Embed Size (px)
Citation preview

Is ubiquitination always the result of mistakes?

The N-end Rule: The N-terminal amino acid determines half-life
Destabilizing N-termini are recognized by a
special E2/E3
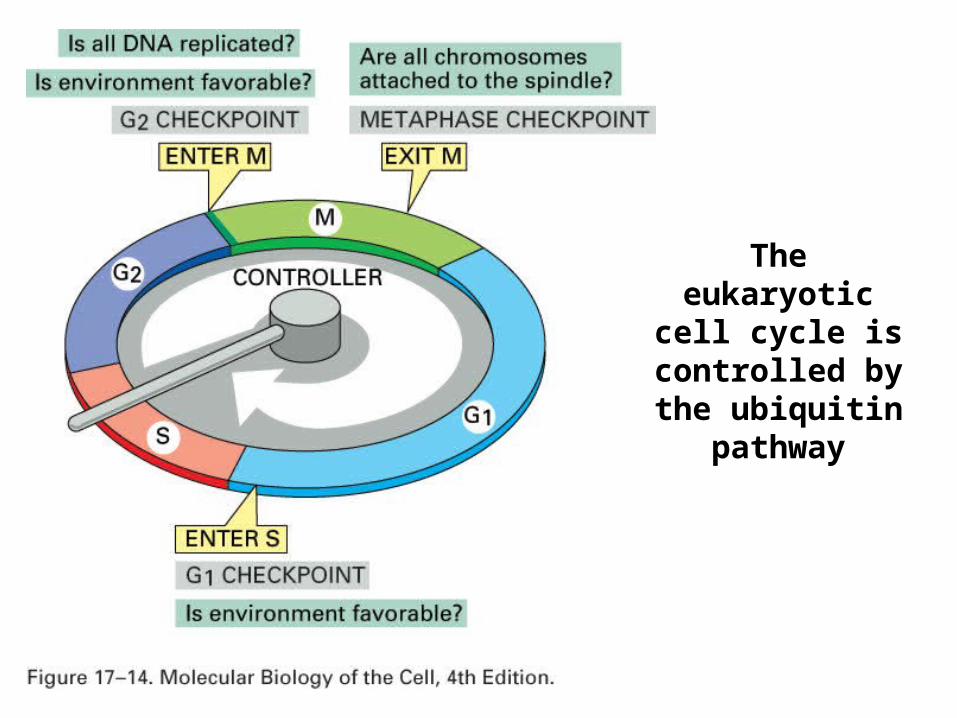
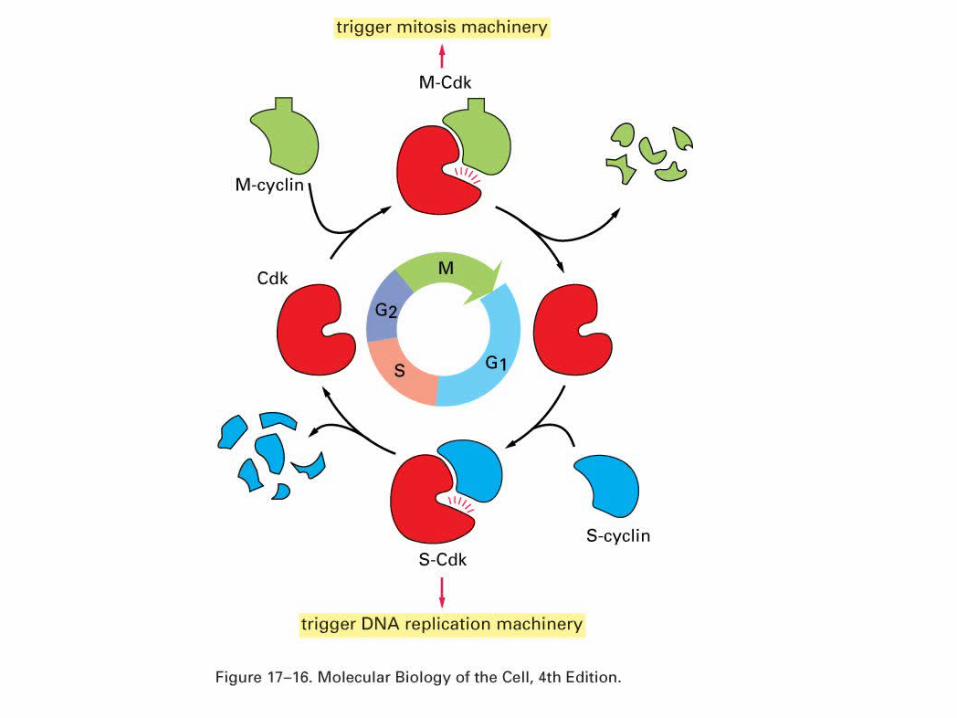
The eukaryotic cell cycle is
controlled by the ubiquitin pathway
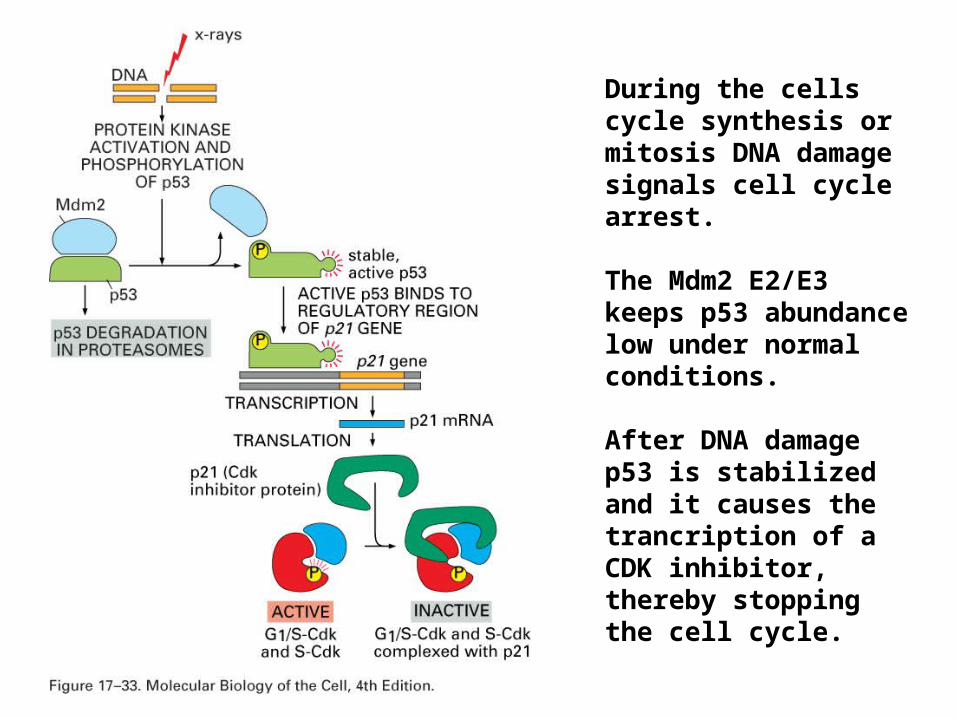
During the cells cycle synthesis or mitosis DNA damage signals cell cycle arrest.
The Mdm2 E2/E3 keeps p53 abundance low under normal conditions.
After DNA damage p53 is stabilized and it causes the trancription of a CDK inhibitor, thereby stopping the cell cycle.
