleic Acids Res 2015 43 D6ndashD17
23 Ye J McGinnis S Madden TL BLAST Improvements for better sequence analysis Nucleic Acids Res 200634 W6ndashW9 [CrossRef] [PubMed]
24 BioEdit Sequence Alignment Editor for Windows 9598NTXPVista7 Available online httpwwwmbioncsuedubioeditbioedithtml (accessed on 2 December 2015)
25 Sievers F Higgins DG Clustal omega accurate alignment of very large numbers of sequencesMethods Mol Biol (Clifton NJ) 2014 1079 105ndash116
26 Saitou N Nei M The neighbor-joining method A new method for reconstructing phylogenetic treesMol Biol Evol 1987 4 406ndash425 [PubMed]
27 Kimura M A simple method for estimating evolutionary rates of base substitutions through comparativestudies of nucleotide sequences J Mol Evol 1980 16 111ndash120 [CrossRef] [PubMed]
28 Tamura K Stecher G Peterson D Filipski A Kumar S MEGA6 Molecular Evolutionary GeneticsAnalysis version 60 Mol Biol Evol 2013 30 2725ndash2729 [CrossRef] [PubMed]
29 Felsenstein J Confidence Limits on Phylogenies An Approach Using the Bootstrap Evolution 1985 39783ndash791 [CrossRef]
30 Bligh EG Dyer WJ A Rapid Method of Total Lipid Extraction and Purification Can J Biochem Physiol1959 37 911ndash917 [CrossRef] [PubMed]
Energies 2017 10 224 14 of 16
31 Yu X Zhao P He C Li J Tang X Zhou J Huang Z Isolation of a novel strain of Monoraphidium spand characterization of its potential application as biodiesel feedstock Bioresour Technol 2012 121 256ndash262[CrossRef] [PubMed]
32 Hartree EF Determination of protein A modification of the lowry method that gives a linear photometricresponse Anal Biochem 1972 48 422ndash427 [CrossRef]
33 DuBois M Gilles KA Hamilton JK Rebers PA Smith F Colorimetric Method for Determination ofSugars and Related Substances Anal Chem 1956 28 350ndash356 [CrossRef]
34 AOAC International AOAC Official Methods of Analysis of the Association of Official Analytical Chemists (AOAC)15th ed AOAC International Washington DC USA 1990 Volume 1
35 Ichihara K Fukubayashi Y Preparation of fatty acid methyl esters for gas-liquid chromatographyJ Lipid Res 2010 51 635ndash640 [CrossRef] [PubMed]
36 Brennan L Owende P Biofuels from microalgaemdashA review of technologies for production processingand extractions of biofuels and co-products Renew Sustain Energ Rev 2010 14 557ndash577 [CrossRef]
37 Raja R Hemaiswarya S Kumar NA Sridhar S Rengasamy R A perspective on the biotechnologicalpotential of microalgae Crit Rev Microbiol 2008 34 77ndash88 [CrossRef] [PubMed]
38 Mata TM Martins AA Caetano NS Microalgae for biodiesel production and other applicationsA review Renew Sustain Energy Rev 2010 14 217ndash232 [CrossRef]
39 De Wilt A Butkovskyi A Tuantet K Leal LH Fernandes TV Langenhoff A Zeeman GMicropollutant removal in an algal treatment system fed with source separated wastewater streamsJ Hazard Mater 2015 304 84ndash92 [CrossRef] [PubMed]
40 Li T Lin G Podola B Melkonian M Continuous removal of zinc from wastewater and mine dumpleachate by a microalgal biofilm PSBR J Hazard Mater 2015 297 112ndash118 [CrossRef] [PubMed]
41 Sioli H The Amazon and its main affluents Hydrography morphology of the river courses and river typesIn Monographiae Biologicae Sioli H Ed Springer Dordrecht The Netherlands 1984 pp 127ndash165
42 Moniz MBJ Kaczmarska I Barcoding of Diatoms Nuclear Encoded ITS Revisited Protist 2010 161 7ndash34[CrossRef] [PubMed]
43 Coleman AW Is there a molecular key to the level of ldquobiological speciesrdquo in eukaryotes A DNA guideMol Phylogenet Evol 2009 50 197ndash203 [CrossRef] [PubMed]
44 Mai JC Coleman AW The Internal Transcribed Spacer 2 Exhibits a Common Secondary Structure inGreen Algae and Flowering Plants J Mol Evol 1997 44 258ndash271 [CrossRef] [PubMed]
45 Abou-Shanab RAI Matter IA Kim S-N Oh Y-K Choi J Jeon B-H Characterization andidentification of lipid-producing microalgae species isolated from a freshwater lake Biomass Bioenergy2011 35 3079ndash3085 [CrossRef]
46 Do Nascimento M Ortiz-Marquez JCF Sanchez-Rizza L Echarte MM Curatti L Bioprospectingfor fast growing and biomass characterization of oleaginous microalgae from South-Eastern Buenos AiresArgentina Bioresour Technol 2012 125 283ndash290 [CrossRef] [PubMed]
47 Araujo GS Matos LJBL Gonccedilalves LRB Fernandes FAN Farias WRL Bioprospecting foroil producing microalgal strains Evaluation of oil and biomass production for ten microalgal strainsBioresour Technol 2011 102 5248ndash5250 [CrossRef] [PubMed]
48 Zhou W Li Y Min M Hu B Chen P Ruan R Local bioprospecting for high-lipid producing microalgalstrains to be grown on concentrated municipal wastewater for biofuel production Bioresour Technol 2011102 6909ndash6919 [CrossRef] [PubMed]
49 Mahmoud EA Farahat LA Abdel Aziz ZK Fatthallah NA Salah El Din RA Evaluation of thepotential for some isolated microalgae to produce biodiesel Egypt J Pet 2015 24 97ndash101 [CrossRef]
50 Ho S Lai Y-Y Chiang C-Y Chen C-NN Chang J-S Selection of elite microalgae for biodieselproduction in tropical conditions using a standardized platform Bioresour Technol 2013 147 135ndash142[CrossRef] [PubMed]
51 Li L Cui J Liu Q Ding Y Liu J Screening and phylogenetic analysis of lipid-rich microalgae Algal Res2015 11 381ndash386 [CrossRef]
52 Rodolfi L Chini Zittelli G Bassi N Padovani G Biondi N Bonini G Tredici MR Microalgae for oilStrain selection induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactorBiotechnol Bioeng 2009 102 100ndash112 [CrossRef] [PubMed]
Energies 2017 10 224 15 of 16
53 Li P Miao X Li R Zhong J In situ biodiesel production from fast-growing and high oil content Chlorellapyrenoidosa in rice straw hydrolysate J Biomed Biotechnol 2011 2011 141207 [CrossRef] [PubMed]
54 Doan TTY Sivaloganathan B Obbard JP Screening of marine microalgae for biodiesel feedstockBiomass Bioenergy 2011 35 2534ndash2544 [CrossRef]
55 Abou-Shanab RAI Hwang J-H Cho Y Min B Jeon B-H Characterization of microalgal speciesisolated from fresh water bodies as a potential source for biodiesel production Appl Energy 2011 883300ndash3306 [CrossRef]
56 Chaichalerm S Pokethitiyook P Yuan W Meetam M Sritong K Pugkaew W Kungvansaichol KKruatrachue M Damrongphol P Culture of microalgal strains isolated from natural habitats in Thailand invarious enriched media Appl Energy 2012 89 296ndash302 [CrossRef]
57 Zhang S Liu P Yang X Hao Z Zhang L Luo N Shi J Isolation and identification by 18S rDNAsequence of high lipid potential microalgal species for fuel production in Hainan Dao Biomass Bioenergy2014 66 197ndash203 [CrossRef]
58 Chiu S-Y Kao C-Y Tsai M-T Ong S-C Chen C-H Lin C-S Lipid accumulation and CO2 utilizationof Nannochloropsis oculata in response to CO2 aeration Bioresour Technol 2009 100 833ndash838 [CrossRef][PubMed]
59 Huerlimann R de Nys R Heimann K Growth lipid content productivity and fatty acid composition oftropical microalgae for scale-up production Biotechnol Bioeng 2010 107 245ndash257 [CrossRef] [PubMed]
60 Liang Y Sarkany N Cui Y Biomass and lipid productivities of Chlorella vulgaris under autotrophicheterotrophic and mixotrophic growth conditions Biotechnol Lett 2009 31 1043ndash1049 [CrossRef] [PubMed]
61 Hempel N Petrick I Behrendt F Biomass productivity and productivity of fatty acids and amino acidsof microalgae strains as key characteristics of suitability for biodiesel production J Appl Phycol 2012 241407ndash1418 [CrossRef] [PubMed]
62 Yeesang C Cheirsilp B Effect of nitrogen salt and iron content in the growth medium and light intensityon lipid production by microalgae isolated from freshwater sources in Thailand Bioresour Technol 2011 1023034ndash3040 [CrossRef] [PubMed]
63 Msanne J Xu D Konda AR Casas-Mollano JA Awada T Cahoon EB Cerutti H Metabolic andgene expression changes triggered by nitrogen deprivation in the photoautotrophically grown microalgaeChlamydomonas reinhardtii and Coccomyxa sp C-169 Phytochemistry 2012 75 50ndash59 [CrossRef] [PubMed]
64 Lin Q Lin J Effects of nitrogen source and concentration on biomass and oil production of aScenedesmus rubescens like microalga Bioresour Technol 2011 102 1615ndash1621 [CrossRef] [PubMed]
65 Sheehan J Dunahay T Benemann J Roessler P A Look Back at the US Department of Energyrsquos AquaticSpecies ProgrammdashBiodiesel from Algae US Department of Energyrsquos Office of Fuels Development Golden COUSA 1998 p 328
66 Vaulot D Olson RJ Merkel S Chisholm SW Cell-cycle response to nutrient starvation in twophytoplankton species Thalassiosira weissflogii and Hymenomonas carterae Mar Biol 1987 95 625ndash630[CrossRef]
67 Olson RJ Chisholm SW Effects of light and nitrogen limitation on the cell cycle of the dinoflagellateAmphidinium carteri J Plankton Res 1986 8 785ndash793 [CrossRef]
68 Li Y Fei X Deng X Novel molecular insights into nitrogen starvation-induced triacylglycerolsaccumulation revealed by differential gene expression analysis in green algae Micractinium pusillumBiomass Bioenergy 2012 42 199ndash211 [CrossRef]
69 Abdelaziz AEM Leite GB Belhaj MA Hallenbeck PC Screening microalgae native to Quebec forwastewater treatment and biodiesel production Bioresour Technol 2014 157 140ndash148 [CrossRef] [PubMed]