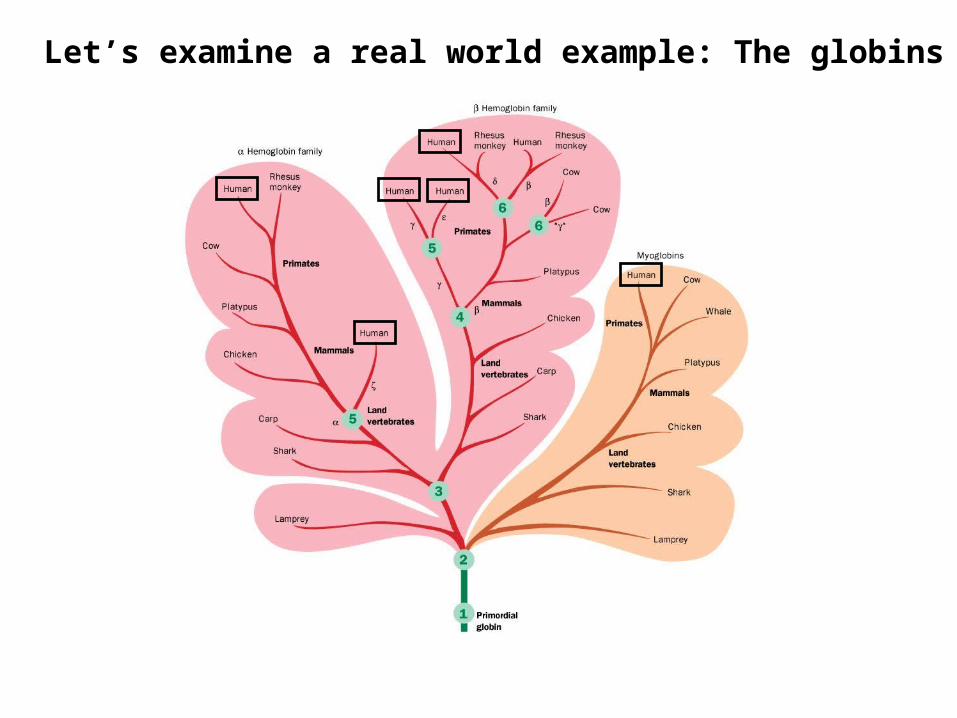
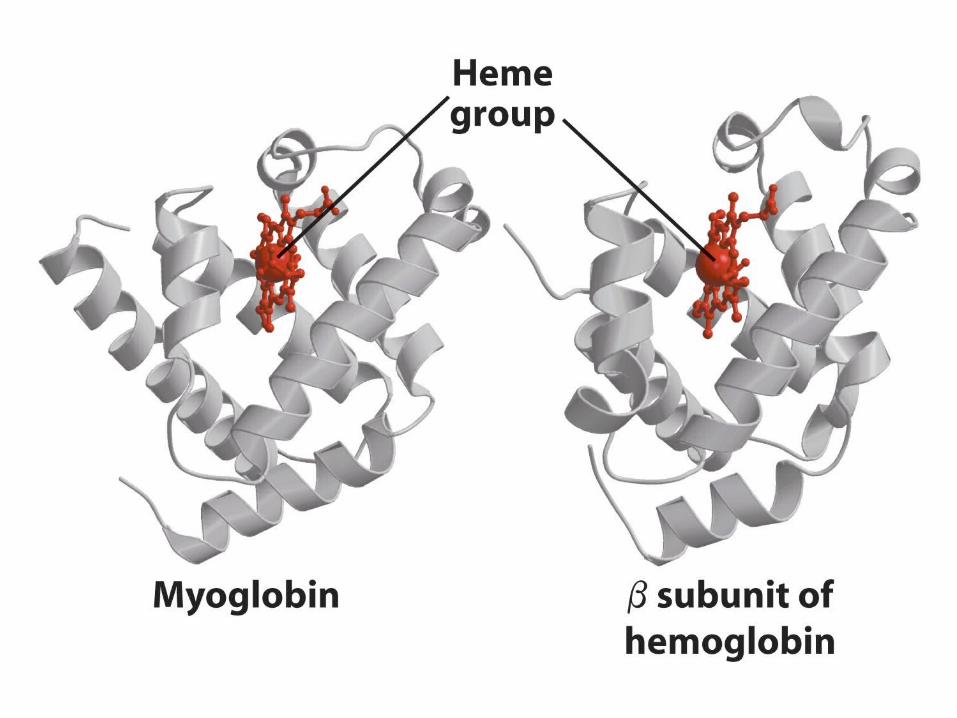
Let’s examine a real world example: The globins
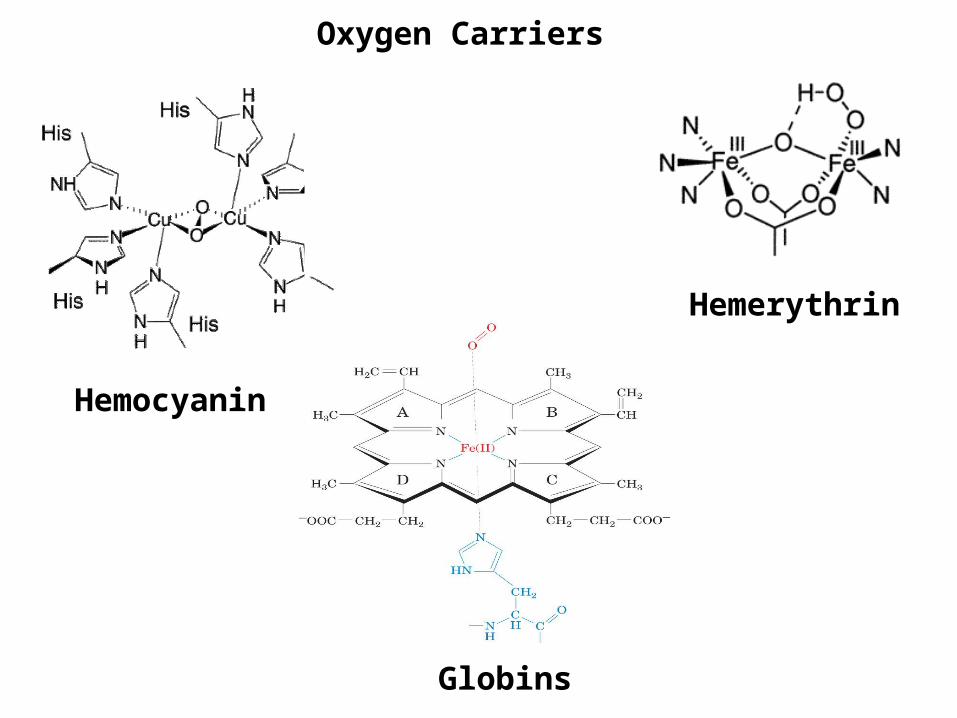
Oxygen Carriers
Hemerythrin
Hemocyanin
Globins

Need metals to bind to oxygen…..why?
Oxygen is a diradical
It has 2 unpaired electrons
1/23O2 + 1X ---> 1XO

The spin restriction limits the chemical reactivity by imposing a
kinetic barrier
This is the oxygen paradox
Singlet oxygen in the excited state is extraordinarily reactive
This is the basis for photodynamic therapy

Fe(II)-O2 Fe(III)-O2-
Fe(III)-O2- + Fe(II) Fe(III)-O2
2--Fe(III)
Fe(III)-O22--Fe(III)
2Fe(IV)=O Fe(III)-O-Fe(III)
2Fe(IV)=O
Metals cause oxygen to become reactive because they are radicals themselves. They
eliminate spin restrictions
Highly reactive!
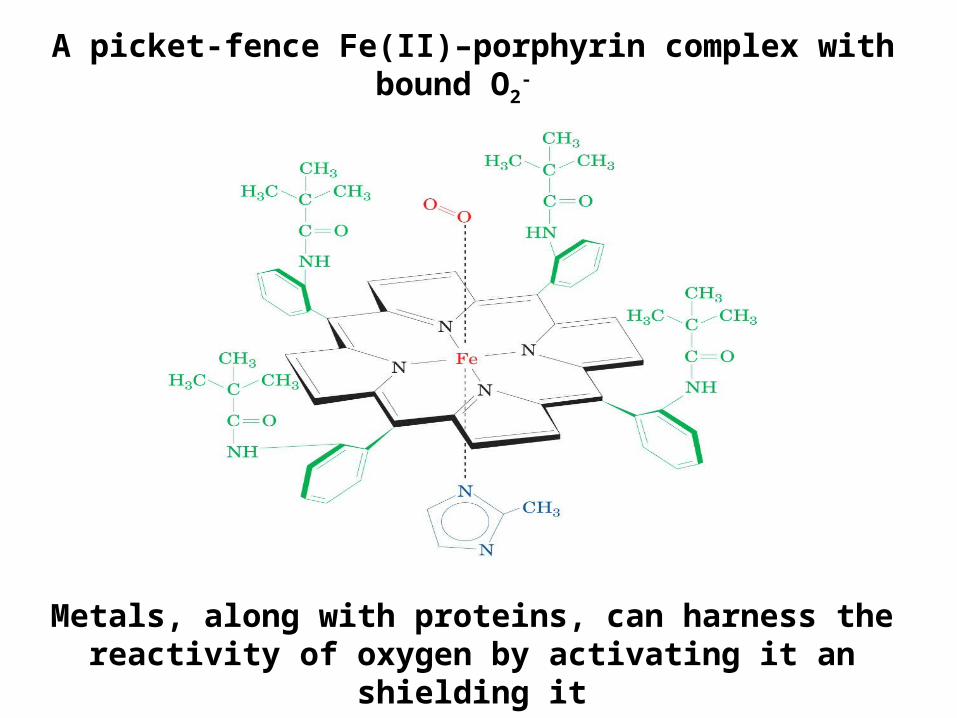
A picket-fence Fe(II)–porphyrin complex with bound O2-
Metals, along with proteins, can harness the reactivity of oxygen by activating it an shielding it

Fe(II) binds dioxygen Fe(III) does notWhy?