70 Valdez-Ojeda R Gonzaacutelez-Muntildeoz M Us-Vaacutezquez R Narvaacuteez-Zapata J Chavarria-Hernandez JCLoacutepez-Adriaacuten S Barahona-Peacuterez F Toledano-Thompson T Garduntildeo-Soloacuterzano G Escobedo-GraciaMedrano RM Characterization of five fresh water microalgae with potential for biodiesel productionAlgal Res 2015 7 33ndash44 [CrossRef]
71 Shrivastav A Mishra SK Suh WI Farooq W Moon M Kim T-H Kumar K Choi G-G Park MSYang J-W Characterization of newly isolated oleaginous microalga Monoraphidium sp for lipid productionunder different conditions Algal Res 2015 12 289ndash294 [CrossRef]
72 Yen H-W Hu I-C Chen C-Y Ho S-H Lee D-J Chang J-S Microalgae-based biorefinerymdashFrombiofuels to natural products Bioresour Technol 2013 135 166ndash174 [CrossRef] [PubMed]
Energies 2017 10 224 16 of 16
73 Li J Liu Y Cheng JJ Mos M Daroch M Biological potential of microalgae in China for biorefinery-basedproduction of biofuels and high value compounds New Biotechnol 2015 32 588ndash596 [CrossRef] [PubMed]
74 El-Sheekh M Abomohra AE-F Hanelt D Optimization of biomass and fatty acid productivity ofScenedesmus obliquus as a promising microalga for biodiesel production World J Microbiol Biotechnol 201329 915ndash922 [CrossRef] [PubMed]
75 Singh P Guldhe A Kumari S Rawat I Bux F Investigation of combined effect of nitrogen phosphorusand iron on lipid productivity of microalgae Ankistrodesmus falcatus KJ671624 using response surfacemethodology Biochem Eng J 2015 94 22ndash29 [CrossRef]
76 Kilham S Kreeger D Goulden C Lynn S Effects of nutrient limitation on biochemical constituents ofAnkistrodesmus falcatus Freshw Biol 1997 38 591ndash596 [CrossRef]
77 Breuer G Lamers PP Martens DE Draaisma RB Wijffels RH The impact of nitrogen starvation on thedynamics of triacylglycerol accumulation in nine microalgae strains Bioresour Technol 2012 124 217ndash226[CrossRef] [PubMed]
78 Breuer G de Jaeger L Artus VG Martens DE Springer J Draaisma RB Eggink G Wijffels RHLamers PP Superior triacylglycerol (TAG) accumulation in starchless mutants of Scenedesmus obliquus (II)evaluation of TAG yield and productivity in controlled photobioreactors Biotechnol Biofuels 2014 7 70[CrossRef] [PubMed]
79 Nigam S Prakash-Rai M Sharma R Effect of Nitrogen on Growth and Lipid Content ofChlorella pyrenoidosa Am J Biochem Biotechnol 2011 7 126ndash131 [CrossRef]
80 Piligaev AV Sorokina KN Bryanskaya AV Peltek SE Kolchanov NA Parmon VN Isolation ofprospective microalgal strains with high saturated fatty acid content for biofuel production Algal Res 201512 368ndash376 [CrossRef]
81 Weiss SB Kennedy EP The enzymatic synthesis of triglycerides J Am Chem Soc 1956 78 3550[CrossRef]
82 Miller R Wu G Deshpande RR Vieler A Gaumlrtner K Li X Moellering ER Zaumluner S Cornish AJLiu B et al Changes in Transcript Abundance in Chlamydomonas reinhardtii following Nitrogen DeprivationPredict Diversion of Metabolism Plant Physiol 2010 154 1737ndash1752 [CrossRef] [PubMed]
83 Rismani-Yazdi H Haznedaroglu BZ Bibby K Peccia J Transcriptome sequencing and annotation of themicroalgae Dunaliella tertiolecta Pathway description and gene discovery for production of next-generationbiofuels BMC Genom 2011 12 148 [CrossRef] [PubMed]
84 Ramos MJ Fernaacutendez CM Casas A Rodriacuteguez L Peacuterez A Influence of fatty acid composition of rawmaterials on biodiesel properties Bioresour Technol 2009 100 261ndash268 [CrossRef] [PubMed]
85 Knothe G Dependence of biodiesel fuel properties on the structure of fatty acid alkyl estersFuel Process Technol 2005 86 1059ndash1070 [CrossRef]
86 Knothe G ldquoDesignerrdquo Biodiesel Optimizing Fatty Ester Composition to Improve Fuel PropertiesEnergy Fuels 2008 22 1358ndash1364 [CrossRef]
87 Smith PC Ngothai Y Dzuy Nguyen Q OrsquoNeill BK Improving the low-temperature properties ofbiodiesel Methods and consequences Renew Energy 2010 35 1145ndash1151 [CrossRef]
88 Dunn RO Effect of antioxidants on the oxidative stability of methyl soyate (biodiesel) Fuel Process Technol2005 86 1071ndash1085 [CrossRef]
89 Knothe G Improving biodiesel fuel properties by modifying fatty ester composition Energy Environ Sci2009 2 759ndash766 [CrossRef]
copy 2017 by the authors licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)

Energies 2017 10 224 12 of 16
characterized in the stationary growth phase and exhibited significant heterogeneity in key parameterssuch as biomass productivity (36ndash316 mgmiddotLminus1middotdminus1) lipid productivity (10ndash64 mgmiddotLminus1middotdminus1) andtotal lipid content (135ndash467) Furthermore three selected strains based on elevated biomass andlipid productivities were cultured under nitrogen-sufficient and nitrogen-deficient medium to boostlipid accumulation Under these culture conditions all strains showed noticeable decreases in totalprotein content and increases in total lipid and total carbohydrate contents In addition these strainspresented modifications in the fatty acids profiles with improvement in biodiesel properties due to apredominance of SFA and MUFA which corresponds to an increase in cetane number The results ofthis research suggest that the isolated native microalgae from the Peruvian Amazon could be suitablerenewable feedstocks for the production of both biodiesel and bioethanol which could contribute tosustainable development at the local regional and national levels as a viable alternative to petroleumexploitation However it will be necessary to develop open closed or hybrid systems for large-scalebiomass production of selected native microalgae strains andor de novo microalgae strains designedto produce biofuels and with the abilities to synthesize useful bioactive compounds to be efficientlyexploited using biorefinery approaches
Supplementary MaterialsThe following are available online at wwwmdpicom1996-1073102224s1 Figure S1 Results of
electrophoretic and spectrophotometric analysis of the genomic DNA isolated from the fourteen microalgaeFigure S2 ITS2-rDNA structure (A) and amplicons obtained (B) from the microalgae isolated Figure S3Microphotographic images of the three microalgae strains analyzed under light and fluorescence stained withNile Red Figure S4 Spectrophotometric scanning for the determination of the maximum absorption peak foreach microalgae strain
Acknowledgments This research was supported by the Peruvian funding agencies Consejo Nacional de CienciaTecnologiacutea e Innovacioacuten Tecnoloacutegica (CONCYTEC) with Grant Contract No 364-CONCYTEC-OAJ and Fondospara la Innovacioacuten Ciencia y Tecnologiacutea (FINCyT) with Grant Contract No 383-PNICP-PIBA-2014
Author Contributions Marianela Cobos and Juan C Castro obtained funds for the research participated inthe study design coordinated activities and participated in the preparation of the manuscript Jae D Paredesparticipated in sample collection microalgae isolation and helped to draft the manuscript J Dylan Maddoxperformed the bioinformatics analysis and participated in the preparation of the manuscript Gabriel Vargas-Aranaand Leenin Flores participated in biochemical analysis and helped to draft the manuscript Carla P Aguilarparticipated in the study design supervised biochemical analysis and helped to draft the manuscriptJorge L Marapara participated in molecular analysis and helped to draft the manuscript
Conflicts of Interest The authors declare no conflict of interest
References
1 BP Global BP Statisical Review of World Energy 2014 Available online httpwwwbpcomcontentdambppdfEnergy-economicsstatistical-review-2014BP-statistical-review-of-world-energy-2014-full-reportpdf (accessed on 20 June 2014)
2 Pachauri RK Reisinger A Cambio Climaacutetico 2007 Informe de Siacutentesis Contribucioacuten de los Grupos de Trabajo III y III al Cuarto Informe de Evaluacioacuten del Grupo Intergubernamental de Expertos Sobre el Cambio Climaacutetico IPCCGeneva Switzerland 2007 p 104
3 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Ley de Promocioacuten del Mercado deBiocombustibles Available online httpwww2congresogobpesicrcendocbibcon4_uibdnsf9E95620CC059138105257C9E005AB2F9$FILE28054pdf (accessed on 12 February 2017)
4 Peruacute Comisioacuten Permanente del Congreso de la Repuacuteblica Reglamento de la Ley de Promocioacuten del Mercadode Biocombustibles Available online httpwww2osinerggobpeMarcoLegaldocrevDS-013-2005-EM-CONCORDADOpdf (accessed on 12 February 2017)
5 Areacutevalo LF Nalvarte J Torres J Ramiacuterez Y Impactos Socio-Econoacutemicos de la Produccioacuten de Biocombustiblesen la Amazoniacutea Peruana Primera SNV-WWF Lima Peruacute 2009
6 Sharma KK Schuhmann H Schenk PM High Lipid Induction in Microalgae for Biodiesel ProductionEnergies 2012 5 1532ndash1553 [CrossRef]
Energies 2017 10 224 13 of 16
7 Schenk PM Thomas-Hall SR Stephens E Marx UC Mussgnug JH Posten C Kruse O Hankamer BSecond Generation Biofuels High-Efficiency Microalgae for Biodiesel Production BioEnergy Res 2008 120ndash43 [CrossRef]
8 Foley JA Asner GP Costa MH Coe MT de Fries R Gibbs HK Howard EA Olson S Patz JRamankutty N et al Amazonia revealed Forest degradation and loss of ecosystem goods and services inthe Amazon Basin Front Ecol Environ 2007 5 25ndash32 [CrossRef]
9 Duong VT Li Y Nowak E Schenk PM Microalgae Isolation and Selection for Prospective BiodieselProduction Energies 2012 5 1835ndash1849 [CrossRef]