Oxygen to metal charge transfer
Fe(II)-O2 Fe(III)-O2-
Fe(II) will also bind NO, CO, S2- , CN-
Fe(III)-O2 Fe(IV)-O2-
Stable
Unstable
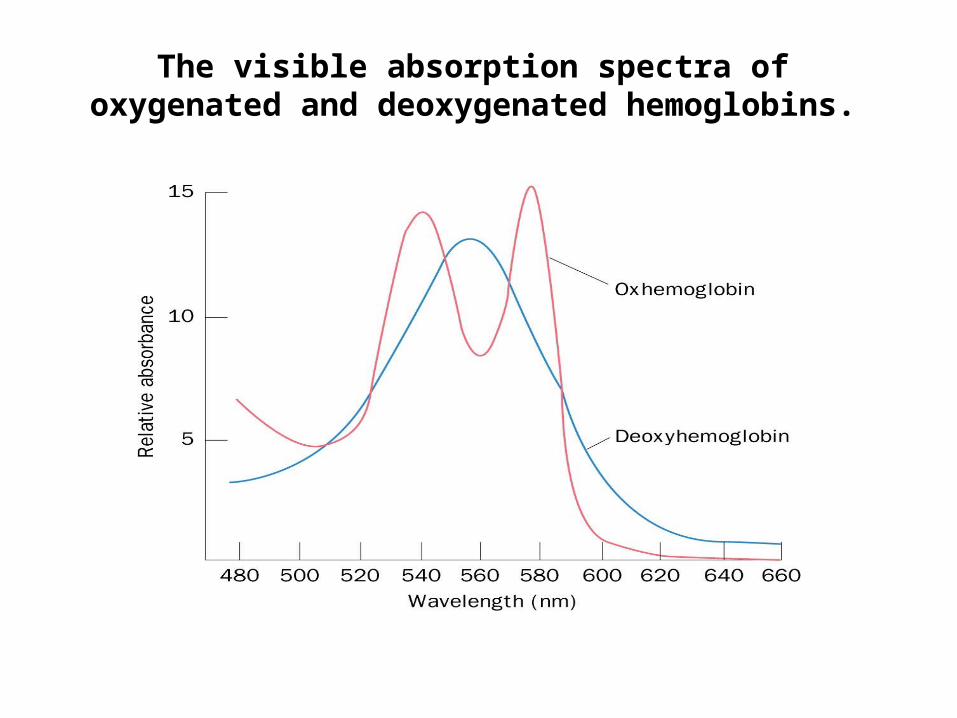
The visible absorption spectra of oxygenated and deoxygenated hemoglobins.
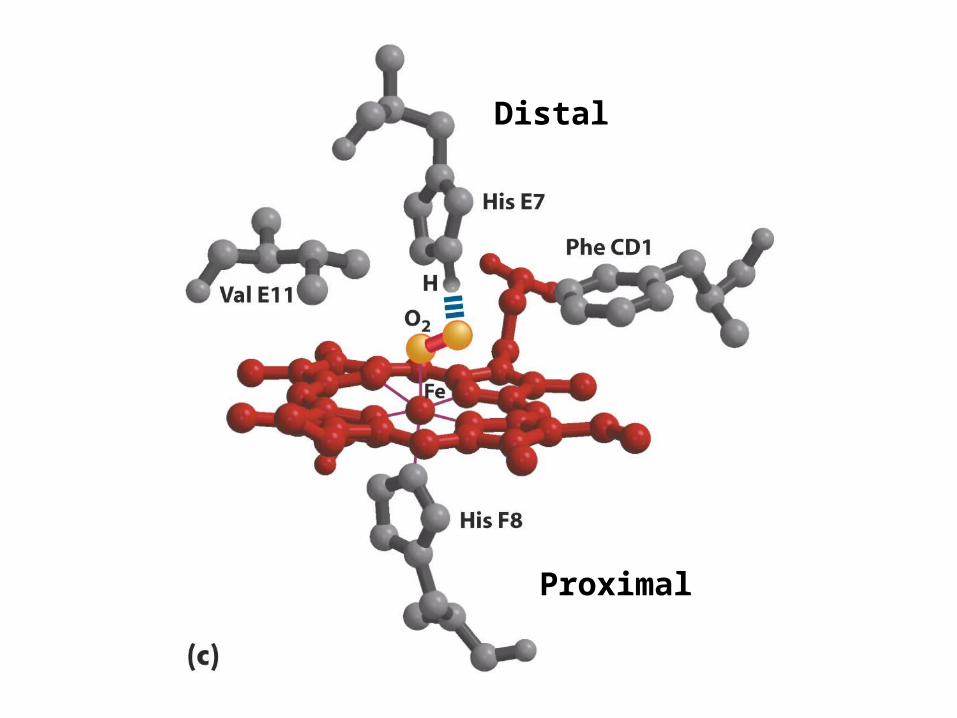
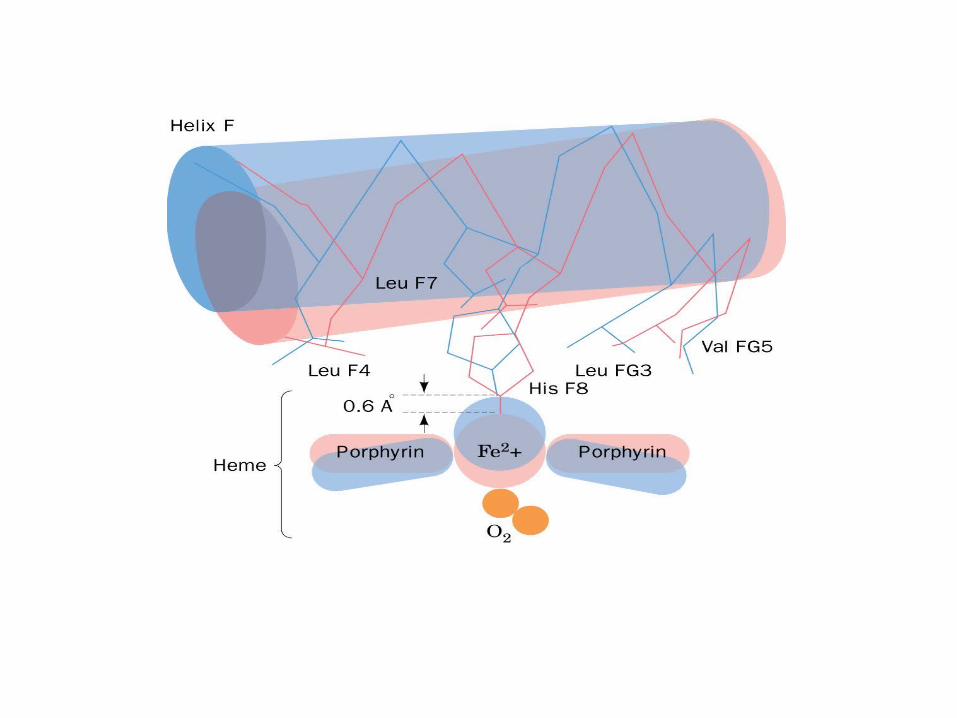
Distal
Proximal
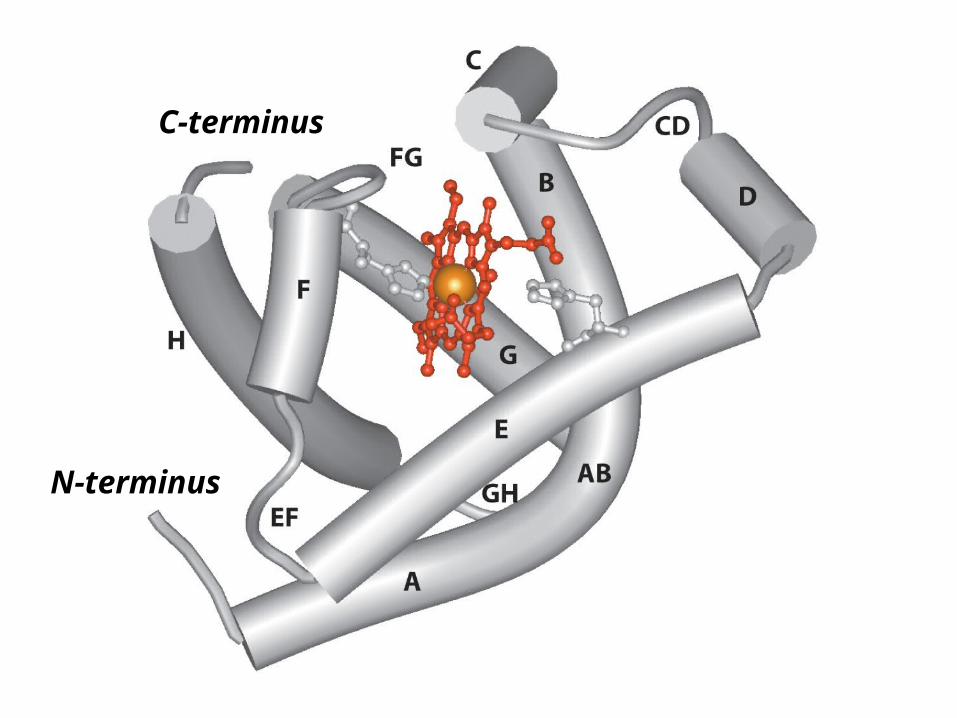
N-terminus
C-terminus
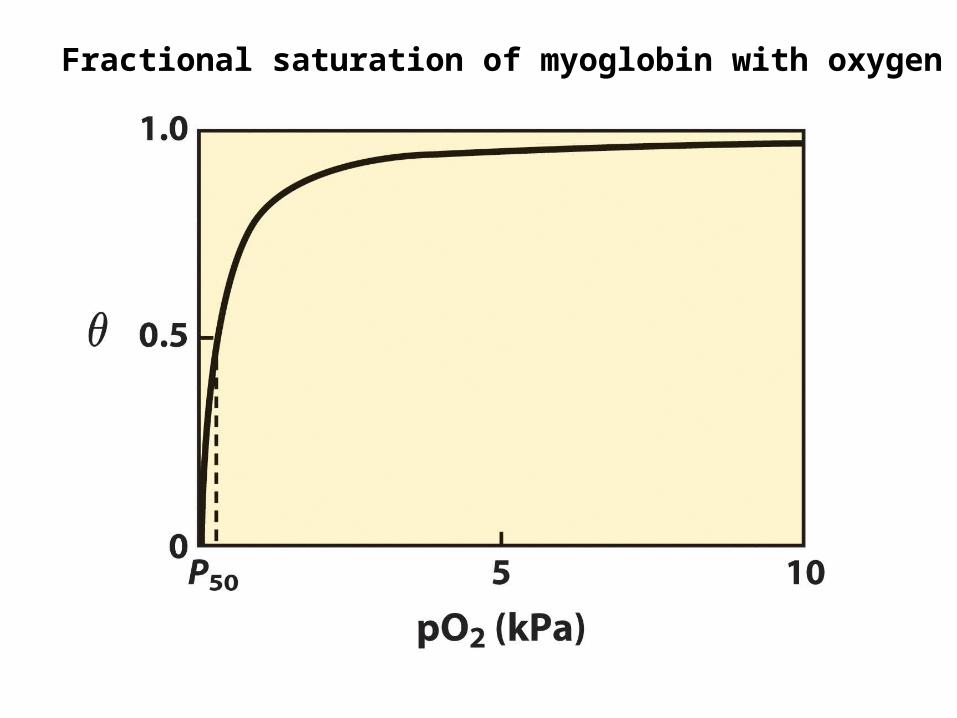
Fractional saturation of myoglobin with oxygen