10 Georgianna DR Mayfield SP Exploiting diversity and synthetic biology for the production of algalbiofuels Nature 2012 488 329ndash335 [CrossRef] [PubMed]
11 Wijffels RH Barbosa MJ An Outlook on Microalgal Biofuels Science 2010 329 796ndash799 [CrossRef][PubMed]
12 Richmond A Handbook of Microalgal Culture Biotechnology and Applied Phycology Blackwell Sci Ltd HudsonNJ USA 2004
13 Menetrez MY An overview of algae biofuel production and potential environmental impactEnviron Sci Technol 2012 46 7073ndash7085 [CrossRef] [PubMed]
14 Khoo HH Sharratt PN Das P Balasubramanian RK Naraharisetti PK Shaik S Life cycle energy andCO2 analysis of microalgae-to-biodiesel Preliminary results and comparisons Bioresour Technol 2011 1025800ndash5807 [CrossRef] [PubMed]
15 Chu SP The Influence of the Mineral Composition of the Medium on the Growth of Planktonic AlgaePart I Methods and Culture Media J Ecol 1942 30 284ndash325 [CrossRef]
16 Carlos EMB Mariatildengela M Gecircneros de Algas de Aacuteguas Continentais do BrasilmdashChave Para Identificaccedilatildeo eDescriccedilotildees 2nd ed Rima Sao Carlos Brasil 2006
17 Greenspan P Mayer EP Fowler SD Nile red A selective fluorescent stain for intracellular lipid dropletsJ Cell Biol 1985 100 965ndash973 [CrossRef] [PubMed]
18 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissuePhytochem Bull 1987 19 11ndash15
19 Sambrook J Fritsch EF Maniatis T Molecular Cloning A Laboratory Manual 2nd ed Cold Spring HarborLaboratory Press New York NY USA 1989
20 Kaur S Sarkar M Srivastava RB Gogoi HK Kalita MC Fatty acid profiling and molecularcharacterization of some freshwater microalgae from India with potential for biodiesel productionNew Biotechnol 2012 29 332ndash344 [CrossRef] [PubMed]
21 Benson DA Cavanaugh M Clark K Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW GenBankNucleic Acids Res 2013 41 D36ndashD42 [CrossRef] [PubMed]
22 NCBI Resource Coordinators Database Resources of the National Center for Biotechnology InformationNucleic Acids Res 2015 43 D6ndashD17
23 Ye J McGinnis S Madden TL BLAST Improvements for better sequence analysis Nucleic Acids Res 200634 W6ndashW9 [CrossRef] [PubMed]
24 BioEdit Sequence Alignment Editor for Windows 9598NTXPVista7 Available online httpwwwmbioncsuedubioeditbioedithtml (accessed on 2 December 2015)
25 Sievers F Higgins DG Clustal omega accurate alignment of very large numbers of sequencesMethods Mol Biol (Clifton NJ) 2014 1079 105ndash116
26 Saitou N Nei M The neighbor-joining method A new method for reconstructing phylogenetic treesMol Biol Evol 1987 4 406ndash425 [PubMed]
27 Kimura M A simple method for estimating evolutionary rates of base substitutions through comparativestudies of nucleotide sequences J Mol Evol 1980 16 111ndash120 [CrossRef] [PubMed]
28 Tamura K Stecher G Peterson D Filipski A Kumar S MEGA6 Molecular Evolutionary GeneticsAnalysis version 60 Mol Biol Evol 2013 30 2725ndash2729 [CrossRef] [PubMed]
29 Felsenstein J Confidence Limits on Phylogenies An Approach Using the Bootstrap Evolution 1985 39783ndash791 [CrossRef]
30 Bligh EG Dyer WJ A Rapid Method of Total Lipid Extraction and Purification Can J Biochem Physiol1959 37 911ndash917 [CrossRef] [PubMed]
Energies 2017 10 224 14 of 16
31 Yu X Zhao P He C Li J Tang X Zhou J Huang Z Isolation of a novel strain of Monoraphidium spand characterization of its potential application as biodiesel feedstock Bioresour Technol 2012 121 256ndash262[CrossRef] [PubMed]
32 Hartree EF Determination of protein A modification of the lowry method that gives a linear photometricresponse Anal Biochem 1972 48 422ndash427 [CrossRef]
33 DuBois M Gilles KA Hamilton JK Rebers PA Smith F Colorimetric Method for Determination ofSugars and Related Substances Anal Chem 1956 28 350ndash356 [CrossRef]
34 AOAC International AOAC Official Methods of Analysis of the Association of Official Analytical Chemists (AOAC)15th ed AOAC International Washington DC USA 1990 Volume 1
35 Ichihara K Fukubayashi Y Preparation of fatty acid methyl esters for gas-liquid chromatographyJ Lipid Res 2010 51 635ndash640 [CrossRef] [PubMed]
36 Brennan L Owende P Biofuels from microalgaemdashA review of technologies for production processingand extractions of biofuels and co-products Renew Sustain Energ Rev 2010 14 557ndash577 [CrossRef]
37 Raja R Hemaiswarya S Kumar NA Sridhar S Rengasamy R A perspective on the biotechnologicalpotential of microalgae Crit Rev Microbiol 2008 34 77ndash88 [CrossRef] [PubMed]
38 Mata TM Martins AA Caetano NS Microalgae for biodiesel production and other applicationsA review Renew Sustain Energy Rev 2010 14 217ndash232 [CrossRef]
39 De Wilt A Butkovskyi A Tuantet K Leal LH Fernandes TV Langenhoff A Zeeman GMicropollutant removal in an algal treatment system fed with source separated wastewater streamsJ Hazard Mater 2015 304 84ndash92 [CrossRef] [PubMed]
40 Li T Lin G Podola B Melkonian M Continuous removal of zinc from wastewater and mine dumpleachate by a microalgal biofilm PSBR J Hazard Mater 2015 297 112ndash118 [CrossRef] [PubMed]
41 Sioli H The Amazon and its main affluents Hydrography morphology of the river courses and river typesIn Monographiae Biologicae Sioli H Ed Springer Dordrecht The Netherlands 1984 pp 127ndash165
42 Moniz MBJ Kaczmarska I Barcoding of Diatoms Nuclear Encoded ITS Revisited Protist 2010 161 7ndash34[CrossRef] [PubMed]
43 Coleman AW Is there a molecular key to the level of ldquobiological speciesrdquo in eukaryotes A DNA guideMol Phylogenet Evol 2009 50 197ndash203 [CrossRef] [PubMed]
44 Mai JC Coleman AW The Internal Transcribed Spacer 2 Exhibits a Common Secondary Structure inGreen Algae and Flowering Plants J Mol Evol 1997 44 258ndash271 [CrossRef] [PubMed]
45 Abou-Shanab RAI Matter IA Kim S-N Oh Y-K Choi J Jeon B-H Characterization andidentification of lipid-producing microalgae species isolated from a freshwater lake Biomass Bioenergy2011 35 3079ndash3085 [CrossRef]
46 Do Nascimento M Ortiz-Marquez JCF Sanchez-Rizza L Echarte MM Curatti L Bioprospectingfor fast growing and biomass characterization of oleaginous microalgae from South-Eastern Buenos AiresArgentina Bioresour Technol 2012 125 283ndash290 [CrossRef] [PubMed]
47 Araujo GS Matos LJBL Gonccedilalves LRB Fernandes FAN Farias WRL Bioprospecting foroil producing microalgal strains Evaluation of oil and biomass production for ten microalgal strainsBioresour Technol 2011 102 5248ndash5250 [CrossRef] [PubMed]
48 Zhou W Li Y Min M Hu B Chen P Ruan R Local bioprospecting for high-lipid producing microalgalstrains to be grown on concentrated municipal wastewater for biofuel production Bioresour Technol 2011102 6909ndash6919 [CrossRef] [PubMed]
49 Mahmoud EA Farahat LA Abdel Aziz ZK Fatthallah NA Salah El Din RA Evaluation of thepotential for some isolated microalgae to produce biodiesel Egypt J Pet 2015 24 97ndash101 [CrossRef]
50 Ho S Lai Y-Y Chiang C-Y Chen C-NN Chang J-S Selection of elite microalgae for biodieselproduction in tropical conditions using a standardized platform Bioresour Technol 2013 147 135ndash142[CrossRef] [PubMed]
51 Li L Cui J Liu Q Ding Y Liu J Screening and phylogenetic analysis of lipid-rich microalgae Algal Res2015 11 381ndash386 [CrossRef]
52 Rodolfi L Chini Zittelli G Bassi N Padovani G Biondi N Bonini G Tredici MR Microalgae for oilStrain selection induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactorBiotechnol Bioeng 2009 102 100ndash112 [CrossRef] [PubMed]
Energies 2017 10 224 15 of 16
53 Li P Miao X Li R Zhong J In situ biodiesel production from fast-growing and high oil content Chlorellapyrenoidosa in rice straw hydrolysate J Biomed Biotechnol 2011 2011 141207 [CrossRef] [PubMed]
54 Doan TTY Sivaloganathan B Obbard JP Screening of marine microalgae for biodiesel feedstockBiomass Bioenergy 2011 35 2534ndash2544 [CrossRef]
55 Abou-Shanab RAI Hwang J-H Cho Y Min B Jeon B-H Characterization of microalgal speciesisolated from fresh water bodies as a potential source for biodiesel production Appl Energy 2011 883300ndash3306 [CrossRef]
56 Chaichalerm S Pokethitiyook P Yuan W Meetam M Sritong K Pugkaew W Kungvansaichol KKruatrachue M Damrongphol P Culture of microalgal strains isolated from natural habitats in Thailand invarious enriched media Appl Energy 2012 89 296ndash302 [CrossRef]
57 Zhang S Liu P Yang X Hao Z Zhang L Luo N Shi J Isolation and identification by 18S rDNAsequence of high lipid potential microalgal species for fuel production in Hainan Dao Biomass Bioenergy2014 66 197ndash203 [CrossRef]
58 Chiu S-Y Kao C-Y Tsai M-T Ong S-C Chen C-H Lin C-S Lipid accumulation and CO2 utilizationof Nannochloropsis oculata in response to CO2 aeration Bioresour Technol 2009 100 833ndash838 [CrossRef][PubMed]
59 Huerlimann R de Nys R Heimann K Growth lipid content productivity and fatty acid composition oftropical microalgae for scale-up production Biotechnol Bioeng 2010 107 245ndash257 [CrossRef] [PubMed]
60 Liang Y Sarkany N Cui Y Biomass and lipid productivities of Chlorella vulgaris under autotrophicheterotrophic and mixotrophic growth conditions Biotechnol Lett 2009 31 1043ndash1049 [CrossRef] [PubMed]
61 Hempel N Petrick I Behrendt F Biomass productivity and productivity of fatty acids and amino acidsof microalgae strains as key characteristics of suitability for biodiesel production J Appl Phycol 2012 241407ndash1418 [CrossRef] [PubMed]