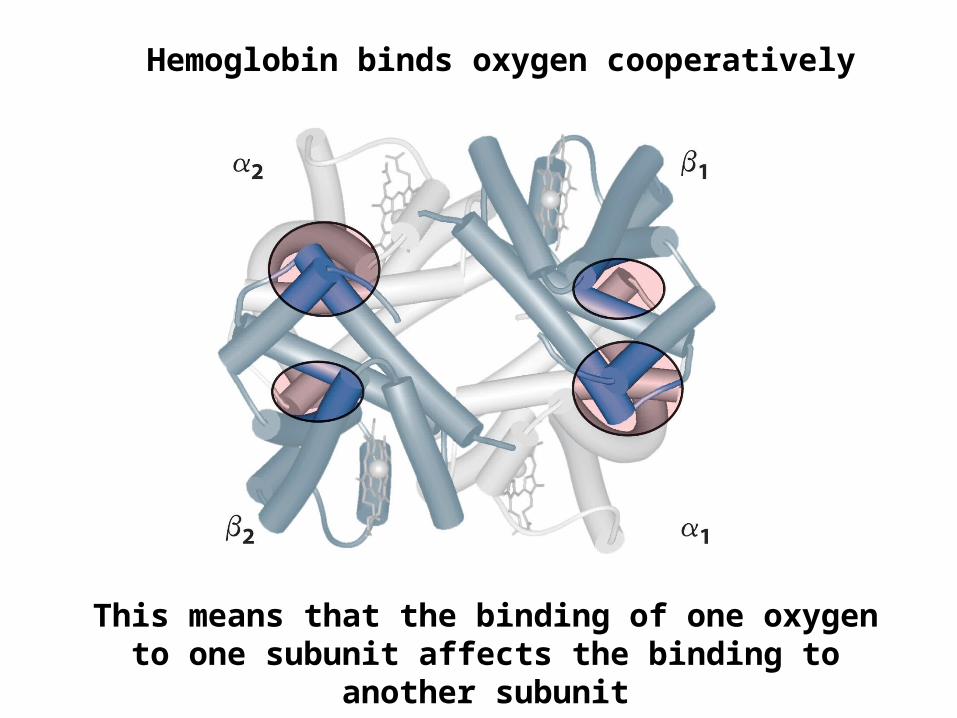
Hemoglobin binds oxygen cooperatively
This means that the binding of one oxygen to one subunit affects the binding to another subunit
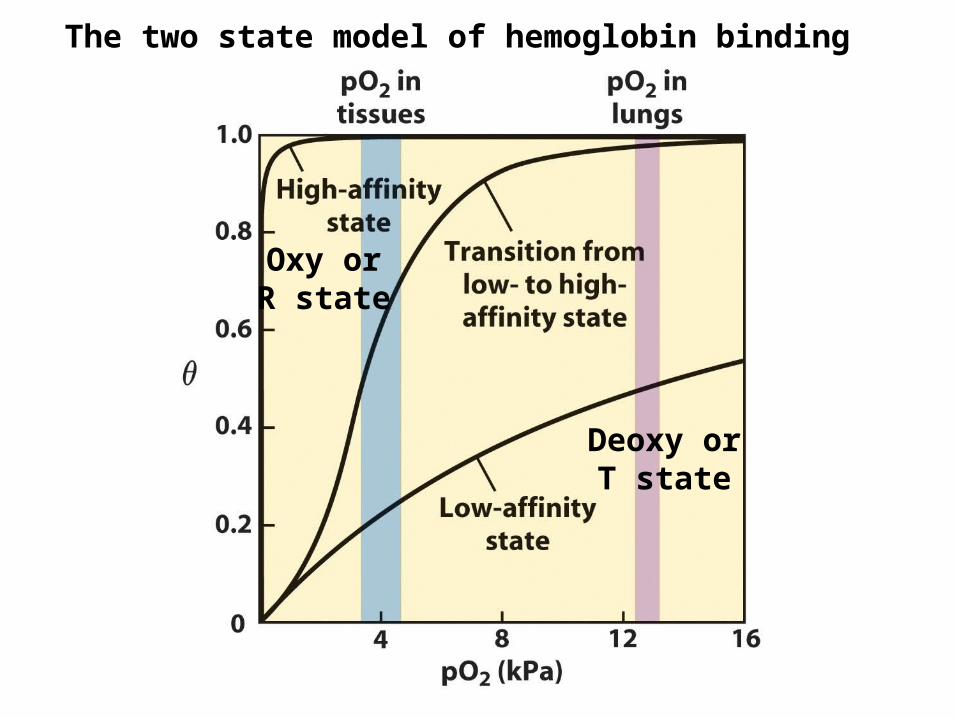
Deoxy orT state
Oxy orR state
The two state model of hemoglobin binding
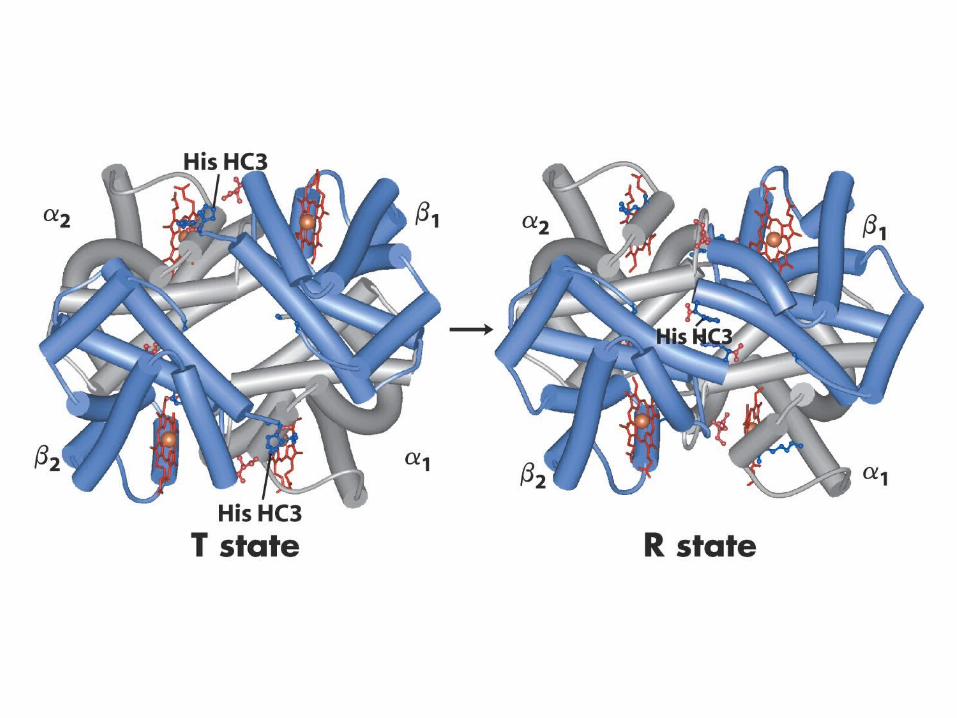

Major Structural differences upon oxidation of hemoglobin
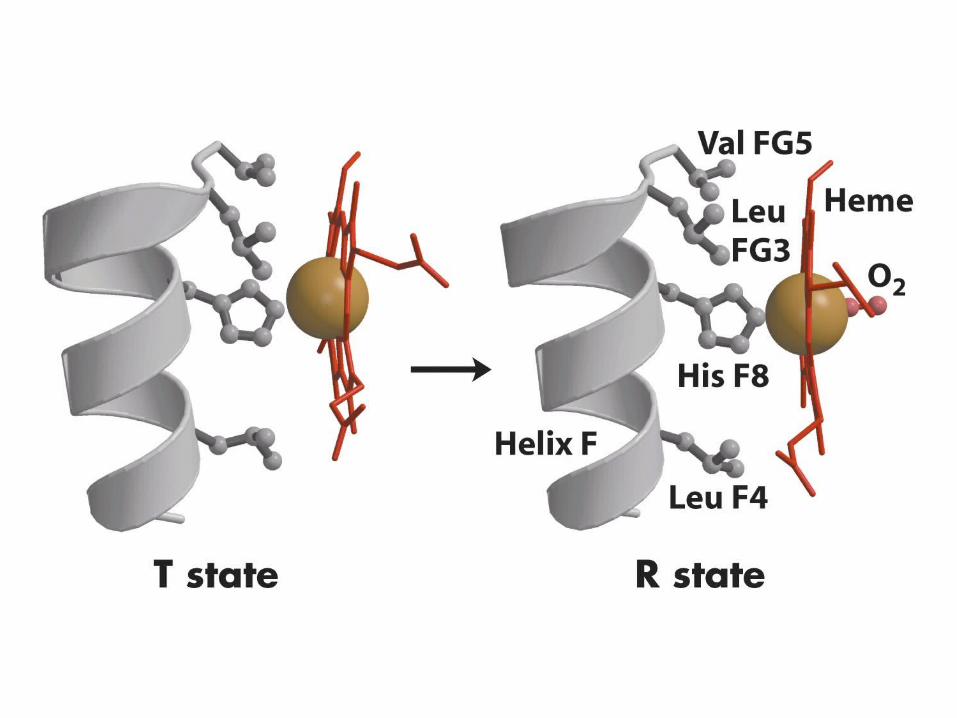
Fe moves from 0.55Å out of the heme plane to 0.22Å out of the plane
Extensive 1-1 contacts unchangedMinimal 1-2 contact altered by as
much as 6 Å
15º offcenter rotation of the protomers

High spin Oh Fe2+
xy
xz yz
x2-y2
z2
Increased radius
Low spin Oh Fe3+
xy
xz yz
x2-y2
z2
Decreased radius
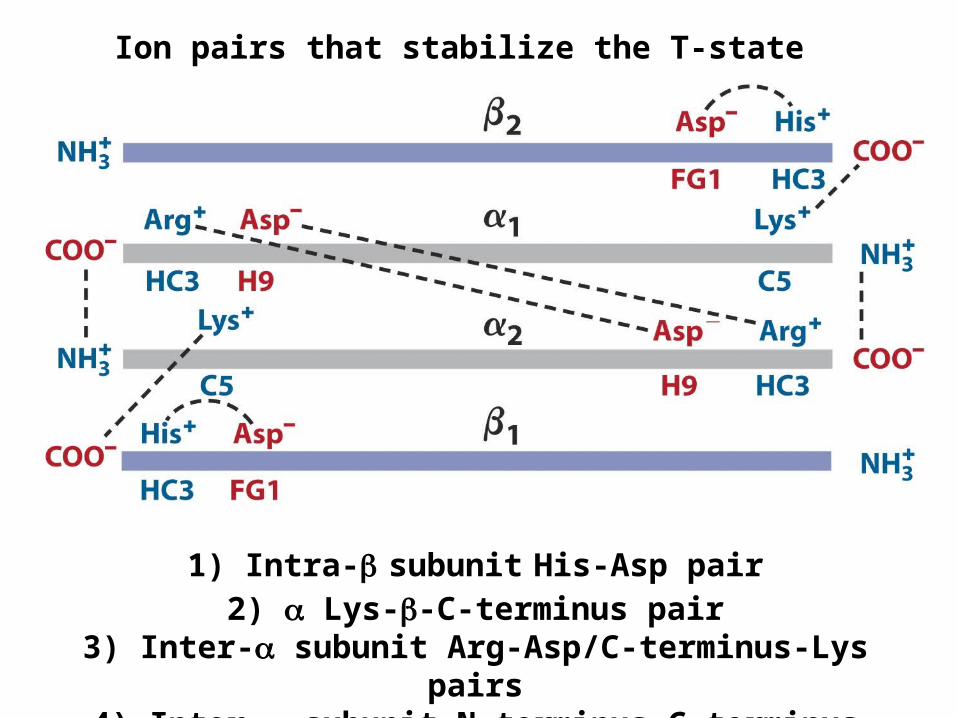
1) Intra- subunit His-Asp pair2) Lys--C-terminus pair
3) Inter- subunit Arg-Asp/C-terminus-Lys pairs4) Inter- subunit N-terminus-C-terminus pair
Ion pairs that stabilize the T-state
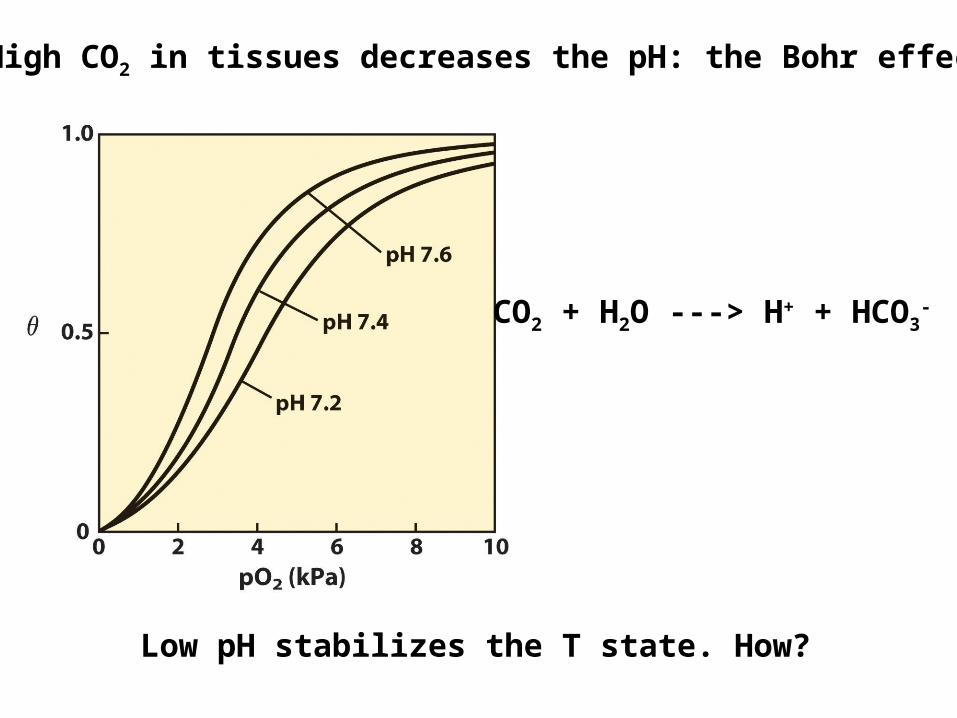
Low pH stabilizes the T state. How?