62 Yeesang C Cheirsilp B Effect of nitrogen salt and iron content in the growth medium and light intensityon lipid production by microalgae isolated from freshwater sources in Thailand Bioresour Technol 2011 1023034ndash3040 [CrossRef] [PubMed]
63 Msanne J Xu D Konda AR Casas-Mollano JA Awada T Cahoon EB Cerutti H Metabolic andgene expression changes triggered by nitrogen deprivation in the photoautotrophically grown microalgaeChlamydomonas reinhardtii and Coccomyxa sp C-169 Phytochemistry 2012 75 50ndash59 [CrossRef] [PubMed]
64 Lin Q Lin J Effects of nitrogen source and concentration on biomass and oil production of aScenedesmus rubescens like microalga Bioresour Technol 2011 102 1615ndash1621 [CrossRef] [PubMed]
65 Sheehan J Dunahay T Benemann J Roessler P A Look Back at the US Department of Energyrsquos AquaticSpecies ProgrammdashBiodiesel from Algae US Department of Energyrsquos Office of Fuels Development Golden COUSA 1998 p 328
66 Vaulot D Olson RJ Merkel S Chisholm SW Cell-cycle response to nutrient starvation in twophytoplankton species Thalassiosira weissflogii and Hymenomonas carterae Mar Biol 1987 95 625ndash630[CrossRef]
67 Olson RJ Chisholm SW Effects of light and nitrogen limitation on the cell cycle of the dinoflagellateAmphidinium carteri J Plankton Res 1986 8 785ndash793 [CrossRef]
68 Li Y Fei X Deng X Novel molecular insights into nitrogen starvation-induced triacylglycerolsaccumulation revealed by differential gene expression analysis in green algae Micractinium pusillumBiomass Bioenergy 2012 42 199ndash211 [CrossRef]
69 Abdelaziz AEM Leite GB Belhaj MA Hallenbeck PC Screening microalgae native to Quebec forwastewater treatment and biodiesel production Bioresour Technol 2014 157 140ndash148 [CrossRef] [PubMed]
70 Valdez-Ojeda R Gonzaacutelez-Muntildeoz M Us-Vaacutezquez R Narvaacuteez-Zapata J Chavarria-Hernandez JCLoacutepez-Adriaacuten S Barahona-Peacuterez F Toledano-Thompson T Garduntildeo-Soloacuterzano G Escobedo-GraciaMedrano RM Characterization of five fresh water microalgae with potential for biodiesel productionAlgal Res 2015 7 33ndash44 [CrossRef]
71 Shrivastav A Mishra SK Suh WI Farooq W Moon M Kim T-H Kumar K Choi G-G Park MSYang J-W Characterization of newly isolated oleaginous microalga Monoraphidium sp for lipid productionunder different conditions Algal Res 2015 12 289ndash294 [CrossRef]
72 Yen H-W Hu I-C Chen C-Y Ho S-H Lee D-J Chang J-S Microalgae-based biorefinerymdashFrombiofuels to natural products Bioresour Technol 2013 135 166ndash174 [CrossRef] [PubMed]
Energies 2017 10 224 16 of 16
73 Li J Liu Y Cheng JJ Mos M Daroch M Biological potential of microalgae in China for biorefinery-basedproduction of biofuels and high value compounds New Biotechnol 2015 32 588ndash596 [CrossRef] [PubMed]
74 El-Sheekh M Abomohra AE-F Hanelt D Optimization of biomass and fatty acid productivity ofScenedesmus obliquus as a promising microalga for biodiesel production World J Microbiol Biotechnol 201329 915ndash922 [CrossRef] [PubMed]
75 Singh P Guldhe A Kumari S Rawat I Bux F Investigation of combined effect of nitrogen phosphorusand iron on lipid productivity of microalgae Ankistrodesmus falcatus KJ671624 using response surfacemethodology Biochem Eng J 2015 94 22ndash29 [CrossRef]
76 Kilham S Kreeger D Goulden C Lynn S Effects of nutrient limitation on biochemical constituents ofAnkistrodesmus falcatus Freshw Biol 1997 38 591ndash596 [CrossRef]
77 Breuer G Lamers PP Martens DE Draaisma RB Wijffels RH The impact of nitrogen starvation on thedynamics of triacylglycerol accumulation in nine microalgae strains Bioresour Technol 2012 124 217ndash226[CrossRef] [PubMed]
78 Breuer G de Jaeger L Artus VG Martens DE Springer J Draaisma RB Eggink G Wijffels RHLamers PP Superior triacylglycerol (TAG) accumulation in starchless mutants of Scenedesmus obliquus (II)evaluation of TAG yield and productivity in controlled photobioreactors Biotechnol Biofuels 2014 7 70[CrossRef] [PubMed]
79 Nigam S Prakash-Rai M Sharma R Effect of Nitrogen on Growth and Lipid Content ofChlorella pyrenoidosa Am J Biochem Biotechnol 2011 7 126ndash131 [CrossRef]
80 Piligaev AV Sorokina KN Bryanskaya AV Peltek SE Kolchanov NA Parmon VN Isolation ofprospective microalgal strains with high saturated fatty acid content for biofuel production Algal Res 201512 368ndash376 [CrossRef]
81 Weiss SB Kennedy EP The enzymatic synthesis of triglycerides J Am Chem Soc 1956 78 3550[CrossRef]
82 Miller R Wu G Deshpande RR Vieler A Gaumlrtner K Li X Moellering ER Zaumluner S Cornish AJLiu B et al Changes in Transcript Abundance in Chlamydomonas reinhardtii following Nitrogen DeprivationPredict Diversion of Metabolism Plant Physiol 2010 154 1737ndash1752 [CrossRef] [PubMed]
83 Rismani-Yazdi H Haznedaroglu BZ Bibby K Peccia J Transcriptome sequencing and annotation of themicroalgae Dunaliella tertiolecta Pathway description and gene discovery for production of next-generationbiofuels BMC Genom 2011 12 148 [CrossRef] [PubMed]
84 Ramos MJ Fernaacutendez CM Casas A Rodriacuteguez L Peacuterez A Influence of fatty acid composition of rawmaterials on biodiesel properties Bioresour Technol 2009 100 261ndash268 [CrossRef] [PubMed]
85 Knothe G Dependence of biodiesel fuel properties on the structure of fatty acid alkyl estersFuel Process Technol 2005 86 1059ndash1070 [CrossRef]
86 Knothe G ldquoDesignerrdquo Biodiesel Optimizing Fatty Ester Composition to Improve Fuel PropertiesEnergy Fuels 2008 22 1358ndash1364 [CrossRef]
87 Smith PC Ngothai Y Dzuy Nguyen Q OrsquoNeill BK Improving the low-temperature properties ofbiodiesel Methods and consequences Renew Energy 2010 35 1145ndash1151 [CrossRef]
88 Dunn RO Effect of antioxidants on the oxidative stability of methyl soyate (biodiesel) Fuel Process Technol2005 86 1071ndash1085 [CrossRef]
89 Knothe G Improving biodiesel fuel properties by modifying fatty ester composition Energy Environ Sci2009 2 759ndash766 [CrossRef]
copy 2017 by the authors licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)

Energies 2017 10 224 13 of 16
7 Schenk PM Thomas-Hall SR Stephens E Marx UC Mussgnug JH Posten C Kruse O Hankamer BSecond Generation Biofuels High-Efficiency Microalgae for Biodiesel Production BioEnergy Res 2008 120ndash43 [CrossRef]
8 Foley JA Asner GP Costa MH Coe MT de Fries R Gibbs HK Howard EA Olson S Patz JRamankutty N et al Amazonia revealed Forest degradation and loss of ecosystem goods and services inthe Amazon Basin Front Ecol Environ 2007 5 25ndash32 [CrossRef]
9 Duong VT Li Y Nowak E Schenk PM Microalgae Isolation and Selection for Prospective BiodieselProduction Energies 2012 5 1835ndash1849 [CrossRef]
10 Georgianna DR Mayfield SP Exploiting diversity and synthetic biology for the production of algalbiofuels Nature 2012 488 329ndash335 [CrossRef] [PubMed]
11 Wijffels RH Barbosa MJ An Outlook on Microalgal Biofuels Science 2010 329 796ndash799 [CrossRef][PubMed]
12 Richmond A Handbook of Microalgal Culture Biotechnology and Applied Phycology Blackwell Sci Ltd HudsonNJ USA 2004
13 Menetrez MY An overview of algae biofuel production and potential environmental impactEnviron Sci Technol 2012 46 7073ndash7085 [CrossRef] [PubMed]
14 Khoo HH Sharratt PN Das P Balasubramanian RK Naraharisetti PK Shaik S Life cycle energy andCO2 analysis of microalgae-to-biodiesel Preliminary results and comparisons Bioresour Technol 2011 1025800ndash5807 [CrossRef] [PubMed]
15 Chu SP The Influence of the Mineral Composition of the Medium on the Growth of Planktonic AlgaePart I Methods and Culture Media J Ecol 1942 30 284ndash325 [CrossRef]
16 Carlos EMB Mariatildengela M Gecircneros de Algas de Aacuteguas Continentais do BrasilmdashChave Para Identificaccedilatildeo eDescriccedilotildees 2nd ed Rima Sao Carlos Brasil 2006
17 Greenspan P Mayer EP Fowler SD Nile red A selective fluorescent stain for intracellular lipid dropletsJ Cell Biol 1985 100 965ndash973 [CrossRef] [PubMed]
18 Doyle JJ Doyle JL A rapid DNA isolation procedure for small quantities of fresh leaf tissuePhytochem Bull 1987 19 11ndash15
19 Sambrook J Fritsch EF Maniatis T Molecular Cloning A Laboratory Manual 2nd ed Cold Spring HarborLaboratory Press New York NY USA 1989
20 Kaur S Sarkar M Srivastava RB Gogoi HK Kalita MC Fatty acid profiling and molecularcharacterization of some freshwater microalgae from India with potential for biodiesel productionNew Biotechnol 2012 29 332ndash344 [CrossRef] [PubMed]
21 Benson DA Cavanaugh M Clark K Karsch-Mizrachi I Lipman DJ Ostell J Sayers EW GenBankNucleic Acids Res 2013 41 D36ndashD42 [CrossRef] [PubMed]
22 NCBI Resource Coordinators Database Resources of the National Center for Biotechnology InformationNucleic Acids Res 2015 43 D6ndashD17
23 Ye J McGinnis S Madden TL BLAST Improvements for better sequence analysis Nucleic Acids Res 200634 W6ndashW9 [CrossRef] [PubMed]
24 BioEdit Sequence Alignment Editor for Windows 9598NTXPVista7 Available online httpwwwmbioncsuedubioeditbioedithtml (accessed on 2 December 2015)
25 Sievers F Higgins DG Clustal omega accurate alignment of very large numbers of sequencesMethods Mol Biol (Clifton NJ) 2014 1079 105ndash116
26 Saitou N Nei M The neighbor-joining method A new method for reconstructing phylogenetic treesMol Biol Evol 1987 4 406ndash425 [PubMed]
27 Kimura M A simple method for estimating evolutionary rates of base substitutions through comparativestudies of nucleotide sequences J Mol Evol 1980 16 111ndash120 [CrossRef] [PubMed]
28 Tamura K Stecher G Peterson D Filipski A Kumar S MEGA6 Molecular Evolutionary GeneticsAnalysis version 60 Mol Biol Evol 2013 30 2725ndash2729 [CrossRef] [PubMed]
29 Felsenstein J Confidence Limits on Phylogenies An Approach Using the Bootstrap Evolution 1985 39783ndash791 [CrossRef]
30 Bligh EG Dyer WJ A Rapid Method of Total Lipid Extraction and Purification Can J Biochem Physiol1959 37 911ndash917 [CrossRef] [PubMed]
Energies 2017 10 224 14 of 16
31 Yu X Zhao P He C Li J Tang X Zhou J Huang Z Isolation of a novel strain of Monoraphidium spand characterization of its potential application as biodiesel feedstock Bioresour Technol 2012 121 256ndash262[CrossRef] [PubMed]
32 Hartree EF Determination of protein A modification of the lowry method that gives a linear photometricresponse Anal Biochem 1972 48 422ndash427 [CrossRef]
33 DuBois M Gilles KA Hamilton JK Rebers PA Smith F Colorimetric Method for Determination ofSugars and Related Substances Anal Chem 1956 28 350ndash356 [CrossRef]
34 AOAC International AOAC Official Methods of Analysis of the Association of Official Analytical Chemists (AOAC)15th ed AOAC International Washington DC USA 1990 Volume 1
35 Ichihara K Fukubayashi Y Preparation of fatty acid methyl esters for gas-liquid chromatographyJ Lipid Res 2010 51 635ndash640 [CrossRef] [PubMed]
36 Brennan L Owende P Biofuels from microalgaemdashA review of technologies for production processingand extractions of biofuels and co-products Renew Sustain Energ Rev 2010 14 557ndash577 [CrossRef]
37 Raja R Hemaiswarya S Kumar NA Sridhar S Rengasamy R A perspective on the biotechnologicalpotential of microalgae Crit Rev Microbiol 2008 34 77ndash88 [CrossRef] [PubMed]
38 Mata TM Martins AA Caetano NS Microalgae for biodiesel production and other applicationsA review Renew Sustain Energy Rev 2010 14 217ndash232 [CrossRef]
39 De Wilt A Butkovskyi A Tuantet K Leal LH Fernandes TV Langenhoff A Zeeman GMicropollutant removal in an algal treatment system fed with source separated wastewater streamsJ Hazard Mater 2015 304 84ndash92 [CrossRef] [PubMed]
40 Li T Lin G Podola B Melkonian M Continuous removal of zinc from wastewater and mine dumpleachate by a microalgal biofilm PSBR J Hazard Mater 2015 297 112ndash118 [CrossRef] [PubMed]
41 Sioli H The Amazon and its main affluents Hydrography morphology of the river courses and river typesIn Monographiae Biologicae Sioli H Ed Springer Dordrecht The Netherlands 1984 pp 127ndash165
42 Moniz MBJ Kaczmarska I Barcoding of Diatoms Nuclear Encoded ITS Revisited Protist 2010 161 7ndash34[CrossRef] [PubMed]
43 Coleman AW Is there a molecular key to the level of ldquobiological speciesrdquo in eukaryotes A DNA guideMol Phylogenet Evol 2009 50 197ndash203 [CrossRef] [PubMed]
44 Mai JC Coleman AW The Internal Transcribed Spacer 2 Exhibits a Common Secondary Structure inGreen Algae and Flowering Plants J Mol Evol 1997 44 258ndash271 [CrossRef] [PubMed]
45 Abou-Shanab RAI Matter IA Kim S-N Oh Y-K Choi J Jeon B-H Characterization andidentification of lipid-producing microalgae species isolated from a freshwater lake Biomass Bioenergy2011 35 3079ndash3085 [CrossRef]
46 Do Nascimento M Ortiz-Marquez JCF Sanchez-Rizza L Echarte MM Curatti L Bioprospectingfor fast growing and biomass characterization of oleaginous microalgae from South-Eastern Buenos AiresArgentina Bioresour Technol 2012 125 283ndash290 [CrossRef] [PubMed]
47 Araujo GS Matos LJBL Gonccedilalves LRB Fernandes FAN Farias WRL Bioprospecting foroil producing microalgal strains Evaluation of oil and biomass production for ten microalgal strainsBioresour Technol 2011 102 5248ndash5250 [CrossRef] [PubMed]
48 Zhou W Li Y Min M Hu B Chen P Ruan R Local bioprospecting for high-lipid producing microalgalstrains to be grown on concentrated municipal wastewater for biofuel production Bioresour Technol 2011102 6909ndash6919 [CrossRef] [PubMed]
49 Mahmoud EA Farahat LA Abdel Aziz ZK Fatthallah NA Salah El Din RA Evaluation of thepotential for some isolated microalgae to produce biodiesel Egypt J Pet 2015 24 97ndash101 [CrossRef]
50 Ho S Lai Y-Y Chiang C-Y Chen C-NN Chang J-S Selection of elite microalgae for biodieselproduction in tropical conditions using a standardized platform Bioresour Technol 2013 147 135ndash142[CrossRef] [PubMed]
51 Li L Cui J Liu Q Ding Y Liu J Screening and phylogenetic analysis of lipid-rich microalgae Algal Res2015 11 381ndash386 [CrossRef]
52 Rodolfi L Chini Zittelli G Bassi N Padovani G Biondi N Bonini G Tredici MR Microalgae for oilStrain selection induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactorBiotechnol Bioeng 2009 102 100ndash112 [CrossRef] [PubMed]
Energies 2017 10 224 15 of 16
53 Li P Miao X Li R Zhong J In situ biodiesel production from fast-growing and high oil content Chlorellapyrenoidosa in rice straw hydrolysate J Biomed Biotechnol 2011 2011 141207 [CrossRef] [PubMed]
54 Doan TTY Sivaloganathan B Obbard JP Screening of marine microalgae for biodiesel feedstockBiomass Bioenergy 2011 35 2534ndash2544 [CrossRef]
55 Abou-Shanab RAI Hwang J-H Cho Y Min B Jeon B-H Characterization of microalgal speciesisolated from fresh water bodies as a potential source for biodiesel production Appl Energy 2011 883300ndash3306 [CrossRef]
56 Chaichalerm S Pokethitiyook P Yuan W Meetam M Sritong K Pugkaew W Kungvansaichol KKruatrachue M Damrongphol P Culture of microalgal strains isolated from natural habitats in Thailand invarious enriched media Appl Energy 2012 89 296ndash302 [CrossRef]
57 Zhang S Liu P Yang X Hao Z Zhang L Luo N Shi J Isolation and identification by 18S rDNAsequence of high lipid potential microalgal species for fuel production in Hainan Dao Biomass Bioenergy2014 66 197ndash203 [CrossRef]
58 Chiu S-Y Kao C-Y Tsai M-T Ong S-C Chen C-H Lin C-S Lipid accumulation and CO2 utilizationof Nannochloropsis oculata in response to CO2 aeration Bioresour Technol 2009 100 833ndash838 [CrossRef][PubMed]
59 Huerlimann R de Nys R Heimann K Growth lipid content productivity and fatty acid composition oftropical microalgae for scale-up production Biotechnol Bioeng 2010 107 245ndash257 [CrossRef] [PubMed]
60 Liang Y Sarkany N Cui Y Biomass and lipid productivities of Chlorella vulgaris under autotrophicheterotrophic and mixotrophic growth conditions Biotechnol Lett 2009 31 1043ndash1049 [CrossRef] [PubMed]
61 Hempel N Petrick I Behrendt F Biomass productivity and productivity of fatty acids and amino acidsof microalgae strains as key characteristics of suitability for biodiesel production J Appl Phycol 2012 241407ndash1418 [CrossRef] [PubMed]
62 Yeesang C Cheirsilp B Effect of nitrogen salt and iron content in the growth medium and light intensityon lipid production by microalgae isolated from freshwater sources in Thailand Bioresour Technol 2011 1023034ndash3040 [CrossRef] [PubMed]
63 Msanne J Xu D Konda AR Casas-Mollano JA Awada T Cahoon EB Cerutti H Metabolic andgene expression changes triggered by nitrogen deprivation in the photoautotrophically grown microalgaeChlamydomonas reinhardtii and Coccomyxa sp C-169 Phytochemistry 2012 75 50ndash59 [CrossRef] [PubMed]
64 Lin Q Lin J Effects of nitrogen source and concentration on biomass and oil production of aScenedesmus rubescens like microalga Bioresour Technol 2011 102 1615ndash1621 [CrossRef] [PubMed]
65 Sheehan J Dunahay T Benemann J Roessler P A Look Back at the US Department of Energyrsquos AquaticSpecies ProgrammdashBiodiesel from Algae US Department of Energyrsquos Office of Fuels Development Golden COUSA 1998 p 328
66 Vaulot D Olson RJ Merkel S Chisholm SW Cell-cycle response to nutrient starvation in twophytoplankton species Thalassiosira weissflogii and Hymenomonas carterae Mar Biol 1987 95 625ndash630[CrossRef]
67 Olson RJ Chisholm SW Effects of light and nitrogen limitation on the cell cycle of the dinoflagellateAmphidinium carteri J Plankton Res 1986 8 785ndash793 [CrossRef]
68 Li Y Fei X Deng X Novel molecular insights into nitrogen starvation-induced triacylglycerolsaccumulation revealed by differential gene expression analysis in green algae Micractinium pusillumBiomass Bioenergy 2012 42 199ndash211 [CrossRef]
69 Abdelaziz AEM Leite GB Belhaj MA Hallenbeck PC Screening microalgae native to Quebec forwastewater treatment and biodiesel production Bioresour Technol 2014 157 140ndash148 [CrossRef] [PubMed]
70 Valdez-Ojeda R Gonzaacutelez-Muntildeoz M Us-Vaacutezquez R Narvaacuteez-Zapata J Chavarria-Hernandez JCLoacutepez-Adriaacuten S Barahona-Peacuterez F Toledano-Thompson T Garduntildeo-Soloacuterzano G Escobedo-GraciaMedrano RM Characterization of five fresh water microalgae with potential for biodiesel productionAlgal Res 2015 7 33ndash44 [CrossRef]
71 Shrivastav A Mishra SK Suh WI Farooq W Moon M Kim T-H Kumar K Choi G-G Park MSYang J-W Characterization of newly isolated oleaginous microalga Monoraphidium sp for lipid productionunder different conditions Algal Res 2015 12 289ndash294 [CrossRef]