High CO2 in tissues decreases the pH: the Bohr effect
CO2 + H2O ---> H+ + HCO3-
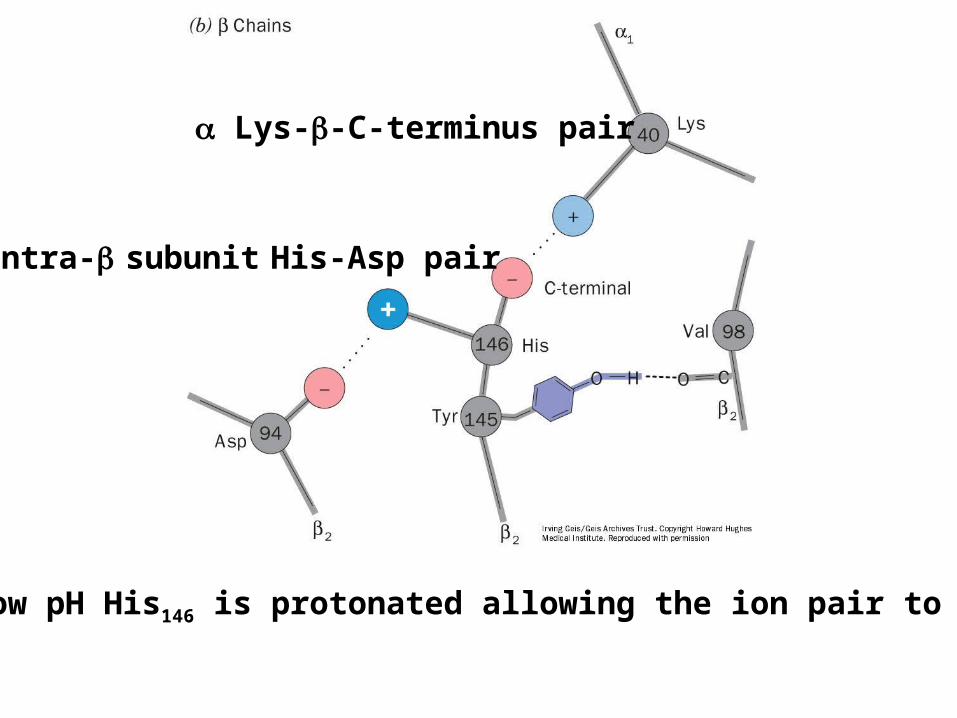
Lys--C-terminus pair
Intra- subunit His-Asp pair
At low pH His146 is protonated allowing the ion pair to form
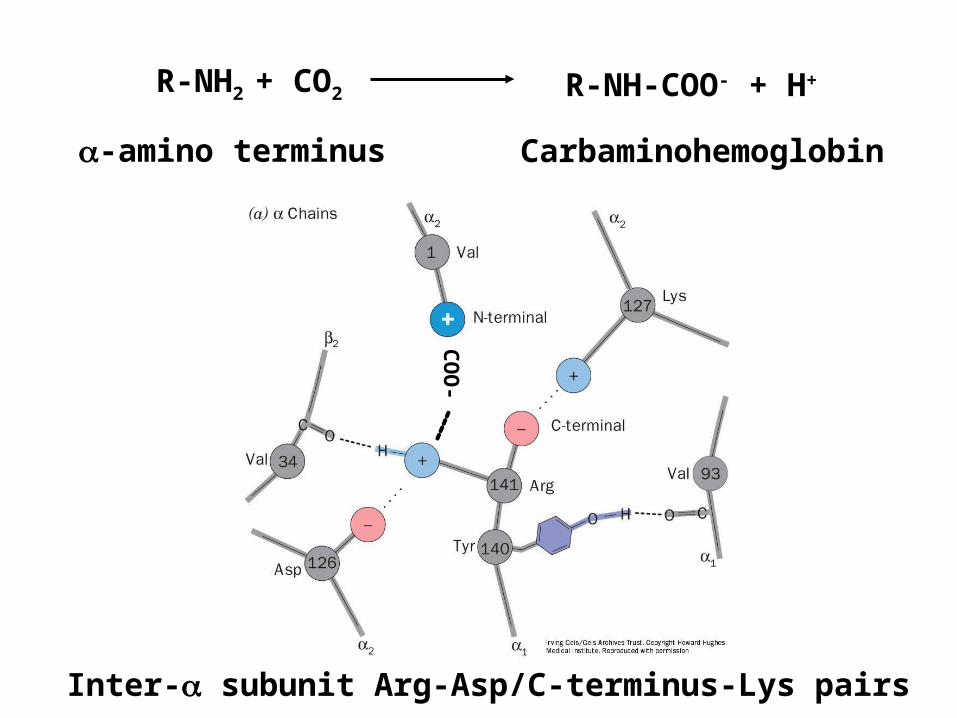
CO
O-
R-NH2 + CO2 R-NH-COO- + H+
Carbaminohemoglobin-amino terminus
Inter- subunit Arg-Asp/C-terminus-Lys pairs
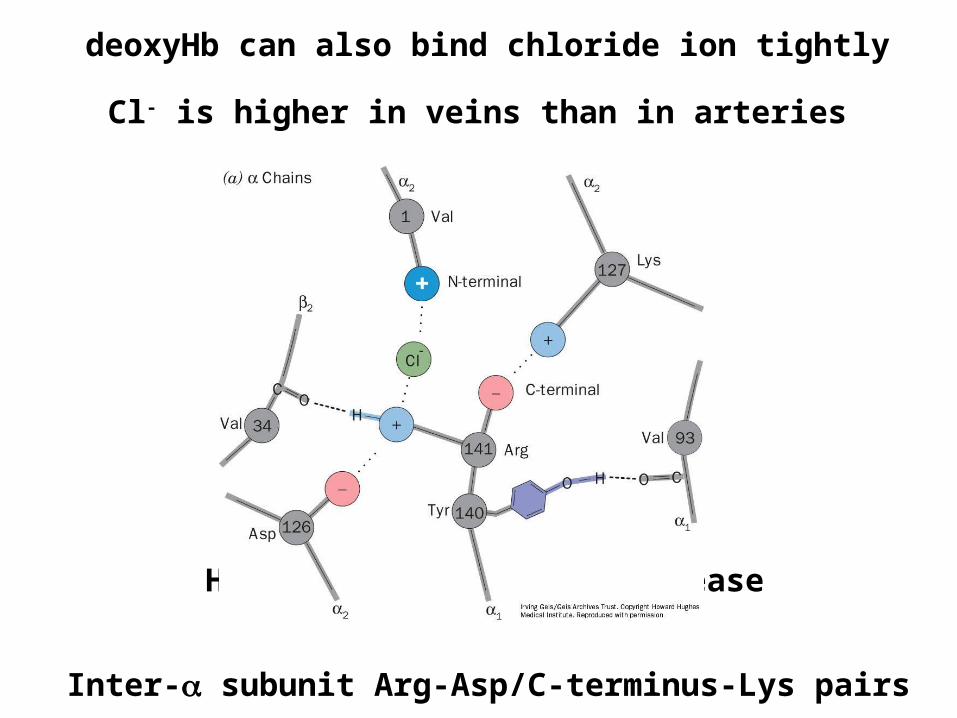
deoxyHb can also bind chloride ion tightly
High Cl- will cause O2 release
Cl- is higher in veins than in arteries
Inter- subunit Arg-Asp/C-terminus-Lys pairs

Thus the T state is stabilized by:Low pH
High CO2
High Cl-
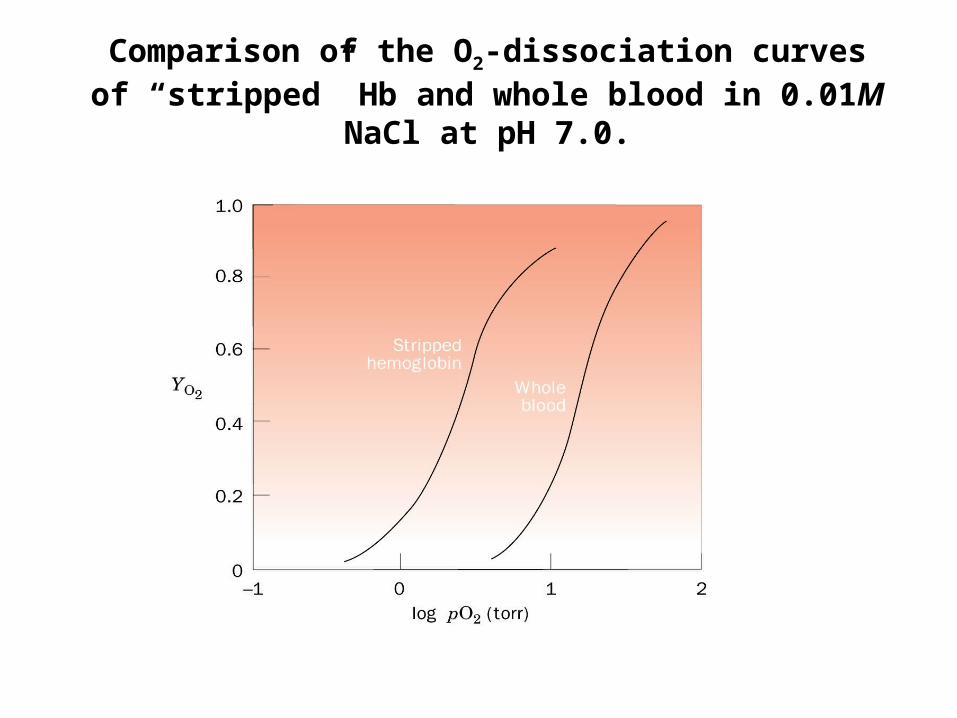
Comparison of the O2-dissociation curves of “stripped” Hb and whole blood in 0.01M NaCl at pH
7.0.