72 Yen H-W Hu I-C Chen C-Y Ho S-H Lee D-J Chang J-S Microalgae-based biorefinerymdashFrombiofuels to natural products Bioresour Technol 2013 135 166ndash174 [CrossRef] [PubMed]
Energies 2017 10 224 16 of 16
73 Li J Liu Y Cheng JJ Mos M Daroch M Biological potential of microalgae in China for biorefinery-basedproduction of biofuels and high value compounds New Biotechnol 2015 32 588ndash596 [CrossRef] [PubMed]
74 El-Sheekh M Abomohra AE-F Hanelt D Optimization of biomass and fatty acid productivity ofScenedesmus obliquus as a promising microalga for biodiesel production World J Microbiol Biotechnol 201329 915ndash922 [CrossRef] [PubMed]
75 Singh P Guldhe A Kumari S Rawat I Bux F Investigation of combined effect of nitrogen phosphorusand iron on lipid productivity of microalgae Ankistrodesmus falcatus KJ671624 using response surfacemethodology Biochem Eng J 2015 94 22ndash29 [CrossRef]
76 Kilham S Kreeger D Goulden C Lynn S Effects of nutrient limitation on biochemical constituents ofAnkistrodesmus falcatus Freshw Biol 1997 38 591ndash596 [CrossRef]
77 Breuer G Lamers PP Martens DE Draaisma RB Wijffels RH The impact of nitrogen starvation on thedynamics of triacylglycerol accumulation in nine microalgae strains Bioresour Technol 2012 124 217ndash226[CrossRef] [PubMed]
78 Breuer G de Jaeger L Artus VG Martens DE Springer J Draaisma RB Eggink G Wijffels RHLamers PP Superior triacylglycerol (TAG) accumulation in starchless mutants of Scenedesmus obliquus (II)evaluation of TAG yield and productivity in controlled photobioreactors Biotechnol Biofuels 2014 7 70[CrossRef] [PubMed]
79 Nigam S Prakash-Rai M Sharma R Effect of Nitrogen on Growth and Lipid Content ofChlorella pyrenoidosa Am J Biochem Biotechnol 2011 7 126ndash131 [CrossRef]
80 Piligaev AV Sorokina KN Bryanskaya AV Peltek SE Kolchanov NA Parmon VN Isolation ofprospective microalgal strains with high saturated fatty acid content for biofuel production Algal Res 201512 368ndash376 [CrossRef]
81 Weiss SB Kennedy EP The enzymatic synthesis of triglycerides J Am Chem Soc 1956 78 3550[CrossRef]
82 Miller R Wu G Deshpande RR Vieler A Gaumlrtner K Li X Moellering ER Zaumluner S Cornish AJLiu B et al Changes in Transcript Abundance in Chlamydomonas reinhardtii following Nitrogen DeprivationPredict Diversion of Metabolism Plant Physiol 2010 154 1737ndash1752 [CrossRef] [PubMed]
83 Rismani-Yazdi H Haznedaroglu BZ Bibby K Peccia J Transcriptome sequencing and annotation of themicroalgae Dunaliella tertiolecta Pathway description and gene discovery for production of next-generationbiofuels BMC Genom 2011 12 148 [CrossRef] [PubMed]
84 Ramos MJ Fernaacutendez CM Casas A Rodriacuteguez L Peacuterez A Influence of fatty acid composition of rawmaterials on biodiesel properties Bioresour Technol 2009 100 261ndash268 [CrossRef] [PubMed]
85 Knothe G Dependence of biodiesel fuel properties on the structure of fatty acid alkyl estersFuel Process Technol 2005 86 1059ndash1070 [CrossRef]
86 Knothe G ldquoDesignerrdquo Biodiesel Optimizing Fatty Ester Composition to Improve Fuel PropertiesEnergy Fuels 2008 22 1358ndash1364 [CrossRef]
87 Smith PC Ngothai Y Dzuy Nguyen Q OrsquoNeill BK Improving the low-temperature properties ofbiodiesel Methods and consequences Renew Energy 2010 35 1145ndash1151 [CrossRef]
88 Dunn RO Effect of antioxidants on the oxidative stability of methyl soyate (biodiesel) Fuel Process Technol2005 86 1071ndash1085 [CrossRef]
89 Knothe G Improving biodiesel fuel properties by modifying fatty ester composition Energy Environ Sci2009 2 759ndash766 [CrossRef]
copy 2017 by the authors licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)

Energies 2017 10 224 14 of 16
31 Yu X Zhao P He C Li J Tang X Zhou J Huang Z Isolation of a novel strain of Monoraphidium spand characterization of its potential application as biodiesel feedstock Bioresour Technol 2012 121 256ndash262[CrossRef] [PubMed]
32 Hartree EF Determination of protein A modification of the lowry method that gives a linear photometricresponse Anal Biochem 1972 48 422ndash427 [CrossRef]
33 DuBois M Gilles KA Hamilton JK Rebers PA Smith F Colorimetric Method for Determination ofSugars and Related Substances Anal Chem 1956 28 350ndash356 [CrossRef]
34 AOAC International AOAC Official Methods of Analysis of the Association of Official Analytical Chemists (AOAC)15th ed AOAC International Washington DC USA 1990 Volume 1
35 Ichihara K Fukubayashi Y Preparation of fatty acid methyl esters for gas-liquid chromatographyJ Lipid Res 2010 51 635ndash640 [CrossRef] [PubMed]
36 Brennan L Owende P Biofuels from microalgaemdashA review of technologies for production processingand extractions of biofuels and co-products Renew Sustain Energ Rev 2010 14 557ndash577 [CrossRef]
37 Raja R Hemaiswarya S Kumar NA Sridhar S Rengasamy R A perspective on the biotechnologicalpotential of microalgae Crit Rev Microbiol 2008 34 77ndash88 [CrossRef] [PubMed]
38 Mata TM Martins AA Caetano NS Microalgae for biodiesel production and other applicationsA review Renew Sustain Energy Rev 2010 14 217ndash232 [CrossRef]
39 De Wilt A Butkovskyi A Tuantet K Leal LH Fernandes TV Langenhoff A Zeeman GMicropollutant removal in an algal treatment system fed with source separated wastewater streamsJ Hazard Mater 2015 304 84ndash92 [CrossRef] [PubMed]
40 Li T Lin G Podola B Melkonian M Continuous removal of zinc from wastewater and mine dumpleachate by a microalgal biofilm PSBR J Hazard Mater 2015 297 112ndash118 [CrossRef] [PubMed]
41 Sioli H The Amazon and its main affluents Hydrography morphology of the river courses and river typesIn Monographiae Biologicae Sioli H Ed Springer Dordrecht The Netherlands 1984 pp 127ndash165
42 Moniz MBJ Kaczmarska I Barcoding of Diatoms Nuclear Encoded ITS Revisited Protist 2010 161 7ndash34[CrossRef] [PubMed]
43 Coleman AW Is there a molecular key to the level of ldquobiological speciesrdquo in eukaryotes A DNA guideMol Phylogenet Evol 2009 50 197ndash203 [CrossRef] [PubMed]
44 Mai JC Coleman AW The Internal Transcribed Spacer 2 Exhibits a Common Secondary Structure inGreen Algae and Flowering Plants J Mol Evol 1997 44 258ndash271 [CrossRef] [PubMed]
45 Abou-Shanab RAI Matter IA Kim S-N Oh Y-K Choi J Jeon B-H Characterization andidentification of lipid-producing microalgae species isolated from a freshwater lake Biomass Bioenergy2011 35 3079ndash3085 [CrossRef]
46 Do Nascimento M Ortiz-Marquez JCF Sanchez-Rizza L Echarte MM Curatti L Bioprospectingfor fast growing and biomass characterization of oleaginous microalgae from South-Eastern Buenos AiresArgentina Bioresour Technol 2012 125 283ndash290 [CrossRef] [PubMed]
47 Araujo GS Matos LJBL Gonccedilalves LRB Fernandes FAN Farias WRL Bioprospecting foroil producing microalgal strains Evaluation of oil and biomass production for ten microalgal strainsBioresour Technol 2011 102 5248ndash5250 [CrossRef] [PubMed]
48 Zhou W Li Y Min M Hu B Chen P Ruan R Local bioprospecting for high-lipid producing microalgalstrains to be grown on concentrated municipal wastewater for biofuel production Bioresour Technol 2011102 6909ndash6919 [CrossRef] [PubMed]
49 Mahmoud EA Farahat LA Abdel Aziz ZK Fatthallah NA Salah El Din RA Evaluation of thepotential for some isolated microalgae to produce biodiesel Egypt J Pet 2015 24 97ndash101 [CrossRef]
50 Ho S Lai Y-Y Chiang C-Y Chen C-NN Chang J-S Selection of elite microalgae for biodieselproduction in tropical conditions using a standardized platform Bioresour Technol 2013 147 135ndash142[CrossRef] [PubMed]
51 Li L Cui J Liu Q Ding Y Liu J Screening and phylogenetic analysis of lipid-rich microalgae Algal Res2015 11 381ndash386 [CrossRef]
52 Rodolfi L Chini Zittelli G Bassi N Padovani G Biondi N Bonini G Tredici MR Microalgae for oilStrain selection induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactorBiotechnol Bioeng 2009 102 100ndash112 [CrossRef] [PubMed]
Energies 2017 10 224 15 of 16
53 Li P Miao X Li R Zhong J In situ biodiesel production from fast-growing and high oil content Chlorellapyrenoidosa in rice straw hydrolysate J Biomed Biotechnol 2011 2011 141207 [CrossRef] [PubMed]
54 Doan TTY Sivaloganathan B Obbard JP Screening of marine microalgae for biodiesel feedstockBiomass Bioenergy 2011 35 2534ndash2544 [CrossRef]
55 Abou-Shanab RAI Hwang J-H Cho Y Min B Jeon B-H Characterization of microalgal speciesisolated from fresh water bodies as a potential source for biodiesel production Appl Energy 2011 883300ndash3306 [CrossRef]
56 Chaichalerm S Pokethitiyook P Yuan W Meetam M Sritong K Pugkaew W Kungvansaichol KKruatrachue M Damrongphol P Culture of microalgal strains isolated from natural habitats in Thailand invarious enriched media Appl Energy 2012 89 296ndash302 [CrossRef]
57 Zhang S Liu P Yang X Hao Z Zhang L Luo N Shi J Isolation and identification by 18S rDNAsequence of high lipid potential microalgal species for fuel production in Hainan Dao Biomass Bioenergy2014 66 197ndash203 [CrossRef]
58 Chiu S-Y Kao C-Y Tsai M-T Ong S-C Chen C-H Lin C-S Lipid accumulation and CO2 utilizationof Nannochloropsis oculata in response to CO2 aeration Bioresour Technol 2009 100 833ndash838 [CrossRef][PubMed]
59 Huerlimann R de Nys R Heimann K Growth lipid content productivity and fatty acid composition oftropical microalgae for scale-up production Biotechnol Bioeng 2010 107 245ndash257 [CrossRef] [PubMed]
60 Liang Y Sarkany N Cui Y Biomass and lipid productivities of Chlorella vulgaris under autotrophicheterotrophic and mixotrophic growth conditions Biotechnol Lett 2009 31 1043ndash1049 [CrossRef] [PubMed]
61 Hempel N Petrick I Behrendt F Biomass productivity and productivity of fatty acids and amino acidsof microalgae strains as key characteristics of suitability for biodiesel production J Appl Phycol 2012 241407ndash1418 [CrossRef] [PubMed]
62 Yeesang C Cheirsilp B Effect of nitrogen salt and iron content in the growth medium and light intensityon lipid production by microalgae isolated from freshwater sources in Thailand Bioresour Technol 2011 1023034ndash3040 [CrossRef] [PubMed]
63 Msanne J Xu D Konda AR Casas-Mollano JA Awada T Cahoon EB Cerutti H Metabolic andgene expression changes triggered by nitrogen deprivation in the photoautotrophically grown microalgaeChlamydomonas reinhardtii and Coccomyxa sp C-169 Phytochemistry 2012 75 50ndash59 [CrossRef] [PubMed]
64 Lin Q Lin J Effects of nitrogen source and concentration on biomass and oil production of aScenedesmus rubescens like microalga Bioresour Technol 2011 102 1615ndash1621 [CrossRef] [PubMed]
65 Sheehan J Dunahay T Benemann J Roessler P A Look Back at the US Department of Energyrsquos AquaticSpecies ProgrammdashBiodiesel from Algae US Department of Energyrsquos Office of Fuels Development Golden COUSA 1998 p 328
66 Vaulot D Olson RJ Merkel S Chisholm SW Cell-cycle response to nutrient starvation in twophytoplankton species Thalassiosira weissflogii and Hymenomonas carterae Mar Biol 1987 95 625ndash630[CrossRef]
67 Olson RJ Chisholm SW Effects of light and nitrogen limitation on the cell cycle of the dinoflagellateAmphidinium carteri J Plankton Res 1986 8 785ndash793 [CrossRef]
68 Li Y Fei X Deng X Novel molecular insights into nitrogen starvation-induced triacylglycerolsaccumulation revealed by differential gene expression analysis in green algae Micractinium pusillumBiomass Bioenergy 2012 42 199ndash211 [CrossRef]
69 Abdelaziz AEM Leite GB Belhaj MA Hallenbeck PC Screening microalgae native to Quebec forwastewater treatment and biodiesel production Bioresour Technol 2014 157 140ndash148 [CrossRef] [PubMed]
70 Valdez-Ojeda R Gonzaacutelez-Muntildeoz M Us-Vaacutezquez R Narvaacuteez-Zapata J Chavarria-Hernandez JCLoacutepez-Adriaacuten S Barahona-Peacuterez F Toledano-Thompson T Garduntildeo-Soloacuterzano G Escobedo-GraciaMedrano RM Characterization of five fresh water microalgae with potential for biodiesel productionAlgal Res 2015 7 33ndash44 [CrossRef]
71 Shrivastav A Mishra SK Suh WI Farooq W Moon M Kim T-H Kumar K Choi G-G Park MSYang J-W Characterization of newly isolated oleaginous microalga Monoraphidium sp for lipid productionunder different conditions Algal Res 2015 12 289ndash294 [CrossRef]
72 Yen H-W Hu I-C Chen C-Y Ho S-H Lee D-J Chang J-S Microalgae-based biorefinerymdashFrombiofuels to natural products Bioresour Technol 2013 135 166ndash174 [CrossRef] [PubMed]
Energies 2017 10 224 16 of 16
73 Li J Liu Y Cheng JJ Mos M Daroch M Biological potential of microalgae in China for biorefinery-basedproduction of biofuels and high value compounds New Biotechnol 2015 32 588ndash596 [CrossRef] [PubMed]
74 El-Sheekh M Abomohra AE-F Hanelt D Optimization of biomass and fatty acid productivity ofScenedesmus obliquus as a promising microalga for biodiesel production World J Microbiol Biotechnol 201329 915ndash922 [CrossRef] [PubMed]
75 Singh P Guldhe A Kumari S Rawat I Bux F Investigation of combined effect of nitrogen phosphorusand iron on lipid productivity of microalgae Ankistrodesmus falcatus KJ671624 using response surfacemethodology Biochem Eng J 2015 94 22ndash29 [CrossRef]
76 Kilham S Kreeger D Goulden C Lynn S Effects of nutrient limitation on biochemical constituents ofAnkistrodesmus falcatus Freshw Biol 1997 38 591ndash596 [CrossRef]
77 Breuer G Lamers PP Martens DE Draaisma RB Wijffels RH The impact of nitrogen starvation on thedynamics of triacylglycerol accumulation in nine microalgae strains Bioresour Technol 2012 124 217ndash226[CrossRef] [PubMed]
78 Breuer G de Jaeger L Artus VG Martens DE Springer J Draaisma RB Eggink G Wijffels RHLamers PP Superior triacylglycerol (TAG) accumulation in starchless mutants of Scenedesmus obliquus (II)evaluation of TAG yield and productivity in controlled photobioreactors Biotechnol Biofuels 2014 7 70[CrossRef] [PubMed]
79 Nigam S Prakash-Rai M Sharma R Effect of Nitrogen on Growth and Lipid Content ofChlorella pyrenoidosa Am J Biochem Biotechnol 2011 7 126ndash131 [CrossRef]
80 Piligaev AV Sorokina KN Bryanskaya AV Peltek SE Kolchanov NA Parmon VN Isolation ofprospective microalgal strains with high saturated fatty acid content for biofuel production Algal Res 201512 368ndash376 [CrossRef]
81 Weiss SB Kennedy EP The enzymatic synthesis of triglycerides J Am Chem Soc 1956 78 3550[CrossRef]
82 Miller R Wu G Deshpande RR Vieler A Gaumlrtner K Li X Moellering ER Zaumluner S Cornish AJLiu B et al Changes in Transcript Abundance in Chlamydomonas reinhardtii following Nitrogen DeprivationPredict Diversion of Metabolism Plant Physiol 2010 154 1737ndash1752 [CrossRef] [PubMed]
83 Rismani-Yazdi H Haznedaroglu BZ Bibby K Peccia J Transcriptome sequencing and annotation of themicroalgae Dunaliella tertiolecta Pathway description and gene discovery for production of next-generationbiofuels BMC Genom 2011 12 148 [CrossRef] [PubMed]
84 Ramos MJ Fernaacutendez CM Casas A Rodriacuteguez L Peacuterez A Influence of fatty acid composition of rawmaterials on biodiesel properties Bioresour Technol 2009 100 261ndash268 [CrossRef] [PubMed]
85 Knothe G Dependence of biodiesel fuel properties on the structure of fatty acid alkyl estersFuel Process Technol 2005 86 1059ndash1070 [CrossRef]
86 Knothe G ldquoDesignerrdquo Biodiesel Optimizing Fatty Ester Composition to Improve Fuel PropertiesEnergy Fuels 2008 22 1358ndash1364 [CrossRef]
87 Smith PC Ngothai Y Dzuy Nguyen Q OrsquoNeill BK Improving the low-temperature properties ofbiodiesel Methods and consequences Renew Energy 2010 35 1145ndash1151 [CrossRef]
88 Dunn RO Effect of antioxidants on the oxidative stability of methyl soyate (biodiesel) Fuel Process Technol2005 86 1071ndash1085 [CrossRef]
89 Knothe G Improving biodiesel fuel properties by modifying fatty ester composition Energy Environ Sci2009 2 759ndash766 [CrossRef]
copy 2017 by the authors licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)

Energies 2017 10 224 15 of 16
53 Li P Miao X Li R Zhong J In situ biodiesel production from fast-growing and high oil content Chlorellapyrenoidosa in rice straw hydrolysate J Biomed Biotechnol 2011 2011 141207 [CrossRef] [PubMed]
54 Doan TTY Sivaloganathan B Obbard JP Screening of marine microalgae for biodiesel feedstockBiomass Bioenergy 2011 35 2534ndash2544 [CrossRef]
55 Abou-Shanab RAI Hwang J-H Cho Y Min B Jeon B-H Characterization of microalgal speciesisolated from fresh water bodies as a potential source for biodiesel production Appl Energy 2011 883300ndash3306 [CrossRef]
56 Chaichalerm S Pokethitiyook P Yuan W Meetam M Sritong K Pugkaew W Kungvansaichol KKruatrachue M Damrongphol P Culture of microalgal strains isolated from natural habitats in Thailand invarious enriched media Appl Energy 2012 89 296ndash302 [CrossRef]
57 Zhang S Liu P Yang X Hao Z Zhang L Luo N Shi J Isolation and identification by 18S rDNAsequence of high lipid potential microalgal species for fuel production in Hainan Dao Biomass Bioenergy2014 66 197ndash203 [CrossRef]
58 Chiu S-Y Kao C-Y Tsai M-T Ong S-C Chen C-H Lin C-S Lipid accumulation and CO2 utilizationof Nannochloropsis oculata in response to CO2 aeration Bioresour Technol 2009 100 833ndash838 [CrossRef][PubMed]
59 Huerlimann R de Nys R Heimann K Growth lipid content productivity and fatty acid composition oftropical microalgae for scale-up production Biotechnol Bioeng 2010 107 245ndash257 [CrossRef] [PubMed]
60 Liang Y Sarkany N Cui Y Biomass and lipid productivities of Chlorella vulgaris under autotrophicheterotrophic and mixotrophic growth conditions Biotechnol Lett 2009 31 1043ndash1049 [CrossRef] [PubMed]
61 Hempel N Petrick I Behrendt F Biomass productivity and productivity of fatty acids and amino acidsof microalgae strains as key characteristics of suitability for biodiesel production J Appl Phycol 2012 241407ndash1418 [CrossRef] [PubMed]
62 Yeesang C Cheirsilp B Effect of nitrogen salt and iron content in the growth medium and light intensityon lipid production by microalgae isolated from freshwater sources in Thailand Bioresour Technol 2011 1023034ndash3040 [CrossRef] [PubMed]
63 Msanne J Xu D Konda AR Casas-Mollano JA Awada T Cahoon EB Cerutti H Metabolic andgene expression changes triggered by nitrogen deprivation in the photoautotrophically grown microalgaeChlamydomonas reinhardtii and Coccomyxa sp C-169 Phytochemistry 2012 75 50ndash59 [CrossRef] [PubMed]
64 Lin Q Lin J Effects of nitrogen source and concentration on biomass and oil production of aScenedesmus rubescens like microalga Bioresour Technol 2011 102 1615ndash1621 [CrossRef] [PubMed]