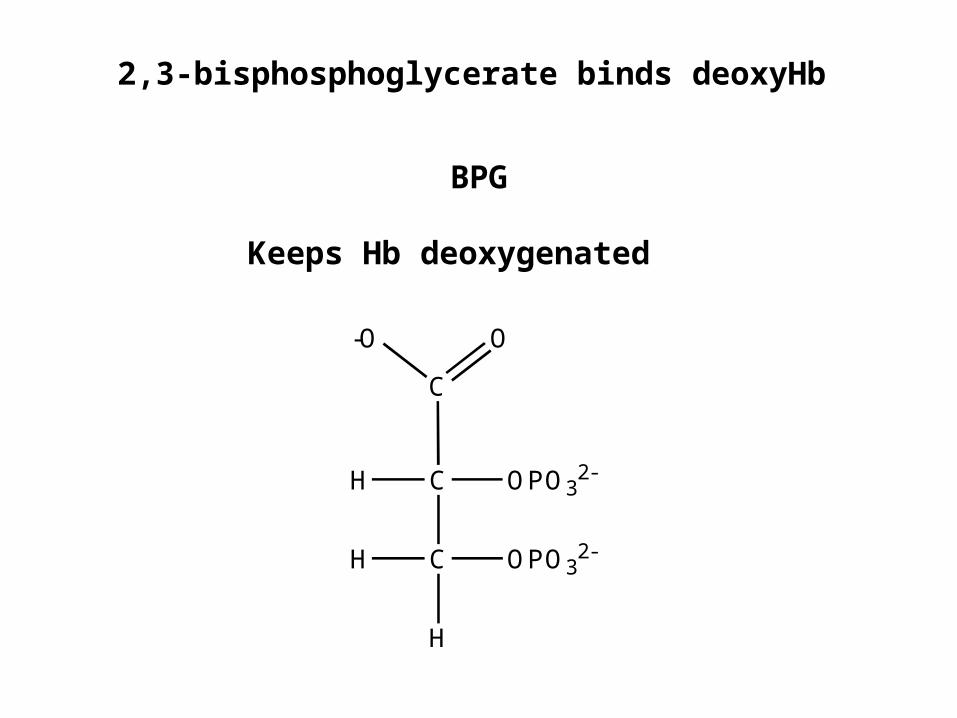
2,3-bisphosphoglycerate binds deoxyHb
BPG
Keeps Hb deoxygenated
-O
C
O
C
C
H
H OPO32-
H OPO32-
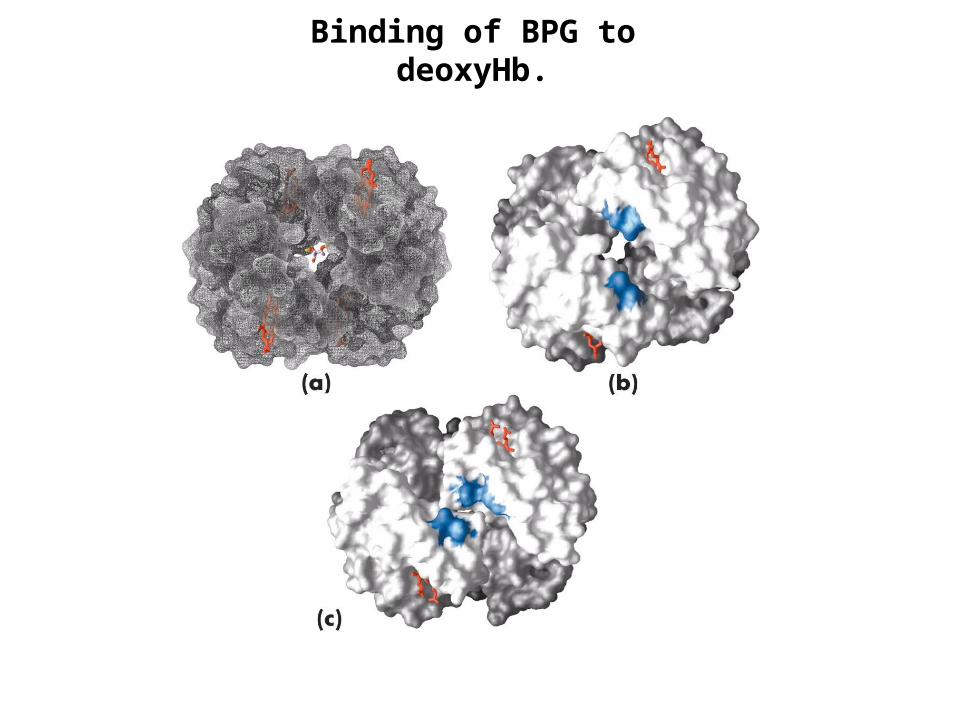
Binding of BPG to deoxyHb.
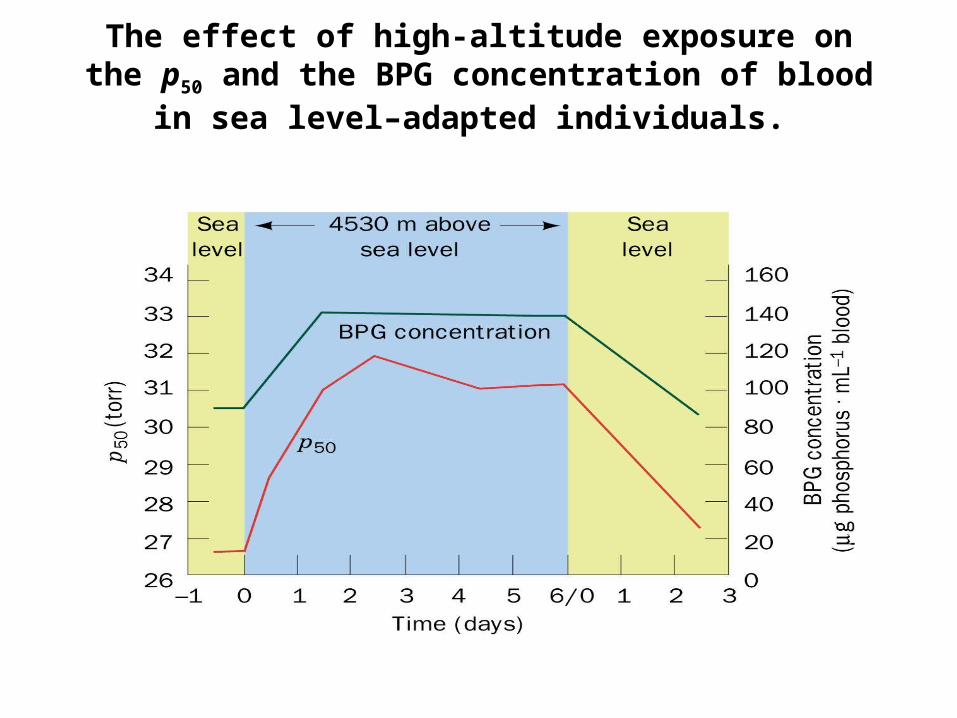
The effect of high-altitude exposure on the p50 and the BPG concentration of blood in sea level–
adapted individuals.