65 Sheehan J Dunahay T Benemann J Roessler P A Look Back at the US Department of Energyrsquos AquaticSpecies ProgrammdashBiodiesel from Algae US Department of Energyrsquos Office of Fuels Development Golden COUSA 1998 p 328
66 Vaulot D Olson RJ Merkel S Chisholm SW Cell-cycle response to nutrient starvation in twophytoplankton species Thalassiosira weissflogii and Hymenomonas carterae Mar Biol 1987 95 625ndash630[CrossRef]
67 Olson RJ Chisholm SW Effects of light and nitrogen limitation on the cell cycle of the dinoflagellateAmphidinium carteri J Plankton Res 1986 8 785ndash793 [CrossRef]
68 Li Y Fei X Deng X Novel molecular insights into nitrogen starvation-induced triacylglycerolsaccumulation revealed by differential gene expression analysis in green algae Micractinium pusillumBiomass Bioenergy 2012 42 199ndash211 [CrossRef]
69 Abdelaziz AEM Leite GB Belhaj MA Hallenbeck PC Screening microalgae native to Quebec forwastewater treatment and biodiesel production Bioresour Technol 2014 157 140ndash148 [CrossRef] [PubMed]
70 Valdez-Ojeda R Gonzaacutelez-Muntildeoz M Us-Vaacutezquez R Narvaacuteez-Zapata J Chavarria-Hernandez JCLoacutepez-Adriaacuten S Barahona-Peacuterez F Toledano-Thompson T Garduntildeo-Soloacuterzano G Escobedo-GraciaMedrano RM Characterization of five fresh water microalgae with potential for biodiesel productionAlgal Res 2015 7 33ndash44 [CrossRef]
71 Shrivastav A Mishra SK Suh WI Farooq W Moon M Kim T-H Kumar K Choi G-G Park MSYang J-W Characterization of newly isolated oleaginous microalga Monoraphidium sp for lipid productionunder different conditions Algal Res 2015 12 289ndash294 [CrossRef]
72 Yen H-W Hu I-C Chen C-Y Ho S-H Lee D-J Chang J-S Microalgae-based biorefinerymdashFrombiofuels to natural products Bioresour Technol 2013 135 166ndash174 [CrossRef] [PubMed]
Energies 2017 10 224 16 of 16
73 Li J Liu Y Cheng JJ Mos M Daroch M Biological potential of microalgae in China for biorefinery-basedproduction of biofuels and high value compounds New Biotechnol 2015 32 588ndash596 [CrossRef] [PubMed]
74 El-Sheekh M Abomohra AE-F Hanelt D Optimization of biomass and fatty acid productivity ofScenedesmus obliquus as a promising microalga for biodiesel production World J Microbiol Biotechnol 201329 915ndash922 [CrossRef] [PubMed]
75 Singh P Guldhe A Kumari S Rawat I Bux F Investigation of combined effect of nitrogen phosphorusand iron on lipid productivity of microalgae Ankistrodesmus falcatus KJ671624 using response surfacemethodology Biochem Eng J 2015 94 22ndash29 [CrossRef]
76 Kilham S Kreeger D Goulden C Lynn S Effects of nutrient limitation on biochemical constituents ofAnkistrodesmus falcatus Freshw Biol 1997 38 591ndash596 [CrossRef]
77 Breuer G Lamers PP Martens DE Draaisma RB Wijffels RH The impact of nitrogen starvation on thedynamics of triacylglycerol accumulation in nine microalgae strains Bioresour Technol 2012 124 217ndash226[CrossRef] [PubMed]
78 Breuer G de Jaeger L Artus VG Martens DE Springer J Draaisma RB Eggink G Wijffels RHLamers PP Superior triacylglycerol (TAG) accumulation in starchless mutants of Scenedesmus obliquus (II)evaluation of TAG yield and productivity in controlled photobioreactors Biotechnol Biofuels 2014 7 70[CrossRef] [PubMed]
79 Nigam S Prakash-Rai M Sharma R Effect of Nitrogen on Growth and Lipid Content ofChlorella pyrenoidosa Am J Biochem Biotechnol 2011 7 126ndash131 [CrossRef]
80 Piligaev AV Sorokina KN Bryanskaya AV Peltek SE Kolchanov NA Parmon VN Isolation ofprospective microalgal strains with high saturated fatty acid content for biofuel production Algal Res 201512 368ndash376 [CrossRef]
81 Weiss SB Kennedy EP The enzymatic synthesis of triglycerides J Am Chem Soc 1956 78 3550[CrossRef]
82 Miller R Wu G Deshpande RR Vieler A Gaumlrtner K Li X Moellering ER Zaumluner S Cornish AJLiu B et al Changes in Transcript Abundance in Chlamydomonas reinhardtii following Nitrogen DeprivationPredict Diversion of Metabolism Plant Physiol 2010 154 1737ndash1752 [CrossRef] [PubMed]
83 Rismani-Yazdi H Haznedaroglu BZ Bibby K Peccia J Transcriptome sequencing and annotation of themicroalgae Dunaliella tertiolecta Pathway description and gene discovery for production of next-generationbiofuels BMC Genom 2011 12 148 [CrossRef] [PubMed]
84 Ramos MJ Fernaacutendez CM Casas A Rodriacuteguez L Peacuterez A Influence of fatty acid composition of rawmaterials on biodiesel properties Bioresour Technol 2009 100 261ndash268 [CrossRef] [PubMed]
85 Knothe G Dependence of biodiesel fuel properties on the structure of fatty acid alkyl estersFuel Process Technol 2005 86 1059ndash1070 [CrossRef]
86 Knothe G ldquoDesignerrdquo Biodiesel Optimizing Fatty Ester Composition to Improve Fuel PropertiesEnergy Fuels 2008 22 1358ndash1364 [CrossRef]
87 Smith PC Ngothai Y Dzuy Nguyen Q OrsquoNeill BK Improving the low-temperature properties ofbiodiesel Methods and consequences Renew Energy 2010 35 1145ndash1151 [CrossRef]
88 Dunn RO Effect of antioxidants on the oxidative stability of methyl soyate (biodiesel) Fuel Process Technol2005 86 1071ndash1085 [CrossRef]
89 Knothe G Improving biodiesel fuel properties by modifying fatty ester composition Energy Environ Sci2009 2 759ndash766 [CrossRef]
copy 2017 by the authors licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)

Energies 2017 10 224 16 of 16
73 Li J Liu Y Cheng JJ Mos M Daroch M Biological potential of microalgae in China for biorefinery-basedproduction of biofuels and high value compounds New Biotechnol 2015 32 588ndash596 [CrossRef] [PubMed]
74 El-Sheekh M Abomohra AE-F Hanelt D Optimization of biomass and fatty acid productivity ofScenedesmus obliquus as a promising microalga for biodiesel production World J Microbiol Biotechnol 201329 915ndash922 [CrossRef] [PubMed]
75 Singh P Guldhe A Kumari S Rawat I Bux F Investigation of combined effect of nitrogen phosphorusand iron on lipid productivity of microalgae Ankistrodesmus falcatus KJ671624 using response surfacemethodology Biochem Eng J 2015 94 22ndash29 [CrossRef]
76 Kilham S Kreeger D Goulden C Lynn S Effects of nutrient limitation on biochemical constituents ofAnkistrodesmus falcatus Freshw Biol 1997 38 591ndash596 [CrossRef]
77 Breuer G Lamers PP Martens DE Draaisma RB Wijffels RH The impact of nitrogen starvation on thedynamics of triacylglycerol accumulation in nine microalgae strains Bioresour Technol 2012 124 217ndash226[CrossRef] [PubMed]
78 Breuer G de Jaeger L Artus VG Martens DE Springer J Draaisma RB Eggink G Wijffels RHLamers PP Superior triacylglycerol (TAG) accumulation in starchless mutants of Scenedesmus obliquus (II)evaluation of TAG yield and productivity in controlled photobioreactors Biotechnol Biofuels 2014 7 70[CrossRef] [PubMed]
79 Nigam S Prakash-Rai M Sharma R Effect of Nitrogen on Growth and Lipid Content ofChlorella pyrenoidosa Am J Biochem Biotechnol 2011 7 126ndash131 [CrossRef]
80 Piligaev AV Sorokina KN Bryanskaya AV Peltek SE Kolchanov NA Parmon VN Isolation ofprospective microalgal strains with high saturated fatty acid content for biofuel production Algal Res 201512 368ndash376 [CrossRef]
81 Weiss SB Kennedy EP The enzymatic synthesis of triglycerides J Am Chem Soc 1956 78 3550[CrossRef]
82 Miller R Wu G Deshpande RR Vieler A Gaumlrtner K Li X Moellering ER Zaumluner S Cornish AJLiu B et al Changes in Transcript Abundance in Chlamydomonas reinhardtii following Nitrogen DeprivationPredict Diversion of Metabolism Plant Physiol 2010 154 1737ndash1752 [CrossRef] [PubMed]
83 Rismani-Yazdi H Haznedaroglu BZ Bibby K Peccia J Transcriptome sequencing and annotation of themicroalgae Dunaliella tertiolecta Pathway description and gene discovery for production of next-generationbiofuels BMC Genom 2011 12 148 [CrossRef] [PubMed]
84 Ramos MJ Fernaacutendez CM Casas A Rodriacuteguez L Peacuterez A Influence of fatty acid composition of rawmaterials on biodiesel properties Bioresour Technol 2009 100 261ndash268 [CrossRef] [PubMed]
85 Knothe G Dependence of biodiesel fuel properties on the structure of fatty acid alkyl estersFuel Process Technol 2005 86 1059ndash1070 [CrossRef]
86 Knothe G ldquoDesignerrdquo Biodiesel Optimizing Fatty Ester Composition to Improve Fuel PropertiesEnergy Fuels 2008 22 1358ndash1364 [CrossRef]
87 Smith PC Ngothai Y Dzuy Nguyen Q OrsquoNeill BK Improving the low-temperature properties ofbiodiesel Methods and consequences Renew Energy 2010 35 1145ndash1151 [CrossRef]
88 Dunn RO Effect of antioxidants on the oxidative stability of methyl soyate (biodiesel) Fuel Process Technol2005 86 1071ndash1085 [CrossRef]
89 Knothe G Improving biodiesel fuel properties by modifying fatty ester composition Energy Environ Sci2009 2 759ndash766 [CrossRef]
copy 2017 by the authors licensee MDPI Basel Switzerland This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (httpcreativecommonsorglicensesby40)











![Gold Nanoparticles with Organic Linkers for …...Synthetic strategies to produce different shapes of Au NPs include use of surfactants (cetyltrimethylammonium bromide [CTAB] and cet-yltrimethylammonium](https://img.pdfslide.us/doc/110x75/5fa97cad1ea82b25516b28a4/gold-nanoparticles-with-organic-linkers-for-synthetic-strategies-to-produce.jpg)

