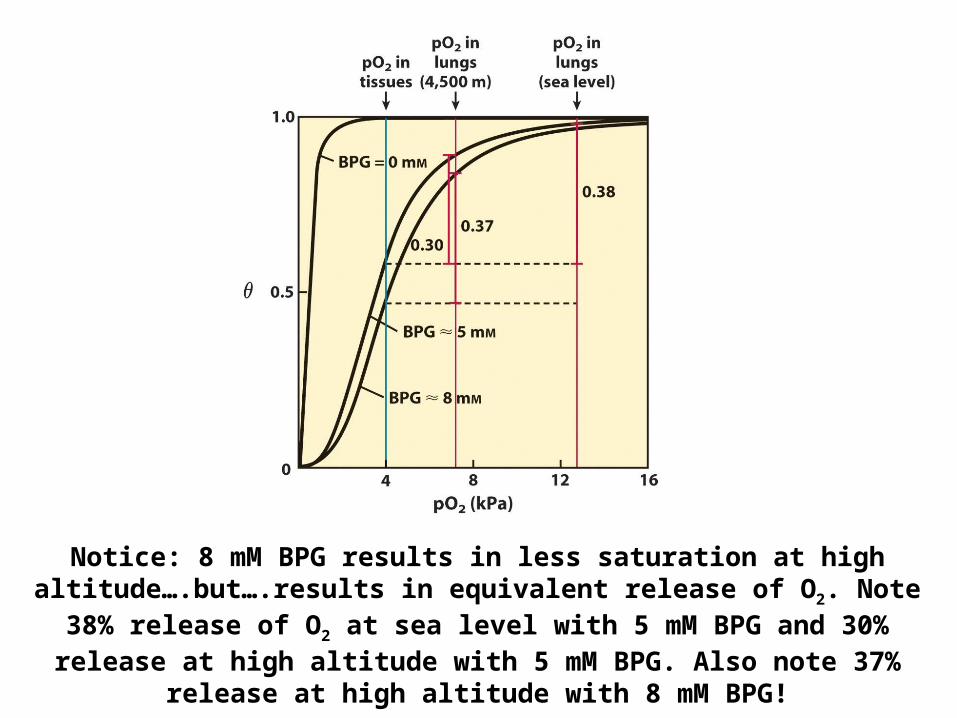
Notice: 8 mM BPG results in less saturation at high altitude….but….results in equivalent release of O2. Note 38% release of O2 at sea level with 5 mM BPG and 30% release at high altitude with 5
mM BPG. Also note 37% release at high altitude with 8 mM BPG!
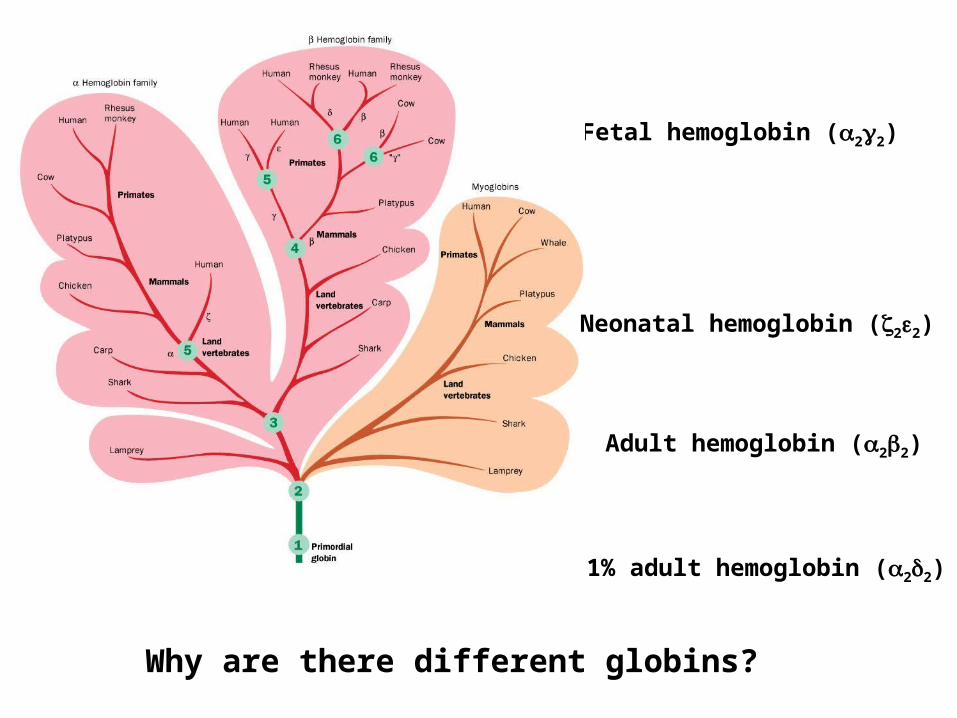
Fetal hemoglobin (22)
Adult hemoglobin (22)
Neonatal hemoglobin (22)
1% adult hemoglobin (22)
Why are there different globins?
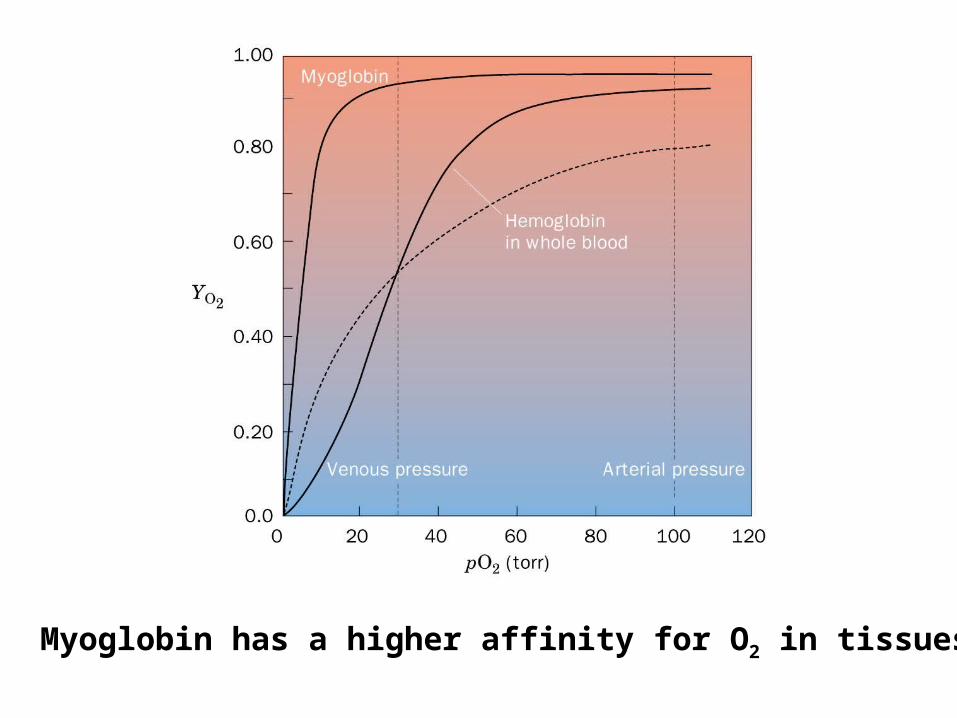
Myoglobin has a higher affinity for O2 in tissues
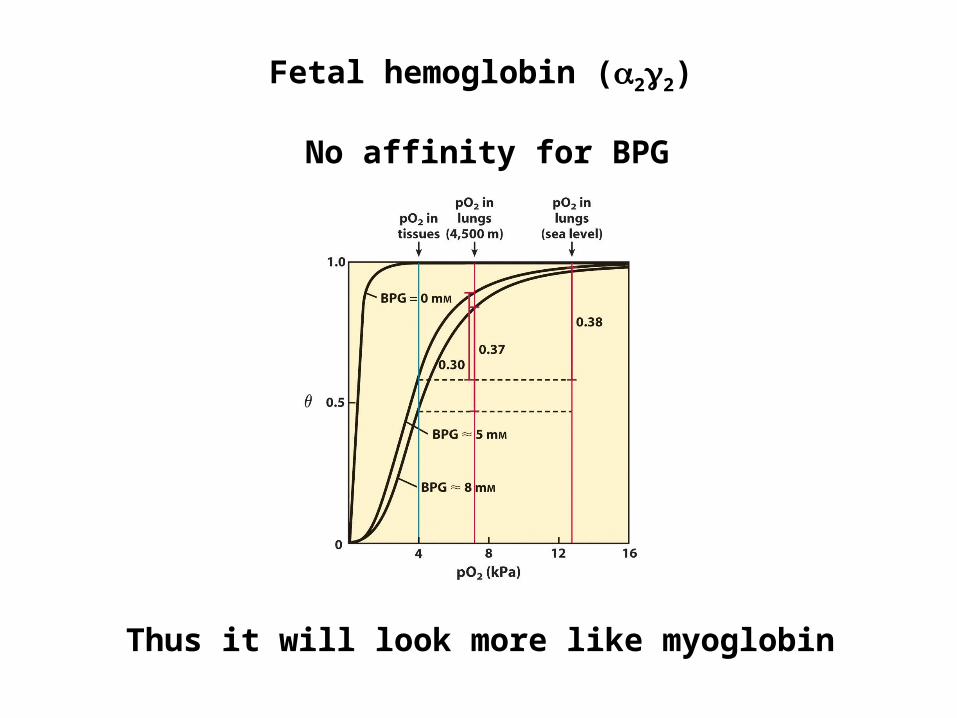
Fetal hemoglobin (22)
No affinity for BPG
Thus it will look more like myoglobin





























